Embed Size (px)
Citation preview

J Cancer Res Clin Oncol (1984) 108:204-213 C 'cer esearch Clinical @neology �9 Springer-Verlag 1984
In Vivo and In Vitro Binding of 1,2-Dibromoethane and 1,2-Diehloroethane to Macromolecules in Rat and Mouse Organs*
G. Arfellini, S. Bartoli 1, A. Colacci, M. Mazzullo, M. C. Galli, G. Prodi, and S. Grilli
Centro di Cancerogenesi Chimica, Istituto di Cancerologia and 1 Istituto diIstologiaeEmbriologiaGenerale, Universit~idiBologna, I-40126 Bologna, Italy
Summary. The comparative interaction of equimolar amounts of 1,2-dichloroethane and 1,2-dibro- moethane with rat and mouse nucleic acids was stud- ied in both in vivo (liver, lung, kidney and stomach) and in vitro (liver microsomal and/or cytosolic fractions) systems. In vivo, liver and kidney DNA showed the highest labeling, whereas the binding to lung DNA was barely detectable. Dibromoethane was more highly reactive than dichloroethane in both spe- cies. With dichloroethane, mouse DNA labeling was higher than rat DNA labeling whatever the organ con- sidered: the opposite was seen for the bioactivation of dibromoethane. RNA and protein labelings were higher than DNA labeling, with no particular pattern in terms of organ or species involvement. In vitro, in addition to a low chemical reactivity towards nucleic acids shown by haloethanes per se, both compounds were bioactivated by either liver microsomes and cytosolic fractions to reactive forms capable of binding to DNA and polynucleotides. UV irra- diation did not photoactivate dibromoethane and dichloroethane. The in vitro interaction with DNA mediated by enzymatic fractions was PB-inducible (one order of magnitude, using rat microsomes). In vi- tro bioactivation of haloethanes was mainly per- formed by microsomes in the case of dichloroethane
* Supported by a grant from Ministero della Sanitfi, Rome, "Piano di ricerca nel campo delle malattie sociali", no. 500.4/RSC/135/L/ 1208
Offprints requests to: Prof. S. Grilli, Centro di Cancerogenesi Chimi- ca, Istituto di Cancerologia dell'Universitgt, viale Filopanti 22, 1-40126 Bologna, Italy Abbreviations: CBI, covalent binding index; EDTA, ethylenedia- minetetraacetate; GSH, glutathione, reduced form; NADP, nicotinamide adenine dinucleotide phosphate; NADPH, nicotina- mide adenine dinucleotide phosphate, reduced fdtr~ PB, phenobar- bitone; poly (A), polyadenylic acid; poly (C), pot~eytidylic acid; poly (G), polyguanylic acid; poly (U), polyuridylic acid; POPOP, 1,4- bis[2-(5-phenyloxazolyl)]-benzene; PPO, diphenyloxazole; Tris, tris(hydroxymethyl)aminomethane; UV, ultraviolet
and by cytosolic fractions in the case of di- bromoethane. When microsomes plus cytosol were used, rat enzymes were more efficient than mouse enzymes in inducing a dibromoethane-DNA inter- action: the opposite situation occurred for di- chloroethane-DNA interaction, and this is in agree- ment with the in vivo pattern. In the presence of both metabolic pathways, addition or synergism occurred. Dibromoethane was always more reactive than dichloroethane. An indication of the presence of a mi- crosomal GSH transferase was achieved for the acti- vation of dibromoethane. No preferential binding in vitro to a specific polynucleotide was found. Polynu- cleotide labeling was higher than (or equal to) DNA binding. The labeling of microsomal RNA and pro- teins and of cytosolic proteins was many times lower than that of DNA or polynucleotides. The in vivo and in vitro data reported above give an unequivocal indi- cation of the relative reactivity of the haloethanes ex- amined with liver macromolecules from the two spe- cies and agree, on the whole, with the relative genotoxicity (DNA repair induction ability, mutage- nicity and carcinogenicity) of the chemicals.
Key words: 1,2-Dibromoethane and 1,2-dichloro- ethane - Nucleic acid interaction In vivo and in vitro systems - Comparison of haloethanes - Mouse and rat
Introduction
Both 1,2-dichloroethane and 1,2-dibromoethane are the subject of great interest and both are produced in remarkable amounts (more than 5 million tons and nearly 200 million kg, respectively). Dibromoethane is used mainly as a gasoline additive (lead scavenger), as a solvent, and as a fumigant for a variety of vegetable,

G. Arfellini et al.: Binding of 1,2-Dibromoethane and 1,2-Dichloroethane to Macromolecules in Rat and Mouse Organs 205
fruit, and gra in crops dur ing storage ( I A R C 1977). Dich loroe thane is ma in ly used in the p roduc t ion of vinylchlor ide m o n o m e r f rom ethylene but , as di- b romoe thane , is also employed as gasoline addit ive and fumigan t ( I A R C 1979). These chemicals are, therefore, diffused to the env i ronment . Their ge- notoxici ty has been tested in terms of metabo l i sm ( R a n n u g 1980), D N A damage and repair (Perocco and Prodi 1981), mutagenic i ty [in phages, micro-or- ganisms, higher plants , Drosophi la , m a m m a l i a n sys- tems ( R a n n u g 1980) and h u m a n cells (Ferreri et al.
1983)], and their abil i ty to induce various tumors (skin, s tomach, lung, spleen, liver, and breast) in rodents ( I A R C 1977, 1979, 1982; R a n n n u g 1980). These last studies showed that d ib romoe thane , which gives rise ma in ly to local tumors , is more active than dichloro- ethane, which induces dis tant tumors , and that mouse is more sensitive to d ichloroethane than rat (Weis- burger 1977; I A R C 1979).
A few studies reported to covalent b ind ing of di- b r o m o e t h a n e or d ich loroe thane to macromolecules ei- ther in vivo (Nachtomi and Sarma 1977; Hill et al. 1978; Reitz et al. 1980) or in vitro (Hill et al. 1978; Banerjee and Van D u u r e n 1979; Banerjee et al. 1980; Guenger ich et al. 1980; Sundhe imer et al. 1982).
In the present work, our a im was a systematic com- par ison of the metabol ic condi t ions of b ind ing of the two haloethanes to macromolecules in re la t ion to their careinogenici ty in the ra t and in the mouse. Therefore, the react ion of equ imolar amoun t s of haloethanes
with D N A , R N A , and proteins of the two species was measured in bo th in vivo and in vitro systems.
Materials and Methods
Chemicals and Radiochemicals
DNA, polynucleotides, NADPH, NADP, glucose-6-phosphate, glu- cose-6-phosphate dehydrogenase, ribonuclease I, sodium dodecyl sulfate, and GSH were purchased from Sigma Chemical Co., St Louis, Mo., USA; phenobarbitone (PB) from Carlo Erba, Milan, Italy; lumasolve from Lumac, Basel, Switzerland; and ready-solv MP from Beckman, Milan, Italy. Other chemicals, all of analytical grade, were obtained from Merck, Drehier, RFG. [U-I~C]I,2- Dichloroethane (23mCi/mmol) and [U-14C]l,2-dibromoethane (14.6 mCi/mmol) were purchased from the Radioehemical Centre, Amersham, UK. The chemical purity of tracers was greater than 99% and their radiochemical purity was greater than 98% according both to the supplier's analysis and to our double check performed be- fore tracer use by chromatography on silica gel 60 (70-230 mesh) (Merck) columns (1.5 • 30 cm) with cold (0 ~ n-pentane-ether (9:1) as eluant. The specific activity of [14C]dichloroethane was lowered to 14.6 mCi/mmol by adding the proper amount of cold haloethane to the tracer.
Animals
Male inbred Wistar rats (about 300 g) and BALB/c mice (22-28 g) were obtained from Morini, S. Polo d'Enza, Italy and Charles River, Calco, Italy, respectively. The animals were housed in macrolon cages at 22 ~ with 12 h light and 12 h darkness, receiving a standard
pellet diet (Vogt-M611er) purchased from Piccioni, Brescia, Italy and drinking water ad libitum.
In Vivo Binding to DNA, RNA, and Proteins
Two rats and eight mice (28 g) received 127 ~tCi (8.7 ~tmol) a4C-la- beled haloethane/kg body wt in ethanol IP (0.28 and 0.05 ml for rat and mouse, respectively). Soon afterwards, animals were kept without food and killed 22 h after injection. To obtain DNA, RNA, and proteins, livers, kidneys, lungs and stomachs were removed, pooled, and processed according to the phenol method of Kinoshita and Gelboin (1972). Fractions were exhaustively washed with or- ganic solvents until no radioactivity was extracted from macromol- ecules. The purity of these macromolecules was tested by colorimet- ric reactions [Biorad dye (Bradford 1976), diphenylamine (Burton 1956), and orcinol (Schneider 1957)] and ultraviolet (UV) absorp- tion: in particular, protein removal from nucleic acids was always achieved. Scintillation liquid (10 ml; 0.4% PPO and 0.005% POPOP in toluene) was added to aliquots of DNA, RNA, and proteins (1- 6 mg) dissolved in 1 ml lumasolve. Duplicate countings were per- formed in an Intertechnique SL 32 spectrometer (Plaisir, France): each vial was counted with an efficiency of about 88% for a long enough time to allow counting of at least 4,000 counts with a back- ground of 32 c.p.m.
In Vitro Interaction with Macromolecules
All the experiments described below were performed in duplicate.
Chemical Reactivity. Interaction was achieved by incubating 1.2 ~Ci 14C-labeled haloethane and 2 mg DNA in a final volume of 3 ml 25 mM Tris buffer, pH 7.0, containing 0.1 M NaC1 and 10 mM ED- TA, made 0.1% ethanol, at 0 ~ 22 ~ and 40 ~ for 2 h. More- over, the reactivity of haloethanes towards DNA and polyribonu- cleotides was assessed under the same conditions of in vitro enzyme- mediated binding (see below), i.e., 1.5 mg DNA and 2.5 ~tCi 14C-la- beled haloethane were allowed to interact in 3 ml 80 m M potassium phosphate buffer, pH 7.7, containing 5 mM MgC12 (hereafter re- ferred to as phosphate-MgCl2), made 0.2% ethanol, at 37 ~ for 90 min in air in the absence of both enzymes and cofactors. DNA and polynucleotides were then precipitated by adding 2-3 vol. cold ethanol to aqueous layers in the presence of sodium acetate, and re- peatedly washed with ethanol and ether until no radioactivity was present in the supernatants: nucleic acid purity and labeling were then measured.
Photoactivation. DNA (2mg) and tracer (2 ~tCi) dissolved in a final volume of 3 ml 25 mM Tris buffer, pH 7.0, containing 0.1 M NaC1 and 10raM EDTA, made 0.17% ethanol, were subjected to UV irradiation with a beam of 310-395 nm (2max=365 nm) produced by a meanpressure mercury vapor lamp (Hanau Q-400, RFG) (Sovirel filter) into a closed spectrophotometric quartz cell (thickness: 1 cm) placed at 35 cm from the irradiation source. The incident fluence rate of the beam, measured by ferric oxalate acti- nometer (Balzani et al. 1968), was about, 1,790J/cmZ/min. The irradiation was performed at 22 ~ for 0, 1, 2, and 3 h in air and the reaction was stopped by quick chilling at 0 ~ As a further control, a 3-h incubation was performed in the dark under otherwise identical experimental conditions. DNA labeling was then deter- mined as described in the previous section.
Enzyme-Mediated A~ztivation
Isolation of L~ver Enzymatic Fractions. Microsomes and cytosol were obtained (Guengerich 1977) from livers of 4 rats and 22 mice (22 g), pretreated or not with PB [100 mg/kg/day, IP, dissolved in sterile 0.9% NaC1 solution (0.5 and 0.2 ml for rat and mouse, respectively)] during the 2 days prior to sacrifice (Hill et al. 1978) to obtain induced
206 G. Arfellini et al.: Binding of 1,2-Dibromoethane and 1,2-Dichloroethane to Macromolecules in Rat and Mouse Organs
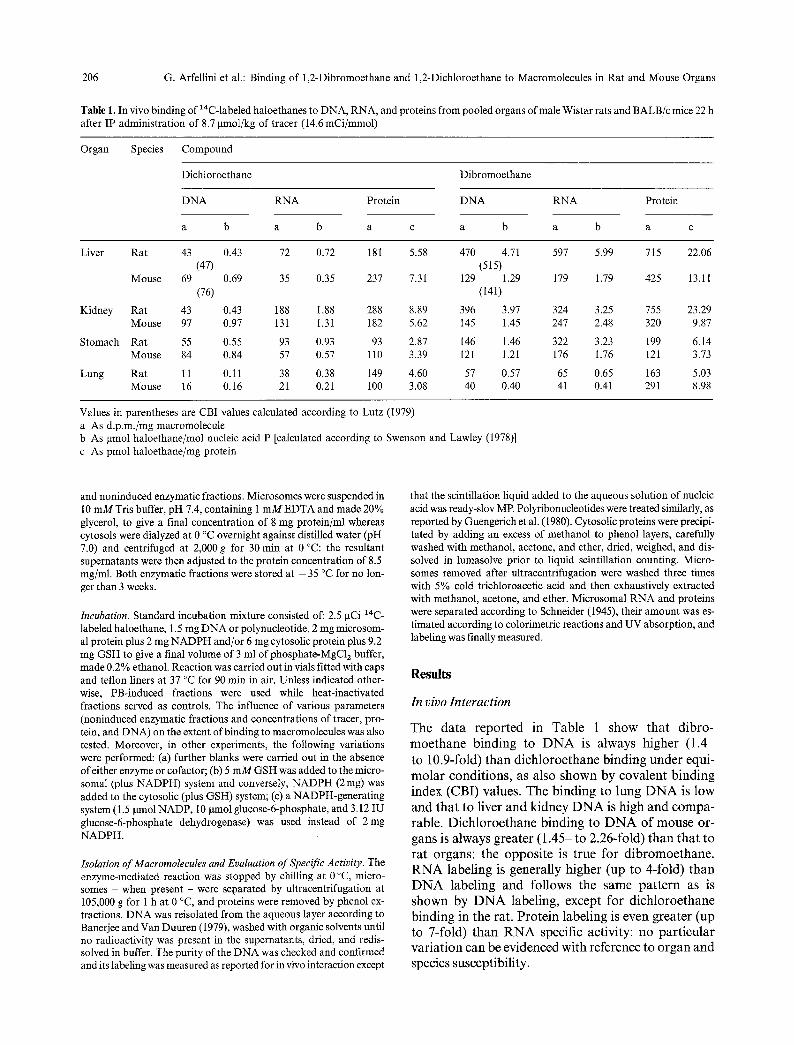
Table 1. In vivo binding of 14C-labeled haloethanes to DNA, RNA, and proteins from pooled organs of male Wistar rats and BALB/c mice 22 h after IP administration of 8.7 gmol/kg of tracer (14.6 mCi/mmol)
Organ Species Compound
Dichloroethane Dibromoethane
DNA RNA Protein DNA RNA Protein
a b a b a c a b a b a c
Liver Rat 43 0.43 72 0.72 181 5.58 470 4.71 597 5.99 715 22.06 (47) (515)
Mouse 69 0.69 35 0.35 237 7.31 129 1.29 179 1.79 425 13.11 (76) (141)
Kidney Rat 43 0.43 188 1.88 288 8.89 396 3.97 324 3.25 755 23.29 Mouse 97 0.97 131 1.31 182 5.62 145 1.45 247 2.48 320 9.87
Stomach Rat 55 0.55 93 0.93 93 2.87 146 1.46 322 3.23 199 6.14 Mouse 84 0.84 57 0.57 110 3.39 121 1.21 176 1.76 121 3.73
Lung Rat 11 0.11 38 0.38 149 4.60 57 0.57 65 0.65 163 5.03 Mouse 16 0.16 21 0.21 I00 3.08 40 0.40 41 0.41 291 8.98
Values in parentheses are CBI values calculated according to Lutz (1979) a As d.p.m./mg macromolecule b As gmol haloethane/mol nucleic acid P [calculated according to Swenson and Lawley (1978)] c As pmol haloethane/mg protein
and noninduced enzymatic fractions. Microsomes were suspended in 10 mM Tris buffer, pH 7.4, containing 1 mM EDTA and made 20% glycerol, to give a final concentration of 8 mg protein/ml whereas cytosols were dialyzed at 0 ~ overnight against distilled water (pH 7.0) and centrifuged at 2,000 g for 30 min at 0 ~ the resultant supernatants were then adjusted to the protein concentration of 8.5 mg/ml. Both enzymatic fractions were stored at - 3 5 ~ for no lon- ger than 3 weeks.
Incubation. Standard incubation mixture consisted off 2.5 gCi 14C- labeled haloethane, 1.5 mg DNA or polynucleotide, 2 mg microsom- al protein plus 2 mg NADPH and/or 6 mg cytosolic protein plus 9.2 mg GSH to give a final volume of 3 ml of phosphate-MgC12 buffer, made 0.2% ethanol. Reaction was carried out in vials fitted with caps and teflon liners at 37 ~ for 90 rain in air. Unless indicated other- wise, PB-induced fractions were used while heat-inactivated fractions served as controls. The influence of various parameters (noninduced enzymatic fractions and concentrations of tracer, pro- tein, and DNA) on the extent of binding to macromolecules was also tested. Moreover, in other experiments, the following variations were performed: (a) further blanks were carried out in the absence of either enzyme or cofactor; (b) 5 mM GSH was added to the micro- soma1 (plus NADPH) system and conversely, NADPH (2mg) was added to the cytosolic (plus GSH) system; (c) a NADPH-generating system (1.5 gmol NADP, 10 gmol glucose-6-phosphate, and 3.12 IU glucose-6-phosphate dehydrogenase) was used instead of 2 mg NADPH.
Isolation of Macromolecules and Evaluation of Specific Activity. The enzyme-mediated reaction was stopped by chilling at 0 ~ micro- somes - when present - were separated by ultracentrifugation at 105,000 g for 1 h at 0 ~ and proteins were removed by phenol ex- tractions. DNA was reisolated from the aqueous layer according to Banerjee and Van Duuren (1979), washed with organic solvents until no radioactivity was present in the supernatants, dried, and redis- solved in buffer. The purity of the DNA was checked and confirmed and its labeling was measured as reported for in vivo interaction except
that the scintillation liquid added to the aqueous solution of nucleic acid was ready-slov MR Polyribonucleotides were treated similarly, as reported by Guengerich et al. (1980). Cytosolic proteins were precipi- tated by adding an excess of methanol to phenol layers, carefully washed with methanol, acetone, and ether, dried, weighed, and dis- solved in lumasolve prior to liquid scintillation counting. Micro- somes removed after ultracentrifugation were washed three times with 5% cold trichloroacetic acid and then exhaustively extracted with methanol, acetone, and ether. Microsomal RNA and proteins were separated according to Schneider (1945), their amount was es- timated according to colorimetric reactions and UV absorption, and labeling was finally measured.
Results
In vivo Interaction
T h e d a t a r e p o r t e d in T a b l e 1 s h o w t h a t d ib ro -
m o e t h a n e b i n d i n g to D N A is a lways h i g h e r (1 .4 -
to 10.9-fold) t h a n d i c h l o r o e t h a n e b i n d i n g u n d e r equ i -
m o l a r c o n d i t i o n s , as a lso s h o w n by c o v a l e n t b i n d i n g
i ndex (CBI ) va lues . T h e b i n d i n g to l u n g D N A is l ow
a n d t h a t to l iver a n d k i d n e y D N A is h i g h a n d c o m p a -
rable . D i c h l o r o e t h a n e b i n d i n g to D N A o f m o u s e or -
gans is a lways g r ea t e r ( 1 . 4 5 - to 2 .26- fo ld) t h a n t h a t to r a t o rgans : the o p p o s i t e is t rue fo r d i b r o m o e t h a n e .
R N A labe l ing is g e n e r a l l y h i g h e r (up to 4- fo ld) t h a n
D N A labe l ing a n d fo l l ows the s a m e p a t t e r n as is
s h o w n by D N A labe l ing , e x c e p t fo r d i c h l o r o e t h a n e
b i n d i n g in the rat . P r o t e i n l abe l ing is e v e n g r ea t e r (up to 7- fo ld) t h a n R N A speci f ic ac t iv i ty : n o p a r t i c u l a r v a r i a t i o n c a n be e v i d e n c e d w i t h r e fe rence to o r g a n a n d
species suscept ib i l i ty .
G. Arfellini et al.: Binding of 1,2-Dibromoethane and 1,2-Dichloroethane to Macromolecules in Rat and Mouse Organs 207
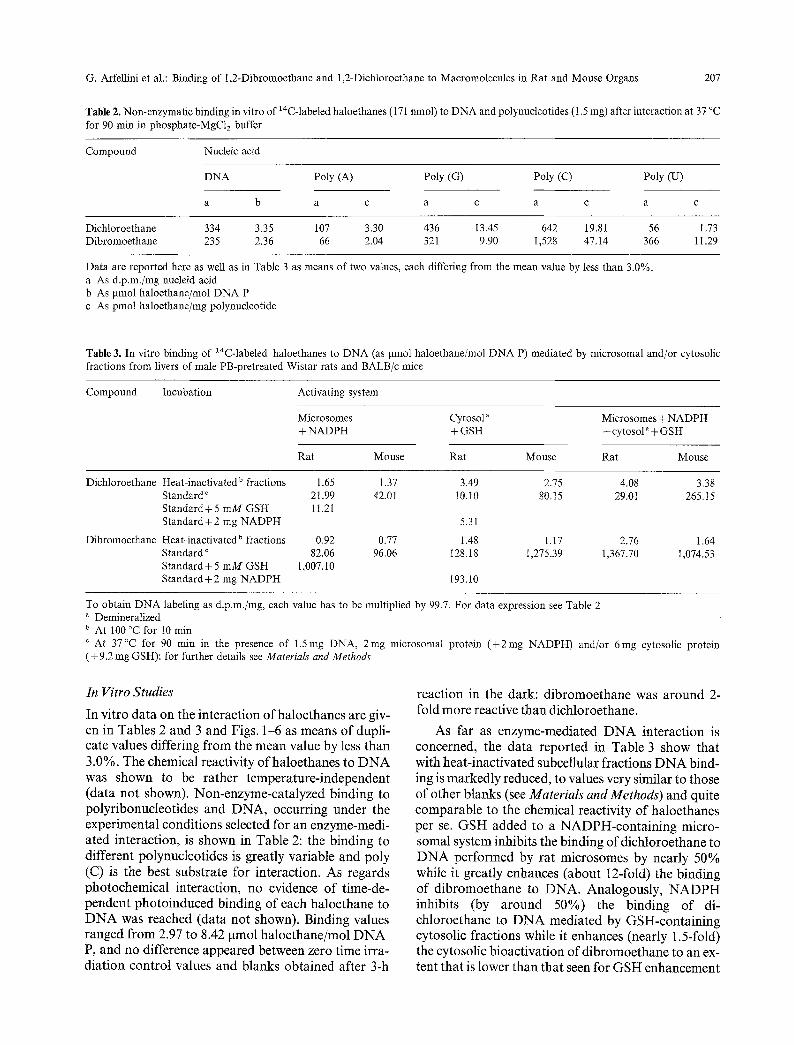
Table 2. Non-enzymatic binding in vitro of 14C-labeled haloethanes (171 nmol) to DNA and polynucleotides (1.5 mg) after interaction at 37 ~ for 90 rain in phosphate-MgCI2 buffer
Compound Nucleic acid
DNA Poly (A) Poly (G) Poly (C) Poly (U)
a b a c a c a c a c
Dichloroethane 334 3.35 107 3.30 436 13.45 642 19.81 56 1.73 Dibromoethane 235 2.36 66 2.04 321 9.90 1,528 47.14 366 11.29
Data are reported here as well as in Table 3 as means of two values, each differing from the mean value by less than 3.0%. a As d.p.m./mg nucleid acid b As pmol haloethane/mol DNA P c As pmol haloethane/mg polynucleotide
Table 3. In vitro binding of 14C-labeled haloethanes to DNA (as gmol haloethane/mol DNA P) mediated by microsomal and/or cytosolic fractions from livers of male PB-pretreated Wistar rats and BALB/c mice
Compound Incubation Activating system
Microsomes Cytosol ~ Microsomes + NADPH + NADPH + GSH + cytosol a § GSH
Rat Mouse Rat Mouse Rat Mouse
Dicbloroethane
Dibromoethane
Heat-inactivated b fractions 1.65 1.37 3.49 2.75 4.08 3.38 Standard ~ 21.99 42.01 10.10 80.15 29.0t 265.15 Standard+5 mM GSH I1.21 Standard+2 mg NADPH 5.31
Heat-inactivated b fractions 0.92 0.77 1.48 1.17 2.76 1.64 Standard c 82.06 96.06 128.18 1,275.39 1,367.70 1,074.53 Standard+5 mM GSH 1,007.10 Standard+2 mg NADPH 193.10
To obtain DNA labeling as d.p.m./mg, each value has to be multiplied by 99.7. For data expression see Table 2 " Demineralized b At 100 ~ for 10 rain ~ At 37~ for 90 min in the presence of 1.5mg DNA, 2mg microsomal protein (+2rag NADPH) and/or 6rag cytosolic protein (+ 9.2 mg GSH): for further details see Materials and Methods
In Vitro Studies
In vitro data on the interaction of haloethanes are giv- en in Tables 2 and 3 and Figs. 1-6 as means of dupli- cate values differing from the mean value by less than 3.0%. The chemical reactivity ofhaloethanes to DNA was shown to be rather temperature-independent (data not shown). Non-enzyme-catalyzed binding to polyribonucleotides and DNA, occurring under the experimental conditions selected for an enzyme-medi- ated interaction, is shown in Table 2: the binding to different polynucleotides is greatly variable and poly (C) is the best substrate for interaction. As regards photochemical interaction, no evidence of time-de- pendent photoinduced binding of each haloethane to DNA was reached (data not shown). Binding values ranged from 2.97 to 8.42 gmol haloethane/mol DNA P, and no difference appeared between zero time irra- diation control values and blanks obtained after 3-h
reaction in the dark: dibromoethane was around 2- fold more reactive than dichloroethane.
As far as enzyme-mediated DNA interaction is concerned, the data reported in Table 3 show that with heat-inactivated subcellular fractions DNA bind- ing is markedly reduced, to values very similar to those of other blanks (see Materials and Methods) and quite comparable to the chemical reactivity of haloethanes per se. GSH added to a NADPH-containing micro- somal system inhibits the binding of dichloroethane to DNA performed by rat microsomes by nearly 50% while it greatly enhances (about 12-fold) the binding of dibromoethane to DNA. Analogously, NADPH inhibits (by around 50%) the binding of di- chloroethane to DNA mediated by GSH-containing cytosolic fractions while it enhances (nearly 1.5-fold) the cytosolic bioactivation of dibromoethane to an ex- tent that is lower than that seen for GSH enhancement

208 G. Arfellini et al.: Binding of 1,2-Dibromoethane and 1,2-Dichloroethane to Macromolecules in Rat and Mouse Organs
A B r 2
~176176 j A RNA proteins RNA proteins A
- - Rat
J C l 2
I [ - - M o u s e - - - - "1
proteins DNA RNA proteins
B r 2 C I 2
" - I" R a t I [ M o u s e I I R a t ] [ - - M o u s e - ' - - - 1 I O 0
13" D N A proteins DNA proteins DNA proteins DNA proteins
Fig. l A, B. Effect of pretreatment with PB (100 mg/kg) on in vitro bioactivation of [14C]dibromoethane (Br2) and [14C]dichloroethane (C12) by rat and mouse liver microsomes (A) and cytosol (B) to forms capable of interacting with calf thymus DNA and macromolecular constituents of subcellular fractions. Data collected with noninduced fractions (BB) are reported as % of net values (total binding minus blanks) obtained with PB-induced (tz) enzymatic fractions (i.e., the standard incubation procedure which is given as 100) from duplicate trials: variation of each single value from the mean was always less than 3.0%. For absolute values of labeling see Table 3. NADH and GSH were added to systems A and B, respectively: for further details see Materials andMethods
of microsomal activation. Enzyme-catalyzed binding of dibromoethane to DNA is higher than that of dichloroethane, and mouse enzymes are generally more efficient than rat enzymes. Dichloroethane- DNA binding is mainly due to the microsomal path- way, whereas the dibromoethane-DNA interaction is mainly mediated by cytosolic enzymes. When the two activating systems are present simultaneously, addi- tion or synergism occurs, according to species or haloethane. Labeling of macromolecules from micro- somes (RNA and proteins) and cytosol (proteins), which is markedly reduced by heating, follows the or- der: microsomal protein > microsomal RNA > cytosolic protein (see Fig. 5). Interestingly, RNA and protein labeling is noticeably reduced in the presence of both metabolic pathways: the opposite is true for DNA interaction leading to much higher (1-2 orders of magnitude) values. Moreover (data not shown), the NADPH-generating system is less effective (by nearly 30%) than standard incubation conditions, where pre- formed NADPH is fully available at time zero.
Figure 1 reports the effectiveness of PB-induced and noninduced subcellular fractions from liver of both species in activating haloethanes to forms ca- pable of interacting with DNA. An enzymatic induc- tion occurs regardless of either haloethane or macro- molecular binding acceptor considered: such an effect is more marked when rat microsomes are used. It should be explained that the data on noninduced fractions reported in Fig. 1 are given as percentages of
the values found with PB-induced fractions, i.e., the standard incubation procedure also used to provide reference values for Figs. 2-4. The increase in the amount of activating enzymes leads to more DNA binding but less binding to RNA and proteins (data not shown).
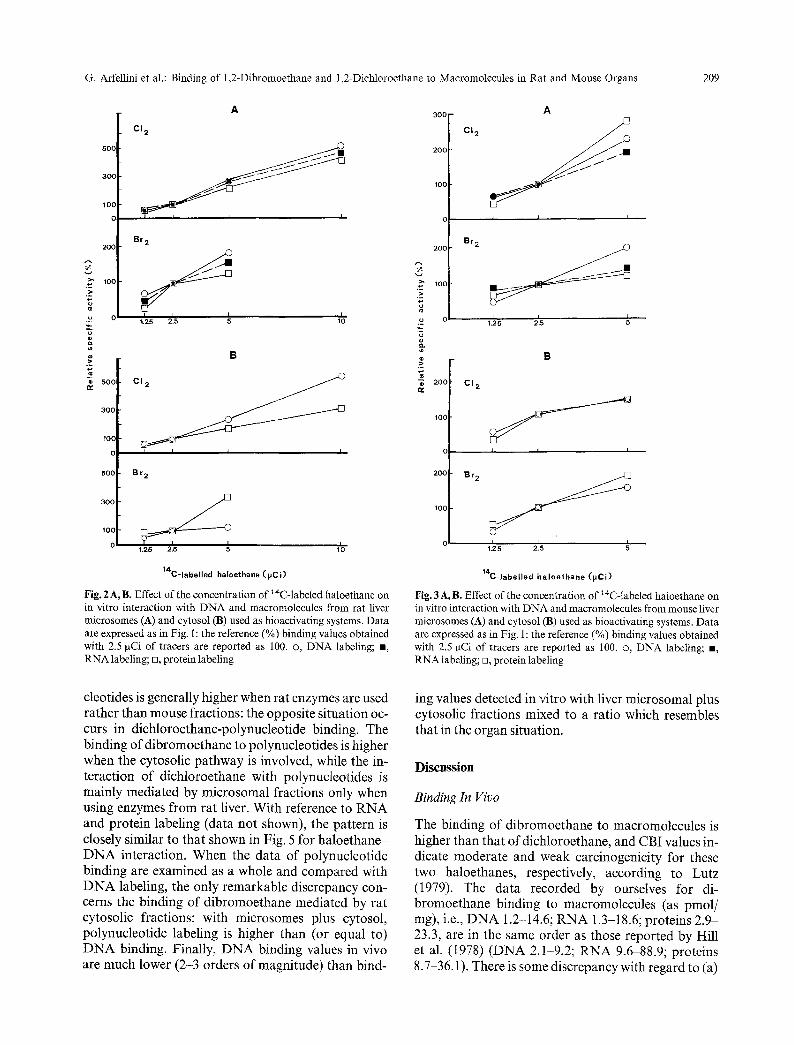
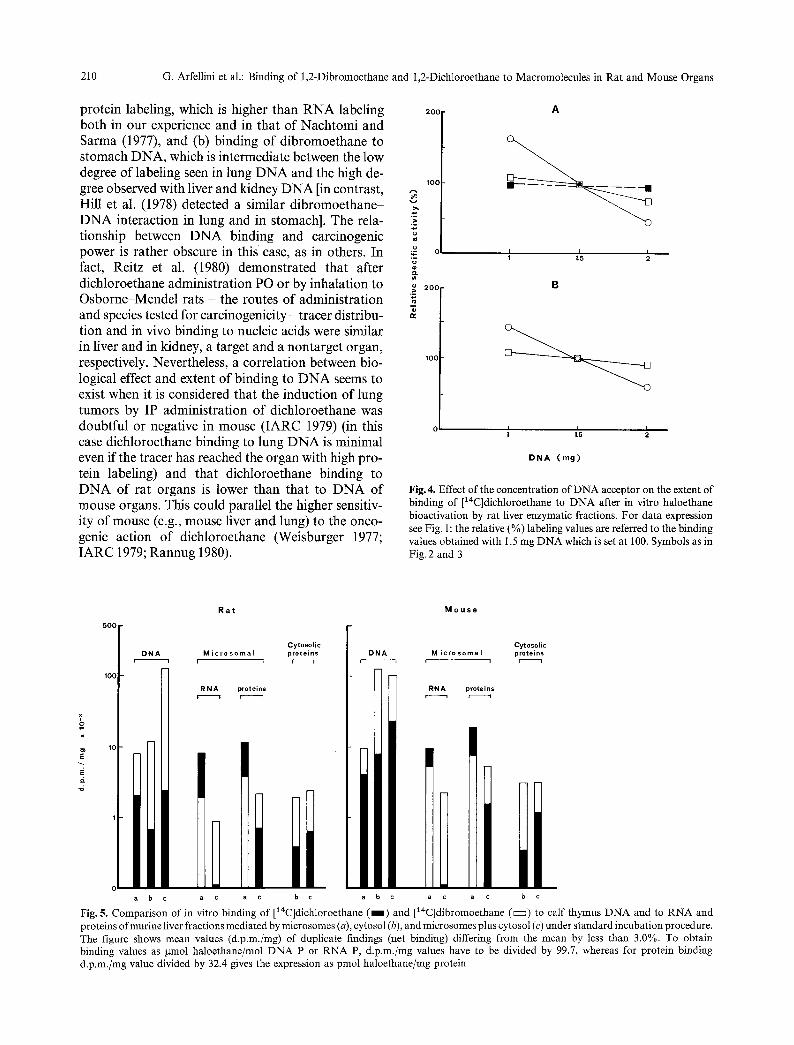
Figure 2 and 3 show that there is a positive correla- tion between extent of binding and tracer concentra- tion, whereas an inverse relationship with the amount of DNA acceptor is observed in Fig. 4.
Figure 5 evidences the whole pattern ofhaloethane binding (as absolute values) to DNA and to macromo- lecular constituents of subcellular fractions, mediated by microsomes and/or cytosol from rat and mouse liver. Although the main results have been summa- rized above, Fig. 5 is included to facilitate the compar- ison on DNA binding values with polynucleotide labelings, which are shown in Fig. 6.
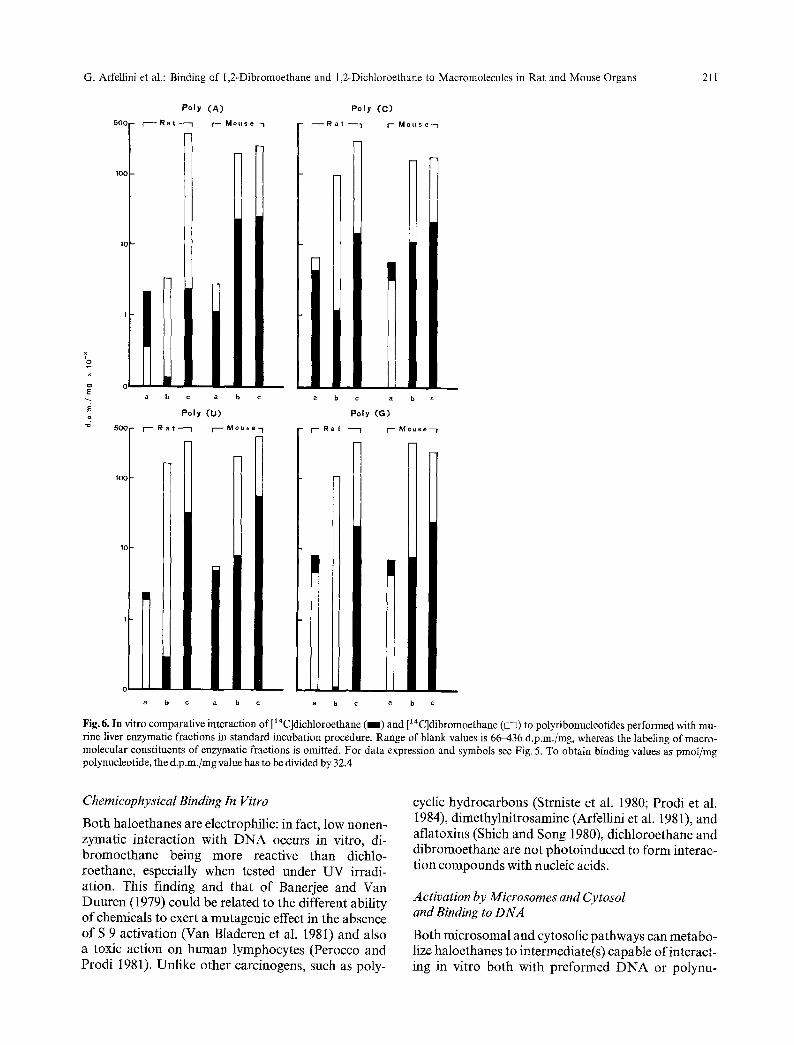
Figure 6 shows that poly (G) is one of the most heavily labeled substrates for microsome-mediated binding. With cytosol, the most heavily labeled polynucleotide varies according to either the haloethane or the species considered. Poly (U) is gen- erally the best binding acceptor in the presence of both microsomes and cytosol. Mouse cytosolic fractions are more efficient than rat cytosolic enzymes. With cytosol alone or with microsomes plus cytosol, the en- zyme-catalyzed binding of dibromoethane is higher than that of dichloroethane. With microsomes plus cytosol, the binding of dibromoethane to polynu-
G. Arfellini et al.: Binding of 1,2-Dibromoethane and 1,2-Dichloroethane to Macromolecules in Rat and Mouse Organs 209
500
30C
A
C l 2
lOC
(] o
B r 2 200 200
10C >" 100
.> ,>
u I I I I " - 1.25 2.5 5 10 .U 0
U U
r ca
500 -~ 200
3 0 0
100
0
500
300
100
B
I I I I
B r 2
1.25 2.5 5 1~3
14C- label led haloethane ( I J C i )
F i g . 2 A , B. E f f e c t o f t h e c o n c e n t r a t i o n o f 14C- l abe l ed h a l o e t h a n e o n
in vitro interaction with DNA and macromolecules from rat liver microsomes (A) and cytosol (B) used as bioactivating systems. Data are expressed as in Fig. 1: the reference (%) binding values obtained with 2.5 gCi of tracers are reported as 100. o, DNA labeling; B, RNA labeling; n, protein labeling
300
CI 2
200
100
Br 2
100
0
200
lOO
C I 2
B r 2
A
I [ P
1;~ 2'.5
1.;5 2!5
14C- labe l led h a l o e t h a n e ( p C i )
Fig. 3 A,B. Effect of the concentration of ~4C-labeled haloethane on in vitro interaction with DNA and macromolecules from mouse liver microsomes (A) and cytosol (B) used as bioactivating systems. Data are expressed as in Fig. 1: the reference (%) binding values obtained with 2.5 rtCi of tracers are reported as 100. o, DNA labeling; e, RNA labeling; n, protein labeling
cleotides is generally higher when rat enzymes are used rather than mouse fractions: the opposite situation oc- curs in dichloroethane-polynucleotide binding. The binding of dibromoethane to polynucleotides is higher when the cytosolic pathway is involved, while the in- teraction of dichloroethane with polynucleotides is mainly mediated by microsomal fractions only when using enzymes from rat liver. With reference to RNA and protein labeling (data not shown), the pattern is closely similar to that shown in Fig. 5 for haloethane- D N A interaction. When the data of polynucleotide binding are examined as a whole and compared with D N A labeling, the only remarkable discrepancy con- cerns the binding of dibromoethane mediated by rat cytosolic fractions: with microsomes plus cytosol, polynucleotide labeling is higher than (or equal to) D N A binding. Finally, D N A binding values in vivo are much lower (2-3 orders of magnitude) than bind-
ing values detected in vitro with liver microsomal plus cytosolic fractions mixed to a ratio which resembles that in the organ situation.
Discuss ion
Binding In Vivo
The binding of dibromoethane to macromolecules is higher than that of dichloroethane, and CBI values in- dicate moderate and weak carcinogenicity for these two haloethanes, respectively, according to Lutz (1979). The data recorded by ourselves for di- bromoethane binding to macromolecules (as pmol/ mg), i.e., DNA 1.2-14.6; RNA 1.3-18.6; proteins 2.9- 23.3, are in the same order as those reported by Hill et al. (1978) (DNA 2.1-9.2; RNA 9.6-88.9; proteins 8.7 36.1). There is some discrepancy with regard to (a)
210 G. Arfellini et al.: Binding of 1,2-Dibromoethane and 1,2-Dichloroethane to Macromolecules in Rat and Mouse Organs
protein labeling, which is higher than RNA labeling both in our experience and in that of Nachtomi and Sarma (1977), and (b) binding of dibromoethane to stomach DNA, which is intermediate between the low degree of labeling seen in lung DNA and the high de- gree observed with liver and kidney DNA [in contrast, Hill et al. (1978) detected a similar dibromoethane- DNA interaction in lung and in stomach]. The rela- tionship between DNA binding and carcinogenic power is rather obscure in this case, as in others. In fact, Reitz et al. (1980) demonstrated that after dichloroethane administration PO or by inhalation to Osborne-Mendel rats - the routes of administration and species tested for carcinogenicity- tracer distribu- tion and in vivo binding to nucleic acids were similar in liver and in kidney, a target and a nontarget organ, respectively. Nevertheless, a correlation between bio- logical effect and extent of binding to DNA seems to exist when it is considered that the induction of lung tumors by IP administration of dichloroethane was doubtful or negative in mouse (IARC 1979) (in this case dichloroethane binding to lung DNA is minimal even if the tracer has reached the organ with high pro- tein labeling) and that dichloroethane binding to DNA of rat organs is lower than that to DNA of mouse organs. This could parallel the higher sensitiv- ity of mouse (e.g., mouse liver and lung) to the onco- genic action of dichloroethane (Weisburger 1977; IARC 1979; Rannug 1980).
2 0 0 A
-g
o
1 0 0
2 0 C
100
,!s ;
B
o 1 ,!~ �9
D N A ( r n g )
Fig. 4. Effect of the concentration of DNA acceptor on the extent of binding of [14C]dichloroethane to DNA after in vitro haloethane bioactivation by rat liver enzymatic fractions. For data expression see Fig. 1: the relative (%) labeling values are referred to the binding values obtained with 1.5 mg DNA which is set at 100. Symbols as in Fig. 2 and 3
5 0 0
100
R a t
C y t o s o l i c D N A M i c r o s o m a l proteins
, i = q i
R N A proteins
Hi a b c a c a c c
M o u s e
C y t o s o l i c D N A M i c r o s o m a I proteins
E - - 1 r 1 i - - 1
RNA proteins
Eli a b c c a c b c
Fig. 5. Comparison of in vitro binding of [14C]dichloroethane ( m ) and [t4C]dibromoethane (c~) to calf thymus DNA and to RNA and proteins ofmurine liver fractions mediated by microsomes (a), cytosol (b), and microsomes plus cytosol (c) under standard incubation procedure. The figure shows mean values (d.p.m./mg) of duplicate findings (net binding) differing from the mean by less than 3.0%. To obtain binding values as gmol haloethane/mol DNA P or RNA P, d.p.m./mg values have to be divided by 99.7, whereas for protein binding d.p.m./mg value divided by 32.4 gives the expression as pmol haloethane/mg protein
G. Arfellini et al.: Binding of 1,2-Dibromoethane and 1,2-Dichloroethane to Macromolecules in Rat and Mouse Organs 211
Poly (A)
5 0 0 1 ~ R a t - - - 1 r - M o u s e 7
/ lOO
l o
i
Poly (C)
F- M o u s e - - I
a b c a b c a b c b c
E P o l y ( U ) P o l y ( G )
"~ 5 0 0 i ~ R a t ~ i ~ M o u s e - t r ~ R a t -----1 f"-MGuse~
l o o
: I' i i i
4
a b c a b c a b c a b c
Fig. 6. In vitro comparative interaction of [14C]dichloroethane ( 1 ) and [~4C]dibromoethane ( ~ ) to polyribonucleotides performed with mu- fine liver enzymatic fractions in standard incubation procedure. Range of blank values is 66M36 d.p.m./mg, whereas the labeling of macro- molecular constituents of enzymatic fractions is omitted. For data expression and symbols see Fig. 5. To obtain binding values as pmoI/mg polynucleotide, the d.p.m./mg value has to be divided by 32.4
Chemicophysical Binding In Vitro
Both haloethanes are electrophilic: in fact, low nonen- zymatic interaction with DNA occurs in vitro, di- bromoethane being more reactive than dichlo- roethane, especially when tested under UV irradi- ation. This finding and that of Banerjee and Van Duuren (1979) could be related to the different ability of chemicals to exert a mutagenic effect in the absence of S 9 activation (Van Bladeren et al. 1981) and also a toxic action on human lymphocytes (Perocco and Prodi 1981). Unlike other carcinogens, such as poly-
cyclic hydrocarbons (Strniste et al. 1980; Prodi et al. 1984), dimethylnitrosamine (Arfellini et al. 1981), and aflatoxins (Shieh and Song 1980), dichloroethane and dibromoethane are not photoinduced to form interac- tion compounds with nucleic acids.
Activation by Microsomes and Cytosol and Binding to DNA
Both microsomal and cytosolic pathways can metabo- lize haloethanes to intermediate(s) capable of interact- ing in vitro both with preformed DNA or polynu-

212 G. Arfellini et al.: Binding of 1,2-Dibromoethane and 1,2-Dichloroethane to Macromolecules in Rat and Mouse Organs
cleotides and with the macromolecular constituents of subcellular fractions. This is in agreement with the data of Guengerich et al. (1980) for dichloroethane and of Su~dheimer et al. (1982) for dibromoethane, but not with those of Banerjee et al. (1980), who de- tected only the microsomal activation of di- chloroethane. PB pretreatment increases liver enzy- matic activity, as previously shown by Hill et al. (1978) in the case ofdibromoethane and by Banerjee and Van Duuren (1979) in the case of dichloroethane [data not confirmed by Guengerich et al. (1980)]. The relation- ship between extent of binding and tracer or acceptor concentration ( P < 0.05 at linear regression analysis) confirms that enzymatic activating pathways are in- volved in binding processes. Inhibition by GSH of microsome-mediated interaction of dichloroethane with DNA performed by mouse lung enzymes was shown to occur in vitro (Banerjee et al. 1980), and the same inhibitory effect is reported in the present paper, where rat liver microsomes are employed: in contrast, synergism was observed in the experience of Guengerich et al. (1980). At the same time, we found that d ibromoethane-DNA interaction mediated by rat liver microsomes is greatly enhanced by GSH. This could be due to the presence of a microsomal GSH transferase activity towards dibromoethane besides the cytosolic activity. It is true that GSH transferase activity not only exerts a protective role against attack from electrophiles, but can also bioactivate di- bromoethane to react with DNA. The influence of N A D P H on cytosol-mediated interaction resembles that of GSH on microsome-mediated binding dis- cussed above, since inhibition and enhancement were found in the case of dichloroethane and dibro- moethane interaction, respectively. The greater ef- fectiveness of the cytosolic pathway in carrying out di- b romoe thane-DNA interaction agrees with the marked increase of the mutagenicity of dibro- moethane on S. typhimurium in the presence of rat liver cytosol plus GSH (Van Bladeren et al. 1981). It should be underlined that the pattern of relative bind- ing of the two haloethanes to DNA obtained in vitro with liver microsomes plus cytosol (working amount deriving from about 300 mg wet tissue) mimics that of the in vivo situation.
Enzyme-Mediated Interaction with Polynucleotides
The only report available in literature (Guengerich et al. 1980) is restricted to dichloroethane and rat liver enzymes: it shows that all polynucleotides are labeled and that poly (U) and poly (G) are the best binding ac- ceptors for microsomal and cytosolic pathway, respec- tively. We do not confirm these findings, although the binding of dichloroethane to D N A mediated by microsomes is 63 pmol/mg in our experience as well as
in that of the authors just quoted (60-72 pmol/mg). The differences described above could be due to the different experimental design and/or to the animal strain utilized: in fact, S 9 fraction from liver of Wistar rats was more active in inducing mutagenicity by dichloroethane towards S. typhimurium (Rannug 1980) than was the same fraction extracted from liver of Sprague-Dawley rats.
In conclusion, a higher reactivity of dibro- moethane than of dichloroethane is systematically evi- denced in in vitro systems involving bioactivation. Such a relative reactivity of haloethanes has also been found in in vivo binding studies (see above), in meta- bolic studies (Livesey and Anders 1979), and in evalu- ation of both the D N A repair induction (Perocco and Prodi 1981) and the mutagenicity on microorganisms, on mammalian cells (Rannug 1980), and also on hu- man (EUE) cells (Ferreri et al. 1983). Moreover, the patterns of in vitro and in vivo binding are in suffi- cient agreement, the involvement of two metabolic pathways capable of differently bioactivating both haloethanes is confirmed, and a correlation among nucleic acid interaction, DNA damage, mutagenicity, and carcinogenicity by haloethanes seems to exist. Im- provement of our awareness of haloethane-nueleic acid interaction will require adducts isolation and characterization. Due to the lack of literature reports, our future research will be focused mainly on this sub- ject.
References
Arfellini G, Prodi G Rocchi P, Grilli S (1981) Photoinduced reaction of dimethylnitrosamine with DNA and polynucleotides. Photo- them Photobio134:645-648
Balzani V, Ballardini R, Sabbatini N, Moggi L (1968) Photochemi- cal reactions of nitropentamine cobalt (III) ion. Inorgan Chem 7:1398-1404
Banerjee S, Van Duuren BL (1979) Binding of carcinogenic halogen- ated hydrocarbons to cell macromolecules. J Natl Cancer Inst 63:707-711
Banerjee S, Van Duuren BL, Oruambo FI (1980) Microsome-medi- ated covalent binding of 1,2-dichloroethane to lung microsomal protein and salmon sperm DNA. Cancer Res 40:2170-2173
Bradford MM (1976) A rapid and sensitive method for the quantita- tion of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248-254
Burton K (1956) A study on the conditions and mechanism of the di- phenylamine reaction for the colorimetric estimation of deoxyribonucleic acid. Biochem J 62:315-323
Ferreri AM, Rocchi P, Capucci A, Prodi G (1983) Induction of diph- theria toxin-resistant mutants in human cells by halogenated compounds. J Cancer Res Clin Onco1105: I 11-112
Guengerich FP (1977) Separation and purification of multiple forms of microsomal cytochrome P-450: activities of different forms of cytochrome P-450 towards several compounds of environmental interest. J Biol Chem 252:3970-3979
Guengerich FP, Crawford WM Jr, Domoradzki JY, Macdonald TL, Watanabe PG (1980) In vitro activation of 1,2-dichloroethane by microsomal and cytosolic enzymes. Toxicol Appl Pharmacol 55:303-317

G. Arfellini et al.: Binding of 1,2-Dibromoethane and 1,2-Dichloroethane to Macromolecules in Rat and Mouse Organs 213
Hill DL, Shih T-W, Johnston TP, Struck RF (1978) Macromolecular binding and metabolism of the carcinogen 1,2-dibromoethane. Cancer Res 38:2438-2442
Kinoshita N, Gelboin HV (1972) Aryl hydrocarbon hydroxylase and polycyclic hydrocarbon tumorigenesis: effect of the enzyme in- hibitor 7,8-benzoflavone on tumorigenesis and macromolecular binding. Proc Natl Acad Sci USA 69:824 828
IARC (1977) Monographs on the Evaluation of the Carcinogenic Risk of Chemicals to Humans 15, Ethylene dibromide, pp. 195- 209
IARC (1979) Monographs on the Evaluation of the Carcinogenic Risk of Chemicals to Humans 20, 1,2-Dichloroethane, pp. 429- 448
IARC (1982) Monographs on the Evaluation of the Carcinogenic Risk of Chemicals to Humans [Suppl] 4, Ethylene dibromide, pp. 124-126
Livesey JC, Anders MW (1979) In vitro metabolism of 1,2-di- haloethanes to ethylene. Drug Metab Dispos 7:199-203
Lutz WK (1979) In vivo covalent binding of organic chemicals to DNA as a quantitative indicator in the process of chemical car- cinogenesis. Mutat Res 65:28%356
Nachtomi E, Sarma DSR (1977) Repair of rat liver DNA in vivo damaged by ethylene dibromide. Biochem Pharmacol 26:1941- 1945
Perocco P, Prodi G (1981) DNA damage by haloalkanes in human lymphocytes cultured in vitro. Cancer Lett 13:213-218
Prodi G, Grilli S, Mazzullo M, Colacci A, Arfellini G (1984) Com- parison between photo-induction and microsomal activation of polycyclic hydrocarbons with different oncogenic power. Toxi- col Pathol 12 (in press)
Rannug U (1980) Genotoxic effects of 1,2-dibromoethane and 1,2- dichloroethane. Mutat Res 76:269-295
Reitz RH, Fox TR, Domoradzki JY, Quast JF, Langvardt P, Watanabe PF (1980) Pharmacokinetics and macromolecular in- teraction of ethylene dichloride: comparison of oral and inhala- tion exposures. In: Ames B, Infante P, Reitz R (eds) Ethylene dichloride. Cold Spring Harbor Laboratory, New York, pp 135-148 (Banbury report no. 5)
Schneider WC (1945) Phosphorus compounds in animal tissues. I. Extraction and estimation of desoxypentose nucleic acid and of pentose nucleic acid. J Biol Chem 161:293-303
Schneider WC (1957) Determination of nucleic acids in tissues by pentose analysis. Methods Enzymol 3:680-684
Shieh J-C, Song P-S (1980) Photochemically induced binding of af- latoxins to DNA and its effects on template activity. Cancer Res 40:68%695
Strniste GF, Martinez E, Martinez AN, Brake RJ (1980) Photoindu- ced reactions of benzo(a)pyrene with DNA in vitro. Cancer Res 40:245-252
Sundheimer DV, White RD, Brendel K, Sipes IG (1982) The bioacti- vation of 1,2-dibromoethane in rat hepatocytes: covalent bind- ing to nucleic acids. Carcinogenesis 3:1129-1133
Swenson DH, Lawley PD (1978) Alkylation of deoxyribonucleic acid by carcinogens dimethyl sulphate, ethyl methanesulphon- ate, N-ethyl-N-nitrosourea and N-methyl-N-nitrosourea. Bio- chem J 171:575-587
Van Bladeren PJ, Breimer DD, Rotteveel-Smijs GMT, de Knijff P, Mohn GR, van Meeteren-W/ilchli B, Buijs W, van der Gen A (1981) The relation between the structure of vicinal dihalogen compounds and their mutagenic activation via conjugation to glutathione. Carcinogenesis 2:499-505
Weisburger EK (1977) Carcinogenicity studies on halogenated hy- drocarbons. Environ Health Perspect 21:7-16
Received June 15, 1983/Accepted February 21, 1984





























