Embed Size (px)
Citation preview

This article was downloaded by: [117.173.186.197]On: 25 March 2014, At: 08:03Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: MortimerHouse, 37-41 Mortimer Street, London W1T 3JH, UK
Italian Journal of ZoologyPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/tizo20
Karyotypic and genetic divergence in theEthiopian Myomys ‐ Stenocephalemys complex(Mammalia, Rodentia)Marco Corti a , Alessandra Scanzani a , Anna Rita Rossi a , Maria Vittoria Civitelli a ,Afework Bekele b & Ernesto Capanna ca Dipartimento di Biologia Animale e dell'Uomo , Universitâ degli Studi ‘La Sapienza’di Roma , via Borelli 50, Roma, I‐00161, Italy E-mail:b Biology Department , Addis Ababa University , Addis Ababa, Ethiopiac Dipartimento di Biologia Animale e dell'Uomo , Universitâ degli Studi ‘La Sapienza’di Roma , via Borelli 50, Roma, I‐00161, ItalyPublished online: 28 Jan 2009.
To cite this article: Marco Corti , Alessandra Scanzani , Anna Rita Rossi , Maria Vittoria Civitelli , Afework Bekele& Ernesto Capanna (1999) Karyotypic and genetic divergence in the Ethiopian Myomys ‐ Stenocephalemys complex(Mammalia, Rodentia), Italian Journal of Zoology, 66:4, 341-349
To link to this article: http://dx.doi.org/10.1080/11250009909356275
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”)contained in the publications on our platform. However, Taylor & Francis, our agents, and our licensorsmake no representations or warranties whatsoever as to the accuracy, completeness, or suitabilityfor any purpose of the Content. Any opinions and views expressed in this publication are the opinionsand views of the authors, and are not the views of or endorsed by Taylor & Francis. The accuracy ofthe Content should not be relied upon and should be independently verified with primary sources ofinformation. Taylor and Francis shall not be liable for any losses, actions, claims, proceedings, demands,costs, expenses, damages, and other liabilities whatsoever or howsoever caused arising directly orindirectly in connection with, in relation to or arising out of the use of the Content.
This article may be used for research, teaching, and private study purposes. Any substantial orsystematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution inany form to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

Ital. J. Zool., 66. 341-349 (1999)
Karyotypic and genetic divergence in theEthiopian Myomys - Stenocephalemyscomplex (Mammalia, Rodentia)
MARCO CORTIALESSANDRA SCANZANIANNA RITA ROSSIMARIA VITTORIA CIVITELLIDipartimento di Biologia Animale e dell'Uomo,Universitâ degli Studi 'La Sapienza' di Roma,via Borelli 50, I-00161 Roma (Italy)E-mail: [email protected]
AFEWORK BEKELEBiology Department, Addis Ababa University, Addis Ababa (Ethiopia)
ERNESTO CAPANNADipartimento di Biologia Animale e dell'Uomo,Universitâ degli Studi 'La Sapienza' di Roma,via Borelli 50, I-00161 Roma (Italy)
ABSTRACT
The Ethiopian Myomys - Stenocephalemys complex has longconstituted a taxonomic and systematic puzzle. The species occurat different altitudes, from 1000 up to 4300 m a.s.l., and show re-markable adaptations to high altitude habitats. The four speciesforming the complex, M. albipes, M. ruppi, S. albocaudata and S.griseicauda, are all endemic to the country and constitute amonophyletic group. We describe their karyotypes, includingtheir G- and C- banding patterns, and their allozyme patterns.Phylogenetic trees based on chromosomal rearrangements and al-lozymes support the current taxonomy. Nonetheless, comparisonswith genetic similarity data and other hypotheses drawn frommtDNA and geometric morphometrics show substantial disagree-ment, and therefore it is not possible to claim the paraphyly ofStenocephalemys which would then include Myomys.
KEY WORDS: Myomys - Stenocephalemys - African rodents -Chromosomes - Allozymes.
ACKNOWLEDGEMENTS
Ato Goitom Redda (Addis Ababa) contributed substantiallywith his experience and friendship during the 1994-1995 Bale andShowa field expeditions, which were organised with the permis-sion of the Ethiopian Wildlife Conservation Organization (EWCO).Many thanks go to the personnel of the Bale Mountains NationalPark. This work benefits from the grants "Caratterizzazione dellabiodiversità in ambiente tropicale: un'analisi genetica, molecolaree morfometrica in Vertebrati", Fondi Ateneo ex 60%, and "Com-missione per i Musei Naturalistic! dell'Accademia Nazionale deiLincei".
(Received 13 July 1999 - Accepted 20 August 1999)
INTRODUCTION
Ethiopia is known for its high number of endemicmammals: among the 70 species of rodents recorded,21% are endemic (Yalden & Largen, 1992; Hillman,1993; Yalden et al, 1996). This is mainly determined bythe diversity characterizing the country, as a conse-quence of high altitudinal ranges, from below sea levelup to more than 4000 m a.s.l. Furthermore, the Ethiopi-an plateau is bisected by the Rift Valley which isolateshigh altitude species on its western and eastern sides.
On the eastern plateau, the Bale mountain region isthe main massif and constitutes an area of high rodentendemism. Different species have been recorded onlyhere, such as Arvicanthis blicki Frick, 1914, Lophuromysmelanonyx Petter, 1972, Lophuromys sp. (Aniskin et al,1997a), Tachyoryctes macrocephalus (Riippel, 1842),Stenocephalemys albocaudata Frick, 1914. Recently, ithas been shown that most of the rodent species occur-ring in this area are also characterized by extensivekaryotype rearrangements (Corti et al., 1995; 1996;Aniskin et al, 1997a, b; Lavrechenko et al, 1997), sug-gesting that isolation and altitude have played a role infavouring the establishment of chromosomal variants.
Among the Ethiopian endemic species, there is thegroup including the genera Stenocephalemys and Myo-mys, which have been considered phylogeneticallyclosely related (Yalden et al, 1976). Stenocephalemyscomprises only two species: S. albocaudata, endemic toBale, and S. griseicauda, which has a broader distribu-tion including Bale and Simien (Yalden et al., 1976).They occur at different altitudes, S. albocaudata beingconfined to the high altitude Afro-Alpine moorlands be-tween 3000 and 4100 m a.s.l., and S. griseicauda morewidespread in scrubby, open habitats between 2400and 3300 m a.s.l. (Yalden & Largen, 1992). Their rangesoverlap to a certain extent in the Bale massif between3000 and 3300 m (Yalden, 1988). Myomys albipes (Rüp-pel, 1842), (Yalden et al, 1976), the other member ofthe group, has a much wider distribution in areas ex-tending between 1500 and 3300 m a.s.l. (AfeworkBekele & Corti, 1997). This species is progressively re-placed altitudinally in Bale by S. griseicauda, althoughit prefers forest habitats (Yalden, 1988).
This complex of species has recently been the subjectof several studies including the sequence analysis of themtDNA gene l6S (Fadda et al., submitted), and three-di-mensional geometric morphometrics of the skull (Fadda& Corti, in press). These studies have argued that thegenus Stenocephalemys may be paraphyletic, so that thecurrent taxonomy may not reflect the systematic rela-tionships between species.
In this paper we describe the karyotypes and the mul-ti-locus allozyme patterns of S. albocaudata and S. gri-seicauda from the Bale Mountains, and of M. albipesfrom the same and other regions of Ethiopia. In an at-tempt to contribute to the clarification of the systematicsof this species complex, we compared their karyotypeswith M. fumatus from Somalia and their allozyme pat-
Dow
nloa
ded
by [
117.
173.
186.
197]
at 0
8:03
25
Mar
ch 2
014

342 M. CORTI, A. SCANZANI, A. R. ROSSI, M. V. CIVITELLI, AFEWORK BEKELE, E. CAPANNA
= M. albipes
IN IN M. fumatus
Fig. 1 - Map of East Africa and the Horn of Africa (enlargement ofthe area boxed), with the approximate ranges of M. albipes andM. fumatus and the location of the samples studied. Ranges of 5.griseicauda, S. albocaudata and D. harringtoni are not shown. 1,Sululta; 2, Dinshu; 3, Sanetti Plateau; 4, Harenna Forest; 5, Man-aghesha; 6, Afgoi.
terns with Desmomys barringtoni from Ethiopia. Thechoice of M. fumatus as the outgroup was consideredreasonably appropriate as this species has been sug-gested by Misonne (1969) as the most ancient memberof the Praomys/Myomys group.
MATERIALS AND METHODS
Specimens were live trapped during Ethiopian field expeditionsin 1994 and 1995 and in Somalia (M. fumatus) in 1979 (Fig. 1).Trapping localities are as follows (the number of individuals is in-dicated for chromosome preparations and allozymes):
Myomys albipes: Ethiopia, Sululta (near Addis Ababa, 09° 15' N- 38° 43' E, 2700 m a.s.l.), chromosomes - 2c?, 1Q; allozymes = 4.Dinshu (Bale Mountains National Park, 07° 06' N - 39° 48' E, 3200m a.s.l.), chromosomes - 2çf, allozymes = 1. Managhesha (08°55' N - 38° 37' E, 2300 m a.s.l.), chromosomes = lcf, 19; al-lozymes = 7. Harenna Forest (Bale Mountains National Park, 06°42' N - 39° 44' E, 2400 m a.s.l.), allozymes = 1.
M. fumatus: Somalia (Afgoi, 02° 90' N - 45° 07 E); chromo-somes = one Q.
Stenocephalemys albocaudata: Ethiopia, Sanetti Plateau (BaleMountains National Park, 06° 52' N - 39° 52' E, 4100 m a.s.l.),chromosomes and allozymes = 2cf, 2Ç>.
5. griseicauda: Ethiopia, Dinshu (Bale Mountains National Park,07° 06' N - 39° 48' E, 3200 m a.s.l.), chromosomes and allozymes= one cf;
Desmomys harringtoni: Ethiopia, Sululta (09° 15' N - 38° 43' E,2700 m a.s.l.) and Menagesha (08° 55' N - 38° 37' E, 2300 ma.s.l.), allozymes, 8 specimens.
We adopted the recent revision of the genus Praomys by Mus-
ser & Carleton (1993) and assigned the name Myomys to allspecies occurring in the Horn and East Africa.
Chromosomes
Chromosome metaphases were obtained from the bone mar-row at the Biology Department, Addis Ababa University, follow-ing Hsu & Patton (1969). Slides were then prepared at the Dipar-timento di Biologia Animale e dell'Uomo, University of Rome 'LaSapienza'.
Metaphases were stained by the Giemsa pH7 method. AgNORswere obtained following Howell & Black (1980). G- and C- bandswere obtained through the methods of Seabright (1971) and Bick-ham (1979), respectively.
Homologue elements in the species were detected on the basisof chromosome morphology and G-banding, and character statetrees (cts) constructed following Corti et al. (1996). Direction ofchromosomal change was established by assuming the M. fuma-tus state as primitive. When a corresponding homologue was notfound in a species, we simply assumed it to be absent, which doesnot alter the construction of cts. The matrix of cts was subjectedto parsimony analysis to find the most parsimonious tree.
Electrophoresis
Muscle and liver were dissected from each specimen in thefield and stored immediately at -80° C. Allozymes were detectedby horizontal starch-gel electrophoresis (Table I), following thestandard staining procedures (Pasteaur et al., 1988). Twenty-fivepresumptive gene loci, encoding 21 enzyme system were re-solved in all samples. Alleles at each locus were designated bytheir anodic mobilities (100) relative to the most frequent allele inthe M. albipes population, designated as 100 (>100 = higher mo-bility, <100 = lower mobility). Individuals from the latter popula-tion were used as controls on all gels.
Allelic frequencies at all loci were calculated. Estimation of ge-netic and phenetic relationships among populations was obtainedcalculating Nei's (1978) genetic distances and generating aphenogram of all samples by means of the UPGMA cluster analy-sis (Sneath & Sokal, 1973).
A phylogenetic reconstruction, using "discrete character parsi-mony analysis" (Crother 1990) was performed. All enzyme elec-tromorphs (alleles) were treated as characters with one of twostates, present (1) or absent (0), for all taxa, and the binary-codedcharacters were used for the analysis. The branch-and-bound al-gorithm of PHYLIP (Felsenstein, 1993) was used to search for thetrees requiring the smallest number of changes (steps), and therobustness at each node was tested by bootstrapping with 1000-dataset replicates.
RESULTS
Karyotype description
Myomys albipes - The diploid number is 2n = 46, theautosomal Fundamental Number (aFN) is 50-53. The au-tosomal set is composed by 17 pairs of acrocentrics de-creasing in size, two pairs of submetacentrics, a pair ofsmall metacentrics, and two pairs showing a polymor-phism, having either an acrocentric or submetacentricshape (Fig. 2). Given the limited number of specimensstudied, it is impossible to ascertain whether this poly-morphism occurs on a geographic basis. The X-chromo-some is a large submetacentric and the Y-chromosomeis a small metacentric (Fig. 2). The Y-chromosome isentirely heterochromatic and the short arm of the X-chromosome is C-positive (Fig. 3c). A maximum num-
Dow
nloa
ded
by [
117.
173.
186.
197]
at 0
8:03
25
Mar
ch 2
014

DIVERGENCE IN MYOMYS AND STENOCEPHALEMYS 343
TABLE I - The enzyme systems tested, with the Enzyme Commission Number (E.C.N.), the loci, the buffer systems and the tissue used.
Enzyme system
Aspartate aminotransferase
Aconitate hydrataseAdenosine deaminaseAlcohol dehydrogenaseAdenylate kinaseCreatine kinaseGlyceraldehyde-3-phosphate dehydrogenase(x-Glycerophosphate dehydrogenaseGlucose-6-phosphate dehydrogenaseGlucosephosphate isomeraseIsocitrate dehydrogenaseLactate dehydrogenase
Malate dehydrogenase
Malic enzymeMannosephosphate isomerasePhosphogluconate dehydrogenasePhosphoglucomutaseGeneral proteins
Superoxide dismutaseXanthine dehydrogenase
E.C.N.
2.6.1.1
4.2.1.33.5.4.41.1.1.12.7.4.32.7.3-21.2.1.121.1.1.81.1.1.495.3.1.91.1.1.421.1.1.27
1.1.1.37
1.1.1.405.3.1.81.1.1.43 ,2.7.5.1
1.15.1.11.2.1.37
Locus
Aat-1*Aat-2*Aco-VAda-1*Adh *Ak-1*Ck-1*Gapd-rGpd*G6pdh*Gpi*Idh *Ldh-VLdh-2 *Ldh-3mMdh *Mdh-1 *Me*Mpi 'Pgd*Pgm-1*Pt-1*Pt-2'Sod'Xdh*
Buffer
4422422242421114443424411
Tissue
MuscleMuscleMuscleMuscleLiverMuscleMuscleMuscleMuscleMuscleMuscleMuscleMuscleMuscleMuscleMuscleMuscleMuscleMuscleMuscleMuscleMuscleMuscleLiverMuscle
1, Tris/citrate (Poulik, 1957); 2, Tris/citrate (Selander, 1971); 3, Tris/versene/borate (Brewer & Sing, 1970); 4, Phosphate/citrate (Harris,1966).
ber of five chromosomes carrying AgNORs was found(Fig. 3b). These NORs have an intercalary position inthe first pair, and a telomeric position in pairs 7 and 15.The G-banded karyotype is shown in Figure 4.
Myomys fumatus - The diploid number is 2n = 36,and the aFN is 38. All autosomes are acrocentric de-creasing in size except for a pair of small metacentrics(Fig. 2). The X-chromosome is a large submetacentric.There is no information on the Y-chromosome, as theonly specimen analysed was a female. C-bands and Ag-NORs were not available for this specimen. The G-banded karyotype is shown in Figure 4.
Stenocephalemys albocaudata - The diploid number is2n = 54, and the aFN is 60. The autosomal set is com-posed of three pairs of large subtelocentric chromo-somes, 22 pairs of acrocentrics decreasing in size, and apair of small metacentrics (Fig. 2). A polymorphism wasfound for the X-chromosome, being either a large meta-centric or a submetacentric (Figs 2, 3c and 4). The Y-chromosome is a medium-size heterochromatic submeta-centric (Fig. 3c). The short arm of the X-chromosome isC-positive (Fig. 3c). A maximum number of six chromo-somes carrying AgNORs was found, occurring on fourdifferent pairs: in a telomeric position on chromosome 3,at the level of the secondary constriction on chromosome14, in a telomeric position on a small acrocentric, and in
a centromeric position on a small acrocentric (Fig. 3a).The G-banded karyotype is presented in Figure 4.
Stenocephalemys griseicauda - The diploid number is2n = 54, and the aFN is 54 (Fig. 2). One large subtelo-centric chromosome, 24 acrocentric pairs decreasing insize and a pair of small metacentric chromosomes con-stitute the autosomal set. The X-chromosome is a largesubmetacentric, and the Y-chromosome is a medium-sized submetacentric. The short arms of the X chromo-some are heterochromatic and the Y chromosome is en-tirely C-positive (Fig. 3c). No information is available forAgNORs. The G-banded karyotype is shown in Figure 4.
Karyotype comparisons
Homologies and chromosomal rearrangements weredetected by comparing karyotypes of G-banded chro-mosomes (Fig. 4). The chromosome numbering ofStenocephalemys was chosen as a reference. The corre-sponding chromosome number of Myomys albipes andM. fumatus follows in parentheses; nf indicates absenceof the homologue; p indicates a polymorphic state.
Character state trees (cts) were defined on the basisof comparisons (S. albocaudata = Sa; S. griseicauda =Sg; M. albipes = Ma; M. fumatus - Mf), and polarised onM. fumatus.
Dow
nloa
ded
by [
117.
173.
186.
197]
at 0
8:03
25
Mar
ch 2
014

344 M. CORTI, A. SCANZANI, A. R. ROSSI, M. V. CIVITELLI, AFEWORK BEKELE, E. CAPANNA
KM
nfl (ÏÙ4 5
nn «ft00 m «s on nn oft 1/ If II ill ilf IS
SS fill nh 00 « A I III
pO ftR itD * H H$l fl ifMyomys albipes
6S fift fiß OS0 8 Oft f?:ü ;(i!OA f)9 Oft
X Y Myomys fumatus X X
I I II fil I I SA 10m no ü «J.an m
Stenocephalemys albocaudata X X Stenocephalemys griseicauda X Y
Fig. 2 - The karyotypes of the four species studied. In the box are enclosed the different forms of chromosome pairs 4 and 5 occurringin M. albipes.
Sa Sa Sg Ma
XFig. 3 - a, b, The chromosomes carrying AgNORs in (a) S. albo-caudata and (b) M. albipes. c, Constitutive heterochromatin insex chromosomes as determined by C-banding. Sa, S. albocauda-ta; Sg, 5. griseicauda; Ma, M. albipes.
Chromosome 1 (1, 1) - This pair shares the same sub-telocentric morphology and banding pattern in S. albo-caudata and S. griseicauda. M. albipes and M. fumatus
show the same banding pattern as the former twospecies, but with an acrocentric morphology. This chro-mosome carries AgNORs in M. albipes (Fig. 3a, b). Cts:Ma,Mf->Sa,Sg.
Chromosomes 2 (8, nf) - This pair presents the samebanding pattern in M. albipes, S. albocaudata and S.griseicauda, but it cannot be recognized in M. fumatus.The chromosome morphology is acrocentric in M. al-bipes and S. griseicauda but subtelocentric in S. albo-caudata. Cts: Mf-»Sg,Ma->Sa.
Chromosomes 3 (5, nf) - This pair presents the samebanding pattern in M. albipes, S. albocaudata and S. gri-seicauda, but it cannot be recognized in M. fumatus. Thechromosome morphology is acrocentric S. griseicaudaand subtelocentric in S. albocaudata. Myomys albipesshows a polymorphism for this chromosome morpholo-gy (see previous section), and for the comparison theacrocentric state has been used. Cts: Mf —>Sg,Ma(p)—»Sa.
Chromosome 4 (4, nf) - This pair presents the samemorphology and banding pattern in M. albipes, S. albo-caudata and S. griseicauda, but it cannot be recognizedin M. fumatus. Myomys albipes shows a polymorphismfor this chromosome morphology (see previous sec-tion), and for the comparison the acrocentric state hasbeen used. Cts: Mf->Sa,Sg,Ma(p).
Dow
nloa
ded
by [
117.
173.
186.
197]
at 0
8:03
25
Mar
ch 2
014

DIVERGENCE IN MYOMYS AND STENOCEPHALEMYS 345
Sa Sg Ma Mf
mi m m IK1 1 1 1 2 2 8 3 3 5 4 4 4 5 5
11 11 9
16 1614
22 22
12 12 12
50 ß181817
.h*23 23
If«6 6 3 7 7 6 3 8 8 179 179 2 2 10 10 7 5
131311 14 1410 15 15
19 1913 8
24 2419
20 20 15
25 25
ftéÛ21 21 18
26 26 26 22
Sa Sa Sg Ma Mf Sa Sg Ma
X Y
Fig. 4 - G-banded comparison of the karyotypes of the four species. The comparison is based on the karyotype of S. albocaudata.Some of the corresponding elements are not recognised in M. albipes and Mfumatus. Sa, S. albocaudata, Sg, 5. griseicauda, Ma, M. al-bipes, Mf, M.fumatus. (See the text for details).
Chromosome 11 (9, nf), 12 (12, nf), 13 (11, nf), 16(14, nf), 18 (17, nf), 20 (15, nf), 21 (18, nf), 24 (19, nf) -These pairs present the same morphology and bandingpattern in M. albipes, S. albocaudata and S. griseicauda,but they cannot be recognized in M. fumatus. Cts:Mf->Sa,Sg,Ma.
Chromosome 5 (nf, nf), 8 (nf, nf), 15 (nf, nf), 22 (nf,nf), 23 (nf, nf), 25 (nf, nf) - The chromosome has thesame morphology and banding pattern in S. albocauda-ta and S. griseicauda, but it cannot be recognized in M.fumatus and M. albipes. Cts: Ma,Mf—»Sa.Sg.
Chromosome 6 (3, nf) - This pair has the same band-ing pattern in M. albipes, S. albocaudata and S. grisei-cauda, but with a different morphology (subtelocentric)
in M. albipes. The chromosome cannot be recognized inM.fumatus. Cts: Mf->Ma->Sa,Sg.
Chromosome 7 (6, 3), 10 (7, 5), 19 (13, 8), 26 (22, 16)- These chromosome pairs have the same morphologyand banding pattern in all four species.
Chromosome 9 (2, 2) - This pair has the same mor-phology and banding pattern in S. albocaudata and S.griseicauda, while M. albipes and M. fumatus show anaddition in the long arm corresponding to chromosome17 of the former two. Moreover, this chromosome issubtelocentric in M. albipes. Cts: Ma->Mf->Sa,Sg.
Chromosome 14 (10, nf) - This pair presents the samemorphology and banding pattern in M. albipes, S. albo-caudata and S. griseicauda, but it cannot be recognized
Dow
nloa
ded
by [
117.
173.
186.
197]
at 0
8:03
25
Mar
ch 2
014

346 M. CORTI, A. SCANZANI, A. R. ROSSI, M. V. CIVITELLI, AFEWORK BEKELE, E. CAPANNA
I I I I I I I I M M 1 I2a 3a 4a 6a 11 12 1314a16 18 20 21 24 X
3p 4p 9a
I I I2b 3b 14b
I I I I I I I I I1 5 6b 8 9b 15 22 23 25
M. fumatus
M. albipes
S. albocaudata
S. griseicauda
Fig. 5 - Phylogenetic tree constructed on the basis of chromosomal rearrangements. The tree has been outgroup rooted on M. fumatusand does not present any homoplasy. Bars on the branches indicate the occurrence of chromosomal rearrangements; p indicates a poly-morphic state.
in M. fümatus. Moreover, it presents an intercalary con-striction in 5. albocaudata (Fig. 2). Cts.- Mf->Sg,Ma->Sa.
X-chromosome - This hétérochromosome has thesame banding pattern in the four species. Myomys fuma-tus presents a different morphology. Cts: Mf-»Sa,Sg,Ma.
The phylogenetic tree constructed on the basis of thecharacter state trees is shown in Figure 5. The tree hasbeen outgroup-rooted on M. fumatus, and it containsno homoplasy. The two species of Stenocephalemysform a monophyletic group, to which M. albipes is thesister group.
Electrophoresis
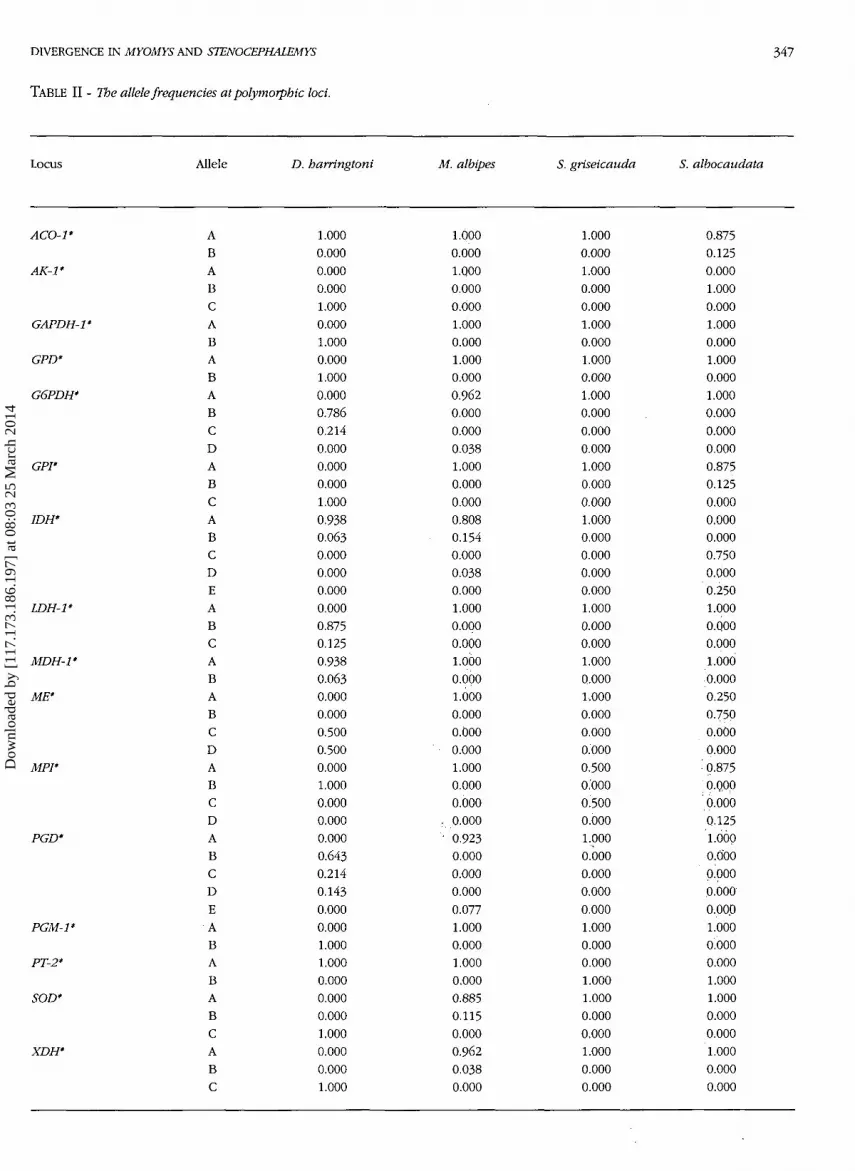
Sixteen out of 25 scored loci are polymorphic. Allelefrequencies are shown in Table II. One locus (PT-2*) isdiagnostic between M. albipes and the two Steno-cephalemys species, showing fixed alleles of differentmobility between the two groups of samples. The spec-imen of S. grisecauda shares the most common allelewith M. albipes at all the remaining loci, while S. albo-caudata samples, although sharing the same allele withM. albipes at 13 out of the 16 polymorphic loci, arecharacterized by private alleles at two loci, i.e. AK-1*and IDH*.
UPGMA dendrogram (not shown) and the consensustree obtained with 1000 data set replicates (Fig. 6)showed different topologies (see Discussion).
DISCUSSION
The Ethiopian species of Myomys and Stenocephale-mys display a considerable amount of difference in thekaryotype with respect to the other Myomys from Soma-lia. Much of it is shared, so that, at least based uponkaryotype rearrangements, a monophyletic hypothesisfor the Ethiopian species is acceptable.
These differences may have a geographic origin. Myo-mys fumatus occurs in the dry, scrubby steppes oflästern Ethiopia, Somalia and northeastern Kenya, whileM. albipes is typical of the highland forests surroundingthe Ethiopian rift, and Stenocephalemys ranges to even
higher altitudes in the Simien and Bale. Geographic iso-lation represents a reasonable cause for the high kary-otypic divergence between Somalian and Ethiopianspecies.
However, there are several karyotype rearrangementsdistinguishing M. albipes from Stenocephalemys, and 5.griseicauda from S. albocaudata. There are no centricfusions/fissions characterizing these karyotypes, usuallyassociated in rodents with fast chromosomal speciation(see King, 1993, for a general review). Rather, it is morelikely that habitat segregation and geographic isolationduring rifting has been the driving force in establishingchromosomal variants. The deepening of the Rift has bi-sected the Ethiopian plateau, preventing gene flow be-tween highland species occurring on its eastern andwestern sides. Moreover, climatic fluctuations since thePleistocene may have altered species' ranges, and theircurrent distribution is likely the result after the postglacial dry phase (Adamson et al, 1980). Chromosomalvariants could have arisen and become fixed in localpopulations, and the current clinal altitudinal distributionmay reflect species adaptation. This has been considereda common phenomenon for other Ethiopian endemics,such as Arvicanthis blicki and A. abyssinicus (Capannaet al., 1996; Corti et al, 1996) and Lophuromysmelanonyx (Corti et al, 1995), and Lophuromys sp. Aand B from Bale Mountains (Aniskin et al., 1997a).
There is a further aspect of the karyotype deservingdiscussion. The diploid and fundamental number of M.albipes has been previously described by Orlov et al(1989). No description of the banding pattern, however,was given nor for the polymorphism of pairs 4 and 5.The origin of the polymorphism remains unsolved, i.e.there is no quantitative information on its occurrenceover the species range which may help in relating it togeography or other causes. It represents, however, avery common phenomenon in rodents.
Apart from the description of the diploid and funda-mental number, and of the banding pattern and AgNORchromosome location, the final question concerns thesystematics and taxonomy of these species, i.e. whetherthe genus Stenocephalemys should be considered para-phyletic or not.
Dow
nloa
ded
by [
117.
173.
186.
197]
at 0
8:03
25
Mar
ch 2
014
DIVERGENCE IN MYOMYS AND STENOCEPHALEMYS 347
TABLE II - The allele frequencies at polymorphic loci.
Locus Allele D. harringtoni M. albipes S. griseicauda S. albocaudata
ACO-1'
AK-1*
GAPDH-1*
GPD*
G6PDH'
GPI"
IDH*
LDH-1*
MDH-1*
ME'
MPI*
PGD*
PGM-1*
PT-2*
SOD*
XDH*
ABABCABABABCDABCABCDEABCABABCDABCDABCDEABABABCABC
1.0000.0000.0000.0001.0000.0001.0000.0001.0000.0000.7860.2140.0000.0000.0001.0000.9380.0630.0000.0000.0000.0000.8750.1250.9380.0630.0000.0000.5000.5000.0001.0000.0000.0000.0000.6430.2140.1430.0000.0001.0001.0000.0000.0000.0001.0000.0000.0001.000
1.0000.0001.Q000.0000.0001.0000.0001.0000.0000.9620.0000.0000.0381.0000.0000.0000.8080.1540.0000.0380.0001.0000.0000.0001.0000.0001.0000.0000.0000.0001.0000.0000.000
..0.000• 0.923
0.0000.0000.0000.0771.0000.0001.0000.0000.8850.1150.0000.9620.0380.000
1.0000.0001.0000.0000.0001.0000.0001.0000.0001.0000.0000.0000.0001.0000.0000.0001.0000.0000.0000.0000.0001.0000.0000.0001.0000.0001.0000.0000.0000.0000.5000.0000.5000.0001.0000.0000.0000.0000.0001.0000.0000.0001.0001.0000.0000.0001.0000.0000.000
0.8750.1250.0001.0000.0001.0000.0001.0000.0001.0000.0000.0000.0000.8750.1250.0000.0000.0000.7500.0000.2501.0000.Q000.0001.0000.0000.2500.750
o.ooo0.000
• 0.8750.Q00
' 0.0000.1251.0000.0000.0000.0000.0001.0000.0000.0001.0001.0000.0000.0001.0000.0000.000
Dow
nloa
ded
by [
117.
173.
186.
197]
at 0
8:03
25
Mar
ch 2
014

348 M. CORTI, A. SCANZANI, A. R. ROSSI, M. V. CIVITELLI, AFEWORK BEKELE, E. CAPANNA
1000
770.5
D. harringtoni
S. ghseicauda
S. albocaudata
M. albipes
Fig. 6 - Consensus tree obtained by discrete character parsimonyanalysis of the allozyme variants. Numbers indicate the percent-age of the 1000-bootsrap replicated in which the taxa attached toeach node occurred among the trees.
Morphology (at least that of the skull) does not helpin solving the puzzle. Fadda & Corti (in press) showedthat even if morphometric relationships support the cur-rent taxonomy, much of the differentiation in shape andsize reflects patterns of adaptation to the very differenthabitats characterising the various altitudinal ranges.However, with morphometry dominated by 'ecologicaleffects', taxonomy based on this kind of character maynot properly reflect systematic relationships.
The chromosome phylogeny in Figure 5 supports thecurrent taxonomy recognizing two genera. The hypoth-esis is also in agreement with the phylogenetic tree(Fig. 6) based upon the presence/absence of the allelesfrom the allozyme analysis. Allozyme data showed thatS. albocaudata samples are genetically well distinctfrom S. griseicauda and M. albipes, as demonstrated bythe presence of loci with private alleles and genetic dis-tance values (D = 0.127). Moreover, the arrangement ofStenocephalemys in the UPGMA dendrogram obtainedon Nei's genetic distances (not shown) shows a closerrelationship between S. griseicauda and M. albipes.However, the value of genetic distances, as well as ofmethods of phenetic reconstruction using them, is ques-tionable when population/species are represented by asingle individual. Thus, the consensus tree, constructedwith an outgroup and not related to allelic frequencies,seems more accurate and thus supports the hypothesisof a monophyletic origin of Stenocephalemys derivedfrom chromosomal data.
There is, however, disagreement with this hypothesisof phylogenetic relationships in Fadda et al. (submitted)who, on the basis of the analysis of a 469 base pairlong 16S rRNA mitochondrial gene fragment, showedthe occurrence of two main clades: one including 5. gri-seicauda and M. albipes, and the other S. albocaudata.On this basis, they concluded that the genus Steno-cephalemys is paraphyletic, as S. griseicauda is phylo-genetically closer to M. albipes than to S. albocaudata.
These different phylogenetic hypotheses challengethe proper use of M. fumatus as the outgroup for chro-mosome analysis, i.e. the polarization of chromosomalchanges. The cladogram in Figure 5 suggests that evolu-tion of this group was accompanied by an increase indiploid number (e.g. from 2n = 36 up to 2n = 54), andby rearrangements such as fissions (e.g. chromosome
9). If one were to accept Fadda et al.'s (submitted) phy-logeny and the pattern of genetic similarities depictedby allozymes (UPGMA), i.e. Stenocephalemys being pa-raphyletic, as representing reliable hypotheses of ances-tral descendant relationships, then character polaritywould change and chromosomal evolution would becharacterised by 2n reduction and arm fusions. This stillleaves the Myomys / Stenocephalemys complex an un-solved systematic puzzle.
REFERENCES
Adamson D. A., Gasse F., Street F. A., Williams M. A. J., 1980 -Late Quaternary history of the Nile. Nature, 288: 50-55.
Afework Bekele, Corti M., 1997 - Forest blocks and altitude as in-dicators of Myomys alpibes (Ruppel 1842) (Mammalia Rodentia)distribution in Ethiopia. Trop. Zool., 10: 295 - 320.
Aniskin V. M., Lavrenchenko A. L., Varshavskii A. A., MilishnikovN. A., 1997a - Karyotypic differentiation of three harsh-furredmouse species of genus Lophuromys (Murinae, Rodentia) fromthe Bale Mountains National Park, Ethiopia. Russ. J. Genet., 33:818-824.
Aniskin V. M., Lavrenchenko A. L., Varshavskii A. A., MilishnikovN. A., 1997b - Karyotypes and chromosomal differentiation oftwo Tachyoryctes species (Rodentia, Tachyoryctinae) fromEthiopia. Russ. J. Genet., 33: 1079-1084.
Bickham J. W., 1979 - Banded karyotypes of 11 species of Ameri-can bats (genus Myotis). Cytologia, 44: 789-797.
Brewer G., Sing C. F., 1970 - Are introduction to isozyme tech-niques. Academic Press, London and New York, 186 pp.
Capanna E., Afework Bekele, Capula M., Castiglia R., Civitelli M.V., Codja J. T. CL., Corti M., Fadda C., 1996 - A multidisciplinaryapproach to the systematics of the genus Arvicanthis Lesson,1842 (Rodentia, Murinae). Mammalia, 60: 677-696.
Corti M., Civitelli M. V., Afework Bekele, Castiglia R., Capanna E.,1995 - The chromosomes of three endemic rodents of the Balemountains, South Ethiopia. Rend. fis. Acc. Lincei, s. 9, 6: 157-164.
Corti M., Civitelli M. N., Castiglia R., Afework Bekele, Capanna E.,1996 - Cytogenetics of the genus Arvicanthis (Rodentia, Muri-dae). 2. The chromosomes of three species from Ethiopia: A.abyssinicus, A. dembeensis and A. blicki. Z. Saugetier., 61: 339-351.
Crother B. I., 1990 - Is "some better than none" or do allele fre-quencies contain phylogenetically useful information? Cladis-tics, 6: 277-281.
Fadda C., Corti M. - Taxonomy and systematics of the EthiopianMyomys - Stenocephalemys complex (Rodentia, Mammalia): anevaluation using three dimensional Procrustes analysis. In M.Corti and L. F. Marcus (eds), Geometric morphometrics in Mam-malogy, Ital. J. Mamm., (in press.).
Fadda C., Corti M., Verheyen E. - Molecular systematics of threeendemic Ethiopian rodent species: Myomys albipes, Steno-cephalemys griseicauda and S. albocaudata and their relation-ships with closely related genera (Murinae, Mammalia). (Sub-mitted)
Felsenstein J., 1993 - PHYLIP (Phylogeny Inference Package) ver-sion 3.5c. Distributed by the author. Department of Genetics,University of Washington, Seattle.
Frick C., 1914 - A new genus and some new species and sub-species of Abyssinian Rodents. Ann. Carnegie Mus., LX, Nos. 1-2.
Harris H., 1966 - Enzyme polymorphism in man. Proc. Roy. Soc.,164: 298-310.
Hillman J. C., 1993 - Ethiopia: compendium of wildlife conserva-tion information. WCI and EWCO, Addis Ababa, 786 pp.
Howell V. M., Black, D. A., 1980 - Controlled silver staining of nu-cleolus organizer regions with a protective colloidal developer:a 1-step method. Experientia, 36: 1014-1015.
Dow
nloa
ded
by [
117.
173.
186.
197]
at 0
8:03
25
Mar
ch 2
014

DIVERGENCE IN MYOMYS AND STENOCEPHALEMYS 349
Hsu T. C., Patton J. L., 1969 - Bone marrow preparation for chro-mosome studies. In: K. Benirsrke (ed.), Comparative mam-malian cytogenetics. Springer-Verlag, New York, pp. 454-460.
King M., 1993 - Species evolution. The role of chromosomechange. Cambridge University Press, pp. 336.
Lavrenchenko L. A., Milishnikov N. A., Aniskin V. M., WarshavskyA. A., Woldegabriel Gebrekidan, 1997 - The genetic diversity ofsmall mammals of the Bale Mountains, Ethiopia. Ethiop. J. Sci.,20: 213-233.
Misonne X., 1969 - African and Indo-Australian Muridae. Evolu-tionary trends. Ann. Mus. Roy. Afr. centr. Sci. Zool., 172: 1-219
Musser G. G., Carleton M. D., 1993 - Family Muridae. In: D. E.Wilson, D. A. M. Reeder (eds), Mammal species of the world. Ataxonomic and geographic reference. Smithsonian Instit. Press.Washington and London, pp. 630-631, 642-644, 664.
Nei M., 1978 - Estimation of average heterozygosity and geneticdistance from a small number of individuals. Genetics, Austin,Texas, 89. 583-590.
Orlov V. N., Bulatova, N. S., Milishinkov A. N., 1989 - Karyotypesof some Mammalian species (Insectivora, Rodentia) in Ethiopia.In: V. E. Sokolov (ed.), Ecological and faunistic studies insouthwestern Ethiopia, pp. 95-109.
Pasteaur N., Pasteur G., Bonhomme F., Catalan J., Britton-Davidi-an J., 1988 - Practical isozyme genetics. Ellis Horwood Limited,215 pp.
Petter F., 1972 - Deux rongeurs nouveaux d'Ethiopie: Steno-
cephalemys griseicauda sp nov. et Lophuromys melanonyx sp.nov. Mammalia, 36: 171-181.
Poulik M. D., 1957 - Starch gel electrophoresis in a discontinuoussystem of buffers. Nature, 180: 1477.
Rüppel E., 1842 - Saugethiere aus der Ordnung der Nager,Beobachtet im Nordoshlichen Afrika. Mus. Senckernberg., 3:99-116.
Seabright M. A., 1971 - A rapid banding technique for humanchromosomes. Lancet, 2: 971-972.
Seiander R. K., Smith M. H., Yang S. Y., Johnson W. E., Gentry J.B., 1971 - Biochemical polymorphism and systematics in thegenius Peromyseus. I. Variation in the old-field mouse (Peromy-seus polionatus). Studies in Genetics VI, Univ. Texas Publ.,7103: 49-90.
Sneath P. A., Sokal R. R., 1973 - Numerical taxonomy. W. H. Free-man and Co, San Francisco, California, 573 pp.
Yalden D. W., 1988 - Small mammals of the Bale mountains,Ethiopia. Afr. J. Ecol., 26: 281-294.
Yalden D. W., Largen M. J., 1992 - The endemic mammals ofEthiopia. Mamm. Rev., 22: 115-150.
Yalden D. W., Largen M. J., Kock D.. 1976 - Catalogue of themammals of Ethiopia 2. Insectivora and Rodentia. Mon. Zool.Ital. (N. S.), Suppl. 8: 1-118.
Yalden D. W., Largen M. J., Kock D., Hillman J. C., 1996 - Cata-logue of the mammals of Ethiopia and Eritrea. 7. Revised check-list, zoogeography and conservation. Trop. Zool., 9: 73-164.
Dow
nloa
ded
by [
117.
173.
186.
197]
at 0
8:03
25
Mar
ch 2
014





























