Embed Size (px)
Citation preview

Immunology 1980 41 303
The induction ofchronic antige-antibody complex disease in selectively bred miceproducing either high or low affinity antibody to protein antigens
MADELEINE E. DEVEY & M. W. STEWARD Immunology Unit, Department of Medical Microbiology,London School ofHygiene and Tropical Medicine, Keppel Street, London
Acceptedfor publication 17 April 1980
Summary. Mice selectively bred to produce either highor low affinity antibody to protein antigens were in-jected daily with bovine serum albumin (BSA) andstudied for the development of chronic antigen-anti-body complex disease. After 41-44 injections of BSA,evidence of circulating antigen-antibody complexesand renal localization of complexes was obtained inboth lines. However, low affinity mice had signifi-cantly higher levels of circulating complexes than highaffinity mice. Impairment of renal function was seen inlow affinity mice but not in high affinity mice and therewas significantly more complex deposition in the glo-meruli of low compared to high affinity mice. Further-more, the pattern of localization of complexes waspredominantly mesangial in high affinity mice but inlow affinity mice the deposition was both mesangialand in the glomerular basement membrane. The roleof antibody affinity in the induction of chronic anti-gen-antibody complex disease is discussed in the lightof these results.
Correspondence: Dr M. E. Devey, Immunology Unit,Department of Medical Microbiology, London School ofHygiene and Tropical Medicine, Keppel Street, LondonWCIE 7HT.0019-2805/80/1000-0303$02.do(© 1980 Blackwell Scientific Publications
INTRODUCTION
There is increasing evidence that the genetically con-trolled production of low affinity antibody which ispoor at immune elimination of antigen is a contribu-tary factor in the development of chronic antigen-antibody complex disease (Soothill & Steward, 1971;Petty, Steward & Soothill, 1972; Steward, 1979). In therabbit model of chronic antigen-antibody complexdisease, the most severe renal lesions, characterized bydiffuse membranous proliferation, occur in animalsproducing non-precipitating antibodies (Pincus,Haberkern & Christian, 1968) of low affinity (Kuri-yama, 1973). Passive transfer to mice of preformedantigen-antibody complexes composed of rabbit anti-bodies of defined affinity has shown that complexescomposed of high affinity antibodies made over a widerange of antigen excess localize predominantly in themesangium whereas those composed of low affinityantibodies form predominantly subendothelial de-posits in the capillary loops (Koyama, Niwa, Shige-matsu, Taniguchi & Tada, 1978; Germuth, Rodriguez,Lorelle, Trump, Milano & Wise, 1979).Antibody affinity is a genetically controlled para-
meter ofthe immune response, which is controlled bymechanisms independent of those governing antibodylevels (Katz & Steward, 1975; Steward & Petty, 1976;Kim & Siskind, 1978). The development by selective
303

Madeleine E. Devey & M. W. Steward
breeding of two lines of TO mice producing similaramounts of antibody of different affinity to proteinantigens injected in saline (Katz & Steward, 1975) hasprovided an ideal model for the study of the role ofantibody affinity in chronic antigen-antibody complexdisease.
In a preliminary report from this laboratory (Stew-ard, 1979), experiments were described in which TOmice producing either high or low affinity antibodywere repeatedly injected with increasing amounts ofhuman serum albumin. In low affinity mice, there wasgreater renal localization of complexes demonstratedby immunofluorescence, more deaths from apparentrenal failure and more antibody was eluted from kid-ney homogenates of low than from high affinity mice.Furthermore, the pattern of renal localization ofcom-plexes was predominantly capillary in low affinity micebut predominantly mesangial in high affinity mice. Inthe work described here, we extend and confirm theseinitial observations using a larger number of animalsand fixed daily doses of antigen, and investigate therelationship between renal function, pattern of renallocalization ofcomplexes and the presence ofcirculat-ing antigen-antibody complexes in high and lowaffinity mice.
MATERIALS AND METHODS
MiceOne hundred age- and sex-matched generation 13 TOmice, selectively bred to produce either high or lowaffinity antibodies, were used.
InjectionsMice were divided into five groups and each groupreceived a daily i.p. injection of BSA in saline 6 or 7days a week. The antigen doses, which remained con-stant throughout the experiment, were 0-25, 0-5, 1-0and 1-5 mg. Control mice of each line were not in-jected. The animals were killed after they had receivedbetween 41 and 44 injections of BSA.
SeraSera taken at the end of the experiment, at least 24 hafter the last injection of BSA, were stored at - 700until tested.
Assessment ofrenalfunctionThe glomerular filtration rate (GFR) was assessedweekly by the measurement of the clearance of 5"Cr-
EDTA from the blood (Knight, Adams & Purves,1977). Results were expressed as the ratio of the tj5'Cr-EDTA for each experimental animal to a mean ofthe control values (x/c).
Measurement ofproteinuriaThe albumip content of urine samples (3 kul) was deter-mined by rocket immunoelectrophoresis using 3%anti-mouse serum albumin in 2% agarose. Knownconcentrations of mouse albumin were included oneach plate as standards and proteinuria was assessedby measurement of the precipitin peaks after stainingwith Coomassie brilliant blue.
Measurement ofantibodiesFree antibody in serum was measured by a doubleisotope ammonium sulphate precipitation methodusing 1251-BSA (Gaze, West & Steward, 1973). Anti-body binding in vivo was also assessed after an i.p.injection of 125I-BSA (0-3 ,uCi/mouse) followed bymeasurement of globulin-bound antigen in serum byprecipitation with 50% saturated ammonium sul-phate.
Measurement ofantigen-antibody complexesCirculating antigen-antibody complexes weremeasured using three solid-phase binding assays inparallel: the conglutinin binding assay (KBA); the Clqbinding assay (ClqBA); and the rheumatoid factorbinding assay (RFBA), which have been described indetail by Devey, Taylor & Steward (1980). Briefly,polystyrene tubes (LP3, Luckman Ltd) were coatedwith Clq, conglutinin or polyclonal rheumatoid fac-tor. For the ClqBA, sera were pre-incubated withEDTA. Duplicate serum samples were diluted in phos-phate-buffered saline containing 0-05% Tween 20 or,for the KBA, in veronal-buffered saline containingCa2+ and Mg2+ and 0.05% Tween 20. After incuba-tion and washing, complex-like material was esti-mated from the binding of affinity-purified1251-labelled anti-mouse immunoglobulin sera to thetubes. Results were expressed as the percentage in-crease of the binding of 1251-labelled anti-mouse im-munoglobulin in the test serum (mean of duplicates)over the mean of that binding in the control sera.Values that were two standard deviations or moreabove the mean of the control values were consideredpositive.
ImmunofluorescenceKidneys were snap frozen in n-hexane cooled inalcohol/dry ice. Four micron cryostat sections were
304

Chronic antigen-antibody complex disease
cut and stained directly with FITC-conjugated anti-sera to either mouse immunoglobulins, mousefIC//3IA or BSA (Nordic). Sections were examinedunder a Zeiss fluorescence photomicroscope and theintensity of fluorescence was scored on a 0 to 3 + scaleby an independent observer. The pattern of localiza-tion of the deposits was also determined. Photographswere taken using 400 ASA Ectachrome film (Kodak).
KneejointsKnee joints were decalcified in 10% formal-saline.Histological processing and examination were kindlycarried out by Professor R. R. A. Coombs and Dr G.Oldham (Division of Immunology, Department ofPathology, Cambridge University).
StatisticsResults were analysed by the Student's t test and by thechi-squared test.
RESULTS
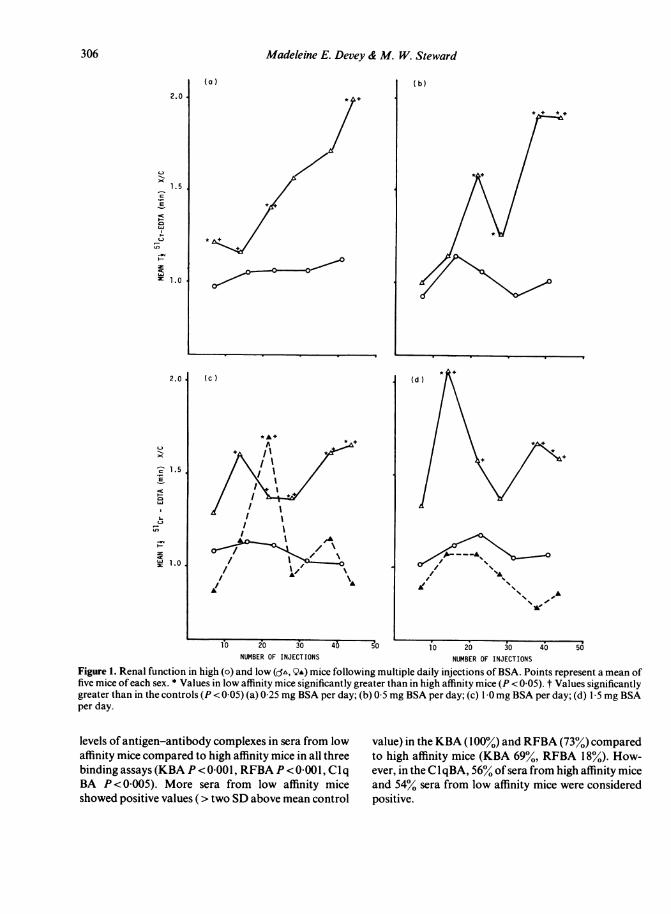
Renal function during chronic antigen administrationA marked decrease in glomerular filtration as assessedby the clearance of5'Cr-EDTA was seen in low affinitymice at all antigen doses, with the exception of femalemice receiving the highest antigen doses. Renal func-tion in high affinity mice however, did not differ signifi-cantly from the controls at any time during the experi-ment in any group. Since there were no significantdifferences in renal function between male and femalemice receiving 0 25 mg and 0-5 mg BSA per day, thevalues were therefore considered together (Fig. 1 a andb). Renal function was significantly impaired (shownby an increased tj for 51Cr-EDTA) in low affinity micecompared to both their controls and high affinity miceon day 7 (P<0-025), day 22 (P<005) and day 44(P<0 02). Although no low affinity mice died fromrenal failure in this experiment, we have subsequentlyshown that 100% mortality from renal failure occurredin low affinity female mice when daily injections of0 25 mg/day HSA were continued for a longer period(Reinhardt & Devey, unpublished observations). Inmice receiving 0-5 mg BSA per day (Fig. I b), the GFRwas again markedly impaired in both male and femalelow affinity mice compared to high affinity mice andthe controls. These differences were statistically sig-nificant on day 22 (P < 0-02), day 28 (P < 0 05), day 38(P= 0005) and on day 44 (P<0-02).
Following daily injections of 1 0 mg BSA there wasa significant decrease in GFR in low affinity male mice
but not in female mice (except at one point) and sothese values are plotted separately (Fig. Ic). The GFRwas significantly decreased in low affinity male micecompared to high affinity mice on days 28 (P < 0 005),38 (P<0 001) and 44 (P<0-02). Low affinity femalemice showed a decrease in GFR only on day 22(P=0 01). Similarly, in mice receiving 1[5 mg BSA perday there was no decrease in the GFR in low affinityfemale mice at any time (Fig. Id).Low affinity male mice, however, did show a signifi-
cant decrease in GFR compared to high affinity miceon days 14 (P < 0-02), 38 (P < 0 007) and 44 (P < 0 01).
ProteinuriaVery few mice showed any indication of severe pro-teinuria after 41-44 injections of BSA; of the 34 lowaffinity line mice tested, only 2 mice gave values ofover100 mg% mouse albumin (1 each in the 0 25 mg and 0 5mg dose groups) and a further 4 mice had values over10 mg%. In the high affinity line, 2 out of 36 mice hadvalues just over 10 mg%.
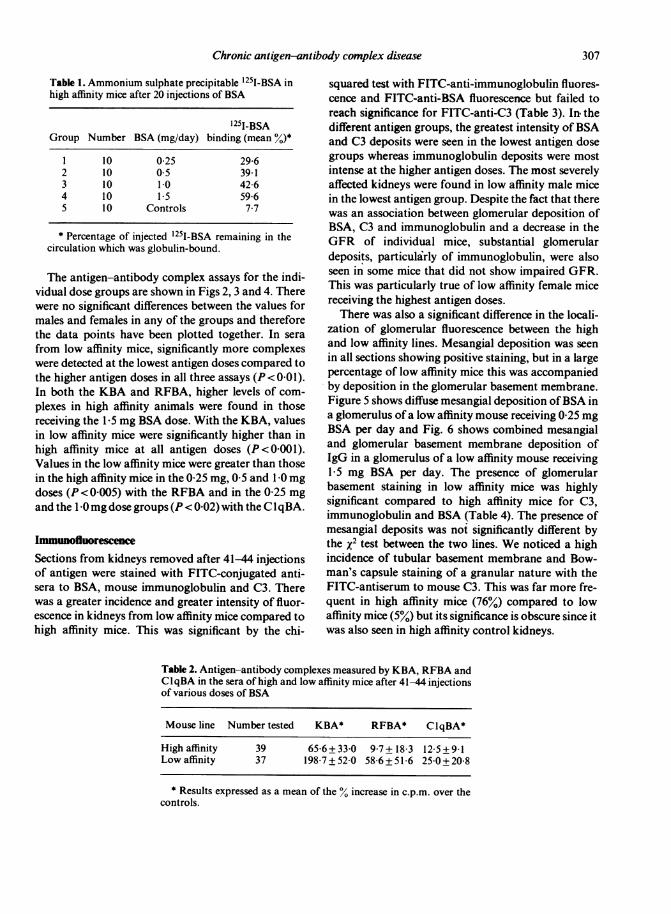
Antibody bindingThe absence of renal impairment in high affinity micemay have resulted from an absence of antigen-anti-body complexes because of tolerance and failure ofantibody synthesis. High affinity mice which hadreceived 20 daily injections of BSA were injected with'25I-BSA. Five hours later serum was obtained and theglobulin-bound antigen determined by ammoniumsulphate precipitation. As shown in Table 1, highaffinity mice in all antigen dose groups had demon-strable globulin binding at levels which were greatestin those mice receiving the highest dose of antigen.Furthermore, at the end of the experiment sera fromboth high and low affinity mice had similar levels offree antibody (data not shown).
Circulating antigen-antibody complexesSeventy-six serum samples from high and low affinitymice from all antigen dose groups were assayed forcirculating antigen-antibody complexes using thesolid-phase KBA, ClqBa and RFBA in parallel. Ineach test, with the exception of the RFBA for highaffinity mice, binding of '251-labelled antibody tomouse immunoglobulin was significantly increased inthe experimental sera of both high and low affinityanimals as compared to the controls (P < 0 00 1). Table2 shows that overall there were significantly higher
305
306 Madeleine E. Devey & M. W. Steward
2.
-1.5._.E-C
LU
TI.-
2:1.0I
.
t-
-C
:E1
(a ) ( b)
(d )
,~~~~~~%e.~~~~~~~~~~'C1 0 20 30 40 00 1i0 20 30 40 50
NUMBER OF INJECTIONS NUMBER OF INJECTIONS
Figure 1. Renal function in high (o) and low (o', QA) mice following multiple daily injections of BSA. Points represent a mean offive mice of each sex. * Values in low affinity mice significantly greater than in high affinity mice (P < 0-05). t Values significantlygreater than in the controls (P < 0 05) (a) 0 25 mg BSA per day; (b) 0 5 mg BSA per day; (c) 1 0 mg BSA per day; (d) 1 5 mg BSAper day.
levels of antigen-antibody complexes in sera from lowaffinity mice compared to high affinity mice in all threebinding assays (KBA P < 0 001, RFBA P < 0-001, ClqBA P< 0 005). More sera from low affinity miceshowed positive values ( > two SD above mean control
value) in the KBA (100%) and RFBA (73%) comparedto high affinity mice (KBA 69%, RFBA 18%). How-ever, in the C qBA, 56% of sera from high affinity miceand 54% sera from low affinity mice were consideredpositive.
(c )
/
*A
Chronic antigen-antibody complex disease
Table 1. Ammonium sulphate precipitable 1251-BSA inhigh affinity mice after 20 injections of BSA
1251-BSAGroup Number BSA (mg/day) binding (mean %)*
I 10 0 25 29-62 10 05 39 13 10 1 0 4264 10 15 5965 10 Controls 7*7
* Percentage of injected 1251-BSA remaining in thecirculation which was globulin-bound.
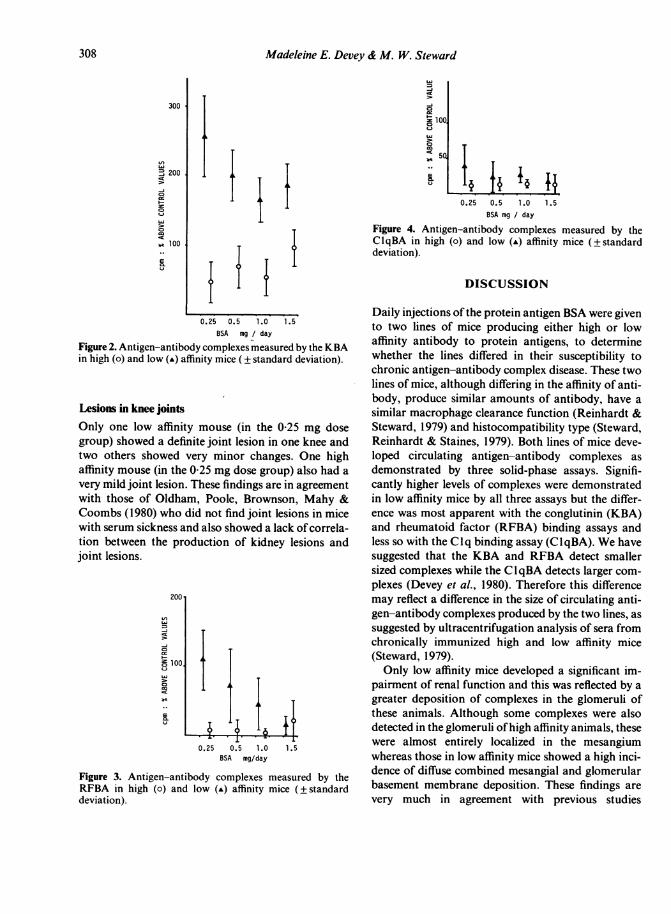
The antigen-antibody complex assays for the indi-vidual dose groups are shown in Figs 2, 3 and 4. Therewere no significant differences between the values formales and females in any of the groups and thereforethe data points have been plotted together. In serafrom low affinity mice, significantly more complexeswere detected at the lowest antigen doses compared tothe higher antigen doses in all three assays (P <00 1).In both the KBA and RFBA, higher levels of com-plexes in high affinity animals were found in thosereceiving the 1[5 mg BSA dose. With the KBA, valuesin low affinity mice were significantly higher than inhigh affinity mice at all antigen doses (P<0001).Values in the low affinity mice were greater than thosein the high affinity mice in the 0 25 mg, 0-5 and I 0 mgdoses (P <0-005) with the RFBA and in the 0 25 mgand the 1 0mg dose groups (P < 0-02) with the CIqBA.
Immunofluorescence
Sections from kidneys removed after 41-44 injectionsof antigen were stained with FITC-conjugated anti-sera to BSA, mouse immunoglobulin and C3. Therewas a greater incidence and greater intensity of fluor-escence in kidneys from low affinity mice compared tohigh affinity mice. This was significant by the chi-
squared test with FITC-anti-immunoglobulin fluores-cence and FITC-anti-BSA fluorescence but failed toreach significance for FITC-anti-C3 (Table 3). In, thedifferent antigen groups, the greatest intensity ofBSAand C3 deposits were seen in the lowest antigen dosegroups whereas immunoglobulin deposits were mostintense at the higher antigen doses. The most severelyaffected kidneys were found in low affinity male micein the lowest antigen group. Despite the fact that therewas an association between glomerular deposition ofBSA, C3 and immunoglobulin and a decrease in theGFR of individual mice, substantial glomerulardeposits, particularly of immunoglobulin, were alsoseen in some mice that did not show impaired GFR.This was particularly true of low affinity female micereceiving the highest antigen doses.There was also a significant difference in the locali-
zation of glomerular fluorescence between the highand low affinity lines. Mesangial deposition was seenin all sections showing positive staining, but in a largepercentage of low affinity mice this was accompaniedby deposition in the glomerular basement membrane.Figure 5 shows diffuse mesangial deposition ofBSA ina glomerulus of a low affinity mouse receiving 0-25 mgBSA per day and Fig. 6 shows combined mesangialand glomerular basement membrane deposition ofIgG in a glomerulus of a low affinity mouse receiving15 mg BSA per day. The presence of glomerularbasement staining in low affinity mice was highlysignificant compared to high affinity mice for C3,immunoglobulin and BSA (Table 4). The presence ofmesangial deposits was not significantly different bythe x2 test between the two lines. We noticed a highincidence of tubular basement membrane and Bow-man's capsule staining of a granular nature with theFITC-antiserum to mouse C3. This was far more fre-quent in high affinity mice (76%) compared to lowaffinity mice (5%) but its significance is obscure since itwas also seen in high affinity control kidneys.
Table 2. Antigen-antibody complexes measured by KBA, RFBA andClqBA in the sera of high and low affinity mice after 41-44 injectionsof various doses of BSA
Mouse line Number tested KBA* RFBA* ClqBA*
High affinity 39 65-6+33-0 9-7+18 3 12 5+9-1Low affinity 37 198-7+52-0 58 6+51 6 25 0+20 8
* Results expressed as a mean of the % increase in c.p.m. over thecontrols.
307
Madeleine E. Devey & M. W. Steward
300
'J 200
-j
co
al 100..
II
LO
=D00
-ix
5..
IIiil
0.25 0.5 1.0 1.5BSA mg / day
Figure 2. Antigen-antibody complexes measured by the KBAin high (o) and low (A) affinity mice (± standard deviation).
Lesions in knee jointsOnly one low affinity mouse (in the 0-25 mg dosegroup) showed a definite joint lesion in one knee andtwo others showed very minor changes. One highaffinity mouse (in the 0 25 mg dose group) also had avery mild joint lesion. These findings are in agreementwith those of Oldham, Poole, Brownson, Mahy &Coombs (1980) who did not find joint lesions in micewith serum sickness and also showed a lack of correla-tion between the production of kidney lesions andjoint lesions.
J)
co
.D
0.25 0.5 1.0BSA mg/day
1.5
Figure 3. Antigen-antibody complexes measured by theRFBA in high (o) and low (-) affinity mice (±standarddeviation).
0.25 0.5 1.0 1.5BSA mg / day
Figure 4. Antigen-antibody complexes measured by theClqBA in high (o) and low (A) affinity mice (±standarddeviation).
DISCUSSION
Daily injections of the protein antigen BSA were givento two lines of mice producing either high or lowaffinity antibody to protein antigens, to determinewhether the lines differed in their susceptibility tochronic antigen-antibody complex disease. These twolines of mice, although differing in the affinity of anti-body, produce similar amounts of antibody, have asimilar macrophage clearance function (Reinhardt &Steward, 1979) and histocompatibility type (Steward,Reinhardt & Staines, 1979). Both lines of mice deve-loped circulating antigen-antibody complexes asdemonstrated by three solid-phase assays. Signifi-cantly higher levels of complexes were demonstratedin low affinity mice by all three assays but the differ-ence was most apparent with the conglutinin (KBA)and rheumatoid factor (RFBA) binding assays andless so with the Clq binding assay (ClqBA). We havesuggested that the KBA and RFBA detect smallersized complexes while the ClqBA detects larger com-plexes (Devey et al., 1980). Therefore this differencemay reflect a difference in the size of circulating anti-gen-antibody complexes produced by the two lines, assuggested by ultracentrifugation analysis of sera fromchronically immunized high and low affinity mice(Steward, 1979).Only low affinity mice developed a significant im-
pairment of renal function and this was reflected by agreater deposition of complexes in the glomeruli ofthese animals. Although some complexes were alsodetected in the glomeruli ofhigh affinity animals, thesewere almost entirely localized in the mesangiumwhereas those in low affinity mice showed a high inci-dence of diffuse combined mesangial and glomerularbasement membrane deposition. These findings arevery much in agreement with previous studies
308
Chronic antigen-antibody complex disease
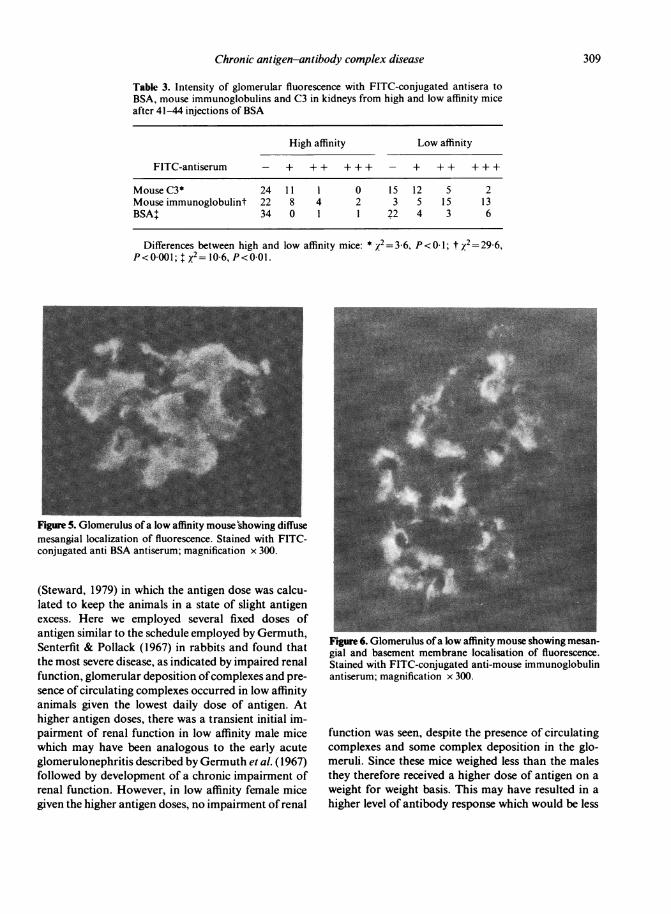
Table 3. Intensity of glomerular fluorescence with FITC-conjugated antisera toBSA, mouse immunoglobulins and C3 in kidneys from high and low affinity miceafter 41-44 injections of BSA
High affinity Low affinity
FITC-antiserum - + + + + + + - + + + + + +
Mouse C3* 24 11 1 0 15 12 5 2Mouse immunoglobulint 22 8 4 2 3 5 15 13BSA+ 34 0 1 1 22 4 3 6
Differences between high and low affinity mice: * X2=3-6,P<0 001; $ x2= 10-6, P<0 01.
Figure 5. Glomerulus of a low affinity mouse'showing diffusemesangial localization of fluorescence. Stained with FITC-conjugated anti BSA antiserum; magnification x 300.
(Steward, 1979) in which the antigen dose was calcu-lated to keep the animals in a state of slight antigenexcess. Here we employed several fixed doses ofantigen similar to the schedule employed by Germuth,Senterfit & Pollack (1967) in rabbits and found thatthe most severe disease, as indicated by impaired renalfunction, glomerular deposition ofcomplexes and pre-sence of circulating complexes occurred in low affinityanimals given the lowest daily dose of antigen. Athigher antigen doses, there was a transient initial im-pairment of renal function in low affinity male micewhich may have been analogous to the early acuteglomerulonephritis described by Germuth et al. (1967)followed by development of a chronic impairment ofrenal function. However, in low affinity female micegiven the higher antigen doses, no impairment of renal
P<O 1; t 2=29-6,
Figure 6. Glomerulus ofa low affinity mouse showing mesan-gial and basement membrane localisation of fluorescence.Stained with FITC-conjugated anti-mouse immunoglobulinantiserum; magnification x 300.
function was seen, despite the presence of circulatingcomplexes and some complex deposition in the glo-meruli. Since these mice weighed less than the malesthey therefore received a higher dose of antigen on aweight for weight basis. This may have resulted in ahigher level of antibody response which would be less
309

310 Madeleine E. Devey & M. W. Steward
Table 4. Localization ofantigen-antibody complexes inthe glomeruli of high and low affinity mice
% of positive sectionsshowing GBM staining
FITC-conjugatedantibody to: High affinity Low affinity
Mouse C3* 0 26Mouse immunoglobulint 13 41BSA$ 0 90
Differences between the two lines: * x2= 5 7, P < 0 05;t x2= 10-2, P<0 01; t x2= 10 6, P<0-01.
likely to cause chronic glomerulonephritis (Germuthet al., 1967). As there were no deaths from renalfailure, limited proteinuria and the absence of severemembranous proliferative glomerulonephritis wehave concluded that, after only 41-44 injections ofantigen, the chronic disease we have described was at arelatively early stage. We have subsequently shownthat daily injection of antigen continued for a longerperiod of time led to a high incidence of deaths in lowaffinity mice accompanied by glomerular deposition ofcomplexes which was predominantly in the glomerularbasement membrane. In this experiment, even after100 antigen injections, high affinity mice did not showany impairment ofrenal function and their glomerularcomplex deposition was predominantly mesangial(Reinhardt & Devey, unpublished observations).Our findings and those of others (Kuriyama, 1973;
Koyama et al., 1978; Germuth et al., 1979; Steward,1979) provide very strong evidence that the produc-tion of antibody of low affinity is a most importantfactor in the development of chronic glomeruloneph-ritis. Low affinity antibody may be more likely to formthe small soluble complexes (Class 1) described byGermuth & Rodriguez (1973) which will localize onthe glomerular basement membrane to cause the mostsevere form of chronic glomerulonephritis. Highaffinity antibody which may form larger, more stablecomplexes (Class II and III) would be more readilycleared by the reticuloendothelial system or by theglomerular mesangium. Modulation of the affinity ofthe antibody response may result in abrogation of thechronic glomerulonephritis we describe here in lowaffinity antibody producing mice.
ACKNOWLEDGMENTS
We thank Professor P. F. Kohler for his help in inter-pretation of the immunofluorescence studies and MissKatherine Bleasdale for excellent technical assistance.Financial support by the Wellcome Trust is gratefullyacknowledged.
REFERENCES
DEVEY M.E., TAYLOR J. & STEWARD M.W. (1980) Measure-ment of antigen-antibody complexes in mouse sera byconglutinin, Clq and rheumatoid factor solid phase bind-ing assays. J. immunol. Meth. 34, 191.
GAZE S., WEST N.J. & STEWARD M.W. (1973) The use of adouble isotope method in the determination of antibodyaffinity. J. Inmunol. Meth. 3, 357.
GERMUTH F.G. & RODRIGUEZ E. (1973) Immunopathology ofthe Renal Glomerulus. Little, Brown and Company,Boston.
GERMUTH F.G., SENTERFIT L.B. & POLLACK A.D. (1967)Immune complex disease. I. Experimental acute andchronic glomerulonephritis. Johns Hopkins med. J. 120,225.
GERMUTH F.G., RODRIGUEZ E., LORELLE C.A., TRUMP E.I.,MILANO L. & WISE O'L. (1979) Passive immune complexglomerulonephritis in mice: models for various lesionsfound in human disease. I. High avidity complexes andmesangio proliferative glomerulonephritis. II. Low avi-dity complexes and diffuse proliferative glomeruloneph-ritis with subepithelial deposits. Lab. Invest. 41, 360.
KATZ F.E. & STEWARD M.W. (1975) The genetic control ofantibody affinity in mice. Immunology, 29, 543.
KIM Y.T. & SISKIND G.W. (1978) Studies on the control ofantibody synthesis. XII. Genetic influences on antibodyaffinity. Immunology, 34, 669.
KNIGHT J.G., ADAMS D.D. & PURVES H.D. (1977) The gene-

Chronic antigen-antibody complex disease 311
tic contribution of the NZB mouse to the renal disease ofthe NZB x NZW hybrid. Clin. exp. Immunol. 28,352.
KOYAMA A., NIWA Y., SHIGEMATSU H., TANIGUCHI M. &TADA T. (1978) Studies on passive serum sickness. II.Factors determining localization of antigen-antibodycomplexes in the murine renal glomerulus. Lab. Invest. 38,253.
KURIYAMA T. (1973) Chronic glomerulonephritis induced byprolonged immunization in the rabbit. Lab. Invest. 28,224.
OLDHAM G., POOLE A.R., BROWNSON J.M., MAHY B.W.J. &CooMBS R.R.A. (1980) Experiments in mice and rats onthe induction ofjoint lesions following injections ofserumand in chronic virus infection. Int. Archs Allergy appl.Immunol. 62, 213.
PETTY R.E., STEWARD M.W. & SOOTHILL J.F. (1972) Theheterogeneity of antibody affinity in inbred mice and itspossible immunopathologic significance. Clin. exp. Im-munol. 12, 231.
PINcus T., HABERKERN R. & CHRISTIAN C.L. (1968) Experi-mental chronic glomerulitis. J. exp. Med. 127, 819.
REINHARDT M.C. & STEWARD M.W. (1979) Antibody affinityand clearance function studies in high and low antibodyaffinity mice. The effect of protein deficiency. Immuno-logy, 38, 735.
SOOTHILL J.F. & STEWARD M.W. (1971) The immunopatho-logical significance of the heterogeneity of antibodyaffinity. Clin. exp. Immunol. 9,193.
STEWARD M.W. (1979) Chronic immune complex disease inmice: the role of antibody affinity. Clin. exp. Immunol. 38,414.
STEWARD M.W. & PETTY R.E. (1976) Evidence for the geneticcontrol of antibody affinity from breeding studies withinbred mouse strains producing high and low affinityantibody. Immunology, 30, 789.
STEWARD M.W., REINHARDT M.C. & STAINES N.A. (1979)The genetic control of antibody affinity. Evidence frombreeding studies with mice selectively bred for either highor low affinity production. Immunology, 37, 697.







![[Monoclonal antibodies in clinical immunology]](https://img.pdfslide.net/doc/110x75/63375eff20d9c9602f0b4441/monoclonal-antibodies-in-clinical-immunology.jpg)





















