Embed Size (px)
Citation preview


LA AVIFAUNA DE LOS BOSQUES DE TRANSICION DEL NOROESTE
DE ARGENTINA
Patricia Capllonch
Trabajo de Tesis presentado a la Facultad de Ciencias Naturales e Instituto
Miguel Lillo, Universidad Nacional de Tucumán, para optar por el título de
Doctora en Ciencias Biológicas orientación Zoología: Director Dr. Ricardo
Alberto Ojeda
1997
San Miguel de Tucumán, Argentina

INDICE
Resumen…………………………………………………………………………….. 4
Abstrack……………………………………………………………………………… 5
Introducción…………………………………………………………………………. 5
Objetivos……………………………………………………………………………..
Introducción a la biogeografía de la Avifauna Neotropical…………………………
Teoría de los Refugios Pleistocénicos……………………………………………….
Historia biogeográfica del Norte de Argentina……………………………………..
Antecedentes zoogeográficos de las Yungas……………………………………….
Composición y distribución de la avifauna de las Yungas………………………….
Grado de endemismo……………………………………………………………….
Variación latitudinal………………………………………………………………….
Materiales y Métodos…………………………………………………………………
Localidades de estudio………………………………………………………..
Inventario de las aves…………………………………………………………..
Monitoreo………………………………………………………………………
Análisis zoogeográfico de afinidades…………………………………………..
Censo de aves nidificantes…………………………………………………….
Estudio de la vegetación………………………………………………………
Estructura trófica de las comunidades………………………………………..
Diversidad de macronichos…………………………………………………..
Biomasa………………………………………………………………………..
Dominancia………………………………………………………………………
Área de estudios intensivos………………………………………………………
Localidad de La Junta……………………………………………………………
Localidad de Dique San Ignacio………………………………………………..
Localidad de Piedra Tendida…………………………………………………..
Resultados……………………………………………………………………………….
Estructura y composición florística de la vegetación……………………………………
Composición y distribución de la avifauna de los bosques de Transición de Argentina…
Diversidad de especies……………………………………………………………………..
Dominancia…………………………………………………………………………………
Análisis zoogeográfico……………………………………………………………………
Distribución de taxones de aves………………………………………………………….
Características poblacionales locales y distribución latitudinal…………………………..
La estacionalidad y su influencia en la avifauna………………………………………….
Dinámica de especies de aves……………………………………………………………..
Migrantes Neárticos……………………………………………………………………….
Migrantes Neotropicales……………………………………………………………………
Migrantes desde el sur de Argentina………………………………………………………….
Migrantes altitudinales o locales……………………………………………………………..
Formación de bandadas mixtas de aves……………………………………………………..
Organización trófica de la avifauna………………………………………………………….
Categorías tróficas………………………………………………………………………
Relación entre las tácticas alimenticias y la biomasa…………………………………
Diversidad de macronichos……………………………………………………………..
Reproducción…………………………………………………………………………
Muda…………………………………………………………………………………..
Discusión………………………………………………………………………………..
Listas de aves……………………………………………………………………………

RESUMEN
Entre 1984 y 1994 realicé estudios de campo sobre las comunidades de aves de los Bosques
Pedemontanos del noroeste de Argentina desde el límite con Bolivia hasta la provincia de
Catamarca. Estos bosques permanecían desconocidos hasta ese momento en cuanto a su
composición taxonómica y dinámica de especies y debido a su acelerada destrucción era
inminente su riesgo de extinción. Como resultado de estos estudios, y de la comparación con
bosques chaqueños y selvas montanas lindantes, llegué a la conclusión de que el Bosque
Pedemontano, Selva Pedemontana o Selva de Transición según los autores, es un bosque con
características ornitológicas propias. No es una simple área ecotonal entre las asociaciones
xerófilas de las planicies y las selvas húmedas de las laderas montañosas. Son además,
bosques imprescindibles para la conservación de una rica avifauna debido a la importancia
que tienen para especies de aves migratorias. Esto es el resultado de su ubicación altitudinal
al pié de altas montañas y su rango de distribución latitudinal (entre los 21º y los 28º de
latitud sur), coincidente con la ruta migratoria de numerosas especies de aves de Argentina.
La avifauna del Bosque Pedemontano de Transición y la de la Selva Montana poseen un
número importante de especies compartidas, aunque con dominancia, abundancia relativa y
constancia de especies diferente. La mayor parte de los géneros de aves perteneces al
Dominio Amazónico y son propios de bosques tropicales húmedos. Muchas especies
chaqueñas ingresan con la alteración de este bosque, pero están prácticamente excluidas
cuando el bosque conserva su estructura de bosque alto con diversidad de estratos.
La principal característica del Bosque Pedemontano es su marcada estacionalidad, seco entre
Mayo y Octubre y casi completamente deciduo entre Junio y fines de Septiembre.
Típicamente existe una contingencia anual con una estación caliente y húmeda y una
templada y seca. La llegada de las lluvias y la abundancia de recursos, especialmente frutos,
impone en la avifauna un ritmo reproductivo anual estacional, similar en su dinámica al que
ocurre en bosques templados. Esto determina también que una parte importante de especies
de aves realicen desplazamientos migratorios y se produzcan asociaciones interespecíficas en
bandadas durante la época de escasez. Esta dinámica compleja es contingente, repitiéndose
cada año de forma casi exacta el arribo de migrantes y el inicio de la temporada de cría para
cada especie en particular.
La mayor parte de las especies cría una sola vez al año, inclusive las residentes, y son
omnívoras durante el otoño e invierno. Estas afirmaciones se basan en las capturas de más de
4000 aves en 25 localidades, en las cuales realicé un estudio del plumaje, especialmente
mudas, condición reproductiva y peso, lo que me permitió establecer el ritmo anual de cada
especie.
Se registraron 357 especies para las Yungas de Argentina y 215 especies para los Bosques
Pedemontanos de Transición, de las cuales 83 eran residentes y por su constancia caracterizan
a estos bosques en todo su rango de distribución. Se registraron dos especies nuevas para la provincia de Tucumán, Elanoides forficatus y Todirostrum plumbeiceps. Se aumentó la
distribución de Pyrrhura molinae, Psaracolius decumanus y Atlapetes torquatus hasta el
límite entre Salta y Tucumán. Se registraron especies raras, poco conocidas o con prioridad
de conservación como Oroaetus isidori, Tigrisoma fasciatum y Aegolius harrisii en varias
localidades. Del total de especies frecuentes, 130 ocurren en todo el rango de extensión de
estos bosques. Un 12% de ellas llegan solo hasta el sudeste de Jujuy, tienen su límite sur de
distribución con el bosque de Palo blanco y no alcanzan al de Tipa y Pacará (Cabrera 1976).
La composición de especies de aves en los Bosques Pedemontanos varía latitudinalmente,
produciéndose un empobrecimiento de especies de norte a sur, esta caída de especies se
produce junto con la desaparición paulatina hacia el sur del Bosque Pedemontano de palo
blanco (Calyicophylum multiflorum) y su reemplazo por el Bosque Pedemontano de tipa y
pacará (Tipuana tipu y Enterolobium contortisiliquum).

ABSTRACT
Between 1984 and 1994 I conducted field studies on bird communities of the piedmont
forests of northwestern Argentina from the border with Bolivia to the province of Catamarca.
These forests remained unknown so far in their taxonomic composition and dynamics of
species, and due to its accelerated destruction was imminent risk of extinction. As a result of
these studies, and compared to neighboring Chaco forests and montane forests, I came to the
conclusion that the Bosque Pedemontano or Selva Pedemontana de Transition, according to
the authors, is a forest with ornithological own characteristics. It is not a simple ecotonal area
between the xeric forest associations of the plains and rain forests of the mountainsides. They
are also essential for the preservation of a rich bird life due to the importance for migratory
bird species. This is the result of their altitudinal location at the foot of high mountains and
latitudinal distribution range (between 21 and 28 degrees south latitude), coinciding with the
migratory route for many species of birds of Argentina.
The avifauna of Bosque Pedemontano de Transición and Selva Montana have a number of
shared species, although they have different dominance, relative abundance and regularity of
species. Most of the genera of birds belong to the Amazonian domain and are typical of
tropical rain forests. Many species invade the Chaco forest where extraction occurs, but are
practically excluded when the forest retains its structure with high diversity of forest strata.
The main feature of piedmont forest is its seasonality, dry between May and October and
almost completely deciduous between June and late September. There is typically an annual
contingency with a hot and wet season and a warm and dry. The arrival of rains and abundant
resources, especially fruits, imposes seasonal annual reproductive rate, similar in its dynamics
to what occurs in temperate forests. This also determines that an important part of species of
birds make migratory movements and interspecific associations occur in flocks during the
lean season. This complex dynamics is contingent, repeating every year almost exactly the
arrival of migrants and the beginning of the breeding season for particular species.
Most species breed once a year, including residents, and are omnivorous during the fall and
winter. These statements are based on catches of more than 4000 birds in 25 locations, where
I conducted a study of plumage, especially molt, weight and reproductive status, which
allowed me to set the annual rate of each species.
I recorded 357 species to the Yungas of Argentina and 215 species for Bosques
Pedemontanos de Transition, of which 83 were residents, and its constant presence
characterize these forests throughout their range.
Two new species for the province of Tucuman, Elanoides forficatus and Todirostrum
plumbeiceps were recorded. The distribution of Pyrrhura molinae, Psaracolius decumanus
and Atlapetes torquatus was increased to the limit between Salta and Tucuman. Rare or little
known species or with conservation priority were recorded, as Oroaetus isidori, Tigrisoma
fasciatum and Aegolius harrisii in several locations. Of all common species, 130 occur
throughout the range extension of these forests. 12% of them come just to the southeast of
Jujuy, has its southern limit of distribution in Palo blanco forest and do not reach the Tipa and
Pacará forest (Cabrera 1976).
The composition of species of birds in the Bosques Pedemontanos varies latitudinally,
resulting in a depletion of species from north to south, the decline of species occurs along
with the gradual disappearance south piedmont forest of palo blanco (Callycophylum
multiflorum) and replacement the piedmont of tipa and pacará Forest (Tipuana tipu and
Enterolobium contortisiliquum).
INTRODUCCION
Los Bosques Subtropicales de Transición (Hueck 1978) se extienden aproximadamente desde
los 21º en Santa Cruz de la Sierra, Bolivia, hasta los 28º en los cordones montañosos que
limitan las provincias de Tucumán y Catamarca en Argentina. Su distribución es irregular a

lo largo de los pedemontes orientales ocupados por las Yungas (Fig. 1), pertenecientes al
Dominio Amazónico (Cabrera y Willink, 1980). Prado (1995), estableció vínculos florísticos
de la Selva Pedemontana con la Caatinga, en el nordeste brasileño, y con la provincia Guajira
del Nordeste de Colombia y Venezuela. De esta manera, ratificó, la ubicación de la Selva
Pedemontana o Selva de Transición dentro del Dominio Amazónico.
.
Fig. 1: Distribución de los Bosques Pedemontanos de Transición en el Noroeste de Argentina
Las denominaciones de este bosque varían según los autores y han causado confusión. Hueck
(1951), lo denomina Distrito de las Selvas de Transición, distinguiendo la Selva de
Transición de Palo blanco y Palo amarillo, que se extiende desde el límite con Bolivia hasta
Jujuy, y la Selva de Transición de Tipa y Pacará en el sur de Salta y Tucumán. Meyer
(1963), lo incluye dentro de la Selva Basal, de características decíduas, distinguiéndola de la
Selva de Mirtáceas, con predominio de perennifolias, que se encuentra a mayor altura.
Cabrera (1976) los ubica dentro de la Provincia Fitogeográfica de las Yungas, con la
denominación de Distrito de las Selvas de Transición, distinguiendo además el Distrito de las
Selvas Montanas y el Distrito de los Bosques Montanos. Vervoorst (1979) los incluyó en los
Bosques Pedemontanos, denominando Bosque Montano Inferior y Bosque Montano Superior
a los pisos superiores de vegetación. Limitan al este con el Distrito Occidental de la Provincia
Fitogeográfica Chaqueña, bosques de características xerófilas con dominio de géneros como
Prosopis, Acacia y Schinopsis (Cabrera 1976, Hueck 1978). Fue considerada por Cabrera
como una zona transicional entre el chaco y la selva. Existe una homogeneidad de su
composición florística a lo largo de su rango latitudinal (Brown, 1986), aunque con dos áreas
de mayor riqueza debido a las abundantes precipitaciones. Estas son las áreas del Parque
Nacional El Rey y la del sur de Tucumán, con precipitaciones superiores a los 2000 mm en
algunos años (Bianchi 1981). Es un bosque empobrecido en el número de especies de la flora,
respecto al de Palo blanco y Palo amarillo de Salta y Jujuy. Según Digilio y Legname
(1966), de las 166 especies arbóreas presentes en las Yungas, 81 habitan desde la frontera con
Bolivia hasta Tucumán, 20 no se encuentran en el sector sur (sur de Salta y Tucumán) y solo
3 son endémicas de este sector.
En general, los Bosques Pedemontanos de las Yungas del noroeste de Argentina son poco
conocidos, aunque tratados en forma descriptiva por algunos autores (Cabrera, 1976;
Vervoorst, 1969, 1979). Los bosques de Tipa y Pacará de la porción más austral de su

extensión son mejor conocidos (Hueck 1954, Meyer 1963, Digilio y Legname 1966, Ayarde
1995). Sus relaciones florísticas, probable origen y características de la vegetación fueron
tratadas por Prado (1995). Su estructura y dinámica ha sido estudiada en una parcela de la
Reserva Provincial de La Florida, Tucumán (Ayarde 1995). Las Selvas y Bosques Montanos
han recibido más atención de los autores (Hueck 1954, Meyer 1963, Cabrera 1976, Grau
1985, Brown 1986, Slanis 1990, Aceñolaza 1991).
Respecto a los Bosques Montanos que se desarrollan por encima, la diferenciación en dos
pisos de vegetación es fisonómica y estructural, no tanto florística (Brown et al. 1985,
Cabrera 1976, Meyer 1963). De acuerdo con Brown et al. (1985), de 40 especies arbóreas
identificadas en 5 hectáreas muestreadas en el Parque Nacional El Rey, el 68% se encuentra
en ambos pisos. Las diferencias mas notables entre ambos pisos la produce la dominancia de
árboles caducifolios y sotobosque enmarañado y muy denso en el piso inferior, y la
abundancia de árboles peremnifolios de la familia Mirtaceae (Myrcianthes mato, M. pungens,
Stenocalix uniflora, Mirrhinium loranthoides) y el escaso desarrollo del estrato arbustivo en
el piso superior.
Respecto a la avifauna, no existe en la bibliografía un estudio completo sobre la estructura y
composición de las comunidades de aves de los Bosques Pedemontanos de Transición, ya que
los estudios existentes (Olrog, 1969; Vides, 1992) no discriminan lo que ocurre en todo el
largo de la angosta franja meridional ocupada por estos bosques. Un análisis biogeográfico
sobre la distribución de especies de aves en los bosques tropicales, subtropicales y templados
fue realizado en forma cualitativa por Olrog (1969). Este mismo autor destacó, sin embargo,
la presencia de algunas especies particularmente en el Bosque de Transición (Olrog, 1978).
Carece también la bibliografía existente de trabajos comparativos de comunidades de aves de
los bosques del noroeste como los realizados para bosques tropicales y templados (Orians,
1969; Karr, 1971), salvo el de Short (1975) para el Chaco y el de Vides Almonacid (1992),
para taxocenosis de aves de la Sierra de San Javier, Tucumán.
Por otro lado, el conocimiento de la distribución geográfica y temporal de aves migratorias en
el sur de Sudamérica es mucho menos completa que para otras áreas del continente
(Dabbene 1910, Fitzpatrick 1980, Myers 1980), y generalmente referida a migrantes desde el
Hemisferio Norte que invernan en el extremo sur de Sudamérica. Los patrones ecológicos de
la distribución espacial y temporal de migrantes en el noroeste de Argentina son
prácticamente desconocidos en la bibliografía, salvo los trabajos de Olrog (1962, 1968, 1969,
1971), pero en ninguno de ellos se destaca la importancia de estos bosques para especies
migratorias. La proporción de especies migratorias y su relación con los residentes no ha
sido aún discutida para las Yungas de Argentina. Existe sin embargo extensa bibliografía
sobre la selección de hábitats boscosos por parte de migrantes para ambientes tropicales
(Karr, 1976; Pearson 1980, Terborgh y Faaborg, 1980; Willis, 1966).
En cuanto a las relaciones entre las especies de aves, bandadas mixtas de especies que
forrajean juntas han sido descriptas para áreas templadas y tropicales (Austin y Smith, 1972;
Moynihan, 1979; Powell, 1985). Las razones o ventajas de su formación fueron interpretadas
de modo diferente por distintos autores, figurando entre estas la reducción a la agresión
interespecífica (Austin y Smith, 1972), disminución al riesgo de predación (Buskirk, 1976), la
explotación del alimento de la manera más económica posible (Rappole y Warnes, 1980) o
por aprendizaje observando a otros individuos. Según Moynihan (1979), las ventajas de cada
especie miembro de la bandada son variables pero la detección de los predadores y el mejor
aprovechamiento del alimento se ve incrementado en beneficio de todas. Aunque bandadas
mixtas son comunes en áreas boscosas de diferentes latitudes (Morse, 1970), es en los
trópicos donde alcanzan su máximo desarrollo y diversidad (Hutto, 1987; Powell, 1985). El
número promedio de especies que las componen varía según los lugares: 4.7 y 18.6 en
bosques del Oeste de México (Hutto, 1987); 3.5 en las Yungas en el Norte de Bolivia y 5.24
en las laderas Este del Machu Pichu (Moynihan, 1979). No existen estudios comparativos de

los que ocurre en las Yungas de Argentina respecto a las asociaciones de aves y su relación
con la variación temporal de los bosques donde habitan, aunque sí aportes puntuales.
La estructura trófica de una comunidad de aves y los patrones del modo en que dichas
especies explotan los recursos, son utilizadas para indicar como, donde y que clase de
alimentos provee dicho hábitat (Holmes y Recher, 1986). Las especies de aves fueron a
menudo asignadas a diferentes categorías tróficas según un conocimiento general de sus
hábitos alimenticios. Pocos estudios han identificado las estructuras tróficas de modo
objetivo o evaluado la relativa importancia de los factores que las definen, y de qué modo
determinan la estructura del ensamble de aves que la componen (Cody, 1974; Holmes et al,
1979; Holmes y Recher, 1986; Sabo y Holmes, 1983).
OBJETIVOS
Los objetivos planteados para la realización de este trabajo fueron:
1-Estudiar la composición específica y estructura de comunidades de aves de los Bosques
Pedemontanos de Transición del noroeste de Argentina.
2-Analizar la dinámica de especies de aves en estos bosques en relación con la oferta
estacional de recursos, los patrones migratorios y las conexiones con otras unidades
biogeográficas neotropicales.
3-Analizar la caída de especies desde el norte de Salta hasta Catamarca, considerando datos
históricos de distribución zoogeográfica y datos obtenidos durante este estudio.
De esta forma he pretendido contribuir al conocimiento de la estructura y funcionamiento del
Bosque Pedemontano de Transición, casi desconocidos desde el punto de vista científicos,
antes de su inminente desaparición.
INTRODUCCION A LA BIOGEOGRAFIA DE LA AVIFAUNA NEOTROPICAL
La avifauna Neotropical es la más rica del mundo, alcanza su mayor diversidad en la
Amazonia de Perú, Bolivia y Brasil, al pie de los Andes. Desde allí decrece en dirección al
Sur y al Este. Sudamérica cuenta con unas 3.000 especies de aves, si se tiene en cuenta los
migrantes desde América del Norte, pertenecientes a 900 géneros y más de 90 Familias, de
las cuales 30 son endémicas de la Región Neotropical y 11 para Sudamérica exclusivamente.
Desde el punto de vista zoogeográfico, los grupos más interesantes de aves Neotropicales son
aquellos restringidos a Centroamérica y Sudamérica (Olrog, 1968; De Schauensee, 1970).
Las Familias endémicas para Sudamérica son: Rheidae, Anhimidae, Opisthocomidae,
Psophidae, Thinocoridae, Cariamidae, Steatornithidae, Conopophagidae, Phytotomidae,
Oxyurincidae y Catablyrhynchidae, representadas por uno o dos géneros o por pocas especies
y de distribución limitada, por lo que deben ser consideradas como relictos de faunas
antiguas. El número de especies se incrementa a 3.300 si consideramos las de América
Central (De Schauensee, 1970).
Las razones para tal diversidad son en parte debidas a las características fitogeográficas de
Sudamérica, dividida en grandes extensiones de bosques tropicales de tierras bajas, zonas
áridas de la cosa Pacífica y en centro y sur del continente y zonas montañosas de los Andes.
Las Familias de más amplia distribución, tanto en Norte como en Sudamérica son:
Trochilidae, Tyrannidae, Thraupidae e Icteridae. La mayoría de los miembros de las Familias
Troglodytidae, Mimidae, Vireonidae, Parulidae y Momotidae, se encuentran tanto en Centro
como en Sudamérica. Algunas Familias poseen distribución mundial en los trópicos como:
Psittacidae, Trogonidae, Capitonidae, Columbidae, Phasianidae, Cuculidae, Corvidae,
Turdidae, etc. Otras son cosmopolitas como Strigidae, Falconidae, Accipitridae, Picidae e
Hirundinidae (Olrog, 1969).
En cuanto a la Región Amazónica, de las 600 o 650 especies de aves, unas 400 son
endémicas (Haffer, 1982), muchas de las cuales forman superespecies con numerosas
haloespecies (Mayr, 1963; Vuilleumier, 1969). Los miembros o haloespecies de una
superespecie se reemplazan unos a otros geográficamente, pudiendo ser alopátridas

(separadas) o parapátridas (estar en contacto). Las haloespecies se excluyen unas a otras por
ser (todavía) fuertes competidoras ecológicas, cercanas en hábitos y fenotipos. La ausencia
de rasgos físicos que demuestren hibridación es considerada como un criterio para establecer
las formas parapátridas. Entre algunos grupos que poseen superespecies están las Familias
Cracidae (Vuilleumieur, 1965), Coerebidae (Moynihan, 1979), Picidae (Short, 1972) y
Ramphastidae (Haffer, 1974 b).
Teoría de los refugios Pleistocénicos
La causa de tan alta diversidad ha tratado de ser explicada por diversos autores. Estudios
zoogeográficos de la distribución de especies y estructura de las poblaciones de aves,
lagartijas y mariposas en la Amazonia, sugieren que muchas especies y subespecies se
originaron a partir de pequeñas poblaciones durante varios períodos de aislamiento
geográfico-ecológico (Haffer, 1974 a). Repetidas fluctuaciones climáticas durante el
Pleistoceno habrían llevado a vastos cambios en la vegetación que cubría Sudamérica.
Numerosas zonas de contacto secundario, como las anteriormente mencionadas (especies
parapátridas), indican la existencia anterior de barreras ecológicas en la Amazonia que luego
desaparecieron. Estas zonas de contacto bien definidas, indican áreas donde las faunas se
superponen y funden como en la Amazonia Central, al sur del Río Amazonas y noroeste de
Colombia.
La avifauna tan variada de las montañas de la zona tropical de Sudamérica probablemente
surgió a partir de formas inmigrantes de las tierras bajas a fines del Terciario y hacia
comienzos del Pleistoceno (Chapman, 1926). Las aves que colonizaron las planicies altas
probablemente inmigraron desde el sur de Sudamérica (Chapman, 1926). Los efectos de las
fluctuaciones climáticas durante el Pleistoceno (últimos 800.000 años), probablemente
causaron una inmensa diferenciación de esta avifauna montana original y llevaron al
desarrollo de numerosas especies y géneros nuevos en las faunas de aves montanas, y
probablemente causaron la extinción de formas intermedias (Haffer, 1970). Los hábitats de
las aves montanas fueron divididos en numerosas porciones pequeñas a lo largo de las altas y
aisladas montañas de los Andes durante los períodos secos y cálidos de los interglaciales.
Las diferentes especies habrían surgido en muchas de las pequeñas porciones montañosas
aisladas (Haffer, 1970). Durante los períodos glaciales húmedos y fríos, las zonas de vida
montanas se localizaron mucho más abajo, a lo largo de las laderas montañosas, haciéndose
más continuas y facilitando la extensión del rango de esas nuevas especies. Este proceso se
repitió varias veces durante el Pleistoceno, influyendo marcadamente las condiciones
ambientales en los trópicos y causando numerosas extinciones en Sudamérica. En las
montañas los pisos de vegetación o zonas de vida fueron comprimidos y expandidos
verticalmente repetidas veces (Haffer, 1969), durante los períodos cálidos y fríos, causando
cambios en las tierras bajas en la distribución de selvas y áreas abiertas. La continuidad que
actualmente vemos en la selva Amazónica parece ser un estadío reciente y temporario en la
historia de la vegetación en Sudamérica. Durante los cambios climáticos pleistocénicos, la
selva amazónica fue subdividida en pequeños bosques aislados unos de otros por vegetación
abierta (Haffer, 1969, 1970, 1974; Vuilleumier, 1969), que sirvieron de refugio para
numerosas poblaciones que derivaron luego en nuevas especies. Las áreas de refugio se
expandieron nuevamente durante períodos húmedos, permitiendo que las nuevas especies
extendieran sus áreas de distribución. Analizando los patrones actuales de distribución de las
aves del dominio amazónico, muchas tienen un rango de distribución restringido, mientras
que otras poseen distribución amplia. Existen centros geográficos con números altos de
especies de distribución restringida al este de Ecuador, sureste de Perú y las Guayanas, y
número decreciente de especies de este tipo hacia el sur del Brasil y hacia la costa norte de
Sudamérica en Colombia. Pueden distinguirse varios centros de endemismo (Haffer, 1982,
Ridgely y Tudor, 1989) (Fig. 2), cada uno caracterizado por 10 a 50 especies. Los grandes
ríos amazónicos hacen de barrera para la distribución de muchas aves. Pero en muchos casos

el aislamiento surgió como producto de la competencia interespecífica y no de la
imposibilidad de cruzar los ríos (Willis, 1976).
Fig 2-Principales centros de endemismo en Sudamérica. A) Centros de endemismo de tierras
bajas, B) Centros de endemismo montanos.
Varios grupos endémicos penetran en los bosques de la costa Atlántica del sur de Brasil,
formando también un importante centro de endemismo, otros pocos grupos continúan por
Bolivia hasta las provincias del norte argentino, donde los endemismos, como veremos más
adelante, son relativamente escasos. En cambio, sí existen numerosos casos de especies
endémicas en la región del sur del Brasil, este de Bolivia, noreste de Argentina y Paraguay.
Estas son propias de áreas abiertas y bosques xerófilos. Hay dos principales centros de
endemismo, uno al sureste de Brasil y otro en las planicies altas del noreste de Argentina, y
en Paraguay y este de Bolivia, comprendiendo el llamado gran Chaco (Short, 1975).
Resumiendo, la vasta región de los bosques tropicales y subtropicales sudamericanos está
caracterizada, desde el punto de vista ornitogeográfico por tres rasgos fundamentales:
a)Existen áreas geográficas con alto grado de endemismo, con numerosas especies de rangos
restringidos.
b)Hay poblaciones disyuntas de especies de poca capacidad de dispersión o especialistas,
separadas por barreras geográficas o de distinto tipo de vegetación.
c)Hay zonas de contacto o hibridación conspicuas entre los centros de endemismo de
especies.
Estos patrones biogeográficos de la teoría de Haffer prueban que la avifauna pleistocénica fue
influida por cambios geográficos de clima y vegetación que determinaron el aislamiento y el
surgimiento de nuevas especies. Aún cuando puedan discutirse algunos aspectos ecológicos
de la teoría, esta ayudó a establecer la historia biogeográfica de parte de Sudamérica,
determinando cuando y porqué se produjo la enorme diversidad de aves de la Amazonia y
bosques andinos.
Historia biogeográfica del norte de Argentina
Resulta difícil establecer con claridad la cronología de los fenómenos glaciales ocurridos en
el noroeste de Argentina debido a la escasez de datos. Sólo se hicieron tentativas generales,

pero no integradas a los resultados de las investigaciones más nuevas, como ocurre con las
áreas mejor conocidas de los Andes Patagónicos. Es posible que sólo pueda definirse con
precisión una época glacial para el noroeste, ubicada a fines del Pleistoceno Superior
(Aceñolaza y Toselli, 1981), época en la que se produjeron englazamientos en los núcleos
montanos por arriba de los 3.000 m de altura.
Fig. 3: Panorama de las Sierras de Tucumán vista desde la ciudad de San Miguel deTucumán,
tomado de Kuhn (1924).
La zona de las Sierras del Aconquija-Cumbres Calchaquíes (Fig. 3), puede ser considerada
como una de las principales zonas de glaciares durante el Cuaternario, pudiendo distinguirse
sus efectos, en la provincia de Tucumán, en los cerros Del Candado, Overo y Pabellón
(Kuhn, 1924; Kuhn y Rohmeder, 1943). Esta etapa glacial, según estos autores, cabría dentro
del período de la máxima glaciación de los Andes o el de la última glaciación con sus
diferentes fases. También se observan signos de glaciación en los nevados de Cachi, en un
amplio sector que va desde la Quebrada de las Arcas en el Sur, hasta el cerro Ciénaga Grande
en los Nevados de Palermo. Sin embargo, la magnitud de estas manifestaciones glaciales
sería menor que la anterior de la provincia de Tucumán. También se observa una morfología
glacial en los nevados de Acay y Chañi, Sierra de Aguilar y de Santa Victoria, y sobre todo el
ambiente puneño por sobre los 5.000 m de altura. La etapa glacial sería, entonces,
relativamente nueva en Argentina, alcanzando su mayor desarrollo en los cordones más altos
del Aconquija y Cumbres Calchaquíes.
Existen evidencias de oscilaciones climáticas, húmedas y secas, en el sur de Bolivia.
Analizando yacimientos Paleontológicos, Marshal et. al. (1984) establecieron que la
formación Ñapua I de Tarija, del Pleistoceno Medio, se acumuló en clima seco de llanura y
contiene numerosos géneros de megafauna actualmente extinta, mientras que Ñapua II, de
finales del Pleistoceno y comienzos del Holoceno, se acumuló durante un período húmedo de
inundación donde actualmente es un hábitat chaqueño. La diferencia entre Ñapua I y Ñapua
II es notable, con la desaparición de grandes mamíferos y la aparición de fauna
contemporánea de menor tamaño.
Registros paleontológicos de Argentina, Brasil y Uruguay (Bombin, 1975) demuestran que la
megafauna de las llanuras desapareció primero de las zonas templadas de Argentina y
sobrevivieron hasta comienzos del Holoceno en refugios tropicales de Brasil. En la
Amazonia había áreas de vegetación abierta o semiabierta, donde abundaban Toxodon,
Pampatherium, un équido y un cérvido durante el Pleistoceno. Luego el clima cambió,
predominando condiciones más secas a finales del Pleistoceno, permaneciendo esta fauna en
refugios de bosque. Géneros como Paleolama y Scelidodon desaparecieron mucho antes de
Argentina. El paisaje se componía de un mosaico de pastizales, sabanas y vegetación
xerófila, entremezclados con bosques húmedos cerca de los cursos de agua. A partir de este
período árido comenzó una época gradualmente más húmeda en el Holoceno, durante la cual
los bosques húmedos se extendieron, situación que sigue ocurriendo actualmente.
Los patrones de distribución actual de la avifauna en el norte de Argentina revelan estas
fluctuaciones pleistocénicas y holocénicas (Vuilleumier, 1969). La presencia de géneros,
especies y subespecies en común en áreas actualmente separadas, como las selvas del noreste

y noroeste, por una barrera de vegetación xerófila, manifiestan que hubo intercambio
faunístico y actualmente presentan, ya sea distribuciones disyuntas o zonas de contacto
secundario o hibridación. Como ejemplos, la Familia Turdidae posee varias especies en el
norte, cada una dominando prácticamente todos los tipos de vegetación en las zonas
montañosas y de llanura; Turdus chiguanco en pastizales por arriba de los 2.000 m; T.
nigriceps en Bosques Montanos; T. rufiventris en Selvas Montanas; T. amaurochalinus en
Bosques Chaqueños. De estas especies, T. nigriceps y T. albicollis poseen distribución
disyunta con subespecies habitando los bosques del NW y NE. Turdus albicollis contemptus
fue encontrada por Olrog (1972) en Bosque de Transición en el norte de la provincia de Salta,
y habita además en el centro y sur de Bolivia; T. a. paraguayensis se encuentra en Misiones,
este de Paraguay y sur y sureste de Brasil. Turdus nigriceps cabanis habita los bosques
húmedos de altura de Jujuy, Salta, Tucumán y Catamarca, además de zonas serranas de
Córdoba y T. subalaris en bosque húmedos en serranías de Misiones, este de Paraguay y
sudeste de Brasil. La distribución actual disyunta denota una vinculación anterior entre
ambas regiones, desde el punto de vista de la flora, hoy separadas por el Bosque Chaqueño.
La presencia de T. n. nigriceps en la provincia de Córdoba indica una vinculación anterior
más estrecha entre esta zona y la del Aconquija. Este tipo de distribución aparece también en
otros grupos con escasa capacidad de migración (considerando la posibilidad de que las
especies nombradas anteriormente pudieron haber migrado), como la de la Familia
Formicaridae. Las especies Batara cinerea y Thamnophillus ruficapillus, típicas del Bosque
de Transición, presentan casos similares de dispersión. En la revisión del status de Pionus
maximiliani, Smith (1960), estableció una zona de contacto secundario, con formas que
denotan una intergradación entre P. m. lacerus y P. m. siy en la provincia de Tucumán, a lo
largo de la zona chaqueña del este de la provincia.
Antecedentes zoogeográficos de las Yungas
Dabbene (1910) dividió a las Yungas en Región del Parque y Región de los Bosques Subtropicales hasta los 1200 m, y Región del Aliso y de los Prados Andinos.
Cabrera et al. (1947), consideraron a las Yungas como Distrito Subtropical de la Subregión
Guyano-Brasileña.
Olrog (1959), siguiendo a Cabrera, subdividió a la Argentina en cuatro zonas zoogeográficas:
Subtropical, Chacopampeana, Andina y Subatlántica. En la Zona Subtropical distingue una
Subzona Subtropical Oriental y una Subzona Subtropical Occidental. Esta última
comprende, según el autor: a) los cerros boscosos del norte de Salta y este de Jujuy, con
selvas de avifauna rica y poco conocida; b) cerros boscosos del sur de Salta y oeste de
Tucumán, más pobre que la anterior; y c) alisales entre 1800 y 2500 m de altura en los cerros,
área pobre en aves, aunque con algunas especies típicas. Para la Subzona Subtropical
Occidental agregó "que sus elementos se mezclan, particularmente con los de la Subzona
Andina y Chacoandina, presentando con frecuencia un panorama peculiar y confuso",
admitiendo que es común a la fauna del sur de Bolivia.
Cabrera y Willink (1973) denominan a las regiones boscosas húmedas del Noroeste como
Provincia Biogeográfica de las Yungas, con una fauna difícil de caracterizar debido a la
mezcla de elementos con la Provincia Amazónica, Chaqueña y de la Prepuna.
Olrog (1984), consideró seis Zonas Ornitogeográficas para las aves argentinas: a) Estepas y
matorrales andinos, b) Matorrales áridos occidentales, c) Estepas húmedas, d) Chaco, e)
Selvas y Bosques húmedos y f) Litoral Atlántico y de alta mar. En las Selvas y Bosques
húmedos distingue las del NO, las del NE y las Australes o Patagónicas.
Olrog y Capllonch (1986), mantuvieron la clasificación de Olrog (1984) caracterizándola con
algunos elementos típicos de las Selvas del NW como: Merganetta armata berlepschi,
Tigrisoma fasciatum, Cinclus schultzi, además de varias especies del Género Penelope, dos
de Columba, y varias de la Familia Psittacidae como Ara caninde y Aratinga mitrata. En ese
trabajo consideran como endémicos a Compsospiza baeri y a Atlapetes citrinellus.

Composición y distribución de la avifauna de las Yungas
La avifauna de las Yungas está conformada por elementos de diferentes orígenes geográficos
y de distintas edades evolutivas y, aunque una prolongación de la amazónica, presenta
características propias a causa de los distintos períodos de expansión y retracción glacial
durante el Pleistoceno, poniéndose en contacto y aislándose del resto de las masas boscosas
sudamericanas. El contacto con la avifauna andina y de los matorrales áridos occidentales a
lo largo de todo su borde occidental y del Chaco en el oriental, le dieron una heterogeneidad
que es una característica marcada desde el punto de vista de la composición taxonómica.
Grado de endemismo
A diferencia de lo que ocurre en los Bosques Húmedos Patagónicos Fueguinos (Vuilleumier,
1967), donde existe un número relativamente alto de endemismos y poca especiación, las
Yungas de Argentina poseen relativamente pocos endemismos y un proceso de especiación
poco evidente, como el que puede observarse en los Páramos y Puna andinos (Vuilleumier,
1969), con géneros multiespecíficos como Muscisaxicola y Agriornis. Según este autor, el
prolongado aislamiento y la uniformidad ecológica durante ese período determinaron en los
Bosque Patagónicos que casi el 50% de las especies fuera endémica o de distribución
restringida y 7 géneros uniespecíficos, lo que favoreció la evolución filética antes que la
especiación.
El piso superior de las Yungas de Salta, Jujuy y Tucumán, constituido por los Bosques
Montanos (alisales y pinares), posee sólo dos especies endémicas de Argentina y que pueden
ser consideradas como típicas (considerando como típicas a aquellas especies que crían sólo
en ese tipo de vegetación aunque realicen desplazamientos migratorios invernales hacia
niveles más bajos); además hay especies y subespecies endémicas compartidas con el sur de
Bolivia, como son:
Leptoptila megalura saturata. Habita los alisales desde el norte de Salta hasta Catamarca.
En la región subtropical del sur de Bolivia se encuentra otra subespecie L. m. megalura (Olrog, 1963). Ambas especies están relacionadas a con L. verreauxi decipiens, de amplia
distribución en selvas montanas y pedemontanas y en el Chaco.
Amazona tucumana. Forma en común con Bolivia austral, cercana a A. pretrei, de las selvas
de Misiones y sur de Brasil, se distribuye por los alisales de todo el norte Argentino.
Geotrygon frenata margaritae. Subespecie conocida sólo para el sudeste de Jujuy, en las
Sierras de Santa Bárbara, donde cría en alisales, y como migratoria en la Selva Montana en
Calilegua (Olrog, 1973). Otras tres subespecies se distribuyen a lo largo de los bosques
tropicales de los Andes, desde Colombia hasta el centro de Bolivia (Hoy et al., 1963).
Scytalopus s. superciliaris. Habita en los alisales de todo el norte de Argentina, otra
subespecie se encuentra en el sur de Bolivia (S. s. zimmer).
Mecocerculus hellmayri argentinus. Sólo ha sido citada para los pinares de Podocarpus del
sudeste de Jujuy en Argentina, pero también se encuentra en la zona subtropical de Perú y
Bolivia (De Schauensee, 1970).
Cinclus schultzi. Es endémica de Argentina y sur de Bolivia, propia de los ríos de los
Bosques Montanos de Jujuy, Salta, Tucumán y montañas de Tarija, en el sur de Bolivia
(Ridgely y Tudor, 1989).
Saltator rufiventris. Fue encontrada en alisales de Alto Calilegua, Jujuy; también habita el
oeste de Bolivia (Olrog, 1972; Olrog y Contino, 1970).
Poospiza e. erythrophrys. Cría en los alisales desde el centro de Bolivia hasta Catamarca
(Ridgely y Tudor, 1989).
Compsospiza baeri. Sólo conocida en Tucumán en pastizales húmedos de altura en el
Aconquija y bordes del Bosque Montano Superior.
En las Selvas Montanas existe una sola especie endémica para Argentina:
Atlapetes citrinellus. De Salta, Jujuy, Tucumán y Catamarca.
Existen, sin embargo, varias endémicas en común con el sur de Bolivia:
Penelope dabbenei. Llega hasta el sudeste de Jujuy desde el centro de Bolivia (Olrog, 1979).

Microstilbon burmeisteri. Del sur de Bolivia y noroeste de Argentina, a lo largo de todas las
Yungas.
Eriocnemis glaucopoides. Es típico de las selvas desde el sur de Bolivia hasta la provincia de
Catamarca.
Veniliornis frontalis. Se distribuye desde el centro de Bolivia hasta los límites entre
Tucumán y Catamarca. Una distribución similar presentan: Synallaxis s. superciliosa y
Elaenia strepera. Atlapetes fulviceps llega desde el sur de Bolivia sólo hasta el sudeste de
Jujuy.
En los Bosques de Transición no se encuentran endemismos salvo para el bosque de palo
blanco de Bolivia austral, norte de Salta y sudeste de Jujuy: Ara auricollis, Ara caninde
(ocasional en el extremo norte de Salta) (Olrog, 1963; Short, 1975) y Picumnus
dorbignyianus, común en la zona del Parque Nacional Calilegua.
Si analizamos la distribución de algunas especies propias del bosque húmedo, muchas de
ellas poseen distribuciones disyuntas a lo largo de los bosques andinos (Buteo albigula, Ara
militaris, Aegolius harrisii, Grallaria albigula, Phyllomyias burmeisteri, Basileuterus
signatus, Catamblyrrhynchus diadema, Sicalis citrina), (De Schauensee, 1970; Olrog, 1968;
Ridgely y Tudor, 1989; Short, 1975), indicando una distribución anterior más extensa y
contínua. La restricción de varias de estas formas evidencia su dependencia de la historia
pleistocénica de los hábitats amazónicos, coincidentes con las expansiones y constricciones
cíclicas planteadas en la teoría de Haffer (1982).
Variación latitudinal de aves de las Yungas de Argentina Desde las Yungas del sur de Bolivia ocurre una pauperización del número de especies de
aves propias de bosques húmedos tropicales a medida que se desciende hacia el sur (De
Schauensee, 1970; Olrog, 1963, 1968). Catorce especies tienen su límite sur de dispersión en
el norte de Salta y otras 37 especies llegan sólo hasta el sudeste de Jujuy.
La variación en la organización de las comunidades de aves, especialmente en lo concerniente a la diversidad, a lo largo de un gradiente latitudinal en las Yungas, puede ser
analizado mediante tres perspectivas fundamentales (Pianka, 1966).
1. Desde un punto de vista evolutivo, la diversidad tiene que ver con los ritmos de
especiación y una larga estabilidad climática (Karr, 1976b), como lo ocurrido en los Bosques
Patagónicos, donde un prolongado aislamiento y estabilidad favorecieron la especiación de
numerosas formas endémicas (Vuilleumier, 1985).
Un buen número de especies de tiránidos está relacionado a la historia evolutiva de los
bosques andinos, distribuyéndose en las selvas y bosques montanos de los Andes
Sudamericanos de donde son endémicos (Ochthoeca leucophrys, Myiotheretes striaticollis,
Sayornis nigricans, Knipolegus aterrimus, K. cabanisi, Contopus fumigatus, Phyrrhomyias
cinnamomea, Mecocerculus leucophrys, M. hellmayri, Pseudocolopteryx acutipennis,
Elaenia albiceps, E. strepera, E. obscura, Phyllomyias sclateri y P. burmeisteri). Knipolegus
cabanisi, M. hellmayri, E. strepera y P. sclateri son endémicos de los bosques andinos del
sur de Perú, oeste de Bolivia y noroeste de Argentina. La historia evolutiva de varios de
estos géneros (Ochthoeca, Myiotheretes, Sayornis), se relaciona a la de los géneros
Muscisaxicola y Agriornis (Vuilleumier, 1971), ambos terrícolas adaptados a las altas
montañas y estepas patagónicas.
2. Desde una perspectiva biogeográfica, la diversidad está determinada por patrones de
intercambio faunístico entre regiones, la inmigración y extinción de especies y el efecto de la
geografía local (Vuilleumier, 1970), existiendo una correlación entre el número de especies y
el tamaño del área ocupada (MacArthur y Wilson, 1967). Esta se determina por la tasa de
inmigración y extinción y, en este caso también, por el grado de aislamiento entre las fuentes
de especiación y el área en cuestión. Aunque la caída de especies de las Yungas puede
explicarse de varias maneras, la expresión geográfica del área ocupada, en forma de una
"cuña" de bosques húmedos, rodeados de vegetación xerófila o desértica, hacen que sufran de
insularidad y de un proceso de invasión de especies propias de áreas de áreas de crecimiento

secundario o abiertas (tanto animales como vegetales). Esto conlleva una modificación del
hábitat original (Jansen, 1983) y la caída de especies especialistas y aumento de especies
generalistas u oportunistas, competidoras más agresivas.
3. Desde un punto de vista ecológico, la diversidad está determinada por la habilidad de las
especies para acomodarse a diversos factores como la disponibilidad estacional y variedad de
recursos, y la complejidad del hábitat (Orians, 1969; Pianka, 1966).
ÁREA DE ESTUDIOS Y MÉTODOS
El trabajo fue integrándose a través de unidades de estudio como: vegetación, números de
especies de aves, dieta, patrones de desplazamientos y uso del hábitat. Paralelamente se
obtuvieron otros datos, como estudios de plumaje y condición reproductiva que permitieron
cronometrar la dinámica de desplazamientos y época reproductiva.
Composición florística de las Selvas de Transición
Aproximadamente la mitad de las especies que se encuentran en el distrito de las Selvas
Montanas (Brown 1986; Cabrera, 1976), están presentes tanto en el bosque de Palo Blanco
como en el de Tipa y Pacará (Grau et al. 1997). Algunos ejemplos son: Juglans australis,
Parapiptadenia excelsa, Phoebe porphyria, Blepharocalix salicifolius, Tipuana tipu, Piper
tucumanum, Allophyllus edulis, Tabebuia avellanedae, Solanum riparium y Jacaranda
mimosifolia.
El Bosque de Transición de palo blanco
Se distribuye en forma actualmente discontinua desde el límite entre Argentina y Bolivia
hasta el Parque Nacional El Rey, en el sur de Salta, donde es reemplazado por el de Tipa y
Pacará. Es un bosque más alto que éste último, con árboles que fácilmente alcanzan los 30 m
de altura. Se extiende desde la llanura desde los 350 m hasta cerca de los 700 m, según las
localidades. Avanza internándose en el chaco a través de bosques de galería, donde especies
como E. contortisiliquum, Piptadenia excelsa, Prosopis nigra, Chorisia insignis, Acacia sp, de gran porte acompañan a los ríos. Calycophyllum. multiflorum y Phyllostilon rhamnoides
son los árboles dominantes en altura y destacables por sus troncos de fuste recto y color claro.
Están acompañados por Tabebuia impetiginosa, Enterolobium contortisiliquum,
Anadenanthera columbrina, Cordia trichotoma, Gleditsia amorphoides, Jacaranda
mimosifolia, Ruprechtia laxiflora, Schinopsis haenkeana, Acacia sp, Schinus sp, Chorisia
insigne, Piptadenia excelsa y Cedrela sp como árboles de mayor porte, árboles menores de
Tecoma stans, Carica quercifolia, Prosopis nigra, Fagara nigrescens y Poliera microphylla.
El estrato arbustivo es denso y enmarañado y está compuesto por Solanum granuloso-
leprosum, S. tucumanense, S. lorentzii, Chamisoa altissima, Verbesina sp, Cnicothamus
azafran, Piper tucumanum, Acacia etilis y Bouganvillea stipitata, entre otras muchas
especies. Abundan las epífitas como Aechmea distichanta y según las localidades están
presentes cactus de gran porte del género Trichocereus.
El Bosque de Transición de tipa y pacará
Se distribuye también en forma actualmente discontinua desde la llanura (aproximadamente
450 m de altura), hasta los 750 m. Durante la estación lluviosa, de Noviembre a Mayo, se
torna exuberante, aunque los árboles de mayor porte como Tipuana tipu, Anadenanthera
colubrina, Cedrela lilloi y Parapiptadenia excelsa, difícilmente sobrepasan los 20 m de
altura, como ocurre con el bosque de Transición de Palo blanco de Salta y Jujuy. La causa es
la explotación a la que se vio sometido este bosque, donde se extrajeron las especies
maderables de mayor porte y se desmontó para ganadería durante los últimos 200 años
(Vervoorst, 1982). El estrato arbóreo superior, formado por las copas de los árboles más
altos, está constituido por R .laxiflora, E. contortisiliquum, Phoebe porphyria, y los ya
mencionados. Los más grandes y frondosos, con gran diámetro de copa son los ejemplares
de T. tipu. El segundo estrato arbóreo está constituido por especies de 10-12 m de altura
como: J australis, F. coco, F. rombifolia, M. pungens, Xilosma pubescens, Scutia buxifolia,
Piper tucumanum, Allophyllus edulis, T. impetiginosa, J. mimosifolia, Rapanea laetenvirens,

Chorisia insigne, Caesalpinia paraguayensis y S. haenqueana. Una especie exótica muy
común es Morus nigra, sobre todo a lo largo de los ríos. Un tercer estrato está formado por
arbustos y arbolitos como Psychotria carthagenensis, Lantana camara, Abutilon sp, Urera
Solanum clavatum, S. riparium, C. azafran, Bacharis microdonta, Vasobia pauriflorus. En
este estrato abundan las enredaderas o plantas apoyantes como Celtis iguanea, C. altissima y
Passiflora misera y algunas espinosas como Rubus boliviensis y Celtis spinosa. El estrato
herbáceo está constituido por varios helechos como Telypteris sp y Adiantum sp y hierbas
como Anagallis arvensis, Oxalis sp, Valeriana sp, Sida sp, Bromis sp, Cyperus sp. Hay
numerosas plantas epifitas como A. distichanta, Vriesia, Tillandsia, Oncidium, Peperomia,
Polypodium y Rhipsalis, además líquenes y musgos.
Localidades de estudio
El trabajo es el resultado de la información obtenida en 50 viajes de campañas entre 1984 y
1997 a 25 localidades (Fig.4). En todas las localidades se realizaron observaciones sobre la
vegetación, como su aspecto, altura y grado de cobertura, identificándose especies arbóreas
dominantes. El estudio se realizó intensivamente en tres sitios principales de inventario y
monitoreo en hábitat equiparables de selvas de Tipa y Pacará en la provincia de Tucumán: La
Junta, Choromoro (26º12S 65º25´W), Piedra Tendida, Burruyacu (26º40´S 64º07W) y Dique
San Ignacio (27º44´S 65º40´W). En ellos se estudió la vegetación a lo largo del año,
distribución de las aves según los estratos, dieta, biomasa y dinámica de las especies,
realizándose un seguimiento cuali y cuantitativo en varias visitas a cada uno a lo largo de dos
años seguidos.
Fig. 4: Mapa de localidades de estudio en el noroeste argentino.

En la selva de palo blanco, se hizo un estudio estacional en dos épocas del año en Piquirenda
Viejo, Salta (22º20´S 63º47´W) y Laguna La Brea, Sierra de Santa Bárbara, Jujuy (23º52´S
64º27´W). Para completar la supervisión y el monitoreo de bosques pedemontanos a nivel
región, se hicieron censos y capturas con redes en Orán (23º22´S 64º09´W), Campo Quijano
(24º40´S 65º02´W), Parque Nacional El Rey (24º30´S 64º30´W), Paso de La Cruz, 43 Km al
E de Lumbreras (25º02´S 64º34´W), Metán (25º30´S 64º57´W), todas localidades de la
Provincia de Salta y en la Reserva Provincial La Florida, Tucumán (27º14´S 65º34´W). En
las localidades de Metán y La Florida se realizaron observaciones sobre dieta y uso del
hábitat y censos para obtener valores de biomasa.
Para establecer la época de reproducción de especies que crían a más altura e invernan en los
bosques pedemontanos fueron estudiadas: Las Aguitas, Taficillo, Sierra de San Javier,
Tucumán (26º42´S 65º20´W), con bosques de Pino del cerro (Podocarpus parlatorei) y
Aliso (Alnus acuminata), alisales en el Km 47, Ruta 307, próximo a La Angostura, Tafí del
Valle, Tucumán (26º56´S 65º41´W) y un sitio también con alisales en la Cuesta del Clavillo,
3 Km SW de La Banderita, Catamarca (27º24´S 66º06´W).
Para comparar las taxocenosis de Selva Montana con las de Transición y para el estudio de la
dinámica de las especies (bandadas mixtas, desplazamientos migratorios) en la Provincia de
Tucumán, se hicieron estudios en Horco Molle, 750 m (26º50´S 65º18´W), Piedras
Coloradas, Ruta 307, 750 m (27º05´S 65º39´W), El Indio, Ruta 307, 1200 m (27º00´S
65º41´W), Reserva Provincial Santa Ana, 800 m (27º28´S 65º41´W), y Cochuna, 1000 m
(27º18´S 65º55´W).
Se muestrearon las siguientes localidades chaqueñas, para recabar información sobre la
distribución y dinámica de especies compartidas y desplazamientos migratorios: Tonono, 30
Km al E de Tartagal, Salta. Bosque de galería de vegetación chaqueña sobre las márgenes del
Río Itiyuro (22º24’S 63º29’W), Finca Santa Cruz, Rosario de la Frontera, Salta. Bosque
chaqueño serrano (25º57’S 64º40’W); Chulca, San Pedro de Colalao, Tucumán. Bosque Chaqueño Serrano (26º11’S 65º30’W); Las Tipas, Parque Biológico Sierra de San Javier.
Bosque Chaqueño Serrano. (26º30’S 65º15’W); Ticucho, Tucumán. Bosque Chaqueño
Serrano (26º31´S 65º15´W); Aguas Saladas, Los Puestos, Tucumán. Bosque Chaqueño
(27º17´S 65º00´W).
Métodos para el estudio de la avifauna
Las aves fueron identificadas por captura, observación y canto. Se utilizó la técnica de
captura mediante redes de 12, 5 m de largo por 2,5 m de alto. En cada localidad se
confeccionó una lista de aves. También se hicieron listas generales para la Selva de
Transición, Bosque Chaqueño, Bosque Chaqueño Serrano, Selva y Bosque Montano. Las
aves fueron determinadas específicamente, consultándose en algunos casos la Colección
Ornitológica Lillo. El análisis de los datos obtenidos, especialmente la significación de la
presencia o ausencia de especies en una u otra localidad se completó mediante una revisión
bibliográfica de las aves de las Yungas de Argentina y sur de Bolivia, determinándose caída
del número de especies, endemismos y especies de distribución restringida. También se
realizó un monitoreo de aves para determinar el status y las tendencias de las poblaciones de
aves residentes y migratorias y de sus hábitat. Fue sistemático, cubriéndose todos los tipos de
hábitat posibles en cada localidad estudiada y todas las aves capturadas, vistas u oídas. Se
determinaron especies raras y de hábitat especiales y se estableció con precisión la época
invernal y la de nidificación. El monitoreo consistió fundamentalmente en determinar
presencia, distribución y abundancia relativa de las aves. Permitió, además, conocer el status
de cada ave como migratoria. Se categorizaron a las aves como: residente, migrante
altitudinal, migrante latitudinal y errática. Para cada ave se estableció: época reproductiva,
época invernal y época de migración en el caso de las migratorias. La condición reproductiva
se determinó mediante la presencia o ausencia de placas incubatrices en las hembras y
protuberancias cloacales en los machos. Las protuberancias cloacales de los machos
comienzan a desarrollarse con el cortejo, al comienzo de la estación reproductiva y alcanzan

su máximo desarrollo para la cópula, disminuyendo progresivamente con el nacimiento de
pichones. La placa incubatriz es una superficie del vientre de la hembra y muchas veces del
macho (según las especies), que pierde el plumaje y acumula líquidos, aumentando la
temperatura en esa zona. Permite incubar los huevos de forma más efectiva y va
desinflamándose al final del período de incubación. Luego de la estación de cría, esta
superficie vuelve a llenarse de plumas de contorno.
Los censos fueron de 3 días de duración, realizándose 2, 4 y 2 horas de censo
respectivamente en cada uno de los días. Se emplearon las dos primeras y las dos últimas
horas de luz en cada uno de los casos, completándose 8 horas de censo en cada localidad.
Durante cada censo, se recorrió lentamente a pié una transecta, anotándose todas las aves
vistas u oídas. En cada censo se obtuvieron datos sobre: número total de individuos y de
especies, alimentación, uso del hábitat y formación de bandadas mixtas.
Diversidad
La diversidad se calculó mediante el Índice de Diversidad de Simpson (D):D= 1- (pi) ,
donde pi es la proporción de individuos de la especie i en la comunidad (Simpson, 1949).
Análisis zoogeográfico de afinidades faunísticas
Para caracterizar los sitios de estudio y establecer sus afinidades se realizó un análisis de
correspondencia (CA) y un Análisis de Coordenadas Principales sobre una Matriz de
Distancia Chi Cuadrado, empleándose la presencia y ausencia de especies según las
localidades y la abundancia relativa de cada una para definir los ejes. Se analizó la
distribución de las especies obtenidas en las listas de aves de cada sitio de muestreo mediante
un Análisis de Agrupamientos sobre una Matriz de Distancia Chi Cuadrado, Técnica
UPGMA, para sitios y para especies (Rohlf, 1990).
Vegetación
En La Junta, Piedra Tendida y San Ignacio, se estudiaron 5 parcelas de 20 m x 20 m. En
ellas se determinaron especies arbóreas dominantes, altura de los árboles y especies arbóreas
o arbustivas fructificantes.
La distribución vertical del follaje, expresada mediante porcentaje de cobertura, fue medida
mediante telémetro de cámara fotográfica, haciendo foco verticalmente en el follaje,
ubicando un punto en el centro del campo. La presencia o ausencia de foco fue establecida
para 4 intervalos (2-3 m, 3-5 m, 5-10 m y 10-20 m). Para el estrato bajo, se contó el número
de contactos de la vegetación con una barra graduada de 2 m de altura. Las diferencias en la
estructura vertical del follaje, fueron expresadas en porcentaje de cobertura (Karr y Roth,
1971). Las diferencias en porcentaje de cobertura en cada estrato fueron promediadas para
todas las categorías de altura. La distribución vertical del follaje, obtenida para la época
decídua y la época lluviosa en las tres localidades, fue correlacionado luego con el número de
especies de aves. Los muestreos de aves se hicieron en las mismas áreas que los estudios de
vegetación.
La época de fructificación, pico de fructificación e identificación de las especies vegetales
más consumidas por las aves, se determinaron mediante observaciones de campo y consulta
bibliográfica.
Biomasa
Se trabajó sobre transectas de 300 m de largo por 30 m de ancho (9.000 m2), caminando
lentamente y contando el número de individuos de cada especie, en un espacio comprendido
por 15 m a cada lado del observador. Se utilizaron sendas previamente hechas, en algunas
ocasiones la mismas usadas para censos y colocación de redes. Se tomaron simultáneamente
los pesos de las aves que eran capturadas en cada localidad mediante las redes. Este método
fue utilizado de modo similar en varias localidades para poder comparar.
Dominancia
Wittaker (1967) definió el índice de dominancia (ID) como la concentración de la influencia
de las dos especies mas abundantes que controlan la composición de especies dentro de una
comunidad. Lo expresó como ID =100(Y1+Y2)/Yt ,donde Y1 y Y2 representan la abundancia

de las especies 1 y 2 e Yt es el número total de especies. Se establecieron valores de ID para
6 localidades de estudio, en algunos casos durante la época invernal y primaveral
(Piquirenda Viejo, La Junta, Piedra Tendida y San Ignacio), y en otros sólo durante la época
invernal (Metán y La Florida).
Estructura trófica de las comunidades
Fue estudiada simultáneamente en viajes de campaña realizados entre Agosto de 1989 y
Diciembre de 1991 a las localidades de La Junta, Piedra Tendida y San Ignacio. Para obtener
datos comparativos sobre dieta y uso del hábitat, se realizaron viajes a Piquirenda Viejo,
Metán y La Florida, localidades pedemontanas con Bosque de Transición.
Las dietas fueron determinadas mediante: a) registros de campo,
b) contenidos estomacales, c) heces colectadas de las aves al ser capturadas, y d) bibliografía.
Se siguieron los procedimientos descriptos por Holmes et al. (1979). Se elaboró previamente
un matriz de datos compuesta por el número de especies participantes y las categorías de
forrajeo.
Para establecer las diferentes categorías de forrajeo se catalogaron primeramente los tipos de
alimentos consumidos: insectos, semillas, frutos, pequeños vertebrados e insectos (carnívoro)
y néctar, detallándose en algunos casos más específicamente el alimento como peces,
mamíferos o aves. Las combinaciones de dietas como: insectos + semillas, néctar + insectos,
frutos + semillas, se corroboraron en base al análisis de algunos contenidos estomacales. Los
contenidos estomacales fueron analizados sólo en forma cualitativa.
Según los métodos utilizados por las aves para la obtención o captura del alimento fueron
clasificadas en los siguientes tipos:
1- Recolector: Aquellas especies que buscan y recogen un tipo de alimento que se encuentra
fijo, estacionario en el sustrato. Pueden ser frutos, insectos o flores. El ave avanza a saltitos o
se encuentra perchada y va tomando lo que encuentra a su paso sin tener que perseguir a su
presa. 2- Atrapador: El ave se abalanza de un salto o vuelo corto desde su percha y arrebata la presa
que puede estar en el suelo, en el follaje o en el agua.
3- Captura aérea de insectos: El ave sale de su percha hacia el aire para alcanzar un insecto o
lo persigue en vuelo. Generalmente eligen percha perchas altas o copas de los árboles.
4- Caza al vuelo rapaz: A la vista de la presa, generalmente aves, el rapaz inicia su
persecución a través de la vegetación, alcanzándola en pleno vuelo o haciéndola bajar al
suelo donde la inmoviliza con las garras.
5- Cazador por revoloteo: El ave revolotea entre el follaje, y con movimientos rápidos y
acrobáticos captura insectos en el aire o entre las hojas y ramitas.
6- Buscador: Método en que las aves buscan insectos en la corteza de los árboles o en la
hojarasca del suelo. Introducen su pico en cavidades ya existentes o levantando la corteza
desprendida.
7- Taladrador: El ave perfora la corteza de ramas y troncos en busca de sus presas,
generalmente larvas y hormigas. Poseen un pico fuerte como un cincel.
8- Planeador: Método utilizado por aves grandes, generalmente carroñeras que utilizan las
corrientes térmicas de aire para elevarse y mantenerse planeando hasta detectar el alimento.
Pero lo utilizan también algunos rapaces grandes que se alimentan de presas vivas.
Los sustratos desde donde los alimentos fueron tomados fueron clasificados como:
a - Suelo. Incluyendo hojarasca, pastos, etc.
b - Troncos. El eje principal de árboles.
c - Ramas. Las ramas secundarias del árbol.
d - Ramitas. Aquellas que soportan directamente las hojas.
e - Hojas. Incluyendo pecíolo y lámina.
f - Corteza. Puede estar desprendida o no del tronco pero todavía suspendida de él o de otras
ramas o follaje.
g - Flores.

h - Cápsulas o semillas.
i - Aire.
Reuniendo los métodos de alimentación, el sustrato y el tipo de alimento consumido se
elaboró una tabla con 19 categorías tróficas.
Los datos invernales fueron tomados durante los meses de Junio, Julio y Agosto, y los de
primavera en Octubre, Noviembre y Diciembre.
Diversidad de macronichos
Se calculó mediante la fórmula de Shannon-Wiener (Margaleff, 1958): H'= S (pi) (Log.pi)
donde pi es la proporción de especies dentro de cada categoría. La sumatoria de todos los
logaritmos da una medida (nats de inform./macronichos) de la diversidad de macronichos. Se
calculó para las localidades mencionadas anteriormente donde se estudió la dieta.
ÁREA DE ESTUDIOS INTENSIVOS
Localidad La Junta
Se ubica en la porción norte del Sistema Orográfico Central o Principal de la provincia de
Tucumán. Este posee cimas de alturas superiores a los 4.000 m y en algunos casos 5000 m.
Presenta en su sección media una larga muralla ininterrumpida de altas cordilleras, con
faldeos anchos y suaves hacia la vertiente oriental (Fig. 5). En la porción norte, el bloque de cordilleras está subdividido por valles longitudinales y sierras, cuya altura disminuye en
dirección al este (Khun y Rohemeder, 1924). El Valle de Choromoro es uno de los tantos
Fig. 5: Panorama de las Cumbres Calchaquíes desde el Taficillo, Sierra de San Javier.
que se intercalan entre las formas altas como el Morro de La Ciénaga, Abra de las Cañas,
Cumbres de Raco y Siambón y Cabra Horco, con alturas que superan los 3000 m. (Fig. 6).
La Junta se ubica entre los cerros Alto de la Totora y Taco Yacu y debe su nombre a la
confluencia de los Ríos Rearte, de las Cuevas y El Potrero, para formar el Río Chuscha o
Choromoro. Este río nace justamente en esta localidad y su cauce corre por un estrecho valle
fluvial rodeado de los cerros antes mencionados hasta desembocar en el Río Salí.
En La Junta, predominan los vientos monzónicos procedentes del norte y el noroeste, con una
mínima influencia de los vientos del cuadrante sur, generalmente débiles y continuos, con
lenta afluencia de aire fresco (Santillán de Andrés y Ricci, 1966).
Las precipitaciones son más abundantes que en el resto de la cuenca de Choromoro, entre los
600 y 800 mm anuales, según el año y la localidad, debido a la exposición distinta de las
estribaciones montañosas secundarias como el Alto de la Totora, Taco Yaco y Cabra Horco.

Es común que las vertientes occidentales y los fondos de los valles reciban menores
precipitaciones que las vertientes orientales (Santillán de Andrés y Ricci, 1966). Las lluvias
son tormentosas y locales en verano y suaves, persistentes y débiles en invierno. Llueve en
Fig. 6: Ubicación de la localidad La Junta en la cuenca del Río Choromoro, Tucumán..
general a partir del mes de Noviembre, acentuándose la pluviosidad entre Diciembre y marzo
y declinando rápidamente en Abril. La estación seca es muy marcada entre Mayo y Octubre,
estableciéndose una secuencia de 6 meses secos y 6 húmedos.
De acuerdo con estos mismos autores, en la cuenca de Choromoro, las temperaturas medias
de verano oscilan entre los 22 y 23ºC y las de invierno entre los 9 y 12ºC .
Localidad Dique San Ignacio
Se ubica en la porción más austral del Sistema Orográfico Central, con faldeos anchos y
suaves hacia el este, próximo a la provincia de Catamarca (Fig. 7). La cordillera describe un
arco abierto hacia el este, cuyo vértice se halla frente a la ciudad de Concepción, reflejando el
rumbo de las rocas antiguas (SSO - NNE en el Aconquija y SE - NO en la parte que se
continúa en la provincia de Catamarca). Los Nevados del Aconquija ocupan el lugar donde
se produce esta vuelta de rumbo, con alturas de 5.500 m. en sus picos más altos. Las sierras
del Aconquija, son condensadoras del vapor de agua de las masas de aire húmedo
provenientes del N y NE. Toda la vertiente oriental de los cordones montañosos de la
provincia está dotada de una red hidrológica bien desarrollada que desemboca en la llanura.
La localidad de estudio se encuentra en la toma que alimenta de agua al Dique San Ignacio,
ubicado unos 3 Kilómetros hacia el este, al pié de las Cumbre de los Llanos. El espejo de
Fig. 7: Panorama de los Nevados del Aconquija y ubicación de la localidad Dique San
Ignacio.

agua del dique tiene dos hectáreas y provee de agua de riego a las poblaciones ubicadas a
menor altura. La distinta ubicación de los cordones montañosos en las tres localidades
estudiadas intensivamente, determina direcciones generales variadas respecto a los vientos.
En San Ignacio, los vientos tienen una dirección predominante SSE. Las quebradas y
pendientes que miran hacia el sur y suroeste, se encuentran en una situación más favorable
respecto a la humedad que reciben, en comparación a las que miran al norte o noroeste. Por
ello, las quebradas muestran una marcada diferencia en la vegetación en sus dos pendientes
(Kuhn, 1924). Es la localidad más húmeda de las tres estudiadas, influida por las altas
montañas a su espalda. Las precipitaciones medias anuales son de 1.200 mm (Bruchmann,
1977). Las lluvias son abundantes y tormentosas a partir de Octubre y hasta Marzo,
declinando luego en Abril y Mayo. Existe una estación seca marcada entre Mayo y
Setiembre. La época lluviosa es de 8 meses, más marcada y prolongada que en la localidad
de La Junta. Las temperaturas medias de verano oscilan entre los 24 y 26ºC y en invierno
son de 12ºC (datos del Departamento de Graneros), (Bruchmann, 1977).
Localidad Piedra Tendida
Se ubica en el Sistema Oriental, conocido como Sierras de Burruyacu, por ocupar gran parte
del Departamento homónimo. Estas sierras se extienden en sentido longitudinal a lo largo de 170 km desde cerca del Río Juramento, en la provincia de Salta, hasta las proximidades de
San Miguel de Tucumán (Fig. 8). Se distinguen claramente dos cordones principales de
rumbo NNE-SSW y casi paralelos entre sí: el Occidental, constituido por las Sierras de
Medina y del Nogalito (2050 m y 1750 m, respectivamente), (Santillán de Andrés et al.,
1967); y el Oriental, formado por las Sierras del Campo y de la Ramada, de 1160 m y 2000 m
respectivamente (Santillán de Andrés et al., 1967). Por entre ambas sierras se ubica la falla
"del Tajamar", por donde corre el río del mismo nombre que atraviesa Piedra Tendida. El
borde sur y oriental de este conjunto montañoso se pierde con suaves estribaciones en la
llanura, mientras que el borde occidental de las Sierras de Medina cae abruptamente hacia la
cuenca del Río Salí. La localidad está influida por vientos del sureste. Los vientos húmedos
del sur en su avance hacia el norte determinan la pluviosidad. De aquí que los mayores
registros pluviométricos par las Sierras de Burruyacu correspondan a las laderas orientales de
las sierras del Campo y de La Ramada. Por otro lado, debido a la poca elevación de la Sierra
de La Ramada, las lluvias son abundantes hasta las cumbres, por lo que estas se encuentran
cubiertas de una tupida vegetación (Santillán de Andrés et al., 1967).
Fig. 8: Sierras del Noroeste de Tucumán y ubicación de la localidad Piedra Tendida.

Las precipitaciones medias anuales son de 970 mm para La Ramada y de 1.000 mm para El
Cajón, próximo a la localidad de estudio. Las heladas ocurren más frecuentemente en las
zonas bajas que en las montañas y en las quebradas el frío es más notorio por ser estas
estrechas, umbrías y profundas. En verano se registran temperaturas medias inferiores que en
el llano, aunque en toda la región, debido a lo variado del relieve, se produce una variedad de
microclimas, tanto en temperatura como en humedad. Las temperaturas medias de verano en
la zona de Piedra Tendida oscilan entre 20 y 25ºC y en invierno entre 13 y 15ºC (Santillán
de Andrés et al., 1967). Esta localidad posee precipitaciones y temperaturas intermedias
respecto a las de La Junta y Dique San Ignacio.
RESULTADOS
ESTRUCTURA Y COMPOSICION FLORISTICA DE LA VEGETACION
El Bosque de Transición de palo blanco se distribuye en forma actualmente discontinua desde
el límite entre Argentina y Bolivia hasta el Parque Nacional El Rey, en el sur de Salta, donde
es reemplazado por el de tipa y pacará. Es un bosque más alto que éste último, con árboles
que fácilmente alcanzan los 30 m de altura. Se extiende desde la llanura (350 m aproximados)
hasta cerca de los 600 m, según las localidades. Avanza internándose en el chaco a través de
bosques de galería, como ocurre en la localidad de Tonono, 30 Km al este de Tartagal, donde
especies como E. contortisiliquum, Piptadenia excelsa, Prosopis nigra, Chorisia insignis,
Acacia sp (garabatos), de gran porte acompañan al Río Itiyuro. La composición florística
está constituida por C. multiflorum y por P. rhamnoides, como árboles dominantes en altura
y destacables por sus troncos de fuste recto y color claro, como bien puede apreciarse en
Laguna La Brea. En esta localidad la vegetación estaba compuesta por T. impetiginosa, E.
contortisiliquum, A. columbrina, Cordia trichotoma, Gleditsia amorphoides, J. mimosifolia,
Ruprechtia laxiflora, Schinopsis haenkeana, Acacia sp, Schinus sp, Chorisia insigne,
Piptadenia excelsa y Cedrela sp como árboles de mayor porte, árboles menores de Tecoma
stans, Carica quercifolia, Prosopis nigra, Fagara nigrescens y Poliera microphylla, y
estrato arbustivo compuesto por Solanum granuloso-leprosum, S. tucumanense, S. lorentzii,
Chamisoa altissima, Verbesina sp, Cnicothamus azafran, Piper tucumanum, Acacia etilis y
Bouganvillea stipitata. Abundan las epífitas como Aechmea distichanta y se encontraban
presentes cactus del Género Trichocereus.
El Bosque de Transición de tipa y pacará se distribuye en forma actualmente discontinua
desde la llanura (aproximadamente 450 m de altura), hasta los 750 m.
Durante la estación lluviosa, de Noviembre a Mayo, se torna exuberante, aunque los árboles
de mayor porte (T. tipu, A. colubrina, Cedrela lilloi, P. excelsa), difícilmente sobrepasan los
20 m de altura, como ocurre con el bosque de Transición de palo blanco de Salta y Jujuy. La
causa podría ser la explotación a la que se vio sometido este bosque, donde se extrajeron las
especies maderables de mayor porte y se desmontó para ganadería (Vervoorst, 1982).
Pueden distinguirse 4 estratos de vegetación en parcelas con bosque maduro:
El estrato arbóreo superior, formado por las copas de los árboles más altos, está constituido
por R. laxiflora, E. contortisiliquum, Phoebe porphyria, y los ya mencionados T. tipu, A.
colubrina, C. lilloi y P. excelsa. Generalmente los ejemplares de T. tipu son los más grandes
y frondosos, con gran diámetro de copa.
El segundo estrato arbóreo está constituido por especies de 10-12 m de altura como : J.
australis, F. coco, F. rombifolia, M. pungens, Xilosma pubescens, Scutia buxifolia, Piper
tucumanum, Allophyllus edulis, T. impetiginosa, J. mimosifolia, Rapanea laetenvirens,
Chorisia insigne, Caesalpinia paraguayensis y S. haenqueana. Una especie exótica muy
común es Morus nigra, sobre todo a lo largo de los ríos.Un tercer estrato está formado por
arbustos y arbolitos como Psychotria carthagenensis, Lantana camara, Abutilon sp, Urera ,
Solanum clavatum, S. riparium, C. azafran, Bacharis microdonta, Vasobia pauriflorus. En

este estrato abundan las enredaderas o plantas apoyantes como Celtis iguanea, C. altissima y
Passiflora misera y algunas espinosas como Rubus boliviensis y Celtis spinosa.
El estrato herbáceo está constituido por varios helechos como Telypteris sp y Adiantum sp y
hierbas como Anagallis arvensis, Oxalis sp, Valeriana sp, Sida sp, Bromis sp, Cyperus sp.
Hay numerosas plantas epifitas como A. distichanta, Vriesia, Tillandsia, Oncidium,
Peperomia, Polypodium y Rhipsalis, además líquenes y musgos.
En las distintas localidades estudiadas de Bosque de Transición de tipa y pacará existe
similitud en cuanto a la composición y estructura de la vegetación, aunque se presentan
algunas asociaciones vegetales particulares según las características e historia del sitio, entre
las que se pueden citar:
a) Zonas con abundancia de tusca (Acacia aroma), en distintos estadíos sucesionales, desde
asociaciones puras hasta estadíos más avanzados junto a arbolitos de A. edulis, X. pubescens,
Eugenia sp y jóvenes de A. colubrina, con diámetros máximos de tronco no mayores de 30
cm y densidades arbóreas de 600 árboles por Ha (árboles de más de 15 cm de diámetro de
tronco). A. aroma actúa como invasora desde el Chaco (Vervoorst, 1982) y se encuentra en
las zonas de borde.
b) Asociaciones casi puras de A. colubrina, junto a P. excelsa, que atraen bandadas enormes
de loros durante el invierno. Por debajo crece un estrato arbustivo denso, favorecido por la
buena iluminación durante los meses de sequía en que estos árboles pierden completamente
sus hojas, abundan en estas parcelas C. iguanea y R. boliviensis, formando una trama
arbustiva espinosa hasta 3-4 m de altura.
c) Bosquecillos con predominancia de J. australis, árbol que pierde tempranamente las hojas
en el mes de Junio, a veces en asociaciones casi puras, de no más de 15 m de altura.
d) Quebradas y laderas muy húmedas con predominancia de P. porphyrya. En el estrato bajo
abunda el helecho P. deflexa, que alcanza 2 m de altura, además hay abundancia de epifitas y
musgos.
e) En quebradas secas y pedregosas crece C. insigne, junto a A. colubrina de escaso porte y
en el estrato arbustivo abundan cactáceas y bromeliáceas, como ocurre en las quebradas de
los ríos San Ignacio y Choromoro.
COMPOSICION Y DISTRIBUCION DE LA AVIFAUNA DE LOS BOSQUES DE
TRANSICION DE ARGENTINA
Diversidad de especies
De acuerdo con observaciones personales, aves marcadas y datos bibliográficos se registraron
357 especies de aves en ambientes de bosques poco alterados de las Yungas de Argentina,
entre Bosque de Transición, Selva Montana y Bosques de Alisos y de Pino del Cerro, desde
los límites con Bolivia hasta la provincia de Catamarca. Esto representa el 36,3% del total de
las formas citadas para el país (Narosky e Yzurieta, 1988; Olrog, 1984).
En los Bosques Pedemontanos de Transición, desde el límite con Bolivia hasta el extremo sur
de su distribución, registré 215 especies de aves durante este estudio (Lista 1), con datos
propios obtenidos en las localidades citadas anteriormente. Es una gran riqueza considerando
que el área ocupada por estos bosques es de aproximadamente 890 Km2 y que la riqueza de
aves representa el 20% del total de las formas registradas para Argentina. Comparando estos
datos con el Chaco (Short, 1975), 409 especies fueron registradas en todo el gran Chaco, un
área más de 10 veces mayor, de aproximadamente 1.000.000 de Km. La Caatinga y el
Cerrado de Brasil con cerca de 200 especies ocupan un área de unos 2.383.000 Km2, que
representa sólo un 13% de la avifauna de Brasil (Sick, 1965).
Entre las 215 especies registradas, 83 especies se encontraron presentes en los censos de
otoño y/o invierno y primavera, por lo que pueden ser consideradas residentes, y por su
constancia, utilizada para caracterizar a estos bosques.

La riqueza de especies, expresada por las especies de aves residentes y las nidificantes
(aunque luego de criar migren) fue de 146 especies. Para las 69 especies restantes, estos
bosques son parte importante de su dinámica de desplazamientos.
Respecto a la diversidad local, en el bosque de palo blanco del extremo norte de Salta
(localidad de Piquirenda Viejo), se encontraban presentes 78 especies, incluyendo censos de
Mayo, Agosto y Noviembre, en un área aproximada de 10 Ha de bosque atravesada por el
Río Itiyuro (Lista 2). De éstas, 34 especies eran residentes, encontrándose presentes en
otoño, invierno y primavera, y 21 especies no se encontraron en el bosque de Tipa y Pacará.
En el bosque de palo blanco del sudeste de Jujuy (localidad de Laguna La Brea), se
registraron 104 especies de aves (Lista 3). Este número tan alto de especies, especialmente
en el mes de Mayo, se ve influido por que el área censada está próxima a bosques chaqueños
y a una laguna natural de agua salobre a que atrae a varias especies de distribución amplia.
El área de estudio, de aproximadamente 10 Ha, se encuentra parcialmente inundado en Mayo,
culminación del período de lluvias que la laguna acumula. Esto es coincidente con el arribo
de especies migratorias desde provincias ubicadas más al sur, que migran a través de estos
bosques. Aún así, esta localidad alberga el mayor número de especies residentes, 43, en
comparación con lo que ocurre en otras localidades de bosques pedemontanos.
Diversidad de aves en Tucumán
En el Bosque de Transición de la provincia de Tucumán se registraron 142 especies, de las
cuales 104 son frecuentes y solo 19 son constantes, están presentes en cada localidad y
pueden ser utilizadas como diagnósticas, estas son: Crypturellus tataupa, Buteo
magnirostris, Pionus maximiliani, Leptoptila verreauxi, Chlorostilbon aureoventris,
Picumnus cirratus, Sittasomus griseocapillus, Synallaxis superciliosa ,Syndactila
rufosuperciliatus, Sayornis nigricans, Phylloscartes ventralis, Troglodytes aedon,
Cyanocorax chrysops, Turdus rufiventris, Thraupis sayaca, Saltator caerulescens,
Cyanocompsa cyanea, Arremon flavirostris y Zonotrichia capensis.
Entre las 142 especies de aves en de estos bosques en las tres localidades estudiadas en
intensidad, sólo 104 especies son frecuentes (Lista 4). Respecto a la diversidad local, se
registraron 71 especies para Piedra Tendida, 65 para La Junta y 68 en San Ignacio. Es
interesante destacar que sólo 11 especies residentes son comunes a las tres localidades y
estaban presentes en los censos de invierno y verano y 8 especies migrantes neotropicales
nidificantes en estos bosques están presentes en los tres sitios durante la primavera.
Comparando las especies registradas durante los censos en las tres localidades en Tucumán,
hay una regularidad notoria en el número de especies de aves en cada localidad. Los
números variaron entre 33 y 38 especies por censo de 8 horas de duración, distribuidas en tres
días, sin importar la época del año (Tablas 1, 2 y 3).
ESPECIE JUL SET OCT Cat. Trófica
Crypturellus tataupa 0.007 0.01 RO
Cathartes aura 0.01 P
Coragyps atratus 0.03 0.01 P
Buteo magnirostris 0.007 0.01 0.01 AC
Polyborus plancus 0.007 P
Merganetta armata 0.01 0.01 RO
Leptoptila verreauxi 0.007 RS
Aratinga mitrata 0.17 RFS
Pionus maximiliani 0.04 RFS
Amazona aestiva 0.05 RFS
Chlorostilbon aureoventris 0.01 0.02 RNI
Amazilia chionogaster 0.03 0.008 RNI
Sappho sparganura 0.01 RNI
Ceryle torquata 0.01 AP
Campephilus leucopogon 0.01 0.008 TT
Picumnus cirratus 0.007 T
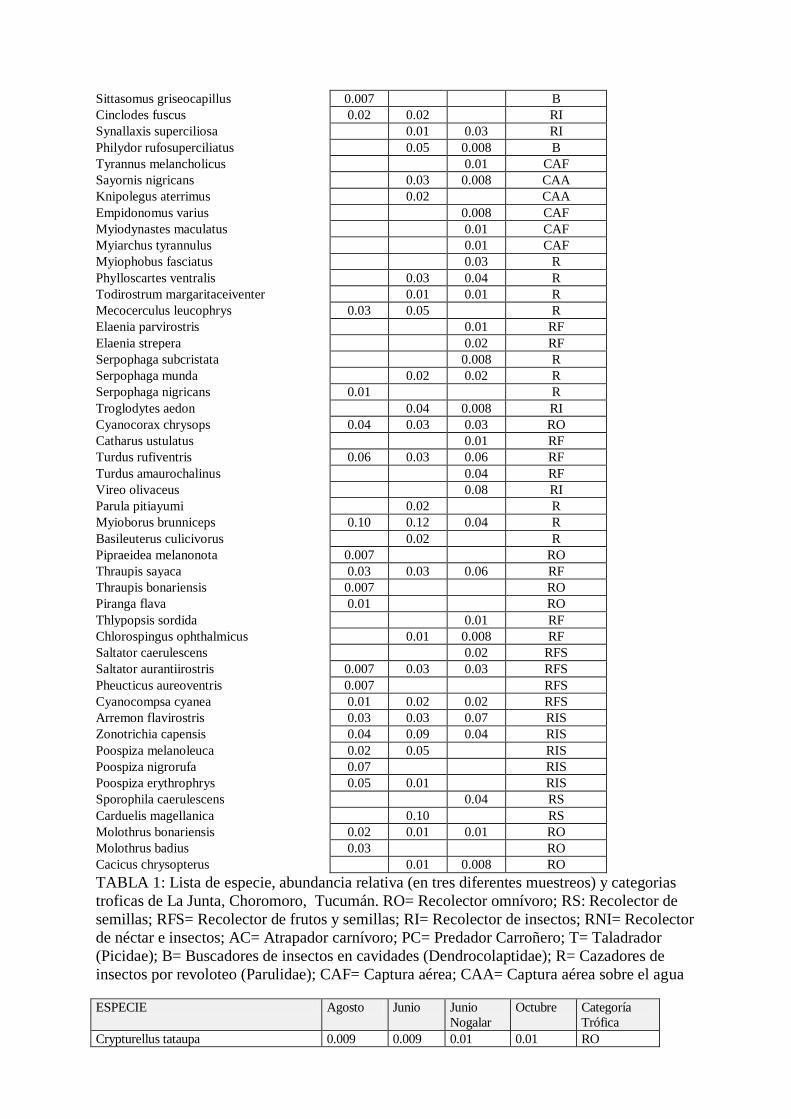
Sittasomus griseocapillus 0.007 B
Cinclodes fuscus 0.02 0.02 RI
Synallaxis superciliosa 0.01 0.03 RI
Philydor rufosuperciliatus 0.05 0.008 B
Tyrannus melancholicus 0.01 CAF
Sayornis nigricans 0.03 0.008 CAA
Knipolegus aterrimus 0.02 CAA
Empidonomus varius 0.008 CAF
Myiodynastes maculatus 0.01 CAF
Myiarchus tyrannulus 0.01 CAF
Myiophobus fasciatus 0.03 R
Phylloscartes ventralis 0.03 0.04 R
Todirostrum margaritaceiventer 0.01 0.01 R
Mecocerculus leucophrys 0.03 0.05 R
Elaenia parvirostris 0.01 RF
Elaenia strepera 0.02 RF
Serpophaga subcristata 0.008 R
Serpophaga munda 0.02 0.02 R
Serpophaga nigricans 0.01 R
Troglodytes aedon 0.04 0.008 RI
Cyanocorax chrysops 0.04 0.03 0.03 RO
Catharus ustulatus 0.01 RF
Turdus rufiventris 0.06 0.03 0.06 RF
Turdus amaurochalinus 0.04 RF
Vireo olivaceus 0.08 RI
Parula pitiayumi 0.02 R
Myioborus brunniceps 0.10 0.12 0.04 R
Basileuterus culicivorus 0.02 R
Pipraeidea melanonota 0.007 RO
Thraupis sayaca 0.03 0.03 0.06 RF
Thraupis bonariensis 0.007 RO
Piranga flava 0.01 RO
Thlypopsis sordida 0.01 RF
Chlorospingus ophthalmicus 0.01 0.008 RF
Saltator caerulescens 0.02 RFS
Saltator aurantiirostris 0.007 0.03 0.03 RFS
Pheucticus aureoventris 0.007 RFS
Cyanocompsa cyanea 0.01 0.02 0.02 RFS
Arremon flavirostris 0.03 0.03 0.07 RIS
Zonotrichia capensis 0.04 0.09 0.04 RIS
Poospiza melanoleuca 0.02 0.05 RIS
Poospiza nigrorufa 0.07 RIS
Poospiza erythrophrys 0.05 0.01 RIS
Sporophila caerulescens 0.04 RS
Carduelis magellanica 0.10 RS
Molothrus bonariensis 0.02 0.01 0.01 RO
Molothrus badius 0.03 RO
Cacicus chrysopterus 0.01 0.008 RO
TABLA 1: Lista de especie, abundancia relativa (en tres diferentes muestreos) y categorias
troficas de La Junta, Choromoro, Tucumán. RO= Recolector omnívoro; RS: Recolector de
semillas; RFS= Recolector de frutos y semillas; RI= Recolector de insectos; RNI= Recolector
de néctar e insectos; AC= Atrapador carnívoro; PC= Predador Carroñero; T= Taladrador
(Picidae); B= Buscadores de insectos en cavidades (Dendrocolaptidae); R= Cazadores de
insectos por revoloteo (Parulidae); CAF= Captura aérea; CAA= Captura aérea sobre el agua
ESPECIE Agosto Junio Junio
Nogalar
Octubre Categoría
Trófica
Crypturellus tataupa 0.009 0.009 0.01 0.01 RO

Coragyps atratus 0.01 PC
Cathartes aura 0.009 PC
Sarcoramphus papa 0.004 0.01 PC
Buteo magnirostris 0.004 0.002 0.01 0.02 AC
Micrastur ruficollis 0.01 CVR
Polyborus plancus 0.009 0.009 0.01 PC
Penelope obscura 0.01 0.01 RO
Tigrisoma fasciatum 0.009 0.009 AC
Aramides cajanea 0.009 0.004 RO
Leptoptila verreauxi 0.004 0.009 RS
Aratinga mitrata 0.009 0.02 RFS
Pionus maximiliani 0.04 0.007 0.02 RFS
Amazona tucumana 0.01 RFS
Amazona aestiva 0.01 RFS
Piaya cayana 0.004 AC
Tyto alba 0.004 AM
Otus choliba 0.004 AC
Aegolius harrisi 0.004 AC
C. aureoventris 0.004 0.009 0.007 0.01 RNI
Amazilia chionogaster 0.01 0.02 0.02 RNI
Eriocnemis glaucopoides 0.004 RNI
Sappho sparganura 0.01 0.004 RNI
Chloroceryle americana 0.004 AP
Picumnus cirratus 0.02 0.004 0.01 T
Piculus rubiginosus 0.01 0.007 TT
Campephilus leucopogon 0.01 TT
Sittasomus griseocapillus 0.004 0.02 BC
Cinclodes fuscus 0.004 RI
Synallaxis superciliosa 0.05 0.04 0.02 0.02 RI
P. rufosuperciliatus 0.03 0.04 0.06 0.01 B
Ochthoeca leucophrys 0.007 CAF
Sayornis nigricans 0.009 0.004 0.01 CAA
Knipolegus cabanisi 0.004 0.02 CAF
Knipolegus aterrimus 0.02 CAA
Empidonomus varius 0.02 CAF
Myiodynastes maculatus 0.05 CAF
Myiarchus swainsoni 0.02 CAF
Myiarchus tyrannulus 0.004 CAF
Tyrannus melancholicus 0.01 CAF
Phylloscartes ventralis 0.05 0.04 0.04 0.02 R
Mecocerculus leucophrys 0.01 0.004 0.05 R
Todirostrum margaritaceiventer 0.01 R
Todirostrum plumbeiceps 0.01 R
Elaenia obscura 0.004 CAF
Troglodytes aedon 0.009 0.01 0.01 RI
Cyanocorax chrysops 0.05 0.03 0.07 0.09 RO
Catharus ustulatus 0.02 RF
Turdus rufiventris 0.01 0.08 0.06 0.04 RO
Turdus amaurochalinus 0.01 RO
Cychlaris gujanensis 0.04 RO
Vireo olivasceus 0.04 R1
Parula pitiayumi 0.01 0.03 0.01 R
Geothlypis aequinoctialis 0.004 R
Myioborus brunniceps 0.07 0.06 0.06 0.02 R
Basileuterus culicivorus 0.009 0.02 0.01 R
Pipraeidea melanonota 0.01 RO
Thraupis sayaca 0.009 0.10 0.03 0.09 RO
Thraupis bonariensis 0.01 RF
Piranga flava 0.03 RO

Chlorospingus ophthalmicus 0.02 0.02 RO
Saltator caerulescens 0.01 0.01 0.05 RFS
Saltator aurantiirostris 0.01 0.01 0.02 RFS
Pheucticus aureoventris 0.004 RFS
Cyanocompsa cyanea 0.01 0.004 0.01 RFS
Coryphospingus cucullatus 0.004 0.02 RIS
Atlapetes citrinellus 0.009 0.01 RIS
Arremon flavirostris 0.06 0.06 0.06 0.02 RIS
Zonotrichia capensis 0.02 0.01 0.07 0.04 RIS
Poospiza erythrophrys 0.01 0.12 RIS
Carduelis magellanicus 0.02 0.02 RS
Molothrus bonariensis 0.01 0.01 RO
Icterus cayanensis 0.004 0.04 RO
Cacicus chrysopterus 0.04 0.01 RO
TABLA 2: Abundancia relativa y categorías tróficas de Piedra Tendida, Burrruyacu,
Tucumán. RO= Recolector omnívoro; RS: Recolector de semillas; RFS= Recolector de frutos
y semillas; RI= Recolector de insectos; RNI= Recolector de néctar e insectos; AC= Atrapador
carnívoro; PC= Predador Carroñero; T= Taladrador (Picidae); B= Buscadores de insectos en
cavidades (Dendrocolaptidae); R= Cazadores de insectos por revoloteo (Parulidae); CAF=
Captura aérea; CAA= Captura aérea sobre el agua; CVR= Captura en vuelo rapaz.
Especie Mayo Octubre Diciembre Categoria
trófica
Crypturellus tataupa 0.02 0.008 0.02 RO
Cathartes aura 0.01 PC
Coragyps atratus 0.02 PC
Buteo magnirostris 0.01 0.009 AC
Buteogallus urubitinga 0.008 AP
Polyborus plancus 0.01 PC
Nycticorax nycticorax 0.01 AP
Aramides cajanea 0.01 0.01 AC
Leptoptila verreauxi 0.01 0.03 RS
Pionus maximiliani 0.04 0.03 0.01 RFS
Amazona aestiva 0.08 RFS
Guira guira 0.009 AC
Otus choliba 0.02 0.008 AC
Chlorostilbon aureoventris 0.01 RNI
Amazilia chionogaster 0.03 0.03 0.01 RNI
Eriocnemis glaucopoides 0.01 0.008 RNI
Sappho sparganura 0.03 RNI
Chloroceryle amazona 0.01 AP
Chloroceryle americana 0.03 AP
Ceryle torquata 0.01 0.02 AP
Picumnus cirratus 0.01 0.01 T
Campephilus leucopogon 0.02 0.009 T
Xiphocolaptes major 0.01 B
Sittasomus griseocapillus 0.01 0.008 B
Cinclodes fuscus 0.04 RI
Synallaxis superciliosa 0.05 0.02 0.03 RI
Philydor rufosuperciliatus 0.01 0.01 B
Tyrannus melancholicus 0.01 CAF
Sayornis nigricans 0.02 0.008 CAA
Myiodynastes maculatus 0.01 0.04 CAF
Pitangus sulphuratus 0.01 0.01 0.03 CAF
Myiophobus fasciatus 0.01 R
Phylloscartes ventralis 0.06 0.01 R
Mecocerculus leucophrys 0.008 R

Elaenia albiceps 0.01 0.04 RF
Elaenia parvirostris 0.009 RF
Elaenia strepera 0.009 RF
Serpophaga munda 0.01 R
Serpophaga nigricans 0.01 R
Troglodytes aedon 0.02 RI
Troglodytes solstitialis 0.02 0.008 RI
Notiochelidon cyanoleuca 0.21 0.24 0.24 CA
Progne modesta 0.02 CA
Cyanocorax chrysops 0.07 0.008 0.04 RO
Catharus ustulatus 0.01 RF
Turdus nigriceps 0.04 RO
Turdus rufiventris 0.01 0.03 0.05 RO
Turdus amaurochalinus 0.05 0.05 RO
Vireo olivaceus 0.01 0.04 R
Parula pitiayumi 0.02 R
Myioborus brunniceps 0.15 R
Basileuterus culicivorus 0.01 R
Pipraeidea melanonota 0.01 RO
Thraupis sayaca 0.09 0.01 0.02 RO
Piranga flava 0.02 RO
Thlypopsis sordida 0.008 0.009 RO
Saltator caerulescens 0.008 0.06 RFS
Saltator aurantiirostris 0.01 RFS
Cyanocompsa cyanea 0.01 0.009 RFS
Coryphospingus cucullatus 0.01 0.01 RIS
Atlapetes citrinellus 0.01 RIS
Arremon flavirostris 0.07 0.08 0.07 RIS
Zonotrichia capensis 0.03 0.05 0.03 RIS
Sporophila caerulescens 0.02 RS
Molothrus bonariensis 0.01 RIS
Molothrus badius 0.01 RIS
TABLA 3: Abundancia relativa y categorías troficas de aves de San Ignacio, Tucumán. RO=
Recolector omnívoro; RS: Recolector de semillas; RFS= Recolector de frutos y semillas; RI=
Recolector de insectos; RNI= Recolector de néctar e insectos; AC= Atrapador carnívoro;
AC= Atrapador piscívoro; PC= Predador Carroñero; T= Taladrador (Picidae); B= Buscadores
de insectos en cavidades (Dendrocolaptidae); R= Cazadores de insectos por revoloteo
(Parulidae); CAF= Captura aérea; CAA= Captura aérea sobre el agua.
En Piedra Tendida, Burruyacu, 20 especies constantes estuvieron presentes todo el año (Tabla
4), mientras que 12 sólo en los censos de primavera.
En La Junta, Choromoro, 22 especies eran residentes (Tabla 4), 15 de ellas coincidentes con
las residentes en Piedra Tendida. En primavera, 12 especies migratorias llegaron para criar o
invernar, 6 de ellas comunes a las de Piedra Tendida.
En Dique San Ignacio, 18 especies eran residentes (Tabla 4) y 18 estuvieron presentes sólo
en época de cría. Entre las residentes, 11 de ellas son compartidas con las residentes en La
Junta y 12 con las residentes en Piedra Tendida.
Comparando la diversidad entre hábitats en muestreos invernales, el Índice de Diversidad de
Simpson, dio como resultado un valor mucho menor para la localidad de Piquirenda Viejo, en
bosque de palo blanco en el extremo norte de Salta (D=0.54) y valores mayores para el
bosque de Tipa y Pacará de las localidades de Piedra Tendida (D=0.96), La Junta (D=0.93) y
San Ignacio (D=0.93). Este índice da más peso a las especies comunes y menos a las raras,
reflejando la probabilidad de que dos individuos tomados al azar en la comunidad sean de la
misma especie, por lo que el valor menor está destacando una mayor diversidad para
Piquirenda Viejo y una caída en la diversidad a lo largo del gradiente latitudinal de

PIEDRA TENDIDA LA JUNTA SAN IGNACIO
Crypturellus tataupa Crypturellus tataupa Crypturellus tataupa
Sarcoramphus papa Coragyps atratus Buteo magnirostris
Buteo magnirostris Buteo magnirostris Pionus maximiliani
Pionus maximiliani Amazilia chionogaster Leptotila verreauxi
Picumnus cirratus Campephilus leucopogon Chlorostilbon aureoventris
Synallaxis superciliosa Synallaxis superciliosa Eriocnemis glaucopoides
Phylidor rufosuperciliatus Phylidor rufosuperciliatus Picumnus cirratus
Ochtoeca leucophrys Sayornis nigricans Sittasomus griseocapillus
Sayornis nigricans Phylloscartes ventralis Synallaxis superciliosa
Phylloscartes ventralis Hemitriccus margaritaceiventer Phylidor rufosuperciliatus
Troglodytes aedon Serpophaga munda Sayornis nigricans
Cyanocorax chrysops Troglodytes aedon Hirundinea ferruginea
Turdus rufiventris Cyanocorax chrysops Phylloscartes ventralis
Myioborus brunniceps Turdus rufiventris Cyanocorax chrysops
Thraupis sayaca Myioborus brunniceps Turdus rufiventris
Saltator caerulescens Thraupis sayaca Thraupis sayaca
Cyanocompsa cyanea Saltator caerulescens Arremon flavirostris
Zonotrichia capensis Cyanocompsa cyanea Zonotrichia capensis
Arremon flavirostris
Zonotrichia capensis
Molothrus bonariensis
Cacicus chrysopterus
Tabla 4: Especies residentes en parcelas de estudios intensivos en localidades de la provincia
de Tucumán.
distribución del Bosque de Transición. Comparando la diversidad de aves dentro del hábitat,
en muestreos invernales en la localidad de Piedra Tendida, se aplicó el Índice de Diversidad
de Simpson para parcelas con dominancia de nogal (J. australis), donde en ciertas partes es
monoespecífico, y para el bosque con diversidad de especies arbóreas aunque con
dominancia de cebil (A. colubrina), dando valores similares (D=0.95 para el primero y
D=0.96 para el segundo). La diversidad de aves en la localidad de La Junta, en un muestreo
invernal, da valores similares para el bosque maduro y para un tuscal existente en las
márgenes del Río Choromoro (D=0.93 para el primero y D=0.92 para el segundo).
Relación entre la diversidad de altura del follaje y la diversidad de aves
No existe correlación entre la Diversidad de Altura del Follaje (DAF) y la Diversidad de
Especies de Aves (DEA) entre la época decídua y la lluviosa en parcelas de las tres
localidades de Bosque de Transición estudiadas de la provincia de Tucumán. El bosque sufre
una compensación estructural de estratos, en la época decídua aumenta la densidad del estrato
arbustivo y disminuye la densidad del estrato arbóreo superior. En primavera y verano el
estrato superior de las copas de los árboles se hace más denso y disminuye el estrato inferior
debido a la poca iluminación, por lo que al graficar la sumatoria de los porcentajes de
cobertura en relación al número de especies de aves, no resultan curvas significativas (Figs. 9
y 10). Por otro lado, la riqueza de especies de aves es similar en diferentes censos, invernales
o estivales, en localidades con bosques de una heterogeneidad estructural similar como lo son
las localidades de Tucumán.

Fig. 9 y 10: Porcentajes de cobertura en relación al número de especies de aves en Piedra
Tendida y San Ignacio.
Dominancia
La dominancia en el bosque de palo blanco de Piquirenda Viejo la constituían Basileuterus
bivittatus y Pyrrhura molinae durante el mes de Mayo y Arremon flavirostris y B. bivittatus
en época de cría. En el bosque de transición de Laguna La Brea, Synallaxis scutatus y
Basileuterus bivittatus eran dominantes en el mes de Julio.
En La Junta, la dominancia durante la época decídua estaba dada por A. flavirostris y
Syndactila rufosuperciliatus dentro del bosque y Carduelis magellanica y Zonotrichia
capensis en áreas abiertas invadidas por tuscales. El bosque pierde, en esta localidad, casi
completamente el follaje durante la época invernal, y la zona tiene influencia de las áreas
cultivadas próximas y áreas xerófilas del Chaco Serrano que se desarrollan a menor altura.
Vireo olivaceus y A. flavirostris dominaban en los censos de primavera.
En Piedra Tendida, durante la estación seca invernal, la dominancia estuvo dada por especies
que formaban bandadas uniespecíficas (Poospiza erythrophrys y Thraupis sayaca). En
Octubre dominaban Cyanocorax chrysops y T. sayaca. En San Ignacio, T. sayaca y
Myiborus brunniceps eran los dominantes en el invierno y A. flavirostris y Saltator
caerulescens, durante el mes de Diciembre (Tabla 5).
LOCALIDAD invierno primavera
Piquirenda Viejo 1.6
Basileuterus bivittatus
Pyrrhura molinae
0.66
Basileuterus bivittatus
Arremon flavirostris
Metán 0.60
Arremon flavirostris
Turdus rufiventris
La Junta 0.50
Arremon flavirostris
Philydor rufosuperciliatus
0.42
Vireo olivaceus
Arremon flavirostris
Piedra Tendida 0.36
Poospiza erythrophrys
Thraupis sayaca
0.54
Thraupis sayaca
Cyanocorax chrysops
San Ignacio 0.66
Myioborus bruniceps
Thraupis sayaca
0.39
Arremon flavirostris
Saltator caerulescens
Florida 0.80
Thraupis sayaca
Turdus rufiventris
TABLA 5: Especies dominantes en varias localidades de Bosque de Transicion.

Dominancia (ID = 100 (Ya+ Yb) / Yt)
ANALISIS ZOOGEOGRAFICO
El análisis zoogeográfico realizado de acuerdo a la presencia y ausencia de las especies de
aves por localidad dio como resultado:
a.- La determinación de una serie de especies importantes para definir cada localidad según el
Análisis de Correspondencia (CA) por filas. En la Fig. 11, en el diagrama de ordenamiento
de CA están representadas las localidades de muestreo, donde la localidad Metán aparece en
un punto intermedio entre las localidades de Salta y Tucumán, aunque más próxima a las de
Tucumán.
Fig. 11: Diagrama de Ordenamiento de CA. Ejes 1 y 2 de CA donde se representan las
localidades de muestreo. JUN: La Junta; IGN: Dique San Ignacio; PIE: Piedra Tendida;
MET: Metán; BRE: Laguna La Brea; PIQ: Piquirenda Viejo.
En la Fig. 12, en el diagrama de ordenamiento de CA se representan las especies de aves,
agrupadas por un número que las identifica. Varios números componen una localidad y
aparecen especies compartidas por dos o más localidades.
b.- El agrupamiento de localidades empleando la técnica UPGMA, separa tres grupos de
localidades, relaciona entre sí a las localidades de Tucumán y a las del norte de Salta y
sudeste de Jujuy, separando ambos grupos y distanciándolos a la vez de la localidad Metán.
En la Fig. 13 el agrupamiento de localidades se hizo sobre una Matriz de Distancia Chi- Cuadrado, y en la Fig. 14 sobre una Matriz de Distancia de Dice (= Coeficiente de Sorensen).
c.- El agrupamiento de especies (N= 190) según el Análisis de Coordenadas Principales sobre
una Matriz Chi- Cuadrado (Fig. 15), donde un grupo importante de especies son compartidas
por las tres localidades de Tucumán, por lo presentan gran similitud, mientras que Piquirenda
Viejo y Laguna La Brea presentan menor similitud, un grupo mayor de especies propias no
compartidas y se refleja la caída de especies. En este análisis no se consideró la localidad
Metán.
d.- El agrupamiento de las especies (N= 190), según el Análisis de Coordenadas Principales
sobre una Matriz Chi, donde un grupo importante de especies son compartidas por las tres
localidades de Tucumán, por lo que presentan gran similitud, mientras que Laguna La Brea y
Piquirenda Viejo presentan menor similitud, un grupo mayor de especies propias no
compartidas y y se refleja la caída de especies (Fig. 14). En este análisis no se consideró a la
localidad Metán.
e.- La determinación de una serie de especies importantes para definir cada localidad, según
el Análisis de Correspondencia (CA), por filas (Fig. 15).

33
Fig. 12: Diagrama de Ordenamiento de CA. Ejes 1 y 2 de CA donde se representan las
especies de aves. Cada número indica un grupo de especies. Referencias:
1.- Elanoides forficatus, Casiornis rufa, Picumnus dorbignianus, Turdus albicollis, Psaracolius
decumanus, Phaethornis pretrei, Suiriri suiriri, Micrastur semitorquatus, Batara cinerea,
Leptopogon amaurocephalus, Satrapa icterophrys, Turdus chiguanco, Caprimulgus saltarius,
Thamnophilus ruficapillus, Myiopagis viridicata.
2.- Ramphastos toco, Falco sparverius, Atlapetes torquatus.
3.- Tigrisoma fasciatum, Penelope obscura.
4.- Buteogallus urubitinga
5.- Claravis pretiosa, Glaucidium brasilianum, Dendrocolaptes picumnus, Pyrrhura molinae,
Pulsatrix perspicillata, Tamnophilus caerulescens, Veniliornis frontalis, Trogon curucui.
6.- Basileuterus bivittatus
7.- Piaya cayana, Icterus cayanensis
8.- Xyphocolaptes major, Turdus nigriceps
9.- Cychlaris gujanensis, Todirostrum plumbeiceps
10.- Herpsilochmus pileatus, Myrmorchilus strigilatus, Taraba major, Tolmomyas sulphurescens,
Tiaris obscura, Lepidocolaptes angustirostris, Falco rufigularis, Accipiter bicolor, Fluvicola
pica, Machethornis rixosa, Polioptila dumicola, Phacellodomus rufifrons, Pachyramphus
viridis, Knipolegus striaticeps, Catharus dryas, Diglossa baritula, Contopus fumigatus,
Certhiaxix pyrrhophia, Caprimulgus parvulus
11.- Columbina picui, Synallaxis frontalis
12.- Piculus rubiginosus, Pitangus sulphuratus, Serpophaga subcristata
13.- Guira guira, Myiarchus tyrannulus
14.- Poospiza melanoleuca
15.- Elaenia strepera, Serpophaga nigricans, Thraupis bonariensis, Molothrus badius, Elaenia
parvirostris
16.- Myioborus brunniceps, Campephilus leucopogon, Saltator caerulescens, Zonotrichia capensis,
Pionus maximiliani, Saltator aurantirostris
17.- Phylloscartes ventralis, Pheucticus aureoventris, Cacicus chrysopterus
18.- Myiarchus swainsoni, Micrastur ruficollis, Chlorospingus ophthalmicus
19.- Picumnus cirratus, Crotophaga ani, Columbina talpacoti

34
20.- Catharus ustulatus, Tyrannus melancholicus, Sayornis nigricans, Myiodynastes maculatus,
Chlorostilbon aureoventris
21.- Aegolius harrisi, Elaenia obscura, Amazona tucumana, Knipolegus cabanisi, Ochthoeca
leucophrys, Tyto alba, Geothlypis aequinoctialis
22.- Aratinga mitrata, Synallaxis superciliosa, Poospiza erythrophrys
23.- Basileuterus culicivorus, Atlapetes citrinellus, Empidonomus varius, Carduellis magellanicus,
Picumnus cirratus thamnophiloides, Eriocnemis glaucopoides, Molothrus bonariensis,
Cinclodes fuscus
24.- Merganetta armata, Poospiza nigrorufa, Nycticorax nycticorax, Troglodytes solstitialis,
Sporophila caerulescens, Notiochelydon cyanoleuca, Myiophobus fasciatus, Elaenia albiceps
25.- Aramides cajanea, Todirostrum margaritaceiventer, Serpophaga munda, Otus choliba,
Knipolegus aterrimus
26.- Piranga flava, Sittasomus griseocapillus, Coryphospingus cucullatus, Thlypopsis sordida
27.- Amazilia chionogaster, Pipraeidea melanonota, Philydor rufosuperciliatus, Vireo olivaceus,
Turdus amaurochalinus, Thraupis sayaca
28.- Amazona aestiva, Arremon flavirostris, Crypturellus tataupa, Turdus rufiventris, Parula
Pitiayumi, Cyanocompsa cyanea, Cyanocorax chrysops, Mecocerculus leucophrys,
Leptoptila verreauxi
Los distintos métodos de análisis identifican claramente las diferencias de la avifauna entre el
Bosque de Transición de palo blanco y el de Tipa y Pacará mostrando una zonación
latitudinal. Distancian claramente entre sí a la localidad del extremo norte de Salta con la del
sudeste de Jujuy y destacan las afinidades de las localidades de Tucumán, aunque agrupando
La Junta con San Ignacio y separando levemente a Piedra Tendida.
Fig. 13: Agrupamiento (UPGMA) de localidades sobre una matriz de Distanchia Chi-
Cuadrado. MET: Metán; IGN: San Ignacio; JUN: La Junta; PIE: Piedra Tendida; BRE:
Laguna La Brea; PIQ: Piquirenda Viejo.
Fig. 14: Agrupamiento (UPGMA) de localidades sobre una matriz de Distancia de Dice
(=Coeficiente de Comunidades de Sorensen). MET: Metám; IGN: San Ignacio; JUN: La
Junta; PIE: Piedra Tendida; BRE: Laguna La Brea; PIQ: Piquirenda Viejo.

35
Fig. 15: Dendrograma (presencia/ausencia) para 6 localidades, similitus de Sorensen,
UPGMA) que muestra el agrupamiento de las especies de aves.
Distribución de taxones de aves
Una parte importante de las aves de los Bosques Pedemontanos de Transición ocurren en
todo el rango de extensión de estos bosques en Argentina, cerca de 130 especies son
frecuentes de observar entre el norte de Salta y Tucumán. Varias especies tropicales como
Aburria pipile, Ara ararauna, Pteroglossus castanotis, Brotogeris versicolorus, Galbula
rufoviridis, Veniliornis passerinus, Xiphorhynchus guttatus, Synallaxis gujanenis,
Thamnophilus doliatus, Piriglena leuconota, Trithorus guarayanus, Basileuterus flaveolus y
Saltator similis, frecuentes en los Bosques de Transición de la Sierra de San José de los
Chiquitos, Departamento Santa Cruz, Bolivia, donde fueron registradas por Olrog (1963), no
alcanzan en su distribución el límite con Argentina.
Los No Passeriformes y los Oscines incluyen cada uno un tercio de los taxones de aves. Los
No Passeriformes representan el 40% del total de taxones, son de mayor tamaño y poseen
rangos de distribución mucho más amplios que los Passeriformes. Este porcentaje es menor
al que ocurre en el Chaco, donde aproximadamente la mitad de los taxones son No
Passeriformes (Short, 1975) y mayor al de las selvas del noreste, donde representan
aproximadamente el 30% (Lucero y Alabarce, 1980). Los rapaces de gran tamaño son
actualmente escasos, salvo las especies carroñeras. Abundan todavía los rapaces de tamaño
mediano y chico que se alimentan de aves, insectos y micromamíferos. Son representativos

36
un inambú (Crypturellus tataupa), tres especies de jotes (Coragyps atratus, Cathartes aura
y Sarcorhamphus papa) un carancho (Polyborus palcus), dos águilas, una de ellas piscívora o
cangrejera (Buteogallus urubitinga), un aguilucho (Buteo magnirostris), un elanio tijereta
(Elanoides forficatus), un esparvero especializado en la caza de aves (Accipiter striatus) y
dos halcones de bosque (Micrastur semitorquatus y M. ruficollis), una pava de monte
(Penelope obscura), varias especies de palomas (Leptoptila verreauxi, Columba cayenensis,
Columbina talpacoti), una maracaná del bosque de palo blanco (Ara auricollis), una cotorra
(Aratinga mitrata) y dos loros (Pionus maximiliani y Amazona aestiva), 3 especies de
lechuzas (Aegolius harrisi, Otus choliba, Glaucidium brasilianum) y dos lechuzones de gran
tamaño, uno de ellos sólo presente en el bosque de palo blanco (Pulsatrix perspicillata y Asio
stygius), 5 especies de picaflores, una de ellas sólo del extremo norte de Salta (Amazilia
chionogaster, Chlorostilbon aureoventris, Microstilbon burmeisteri, Eriocnemis
glaucopoides y Phaethornis pretrei) un surucuá, un burgo y un tucán que no llegan en su
distribución hasta Tucumán (Trogon curucui, Momotus momota y Ramphastos toco) y 6
especies de carpinteros, una de ellas característica sólo del sudeste de Jujuy (Picumnus
dorbignyanus, Picumnus cirratus, Veniliornis frontalis, Campephilus leucopogon y Piculus
rubiginosus). Las especies acuáticas presentes en los pocos espejos de agua existentes en
estos bosques (El Rey, Laguna La Brea), tienen todas una distribución amplia en los trópicos
de Sudamérica (Podylimbus podiceps, Podiceps dominicus, Anhinga anhinga, Phalacrocorax
olivaceus, Amazonetta brasiliensis, Cairina moschata).
Una característica importante del Bosque Pedemontano es la escasez de Suboscines
furnaroides en general (Familias Dendrocolaptidae, Furnaridae y Formicaridae), dominantes
en el noroeste de Sudamérica (Olrog, 1984).
La escasez de formicáridos distinguen a este bosque de los demás bosques del oeste de
Sudamérica (63 especies en Bolivia, más de 100 en Perú y 130 entre Ecuador y Colombia), (Olrog, 1984).
En cambio, en las selvas del noreste de Argentina (Lucero y Alabarce, 1980), abundan los
furnáridos Sclerurus scansor, Heliobletus contaminatus, varias especies del género Philydor
y Synallaxis y los formicáridos Mackenziaena leachii, Dysithamnus mentalis, dos especies
del Género Thamnophilus, dos especies del Género Drymophila, Pyriglena leucoptera y
Conopophaga lineata.
Los suboscines furnaroides comprenden el 14% del total de la avifauna. Sólo 17 especies han
sido registradas, 4 especies de dendrocoláptidos trepadores (Sittasomus griseocapillus,
Xiphocolaptes major, Lepidocolaptes angustirostris y Dendrocolaptes picumnus), una
remolinera (Cinclodes fuscus), 3 pijuies (Synallaxis superciliosa, S. frontalis y S. scutatus),
dos furnáridos trepadores (Certhiaxis pyrrhophia y Philydor rufosuperciliatus), un espinero
(Phacellodomus rufrifrons), dos bataráes y tres bataracitos (Batara cinerea, Taraba major,
Herpsilochmus pileatus, Thamnophilus ruficapillus y T. caerulescens) y un hormiguero
terrícola (Myrmorchilus strigilatus). Este número se reduce drásticamente desde el extremo
norte de Salta y sudeste de Jujuy, donde representan el 11 y el 14% del total de taxones
respectivamente, hacia el sur (9% en Metán, 7.5% en San Ignacio y 5% en Piedra Tendida).
En Tucumán D. picumnus, S. scutatus, B. cinerea, H. pileatus y M. strigilatus) no se
encuentran presentes.
Los Suboscines tiranoides son más numerosos, gracias a la abundancia de miembros de la
Familia Tyrannidae. Representan el 20% del total de especies, sin diferencias significativas a
lo largo del rango latitudinal de distribución de estos bosques. Están conformados por 3
anambés (Familia Cotingidae) y 40 especies de tiránidos (Familia Tyrannidae), el 32% de las
especies presentes en Argentina. Pocas especies de tiránidos son residentes en los Bosques
Pedemontanos de Argentina: Sayornis nigricans, de los cursos de agua; Todirostrum
plumbeiceps y T. margaritaceiventer, tiránidos pigmeos del estrato arbustivo denso;
Phylloscartes ventralis; Tolmomyias sulphurescens, migratorio en la parte sur de su rango de

37
distribución y Elaenia obscura, único fío-fío que permanece en estos bosques. Son todas
especies típicamente selváticas y con especies cogenéricas amazónicas. Otro grupo
importante de especies selváticas es migratorio, entre ellas Empidonax euleri, 3 especies del
Género Elaenia (E. strepera, E. albiceps y E. parvirostris) y Myiodynastes maculatus. Un
tercer grupo está relacionado a ambientes abiertos o bosques xerófilos sudamericanos como
Tyrannus melancholicus, Suiriri suiriri, Myiarchus swainsoni y Empidonomus varius.
Los Oscines son numerosos, especialmente hacia el sur de la distribución de los Bosques
Pedemontanos. Varían entre un 29% en el norte de Salta y un 40% en el sur de Tucumán,
conformándose principalmente por especies semilleras de la Familia Emberizidae como
pepiteros, monteritas, una especie de cabecita negra, dos jilgueros, un corbatita, y dos
arbusteros. Algunas de estas especies están estrechamente relacionadas con especies de
distribución tropical en el estrato bajo de selvas (Géneros Saltator, Coryphospingus,
Cyanocompsa, Arremon, Poospiza). Otros emberízidos, como Sicalis flaveola, S. luteola y
Sporophila caerulescens, evolutivamente más modernas, se encuentran en proceso de
radiación (Olrog, 1978), con la mayoría de las poblaciones subespecíficas entre los paralelos
30 y 45º de latitud sur, ocupando montañas bajas, valles y planicies. En el caso de las
cogenéricas Atlapetes torquatus, especie común en el Bosque Pedemontano de palo blanco, y
Atlapetes citrinellus, especie que aunque nidifica en el Bosque Montano pasa varios meses en
los pedemontes, se reemplazan geográficamente en el Bosque de Transición, siendo la
segunda común solo en la porción austral de estos bosques.
Características poblacionales locales y distribución de especies a lo largo del gradiente
latitudinal
En cada localidad existen poblaciones locales de alguna especie en particular que no estaba
presente en los otros sitios. En el extremo norte de Salta, en la localidad de Piquirenda Viejo,
es destacable la presencia de Sarcoramphus papa, Tigrisoma fasciatum, Micrastur semitorquatus, Pulsatrix perspicillata, Phaethornis pretrei, Trogon curucui, Turdus albicollis
y Psaracolius decumanus como especies residentes. Son éstas especies muy especializadas y
típicas de ambientes boscosos húmedos tropicales. Sólo las dos primeras se encuentran
también como especies residentes en Tucumán.
El 12% de las especies registradas llegan en su distribución hacia el sur sólo hasta el sudeste
de Jujuy como E. forficatus, M. semitorcuatus, A. auricollis, P. pretrei, P. dorbignyanus, D.
picumnus, Contopus cinereus, L. amaurocephalus, S. scutatus. Nueve especies observadas o
capturadas en el bosque de palo blanco de la Localidad de Laguna La Brea no llegan en su
distribución hasta la provincia de Tucumán. Entre las residentes, son destacables, Ara
auricollis, Pyrrhura molinae, Herpsilochmus pileatus, Myrmorchilus strigilatus y Catharus
dryas, especies tropicales que no se encuentran en el bosque de Tipa y Pacará.
Varias especies tienen su límite sur de distribución donde termina el bosque de palo blanco y
comienza el de Tipa y Pacará: Psaracolius decumanus fue observado hasta la localidad de
Lumbreras, Todirostrum plumbeiceps, Atlapetes torquatus, B. bivittatus y Ramphastos toco
son abundantes hasta Metán, S. scutatus llega hasta el Parque Nacional El Rey.
Las especies residentes constantes del Bosque de Transición de Tucumán, presentes en cada
uno de los censos eran: Entre las migratorias, 8 especies migrantes neotropicales estaban
presentes en los tres sitios, donde arriban a criar en primavera (Tyrannus melancholicus,
Myiodinastes maculatus, Elaenia strepera, E. parvirostris, Myiophobus fasciatus, Vireo
olivaceus, Turdus amaurochalinus y Sporophila caerulescens). Una llegó en forma regular
en primavera como migratoria desde el Hemisferio Norte a las tres localidades (Catharus
ustulatus). Este ensamble de 19 especies constantes puede considerarse como diagnóstico
para este tipo de bosque en la provincia de Tucumán, y la acerca mas a un bosque templado
que a uno tropical donde un número importante de especies son residentes.
Piedra Tendida tiene una población permanente del jote o cóndor real (Sarcoramphus papa),
con jóvenes y adultos. Esta especie no ha sido encontrada en las otras localidades de Bosque

38
de Transición, aunque sí en Horco Molle y en la Reserva Provincial de La Florida. La
garza de los torrentes (Tigrisoma fasciatum) estuvo presente en el Río Tajamar, en esa misma
localidad, en los censos de Junio y Agosto. Es una especie frecuente en los ríos en el bosque
pedemontano de la provincia de Tucumán, donde se ubica en zonas donde el río hace un
recodo o remanso y está cubierto por la sombra de los árboles. Ha sido observada en la
Reserva Provincial de La Florida (Río El Saltón), Horco Molle (Río Piedras), Piedras
Coloradas (Río Los Sosa), El Indio (en un afluente del Río Los Sosa). Es interesante la
presencia de Ochthoeca leucophrys, en todos los censos en esta localidad, aún en los de
primavera, cuando debería estar criando en el Bosque Montano Superior.
Durante este estudio, se capturó en Piedra Tendida una nueva especie para la provincia,
Todirostrum plumbeiceps viridiceps (Salvadori). Este pequeño tiránido es típico de la
vegetación arbustiva densa de bosques húmedos de Jujuy, Salta y sur de Bolivia (Olrog,
1978). También fue capturado en Piquirenda Viejo, Parque Nacional El Rey, Lumbreras y
Metán, todas en la provincia de Salta, y en Laguna La Brea, provincia de Jujuy. Es
interesante su presencia por primera vez para Tucumán, ya que es un ave residente, de
pequeño tamaño (8 gramos), con poca capacidad de vuelo. Las Sierras de Medina se
encuentran actualmente aisladas del resto de las masas boscosas húmedas del norte y del
oeste por vegetación xerófila y campos cultivados, lo que hace pensar en una población
relictual aislada que ha permanecido en estas sierras y no fue notada anteriormente.
La Junta posee poblaciones permanentes de Merganetta armata, Campephilus leucopogon y
Cacicus chrysopterus, que están ausentes de las otras localidaes. La presencia del pato de
los torrentes no había sido registrada con anterioridad para esta localidad, aumentándose su
distribución conocida para la provincia de Tucumán. Ha sido capturado un ejemplar macho
joven en Agosto y se han observado parejas en los Ríos Rearte y Choromoro durante el
invierno y la primavera. Debido a los tuscales que crecen en el amplio cauce del Río Choromoro, son frecuentes las bandadas de Carduelis magellanica, Coryphospingus
cucullatus, Poospiza nigrorufa y P. melanoleuca durante la época invernal. Además, se
registraron especies que no fueron observadas en las otras localidades de Bosque de
Transición de Tucumán como Charadrius collaris, Certhiaxis pyrrhophia y Cinclodes
atacamensis. Fue capturada en Agosto en esta localidad Zonotrichia capensis australis, raza
patagónica del chingolo que cría en Chubut, Santa Cruz y Tierra del Fuego, migrando en
otoño por el oeste hasta Salta, Jujuy y Bolivia (Olrog, 1979).
En Dique San Ignacio existen poblaciones permanentes del águila negra pescadora
Buteogallus urubitinga, de los picaflores Eriocnemis glaucopoides y Microstilbon
burmeisteri y Xiphocolaptes major, el mayor de los dendrocoláptidos argentinos. Llega todos
los años a criar la golondrina negra Progne modesta, que ha sido capturada en condición
reproductiva en el mes de Diciembre. Existe en el lugar una población de media centena de
individuos que anidan en una vivienda abandonada. El loro alisero, Amazona tucumana, ha sido observado durante el invierno en pequeños grupos. Los martín - pescadores Ceryle torquata y Chloroceryle americana se encuentran en forma permanente y C. amazona sólo durante el verano.
Es la localidad donde arriban a criar en primavera la mayor cantidad de especies migratorias
latitudinales, entre ellas 3 especies del Género Elaenia. San Ignacio fue la localidad de
Tucumán con mayor abundancia en número de especies durante la estación reproductiva.
LA ESTACIONALIDAD Y SU INFLUENCIA EN LA AVIFAUNA
El clima, que determina una estación seca, de Mayo a Octubre, y una lluviosa, influye en la
marcada estacionalidad que tiene el Bosque de Transición, donde la mayor parte de los
árboles pierden total o parcialmente el follaje (Lista 5) durante el invierno. Las especies
perennifolias están representadas por árboles o arbolitos del estrato medio como E. uniflora,
A. edulis, S. buxifolia y E. pungens. Durante la estación seca, el suelo se cubre de hojarasca y
hay una buena iluminación del estrato arbustivo y herbáceo que le permite desarrollarse. La

39
cobertura arbórea es entonces escasa y está representada por árboles de altura media. Las
Figs. 16 y 17 muestran comparativamente el porcentaje de cobertura en época seca y lluviosa
de parcelas de las tres localidades estudiadas. La característica más marcada es la densidad
que adquiere el estrato arbustivo y herbáceo durante la época decídua. Varias especies de
estos estratos florecen (S. verbascifolium, S. clavatum, S. tucumanum, Cnicothamus sp, R.
boliviensis) y fructifican (Fragaria sp, C. iguanae, P. carthagenensis) en la estación seca.
Durante el verano la cobertura más densa ocurre entre los 10 - 20 m y el estrato arbustivo es
proporcionalmente menos denso y más sombreado que durante le invierno. La floración de la
mayor parte de las especies arbóreas ocurre entre Agosto y Octubre, y el período de
fructificación comienza en Octubre, alcanza su máximo en Noviembre y declina a partir de
Enero.
Shinopsis haenkeana* Horco quebracho
Jacaranda mimosifolia* Tarco
Tabebuia impetiginosa* Lapacho
Tecoma stans* Guarán
Cordia trichotoma* Afata grande
Patagonula americana. Guayaibí
Terminalia triflora Lanza amarilla
Caesalpinia paraguarensis* Guayacán
Gleditsia amorphoides Espina de corona
Acacia etilis* Garabato guiador
Anadenanthera colubrina* Cebil colorado
Enterolobium contorstisiliquum* Pacará
Parapiptadenia excelsa* Horco cebil
Prosopis nigra* Algarrobo negro
Amburana cearensis* Roble del país
Tipuana tipu* Tipa
Phoebe porphyria Laurel de la falda
Cedrela lilloi* Cedro
Cedrela odorata* Cedro salteño
Rapanea laetenvirens Falso San Antonio
Eugenia uniflora Arrayán
Myrcianthes mato Horco mato
Myrcianthes pungens Mato
Piper tucumanum Lata de pobre
Ruprechtia laxiflora virarú
Scutia buxifolia Coronillo
Calycophyllum multiflorum* Palo blanco
Solanum granuloso* Tabaquillo
Vassobia parviflorus Candelilla
Heliocarpus popayensis* Afata blanca
Phyllostylon rhamnoides* Palo amarillo
Juglans australis* Nogal criollo
Lista 5: Especies arbóreas presentes en los Bosques Pedemontanos de Transición.
* Especies decíduas.
Las especies con frutos secos (T. tipu, T. triflora), de dispersión anemófila fructifican
durante la época seca y forman el dosel del bosque. Los árboles predados por loros (A.
colubrina, P. excelsa), fructifican durante el verano pero las vainas secas permanecen en las
plantas durante todo el año y son intensamente consumidas.

40
Fig. 16: Porcentaje de cobertura en diferentes estratos en época decidua y lluviosa en el
Bosque de Transición de San Ignacio.
Fig. 17: Porcentaje de cobertura en diferentes estratos en época decidua y lluviosa en el
Bosque de Transición de Piedra Tendida.
En los tres sitios estudiados en Tucumán, además de Horco Molle con Bosque Pedemontano
Húmedo, había parches con abundancia de P. carthagenensis en el estrato arbustivo. Este
arbusto o arbolito de hasta 4 m de altura, comienza a producir frutos todavía verdes a partir
de Mayo y va fructificando durante el otoño e invierno paulatinamente, alcanzando su pico
de fructificación en Julio - Agosto, dando gran cantidad de frutos pequeños drupáceos que
son intensamente consumidos por aves. Otra especie de frutos carnosos también importantes
para las aves durante el invierno es C. iguanae, planta apoyante de frutos anaranjados de 2
cm de diámetro, que fructifica abundantemente durante el mes de Junio.
El brote de las hojas ocurre en Setiembre, dependiendo esto del clima (lluvias, temperatura),
pero no determina cambios fundamentales en el juego de especies de aves. A fines de
Setiembre, comienzos de Octubre, con la llegada de las lluvias, comienzan a fructificar
numerosas especies del estrato arbustivo y arbóreo (R. boliviensis, Morus nigra), alcanzando

41
el pico de fructificación en Noviembre - Diciembre para el Bosque de Transición de la
provincia de Tucumán (Fig. 18). Las especies que son consumidas por aves entre Octubre y
Diciembre pueden verse en la Fig. 19, de ellas las más importantes son R. boliviensis y M.
nigra en Octubre y Vassobia parviflorus, A. edulis y R. boliviensis en Diciembre.
Fig. 18: Número de especies de plantas fructificantes, picos de fructificación y fecha de
arribo de migranttes latitudinales.
. 19: Epoca de fructificación de algunas especies comunes en el Bosque de Transición.
*Especies de frutos consumidos por aves.
DINAMICA DE ESPECIES DE AVES EN LOS BOSQUES PEDEMONTANOS DE
TRANSICION
Una buena parte de la avifauna del Bosque de Transición en la provincia de Tucumán
presenta desplazamientos migratorios, distinguiéndose dos tipos de migrantes: migrantes
latitudinales y migrantes altitudinales o locales. Dentro de los migrantes latitudinales son
considerados en este trabajo tres grupos de especies según el área donde crían y el uso que
hacen del Bosque de Transición: a) Migrantes Neotropicales que crían en el Sur de

42
Argentina e invernan en las provincias norteñas y países limítrofes, b) Migrantes
Neotropicales que crían en el Bosque de Transición del noroeste Argentino y migran luego
hacia el norte, la Amazonia o las Yungas de Perú, Bolivia y Ecuador, c) Migrantes Neárticos
que crían en el Hemisferio Norte e invernan en el Neotrópico, llegando en su distribución
hacia el sur hasta Argentina.
Migrantes latitudinales Neárticos
-Falco peregrinus anatum Bonaparte. Llega cada año al norte de Argentina, desde sus
áreas de cría en Canadá y Estados Unidos (Wetmore, 1965). Durante este estudio ha sido
registrado en Campo Quijano, Salta, observado regularmente en la localidad de Horco
Molle y en una oportunidad en la represa de San Ignacio.
-Elanoides forficatus forficatus (Linné). Nidifica en gran parte del este de Estados Unidos,
especialmente en la costa Atlántica sur y en la del Golfo de Méjico, siendo abundante en el
sur de Florida (Wetmore, 1965). Las poblaciones de la parte norte de su área de nidificación
migran regularmente hacia el sur, llegando hasta Argentina. Ha sido observado en
Piquirenda Viejo en un grupo de 4 individuos en el mes de Noviembre.
-Empidonax trailii trailii (Audubon). Cría en América del Norte, desde Alaska y Nueva
Escocia hasta Baja California y Virginia. Inverna desde el sur de Méjico hasta Argentina.
Durante este estudio ha sido registrado únicamente en Piquirenda Viejo en el mes de
Noviembre.
-Catharus ustulatus swainsoni (Tschudi). Este pequeño zorzal cría en los bosques del Este
de Estados Unidos y Canadá (Rappole y Warnes, 1980) y arriba al Bosque de Transición a
fines de Octubre (años 1989, 1990) o comienzos de Noviembre (año 1988). Los datos
obtenidos revelaron que permanece en la provincia hasta comienzos de Febrero en que
desaparece. Encontramos que es una especie de presencia regular en los pedemontes de la
provincia de Tucumán año a año, y que es común a las tres localidades estudiadas en intensidad.Su arribo, en pequeños grupos de 8 - 10 individuos fue observado en Burruyacu
el 17 de Octubre de 1989, capturándose 4 ejemplares que pesaron 25 gr., peso inferior en 6
gramos al peso de individuos capturados en otras áreas de Tucumán, Salta y Jujuy en
Noviembre, Diciembre y Enero. Las aves recién arribadas dependían inmediatamente de los
frutos de R. boliviensis y M. nigra en Burruyacu y de V. pauriflora y R. bolivienis en
Diciembre en San Ignacio. Durante su permanencia en el Bosque de Transición son
solitarios y aparentemente territoriales. Un ave marcada el 16 de Diciembre en San Ignacio
fue vista en los dos días subsiguientes en un árbol de V. pauriflora. En 6 horas de censo, las
visitas de esta especie a este árbol representaron el 2.5% de la explotación total por parte del
resto de los frugívoros (Turdus rufiventris, T. amaurochalinus, E. albiceps, E. strepera, T.
sayaca, Thlypopsis sordida, V. olivaceus, Saltator caerulescens, Arremon flavirostris,
Zonotrichia capensis) durante el día.
En el Bosque de Transición de palo blanco del Norte de la provincia de Salta fueron
registradas Empidonax traillii, C. ustulatus y Elanoides forficatus. Arriban para invernar en
la misma época, aunque en el caso de E. forficatus, existe también una población residente.
Migrantes Latitudinales Neotropicales
En el Bosque de Transición, los migrantes que allí nidifican y que realizan largos
desplazamientos hacia el norte, que denomino migrantes neotropicales, arriban en forma
sincrónica justo antes que se produzca el pico de fructificación de las especies vegetales de
las cuales se alimentan (Fig. 20). En las tres localidades estudiadas en la provincia de
Tucumán, existe una constancia en cuanto al uso del bosque por parte de los migrantes,
tanto por el número de especies visitantes, como por el tiempo de utilización. La llegada y
partida de migrantes, tanto los que invernan como los que crían en el Bosque de Transición,
es sincrónica a lo largo de los pedemontes en la provincia de Tucumán, variando año a año
según las condiciones climáticas imperantes en la región.

43
Fig. 20: Número de plantas fructificantes, pico de fructificación y fechas de arribo y partida
de migrantes.
La relación de las especies residentes versus las migratorias puede verse en las Figs. 21, 22
y 23, donde se manifiestan dos picos importantes de impacto de especies que arriban a
invernar o a criar, uno en invierno y otro en primavera (Figs 22 y 23). En la localidad San
Ignacio, la afluencia invernal de migrantes altitudinales es menos marcada que en las otras
dos localidades (Fig. 21), que son más importantes como lugar de invernada.
Fig. 21: Relación Residentes versus Migrantes en la Localidad del Dique San Ignacio.
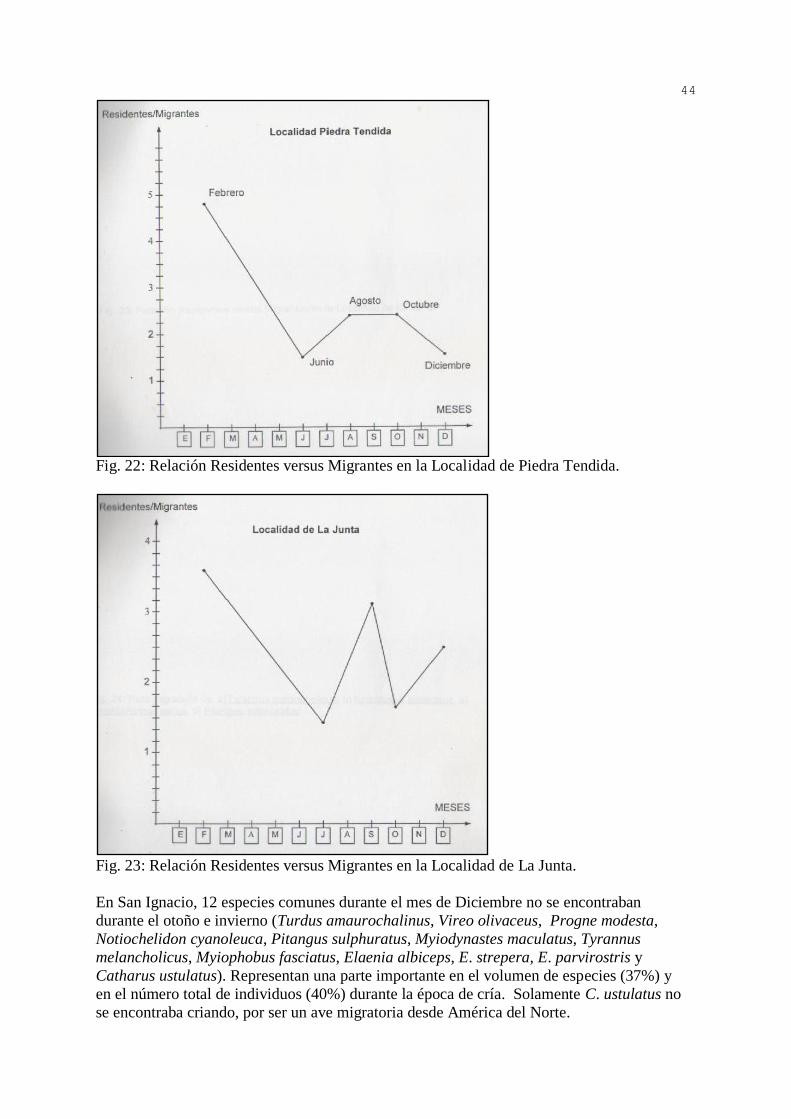
44
Fig. 22: Relación Residentes versus Migrantes en la Localidad de Piedra Tendida.
Fig. 23: Relación Residentes versus Migrantes en la Localidad de La Junta.
En San Ignacio, 12 especies comunes durante el mes de Diciembre no se encontraban
durante el otoño e invierno (Turdus amaurochalinus, Vireo olivaceus, Progne modesta,
Notiochelidon cyanoleuca, Pitangus sulphuratus, Myiodynastes maculatus, Tyrannus
melancholicus, Myiophobus fasciatus, Elaenia albiceps, E. strepera, E. parvirostris y
Catharus ustulatus). Representan una parte importante en el volumen de especies (37%) y
en el número total de individuos (40%) durante la época de cría. Solamente C. ustulatus no
se encontraba criando, por ser un ave migratoria desde América del Norte.

45
En Piedra Tendida, 8 especies abundantes a fines del mes de Octubre, no estaban presentes
durante la época invernal (T. melancholicus, Empidonomus varius, M. maculatus,
Myiarchus swainsoni, T. amaurochalinus, V. olivaceus, Carduelis magellanicus y C.
ustulatus). Representaban el 23,4% de las especies presentes y el 26, 5% del número total
de individuos.
En La Junta, el 35% de las especies comunes durante el mes de Octubre, migran después de
criar y desaparecen hasta la primavera siguiente (T. melancholicus, E. varius, M. maculatus,
Myiarchus tyrannulus, M. fasciatus, E. parvirostris, E. strepera, Serpophaga subcristata, T.
amaurochalinus, V. olivaceus, Sporophila carulescens) o luego de invernar como C.
ustulatus. Representan el 33,9% del número total de individuos. Los datos obtenidos en
este trabajo indican que el grueso de las especies migratorias neotropicales de largo
recorrido, residentes de verano en el Bosque de Transición, está constituida por miembros
de la Familia Tyrannidae.
Localidades La Junta Piedra Tendida San Ignacio
INVIERNO
N° de especies 33 36 36
Migrantes altitudinales 9 (27%) 8 (22%) 6 (16.6%)
PRIMAVERA
N° de especies 36 33 33
Migrantes latitudinales (ML) 11 (35%) 8 (23.4%) 12 (37%)
% de individuos ML 33.9% 26.5% 40.0%
Proporción de migrantes altitudinales y latitudinales en la provincia de Tucumán
Sólo unas pocas son residentes, la mayoría migra hacia el Norte luego de criar, correspondiendo a las siguientes especies: FAMILIA TYRANNIDAE
Tyrannus melancholicus melancholicus Vieillot. Arriba a criar a fines de Setiembre y
permanece hasta fines de Abril, en que migra. Durante su permanencia en el Bosque de
Transición es abundante en el estrato alto de vegetación, en las copas de los árboles.
Respecto a los lugares donde inverna, ha sido registrada en el Oeste del Matto Grosso
(Willis y Oniki, 1990), de donde desaparece a comienzos de Agosto, y por Fitzpatrick para
Perú (1980). Ha sido capturada en Río das Mortes, Brasil Central a fines de Agosto y
comienzos de Setiembre (Pinto y De Camargo, 1952). No fue registrada para Bolivia ni por
Olrog (1963) ni por Pearson (1980) durante el invierno ni primavera, por lo que pareciera
migrar a través del Chaco hacia el Pantanal de Brasil (Fig. 24 a).
Knipolegus striaticeps (D Orbignyi y Lafresnaye). Es un ave de distribución chaqueña, en
Argentina cría desde La Rioja y Córdoba, por todo el Norte, inclusive el Chaco paraguayo
(Short, 1975). En Tucumán, es común en el Bosque de Transición sólo a lo largo de la
vegetación arbustiva de los ríos. Fue encontrada criando en El Cadillal y en Piedra Tendida
entre Octubre y Diciembre. Migra luego al norte a Bolivia, Paraguay y Brasil (Olrog, 1984),
(Fig. 24 b) Ha sido registrada para el pantanal de Mato Grosso por Willis y Oniki (1990).
Se la capturó entre Octubre y Marzo en el Bosque de Transición. Un ejemplar capturado en
Mayo en El Cadillal, probablemente sea parte de una población más austral que migre hacia
las provincias del Norte Argentino.
Empidonomus varius varius (Vieillot). Es una especie común en el Bosque de Transición y
áreas chaqueñas aledañas a partir de Setiembre. Tiene amplia distribución en Argentina en
sabanas arboladas y bosques abiertos. En los bosques pedemontanos de la provincia
permanece hasta Marzo. Hay registros de su permanencia invernal en Bolivia, Perú y
Ecuador, de donde desaparece en verano (Pearson, 1980). Llega hasta Colombia donde se la
capturó en los Llanos Orientales y en la Amazonia colombiana en Mayo y Agosto (Olivares,

46
Fig. 24: Ruta migratoria de: a)Tyrannus melancholicus, b) Knipolegus striaticeps, c)
Empidonomus varius, d) Pitangus sulphuratus.
Fig. 25: Ruta migratoria de: a) Myiarchus tyrannulus, b) Myiarchus swainsoni, c)
Empidonax euleri, d) Myiodynastes maculatus.
Fig. 26: Ruta migratoria de: a) Elaenia albiceps, b) Elaenia strepera, c) Elaenia parvirostris,
d) Myiophobus fasciatus.
Fig: 27: Ruta migratoria de: a) Vireo olivaceus, b) Notiochelidon cyanoleuca, c) Progne
modesta, d) Turdus amaurochalinus.

47
1959). Ha sido registrada para el Estado de Mato Grosso como visitante de invierno (Pinto
y De Camargo, 1948). Fig. 24 c.
Pitangus sulphuratus bolivianus (Lafresnaye). Su migración ha sido probada mediante
recuperación de un ave anillada. Olrog (1963), marcó ejemplares en el Bañado de Figueroa,
Santiago del Estero, uno de los cuales fue recuperado en Santa Catalina, Brasil, luego de 6
años (Lucero, 1982). Es un ave común durante el invierno en el Oeste del Matto Grosso
(Willis y Oniki, 1990), de donde desaparece en verano. En el Bosque de Transición en la
Provincia de Tucumán lo hemos encontrado todo el año, aunque en mucha menor
proporción durante el invierno. Probablemente distintas poblaciones tengan distintos
patrones migratorios, con desplazamientos altitudinales y latitudinales y seguramente
algunas poblaciones del centro de Argentina invernan en Tucumán. Fig. 24 d.
Myiarchus tyrannulus tyrannulus (P.L.S. Muller). Llega a criar a mediados de Setiembre,
tornándose por unos días muy abundante en plantas fructificantes de M. nigra, R. boliviensis
y P. carthagenensis. Permanece hasta Abril, en que desaparece, de manera menos
perceptible que su arribo, que es notorio. Fue observada en migración en Agosto en el Sur
de Bolivia por Olrog (1963). Fue observada por Willis (1975) en bosques del oeste de Mato
Grosso en el Río Paraguay superior en Julio. Posteriormente, Willis y Oniki (1990) lo citan
como un visitante invernal en Matto Grosso. Fue capturado en Julio en el Sudeste de Jujuy
en Laguna La Brea y en Agosto en Orán, provincia de Salta. Estos datos establecen con
seguridad que poblaciones del centro del país invernan en Salta y Jujuy y probablemente las
del Norte Argentino lo hagan en Bolivia, Paraguay y Brasil. Fig. 25 a.
Myiarchus swainsoni swainsoni Cabanis y Heine. Arriba a criar a fines de Setiembre y
permanece hasta Febrero en la provincia de Tucumán. Al igual que la especie anterior, gran
parte de su dieta es frugívora. Llega en migración hasta Venezuela, donde ha sido capturada
en Bolívar en el mes de Junio y hasta Colombia donde fue coleccionada en la Sabana de Bogotá en el mes de Junio (Olivares, 1959), (Fig. 25 b). Fue capturada por Olrog (1963) en
Bolivia a comienzos de Setiembre y observada en comportamiento migratorio en zona de
Bosque de Transición en el Sur de Bolivia. Durante este estudio fue capturada a fines de
Agosto en Orán, Salta, y por su abundancia mostraba estar en migración.
Empidonax euleri argentinus (Cabanis). Cría en el Bosque de Transición y en la Selva
Montana. En el primero fue capturado a partir de mediados de Octubre y hasta Febrero,
aunque no es una especie abundante en los pedemontes, prefiriendo la Selva Montana,
donde sí es abundante en las zonas arbustivas densas cercanas a los ríos en verano. Fue
reportada como migrante invernal por Pearson (1980) en Bolivia y Perú, y por Fitzpatric
(1980) en el Parque Nacional Manu, Perú, por lo que puede establecerse ya una ruta
migratoria a lo largo de las Yungas (fig. 25c).
Myiodynastes maculatus solitarius (Vieillot). Es una especie muy común a partir de
mediados de Octubre y hasta Marzo. Arriba los primeros días de Octubre a Burruyacu y
San Ignacio. Depende también de frutos de R. boliviensis, R. laetenvirens, M. nigra y P.
porphyria. Migra luego de criar a Bolivia, Perú y Matto Grosso (Pearson, 1980; Fitzpatric,
1980; Willis y Oniki, 1990). También llega en migración hasta Colombia, donde ha sido
colectada en el mes de Junio en los Llanos del Meta en el mes de Junio (Nicéforo María,
1948: 208). Fig. 25 d.
Elaenia albiceps albiceps (D Orbignyi y Lafresnaye). Ha sido capturada durante este
estudio a partir de fines de Setiembre en Bosques de Transición y Selva Basal en la
provincia de Tucumán. Llegan en forma sincrónica junto a otras especies de Elaenia.
Utiliza los Bosques Pedemontanos sólo como lugar de paso, ya que asciende a criar al
Bosque Montano Superior, donde lo hace en Diciembre y Enero. Es muy abundante en los
alisales hasta Marzo, en que desaparece. Migra a la Amazonia de Brasil (Fig. 26 a). Ha
sido colectada en Río Taquarucu (Pinto, 1950) y registrada por Willis y Oniki (1990) para
la Serra das Araras en el Sudoeste de Mato Grosso.

48
Elaenia strepera Cabanis. Es común en el Bosque de Transición y la Selva Basal durante
el verano, prefiere el estrato bajo y arbustivo denso, donde cría. Ha sido encontrada criando
en San Ignacio y Horco Molle. Arriba a nidificar a fines de Setiembre o comienzos de
Octubre. Migra luego por las Yungas hacia Bolivia y Perú, donde inverna (Fig. 26 b). Fue
registrada como ave de paso en Tumi Chucua, Bolivia, en el mes de Setiembre, y como
residente de invierno en Yarinacocha, Perú (Pearson, 1980).
Elaenia parvirostris Pelzeln. Es común en las zonas ecotonales entre el Bosque de
Transición y el Chaco. Arriba a fines de Setiembre o comienzos de Octubre, según los años.
Llega en migración hasta el Norte de Sudamérica (Fig. 26 c). Ha sido colectada en Cali,
Colombia, en el mes de Mayo por Olivares (1959) y el autor menciona que fue capturada en
otras localidades de ese país entre Abril y Octubre. Fue coleccionada también en el sur de
Venezuela, en Puerto Ayacucho, en el mes de Mayo (Friedman, 1948); y en el oeste del
Mato Grosso, en el mes de Julio por Willis (1975).
Myiophobus fasciatus flammiceps (Temminck). Es el tiránido más común en zonas con
pastizales, arbustivas o con tuscales o matorrales cerca del agua en el Bosque de Transición.
Estas son áreas de bordes, alteradas o junto a campos cultivados. Llega a criar a comienzos
de Octubre y ha sido capturado hasta fines de Marzo. Migra hacia Brasil y Perú (Fig. 26 d).
Ha sido colectada en Río das Mortes, Mato Grosso (Pinto y De Camargo, 1952), y en el
Pantanal de Brasil (Willis y Oniki, 1990). Fitzpatrick (1980), la reportó como poco común
para la Estación Biológica Cocha Cashu, Parque Nacional Manu, Perú, en áreas arbustivas
abiertas de bordes o márgenes arbustivas de ríos sólo durante el invierno. FAMILIA VIREONIDAE
Vireo olivaceus chivi (Vieillot). Es muy común durante el verano en todas las localidades
estudiadas. Fue observado su arribo en migración el 6 de Octubre de 1989 en Burruyacu,
donde se capturaron 25 ejemplares. Un ejemplar anillado ese día fue recuperado el 15 de
Diciembre en San Ignacio (unos 200 km hacia el sur), indicando la dirección de la
migración. En San Ignacio se encontraba criando. Durante el invierno desaparece
totalmente de Tucumán, no así del Bosque de Transición de Palo Blanco de Salta y Jujuy,
donde es abundante y se lo ha observado formando parte de bandadas mixtas de aves en
Laguna La Brea, Jujuy. Probablemente las poblaciones más australes de Argentina invernan
a lo largo del Bosque de Transición de Salta, Jujuy y Bolivia, donde Olrog lo cita para las
Yungas durante el invierno (1963). Llega en migración hasta Colombia (Fig. 27 a), donde
ha sido capturada en el mes de Mayo en los LLanos Orientales (Olivares, 1959). FAMILIA HIRUNDINIDAE
Notiochelidon cyanoleuca cyanoleuca (Vieillot). Pertenece a la familia Hirundinidae.
Grandes bandadas arriban a Tucumán a fines de Setiembre y es la golondrina más común
durante todo el verano en todos los ambientes boscosos húmedos de la provincia de
Tucumán. Durante el invierno también se encuentra presente, aunque en menor número.
Según Olrog (1979), migran luego de criar a la Amazonia (Fig. 27 b). Willis y Oniki
(1990), observaron su migración hacia el sur el 6 de Agosto de 1987 y días subsiguientes en
el Pantanal de Matto Grosso.
Progne modesta elegans Baird. Una colonia nidificante fue encontrada en una vivienda
abandonada en Dique San Ignacio. Unos 30 individuos fueron registrados en los censos de
Octubre y Diciembre en dicha localidad. Según Narosky e Yzurieta (1987), migran luego
de criar hasta Perú, Bolivia, Paraguay, Brasil y Guyanas (Fig. 27 c). Ha sido registrada
como residente de invierno en Yarinacocha, Perú y como pasajera en Tumi Chucua, Bolivia,
por Pearson (1980). FAMILIA TURDIDAE
Turdus amaurochalinus Cabanis. Tiene una distribución amplia. Cría en el Bosque de
Transición y en el Bosque Chaqueño. Fue encontrada nidificando en La Junta, Piedra
Tendida y Dique San Ignacio en primavera. En el Bosque de Transición de Tucumán, Salta
y Jujuy es migratoria, ya que estuvo ausente de los censos del otoño e invierno. No así del

49
Bosque Chaqueño, donde estaba presente durante el invierno, especialmente cerca de ríos y
aguadas. Fue capturada en Junio en Antillas, sur de la provincia de Salta; en Julio en
Tartagal, provincia del Chaco, en las márgenes del Río Teuco; y en Agosto en Tonono, al
norte de la provincia de Salta. La primera de estas localidades pertenece al Chaco Serrano y
las restantes son chaqueñas. Se distribuye también por Brasil, donde fue encontrada
nidificando en el Pantanal y Passárgada, sudoeste de Mato Grosso, y como migratoria en
Monte Alegre, Estado de Sao Paulo (Pinto, 1949); Río Tacuarucu (Pinto, 1950); Río
Paraguay Superior (Willis, 1975); y Río de Cágado y Campos do Encanto, en el sudoeste de
Mato Grosso (Willis y Oniki, 1990), donde formaba bandadas de invierno. Según Olrog
(1978), se distribuye por Uruguay, Paraguay, Bolivia, sudeste de Perú y Brasil, llegando
hasta Amazonas como migratoria (Fig. 27 d). FAMILIA EMBERIZIDAE
Sporophila caerulescens caerulescens (Vieillot). Cría en el bosque pedemontano en áreas
arboladas próximas a campos cultivados. Se encontraba presente en primavera y verano en
las tres localidades estudiadas, aunque era común sólo en San Ignacio. Cría a partir de
Octubre y hasta fines de Marzo. Encontramos machos con protuberancias cloacales y canto
completo a mediados de Marzo. Probablemente tengan más de una puesta durante la época
de cría. Migran a mediados de Abril hacia el norte, desapareciendo de los Bosques de
Transición. Esta especie cría desde el norte y este de Bolivia y el centro de Brasil hacia el
sur, a través de Paraguay, Uruguay y el norte y centro de Argentina. Las poblaciones de
Argentina son migratorias, moviéndose hacia el norte en el otoño. Pequeños grupos se
encuentran durante este tiempo en la Amazonia (este de Perú, extremo sureste de Colombia
y Amazonia central de Brasil), (Ridgely y Tudor, 1989). En Tucumán arriban en migración
a fines de Setiembre.
Migrantes Neotropicales desde el sur de Argentina
Varias especies migratorias desde el Sur de Argentina fueron registradas en áreas abiertas o
de borde, campos cultivados cercanos, playas de diques y lagunas y a lo largo de ríos, pero
nunca dentro del bosque propiamente dicho, formando parte o integrándose a la comunidad
de aves residentes. Durante el mes de Mayo en zonas cercanas al Dique El Cadillal se
registró la presencia de Lessonia rufa, Anthus sp y Agriornis murina en bandadas mixtas de
más de 100 individuos, en claro comportamiento migratorio. Un grupo de Knipolegus
aterrimus, de 15 individuos, fue observado en el borde del espejo de agua del Dique San
Ignacio a fines de Mayo por un día. Bandadas enormes de Sturnella loica fueron
observadas en campos cultivados de Horco Molle durante una semana a comienzos de Abril,
desapareciendo luego. Bandaditas de Cinclodes fuscus son comunes durante el otoño a lo
largo de los ríos de la llanura tucumana. Mimus triurus comienza a hacerse común en Mayo
y permanece hasta Junio aunque siempre en áreas abiertas arboladas o de borde con el
chaco, fue observada en San Ignacio y Piedra Tendida. Embernagra platensis es un ave
observada en las zonas de borde del bosque de Transición, pastizales y arbustales abiertos, a
comienzos del otoño en grupos conspicuos. Bandadas enormes de Sicalis luteola, de más de
500 ejemplares, fueron observadas en Agosto en la Reserva Provincial La Florida, en claro
comportamiento migratorio.
Los resultados del monitoreo de aves durante todo el año en las tres localidades estudiadas
en Tucumán dan como resultado que los Bosques Pedemontanos son zonas de paso para
migrantes al comienzo del otoño (Pitangus sulphuratus, Myiarchus tyrannulus, Turdus
amaurochalinus, Myioborus brunniceps, Parula pitiayumi, Thraupis bonariensis), por el
comportamiento grupal en bandaditas de algunas especies. Seguramente pertenecen a
poblaciones más australes, pero no fue posible conocer con seguridad si es que estos
migrantes provienen desde el sur de Argentina o forman parte del ensamble de especies de
bosques a más altura.

50
Migrantes altitudinales o locales
Son aves que crían por lo general en el Bosque Montano Superior y pastizales húmedos de
altura por arriba de los 1500 m y descienden luego de criar hacia niveles más bajos. Estas
especies utilizan al bosque de Transición como área de invernada, arribando en forma
menos sincrónica que los migrantes latitudinales a partir del mes de Abril, y permanecen en
estos bosques hasta Setiembre en que comienzan a ascender hasta los pisos superiores de
vegetación en los cerros (Fig. 28).
Fig. 28: Migrantes altitudinales en el Bosque de Transición
Los migrantes altitudinales se incorporan al ensamble de especies durante el invierno,
mostrando inclusive fidelidad al sitio donde pasan el invierno cada año. Como resultado del
marcado intensivo se recuperaron aves anilladas en temporadas invernales anteriores en las
localidades de Horco Molle y Piedra Tendida de tres especies migrantes altitudinales
(Mecocerculus leucophrys, Poospiza erythrophrys y Atlapetes citrinellus). Un zorzal
pecho-colorado (Turdus rufiventris) marcado en Horco Molle en primavera, se recuperó en
San Miguel de Tucumán durante el invierno, mostrando que esta especie que
considerábamos residente también realiza desplazamientos altitudinales, aunque es el único
dato con que contamos.
Las especies cuyos desplazamientos altitudinales fueron comprobados durante este estudio
se detallan a continuación: FAMILIA ANATIDAE
Merganetta armata berlepschi Hartert. Ha sido registrada en los ríos Cochuna, Pueblo
Viejo, Los Sosa y Choromoro (este último atraviesa la localidad de La Junta). No fue
registrada en las Sierras de Burruyacu ni en San Ignacio. Durante el invierno es una ave
frecuente en los ríos mencionados por debajo de los 800 m, mientras que en primavera y verano se la encuentra en niveles más altos, donde es común en el Bosque Montano. FAMILIA PSITTACIDAE
Aratinga mitrata mitrata (Tschudi). Cría en la Selva montana, tanto en el Aconquija, como
en las Sierras de Burruyacu. En Mayo desciende hacia el Bosque de Transición y el Chaco
Serrano, formando grandes bandadas (se observó una bandada de más de 100 individuos en
Piedra Tendida). Es una especie errática durante el invierno en los pedemontes,

51
alimentándose de las vainas del cebil. Fue encontrada en las tres localidades, aunque
números mayores se observaron en Burruyacu y La Junta.
Amazona tucumana (Cabanis). Cría en el Bosque Montano Superior en alisales. En
invierno desciende hacia los pedemontes. No se observaron números tan grandes como los
de la especie anterior. En Burruyacu la bandada más numerosa no sobrepasaba los 20
individuos. Se alimentan de cebiles y también tienen un comportamiento errático durante el
invierno. Es frecuente también en Dique San Ignacio.
Amazona aestiva xanthopterix (Berlepsch). Realiza desplazamientos entre el Chaco, donde
cría y el Bosque de Transición y Selva Montana, donde inverna. En otoño e invierno se
alimenta de cebiles y citrus cultivados en fincas cercanas. Utiliza el bosque para dormir y
realiza desplazamientos diarios a los lugares donde se alimentan. En invierno se observan
bandadas de 40 a 100 individuos, en época de cría desaparece de los pedemontes. FAMILIA TROCHILIDAE
Leucippus chinogaster hypoleucus (Gould). Es el picaflor más común durante el otoño-
invierno en el Bosque de Transición. Según Olrog (1979, 1984) cría en las zonas arbustivas
áridas en los cerros, sin embargo los datos de este trabajo demuestran que cría a niveles más
bajos, en la Selva y Bosque Montano. Se lo ha capturado durante todo el año en Transición,
aunque en mucho menor proporción en la época de cría.
Eriocnemis glaucopoides (D Orbignyi). Es común durante los meses de otoño e invierno,
aunque desaparece en el verano. Se lo ha capturado en áreas abiertas cultivadas durante el
invierno e inclusive en Chaco Serrano (Las Talas, Antillas, Ticucho).
Sappho sparganura sappho (Lesson). Cría en alisales a partir de Noviembre. En Mayo
aparece en el Bosque de Transición, habiéndoselo capturado hasta Octubre. Ha sido
observado en el Bañado de Figueroa, en el centro de la provincia de Santiago del Estero, en
Agosto. Es particularmente abundante en Dique San Ignacio entre Mayo y Octubre.
Microstilbon burmeisteri (Sclater). Cría en la Selva de Mirtáceas, durante el otoño e
invierno desciende hacia el Bosque de Transición y zonas abiertas cultivadas de la llanura
tucumana. Se lo observó en los pedemontes cultivados en Abril y Mayo donde frecuentaba
las flores de los árboles de Eucaliptus sp. y es constante en Dique San Ignacio en invierno. FAMILIA FURNARIIDAE
Cinclodes fuscus tucumanus Chapman. Habita en ambientes con pajonal cerca de ríos entre
los 1.000 y 3.000 m de altura, en las provincias de Tucumán, Catamarca y La Rioja (Olrog,
1974). En otoño desciende hacia niveles más bajos, donde es común desde el mes de Mayo
en las márgenes de los ríos a lo largo de los pedemontes, permaneciendo hasta Setiembre.
Se lo encuentra solitario o en parejas. Ha sido registrada para Horco Molle, La Junta, Piedra
Tendida, Dique San Ignacio y Cochuna durante el invierno. En verano ha sido encontrada
sólo en Dique San Ignacio.
Cinclodes atacamensis atacamensis (Philippi). Cría por arriba de los 2.000 metros de
altura en los cerros, preferentemente en lugares pedregosos y de pajonal cercanos a cursos
de agua. En Mayo desciende a los valles intermontanos como Tafí del Valle y quebradas de
montaña (Olrog, 1949). Ha sido capturado en el Río Choromoro, en La Junta en el mes de
Julio. No ha sido registrada en otras localidades pedemontanas. FAMILIA TYRANNIDAE Ochthoeca leucophrys tucumana Berlepsch. Es un visitante regular en los pedemontes
durante el invierno. A sido capturado en La Junta, Piedra Tendida y Horco Molle entre
Junio y Setiembre. Cría en los pastizales húmedos de altura, en quebradas arbustivas de
pajonal cerca de los cursos de agua (Olrog, 1978) entre Noviembre y Abril. Ha sido
registrado para Tafí del Valle entre Noviembre y Febrero.
Knipolegus cabanisi Schultz. Fue capturado y observado en Junio y Agosto en Piedra
Tendida, donde era común, y observado regularmente cerca de los ríos en Horco Molle
durante el invierno. Ha sido registrado también para Potrerillo, provincia de Salta, en
bosque pedemontano para el mes de Mayo y para Cochuna para el mes de Setiembre. No ha

52
sido encontrado en los pedemontes durante el verano. Ha sido registrada para El Indio,
Ruta 307 para el mes de Octubre. Es común en el Bosque Montano Superior cerca de los
ríos durante el verano. En La Banderita, Catamarca, los machos tenían protuberancias
cloacales a fines del mes de Noviembre.
Mecocerculus leucophrys leucophrys (D Orbignyi y Lafresnaye). Es un ave común en el
bosque de alisos y pino del cerro. Se encontraba criando a mediados de Octubre en
Cochuna en selva montana y a fines de Noviembre en La Banderita, Catamarca y en el Km
49, ruta 307, en bosques de alisos. Desciende a partir de Marzo a los pedemontes, donde es
constante durante el invierno, y donde participa en la formación de bandadas mixtas de
aves. Tiene este mismo comportamiento migratorio a lo largo de todas las yungas de
Argentina. Ha sido registrado en Piquirenda Viejo, Salta, en bosque pedemontano durante
el mes de Agosto; en Laguna La Brea, Jujuy, en Bosque de Transición de Palo Blanco
durante el mes de Julio; y en las tres localidades de Bosque de Transición estudiadas en la
provincia de Tucumán durante todo el otoño e invierno. Seguramente se interna algo en el
Chaco por los bosques de galería de los ríos, ya que fue observado en Tonono, Salta, en el
bosque que rodeaba al Río Itiyuro, durante el mes de Agosto. También es un visitante
regular en los bosques de Chaco Serrano. Fue observado en Las Tipas, Tucumán, y en
Antillas, Salta, en Agosto y Junio respectivamente. FAMILIA TURDIDAE
Turdus nigriceps nigriceps Cabanis. Es un ave común en las selvas y bosques montanos de
la provincia desde fines de Setiembre y durante todo el verano. Su comportamiento
migratorio todavía no está claro. Se capturaron jóvenes durante el mes de Mayo en bosque
pedemontano de San Ignacio donde, por su abundancia y comportamiento, estaban en
migración. Desaparece luego del Bosque de Transición hasta fines de Setiembre, comienzos
de Octubre (Fig. 29), donde fue observado su arribo en grupos compactos de 20-30
individuos a los pedemontes. Cantan estridentemente durante Octubre en Horco Molle y
otras localidades pedemontanas y van ascendiendo paulatinamente aprovechando los
parches con abundantes frutos de moras y zarzamoras. Los machos compiten con el canto
desde el arribo a los pedemontes hasta llegar al Bosque Montano Superior donde crían a
fines de Noviembre.
Fig. 29: Análisis acumulativo de especies que arriban en primavera al Bosque Pedemontano
de Piedra Tendida desde latitudes más al norte.

53
Probablemente esta especie inverne en el Chaco. Se la encontró en relativa abundancia en
el mes de Setiembre en el Bañado de Figueroa, Santiago del Estero. FAMILIA COEREBIDAE
Diglossa baritula sittoides (D Orbignyi y Lafresnaye). Es un visitante ocasional en los
pedemontes y Bosque de Transición durante el invierno. Ha sido observada en Piedra
Tendida y en Horco Molle en el mes de Junio. Cría en Noviembre en el bosque de aliso.
Fue encontrada abundante sólo en La Banderita, Catamarca, donde las hembras estaban
incubando a fines de Noviembre. FAMILIA THRAUPIDAE
Thraupis sayaca obscura Naumburg. Es una especie muy móvil durante el invierno en el
Bosque de Transición, sus desplazamientos en ésta época parecen erráticos y se producen
por lo general en bandadas uniespecíficas. A veces es superabundante en algunas parcelas
con frutos dentro del bosque, pero por lo general se desplaza por las copas de los árboles en
pequeños grupos. Seguramente realizan desplazamientos altitudinales diarios en busca de
parches con frutos, descendiendo a los campos cultivados con citrus y otros frutales y hacia
los bosques xerófilos del Este de la provincia.
Thraupis bonariensis schulzei Brodkorb. No es común en el bosque de Transición, prefiere
el Chaco y fincas con frutales. Su comportamiento migratorio también responde a un patrón
errático durante el invierno, pero sus desplazamientos no parecen ser tan marcadamente
altitudinales como la especie anterior sino más bien a lo largo de la llanura tucumana, donde
prefiere los bosques xerófilos. Se lo observó tanto en invierno como en verano en Piedra
Tendida y en La Junta.
Thlypopsis sordida sordida (D Orbignyi y Lafresnaye). Es un típico migrante altitudinal,
desplazándose por las copas de los árboles sin formar agrupaciones conspicuas. Nunca llega
a ser abundante como T. sayaca, pero su presencia es regular en los pedemontes y aún en el Chaco durante el invierno. Cría en primavera en la selva y en el bosque montano,
convirtiéndose en escasa durante el invierno.
Piranga flava flava Vieillot. Inverna en el bosque de Transición y en la selva montana. Se
la encuentra siempre en parejas solitarias y no muestran el tipo de desplazamiento erráticos
de T. sayaca. Durante la época de cría es más abundante en las zonas chaqueñas de la
provincia, por lo que pareciera preferir criar allí. En Horco Molle una pareja marcada
permaneció durante casi todo el invierno en la misma área. FAMILIA EMBERIZIDAE
Pheucticcus aureoventris (D Orbignyi y Lafresnaye). Es común en el Bosque de
Transición sólo durante el otoño e invierno, desapareciendo en Octubre. Cría a más altura,
por encima de los 1500 m de altura en los cerros, en matorrales y alisales durante los meses
de Noviembre y Diciembre. Llega a ser común en algunas localidades del bosque montano
superior como La Banderita, provincia de Catamarca. En sus desplazamientos invernales
llega al Chaco, donde grupos de jóvenes fueron observados en Tonono y Antillas, Salta, en
Agosto y Junio respectivamente. Durante este estudio fue registrado sólo durante el
invierno en La Junta y Piedra Tendida.
Atlapetes citrinellus (Cabanis). Cría preferentemente por arriba de los 1200 m de altura en
los cerros entre Noviembre y Marzo.Es particularmente abundante en el bosque de alisos
durante el verano.En invierno es constante en los bosques pedemontanos. En Horco Molle
es común entre Febrero y Setiembre, formando parte de bandadas mixtas de aves. En las
localidades estudiadas de Bosque de Transición su presencia es regular, aunque abundante
sólo en San Ignacio y su participación en bandadas mixtas en Burruyacu fue casi nula.
Coryphospingus cucullatus fargoi Brodcorb. Son frecuentes las bandaditas en las zonas de
borde del Bosque de Transición con el Chaco durante el invierno. Durante el verano
prácticamente desaparece. Es una especie típicamente chaqueña. Probablemente se mueva
entre el Chaco y el Bosque de Transición con comportamiento errático, parcialmente

54
migratorio, durante la época de escasez. Durante este estudio fue registrada en La Junta y
San Ignacio sólo en invierno y comienzos de primavera.
Poospiza erythrophrys. De las especies migrantes altitudinales es la de mayor significación
como visitante invernal por su abundancia, regularidad y participación en agrupaciones
interespecíficas durante el invierno en el Bosque de Transición. Es frecuente entre Febrero
y Setiembre en los pedemontes de la provincia de Tucumán. Cría en el Bosque Montano
Superior, donde las hembras incuban a partir de Noviembre. Es frecuente en las zonas de
Chaco Serrano de Tapia y Burruyacu, observándosela en bandaditas uniespecíficas.
FORMACIÓN DE BANDADAS MIXTAS DE AVES
Las especies que regularmente forman bandadas mixtas en los bosques pedemontanos del
noroeste de Argentina lo hacen sólo durante la época otoñal e invernal (Tabla 6). Son en su
mayoría propias de los estratos arbustivo y bajo (Fig. 31), semilleros arbusteros, arañeros y
pequeñas mosquetas y piojitos tiránidos insectívoros (Tabla 7). En la provincia de Tucumán,
tres de ellas son migrantes altitudinales desde el Bosque Montano Superior (M. leucophrys,
A. citrinellus y P. erythrophrys).
Fueron encontradas asociaciones interespecíficas en forma de bandadas en todos los pisos
altitudinales de las Yungas de la provincia de Tucumán durante el otoño e invierno. Existen
diferencias en la composición y frecuencia de especies presentes en bandadas mixtas entre la
selva montana y el bosque pedemontano.
Bandadas mixtas son distinguibles a partir de Mayo en la Selva Montana de la localidad de
Horco Molle. Las bandadas están cohesivamente formadas a partir de las 8.30 h durante el
invierno, disminuyendo en general sus actividades de forrajeo entre las 11.30 y 14.30 h, en
que sus miembros descansan en sus lugares preferidos. Al reagruparse, los grupos
continúan hasta que oscurece (19.30 h aproximadamente). En Horco Molle, Philydor rufosuperciliatus tiene un papel preponderante en dar el llamado que forma la bandada y
como emisor de voces de alarma que alertan al grupo. Myioborus brunniceps, conspicua por
coloración y comportamiento inquieto, también emite voces de alerta, entra y sale de las
bandadas y desarrolla gran actividad. Synallaxis superciliosa, Arremon flavirostris y
Chlorospingus ophthalmicus emiten permanentemente voces que podrían servir para
mantener unidos a los miembros aunque no tengan contacto visual entre ellos. Arremon
flavirostris y Basileuterus culicivorus son nucleantes pasivos (Moynihan, 1962) en esa
localidad, siendo seguidas más frecuentemente de lo que ellas siguen a otras especies, se
encuentran en porcentajes elevados en bandadas y generalmente una o dos están presentes
en cada agregación interespecífica, actuando como aglutinantes de las demás especies.Al
representar la Abundancia Relativa (AR) en función de la Frecuencia (Fe) para cada especie
(Fig. 30), destacan 4 de importancia alta (A. flavirostris y B. culicivorus, Myioborus
brunniceps y Poospiza erythrophrys), mientras que Philydor rufosuperciliatus, S.
superciliosa, y A. citrinellus, se agrupan alrededor de los nucleantes pasivos, siguiéndolas
más frecuentemente de lo que son seguidas y encontrándose en porcentajes mucho menores
que ellas en bandadas.

55
Fig. 30: Abundancia Relativa en función de la Frecuencia para cada especie que formaba
parte de bandadas mixtas en Bosque Montano Inferior de Horco Molle. Ver Tabla 7 para las
abreviaciones de los nombres de las especies.
En general los desplazamientos de los grupos son lentos (X = 10 m en 5 minutos) y sus
miembros se ubican, sobre todo las especies de mayor significación, en el estrato arbustivo y
en el suelo. El mayor número de especies se desenvuelve hasta los 4 m de altura (Fig. 31).
Fruteros y especies trepadoras (Picumnus cirratus y P. rufosuperciliatus) alcanzan el estrato
alto. De las dos nucleantes pasivas A. flavirostris es prácticamente terrícola y B. culicivorus
del estrato bajo. T. sordida, P. melanonota y Tharupis sayaca entran y salen de las
bandadas. P. cirratus y P. rufosuperciliatus realizan desplazamientos verticales por troncos.
Zorzales (T. rufiventris), pepiteros (S. caerulescens y S. aurantiirostris), urracas
(Cyanocorax chrysops), picaflores (salvo S. sparganura) y ratonas (Troglodytes solstitialis
y T. aedon), no participan en bandadas mixtas y son indiferentes al desplazamiento de los
grupos. Las especies que fueron observadas participando en bandadas mixtas representan el
50% del total de especies frecuentes durante el invierno en Horco Molle.
Las tácticas alimenticias empleadas por los miembros de las bandadas son por lo general la
recolección (Tabla 7), donde las aves toman un alimento estacionario del sustrato. Forman

56
Fig. 31: Utilización de estratos de vegetación de especies que participan en bandadas mixtas.
un grupo de especies exclusivamente insectívoras (S. superciliosa), frugívoras - insectívoras
(Ch. ophthalmicus, A. citrinellus) y semilleras - insectívoras (A. flavirostris, P.
erythrophrys). Capturas aéreas, a modo de revoloteo entre el follaje realizan tiránidos y
parúlidos. Algunas especies emplean una combinación de tácticas como salidas aéreas para
atrapar insectos con recolección de frutos e insectos (P. melanonota, T. sayaca).
Un promedio de 3.75 especies y 5.9 individuos por bandada resultan de 46 bandadas
observadas entre Mayo y Octubre en Horco Molle.A partir del mes de Octubre, en que la
mayor parte de las aves residentes comienzan a criar, se produce un cambio en la cohesión y
regularidad de las agrupaciones interespecíficas y durante todo el verano no se observan
bandadas mixtas significativas. Probablemente esto se deba a las exigencias de la
nidificación (Morse, 1970), pero también a la variación estacional del alimento disponible y
a los cambios en la composición de la avifauna, con el arribo de especies migratorias que
crían en estos bosques, y a la partida de migrantes altitudinales.
En los Bosques de Transición, 15 especies participaban de bandadas (Lista 6).Sobre 16
bandadas observadas en los censos de Junio y Agosto en Piedra Tendida, Burruyacu, al
graficar la AR/F, se observan 3 especies de importancia alta. S. superciliosa y A. flavirostris,
eran nucleantes pasivos alrededor de los cuales se agrupaban las demás especies, y P.
erythrophrys, migrante altitudinal, era el que mayor importancia tenía tanto en el número de
individuos por bandada como en la Frecuencia en que participaba (Fig. 32). M. brunniceps,
que en Horco Molle era un activo participante, tenía relativamente poca importancia,
apareciendo también como errático, con desplazamientos rápidos, sin formar parte por largo
tiempo de los grupos. En Burruyacu A. citrinellus era poco abundante, quizás a esto se deba
su pobre participación en la constitución de las bandadas. En el Bosque de Transición, al igual que en Horco Molle, las bandadas mixtas frecuentan el
estrato bajo, varias especies prácticamente se alimentan en el suelo (A. flavirostris, S.
superciliosa, P. rufosuperciliosa, Zonotrichia capensis). Las bandadas son más silenciosas
en Piedra Tendida, con menos vocalizaciones que en Horco Molle, quizás debido a que el
bosque es más abierto y hay mayor visibilidad. S. superciliosa es la que vocaliza más,
manteniendo la cohesión de los grupos, P. rufosuperciliatus es más silencioso que en Horco

57
Molle. Debido a la hojarasca seca, las bandadas son detectadas por el ruido que producen
las aves al alimentarse y avanzar.
Fig. 32: Abundancia Relativa en función de la Frecuencia para cada especie que formaba
parte de bandadas mixtas en Bosque Pedemontano Inferior de Piedra Tendida. Ver Tabla 7
para las abrevaciones de los nombres de las especies. ESPECIE BM TP PB
Picumnus cirratus (Pc) + + +
Sittasomus griseocapillus +
Synallaxis scutatus +
Synallaxis frontalis +
Synallaxis superciliosa (Ss) + +
Philydor rufosuperciliatus (Pr) + + +
Thamnophilus caerulescens +
Mecocerculus leucophrys (Ml) + + +
Phylloscartes ventralis (Pv) + + +
Todirostrum plumbeiceps +
Tolmomyias sulphurescens +
Vireo olivaceus + +
Parula pitiayumi (Pp) + + +
Myioborus brunniceps (Mb) + + +
Basileuterus bivittatus +
Basileuterus culicivorus (Bc) + +
Thraupis sayaca (Ty) + + +
Thlypopsis sordida (Ts) + +
Pipraeidea melamonota (Pm) + +
Chlorospingus ophthalmicus (Co) + +
Arremon flavirostris (Af) + + +
Atlapetes citrinellus (Ac) + +
Poospiza erythrophrys (Pe) + + +
Coryphospingus cucullatus + +
Zonotichia capensis (Zc) + +
Cyanocompsa cyanea + +
TABLA 7: Especies que participan en bandadas mixtas en el Bosque Pedemontano de
Transicion de Argentina. BM= Bosque Montano Inferior; TP= Bosque de Transición de tipa
y pacará; PB= Bosque de Transición de palo blanco (Piquirenda, Salta).

58
La alimentación es generalmente omnívora o insectívora entre los miembros de las
bandadas mixtas. Parúlidos y tiránidos emplean tanto el revoloteo entre el follaje o captura
aérea, como la recolección de presas fijas entre las hojas o ramas.
Las bandadas en Burruyacu estaban constituidas por 5 especies y 8.3 individuos promedio
por bandada. Las especies que participaban en la formación de bandadas mixtas en esta
localidad representaban el 37.5% del total de especies presentes.
En los Bosques Pedemontanos de Palo Blanco, 19 especies participaban de bandadas mixtas
de aves. Los integrantes más regulares en la formación de bandadas en Laguna La Brea
eran Synallaxis scutatus, Basileuterus bivittatus, Poospiza mleanoleuca, P.
rufosuperciliatus, A. flavirostris y Todirostrum plumbeiceps. En Piquirenda Viejo, S.
scutatus, B. bivittatus, P. rufosuperciliatus, Sittasomus griseocapillus, Thamnophillus
caerulescens y Atlapetes torquatus forrajeaban frecuentemente juntos, inclusive en el suelo
debido a lo decíduo del bosque.
ORGANIZACION TROFICA DE LA AVIFAUNA
La estructura trófica de las comunidades de aves del Bosque de Transición presenta un
patrón que responde a las condiciones de un bosque subtropical con una estación fría y seca
y una cálida y húmeda. Típicamente se produce un pico de productividad de frutos a fines
de primavera y comienzos del verano y un período de escasez durante la época seca
invernal. Durante este último período la estructura de la vegetación del bosque cambia, los
árboles, decíduos en su mayoría, pierden totalmente el follaje. Este patrón acerca al Bosque
de Transición más a las características de los bosques templados que a los tropicales y
determina la estructura trófica de las comunidades.
El grueso de las especies de aves está constituido por aves de tamaño pequeño, entre 3 y 15
gramos (40%) (Tabla 8). El número de especies va decreciendo progresivamente en las
categorías de más peso. En un análisis realizado sobre 61 especies frecuentes en estos
bosques, 35 son pequeñas (Fig. 33) y se alimentan exclusivamente de insectos que obtienen
mediante diferentes tácticas como recolección, revoloteo, captura aérea e inclusive una
taladradora. Otras 6 especies son recolectoras de néctar e insectos, una de ellas de sólo 3
gramos (Microstilbon burmeisteri), y 5 especies son recolectoras de insectos y semillas.
Fig. 33: Categorías de pesos de aves frecuentes en Bosques Pedemontanos.
A-Recolectores
Gran parte de las especies de aves del bosque de Transición son recolectoras. Es un gremio
taxonómicamente diverso, incluye zorzales, fruteros, tordos, una gallineta, una pava de

59
monte y un inambú. El modo de forrajear es el mismo, lo que varía es el sustrato, desde el
suelo hasta la copa de los árboles. Son especies generalistas que varían la dieta según la
oferta a lo largo del año. Se observa un aumento notable de la omnivoría durante el invierno
y de la frugivoría hacia fines de la primavera y el verano.
Recolectores frugívoros: El grado de frugivoría difiere significativamente entre las especies
de aves a lo largo del año. Esta se correlaciona con una contingencia estacional, con un pico
de fructificación claro en la primavera. Por este motivo se les considera en el análisis como
frugívoros desde Octubre hasta Enero y omnívoros el resto del año. No existen aves
dependientes de una sola especie de planta fructificante, son todas generalistas, con la
excepción de Euphonia chlorotica. Otras especies típicamente semilleras pero que
consumen frutos dentro del bosque son emberícidos como Saltator aurantiirostris,
Pheucticcus aureoventris y Cyanocompsa cyanea. El segundo grupo está formado por
miembros de la Familia Tyrannidae y considerados como atrapamoscas que realizan captura
aérea sobre el follaje. Son migratorios y permanecen sólo 4 o 5 meses en estos bosques.
Recolectores insectívoros: Comprende en general a aves de las Familias Furnariidae y
Troglodytidae. Pocas especies de furnáridos son comunes en el Bosque de Transición de
Tucumán, una de las cuales está especializada para trepar por troncos. Dos especies
cogenéricas de pijuíes son típicas del estrato arbustivo denso, Synallaxis superciliosa del
bosque maduro y S. frontalis de los ecotonos con el bosque chaqueño y áreas arbustivas.
Otro furnárido recolector, pero de los cursos de agua es Cinclodes fuscus, presente
únicamente en otoño e invierno.Dos especies de Troglodítidos, Troglodytes aedon y
Troglodytes solstitialis, son comunes en el estrato arbustivo bajo. La segunda muestra una
gran especialización ya que se alimenta en las hojas muertas suspendidas. Esta práctica es
más notable durante la época decídua. Es un ave típicamente selvática, mientras que la otra
es de amplia distribución. Recolectores nectarívoros: Hay 5 especies de picaflores constantes en el Bosque de
Transición, Chlorostilbon aureoventris, Leucippus chinogaster, Microstilbon burmeisteri,
Eriocnemis glaucopoides y Sappho sparganura, las tres primeras residentes y las dos
últimas migrantes altitudinales que están presentes sólo en los meses invernales
Recolectores granívoros: Varias especies de la Familia Emberizidae, típicamente granívora,
son en realidad omnívoros o semilleros-insectívoros durante gran parte del año. El análisis
de contenidos estomacales de A. citrinellus, dio como resultado la presencia de varios tipos
de coleópteros, semillas de P. carthagenensis y piedritas. A. flavirostris tiene una
alimentación típicamente insectívora-granívora. En sus estómagos se encontraron
himenópteros (hormigas y avispas), coleópteros, semillas, espigas y piedritas. Una
alimentación similar presenta Z. capensis . P. melanoleuca y C. cucullatus , frecuentes sólo
en las zonas alteradas, arbustales y tuscales buscan insectos entre la vegetación y se
alimentan también de pequeñas semillas. P. erythrophrys es también muy insectívora,
aunque también come frutos de P. carthagenensis) y semillas.
Observaciones sobre S. aurantiirostris dieron como resultado una dieta muy variada a lo
largo del año, semillas de Acacia aroma, frutos de Urera, P. carthagenensis, y brotes de
hojas de Justicia sp. Cyanocompsa cyanea preda, al igual que la especie anterior, sobre
semillas de A. aroma y se alimenta de una variedad de frutos como C. iguanea y A. edulis.
B. Atrapadores
Comprende un grupo variado de No Passeriformes, algunos terrícolas, otros arborícolas y
unos cuantos de los cursos de agua. En cada localidad muestreada siempre estuvieron
presentes de uno a tres atrapadores. Las especies de rapaces que emplean este modo de
alimentación son:
-Buteo magnirostris. Es el aguilucho más común en las zonas boscosas del norte de
Argentina. Diurno, se alimenta principalmente de insectos grandes, aunque también de aves
y mamíferos pequeños como ratones. En ocasiones mata las aves que quedan atrapadas en

60
las redes de muestreo. Permanece quieto en una rama o poste por horas, cayendo sobre las
presas para comerlas en el suelo.
-Otus choliba. Es la lechucita más común en los Bosques pedemontanos. Nocturna, se
alimenta principalmente de pequeños mamíferos, roedores y también marmosas. Vuela
desde los árboles para caer sobre el suelo o una rama sobre la presa, sujetándola con las
garras. Estuvo presente en cada una de las localidades muestreadas, sólo que no aparece en
los censos debido a sus hábitos. Los datos obtenidos fueron por captura y al escuchar sus
sonidos u observarla durante la noche.
-Aegolius harrisii. Tiene un comportamiento similar a la anterior. Este estudio ha revelado
su presencia como mucho más común de lo que se pensaba (Olrog, 1985). Ha sido
capturada en Piedra Tendida, Piedras Coloradas (al lado del Río Los Sosa a 750 m de altura)
y Horco Molle. Estas tres localidades son nuevas para la especie en Tucumán. Parece ser
un habitante regular de los Bosques pedemontanos, al menos durante el invierno ya que no
ha sido capturada en verano. Es nocturna y se alimenta de aves y mamíferos.
Otras aves no rapaces que emplean esta táctica de forrajeo son las garzas, martín pescadores
y cucos. En Tucumán, se encuentran presentes dos garzas en los riachos del bosque,
Nycticorax nycticorax, nocturna y Tigrisoma fasciatum, diurna. La primera era común en
San Ignacio y la segunda fue observada en las localidades de Piedra Tendida, Piedras
Coloradas y La Florida
Las tres especies de martín pescador se encuentran en los ríos de los pedemontes. Por lo
general usan una percha sobre el río, una rama o piedra para lanzarse al agua y levantar la
presa con el pico.
Varias especies de la Familia Cuculidae se encuentran en los bosques de norte de Argentina,
pero sólo tres son comunes en el Bosque de Transición de la provincia de Tucumán, Guira
guira, ave social presente exclusivamente junto a cultivos y zonas abiertas arboladas, Tapera naevia, ave solitaria de las zonas alteradas (tuscales) o más xerófilas y Piaya
cayana, también solitaria que prefiere las zonas arbustivas densas. Esta última ha sido
encontrada sólo en Burruyacu como frecuente. Son especies atrapadoras de roedores y
pequeños reptiles, aunque también recolectoras de insectos y huevos de aves.
C. Cazadores por revoloteo
Es una práctica común entre los miembros de la Familia Parulidae y de un grupo de especies
de la Familia Tyrannidae. En todos los casos son aves que prefieren la vegetación densa, de
pequeño tamaño y movimientos rápidos y /o acrobáticos. Se alimentan de insectos
voladores y artrópodos blandos como arañas, cochinillas, larvas y orugas. Los obtienen
revisando cuidadosamente el haz y envés de hojas y tallo. Pero típicamente realizan
movimientos rápidos con las alas volando de una rama para volver rápidamente, estas
prácticas espantan insectos que se encuentran posados lo que les permiten ubicarlos. Todas
las especies están relacionadas con hábitats selváticos, son pequeñas (menores a 15 gramos)
y predominan las coloraciones verdosas o amarillentas. Son propias del estrato bajo y
medio denso. Algunas se encuentran siempre cerca de los cursos de agua (Geothlypis
aequinoctialis y Serpophaga nigricans). Las más comunes en los bosques de Transición de
Tucumán son: Myiophobus fasciatus, Phylloscartes ventralis, Todirostrum
margaritaceiventer, Mecocerculus leucophrys, Serpophaga subcristata, S. munda, Parula
pityayumi, Myioborus brunniceps y Basileuterus culicivorus. Esta última especie es
reemplazada desde Metán hacia el norte por B. bivitatus. En Burruyacú ha sido encontrada
otra especie, ya mencionada, Todirostrum plumbeiceps, nueva para la provincia.
D. Cazadores por captura aérea
Es un método utilizado principalmente por miembros de la Familia Tyrannidae, vulgarmente
conocidos como atrapamoscas, aunque también ocasionalmente la realizan otros grupos de
aves como fruteros (Familia Thraupidae). Necesita de espacio para poder realizarse por lo
que lo utilizan aves propias del estrato superior o de las copas de los árboles. Las aves más

61
comunes, todas ellas migratorias, que emplean este método son: Ochthoeca leucophrys,
Tyrannus melancholicus, Empidonomus varius, Myiodinastes maculatus, Pitangus
sulphuratus, Myiarchus swainsoni y M. tyrannulus. Este grupo es de distribución chaqueña
o de áreas abiertas, se alimentan de este modo típicamente tiránido, con salidas aéreas, pero
también algunas especies consumen frutos (T. melancholicus, M. maculatus). Lo hacen
preferentemente en las copas de los árboles o en claros del bosque. Otro grupo penetra
únicamente por los ríos, también migratorios, aunque exclusivamente insectívoros. Realizan
salidas aéreas sobre el agua, no sobre el follaje. Son éstos: Sayornis nigricans, Knipolegus
cabanisi, K. striaticeps y K. aterrimus. Hirundinea ferruginea, se encuentra exclusivamente
en profundas quebradas y a gran altura sobre el río. Estaba presente sólo en San Ignacio.
E. Caza al vuelo rapaz
Entran dentro de esta categoría un halcón de bosque (Micrastur ruficollis) y golondrinas. La
primera es una especie común en los bosques húmedos de Tucumán. En los bosques del
norte de Salta es común otra especie similar pero de mayor tamaño (M. semitorquatus). Se
alimentan exclusivamente de aves que persiguen entre el follaje y hacen caer al suelo para
comerlas. M. ruficollis ha sido observado en varias oportunidades alimentándose de
zorzales y palomas, sus presas más comunes, y también atacó aves que estaban atrapadas en
las redes. De esta manera ha podido ser capturado y marcado. M. semitorquatus fue
observado dando caza a pavas del monte (P. obscura) en Piquirenda Viejo.
Sólo dos especies de golondrinas aparecen en el estudio como comunes. La golondrina
negra (Progne modesta), en los ecotonos con zonas abiertas y próximos a casas
abandonadas (San Ignacio); y la barranquera azul, Notiochelidon cyanoleuca, muy común
sobre las selvas del noroeste argentino.
F.- Buscadores
Son especies muy especializadas que aprovechan la exfoliación de la corteza y cavidades ya existentes en los árboles para buscar su alimento. El pico de estas especies es relativamente
blando, por lo que están imposibilitadas de taladrar la madera. Su alimento está compuesto
por larvas de coleópteros que viven en la corteza y artrópodos que se encuentran posados o
en cavidades, generalmente no voladores. Típicamente estas aves ascienden por los troncos
y ramas como lo hacen los carpinteros y son de coloración pardo rojiza. Dos especies de la
Familia Dendrocolaptidae son comunes en los bosques pedemontanos de Tucumán, una de
gran tamaño, Xiphocolaptes major y una pequeña, Syttasomus griseocapillus. Una sola
especie de la Familia Furnariidae, Philydor rufosuperciliatus, tiene este comportamiento y
es la más común en cada localidad estudiada. En Salta y Jujuy, se incorpora
Dendrocolaptes picumnus, que está ausente de Tucumán. El número de especies buscadoras
o trepadoras es mayor en los bosques xerófilos más al este, donde 4 o 5 especies están
presentes en cada localidad.
G. Taladradores
A diferencia de las anteriores, tienen un pico fuerte que les permite hacer agujeros en la
corteza. Pertenecen a la Familia Picidae, con 6 especies representadas en los Bosques
Pedemontanos del noroeste de Argentina. En Tucumán se encontraron sólo tres especies,
Picumnus cirratus, de 9 gramos que busca alimento principalmente en las ramas delgadas,
es la más común y residente en los pedemontes; Piculus rubiginosus, de tamaño mediano,
sólo en bosques maduros ya que prefiere los hábitats de Selva Montana como Horco Molle y
Piedras Coloradas; y Campephilus leucopogon, de gran tamaño, taladradora de troncos.
Veniliornis frontalis, típico de la Selva Montana, no ha sido encontrada durante el estudio en
Bosque de Transición en Tucumán. Estos carpinteros se alimentan de larvas de coleópteros,
hormigas y termitas. Picumnus dorbignyanus era común sólo en Piquirenda Viejo.

62
RELACIÓN ENTRE LAS TÁCTICAS ALIMENTICIAS Y LA BIOMASA
En Piedra Tendida la biomasa por hectárea durante el invierno variaba entre 800 y 1.500 gr.
La mayor parte estaba constituida por especies omnívoras medianas y grandes, como
zorzales, urracas y pavas de monte, cuyos pesos variaban entre 60 y 1.200 gr. (Tabla 8).
Los recolectores de insectos o de insectos y semillas constituían otra categoría trófica
importante por su biomasa durante los meses de invierno. Pero, a diferencia de los
omnívoros, comprenden a pequeñas aves de entre 5 y 30 gr. como gran cantores y chingolos
(Fig. 33). ESPECIE PESO RANGO
Merganetta armata 485.00 450 - 500
Micrastur ruficollis 270.00
Buteo magnirostris 389.00 380 - 400
Otus choliba 138.00 120 - 174
Leptoptila verreauxi 220.00 210 - 230
Chlorostilbon aureoventris 4.25 4 - 4.50
Leucipus chinogaster 4.87 4.50 - 5
Eriocnemis glaucopoides 3.25 3 - 3.5
Sappho sparganura 6.16 5 - 7
Microstilbon burmeisteri 3.00
Ceryle torquata 308.50 307 - 310
Chloroceryle americana 32.00 31 - 33
Picumnus cirratus 9.40 9 - 10
Veniliornis frontalis 35.00
Sittasomus griseocapillus 11.30 11 - 12
Cinclodes fuscus 27.30 26 -28
Cinclodes atacamentsis 51.00
Certhiaxis pyrrhophya 13.00
Synallaxis superciliosa 12.00 11 - 14
Philydor rufosuperciliatus 26.50 22 - 30
Thamnophillus caerulescens 21.33 18 - 23
Thamnophillus ruficapillus 20.00
Ochthoeca leucophrys 24.00
Knipolegus striaticeps 12.00
Knipolegus aterrimus 20.00
Knipolegus cabanisi 19.25 18 - 21
Myiarchus tyrannulus 31.50 31 - 30
Myiarchus swainsoni 25.00
Empidonax euleri 11.00
Myiophobus fasciatus 11.50 10 - 11
Phylloscartes ventralis 8.50 8 - 10
Hemitriccus margaritaceiventer 7.80 7 - 9
Mecocerculus lecucophrys 10.25 9 - 12
Elaenia albiceps 15.16 14 - 16
Elaenia parvirostris 13.50 13 - 14
Elaenia strepera 20.75 20 - 22
Elaenia obscura 26.28 25 - 28
Serpophaga subcristata 6.00
Serpophaga munda 5 50
Serpophaga nigricans 8.83 8.5 - 9
Phytotoma rutila 35.00
Troglodytes aedon 10.00
Troglodytes solstitialis 10.00
Motiochelidon cyanoleuca 13.20 12 - 14
Cyanocorax chrysops 150.00 125 - 181
Catharus ustulatus 29.43 25.5 - 32
Turdus nigriceps 50.44 41 - 59
Turdus rufiventris 64.50 59 - 73

63
Turdus amaurochalinus 61.60 59 - 66
Cychlaris gujanensis 30.00
Vireo olivaceus 14.91 14 - 15.5
Parula pitiayumi 6.33 6 - 7
Geothlypis aequinoctialis 9.00
Myioborus brunniceps 8.77 8 - 10
Basileuterus culicivorus 8.83 8 - 9
Basileuterus bivittatus 14.00 12 - 16
Diglossa baritula 10.50 10 - 11
Pipraeidea melanonota 23.00 22 - 27
Thraupis sayaca 31.60 31 - 33
Thlypopsis sordida 14.30 12 - 16
Chlorospingus ophthalmicus 16.42 15 - 17
Saltator aurantiirostris 43.70 36.5 - 48
Pheucticus aureoventris 46.50 46 - 47
Cyanocompsa cyanea 27.50 27 - 28
Sporophila caerulescens 11.00
Coryphospingus cucullatus 13.00
Atlapetes citrinellus 28.10 25 - 31
Atlapetes torquatus 35.00
Arremon flavirostris 25.50 24 - 28
Zonotrichia capensis 19.40 18 - 21
Poospiza melanoleuca 12.00
Poospiza erythrophrys 14.80 14 - 16
Carduelis magellanica 11.00
Caccicus chrysopterus 36.25 30 - 41.5
TABLA 8: Pesos de aves del Bosque Pedemontano de Transición.
Las especies recolectoras de frutos y semillas como pepiteros y reina moras, por lo general
de tamaño pequeño, entre 25 y 45 gr., tenían incidencia en el aumento de la biomasa sólo en
algunos casos, por la presencia de loros que se alimentaban de cebiles con frutos en ciertas
parcelas. La biomasa por hectárea en censos realizados en Octubre en la misma localidad era
levemente mayor, entre 1.200 y 1.800 gr. Los recolectores de frutos como tiránidos,
tráupidos y túrdidos, constituían la mayor parte de la biomasa, junto a los recolectores de
frutos y semillas como loros y pepiteros. Entre los insectívoros, los que realizaban captura
aérea sobre el follaje, en general tiránidos, eran más significativos que los miembros de
otros gremios como buscadores, recolectores de insectos y captura por revoloteo. Estas
especies insectívoras son en todos los casos pequeñas.
Los valores de biomasa por hectárea de la localidad de San Ignacio son similares, entre 800
y 1.600 gr., sin diferencias significativas entre invierno y verano. Los valores de la Reserva
La Florida de Agosto, son mayores, entre 2000 y 2500 gr. por ha, similares a los del bosque
de palo blanco. En La Florida, la presencia de loros en los censos (Pionus maximiliani y
Amazona aestiva) y la gran altura del bosque (más de 20 m), eran determinantes.
DIVERSIDAD DE MACRONICHOS
Las diferencias entre los Índices de Diversidad de Macronichos (Indice de Diversidad de
Shannon-Wiener) no son significativos entre las distintas localidades con Bosque
Pedemontano de Transición estudiadas (Tabla 9). Son similares para Piquirenda Viejo, San
Ignacio y Piedra Tendida, entre 0.80 y 1 nats inf/macronichos (N=12; X= 0.93). No existen
diferencias significativas entre los meses de invierno y primavera en una misma localidad.
Durante la primavera aumentan notablemente los recolectores frugívoros y los recolectores
omnívoros durante el otoño e invierno (Tablas 10, 11, 12 y 13). Los valores de diversidad
aumentan cuando hay una distribución más equitativa en el número de especies en cada
categoría y cuanto mayor es el número de categorías alimenticias ocupadas. La Junta

64
presenta valores levemente menores (entre 0.79 y 0.84 nats inf/macronichos) debido a una
menor riqueza de especies.Los resultados de este estudio indican que la estructura trófica de
estas comunidades de aves varía poco de sitio a sitio respecto a la diversidad de categorías LOCALIDAD MES NATS INF /
MACRONICHOS
NUMERO
SP DE AVES
Piquirenda Viejo Agosto 0.95 54
Piquirenda Viejo Noviembre 0.93 45
San Ignacio Mayo 1.08 37
San Ignacio Octubre 0.98 38
San Ignacio Diciembre 0.94 34
La Junta Julio 0.81 32
La Junta Setiembre 0.84 34
La Junta Octubre 0.80 37
Piedra Tendida Junio 0.95 35
Piedra Tendida Agosto 0.99 36
Piedra Tendida Octubre 1 33
Piedra Tendida, Bosque con
nogalar
Junio 0.92 46
TABLA 9: diversidad de macronichos en aves de bosques pedemontanos
Indice de diversidad H’ = -pi) (log 2 pi)
CATEGORIA
MAYO
OCTUBRE DICIEMBRE
RECOLECTOR
RO omnívoro 6 2 2
RI insectos 3 3 3
RS semillas 1 1
RIS insectos y semillas 4 3 4
RFS frutos y semillas 2 4 3
F frugívoro 3 8
RNI néctar e insectos 4 2 1
ATRAPADOR
AC carnívoro 1 2
AM mamíferos 2
AP peces 3 1
CAPTURA AEREA
A sobre el agua 1 1 1
CAF sobre el follaje 2 3 3
CAZA AL VUELO RAPAZ
0CVR aves
REVOLOTEO
R insectos 2 5 2
BUSCADOR
BC en corteza 2 1
BS en el suelo
TALADRADOR
T en ramas 1 1
TT en troncos 1 1
PLANEADOR
Pca carnívoro 1
PC carroña 1 3
TABLA 10: Categorías tróficas de aves de San Ignacio, Tucumán.
tróficas, aunque cambien las categorías según la estación del año debido a la disponibilidad
estacional de alimentos. Típicamente, las especies que permanecen todo el año sin migrar
varían su dieta a lo largo del año según la disponibilidad y abundancia estacional de
diferentes tipos de alimentos. Predominan las especies omnívoras, existen pocas especies

65
residentes especialistas, en todos los casos son insectívoras, y las frugívoras son
migratorias. Los omnívoros son dominantes inclusive en las localidades de Salta y Jujuy
durante el
CATEGORIA
AGOSTO
JUNIO
JUNIO
nogalar
OCTUBRE
RECOLECTOR
RO omnívoro 7 8 11 4
RI insectos 2 2 2 2
RS semillas 1 2 1
RIS insectos y
semillas
3 4 4 3
RFS frutos y
semillas
4 4 5 3
F frugívoro 7
RNI néctar e
insectos
4 2 2 2
ATRAPADOR
AC carnívoro 4 2 2 1
AM mamíferos 1
AP peces 1
CAPTURA AEREA
CAA sobre el agua 1 1 1 2
CAF sobre el
follaje
1 2 4
CAZA AL VUELO RAPAZ
CVR aves 1
REVOLOTEO
R insectos 5 5 6
BUSCADOR
BC en corteza 1 1 1
BS en el suelo 1 1 1
TALADRADOR
T en ramas 1 1 1
TT en troncos 1 2
PLANEADOR
PC carroña 4 1 1 1
TABLA 11: Categorías tróficas de aves de Piedra Tendida, Burruyacu, Tucuman.
CATEGORÍAS 0 I S IS RFS RF RNI C A Ca
Recolector 10 4 3 6 5 1
Atrapador 3
Caza al vuelo rapaz 1
Cap. aérea s/ follaje 1
Revoloteo 5
Buscador 2
Taladrador 1
Planeador 2
TABLA 12: Categorías tróficas de aves de Metán, Salta.
invierno, mientras que las especies semilleras o insectívoras semilleras son escasas y su
abundancia se mantiene constante.

66
Categorías 0 I S IS RFS RF RNI C A Ca
Recolector 4 2 2 2 3 3
Atrapador 3
Caza al vuelo rapaz 2
Cap. aérea s/ follaje 6
Cap. aérea s/agua 1
Revoloteo 3
Taladrador 1
Buscador 3
TABLA 13: Categorías tróficas de aves de Piquirenda Viejo, Salta.
REPRODUCCION
La mayor parte de las especies de Passeriformes del Bosque Pedemontano de la provincia de
Tucumán se reproducen entre Octubre y Noviembre (Tabla 14), comenzando en Setiembre
el establecimiento de los territorios de cría. Pocas especies criaron dos veces durante este
estudio en la misma temporada (Columbina picui, T. aedon, Z. capensis, S. caerulescens) y
sólo se encontró criando a Molothrus badius (con pichones en el nido) en Marzo y Abril.
Algunas especies, particularmente las migratorias, utilizaron sólo Noviembre y Diciembre
para criar (T. melancholicus, E. varius, M. tyrannulus, M. fasciatus, E. strepera, E.
parvirostris, S. munda, V. olivaceus, P. pitiayumi). El Tamaño de los territorios de invierno
y verano se modifica, achicándose notablemente en verano para algunas especies de
Passeriformes. El tamaño de territorio de A. flavirostris, estudiado en Piedra Tendida varió
de 1 Ha en Junio, donde se desenvolvía mayormente dentro de una bandada mixta, a
aproximadamente 600 m en el mes de Octubre (Fig. 34).
Fig. 34: Territorio de cría y territorio de invierno de Arremon flavirostris en Piedra Tendida.

67
Este espacio es pequeño dado el comportamiento terrícola de esta especie. El territorio de
cría de B. culicivorus no superaba 10 m2 durante el mes de Octubre en San Ignacio, aunque
frecuentando dos estratos e infinidad de epífitas. Unas 10 Ha eran forrajeadas por Especie E F M A M J J A S O N D Observaciones
Merganetta armata Bosque Montano
superior Tigrisoma fasciatum
Leucippus chinogaster
Ceryle torquata
Chloroceryle americana
Veniliornis frontalis Bosque Montano
superior Synallaxis superciliosa
Cinclodes fuscus Bosque Montano
superior Philydor rufosuperciliatus
Sayornis nigricans
Tyrannus melancholicus
Empidonomus varius
Empidonomus
aurantioatrocristatus
Chaco
Myiodynastes maculatus Migratoria
Pitangus sulphuratus Migratoria
Myiarchus tyrannulus Migratoria
Myiophobus fasciatus Migratoria
Mecocerculus leucophrys Bosque Montano
superior Phylloscartes ventralis Migratoria
Elaenia albiceps Migratoria
Elaenia strepera Migratoria
Elaenia parvirostris
Elaenia obscura
Serpophaga munda
Troglodytes aedon
Notiochelidon cyanoleuca
Cyanocorax chrysops
Turdus nigriceps
Turdus rufiventris
Vireo olivaceus
Parula pitiayumi
Geothlypis aequinoctial
Myioborus brunniceps
Basileuterus culicivorus Muda en cabeza
Pipraeidea melanonota
Piranga flava
Thraupis sayaca
Thlypopsis sordida
Chlorospingus ophtalmicus
Saltator caerulescens
Saltator aurantiirostris
Pheucticus aureoventris Bosque Montano
superior Cyanocompsa cyanea Muda en cola
Sporophila caerulescens
Coryphospingus cucullatus Chaco
Atlapetes citrinellus Bosque Montano
superior Arremon flavirostris
Zonotrichia capensis
Poospiza melanoleuca Chaco
Poospiza erythrophrys Bosque Montano
superior Carduelis magellanica
Molothrus bonariensis
Molothrus badius
Juvenil
Muda
Epoca de
cría
TABLA 14: Epoca de reproducción y muda en aves frecuentes en Bosque de Transición

68
Cyanocorax chrysops en grupos de 7 a 11 individuos durante el otoño en Burruyacu. Estos
territorios de cría eran mantenidos mediante cantos (T. solstitialis, T. aedon, T. rufiventris,
V. olivaceus, C. gujanensis, M. brunniceps, B. culicivorus, G. aequinoctilais, C. cyanea, S.
aurantiirostris, A. flavirostris, Z. capensis. T. sayaca, P. melanonota, P. flava, T. sordida,
M. bonariensis, M. badius) o voces territoriales (P. sulphuratus, M. maculatus, T.
melancholicus, M. tyrannulus) en los Passeriformes. Las manifestaciones agresivas
(agonísticas) eran frecuentes en varias especies, presentándose al final de la estación de cría
el plumaje de la cola típicamente gastado o casi inexistente en varias de ellas (P.
rufosuperciliatus, T. rufiventris, T. sayaca, A. flavirostris, S. aurantiirostris). Junto al canto
comienzan en Setiembre a desarrollarse protuberancias cloacales en los machos, mientras
que las placas incubatrices de las hembras aparecen en Octubre y están plenamente
desarrolladas en Noviembre.
Fueron encontrados nidos preferentemente en el estrato bajo, de Myophobus fasciatus,
Elaenia strepera, E. parvirostris, Serpophaga munda y Vireo olivaceus en arbustos densos,
de Empidonax euleri, Pipraeidea melanonota y V. olivaceus a un metro del suelo, de
Sporophila caerulescens y Zonotrichia capensis en gramíneas, y de Crypturellus tataupa y
Arremon flavirostris en el suelo. Pitangus sulphuratus, Myiodynastes maculatus, Myiarchus
swainsoni y Cyanocorax chrysosps tenían nidos construidos a gran altura, mientras que
Turdus amaurochalinus, Turdus rufiventris y Basileuterus culicivorus tenían sus nidos en el
estrato medio. Un nido de éste último se encontraba entre musgos colgantes a 5 m del suelo
en San Ignacio. Otus choliba, Aratinga mitrata, Pionus maximiliani, Campephilus
leucopogon, Picumnus cirratus, Xiphocolaptes major, Sittasomus griseocapillus, Philydor
rufosuperciliatus y Molothrus badius fueron encontrados anidando en troncos. Otus choliba se encontraba nidificando en un hueco a 2 m de altura en Las Tipas, Sierra de San Javier, a 4
m del suelo en Piedra Tendida y a 8 m en Horco Molle. Dos nidos de A. mitrata se
encontraron a más de 10 m de altura en San Ignacio y Cochuna. P. maximiliani tenía su
nido a 10 m de altura en Piedra Tendida. Un nido de P. rufosuperciliatus en un tronco
caído tenía dos jóvenes en Noviembre en Horco Molle. M. badius fue encontrado
nidificando en Marzo en la misma localidad, en un hueco a 10 m de altura (probablemente
un nido abandonado de otra especie).
Jóvenes voladores de casi todas las especies de passeriformes se capturaron por lo general
en Enero y Febrero (Tabla 14), entre las que crían en el Bosque Pedemontano, aunque
jóvenes de varias especies que nidifican en el Bosque Montano Superior son abundantes en
Mayo (T. nigriceps, S. aurantirostris, Pheucticus aureoventris, P. flava).
He obtenido relativamente pocos datos de No Passeriformes, debido a la poca posibilidad de
capturarlos. He capturado jóvenes buenos voladores de Merganeta armata entre Mayo y
Agosto (La Junta y Piedras Coloradas), y por datos obtenidos en el Bosque Montano
Superior, crían entre Diciembre y Enero. Jóvenes de Tigrisoma fasciatum fueron
observados en Junio y Agosto en Piedra Tendida, un subadulto fue observado en Agosto en
La Florida. Un joven de Sarcoramphus. papa fue observado en Junio y Agosto en la misma
localidad. Buteo magnirostris realiza manifestaciones de cortejo en Octubre y se observaron
jóvenes entre Abril y Junio. Otus choliba fue encontrada nidificando en Marzo y con
jóvenes en el nido en Mayo y Junio en Horco Molle. Los picaflores A. chionogaster y C.
aureoventris estaban incubando en Octubre en Piedra Tendida. Leptoptila verreauxi
nidifica entre Octubre y Enero, mientras que Columbina picui, extiende su temporada de
cría hasta Abril. Los estridentes llamados para reproducción de Aramides cajanea se
escuchan en Noviembre y Diciembre y se observaron adultos con jóvenes en Febrero y
Marzo en Horco Molle.

69
MUDA
La presencia de muda masiva en cuerpo, remeras, timoneras y cobertoras ocurre en Marzo y
menos intensamente en Abril, tanto en los No Passeriformes como en los Passeriformes
estudiados (Tabla 14). Corresponde ésta a la muda postnupcial, y en el caso de las especies
migratorias ya mencionadas anteriormente no fue posible de observar ya que abandonan sus
territorios de cría para migrar. En Passeriformes esta muda es paulatina, mudándose una a
una las plumas del vuelo y parte del plumaje de contorno del cuerpo. En vireónidos,
parúlidos y traúpidos (V. olivaceus, P. pitiayumi, G. aequinoctialis, M. brunniceps, B.
culicivorus, P. melanonota, P. flava, T. sayaca, T. sordida, C. ophthalmicus), estuvo
perfectamente sincronizada, se realiza en el mes de Marzo, presentando el plumaje cañones
de plumas nuevas prácticamente en todo el cuerpo. Los furnáridos P. rufosuperciliatus, S.
superciliosa y C. fuscus mudan paulatinamente entre Febrero y Abril. Los tiránidos S.
munda y E. obscura mudan su plumaje en Marzo, mientras que P. ventralis muda en el mes
de Abril.
La muda prenupcial no se presenta en las aves del Bosque Pedemontano como una muda
masiva sino de ciertas partes del cuerpo, alas o cola. Ocurre en Agosto y Setiembre y fue
registrada en pocas especies: C. cyanea y P. aureoventris en cola, M. brunniceps en cabeza
y corona, P. melanonota en plumas de contorno. Los jóvenes de la mayor parte de los
Passeriformes también presentaron un recambio completo del plumón por plumas
definitivas en el mes de Marzo.
DISCUSION
Los Bosques Pedemontanos de Transición poseen características típicas o propias respecto a
la avifauna y no deben ser considerados como una simple mezcla o ecotono entre las
taxocenosis chaqueñas y yungueñas. Existen algunos elementos importantes para ser tratado como un bosque con una identidad ornitológica propia:
a. Existe una constancia en la presencia de un grupo importante de especies yungueñas, que
preferentemente nidifican en ellos y no en las Selvas Montanas (Pionus maximiliani,
Elaenia strepera, Elaenia parvirostris, Phacellodomus rufifrons, Batara cinerea,
Thamnophillus ruficapillus, Vireo olivaceus, Cyanocompsa cyanea, Icterus cayanensis).
b. Existe un ensamble de especies diagnóstico por su constancia y abundancia relativa, con
especies dominantes típicas, que son distintas a las dominantes en el Chaco y en los bosques
húmedos lindantes (Tabla 5). Las 83 especies residentes tambien pueden ser utilizadas
como un caracter definitorio (lista6).
c. Reciben en forma sincrónica migrantes que llegan a nidificar o a invernar dentro de una
llamativa contingencia de patrones estacionales. Es improbable que esto ocurra sino por un
largo tiempo evolutivo de adecuación. Muchas de estas aves tienen largas rutas migratorias,
destacándose dos rutas principales por la cantidad de especies involucradas.
Presenta una dinámica muy intensa de especies migratorias que involucra a más de 100
especies, si descartamos las residentes constantes (83). Dentro de los migrantes latitudinales
(31% del total de especies), pueden distinguirse tres grupos de especies según el área donde
crían y el uso que hacen del Bosque de Transición:
1. Migrantes Neotropicales que crían en el Sur de Argentina e invernan en las provincias
norteñas y países limítrofes.
2. Migrantes Neotropicales que crían en Transición y migran luego hacia el Norte, la
Amazonia o las Yungas de Perú, Bolivia y Ecuador.
3. Migrantes Neárticos desde el Hemisferio Norte que invernan en Argentina.
El Pantanal de Matto Grosso es la principal área de recepción de migrantes que crían en los
Bosques Pedemontanos de Transición del noroeste argentino y atraviesan el Chaco en
migración, mientras que el resto de las especies migran a lo largo de las Yungas por Bolivia,
Perú, Ecuador, Colombia y Venezuela.

70
Estos bosques no sufren el impacto de especies migratorias neárticas, como ocurre en el
norte y centro de Sudamérica donde los migrantes, por su cantidad, compiten por recursos
con las especies residentes. La presencia estacional del zorzal norteamericano, única especie
significativa por su abundancia, cuando los recursos de frutos están en su nivel máximo
(Noviembre - Enero), es de poca consecuencia como factor que pudiese afectar la
disponibilidad de frutos para las especies residentes o nidificantes en el Bosque de
Transición.
d. Es un bosque altamente decíduo, más del 50% de las especies arbóreas (J. australis, A.
colubrina, T. tipu, E. contortisiliquum, P. nigrescens, J. mimosifolia, T. avellanedae, entre
otras) pierden total o parcialmente el follaje, y a pesar de tener temperaturas medias
anuales relativamente altas (entre 16 y 18 ºC), se comporta respecto a la avifauna, como un
bosque templado ya que una tercera parte de su avifauna migra hacia el norte luego de criar.
e. La coexistencia de especies en la época de estrés se ve favorecida por la formación de
bandadas mixtas de aves, con una similaridad en el número e identidad de las especies que
la componen a lo largo del rango de extensión de estos bosques en el norte de Argentina.
Esto, junto al hecho de estar compuestas por aves residentes y migrantes que manifiestan
fidelidad al sitio donde invernan, es otro factor que habla del largo tiempo de adecuación
que necesariamente debe haber existido para que se produjeran estas asociaciones
interespecíficas que sorprenden por su constancia.
f. La estacionalidad determina que la mayoría de las especies arbóreas y arbustivas con
frutos carnosos fructifiquen en la época lluviosa y sean escasos durante la época seca.
Probablemente sean las lluvias y no las temperaturas las que determinan la estacionalidad,
ya que estos bosques están prácticamente libres de heladas. Esto controla la población de
frugívoros, las especies residentes son frugívoras generalistas u omnívoras, muchas de ellas
erráticas, aprovechando parches con abundancia de frutos. Otros frugívoros son migratorios y arriban a criar en época de abundancia. Muchas especies son erráticas en la época de
escasez, es común observar durante el otoño e invierno individuos o grupos de individuos de
distintas especies que están de paso y no son vistas nuevamente en el sitio. Esta limitación
en la oferta de frutos, con una marcada estacionalidad en la disponibilidad es la causante de
que no se instalasen en las Yungas de Argentina grupos o taxones típicamente frugívoros
como Ramphastidae o Pipridae, de amplia distribución en los bosques tropicales de
Sudamérica. Por otro lado, las plantas que fructifican durante la época de escasez invernal,
juegan un rol muy importante para la comunidad entera de frugívoros en el Bosque de
Transición. El tipo de desplazamientos de varias especies de aves, que parecen ser erráticos,
es una respuesta a la distribución en parches de P. carthagenensis y C. iguanae, como
ocurre en Thraupis sayaca, Pipraeidea melanonota, Thlypopsis sordida, Atlapetes
citrinellus y Poospiza erythrophrys. Varias especies realizan desplazamientos entre el
Chaco y las Selvas Montanas, los más conspicuos son Amazona aestiva y Ramphastos toco,
pero hay un número grande de especies más pequeñas que dependen de estos
desplazamientos para sobrevivir (Saltator aurantiirostris, Pheucticus aureoventris,
Euphonia chlorotica). Estas especies se ven afectadas al ser cortada la vía natural de
migración altitudinal debido a la deforestación de los pedemontes.
g. La característica de la estructura trófica de la avifauna de estos bosques es la
preponderancia de la omnivoría durante el invierno y de la frugivoría entre Octubre y Enero.
h. Se produce desde los límites con Bolivia, un empobrecimiento en el número de especies
aves como ocurre con otros pisos altitudinales de las Yungas. Existe un paulatino
aislamiento del bosque de palo blanco con el de Tipa y Pacará, distribuyéndose actualmente
en forma discontinua. La distribución de varias especies (R. toco, P. decumanus, A.
torquatus, B. bivittatus), llega hasta Metán y Lumbreras, en el sur de la provincia de Salta.
Este paulatino aislamiento de masas boscosas anteriormente continuas me permite
interpretar la presencia de Todirostrum plumbeiceps (nueva especie agregada a la provincia

71
de Tucumán) como una población relictual en las Sierras de Medina, serranías aisladas del
resto de las masas boscosas húmedas, por ser ésta una especie residente, sin capacidad de
desplazamientos migratorios.
La caída en el número de especies desde el bosque de palo blanco hacia el sur, puede
fundamentarse en varias razones:
- Debido a variaciones en el medio ambiente, factor reconocido como limitante de la
diversidad en la ecología de comunidades (MacArthur, 1969), en este caso producto de la
estacionalidad de los recursos y determinante en la porción más austral de las Yungas de la
pauperización de la avifauna. El análisis de las formas vegetales y animales de las Yungas
de Argentina, revela una alta diversidad de formas vivientes y una estructura de vegetación
propia, en verano, de un bosque tropical húmedo. Pero se encuentran sometidas a un
régimen climático con inviernos rigurosos, en ocasiones nevados en sus partes altas.
Algunos caracteres propios de los bosques templados aparecen junto a tan alta diversidad,
como la dominancia de ciertas especies arbóreas. El aumento proporcional de grupos
generalistas hacia el sur (oscines y suboscines tiránidos), puede también relacionarse con
cambios estacionales en los recursos, ya que son grupos resistentes a cambios climáticos y
más plásticos que los grupos especializados (suboscines furnaroides), pudiendo migrar o
cambiar de dieta según las presiones a las que se ven sometidos.
- Debido a la complejidad estructural del bosque, relacionada con el número de especies
que puede soportar. Orians (1969), sugirió que uno de los posibles factores que contribuyen
al mantenimiento de un gran número de especies en los trópicos es debido a las numerosas
subdivisiones posibles en el plano horizontal en estos bosques. Además, hay una estrecha
relación entre la complejidad estructural del ambiente y la diversidad, densidad y biomasa
de la avifauna (MacArthur y MacArthur, 1961; Rotemberry y Wiens, 1980). Existe una
disminución en el número de especies vegetales en las Yungas, desde Bolivia hacia el sur, ya tratada anteriormente, y también una mayor variación temporal en la estructura de la
vegetación a lo largo del año. MacArthur (1970, 1972), estableció que ante el aumento en
los niveles de productividad, aumentaba la diversidad mediante un proceso de compactación
de especies, seguido por un incremento en la especialización y/o un incremento en la
superposición de las preferencias de cada especie (MacArthur, 1970). A través de un
incremento en los recursos aumentaba el número de nichos posibles en la comunidad.
Acaso, el incremento o disminución en la especialización en las aves depende directamente
de la disponibilidad y abundancia de los recursos durante todo el año. La adición de formas
de grupos generalistas hacia el sur (Tyrannidae, Thraupidae, Emberizidae), estaría
determinada por los cambios estacionales en la disponibilidad de recursos. Si analizamos
los taxones de aves presentes en los bosques subtropicales del sur de Bolivia y zonas
limítrofes del norte de Salta (Olrog, 1963, 1984), y que desaparecen hacia el sur, o que
disminuyen en el número de especies, vemos que tienen en común el ser aves de bosques
tropicales, insectívoros o frugívoros especializados (Trogonidae, Momotidae, Bucconidae,
Ramphastidae, Trochilidae, Picidae, Dendrocolaptidae, Formicariidae y Coerebidae).
Willis (1975), estudió la respuesta de la avifauna de un bosque amazónico en el oeste de
Matto Grosso al paso de un frente frío, observando que la actividad de los insectívoros se
redujo como consecuencia de la inactividad de insectos, en cambio los frugívoros y
omnívoros se hicieron más conspicuos. El autor sugirió que ante un cambio drástico en las
condiciones climáticas, los insectívoros especializados se ven inhibidos, aumentando el
porcentaje de omnívoros. Es más probable que estos cambios sucedan más comúnmente en
los bordes o áreas angostas de bosque que en las grandes extensiones uniformes de selvas
amazónicas. Como resultado, los suboscines furnaroides (Familias Furnariidae,
Dendrocolaptidae y Formicariidae), insectívoros especializados, además de otros grupos
como picaflores, van siendo empujados hacia dentro de las selvas, reemplazados por
Oscines omnívoros o por Suboscines tiranoides frugívoros-insectívoros (Tyrannidae,

72
Thraupidae, Cotingidae), omnívoros en su mayoría o que pueden migrar en períodos
desfavorables. Típicamente, los Suboscines furnaroides dominan en las selvas amazónicas y
los Suboscines tiranoides en las zonas secundarias o periféricas (Novaes, 1973). Pocas
especies de furnáridos, todos ellos muy especializados como S. scutatus, terrícola
insectívoro; S. superciliosa, del estrato arbustivo denso y P. rufosuperciliatus, especie
trepadora insectívora, son residentes típicos en los Bosques de Transición de Argentina.
Relativamente más dendrocoláptidos se encuentran presentes, aunque sin alcanzar la
cantidad de especies del Chaco Serrano lindante.
La fauna más diversa de las Yungas de Bolivia y del norte de Salta, con respecto a la que se
encuentra hacia el sur, es consecuencia de una mayor riqueza de recursos, de una mayor
disponibilidad a lo largo del año y de estar más próxima a las áreas amazónicas, permitiendo
un mejor intercambio faunístico.
La historia biogeográfica del noroeste Argentino está influida por el mismo tipo de
fenómenos pleistocénicos que ocurrieron en la Amazonia. Los principales rasgos en los
patrones de distribución de las aves de las Yungas son:
1.- La avifauna está relacionada estrechamente con la Amazónica, manifestándose como una
versión empobrecida de la misma.
2.- La avifauna del noroeste y noreste de Argentina estuvieron en contacto en épocas más
húmedas, con mayor desarrollo de las selvas, por lo que contienen numerosos elementos
comunes.
Al igual que el resto de las Yungas, avifauna de los Bosques Pedemontanos de Transición
de Argentina tiene estrecha relación con la avifauna de la zona tropical húmeda de
Sudamérica y aunque estos bosques tienen una distribución subtropical, la mayor parte de
los Géneros son compartidos con el Dominio Amazónico y especialmente con el resto de las
Yungas de Sudamérica (Leptoptila, Penelope, Claravis, Ara, Aratinga, Pyrrhura, Amazona, Piaya, Pulsatrix, Trogon, Momotus, Ramphastos, Piculus, Picumnus, Sittasomus,
Veniliornis, Xiphocolaptes, Philydor, Synallaxis, Thamnophilus, Pachyramphus,
Todirostrum, Legatus, Elaenia, Tolmomyias, Catharus, Turdus, Cychlaris, Vireo,
Basileuterus, Thraupis, Piranga, Cyanocompsa, Arremon, Poospiza y Cacicus).
Con la Caatinga y el Cerrado de Brasil, comparte pocos géneros comunes como
Crypturellus, Lepidocolaptes, Ara, Tyrannus, Philydor, Phacellodomus, Herpsilochmus,
Cyanocompsa y Sicalis (Sick, 1965), habiendo más afinidades entre ellos y el Chaco, con
muchas especies comunes como Aratinga acuticaudata, Myrmorchilus strigilatus,
Schoeniophylax phryganophila, Xolmis irupero, Stigmatura budytoides, Ammodramus
humeralis, Carduelis magellanica, Molothrus badius (Short, 1975), y casi todos los
géneros.
Debido a su contacto en su lado oriental con el Chaco a lo largo de todo su rango de
distribución, muchas especies de los bosques xerófilos ingresan en zonas de bordes o
alteradas, aunque son raras en el bosque maduro (Ortalis canicollis, Cariama cristata,
Chunga burmeisteri, Picumnus cirratus pilcomayensis, Taraba major, Myrmorchilus
strigilatus, Polioptila dumicola, Coryphospingus cucullatus, Poospiza melanoleuca,
Sicalis luteola). Las especies chaqueñas no forman parte del ensamble de especies propias
del bosque no alterado, por lo que éste no puede ser considerado como una mezcla de
elementos del Chaco y de la Selva Montana.
El Bosque Pedemontano es, de los bosques de las Yungas, el que más ha sufrido el impacto
del poblamiento en el noroeste de Argentina. A mediados del siglo XVI se establecieron los
primeros colonos españoles y se introdujeron los primeros animales domésticos
(Santamarina, 1966). A fines del siglo XVI había ya 4 ciudades ubicadas a lo largo de su
rango de distribución (San Salvador de Jujuy, Salta, Esteco e Ibatín). Esta degradación
constituida principalmente por el clareo del bosque para pastaje y extracción de especies

73
maderables se convirtió en transformación con la introducción, a mediados del siglo XIX,
de la industria azucarera.
Como afectó esta transformación a nivel de comunidades de aves es algo que no se conocerá
nunca completamente, pero sin duda la extinción casi total en la parte central de su rango de
distribución afectó a los pisos superiores. El tremendo impacto que significó la eliminación
de la continuidad entre los bosques montanos húmedos y los xerófilos de las llanuras del
este, puede inferirse por la desaparición de algunas especies muy conspicuas de la provincia
de Tucumán, en forma coincidente con el desarrollo de esta etapa industrial. Algunos
mamíferos como el tapir y el jaguar, boas, grandes loros, el tucán, aves especializadas como
formicáridos o grandes rapaces como águilas realizaban desplazamientos altitudinales, eran
típicos del estrato bajo y denso de estos bosques o poseían grandes territorios (Martín De
Moussy, 1864). La presencia de dos o tres especies de rapaces carroñeros en cada censo
(jotes y caranchos), y la rareza de especies de verdaderas aves de presa tiene relación directa
al manejo de la naturaleza por parte del hombre. Sin el grupo de aves dependiente de
carroña, la abundancia de "verdaderos rapaces" decrece con el incremento de las influencias
humanas en los ambientes naturales (Reichholf, 1974). En general, todo el proceso de
degradación se evidencia en una decreciente diversidad de especies, que fue corroborado
aplicando el Indice de Simpson que remarca la abundancia de especies poco frecuentes,
dando como resultado valores de una elevada diversidad específica para el sector norte del
Bosque Pedemontano. Varias especies que dependen del Bosque de Transición para su
supervivencia, por su creciente rareza, probablemente están cercanas a la extinción como
Buteo nitidus, Buteo leucorrhous, Amazona tucumana, Amazona aestiva, Ara auricollis,
Piaya cayana, Batara cinerea, Pachyrhamphus viridis y Psaracolius decumanus.
Los bosques de Yungas del noroeste argentino se encuentran razonablemente representados
por áreas naturales protegidas (Parques Nacionales Baritú, Calilegua, El Rey y Los Alisos), sin embargo, ninguno incluye áreas de tamaño suficientemente apropiado de Bosque
Pedemontano bajo medidas estrictas de conservación. Por este motivo, sería recomendable
iniciar gestiones para aumentar áreas pedemontanas protegidas a lo largo del estrecho rango
de distribución de los Bosques de Transición del noroeste argentino.
LITERATURA CITADA
Aceñolaza, F. y A. Toselli. 1981. Geología del noroeste argentino. Universidad Nacional
de Tucumán, 212 p.
Adamoli, J. 1972. Frecuencia, confinamiento, y transgresividad de especies del Chaco
Argentino. Resúmenes de la I Reunión Argentina de Ecología.
Austin, G. T. y E. L. Smith 1972. Winter foraging ecology of mixed insectivorous birds
flocks in oak woodland in southern Arizona. The Condor, 74:17-24.
Ayarde, H. R. 1995. Estructura de un sector de selva pedemontana. Reserva Fiscal Parque
La Florida, Tucumán (Argentina), Pp. 19-52. En: Investigación, conservación y
desarrollo en selvas subtropicales de montaña. Proyecto de Desarrollo Agroforestal.
L. I. E. Y. (A. D. Brown y H. R. Grau, eds.).
Bombin. 1975. Afinidade paleoecológica, cronológica e estratigráfica do componente de
megamamíferos na biota de Quaternario terminal da provincia de Buenos Aires
(Argentina), Uruguai e Rio Grande do Sul (Brasil). Comun. Mus. Ci., PUCRGS,
Porto Alegre, 9: 1-28.
Brown, A. D. 1986. Autoecología de bromeliáceas epífitas del noroeste argentino y su
relación con Cebus apella (Primates). Tesis Doctoral no publicada presentada a la
Facultad de Ciencias Naturales y Museo de la Universidad Nacional de La Plata.
Brown, A.; S. C. Chalukian y L.M. Malmierca 1985. Estudio florístico-estructural de un
sector de selva semicaducifolia del noroeste argentino. I. Composición florística,
densidad y diversidad. Darwiniana, 26:27-41.

74
Buskirk, W. H. 1976. Social systems in a tropical forest avifauna. Amer. Nat.,
110:293-310.
Cabrera, A. L. 1976. Regiones Fitogeográficas Argentinas. Enciclopedia Argentina de
Agricultura y Jardinería, 2:1-85.
Cabrera, A. L. ,. y A. Willink. 1973. Biogeografía de América Latina. Serie de Biología.
O. E. A., 13:120.
Cabrera, A.; J. Yepes y M. Doello Jurado. 1947. Zoogeografía de la República Argentina.
En Geografía de la República Argentina (G.A.E.A.), T. 8:347-483.
Chapman, F. M. 1926. The distribution of bird life in Ecuador. Bull. Amer. Mus. Nat.
Hist., 55: 1-784.
Cody, M. L. 1974. Competition and the structure of bird communities. Princeton
University Press, Princeton, New Yersey.
De Schauensee, R. M. 1970. A guide to the birds of South America. The Academy of
Natural Sciences of Philadelphia, Livingston Publishing Company, Penn, 470 pp.
Dabbene, R. 1910. Ornitología Argentina. An. Mus. Nac. Bs. As. 18, (1). Buenos Aires.
Digilio, A. P. y. P. R. Legname. 1966. Los árboles indígenas de la provincia de Tucumán.
Opera Lilloana, 15:1-29.
Fitzpatrick, J. W. 1980. Wintering of North American tyrant flycatchers in the Neotropics.
in: Migrant Bird in the Neotropics. A. Keast and E.S. Morton Eds. Smithsonian
Institution Press, :67-78. Sonderdruc aus: Bonn. Zool. Beitr., 25:87-117.
Haffer, J. 1969. Speciation in Amazonian forest birds. Science, 165:131-136.
Haffer, J. 1970. Entstehung und Ausbreitung nord -Andiner Bergvögel. Zool. Jb. Syst. Bd.,
97:301-337.
Haffer, J. 1974a. Pleistozane differenzierung der amazonischen vogelfauna.
Haffer, J. 1974b. Avian speciation in tropical South America, with systematic survey of the toucans (Ramphastidae) and jacamars (Galbulidae). Cambridge, Massachussets,
Nuttal Ornithological Club Publications, 14.
Haffer, J. 1982. General aspects of the refuge theory. In: biological diversification in the
tropics, pp. 6-24 (Gt. Prance, Ed.). New York, Columbia University Press.
Holmes, R. T. Jr. Bonney y S. W. Pacala. 1979. Guild structure of the Hubbard Brook bird
community: a multivariate aproach. Ecology, 60:512-520.
Holmes, R. T. y H. F. Recher. 1986. Determinants of guild structure in forest communities:
an intercontinental comparison. The Condor, 88:427-439.
Holmes, R. T. y H. F. Recher. 1986. Search tactics of insectivorous birds foraging in an
Australian eucalypt forest. Auk, 103:515-530.
Hoy, G., F. Contino y E. Blake. 1963. Addendae a la avifauna argentina. Bol. Acad. Nac.
Cienc. Córdoba.
Hueck, K. 1951. Die Sudgrenze des Tucumanisch-Bolivianischen Waldebieted in Nordues
Argentinient. Mit. Albildungen. Erdkunden Ban V. Bonn, 7.
Hueck, K. 1954. Waldbäume und waldtypen aus NW-Argentinien. Berlín -
Grunewald, Fritz Haller Verlag., 31 pp.
Hueck, K. 1978. Los bosques de sudamérica. Ecología, composición e importancia
económica. Sociedad Alemana de Cooperación Técnica, Göttingen, Alemania.
Hutto, R. L. 1987. A description of mixed-species insectivorous bird flocks in western
Mexico Condor 89:282-292.
Grau, H. R., M. F. Arturi, A. D. Brown y P. G. Aceñolaza. 1997. Floristic and structural
pattern along a chronosequence of secondary forest succession in Argentinean
subtropical montane forests. Forest Ecology and Management 95:161-171.
Jansen, D. H. 1983. No Park is an island: increase interference from outside as Park size
decreases. Oikos, 41:402-410.

75
Karr, J. R. 1971. Structure of avian communities in selected Panama and Illinois habitats.
Ecological Monographs, 41:207-228.
Karr, J. R. 1976. On the relative abundance of migrants from the North Temperate Zone in
tropical habitats. Wils. Bull., 88: 433-458.
Karr, J. R. 1976. Withing and betwen-habitat avian diversity in African and Neotropical
lowlands habitats. Ecological Monographs, 46:457-481.
Karr, J. R. y R. R. Roth. 1971. Vegetation structure and avian diversity in several New
World áreas. Amer. Nat., 105.
Kuhn, F. 1924. Estudio fisiográfico de las Sierras de Tucumán. Fac. de Fil. y Letras.
U.N.T.
Kuhn, F. y Rohmeder, G. 1924. Estudio fisiográfico de las Sierras de Tucumán. Instituto
de Estudios Geográficos de la U.N.T., Monografía Nº 3,
Lucero, M. M. y E. A. Alabarce. 1980. Frecuencia de especies e individuos en una parcela
de la selva misionera. Revista del Museo Argentino de Ciencias Naturales
Bernardino Rivadavia. Tomo II, Nº 7.
Mac Arthur, R. H. 1972. Geographical Ecology. Harper and Row, New York.
Mac Arthur, R. H. y.J. W. Mac Arthur. 1961. On birds species diversity. Ecology,
42:594-598.
Mac Arthur, R. H. 1970. Species packingand competitive equilibrium for many species.
Theoretical Population Biology, 1: 1-11.
Mac Arthur, R. H. y E. O. Wilson. 1967. The theory of island biogeography. Princeton
University Press. Princeton. New Yersey,
Malizia, L.
Margaleff, R. 1958. Information Theory in ecology. Gen. Syst., 3:36-71.
Marshal, L. G., A. Berta, R. Hoffstetter, R. Pascual, O. A. Reig, M. Bombin y A. Mones. 1984. Mammals and stratigraphy: geochronology of South America.
Paleovertebrata, Montpellier, Mém. Extr., pp. 1-76.
Martin De Moussy, V. 1864. Description geographique et statistique de la Confédération
Argentine. Librairie de Firmin Didot Fréres, Fils et Cie., Vol. 3.
Mayr, E. 1963. Animal species and evolution. Harvard University Press. Cambridge,
Mass.,
Meyer, T. 1963. Estudio sobre la Selva Tucumana. La Selva de Mirtáceas de Las Pavas.
Opera Lilloana. Instituto Miguel Lillo. U.N.T., 10
Morse, D. H. 1970. Ecological aspects of some mixed-species foraging flocks of birds.
Ecol. Monogr., 40:310-344.
Moynihan, M. 1979. Geografic variation in social behavior and in adaptations to
competition among Andean birds. Publ. Nutall Ornithol. Club, 18:1-162.
Moynihan, M. 1962. The organization and probable evolution of some mixed species
flocks of Neotropical birds. Smithson. Misc. Collect., 143(7):1-140.
Munn, C. A. y.J. W. Terborgh. 1979. Multi-species territoriality in neotropical foraging
flocks. The Condor, 81:338-347.
Myers, J. P. 1980. The pampas shorebird community: interactions between breeding and
nonbreeding members, pp. 37-49. En: Migrant Bird in the Neotropics. (A. Keast
and E.S. Morton Eds.) Smithsonian Institution Press.
Narosky, T. y D. Izurieta. 1988. Guía para la identificación de las aves de Argentina y
Uruguay. Asoc. Ornit. del Plata, 345 pp.
Nicéforo María, H. 1947. Notas sobre aves de Colombia, II. Caldasia, 4(19):317-377.
Novaes, F. C. 1973. Aves de uma vegetacao secundaria na foz do Amazonas. Publ.
Avulsas, Museu Goeldi, 21:1-88.
Olivares, A. 1959. Aves migratorias en Colombia, II. Revista Academia Colombiana de
Ciencias, 10:41.

76
Olrog, C. C. 1959. Las aves argentinas. Una guía de campo. Tucumán, Instituto Miguel
Lillo.
Olrog, C. C. 1962. El anillado de aves en Argentina. 1961 - 1962. Segundo Informe.
Suplemento de Neotrópica. 8:27.
Olrog, C. C. 1963. Notas sobre aves bolivianas. Acta Zoológica Lilloana, XIX:407-478.
Olrog, C. C. 1968. El anillado de aves en Argentina. 1964 - 1966. Quinto Informe.
Neotrópica. Vol 14, Nº 43.
Olrog, C. C. 1968. Las aves sudamericanas: una guía de campo. U.N.T.
F.-I.M.L., Tomo Primero.
Olrog, C. C. 1969. Biogeography and Ecology in South America. Edited by E. J. Fittkau, J.
Illies, H. klinge, J. H. Schwabe y H. Sioli. Publishers The Hague.
Olrog, C. C. 1971. El anillado de aves en Argentina. 1961-1971. Séptimo Informe.
Neotrópica, 17(53):97-100.
Olrog. C. C. 1972. Adiciones a la avifauna argentina. Acta Zoológica Lilloana, Tomo
XXVI:17.
Olrog, C. C. 1974. Notas ornitológicas sobre la colección del Instituto Miguel Lillo. Acta
Zoológica Lilloana, XXXI:8:69-76.
Olrog, C. C. 1978. A comparison of Suboscines and Oscine radiation. Symposium on
speciation in south american birds. Separatum ex Actis XVII Congressus
Internationalis Ornithologici, Berlín, pp. 1262-1267.
Olrog, C. C. 1979. Nueva lista de la avifauna argentina. Opera Lilloana, XXVII pp 324.
Olrog, C. C. 1984. Las aves argentinas: "Una nueva guía de campo". Administración de
Parques Nacionales, 352 pp.
Olrog, C. C. 1984. La dispersión en Sudamérica de Suboscines versus Oscines. Rev. del
Mus. Arg. de Ciencias Nat. Bernardino Rivadavia. Tomo XIII, Nº18. Olrog, C. C. 1985. Status of wet forest raptors in northwestern Argentina. ICBP Technical
Publication Nº 5.
Olrog, C. C. y P. Capllonch. 1986. Biornitología argentina. Supl. Historia Natural 2:1-41.
Olrog, C. C. y F. Contino. 1970. Dos especies nuevas para la avifauna argentina.
Neotrópica 16 (48):95.
Orians, G. 1969. The number of birds species in some tropical forests. Ecology,
50:5:783-797.
Pearson, D. L. 1980. Bird migration in amazonian Ecuador, Perú and Bolivia. in: Migrant
Bird in the Neotropics. A. Keast and E.S. Morton Eds. Smithsonian Institution
Press, 273-283.
Pianka, E. R. 1966. Latitudinal gradients in species diversity: a review of concepts.
American Naturalist, 100:33-46.
Pinto, O. 1944. Algunas adendas a avifauna de Monte Alegre. Papéis Avulsos do Dep. de
Zool. Secretaría da Agricultura, Sao Paulo, Brasil.
Pinto, O. 1950. Peter W. Lund e sua contribucao a ornitología brasileira. Papéis
Avulsos do Dep. de Zool., IX, N 18:269-283.
Pinto, O y E. De Camargo. 1948. Sobre uma colecao de aves do Rio Das Mortes (Estado de
Mato Grosso). Papéis Avulsos do Dep. de Zool. Secretaría da Agricultura, Sao
Paulo, Brasil, 8(26):287- 336.
Pinto, O. y E. De Camargo. 1952. Nova contribucao a ornotologia do Rio Das Mortes. Dep.
de Zool., Secretaría da Agricultura, Sao Paulo, Brasil, 10(11):213-234.
Powell, G. V. N. 1980. Migrant participation in Neotropical mixed species flocks, pp.
477-483. In: migrants birds in the Neotropics (A. Keast and E.S. Morton Eds.),
Smithsonian Institution Press.

77
Powell, G. V. N. 1985. Sociobiology and the adaptative significance of interspecific
foraging flocks in the Neotropics. Ornith. Monog. American Ornithologists Union,
36.
Prado, D. E. 1995. Selva Pedemontana: contexto regional y lista florística de un ecosistema
en peligro. Pp. 19-52. En: Investigación, conservación y desarrollo en selvas
subtropicales de montaña (Brown, A. D. y H. R. Grau, eds), Proyecto de Desarrollo
Agroforestal. L. I. E. Y.
Rappole, J. H. y.D. W. Warnes. 1980. Ecological aspects of migrants birds
behavior in Veracruz, Mexico, pp. 353-393. in: Migrant birds in the Neotropics.(A.
Keast and E.S. Morton Eds.) Smithsonian Institution Press.
Reichholf, J. 1974. Artenreichtum, häufigkeit und diversität der greifvögel in einigen
gebieten von Südamerika. Journal für Ornithologie, 115 (4):381-397.
Ridgely, R.S. y G. Tudor. 1989. The birds of South America. Vol. I The oscines
passerines. University of Texas Press, Austin; 516 pp.
Rohlf, F. J. 1990. NTSYS - PC. Numerical Taxonomy and Multivariate Analysis System.
Dep. of Ecol. and Evol. State Univ. New York.
Rotenberry, J. T. y J. A. Wiens. 1980. Habitat structure, patchiness and avian communities
in North American steppe vegetation: a multivariate analysis. Ecology,
61(5):1228-1250.
Sabo, S. R. y R. T. Holmes. 1983. Foraging niches and the structure of forest birds
communities in contrasting montane habitats. The Condor, 85:121-138.
Santillán de Andrés, S. E. 1966. Reseña de la evolución del paisaje rural en la provincia de
Tucumán. Humanitas, Rev. de la Fac. de Fil. y Letras. U.N.T. 19:59-65.
Santillán de Andrés, S. E. y T. R. Ricci. 1966. La Región de la cuenca Tapia-Trancas.
Serie Monográfica. Fac. de Fil. y Letras. Dep. de Geografía. U.N.T., 15:1-69. Santillán de Andrés, S. E., E. B. De Santamarina, T. R. Ricci y E. J. Wurschmidt. 1967.
La región de las sierras del nordeste de la provincia de Tucumán. Serie
Monográfica N 16. Fac. de Fil. y Letras. Dep. de Geografía. U.N.T.
Short, L. 1972. Systematics and behavior of South American flickers (Aves: Colaptes).
Bull. Amer. Mus. Nat. Hist., 149:1-110.
Short, L. 1975. A zoogeographic analysis of the South American Chaco avifauna. Bull.
Amer. Mus. Nat. Hist., 154:Nº3.
Sick, H. 1965. A fauna do Cerrado. Arquivos de Zoologia, Dep. de Zool. Sao Paulo 12:71-
93.
Simpson, E. H. 1949. Measurement of diversity. Nature 163: 688 pp.
Smith, W. J. y J. Vuilleumieur. 1971. Evolutionary relationships of some South American
ground tyrants. Bull. Mus. Comp. Zool., Harvard Univ. Cambridge, Mass. U.S.A.,
141(5):181-232.
Terborgh, J. y J. Faaborg. 1980. Factors affecting the distribution and abundance of North
American migrants in the Eastern Caribean Region. In: Migrant birds in the
Neotropics. Ecology, behavior, distribution and conservation. (A. Keast and E.
Morton Eds.), Smithsonian Institution.
Torres Bruchmann, E. 1973. Atlas agroclimático y bioclimático de Tucumán. Segunda
Parte. Public. Especial. Fac. de Agronomía y Zootecnia. U.N.T., 10.
Vides Almonacid, R. 1992. Estudio comparativo de las taxocenosis de aves de los Bosques
Montanos de la Sierra de San Javier, Tucumán: bases para su manejo y
conservación. Trabajo de Tesis Doctoral no publicada. Facultad de Ciencias
Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán.
Vervoorst, F. 1979. La vegetación del noroeste de Argentina y su degradación. Serie
conservación de la Naturaleza 1:1-9. Ministerio de Cultura y Educación. Fundación
Miguel Lillo. Tucumán.

78
Vervoorst, F. 1982. Noroeste. Serie Conservación de la Naturaleza, 2:9-24.
Vuilleumier, F. 1965. Relantionships and evolution within the Cracidae (Aves,
Galliformes). Bull. Mus. Comp. Zool., Harvard University. 134:Nº 1.
Vuilleumier, F. 1969. Pleistocene speciation in birds living in the high Andes. Nature,
223:1179-1180.
Vuilleumieur, F. 1969. Systematics and evolution in Diglossa (Aves, Coerebidae). Amer.
Mus. Novit. Nº2381, 44p.
Vuilleumier, F. 1970. Insular biogeography in continental regions. I. The northern Andes
of South America. The Amer. Nat., Vol 104, Nº938.
Vuilleumier, F. 1971. Evolutionary relantionships of some south american ground tyrants.
Bull. of Mus. of Comp. Zool. Harvard University, Cambridge, Mass. Vol 141, Nº5.
Vuilleumieur, F. 1980. Speciation in birds in the high Andes. Simposium on speciation in
South American birds. XVII Congressus Internationalis Ornithologsci. Berlin.
Vuilleumier, F. 1985. Forest birds of Patagonia: ecologycal, geography, speciation,
endemism, and faunal history. In: Neotropical Ornothology: 255-304. (P. A.
Buckley; M. S. Foster; E. S. Morton; R. S. Ridgely; F. G. Buckley, Eds.). American
Ornothologists Union Monographs 36.
Wetmore, A. 1965. Water, prey, and game birds of North America. National Geographic
Society. Washington, D. C.
Whittaker, R. H. 1967. Gradient analysis of vegetation. Biol. Rev., 42[207-246.
Wiens, J. A. and J. T. Rotemberry. 1980. Bird community structure in cold shrub deserts:
competition or chaos. Proc. XVII Int. Congr. (Berlin), :1063-1070.
Willis E. O. 1966. The role of migrant birds at swarms of army ants. Living Bird,
5:187-231.
Willis, E. O. 1975. Aves observadas en bosques en el oeste de Mato Grosso. Cta Amazonica, 6(3):339-394.
Willis, E. O. 1976. Effects of a cold wave on an Amazonian avifauna in the upper
Paraguay drainage, Western Mato Grosso, and suggestions on Oscine-Suboscine
relationships. Acta Amazonica, 6(3):379-394.

Lista 1: Lista general de las aves de los Bosques Pedemontanos de Transición de Argentina
Crypturellus t. tataupa (Temminck ) inambú del monte
Podiceps dominicus speciosus (Lynch) macá chico
Podylimbus podiceps antarcticus (Lesson) macá de pico grueso
Anhinga a. anhinga (Linné) anhinga
Phalacrocorax o. olivaceus (Humboldt) biguá
Egretta alba egretta (Gmelin) garza blanca
Egretta t. thula (Molina) garcita blanca
Egretta i. ibis (Linné) garcita bueyera
Syrigma s. sibilatrix (Temminck) chiflón
Nycticorax nycticorax hoactli (Gmelin) garza bruja o cuaco
Tigrisoma fasciatum pallescens Olrog hocó oscuro
Theristicus c. caudatus (Boddaert) bandurria común
Merganetta armata berlepschi Hartert pato de los torrentes
Amazonetta brasiliensis (Gmelin) pato cutirí
Cairina moschata (Linné) pato criollo
Oxyura vittata (Philippi) pato zambullidor chico
Cathartes aura jota (Molina) jote de cabeza roja
Coragyps atratus (Beckstein) jote de cabeza negra
Sarcoramphus papa (Linné) jote real
Accipiter bicolor guttifer Hellmayr esparvero bicolor
Accipiter striatus erythronemius Kaup esparvero chico
Buteo nitidus pallidus (Todd) agulicho gris
Buteo magnirostris saturatus (Sclater y Salvien) garganchillo
Buteo a. albicaudatus Vieillot aguilucho caeza negra
Buteogallus u. urubitinga (Gmelin) águila negra
Spizastur melanoleucus (Vieillot) águila calzada
Elanoides forficatus yetapa (Vieillot) elanio tijereta
Elanoides f. forficatus (Linné) elanio tijereta
Gampsonyx s. swainsoni Vigors milanito blanco
Micrastur s. semitorquatus (Vieillot) halcón semiacollarado
Micrastur ruficollis olrogi Amadon halcón de cuello rojo
Polyborus plancus brasiliensis (Gmelin) carancho
Polyborus ch. chimango Vieillot chimango
Falco peregrinus anatum Bonaparte halcón peregrino
Falco rufigularis ophryophanes (Salvadori) halcón plomizo mayor
Falco sparverius cinnamominus Swainson halconcito común
Ortalis c. canicollis (Wagler) charata
Penelope obscura bridgesi (G.R.Gray) pava de monte
Rallus s. sanguinolentus Swainson gallineta común
Aramides c. cajanea (P.L.S. Muller) chiricote
Gallinula chloropus galeata (Lichtenstein) polla de agua negra
Fulica leucoptera Vieillot gallareta frenteamarilla
Fulica rufifrons Philippi y Landbeck gallareta frenterroja
Cariama cristata (Linné) chuña de patas rojas
Jacana j. jacana (Linné) gallito de agua
Charadrius collaris Vieillot chorlo de collar
Columba picazuro Temminck paloma picazura
Columba cayenensis sylvestris Vieillot paloma colorada
Zenaida auriculata chrysauchenia (Reichenbach) torcaza
Columbina p. picui (Temminck) palomita común
Columbina t. talpacoti (Temminck) palomita castaña
Claravis pretiosa (Ferrari-Pérez) tórtola azul
Leptoptila verreauxi decipiens (Salvadori) bumbuna

80
Ara auricollis Cassin Maracaná de cuello dorado
Aratinga m. mitrata (Tschudi) cotorra de cara roja
Aratinga a. aurea (Gmelin) cotorra de frente dorada
Pyrrhura molinae australis Todd chiripepe salteño
Pionus maximiliani siy Souancé loro choclero
Pionus m. lacerus (Heine) loro choclero tucumano
Amazona tucumana (Cabanis) loro alisero
Amazona aestiva xanthopterix (Berlepsch) loro hablador
Piaya cayana mogenseni Peters alma de gato
Guira guira Gmelin machilo
Tapera naevia chochi (Vieillot) crespín
Tyto alba tuidara (J.E. Gray) lechuza de campanario
Otus ch. choliba (Vieillot) lechucita común
Pulsatrix perspicillata boliviana L. Kelso lechuzón anteojo
Glaucidium b. brasilianum (Gmelin) caburé
Asio s. stygius (Wagler) lechuzón negrusco
Aegolius harrisi dabbenei Olrog lechucita acanelada
Caprimulgus s. saltarius Olrog atajacaminos oscuro
Caprimulgus p. parvulus Gould atajacaminos chico
Hydropsalis brasiliana furcifera (Vieillot) atajacaminos coludo
Chaetura andrei meridionalis Hellmayr vencejo de garganta blanca
Phaethornis pretrei (Lesson y DeLattre) ermitaño de frente negra
Chlorostilbon a. aureoventris (D Orbigny y Lafresnaye) picaflor común
Amazilia chionogaster hypoleucus (Gould) picaflor de vientre blanco
Eriocnemis glaucopoides (D Orbigny y Lafresnaye) picflor de frente violeta
Sappho sparganura sappho (Lesson) picaflor coludo rojo
Microstilbon burmeisteri (Sclater) picaflor enano
Trogon curucui benhi Gould surucuá purpúreo
Ceryle t. torquata (Linné) martín pescador grande
Chloroceryle a. amazona (Latham) martín pescador mediano
Chloroceryle americana nathewsi (Laubmann) martín pescador chico
Momotus momota nattereri Sclater burgo
Ramphastos t. toco P. L. S. Müller tucán grande
Picumnus dorbignyanus Lafresnaye carpinterito manchado
Picumnus cirratus pilcomayensis Hargitt carpinterito común
Picumnus cirratus thamnophiloides Bond y Schauensee carpinterito común
Piculus rubiginosus tucumanus Cabanis carpintero dorado verde
Veniliornis frontalis (Cabanis) carpintero de frente negra
Campephilus l. leucopogon (Valenciennes) carpintero negro dorso crema
Campephilus l. major (Olrog) carpintero negro dorso crema
Sittasomus g. griseocapillus (Vieillot) trepadorcito común
Xiphocolaptes major castaneus (Ridgway) trepador grande colorado
Dendrocolaptes picumnus casaresi (Steullet y Deautier) trepadorcito mediano común
Lepidocolaptes angustirostris dabbenei (Esteban) trepador chico picolargo
Cinclodes fuscus tucumanus Chapman remolinera parda
Synallaxis f. frontalis Pelzeln pijuí de frente gris
Synallaxis s. superciliosa Cabanis pijuí de ceja canela
Synallaxis scutatus whitii Sclater pijuí anaranjado
Certhiaxis p. pyrrhophia (Vieillot) curutié de vientre blanco
Phacellodomus rufifrons sincipitalis Cabanis espinero frentirufo
Philydor rufosuperciliatus oleaginea (Sclater) titirí
Batara cinerea argentina Shipton batará grande
Taraba m. major (Vieillot) batará blanco y negro
Thamnophilus r. ruficapillus Vieillot bataracito pardo

81
Thamnophilus caerulescens connectens Berlepsch bataracito común
Herpsilochmus pileatus atricapillus Pelzeln tiluchí de corona negra
Myrmorchilus strigilatus (Wied) hormiguero moteado
Pachyramphus v. viridis (Vieillot) anambé verde
Pachyramphus polychopterus spixii (Swainson) anambé común
Platypsaris rufus audax (Cabanis) anambé copetón
Myiotheretes striaticollis pallidus (Berlepsch) monjita rojiza
Ochthoeca leucophrys tucumana Berlepsch pitajo gris
Sayornis nigricans latirostris Cabanis pitajo negruzco
Knipolegus a. aterrimus Kaup viudita negra
Knipolegus cabanisi Schulz viudita plomiza
Knipolegus striaticeps D Orbigny y Lafresnaye viudita gris
Pyrocephalus r. rubinus (Boddaert) viudita roja
Satrapa icterophrys (Vieillot) viudita amarilla
Machetornis r. rixosa (Vieillot) picabuey
Tyrannus s. savanna Vieillot tijereta
Tyrannus m. elancholicus Vieillot pitirre real
Empidonomus v. varius (Vieillot) tuquito manchado
Legatus l. leucophaius (Vieillot) tuquito pirata
Myiodynastes maculatus solitarius (Vieillot) pitirre rayado
Pitangus sulphuratus bolivianus (Lafresnaye) quetupí
Casiornis rufa (Vieillot) burlisto rufo
Myiarchus t. tyrannulus (P.L.S. Müller) burlisto colicastaño
Myiarchus swainsoni ferocior Cabanis burlisto pardo
Myiarchus tuberculifer atriceps (Cabanis) burlisto de cabeza negra
Contopus cinereus pallescens (Hellmayr) frailecillo plomizo
Contopus fumigatus brachyrhynchus Cabanis frailecillo copetón
Empidonax t. traillii (Audubon) frailecillo norteamericano
Empidonax euleri argentinus (Cabanis) frailecillo de ala rufa
Myiophobus fasciatus flammiceps (Temminck) mosqueta de pecho rayado
Hirundinea ferruginea pallidor (Hartert y Goodson ) birro
Tolmomyias sulphurescens pallescens (Hartert y Goodson) pico chato sulphuroso
Todirostrum plumbeiceps viridiceps (Salvadori) mosqueta cabeza canela
Todirostrum margaritaceiventer (D Orbigny y Lafresnaye) mosqueta común
Phylloscartes ventralis tucumanus Zimmer mosquetita verde
Serpophaga subcristata Vieillot piojito de vientre amarillo
Serpophaga munda Berlepsch piojito de vientre blanco
Serpophaga nigricans (Vieillot) piojito plomizo
Mecocerculus l. leucophrys (D Orbigny y Lafresnaye) piojito de garganta blanca
Elaenia a. albiceps (D Orbigny y Lafresnaye) fío fío común
Elaenia parvirostris Pelzeln fío fío picocorto
Elaenia strepera Cabanis fío fío plomizo
Elaenia o. obscura (D Orbigny y Lafresnaye) fío fío obscuro
Myiopagis viridicata (Vieillot) fío fío copete amarillo
Suiriri s. suiriri (Vieillot) suiriri
Camptostoma obsoletum bolivianum Zimmer bobito chiquito
Leptopogon a. amaurocephalus Tschudi movedizo gorra sepia
Progne modesta elegans Baird golondrina negra
Notiochelidon c. cyanoleuca (Vieillot) golondrina barranquera
Stelgidopteryx r. ruficollis (Vieillot) golondrina de ribera
Cyanocorax ch. chrysops (Vieillot) urraca
Cyanocorax ch. tucumanus Cabanis urraca
Troglodytes aedon rex Berlepsch y Leverkühn carrasquita
Catharus dryas blakei (Olrog) zorzalito overo

82
Catharus ustulatus swainsoni (Tschudi) zorzalito norteamericano
Turdus n. nigriceps Cabanis zorzal plomizo
Turdus r. rufiventris Vieillot zorzal pecho colorado
Turdus amaurochalinus Cabanis chalchalero
Turdus albicollis paraguayensis Chubb zorzal de cuello blanco
Polioptila d. dumicola (Vieillot) tacuarita azul
Cyclarhis gujanensis viridis (Vieillot) juan chiviro
Vireo olivaceus chivi Vieillot chiví común
Parula p. pitiayumi (Vieillot) pitiayumi
Geothlypis aequinoctialis velata (Vieillot) arañero de cara negra
Myioborus b. brunniceps (D Orbigny y Lafresnaye) arañero cabeza castaña
Basileuterus bivittatus argentinae Zimmer arañero ceja negra
Basileuterus culicivorus viridiscens Todd arañero chico
Diglossa baritula sittoides (D Orbigny y Lafresnaye) mielero payador
Euphonia musica aureata (Vieillot) curruñatá corona azul
Euphonia chlorotica serrirostris D Orbigny y Lafresnaye curruñatá común
Pipraeidea melanonota venezuelensis (Sclater) viuva
Thraupis sayaca obscura Naumburg celestino
Thraupis bonariensis schultzei Brodkorb naranjero
Piranga f. flava Vieillot fueguero
Thlypopsis s. sordida (D Orbigny y Lafresnaye) pioró cabeza anaranjada
Chlorospingus ophthalmicus argentinus Hellmayr fruterito ojo blanco
Saltator c. caerulescens Vieillot pepitero plomizo
Saltator a. aurantiirostris Vieillot pepitero pico dorado
Paroaria c. capitata (D Orbigny y Lafresnaye) cardenal sin copete
Cyanocompsa cyanea argentina (Sharpe) reina mora
Tiaris o.obscura (D Orbigny y Lafresnaye) tordillo pardo
Sporophila c. caerulescens (Vieillot) corbatita
Sicalis flaveola pelzelni Sclater jilguero dorado
Sicalis luteola luteiventris (Meyer) jilguero chico
Coryphospingus cucullatus fargoi Brodkorb brasita de fuego
Atlapetes citrinellus (Cabanis) arbustero yungueño
Atlapetes torquatus borelli (Salvadori) arbustero de vientre blanco
Arremon flavirostris dorbignii Sclater gran cantor
Zonotrichia capensis hypoleuca (Todd) chingolo
Zonotrichia capensis pulacayensis (Ménégaux) chingolo
Poospiza erythrophrys Sclater
Poospiza melanoleuca (D Orbigny y Lafresnaye) monterita común
Poospiza nigrorufa whitei Sclater monterita ocrácea
Carduelis magellanica alleni (Ridgeway) cabecita negra común
Molothrus b. bonariensis (Gmelin) tordo común
Molothrus b. badius (Vieillot) tordo músico
Psaracolius decumanus maculosus (Chapman) conoto negro
Cacicus chrysopterus (Vigors) boyero ala amarilla
Icterus cayenensis pyrrhopterus Vieillot toche de cobija canela

83
Lista 2: Lista de aves del Bosque de Transición de palo blanco de Piquirenda Viejo,
extremo norte de la provincia de Salta.
* Especies que no llegan en su distribución al bosque de Tipa y Pacará.
> Migrantes desde el Hemisferio Norte.
ESPECIE MAYO AGOSTO NOVIEMBRE
Crypturellus tataupa x x x
Tigrisoma fasciatum x x x
Ardea cocoi x
Cathartes aura x x x
Sarcoramphus papa x
Buteo magnirostris x x x
Buteogallus urubitinga x
Falco sparverius x x
Elanoides forficatus * > x
Micrastur semitorquatus * x x
Penelope obscura x x x
Aramides cajanea x x x
Claravis pretiosa x
Leptoptila verreauxi x x x
Amazona aestiva x x
Pyrrhura molinae * x x x
Piaya cayana x x x
Glaucidium brasilianum x
Pulsatrix perspicillata * x
Chaetura andrei * x
Phaethornis pretrei * x x x
Chlorostilbon aureoventris x x x
Amazilia chionogaster x x
Otus choliba x x
Caprimulgus saltarius * x
Trogon curucui x x
Ramphastos toco * x x x
Picumnus dorbignianus * x x x
Campephilus leucopogon x
Veniliornis frontalis x x
Xiphocolaptes major x
Dendrocolaptes picumnus * x x x
Sittasomus griseocapillus x x x
Synallaxis scutatus x x x
Philydor rufosupeciliatus x x x
Batara cinerea x
Thamnophilus caerulescens x x
Thamnophilus ruficapillus x
Sayornis nigricans x x
Knipolegus aterrimus x
Satrapa icterophrys x
Empidonax traillii *> x
Myiodynastes maculatus x
Hirundinea ferruginea x x x
Phylloscartes ventralis x x x
Serpophaga munda x
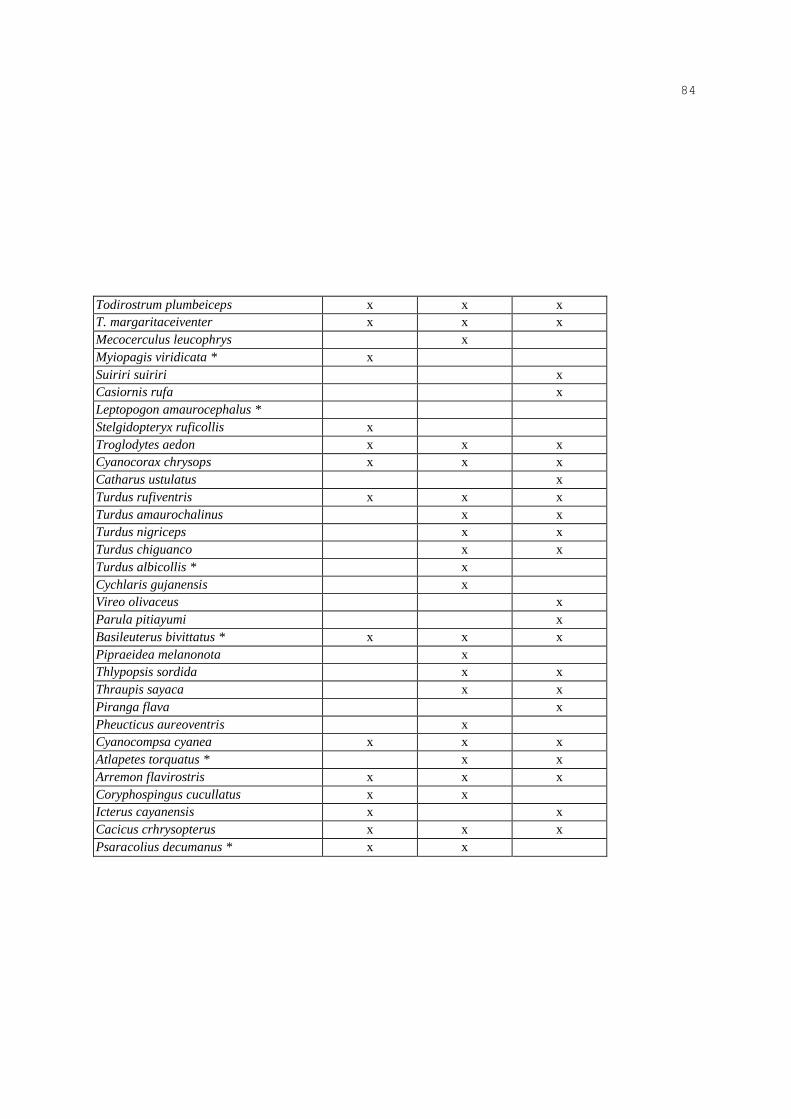
84
Todirostrum plumbeiceps x x x
T. margaritaceiventer x x x
Mecocerculus leucophrys x
Myiopagis viridicata * x
Suiriri suiriri x
Casiornis rufa x
Leptopogon amaurocephalus *
Stelgidopteryx ruficollis x
Troglodytes aedon x x x
Cyanocorax chrysops x x x
Catharus ustulatus x
Turdus rufiventris x x x
Turdus amaurochalinus x x
Turdus nigriceps x x
Turdus chiguanco x x
Turdus albicollis * x
Cychlaris gujanensis x
Vireo olivaceus x
Parula pitiayumi x
Basileuterus bivittatus * x x x
Pipraeidea melanonota x
Thlypopsis sordida x x
Thraupis sayaca x x
Piranga flava x
Pheucticus aureoventris x
Cyanocompsa cyanea x x x
Atlapetes torquatus * x x
Arremon flavirostris x x x
Coryphospingus cucullatus x x
Icterus cayanensis x x
Cacicus crhrysopterus x x x
Psaracolius decumanus * x x

85
Lista 3: Lista de aves del Bosque de Transición de palo blanco de Laguna La Brea, sudeste
de la provincia de Jujuy.
ESPECIE MAYO JULIO NOVIEMBRE
Crypturellus tataupa x x x
Podiceps dominicus x x x
Podilymbus podiceps x
Anhinga anhinga x x
Phalacrocorax olivaceus x x
Egretta alba x
Egretta thula x
Egretta ibis x
Cairina moschata x x
Cathartes aura x
Coragyps atratus x x x
Sarcoramphus papa x
Spizastur melanoleucus x
Accipiter bicolor x x
Buteo magnirostris x x x
Polyborus plancus x x x
Falco rufigularis x
Aramides cajanea x x x
Gallinula chloropus x
Jacana jacana x x x
Columbina picui x x x
Leptoptila verreauxi x x x
Claravis pretiosa x
Ara auricollis x
Pyrrhura molinae x x x
Amazona aestiva x x
Pionus maximiliani x x x
Guira guira x x x
Piaya cayana x x x
Otus choliba x x x
Glaucidium brasilianum x
Pulsatrix perspicillata x
Caprimulgus parvulus x x
Sappho sparganura x
Amazilia chinogaster x x
Trogon curucui x
Ceryle torquata x
Chloroceryle americana x x x
Picumnus cirratus x x x

86
Campephillus lecucopogon x x x
Piculus rubiginosus x
Veniliornis frontalis x x
Sittasomus griseocapillus x x x
Xiphocolaptes major x
Lepidocolaptes angustirostris x x x
Dendrocolaptes picumnus x
Synallaxis frontalis x
Synallaxis scutatus x x x
Certhiaxis pyrrhopia x
Phacellodomus rufifrons x
Philydor rufosuperciliatus x x x
Taraba major x
Thamnophillus caerulescens x x x
Myrmorchilus strigilatus x x x
Herpsilochmus pileatus x
Pachyramphus viridis x
Knipolegus striaticeps x
Knipolegus aterrimus x
Fluvicola pica x
Machetornis rixosa x
Pitangus sulphuratus x
Myiarchus tyrannulus x
Contopus fumigatus x
Tolmomyas sulphurescens x
Todirostrum plumbeiceps x x x
T. margaritaceiventer x x x
Serpophaga nigricans x
Serpophaga munda x
Serpophaga subcristata x x
Mecocerculus leucophrys x x
Elaenia strepera x
Elaenia parvirostris x
Camptostoma obsoletum x x x
Troglodytes aedon x x x
Stelgidopteryx ruficollis x
Cyanocorax chrysops x x x
Catharus dryas x
Turdus rufiventris x x x
Turdus nigriceps x
Turdus amaurochalinus x
Polioptila dumicola x
Cychlaris gujanensis x
Vireo olivaceus x x
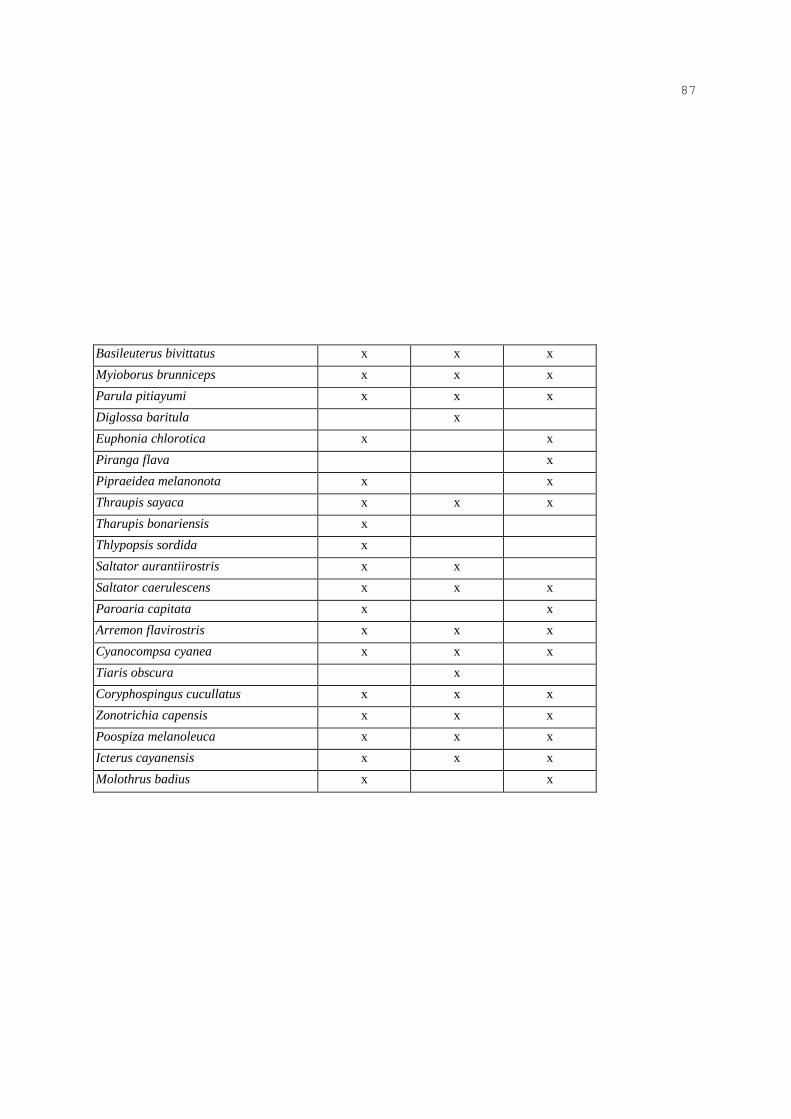
87
Basileuterus bivittatus x x x
Myioborus brunniceps x x x
Parula pitiayumi x x x
Diglossa baritula x
Euphonia chlorotica x x
Piranga flava x
Pipraeidea melanonota x x
Thraupis sayaca x x x
Tharupis bonariensis x
Thlypopsis sordida x
Saltator aurantiirostris x x
Saltator caerulescens x x x
Paroaria capitata x x
Arremon flavirostris x x x
Cyanocompsa cyanea x x x
Tiaris obscura x
Coryphospingus cucullatus x x x
Zonotrichia capensis x x x
Poospiza melanoleuca x x x
Icterus cayanensis x x x
Molothrus badius x x

88
Lista 4: Lista de aves del Bosque de Transición de la provincia de Tucumán.
Crypturellus t. tataupa (Temminck) inambú del monte
Phalacrocorax o. olivaceus (Humboldt) biguá
Nycticorax nycticorax hoactli (Gmelin) garza bruja o cuaco
Tigrisoma fasciatum pallescens Olrog hocó oscuro
Merganetta armata berlepschi Hartert pato de los torrentes
Cathartes aura jota (Molina) jote de cabeza roja
Coragyps atratus (Beckstein) jote de cabeza negra
Sarcoramphus papa (Linné) jote o cóndor real
Buteo magnirostris saturatus (Sclater y Salvin) garganchillo
Buteogallus u. urubitinga (Vieillot) águila negra
Micrastur ruficollis olrogi Amadon halcón de cuello rojo
Polyborus plancus brasiliensis (Gmelin) carancho
Penelope obscura bridgesi G.R. Gray pava de monte
Aramides c. cajanea (P.L.S.Muller) chiricote
Leptoptila verreauxi decipiens (Salvadori) bumbuna
Aratinga m. mitrata (Tschudi) cotorra de cara roja
Pionus maximiliani lacerus (Heine) loro choclero
Amazona tucumana (Cabanis) loro alisero
Amazona aestiva xanthopteryx (Berlepsch) loro hablador
Piaya cayana mogenseni Peters alma de gato
Guira guira Gmelin machilo o pirincho
Tyto alba tuidara (J.E. Gray) lechuza de campanario
Otus c. choliba (Vieillot) lechucita común
Aegolius harrisi dabbenei Olrog lechucita acanelada
Chlorostilbon a. aureoventris (D Orbignyi y Lafresnaye) picaflor verde común
Leucippus chinogaster hypoleucus (Gould) picaflor de vientre blanco
Eriocnemis glaucopoides (D Orbignyi y Lafresnaye) picaflor de frente violeta
Sappho sparganura sappho (Lesson) picaflor coludo rojo
Ceryle t. torquata (Linné) martín pescador grande
Chloroceryle a. amazona (Latham) martín pescador mediano
Chloroceryle americana mathewsi Laubmann martín pescador chico
Picumnus cirratus thamnophiloides Bond y Schauensee carpinterito común
Piculus rubiginosus tucumanus Cabanis carpintero dorado verde
Campephilus leucopogon major (Olrog) carpintero negro dorso crema
Xiphocolaptes m. major (Vieillot) trepador grande colorado
Sittasomus g. griseocapillus (Vieillot) trepadorcito común
Cinclodes fuscus tucumanus Chapman remolinera parda
Cinclodes a. atacamensis (Philippi) remolinera grande
Synallaxis s. superciliosa Cabanis pijuí de ceja canela
Synallaxis f. frontalis Pelzeln pijuí de frente gris
Phacellodomus rufifrons sincipitalis Cabanis espinero de frente rufa
Philydor rufosuperciliatus oleaginea (Sclater) titirí
Thamnophilus ruficapillus cochabambae (Chapman) bataracito pardo
Thamnophilus caerulescens connectens Berlepsch bataracito común
Ochthoeca leucophrys tucumana Berlepsch pitajo gris
Tyrannus m. melancholicus (Vieillot) pitirre real
Sayornis nigricans latirostris Cabanis pitajo negruzco
Knipolegus a.aterrimus Kaup viudita negruzca
Knipolegus cabanisi Schultz viudita plomiza
Knipolegus striaticeps viudita gris
Empidonomus v. varius (Vieillot) tuquito manchado
Myiodynastes maculatus solitarius (Vieillot) pitirre rayado

89
Pitangus sulphuratus bolivianus (Lafresnaye) ketupí
Myiarchus t. tyrannulus (P.L.S. Muller) burlisto cola castaña
Myiophobus fasciatus flammiceps (Temminck) mosqueta de pecho rayado
Phylloscartes ventralis tucumanus Zimmer mosquetita verde
Hemitriccus m. margaritaceiventer (D Orbignyi y Lafresnaye) titirijí común
Mecocerculus l. leucophrys (D Orbignyi y Lafresnaye) piojito arbustero
Elaenia a. albiceps (D Orbignyi y Lafresnaye) fiofío común
Elaenia parvirostris (Pelzeln) fiofío de pico corto
Elaenia strepera Cabanis fiofío plomizo
Elaenia o. obscura (D Orbignyi et Lafresnaye) fiofío grande
Serpophaga subcristata Vieillot piojito de vientre amarillo
Serpophaga munda Berlepsch piojito de vientre blanco
Serpophaga nigricans (Vieillot) piojito plomizo
Troglodytes aedon rex Berlepsch y Leverkuhn carrasquita
Troglodytes solstitialis auricularis Cabanis carrasquita de ceja blanca
Notiochelidon c. cyanoleuca (Vieillot) golondrina barranquera
Cyanocorax chrysops tucumanus Cabanis urraca
Catharus ustulatus swainsoni (Tschudi) zorzalito norteamericano
Turdus n. nigriceps Cabanis zorzal plomizo
Turdus r. rufiventris Vieillot zorzal pechocolorado
Turdus amaurochalinus Cabanis chalchalero
Cychlarhys gujanensis viridis (Vieillot) juan chiviro
Vireo olivaceus chivi Tschudi chiví
Parula pitiayumi (Vieillot) pitiayumí
Geothlypis aequinoctialis (D Orbignyi) arañero de cara negra
Myioborus b. brunniceps (D Orbignyi et Lafresnaye) arañero cabeza castaña
Basileuterus culicivorus viridescens Tood arañero chico
Euphonia chlorotica serriorostris D Orbignyi el Lafresnaye curruñata común
Pipraeidea melanonota venezuelensis (Sclater) viuva
Thraupis sayaca obscura Naumburg celestino
Thraupis bonariensis schulzei Brodkorb naranjero
Piranga flava Vieillot fueguero
Thlypopsis s. sordida (D Orbignyi et Lafresnaye) pioró cabeza amarilla
Chlorospingus ophthalmicus argentinus Hellmayr fruterito ojo blanco
Saltator c. caerulescens Vieillot pepitero plomizo
Saltator a. aurantiirostris Vieillot pepitero de collar
Pheucticus aureoventris (D Orbignyi et Lafresnaye) rey del bosque
Cyanocompsa cyanea argentina (Sharpe) reina mora
Sporophila c. caerulescens (Vieillot) corbatita doble collar
Coryphospingus cucullatus fargoi Brodkorb brasita de fuego
Atlapetes citrinellus (Cabanis) arbustero oliváceo
Arremon flavirostris dorbignii Sclater gran cantor
Zonotrichia capensis pulacayensis (Ménégaux) chingolo
Poospiza melanoleuca (D Orbignyi et Lafresnaye) monterita común
Poospiza nigrorufa whitei Sclater monterita castaña
Poospiza e. erythrophrys Sclater monterita alisera
Carduelis magellanica tucumana (Todd) cabecita negra común
Molothrus b. bonariensis (Gmelin) tordo común
Molothrus b. badius (Vieillot) tordo músico
Icterus cayanensis pyrrhopterus Vieillot toche de cobija canela
Cacicus chrysopterus (Vigors) boyero de ala amarilla

90
LISTA 6. Especies Residentes del Bosque Pedemonano de Transición
1. Crypturellus tataupa
2. Phalacrocorax olivaceus
3. Tigrisoma fasciatum
4. Sarcoramphus papa
5. Cathartes aura
6. Coragyps atratus
7. Micrastur ruficollis
8. Micrastur semitorquatus
9. Buteo magnirostris
10. Falco sparverius
11. Polyborus plancus
12. Penelope obscura
13. Aramides cajanea
14. Columbina picui
15. Leptoptila verreauxi
16. Pyrrhura molinae
17. Pionus maximiliani
18. Piaya cayana
19. Guira guira
20. Phaethornis pretrei
21. Chlorostilbon aureoventris
22. Amazilia chionogaster
23. Eriocnemis glaucopoides
24. Otus choliba
25. Caprimulgus parvulus
26. Trogon curucui
27. Chloroceryle americana
28. Ceryle torquata
29. Ramphastos toco
30. Picumnus dorbignianus
31. Picumnus cirratus
32. Campephilus leucopogon
33. Veniliornis frontalis
34. Dendrocolaptes picumnus
35. Lepidocolaptes angustirostris
36. Sittasomus griseocapillus
37. Phacellodomus rufifrons
38. Synallaxis scutatus
39. Synallaxis superciliosa
40. Philydor rufosuperciliatus
41. Thamnophilus caerulescens
42. Thamnophilus ruficapillus
43. Mirmorchilus strigilatus
44. Sayornis nigricans
45. Hirundinea ferruginea
46. Pitangus sulphuratus
47. Phylloscartes ventralis
48. Todirostrum plumbeiceps
49. Todirostrum margaritaceiventer
50. Serpophaga munda
51. Camptostoma obsoletum
52. Troglodytes aedon
53. Troglodytes solstitialis
54. Notiochelidon cyanoleuca
55. Cyanocorax chrysops
56. Turdus amaurochalinus
57. Turdus nigriceps
58. Turdus rufiventris
59. Vireo olivaceus
60. Basileuterus bivittatus
61. Basileuterus culicivorus
62. Myoborus brunniceps
63. Parula pitiayumi
64. Euphonia chlorotica
65. Thraupis sayaca
66. Thlypopsis sordida
67. Pipraeidea melanonota
68. chlorospingus ophthalmicus
69. Saltator caerulescens
70. Saltator aurantiirostris
71. Cyanocompsa cyanea
72. Arremon flavirostris
73. Atlapetes torquatus
74. Paroaria capitata
75. Coryphospingus cucullatus
76. Zonotrichia capensis
77. Poospiza melanoleuca
78. Poospiza erythrophrys
79. Carduelis magellanica
80. Molothrus badius
81. Molothrus bonariensis
82. Icterus cayanensis
83. Cacicus chrysopterus
84. Psaracolius decumanus

91





























