Embed Size (px)
Citation preview

Direct somatic embryo formation (without intervening callus) from garlic clove basal tissue was induced
in which the influence of plant growth regulators (PGRs) on various explants was examined. Medium
added with 2.0 mg/l 6-benzylaminopurine (BAP) and 0.5 mg/l 2,4-dichlorophenoxyacetic acid (2,4-D)
were the most effective PGR combination for somatic embryo induction. It induced embryos directly in
85.5% of the basal clove explant. Callus induction was also obtained from other parts of explant and 2.0
mg/l 2,4-D induced callusing in 86.5% of the inoculated explants. Protein, amino acid and alliin content
were measured in callus and in embryos. Somatic embryos had more soluble protein and free amino acid
compared to callus. HPTLC analysis revealed that alliin was significantly high in somatic embryos com-
pared to undifferentiated callus tissue; the content was even more in older embryos. The present study of
Allium indicates that the event of morphogenetic development including in vitro embryogeny can effec-
tively be analysed by monitoring the changes of biochemical profiles.
Keywords: Allium sativum – alliin – callus – HPTLC – somatic embryo
INTRODUCTION
Garlic (Allium sativum) is an important culinary and medicinal plant used worldwide.
It shows several biological activities such as antibiotic, antitumour, antiatheroscle-
rotic [6, 7], cholesterol-lowering effect [46] and it also prevents cardiovascular dis-
orders [37]. Propagation of garlic is mainly accomplished by vegetative methods
Acta Biologica Hungarica 60 (4), pp. 441–454 (2009)
DOI: 10.1556/ABiol.60.2009.4.10
0236-5383/$ 20.00 © 2009 Akadémiai Kiadó, Budapest
IMPROVED ALLIIN YIELD IN SOMATIC EMBRYOS
OF ALLIUM SATIVUM L.
(CV. YAMUNA SAFED) AS ANALYZED BY HPTLC
S. A. NASIM, A. MUJIB,* K. RASHMI,
F. SAMAR, A. JUNAID and MAHMOODUZZAFAR
Department of Botany, Cellular Differentiation and Molecular Genetics Section, Hamdard University,
New Delhi 110 062, India
(Received: November 17, 2008; accepted: January 26, 2009)
* Corresponding author; e-mail: [email protected]
Abbrevations: 2,4-dichlorophenoxyacetic acid (2,4-D); naphthalene acetic acid (NAA); 6-benzy-
laminopurine (BAP); 6-furfuryl aminopurine (KIN); plant growth regulator (PGR); somatic embryogen-
esis (SE); Least Significant Difference (LSD), High Pressure Thin Layer Chromatography (HPTLC).

442 S. A. NASIM et al.
Acta Biologica Hungarica 60, 2009
which demonstrate a low coefficient of multiplication [32]; therefore it takes many
years to produce sufficient number of seed bulbs for practical cultivation of new elite
variety [31]. Similarly, the crop improvement by cross fertilization is limited as gar-
lic shows sexual incompatibility [27]. In Allium, callus culture and in vitro morpho-
genesis have been achieved from various plant parts [3, 30, 39, 42] but the rate of
multiplication and the number of plantlets regenerated per explant were not always
significantly high [38]. Plant regeneration through somatic embryogenesis is rare but
is not uncommon in Allium [42]. It has several advantages over organogenesis and
appears to be the most promising technique for fast propagation of plants [17]. The
developmental protocols to establish embryonic cultures with synchronous embryo
forming ability may able to eliminate many of the problems associated with zygotic
embryo development.
Garlic, like many other members of Alliums, contains high organic sulphur com-
pounds in the form of alkylcysteine sulphoxides and γ-glutamyl peptides. On tissue
damage and with alliinase enzyme’s activity, the alkyl cysteine sulphoxide releases
compounds that give unique Allium’s odour and flavour. In garlic, the major flavour
precursor is alliin while in onion it is isoalliin. Alliin has been proposed to be syn-
thesized by two pathways, in one, serine combines with allyl thiol source to produce
allyl cysteine which on oxidation produce alliin while in the other, glutathione
derived cysteine unites with an allyl source, forming an intermediate allyl glu-
tathione, later converts into allyl cysteine which finally give rise to allin [23].
An early biochemical marker(s) for the identification of embryonic potency would
be of immense value in studying efficient plant regeneration. It has earlier been noted
that the morphogenetic pattern is associated with specific biochemical changes that
can be used as markers [45]. Several quantitative and qualitative studies were con-
ducted and their developmental pattern were analysed during somatic embryogene-
sis and organogenesis events [21, 24]. Dave and Batra [10] studied the metabolic fac-
tors which promote to induce somatic embryos in culture and the embryogenic tis-
sues were compared with non-embryogenic callus. Earlier, biochemical differences
between embryonic and non-embryonic callus cultures with respect to polypeptide
pattern were also investigated in rice [8]. But, there are only a few biochemical
reports that discussed events during differentiation in medicinal plants [18]. In this
communication, a simple high frequency direct somatic embryogenic system is
reported from basal part of clove in Allium sativum cv. Yamuna Safed in which we
investigated the role of BAP and 2,4-D in embryogenesis. Attention has also been
paid to identify the biochemical differences that existed between callus and embryo-
genic tissues/embryos in Allium sativum during plant regeneration.

MATERIALS AND METHODS
Plant material
Local Indian garlic (Allium sativum L. c.v Yamuna safed, voucher specimen IC-
375117, deposited at NBPGR, New Delhi) a moderate maturing variety, was used as
experimental material. It was procured from National Horticultural Research and
Development Foundation, Nasik (Maharashtra). Bulbs were harvested from 4–5
months old plants and stored at 15 °C, and individual clove of bulb was used as
explant.
Explant preparation
Healthy garlic cloves were selected and washed under running water for 1 h, surface
sterilized with 0.1% HgCl2 for 15 min and 70% ethanol for 30 sec and washed sev-
eral times with sterilized distilled water. The clove was divided into basal, middle
and tip part; and inoculated in MS [29] medium. Nearly 3–5 mm cloves were trans-
ferred to culture tubes containing 15 ml of solidified basal medium supplemented
with various concentrations of BAP and 2,4-D.
Medium and culture conditions
The MS medium was used that was added with various plant growth regulators,
PGRs such as 2,4-D, BAP and KIN (6-furfuryl amino purine). The medium pH was
adjusted to 5.6 prior to autoclaving at 121 °C for 20 min. All the cultures were incu-
bated at 25 ± 2 °C in a culture room under 16-h photoperiod provided by cool white
fluorescent light at intensity of 40 Wm–2.
The frequency of callusing, i.e. the number of explants that produced callus was
counted after six weeks of incubation. Similarly, the frequency of globular somatic
embryos and the mean number of embryos per explant were recorded after two
months of incubation. Each treatment had at least 10 explants with 5 replicates.
Values were represented as mean ± standard error at 5% significance level.
Biochemical analyses
Estimation of protein
Bradford method [5] was used to estimate the protein content, 0.5 g tissue was
ground in prechilled mortar and pestle with 1.0 ml (0.1M) phosphate buffer (pH 7.0)
on ice and centrifuged at 5,000 rpm for 10 min at 4 °C. TCA (0.5 ml) was added and
the sample was again centrifuged at 5,000 rpm for 10 min. The supernatant was dis-
Plant growth regulators and somatic embryogenesis 443
Acta Biologica Hungarica 60, 2009

444 S. A. NASIM et al.
Acta Biologica Hungarica 60, 2009
carded and the pellet was dissolved in 1.0 ml of 0.1N NaOH after washing with dou-
ble-distilled water. After adding 5.0 ml of Bradford reagent, the optical density was
measured at 595 nm.
Estimation of free amino acids
Free amino acids content was estimated by the method of Lee and Takahashi [24]. In
brief, 0.1 g tissue was incubated overnight in 70% ethanol followed by washing with
double-distilled water. Then, 1.5 ml of 55% glycerol and 0.5 ml ninhydrin solution
were added, boiled at 100 °C for 20 min and cooled down. The final volume was
made up to 6 ml with double-distilled water and the optical density was measured at
570 nm.
Quantification of alliin
Extraction of garlic material
Arnault et al. [1] method was used for the extraction of garlic. One gram of garlic in
the form of dry powder was extracted at room temperature using 10 ml methanol –
water (80 : 20, v/v) plus 0.05% formic acid at pH < 3.
Quantification of alliin by HPTLC
The content of alliin in in vitro cultures was estimated using alliin (Sigma Chemical,
MI, USA) as standard.
Standard Alliin: One mg/ml solution of standard alliin as prepared in methanol
and 1 ml of this solution was diluted to 10 ml with methanol to get 100 µg/ml solu-
tion, these solutions were used for application on TLC plate for preparation of stan-
dard plot. The UV spectrum of standard alliin solution in methanol (50 µg/ml) was
recorded using UV spectrophotometer (UV-1601, Shimadzu Japan) for authentica-
tion. The λmax was matched with standard alliin.
HPTLC instrument and conditions
The samples were spotted in the form of bands (width 4 mm) with Camag microlitre
syringe on precoated silica gel aluminium plate 60F-254 (20 cm × 10 cm × 0.2 mm
thickness, E. Merck, Germany) using a Camag Linomat V (Switzerland). A constant
application rate of 80 nl/s was employed and space between two bands was 5.2 mm.
The slit dimension was kept a 4 mm × 0.1 mm; and a 20 mm/sec scanning speed was
employed. The mobile phase consisted of n-butanol : methanol : ammonia (8 : 1 : 1.5,
v/v). Linear ascending development was carried out in twin through glass chamber,

saturated with mobile phase. The optimum chamber saturation time for mobile phase
was 30 min at room temperature. The length of chromatogram run was 65 mm. TLC
plates were dried with air current provided by an air dryer. Densitometric scanning
was performed on Camag TLC scanner IV (absorbance mode 510 nm) after spraying
with anisaldehyde sulphuric acid and dried at 100C. Deuterium and tungsten lamps
were used as radiation sources.
Calibration curve of alliin
Different volumes of standard solution (0.5 – 6 µl) were spotted in triplicate on TLC
plate to obtain 50–1000 ng per spot of alliin. The data of peak area vs. alliin con-
centration were treated by linear least square regression and the regression equation
thus obtained from standard curve, was used to estimate alliin in different samples.
Quantification of alliin
Sample (5 µl) of in vitro cultures (calli and embryos) was applied in triplicate on
TLC plate. The alliin yield was quantified by using regression equation of calibration
curve.
Statistical analysis
All data are expressed as mean ± standard error. Mean number of somatic embryos
per explant and mean number of explant showing callus were determined using
ANOVA followed by LSD test at P= 0.05.
RESULTS AND DISCUSSION
Explant type and effect of PGRs on callusing and embryogenesis
Various sections of garlic clove (basal, middle and tip part) were inoculated in MS
added with 2,4-D and BAP. Of the several implanted explants, the tip and the middle
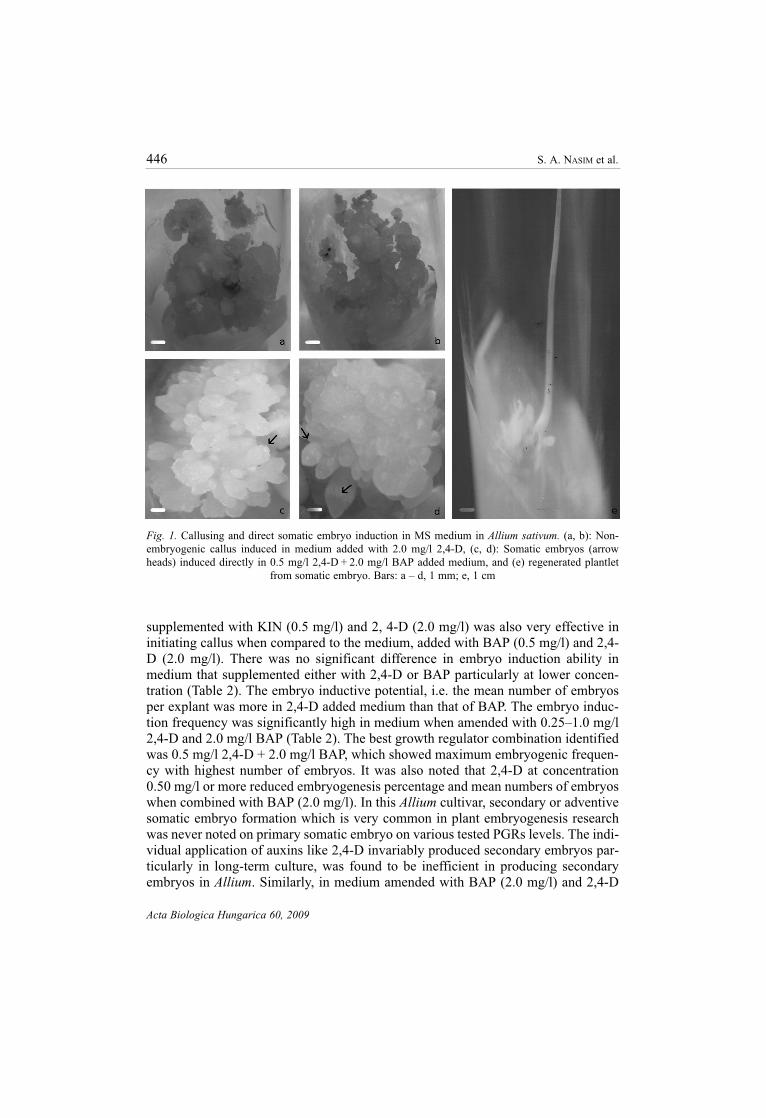
part of the clove developed callus (Fig. 1a, b) but did not produce embryos in cul-
ture. High frequency somatic embryos were only observed at the basal part of the
clove (Fig. 1c, d) which proliferated and produced more number of embryos on sub-
sequent passages.
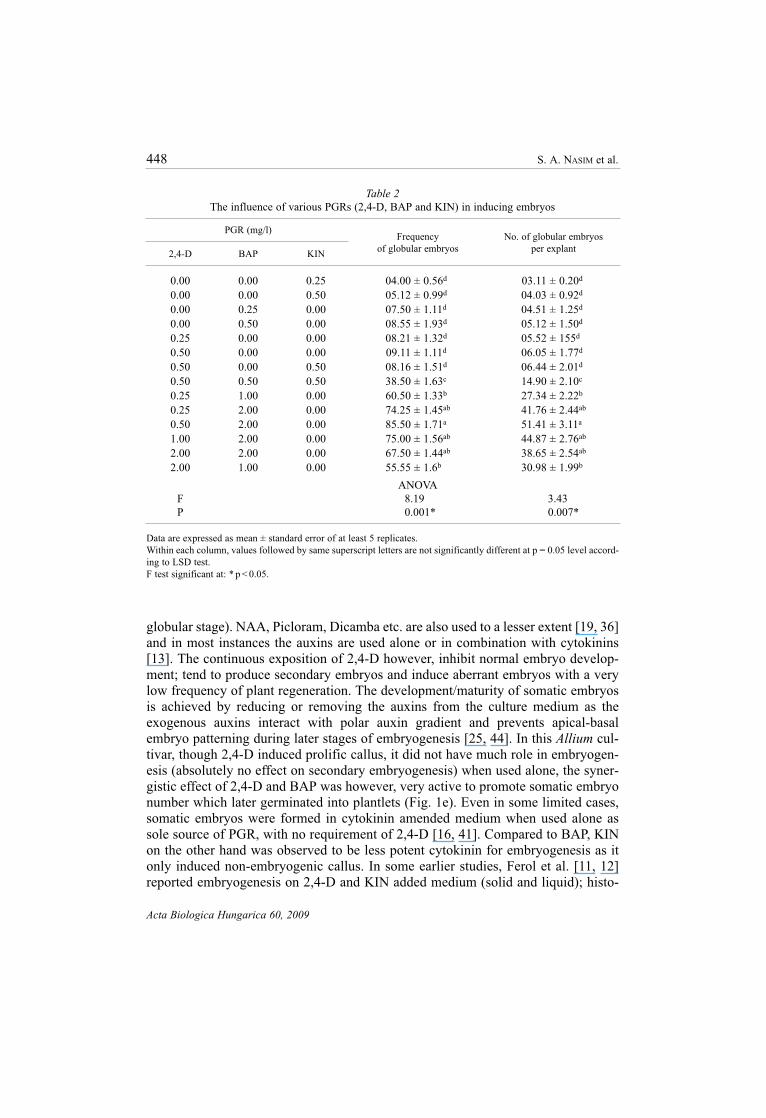
The role of different concentrations of 2,4-D, BAP and KIN in producing callus
and to induce embryos was presented in Tables 1 and 2. It was observed from the
tables that 2,4-D at 2.0 mg/l was the most effective concentration for callus forma-
tion and percent callus induction was least on BAP added medium (Table 1). Medium
Plant growth regulators and somatic embryogenesis 445
Acta Biologica Hungarica 60, 2009
446 S. A. NASIM et al.
Acta Biologica Hungarica 60, 2009
supplemented with KIN (0.5 mg/l) and 2, 4-D (2.0 mg/l) was also very effective in
initiating callus when compared to the medium, added with BAP (0.5 mg/l) and 2,4-
D (2.0 mg/l). There was no significant difference in embryo induction ability in
medium that supplemented either with 2,4-D or BAP particularly at lower concen-
tration (Table 2). The embryo inductive potential, i.e. the mean number of embryos
per explant was more in 2,4-D added medium than that of BAP. The embryo induc-
tion frequency was significantly high in medium when amended with 0.25–1.0 mg/l
2,4-D and 2.0 mg/l BAP (Table 2). The best growth regulator combination identified
was 0.5 mg/l 2,4-D + 2.0 mg/l BAP, which showed maximum embryogenic frequen-
cy with highest number of embryos. It was also noted that 2,4-D at concentration
0.50 mg/l or more reduced embryogenesis percentage and mean numbers of embryos
when combined with BAP (2.0 mg/l). In this Allium cultivar, secondary or adventive
somatic embryo formation which is very common in plant embryogenesis research
was never noted on primary somatic embryo on various tested PGRs levels. The indi-
vidual application of auxins like 2,4-D invariably produced secondary embryos par-
ticularly in long-term culture, was found to be inefficient in producing secondary
embryos in Allium. Similarly, in medium amended with BAP (2.0 mg/l) and 2,4-D
Fig. 1. Callusing and direct somatic embryo induction in MS medium in Allium sativum. (a, b): Non-
embryogenic callus induced in medium added with 2.0 mg/l 2,4-D, (c, d): Somatic embryos (arrow
heads) induced directly in 0.5 mg/l 2,4-D + 2.0 mg/l BAP added medium, and (e) regenerated plantlet
from somatic embryo. Bars: a – d, 1 mm; e, 1 cm

(0.25–1.0 mg/l), where primary somatic embryos occurred more frequently, no sec-
ondary embryo was formed in this very medium in Allium.In this present investigation, different parts of the explant (basal, middle and tip of
clove) were used in which callus was induced from middle and tip part of the clove
while basal clove directly formed embryos; we also noted variable callus yield in dif-
ferent parts of the clove explant (data not shown). The PGRs such as 2,4-D, BAP and
KIN were active but the callusing responses varied from one PGR to other. Similar
observation of differential requirement was previously observed in other groups of
plants and this may be due to different physiological gradient, photosynthetic reserve
differences, endogenous PGRs level etc. that existed in parts (proximal and distal) of
the same clove explant [20]. In the present investigation of Allium, we observed that
the calli from two or more sources were different not only morphologically; their
morphogenetic response was also not the same. In callus developed from the basal
part, cells acquired embryogenic competence, embryos were induced, later germi-
nated into plantlets, this embryogeny is very similar to zygotic embryo development,
while in callus induced from middle or tip part of the clove this embryogenic signal
is altogether absent. The type and concentration of auxins employed is critical for the
induction and development of somatic embryos [14]. The presence of 2,4-D in par-
ticular has been found to be necessary for the induction of embryo (perhaps up to
Plant growth regulators and somatic embryogenesis 447
Acta Biologica Hungarica 60, 2009
Table 1
Effect of various concentrations of PGRs on callusing
PGR (mg/l)
% Callus induction
2,4-D BAP KIN
0.00 0.00 0.25 45.00 ± 15.00c
0.00 0.00 0.50 51.10 ± 12.50bc
0.00 0.25 0.00 19.00 ± 5.10d
0.00 0.50 0.00 21.50 ± 8.00d
0.25 0.00 0.00 42.80 ± 11.50c
0.50 0.00 0.00 61.00 ± 11.10b
0.50 0.50 0.50 23.20 ± 15.50cd
1.00 0.00 0.00 70.50 ± 15.30ab
2.00 0.00 0.00 86.50 ± 9.50a
3.00 1.00 0.00 59.20 ± 12.20b
2.00 0.50 0.00 48.20 ± 8.50c
2.00 0.00 0.50 75.40 ± 10.50ab
ANOVA
F 9.13
P 0.002*
Data are expressed as mean ± standard error of at least 5 replicates.
Within each column, values followed by same superscript letters are not signifi-
cantly different at p = 0.05 level according to LSD test.
F test significant at: *p < 0.05.
448 S. A. NASIM et al.
Acta Biologica Hungarica 60, 2009
globular stage). NAA, Picloram, Dicamba etc. are also used to a lesser extent [19, 36]
and in most instances the auxins are used alone or in combination with cytokinins
[13]. The continuous exposition of 2,4-D however, inhibit normal embryo develop-
ment; tend to produce secondary embryos and induce aberrant embryos with a very
low frequency of plant regeneration. The development/maturity of somatic embryos
is achieved by reducing or removing the auxins from the culture medium as the
exogenous auxins interact with polar auxin gradient and prevents apical-basal
embryo patterning during later stages of embryogenesis [25, 44]. In this Allium cul-
tivar, though 2,4-D induced prolific callus, it did not have much role in embryogen-
esis (absolutely no effect on secondary embryogenesis) when used alone, the syner-
gistic effect of 2,4-D and BAP was however, very active to promote somatic embryo
number which later germinated into plantlets (Fig. 1e). Even in some limited cases,
somatic embryos were formed in cytokinin amended medium when used alone as
sole source of PGR, with no requirement of 2,4-D [16, 41]. Compared to BAP, KIN
on the other hand was observed to be less potent cytokinin for embryogenesis as it
only induced non-embryogenic callus. In some earlier studies, Ferol et al. [11, 12]
reported embryogenesis on 2,4-D and KIN added medium (solid and liquid); histo-
Table 2
The influence of various PGRs (2,4-D, BAP and KIN) in inducing embryos
PGR (mg/l)Frequency No. of globular embryos
of globular embryos per explant2,4-D BAP KIN
0.00 0.00 0.25 04.00 ± 0.56d 03.11 ± 0.20d
0.00 0.00 0.50 05.12 ± 0.99d 04.03 ± 0.92d
0.00 0.25 0.00 07.50 ± 1.11d 04.51 ± 1.25d
0.00 0.50 0.00 08.55 ± 1.93d 05.12 ± 1.50d
0.25 0.00 0.00 08.21 ± 1.32d 05.52 ± 155d
0.50 0.00 0.00 09.11 ± 1.11d 06.05 ± 1.77d
0.50 0.00 0.50 08.16 ± 1.51d 06.44 ± 2.01d
0.50 0.50 0.50 38.50 ± 1.63c 14.90 ± 2.10c
0.25 1.00 0.00 60.50 ± 1.33b 27.34 ± 2.22b
0.25 2.00 0.00 74.25 ± 1.45ab 41.76 ± 2.44ab
0.50 2.00 0.00 85.50 ± 1.71a 51.41 ± 3.11a
1.00 2.00 0.00 75.00 ± 1.56ab 44.87 ± 2.76ab
2.00 2.00 0.00 67.50 ± 1.44ab 38.65 ± 2.54ab
2.00 1.00 0.00 55.55 ± 1.6b 30.98 ± 1.99b
ANOVA
F 8.19 3.43
P 0.001* 0.007*
Data are expressed as mean ± standard error of at least 5 replicates.
Within each column, values followed by same superscript letters are not significantly different at p = 0.05 level accord-
ing to LSD test.
F test significant at: * p < 0.05.
logical analysis was also conducted to confirm the somatic embryogenesis based
plant regeneration in Allium sativum.
Amino acid, protein and alliin content in callusand in somatic embryos
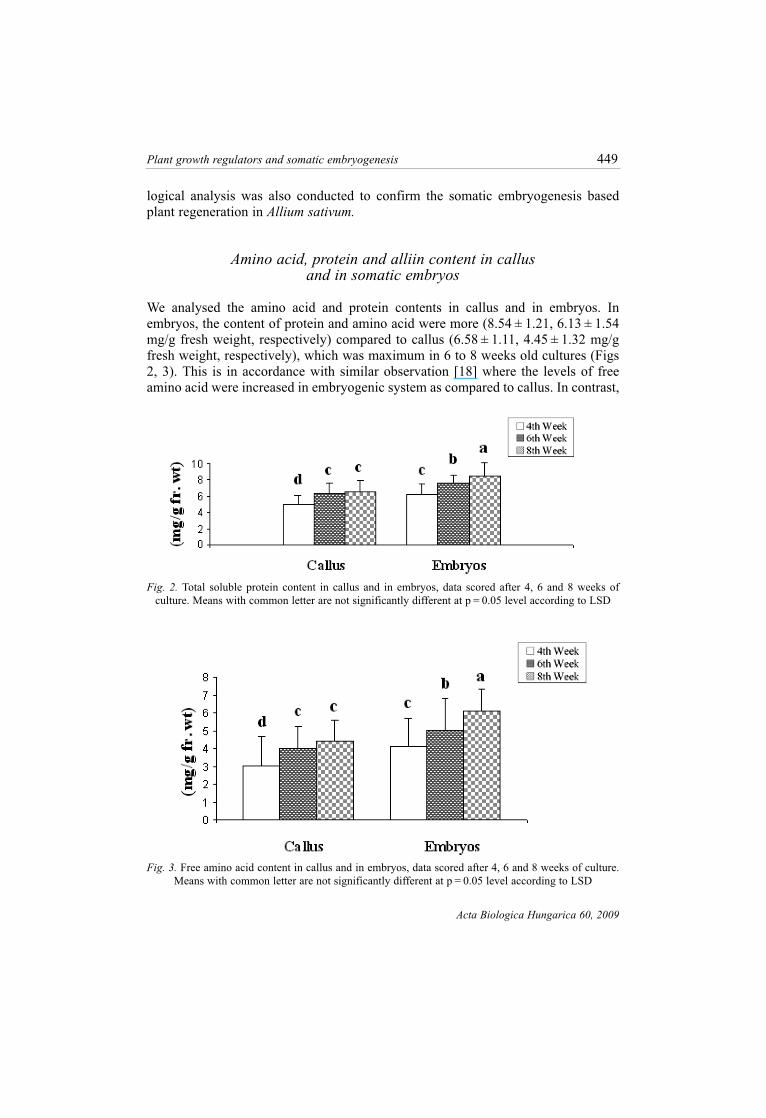
We analysed the amino acid and protein contents in callus and in embryos. In
embryos, the content of protein and amino acid were more (8.54 ± 1.21, 6.13 ± 1.54
mg/g fresh weight, respectively) compared to callus (6.58 ± 1.11, 4.45 ± 1.32 mg/g
fresh weight, respectively), which was maximum in 6 to 8 weeks old cultures (Figs
2, 3). This is in accordance with similar observation [18] where the levels of free
amino acid were increased in embryogenic system as compared to callus. In contrast,
Plant growth regulators and somatic embryogenesis 449
Acta Biologica Hungarica 60, 2009
Fig. 3. Free amino acid content in callus and in embryos, data scored after 4, 6 and 8 weeks of culture.
Means with common letter are not significantly different at p = 0.05 level according to LSD
Fig. 2. Total soluble protein content in callus and in embryos, data scored after 4, 6 and 8 weeks of
culture. Means with common letter are not significantly different at p = 0.05 level according to LSD
450 S. A. NASIM et al.
Acta Biologica Hungarica 60, 2009
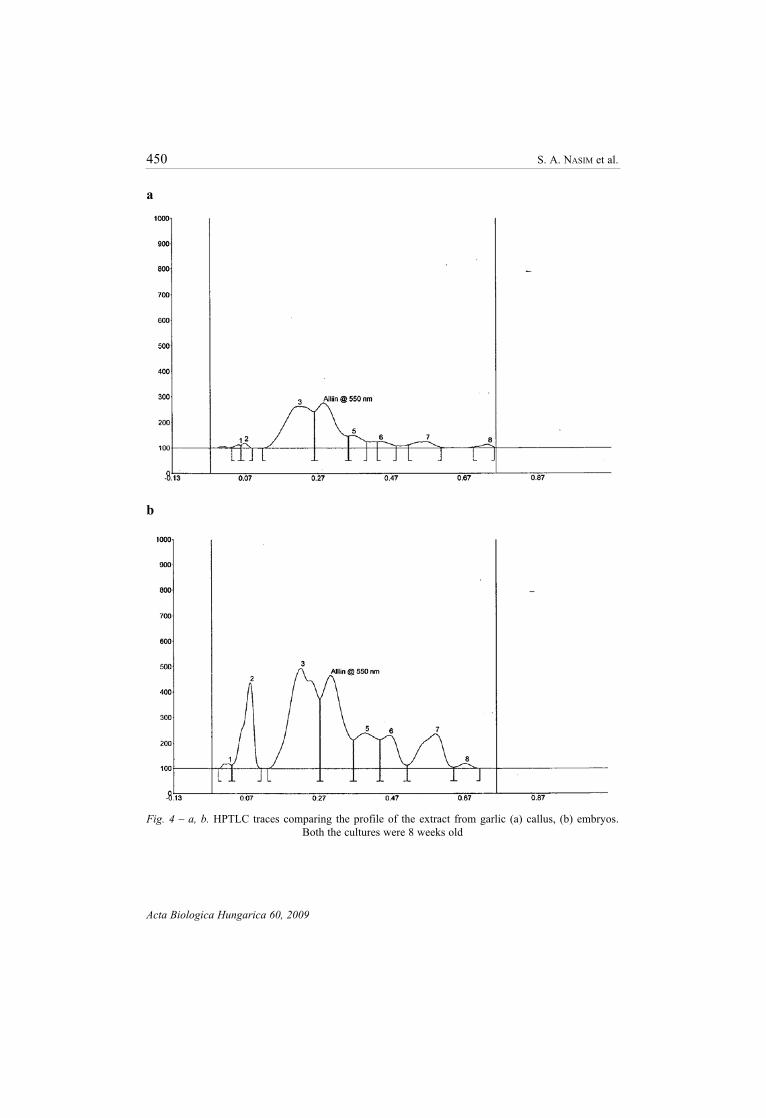
Fig. 4 – a, b. HPTLC traces comparing the profile of the extract from garlic (a) callus, (b) embryos.
Both the cultures were 8 weeks old
a
b
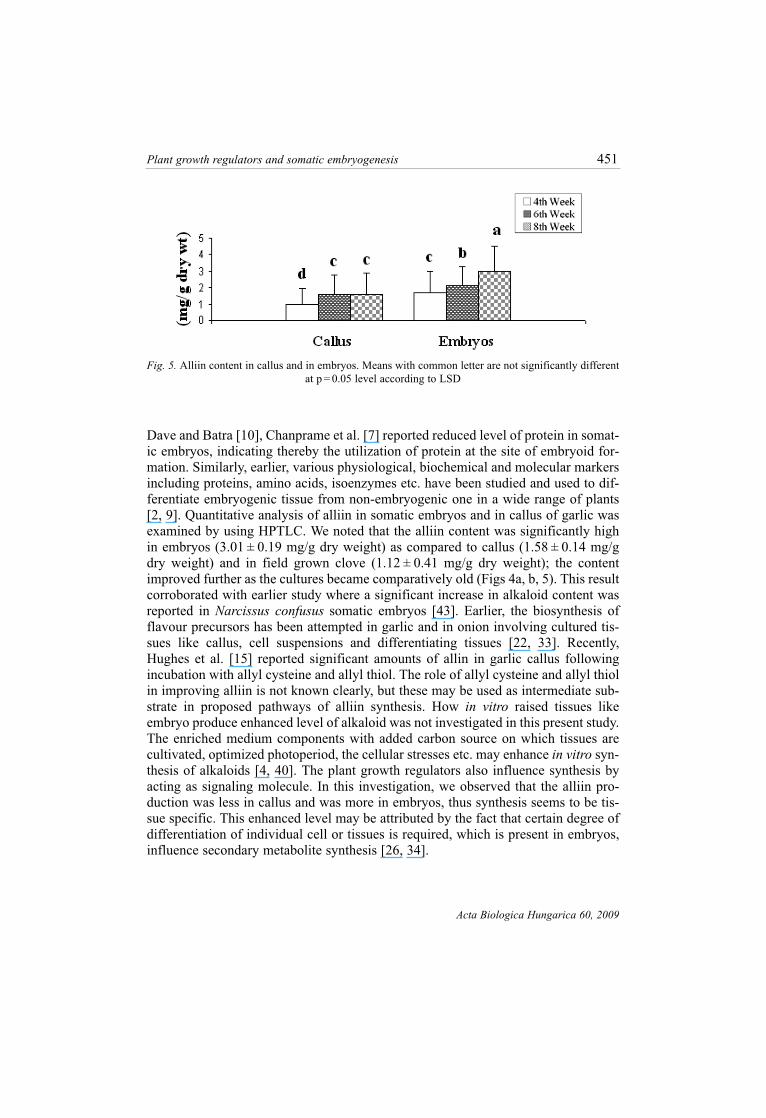
Dave and Batra [10], Chanprame et al. [7] reported reduced level of protein in somat-
ic embryos, indicating thereby the utilization of protein at the site of embryoid for-
mation. Similarly, earlier, various physiological, biochemical and molecular markers
including proteins, amino acids, isoenzymes etc. have been studied and used to dif-
ferentiate embryogenic tissue from non-embryogenic one in a wide range of plants
[2, 9]. Quantitative analysis of alliin in somatic embryos and in callus of garlic was
examined by using HPTLC. We noted that the alliin content was significantly high
in embryos (3.01 ± 0.19 mg/g dry weight) as compared to callus (1.58 ± 0.14 mg/g
dry weight) and in field grown clove (1.12 ± 0.41 mg/g dry weight); the content
improved further as the cultures became comparatively old (Figs 4a, b, 5). This result
corroborated with earlier study where a significant increase in alkaloid content was
reported in Narcissus confusus somatic embryos [43]. Earlier, the biosynthesis of
flavour precursors has been attempted in garlic and in onion involving cultured tis-
sues like callus, cell suspensions and differentiating tissues [22, 33]. Recently,
Hughes et al. [15] reported significant amounts of allin in garlic callus following
incubation with allyl cysteine and allyl thiol. The role of allyl cysteine and allyl thiol
in improving alliin is not known clearly, but these may be used as intermediate sub-
strate in proposed pathways of alliin synthesis. How in vitro raised tissues like
embryo produce enhanced level of alkaloid was not investigated in this present study.
The enriched medium components with added carbon source on which tissues are
cultivated, optimized photoperiod, the cellular stresses etc. may enhance in vitro syn-
thesis of alkaloids [4, 40]. The plant growth regulators also influence synthesis by
acting as signaling molecule. In this investigation, we observed that the alliin pro-
duction was less in callus and was more in embryos, thus synthesis seems to be tis-
sue specific. This enhanced level may be attributed by the fact that certain degree of
differentiation of individual cell or tissues is required, which is present in embryos,
influence secondary metabolite synthesis [26, 34].
Plant growth regulators and somatic embryogenesis 451
Acta Biologica Hungarica 60, 2009
Fig. 5. Alliin content in callus and in embryos. Means with common letter are not significantly different
at p = 0.05 level according to LSD

452 S. A. NASIM et al.
Acta Biologica Hungarica 60, 2009
CONCLUSIONS
In conclusion, the present study demonstrates that in vitro cultures provide a unique
tool to study the biochemical difference(s) associated with morphological develop-
ment. It also indicates that in embryos, the content of primary compounds like solu-
ble amino acid and protein are high, so is alliin, which could both be used as mark-
ers to distinguish callus from comparatively complex tissues like developing
embryos.
REFERENCES
1. Arnault, J. P., Christidès, N., Mandon, T., Haffner, R., Kahane, J., Auger, J. (2003) High-performance
ion-pair chromatography method for simultaneous analysis of alliin, deoxyalliin, allicin and dipep-
tide precursors in garlic products using multiple mass spectrometry and UV detection. J. Chromato-
graphy 991, 69–75.
2. Bagnoli, F., Capuana, M., Racchi, M. L. (1998) Developmental changes of catalase and superoxide
dismutase isoenzimes in zygote and somatic embryos of horse chestnut. Aust. J. Plant Physiol. 25,
909–913.
3. Barandiaran, X., Die Pietro, A., Martin, J. (1998) Biolistic transfer and expression of a uidA reporter
gene in different tissues of Allium sativum L. Plant Cell Rep. 17, 737–741.
4. Bhat, M. A., Ahmad, S., Junaid, A., Mujib, A., Mahmooduzzafar (2008) Salinity stress enhanced pro-
duction of solasodine in Solanum nigrum L. Chem. Pharm. Bull. 56, 17–21.
5. Bradford, M. M. (1976) A rapid and sensitive method for quantification of microgram and quantities
of protein utilizing the principle of protein dye binding. Anal. Biochem. 72, 248–254.
6. Campbell, J. H., Efendy, J. L., Smith, N. J., Campbell, G. R. (2001) Molecular basis by which garlic
suppresses atherosclerosis. J. Nutr. 131, 1006S–1009S.
7. Chanprame, S., Kuo, T. M., Widholm, J. M. (1998) Soluble carbohydrate content of soybean (Glycine
max L.) Merr. Somatic and zygotic embryos during development. In Vitro Cell. Dev. Biol. Plant 34,
64–68.
8. Chen, L. J., Luthe, D. S. (1987) Analysis of protein from embryogenic and non-embryogenic rice
(Oryza sativus L.) calli. Plant Sci. 48, 181–188.
9. Chugh, A., Khurana, P. (2002) Gene expression during somatic embryogenesis-recent advances.
Current Sci. 83, 715–730.
10. Dave, A., Batra, A. (1995) Role of protein metabolism constituents in somatic embryo formation in
cumin. Indian J. Plant Physiol. 38, 25–27.
11. Ferol, L., Chovelon, V., Causse, S., Michaux- Ferriere, N., Kahane, R. (2002) Evidence of a somat-
ic embryogenesis process for plant regeneration in garlic (Allium sativum L.). Plant Cell Rep. 21,
197–203.
12. Ferol, L., Chovelon, V., Causse, S., Triaire, D., Arnault, I., Auger, J., Kahane, R. (2005)
Establishment of embryogenic cell suspension cultures of garlic (Allium sativum L.), plant regenera-
tion and biochemical analyses. Plant Cell Rep. 24, 319–324.
13. Gaj, M. D. (2004) Factors influencing somatic embryogenesis induction and plant regeneration with
particular reference to Arabidopsis thaliana (L.) Heynh. Plant Growth Reg. 43, 27–47.
14. Halperin, W. (1995) In vitro embryogenesis: Some historical issues and unresolved problems. In:
Thorpe, T. A. (ed.) In Vitro Embryogenesis in Plants: Current Plant Science and Biotechnology in
Agriculture. Vol. 20, Kluwer Academic Publishers, Dordrecht, Netherlands, pp. 1–15.
15. Hughes, J., Tregova, A., Tomsett, A. B., Jones, M. G., Cosstick, R., Collin, H. A. (2005) Synthesis
of the flavour precursor, alliin, in garlic tissue cultures. Phytochemistry 66, 187–194.

16. Iantcheva, A., Vlahova, M., Bakalova, E., Kondorosi, E., Elliott, M. C., Atanassov, A. (1999)
Regeneration of diploid annual medics via direct somatic embryogenesis promoted by thidiazuron
and benzylaminopurine. Plant Cell Rep. 18, 904–910.
17. Ignacimuthu, S. (1995) Plant cell and tissue culture. In: Ignacimuthu, S. (ed.), Basic Biotechnology,
TATA McGraw-Hill Pub. Co Ltd., New Delhi, pp.100–142.
18. Jeyaseelan, M., Rao, M. V. (2005) Biochemical studies of embryogenic and non-embryogenic callus
of Cardiospermum halicacabum L. Indian J. Exp. Biol. 43, 555–560.
19. Jimenez, V. M., Thomas, C. (2006) Participation of plant hormones in determination and progression
of somatic embryogenesis. In: Mujib, A., Samaj, J. (eds) Plant Cell Monograph (Somatic embryoge-
nesis), Vol. 2, Springer-Verlag, Berlin Heidelberg, pp. 103–118.
20. Kintzios, S., Brossopoulos, J., Shortsianitis, E., Peppes, D. (2000) Induction of somaic embryogene-
sis from young fully expanded leaves of chilli pepper (Capsicum annuum L.) effect of leaf position,
illumination and explant pretreatment with high cytokinin concentrations. Sci. Hort. 85, 137–144.
21. Komamine, A., Kawara, R., Matsumato, M., Sunabori, S., Toya, T., Fujimora, T. (1992) Mechanism
of somatic embryogenesis in cell cultures: physiology, biochemistry and molecular biology. In Vitro
Cell Dev. Biol. 28, 11–14.
22. Lancaster, J. E., Shaw, M. L. (1991) Metabolism of γ glutamyl peptides during development, storage
and sprouting of onion bulbs. Phytochemistry 30, 2857–2859.
23. Lawson L. D. (1996) The composition and chemistry of garlic cloves and processed garlic. In: Koch,
H. P., Lawson, L. D. (eds), Garlic: The Science and Therapeutic applications of Allium sativum L.
and Related Species. Second ed. Williums and Wilkins, Baltimore, USA, pp. 37–108.
24. Lee, Y. P., Takahashi, T. (1996) Improved calorimetric determination of amino acids with the use of
ninhydrin. Anal. Biochem. 14, 71–77.
25. Liu, C. M., Xu, Z. H., Chua, N. H. (1993) Auxin polar transport is essential for the establishment of
bilateral symmetry during early plant regeneration. Plant Cell 5, 621–630.
26. Ma, R., Ritala, A., Caldentey, K. M. O., Rischer, H. (2006) Development of in vitro techniques for
the important medicinal plant Veratrum californicum. Planta Medica 72, 1142–1148.
27. Masanori, A., Kenji, T., Shinichiro, S. (1995) Regeneration of whole plant from tissue-cultured shoot
primodia of garlic (Allium sativum L.). Plant Cell Rep. 15, 17–21.
28. Milner, J. A. (2001) A historical perspective on garlic and cancer. J. Nutr. 131, 1027S–1031S.
29. Murashige, T., Skoog, F. (1962) A revised medium for rapid growth and bioassays with tobacco tis-
sue cultures. Physiol. Plant 15, 473–497.
30. Myers, J. M., Simon, P. W. (1998) Continuous callus production and regeneration of garlic (Allium
sativum L.) using root segments from shoot tip-derived plants. Plant Cell Rep. 17, 726–730.
31. Nagakubo, T., Nagasawa, A., Ohkawa, H. (1993) Micropropagation of garlic through in vitro bulblet
formation. Plant Cell Tiss. Org. Cult. 32, 175–183.
32. Novak, F. J. (1990) Allium tissue culture. In: Rabinowitch, H. D., Brewster, J. L. (eds), Onion and
allied crops. CRC Boca Raton, Florida, pp. 767–768.
33. Ohsumi, C., Hayashi, T., Sano, K. (1993) Formation of alliin in the cultured tissues of Allium
sativum. Oxidation of S-allyl-L-cysteine. Phytochemistry 33, 107–111.
34. Pande, D., Purohit, M., Srivastava, P. S. (2002) Variation in xanthotoxin content in Ammi majus L.
cultures during in vitro flowering and fruiting. Plant Sci. 162, 583–587.
35. Pedroso, M. C., Pais, M. S. (1995) Factors controlling somatic embryogenesis. Plant Cell Tiss. Organ
Cult. 43, 147–154.
36. Raemakers, C. J. J. M., Jacobsen, E., Visser, R. G. F. (1995) Secondary somatic embryogenesis and
applications in plant breeding. Euphytica 81, 93–107.
37. Rahman, K. (2001) Historical perspective on garlic and cardiovascular disease. J. Nutr. 131,
977S–979S.
38. Rauber, M., Grunewaldt, J. (1988) In vitro regeneration in Allium species. Plant Cell Rep. 7,
426–429.
39. Robledo-Paz, A., Villalobos-Arambula, V. M., Jofre-Garfias, A. E. (2000) Efficient plant regenera-
tion of garlic (Allium sativum L.) by root tip culture. In Vitro Cell Dev. Biol. Plant 36, 416–419.
Plant growth regulators and somatic embryogenesis 453
Acta Biologica Hungarica 60, 2009

454 S. A. NASIM et al.
Acta Biologica Hungarica 60, 2009
40. Rocha, L. K., Oliveira, A. J. B., Mangolin, C. A., Machado, M. F. P. S. (2005) Effect of different cul-
ture medium components on production of alkaloids in callus tissues of Cereus peruvianus
(Cactaceae). Acta Sci. Biol. Sci. 27, 37–41.
41. Sagare, A. P., Lee, Y. L., Lin, T. C., Chen, C. C., Tsay, H. S. (2000) Cytokinin-induced somatic
embryogenesis and plant regeneration in Corydalis yanhusuo (Fumariaceae) – a medicinal plant.
Plant Sci. 160, 139–147.
42. Sata, S. J., Bagatharia, B., Thaker, V. S. (2001) Induction of direct embryogenesis in garlic (Allium
sativum L.). Methods Cell Sci. 22, 299–304.
43. Selles, M., Viladomat, F., Bastida, J., Codina, C. (1999) Callus induction, somatic embryogenesis and
organogenesis in Narcissus confusus: Correlation between the state of differentiation and the content
of galanthamine and related alkaloids. Plant Cell Rep. 18, 646–651.
44. Tokuji, Y., Kuriyama, K. (2003) Involvement of gibberellin and cytokinin in the formation of
embryogenic cell clumps in carrot (Daucus carota). J. Plant Physiol. 160, 133–141.
45. Tran Thanh Van, K., Gendy, C. (1993) Relation between some cytological, biochemical, molecular
markers and plant morphogenesis. In: Roudelakis-Angelakis, K. A., Tran Thanh Van, K. (eds),
Morphogenesis in Plants, Molecular Approaches. Plenum Press, New York, pp. 39–45.
46. Yeh, Y. Y., Liu, L. (2001) Cholesterol-lowering effect on garlic extracts and organosulfur compounds:
human and animal studies. J. Nutr. 131, 989s–993s.





























