Embed Size (px)
Citation preview

In Silico Analyses of Proteomic Data Suggest a Role for HeatShock Proteins in Umbilical Cord Blood HematopoieticStem Cells
Angelo D’Alessandro & Giuliano Grazzini &Bruno Giardina & Lello Zolla
Published online: 10 August 2010# Springer Science+Business Media, LLC 2010
Abstract Umbilical-cord blood (UCB) has growingly becomean accepted alternative source of hematopoietic stem cells fortransplantation purposes. However, the low cell dose limitwithin a single unit is still an obstacle hindering the way of abroader diffusion. The real deal is the lack of knowledge aboutthe molecular processes governing the events of expansion anddifferentiation of these cells. In order to fill this void, severalstudies were focused on the identification of the peculiar wholeprotein profile of UCB-derived hematopoietic stem cells. In thisreview article we provide a referenced list of overall proteinsfrom UCB-derived hematopoietic stem and progenitor cells.This list has been elaborated for pathway and network analyses,along with GO term enrichment for biological and molecular
functions, in order to individuate main classes of proteinsgoverning functioning of these cells. From these analyses itseems to emerge a central role for heat shock proteins inimmature hematopoietic stem cells. Their role might berelevant in protecting crucial transcription factors which driveproliferation and differentiation towards a specific lineage (e.g.erythroid, myeloid). Hereby we also stress the helpfulness ofinteractomics elaboration in providing a unified overview ofindependent proteomics data. It appears that maturation, otherthan representing a bottleneck to protein expression, couldsculpt interaction maps via reducing complexity of immatureinteractomics profiles.
Keywords Umbilical cord blood .
CD34+ stem and progenitor cells . Network analysis .
Pathway analysis . Gene ontology
AbbreviationsESC embryonic stem cellFACS fluorescent activated cell sortingGE gel electrophoresisGVHD Graft versus host diseaseHLA human leukocyte antigenHSC hematopoietic stem cellHSP heat shock proteinIPA Ingenuity Pathway AnalysisMACS magnetic-activated cell sortingSCF stem cell factorUCB Umbilical cord blood
Introduction
Since 1988, when Gluckman’s group performed the firstsuccessful cord cell transplant to a sibling with Fanconi’s
All the authors disclose any financial involvement and conflict ofinterest with the subject treated in this review.
Electronic supplementary material The online version of this article(doi:10.1007/s12015-010-9180-z) contains supplementary material,which is available to authorized users.
A. D’Alessandro : L. ZollaDepartment of Environmental Sciences, Tuscia University,Viterbo, Italy
G. GrazziniItalian National Blood Centre, Istituto Superiore di Sanità,Rome, Italy
B. GiardinaIstituto di Biochimica e di Biochimica Clinica,Università Cattolica,Milan, Italy
B. GiardinaIstituto per la Chimica del Riconoscimento Molecolare, CNR,Roma, Italy
L. Zolla (*)Tuscia University - Largo dell’Università snc,01100 Viterbo, Italye-mail: [email protected]
Stem Cell Rev and Rep (2010) 6:532–547DOI 10.1007/s12015-010-9180-z

anaemia [1], umbilical cord blood (UCB)-derived hemato-poietic stem cells (HSCs) have been growingly attractingthe transplantation endeavour, especially upon observationson their suitability for unrelated transplants [2–5]. Indeed,HSC transplantations have become to be considered theeligible treatment for an increased number of malignancies,such as benign and dysplastic hematologic disorders as wellas of several inherited diseases [6–8]. UCB is one of themain sources of HSCs for transplantation purposes,although they are mainly derived from mobilized peripheralblood and bone marrow. Nonetheless, UCB-derived HSCsare growingly gaining momentum [1] (Fig. 1), due to aseries of relevant considerations, including the harmlesscollection procedure and minor ethical concerns whencompared against stem cells from embryonic sources(Table 1).
Unfortunately, some concerns still dampen the enthusiasmsaround UCB as the principal source of HSCs. To begin with,the volumes of UCB and bone marrow collected from a singledonor are not equivalent. Therefore, it frequently happens thatthe UCB nucleated cell dose does not reach the loweracceptable limit for an adult, though making it suitable formost of pediatric patients [9]. Several approaches have beenproposed to cope with this inconvenience, viz infusion ofunits from multiple donors [10–12], ex vivo or in vitro cellculturing prior to transplantation [13–19], non-myeloablativeconditioning regimens [20], intra-bone inoculation [21].
Since the main drawback of UCB is the low nucleated celldose/unit, the attempts to dissect the processes which regulatethe induction of proliferation and, eventually, differentiation ofUCB-derived HSCs are currently under thorough investigation.In this respect, valuable preliminary knowledge has beenaccumulated through proteomics studies on UCB-derivedHSCs, as it has been recently reviewed [22]. Minor concernshave arisen regarding proteomics analyses, as sample collec-tion and preparation methods have been highly standardized,in order to reduce the bias of the experimental approach
towards a specific output. The first issue to address is tohomogenize cell populations from UCB by exploiting theirstage of differentiation as a discriminating criterion. To thisend, several markers have been considered for evaluationamong the ones related to various phases of commitment.Among them, the CD34 antigen is a sialomucin type Itransmembrane protein with a molecular weight of 105–120 kDs, which is expressed by highly immature andproliferating cells and is a widely-accepted marker ofhematopoietic stem and progenitor cell fractions [23].
Although antibody-based approaches for separation ofCD34+ populations (mainly FACS and MACs), along withdelicate handling procedures have been suggested to partiallyinfluence the molecular profiles of these cells [24], proteomicsstudies on UCB-derived CD34+ in literature have addressedthese cells through complementary approaches and thusconveyed an overall portrait which appears to be reliableenough [25–28]. As these studies have been thoroughlyreviewed [22], we will base on their data to perform an insilico analysis which is based on gene ontology (GO) termenrichment for biological and molecular functions, along withpathway and network analyses, in order to trace out apanoramic profile of the overall proteins individuated exper-imentally ([25–28]) and to dissect specific classes of proteinsaccounting for statistically significant functions. The goal is toretrieve biologically-relevant observations to suggest furtherdirections for experimental analysis on the CD34+ cellpopulations from cord blood sources.
Pathway and Network Analysis, GO Term Enrichmentfor Biological and Molecular Functions
Pathway and network analyses were performed, as previ-ously reported [29–31].
Data from independent studies were regrouped in 5 lists (4including independent studies and one for merged data) anduploaded as .xls files (Microsoft Excel Spreadsheet) forelaboration to the Ingenuity Pathway Analysis (IPA) software(Version 8.5; Ingenuity® Systems, www.ingenuity.com) [32].The software compared the submitted datasets, in whichproteins were expressed through various annotations (e.g.UniProt, GenBank) to the internal database (Ingenuity Path-ways Knowledge Base) in order to compile a uniform list ofproteins prior to further in silico elaboration. IPA software wasinitially developed for the study of pathologies and mainlyfocused on cancer; thus, many of the retrieved pathways arerelated to diseases, in particular to cancer. Basing on IPAresults alone, it could be questioned whether the results arereliable or rather biased towards specific outputs. In order toconfirm IPA observations, IPA or equivalent software werefirstly used to establish protein groups. Then, proteins werealso examined for additional functional annotations in
Fig. 1 The recent trend of donations reveals that cord blood hasrapidly grown in importance during the last ten years as far as itconcerns the sources of hematopoietic stem cells. Adapted from Brandet al. 2008 [1]
Stem Cell Rev and Rep (2010) 6:532–547 533

available database with the aim to establish some hypothesesconcerning their function. In order to validate observationsfrom IPA analysis, functional enrichment of gene ontologies(GOs) was performed exploiting Babelomics tools, such asFatiGO [33–35].
The significance of the association between the data set andthe canonical and disease/toxicity pathways was measured bycalculating either: i) the ratio of the number of proteins fromthe data set that map to the pathway with respect to the totalnumber of proteins that map to the canonical pathway; ii) theFischer’s exact test p-value determining the probability thatthe association between the proteins in the dataset and thecanonical pathway was explained by chance alone. Analo-gous parameters were adopted to calculate network scores,based on the correspondence with the pre-loaded networks inthe database and Fischer’s exact test for p-value determina-tion. Highest scores were proportional to a lower probabilityof casual association. The IPA software allowed us toperform an unbiased elaboration of the available data, withthe final aim to focus subsequent analyses and discussion onthe pivotal pathways and networks. In the end, the softwaredetermines and graphs unbiased networks, in which geneproducts (proteins) are represented as nodes, and thebiological relationship between two nodes is represented asan edge (line). All edges are supported by at least 1 referencefrom the literature, from a textbook, or from canonicalinformation stored in the Ingenuity Pathways KnowledgeBase. Continuous lines (edges) represent direct interactions,while indirect ones are represented by interrupted lines.Circular lines around one node describe a feed-back loop ofactivity of that node on itself (e.g. by self-modulating itsactivity or expression). Gray edges represent interactionswithin a single network, while orange edges cross-link nodesfrom multiple interacting networks.
This kind of approach is known as interactomics. Interac-tomics is a fusion science of biology, informatics andengineering which provides a global view of protein familyinteraction networks [36]. It involves the study of both theinteractions and the consequences of those interactionsbetween and among proteins, and other molecules within acell and can be used to compare networks of interactionbetween and within species to see how the traits of suchnetworks are varied and conserved. The interactome networkincludes certain calculated parameters that weigh the reliabil-ity of a given interaction (i.e. the ‘edges’ of the interactomenetwork) between two proteins, and also qualify the functionalenvironment around any given protein (i.e. the ‘nodes’ of theinteractome network).
In parallel, FatiGO/Babelomics software was used toanalyze the list of gene products and to compare it against therest of the human genome upon conversion of the proteinentries into a list of GO terms, using the corresponding gene-GO association table. Then, Fisher’s exact test was used tocheck for significant over-representation of GO terms in thesubmitted dataset against the rest of the genome (p<0.05). Inaddition to GO [35] terms, FatiGO was used to simultaneous-ly test the dataset for KEGG [37] and BioCarta [38] pathwaysand InterPRO motifs [39].
In Silico Elaboration of Data from IndependentProteomic Studies on UCB-Derived Hematopoietic Stemand Progenitor Cells
Exploiting data from the 4 main proteomics studies onUCB-derived HSCs [46–48, 52], we managed to individ-uate the pivotal networks within these immature fractions(Fig. 2, panels from a to d).
Table 1 Advantages and disadvantages of UCB-derived HSCs for allogeneic transplantation
Advantages Disadvantages
Abundant source of stem cells Low nucleated cell dose in a single unit restrains UCB use to pediatricpatients only while it limits UCB wide-spread adoption in adult patientsEasily available (harmless for mother and child, almost costless)
No ethical concern (in comparison to embryonic stem cells) Future development of potential abnormalities of the newborn donor’s HSCinto adult life and their effects on the recipient, is unknown at the time oftransplant.
Minor populations could be better represented
Low viral contamination (e.g. cytomegalovirus, Epstein-Barr virus) Only one infusion is allowed for patients experiencing graft failure
Actual banking, no more virtual registries: UCB units are rapidlyavailable on demand
Never registered malignant transformations after infusion so far
T-cell immaturity (less cytokine production and aggressivecapacity against the recipient—infrequent acute or severe GVHD)
Partial HLA-matched infusion allows related and unrelated trans-plantations
Frozen storage prolongs shelf-life
No donor attrition or last-time consent refusal
Brand et al. [1]
534 Stem Cell Rev and Rep (2010) 6:532–547

The preliminary concern regarded the matching scoresbetween the experimental data from these papers whencompared against the software database (Ingenuity PathwayKnowledge Base). As a result, we observed an average of
69.20% (standard deviation 0.14) of proteins matching with thedatabase.
Network analysis of data from Zenzmaier et al. (2003) [25]determined the centrality of the “Cellular assembly and
Fig. 2 a Interactomics elaboration of data from Zenzmaier et al. 2003 [25]by means of Ingenuity Pathway Analysis (Ingenuity® Systems, www.ingenuity.com). The software determined and graphed the canonicalpathway with the highest score: the Cellular assembly and Organizationpathway. Several heat shock proteins (Hsp70, Hsp27, HspA3, Hsp90)represent the fulcrum of this pathway, locating in the centre of thenetwork and displaying a high number of interactions (edges) with otherproteins within the network. b Interactomics elaboration of data fromZenzmaier et al. 2003 [26] by means of Ingenuity Pathway Analysis(Ingenuity® Systems, www.ingenuity.com). The software determined andgraphed the canonical pathway with the highest score: the Moleculartransport, cellular trafficking, lipid metabolism pathway. Notably, HspB1(a 27 kDa Hsp protein) interacts with two regulators of proliferationevents, the epidermal growth factor receptor (EGFR) and tumor necrosisfactor (TNF), suggestive of a likely role of this protein in this pivotalbiological process. c Interactomics elaboration of data from Tao et al.2004 [27] by means of Ingenuity Pathway Analysis (Ingenuity® Systems,www.ingenuity.com). Noteworthy, the analyzed dataset included 45proteins which belonged only to the immature fraction (CD34+) but not
to the myeloid committed one (CD15+).The software determined andgraphed the canonical pathway with the highest score: the Cell death andCancer pathway. Notably, once again, HSPs represent the core of theprotein network (9 HSP in the graph), strengthening the idea that thisclass of proteins plays a relevant role in the immature fraction (while notin more committed ones). Further studies are mandatory. d Interactomicselaboration of data from Liu et al. 2006 [28] by means of IngenuityPathway Analysis (Ingenuity® Systems, www.ingenuity.com). Thesoftware determined and graphed the canonical pathway with the highestscore: the Cell morphology, cell-to-cell signaling and interaction pathway.The graph confirms evidences from previously mentioned studies [25–27]in which HSPs represented the protein core of the most relevantindividuated pathway. HSPs play various biological roles (protein foldingcontrol, responses to stress, and so forth) and it is worthwhile to underlinethat computerized analysis evidenced their centrality in UCB-derivedHSCs even when the most relevant pathway varied. Grey nodes: proteinsfrom the dataset having a match in the database; White nodes: proteinsfrom the database which were not identified (if present) upon theexperimental phase
Stem Cell Rev and Rep (2010) 6:532–547 535

organization” network (p-value: 1.38 E-05). Core nodes ofthis network are represented by proteins involved in actincytoskeleton (mainly actin isoforms) and heat shock protein(HSP) 70, which cooperates with an actual machinery of otherHSPs mainly involved in protein degradation activities, asunderlined by the presence of the “proteasome” node(Fig. 2a).
Network analysis of data from Zenzmaier et al. (2005) [26]evidenced the centrality of the “Molecular transport, cellulartrafficking and lipid metabolism” pathway (p-value 1.21 E-05), in which HSP27 interacts with two proliferationregulatory pathways involving epidermal growth factorreceptor (EGFR) and TNF (tumor necrosis factor) (Fig. 2b).This is suggestive of a possible role for HSP27 in themodulation of proliferation signaling. However, the elevatednumber of white nodes in this network indicates that thesoftware introduced a series of protein interactors which havenot been individuated in the experimental analysis performedby the authors [26]. Indeed, the 2005 study by Zenzmaier etal., which was based on the standard 2DE, LC-MS approachyielded rather limited, albeit complementary, results whencompared with the MudPIT approach from the same groupdating back 2003 [25].
When observing Fig. 2c, which graphs the pathway withthe highest score upon Pathway Analysis of CD34+-onlyproteins (Cell death, cancer and gastrointestinal diseases)(p-value=2.23 E-06), it is immediately evident that HSPslikely play a relevant role in more immature fractions (andthose only) from the comparative study by Tao et al. (2004)[27]. These data confirm the observations from in silicoresults based on Zenzmaier et al. and suggest that, whetherHSPs play a pivotal role in immature cells, their expression isdepressed, and their role dwindled, in more mature CD15+
cells. Indeed, multiple HSPs and chaperonins, includingmembers of the 60-kDa and 70-kDa heat shock proteinfamilies, chaperonin-containing TCP1 complex proteins,stress-induced phosphoprotein 1 (Hsp70/Hsp90- organizingprotein), and hypothetical protein DKFZ p761K0511, wereexpressed predominantly in CD34+ cells. This indicates thatCD34+ stem/progenitor cells possess the attributes of cellsunder stress, which is consistent with findings from genomicstudies of highly purified murine HSCs [40].
Network analysis of data from Liu et al. [28] evidenced apivotal network involved in “Cell morphology, cell-to-cellsignaling and interaction” (p-value 3,00 E-05) (Fig. 2d) inwhich HSPs represented, once again, the protein core.Besides, it was individuated another pathway with a highscore, namely “protein synthesis, folding and post-translational modification interaction network”, referring tofundamental cellular functions which, again, recall HSPbiological functions.
Some proteins were enlisted in top score networks frommultiple studies, although they had not been experimentally-
individuated but in one of them. For example, we observed thatsome proteins/nodes accounted for proteins which had beenexperimentally-individuated and thus graphed as “grey nodes”in one map, while they had not been experimentally-identifiedin the other study and thus they were represented as “whitenodes” (for example HSP70 in top networks from [26] Fig. 2aand [28] d). This is relevant in the light of the program bias tocomplement the submitted datasets with protein entries whichhave been registered as interactors in the software database. Ifa protein “artificially” introduced in the one top score networkfrom one study has been actually individuated in anotherexperimental approach to the same cell line, it could meanthat, whether a bias from the software exists, it is reliableenough.
Global Analyses from Merged Data
Given these observations, we decided to perform a unitaryanalysis bymerging of all the data from each independent study[25–28]. Since some proteins might have been omitted fromthe dataset, due to the incompleteness or partial mismatchwith the Ingenuity Knowledge Base (only 69% of the proteinsfrom independent studies were automatically individuated),manual verification of each protein entry was performed andaccuracy of the analyzed dataset manually assessed. Amanually-validated list was compiled excluding redundantentries while including protein isoforms, as to yield a total of465 unique gene products. The list is reported in Supplemen-tary Table 1, in which proteins are indicated along with theirUniProt IDs, UniProt-retrieved general biological function,and reference from which they were gathered.
Notably enough, a series of proteins were individuated ineach one of the independent studies accounting for biologicalfunctions which are not closely-related to hematopoietic stemand progenitor cells (Fig. 3). Some proteins in the list, mainlyfrom [26] and [28], are apparently ectopically expressed. Anexample is cyclin I—CCNI which is related to spermatogen-esis [41], but it is also linked to terminal differentiation of thepost-natal mouse heart [42]. Analogously, the human DEAD-box Y RNA helicase DBY (DDX3Y) gene is located in the(AZFa) region on the human Y chromosome (Yq11.21) and ispredominantly expressed during spermatogonia, playing a rolein RNA metabolism during human spermatogenesis [43].Similar considerations could be made for tesmin (MTL5), ametallothionein-like protein, and zonadhesin (ZAN), bothinvolved in gamete-related functions such as spermatogenesis[44] or gamete recognition [45].
Other proteins in the merged list likely exert biologicalfunctions related to neuronal development. A few examples arerepresented by the neurogenic differentiation factor 6(NEUROD6), which is involved in early differentiation stagesof neurons [46], or astrotactin 1 (ASTN1), a neuronal adhesion
536 Stem Cell Rev and Rep (2010) 6:532–547

molecule that is required for glial-guided migration of youngpostmitotic neuroblasts in cortical regions of developing brain,including cerebrum, hippocampus, cerebellum and olfactorybulb [47]. PAX6 is long known to take part in organmorphogenesis, namely eye-development [48], while CRYGDbelongs to the family of gamma-crystallins, the mainstructural constituents of the eye lens [49]. While a role forthese proteins in HSCs is not to be excluded, albeit nothitherto assessed, these molecules could represent a symptomof the immatureness of CD34+. In parallel, their individuationin immature cells of potential hematopoietic lineages could beinterpreted as a symptom of immaturity of the protein profileas well, whereas this results in a widespread protein
expression which ends up to be sculpted towards specificityas the cell progresses in its way to final commitment. In otherterms, it could be suggested that maturation of stem andprogenitor cells towards specific lineages could represent abottleneck to protein expression. A minor confirmation of thishypothesis could be obtained from the study by Tao et al.[27], the only one comparing CD34+ hematopoietic stem andprogenitor cells to myeloid-committed CD15+ cells, in whichthe latter population did not show any relevant marker ofimmaturity. This could represent an interesting area for furtherresearch, as its results could make the concept of guiding cellsthrough differentiation evolve from turning on the expressionof specific genes to turning off the “ectopic” ones. In parallel,
Fig. 3 A brief representationsof the proteins individuated ex-perimentally which are notclosely related with hematopoi-etic stem and progenitor cells,rather they relate with moremature blood cell components orwith more immature cells fromother physiological districts(embryionic development, neu-ral and eye development, sper-matogenesis). Only gene namesare reported (extended indica-tions can be found in Supple-mentary Table 1)
Stem Cell Rev and Rep (2010) 6:532–547 537

the protein list from merged proteomic data on UCB-derivedHSCs included proteins which are related to terminally-committed mature blood cells, such as hemoglobins (HBA1,HBB, HBE, HBG1, HBG2) or T-lymphocyte specificreceptors (TCRA, TCRB).
Pathway Analysis and GO
From the integrated analysis of pathways and networks, it couldbe deduced whether a certain class of proteins (pathways) andtheir interactors (networks) likely play a more determinant rolein the biological sample under examination or organicallymerge independent experimental data as to obtain a singlepanoramic view of the whole protein network.
Pathway analysis through IPA of the 465 gene productsfrom the merged datasets yielded a total of 69 main pathways,which could be further divided in 790 subpathways, rangingfrom tumorigenesis (6.50 E-15) to embryonic development(1.29 E-02).
Top 25 score results are enlisted in Supplementary Table 1are reported in Fig. 4a, in which results are graphed asnegative logarithm of the probability scores ( the moststatistically significant pathways have the highest absolutevalue in the graph, clockwise progression). The analysisindicated that top 25 score pathways accounted for proteinclasses involved in cellular proliferation (tumorigenesis,proliferation of ekaryotic cells), cell cycle regulation (celldeath of eukaryotic cells, cell death, cell death of normal cells,apoptosis of eukaryotic cells, apoptosis), cell growth anddevelopment, hexose metabolism, transport, folding andmodification of proteins, repairing mechanisms (doublestranded-DNA break repair of cells, repair of cells) andprocessing of genetic information (modification of RNA).
These results partly overlap with the output from GO termenrichment for biological functions (Fig. 4b), from which itemerges a role for those classes of proteins involved inmonosaccharide/carbohydrate metabolic and catabolic process(GO:0005996, 0005975, 0009056, respectively), proteinfolding/response to stress/protein depolimerization/responseto unfolded proteins (GO:0006457, 0006950, 0051261,0006986, respectively), cell cycle (GO:0007049), along withcytoskeleton organization and biogenesis (GO:0007010).
Regulation of cytokeletal structure (hsa04810), modulationof cell to cell communication (hsa01430) and of glycolysis(hsa00010): these pathways have been also individuated asstatistically significant pathways upon interrogation of KEGGdatabase (Table 2). This is coherent with results from IPA andFatiGO/Babelomics, as well as with BioCarta, which deter-mined glycolysis, cell to cell communications as pivotalpathways, while including DNA damage fragment pathway aswell (Table 2).
The indication that UCB-derived HSCs mainly rely onglycolytic metabolism is on line with analogous observation on
murine bone-marrow-derivedHSCs [50], in which quantitativeproteomics approaches revealed high expression of glycolyticenzymes and oxidative stress-related enzymes for damagerepairing. This apparently is a characteristic of highlyproliferating cells, either healthy or diseased (such as cancercells), which is long known as the Warburg effect [51].Warburg effect hypothesis could also explain the indicationthat most of the statistically significant pathways andontologies include proteins involved to stress response (eitherto proteins or DNA).
All these observations were confirmed by GO termenrichment for molecular functions (Fig. 4c), which describedproteins according to their molecular activity that, in this verycase, could be enlisted as follows: cytoskeletal protein binding(GO:0008092), actin binding (GO:0003779), both includingstructural components of cytoskeleton (GO:0005200); inparallel, unfolded protein binding (GO:0051082), and proteinbinding-bridging (GO:0030674), as well as other functionsrecalling DNA repairing and RNA processing (GO:0000166).Concordingly, top three recurring molecular motifs throughInterPro motifs analysis stressed the relevance of spectrinrepeat (IPR002017) and chaperonin-related molecular motifs(IPR002423 (Chaperonin Cpn60/TCP-1) and IPR001023(heat shock protein Hsp70)—Table 2).
Network Analysis
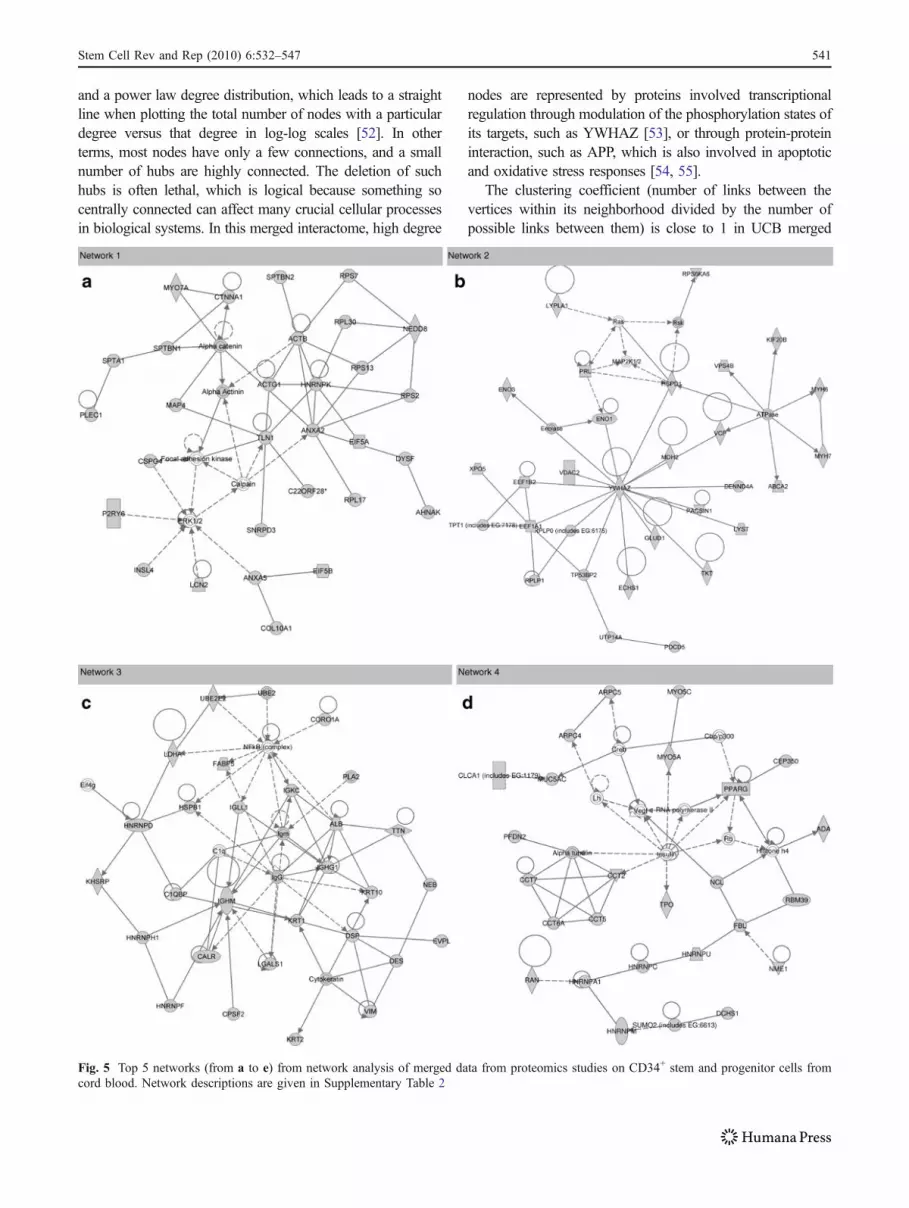
In order to provide visual evidence of these elaborations, anetwork analysis was performed with IPA as to obtain 25distinct networks (Supplementary Table 2).
Top five networks are reported in Fig. 5 (panels from a to e).Top score network appears to be devoted to “ProteinSynthesis” (Fig. 5a), as most of the proteins enlisted accountfor functions related to protein translation (form eukaryoticinitiation factors such as EIF5A and EIF5B, to ribosomalcomponents—RPL17, RPL30, RPS2, RPS7, RPS13), al-though some core nodes are related to actin cytoskeleton(ACTB, ACTG1, alpha actinin).
Network 2 regrouped proteins involved in “MolecularTransport, Metabolism, Small Molecule Biochemistry”(Fig. 5b). Among these proteins, there could be foundglycolytic enzymes (ENO1) and molecular transporters (e.g.GLUD1, XPO5, MYH7, VCP) which were coordinated byYWHAZ in a node-centered high-degree structure. Thedegree of a node indicates the number of links connected to1 vertex. High degree nodes are the most relevant ones ininteractome networks, as they represent the fulcrum ofmultiple signaling pathways, and their positive/negativemodulation could result in alterations of the activities of theirlikely interactors.
Top network 3 has been named by the software as the “Hairand Skin Development and Function, Organ Development,Dermatological Diseases and Conditions” network (Fig. 5c),
538 Stem Cell Rev and Rep (2010) 6:532–547

although it mainly includes proteins involved in the immunesystem (C1q, C1qBP, IgG, IGHG1, IGHM, IGKC, IGLL1,Igm), RNA splicing (HNRNPD, HNRNPF, HNRNPH1) andprotein folding/degradation (HSPB1, UBE2, UBE2E2). In
this case, no relevant node-centered structure could beobserved, rather a geometric pattern occupied the center ofthe network. Geometric patterns including molecules sharingsimilar functions (such as the catalytic ring which has been
Fig. 4 Top 25 results frompathway Analysis with Ingenui-ty Pathway Analysis (a), show-ing negative log10 of scoreprobabilities for the individuatedpathways (the highest scorepathways in the graph are themost statistically significant);Top 15 results from Babelomics/Fatigo analyses for Gene Ontol-ogies in relation to biological (b)and molecular (c) functions(normalized data are reported aspercentages)
Stem Cell Rev and Rep (2010) 6:532–547 539

individuated in the protein core of red blood cells [29]) are analternative to node-centered structures in protein-proteininteraction maps, as they guarantee that the molecular path-ways in which they are involved are maintained notwith-standing eventual alterations disrupting one single node.
“Cell Death, Cell Morphology, Molecular Transport” is thetop network 4 (Fig. 5d), in which the geometric structure ofchaperonin containing TCP1 subunit (CCT) isoforms (CCT2,CCT5, CCT7, CCT6A) represent almost a distinct entitydevoted to protein degradation, while other proteins areincluded in linear cascades referring to translational control(HNRNPA1, HNRNPC, HNRNPM, HNRNPU). Linearcascades differ from node-centered structures and geometricpatterns, in that each single node in the cascade is relevant inthe specific pathway, and interruptions of the cascade likelycompromise the biological activity exerted by these moleculesin a time- or space-dependent regulation of interactions.
Network number 5 (“Cancer, Post-Translational Modifica-tion”) mainly relies on a geometric pattern of heat shockproteins interacting among each other (HSP, Hsp27, Hsp70,Hsp90, HSP90AA1, HSP90AB1, HSPA6, HSPA8, HSPA9) asto guarantee correct folding of nascent proteins or refolding ofunfolded/misfolded ones.
Other protein groups involved in top score biological/molecular functions and pathways, such as proteins involved
in DNA damage repairing (ATM, RAD50, RAD51L) or cellcycle regulation (e.g. cyclins, CDKN3), although they did notmake the top 5 score networks, they were nonethelessindividuated in network 8 (top 10) or 13 (top 15), respectively(Supplementary Table 2). Therefore, it is reasonable toconclude that the information retrieved from network analysisis on line with other in silico results.
In parallel, a unique map was retrieved from mergednetworks as reported in Supplementary Figure 1. At a firstglance, it is evident that merging networks from the wholenon-redundant hypothetic proteome of UCB-derived HSCsproduces a burst of complexity, especially when compared towell established interactomes of more mature blood cells inliterature (for example, red blood cells [29]). This could beexplained as a symptom of “immaturity” of those cells, notonly by a mere proteomics point of view through an increasedprotein variability (see the previous paragraph and [27]), butalso through a not-yet well organized protein arrangementwhich prevents more ordered network organizations. Thediameter of the network (the maximum distance between anytwo nodes) is definitely higher than the diameter observed forred blood cell top score merged networks [29], even whenaddressing a higher number of nodes. The degree is high, as itis expected for biological networks, which are scale-free.Scale-free networks are characterized by high-degree nodes
Table 2 KEGG pathway and BioCarta analysis of umbilical cord blood-derived CD34+ stem and progenitor cells (vs the rest of the humangenome)
#Index Term #1 vs #2 p value Proteins KEGG
1 Glycolysis / Gluconeogenesis (hsa00010)
86.32% 13.68% 5.27e-3 ALDOA PGAM1GAPDH PDHA1
2 Cell comunication (hsa01430) 79.99% 20.01 2.14e-2 THBS2 VIM ACTBDES
3 Regulation of actin cytoskeleton (hsa04810)
71.84% 28.16% 7.78e-2 MYH10 ITGA2BMRLC2 ACTN2
BioCarta1 h_cell2cellPathway 86.03% 13.97% 5.93e-2 CTNNA1 ACTN2
2 h_DNAfragmentPathway 75.49% 24.51 3.09e-1 HMGB1
3 h_glycolysisPathway 86.03% 13.97 1.9e-1 GAPDH
InterPro Motifs1 IPR002017 (spectrin repeat) 94.45% 5.55 3.3e-10 ACTN2 RAD50
SPTBN1 SPTBN2ACTN4 MACF1 KRT1DMD UTRN SYNE2EVPL MYH13 PLEC1SPTA1 KTN1 DSP
2 IPR002423 (Chaperonin Cpn60/TCP-1)
95.44% 4.56% 2.64e-7 CCT7 CCT5 HSPD1TCP1 CCT2 CCT6AMKKS
3 IPR001023 (heat shock protein Hsp70)
90.11% 9.89% 1.56e-3 HSPA6 HSPA5HSPA8 HSPA9
540 Stem Cell Rev and Rep (2010) 6:532–547
and a power law degree distribution, which leads to a straightline when plotting the total number of nodes with a particulardegree versus that degree in log-log scales [52]. In otherterms, most nodes have only a few connections, and a smallnumber of hubs are highly connected. The deletion of suchhubs is often lethal, which is logical because something socentrally connected can affect many crucial cellular processesin biological systems. In this merged interactome, high degree
nodes are represented by proteins involved transcriptionalregulation through modulation of the phosphorylation states ofits targets, such as YWHAZ [53], or through protein-proteininteraction, such as APP, which is also involved in apoptoticand oxidative stress responses [54, 55].
The clustering coefficient (number of links between thevertices within its neighborhood divided by the number ofpossible links between them) is close to 1 in UCB merged
Fig. 5 Top 5 networks (from a to e) from network analysis of merged data from proteomics studies on CD34+ stem and progenitor cells fromcord blood. Network descriptions are given in Supplementary Table 2
Stem Cell Rev and Rep (2010) 6:532–547 541

network, while it is definitely lower in its red blood cellcounterpart [29]. Therefore, it is of note that the networkgraphing the top 500 nodes in red blood cells has a clearertopology (lower degree, diameter and clustering coefficient)than the 465 nodes of UCB-derived HSCs. It is as ifmaturation could represent a bottleneck not only to proteinexpression, but also to protein-protein interactions through amechanism of “interaction maturation” which leads toprogressive sculpting of specific pathways at the expenses ofprotein expression variability (immature cells often expressproteins which are characteristic of other tissues [28]).Hopefully, further studies will shed light on this hypothesisas well.
A deeper analysis suggests that the center (and south-center) portion of the map is dominated by proteins involvedin cell death inhibition (inhibition of apoptosis) as well as inprotein folding and cancer markers, which implies that thecenter of the map also harbors those proteins belonging to thehighest score pathways from pathway and GO term enrich-ment analyses. HSP hubs show the highest closenesscentrality score as they garrison this region of the interactomeand project their edges towards both periphery and the veryheart of the map. In topology and related areas in mathemat-ics, closeness is one of the basic concepts in a topologicalspace. Intuitively we say two sets are close if they arearbitrarily near to each other. The concept can be definednaturally in a metric space where a notion of distance betweenelements of the space is defined, but it can be generalized totopological spaces where we have no concrete way tomeasuredistances. In graph theory closeness is a centrality measure ofa vertex within a graph. Vertices that are 'shallow' to othervertices (that is, those that tend to have short geodesic
distances to other vertices with in the graph) have highercloseness. Closeness is preferred in network analysis tomean shortest-path length, as it gives higher values to morecentral vertices, and so is usually positively associated withother measures such as degree.
As furtherly pinpointed by the statistical analysis (previousparagraph), this constitutes the fulcrum of the mergednetwork, thus reinforcing evidences from top score networkgraphs of each independent proteomics study [25–28]. Asimplified view of this region is given in Fig. 6, which showsa portion of the Protein Folding/Post-translational modifica-tion and Cancer top score network 5.
In the light of these considerations, an overview onHSPs will be given as follows, in order to elucidate whythey are rapidly becoming to be considered an eligiblemarker of proliferating cells and how this relates to UCB-derived HSCs investigations.
Heat Shock Proteins: the Role in Cancer/Stem CellProliferation and Hematopoietic Differentiation
Stress or heat shock proteins (HSPs) were first discoveredin 1962 [56] as a set of evolutionary ancient and highlyconserved proteins whose expression was induced bydifferent kinds of stresses. These intracellular molecularchaperones belong to the protein-folding and qualitycontrol systems: their biological functions include arelevant role in post-translational checkpoints which areneeded to ensure fidelity and regulation of eukaryoticprotein expression [57, 58]. Well-known is their role in thecatalysis of the proper folding either of nascent ormisfolded proteins.
Although most of them are constitutively expressed,their expression is upregulated by various physiologicalperturbations and stressors (oxidation, hypoxia, ischemia,heavy metals, elevated temperature, radiation, calciumincrease, glucose deprivation or starvation, cancer andmicrobial infection) [59] and are catalogued into severalclasses, according to their molecular weight (e.g. HSPs 27,60, 70, 90) [60].
The most studied HSPs are HSP 70, 90 and the smallHSP27, the latter being ATP-independent in its chaperoneactivity, mainly devoted to prevent protein aggregation ratherthan controlling protein folding [61].
It is worthwhile to recall that independent (Figs. 2, 3, 4 and5) and merged (Fig. 6) interactomics analyses of proteomicsdata from literature have stressed the centrality of HSPs notonly in protein-folding/post-translational modification net-works, but also in cancer and cell death inhibition top scorepathways (Fig. 6). This is in agreement either with findingsabout HSP involvement in stem cell proliferation, hematopoi-etic differentiation, inhibition of apoptosis and cancer [62–87].
Fig. 5 (continued)
542 Stem Cell Rev and Rep (2010) 6:532–547

Stem Cell Proliferation and Hematopoietic Differentiation
A role for HSPs in proliferation and self-renewal of stemcells has been recently proposed by Prinsloo’s group [73].Indeed, whereas their role in mature differentiated-cells iswell-known, it still remains unclear how do they act withinstem cells. Embryonic stem cells actually exhibit anincreased stress-tolerance and concomitant high levels ofchaperone expression [73]. Prinsloo and Colleagues sug-gested that HSPs might directly trigger or protect signalingintermediates in essential proliferative/differentiation cas-cades within immature cells, in like fashion to HSP90taking part in the JAK/STAT3 pathway in somatic andmouse embryonic stem cell lines [74, 75].
HSPs have been lately related to differentiation events ofhematopoietic progenitors towards erythroid [76], myeloid[77], and lymphoid [78] lineages through a protectivemechanism of peculiar transcription factors which are ableto direct differentiation events in immature HSCs. In thisview, a recent Nature paper has stressed the role of HSP 70in the prevention of the inactivation of transcription factor
GATA-1 operated by a caspase-mediated proteolysis,indicating that HSP 70 might indirectly trigger erythroiddifferentiation [76]. Viceversa, HSP70 expression levels areindirectly influenced upon chemical induction of erythroiddifferentiation in K562 cells [79]. HSP70 is also induced inother models of hematopoietic differentiation [80–83].Notably, HSP70 constitutes the central node in the Proteinfolding/Post-translational modification and cancer mergednetworks in Fig. 6.
HSPA8, also known as Hsc70 has been demonstrated toplay a role in cytokine-mediated HSC survival and tonegatively influence the stability of pro-apoptotic BimmRNA, thus preventing apoptosis in hematopoiesis andleukemogenesis [84].
Loss of HSPA9B, also known as HSP 70 kDa 9 ormortalin, recapitulates the ineffective hematopoiesis ofmyelodysplastic syndrome in zebrafish [85].
HSP27 [81] and HSP60 [77] have been instead related tomyeloid commitment via an unexpected role they seem todirectly play as cognate triggers for crucial monocyte-macrophage receptors, which induce activation and prolif-eration of these cells, such as CD15+ [77]. HSP28 has beenalso suggested as a unique marker of growth arrest duringmacrophage differentiation of HL-60 cells [86].
Therefore, other than realistically playing a role inproliferation, HSPs could be also involved in differentiationevents of UCB-derived HSCs and thus might represent akey target either for future investigations or for approachesaddressing the low cell dose limit through induction ofproliferation and modulation of differentiation events.
Apoptosis
In addition to their chaperone activity, HSPs are long-known to counteract cellular senescence and programmedcell death (apoptosis). HSPs have demonstrated a protectiveinfluence toward cell death [62]. Overexpression of HSP27,HSP70 or HSP90 prevents apoptosis triggered by variousstimuli, including hyperthermia, oxidative stress, stauro-sporine, ligation of the Fas/Apo-1/CD95 death receptor oranticancer drugs [61]. Conversely, downregulation orinhibition of HSP27, HSP70 or HSP90 have been shownto be enough to sensitize a cell to apoptosis [68]. The anti-apoptotic effects of HSPs involve protein–protein interac-tions that seem not to be directly related to their chaperonefunctions [63]. For example, HSP27 interacts with cyto-chrome c when released from the mitochondria whereasHSP70 or HSP90 interact with Apaf-1, which in all casesnegatively interferes with the post-mitochondrial pathwayto caspase activation by inhibiting the formation of theapoptosome [64].
HSP70 also has the capacity to interact with theflavoprotein (apoptosis-inducing factor) to negatively inter-
Fig. 6 Simplified view of the top score networks from the wholeinteractome of UCB-derived HSCs basing on the protein datasets fromthe available proteomics studies [25–28]. Protein folding/Post-trans-lational modification and Cancer networks are merged and displayed.HSP70 visibly occupies the center of these merged networks, likelymeaning that it plays a pivotal role in those pathways. Grey nodes:proteins from the dataset having a match in the database; White nodes:proteins from the database which were not identified (if present) uponthe experimental phase; Grey edges: interactions within a network;Light blue edges: interactions involving HSP70
Stem Cell Rev and Rep (2010) 6:532–547 543

fere with caspase-independent apoptosis [63]. HSP70 is aquintessential regulator of apoptosis that can interfere withall main apoptotic pathways: at the pre-mitochondrial level,HSP70 binds to and blocks c-Jun N-terminal Kinase(JNK1) activity; at the mitochondrial level, HSP70 inhibitsBax translocation and insertion into the outer mitochondrialmembrane; at the post-mitochondrial level HSP70 has beendemonstrated to bind directly to Apaf-1, thereby preventingthe recruitment of procaspase-9 to the apoptosome [61].
Cancer
More recently, literature has been extensively focused on thecontroversial role of HSPs in proliferation and cancer progres-sion. Many cells under a transforming pressure, whethermalignant, neurodegenerative, or infective, co-opt specificHSPs, such as HSP90, in order to maintain the stability andfolding of aberrant, transformation-driving proteins, and toregain a ‘pseudo-stable’ state [88]. As these effects occur in acell-specific and transformation-specific manner, one mayenvision that by inhibition of one protein—again, HSP90—biologic activity may be obtained in a wide-range of diseases.
In this scenario, HSPs have recently become valuabletargets for promising cancer treatments [65]. Indeed, earlyanimal studies have clearly demonstrated that purifiedtumor-derived HSP-peptide complexes can elicit a strongtumor specific immune response and act as a potentmultivalent vaccine [66]. Several key players in cell cyclecontrol systems are known to undergo transient associationwith chaperones [67]. However, it is not yet definitelyassessed whether this association may stimulate or hamperproliferation: for example, transfection of HSP27 to a breastcarcinoma cell line led to a slower proliferation [68],whereas antisense inhibition of HSP27 in MCF-7 cellsinhibited growth [69], and induced the acquisition of asecretory phenotype [70]. Besides, overexpression ofHSP27 in rat colon carcinoma cell clones [71] and inbreast cancer cells [72] enhanced their tumorigenicity, asextensively reviewed by Helmbrecht and Colleagues andbriefly resumed in Table 3 [67].
Alternative strategies involve direct [88] or indirect inhibi-tion of specific HSPs, such as pyrimidinone-peptoid hybrid
molecules with distinct negative modulatory effects on Jdomain stimulation of HSP70 and subsequently observedanti-proliferative activity [89]. Small molecule HSP70 mod-ulators have been suggested to be used for the treatment ofspecific cancers, infections, and protein conformationaldiseases [90].
In the near future, proteomics investigation couldaddress cancer stem cells, specific cells which are foundwithin tumors or hematological cancers and possesscharacteristics associated with normal stem cells, specifi-cally the ability to give rise to all cell types found in aparticular cancer sample. These tumorigenic (tumor-form-ing) cells might share relevant features with UCB-derivedHSCs, such as high levels of HSPs, in parallel to the yetwell established Warburg effect [91].
This brief overview supports findings from pathway andnetwork analyses (previous sections) in which top scorenetworks and pathways (Cell death, tumorigenesis/cancerand Protein folding) enlist a long series of HSPs.
Conclusion
Currently, the main clinical relevance of UCB-derived HSCs isstill closely tied to the field of transplantation, where they arerapidly emerging as a fruitful resource. Nevertheless, enthusi-asms are tempered by the low cell dose available in a single unitand, notwithstanding recent rapid advancements [10–21],solution is far from being at hand. The real deal is the lackof knowledge of the molecular processes which govern cellself-renewal and commitment properties, thus further studiesare required to bridge this gap. In this respect, proteomics hasemerged as a valuable tool, as it has been recently reviewed[22, 92, 93].
A limited number of proteomic studies have recentlyattempted to pave the way for a deeper understanding of the“immature” hematopoietic cell fractions, which have beentargeted through the isolation of CD34+ cells from UCB [25–28]. In integrated transcriptomic and proteomic studies,detected proteins often lacked their corresponding mRNAs,probably meaning that “more immature” proliferating cells donot synthesize many new proteins but focus on preserving yet
Table 3 Roles of HSPs in cell/cycle regulation
Increased amounts of HSPs in proliferating(and transformed) cells
Stimulation of the cell cycle by overexpression, inhibition by suppression of HSP synthesis
Serum/growth factor-induced synthesis Essential function in the initiation of DNA replication by binding to parts of the preinitiationcomplex, similar to what is known for bacteriophages
Protooncogene/oncogene-dependentinduction
Auxiliary function in stabilizing the conformation of active/inactive forms of essential cell cycle orsignal cascade proteins such as Cdk4, pRb, p27, Wee-1, Src, Raf
Virus-stimulated expression Cell cycle phase-dependent synthesis and translocation
Helmbrecht et al. [67]
544 Stem Cell Rev and Rep (2010) 6:532–547

existing ones from denaturing stresses (such as oxidation,starvation) [28]. In this review, integrated and complementaryin silico analyses (pathway and network analyses, GO termenrichment for biological and molecular functions) suggesteda central role for a core of proteins, namely HSPs, chaperones,glycolytic enzymes, cytoskeletal and vesicular transportproteins. These proteins also seem to play a controversialrole in proliferation and self-renewal events in cancer [67] andembryonic stem cells [73]. HSPs are likely involved indifferentiation events as well, since HSPs have been recentlyrelated to protective mechanisms which ultimately lead tohematopoietic progenitor differentiation towards erythroid[76], myeloid [77, 80], or lymphoid lineages [78]. These datacould mean that, in the future, we might try to use them asmore or less direct triggers to switch or bias the proliferationor differentiation programs towards the desired lineage andpopulation volume, in likewise fashion to the recent adoptionof HSP-based autologous cancer treatments/vaccines or HSPsmall molecule inhibitors [90, 91], which are currently underexperimentation on animal models with optimistic results.However, this hypothesis just stands still in the field ofspeculations at the moment.
Basic knowledge has been ultimately expanded by thesepreliminary proteomics investigations, since we are nowallowed to trace out maps of proteins which likely constitutethe molecular fulcrum of the biology in these cells. In thisregard, interactomics analysis of data available from literaturehas proven out a fruitful approach to regroup and analyzeproteomic data from independent studies in an organic andunitary way. Moreover, a unitary map of the interactome hasbeen elaborated upon pooling of data derived from studiesavailable in literature (Supplementary Figure 1), whichsuggests that “immature” cells exhibit “immature” interac-tomic profiles, as the whole network appears to be far morecomplex than the single networks alone. It is very likely thatmore mature cells could end up diminishing not only theiroverall protein expression, as it has already been observed andreviewed [22, 27], but their protein-protein interactions couldbe progressively sculpted as well, in like fashion to the eventswhich mould the early central nervous system [94].
References
1. Brand, A., Rebulla, P., Engelfriet, C. P., et al. (2008). Cord bloodbanking. Vox Sanguinis, 95, 335–348.
2. Gluckman, E., Broxmeyer, H. A., Auerbach, A. D., et al. (1989).Hematopoietic reconstitution in a patient with Fanconi’s anemiaby means of umbilical-cord blood from an HLA identical sibling.The New England Journal of Medicine, 321, 1174–1178.
3. Gluckman, E., Rocha, V., & Boyer-Chammard, A. (1997).Outcome of cord-blood transplantation from related and unre-lated donors. The New England Journal of Medicine, 337, 373–381.
4. Rubinstein, P., Carrier, C., & Scaradavou, A. (1998). Outcomes among562 recipients of placental-blood transplants from unrelated donors.The New England Journal of Medicine, 339, 1565–1577.
5. Rubinstein, P., Rosenfeld, R. E., Adamson, J. W., & Stevens, C. E.(1993). Stored placental blood for unrelated bone marrow reconstitu-tion. Blood, 81, 1679–1690.
6. Armitage, J. O. (1995). Bone marrow transplantation. The NewEngland Journal of Medicine, 330, 827–838.
7. Kurtzberg, J. (2004). Cord blood transplantation in genetic disorders.Biology of Blood and Marrow Transplantation, 10, 735–736.
8. Rocha, V., & Gluckman, E. (2006). Clinical use of umbilical cordblood hematopoietic stem cells eurocord and european blood andmarrow transplant group. Biology of Blood and MarrowTransplantation, 12, 34–41.
9. Brunstein, C. G., Setubal, D. C., & Wagner, J. E. (2007).Expanding the role of umbilical cord blood transplantation.British Journal Haematology, 137, 20–35.
10. Ballen, K., Nademanee, A., & Graham, M. (2007). Outcomes ofdouble cord blood transplantation (DCBT) in a diverse popula-tion. Biology of Blood and Marrow Transplantation, 13, 122.
11. Ballen, K. (2006). Double cord blood transplantation in adults using areduced intensity chemotherapy only conditioning regimen. Biology ofBlood and Marrow Transplantation, 12, 1219.
12. Rubinstein, P. (2006). Why cord blood? Human Immunology, 67,398–404.
13. Xu, R., DeLuca, K., & Porter, W. (2005). In vitro umbilical cordblood expansion resulting in unique CD34Bright cell populationthat engrafts in NOD/SCID mice. Biology of Blood and MarrowTransplantation, 11, 69.
14. Madlambayan, G. J., Rogers, I., Purpura, K. A., et al. ClinicallyRelevant Expansion of Hematopoietic Stem Cells with ConservedFunction in a Single-Use, Closed-System Bioprocess. Biol BloodMarrow Transplant, 12, 1020–1030.
15. Rogers, I. M., Yamanaka, N., & Casper, R. F. (2008). A simplifiedprocedure for hematopoietic stem cell amplification using aserum-free, feeder cell-free culture system. Biology of Blood andMarrow Transplantation, 14, 927–937.
16. Robinson, S. N., Ng, J., Niu, T., et al. (2006). (2006) Superior exvivo cord blood expansion following co-culture with bonemarrow-derived mesenchymal stem cells. Bone MarrowTransplantation, 37, 359–366.
17. Jaroscak, J., Goltry, K., Smith, A., et al. (2003). Augmentation ofumbilical cord blood (UCB) transplantation with ex vivo-expanded UCB cells: results of a phase 1 trial using theAastromReplicell System. Blood, 101, 5061–5067.
18. Pecora, A. L., Stiff, P., Jennis, A., et al. (2000). Prompt anddurable engraftment in two older adult patients with high riskchronic myelogenous leukemia (CML) using ex vivo expandedand unmanipulated unrelated umbilical cord blood. Bone marrowTranplantation, 25, 797–799.
19. Hofmeister, C. C., Zhang, J., Knight, K. L., & Stiff, P. J. (2007).Ex vivo expansion of umbilical cord blood stem cells fortransplantation: growing knowledge from the hematopoietic niche.Bone Marrow Transplantation, 39, 11–23.
20. Koh, L. P., & Chao, N. J. (2004). Umbilical cord bloodtransplantation in adults using myeloablative and nonmyeloabla-tive preparative regimens. Biology of Blood and MarrowTransplantation, 10, 1–22.
21. Brunstein, C.G., Barker, J.N., Weisdorf, D.J., et al. (2009) Intra-BMinjection to enhance engraftment after myeloablative umbilical cordblood transplantation with two partially HLA-matched units. BoneMarrow Transplant. 2009, doi:10.1038/bmt.2008.417.
22. D'Alessandro, A., Liumbruno, G., Grazzini, G., Pupella, S.,Lombardini, L., & Zolla, L. (2009) Umbilical cord blood stemcells: Towards a proteomic approach. J Proteomics doi:10.1016/j.jprot.2009.06.009.
Stem Cell Rev and Rep (2010) 6:532–547 545

23. Benedetti, F. (1996). CD34+ cells: biological aspects. Tumori, 82, 3–13.
24. Lang, P., Bader, P., Schumm, M., et al. (2004). Transplantation of acombination of CD133+ and CD34+ selected progenitor cells fromalternative donors. British Journal Haematology, 124, 72–79.
25. Zenzmaier, C., Kollroser, M., Gesslbauer, B., Jandrositz, A., Preiseg-ger, K. H., & Kungl, A. J. (2003). Preliminary 2-D chromatographicinvestigation of the human stem cell proteome. Biochemical andBiophysical Research Communications, 310, 483–490.
26. Zenzmaier, C., Gesslbauer, B., Grobuschek, N., Jandrositz,A., Preisegger, K. H., & Kungl, A. J. (2005). Proteomicprofiling of human stem cells derived from umbilical cordblood. Biochemical and Biophysical Research Communica-tions, 328, 968–972.
27. Tao, W., Wang, M., Voss, E. D., Cocklin, R. R., Smith, J. A.,Cooper, S. H., et al. (2004). Comparative proteomic analysis ofhuman CD34+ stem/progenitor cells and mature CD15+ myeloidcells. Stem Cells, 22, 1003–1014.
28. Liu, F., Lu, J., Fan, H. H., et al. (2006). Insights into human CD34+hematopoietic stem/progenitor cells through a systematically proteo-mic survey coupled with transcriptome. Proteomics, 6, 2673–2692.
29. D’Alessandro, A., Righetti, P. G., & Zolla, L. (2010). The redblood cell proteome and interactome: an update. Journal ofProteome Research, 9(1), 144–163.
30. Timperio, A. M., D’Alessandro, A., Pariset, L., D’Amici, G. M.,Valentini, A., & Zolla, L. (2009). Comparative proteomics andtranscriptomics analyses of livers from two different Bos taurusbreeds: “Chianina and Holstein Friesian”. Journal of Proteomics, 73(2), 309–322.
31. D’Alessandro, A., Righetti, P.G., Fasoli, E., Zolla, L. (2010) Theegg white and yolk interactomes as gleaned from extensiveproteomic data. J Proteomics. doi:10.1016/j.jprot.2010.01.002.
32. Ingenuity Pathway Analysis (Ingenuity® Systems, www.ingenuity.com).
33. Al-Shahrour, F., Minguez, P., Tárraga, J., et al. (2007). FatiGO+: afunctional profiling tool for genomic data. Integration of func-tional annotation, regulatory motifs and interaction data withmicroarray experiments. Nucleic Acids Research, 35, 91–96.
34. Al-Shahrour, F., Minguez, P., Vaquerizas, J. M., Conde, L., &Dopazo, J. (2005). BABELOMICS: a suite of web-tools forfunctional annotation and analysis of group of genes in high-throughput experiments. Nucleic Acids Research, 33, 460–464.
35. Al-Shahrour, F., Díaz-Uriarte, R., & Dopazo, J. (2004). FatiGO: aweb tool for finding significant associations of gene ontologyterms with groups of genes. Bioinformatics, 20, 578–580.
36. Zolla, L. (2008). Proteomics studies reveal important informationon small molecule therapeutics: a case study on plasma proteins.Drug Discovery Today, 13, 1042–1051.
37. Kanehisa, M., Goto, S., Kawashima, S., Okuno, Y., & Hattori, M.(2004). The KEGG resource for deciphering the genome. NucleicAcids Research, 32, 277–280.
38. BioCarta – charting pathways of life. http://www.biocarta.com/San Diego, CA; USA. Last accessed on the 4th May, 2010.
39. Mulder, N. J., Apweiler, R., Attwood, T. K., et al. (2003). TheInterPro Database, 2003 brings increased coverage and newfeatures. Nucleic Acids Research, 31(1), 315–318.
40. Ramalho-Santos, M., Yoon, S., Matsuzaki, Y., Mulligan, R. C., &Melton, D. A. (2002). “Stemness”: transcriptional profiling ofembryonic and adult stem cells. Science, 298, 597–600.
41. Zhu, X., & Naz, R. K. (1998). Expression of a novel isoform ofcyclin I in human testis. Biochemical and Biophysical ResearchCommunications, 249(1), 56–60.
42. Liu, Y., Tang, M. K., Cai, D. Q., Li, M., Wong, W. M., Chow, P.H., et al. (2007). Cyclin I and p53 are differentially expressedduring the terminal differentiation of the postnatal mouse heart.Proteomics, 7(1), 23–32.
43. Ditton, H.J., Zimmer, J., Kamp, C., Rajpert-De Meyts, E., Vogt, P.H. (2004) The AZFa gene DBY(DDX3Y) is widely transcribedbut the protein is limited to the male germ cells by translationcontrol. Hum Mol Genet. 1, 13(19), 2333–2341.
44. Sugihara, T., Wadhwa, R., Kaul, S. C., & Mitsui, Y. (1999). A noveltestis-specific metallothionein-like protein, tesmin, is an early markerof male germ cell differentiation. Genomics, 57(1), 130–136.
45. Gao, Z., Harumi, T., & Garbers, D. L. (1997). Chromosomelocalization of the mouse zonadhesin gene and the humanzonadhesin gene (ZAN). Genomics, 41(1), 119–122.
46. Kathleen Baxter, K., Uittenbogaard, M., Yoon, J., & Chiar-amello, A. (2009) The neurogenic basic helix-loop-helixtranscription factor NeuroD6 concomitantly increases mito-chondrial mass and regulates cytoskeletal organization in theearly stages of neuronal differentiation. ASN Neuro. 1(4). pii:e00016. doi:10.1042/AN20090036.
47. Fink, J. M., Hirsch, B. A., Zheng, C., et al. (1997). Astrotactin(ASTN), a gene for glial-guided neuronal migration, maps tohuman chromosome 1q25.2. Genomics, 40(1), 202–205.
48. Azuma, N., Yamaguchi, Y., Handa, H., Hayakawa, M., Kanai,A., & Yamada, M. (1999). Missense mutation in the alternativesplice region of the PAX6 gene in eye anomalies. AmericanJournal of Human Genetics, 65(3), 656–663.
49. Salim, A., & Zaidi, Z. H. (2003). Homology models of humangamma-crystallins: structural study of the extensive chargenetwork in gamma-crystallins. Biochemical and BiophysicalResearch Communications, 300(3), 624–630.
50. Unwin, R. D., Smith, D. L., Blinco, D., et al. (2006).Quantitative proteomics reveals posttranslational control as aregulatory factor in primary hematopoietic stem cells. Blood,107, 4687–4694.
51. Gogvadze, V., Zhivotovsky, B., & Orrenius, S. (2009). TheWarburg effect and mitochondrial stability in cancer cells. MolAspects Med. doi:10.1016/j.mam.2009.12.004.
52. Hernandez-Toro, J., Prieto, C., & De Las Rivas, J. (2007).APID2NET: unified interactome graphic analyzer. Bioinformatics,23, 2495–2497.
53. Olsen, J. V., Blagoev, B., Gnad, F., Macek, B., Kumar,C., Mortensen,P., et al. (2006). Global, in vivo, and site-specific phosphorylationdynamics in signaling networks. Cell, 127(3), 635–648.
54. Kajkowski, E. M., Lo, C. F., Ning, X., Walker, S., Sofia, H. J., Wang,W., et al. (2001). beta -Amyloid peptide-induced apoptosis regulatedby a novel protein containing a g protein activation module. TheJournal of Biological Chemistry, 276(22), 18748–18756.
55. Kontush, A. (2001). Alzheimer’s amyloid-beta as a preventiveantioxidant for brain lipoproteins. Cellular and MolecularNeurobiology, 21(4), 299–315.
56. Ritossa, F. (1962). A new puffing pattern induced by heat shockand DNP in Drosophila. Experimentia, 18, 571–573.
57. Wickner, S., Maurizi, M. R., &Gottesman, S. (1999). Posttranslationalquality control: folding, refolding, and degrading proteins. Science,286, 1888–1893.
58. Ellgaard, L., Molinari, M., & Helenius, A. (1999). Setting thestandards: quality control in the secretory pathway. Science, 286,1882–1888.
59. Robert, J. (2003). Evolution of heat shock protein and immunity.Developmental and Comparative Immunology, 27, 449–464.
60. Lindquist, S., & Craig, E. A. (1988). The heat shock proteins.Annual Review of Genetics, 22, 631–637.
61. Lanneau, D., de Thonel, A., Maurel, S., Didelot, C., & Garrido, C.(2007). Apoptosis versus cell differentiation: role of heat shockproteins HSP90, HSP70 and HSP27. Prion, 1, 53–60.
62. Garrido, C., & Solary, E. (2003). A role of HSPs in apoptosis through“protein triage”? Cell Death and Differentiation, 10, 619–620.
63. Garrido, C., Gurbuxani, S., Ravagnan, L., & Kroemer, G.(2001). Heat shock proteins: endogenous modulators of
546 Stem Cell Rev and Rep (2010) 6:532–547

apoptotic cell death. Biochemical and Biophysical ResearchCommunications, 286, 433–442.
64. Parcellier, A., Gurbuxani, S., Schmitt, E., Solary, E., & Garrido,C. (2003). Heat shock proteins, cellular chaperones that modulatemitochondrial cell death pathways. Biochemical and BiophysicalResearch Communications, 304, 505–512.
65. Calderwood, S. K., Khaleqye, A., & Sawyer, D. B. (2006). Heatshock proteins in cancer: chaperones of tumorigenesis. Trends inBiochemical Sciences, 31, 164–72.
66. Wang, H. H., Mao, C. Y., Teng, L. S., & Cao, J. (2006). Recentadvances in heat shock protein-based cancer vaccines. Hepatobil-lary & Pancreatic Diseases International, 5, 22–27.
67. Helmbrecht, K., Zeise, E., & Rensing, L. (2000). Chaperones incell cycle regulation and mitogenic signal transduction: a review.Cell Proliferation, 33, 341–365.
68. Kindas-Mugge, I., Micksche, M., & Trautinger, F. (1998).Modification of growth in small heat shock (HSP27) genetransfected breast carcinoma. Anticancer Research, 18, 413–418.
69. Mairesse, N., Horman, S., Mosselmans, R., & Galand, P. (1996).Antisense inhibition of the 27 kDa heat shock protein productionaffects growth rate and cytoskeletal organization in MCF-7 cells.Cell Biology International, 20, 205–212.
70. Horman, S., Fokan, D., Mosselmans, R., Mairesse, N., & Galand,P. (1999). Anti-sense inhibition of small heat-shock protein(HSP27)expression in MCF-7 mammary-carcinoma cells inducestheir spontaneous acquisition of a secretory phenotype. Interna-tional Journal of Cancer, 12, 574–582.
71. Garrido, C., Fromentin, A., Bonnotte, B., Favre, N., Moutet, M.,Arrigo, A. P., et al. (1998). Heat shock protein 27 enhances thetumorigenicity of immunogenic rat colon carcinoma cell clones.Cancer Research, 58, 5495–5499.
72. Lemieux, P., Osterrich, S., Lawrence, J. A., Steeg, P. S.,Hilsenbeck, S. G., Harvey, J. M., et al. (1997). The small heatshock protein HSP27 increases invasiveness but decreasesmotility of breast cancer cells. Invasion & Metastasis, 17, 113–123.
73. Prinsloo, E., Setati, M. M., Longshaw, V. M., & Blatch, G. L.(2009). Chaperoning stem cells: a role for heat shock proteins inthe modulation of stem cell self-renewal and differentiation?Bioessays, 31, 370–377.
74. Sato, N., Yamamoto, T., Sekine, Y., et al. (2003). Involvementofheat-shockprotein90in the interleukin-6-mediatedsignallingpathway through STAT3. Biochemical and Biophysical ResearchCommunications, 300, 847–852.
75. Longshaw, V. M., Baxter, M., Prewitz, M., & Blatch, G. L.(2009). Knockdown of the co-chaperone Hop promotes extranu-clear accumulation of Stat3 in mouse embryonic stem cells.European Journal of Cell Biology, 88(3), 153–166.
76. Ribeil, J. A., Zermati, Y., Vandekerckhove, J., et al. (2006). Hsp70regulates erythropoiesis by preventing caspase-3-mediated cleav-age of GATA-1. Nature, 445, 102–105.
77. Kol, A., Lichtman, A. H., Finberg, R. W., Libby, P., & Kurt-Jones,E. A. (2000). Cutting edge: heat shock protein (HSP) 60 activatesthe innate immune response: CD15 is an essential receptor forHSP60 activation of mononuclear cells. Journal of Immunology,164, 13–17.
78. Ripley, B. J., Stephanou, A., Isenberg, D. A., & Latchman, D. S.(1999). Interleukin-10 activates heat-shock protein 90beta geneexpression. Immunology, 97, 226–231.
79. Moore, D. C., Carter, D. L., & Studzinski, G. P. (1992). Inhibition by 1,25 dihydroxyvitamin D3 of c-myc down-regulation and DNAfragmentation in cytosine arabinoside-induced erythroid differentiationof K562 cells. Journal of Cellular Physiology, 151, 539–548.
80. Sistonen, L., Sarge, K., Phillips, B., Abravayad, K., & Morimoto,R. (1992). Activation of heat shock factor 2 during hemininduceddifferentiation of human erythroleukemia cells. Molecular andCellular Biology, 12, 4104–4111.
81. Garcia-Bermejo, L., Vilaboa, N. C., Perez, C., deBlas, E., Calle, C., &Aller, P. (1995). Modulation of hsp70 and hsp27 gene expression bythe differentiation inducer sodium butyrate in U-937 human promon-ocytic leukemia cells. Leukemia Research, 19, 713–718.
82. Teshima, S., Rokutan, K., Takahashi, M., Nikawa, T., & Kishi, K.(1996). Induction of heat shock proteins and their possible roles inmacrophages during activation by macrophage colony-stimulatingfactor. The Biochemical Journal, 315, 497–504.
83. Leppa, S., Pirkkala, L., Saarento, H., Sarge, K., & Sistonen,L. (1997). Overexpression of HSF2-b inhibits hemin-inducedheat shock gene expression and erythroid differentiation inK562 cells. The Journal of Biological Chemistry, 272, 15293–15298.
84. Matsui, H., Asou, H., & Inaba, T. (2007). (2007) Cytokines directthe regulation of Bim mRNA stability by heat-shock cognateprotein 70. Molecular Cell, 25, 99–112.
85. Craven, S. E., French, D., Ye, W., de Sauvage, F., & Rosenthal, A.(2005). Loss of Hspa9b in zebrafish recapitulates the ineffectivehematopoiesis of the myelodysplastic syndrome. Blood, 105,3528–3534.
86. Spector, N. L., Ryan, C., Samson, W., Levine, H., Nadler, L. M.,& Arrigo, A. P. (1993). Heat shock protein is a unique marker ofgrowth arrest during macrophage differentiation of HL-60 cells.Journal of Cellular Physiology, 156, 619–625.
87. Ahlers, A., Engel, K., Sott, C., Gaestel, M., Herrmann, F., &Brach, M. A. (1994). Interleukin-3 and granulocyte-macrophagecolony-stimulating factor induce activation of the MAPKAPkinase 2 resulting in in vitro serine phosphorylation of the smallheat shock protein (Hsp 27). Blood, 83, 1791–1798.
88. Solit, D. B., & Chiosis, G. (2008). Development and applicationof Hsp90 inhibitors. Drug Discovery Today, 13(1–2), 38–43.
89. Wright, C. M., Chovatiya, R. J., Jameson, N. E., et al. (2008).Pyrimidinone-peptoid hybrid molecules with distinct effects onmolecular chaperone function and cell proliferation. Bioorganic &Medicinal Chemistry, 16(6), 3291–3301.
90. Brodsky, J. L., & Chiosis, G. (2006). Hsp70 molecular chaper-ones: emerging roles in human disease and identification of smallmolecule modulators. Current Topics in Medicinal Chemistry, 6(11), 1215–1225.
91. Scatena,R.,Bottoni,P.,Pontoglio,A.,&Giardina,B. (2010).Revisitingthe Warburg effect in cancer cells with proteomics. The emergence ofnew approaches to diagnosis, prognosis and therapy. Proteomics-Clinical Application, 2(43), 143–158.
92. Gesslbauer, B., Krenn, E., Zenzmaier, C., Preisegger, K. H., &Kungl, A. J. (2006). Lessons from the stem cell proteome. CurrentStem Cell Research & Therapy, 1, 395–409.
93. Zolla, L. (2008). Proteomics and transfusion medicine. BloodTransfusion, 6, 67–69.
94. Johnston, M. V., Nishimura, A., Harum, K., Pekar, J., & Blue, M.E. (2001). Sculpting the developing brain. Advances in Pediatrics,48, 1–38.
Stem Cell Rev and Rep (2010) 6:532–547 547





























