Embed Size (px)
Citation preview

AD-A249 035 ) l lCPROGRESS REPORT
For the Period Ending January 1, 1992VISUAL PERCEPTION OF DEPTH-FROM-OCCLUSION: A NEURAL NETWOR
MODELContract # N00014-90-J-1864 bI
Dp P.!. - Leif H. Finkel FCTDepartment of Bioengineering andInstitute of Neurological Sciences APR 0 11992
University of Pennsylvania
220 South 33rd Street SPhiladelphia, PA 19104
During this period, we have made substantial progress in understanding how objects arediscriminated by the visual system. A number of papers describing our results are enclosed.Major goals of the project have been accomplished in several areas:
5 Completion of the beta release version of a novel neural network simulator, NEXUS
) Development of a parallel version of the NEXUS simulator which allows anatomicallyinterconnected networks to be simultaneously simulated on different workstations linkedby ethernet connections'
3 Continuing research using our model of how the visual system extracts depth-from-occlusion.Considerable progress with regard to how surfaces may be represented. Numerous simula-tions of resp,,..ses to both real and illusory objectsj A N-)
4 Development of related cortically-based models of color visual processing and texture dis-crimination. Color and texture are used to detect contours in images, and these contourscan then be used by our depth-from-occlusion model to determine the relative depth of thecolored or textured surfaces.
The N XUS Neural Simulator
We have now completed the beta release version of the NEXUS simulator. NEXUS is an in-teractive simulator which allows the user to construct large-scale (10 neurons, 106 connections)neural networks. The simulator supports both biologically-based and artificial network architec-tures. In addition, NEXUS allows the user to create hybrid neural systems in which differenttypes of networks are interconnected. Thus, for example, one can create a biologically-basedearly vision system which feeds into a PDP object recognition network.
NEXUS is currently being tested in several research laboratories at Penn. We are also testingit as a tool in a graduate course in Computational Neuroscience (enrollment of 38 students). Thestudents use NEXUS to carry out eight simulations. The simulations include, for example, aPDP model of classifying tree leaves as to their species (oak, maple, beech, etc.), a model for theformation of ocular dominance columns, and analysis of the Adelson-Bergen Energy models ofmotion detection and texture discrimination. This summer, we will use NEXUS to run a short-course on neural simulation at the McDonnell Summer Institute for Cognitive Neuroscience atDartmouth.
After these tests of the simulator are complete we plan to make NEXUS available to allinterested users.
We have also completed a preliminary version of pNEXUS-a parallel version of NEXUS whichallows interconnected networks to be simulated on different machines linked by ethernet connec-tions. The program allows NEXUS to be simultaneously run on a number of workstations, with
...... .... ........... . . . . . .. 9 - 7 29 2~ i' 3 2 I 018 Il92-07825
92 327 018 IRIIIIlIIII'(

different networks defined on each machine. Information is shared between networks by meansof common access to stored files. In this way, very large multi-network models can be simulated.For example, a model of visual cortical function might include several interconnected modelsof shape, color, motion, and depth processing, each of which runs on a separate workstation.pNEXUS allows communication and integration between these modules.
Model of Depth-from-Occlusion
Our model of depth-from-occlusion addresses the problem of how the visual system candiscriminate objects in a complex scene. There are two fundamental problems at the coreof object discrimination: binding and segmentation. The binding problem addresses how theattributes of an object-shape, color, motion, depth-are linked to create an individ,!al object. Itcan also be applied to how the parts of an object are linked or bound together (e.g. why thehorse's tail is perceived as part of the horse). Segmentation deals with the converse problem ofhow attributes of separate objects are distinguished. The process of determining depth-from-occlusion is a paradigmatic example of how the visual cortex may carry out the binding andsegmentation processes. Namely, when two surfaces overlap, there is an ambiguity as to which ofthe surfaces "owns" the common border. Consider, for example, a tree branch crossing in front ofour view of the moon. Since the tree branch is, in fact, in front of the moon, the common borderbelongs to the branch. However, if the "half-moons" were actually two separate objects, thenthe common border would belong to them as well. The determination of which surface "owns"the border determines the occlusion relationship. The extraction of depth-from-occlusion thusprovides a simple but powerful paradigm for studying how objects are defined, discriminated,stratified, and linked.
Our present simulations consist of 10 interconnected networks, each of which contains oneor more topographically organized arrays of 64x64 units (the system contains a total of 1.7x105
units). This total includes both conventional neuronal units, and a new type of network unitcalled PGN (programmable generalized neural) units which execute arbitrary functions or algo-rithms. A single PGN unit can emulate the function of a small circuit or assembly of standardunits. PGN units are particularly useful in situations in which an intensive computation is beingperformed but the anatomical and physiological details of the how the operation is performedin vivo are unknown. Alternatively, PGN units can be used to carry out functions in a compu-tationally efficient manner; for example, to implement a one-step winner-take-all algorithm.
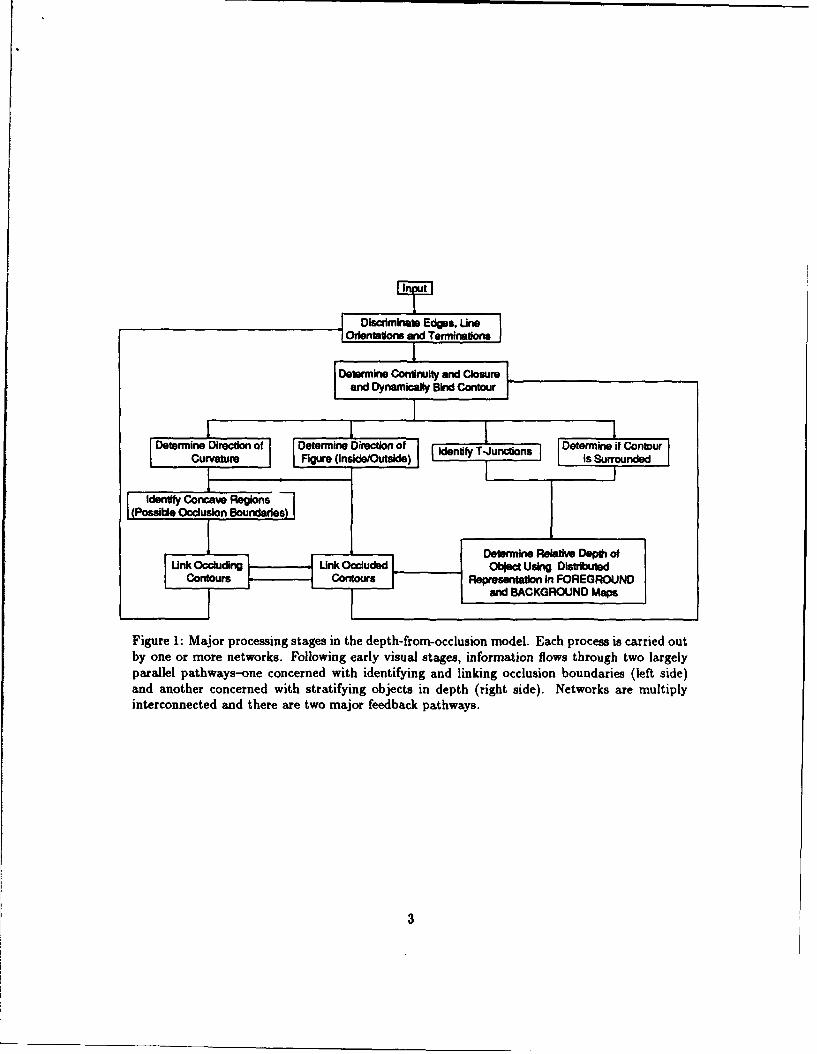
Figure 1 shows the major processes carried out by the network system. Early visual process-ing involves networks specialized for detecting edges, orientation, endstopping, curvature, andjunctions. All of these processes, and all of the processes discussed below, are carried out usingeither known or plausible neural architectures. After contour extraction, more global propertiesare determined, including the determination of closure and the discrimination of the inside-vs.-outside of a contour. For example, figure 2 illustrates the neural mechanism used to determinethe inside-vs.-outside of a contour. Orientation-tuned cells determine the local orientation of acontour. These orientation cells inhibit their nearest neighbors, except for cells located roughlyperpendicular to the local orientation of the contour (see figure 2). Units extend dendrites in astellate pattern, i.e., in all directions, and these dendrites are activated when they intersect thecontour. An important aspect of the mechanism is that each unit responding to the contour isbound with other such units by means of a local tag. This tag could be implemented in vivo byphase-locked firing of units[3], but the mechanism does not depend upon the biophysical imple-mentation of the tag. The tag serves the following function: if the stellate dendrites intersectother contours in the scene (i.e., those with tags which differ from the tag detected by the neigh- L_
boring orientation-tuned unit), these inputs do not activate the unit. The function of the circuitcan then be intuitively understood as follows. If a unit is inside the contour, all of its dendrites
Statement A per telecon ... .Dr. Harold Hawkins ONR/Code 1142 Dis vati'alorArlington, VA 22217-5000
NWW 3/31/92
i Discmalte Edges, U n fOriented"n and Terming;s
prlelpmm Continuity and Closure iand Dynamically Bind Contour id
iDermine Dircton of h Determine o irection pathways.CurvatureF
(Possible Occlusion Boundaes
I Deamine Relative Depth of
Unk Occluding UnkOccluded Obt Using DistributedContours I Contours Representation in FOREGROUND
l t and BACKGROUND Maps
Figure 1: Major processing stages in the depth-from-occlusion model. Each process is carried outby one or more networks. Following early visual stages, information flows through two largelyparallel pathways-one concerned with identifying and linking occlusion boundaries (left side)and another concerned with stratifying objects in depth (right side). Networks are multiplyinterconnected and there are two major feedback pathways.
3

conur bndg
drelction of figur
Figure 2: Neural circuit for determining direction of figure (inside vs. outside). Hypotheticalinput stimulus consists of two closed contours (bold curves). The central unit of 3x3 array(shown below) determines the local orientation of the contour. Surrounding units representpossible directions (indicated by arrows) of the inside of the figure relative to the contour. Allsurrounding units are inhibited (black circles) except for the two units located perpendicular tolocal orientation of the contour. Units receive inputs from the contour binding map via dendritesthat spread out in a stellate configuration, as indicated by clustered arrows (dendrites extend overlong distances in map). Units inside the figure will receive more inputs than those located outsidethe figure. The two uninhibited units compete using a winner-take-all architecture. Note thatinputs from separate objects are not confused due to the tags generated in the contour bindingmap.
will intersect the contour; if the unit is outside, then only some of its dendrites will intersectthe contour. A simple "winner-take-all" circuit can then pick the unit which is on the inside.Using this mechanism, we can determine which side of the contour is the inside in one cycle ofiteration (we use PGN cells to implement a one-step winner-take-all). Note that the problem ofinside/outside has traditionally been considered a difficult computational process, one requiringglobal information about the curve; yet using neural architectures, we have found a mechanismfor determining the solution in one cycle of network operation.
We have similarly used a number of other simple mechanisms, based on known or plausibleneural architectures to carry out all of the visual processes required by the model. The neuralmechanisms include:
" feedback connections from higher to lower cortical areas which serve to integrate visualperception
" a distributed representation of relative depth
* a new role for phase-locked cortical firing
* a neural mechanism for detecting T-junctions and for shuffling objects in relative depth
4

9 neural mechanism for linking objects across occlusion barriers
Details of network construction and extensive results are presented in the enclosed papers.We briefly summarize some of the major results below.
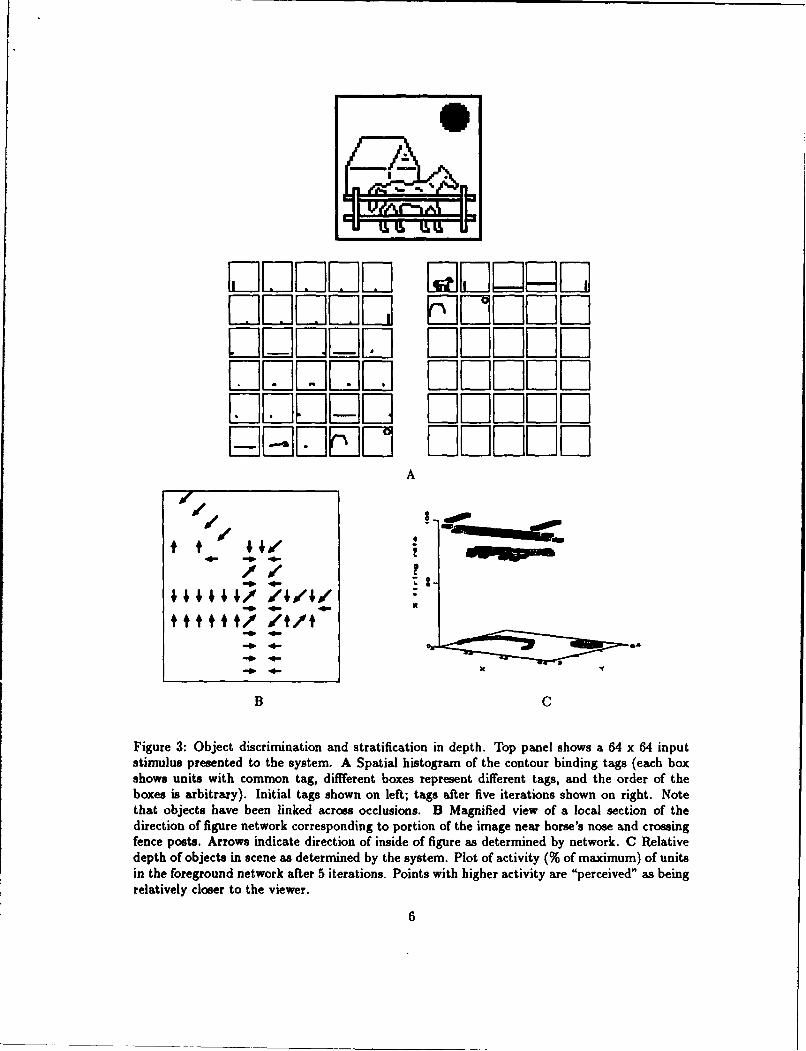
Figure 3 shows a typical visual scene presented to the system. The early networks discriminatethe edges, lines, terminations, and junctions present. Figure 3A displays how contours are boundin a visual scene. On the first cycle of activity, discontinuous segments of contours are boundseparately. These contours are later bound together as a result of feedback from the linkingprocesses.
Figure 3B shows the determination of inside-vs.-outside (we call this the "direction of figure")for a portion of the scene. The direction of the arrows indicates the direction of the "inside" asdetermined by the network.
The presence of T-junctions (e.g., between the horse and the fence) are used by the systemto force various objects into different depth planes. Results of this process are displayed in figure3C which plots the firing rate of units in the foreground network-this indicates the relative depthof the objects. The system has successfully stratified the fence, horse, house and sun.
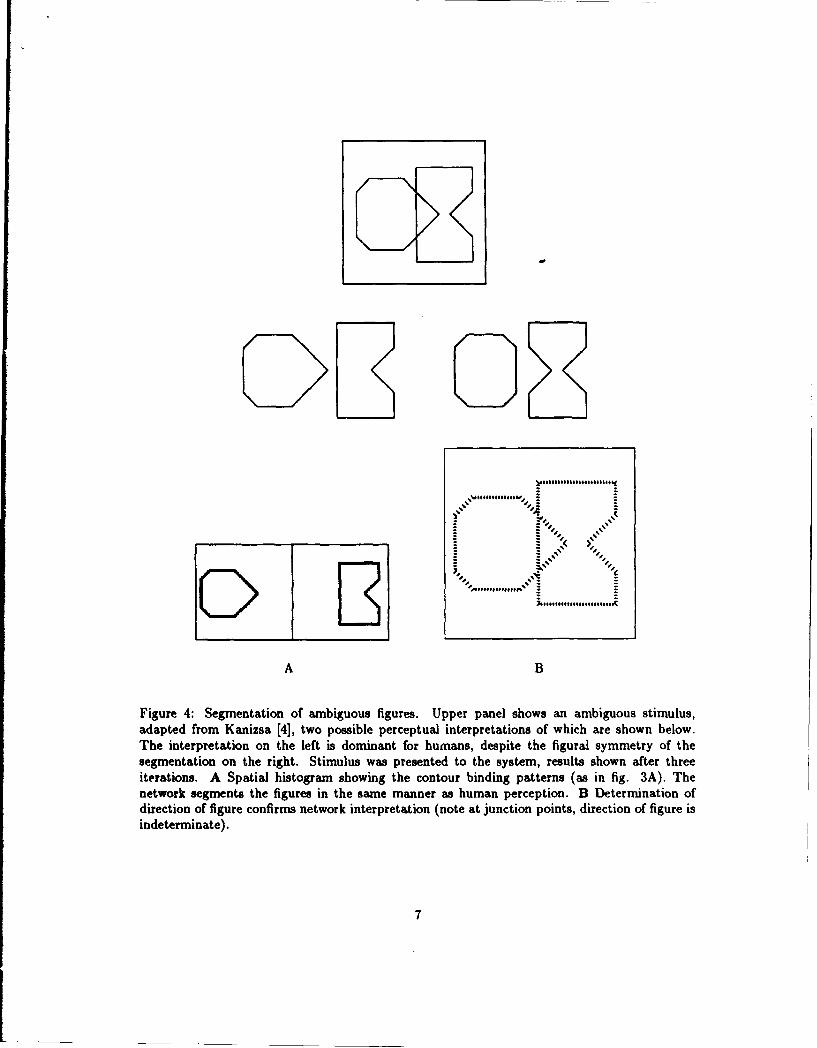
Figure 4 shows a stimulus, adapted from Kanizsa [4], in which there are two possible percep-tual interpretations (middle panels)-on the left, the two figures respect local continuity (this isthe dominant human perception); on the right, the figures respect global symmetry. Figure 4Ashows the contour binding tags, and figure 4B shows the direction of figure determined by thesystem. Both results indicate that the network makes the same perceptual interpretation as ahuman observer.
As this example illustrates, the model obeys several of the classical Gestalt laws of perceptualorganization. The results of figure 4 correspond to the law of "good continuation" [6], but othersimulations have demonstrated effects consistent with the general notion of Priignanz. It is thuspossible that the Gestalt laws arise from intrinsic neural constraints, for example, the anatomicconnection patterns in striate and extrastriate cortex. In our simulations, we used implicitconnectivity patterns based on the anatomical data of Rockland and Lund (9]. These connectionschemes were directly responsible for generating proximity and good continuation effects.
The final simulation is, again, adapted from Kanizsa [4], and shows a perceptually vivid,illusory white square in a field of black discs. The illusory square appears closer than thebackground, and the four black discs inside its borders appear even closer than the square. Thisis an example of what we call "occlusion capture", an effect related to Ramachandran's capturephenomenon. In this case, the illusory square has "captured" the discs within its borders andpulled them into the foreground.
Figure 5A shows the contour binding tags after one (left) and three (right) cycles of activity.Initially, each disc is bound separately. After several cycles, responses to the illusory square aregenerated and the square is given a common tag. Note that the edges of the discs occluded by theillusory square are now bound with the square, not with the discs. This change in "ownership"of the edges is the critical step in discriminating the illusory square as an object. For example,Figure 5B shows determination of the direction of figure after one and three cycles of activity.The change in which surface "owns" the edge is reflected by a change in the direction of "inside".
Figure 5C displays the firing rate of units in the foreground network (as in 30), thus showingthe relative depths discriminated by the system. The discs are placed in the background, theillusory square at an intermediate depth, and the discs located within the borders of the illusorysquare are located closest to the viewer. In this case, the depth cue which forces the internaldiscs to the foreground is not due to T-junctions, but rather to another network mechanism wecall "surround occlusion". Thus the system demonstrates occlusion capture corresponding tohuman perceptions of this stimulus.
5
IDDDD I __WMD
EI ---]rF-]-71 r-I I FIFEDIDDI DDZIILIZ9-Pl-II-I IE1-1-1II -1
A
-f N
B C
Figure 3: Object discrimination and stratification in depth. Top panel shows a 64 x 64 inputstimulus presented to the system. A Spatial histogram of the contour binding tags (each boxshows units with common tag, different boxes represent different tags, and the order of theboxes is arbitrary). Initial tags shown on left; tags after five iterations shown on right. Notethat objects have been linked across occlusions. B Magnified view of a local section of thedirection of figure network corresponding to portion of the image near horse's nose and crossingfence posts. Arrows indicate direction of inside of figure as determined by network. C Relativedepth of objects in scene as determined by the system. Plot of activity (% of maximum) of unitsin the foreground network after 5 iterations. Points with higher activity are "perceived" as beingrelatively closer to the viewer.
6
C 0
............. '* - S l *IH Q t tItI
A B
Figure 4: Segmentation of ambiguous figures. Upper panel shows an ambiguous stimulus,adapted from Kanizsa [4], two possible perceptual interpretations of which are shown below.The interpretation on the left is dominant for humans, despite the figural symmetry of thesegmentation on the right. Stimulus was presented to the system, results shown after threeiterations. A Spatial histogram showing the contour binding patterns (as in fig. 3A). Thenetwork segments the figures in the same manner as human perception. B Determination ofdirection of figure confirms network interpretation (note at junction points, direction of figure isindeterminate).
7

WR I1-IW--WEull. Ir I ll [-l -"l -I I E I:F 1
AD: 0 0, 0: 0
B ) CI'- , I I.'""'
-" ,,,, I , ,,
B C
Figure 5: Occlusion capture. Upper panel shows stimulus (adapted from Kanizsa [4]) in which weperceive a white illusory square. Note that the four black discs inside the illusory square appearcloser than the background. A 64 x 64 discrete version of stimulus wa- presented to the network.A Spatial histogram (as in fig. 2A) of the initial and final (after 3 iterations) contour bindingtags. Note that the illusory square is bound as an object. B Direction of figure determined bythe system. Insets show a magnified view of the initial (left) and final (right) direction of figure(region of magnification is indicated). Note that the direction of figure of the "mouth" of thepac-man flips once the illusory contour is generated. C Activity in the foreground network (% ofmaximum) demonstrates network stratification of objects in relative depth. The illusory squarehas "captured" the background texture. 8

There are a number of points of agreement between our model of depth-from-occlusion andrecent theories of visual perception by Nakayama [8] and Kellman and Shipley [5]. In fact, weprovide neural mechanisms which, when simulated, display many of the psychophysical effectsdescribed by these authors. Our model differs from previous neural models in that it discrimi-nates objects-not just contours. The difference is critical: a network which generates responsesto the three sides of the Kanizsa triangle, for example, is not representing a triangle (the ob-ject) per se. To represent the triangle it is necessary to link these three contours into a singleobject boundary, to represent the surface of the triangle (including surface properties such asdepth, color, texture, etc.), and finally to bind all these attributes into a whole. The proposedmodel demonstrates that one can build a self-contained system for discriminating objects basedon occlusion relationships. The model is successful at stratifying simple visual scenes, for link-ing the representations of occluded objects, and at generating responses to illusory objects in amanner consistent with human perceptual responses. The model uses neural circuits that arebiologically-based, and conforms to general neural principles, such as the use of a distributedrepresentation for depth. The system can be tested in psychophysical paradigms and the re-sults compared to human and animal results. In this manner, a computational model whichis designed based on physiological data and tested with psychophysical data offers a powerfulparadigm for bridging the gap between neuroscience and perception.
Extraction of Texture and Color Boundaries
In real images, there are multiple cues to object boundaries beyond the simple luminancegradients considered above. Two of the most useful such cues are discontinuities in color andtexture. Color and texture also serve as prime examples of "surface" properties which mustbe bound to an object. Proceeding towards our goal of discriminating complex objects in realimages, we have begun to develop neural models of both color and texture discrimination. Thecolor model is based on the anatomy and physiology of the color pathway from the retina toarea V4. It primarily addresses the problem of color constancy and color contrast.' Recentresults of Desimone and Schein [10] recording in macaque visual cortex have implicated the large"silent" surrounds of V4 neurons in this process. We have shown that such a mechanism canqualitatively account for a number of psychophysical resports in the literature [2] [11]. However,we have found it necessary to propose that V4 uses both positive an negative contrast signalsin each color channel in addition to opponent-channel responses (provided by type II cells inarea Vi). The contrast signals reflect the difference between the center and (large) surround ascalculated by V4 cells. (Negative contrast includes those cases when surround activity exceedsactivity in the center of the receptive field). We use a simple neural circuit in which the two"contrast" signals modulate the response of the direct feedfoward "opponent" signal. We canthen account for a wide range of both color and luminance contrast and constancy effects. Weare currently refining and testing the ability of the model to quantitatively account for humanpsychophysical results.
We have also recently begun to test several models of texture discrimination. We haveused NEXUS to reproduce recent models proposed by Adelson and Bergen [1] and by Malikand Peronna [7]. We have successfully used the outputs of these models (i.e., the locations ofdiscriminated texture boundaries) as inputs to our depth-from-occlusion model. In this case, thetextured regions are discriminated, the surfaces are linked to the boundaries, and the textured
I Color constancy refers to the fact that the perceived color of an object remains relatively invariant as thewavelength of the incident light is changed, i.e., an orange looks "orange" under sunlight, fluorescent light,incandescent light, etc. despite the fact that the amount of low and middle wavelength light reflected from theorange differs greatly in the different cases. Color contrast is the related effect in which the perceptual appearanceof a colored object changes when it is viewed against differently colored backgrounds, i.e., an orange looks morered on a yellow background than on a grey background.
9

surfaces are ordered in depth depending upon cues to occlusion. In the same manner, boundariesdiscriminated by other early vision systems could by fed into the occlusion model and used tobind surfaces and order them in depth. It is important to note, however, that color and textureare probably processed by separate systems from those concerned with occlusion. This implicitdifference (which may be relegated to the magnoceliular and parvocellular divisions of the visualprojection) is reflected in the parallel pathways of the depth-from-occlusion model (see figure 1).
Conclusions
We have made substantial progress in a number of areas-the NEXUS simulator, developmentof a theoretical model of occlusion that integrates anatomical, physiological and psychophysicaldata, testing of the model through extensive computer simulations on teal and illusory stimuli,and the development of related models in color and texture discrimination.
We plan to vigorously pursue these findings and to consider a number of additional topics.For example, we are currently testing the ability of the model to account for the perceptualvividness of different illusory contour configurations, the ability to account for the perception oftransparency, and extending the model to handle motion-related occlusion information as occursin the accretion and deletion of texture near a moving occlusion boundary. We are beginningto run preliminary tests of the pNEXUS program in which several different cortical areas aresimultaneously simulated, and interconnections between the areas mediate integration of functionin parallel. Finally, we are attempting to set up a facility for analyzing real video images so asto test the model on a more complex set of stimuli.
Enclosures
Papers:
. L.H. Finkel and P. Sajda (1992) Object Discrimination based on Depth-from-Occlusion,Neural Computation [in press]
2. P. Sajda and L.H. Finkel (1992) NEXUS: A simulation environment for large-scale neuralsystems. Simulation [in press]
3. P. Sajda and L.H. Finkel (1992) A neural network model of object segmentation andfeature bindiug in visual cortex. Proceedings of the Int. Joint Conf. on Neural Networks,(submitted]
4. K. Sakai, P. Sajda, and L.H. Finkel (1992) Texture discrimination and binding by a mod-ified energy model, Proceedings Int. Joint Conf. Neural Networks [submitted]
5. '-.M. Courtney, G. Buchsbaum, and L.H. Finkel (1992) Biologically-based neural networkmodel of color constancy and color contrast, Proceedings Int. Joint Conf. Neural Networks(submitted]
6. L.H. Finkel (1991) Neural Networks, in Encyclopedia of Human Biology, vol. 5, 387-392.
7. L.H. Finkel, G.N. Reeke, and G.M. Edelman (1992) A cortically based model for integrationin visual perception, in Neural Networks for Perception, vol. 1, H. Wechsler, ed., pp. 41-63,Academic Press, Boston.
8. L.H. Finkel and G.M. Edelman (1991) Models of somatotopic map organization, in Lecturesin Complex Systems, vol. III, L. Nadel and D. Stein, eds., pp. 269-284, Addison-Wesley,Redwood City, Ca.
10

*
Abstracts:
1. P. Sajda anu L.H. Finkel (1992) Extraction of depth-from-occlusion by a physiologicallybased network, ARVO Proceedings [in press]
2. S. ".. Courtney, G. Buchsbaum, and L.H. Finkel (1992) Color constancy and color contrastin a physiologically-based network model, ARVO Proceedings [in press]
Technical Reports:
1. P. Sajda, K. Sakai, and L.H. Finkel (1992) NEXUS User Manual, beta release-version 0.8,University of Pennsylvania Technical Report
References
[11 J. Bergen and E. Adelson. Early vision and texture perception. Nature, 333:33-364, 1988.
[2] K.T. Blackwell and G. Buchsbaum. Quantitative studies of color constancy. J. Optical Soc.America, 5:1772-1780, 1988.
[3] C. M. Gray and W. Singer. Neuronal oscillations in orientation columns of cat visual cortex.
Proceedings of the National Academy of Science USA, 86:1698-1702, 1989.
[4] G. Kanizsa. Organization in Vision. Praeger, New York, 1979.
[5] P.J. Kellman and T.F. Shipley. A theory of visual interpolation in object perception. Cog-nitive Psychology, 23:141-221, 1991.
[61 K. Koffka. Principles of Gestalt Psychology. Harcourt, Brace, New York, 1979.
[7] J. Malik and P. Petrona. Preattentive texture discrimination with early vision mechanisms.J. Optical Soc. America, 7:923-932, 1990.
(8] K. Nakayama and S. Shimojo. Toward a neural understanding of visual surface representa-tion. Cold Spring Harbor Symp. Quant. Biol., LV:911-924, 1990.
[9] K. S. Rockland and J. S. Lund. Widespread periodic intrinsic connections in the tree shrewvisual cortex. Science, 215:1532-1534, 1982.
[10] S.J. Schein and R. Desimone. Spectral properties of V4 neurons in the macaque. J. Neuro-science, 10:3370-3389, 1990.
[11] J. Walraven. Discounting the background: the missing link in the explanation of chromaticinduction. Vision Research, 16:289-295, 1976.
11





























