Embed Size (px)
Citation preview

Intratumoral cytokines/chemokines/growth factors and tumorinfiltrating dendritic cells: friends or enemies?
Michael R. Shurin & Galina V. Shurin & Anna Lokshin &
Zoya R. Yurkovetsky & Dmitry W. Gutkin &
Gurkamal Chatta & Hua Zhong & Baohui Han &
Robert L. Ferris
Published online: 23 September 2006# Springer Science + Business Media, LLC 2006
Abstract The tumor microenvironment consists of avariable combination of tumor cells, stromal fibroblasts,endothelial cells and infiltrating leukocytes, such as macro-phages, T lymphocytes, and dendritic cells. A variety ofcytokines, chemokines and growth factors are produced inthe local tumor environment by different cells accountingfor a complex cell interaction and regulation of differenti-
ation, activation, function and survival of multiple celltypes. The interaction between cytokines, chemokines,growth factors and their receptors forms a comprehensivenetwork at the tumor site, which is primary responsible foroverall tumor progression and spreading or induction ofantitumor immune responses and tumor rejection. Althoughthe general thought is that dendritic cells are among the firstcells migrating to the tumor site and recognizing tumorcells for the induction of specific antitumor immunity, theclinical relevance of dendritic cells at the site of the tumorremains a matter of debate regarding their role in thegeneration of successful antitumor immune responses inhuman cancers. While several lines of evidence suggest thatintratumoral dendritic cells play an important role inantitumor immune responses, understanding the mecha-nisms of dendritic cell/tumor cell interaction and modula-tion of activity and function of different dendritic cellsubtypes at the tumor site is incomplete. This review islimited to discussing the role of intratumoral cytokinenetwork in the understanding immunobiology of tumor-associated dendritic cells, which seems to possess differentregulatory functions at the tumor site.
Keywords Intratumoral cytokines . Chemokines . Growthfactors . Dendritic cells . Immunosuppression
AbbreviationsAPC antigen-presenting cellsDC dendritic cell(s)EGF epidermal growth factorGM-CSF granulocyte-macrophage colony-stimulating
factorHGF hepatocyte growth factorHNSCC head and neck squamous-cell carcinomaICAM-1 intercellular adhesion molecule 1
Cancer Metastasis Rev (2006) 25:333–356DOI 10.1007/s10555-006-9010-6
M. R. Shurin :G. V. ShurinDepartment of Pathology, University of Pittsburgh Medical Centerand Cancer Institute,Pittsburgh, PA, USA
M. R. Shurin :R. L. FerrisDepartment of Immunology, University of Pittsburgh MedicalCenter and Cancer Institute,Pittsburgh, PA, USA
A. Lokshin : Z. R. Yurkovetsky :G. ChattaDepartment of Medicine, University of Pittsburgh Medical Centerand Cancer Institute,Pittsburgh, PA, USA
R. L. FerrisDepartment of Otolaryngology, University of Pittsburgh MedicalCenter and Cancer Institute,Pittsburgh, PA, USA
D. W. GutkinVA Pittsburgh Healthcare System,Pittsburgh, PA, USA
H. Zhong : B. HanShanghai Chest Hospital,Shanghai, China
M. R. Shurin (*)Clinical Immunopathology,5725 CHP-MT, 200 Lothrop St.,Pittsburgh, PA 15213, USAe-mail: [email protected]

IDO indoleamine-2,3-deoxygenaseIP-10 interferon-gamma-inducible protein 10;
CXCL10LC Langerhans cellsLCM Laser Capture MicrodissectionLN lymph node(s)LSC Laser Scanning CytometryMCP-1 monocyte chemotactic protein 1; CCL2M-CSF macrophage colony-stimulating factorMIP-3α macrophage inflammatory protein-3α, CCL20NK natural killerNSCLC non-small cell lung cancerPHA phytohemagglutininRANTES regulated upon activation, normal T cell
expressed and secreted chemokine; CCL5TA tumor antigen(s)TADC tumor-associated dendritic cellsTAM tumor-associated macrophagesTGF-β transforming growth factor-βTIL tumor-infiltrating lymphocytesTLR toll-like receptorTNF-α tumor necrosis factor-αSLN sentinel lymph node(s)VEGF vascular endothelial growth factor
1 Introduction: The tumor microenvironment
The tumor microenvironment consists of a variable combi-nation of tumor cells, stromal fibroblasts, endothelial cellsand infiltrating leukocytes, such as macrophages, Tlymphocytes, and dendritic cells (DC); in addition, eosino-
phils, granulocytes, natural-killer (NK) cells and B cells arealso found in some tumor types. A variety of cytokines,chemokines and growth factors are produced in the localtumor environment by different cells accounting for acomplex cell interaction and regulation of differentiation,activation, function and survival of multiple cell types. Theinteraction between cytokines, chemokines, growth factorsand their receptors forms a comprehensive network at thetumor site, which is primary responsible for overall tumorprogression and spreading or induction of antitumorimmune responses and tumor rejection (Fig. 1).
The recognition of an important role for inflammatorycells and their factors in the etiology and pathogenesis of atumor has a long history starting from the 19th centuryworks of Rudolf Ludwig Karl Virchow suggested that“lymphoreticular infiltrate” reflects the origin of cancer atsites of chronic inflammation (“Die Cellularpathologie inihrer Begründung auf physiologische und pathologischeGewebelehre”, 1858). However, the question as to whetherthe inflammatory infiltrate helps or hinders tumors is stillopen [1]. There are several excellent reviews focusing onthe clinical and experimental data implicating proinflam-matory cytokines and chemokines as important potentiatorsof carcinogenesis [1–4]. At the same time, there is agrowing body of evidence demonstrating the power ofimmunological protection from tumor growth, i.e., recog-nition and elimination of cells undergoing malignanttransformation by the immune system.
Several lines of evidence from recent years support theexistence of cancer immunosurveillance, especially studies ofNK cells, the IFN-γ pathway, and molecular regulation of Tand B cell differentiation. In addition, more recent data from
Endothelialcells
Neovascula-rization
TILTADC
Inhibition oftumor recognition &antitumor immunity
TAM (M2)
Promotion of tumorprogression & suppression
of antitumor immunity
Stroma &surrounding
tissues
Tumorspreading
Metastases
Tumor cellsTumor
progression
Pastimulation Autocrine
s
Intracrine
Tumor Endothelialcells
TILTADC
Inhibition oftumor recognition &antitumor immunity
TAM (M2)
o
mor i mu y
Stroma &surrounding
tissues
g
Metastases
Tumor cellsTumor
progression
Paracrine
stimulation
stimulation
Tumor
Fig. 1 Regulatory role oftumor-derived cytokines,chemokines, and growth factorsin the tumor microenvironment.TIL, tumor-infiltrating lympho-cytes; TADC, tumor-associateddendritic cells; TAM, tumor as-sociated macrophages
334 Cancer Metastasis Rev (2006) 25:333–356

several large epidemiological long-term follow-up studies ofcancer incidence in transplant patients on immunosuppressivetherapy have shown a three- to five-fold increase in theincidence of a broad range of cancers, including epithelialcancers such as colorectal, lung and prostate cancer, with anincidence curve that rises in a linear fashion with time [5]. Forinstance, a cohort of renal and cardiac allograft recipients inNorway was found to be at increased risk for SCC (65-foldincrease), malignant melanoma (3-fold) and Kaposi’s sarco-ma (84-fold) when compared with the general population [6].Successful growth of tumors is considered as having“escaped” from this immunosurveillance, generating thebasis for immunoediting or immunosculpturing theory [7,8]. These early surveillance mechanisms are viewed as“shaping” the tumor’s immunological phenotype [9]. Anoth-er model that explains why tumors grow and do not recognizedby the immune system is that tumors behave as immunolog-ically normal tissue. In this theory, tumor cells emergeimmunologically as normal growing cells that do not providedanger signals to activate the immune system because theyexpress neither microbial immune-recognition patterns norliberate stress signals to alarm the innate immune cells [10].
However, immune suppression is clearly observed incancer patients and tumor-bearing animals as well [11].Tumor cells have been reported to impair the function ofimmune system through various mechanisms, includinghiding from the immune system in immune sanctuaries,immune ignorance, production of immunomodulatory cyto-kines, inhibition of the function and activation of immunecells, protection against lytic activity of immune effectorsand inhibition of DC differentiation, function or survival (forreview, see [7, 12, 13]. Th1/Th2/Th3 cytokine disbalance iscommonly seen in serum in cancer patients, which play a
significant role in the insufficient induction of specific cell-mediated antitumor immunity (reviewed in [14–16]). Forinstance, in pleural effusion as well as in tumor tissue ofNSCLC patients, the mRNA expression of IL-4, IL-10,TGF-α, and TGF-β1 was significantly higher than that ofIL-2, IL-12, IL-18 and INF-γ [17]. This predominantexpression of immunosuppressive type II cytokines mirrorsan immunosuppressive state in the immunological microen-vironment. Another opportunity is that the immunologicaltumor microenvironment might contribute to tumor progres-sion by creating unique immunological pressure conditions,which could support selection of novel unrecognizable tumorvariants or even enhance spontaneous mutagenesis.
Thus, the relationship between the immune system andcancer is complex and cytokines, chemokines, and growthfactors in the tumor environment play a key role in thisinteraction (Fig. 2). In this review article, we address the roleof the tumor-derived cytokines, chemokines, and growthfactors, which are involved in attraction of dendritic cells(DC), regulation of DC activity and function, and modula-tion of DC survival in the local tumor microenvironment.Pro- and anti-tumorigenic role of inflammatory cell-derivedcytokines in the tumor microenvironment is discussed else-where [18–20] and in other reviews in this issue.
2 Intratumoral cytokines, chemokines, and growthfactors
Production of tumor-specific and non-specific biologicallyactive molecules by tumor cell lines and primary tumors hasbeen well documented and includes hormones, prostaglan-dins, bioamines, NO, lactic acid, neuropeptides, gangliosides,
Therapeutic C/Ch/Gf
Therapeutic C/Ch/Gfantagonists, inhibitorsor blocking antibodies
Tumor sensitivity to immunologic pressure
TUMOR GROWTH / TUMOR REGRESSION
Anti-tumorigenic factors:• M1-derived C/Ch/Gf• DC1-derived cytokines• Th1 cytokines
Pro-tumorigenic factors:• Tumor-derived C/Ch/Gf• M2-derived C/Ch/Gf• DC2-derived cytokines• Th2 cytokines
TUMOR MICROENVIRONMENT
Therapeutic C/Ch/Gf
Therapeutic C/Ch/Gfantagonists, inhibitorsor blocking antibodies
Tumor sensitivity to immunologic pressure
TUMOR GROWTH / TUMOR REGRESSION
Anti-tumorigenic factors:• M1-derived C/Ch/Gf• DC1-derived cytokines• Th1 cytokines
Pro-tumorigenic factors:• Tumor-derived C/Ch/Gf• M2-derived C/Ch/Gf• DC2-derived cytokines• Th2 cytokines
TUMOR MICROENVIRONMENTFig. 2 Role of intratumoralcytokines, chemokines, andgrowth factors in directing tu-mor progression versus antitu-mor immunity. C/Ch/Gf,cytokines, chemokines, growthfactors; DC, dendritic cells;M, macrophages
Cancer Metastasis Rev (2006) 25:333–356 335
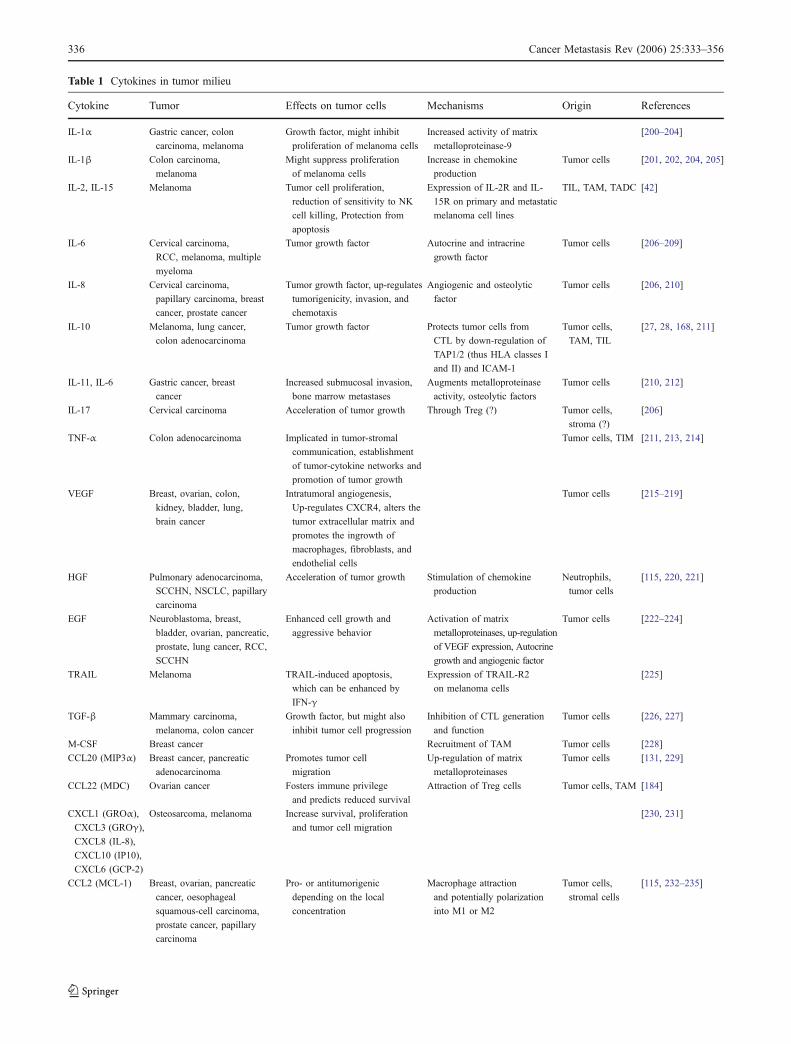
Table 1 Cytokines in tumor milieu
Cytokine Tumor Effects on tumor cells Mechanisms Origin References
IL-1α Gastric cancer, colon
carcinoma, melanoma
Growth factor, might inhibit
proliferation of melanoma cells
Increased activity of matrix
metalloproteinase-9
[200–204]
IL-1β Colon carcinoma,
melanoma
Might suppress proliferation
of melanoma cells
Increase in chemokine
production
Tumor cells [201, 202, 204, 205]
IL-2, IL-15 Melanoma Tumor cell proliferation,
reduction of sensitivity to NK
cell killing, Protection from
apoptosis
Expression of IL-2R and IL-
15R on primary and metastatic
melanoma cell lines
TIL, TAM, TADC [42]
IL-6 Cervical carcinoma,
RCC, melanoma, multiple
myeloma
Tumor growth factor Autocrine and intracrine
growth factor
Tumor cells [206–209]
IL-8 Cervical carcinoma,
papillary carcinoma, breast
cancer, prostate cancer
Tumor growth factor, up-regulates
tumorigenicity, invasion, and
chemotaxis
Angiogenic and osteolytic
factor
Tumor cells [206, 210]
IL-10 Melanoma, lung cancer,
colon adenocarcinoma
Tumor growth factor Protects tumor cells from
CTL by down-regulation of
TAP1/2 (thus HLA classes I
and II) and ICAM-1
Tumor cells,
TAM, TIL
[27, 28, 168, 211]
IL-11, IL-6 Gastric cancer, breast
cancer
Increased submucosal invasion,
bone marrow metastases
Augments metalloproteinase
activity, osteolytic factors
Tumor cells [210, 212]
IL-17 Cervical carcinoma Acceleration of tumor growth Through Treg (?) Tumor cells,
stroma (?)
[206]
TNF-α Colon adenocarcinoma Implicated in tumor-stromal
communication, establishment
of tumor-cytokine networks and
promotion of tumor growth
Tumor cells, TIM [211, 213, 214]
VEGF Breast, ovarian, colon,
kidney, bladder, lung,
brain cancer
Intratumoral angiogenesis,
Up-regulates CXCR4, alters the
tumor extracellular matrix and
promotes the ingrowth of
macrophages, fibroblasts, and
endothelial cells
Tumor cells [215–219]
HGF Pulmonary adenocarcinoma,
SCCHN, NSCLC, papillary
carcinoma
Acceleration of tumor growth Stimulation of chemokine
production
Neutrophils,
tumor cells
[115, 220, 221]
EGF Neuroblastoma, breast,
bladder, ovarian, pancreatic,
prostate, lung cancer, RCC,
SCCHN
Enhanced cell growth and
aggressive behavior
Activation of matrix
metalloproteinases, up-regulation
of VEGF expression, Autocrine
growth and angiogenic factor
Tumor cells [222–224]
TRAIL Melanoma TRAIL-induced apoptosis,
which can be enhanced by
IFN-γ
Expression of TRAIL-R2
on melanoma cells
[225]
TGF-β Mammary carcinoma,
melanoma, colon cancer
Growth factor, but might also
inhibit tumor cell progression
Inhibition of CTL generation
and function
Tumor cells [226, 227]
M-CSF Breast cancer Recruitment of TAM Tumor cells [228]
CCL20 (MIP3α) Breast cancer, pancreatic
adenocarcinoma
Promotes tumor cell
migration
Up-regulation of matrix
metalloproteinases
Tumor cells [131, 229]
CCL22 (MDC) Ovarian cancer Fosters immune privilege
and predicts reduced survival
Attraction of Treg cells Tumor cells, TAM [184]
CXCL1 (GROα),
CXCL3 (GROγ),
CXCL8 (IL-8),
CXCL10 (IP10),
CXCL6 (GCP-2)
Osteosarcoma, melanoma Increase survival, proliferation
and tumor cell migration
[230, 231]
CCL2 (MCL-1) Breast, ovarian, pancreatic
cancer, oesophageal
squamous-cell carcinoma,
prostate cancer, papillary
carcinoma
Pro- or antitumorigenic
depending on the local
concentration
Macrophage attraction
and potentially polarization
into M1 or M2
Tumor cells,
stromal cells
[115, 232–235]
336 Cancer Metastasis Rev (2006) 25:333–356

as well as cytokines, chemokines, and growth factors.Functional significance of some of tumor-derived factors fortumor progression or immunosuppression has been deter-mined, while the biological function of many other substancesin the tumor environment remains to be described. Tumorcells synthesize and release a variety of cytokines forming acomprehensive network at the tumor site and regulatingdifferentiation, function, and survival of both tumor cells andall stromal elements.
The list of tumor-derived cytokines/chemokines/growthfactors is constantly growing and includes, but not limited to,IL-1, IL-2, IL-4, IL-6, IL-8, IL-10, IL-15, VEGF, TFG-β,TNF-α, EGF, FGF, HGF, MIP (Table 1). A number ofexcellent reviews focus on the role and significance of theseand other biologically active factors for tumor growth andprogression [21–26], and only a few examples are givenhere.
IL-10 and TGF-β seem to be the best characterizedtumor-derived cytokines with well-defined immunosuppres-sive function. IL-10 is produced by many human and murinetumor cells and involved in regulating tumor cell prolifera-tion, protection from immune recognition and immunosup-pression. For instance, IL-10 may protect tumor cells fromCTL by down-regulation of HLA classes I and II andICAM-1 expression [27]. The loss of HLA class I expres-sion could be due to IL-10-mediated down-regulation ofTAP1 and TAP2 proteins in tumor cells [28] It has beenshown that the microenvironment of Burkitt’s lymphoma isrich in the IL-10, which can be produced by both tumorcells and macrophages, and that IL-10-activated humanmacrophages have enhanced capacity to engulf apoptoticcells, an anti-inflammatory process known to suppressimmune responses [29]. Furthermore, IL-10-activated mac-rophages, as well as TAM, were found to produce markedlyhigher levels of the B cell survival factor, B cell-activatingfactor of the TNF family/B lymphocyte stimulator (BAFF/BLyS) and thus promote Burkitt’s lymphoma survival andpathogenesis. In addition to B-cell lymphoma, IL-10 hasbeen shown to be a growth factor for human melanoma celllines [27]. Furthermore, autocrine production of IL-4 andIL-10 has been reported to promote thyroid tumor cellprogression and resistance to chemotherapy through the up-
regulation of anti-apoptotic proteins [30]. Finally, it has alsobeen widely speculated that IL-10 could favor the devel-opment of tumors through immunosuppressive mecha-nisms, including the modulation of APC and especiallyDC functions, and the development of Treg cell activity[31].
High concentrations of TGF-β are also frequently foundin cancer patients and are associated with disease progres-sion [32] and poor responses to immunotherapy [33]. Thelevels of TGF-β are higher in patients with disseminatedmelanoma when compared to those with localized disease[34]. In addition to being produced by some tumor celllines, TGF-β may also be released by cells dying apoptoti-cally [35]. TGF-β plays an important role in regulatingactivation, proliferation and activity of T lymphocytes andDC in vivo in the tumor environment [36, 37].
Most tumors have a complex chemokine network, whichcan direct immune responses to the tumor, regulate the leveland cellular composition of the leukocyte infiltrate and playa role in angiogenesis, as well. Tumor cells can also hijackthe chemokine system and gain expression of chemokinereceptors thus responding to specific chemokine gradients[38, 39]. Chemokine receptors, both CC and CXC havebeen detected on tumor cells and the correspondingchemokines are also expressed at the tumor site and atsites of tumor spread, suggesting a role for the chemokinefamily in malignant growth and metastasis [38, 40]. Inaddition, chemokines contribute to TH2 polarization inmany different tumors, and this strategy might help thetumor to subvert the immune system by establishing amicroenvironment of immune cells and cytokines thatsuppress any specific antitumor responses [3]. Secretionof CCL by tumor cell themselves [39] (Ferris et al.,submitted) may maintain an altered phenotype and impairappropriate migration of intratumoral, antigen-bearing DCto draining lymph nodes. Although it seems that chemokineproduction in cancers has a direct relationship with thenature and extent of the leukocyte infiltrate, there is noclear picture of the overall chemokine repertoire of anindividual human cancer type [3]. Many more additionalstudies are required to reveal multivariable role of tumor-derived chemokines in regulating attraction, activation, and
CCL5 (RANTES),
CCL25 (TECK)
NSCLC, prostate cancer Positive predictor of
survival
Up-regulate proliferation
and invasiveness
Tumor cells [236, 237, 238]
CXCL12 (SDF-1) Glioma, astrocytoma, melanoma,
SCLC, ovarian, prostate,
pancreatic, renal and thyroid
cancers, follicular lymphoma
Stimulates the proliferation
and/or survival of CXCR4-
expressing cancer cells
Attraction of Treg cells,
Up-regulation of matrix
metalloproteinases
Tumor cells,
stromal cells
[239, 240, 241,
242, 243]
Table 1 (continued)
Cytokine Tumor Effects on tumor cells Mechanisms Origin ReferencesCytokine Tumor Effects on tumor cells Mechanisms Origin References
Cancer Metastasis Rev (2006) 25:333–356 337

survival of different types of immune cells in the localtumor microenvironment.
The effect of cytokines and chemokines on cells in thetumor microenvironment depends not only on a specificcytokine or chemokine concentration, but also on a numberof other factors. For instance, IL-6 might both support orinhibit tumor cell migration depending on the extracellularmatrix [41], suggesting that different degrees of invasive-ness at the tumor–host interface are not necessarily relatedto specific genetic alterations but might be related todifferent environmental conditions as well. IL-2 and IL-15receptors, which are normally mediate activation of NKcells and T lymphocytes, have been detected on somemurine and human neoplasms. For instance, analysis ofdifferent human uveal melanoma cell lines and cell linesfrom uveal melanoma metastases cultured, with or withoutrecombinant human IL-2 or IL-15 revealed that bothcytokines induced proliferation of tumor cell lines andreduced melanoma cell sensitivity to NK-cell-mediatedcytolysis and cisplatin-induced apoptosis [42]. Thus, theseresults suggest that IL-2 and IL-15 elaborated by tumor-infiltrating lymphocytes, macrophages and DC may affectthe malignant behavior of melanoma by stimulating pro-liferation and reducing melanoma cell susceptibility to NK-cell-mediated cytolysis and induction of apoptosis.
Tumor-associated macrophages (TAM), lymphocytes,fibroblasts, and endothelial cells are also involved in gen-erating specific cytokine network in the tumor microenvi-ronment. For instance, TAM play important roles insupporting malignant outgrowth by producing factors thatpromote angiogenesis, tumor cell survival, and proliferationas well as suppressing innate and adaptive antitumorimmune responses [43–45]. In addition to being immuno-suppressive, infiltrating leukocytes might contribute totumor progression by producing matrix metalloproteinases(MMP) as well as growth and angiogenic factors. Forexample, stimulation of macrophage/tumor cell cocultureswith endothelin-2 led to increased matrix metalloproteinase(MMP)-2 and -9 production by macrophages and a markedincrease in invasion of tumor cells [46].
Tumors can also be infiltrated by eosinophils. Eosinophilcell infiltrates, despite being infrequent, have been indicatedas a marker of better prognosis [47]. In esophageal squamouscell carcinoma, eosinophil cell infiltrates have been inverselyrelated to metastatic potential by unknown mechanisms [48].However, as IL-5-transfected tumors induce the infiltrationof large amounts of eosinophils in vivo without anytherapeutic effect [49] the exact role of these cells and theirsoluble products need still to be determined.
Thus, the primary function of tumor-derived cytokines,chemokines, and growth factors is to aid tumor cellsproliferation, survival, spreading and progression (Figs. 1and 2). Although increasing evidence suggests that altered
immune function accompanies, and indeed may facilitate,cancer progression, one point to consider is that tumorsmay not necessarily produce immunosuppressive cytokinesas escape mechanisms: the immunosuppressive functionsmay be mere side-effects of the angiogenic and growthfactor functions of these cytokines [9]. Finally, a network ofinflammatory cytokines and chemokines in cancer contrib-ute not only to tumor cell growth and survival, but also tocommunication between the tumor cells and stromalelements [50]. Good example is tumor cell production ofthe TNF-α, a critical factor in this autocrine and paracrinenetwork. TNF-α may also initiate and sustain production ofother cytokines and chemokines. Understanding the com-plexity of cytokine/chemokines/growth factor network inthe tumor environment in the recent years reinforced theinterest of tumor immunologist to the tumor cell–immunecell interactions inside of progressing tumors and stimulat-ed the development of improved and novel strategies formeasuring biologically active molecules in the local tumormicroenvironment.
3 Detection of intratumoral cytokines, chemokines,and growth factors
Evaluation of cytokine, chemokines, and growth factorexpression at the tumor site in vivo is based on severalmethodological approaches. Historically, immunohisto-chemical strategies were the most common and widelyused [51–53] in spite of a very limited quantitativeapplicability. For example, using this approach, loss ofexpression of CXCL14 in HNSCC has been recently re-ported [54]. Together with in situ PCR on tumor specimens,immunohistochemistry allow evaluation of cytokine distri-bution in tumor mass and co-localization with specific cellsubtypes. Gene expression profiling is another popularapproach to determine potential alterations of cytokines/cytokine receptors on tumor cells, stromal elements andinfiltrating leukocytes [55–57].
Isolation of tumor cells from fresh tumor implants byenzymatic digestion provides a possibility for a quantitativedetermination of cytokine production by ELISA or flowcytometry. An alternative approach for analyzing a verylimited amount of isolated tumor cells is Laser ScanningCytometry (LSC). Although LSC technology has a flowcytometry heritage, it is not limited to analyzing cells influids. Instead, the technology allows automated analysis ofsolid-phase samples, including adherent cultured or cyto-span cells, tissue sections, and tissue imprints. Cellsscanned with lasers are also available for visual inspection,unlike a flow cytometer. Significantly less sampling isrequired than for flow cytometry and slides may be readrepeatedly. Because the cells are adhered to the slide, the
338 Cancer Metastasis Rev (2006) 25:333–356

LSC is able to record the position for each cell analyzed forpossible re-staining and re-analysis.
Small amounts of tumor cells might be also extracted fromparaffin-embedded tumor slides by Laser Capture Microdis-section (LCM). LCM provides a mean to isolate purepopulation of cells from heterogeneous tissue specimens.These “captured” cells can then be used in a wide range ofdownstream assays such as gene expression analysis at themRNA level or in a range of proteomic assays.
Several approaches have been used to determine cytokinemilieu in LN and SLN. Direct measurement of cytokine levelsis striking but difficult due to low levels of cytokines in smallLN fragments. For instance Leong et al. attempted directassess of cytokines from SLN of melanoma patients by usingELISPOT assay but were unsuccessful except when LN-derived lymphocytes were stimulated with enterotoxin A [58].Unfortunately, this type of assay does not correctly reflect invivo cytokine production. Lee et al. used quantitative RT-PCR to investigate cytokine profiles in the SLN and non-SLN [59]. To ensure in vivo fidelity, they used freshlypreserved tissue taken uniformly from the tissue collected forroutine pathologic analysis. In addition, RNA from eachsample was assessed for purity, quality, and integrity. Theauthors used commercially available forward/reverse primersets and probes for IL-2, IL-10, IL-12, and IFN-γ. ThemRNA segments were amplified on a panel of cDNAderived from PHA-stimulated lymphocytes by using RT-PCR. RNA from the LN samples was reverse-transcribedinto cDNA and then evaluated by quantitative RT-PCR usingthe ABI Prism 7000 sequence detection system and theTaqman assay. The results of this study indicate that if re-sidual melanoma was present at the time of sentinel lym-phadenectomy, the SLN have significantly different cytokinemicroenvironment when compared with neighboring non-SLN: expression levels of IL-10 and IFN-γ genes weresignificantly higher in the SLN when residual melanoma waspresent, suggesting that melanoma interacts with the SLN toincrease expression of these genes. However, SLN expres-sion of IL-2 and IL-12 genes was not changed by thepresence of residual melanoma, suggesting that SLN have arather specific cytokine response to neighboring melanoma[59].
Kusuda et al. (2005) have examined mRNA expressionlevels of Th1 (TNF-α, IFN-γ, and IL-12p40) and Th2 (IL-6and IL-10) cytokines for any association with clinicopatholog-ical characteristics of epithelial ovarian cancer using real-timePCR technique and 40 samples of epithelial ovarian cancers[60]. TNF-α and IFN-γ expression levels were significantlyhigher in serous adenocarcinoma than in non-serous adeno-carcinoma and high TNF-α mRNA expression significantlycorrelated with longer survival in advanced stage (II/III/IV)ovarian carcinomas. In examining all combinations of Th1/Th2 expression values, the most significant association was
between high IFN-γ/IL-12p40/IL-6 expression levels andbetter prognosis in advanced stage ovarian carcinomas.
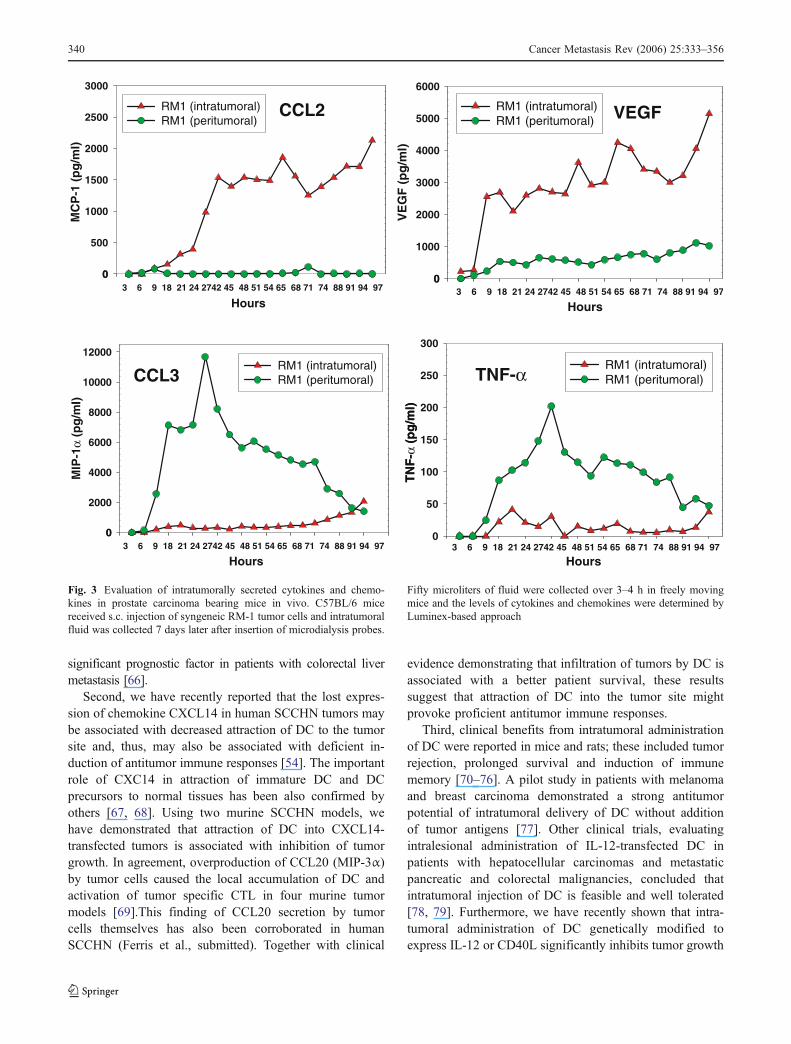
We have recently developed a novel approach that allowsevaluating secretion of intratumoral cytokines in dynamic forseveral days in live freely moving mice. The method utilizesintratumoral insertion of a specially designed microdialysisprobe connected with a micropump and a sample collectiondevise. The whole system is maintained on several uniquebrackets allowing the animal movement in a special cage. Allsoluble components of extracellular fluid from the tumor massof less than 100 kDa size are collected at a speed of 8 μl/h for4 h and analyzed by Multiplex Luminex-based techniques forthe presence of more than 30 cytokines, chemokines andgrowth factors in each sample. Figure 3 represents an ex-ample of multiple cytokine analysis in the murine prostatecancer tumors and adjunct normal tissues using describedabove technique. As shown, different cytokines demonstratedifferent patterns of production, which may reflect tumorprogression or development of antitumor immune responses.Comparative analysis of several tumors at different stages oftumor progression is in progress in our laboratory.
4 Tumor-associated dendritic cell
Little is known about the role of DC in the natural im-munosurveillance of cancer, although the general thought isthat DC are among the first cells migrating to the tumor siteand recognizing tumor cells for the induction of specificantitumor ammunity. The clinical relevance of DC at the siteof the tumor remains, thus, a matter of debate regarding theirrole in the generation of successful anitumor immune responsein human cancers. At the time the significance of peritumoralinfiltration of activated DC in antitumor activity and clinicalprognosis has been shown in various careers [61].
However, several lines of evidence suggest that tumor-associated DC (TADC) or tumor-infiltrating DC (TIDC) playan important role in antitumor immune responses. First,increased numbers of tumor-infiltrating DC are associated withbetter outcome in cancer patients with a variety of tumors [13,62]. For instance, evaluating the prognostic significance ofDC in 132 specimens from patients with primary SCCHN,Reichert et al. reported that a low number of tumor-infiltratingDC was more predictive of poor survival than lymph node(LN) involvement [63]. Goldman et al. concluded that sur-vival and recurrence rates for patients with SCC of the tonguecorrelate with the degree of DC infiltration of the primarytumor [64]. Analyzing the distribution of DC in the primarytumor, adjacent tissue, and regional LN, Kikuchi et al.revealed that the number of DC in the tumor predicts overallsurvival, disease-free survival, and time to disease recurrencein patients with SCCHN [65]. Similarly, the number ofCD83 + DC in the cancer invasive margin was shown to be a
Cancer Metastasis Rev (2006) 25:333–356 339
significant prognostic factor in patients with colorectal livermetastasis [66].
Second, we have recently reported that the lost expres-sion of chemokine CXCL14 in human SCCHN tumors maybe associated with decreased attraction of DC to the tumorsite and, thus, may also be associated with deficient in-duction of antitumor immune responses [54]. The importantrole of CXC14 in attraction of immature DC and DCprecursors to normal tissues has been also confirmed byothers [67, 68]. Using two murine SCCHN models, wehave demonstrated that attraction of DC into CXCL14-transfected tumors is associated with inhibition of tumorgrowth. In agreement, overproduction of CCL20 (MIP-3α)by tumor cells caused the local accumulation of DC andactivation of tumor specific CTL in four murine tumormodels [69].This finding of CCL20 secretion by tumorcells themselves has also been corroborated in humanSCCHN (Ferris et al., submitted). Together with clinical
evidence demonstrating that infiltration of tumors by DC isassociated with a better patient survival, these resultssuggest that attraction of DC into the tumor site mightprovoke proficient antitumor immune responses.
Third, clinical benefits from intratumoral administrationof DC were reported in mice and rats; these included tumorrejection, prolonged survival and induction of immunememory [70–76]. A pilot study in patients with melanomaand breast carcinoma demonstrated a strong antitumorpotential of intratumoral delivery of DC without additionof tumor antigens [77]. Other clinical trials, evaluatingintralesional administration of IL-12-transfected DC inpatients with hepatocellular carcinomas and metastaticpancreatic and colorectal malignancies, concluded thatintratumoral injection of DC is feasible and well tolerated[78, 79]. Furthermore, we have recently shown that intra-tumoral administration of DC genetically modified toexpress IL-12 or CD40L significantly inhibits tumor growth
Hours
MC
P-1
(p
g/m
l)
0
500
1000
1500
2000
2500
3000
RM1 (intratumoral)RM1 (peritumoral)
3 6 9 18 21 24 2742 45 48 51 54 65 68 71 74 88 91 94 97
CCL2
Hours
VE
GF
(p
g/m
l)
0
1000
2000
3000
4000
5000
6000
RM1 (intratumoral)RM1 (peritumoral)
3 6 9 18 21 24 2742 45 48 51 54 65 68 71 74 88 91 94 97
VEGF
Hours
MIP
-1
(p
g/m
l)
0
2000
4000
6000
8000
10000
12000RM1 (intratumoral)RM1 (peritumoral)
3 6 9 18 21 24 2742 45 48 51 54 65 68 71 74 88 91 94 97
CCL3
Hours
TN
F-
(p
g/m
l)
0
50
100
150
200
250
300
RM1 (intratumoral)RM1 (peritumoral)
3 6 9 18 21 24 2742 45 48 51 54 65 68 71 74 88 91 94 97
TNF-
0
RM1 (intratumoral)RM1 (peritumoral)
CCL2
0
RM1 (intratumoral)RM1 (peritumoral) VEGF
α
l
0
RM1 (intratumoral)RM1 (peritumoral)CCL3
TN
F- α
(pg
/ml)
RM1 (intratumoral)RM1 (peritumoral)TNF- α
Fig. 3 Evaluation of intratumorally secreted cytokines and chemo-kines in prostate carcinoma bearing mice in vivo. C57BL/6 micereceived s.c. injection of syngeneic RM-1 tumor cells and intratumoralfluid was collected 7 days later after insertion of microdialysis probes.
Fifty microliters of fluid were collected over 3–4 h in freely movingmice and the levels of cytokines and chemokines were determined byLuminex-based approach
340 Cancer Metastasis Rev (2006) 25:333–356

or causes complete tumor rejection in murine prostate,colon and breast carcinoma models [74, 75]. Evidence hasbeen obtained showing that intratumoral DC can captureand process tumor antigens to be presented to T lympho-cytes [80]. Interestingly, DC were recently found to becytotoxic for several tumor cell lines suggesting that thismay have important consequences for their ability tostimulate tumor-specific CTL [81–83].
A recent study albeit performed in a limited series ofpatients, also addressed the clinical relevance of DC in-filtration on regressing tumor lesions and SLN in melanomapatients and pointed to a pivotal role of mature DC in thelocal expansion of efficient antitumor T-cell-mediated im-mune responses at the initial sites of metastasis [61].Analyzing metastatic melanoma patients’ specimens, theauthors reported that the accumulation of DC-Lamp+ DC,clustered with tumor cells and lymphocytes, was associatedwith local expansion of antigen-specific memory effectorCTL. They also demonstrated a significant correlationbetween the density of DC-Lamp+ DC infiltrates in SLNwith the absence of metastasis in downstream LN [61].
Thus, the infiltration of tumors by DC is of great im-portance in initiating the primary antitumor immuneresponse and was confirmed as an independent, immuno-logic prognostic parameter for survival in patients withcancer. While several lines of evidence suggest that intra-tumoral DC play an important role in antitumor immuneresponses, understanding the mechanisms of DC/tumor cellinteraction is limited. This review is limited to discussingthe role of intratumoral cytokine network in the under-standing immunobiology of TADC, which seems to possessdifferent regulatory functions at the tumor site. Although itis possible that professional ‘regulatory’ DC exist, there isanother view that a regulatory function is not the intrinsicnature of a distinct DC subset and that DC recruited to thetumor site undergo changes that endow them with regula-tory functions favorable for the tumor [12].
5 DC in metastatic lymph nodes
There is a general thought that many primary tumors invadethe local lymphatic system before spreading to distant sitesbecause most patients have LN metastases prior to develop-ment of distant disease. It has been suggested that immuno-suppression of nodes nearest to tumor may facilitate the earlyestablishment of metastases. Recent evidence suggests thatregional LN, especially the SLN, are immunosuppressedwhen compared with neighboring non-SLN in patients withprimary tumors [84, 85]. For instance, lymphocytes fromLN closest to the primary melanoma show diminishedproliferation in response to mitogens and IL-2. These LNalso generate migration-inhibitory chemokines and contain
increased numbers of suppressor cells. APC function of DCwas also significantly decreased in these LN [85, 86].Examining distribution of S100+ DC in the tumor tissuesand regional LN of 60 patients with SCCHN, Deng et al.reported that the DC density in tumor tissues was correlatedwith tumor histological grade and the density of DC wassignificantly higher in reginal lymph nodes without tumorthan those with metastases [87]. Analyzing infiltration ofDC into SLN in oral cavity cancer patients, Sakakura et al.concluded that more significant immune responses againstcancer occur in SLN than in non-SLN [88]. Additionalevidence has been provided that SLN immunosuppressionis induced by melanoma and is mediated by local cyto-kines. The resulting SLN cytokine milieu favors immu-nosuppressive DC and SLN immune dysfunction [59].These changes are evident even in the absence of SLNmetastasis, suggesting that primary melanoma interactswith the SLN to create an immunosuppressive microenvi-ronment that favors tumor metastasis and growth.
Two main DC types in human LN are myeloid andplasmacytoid. Myeloid DC are found in most peripheraltissues and, following antigen exposure, will mature andmigrate to LN and secrete high levels of IL-12. PlasmacytoidDC can be identified by positive surface staining for HLA-DR, bright expression of CD123 (IL-3 receptor α-chain), andlack of lineage-specific markers (CD3, CD19, CD56, CD11c,CD13, and CD33). Plasmacytoid DC are found around highendothelial venules in the T-cell areas of LN [89] and may beinvolved in the induction of tolerance by polarizing towardthe production of Th2 cytokines, such as IL-4 and IL-10.Plasmacytoid DC have been localized to the peritumoralareas of melanomas [90, 91] and may be functionallyimpaired in the tumor tissue and draining LN of patientswith head and neck squamous cell carcinoma [92]. Further-more, using four-color flow cytometry Cox et al. has recentlyevaluated myeloid DC and plasmacytoid DC subpopulationsand intra-cellular cytokine expression of IL-4, IL-10, IL-12,and IFN-γ in these DC subsets in tumor-draining LN ofpatients with breast cancer [93]. Although both myeloid DCand plasmacytoid DC could be easily identified in LN,cytokine expression was primarily found in the plasmacytoidDC subset. In LN draining tumors categorized as poorprognosis, the authors also report a relative increase in IL-10and IL-4 expression by plasmacytoid DC [93]. Thus, theseresults suggest that plasmacytoid DC are major producers ofDC cytokines in human LN and may influence local tumorimmunity in patients with breast cancer.
The recent discovery of indoleamine 2,3-dioxygenase(IDO)-expressing DC in the SLN of melanoma patients pro-vides insight into the relationship between SLN immunosup-pression and local cytokine microenvironment. A subset ofCD123+CCR6+ DC expressing IDO [94], which is a rate-limiting enzyme in tryptophan catabolism [95], has shown to
Cancer Metastasis Rev (2006) 25:333–356 341

be immunosuppressive, as proven by their inhibition of T-cell proliferation in vitro [96] and diminution of T-cell re-sponse in vivo [97]. Importantly, induction of immunosup-pressive IDO-expressing DC can be regulated by IL-10 [94]and IDO can also be induced by stimulation of immatureCD123+CCR6+ DC with IFN-γ, resulting in subsequentreduction in T-cell proliferation [94]. Whereas stimulation ofmature DC with IFN-γ causes inhibition of IDO activity,addition of IFN-γ to DC matured in the presence of IL-10results in enhancement of IDO expression. In other words,the presence of IL-10 can turn a proinflammatory signal ofIFN-γ into an immunosuppressive signal [94].
The relevance of IDO in local down-regulation of im-mune function in patients with early-stage melanoma hasbeen suggested by the presence of IDO-expressing DC inmelanoma SLN [94]. Thus, it is likely that primary tumormight interact with the locoregional immune system,especially the SLN, inducing local immunosuppressionand thereby creating a microenvironment conducive forlymphatic metastasis and tumor growth. Alteration of DCmaturation and function by LN-derived and tumor-derivedcytokines in SLN might play a key role in directing thedevelopment of antitumor immunity or its failure.
6 Regulation of intratumoral dendritic cellsby cytokines, chemokines, and growth factors
One reason for the limited efficacy of immunotherapy incancer patients could be due to local or systemic suppres-sion of the DC system, resulting in inhibition of antitumorimmune responses. In evaluating the immunological statusof cancer patients, moderate immunosuppression was noted
in the presence of relatively confined tumors, and signifi-cant impairment of the immune system was found in theadvanced stages of cancer [98–103]. Although the exactnature of immunodeficiency in these patients is unclear, andthe potential involvement of the DC system as a basis forthe immunosuppression is controversial, numerous evi-dence has been presented recently suggesting that DC aredeficient and not fully functional in the tumor microenvi-ronment (Table 2).
Owning to DC having vital roles in the generation of T-cell-mediated immune response and in the maintenance ofcentral and peripheral tolerance, it is more likely thataltered immunity during tumor progression reflects changesin the phenotype and/or function of DC.
Examination of circulating and tumor-infiltrating DC intumor-bearing animals and in cancer patients has revealed thatDC are functionally impaired in their ability to induce effectorT-cell responses [104–109]. For instance, the percentage ofmyeloid CD11c+, but not plasmacytoid CD123+ DC, wassignificantly lower and expression of HLA-DR was signif-icantly decreased in total and myeloid DC in circulation ofSCCHN patients compared to healthy donors [110]. Simi-larly, the percentage of both DC populations in peripheralblood was significantly lower in patients with NSCLC thanin the control group, while in patients with breast cancer, thenumber of ‘lymphoid’ DC was significantly higher than inNSCLC patients [111]. In a cohort of 54 patients affected bycolorectal cancer, investigation of the number of peripheralblood DC type 1 (DC1, myeloid DC, conventional DC) andtype 2 (DC2, lymphoid DC, plasmacytoid DC) revealed thatin comparison to healthy controls, patients presented reducedDC1 and DC2 numbers [112]. Moreover, in cancer patients,DC showed low levels of HLA DR, CD11c, CD83, CD86,
Table 2 Tumor-related cytokines, chemokines, and growth factors, which are able to modulate DC maturation, function and/or longevity
Cytokine Tumor Effect on DC References
IL-1 GI cancers, melanoma DC maturation and mobilization from inflamed tissue to draining LN [244, 245]IL-6 Cervical carcinoma, RCC,
melanoma, lung cancerInhibition of DC differentiation and maturation [146, 163,
246, 247]IL-8 Hepatocellular carcinoma,
colorectal and pancreaticcancer, cervical carcinoma
Retention of DCs inside human malignant lesions [78]
IL-10 Melanoma, colon adenocarcinoma,prostate cancer, lung cancer, SCC
Induction of tolerogenic DC, inhibition of DC maturation andlongevity, suppression of IL-12 production, and antigen presentation,modulation of CCR expression, up-regulation of B7-H1 expression on DC
[105, 133, 168,175, 248,249, 250, 251]
VEGF Lung, brain, colon, stomach,breast, prostate cancers
Inhibition of DC differentiation and maturation, up-regulation ofB7-H1 expression on DC
[175, 252, 253]
HGF SCCHN, NSCLC Suppression of the Ag-presenting capacity of DC [254]M-CSF RCC, breast cancer Inhibition of DC differentiation and maturation, generation of
tolerogenic DC[163, 228, 255]
CXCL12(SDF-1)
Ovarian cancer Attraction of plasmacytoid DC, which induce regulatory CD8+ T cellsand induce angiogenesis, CXCL12 protects tumor plasmacytoid DCfrom apoptosis
[184, 199]
342 Cancer Metastasis Rev (2006) 25:333–356

and Mannose receptor, an up-regulation of CXCR4 and areduced T-cell stimulation capability [112]. Analyzing DCsubsets in children with cancer Vakkila et al. (2004) haverecently reported that patients with cancer have significantlyless circulating monocytoid DC than controls, whereasabsolute plasmacytoid DC counts did not differ significantlybetween the study groups [113]. However, clinical outcomeof the children with cancer correlated with plasmacytoid DCcount: children with high plasmacytoid DC counts atdiagnosis survived significantly worse than those with lowcounts. Thus, circulating DC subsets are changed in cancerdevelopment and might served as prognostic markers forcancer patients.
Interestingly, Bell et al. demonstrated two levels ofheterogeneity of DC infiltrating breast carcinoma tissue:immature CD1a+ DC were retained within the tumor bed,while mature CD83+DC-Lamp+ DC were visible in peritu-moral areas [114]. Unfortunately, the authors presented noclinical correlates in their study. Similar analysis of tissuedistribution of DC in papillary carcinoma of the thyroid alsorevealed that most DC had an immature phenotype (CD1a++
CD11c+CD40+CD86HLA-DR) and were located at the
invasion edge of the tumor [115]. Recently, in vitro studieshave shown that repetitive stimulation of naïve T cells withimmature DC results in the generation of regulatory T-cellactivity [116]. Other studies suggest that DC-mediatedperipheral tolerance may involve the induction and/orexpansion of regulatory T cells [117]. These observationslead to the hypothesis that tumor cells may progressivelyalter the phenotype and/or function of DC, thus facilitatingthe accumulation of regulatory T cells. These regulatory Tcells play a critical role in suppressing immune responsive-ness and thus allow tumor growth to continue. In fact, Tienet al. have recently reported that prostate tumor cells alter thephenotype and frequency of DC and CD4+CD25+ regulatoryT cells in the tumor microenvironment in vivo and alsoexpand in the LN draining the prostate [118].
Since DC perform a pivotal role in the initiation andregulation of both innate and adaptive immunity, includingantitumor responses, it is, therefore, not surprising that manytumors may have evolved mechanisms that adversely impactDC function (Table 2). To evade host immunity, tumors usenumerous strategies to hinder normal DC differentiation,trafficking, activity, and longevity (Fig. 4).
Maturation of
conventional DC
Function of
conventional DC
Attraction of
conventional DC
(lost of CXCL14)
IL-12, IFN- , IP-10MotilityEndocytosisAPM componentexpressionAntigen processingAntigen presentationB7-1/2
Longevity of
conventional DC
Attraction of
plasmocytoid or
regulatory DC
Differentiation
of endothelial-
like DC
Neovascula-rization
IDOB7-H1/H4IL-10
T U
M O
R
TNF- IL-8
CD4+ TregCD8+ Treg
TA-specific T cells
TU
MO
R P
RO
GR
ESS
ION
Tumor-derived
Cytokines/Chemokines/Growth factors
Tumor-derived
Cytokines/Chemokines/Growth factors
D E
N D
R I
T I
C
C E
L L
S
Maturation of
conventional DC
Function of
conventional DC
Attraction of
conventional DC
(lost of CXCL14)
↓
↓
↓
↓
↓↓↓
↓↓↓
↓↓↓
↓↓
IL-12, IFN- γ , IP-10MotilityEndocytosisAPM componentexpression
oc sn pres nt on
Longevity of
conventional DC
Attraction of
plasmocytoid or
regulatory DC
Differentiation
of endothelial-
like DC
Neovascula-rization
IDO
IL-10
α
CD4+ TregCD8+ Treg
TA-specific T cells
ed
C t
D E
N D
R I
T I
C
C E
L L
S
↓
↓
↓
↓
↓
↓
Fig. 4 Modulation of the dendritic cells system at the tumor site by tumor-derived cytokines, chemokines and growth factors
Cancer Metastasis Rev (2006) 25:333–356 343

6.1 Regulation of DC migration and homingat the tumor site
Localization in tissues, including tumor tissue, and migrationto lymphoid organs are essential steps in DC immunobiologythat are linked to the differentiation and T-cell stimulatoryfunction. Chemokines play an important role in regulation ofDC trafficking, which is also affected by differential regula-tion of the repertoire of chemokine receptors expressed byimmature and mature DC [119]. During their migration fromblood to peripheral tissues, CCR6+ DC appear to be attractedby a CCL20 gradient that is established by local inflamma-tion [120, 121], secreted in part by tumor cells, themselves(Ferris et al., unpublished data). Immature DC may then un-dergo a maturation process characterized by several mod-ifications, including the reciprocal loss of inflammatorychemokine receptors (such as CCR1, CCR2 and CCR5), aswell as CCR6, and acquisition of CCR7, with resulting re-sponsiveness to CCL19/CCL21. However, whether DCchemoattraction to LN and DC maturation are concomitantprocesses remains controversial [122, 123].
It has been suggested that the CC-chemokine network incancer is controlled at the level of CC-chemokine receptorsand this may account for the phenotypes of infiltrating cellsfound in these tumors. At the same time, the leukocyteinfiltrate may contribute to tumor growth and spread byproviding growth survival factors, angiogeneic stimulationand matrix metalloprotease activation. For instance, recentstudies suggest that tumor cell variants that escaped immunerecognition have a significantly reduced ability to secrete pro-inflammatory cytokines and the CCL5 chemokine, and toexpress immunostimulatory chaperones, i.e. have a reducedability to induce “danger signals” [124]. It has been recentlydemonstrated that many tumor cell lines loss the expressionof CXCL14 [125]. We and other reported that CXCL14 is apotent chemoattractant for immature DC [54, 67] suggestingthat loss expression of CXCL14 at the tumor site might beresponsible, at least in part, for low attraction of DC and thusdeficient initiation of antitumor immune responses.
It is important to point here that chemokines may also up-regulate functional activity of DC. For instance, CXCL14not only stimulates DC maturation and activation [54], but isable to up-regulate expression of antigen-processing ma-chinery components in DC and thus, antigen processingand presentation to T cells (Shurin, unpublished data).Similar results were reported for APC function of macro-phages: macrophages generated from bone marrow progen-itor cells in vitro with M-CSF were defective in APCfunction, while macrophages generated in the presence ofthe CC chemokines Lkn-1, MIP-1α, and RANTES togetherwith M-CSF exhibited much stronger APC function [126].In another study, the ability of tumor cells to release che-motactic factors active on DC was investigated in primary
cultures of papillary carcinoma and was compared to that ofthe corresponding normal thyroid cells obtained from thetumor-free contralateral lobe [115]. It was found thatpapillary carcinoma cells were active in attracting DC andthis chemotactic activity was significant increased byaddition of HGF or IL-1β. Further analysis demonstratedthat unstimulated tumor cells contain large amounts of IL-8and MCP-1, while stimulation with HGF and IL-1β inducedexpression of RANTES (CCL5), MIP-3α (CCL20), IP-10(CXCL10), and, to a lesser extent, MIP-1α (CCL3) andMIP-1β (CCL4). Interestingly, HGF + tumor cell nests wereinfiltrated by CD1a+ DC, suggesting that HGF stimulationof tumor cells is one of the molecular mechanisms involvedin the recruitment of immature DC [115]. Similarly, ex-pression of CCL2, which is a chemoattractant and activatorof macrophages and is a key determinant of the macro-phage infiltrate into tumors, has been reported to besignificantly reduced in ovarian adenocarcinoma [127].Thus, not only the presence of certain chemokines in thetumor milieu, but the absence of DC-stimulating cytokinessuch as CXCL14 [54, 68], may results in impaired functionof DC and deficient induction of antitumor immunity.
Different tumors use different approaches to regulatemigration and homing of immune cells at the tumor bed. Incontrary to the described above down-regulation of DCchemoattracting chemokines by SCCHN and prostate carci-noma cells, melanoma and breast cancer cells use a differentapproach. It has been reported that melanoma cell lines caneffectively chemoattract DC, modulate their phenotype and,eventually, severely damage DC mobility: melanoma-condi-tioned DC exhibited an increased adhesion capacity to a mel-anoma cell lines in vitro and did not migrate in response tochemokines [128]. Analyzing the results of different DC-based immunotherapeutic trials, Soruri and Zwirner (2005)summarized that there is now increasing experimentalevidence demonstrating that DC generated in vitro fromcancer patients’ monocytes in the presence of GM-CSF andIL-4 may be hampered in their ability to migrate in responseto inflammatory as well as homeostatic chemataxins [129].The authors concluded that the “classical” cancer-patient-derived DC may not represent the equivalent of migratoryDC in vivo limiting their use as magic bullets in tumorimmunotherapy. Likewise, reduced migratory potential ofDC was observed if cells were generated from patients withchronic myeloid leukemia [130].
Breast carcinoma cells have been shown to producesoluble factors, which may attract DC and their precursorsin vivo, and promote the differentiation of the latter into LCand immature DC with altered functional capacities.Thomachot et al. have reported that breast carcinoma cellsproduce soluble factors, including MIP-3α (CCL20), whichmay attract DC and their precursors in vivo, and promotethe differentiation of the latter into Langerhans cells (LC)
344 Cancer Metastasis Rev (2006) 25:333–356

and immature DC with altered functional capacities [131].In vitro, breast carcinoma cell lines as well as theirconditioned media promoted CD34+ cell differentiation into2 different CCR6 + subpopulations of DC: (a) with a LCphenotype (CD1a+Langerin+CD86+) and (b) an immatureDC phenotype (CD1a++Langerin−CD86−HLA-DRlowC-D40low). This phenomenon was partly driven by a TGFβ-dependent mechanism since TGFβ neutralizing antibodyblocks breast carcinoma-induced LC differentiation andimmature DC development. These DC failed to maturate inresponse to CD40L or LPS and have a reduced T-cellstimulatory capacity [131].
Thus, certain tumors, including melanoma and breast car-cinoma, may attract or trap immunocompetent cells. Conse-quently, the infiltration of tumor mass by DC differentiatedin the tumor environment may contribute to the impairedimmune response against the tumor. Although the clinicalrelevance of these observations remains largely unknown, itis clear that immune cells infiltrating the tumors are notcapable to prevent tumor cell growth in vivo.
Modulation of DC phenotype and motility by both tumor-derived and immune cell-derived factors takes place not onlyat the tumor site, but in tumor-draining LN. Indeed, thefunctional activity of DC in LN plays a key role in DC/T-cellinteraction and initiation of antigen-specific immune respon-ses. In the metastatic LN, it was found that CCR6 was ex-pressed predominantly within tumor areas by cells exhibitinga dendritic morphology. Tumor cells and macrophagesstrongly expressed the CCR6 ligand CCL20 [114] and clearlycolocalized with CCR6+ DC, while large dendrite-shapedCCR7bright cells were detected occasionally in the T-cell-richareas without tumor cell infiltration [61]. The findings ofdominant CCR6/CCL20 expression suggest that the tumor-invaded LN should be considered as an inflammatory siterather than a typical lymphoid organ [61]. Therefore, in themetastatic LN, maturing DC might indeed display significantheterogeneity regarding expression of CCR receptors and couldeven transiently co-express CCR6 andCCR7 in an intermediatestage of maturation, as has been reported recently [132].Altogether, these data point to a potential role of MIP-3α/CCL20 in the local recruitment of DC in metastatic LN.
Finally, in addition to be modified by tumor-derived fac-tors, DCmotility could be also regulated by cytokines specificfor different tumor milieu. Systemic and local intratumoralcytokines also play an important role in regulating DCimmigration, homing, and emigration from the tumor bed.For instance, IL-10 down-regulates the expression of CCR7and up-regulates the expression of CCR5 transcripts in DC[133]. These changes in CCR7 and CCR5 expression wereassociated with inhibition and augmentation, respectively, ofDC chemotaxis toward their respective agonists, MIP-3β(CCL19) and MIP-1α (CCL3), while in vivo homing of DCfrom peripheral sites to secondary lymphoid tissue in mice
was significantly impaired. Furthermore, Th2-types cyto-kines IL-13 and IL-4 strongly increased CXCR1 andCXCR2 chemokine receptor expression in human DC andin turn, may contribute to the accumulation and positioningof DC in Th2-dominated tumor millennia [134].
Another immunosuppressive cytokine, TGF-β1, that, likeIL-10, suppresses DC maturation and function, inhibits theexpression of CCR7 transcripts in murine DC and their pre-cursors, which is associated with suppressed migration inresponse to MIP-3β (CCL19) [135, 136]. In addition, TGF-β1 enhances CCR5 expression on human monocyte-derivedDC, concomitant with enhanced chemotactic migratory re-sponses to its inflammatory ligand, RANTES [137]. Kobie etal. have demonstrated that TGF-β inhibits DC migration toLN and diminishes their capacity to stimulate IFN-γsecretion by tumor-sensitized T lymphocytes in vivo [136].Thus, the impaired homing capacity of IL-10 and/or TGF-β-treated DC to secondary lymphoid tissue, which is probablylinked to decreased expression of CCR7, may be anothermechanism of the deficient induction of antitumor immunityby DC that had been exposed to different cytokines in thetumor microenvironment.
6.2 Regulation of TADC maturation and functionat the tumor site
The ability of DC to pick up and process tumor antigens, aswell as their capacity to leave the tumor mass and migrateto the regional LN for presenting antigen and providing co-stimulation for T-cells, are all crucial elements of efficientinduction of antitumor immunity. All key functions of DCdepend on the state pf DC differentiation and activation andthus, are regulated by cytokine, growth factors, and che-mokines. It has been well documented that a local cytokinemilieu can influence and even reverse monocyte differen-tiation as well as DC maturation [138], suggesting that thelocal cytokine milieu can have a significant impact onactivation and subsequent function of DC. In fact, Troy etal. indicate that minimally activated DC infiltrate the tumormass of RCC and prostate cancer patients [139, 140]. Otherstudies suggest that the tumor-infiltration DC have animmature phenotype and could induce a partial state of T-cell tolerance to the tumor cells, suggesting that alterationsin DC maturation in the tumor microenvironment mayprevent them from activating antigen-specific T cells [141,142]. It is well known that if antigen-presenting cells do notexpress costimulatory molecules CD80 or CD86, andthereby fail to provide an appropriate second signal for Tcells, tolerance pr anergy may develop [143, 144]. Theability of DC from cancer patients to stimulate T lympho-cytes is profoundly suppressed [104, 145–147], explainingthe deficiency of antitumor immunity in these patients.Furthermore, it has been recently reported that other
Cancer Metastasis Rev (2006) 25:333–356 345

functions of DC may also be inhibited by tumors. Forinstance, Tas et al. noticed a decreased ability of peripheralblood DC to form cluster with T cells in patients withSCCHN [148]. Using different murine and human tumormodels and DC/tumors cell co-culture conditions, it hasbeen shown that DC obtained from tumor bearing animalsor generated in vitro in the presence of tumor derived fac-tors have a significantly impaired ability to produced IL-12[105, 148]. Thrunher et al. observed that DC obtained fromRCC had reduced potential top capture coluble antigen[150]. Thus, in additional to inhibiting DC differentiationand maturation, tumor-derived activity of DC and suppres-sion of DC function in career patients is thought tocontribute to the inhibition of immune responses anddisease progression. A number of tumor types have beenshown to disbalance DC maturation and function, includingmelanoma, prostate cancer, lung cancer, breast cancer,gastric cancer, hepatoma, RCC and neuroblastoma [105,151–159] (in addition, see reviews [11, 13, 160]).
There is direct and indirect evidence supporting the notionthat intratumoral cytokines are involved in regulating DCmaturation and activation in vivo. For instance, immunohis-tochemical staining of gastric carcinoma tissues revealed aninverse correlation between the density of DC and VEGFexpression. This finding was also associated with poorprognosis [161]. In patients with lung, head and neck, andbreast cancers, there was a decrease in the function andnumber of mature DC, which was associated with increasedplasma concentrations of VEGF [162]. Furthermore, IL-6and M-CSF derived from tumor cells and macrophages fromthe tumor microenvironment were shown to switch DCdifferentiation towards macrophage lineage [163]. Anotherfactor produced within the tumor microenvironment thatmight interfere with DC functions is TGF-β. TGF-β is apleiotropic cytokine produced by malignant cells of differenthistological types [164–166]. Among the plethora ofimmunosuppressive effects of TGF-β is the capacity tointerfere with DC maturation and function [136]. These alsoinclude down-regulation of cell surface MHC antigens, co-stimulatory molecules, chemokine receptors, as well asimpairment of in vitro chemotaxis [135, 137].
In addition to secreting their own suppressive mediators,tumor cells may also signal surrounding inflammatory cellsto release suppressive cytokines, such as IL-10. It has beenreported that human lung cancers potently induce Tlymphocyte IL-10 production [167] and that increased levelsof T-cell-derived IL-10 severely impair antitumor immunityin vivo, due to defects in IL-12 production and antigen pre-sentation by DC [168]. Consequently, tumor cells, tumor-associated macrophages and Treg cells often produce IL-10,which suppress DC maturation and function. Elevatedconcentrations of IL-10 within the tumor environment havebeen suggested to limit access of DC to the tumor site
[169], decrease maturation/activation of DC [105], lowerIL-12 synthesis [170], confers tolerogenic properties on DC[171], and promote DC apoptosis [172], as well assusceptibility to autologous NK cell lysis [173].
Although alterations of DC maturation and function in thetumor environment are now well documented, little is knownabout the mechanisms responsible for DC-mediated inhibitionof immune responses or inability of DC to induce TA-specificimmune reaction. Decrease presentation of TA to T cells maybe explained by recently demonstrated inhibition of antigen-processing machinery component expression in DC inducedby tumor-derived factors [174]. Co-culture of human mono-cyte-derived DC with different SCCHN cell lines caused aprofound down-regulation of LMP2, LMP10, MB1, TAP1/2,and other antigen processing pathway proteins in DC, whichwas associated with a decreased presentation of an antigen toantigen-specific autologous CD8+ T cells. Next, cytokineswithin the tumor microenvironment might selectively suppressthe expression of co-stimulatory molecules (B7.1 and B7.2)and trigger the expression of co-inhibitory molecules (B7-H1and B7-H4) on DC, contributing to immune evasion by thetumor [12]. For instance, virtually all myeloid DC isolatedfrom the tissues or draining LN of ovarian carcinomasexpress B7-H1 [175], which selectively induces IL-10 duringpriming of T cells, promotes programmed cell death of effec-tor T cells, and might inhibit T-cell growth by ligation of thePD-1 receptor [176–178]. Interestingly, expression of B7-H1on TADC was significantly increased by IL-10 and VEGFand B7-H1 expressing DC produced low levels of IL-12,secreted increased amount of IL-10 and were unable tostimulate proliferation of T cells, which subsequently leadedto impaired T-cell antitumor immunity [175].
Recently it has been shown that ligation of the co-stimulatory molecules B7-1/B7-2 of DC modulates the IDOactivity in DC [179]. IDO+ DC reduce access to freetryptophan and thus may block the cell-cycle progressionof T cells resulting in inhibition of the clonal expansion of Tcells, activation of T-cell apoptosis, induction of anergy orimmune deviation. IDO+ DC have been detected in breasttumor tissue and tumor-draining lymph nodes in patientswith melanoma and patients with breast, colon, lung andpancreatic cancers [97]. Notably, IDO expression by DCmight be up-regulated by tumor-associated CTLA4+CD4+
CD25+ Treg cells, and effectively convert them into regula-tory DC [12]. Interestingly, it has been shown that mousetumor-draining LN contained a subset of DC that constitu-tively expressed immunosuppressive levels of IDO anddespite comprising only 0.5% of LN cells, these plasmacytoidIDO+ DC potently suppressed T-cell responses to antigenspresented by the plasmacytoid DC themselves and also, in adominant fashion, suppressed T-cell responses to third-partyantigens presented by non-suppressive APC [180]. Thus, onemechanism contributing to immunologic unresponsiveness
346 Cancer Metastasis Rev (2006) 25:333–356

toward tumors may be presentation of tumor antigens bytolerogenic host APC, including plasmacytoid DC andcreation of a local microenvironment that is potentlysuppressive of host antitumor T-cell responses.
New studies began revealing the role of different DC sub-populations in the tumor microenvironment. For instance, Weiet al. reported that functional plasmacytoid DC, but notfunctionally mature myeloid DC, contribute to the tumorenvironmental immunosuppressive network [181]. The authorsshowed that plasmacytoid DC presented in ovarian tumorascites induced IL-10+CCR7+CD45RO+CD8+ regulatory Tcells, which significantly suppress myeloid DC-mediatedtumor antigen specific T-cell effector functions throughIL-10. Plasmacytoid DC were also detected in melanomasand head and neck squamous-cell carcinomas [91, 92].Interestingly, plasmacytoid DC within the tumor microenvi-ronment show reduced expression of TLR9 [92], which isinvolved in DC activation, IFN-α production, and thuspromotion of adaptive Th1-type responses [182]. Togetherwith the data demonstrating that TLR9 ligands mightdecrease plasmacytoid DC ability to migrate in response toCXCR3 ligands and increase their response to CCR7 ligands[183], this indicates that plasmacytoid DC can also bephenotypically and functionally modulated in the tumormicroenvironment.
Curiel et al. have recently suggested a new role for DC in thetumor microenvironment—regulation of tumor angiogenesis[184]. They revealed that high numbers of plasmacytoid DCin ovarian cancer, attracted by CXCL-12/SDF-1, inducedangiogenesis in vivo through production of TNF-α and IL-8.By contrast, myeloid DC, which could suppress angiogenesisin vivo through production of IL-12, were absent frommalignant ascites. Thus, the tumor may attract plasmacytoidDC to augment angiogenesis while excluding myeloid DC toprevent angiogenesis inhibition, demonstrating a novel mech-anism for modulating tumor neovascularization. Similarly, in amurine system, Conejo-Garcia et al. demonstrated thatexpression of β-defensins recruited DC precursors to tumorsand enhanced tumor vascularization and growth in thepresence of VEGF-A expression [185]. β-defensins andVEGF-A cooperate to promote tumor vasculogenesis bycarrying out distinct tasks: β-defensins chemoattract DCprecursors through CCR6, whereas VEGF-A primarily indu-ces their endothelial-like specialization and migration tovessels, which is mediated by VEGFR-2. Therefore, DC arepertinent not only in tumor immunopathogenesis, but also intumor vascularization.
Thus, abundance of VEGF, IL-6, M-CSF, TGF-β, andIL-10, versus negligible GM-CSF, IL-4, IL-12, and IFN-γ(i.e., DC differentiation cytokines) causes an aberrant cyto-kine pattern in the tumor microenvironment and results inimbalance of DC differentiation and maturation. In addition,DC responsiveness to DC differentiation cytokines may be
strongly attenuated by tumor-derived factors. Indeed, TNF-αfailed to up-regulate the expression of HLA-DR and co-stimulatory molecule CD86 by DC that were cultured withneuroblastoma cells [186]. Therefore, immature or partiallydifferentiated myeloid DC can function as regulatory DCand are an important component of the immunosuppressivenetworks in the tumor microenvironment [12].
6.3 Regulation of DC longevity in the tumormicroenvironment
The role of apoptosis in regulating DC differentiation andelimination in cancer, as well as the mechanistic basis of thisregulation, remain to be addressed. Others and we havedemonstrated that elimination of DC or DC precursors in thetumor microenvironment is an important aspect of tumor-induced immunosuppression [13, 152, 158, 187]. Tumor-induced apoptosis of DC has been reported for melanoma,prostate cancer, lung cancer, neuroblastoma and other typesof cancer [186, 188]. It might be responsible for a deficientantitumor immune response in patients with cancer, giventhat tumor cells are known to express a number of pro-apoptotic factors for DC such as IL-10, NO, reactiveoxygen species, hyaluronan, certain gangliosides andothers. For instance, gangliosides GM3 and GD3 frommelanoma cells decreased the viable cell yield in monocyte-derived DC cultures and induced significant DC apoptosis[189]. Exposure of monocyte-derived DC to VEGF hasbeen recently reported to result in induction of DCapoptosis. Interestingly, patients with abnormally elevatedblood concentrations of VEGF showed significantly lowervalues of DC [190]. Lung cancer-derived bombesin-likepeptides might also decrease longevity of DC [153].
However, the intracellular mechanisms responsible for theregulation of cell death or protection of DC from the deathsignals remain poorly understood. Our data suggest that theBcl-2 family of pro- and anti-apoptotic proteins plays animportant role in DC death in cancer [191, 192], although bothmitochondria-dependent and independent apoptotic pathwaysmight be involved in tumor-induced premature apoptosis ofDC [193]. It has been also shown that down-regulation ofphosphoinositide 3-kinase (PI3K) could be an important aspectof tumor-induced apoptosis of DC since tumor supernatantssuppressed Akt, NF-κB, and Bcl-xL in DC [194].
Thus, certain tumor-derived factors, including cytokines[112, 172], may be responsible for shortening DC survivalat the tumor site and failure to initiate antitumor immunity.At the same time, the absence of certain cytokine-mediatesignaling might also be responsible for DC apoptosis in thetumor mass. For instance, inhibition of FLT3 signaling, aknown DC growth factor [195], induces apoptosis in bothmouse and human DC [196]. It will be no completeappreciation of the complexity of DC survival in tumors
Cancer Metastasis Rev (2006) 25:333–356 347

without realizing that many cytokines are able to protectDC from tumor-induced death. CD40 ligation, TNF-α,IL-12, and IL-15 were shown to increase DC resistance tomelanoma and prostate adenocarcinoma mediated apoptosis[191, 192, 197, 198]. Furthermore, chemokine CXCL12was demonstrated to protect tumor plasmacytoid DC fromapoptosis [199]. These data are thus consistent with thehypothesis that a balance of stimulatory vs. inhibitory DCcytokines is responsible for DC longevity at the tumor site.
7 Conclusion remarks
1. Different tumors may suppress DC survival or inhibit DCfunction and this maybe an important mechanism,whereby tumors escape immune recognition in cancerpatients.
2. Monocyte-derived DC generated from patients withcancer might display multiple functional abnormalities.Appropriate correction of DC function ex vivo should bedeveloped when considering vaccination with mono-cyte-derived DC.
3. Tumor-induced immunosuppression of DC can also beseen in involved SLN. It might be mediated in part bycytokines such as IL-10 and IFN-γ, and induction ofIDO in DC. SLN cytokine microenvironment is similarto that of non-SLN in the absence of residual tumor cells,suggesting that immunosuppressive SLN cytokine mi-croenvironment can be reversed.
4. Manipulation of local cytokine microenvironment atthe tumor site by using cytokines, such as GM-CSF,IL-12, IL-2, IL-4 can reverse the evidence of immunedysfunction both in the tumor and SLN.
5. Growing evidence suggests that a single cytokine, e.g.,IFN-γ, can neither be classified solely as “proinflamma-tory” or “anti-inflammatory” in its function nor be reliablyused as a measure of desired enhanced immune responsefollowing an immunotherapy. Different temporal andcombined cytokine exposures can significantly alter itseffects on a given group of target cells, including DC.
6. Numerous data proved the potential usefulness of atherapeutic approach to eliminate immunosuppressivetumor-derived factors to improve the effectiveness ofDC-based vaccines.
8 Key unanswered questions
The challenge for the tumor immunologists now is tounderstand the mechanisms by which tumors becomerefractory to immune modulation. Achieving this goal islikely to be dependent upon continuing research to improveour understanding of the heterogeneity and complexity of the
tumor microenvironment. There are several under-developedareas of research and a number of unanswered questions,which may accelerate our move to our main goal—develop-ment of effective means to prevent or treat malignant diseases.For example,
– The inflammatory microenvironment is thought to playa pivotal part in tumorigenesis. Based on recentdevelopments in psychoneuroimmunology, it seemsquite possible that the peripheral nervous systemparticipates in informing the brain about tumorigenesisby transmitting information to the brain about tumor-associated proinflammatory cytokines. This axis mightbe associated with a more effective induction ofantitumor immunity. But this hypothesis requires exper-imental evidence.
– The role of cytokines, chemokines, and growth factorsin the interaction between residual dormant tumor cellsand surrounding immune cells should be investigated.
– To date, no comprehensive analyses of immune cellphenotype and function have been carried out in patientswith many types of cancer. Such information is of criticalimportance for designing effective immune therapeuticstrategies for both early- and late-stage disease.
– Further investigation is clearly required to determinehow the complex interaction among DC, T cells, andtumor cells results in immune suppression.
– How do different cytokines, chemokines and growth fac-tors interact with multiple differentiation, growth, andsurvival signaling pathways in tumor cells and tumorinfiltrating leukocytes?
– How to distinguish between age-dependent and cancer-mediated alterations of the DC system? Does the cyto-kine network may be changed with age in the tumorenvironment as it does in the immune system?
Acknowledgments This work was support in part by 2RO1CA084270 (M.R.S.) DOD PC050252 (M.R.S.), and 1RO1 CA115902(to RLF).
References
1. Brigati, C., Noonan, D. M., Albini, A., & Benelli R. (2002).Tumors and inflammatory infiltrates: friends or foes? Clinical &Experimental Metastasis, 19, 247–258.
2. Robinson, S. C., & Coussens, L. M. (2005). Soluble mediatorsof inflammation during tumor development. Advances in CancerResearch, 93, 159–187.
3. Balkwill, F. (2004). The significance of cancer cell expression ofthe chemokine receptor CXCR4. Seminars in Cancer Biology,14, 171–179.
4. Mantovani, A. (2004). Chemokines in neoplastic progression.Seminars in Cancer Biology, 14, 147–148.
348 Cancer Metastasis Rev (2006) 25:333–356

5. Buell, J. F., Gross, T. G., & Woodle, E. S. (2005). Malignancyafter transplantation. Transplantation, 80, S254–S264.
6. Jensen, P., Hansen, S., Moller, B., Leivestad, T., Pfeffer, P.,Geiran, O., et al. (1999). Skin cancer in kidney and hearttransplant recipients and different long-term immunosuppressivetherapy regimens. Journal of the American Academy ofDermatology, 40, 177–186.
7. Dunn, G. P., Old, L. J., & Schreiber, R. D. (2004). Theimmunobiology of cancer immunosurveillance and immunoedit-ing. Immunity, 21, 137–148.
8. Dunn, G. P., Ikeda, H., Bruce, A. T., Koebel, C., Uppaluri, R.,Bui, J., et al. (2005). Interferon-gamma and Cancer Immunoedit-ing. Immunologic Research, 32, 231–246.
9. Khong, H. T., & Restifo, N. P. (2002). Natural selection of tumorvariants in the generation of “tumor escape” phenotypes. NatureImmunology, 3, 999–1005.
10. Restifo, N. P., Antony, P. A., Finkelstein, S. E., Leitner, W. W.,Surman, D. P., Theoret, M. R., et al. (2002). Assumptions of thetumor ‘escape’ hypothesis. Seminars in Cancer Biology, 12, 81–86.
11. Yang, L., & Carbone, D. P. (2004). Tumor-host immuneinteractions and dendritic cell dysfunction. Advances in CancerResearch, 92, 13–27.
12. Zou, W. (2005). Immunosuppressive networks in the tumourenvironment and their therapeutic relevance. Nature ReviewsCancer, 5, 263–274.
13. Shurin, M. R., & Gabrilovich, D. I. (2001). Regulation ofdendritic cell system by tumor. Cancer Research, Therapy andControl, 11, 65–78.
14. Shurin, M. R., Lu, L., Kalinski, P., Stewart-Akers, A. M., &Lotze, M. T. (1999). Th1/Th2 balance in cancer, transplantationand pregnancy. Springer Seminars in Immunopathology, 21,339–359.
15. Moss, R. B., Moll, T., El-Kalay, M., Kohne, C., Soo Hoo, W.,Encinas, J. et al. (2004). Th1/Th2 cells in inflammatory diseasestates: therapeutic implications. Expert Opin Biol Ther, 4, 1887–1896.
16. Murakami, H., Ogawara, H., & Hiroshi, H. (2004). Th1/Th2cells in patients with multiple myeloma. Hematology, 9, 41–45.
17. Li, R., Ruttinger, D., Si, L. S., & Wang, Y. L. (2003). Analysis ofthe immunological microenvironment at the tumor site inpatients with non-small cell lung cancer. Langenbecks ArchSurg, 388, 406–412.
18. Lin, E. Y., & Pollard, J. W. (2004). Role of infiltrated leucocytesin tumour growth and spread. British Journal of Cancer, 90,2053–2058.
19. Pollard, J. W. (2004). Tumour-educated macrophages promotetumour progression and metastasis. Nature Reviews Cancer, 4,71–78.
20. Mantovani, A., Allavena, P., & Sica, A. (2004). Tumour-associated macrophages as a prototypic type II polarisedphagocyte population: role in tumour progression. EuropeanJournal of Cancer, 40, 1660–1667.
21. Danielpour, D. (2005). Functions and regulation of transforminggrowth factor-beta (TGF-beta) in the prostate. European Journalof Cancer, 41, 846–857.
22. Presta, M., Dell'Era, P., Mitola, S., Moroni, E., Ronca, R., &Rusnati, M. (2005). Fibroblast growth factor/fibroblast growthfactor receptor system in angiogenesis. Cytokine & GrowthFactor Reviews, 16, 159–178.
23. Loureiro, R. M., & D'Amore, P. A. (2005). Transcriptionalregulation of vascular endothelial growth factor in cancer.Cytokine & Growth Factor Reviews, 16, 77–89.
24. Culig, Z., Steiner, H., Bartsch, G., & Hobisch, A. (2005).Interleukin-6 regulation of prostate cancer cell growth. Journalof Cellular Biochemistry, 95, 497–505.
25. Kay, N. E., & Pittner, B. T. (2003). IL-4 biology: impact onnormal and leukemic CLL B cells. Leukemia & Lymphoma, 44,897–903.
26. Apte, R. N., & Voronov, E. (2002). Interleukin-1—a majorpleiotropic cytokine in tumor–host interactions. Seminars inCancer Biology, 12, 277–290.
27. Yue, F. Y., Dummer, R., Geertsen, R., Hofbauer, G., Laine, E.,Manolio, S., et al. (1997). Interleukin-10 is a growth factor forhuman melanoma cells and down-regulates HLA class-I, HLAclass-II and ICAM-1 molecules. International journal of Cancer,71, 630–637.
28. Salazar-Onfray, F., Charo, J., Petersson, M., Freland, S., Noffz,G., Qin, Z., et al. (1997). Down-regulation of the expression andfunction of the transporter associated with antigen processing inmurine tumor cell lines expressing IL-10. Journal of Immunol-ogy, 159, 3195–3202.
29. Ogden, C. A., Pound, J. D., Batth, B. K., Owens, S.,Johannessen, I., Wood, K. et al. (2005). Enhanced apoptoticcell clearance capacity and B cell survival factor production byIL-10-activated macrophages: implications for Burkitt's lympho-ma. Journal of Immunology, 174, 3015–3023.
30. Stassi, G., Todaro, M., Zerilli, M., Ricci-Vitiani, L., Di Liberto,D., Patti, M., et al. (2003). Thyroid cancer resistance tochemotherapeutic drugs via autocrine production of interleukin-4 and interleukin-10. Cancer Research, 63, 6784–6790.
31. Vicari, A. P., & Trinchieri, G. (2004). Interleukin-10 in viraldiseases and cancer: exiting the labyrinth? ImmunologicalReviews, 202, 223–236.
32. Gorsch, S. M., Memoli, V. A., Stukel, T. A., Gold, L. I., &Arrick, B. A. (1992). Immunohistochemical staining for trans-forming growth factor beta 1 associates with disease progressionin human breast cancer. Cancer Research, 52, 6949–6952.
33. Doran, T., Stuhlmiller, H., Kim, J. A., Martin, E. W. Jr., &Triozzi, P. L. (1997). Oncogene and cytokine expression ofhuman colorectal tumors responding to immunotherapy. Journalof Immunotherapy, 20, 372–376.
34. Krasagakis, K., Tholke, D., Farthmann, B., Eberle, J., Man-smann, U., & Orfanos, C. E. (1998). Elevated plasma levels oftransforming growth factor (TGF)-beta1 and TGF-beta2 inpatients with disseminated malignant melanoma. British Journalof Cancer, 77, 1492–1494.
35. Chen, W., Frank, M. E., Jin, W., & Wahl, S. M. (2001). TGF-beta released by apoptotic T cells contributes to an immunosup-pressive milieu. Immunity, 14, 715–725.
36. Letterio, J. J. (2005). TGF-beta signaling in T cells: rolesin lymphoid and epithelial neoplasia. Oncogene, 24, 5701–5712.
37. Chang, C. J., Liao, C. H., Wang, F. H., & Lin, C. M. (2003).Transforming growth factor-beta induces apoptosis in antigen-specific CD4+ T cells prepared for adoptive immunotherapy.Immunology Letters, 86, 37–43.
38. Wang, J., Xi, L., Gooding, W., Godfrey, T. E., & Ferris, R. L.(2005). Chemokine receptors 6 and 7 identify a metastaticexpression pattern in squamous cell carcinoma of the head andneck. Advances in Oto-Rhino-Laryngology, 62, 121–133.
39. Wang, J., Zhang, X., Thomas, S. M., Grandis, J. R., Wells, A.,Chen, Z. G. et al. (2005). Chemokine receptor 7 activatesphosphoinositide-3 kinase-mediated invasive and prosurvivalpathways in head and neck cancer cells independent of EGFR.Oncogene, 24, 5897–5904.
40. Kulbe, H., Levinson, N. R., Balkwill, F., & Wilson, J. L. (2004).The chemokine network in—much more than directing cellmovement. International Journal of Developmental Biology, 48,489–496.
41. Kobel, M., Budianto, D., Schmitt, W. D., Borsi, L., Siri, A., &Hauptmann, S. (2005). Influence of various cytokines on
Cancer Metastasis Rev (2006) 25:333–356 349

adhesion and migration of the colorectal adenocarcinoma cellline HRT-18. Oncology, 68, 33–39.
42. He, Y. G., Mayhew, E., Mellon, J., & Niederkorn, J. Y. (2004).Expression and possible function of IL-2 and IL-15 receptors onhuman uveal melanoma cells. Invest Ophthalmol Vis Sci, 45,4240–4246.
43. Mantovani, A., Bottazzi, B., Colotta, F., Sozzani, S., & Ruco, L.(1992). The origin and function of tumor-associated macro-phages. Immunology Today, 13, 265–270.
44. Sica, A., Saccani, A., & Mantovani, A. (2002). Tumor-associatedmacrophages: a molecular perspective. Int Immunopharmacol, 2,1045–1054.
45. Wyckoff, J., Wang, W., Lin, E. Y., Wang, Y., Pixley, F., Stanley,E. R., et al. (2004). A paracrine loop between tumor cells andmacrophages is required for tumor cell migration in mammarytumors. Cancer Research, 64, 7022–7029.
46. Grimshaw, M. J., Hagemann, T., Ayhan, A., Gillett, C. E.,Binder, C., & Balkwill, F. R. (2004). A role for endothelin-2 andits receptors in breast tumor cell invasion. Cancer Research, 64,2461–2468.
47. Iwasaki, K., Torisu, M., & Fujimura, T. (1986). Malignant tumorand eosinophils. I. Prognostic significance in gastric cancer.Cancer, 58, 1321–1327.
48. Ohashi, Y., Ishibashi, S., Suzuki, T., Shineha, R., Moriya, T.,Satomi, S. et al. (2000). Significance of tumor associated tissueeosinophilia and other inflammatory cell infiltrate in earlyesophageal squamous cell carcinoma. Anticancer Research, 20,3025–3030.
49. Kruger-Krasagakes, S., Li, W., Richter, G., Diamantstein, T., &Blankenstein, T. (1993). Eosinophils infiltrating interleukin-5gene-transfected tumors do not suppress tumor growth. Europe-an Journal of Immunology, 23, 992–995.
50. Szlosarek, P., & Balkwill, F. (2004). The inflammatory cytokinenetwork of epithelial cancer: therapeutic implications. NovartisFoundation Symposium, 256, 227-37; discussion 237–40, 259–269.
51. Tsao, M. S., Liu, N., Nicklee, T., Shepherd, F., & Viallet, J.(1997). Angiogenesis correlates with vascular endothelial growthfactor expression but not with Ki-ras oncogene activation in non-small cell lung carcinoma. Clin Cancer Research, 3, 1807–1814.
52. Anderson, P. S., Smith, H. O., Goldberg, G. L., Fields, A. L.,Runowicz, C. D., & Pollard, J. W. (1999). Colony-stimulatingfactor-1 and its receptor do not have a role in the pathogenesis ofuterine sarcomas. Gynecologic Oncology, 74, 202–207.
53. Hemmerlein, B., Markus, A., Wehner, M., Kugler, A., Zschunke,F., & Radzum, H. J. (2000). Expression of acute and late-stageinflammatory antigens, c-fms, CSF-1, and human monocyticserine esterase 1, in tumor-associated macrophages of renal cellcarcinomas. Cancer Immunology and Immunotherapy, 49, 485–492.
54. Shurin, G. V., Ferris, R., Tourkova, I. L., Perez, L., Lokshin, A.,Balkir, L., et al. (2005). Loss of new chemokine CXCL14 intumor tissue is associated with low infiltration by dendritic cells(DC), while restoration of human CXCL14 expression in tumorcells causes attraction of DC both in vitro and in vivo. Journal ofImmunology, 174, 5490–5498.
55. Allinen, M., Beroukhim, R., Cai, L., Brennan, C., Lahti-Domenici, J., Huang, H., et al. (2004). Molecular characteriza-tion of the tumor microenvironment in breast cancer. CancerCell, 6, 17–32.
56. Fujii, A., Oshima, K., Hamasaki, M., Utsunomiya, H., Okazaki,M., Kagami, Y., et al. (2005). Differential expression ofcytokines, chemokines and their receptors in follicular lympho-ma and reactive follicular hyperplasia: assessment by comple-mentary DNA microarray. Oncology Reports, 13, 819–824.
57. Breuhahn, K., Vreden, S., Haddad, R., Beckebaum, S., Stippel,D., Flemming, P., et al. (2004). Molecular profiling of humanhepatocellular carcinoma defines mutually exclusive interferonregulation and insulin-like growth factor II overexpression.Cancer Research, 64, 6058–6064.
58. Leong, S. P., Peng, M., Zhou, Y. M., Vaquerano, J. E., & Chang,.J. W. (2002). Cytokine profiles of sentinel lymph nodes drainingthe primary melanoma. Annals of Surgical Oncology, 9, 82–87.
59. Lee, J. H., Torisu-Itakara, H., Cochran, A. J., Kadison, A.,Huynh, Y., Morton, D. L., et al. (2005). Quantitative analysis ofmelanoma-induced cytokine-mediated immunosuppression inmelanoma sentinel nodes. Clinical Cancer Research, 11, 107–112.
60. Kusuda, T., Shigemasa, K., Arihiro, K., Fujii, T., Nagai, N., &Ohama, K. (2005). Relative expression levels of Th1 and Th2cytokine mRNA are independent prognostic factors in patientswith ovarian cancer. Oncology Reports, 13, 1153–1158.
61. Movassagh, M., Spatz, A., Davoust, J., Lebecque, S., Romero,P., Pittet, M., et al. (2004). Selective accumulation of mature DC-Lamp+ dendritic cells in tumor sites is associated with efficientT-cell-mediated antitumor response and control of metastaticdissemination in melanoma. Cancer Research, 64, 2192–2198.
62. Becker, Y. (1992). Anticancer role of dendritic cells (DC) inhuman and experimental cancers—a review. Anticancer Re-search, 12, 511–250.
63. Reichert, T. E., Scheuer, C., Day, R., Wagner, W., & Whiteside,T. L. (2001). The number of intratumoral dendritic cells and zeta-chain expression in T cells as prognostic and survival biomarkersin patients with oral carcinoma. Cancer, 91, 2136–2147.
64. Goldman, S. A., Baker, E., Weyant, R. J., Clarke, M. R., Myers,J. N., & Lotze, M. T. (1998). Peritumoral CD1a-positivedendritic cells are associated with improved survival in patientswith tongue carcinoma. Archives of Otolaryngology, Head &Neck Surgery, 124, 641–646.
65. Kikuchi, K., Kusama, K., Taguchi, K., Ishikawa, F., Okamoto,M., Shimada, J., et al. (2002). Dendritic cells in humansquamous cell carcinoma of the oral cavity. AnticancerResearch, 22, 545–557.
66. Miyagawa, S., Soeda, J., Takagi, S., Miwa, S., Ichikawa, E., &Noike, T. (2004). Prognostic significance of mature dendriticcells and factors associated with their accumulation in metastaticliver tumors from colorectal cancer. Hum Pathol, 35, 1392–1396.
67. Shellenberger, T. D., Wang, M., Gujrati, M., Jayakumar, A.,Strieter, R. M., Burdick, M. D., et al. (2004). BRAK/CXCL14 isa potent inhibitor of angiogenesis and a chemotactic factor forimmature dendritic cells. Cancer Research, 64, 8262–8270.
68. Schaerli, P., Willimann, K., Ebert, L. M., Walz, A., & Moser,B. (2005). Cutaneous CXCL14 targets blood precursors toepidermal niches for Langerhans cell differentiation. Immunity,23, 331–342.
69. Fushimi, T., Kojima, A., Moore, M. A., & Crystal, R. G. (2000).Macrophage inflammatory protein 3alpha transgene attractsdendritic cells to established murine tumors and suppressestumor growth. Journal of Clinical Investigation, 105, 1383–1393.
70. Ahmed, S. U., Okamoto, M., Oshikawa, T., Tano, T., Sasai, A.,Kan, S., et al. (2004). Anti-tumor effect of an intratumoraladministration of dendritic cells in combination with TS-1, anoral fluoropyrimidine anti-cancer drug, and OK-432, a strepto-coccal immunopotentiator: involvement of toll-like receptor 4.Journal of Immunotherapy, 27, 432–441.
71. Castellano, G., Woltman, A. M., Nauta, A. J., Roos, A., Trouw,L. A., Seelen, M. A., et al. (2004). Maturation of dendritic cellsabrogates C1q production in vivo and in vitro. Blood, 103,3813–3820.
350 Cancer Metastasis Rev (2006) 25:333–356

72. Ehtesham, M., Kabos, P., Gutierrez, M. A., Samoto, K., Black,K. L., & Yu, J. S. (2003). Intratumoral dendritic cell vaccinationelicits potent tumoricidal immunity against malignant glioma inrats. Journal of Immunotherapy, 26, 107–116.
73. Melcher, A., Todryk, S., Bateman, A., Chong, H., Lemoine, N.R., & Vile, R. G. (1999). Adoptive transfer of immature dendriticcells with autologous or allogeneic tumor cells generatessystemic antitumor immunity. Cancer Research, 59, 2802–2805.
74. Pirtskhalaishvili, G., Shurin, G. V., Gambotto, A., Esche, C.,Wahl, M., Yurkovetsky, Z. R., et al. (2000). Transduction ofdendritic cells with Bcl-xL increases their resistance to prostatecancer-induced apoptosis and antitumor effect in mice. Journalof Immunology, 165, 1956–1964.
75. Yurkovetsky, Z. R., Shurin, G. V., Barry, D. A., Schuh, A. C.,Shurin, M. R., & Robbins, P. D. (2006). Comparative analysis ofantitumor activity of CD40L, RANKL, and 4-1BBL in vivofollowing intratumoral administration of viral vectors or trans-duced dendritic cells. Journal of Gene Medicine, 8, 129–137.
76. Tong, Y., Song, W., & Crystal, R. G. (2001). Combinedintratumoral injection of bone marrow-derived dendritic cellsand systemic chemotherapy to treat pre-existing murine tumors.Cancer Research, 61, 7530–7535.
77. Triozzi, P. L., Khurram, R., Aldrich, W. A., Walker, M. J., Kim,J. A., & Jaynes, S. (2000). Intratumoral injection of dendriticcells derived in vitro in patients with metastatic cancer. Cancer,89, 2646–2654.
78. Feijoo, E., Alfaro, C., Mazzolini, G., Serra, P., Penuelas, I.,Arina, A., et al. (2005). Dendritic cells delivered inside humancarcinomas are sequestered by interleukin-8. InternationalJournal of Cancer, 116, 275–281.
79. Mazzolini, G., Alfaro, C., Sangro, B., Feijoo, E., Ruiz, J.,Benito, A., et al. (2005). Intratumoral injection of dendritic cellsengineered to secrete interleukin-12 by recombinant adenovirusin patients with metastatic gastrointestinal carcinomas. Journalof Clinical Oncology, 23, 999–1010.
80. Melero, I., Vile, R. G., & Colombo, M. P. (2000). Feedingdendritic cells with tumor antigens: self-service buffet or a lacarte? Gene Therapy, 7, 1167–1170.
81. Sun, J., Zhang, J., Chen, J., Chen, H., & Chew, Y. (2001). Invitro study on the morphology of human blood dendritic cellsand LPAK cells inducing apoptosis of the hepatoma cell line.Chinese Medical Journal (Engl), 114, 600–605.
82. Vanderheyde, N., Aksoy, E., Amraoui, Z., Vandenabeele, P.,Goldman, M., & Willems, F. (2001). Tumoricidal activity ofmonocyte-derived dendritic cells: evidence for a caspase-8-dependent, Fas-associated death domain-independent mecha-nism. Journal of Immunology, 167, 3565–3569.
83. Yang, R., Xu, D., Zhang, A., & Gruber, A. (2001). Immaturedendritic cells kill ovarian carcinoma cells by a FAS/FASLpathway, enabling them to sensitize tumor-specific CTLs.International Journal of Cancer, 94, 407–413.
84. Hoon, D. S., Bowker, R. J., & Cochran, A. J. (1987). Suppressorcell activity in melanoma-draining lymph nodes. Cancer Re-search, 47, 1529–1533.
85. Cochran, A. J., Morton, D. L., Stern, S., Lana, A. M., Essner, R.,& Wen, D. R. (2001). Sentinel lymph nodes show profounddownregulation of antigen-presenting cells of the paracortex:implications for tumor biology and treatment. Modern Patholo-gy, 14, 604–608.
86. Lana, A. M., Wen, D. R., & Cochran, A. J. (2001). Themorphology, mmunophenotype and distribution of paracorticaldendritic leucocytes in lymph nodes regional to cutaneousmelanoma. Melanoma Research, 11, 401–410.
87. Deng, Y., Yuan, X., & Chen, Z. (1997). Immunobiologicalsignificance of S-100 protein positive dendritic cells (S-100+DC)
in patients with oral squamous cell carcinoma. Zhonghua KouQiang Yi Xue Za Zhi, 32, 174–176.
88. Sakakura, K., Chikamatsu, K., Takahashi, K., Whiteside, T. L.,& Furuya, N. (2005). Maturation of circulating dendritic cellsand imbalance of T-cell subsets in patients with squamous cellcarcinoma of the head and neck. Cancer Immunology andImmunotherapy, 55, 151–159.
89. Cella, M., Jarrossay, D., Facchetti, F., Alebardi, O., Nakajima,H., & Lanzavecchia, A. (1999). Colonna M, Plasmacytoidmonocytes migrate to inflamed lymph nodes and produce largeamounts of type I interferon. Natural Medicines, 5, 919–923.
90. Vermi, W., Bonecchi, R., Facchetti, F., Bianchi, D., Sozzani, S.,Festa, S., et al. (2003). Recruitment of immature plasmacytoiddendritic cells (plasmacytoid monocytes) and myeloid dendriticcells in primary cutaneous melanomas. Journal of Pathology,200, 255–268.
91. Salio, M., Cella, M., Vermi, W., Facchetti, F., Palmowski, M. J.,Smith, C. L., et al. (2003). Plasmacytoid dendritic cells primeIFN-gamma-secreting melanoma-specific CD8 lymphocytes andare found in primary melanoma lesions. European Journal ofImmunology, 33, 1052–1062.
92. Hartmann, E., Wollenberg, B., Rothenfusser, S., Wagner, M.,Wellisch, D., Mack, B., et al. (2003). Identification andfunctional analysis of tumor-infiltrating plasmacytoid dendriticcells in head and neck cancer. Cancer Research, 63, 6478–6487.
93. Cox, K., North, M., Burke, M., Singhal, H., Renton, S., Aqel,N., et al. (2005). Plasmacytoid dendritic cells (PDC) are themajor DC subset innately producing cytokines in human lymphnodes. Journal of Leukocyte Biology, 78, 1142–1152.
94. Munn, D. H., Sharma, M. D., Lee, J. R., Jhaver, K. G., Johnson,T. S., Keskin, D. B., et al. (2002). Potential regulatory functionof human dendritic cells expressing indoleamine 2,3-dioxyge-nase. Science, 297, 1867–1870.
95. Taylor, M. W., & Feng, G. S. (1991). Relationship betweeninterferon-gamma, indoleamine 2,3-dioxygenase, and tryptophancatabolism. FASEB Journal, 5, 2516–2522.
96. Frumento, G., Rotondo, R., Tonetti, M., Damonte, G., Benatti,U., & Ferrara, G. B. (2002). Tryptophan-derived catabolites areresponsible for inhibition of T and natural killer cell proliferationinduced by indoleamine 2,3-dioxygenase. Journal of Experimen-tal Medicine, 196, 459–468.
97. Munn, D. H. (2002). Tolerogenic antigen-presenting cells.Annals of the New York Academy of Sciences, 961, 343–345.
98. Krant, M. J., Manskopf, G., Brandrup, C. S., & Madoff, M. A.(1968). Immunologic alterations in bronchogenic cancer. Se-quential study. Cancer, 21, 623–631.
99. Brugarolas, A., & Takita, H. (1973). Immunologic status in lungcancer. Chest, 64, 427–430.
100. Wustrow, T. P., & Mahnke, C. G. (1996). Causes of immuno-suppression in squamous cell carcinoma of the head and neck.Anticancer Research, 16, 2433–2468.
101. Chen, H. L., Girgis, K. R., Cunningham, H. T., Meny, G. M.,Nadaf, S., Kavanaugh, D., et al. (1996). Immunologic dysfunc-tion in cancer. Nature Medicine, 2, 1096–1103.
102. Wojtowicz-Praga, S. (1997). Reversal of tumor-induced immu-nosuppression: a new approach to cancer therapy. Journal ofImmunotherapy, 20, 165–177 (see comments).
103. Ferrigno, P., Henry, M., Kahana, J., Koepp, D., Lee, M., Nguyen,L., et al. (1996). Regional immunosuppression in esophagealsquamous cancer: evidence from functional studies with matchedlymph nodes. Experimental Cell Research, 229, 212–216.
104. Gabrilovich, D. I., Chen, H. L., Girgis, K. R., Cunningham, H. T.,Meny, G. M., Nadaf, S., et al. (1996). Production of vascularendothelial growth factor by human tumors inhibits the functionalmaturation of dendritic cells. Natural Medicines, 2, 1096–1103.
Cancer Metastasis Rev (2006) 25:333–356 351

105. Shurin, M. R., Yurkovetsky, Z. R., Tourkova, I. L., Balkir, L., &Shurin, G. V. (2002). Inhibition of CD40 expression and CD40-mediated dendritic cell function by tumor-derived IL-10.International Journal of Cancer, 101, 61–68.
106. Lissoni, P., Vigore, L., Ferranti, R., Bukovec, R., Meregalli, S.,Mandala, M., et al. (1999). Circulating dendritic cells in earlyand advanced cancer patients: diminished percent in themetastatic disease. Journal of Biological Regulators and Ho-meostatic Agents, 13, 216–219.
107. Yanagimoto, H., Takai, S., Satoi, S., Toyokawa, H., Takahashi,K., Terakawa, N., et al. (2005). Impaired function of circulatingdendritic cells in patients with pancreatic cancer. ClinicalImmunology, 114, 52–60.
108. Neves, A. R., Ensina, L. F., Anselmo, L. B., Leite, K. R., Buzaid,A. C., Camara-Lopes, L. H., et al. (2005). Dendritic cells derivedfrom metastatic cancer patients vaccinated with allogeneicdendritic cell-autologous tumor cell hybrids express moreCD86 and induce higher levels of interferon-gamma in mixedlymphocyte reactions. Cancer Immunology and Immunotherapy,54, 61–66.
109. Pedersen, A. E., Thorn, M., Gad, M., Walter, M. R., Johnsen, H.E., Gaarsdal, E., et al. (2005). Phenotypic and functionalcharacterization of clinical grade dendritic cells generated frompatients with advanced breast cancer for therapeutic vaccination.Scandinavian Journal of Immunology, 61, 147–156.
110. Sakakura, K., Chikamatsu, K., Sakurai, T., Takahashi, K.,Murata, T., Oriuchi, N., et al. (2005). Infiltration of dendriticcells and NK cells into the sentinel lymph node in oral cavitycancer. Oral Oncology, 41, 89–96.
111. Wojas, K., Tabarkiewicz, J., Jankiewicz, M., & Rolinski, J.(2004). Dendritic cells in peripheral blood of patients with breastand lung cancer—a pilot study. Folia Histochemica et Cytobio-logica, 42, 45–48.
112. Della Porta, M., Danova, M., Rigolin, G. M., Brugnatelli, S.,Rovati, B., Tronconi, C., et al. (2005). Dendritic cells andvascular endothelial growth factor in colorectal cancer: correla-tions with clinicobiological findings. Oncology, 68, 276–284.
113. Vakkila, J., Thomson, A. W., Vettenranta, K., Sariola, H., &Saarinen-Pihkala, U. M. (2004). Dendritic cell subsets inchildhood and in children with cancer: relation to age anddisease prognosis. Clinical and Experimental Immunology, 135,455–461.
114. Bell, D., Chomarat, P., Broyles, D., Netto, G., Harb, G. M.,Lebecque, S., et al. (1999). In breast carcinoma tissue, immaturedendritic cells reside within the tumor, whereas mature dendriticcells are located in peritumoral areas. Journal of ExperimentalMedicine, 190, 1417–1426.
115. Scarpino, S., Stoppacciaro, A., Ballerini, F., Marchesi, M., Prat,M., Stella, M. C., et al. (2000). Papillary carcinoma of thethyroid: hepatocyte growth factor (HGF) stimulates tumor cellsto release chemokines active in recruiting dendritic cells.American Journal of Pathology, 156, 831–837.
116. Jonuleit, H., Schmitt, E., Schuler, G., Knop, J., & Enk, A. H.(2000). Induction of interleukin 10-producing, nonproliferatingCD4(+) T cells with regulatory properties by repetitive stimula-tion with allogeneic immature human dendritic cells. Journal ofExperimental Medicine, 192, 1213–1222.
117. Oldenhove, G., de Heusch, M., Urbain-Vansanten, G., Urbain, J.,Maliszewski, C., Leo, O., et al. (2003). CD4+ CD25+ regulatoryT cells control T helper cell type 1 responses to foreign antigensinduced by mature dendritic cells in vivo. Journal of Experi-mental Medicine, 198, 259–266.
118. Tien, A. H., Xu, L., & Helgason, C. D. (2005). Altered immunityaccompanies disease progression in a mouse model of prostatedysplasia. Cancer Research, 65, 2947–2955.
119. Sozzani, S., Allavena, P., Vecchi, A., & Mantovani A. (2000).Chemokines and dendritic cell traffic. Journal of ClinicalImmunology, 20, 151–160.
120. Banchereau, J., Briere, F., Caux, C., Davoust, J., Lebecque, S.,Liu, Y. J., et al. (2000). Immunobiology of dendritic cells.Annual Review of Immunology, 18, 767–811.
121. Dieu, M. C., Vanbervliet, B., Vicari, A., Bridon, J. M., Oldham,E., Ait-Yahia, S., et al. (1998). Selective recruitment of immatureand mature dendritic cells by distinct chemokines expressed indifferent anatomic sites. Journal of Experimental Medicine, 188,373–386.
122. Sozzani, S., Luini, W., Borsatti, A., Polentarutti, N., Zhou, D.,Piemonti, L., et al. (1997). Receptor expression and responsive-ness of human dendritic cells to a defined set of CC and CXCchemokines. Journal of Immunology, 159, 1993–2000.
123. Randolph, G. J. (2002). Is maturation required for Langerhans cellmigration? Journal of Experimental Medicine, 196, 413–416.
124. Manjili, M. H., Arnouk, H., Knutson, K. L., Kmieciak, M.,Disis, M. L., Subjeck, J. R., et al. (2005). Emergence of immuneescape variant of mammary tumors that has distinct proteomicprofile and a reduced ability to induce “danger signals”. BreastCancer Research and Treatment, 1–9.
125. Frederick, M. J., Henderson, Y., Xu, X., Deavers, M. T., Sahin,A. A., Wu, H., et al. (2000). In vivo expression of the novelCXC chemokine BRAK in normal and cancerous human tissue.American Journal of Pathology, 156, 1937–1950.
126. Lee, J. K., Kim, J. K., Lee, Y. R., Kim, H. S., Im, S. A., Kim, K.,et al. (2005). Exposure to chemokines during maturationmodulates antigen presenting cell function of mature macro-phages. Cellular Immunology, 234, 1–8.
127. Arnold, J. M., Huggard, P. R., Cummings, M., Ramm, G. A., &Chenevix-Trench, G. (2005). Reduced expression of chemokine(C-C motif) ligand-2 (CCL2) in ovarian adenocarcinoma. BritishJournal of Cancer, 92, 2024–2031.
128. Remmel, E., Terracciano, L., Noppen, C., Zajac, P., Heberer, M.,Spagnoli, G. C., et al. (2001). Modulation of dendritic cellphenotype and mobility by tumor cells in vitro. HumanImmunology, 62, 39–49.
129. Soruri, A., & Zwirner, J. (2005). Dendritic cells: limited potentialin immunotherapy. International Journal of Biochemistry & CellBiology, 37, 241–245.
130. Eisendle, K., Wolf, D., Gastl, G., & Kircher-Eibl, B. (2005).Dendritic cells from patients with chronic myeloid leukemia:functional and phenotypic features. Leukemia & Lymphoma, 46,663–670.
131. Thomachot, M. C., Bendriss-Vermare, N., Massacrier, C., Biota,C., Treilleux, I., Goddard, S., et al. (2004). Breast carcinomacells promote the differentiation of CD34+ progenitors towards 2different subpopulations of dendritic cells with CD1a(high)CD86(−)Langerin- and CD1a(+)CD86(+)Langerin+ phenotypes. Inter-national Journal of Cancer, 110, 710–720.
132. Fleming, M. D., Pinkus, J. L., Fournier, M. V., Alexander, S. W.,Tam, C., Loda, M., et al. (2003). Coincident expression of thechemokine receptors CCR6 and CCR7 by pathologic Langerhanscells in Langerhans cell histiocytosis. Blood, 101, 2473–2475.
133. Takayama, T., Morelli, A. E., Onai, N., Hirao, M., Matsushima,K., Tahara, H., et al. (2001). Mammalian and viral IL-10 enhanceC-C chemokine receptor 5 but down-regulate C-C chemokinereceptor 7 expression by myeloid dendritic cells: impact onchemotactic responses and in vivo homing ability. Journal ofImmunology, 166, 7136–7143.
134. Bonecchi, R., Facchetti, F., Dusi, S., Luini, W., Lissandrini, D.,Simmelink, M., et al. (2000). Induction of functional IL-8 receptors by IL-4 and IL-13 in human monocytes. Journal ofImmunology, 164, 3862–3869.
352 Cancer Metastasis Rev (2006) 25:333–356

135. Ogata, M., Zhang, Y., Wang, Y., Itakura, M., Zhang, Y. Y.,Harada, A., et al. (1999). Chemotactic response toward chemo-kines and its regulation by transforming growth factor-beta1 ofmurine bone marrow hematopoietic progenitor cell-deriveddifferent subset of dendritic cells. Blood, 93, 3225–3232.
136. Kobie, J. J., Wu, R. S., Kurt, R. A., Lou, S., Adelman, M. K.,Whitesell, L. J., et al. (2003). Transforming growth factor betainhibits the antigen-presenting functions and antitumor activityof dendritic cell vaccines. Cancer Research, 63, 1860–1864.
137. Sato, K., Kawasaki, H., Nagayama, H., Enomoto, M., Morimoto,C., Tadokoro, K., et al. (2000). TGF-beta 1 reciprocally controlschemotaxis of human peripheral blood monocyte-derived den-dritic cells via chemokine receptors. Journal of Immunology,164, 2285–2295.
138. Palucka, K. A., Taquet, N., Sanchez-Chapuis, F., & Gluckman, J.C. (1998). Dendritic cells as the terminal stage of monocytedifferentiation. Journal of Immunology, 160, 4587–4595.
139. Troy, A., Davidson, P., Atkinson, C., & Hart, D. (1998)Phenotypic characterisation of the dendritic cell infiltrate inprostate cancer. Journal of Urology, 160, 214–219.
140. Troy, A., Davidson, P., Atkinson, C., & Hart, D. (1999). Renalcell carcinoma and prostate cancer inhibit dendritic cell activa-tion. Australian and New Zealand Journal of Surgery, 69, A111–A112.
141. Ciavarra, R. P., Holterman, D. A., Brown, R. R., Mangiotti, P.,Yousefieh, N., Wright, G. L. Jr., et al. (2004). Prostate tumormicroenvironment alters immune cells and prevents long-termsurvival in an orthotopic mouse model following flt3-ligand/CD40-ligand immunotherapy. Journal of Immunotherapy, 27,13–26.
142. Chaux, P., Favre, N., Martin, M., & Martin, F. (1997). Tumor-infiltrating dendritic cells are defective in their antigen-present-ing function and inducible B7 expression in rats. InternationalJournal of Cancer, 72, 619–624.
143. Harding, F. A., McArthur, J. G., Gross, J. A., Raulet, D. H., &Allison, J. P. (1992). CD28-mediated signalling co-stimulatesmurine T cells and prevents induction of anergy in T-cell clones.Nature, 356, 607–609.
144. Reiser, H., Freeman, G. J., Razi-Wolf, Z., Gimmi, C. D.,Benacerraf, B., & Nadler, L. M. (1992). Murine B7 antigenprovides an efficient costimulatory signal for activation ofmurine T lymphocytes via the T-cell receptor/CD3 complex.Proceedings of the National Academy of Sciences of the UnitedStates of America, 89, 271–275.
145. Nestle, F. O., Burg, G., Fah, J., Wrone-Smith, T., & Nickoloff, B.J. (1997). Human sunlight-induced basal-cell-carcinoma-associ-ated dendritic cells are deficient in T cell co-stimulatorymolecules and are impaired as antigen-presenting cells. Ameri-can Journal of Pathology, 150, 641–651.
146. Ratta, M., Fagnoni, F., Curti, A., Vescovini, R., Sansoni, P.,Oliviero, B., et al. (2002). Dendritic cells are functionallydefective in multiple myeloma: the role of interleukin-6. Blood,100, 230–237.
147. Orsini, E., Guarini, A., Chiaretti, S., Mauro, F. R., & Foa, R.(2003). The circulating dendritic cell compartmen in patientswith chronic lymphocytic leukemia is severely defective andunable to stimulate an effective T-cell response. CancerResearch, 63, 4497–4506.
148. Tas, M., Simons, P., Balm, F., & Drexhage, H. (1993). Depressedmonocyte polarization and clustering of dendritic cells in patientswith head and neck cancer: in vitro restoration of thisimmunosuppression by thymic hormones. Cancer Immunologyand Immunotherapy, 36, 108–114.
149. Gottfried, E., Kunz-Schughart, L. A., Ebner, S., Mueller-Klieser,W., Hoves, S., Andreesen, R., et al. (2005). Tumor-derived lactic
acid modulates dendritic cell activation and antigen expression.Blood.
150. Thurnher, M., Radmayar, C., Ramoner, R., Ebner, S., Bock, G.,Klocker, H., et al. (1996). Human renal-cell carcinoma tissuecontains dendritic cells. International Journal of Cancer, 67, 1–7.
151. Aalamian, M., Pirtskhalaishvili, G., Nunez, A., Esche, C.,Shurin, G. V., Huland, E., et al. (2001). Human prostate cancerregulates generation and maturation of monocyte-derived den-dritic cells. Prostate, 46, 68–75.
152. Katsenelson, N. S., Shurin, G. V., Bykovskaia, S. N., Shogan, J.,& Shurin, M. R. (2001). Human small cell lung carcinoma andcarcinoid tumor regulate dendritic cell maturation and function.Modern Pathology, 14, 40–45.
153. Makarenkova, V. P., Shurin, G. V., Tourkova, I. L., Balkir, L.,Pirtskhalaishvili, G., Perez, L., et al. (2003). Lung cancer-derivedbombesin-like peptides down-regulate the generation and func-tion of human dendritic cells. Journal of Neuroimmunology, 145,55–67.
154. Shurin, G. V., Aalamian, M., Pirtskhalaishvili, G., Bykovskaia,S., Huland, E., Huland, H., et al. (2001). Shurin MR, Humanprostate cancer blocks the generation of dendritic cells from cd34+ hematopoietic progenitors. European Urology, 39 Suppl 4, 37–40.
155. Shurin, G. V., Shurin, M. R., Bykovskaia, S., Shogan, J., Lotze,M. T., & Barksdale, E. M. Jr. (2001). Neuroblastoma-derivedgangliosides inhibit dendritic cell generation and function.Cancer Research, 61, 363–369.
156. Shurin, M. R. (1999). Regulation of dendropoiesis in cancer.Clinical Immunology Newsletter, 19, 135–139.
157. Song, E. Y., Shurin, M. R., Tourkova, I. L., Chatta, G., & Shurin,G. V. (2004). Human renal cell carcinoma inhibits dendritic cellmaturation and functions. Urologe A, 43 Suppl 3, 128–130.
158. Kiertscher, S. M., Luo, J., Dubinett, S. M., & Roth, M. D.(2000). Tumors promote altered maturation and early apoptosisof monocyte-derived dendritic cells. Journal of Immunology,164, 1269–1276.
159. Lee, W. C., Chiang, Y. J., Wang, H. C., Wang, M. R., Lia, S. R.,& Chen, M. F. (2004). Functional impairment of dendritic cellscaused by murine hepatocellular carcinoma. Journal of ClinicalImmunology, 24, 145–154.
160. Pinzon-Charry, A., Maxwell, T., & Lopez, J. A. (2005).Dendritic cell dysfunction in cancer: a mechanism for immuno-suppression. Immunology and Cell Biology, 83, 451–461.
161. Saito, H., Tsujitani, S., Ikeguchi, M., Maeta, M., & Kaibara, N.(1998). Relationship between the expression of vascular endo-thelial growth factor and the density of dendritic cells in gastricadenocarcinoma tissue. British Journal of Cancer, 78, 1573–1577.
162. Almand, B., Resser, J. R., Lindman, B., Nadaf, S., Clark, J. I.,Kwon, E. D., et al. (2000). Clinical significance of defectivedendritic cell differentiation in cancer. Clinical Cancer Research,6, 1755–1766.
163. Menetrier-Caux, C., Montmain, G., Dieu, M. C., Bain, C.,Favrot, M. C., Caux, C., et al. (1998). Inhibition of thedifferentiation of dendritic cells from CD34(+) progenitors bytumor cells: role of interleukin-6 and macrophage colony-stimulating factor. Blood, 92, 4778–4791.
164. Stampfer, M. R., Yaswen, P., Alhadeff, M., & Hosoda, J. (1993).TGF beta induction of extracellular matrix associated proteins innormal and transformed human mammary epithelial cells inculture is independent of growth effects. Journal of CellularPhysiology, 155, 210–221.
165. Rodeck, U., Bossler, A., Graeven, U., Fox, F. E., Nowell, P. C.,Knabbe, C., et al. (1994). Transforming growth factor beta
Cancer Metastasis Rev (2006) 25:333–356 353

production and responsiveness in normal human melanocytesand melanoma cells. Cancer Research, 54, 575–581.
166. Fischer, J. R., Darjes, H., Lahm, H., Schindel, M., Drings, P., &Krammer, P. H. (1994). Constitutive secretion of bioactivetransforming growth factor beta 1 by small cell lung cancer celllines. European Journal of Cancer, 30A, 2125–2129.
167. Huang, M., Sharma, S., Mao, J. T., & Dubinett, S. M. (1996).Non-small cell lung cancer-derived soluble mediators andprostaglandin E2 enhance peripheral blood lymphocyte IL-10transcription and protein production. Journal of Immunology,157, 5512–5520.
168. Sharma, S., Stolina, M., Lin, Y., Gardner, B., Miller, P. W.,Kronenberg, M., et al. (1999). T cell-derived IL-10 promoteslung cancer growth by suppressing both T cell and APCfunction. Journal of Immunology, 163, 5020–5028.
169. Qin, Z., Noffz, G., Mohaupt, M., & Blankenstein, T. (1997).Interleukin-10 prevents dendritic cell accumulation and vaccina-tion with granulocyte-macrophage colony-stimulating factorgene-modified tumor cells. Journal of Immunology, 159, 770–776.
170. De Smedt, T., Van Mechelen, M., De Becker, G., Urbain, J., Leo,O., & Moser, M. (1997). Effect of interleukin-10 on dendriticcell maturation and function. European Journal of Immunology,27, 1229–1235.
171. Steinbrink, K., Wolfl, M., Jonuleit, H., Knop, J., & Enk, A. H.(1997). Induction of tolerance by IL-10-treated dendritic cells.Journal of Immunology, 159, 4772–4780.
172. Ludewig, B., Graf, D., Gelderblom, H. R., Becker, Y., Kroczek,R. A., & Pauli, G. (1995). Spontaneous apoptosis of dendriticcells is efficiently inhibited by TRAP (CD40-ligand) and TNF-alpha, but strongly enhanced by interleukin-10. EuropeanJournal of Immunology, 25, 1943–1950.
173. Carbone, E., Terrazzano, G., Ruggiero, G., Zanzi, D., Ottaiano,A., Manzo, C., et al. (1999). Recognition of autologous dendriticcells by human NK cells. European Journal of Immunology, 29,4022–4029.
174. Tourkova, I. L., Shurin, G. V., Chatta, G. S., Perez, L., Finke, J.,Whiteside, T. L., et al. (2005). Restoration by IL-15 of MHCclass I antigen processing machinery in human dendritic cellsinhibited by tumor-derived gangliosides. Journal of Immunology,175, 3045–3052.
175. Curiel, T. J., Wei, S., Dong, H., Alvarez, X., Cheng, P., Mottram,P., et al (2003). Blockade of B7-H1 improves myeloid dendriticcell-mediated antitumor immunity. Natural Medicines, 9, 562–567.
176. Dong, H., Zhu, G., Tamada, K., & Chen, L. (1999). B7-H1, athird member of the B7 family, co-stimulates T-cell proliferationand interleukin-10 secretion. Natural Medicines, 5, 1365–1369.
177. Freeman, G. J., Long, A. J., Iwai, Y., Bourque, K., Chernova, T.,Nishimura, H., et al. (2000). Engagement of the PD-1 immu-noinhibitory receptor by a novel B7 family member leads tonegative regulation of lymphocyte activation. Journal of Exper-imental Medicine, 192, 1027–1034.
178. Dong, H., Strome, S. E., Salomao, D. R., Tamura, H., Hirano, F.,Flies, D. B., et al. (2002). Tumor-associated B7-H1 promotes T-cell apoptosis: a potential mechanism of immune evasion.Natural Medicines, 8, 793–800.
179. Munn, D. H., Sharma, M. D., & Mellor, A. L. (2004). Ligationof B7-1/B7-2 by human CD4+ T cells triggers indoleamine 2,3-dioxygenase activity in dendritic cells. Journal of Immunology,172, 4100–4110.
180. Munn, D. H., Sharma, M. D., Hou, D., Baban, B., Lee, J. R.,Antonia, S. J., et al. (2004). Expression of indoleamine 2,3-dioxygenase by plasmacytoid dendritic cells in tumor-draininglymph nodes. Journal of Clinical Investigation, 114, 280–290.
181. Wei, S., Kryczek, I., Zou, L., Daniel, B., Cheng, P., Mottram, P.,et al. (2005). Plasmacytoid dendritic cells induce CD8+regulatory T cells in human ovarian carcinoma. Cancer Re-search, 65, 5020–5026.
182. Kaisho, T., & Akira, S. (2003). Regulation of dendritic cellfunction through toll-like receptors. Current Molecular Medi-cine, 3, 759–771.
183. Asselin-Paturel, C., Brizard, G., Chemin, K., Boonstra, A., O'Garra,A., Vicari, A., et al. (2005). Trinchieri G, Type I interferondependence of plasmacytoid dendritic cell activation and migration.Journal of Experimental Medicine, 201, 1157–1167.
184. Curiel, T. J., Cheng, P., Mottram, P., Alvarez, X., Moons, L.,Evdemon-Hogan, M., et al. (2004). Dendritic cell subsetsdifferentially regulate angiogenesis in human ovarian cancer.Cancer Research, 64, 5535–5538.
185. Conejo-Garcia, J. R., Benencia, F., Courreges, M. C., Kang, E.,Mohamed-Hadley, A., Buckanovich, R. J., et al. (2004). Tumor-infiltrating dendritic cell precursors recruited by a beta-defensincontribute to vasculogenesis under the influence of Vegf-A.Natural Medicines, 10, 950–958.
186. Chen, X., Doffek, K., Sugg, S. L., & Shilyansky, J. (2003).Neuroblastoma cells inhibit the immunostimulatory function ofdendritic cells. Journal of Pediatric Surgery, 38, 901–905.
187. Esche, C., Lokshin, A., Shurin, G. V., Gastman, B. R.,Rabinowich, H., Watkins, S. C., et al. (1999). Tumor's otherimmune targets: dendritic cells. Journal of Leukocyte Biology,66, 336–344 (In Process Citation).
188. Shurin, M. R., Esche, C., Lokshin, A., & Lotze, M. T. (1999).Apoptosis in Dendritic Cells, in M. T. Lotze & A. W. Thomson(Eds.), Dendritic Cells: Biology and Clinical Applications (p.673–692). San Diego: Academic.
189. Peguet-Navarro, J., Sportouch, M., Popa, I., Berthier, O.,Schmitt, D., & Portoukalian, J. (2003). Gangliosides fromhuman melanoma tumors impair dendritic cell differentiationfrom monocytes and induce their apoptosis. Journal of Immu-nology, 170, 3488–3494.
190. Lissoni, P., Malugani, F., Bonfanti, A., Bucovec, R., Secondino,S., Brivio, F., et al. (2001). Abnormally enhanced bloodconcentrations of vascular endothelial growth factor (VEGF) inmetastatic cancer patients and their relation to circulatingdendritic cells, IL-12 and endothelin-1. Journal of BiologicalRegulators and Homeostatic Agents, 15, 140–144.
191. Esche, C., Shurin, G. V., Kirkwood, J. M., Wang, G. Q.,Rabinowich, H., Pirtskhalaishvili, G., et al. (2001). Tumornecrosis factor-alpha-promoted expression of Bcl-2 and inhibi-tion of mitochondrial cytochrome c release mediate resistance ofmature dendritic cells to melanoma-induced apoptosis. ClinicalCancer Research, 7, 974s–979s.
192. Pirtskhalaishvili, G., Shurin, G. V., Esche, C., Trump, D. L., &Shurin, M. R. (2001). TNF-alpha protects dendritic cells fromprostate cancer-induced apoptosis. Prostate Cancer ProstaticDisorder, 4, 221–227.
193. Balkir, L., Tourkova, I. L., Makarenkova, V. P., Shurin, G. V.,Robbins, P. D., Yin, X. M., et al. (2004). Comparative analysis ofdendritic cells transduced with different anti-apoptotic mole-cules: sensitivity to tumor-induced apoptosis. Journal of GeneMedicine, 6, 537–544.
194. Kanto, T., Kalinski, P., Hunter, O. C., Lotze, M. T., & Amoscato,A. A. (2001). Ceramide mediates tumor-induced dendritic cellapoptosis. Journal of Immunology, 167, 3773–3784.
195. Shurin, M. R., Esche, C., & Lotze, M. T. (1998) FLT3: receptorand ligand. Biology and potential clinical application. Cytokine& Growth Factor Reviews, 9, 37–48 (In Process Citation).
196. Whartenby, K. A., Calabresi, P. A., McCadden, E., Nguyen, B.,Kardian, D., Wang, T., et al. (2005). Inhibition of FLT3 signaling
354 Cancer Metastasis Rev (2006) 25:333–356

targets DCs to ameliorate autoimmune disease. Proceedings ofthe National Academy of Sciences of the United States ofAmerica, 102, 16741–16746.
197. Pirtskhalaishvili, G., Gambotto, A., Esche, C., Yurkovetsky, Z.R., & Lotze, M.R.M.R. S. (2000). IL-12 and Bcl-xL genetransfection of murine dendritic cells protects them fromprostate-cancer induced apoptosis and improves their antitumoractivity. in AUA 95th Annual Meeting, April 29–May 4. Atlanta,Georgia: Journal of Urology.
198. Tourkova, I. L., Yurkovetsky, Z. R., Gambotto, A., Makaren-kova, V. P., Perez, L., Balkir, L., et al. (2002). Increased functionand survival of IL-15-transduced human dendritic cells aremediated by up-regulation of IL-15Ralpha and Bcl-2. Journalof Leukocyte Biology, 72, 1037–1045.
199. Zou, W., Machelon, V., Coulomb-L'Hermin, A., Borvak, J.,Nome, F., Isaeva, T., et al. (2001). Stromal-derived factor-1 inhuman tumors recruits and alters the function of plasmacytoidprecursor dendritic cells. Natural Medicines, 7, 1339–1346.
200. Ito, R., Kitadai, Y., Kyo, E., Yokozaki, H., Yasui, W., Yamashita,U., et al. (1993). Interleukin 1 alpha acts as an autocrine growthstimulator for human gastric carcinoma cells. Cancer Research,53, 4102–4106.
201. Abdul, M., & Hoosein, N. (2002). Relationship of the interleu-kin-1 system with neuroendocrine and exocrine markers inhuman colon cancer cell lines. Cytokine, 18, 86–91.
202. Satomi, H., Wang, B., Fujisawa, H., & Otsuka, F. (2002).Interferon-beta from melanoma cells suppresses the prolifera-tions of melanoma cells in an autocrine manner. Cytokine, 18,108–115.
203. Han, Y. P., Downey, S., & Garner, W. L. (2005). Interleukin-1alpha-induced proteolytic activation of metalloproteinase-9 byhuman skin. Surgery, 138, 932–939.
204. Song, X., Voronov, E., Dvorkin, T., Fima, E., Cagnano, E.,Benharroch, D., et al. (2003). Differential effects of IL-1 alphaand IL-1 beta on tumorigenicity patterns and invasiveness.Journal of Immunology, 171, 6448–6456.
205. Voronov, E., Shouval, D. S., Krelin, Y., Cagnano, E., Benhar-roch, D., Iwakura, Y., et al. (2003). IL-1 is required for tumorinvasiveness and angiogenesis. Proceedings of the NationalAcademy of Sciences of the United States of America, 100,2645–2650.
206. Tartour, E., Fossiez, F., Joyeux, I., Galinha, A., Gey, A., Claret,E., et al. (1999). Interleukin 17, a T-cell-derived cytokine,promotes tumorigenicity of human cervical tumors in nude mice.Cancer Research, 59, 3698–3704.
207. Alberti, L., Thomachot, M. C., Bachelot, T., Menetrier-Caux, C.,Puisieux, I., & Blay, J. Y. (2004). IL-6 as an intracrine growthfactor for renal carcinoma cell lines. International Journal ofCancer, 111, 653–661.
208. Lu, C., & Kerbel. R. S. (1993). Interleukin-6 undergoestransition from paracrine growth inhibitor to autocrine stimulatorduring human melanoma progression. Journal of Cell Biology,120, 1281–1288.
209. Portier, M., Zhang, X. G., Caron, E., Lu, Z. Y., Bataille, R., &Klein, B. (1993). Gamma-interferon in multiple myeloma:inhibition of interleukin-6 (IL-6)-dependent myeloma cellgrowth and downregulation of IL-6-receptor expression in vitro.Blood, 81, 3076–3082.
210. Guise, T. A., Kozlow, W. M., Heras-Herzig, A., Padalecki, S. S.,Yin, J. J., & Chirgwin, J. M. (2005). Molecular mechanisms ofbreast cancer metastases to bone. Clin Breast Cancer, 5 Suppl,S46–S53.
211. Csiszar, A., Szentes, T., Haraszti, B., Zou, W., Emilie, D.,Petranyi, G., et al. (2001). Characterisation of cytokine mRNAexpression in tumour-infiltrating mononuclear cells and tumour
cells freshly isolated from human colorectal carcinomas. Euro-pean Cytokine Network, 12, 87–96.
212. Howlett, M., Judd, L. M., Jenkins, B., La Gruta, N. L., Grail, D.,Ernst, M., et al. (2005). Differential regulation of gastric tumorgrowth by cytokines that signal exclusively through thecoreceptor gp130. Gastroenterology, 129, 1005–1018.
213. Moore, R. J., Owens, D. M., Stamp, G., Arnott, C., Burke, F.,East, N., et al. (1999). Mice deficient in tumor necrosis factor-alpha are resistant to skin carcinogenesis. Natural Medicines, 5,828–831.
214. Szlosarek, P. W., & Balkwill, F. R. (2003). Tumour necrosisfactor alpha: a potential target for the therapy of solid tumours.Lancet Oncology, 4, 565–573.
215. Bachelder, R. E., Wendt, M. A., & Mercurio, A. M. (2002).Vascular endothelial growth factor promotes breast carcinomainvasion in an autocrine manner by regulating the chemokinereceptor CXCR4. Cancer Research, 62, 7203–7206.
216. Toi, M., Kondo, S., Suzuki, H., Yamamoto, Y., Inada, K.,Imazawa, T., et al. (1996). Quantitative analysis of vascularendothelial growth factor in primary breast cancer. Cancer, 77,1101–1106.
217. Brown, L. F., Berse, B., Jackman, R. W., Tognazzi, K., Manseau,E. J., Dvorak, H. F., et al. (1993). Increased expression ofvascular permeability factor (vascular endothelial growth factor)and its receptors in kidney and bladder carcinomas. AmericanJournal of Pathology, 143, 1255–1262.
218. Senger, D. R., Van de Water, L., Brown, L. F., Nagy, J. A., Yeo,K. T., Yeo, T. K., et al. (1993). Vascular permeability factor(VPF, VEGF) in tumor biology. Cancer Metastasis Review, 12,303–324.
219. Takahashi, Y., Kitadai, Y., Bucana, C. D., Cleary, K. R., & Ellis,L. M. (1995). Expression of vascular endothelial growth factorand its receptor, KDR, correlates with vascularity, metastasis,and proliferation of human colon cancer. Cancer Research, 55,3964–3968.
220. Wislez, M., Rabbe, N., Marchal, J., Milleron, B., Crestani, B.,Mayaud, C., et al. (2003). Hepatocyte growth factor productionby neutrophils infiltrating bronchioloalveolar subtype pulmonaryadenocarcinoma: role in tumor progression and death. CancerResearch, 63, 1405–1412.
221. Siegfried, J. M., Weissfeld, L. A., Singh-Kaw, P., Weyant, R. J.,Testa, J. R., & Landreneau, R. J. (1997). Association ofimmunoreactive hepatocyte growth factor with poor survivalin resectable non-small cell lung cancer. Cancer Research, 57,433–439.
222. Ho, R., Minturn, J. E., Hishiki, T., Zhao, H., Wang, Q., Cnaan,A., et al. (2005). Proliferation of human neuroblastomasmediated by the epidermal growth factor receptor. CancerResearch, 65, 9868–9875.
223. Herbst, R. S. (2004). Review of epidermal growth factor receptorbiology. International Journal of Radiation Oncology, Biology,Physics, 59, 21–26.
224. Janmaat, M. L., & Giaccone, G. (2003). The epidermal growthfactor receptor pathway and its inhibition as anticancer therapy.Drugs Today (Barc), 39, Suppl C, 61–80.
225. Ren, D. H., Mayhew, E., Hay, C., Li, H., Alizadeh, H., &Niederkorn, J. Y. (2004). Uveal melanoma expression of tumornecrosis factor-related apoptosis-inducing ligand (TRAIL) recep-tors and susceptibility to TRAIL-induced apoptosis. InvestOphthalmol Vis Sci, 45, 1162–1168.
226. Wu, S., Boyer, C. M.,Whitaker, R. S., Berchuck, A., Wiener, J. R.,Weinberg, J. B., et al. (1993). Tumor necrosis factor alpha as anautocrine and paracrine growth factor for ovarian cancer: mono-kine induction of tumor cell proliferation and tumor necrosisfactor alpha expression. Cancer Research, 53, 1939–1944.
Cancer Metastasis Rev (2006) 25:333–356 355

227. Mooradian, D. L., Purchio, A. F., & Furcht, L. T. (1990).Differential effects of transforming growth factor beta 1 on thegrowth of poorly and highly metastatic murine melanoma cells.Cancer Research, 50, 273–277.
228. Lin, E. Y., Gouon-Evans, V., Nguyen, A. V., & Pollard, J. W.(2002). The macrophage growth factor CSF-1 in mammary glanddevelopment and tumor progression. Journal of MammaryGland Biology and Neoplasia, 7, 147–162.
229. Campbell, A. S., Albo, D., Kimsey, T. F., White, S. L., & Wang,T. N. (2005). Macrophage inflammatory protein-3alpha promotespancreatic cancer cell invasion. Journal of Surgical Research,123, 96–101.
230. Proost, P., De Wolf-Peeters, C., Conings, R., Opdenakker, G.,Billiau, A., & Van Damme, J. (1993). Identification of a novelgranulocyte chemotactic protein (GCP-2) from human tumorcells. In vitro and in vivo comparison with natural forms ofGRO, IP-10, and IL-8. Journal of Immunology, 150, 1000–1010.
231. Dhawan, P., & Richmond, A. (2002). Role of CXCL1 intumorigenesis of melanoma. Journal of Leukocyte Biology, 72,9–18.
232. Monti, P., Leone, B. E., Marchesi, F., Balzano, G., Zerbi, A.,Scaltrini, F., et al. (2003). The CC chemokine MCP-1/CCL2 inpancreatic cancer progression: regulation of expression andpotential mechanisms of antimalignant activity. Cancer Re-search, 63, 7451–7461.
233. Saji, H., Koike, M., Yamori, T., Saji, S., Seiki, M., Matsushima,K., et al. (2001). Significant correlation of monocyte chemo-attractant protein-1 expression with neovascularization andprogression of breast carcinoma. Cancer, 92, 1085–1091.
234. Ohta, M., Kitadai, Y., Tanaka, S., Yoshihara, M., Yasui, W.,Mukaida, N., et al. (2002). Monocyte chemoattractant protein-1expression correlates with macrophage infiltration and tumorvascularity in human esophageal squamous cell carcinomas.international journal of Cancer, 102, 220–224.
235. Mazzucchelli, L., Loetscher, P., Kappeler, A., Uguccioni, M.,Baggiolini, M., Laissue, J. A., et al. (1996). Monocyte chemo-attractant protein-1 gene expression in prostatic hyperplasia andprostate adenocarcinoma. American Journal of Pathology, 149,501–509.
236. Moran, C. J., Arenberg, D. A., Huang, C. C., Giordano, T. J.,Thomas, D. G., Misek, D. E., et al. (2002). RANTES expressionis a predictor of survival in stage I lung adenocarcinoma.Clinical Cancer Research, 8, 3803–3812.
237. Vaday, G. G., Peehl, D. M., Kadam, P. A., & Lawrence, D. M.(2005). Expression of CCL5 (RANTES) and CCR5 in prostatecancer. Prostate, 66, 124–134.
238. Singh, S., Singh, U. P., Stiles, J. K., Grizzle, W. E., & Lillard, J.W. Jr. (2004). Expression and functional role of CCR9 inprostate cancer cell migration and invasion. Clinical CancerResearch, 10, 8743–8750.
239. Zhou, Y., Larsen, P. H., Hao, C., & Yong, V. W. (2002). CXCR4is a major chemokine receptor on glioma cells and mediates theirsurvival. Journal of Biological Chemistry, 277, 49481–49487.
240. Scotton, C. J., Wilson, J. L., Scott, K., Stamp, G., Wilbanks, G.D., Fricker, S., et al. (2002). Multiple actions of the chemokineCXCL12 on epithelial tumor cells in human ovarian cancer.Cancer Research, 62, 5930–5938.
241. Sun, Y. X., Wang, J., Shelburne, C. E., Lopatin, D. E.,Chinnaiyan, A. M., Rubin, M. A., et al. (2003). Expression of
CXCR4 and CXCL12 (SDF-1) in human prostate cancers (PCa)in vivo. Journal of Cellular Biochemistry, 89, 462–473.
242. Balkwill, F. (2004). Cancer and the chemokine network. NatureReviews Cancer, 4, 540–550.
243. Singh, S., Singh, U. P., Grizzle, W. E., & Lillard, J. W. Jr. (2004).CXCL12–CXCR4 interactions modulate prostate cancer cellmigration, metalloproteinase expression and invasion. Laborato-ry Investigation, 84, 1666–1676.
244. Iizasa, H., Yoneyama, H., Mukaida, N., Katakoka, Y., Naito, M.,Yoshida, N., et al. (2005). Exacerbation of granuloma formationin IL-1 receptor antagonist-deficient mice with impaired dendrit-ic cell maturation associated with Th2 cytokine production.Journal of Immunology, 174, 3273–3280.
245. Wesa, A., & Galy, A. (2002). Increased production of pro-inflammatory cytokines and enhanced T cell responses afteractivation of human dendritic cells with IL-1 and CD40 ligand.BMC Immunol, 3, 14.
246. Park, S. J., Nakagawa, T., Kitamura, H., Atsumi, T., Kamon, H.,Sawa, S., et al. (2004). IL-6 regulates in vivo dendritic celldifferentiation through STAT3 activation. Journal of Immunolo-gy, 173, 3844–3854.
247. Hegde, S., Pahne, J., & Smola-Hess, S. (2004). Novel immuno-suppressive properties of interleukin-6 in dendritic cells: inhibi-tion of NF-kappaB binding activity and CCR7 expression.FASEB Journal, 18, 1439–1441.
248. Kim, J., Modlin, R. L., Moy, R. L., Dubinett, S. M., McHugh, T.,Nickoloff, B. J., et al. (1995). IL-10 production in cutaneousbasal and squamous cell carcinomas. A mechanism for evadingthe local T cell immune response. Journal of Immunology, 155,2240–2247.
249. Huang, M., Wang, J., Lee, P., Sharma, S., Mao, J. T., Meissner,H.,et al. (1995). Human non-small cell lung cancer cells express atype 2 cytokine pattern. Cancer Research, 55, 3847–3853.
250. Steinbrink, K., Jonuleit, H., Muller, G., Schuler, G., Knop, J.,& Enk, A. H. (1998). Interleukin-10-treated human dendriticcells induce a melanoma-antigen-specific anergy in CD8(+) Tcells resulting in a failure to lyse tumor cells. Blood, 93, 1634–1642.
251. Muller, G., Muller, A., Tuting, T., Steinbrink, K., Saloga, J.,Szalma, C., et al. (2002). Interleukin-10-treated dendritic cellsmodulate immune responses of naive and sensitized T cells invivo. Journal of Investigative Dermatology, 119, 836–841.
252. Gabrilovich, D. I., Corak, J., Ciernik, I. F., Kavanaugh, D., &Carbone, D. P. (1997). Decreased antigen presentation bydendritic cells in patients with breast cancer. Clinical CancerResearch, 3, 483–490.
253. Takahashi, A., Kono, K., Ichihara, F., Sugai, H., Fujii, H., &Matsumoto, Y. (2004). Vascular endothelial growth factorinhibits maturation of dendritic cells induced by lipopolysaccha-ride, but not by proinflammatory cytokines. Cancer Immunologyand Immunotherapy, 53, 543–550.
254. Okunishi, K., Dohi, M., Nakagome, K., Tanaka, R., Mizuno, S.,Matsumoto, K., et al. (2005). A novel role of hepatocyte growthfactor as an immune regulator through suppressing dendritic cellfunction. Journal of Immunology, 175, 4745–4753.
255. Li, G., Kim, Y. J., & Broxmeyer, H. E. (2005). Macrophagecolony-stimulating factor drives cord blood monocyte differen-tiation into IL-10(high)IL-12absent dendritic cells with tolero-genic potential. Journal of Immunology, 174, 4706–4717.
356 Cancer Metastasis Rev (2006) 25:333–356





























