Embed Size (px)
Citation preview

Journal of Immunological Methods, 67 (1984) 157-165 157 Elsevier
JIM 02958
Isolation of Guinea Pig Basophils Using Anti-Leukocyte Antibody and Density Gradient
Centrifugation on Percoll 1,2
Frank M. Graziano 3,4, William Lipham, Neela Swaminathan, John Brendel, Frank M. Graziano, Jr. and Thomas Warner
Departments of Medicine and Pathology, University of Wisconsin Hospital, Madison, WI 53792, U.S.A.
(Received 31 August 1983, accepted 7 October 1983)
Peripheral blood basophils were isolated from guinea pigs using a 2-step procedure. Initial enrichment of basophils was achieved by treating leukocyte preparations obtained from whole blood with antibody to contaminating granulocytes and mononuclear cells. Basophil populations of 82% purity (mean; range 65-99%) were obtained by subsequent application of the antibody treated preparation to discontinuous Percoli gradients. The isolated basophils were viable and shown to be functional by histamine release upon stimulation with various secretogogues.
Key words: guinea p i g - basophils - cell separation - anti.leukocyte antibody - Percoll density gradients
Introduction
The basophil is potentially an important participant in immediate (Lichtenstein et al., 1978) and delayed hypersensitivity (Dvorak, 1976; Askenase, 1977) reactions. Attempts to clarify its functional role in the pathogenesis of these processes has been greatly limited by the low number of these cells found in peripheral blood. In the past, success with basophil isolation was modest from both human and animal blood (Day, 1972; Ishizaka et al., 1972). Recently, success in achieving high levels of purity and yield of basophils have been accomplished in humans with affinity chromatog- raphy (MacGlashan and Lichtenstein, 1980) density gradients (Raghuprasad, 1982), and flow microfluorometric techniques (Weil et al., 1983). In the guinea pig,
1 Supported in part by a grant from the American Lung Association and an Allergic Disease Center Grant AI-10404-12. 2 Presented in part at the National Meeting of the American Academy of Allergy, Hollywood, FL, March 1983. 3 Recipient of a National Institutes of Health Young Investigator Award AI-16331. 4 Address reprint requests to: Frank M. Graziano, M.D., Ph.D., University of Wisconsin Hospital, Department of Medicine, 600 Highland Avenue, Madison, WI 53792, U.S.A.
0022-1759/84/$03.00 © 1984 Elsevier Science Publishers B.V.

158
peripheral blood basophil enrichment has been obtained with Ficoll density gradi- ents (Dvorak et al., 1974).
In the following report, we would like to describe a new method for separation of highly purified guinea pig basophils that are viable, and functional. A combination of anti-leukocyte antibody and Perco]l density gradients was utilized, and the basophils obtained were not directly manipulated as a result of the isolation procedure.
Materials and Methods
Animals Outbred Hartley albino guinea pigs were obtained from Bio-Lab Corp. (St. Paul,
MN). Female animals weighing 250-350 g were used throughout the study. New Zealand White rabbits were obtained from Klubertanz Farms (Edgerton, WI). All animals were maintained in the Research Animal Resource Center at the University of Wisconsin.
Reagents Oxazolone (Ox; 4-ethoxymethylene-2-phenyl oxazolone) coupled to human serum
albumin (Ox40-HSA) and to guinea pig albumin (Ox28-GPA) were prepared by methods previously described (Askenase," 1973). Concanavalin A and the calcium ionophore A23187 were obtained from Calbiochem-Behring Corp. (La Jolla, CA). Percoll used for density gradients was obtained from Pharmacia Fine Chemicals (Uppsala). Complete Freund's adjuvant (CFA) was obtained from Difco Laborato- ries (Detroit, MI).
Antiserum to Ox-GPA was produced by immunizing guinea pigs with a 100 ~g priming dose of the antigen emulsified in CFA and distributed subcutaneously among the 4 footpads in 0.1 ml volumes. Seven days after this primary injection, the animals were skin tested with 100/~g and 200 ~g of Ox-GPA in separate sites. Five days later the animals were bled and serum was stored in aliquots at - 9 0 ° C . Antiserum to guinea pig leukocytes was prepared in rabbits. Briefly, a guinea pig leukocyte preparation (< 0.5% basophils) was obtained from normal animals (see below) and 8-10 × 10 6 cells were emulsified in CFA and distributed into the 4 footpads of the animals as a primary immunization. Monthly thereafter, a similar number of leukocytes ( < 0.5% basophils) emulsified in IFA were injected in multiple sites to boost antibody levels. The animals were bled monthly and aliquoted serum was stored at - 90°C . Antibody to guinea pig IgG 1 was also prepared in rabbits. The heavy chain specificity of this antibody was confirmed as previously described (Graziano et al., 1981), and protein determination was performed according to the method of Lowry (Lowry et al., 1951).
Buffers used in the study were the following: Hepes-buffered saline containing 10 mM N-2-hydroxethyl piperazine-N'-2-ethanesulfonic acid (Hepes), 137 mM NaC1, and 5 mM KC1 adjusted to pH 7.4 with NaOH. H G consisted of Hepes-buffered saline plus 0.5 mg/ml gelatin and 1 mg/ml dextrose. HGCM was H G containing 2 mM CaC12 and 1 mM MgC12.

159
Percoll stock (stk) was prepared by mixing 90 ml commercial Percoll solution with 8.3 ml of 10 × Hepes, and 1.7 ml distilled water. The osmolality of the resultant solution was 285 mosmolal/kg H20. The desired densities of Percoll were prepared by mixing HG with stk according to the following formula:
X (ml HG) = Y(ml-stk) x density (stk) - density (desired)
density (desired) - density (HG)
Cell isolation procedures Guinea pigs normally have basophil concentrations as < 1% of their circulating
leukocytes. Therefore prior to isolation procedures a basophilia was induced by daily intraperitoneal (i.p.) injection of sheep blood diluted 1 : 1 with saline (Dvorak et al., 1974). Thirteen days later guinea pigs were anesthetized and exsanguinated. The blood from individual animals was mixed with sufficient ethylenediamine tetraacetic acid (EDTA) to provide a final concentration of 10 mM. A leukocyte preparation was obtained from the whole blood using a modification of procedures previously described (Lett-Brown et al., 1981). In brief, EDTA containing blood was mixed with 3% gelatin (2 parts blood to 1 part gelatin) and placed at an angle of 30 ° in a 37°C water bath. After incubation for 45 min, the leukocyte-rich plasma was withdrawn and centrifuged at 750 rpm (25°C) for 30 min. In some experiments, pelleted cells were washed once in HG containing 4 mM EDTA and residual erythrocytes lysed with hypotonic shock. These cells were then washed with HG, resuspended in 1 ml of a mixture of HG and Percoll (1.06 g/ml 1 : 1) and applied to a discontinuous Percoll gradient (1.06, 1.07_, 1.08, 1.08 g/ml) over a cushion of Percoll density 1.12 g/ml. In other experiments, the peripheral blood leukocyte preparation was incubated (37°C for 30 min) with 2 ml of a 1 : 150 dilution of rabbit anti-guinea pig leukocyte antibody. At the end of the incubation period, 4.5% dextran was added to the antibody-leukocyte mixture (1 part leukocytes to 5 parts dextran), and further incubated (25°C) for 45 min. The leukocyte rich supernatant was removed, washed, applied to Percoll gradients (described above) and spun at 2000 rpm (25°C) for 20 min. Cell bands in the Percoll were then carefully removed, washed with HG, and counted.
Basophil and total cell counts were performed on whole blood, leukocyte prepara- tions, and gradient cell bands. Basophil counts were performed using the Alcian blue method (Gilbert and Ornstein, 1975). Differential counts were done on cytocentri- fuged (Cytospin, Shandon Elliott Corp.) smears stained with Wright's stain. Cell viability was evaluated by trypan blue dye exclusion. Where appropriate, the statistical significance of differences between cell counts was ascertained by using Student's unpaired 2-tailed t-test. A P value of < 0.05 was regarded as a statistically significant difference.
Electron microscopic examination of basophil preparations was performed by fixing pelleted cells with 4% glutaraldehyde in sodium cacodylate buffer (pH 7.4; 1 h at 4°C). The pellet was washed and then fixed with osmium tetroxide in collidine buffer (pH 7.2; 30 min at 25°C). After dehydration the cell pellet was embedded in

160
Epon-Araldite and thin sections (80-90 nm) were cut with an Ultramicrotome (LKB, Broma). Sections were stained with uranyl acetate and lead citrate, and examined in a Hitachi H 500 electron microscope.
In vitro histamine release Histamine release from isolated basophils was evaluated with methods previously
described (Lett-Brown et al., 1983). The secretogogues used were the following: calcium ionophore A23187, concanavalin A, anti-IgG~ antibody, and Ox-HSA conjugates. Briefly, varying concentrations of these agents in HGCM were added in 0.3 ml aliquots to test tubes and equilibrated for 10 min in a 37°C water bath. Enriched basophil suspensions (0.3 ml) were then added to these tubes and in- cubated (37°C) in a shaking water bath for 45 min. At the end of this time period, the tubes were centrifuged, and the supernatant fluid assayed for histamine content using an automated fluorometric technique (Siraganian, 1975). Total histamine content was estimated by treating separate cell suspensions with 6% perchloric acid. Tubes containing cells and buffer alone, and cells and an extraneous antigen (ovalbumin) estimated spontaneous and non-specific histamine release respectively. All histamine assays were done in duplicates.
Basophil-rich suspensions were passively sensitized in vitro with anti-oxazolone antibody by incubating 1-4 x 106 washed and pelleted cells with 1 ml of anti-Ox antibody containing 4 mM EDTA for 60 min a't 37 ° C in a shaking water bath. At the end of this period the cells were washed, resuspended in HGCM and histamine release performed as described above.
Results
Basophil isolation Normal adult guinea pigs have peripheral blood basophil counts of approximately
2 x 104/ml (Dvorak et al., 1974). As noted by others (Dvorak et al., 1974), we were able to increase the concentration of circulating basophils by 12 daily i.p. injections of sheep blood. This procedure in our study led to a mean peripheral blood basophilia of 6% with a range of 2%-12%.
In initial experiments, we localized basophil migration in Percoll gradients by application of leukocyte preparations obtained from basophilic animals to various discontinuous and preformed gradients. Peak basophil counts and histamine con- centration were found at a density of 1.08 g /ml (range 1.07-1.09 g/ml). In our next series of experiments, we applied leukocyte preparations (_< 50 x 10 6 cells per gradient) to discontinuous Percoll gradients with densities of 1.06-1.09 g /ml (at 0.01 g /ml increments). Table I illustrates the results of one such experiment in which a leukocyte preparation containing 6% basophils obtained from 2 animals was layered onto Percoll. As can be observed basophil enrichment to 40% purity was achieved. Though basophil enrichment on Percoll was consistently observed with this method, additional experiments showed the variability to be great. Because of this, in further experiments leukocyte preparations were treated with an anti-leuko-

161
TABLE I
COMPARISON OF BASOPHIL ENRICHMENT FROM TWO INDIVIDUAL EXPERIMENTS USING PERCOLL ALONE OR WITH AN ANTI-LEUKOCYTE ANTIBODY
Procedure a Purification step Total no. cells Basophils Basophil
Basophils WBC (%) yield
Percoll alone
Anti-leukocyte Antibody + Percoll
Whole blood b 8.4 ×106 1.4x 10 s 6 Leukocyte prep. 4.9 x 106 8.2 x 107 6 58 Percoll (1.0 g /ml) 2.5 x 106 6.3 × 106 40 30
Whole blood b 9.2 x 106 1.5 × I0 s 6
Leukocyte prep. 5.2 × 106 1.9 × 107 27 57 Percoll (1.08 g /ml) 3.78 x 106 3.8 x 106 99 41
a The blood from 2 animals was used for each procedure. b Starting volume of blood was 30 ml.
cyte antibody to decrease the number of potentially contaminating cells prior to applying the cells to Percoll.
Rabbit anti-leukocyte serum was heat inactivated (at 56°C for 0.5 h), adsorbed with guinea pig whole serum, and with highly purified preparations of basophils (85-90%) that were occasionally obtained with Percoll alone. We confirmed specific- ity of the antibody for non-basophil leukocytes with immunofluorescence using an (Fab') 2 goat anti-rabbit IgG in a sandwich technique, and by the absence of histamine release using the anti-leukocyte antibody in dilutions of 1 : 50 to 1 : 250. The optimal dilution of anti-leukocyte antibody for basophil enrichment was found to be 1:150. Table I demonstrates the results of a single experiment in which antibody and Percoll were used. Enrichment of basophils (27%) significantly above the level in whole blood was observed in the leukocyte preparation treated with anti-leukocyte antibody. Clumped leukocytes and only rare basophils were seen in the dextran sedimented cells. The leukocyte suspension treated with antibody was then layered onto a discontinuous gradient as described above and the basophils were further purified to 99% with 41% of the cells being recovered from whole blood.
Electron microscopic examination of isolated basophils showed typical large oval granules with a parallel array of granular material and a rim of less-dense material surrounding the granular matrix and separating it from the granular membrane (Fig. 1A and B).
Table II summarizes our data for basophil enrichment using Percoll alone or in combination with an anti-leukocyte antibody. Basophil enrichment was significantly greater (P < 0.05) in antibody treated leukocyte preparations. After Percoll gradient centrifugation of these antibody treated leukocyte preparations further enrichment of basophils to an average purity of 82% was obtained. This was also significantl2¢ different from that seen with Percoll alone (P < 0.001). In 3 out of 40 experiments using Percoll alone, and 16 out of 30 experiments using both anti-leukocyte antibody

162
A
Q l b
Fig. 1. A: electron micrograph of a purified guinea pig basophil preparation, x 13,200. B: typical large oval guinea pig basophil granules with a parallel array of granular material and rim of less-dense material surrounding the granular matrix and separating it from the peri-granular membrane. × 22,000.
163
TABLE II
C U M U L A T I V E DATA FOR BASOPHIL E N R I C H M E N T USING PERCOLL A L O N E OR IN COM- BINATION WITH AN ANT I -L E UKOC YT E ANTIBODY
Procedure Percent basophils + SD Basophil
(no. experiments) Leukocyte Percoll yield
preparation 1.08 g m / m l (%)
Percoll 6 + 4 39 5:24 22 + 12 (40) ( 2 - 1 7 ) " (2 -95 ) (5-40)
Anti-leukocyte 19 + 6 82 5-10 39 + 17 an tibody + Percoll ( 10- 31 ) (65 - 99) ( 20 - 70) (30)
a Values in parentheses indicate the range of basophil percent or yield seen for all experiments in a group.
and Percoll, basophil purities of > 85% were obtained. In the later group, 10 experiments produced purities of 90% or greater. The yield of basophils in the antibody and Percoll group was approximately 40% of the starting basophils in whole blood. Contaminating cells in the-basophil preparations were shown to be primarily neutrophils and eosinophils were also observed. Viability by trypan blue dye exclusion was greater than 90%
I00
m 6O
~4o ~ 20 w
I
,-I, re I,U
~ 5 0 -
B z i,i ~ 3 0 - u,I a. 2 0 -
IO--
O , i0-2
z I 1 I z <~ Oio-2 IO-' 10 ° IO' IO 2
CONCENTRATION CALCIUM IONOPHORE I - A23187 (/~g/ml ) ¢0
5 0 -
4 0 -
C
z
~ 3o w a. 20
30 --
20 - -
CONCENTRATION OXAZOLONE-HSA 50 (pg/ml)
Io
I I03
I I I I n i I I [ I I 0 -P I0 ° I0 ' 10 2 - I0- ' I0 ° I0' I 0 ~ 10 3
CONCENTRATION CONCANAVALIN A CONCENTRATION ANTI-IgG,(/~g/ml) (/~g/mi)
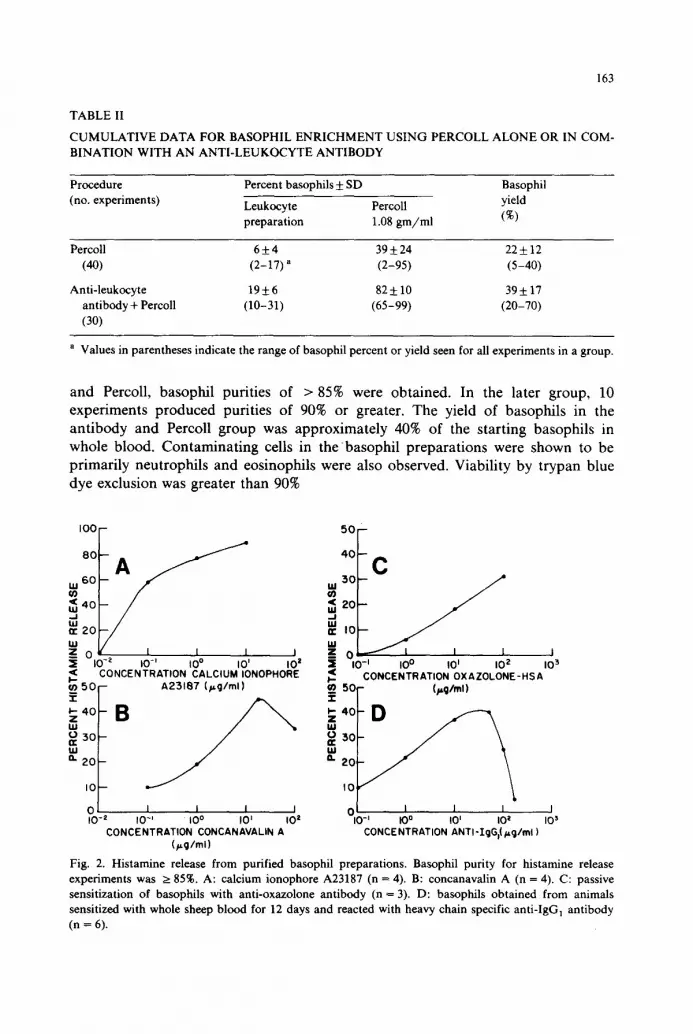
Fig. 2. Histamine release from purified basophil preparations. Basophil purity for histamine release experiments was >__ 85%. A: calcium ionophore A23187 (n = 4). B: concanavalin A (n = 4). C: passive sensitization of basophils with anti-oxazolone antibody (n = 3). D: basophils obtained from animals sensitized with whole sheep blood for 12 days and reacted with heavy chain specific anti-IgG] antibody (n = 6).

164
Histamine release To provide evidence that isolated basophils were functional, we studied their
ability to release histamine upon stimulation. A number of different secretogogues were used. The calcium ionophore A23187, a non-specific histamine releasing agent, stimulated a peak of 90% histamine release at a dose of 10/~g (Fig. 2A). The curve of concanavalin A induced release followed that which has been previously published for heterogeneous populations of guinea pig basophils (Lett-Brown et al., 1981). Peak histamine release was seen at 25/~g of Con A (Fig. 2B). Taking advantage of unoccupied Fc receptors on isolated basophils, the cells were passively sensitized with anti-oxazolone antibody. Upon challenge with Ox-HSA an antigen-induced dose-dependent histamine release was observed "(Fig. 2C), Finally, reaction of purified basophils with varying concentrations of heavy chain specific anti-guinea pig IgG 1 (0.1-100/~g) also resulted in dose-dependent histamine release (Fig. 2D). Histamine release was greatly decreased at antibody concentrations of 100 #g or greater.
Detectable histamine release was observed with 2 x 10 4 purified basophils using the calcium ionophore A23187 as secretogogue. Spontaneous and non-specific histamine release in all experiments ranged between 3-8%. Estimates of histamine content in basophils ranged from 0.6 to 2 pg/cell with an average 0.9 pg/cell.
Discussion
Percoll density gradients have been used to purify various cell populations, viruses, and organelles (Pertoft and Laurent, 1977). In our study, basophil enrich- ment was observed when leukocyte suspensions from whole blood were applied directly to this density gradient material. The purity and yield of basophils was significantly enhanced, however, if the leukocyte suspension was treated with anti-leukocyte antibody prior to application to Percoll. With this latter method a mean basophil purity of 82% and yield of 39% was observed.
For this procedure to be successful a number of steps are important. First, it was necessary to produce basophilic animals (Dvorak et al., 1974). If circulating basophils were less then 1%, basophil enrichment was unsuccessful. Second, it was necessary to adsorb (whole guinea pig serum and enriched populations of basophils) and heat inactivate the anti-leukocyte serum. This assured that antibody to cell-bound im- munoglobulin or basophils was not present, and that complement components would not affect the functional integrity of the basophils. Treatment of leukocyte preparations obtained from whole blood with this antibody gave an initial enrich- ment for basophils up to 30%. This step is critical to the isolation procedure for we have noted that higher basophil percentages in leukocyte preparations before Percoll lead to greater purity of basophils obtained with the Percoll density gradients. Third, the density and osmolality of the Percoll must be regulated. Best results have been obtained with a density of 1.080 g /ml and an osmolality of 285-290 mosmolal /kg H20. Furthermore, extreme care is also required during aspiration of cells and Percoll from above the basophil band. Creation of vortices during the aspiration,

165
forces contaminat ing cells into the basophil band and dramatically lowers the degree of purity.
The functional nature of the isolated basophils was verified by the stimulated cells ability to release histamine. Since guinea pig platelets contain histamine, contamina- tion with this cell was minimized by incubating the gelatin-whole blood mixture at an angle of 30 ° and low speed centrifugation (750 rpm) of the resultant leukocyte-rich plasma (Lett-Brown et al., 1981). Platelet contaminat ion was estimated by coulter counter and microscopic examination. When studying histamine release a number of secretogogues were used. Thus, demonstrat ing that the basophils obtained with our isolation procedure could respond to stimuli acting through a wide variety of membrane activators. Impor tan t in this regard is the fact that the basophils were not specifically treated with ant ibody for cell isolation. Therefore, unoccupied Fc receptors were present and the cells could be passively sensitized with ant ibody to s tudy antigen dependent histamine release.
We conclude that the use of anti-leukocyte and Percoll density gradients is a relatively fast and easy method of isolating guinea pig basophils f rom whole blood in high purity and yield. Funct ional ly iritact basophils 1 -5 x 10 6 in number can be isolated from single animals. This method can perhaps facilitate the study of the role of the basophil in inflammation.
References
Askenase, P.W., 1973, J. Exp. Med. 138, 1144. Askenase, P.W., 1977, Prog. Allergy 23, 199. Day, R.P., 1972, Clin. Allergy 2, 205. Dvorak, H.F., 1976, J. Allergy Clin. Immunol. 58, 229. Dvorak, H.F., S.S. Selvaggio, A.M. Dvorak, R.B. Colvin, D.B. Lean and J. Rypysc, 1974, J. Immunol.
113, 1694. Gilbert, H.S. and L. Ornstein,. 1975, Blood 46, 279. Graziano, F.M., C. Haley, L. Gunderson and P.W. Askenase, 1981, J. Immunol. 127, 1067. lshizaka, T.R., R. DeBernardo, H. Tomioka, L.M. Lichtenstein and K. Ishizake, 1972, J. Immunol. 108,
1000. Lett-Brown, M.A., D.O. Thueson and J.A. Grant, 1981, Int. Arch. Allergy Appl. Immunol. 64, 241. Lett-Brown, M.A., E.B. Mitchell and P.W. Askenase, 1983, J. Immunol. 130, 2104. Lichtenstein, L.M., G. Marone, L.L. Thomas and F.V. Malveaux, 1978, J. Invest. Dermatol. 71, 65. Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951, J. Biol. Chem. 193, 265. MacGlashan, D.W. and L.M. Lichtenstein, 1980, J. Immunol. 124, 2519. Pertoft, H. and T.C. Laurent, 1977, in: Methods of Cell Separation, Vol. 1, ed. N. Catsimpoolis (Plenum
Press, New York) pp. 25-65. Raghuprasad, P.K., 1982, J. Immunol. 129, 2128. Siraganian, R.P., 1975, J. Immunol. Methods 7, 283. Weil, G.J., W.M. Leiserson and T.M. Chused, 1983, J. Immunol. Methods 58, 359.





























