Embed Size (px)
Citation preview

LA IMPORTANCIA DE LA DISPERSIÓN DE SEMILLAS EN LA
RECUPERACIÓN DEL BOSQUE MESÓFILO DE MONTAÑA
DEL CENTRO DE VERACRUZ, MÉXICO
TESIS QUE PRESENTA STÉPHANIE BALTAZAR HERNÁNDEZ
PARA OBTENER EL GRADO DE MAESTRA EN CIENCIAS
Xalapa, Veracruz, México 2014

INSTITUTO DE ECOLOGIA, A.C.
Aprobación final del documento de tesis de grado:
"La importancia de la dispersión de semillas en la recuperación del bosque mesófilo de
montaña del centro de Veracruz, México"
Nombre
Codirección Dr. Octavio R. Rojas Soto
Dra. Martha Bonilla Moheno
Comité Tutorial Dra. Fabiola López Barrera
Dr. Fernando Puebla Olivares
Jurado Dr. Francisco Javier Laborde Dovalí
Dr. Jorge Rodrigo Galindo González --=-'7f'!=~IIiH~=----'--
, , ."..

RECONOCIMIENTOS
A CONACyT por otorgarme la beca para realizar mis estudios de Maestría.
Al Instituto de Ecología A. C. por darme la oportunidad de cursar el posgrado.
A mis directores de tesis Dr. Octavio R. Rojas Soto y Dra. Martha Bonilla Moheno, por su apoyo,
enseñanzas y consejos a lo largo de todo el proyecto de tesis.
A los integrantes de mi comité Dra. Fabiola López Barrera y Dr. Fernando Puebla Olivares por
sus aportaciones y sugerencias a la tesis.
A los investigadores y técnico Dr. Gonzalo Castillo, Dr. Diego Santiago Alarcón, Dr. Antonio
Guillén, Javier Tolome y Dr. Vinicio Sosa por su apoyo en el trabajo de campo, laboratorio, la
identificación de ejemplares y préstamo de la literatura.
A los miembros de mi jurado de examen Dr. Javier Laborde, Dr. Jorge Galindo, por sus valiosos
comentarios y observaciones.
A la Dra. Guadalupe Williams por proporcionar el material y facilitar la elaboración de las
trampas de semillas.
A los investigadores Dr. Jairo Pérez, Dr. Armando Aguirre, Dra. Cecilia Díaz, Dra. Simoneta
Negrete y Dr. Victor Steinmann por sus útiles enseñanzas.
A mis compañeros M. en C. Karla Parra, Biól. Patricia Tellez, M. en C. Claudio Mota, M. en C.
Diana Vázquez, Biól. Natali Novelo y M. en C. Yocupitzia Ramírez, quienes me apoyaron con el
trabajo de campo y laboratorio.
A mis colegas M. en C. Cristian Martínez y Biól. Pedro Maldonado por su apoyo en estadística y
redacción.
A mis compañeros Ecol. Carolina Hernández, Biól. Carolina Solís, M. en C. Angelina Ruiz, M.
en C. Rafael Rueda, M. en C. Omar Hernández, Biól. Jorge Cortinas, M. en C. Omar Suarez,
Biól. Luis Huerta, Biól. Edel Atondo y Petra Galindo quienes me acompañaron a las salidas de
campo.
A los administrativos Enrique Salinas, Mónica Enríquez, Ema Gómez y Bertha Ulloa por su
apoyo en cada tutoral y los trámites de titulación.
A mis compañeros de laboratorio Biól. Norma Hernández, M. en C. Mauricio Ortega y M. en C.
David Prieto por su ayuda y consejos.
A mis amigos Adriana Arguelles, Priscila Bocanegra y Edwin Batista por su apoyo a lo largo de
todo el trabajo de tesis.

DEDICATORIA
A mis padres Carlos Baltazar Sánchez y Cintia Josefina Hernández Moreno, por apoyarme en
todos mis proyectos. Ustedes son mi sabiduría y fortaleza.
A mi hermana Frida Baltazar Hernández, por sus consejos tan objetivos, oportunos y sinceros.
Tú eres mi coraje y templanza.
A la familia Gutiérrez Jiménez por brindarme su apoyo y cariño incondicional como a un
miembro más de su familia. Ustedes son mi balance.

DECLARACiÓN
Excepto cuando es explícitamente indicado en el texto, el trabajo de investigación
contenido en esta tesis fue efectuado por Stéphanie Baltazar Hernández como estudiante
de la carrera de Maestra en Ciencias entre agosto de 2012 y agosto del 2014, bajo la
supervisión de Dr. Octavio R. Rojas Soto y Dra. Martha Bonilla Moheno.
Las investigaciones reportadas en esta tesis no han sido utilizadas anteriormente
Candidato: Stéphanie Baltazar Hernández
Director de tesis: Dr. Octavio R. Rojas Soto
Dra. Martha Bonilla Moheno
para obtener otros grados académicos, ni serán utilizadas para tales fines en el futuro.

11
ÍNDICE
LISTA DE FIGURAS, CUADROS Y ANEXOS .......................................................................... 13
FIGURAS .................................................................................................................................. 13
CUADROS ................................................................................................................................ 15
ANEXOS ................................................................................................................................... 17
RESUMEN ................................................................................................................................ 19
I. INTRODUCCIÓN GENERAL .............................................................................................. 21
II. EL PAPEL DE LAS AVES DISPERSORAS DE SEMILLAS EN LA RESTAURACIÓN
DEL BOSQUE MESÓFILO DE MONTAÑA EN UN POTRERO ABANDONADO ................ 27
RESUMEN ................................................................................................................................ 29
INTRODUCCIÓN .................................................................................................................... 31
MÉTODOS ............................................................................................................................... 33
RESULTADOS ......................................................................................................................... 35
DISCUSIÓN .............................................................................................................................. 37
CUADROS Y FIGURAS ......................................................................................................... 43
ANEXOS ................................................................................................................................... 55
III. LA LLUVIA DE SEMILLAS COMO CATALIZADORA DE LA RESTAURACIÓN
DEL BOSQUE MESÓFILO DE MONTAÑA .............................................................................. 57
RESUMEN ................................................................................................................................ 59
INTRODUCCIÓN .................................................................................................................... 61
MÉTODOS ............................................................................................................................... 63
RESULTADOS ......................................................................................................................... 66
DISCUSIÓN .............................................................................................................................. 69
CUADROS Y FIGURAS ......................................................................................................... 73
ANEXOS ................................................................................................................................... 87
IV. CONCLUSIONES GENERALES ..................................................................................... 91
LITERATURA ......................................................................................................................... 95

13
LISTA DE FIGURAS, CUADROS Y ANEXOS
FIGURAS
Figura 1. Área de estudio. En el mapa se muestra el potrero delimitado por el polígono negro
ubicado al sureste de la ciudad de Xalapa en la zona centro del estado de Veracruz. 43
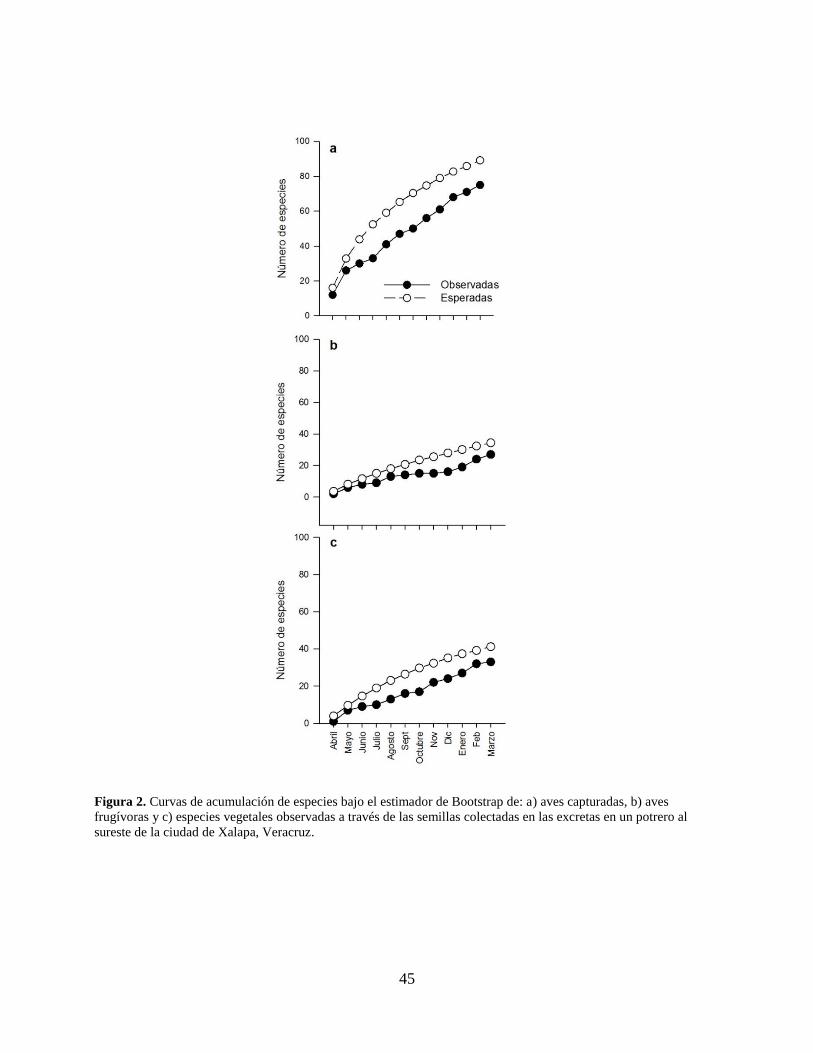
Figura 2. Curvas de acumulación de especies bajo el estimador de Bootstrap de: a) aves
capturadas, b) aves frugívoras y c) especies vegetales observadas a través de las
semillas colectadas en las excretas en un potrero al sureste de la ciudad de Xalapa,
Veracruz. ..................................................................................................................... 45
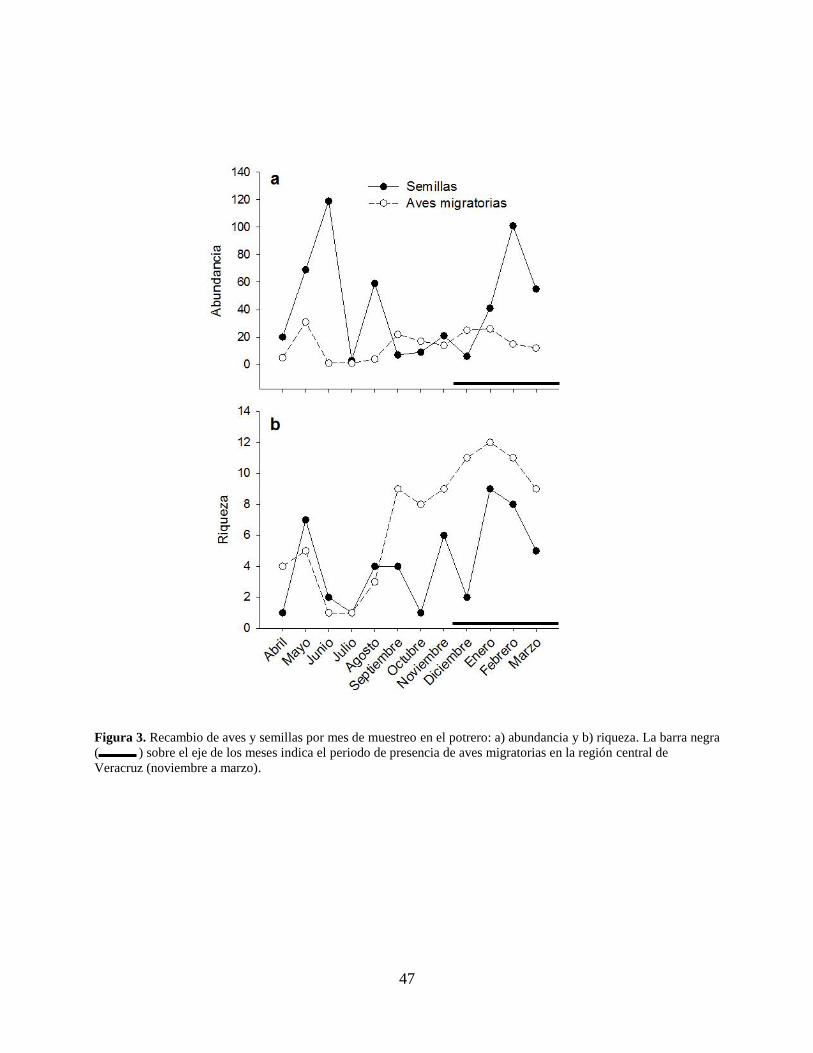
Figura 3. Recambio de aves y semillas por mes de muestreo en el potrero: a) abundancia y b)
riqueza. La barra negra ( ) sobre el eje de los meses indica el periodo de
presencia de aves migratorias en la región central de Veracruz (noviembre a marzo).
..................................................................................................................................... 47
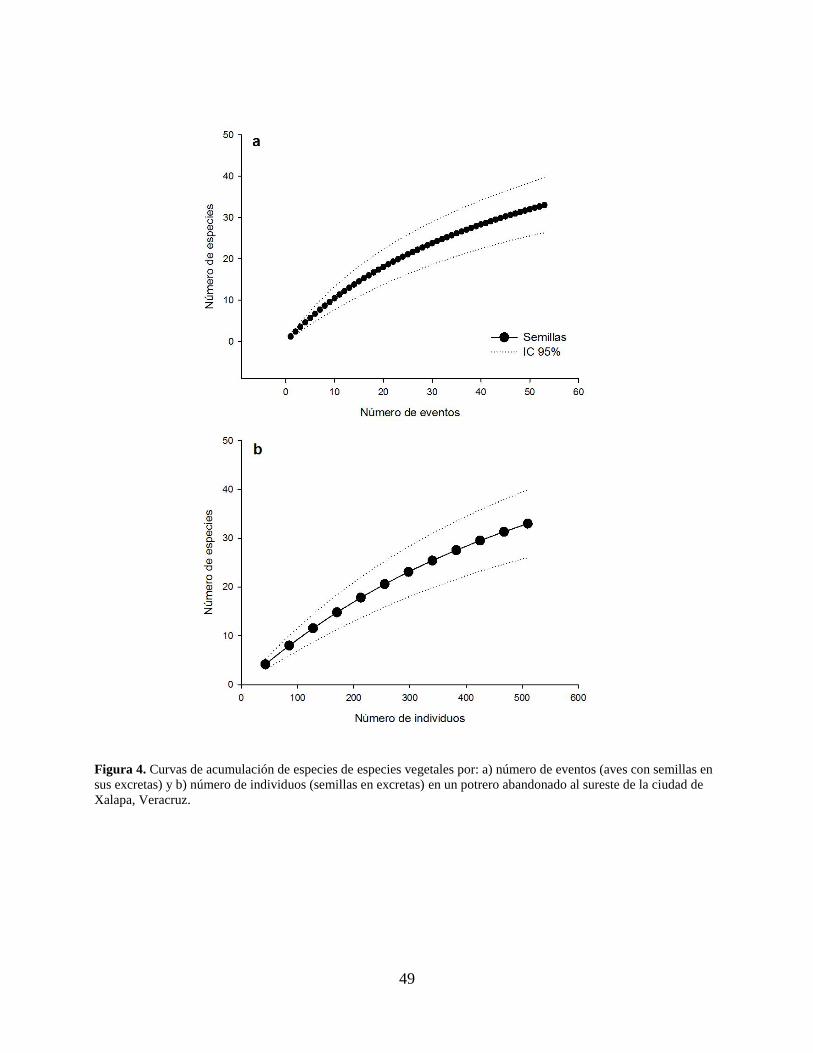
Figura 4. Curvas de acumulación de especies de especies vegetales por: a) número de eventos
(aves con semillas en sus excretas) y b) número de individuos (semillas en excretas)
en un potrero abandonado al sureste de la ciudad de Xalapa, Veracruz. .................... 49
Figura 5. Área de estudio. En el mapa se muestra el potrero (polígono con trama en diagonal) y el
BMM (polígono con trama en horizontal) ubicados al sureste de la ciudad de Xalapa
en la zona centro del estado de Veracruz. ................................................................... 73
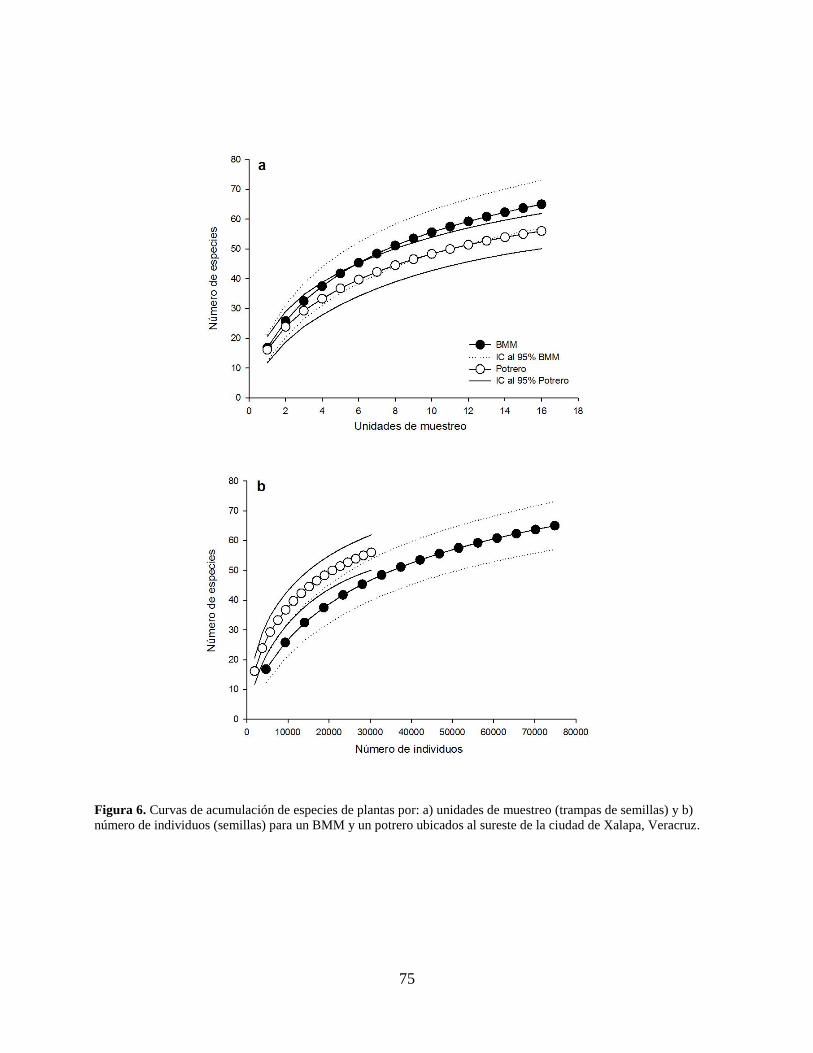
Figura 6. Curvas de acumulación de especies de plantas por: a) unidades de muestreo (trampas de
semillas) y b) número de individuos (semillas) para un BMM y un potrero ubicados al
sureste de la ciudad de Xalapa, Veracruz. ................................................................... 75
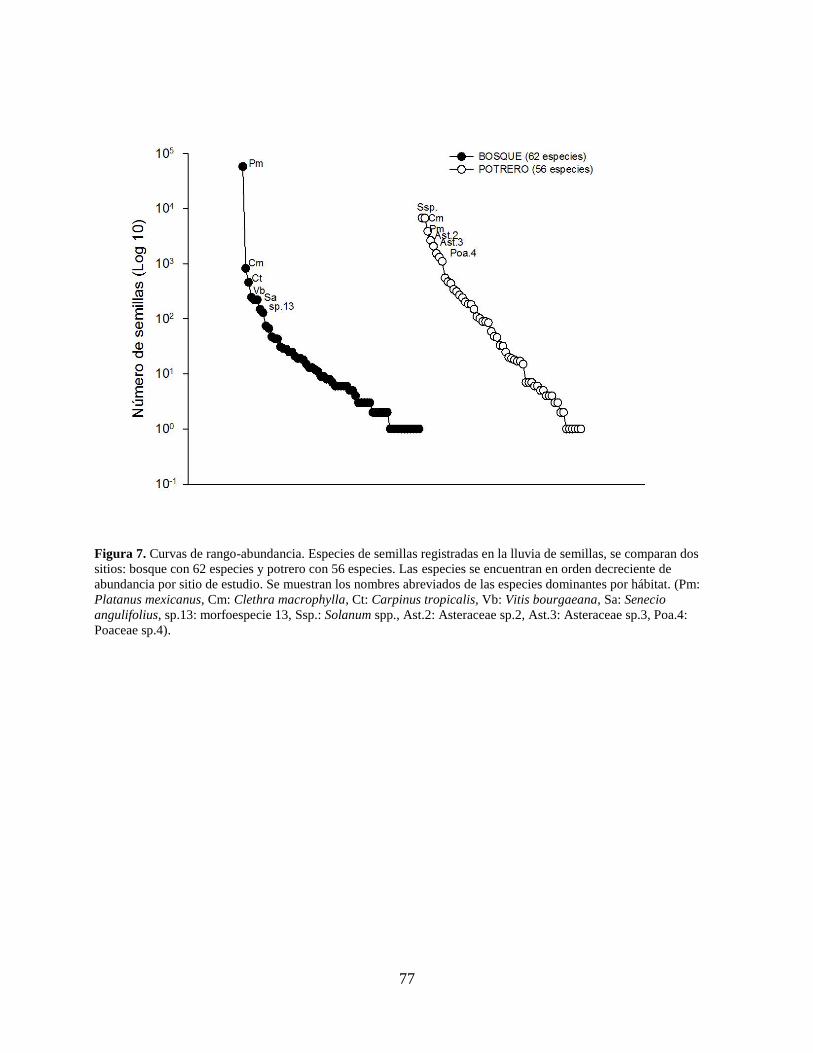
Figura 7. Curvas de rango-abundancia. Especies de semillas registradas en la lluvia de semillas,
se comparan dos sitios: bosque con 62 especies y potrero con 56 especies. Las
especies se encuentran en orden decreciente de abundancia por sitio de estudio. Se
muestran los nombres abreviados de las especies dominantes por hábitat. (Pm:
Platanus mexicanus, Cm: Clethra macrophylla, Ct: Carpinus tropicalis, Vb: Vitis
bourgaeana, Sa: Senecio angulifolius, sp.13: morfoespecie 13, Ssp.: Solanum spp.,
Ast.2: Asteraceae sp.2, Ast.3: Asteraceae sp.3, Poa.4: Poaceae sp.4). ....................... 77
Figura 8. Abundancia mensual de semillas por síndrome de dispersión: a) anemócoras, b)
zoócoras y c) barócoras. Las líneas punteadas representan la abundancia de semillas

14
del potrero, mientras que las líneas sólidas representan la abundancia de semillas en
el BMM. ...................................................................................................................... 79
Figura 9. Abundancia mensual de semillas por hábito de crecimiento: a) pastos, b) herbáceas, c)
leñosas. Las líneas punteadas representan la abundancia de semillas del potrero,
mientras que las líneas sólidas representan la abundancia de semillas en el BMM. ... 81
Figura 10. Abundancia mensual de semillas por etapa sucesional: a) pioneras, b) intermedias, c)
tardías. Las líneas punteadas representan la abundancia de semillas del potrero,
mientras que las líneas sólidas representan la abundancia de semillas en el BMM. ... 83

15
CUADROS
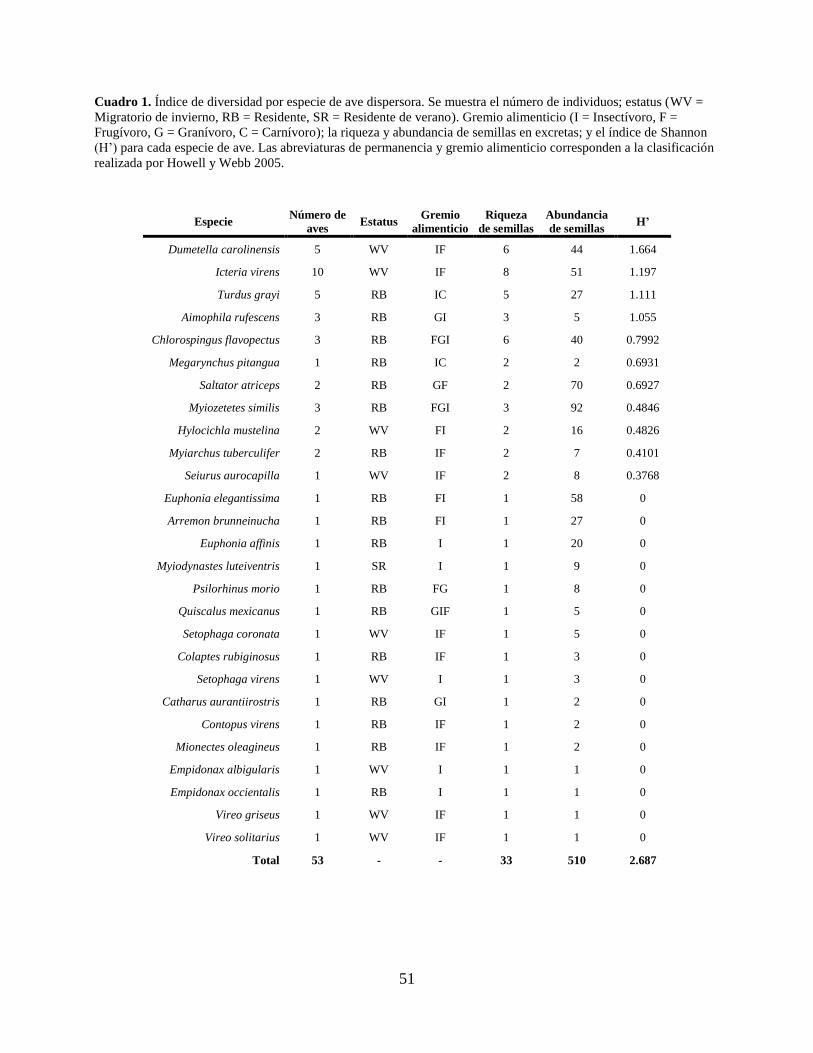
Cuadro 1. Índice de diversidad por especie de ave dispersora. Se muestra el número de
individuos; estatus (WV = Migratorio de invierno, RB = Residente, SR = Residente
de verano). Gremio alimenticio (I = Insectívoro, F = Frugívoro, G = Granívoro, C =
Carnívoro); la riqueza y abundancia de semillas en excretas; y el índice de Shannon
(H’) para cada especie de ave. Las abreviaturas de permanencia y gremio alimenticio
corresponden a la clasificación realizada por Howell y Webb 2005. ......................... 51
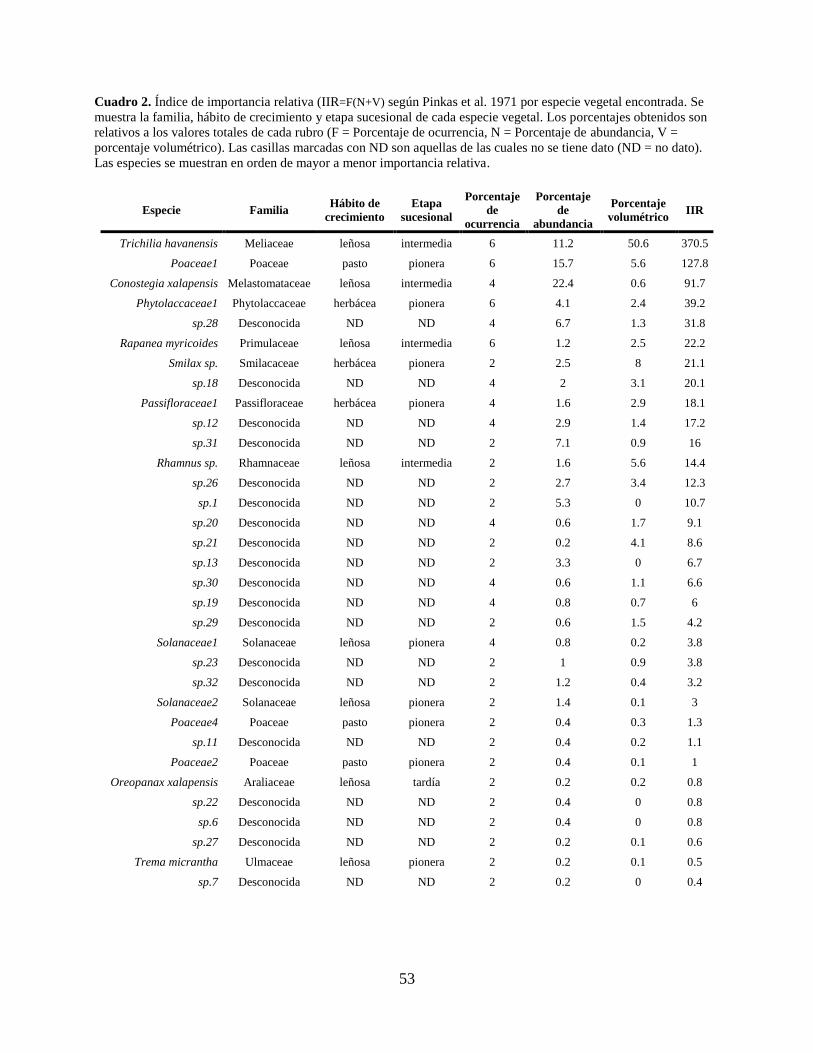
Cuadro 2. Índice de importancia relativa (IIR=F(N+V) según Pinkas et al. 1971 por especie
vegetal encontrada. Se muestra la familia, hábito de crecimiento y etapa sucesional de
cada especie vegetal. Los porcentajes obtenidos son relativos a los valores totales de
cada rubro (F = Porcentaje de ocurrencia, N = Porcentaje de abundancia, V =
porcentaje volumétrico). Las casillas marcadas con ND son aquellas de las cuales no
se tiene dato (ND = no dato). Las especies se muestran en orden de mayor a menor
importancia relativa. .................................................................................................... 53
Cuadro 3. Número promedio de semillas por trampa (± DE), número de especies y valores del
índice Chao-Jaccard en el BMM y el potrero; el número promedio de semillas por
trampa. Los asteriscos en los nombres de las categorías indican los resultados que
tuvieron diferencias significativas en la prueba de U de Mann-Whitney (P<0.05). ... 85

17
ANEXOS
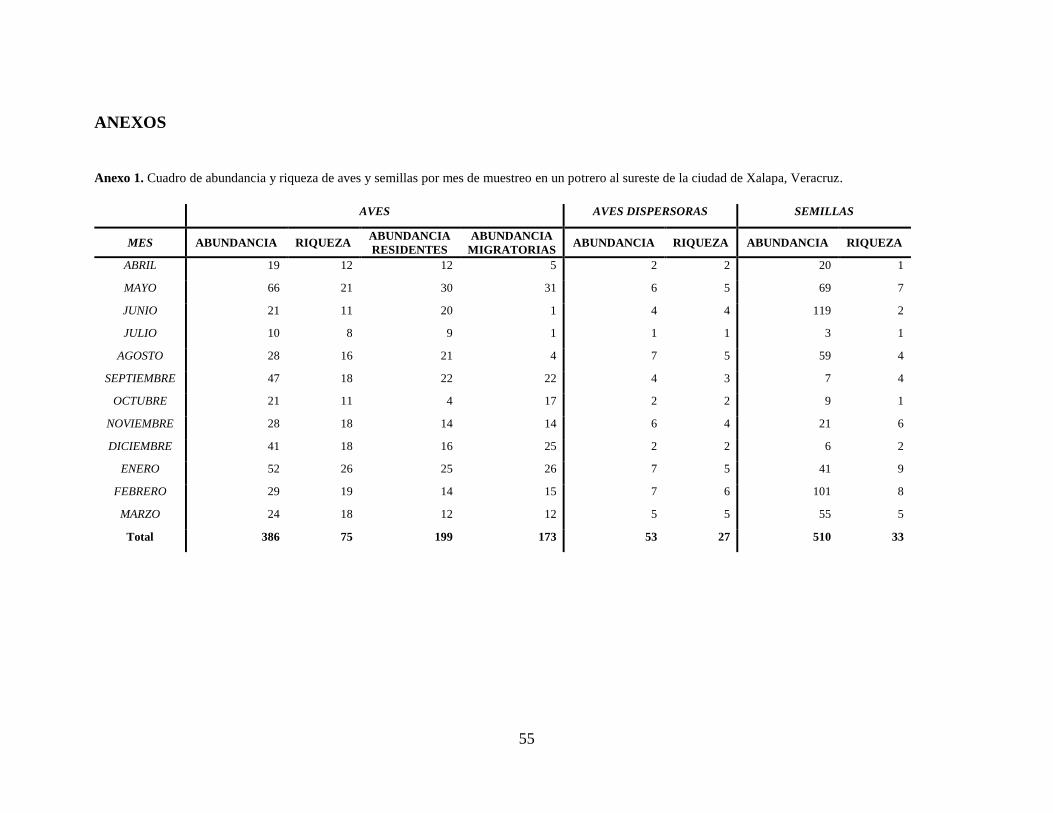
Anexo 1. Cuadro de abundancia y riqueza de aves y semillas por mes de muestreo en un potrero
al sureste de la ciudad de Xalapa, Veracruz. ............................................................... 55
Anexo 2. Abundancia total de especies registradas en lluvia de semillas en cada sitio de muestreo.
Se muestran también la clasificación por hábito de crecimiento, etapa sucesional
(Rzedowski 1978; Castillo-Campos y Medina-Abreo 2005; Rodríguez-Velázquez et
al. 2009), y síndrome de dispersión (Niembro et al. 2010; Hernández-Ladrón de
Guevara et al. 2012; características morfológicas) (NA= No Aplica y ND= No Dato).
..................................................................................................................................... 87

19
RESUMEN
La transformación del bosque mesófilo de montaña (BMM) a otros tipos de cobertura de suelo ha
causado una gran pérdida de su área original. Ante la necesidad de revertir este proceso, es
importante conocer aquellas interacciones que permiten la restauración de este ecosistema, como
lo es la dispersión de semillas. En el presente trabajo (1) se caracterizó la comunidad de aves
dispersoras y las especies de plantas que dispersan en un potrero y (2) se comparó la lluvia de
semillas del potrero con la de un fragmento de BMM aledaño. La captura de aves se realizó con
redes de niebla a lo largo de un año. La colecta de lluvia de semillas se realizó con colectores
cónicos durante nueve meses. Las especies vegetales se clasificaron por síndrome de dispersión,
hábito de crecimiento y etapa sucesional. Se registraron 27 especies de aves que dispersaron 33
especies vegetales. Dumetella carolinensis fue la especie ave que mayor diversidad de semillas
dispersó (H’=1.66), mientras que Trichilia havanensis fue la especie vegetal con el índice de
importancia relativa más alto (IIR=370.5). También se identificaron 81 especies de plantas en la
lluvia de semillas; Platanus mexicana fue la más abundante para ambos sitios. Se encontró una
alta similitud de composición de especies con dispersión anemócora (Ĵabd=0.919), así como de
plantas leñosas (Ĵabd=0.981) y de sucesión tardía (Ĵabd=0.996) entre los dos sitios. Los resultados
de abundancia mostraron diferencias significativas entre ambos sitios, así como para las especies
zoócoras, las plantas herbáceas, pastos, las especies pioneras, intermedia y tardía. Los resultados
sugieren que las aves migratorias juegan un papel importante en el proceso de dispersión de
semillas y que, dadas las características de la lluvia de semillas, el potrero tiene un alto potencial
de recuperar la composición vegetal propia de un BMM.

21
I. INTRODUCCIÓN GENERAL

23
INTRODUCCIÓN GENERAL
Los bosques mesófilos de montaña (BMM) son considerados centros de biodiversidad (Williams-
Linera et al. 2002; Manson 2004); a pesar de ello, son los que presentan la tasa de deforestación
más alta a nivel mundial de entre todos los tipos de bosques (Williams-Linera et al. 2002;
Manson 2004; Martínez et al. 2009; González-Espinoza et al. 2012). En la actualidad, casi la
mitad de la superficie del BMM de México ha sido transformada por actividades antrópicas
(Challenger 1998; Toledo-Aceves et al. 2011; González-Espinoza et al. 2012). Aún con el nivel
de transformación que ha sufrido el sistema, existen procesos que favorecerían su recuperación,
por ejemplo, las diversas estrategias de colonización empleadas por las especies vegetales (Howe
y Smallwood 1982), cuyo alcance puede ser estudiado a través de la dispersión de semillas.
La dispersión de semillas es un proceso ecológico importante para el mantenimiento y la
recuperación de los ecosistemas (Howe y Smallwood 1982; Wang y Smith 2002; Ingle 2003,
Laborde et al. 2008), ya que éste completa el ciclo reproductivo de las plantas adultas a través del
establecimiento de la progenie (Wang y Smith 2002) cuando este sucede. En numerosos casos, la
dispersión de semillas es a corta distancia de las plantas progenitoras, lo que podría dificultar el
establecimiento; sin embargo, las plantas han desarrollado estrategias que compensan su falta de
movilidad y promueven la recuperación de sus poblaciones (Janzen 1970; Connell et al. 1984) en
áreas que han sido perturbadas.
Cuando un ecosistema ha sido perturbado, la recuperación puede realizarse por medio de
su regeneración natural, en donde las fuentes de propágulos son un elemento de vital importancia
(Gubb 1977; Purata 1986; Aide et al. 1996; Aide et al. 2000; Holl et al. 2000). En este sentido, la
presencia y tipo de dispersores influirá en el proceso de colonización de las plantas (Ramírez-
Marcial et al. 1992; Wang y Smith 2002) y por lo tanto, tendrá implicaciones para la
recuperación del sistema. La dispersión de semillas por animales es una de las estrategias más

24
efectivas, sobre todo aquella realizada por vertebrados voladores (aves y murciélagos) (Gorchov
et al. 1995; Galindo-González et al. 2000).
Debido a que los frutos forman parte importante de la dieta de algunas especies de aves y
murciélagos, estos dos grupos de vertebrados voladores son importantes movilizadores de
semillas de las especies de plantas con frutos carnosos en ecosistemas tropicales y templados
(Howe 1977; Snow 1981; Howe 1984a; Guevara et al. 1986; Wunderle 1997; Corlett 1998;
Ortiz-Pulido et al. 2000; Holl 2002; Cole et al. 2010). Un aspecto importante de la dieta de estos
vertebrados frugívoros es que las especies que forma parte de la dieta cotidiana de un grupo, no
necesariamente forman parte de la dieta del otro o solo ocurren de forma ocasional (Palmeirim et
al. 1989), por lo que el papel como dispersores suele ser complementario entre ambos grupos
(Gorchov et al. 1995; Galindo-González et al. 2000, Vásquez-Balbuena 2014).
También el comportamiento es un aspecto a considerar, ya que los murciélagos son
animales que defecan al vuelo, mientras que las aves suelen defecar o regurgitar las semillas
mientras perchan; esta diferencia en la deposición de las semillas provoca que las semillas
dispersadas por murciélagos caigan mayormente en lugares abiertos, mientras que aquellas que
son dispersadas por las aves terminen a la sombra de alguna planta y por tanto en un microclima
con distintas condiciones (Gorchov et al. 1993). Es por lo anterior, que los vertebrados voladores
desempeñan un papel importante y complementario en los procesos de sucesión y la capacidad de
resilencia de los sistemas naturales (Gálvez 2002; Amico y Aizen 2005; Rodríguez-Santamaría et
al. 2006; Reid et al. 2014).
Sin embargo, a pesar de la importancia de este proceso en la regeneración de ecosistemas,
la mayoría de los estudios realizados sobre la dispersión de semillas se han realizado en bosques
tropicales de zonas bajas (Guevara et al. 1992; Ramírez-Marcial et al. 1992; Holl 1999; Galindo-
González et al. 2000; Cubiña y Aide 2001; Rodríguez-Santamaría et al. 2006), prestando poca

25
atención a otros ecosistemas como el BMM. El BMM de la región central de Veracruz cuenta con
una alta diversidad y un importante número de especies endémicas con respecto a su área
(Rzedowski 2006; Williams-Linera 2002; Cruz et al. 2010). Por tanto, el estudio de la dispersión
de semillas permitirá entender uno de los procesos fundamentales que operan en la colonización
de especies y la recuperación de este ecosistema (Ramírez-Marcial et al. 1992; Wang y Smith
2002; Rodríguez-Santamaría et al. 2006). Por ello, el presente trabajo se llevó a cabo con la
finalidad de caracterizar la riqueza y abundancia de la comunidad de aves y de las semillas que
dispersan en un potrero, así como analizar y comparar la lluvia de semillas entre el potrero y un
fragmento de BMM aledaño.

27
II. EL PAPEL DE LAS AVES DISPERSORAS DE SEMILLAS EN LA
RESTAURACIÓN DEL BOSQUE MESÓFILO DE MONTAÑA EN UN
POTRERO ABANDONADO

29
EL PAPEL DE LAS AVES DISPERSORAS DE SEMILLAS EN LA
RESTAURACIÓN DEL BOSQUE MESÓFILO DE MONTAÑA EN UN
POTRERO ABANDONADO
RESUMEN
En la región central de Veracruz, el Bosque Mesófilo de Montaña (BMM) ha disminuido su
extensión de forma considerable en las últimas décadas, por lo que es fundamental entender los
procesos ecológicos que modulan la recuperación de este ecosistema, como la dispersión de
semillas por vertebrados (zoocoría). El objetivo de este trabajo fue caracterizar la composición de
la comunidad de aves dispersoras de semillas, así como la riqueza y abundancia de las semillas
que dispersan dentro de un potrero abandonado. Para ello, se realizaron capturas mensuales con
redes de niebla, identificando tanto el recambio estacional de las especies de aves, como el de las
semillas dispersadas. Se identificaron 27 especies de aves que dispersaron 33 especies vegetales,
siendo Dumetella carolinensis el ave con mayor diversidad de semillas dispersadas (H’=1.66).
Mientras que la especie de planta con el mayor Índice de Importancia Relativa fue Trichilia
havanensis (IIR=370.5). La mayor riqueza de especies de semillas se observó durante enero (9
especies) y febrero (8 especies), dispersadas principalmente por aves migratorias. Por lo que las
especies de aves migratorias parecen tener un papel importante en la dispersión de semillas y por
tanto en la recuperación del BMM.
Palabras clave: recuperación, recambio estacional, aves migratorias.

31
INTRODUCCIÓN
El bosque mesófilo de montaña (BMM) es un ecosistema sumamente amenazado por actividades
humanas, con la tasa de deforestación más alta entre todos los tipos de bosques a escala global
(Doumenge et al. 1995; Aldrich et al. 2000; Williams-Linera et al. 2002; Toledo-Aceves 2011), a
pesar de su importancia como centro de diversidad y por la cantidad de servicios ambientales que
provee (Williams-Linera et al. 2002; Manson 2004; Martínez et al. 2009; González-Espinosa et
al. 2012). En el BMM de la región central de Veracruz, se ha encontrado una alta diversidad y un
importante número de especies endémicas (Williams-Linera 2002; Rzedowski 2006; Cruz et al.
2010). Sin embargo, el cambio de uso del suelo durante las últimas décadas ha mermado de
manera alarmante su extensión (Ramírez-Marcial et al. 1992), por lo que entender la dinámica de
los diversos fenómenos ecológicos que mantienen este ecosistema es fundamental para promover
su conservación y recuperación.
Uno de los procesos fundamentales en la recuperación de ecosistemas perturbados es la
dispersión de semillas por vertebrados (Guevara y Laborde 1993). Los vertebrados juegan un
papel relevante en el mantenimiento de la biodiversidad de las comunidades vegetales (Herrera
1988; Herrera et al. 1994; Jackson et al. 1995), ya que estos dispersan las semillas de más del
80% de las especies leñosas en los trópicos (Howe y Smallwood 1982). Además, se ha observado
que las comunidades animales tienen un impacto positivo en los procesos de sucesión y
resiliencia de los ecosistemas, gracias a su función como vectores de dispersión de semillas
(síndrome de dispersión zoocoro; Gálvez 2002; Rodríguez-Santamaría et al. 2006; Reid et al.
2014).
De los distintos síndromes de dispersión, la zoocoría se ha identificado como la forma
más importante para el transporte de propágulos de especies vegetales en ecosistemas como el

32
BMM. Aunque existe variación dependiendo del ecosistema, se calcula que entre el 50 y el 90
por ciento de las plantas vasculares de un determinado ecosistema producen frutos carnosos que
dependen de la dispersión animal (Howe y Smallwood 1982; Estrada y Fleming 1986), lo que a
su vez implica un transporte de semillas a mayor distancia en comparación con las semillas
dispersadas por otros síndromes (Ingle 2003; Viana et al. 2013). Además, la zoocoría reduce la
mortalidad conspecífica al movilizar los propágulos lejos de la planta madre (Janzen 1970; Hyatt
et al. 2003), transportando las semillas a sitios con condiciones microclimáticas distintas,
potencialmente favorables y, en algunos casos, facilitando el establecimiento de las plántulas
(Howe y Smallwood 1982).
Se sabe que las aves juegan un papel importante como dispersoras de especies vegetales
en ecosistemas tropicales y templados (e.g. Howe 1977; Snow 1981; Howe 1984a; Guevara et al.
1986; Wunderle 1997; Corlett 1998; Ortiz-Pulido et al. 2000; Holl 2002; Cole et al. 2010). A
pesar de que en los últimos años se han realizados estudios que evalúan el papel de las aves como
dispersoras de semillas en el BMM (e.g. Figueroa y Castro 2002; Amico y Aizen 2005; Salvande
et al. 2011; Hernández-Ladrón de Guevara et al. 2012), aún quedan interrogantes respecto a su
aporte en la recuperación del ecosistema, en particular en cuanto a las especies que están
dispersando y a la variación temporal de su papel como dispersoras, por lo que el presente trabajo
tuvo por objetivos: (1) caracterizar la riqueza y abundancia de la comunidad de aves, (2)
caracterizar la riqueza y abundancia de las especies vegetales dispersadas, (3) evaluar el papel de
las aves en el proceso de dispersión de semillas y su aporte estacional, en un potrero abandonado
en la zona central de Veracruz.

33
MÉTODOS
El estudio se realizó en un potrero recientemente abandonado (ca. 2011), con una superficie de 3
ha, donde se ha establecido un monitoreo de la sucesión secundaria. El sitio se encuentra bajo el
resguardo del Instituto de Ecología, A. C. Está ubicado en los límites de la ciudad de Xalapa y el
municipio de Coatepec, Veracruz (19º30’35”-19º30’40” N, 96º56’49”-96º56’42” W), a una
altitud de 1,300 msnm, con una temperatura media anual de 18º C y una precipitación anual de
2000 mm (Williams-Linera 2012). Este sitio tuvo un uso de suelo como potrero durante un
periodo de al menos 60 años (Fig. 1).
Para caracterizar la comunidad de aves dispersoras de semillas, se realizaron capturas
mensuales con 10 redes de niebla de 12 × 2 m teniendo un total de 240m2 de área total de captura
(Ralph et al. 1996), las cuales fueron instaladas de forma aleatoria entre las 0700 y las 1300 hr.,
con un esfuerzo de muestreo de 2100 horas/red, entre abril de 2013 a marzo de 2014. Cada ave
capturada fue extraída de la red de niebla y colocada en un saco ornitológico hasta que defecara;
posteriormente se recolectaron las excretas directamente del saco, las cuales fueron colocadas en
tubos Eppendorf de 1.5 ml y refrigeradas a -4ºC hasta su procesamiento. La identificación de las
aves se realizó con ayuda de guías de campo (e.g. Howell y Webb 1995; Sibley 2000; Peterson et
al. 2008). Para la identificación de las especies de semillas encontradas en las excretas, se realizó
una colecta de plantas en fructificación dentro del sitio de muestreo y el bosque de niebla
aledaño. Cada ejemplar fue prensado y secado para su posterior identificación en el Herbario
XAL. Con las semillas colectadas se elaboró una colección de semillas que facilitó la
identificación de las especies encontradas en las muestras.
De las especies de aves capturadas, fueron determinadas como dispersoras aquellas que
presentaron semillas en sus excretas en al menos un individuo, sin que esto necesariamente

34
implicara la viabilidad de las semillas. El análisis de las excretas se realizó mediante la
separación de los distintos elementos contenidos dentro de las excretas con la ayuda de agujas y
alcohol al 70%. Para preservar la muestra obtenida, la excreta fue fijada a un portaobjetos con
mucílago cristalino. Posteriormente, con un microscopio estereoscópico, las semillas fueron
identificadas por comparación visual con ayuda de la colección de referencia, literatura
especializada, consulta a expertos y ejemplares de herbario.
Para evaluar el esfuerzo de muestreo tanto de aves como de semillas dispersadas, los
datos de abundancia por mes se utilizaron para construir curvas de acumulación de especies por
mes para aves y semillas, y por número de individuos y por número de eventos solamente para
semillas con el programa EstimateS v.9.1.0 con un valor de aleatorizaciones de 100 repeticiones
sin remplazo para el estimador de Bootstrap (Colwell 2013). Para conocer la diversidad de
semillas dispersadas por especie de ave, se calculó el índice de Shannon (PAST v.1.89; Hammer
et al. 2001). Para conocer la fluctuación e influencia de las aves migratorias, se consideró el
periodo de migración de invierno (finales de noviembre a principio de marzo; com. pers. Octavio
Rojas) y se graficaron la riqueza y abundancia tanto para las aves migratorias como para las
semillas dispersadas a lo largo del año. Para evitar el sesgo ocasionado por la variación del
número de semillas por especie de fruto y conocer el aporte relativo de cada una de las especies
de plantas encontradas en la dieta de las aves, se calculó el Índice de Importancia Relativa (IIR;
Pinkas et al. 1971), originalmente utilizado para evaluar dieta de peces, pero que ya ha sido
utilizado para evaluar dieta de aves acuáticas (Beltzer 1985; Olguin et al. 2012) y terrestres
(Beltzer 1983; 1995; Aramburú y Corbalán 2000). El IIR fue basado en la ocurrencia relativa
(porcentaje de muestras con una determinada especie vegetal), abundancia relativa (porcentaje de
semillas de una determinada especie vegetal en relación al total de semillas registradas) y
volumen (porcentaje de volumen de las semillas de una determinada especie vegetal). En el caso

35
del volumen de las semillas, este se obtuvo midiendo el alto, largo y ancho de cada semilla con
un vernier y calculando el valor promedio por especie de planta.
RESULTADOS
Aves
Durante los doce meses de muestreo se capturaron un total de 386 aves pertenecientes a 75
especies de las cuales 45 especies están catalogadas como frugívoras por Howell y Webb (2005).
Del total de aves capturadas, se encontró presencia de semillas en las excretas de 53 individuos
pertenecientes a 27 especies, por lo que estas aves se consideraron como dispersoras (Cuadro 1).
De las especies de aves con semillas en sus excretas, 18 están catalogadas como residentes, ocho
como migratorias de invierno y solo una como residente de verano (Howell y Webb 1995). La
curva de acumulación de especies de las aves capturadas mensualmente indicó que el muestreo
representó el 84% del total de especies de aves esperadas (Fig. 2a), mientras que el muestreo de
aves dispersoras representó el 78.5% del total de especies esperadas (Fig. 2b).
Los meses con mayor abundancia de individuos capturados fueron mayo (52 individuos) y
enero (66 individuos). Los meses con mayor número de aves con semillas en excretas fueron
agosto, enero y febrero con siete capturas cada mes (Anexo 1). En cuanto a la riqueza de estas
aves a lo largo del año, febrero fue el mes con mayor número de especies (6), seguido por mayo,
agosto, enero y marzo con cinco especies en cada mes. La curva de abundancia de aves
migratorias a lo largo del año, mostró que la fluctuación va de un mínimo de un individuo en
junio y julio a un máximo de 31 en mayo (Fig. 3a). En cuanto a la riqueza de aves migratorias la
fluctuación va de un mínimo de una especie en junio y julio, hasta un máximo de 12 especies en

36
enero, comprendido entre los meses de diciembre y febrero con 11 especies de aves migratorias
cada uno, lo que coincide con la época de migración (Fig. 3b).
Semillas
Se colectaron un total de 510 semillas pertenecientes a 33 especies de 11 familias (Cuadro 2), de
las cuales Rapanea myricoides y Trema micrantha son especies comunes del BMM perturbado,
mientras que Oreopanax xalapensis y Rhamnus sp pertenecen los estratos arbustivo y mediano
respectivamente (Castillo-Campos 1991). La especie más abundante fue el arbusto Conostegia
xalapensis (Melastomataceae) con un total de 114 semillas, seguido del árbol Trichilia
havanensis (Meliaceae) con 57 semillas (Anexo 1). La curva de acumulación de especies de
plantas y el estimador de Bootstrap mostraron que el número de especies obtenidas representa el
80% de las especies esperadas (Fig. 2c); sin embargo, la curva de acumulación de especies por
evento (i.e. ave capturada con semillas en excreta; Fig. 4a) y por individuo (i.e. número de
semillas; Fig. 4b) muestran que le muestreo realizado no fue suficiente para alcanzar una asíntota.
Los meses con mayor número de semillas encontradas en aves fueron febrero (101) y
junio (119). La especie de ave con mayor riqueza de semillas en sus excretas fue Icteria virens
(8), seguida por Chlorospingus flavopectus (6) y Dumetela carolinensis (6); mientras que las
especies que defecaron la mayor abundancia de semillas fueron Myiozetetes similis (92) seguida
por Saltator atriceps (70). La diversidad de semillas por especie de ave fue mayor en Dumetela
carolinensis (H’=1.66), seguido por Icteria virens (H’=1.19), Turdus grayi (H’=1.11) y
Aimophila ruficeps (H’=1.05; Cuadro 1). En cuanto a la curva obtenida para la abundancia de
semillas dispersadas, el mes con menor abundancia fue en julio (3 semillas), mientras que los
meses con mayor abundancia fueron junio (119 semillas) y febrero (101 semillas; Fig. 3a). Los
meses con menor riqueza fueron abril, julio y octubre con una especie en cada mes. La mayor

37
riqueza de especies de semillas se observó en enero (9 especies) y febrero (8 especies), que
corresponden al periodo de migración (Fig. 3b).
Respecto al IIR, el análisis de porcentaje de frecuencia de ocurrencia mostró que las
cuatro especies con el porcentaje más alto (6%) fueron T. havanensis, R. myricoides, una especie
de la familia Poaceae (categorizada como Poaceae1; Cuadro 2) y otra de la familia
Phytolacaceae. En cuanto al porcentaje de abundancia, las tres especies con mayor importancia
fueron C. xalapensis (22.4%), una especie de la familia Poaceae (Poaceae1; Cuadro 2; 15.7%) y
T. havanensis (11.2%). En el caso del porcentaje volumétrico las especies más importantes
fueron T. havanensis (50.6%), seguida por Smilax sp. (8.0%), Rhamnus sp. (5.6%) y una especie
de la familia Poaceae (Poaceae1; Cuadro 2; 5.6%). La especie con el IIR más alto fue T.
havanensis con un valor de 370.5 (4 semillas promedio por fruto), seguido por una especies de la
familia Poaceae (Poaceae1; Cuadro 2) con un valor de 127.8 y en tercer lugar se encuentra C.
xalapensis con un valor de 91.7 (numerosas semillas por fruto) para este índice (Cuadro 2).
DISCUSIÓN
A partir del reconocimiento de la importancia de las aves en la dispersión de semillas (Ridley
1930; Van der Pijl 1972), estas han sido catalogadas como efectivas dispersoras que juegan un
papel preponderante en la regeneración de los bosques tropicales (Snow 1965; Howe 1977; Howe
1984a; Howe 1984b; Hughes 1988; Holl 1998). Pese a ello, este proceso ha sido poco estudiado
en el BMM de México (Figueroa y Castro 2002; Hernández-Ladrón de Guevara et al. 2012). En
el presente trabajo, el esfuerzo de muestreo no fue suficiente, probablemente debido al sesgo que
representa el método de muestreo con redes colocadas dentro de un potrero, el cual se enfoca en
la captura de aves de talla pequeña. Sin embargo, el total de especies capturadas en las redes (75)

38
representa alrededor del 14% de las especies reportadas para el centro del estado de Veracruz
(528; Ruelas-Inzunza 2006) y el 23% de las especies reportadas para el municipio de Xalapa
(329; González-García et al. 2014).
A pesar de que este estudio fue realizado en un potrero cercano a una zona urbana, el
número de especies capturadas que han sido catalogadas por la literatura como frugívoras (45)
representan el 27% de las especies de aves frugívoras reportadas para el municipio de Xalapa
(167; Howell y Webb 2005; González-García et al. 2014), superando a las reportadas en bosques
conservados de la Sierra Madre Oriental como en el estado de Hidalgo (36; Martínez Morales
2007) y el estado de Puebla (38; Villa-Bonilla et al. 2009). En cuanto a las aves dispersoras (27
especies con presencia de semillas en las excretas), la riqueza fue similar a lo reportado en un
potrero de la región inmerso en un paisaje menos perturbado de BMM (23; Hernández-Ladrón de
Guevara et al. 2012); y a lo observado en un matorral de la zona costera de la región central de
Veracruz (27; Vázquez-Balbuena 2014).
En cuanto a las especies vegetales encontradas en las excretas de las aves, estas
pertenecen en su gran mayoría a especies de etapas iniciales o intermedias, sin embargo las
especies de etapas intermedias fueron las más abundantes, similar a lo encontrado en otros
estudios (Vandermeer 1990; Hernández-Ladrón de Guevara et al. 2012). Cabe destacar que la
especie más abundante fue el arbusto C. xalapensis, que ha sido identificada como una especie
importante para la recuperación de bosques, ya que genera micro-hábitats favorables para el
establecimiento de otras plantas leñosas y provee perchas naturales para las aves dispersoras
(Almeda 1993). La presencia de C. xalapensis en la zona, la cual también ha sido reportada como
muy abundante en otros sitios de la región (Hernández-Ladrón de Guevara et al. 2012), sugiere
un alto potencial de recuperación del sitio. Las semillas de esta especie de planta, fue encontrada

39
principalmente en excretas de Myiozetetes similis, lo que sugiere que para el pastizal en estudio,
esta ave está actuando como una importante dispersora de esta especie vegetal en particular.
Las especies de aves que presentaron más riqueza y abundancia de semillas fueron:
Icteria virens y Dumetela carolinensis respectivamente, ambas son especies migratorias, lo que
sugiere que su papel es preponderante como dispersoras, particularmente durante los meses de
enero y febrero. A pesar de que en este trabajo, la comparación de la abundancia de las aves
migratorias y las semillas en las excretas parece no seguir un patrón o tener alguna relación a lo
largo del año, la comparación de la riqueza, se observa que durante el periodo de migración
(noviembre-marzo) el número de aves migratorias incremento y por ende también el número de
semillas en las excretas de los individuos migratorios. Se sugiere que las aves migratorias podrían
tener un efecto positivo y complementario al de las especies residentes sobre el proceso
dispersión de semillas, ya que durante su estancia se incrementa la abundancia y riqueza de
semillas excretadas.
Debido a la fuerte interacción mutualista que representa la relación planta-ave (Herrera
1984), la variación estacional de las especies de aves frugívoras podría tener una influencia
determinante en la comunidad vegetal; dado que el fenómeno de migración tiene una la
temporalidad muy marcada. Probablemente las especies de plantas beneficiadas por la presencia
de las aves migratorias sean aquellas cuya fructificación se suscita durante este periodo, lo que
tendría una fuerte influencia a nivel de paisaje al favorecer la dispersión de estas especies.
Debido a que la comunidad de aves responde a los distintos contextos de paisaje (Reid et al.
2014), sería importante realizar monitoreos a mediano y largo plazo con el fin de conocer el
proceso de colonización de las especies vegetales dispersadas.
En general, la diversidad de especies de plantas consumidas por especie de ave tendió a
ser baja, mientras que la comunidad de aves frugívoras en conjunto tuvo una diversidad dentro de

40
los rangos establecidos por Pla (2006) como normales. Sin embargo, en un trabajo cerca del área
de estudio en el que se realizó revisión de excretas se encontraron 17 especies de semillas
(Hernández-Ladrón de Guevara 2012); mientras que en otro trabajo de la zona costera del centro
de Veracruz en donde se realizaron avistamientos, se reportó que las aves consumieron 33
especies de plantas (Ortiz-Pulido et al. 2000), por lo que se considera que en el actual estudio, la
riqueza de semillas dispersadas (33) fue importante dadas las condiciones del sitio (i.e. un potrero
abandonado). A pesar de ello, es de considerar que este resultado podría estar limitado por el
método de muestreo (la captura por redes se enfoca principalmente en aves pequeñas), así como
al tiempo del estudio limitado a un año y a la frecuencia de las colectas en redes, las cuales de
acuerdo a las curvas de acumulación de especies fueron insuficientes para detectar todas las
especies de plantas cuyas semillas son consumidas y llevadas por aves al potrero. Sin embargo,
aún con estas limitantes de muestreo se registró un importante número de eventos de dispersión
hacia el potrero.
A pesar de que C. xalapensis fue la especie vegetal con mayor número de semillas
registradas en este estudio, los resultado del IIR mostraron que T. havanensis es la especie más
importante encontrada en las excretas de las aves capturadas. Resulta interesante que ambas son
especies leñosas que forman parte de la vegetación primaria y secundaria del BMM (Rzedowski
1993); y a su vez generan alimento, resguardo y perchas que propician la dispersión de semillas
por otros dispersores voladores (Guevara y Laborde 1993; Wunderle 1997; Galindo-González et
al. 2000), por lo que resultaría interesante monitorear el establecimiento de estas especies en el
potrero. Aunque existe evidencia de que el paso de las semillas por el tracto digestivo de las aves
modifica de forma favorable las condiciones de germinación (Howe y Smallwood 1982; Howe
1984a), para el caso particular de C. xalapensis, el estudio de Hernández-Ladrón de Guevara et
al. (2012) no es concluyente sobre si la germinación de esta especie está siendo beneficiada por la

41
ingesta de las aves, es por ello que se sugiere continuar con los análisis de viabilidad y
germinación en las semillas dispersadas. Sin embargo es un hecho que las aves están movilizando
las semillas de esta especie, favoreciendo así su dispersión.
En conclusión, los resultados sugieren que a pesar de la cercanía del sitio de estudio con
la ciudad de Xalapa, la comunidad de aves dispersoras del potrero estudiado es diversa. Con
respecto a este gremio de aves en particular, este estudio muestra que son importantes
movilizadoras de semillas, tanto de especies de plantas de etapas sucesionales pioneras como de
especies intermedias, las cuales tienen el potencial de contribuir al proceso de sucesión y
recuperación del BMM en el potrero abandonado. Además, en este sito el fenómeno de
dispersión se ve potenciado por la llegada de aves migratorias al incrementar la riqueza de
semillas dispersadas. Esta investigación sienta las bases para estudios relacionados con el
acontecer de las semillas dispersadas y la dinámica del proceso de recuperación en sitios
perturbados de BMM de la zona centro del estado de Veracruz.
AGRADECIMIENTOS
Agradezco a CONACyT por haber otorgado el financiamiento para los estudios de posgrado y al
posgrado del Instituto de Ecología A. C. Al Dr. Diego Santiago por su apoyo en la identificación
de aves en campo y al Dr. Gonzalo Castillo por su apoyo en la identificación de las especies
vegetales colectadas. A los compañeros de laboratorio Patricia Téllez, Patricia Parra, Claudio
Mota, Natali Novelo, Mauricio Ortega, David Prieto y Norma Hernández por su ayuda en el
trabajo de laboratorio. A quienes apoyaron con el trabajo de campo Diana Balbuena, Omar
Hernández, Cristian A. Martínez, Omar Suárez, Carolina Hernández, Carolina Solís, Angelina
Ruiz, Rafael A. Rueda, Jorge L. Cortinas.

43
CUADROS Y FIGURAS
Figura 1. Área de estudio. En el mapa se muestra el potrero delimitado por el polígono negro ubicado al sureste de la
ciudad de Xalapa en la zona centro del estado de Veracruz.
45
Figura 2. Curvas de acumulación de especies bajo el estimador de Bootstrap de: a) aves capturadas, b) aves
frugívoras y c) especies vegetales observadas a través de las semillas colectadas en las excretas en un potrero al
sureste de la ciudad de Xalapa, Veracruz.
47
Figura 3. Recambio de aves y semillas por mes de muestreo en el potrero: a) abundancia y b) riqueza. La barra negra
( ) sobre el eje de los meses indica el periodo de presencia de aves migratorias en la región central de
Veracruz (noviembre a marzo).
49
Figura 4. Curvas de acumulación de especies de especies vegetales por: a) número de eventos (aves con semillas en
sus excretas) y b) número de individuos (semillas en excretas) en un potrero abandonado al sureste de la ciudad de
Xalapa, Veracruz.
51
Cuadro 1. Índice de diversidad por especie de ave dispersora. Se muestra el número de individuos; estatus (WV =
Migratorio de invierno, RB = Residente, SR = Residente de verano). Gremio alimenticio (I = Insectívoro, F =
Frugívoro, G = Granívoro, C = Carnívoro); la riqueza y abundancia de semillas en excretas; y el índice de Shannon
(H’) para cada especie de ave. Las abreviaturas de permanencia y gremio alimenticio corresponden a la clasificación
realizada por Howell y Webb 2005.
Especie Número de
aves Estatus
Gremio
alimenticio
Riqueza
de semillas
Abundancia
de semillas H’
Dumetella carolinensis 5 WV IF 6 44 1.664
Icteria virens 10 WV IF 8 51 1.197
Turdus grayi 5 RB IC 5 27 1.111
Aimophila rufescens 3 RB GI 3 5 1.055
Chlorospingus flavopectus 3 RB FGI 6 40 0.7992
Megarynchus pitangua 1 RB IC 2 2 0.6931
Saltator atriceps 2 RB GF 2 70 0.6927
Myiozetetes similis 3 RB FGI 3 92 0.4846
Hylocichla mustelina 2 WV FI 2 16 0.4826
Myiarchus tuberculifer 2 RB IF 2 7 0.4101
Seiurus aurocapilla 1 WV IF 2 8 0.3768
Euphonia elegantissima 1 RB FI 1 58 0
Arremon brunneinucha 1 RB FI 1 27 0
Euphonia affinis 1 RB I 1 20 0
Myiodynastes luteiventris 1 SR I 1 9 0
Psilorhinus morio 1 RB FG 1 8 0
Quiscalus mexicanus 1 RB GIF 1 5 0
Setophaga coronata 1 WV IF 1 5 0
Colaptes rubiginosus 1 RB IF 1 3 0
Setophaga virens 1 WV I 1 3 0
Catharus aurantiirostris 1 RB GI 1 2 0
Contopus virens 1 RB IF 1 2 0
Mionectes oleagineus 1 RB IF 1 2 0
Empidonax albigularis 1 WV I 1 1 0
Empidonax occientalis 1 RB I 1 1 0
Vireo griseus 1 WV IF 1 1 0
Vireo solitarius 1 WV IF 1 1 0
Total 53 - - 33 510 2.687
53
Cuadro 2. Índice de importancia relativa (IIR=F(N+V) según Pinkas et al. 1971 por especie vegetal encontrada. Se
muestra la familia, hábito de crecimiento y etapa sucesional de cada especie vegetal. Los porcentajes obtenidos son
relativos a los valores totales de cada rubro (F = Porcentaje de ocurrencia, N = Porcentaje de abundancia, V =
porcentaje volumétrico). Las casillas marcadas con ND son aquellas de las cuales no se tiene dato (ND = no dato).
Las especies se muestran en orden de mayor a menor importancia relativa.
Especie Familia Hábito de
crecimiento
Etapa
sucesional
Porcentaje
de
ocurrencia
Porcentaje
de
abundancia
Porcentaje
volumétrico IIR
Trichilia havanensis Meliaceae leñosa intermedia 6 11.2 50.6 370.5
Poaceae1 Poaceae pasto pionera 6 15.7 5.6 127.8
Conostegia xalapensis Melastomataceae leñosa intermedia 4 22.4 0.6 91.7
Phytolaccaceae1 Phytolaccaceae herbácea pionera 6 4.1 2.4 39.2
sp.28 Desconocida ND ND 4 6.7 1.3 31.8
Rapanea myricoides Primulaceae leñosa intermedia 6 1.2 2.5 22.2
Smilax sp. Smilacaceae herbácea pionera 2 2.5 8 21.1
sp.18 Desconocida ND ND 4 2 3.1 20.1
Passifloraceae1 Passifloraceae herbácea pionera 4 1.6 2.9 18.1
sp.12 Desconocida ND ND 4 2.9 1.4 17.2
sp.31 Desconocida ND ND 2 7.1 0.9 16
Rhamnus sp. Rhamnaceae leñosa intermedia 2 1.6 5.6 14.4
sp.26 Desconocida ND ND 2 2.7 3.4 12.3
sp.1 Desconocida ND ND 2 5.3 0 10.7
sp.20 Desconocida ND ND 4 0.6 1.7 9.1
sp.21 Desconocida ND ND 2 0.2 4.1 8.6
sp.13 Desconocida ND ND 2 3.3 0 6.7
sp.30 Desconocida ND ND 4 0.6 1.1 6.6
sp.19 Desconocida ND ND 4 0.8 0.7 6
sp.29 Desconocida ND ND 2 0.6 1.5 4.2
Solanaceae1 Solanaceae leñosa pionera 4 0.8 0.2 3.8
sp.23 Desconocida ND ND 2 1 0.9 3.8
sp.32 Desconocida ND ND 2 1.2 0.4 3.2
Solanaceae2 Solanaceae leñosa pionera 2 1.4 0.1 3
Poaceae4 Poaceae pasto pionera 2 0.4 0.3 1.3
sp.11 Desconocida ND ND 2 0.4 0.2 1.1
Poaceae2 Poaceae pasto pionera 2 0.4 0.1 1
Oreopanax xalapensis Araliaceae leñosa tardía 2 0.2 0.2 0.8
sp.22 Desconocida ND ND 2 0.4 0 0.8
sp.6 Desconocida ND ND 2 0.4 0 0.8
sp.27 Desconocida ND ND 2 0.2 0.1 0.6
Trema micrantha Ulmaceae leñosa pionera 2 0.2 0.1 0.5
sp.7 Desconocida ND ND 2 0.2 0 0.4
55
ANEXOS
Anexo 1. Cuadro de abundancia y riqueza de aves y semillas por mes de muestreo en un potrero al sureste de la ciudad de Xalapa, Veracruz.
AVES AVES DISPERSORAS SEMILLAS
MES ABUNDANCIA RIQUEZA ABUNDANCIA
RESIDENTES
ABUNDANCIA
MIGRATORIAS ABUNDANCIA RIQUEZA ABUNDANCIA RIQUEZA
ABRIL 19 12 12 5 2 2 20 1
MAYO 66 21 30 31 6 5 69 7
JUNIO 21 11 20 1 4 4 119 2
JULIO 10 8 9 1 1 1 3 1
AGOSTO 28 16 21 4 7 5 59 4
SEPTIEMBRE 47 18 22 22 4 3 7 4
OCTUBRE 21 11 4 17 2 2 9 1
NOVIEMBRE 28 18 14 14 6 4 21 6
DICIEMBRE 41 18 16 25 2 2 6 2
ENERO 52 26 25 26 7 5 41 9
FEBRERO 29 19 14 15 7 6 101 8
MARZO 24 18 12 12 5 5 55 5
Total 386 75 199 173 53 27 510 33

57
III. LA LLUVIA DE SEMILLAS COMO CATALIZADORA DE LA
RESTAURACIÓN DEL BOSQUE MESÓFILO DE MONTAÑA

59
LA LLUVIA DE SEMILLAS COMO CATALIZADORA DE LA
RESTAURACIÓN DEL BOSQUE MESÓFILO DE MONTAÑA
RESUMEN
El bosque mesófilo de montaña (BMM) es un ecosistema de gran relevancia debido a que alberga
una gran diversidad de especies en proporción a su área; sin embargo, este es el ecosistema
terrestre más amenazado a nivel mundial. En el presente trabajo se caracterizó y comparó la
composición de lluvia de semillas en un BMM y un potrero adyacente durante nueve meses. La
colecta de semillas se realizó con 32 trampas, 16 en cada sitio. Las semillas se clasificaron por
hábito de crecimiento, síndrome de dispersión y etapa sucesional. Se colectaron un total de
105,163 semillas de 81 especies. En la clasificación por síndrome de dispersión, la anemocoría
fue el síndrome con mayor similitud entre los dos hábitats (Ĵabd=0.919), mientras que la zoocoría
mostró diferencias significativas entre sitios en cuanto a la abundancia (U=63.50; gl=30;
P=0.015). En la clasificación por hábito de crecimiento, las especies leñosas fueron las de mayor
abundancia y riqueza (BMM: 59,556 semillas; 18 especies; potrero: 19,161 semillas; 15 especies)
con el mayor valor de similitud (Ĵabd=0.981). En cuanto a la clasificación por etapas sucesionales,
se observaron diferencias significativas entre ambos sitios para todas las clasificaciones, mientras
que la mayor similitud se registró para las especies de sucesión tardía (Ĵabd=0.996). Los
resultados sugieren que el potrero estudiado tiene un importante potencial de regeneración
forestal dadas las características de la lluvia de semillas y la cercanía con el BMM.
Palabras clave: potrero, síndromes de dispersión, etapas sucesionales, formas de vida.

61
INTRODUCCIÓN
La recuperación natural de un bosque que ha sido perturbado está determinada por la duración,
intensidad y frecuencia del disturbio previo (Lamb et al. 2005; Muñiz-Castro et al 2006), así
como por los mecanismos de regeneración de la vegetación circundante. La recuperación de estos
sitios puede ocurrir por diversos medios, por ejemplo a través de la interrupción del disturbio o
del control de las especies invasoras; no obstante, las fuentes de propágulos disponibles en el área
aledaña son un elemento indispensable para que este tipo de recuperación se lleve a cabo (Gubb
1977; Purata 1986; Aide et al. 1996; Aide et al. 2000; Holl et al. 2000; Lamb y Gilmour 2003).
De esta forma, la dispersión de semillas forma parte del ciclo de regeneración natural y de la
sucesión de las comunidades forestales que mantienen y regeneran los ecosistemas perturbados
(Peña- Domene et al. 2013). En este sentido, la capacidad de las plantas para dispersarse y
establecerse, influirá de forma crucial en el proceso de colonización y por tanto en la consecuente
recuperación del ecosistema (Ramírez-Marcial et al. 1992; Wang y Smith 2002; Rodríguez-
Santamaría et al. 2006).
La dispersión de semillas es el proceso que une el ciclo reproductivo de las plantas con el
establecimiento de su progenie (Wang y Smith 2002). Este proceso de desplazamiento de las
unidades reproductivas de las plantas tiene la finalidad de incrementar las probabilidades de
establecimiento de la especie (Janzen 1970; Connell et al. 1984). El estudio de este movimiento
puede realizarse a través del análisis de la lluvia de semillas y del banco de semillas (Dalling et
al. 1998; Ramírez-Marcial et al. 1992). En el caso de la lluvia de semillas, su composición es el
reflejo de la fenología de las especies vegetales de un ecosistema así como del comportamiento o
desempeño de los vectores de dispersión, mientras que la permanencia de las especies en el banco
de semillas depende de la latencia y viabilidad de las semillas, así como de las condiciones

62
ambientales que pueden o no ser propicias para la germinación de las plantas (Marlette y
Anderson 1986; Henderson et al. 1988; Levassor et al. 1990; Bedoya-Patiño et al. 2010). Sin
embargo, factores relacionados con el estado sucesional del ecosistema, como la cobertura
vegetal, la humedad, o la incidencia de luz, también influirán de forma importante en el
establecimiento de las especies vegetales que ahí se depositen (Grubb 1977; Denslow y Guzmán
2000).
Por otro lado, las formas o síndromes de dispersión a través de los cuales las semillas
serán transportadas a sitios potenciales de establecimiento, serán determinados por los vectores
físicos (viento, anemocoría; agua, hidrocoría; o gravedad, barocoría), o biológicos (a través de
algún animal; zoocoría; por expulsión mediante las paredes del fruto o partes del fruto,
autocoría), a partir de los cuales serán nombrados (Tiffney 2004). Además de los síndromes de
dispersión, existen otras características importantes de la comunidad de semillas dispersadas a
tomar en cuenta, como el hábito de crecimiento y la etapa sucesional a la que pertenecen las
especies dispersadas, ya que dichas características pueden ayudar a reconocer el potencial de
recuperación de un sitio perturbado (Grubb 1977). Por ejemplo, la presencia de plantas pioneras
es importante para la formación de condiciones que faciliten el establecimiento de otras especies,
puesto que la llegada y establecimiento de especies de sucesión intermedia y tardía dependerá de
las condiciones generadas por especies pioneras (Posada et al. 2000; Holl et al. 2000; Peña-
Domene et al. 2013).
Los estudios de dispersión de semillas por vertebrados frugívoros en México, se han
realizado mayormente en selvas (e.g. Guevara y Laborde 1993; Estrada et al. 1993; Estrada y
Coates-Estrada 1996; Galindo-González et al. 2000). Sin embrago, poco se conoce en general
sobre el proceso de dispersión del Bosque Mesófilo de Montaña (BMM; Del Castillo y Pérez
Ríos 2008; Hernández-Ladrón de Guevara et al. 2012). El área remanente del BMM de la región

63
central de Veracruz es pequeña; pero en comparación con otras zonas del país posee una alta
diversidad botánica y un importante número de especies endémicas (Williams-Linera 2002; Cruz
et al. 2010). A pesar de su importancia, en la actualidad, casi la mitad de la superficie del BMM
en esta región ha sido transformada por actividades antrópicas (Challenger 1998, González-
Espinoza et al. 2012; Toledo-Aceves et al. 2011). Por ello, conocer la composición de la lluvia de
semillas, así como los síndromes de dispersión dominantes en sitios con distinto grado de
perturbación del BMM, ayudará a entender sobre los procesos que participan en el
establecimiento y regeneración forestal de este ecosistema. El objetivo del presente estudio fue
caracterizar y comparar la composición de la lluvia de semillas (síndromes de dispersión, hábito
de crecimiento y etapa sucesional) en un remanente de BMM y un potrero cercano recientemente
abandonado, evaluando el potencial del potrero para recuperar la composición del BMM a partir
de la lluvia de semillas.
MÉTODOS
El estudio se realizó en un potrero en recuperación y un fragmento de BMM, ambos ubicados en
predios pertenecientes al Instituto de Ecología, A. C, Xalapa, Veracruz (19º30’39”-
19°31’16”N,96°56’24”-96°56’08”; 1,400 msnm), la temperatura media anual en el área de
estudio es de 18º C y la precipitación anual promedio es de 2,000 mm con tres estaciones
relativamente bien definidas: estación fría y seca (noviembre a marzo), cálida y seca (abril a
mayo), cálida y húmeda (junio a octubre) (Ortega-Pieck et al. 2011).
El BMM se ubica en el área protegida “Santuario de Bosque de Niebla” el cual tuvo un
uso previo como cafetal, pero actualmente tiene un tiempo de recuperación de 80 años. El potrero
es un área que anteriormente formó parte del BMM, pero que fue transformado a un uso de suelo

64
como cultivo de naranja y ganadería durante un periodo de 60 años. Desde hace tres años se
eliminó la presencia de ganado y se iniciaron proyectos de restauración. El área seleccionada de
cada sitio de estudio fue de 3 ha (Fig. 5).
Para caracterizar y comparar la composición de la lluvia de semillas entre los dos tipos de
hábitat, se utilizaron 32 trampas cónicas de semillas (0.60 m de diámetro, 1.20 m de altura), con
una funda de tela tergal strech (con apertura de malla cerrada para atrapar semillas pequeñas),
que fue remplazada cada mes, teniendo un área total muestreada de 4.48 m2 por hábitat. Las
trampas de semillas se colocaron por pares en ocho parcelas (15 × 15 m) separadas por 30 m cada
par, 16 en potrero y 16 en BMM. Las muestras fueron colectadas y analizadas mensualmente
durante un periodo de 9 meses, comprendido de mayo 2013 a enero 2014. Cuando la revisión de
las muestras no pudo realizarse inmediatamente, las muestras se mantuvieron en refrigeración (-
4ºC) para evitar la descomposición y contaminación.
La identificación de las especies de semillas encontradas en las trampas, se realizó
mediante comparación directa a una colección de referencia que se elaboró en este estudio por
medio de la colecta sistemática de plantas en fructificación dentro y en los alrededores de los
sitios de muestreo. A cada ejemplar colectado le fue removida una muestra de sus frutos y
semillas antes de ser prensado y secado para su incorporación al herbario. La identificación fue
realizada con ayuda del listado botánico del municipio de Xalapa (Castillo-Campos 1991) y los
ejemplares del Herbario XAL. Cuando fue posible, las semillas encontradas en los colectores
fueron identificadas hasta especie usando los ejemplares de referencia y la literatura especializada
(Castillo-Campos y Medina-Abreo 2005; Rodríguez-Velázquez et al. 2009; Niembro et al. 2010).
Para comparar las diferencias en la composición de especies de la lluvia de semillas entre
cada hábitat, estas se caracterizaron con base en: su hábito de crecimiento (pastos, leñosas y
herbáceas); su etapa sucesional (pionera, intermedia, tardía); y su forma de dispersión

65
(anemócoras, barócoras, zoócoras). La clasificación de hábito de crecimiento y etapa sucesional
en las especies identificadas, se hizo de acuerdo a lo reportado en la literatura (Rzedowski 1978;
Castillo-Campos y Medina-Abreo 2005; Rodríguez-Velázquez et al. 2009). La clasificación de
las semillas anemócoras se realizó por la presencia de estructuras como verilo, alas y brácteas
modificadas que se sabe le facilitan a la semilla ser transportadas por el viento. Las semillas
barócoras se clasificaron según la literatura usada para la identificación de las especies
(Rodríguez et al. 2009. Gómez-Pompa et al. 2010). Cuando se encontraron frutos completos, se
clasificaron como zoócoros por los colores propios del síndrome (amarillo, naranja, rojo, negro,
púrpura, morado) y la constitución propia de un fruto carnoso. Esta última clasificación se
complementó con literatura (Niembro et al. 2010), así como las especies vegetales previamente
identificadas en excretas de aves (Hernández-Ladrón de Guevara et al. 2012; Capítulo 1).
Para compara la lluvia de semillas entre ambos hábitats se elaboraron curvas de
acumulación de especies por unidades de muestreo (i.e. trampas de semillas) y por número de
individuos (i.e. número de semillas). Con la finalidad de conocer a las especies dominantes
numéricamente y su mecanismo de dispersión en cada hábitat, se identificaron las semillas
colectadas en las trampas y se obtuvieron curvas de rango-abundancia. Se analizaron la
abundancia y la riqueza solo de las especies identificadas (Anexo 2). Para comparar la similitud
de especies de plantas entre los dos hábitats se calculó el índice Chao-Jaccard usando el programa
EstimateS v.9.1.0; esta comparación se hizo tanto para el total de especies, así como por hábito
de crecimiento, tipo de dispersión y etapa sucesional (Chao et al. 2005). Para evaluar las
diferencias en la abundancia de semillas por hábito de crecimiento, tipo de dispersión y etapa
sucesional entre los dos tipos de hábitat, se hizo una prueba no paramétrica de U de Mann-
Whitney (Systat Ver. 12).

66
RESULTADOS
Se capturaron un total de 105,163 semillas pertenecientes a 81 especies, de las cuales 17% fueron
identificadas a nivel de especie, 6% a nivel de género, 25% a nivel de familia, 52% a nivel de
morfoespecie, es decir un 11% de semillas no identificadas (11,701 semillas con identidad
desconocida). El mes con mayor abundancia y riqueza fue noviembre (51,488 semillas, 44
especies; Anexo 2) y la especie más abundante a lo largo de todo el muestreo fue Platanus
mexicana (61,793 semillas, 59%). La diferencia de abundancia de semillas entre ambos sitios fue
significativa (U=67.0; gl=30; P=0.021) siendo mayor en el BMM, el índice Chao-Jaccard fue de
Ĵabd=0.755 para ambos sitios (Cuadro 3).
En el sitio de BMM se registró un total de 74,892 semillas pertenecientes a 62 especies de
las cuales 28 fueron encontradas exclusivamente en este hábitat, las especies raras fueron 11
(18%). Del total de especies de semillas del BMM, 19% fueron identificadas a nivel de especie,
8% a nivel de género, 26% a nivel de familia, 47% a nivel de morfoespecie (Anexo 2). La
densidad promedio de semillas fue de 37,279 semillas·m2·mes-1 y el mes con mayor abundancia
y riqueza fue noviembre (46,831 semillas, 63%; 26 especies; Anexo 2).
En el potrero se registró un total de 30,271 semillas pertenecientes a 56 especies de las
cuales 19 fueron encontradas exclusivamente en este hábitat, las especies raras fueron 6 (11%).
Del total de especies de semillas del potrero, 18% fueron identificadas a nivel de especie, 5% a
nivel de género, 32% a nivel de familia, 45% a nivel de morfoespecie (Anexo 2). La densidad
promedio de semillas fue de 15,068 semillas·m2·mes-1; el mes con mayor abundancia fue enero
(9,249 semillas, 31%) y el de mayor riqueza fue agosto (27 especies).
La curva de acumulación de especies por unidad de muestreo presentó una mayor
densidad total de especies en el BMM que en el potrero; sin embargo, el índice de confianza de

67
ambos sitios se sobrepone, por lo que la riqueza acumulada por unidad de muestreo para ambos
sitios es equiparable (Fig. 6a). La curva de acumulación de especies por individuos presento una
mayor riqueza acumulada en el potrero que en el BMM, pero de igual manera se sobrepone el
índice de confianza por lo que la riqueza acumulada por número de individuos para ambos sitios
también es equiparable (Fig. 6b). Las curvas de rango-abundancia muestran que en el BMM
Platanus mexicana (57,924 semillas) fue la especie dominante numéricamente con un pico de
abundancia en noviembre (31,711 semillas), seguida por Clethra macrophylla (822 semillas) y
Carpinus tropicalis (457 semillas), que numéricamente fueron menos dominantes. En el potrero
se registraron dos especies con dominancias muy similares, Solanum sp. (6,770 semillas) y
Clethra macrophylla (6,707 semillas), seguidas por Platanus mexicana y dos especies de la
familia Asteraceae (Fig. 7). En el caso de Solanum sp., el pico de abundancia fue en agosto
(4,054 semillas), mientras que para C. macrophylla su mayor abundancia fue en enero (6,132
semillas).
En la clasificación por síndrome de dispersión (Anexo 2), la anemocoría fue el síndrome
más abundante (BMM: 59,613 semillas; 75%; potrero: 19,908 semillas; 25%). El pico de
abundancia fue en el mes de noviembre (36,127 semillas; 46%) que se debió al importante aporte
de P. mexicana (Fig. 8a). En el caso de la zoocoría (BMM: 8,692; 94.5%; potrero: 510; 5.5%), el
pico de abundancia se mostró en el mes de abril (4,203 semillas; 46%) debido a un importante
aporte de Solanum sp. en el potrero (Fig. 8b). Para la barocoría, el pico de abundancia se presentó
en el mes de noviembre (225 semillas; 51%), la especie más abundante de éste síndrome fue
Leucaena pulverulenta, la cual se presentó solamente en el potrero (Fig. 8c). En cuanto a la
comparación de la abundancia de semillas por síndrome de dispersión, la anemocoría (U=78.00;
gl=30; P=0.059) y la barocoría (U=147.00; gl=30; P=0.437) no presentaron diferencias
significativas en el promedio de semillas por trampa entre los sitios; mientras que la zoocoría

68
mostró diferencia significativa (U=63.50; gl=30; P=0.015), siendo mayor el promedio de semillas
zoócoras en el potrero que en el BMM. En cuanto a la riqueza, el síndrome de dispersión que
mayor número de especies presentó fue la zoocoría con 18 especies en ambos sitios, seguida por
la anemocoría con 16 y finalmente la barocoría con 5. El valor más alto para el índice de
similitud lo presentó la anemocoría (Ĵabd=0.919), seguida por la zoocoría (Ĵabd=0.765). En cuanto
a la barocoría, no hubo especies compartidas entre ambos sitios (Ĵabd=0; Cuadro 3).
La clasificación por hábito de crecimiento (Anexo 2), mostró que en ambos sitios las
especies leñosas fueron las de mayor abundancia y riqueza (BMM: 59,556 semillas; 76%; 18
especies. Potrero: 19,161 semillas; 24%; 15 especies). El pico de abundancia fue en el mes de
noviembre (35,951 semillas; 46%), principalmente en el BMM por la presencia de P. mexicana
(Fig. 9c). En el caso de las plantas herbáceas (BMM: 577; 7.5%; potrero: 7,115; 92.5%) el pico
de abundancia se observó en el mes de diciembre (3,374 semillas; 44%), debido a la presencia de
semillas de la familia Asteraceae. Para los pastos (BMM: 85; 4%; potrero: 2,097; 96%) el pico de
abundancia se observó en diciembre (716 semillas; 33%) principalmente en el potrero (Fig. 9a y
9b) por la especie Poaceae 4 (Anexo 2). La comparación de la abundancia por sitio mostró
diferencias significativas para las plantas herbáceas (U=35.50; gl=30; P<0.001) y los pastos
(U=63.50; gl=30; P=0.014), no así para las plantas leñosas (U=79.00; gl=30; P=0.065). El valor
más alto en el síndrome de similitud entre sitios fue para las plantas leñosas (Ĵabd=0.981), seguido
por las especies de plantas herbáceas (Ĵabd=0.924) y finalmente los pastos (Ĵabd=0.156; Cuadro 3).
Para la clasificación por etapa sucesional (Anexo 2), las especies se caracterizaron de
acuerdo con Rzedowski (1978), Castillo-Campos y Medina-Abreo (2005) y Rodríguez-
Velázquez et al. (2009). Las especies de sucesión tardía fueron las más abundantes en ambos
sitios (BMM: 58,597 semillas; 94%; potrero: 3,887 semillas; 6%), con un pico en el mes de
noviembre (33,869 semillas; 54%), principalmente en el BMM (Fig. 10c) por la presencia de P.

69
mexicana (Anexo 2). El pico de abundancia para las especies pioneras (BMM: 662; 7%; potrero:
9,212; 93%) se presentó en el mes de diciembre (4,090 semillas; 41%) principalmente en el
potrero (Fig. 10a) por la presencia de la especie Asteraceae 2 (Anexo 2). En el caso de las
especies de sucesión intermedia (BMM: 974; 6%; potrero: 15,292; 94%), el pico se presentó en el
mes de enero (6,402 semillas; 39%) principalmente en el potrero (Fig. 10b) por la presencia de
Clethra macrophylla (Anexo 2). Considerando la etapa sucesional, se observaron diferencias
significativas entre ambos sitios: pioneras (U=26.00; gl=30; P<0.001), intermedias (U=13.00;
gl=30; P<0.001) y tardías (U=189.00; gl=30; P=0.021). Las especies pioneras fueron las que
mostraron mayor riqueza en ambos sitios (BMM: 13 especies; potrero: 14 especies) y el valor de
índice de similitud más alto fue para las especies de sucesión tardía (Ĵabd=0.996), seguido por las
especies de sucesión intermedia (Ĵabd=0.979) y finalmente las especies pioneras (Ĵabd=0.791;
Cuadro 3).
DISCUSIÓN
Una de las principales barreras para la recuperación de sitios que han sido perturbados es la
llegada de propágulos que puedan germinar y establecerse, por lo que es importante conocer la
riqueza y abundancia de las semillas que llegan a estos sitios (Hughes y Fahey 1988; Aide et al.
2000; Holl et al. 2000). Al comparar el BMM y el potrero por unidad de muestreo, el BMM
presentó la mayor riqueza, mientras por número de individuos el potrero presentó el mayor
número de especies acumuladas. Sin embrago, en ambos caso se observó un sobrelapamiento de
los índices de confianza, lo que sugiere que la diversidad de la lluvia de semillas es equiparable
en ambos sitios.

70
La especie dominante en todo el muestreo, Platanus mexicana, es una especie de fácil
establecimiento en su área de distribución, localmente abundante y exenta de peligro de extinción
(Carranza 1994). La segunda especie dominante, Clethra macrophylla, es una especie escasa en
su área de distribución y propia del BMM (González-Villareal 1996). Ambas especies son plantas
leñosas de dispersión anemócora, por lo que alcanzan grandes distancias de dispersión y se
encuentran presentes gran parte del año, como es propio (dependiendo de la especie) de las
plantas con este síndrome de dispersión (Howe y Smallwood 1982; Cole et al. 2010; Martínez-
Garza et al. 2011); éstas características explican la abundancia y representación en el paisaje de
estas dos especies. La presencia de las semillas de ambas especies en el potrero coincide con
reportes que indican que las plantas de dispersión anemócora son dominantes en la colonización
inicial de los mismos (Aide et al. 1996; Holl 1999), por lo que son especies importantes para la
restauración (Castillo-Campos 1991; Williams-Linera 2012).
En el presente estudio, la zoocoría fue el segundo síndrome más abundante para ambos
sitios debido a que la tercer especie dominante fue Solanum sp. Sin embargo, los resultados por
abundancia mostraron que este síndrome de dispersión es significativamente más abundante en el
potrero que en el BMM, lo que podría favorecer al establecimiento de estas y otras especies en el
potrero. Es importante resaltar que la alta abundancia y riqueza de especies zoócoras en el potrero
podría deberse al contexto de paisaje local, como lo descrito por Cole et al. (2010), donde se
menciona que la ubicación de un sitio preservado adyacente a uno perturbado juega un papel
preponderante para la restauración del sitio.
El género Solanum está principalmente constituido de plantas leñosas pioneras, las cuales
han sido descritas como importantes para la regeneración en bosques tropicales (Guevara y
Laborde 1993; Wunderle 1997; Jordano 2000; Vieira y Scariot 2006; Quesada et al. 2009; ver
Capítulo 1). Las especies que pertenecen a este género, suelen ser abundantes en áreas abiertas o

71
perturbadas y de fácil establecimiento (Nee 1986), debido a que está asociado a la dispersión
zoócora mediada principalmente por murciélagos. Ésta característica podría explicar su
abundante presencia en sitios como el potrero estudiado ya que los murciélagos tienden a excretar
al vuelo y en sitios abiertos (Gorchov et al. 1993). Es probable que los arbustos aislados de esta
especie en el potrero provean de sitios de resguardo y forrajeo para los organismos dispersores
propiciando el proceso de dispersión y en consecuencia la sucesión (Guevara y Laborde 1993;
Galindo-González et al. 2000).
La alta abundancia de semillas de plantas herbáceas y pastos en el potrero, durante la
estación de invierno denota la capacidad del sitio perturbado para propiciar el establecimiento
especies heliófilas que podrían dificultar el establecimiento de otras especies nativas y leñosas.
La invasión por el pasto africano, Cynodon plectostachyus que fue introducido para el forrajeo
del ganado (Ortega-Pieck 2011; Román-Dañobeytia et al. 2012), en ausencia de ganado provoca
una acumulación de biomasa tanto aérea como subterránea que genera una fuerte competencia
por los recursos, dificultando el establecimiento de otras especies importantes para la
recuperación del BMM (e.g. Quercus xalapensis; Ortega-Pieck et al. 2011). En sitios dominados
por especies invasoras, la colonización y establecimiento de especies de sucesión intermedia y
tardía, principalmente de árboles y arbustos puede resultar menos efectiva. Para restaurar la
vegetación forestal del sitio es necesario eliminar a la especie exótica invasora y promover el
establecimiento de las especies nativas (Muñiz-Castro 2008; Ortega-Pieck 2011; Román-
Dañobeytia et al. 2012).
Los resultados de este trabajo deben tomarse con precaución, ya que no todas las especies
fueron identificadas, sin embargo, sugieren que el BMM tiene un alto potencial de recuperación
en el potrero estudiado, dada la abundancia y la riqueza de las semillas que llegan al potrero, lo
cual es propiciado por el contexto del paisaje y la cercanía del bosque. A pesar de la

72
transformación sufrida en el sitio, la llegada de propágulos no será la limitante para que este sitio
pueda alcanzar una composición similar al BMM. Sin embargo, un manejo activo del sitio (i.e.,
deteniendo el crecimiento del pasto exótico), facilitaría la recuperación de la composición de
especies típica del BMM.
AGRADECIMIENTOS
Se agradece a CONACyT por haber otorgado el financiamiento para los estudios de posgrado y al
posgrado del Instituto de Ecología A.C. Al Dr. Gonzalo Castillo por su valioso apoyo en la
identificación de las especies vegetales. Al Dr. Javier Laborde por sus sugerencias. A Patricia
Téllez, Natali Novelo, Patricia Parra, Claudio Mota, Mauricio Ortega, David Prieto y Norma
Hernández por la ayuda en el trabajo de laboratorio y a Javier Tolome, Cristian Martínez, Edel
Atondo, Petra Galindo y Jorge L. Cortinas quienes apoyaron con el trabajo de campo.

73
CUADROS Y FIGURAS
Figura 5. Área de estudio. En el mapa se muestra el potrero (polígono con trama en diagonal) y el BMM (polígono
con trama en horizontal) ubicados al sureste de la ciudad de Xalapa en la zona centro del estado de Veracruz.
75
Figura 6. Curvas de acumulación de especies de plantas por: a) unidades de muestreo (trampas de semillas) y b)
número de individuos (semillas) para un BMM y un potrero ubicados al sureste de la ciudad de Xalapa, Veracruz.
77
Figura 7. Curvas de rango-abundancia. Especies de semillas registradas en la lluvia de semillas, se comparan dos
sitios: bosque con 62 especies y potrero con 56 especies. Las especies se encuentran en orden decreciente de
abundancia por sitio de estudio. Se muestran los nombres abreviados de las especies dominantes por hábitat. (Pm:
Platanus mexicanus, Cm: Clethra macrophylla, Ct: Carpinus tropicalis, Vb: Vitis bourgaeana, Sa: Senecio
angulifolius, sp.13: morfoespecie 13, Ssp.: Solanum spp., Ast.2: Asteraceae sp.2, Ast.3: Asteraceae sp.3, Poa.4:
Poaceae sp.4).
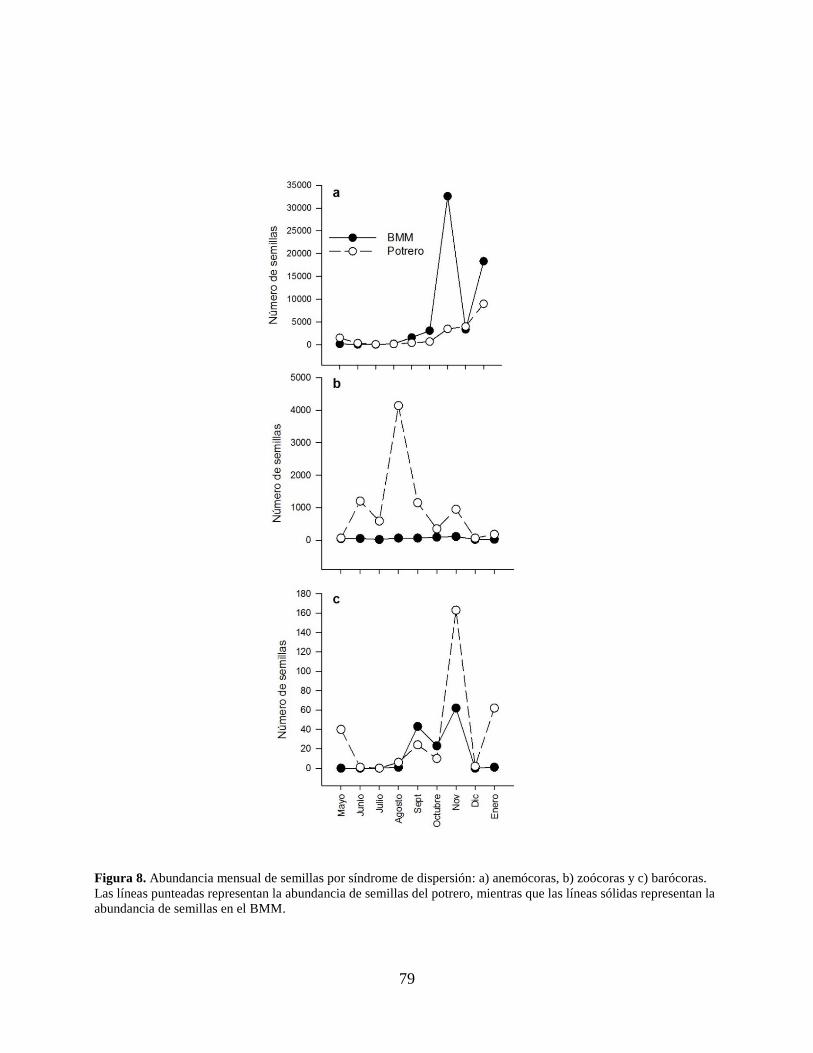
79
Figura 8. Abundancia mensual de semillas por síndrome de dispersión: a) anemócoras, b) zoócoras y c) barócoras.
Las líneas punteadas representan la abundancia de semillas del potrero, mientras que las líneas sólidas representan la
abundancia de semillas en el BMM.
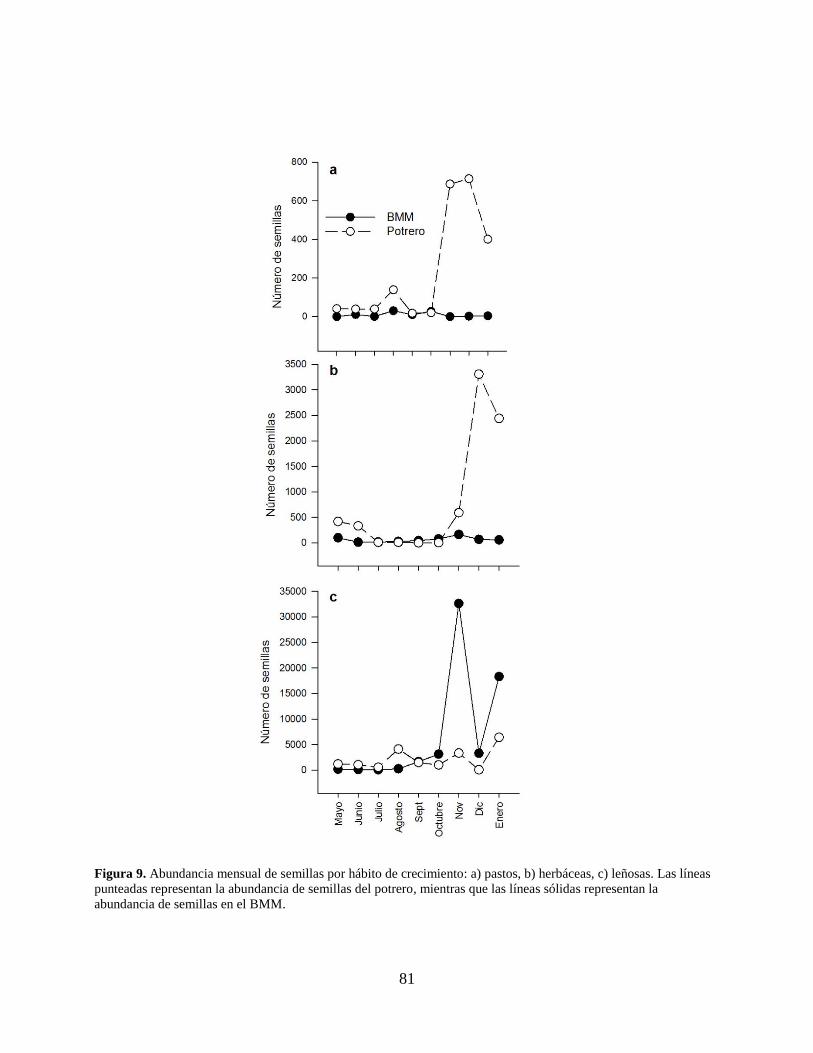
81
Figura 9. Abundancia mensual de semillas por hábito de crecimiento: a) pastos, b) herbáceas, c) leñosas. Las líneas
punteadas representan la abundancia de semillas del potrero, mientras que las líneas sólidas representan la
abundancia de semillas en el BMM.
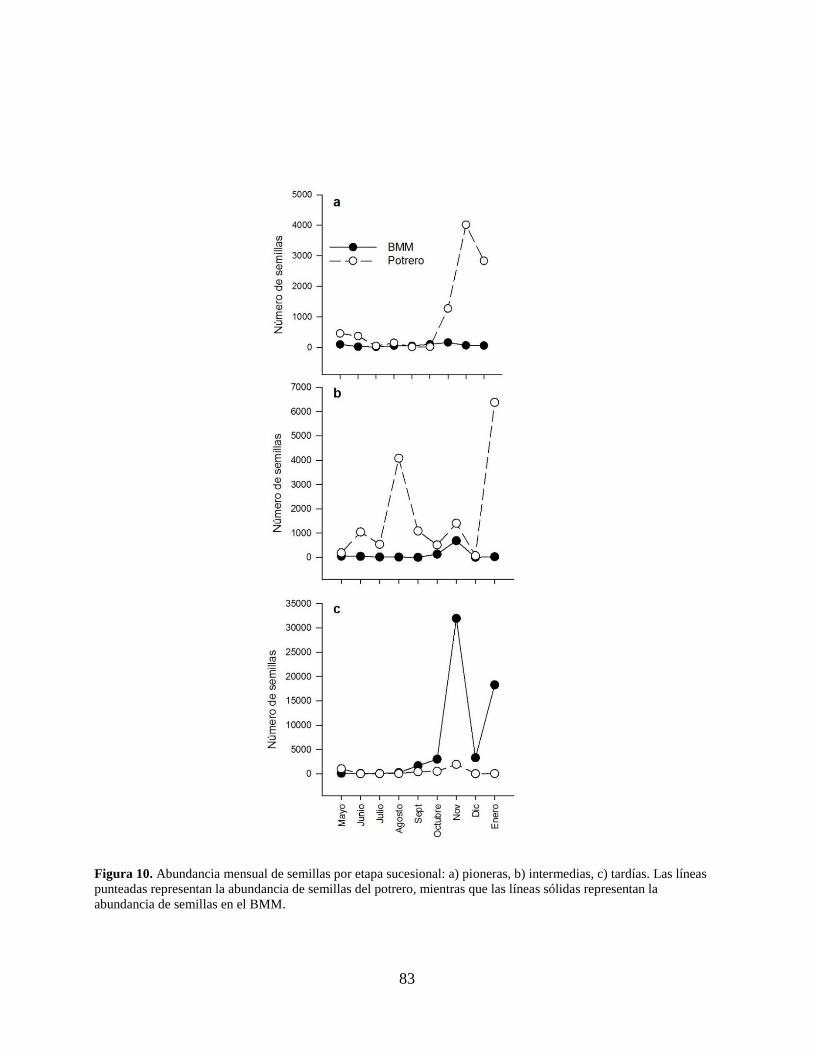
83
Figura 10. Abundancia mensual de semillas por etapa sucesional: a) pioneras, b) intermedias, c) tardías. Las líneas
punteadas representan la abundancia de semillas del potrero, mientras que las líneas sólidas representan la
abundancia de semillas en el BMM.
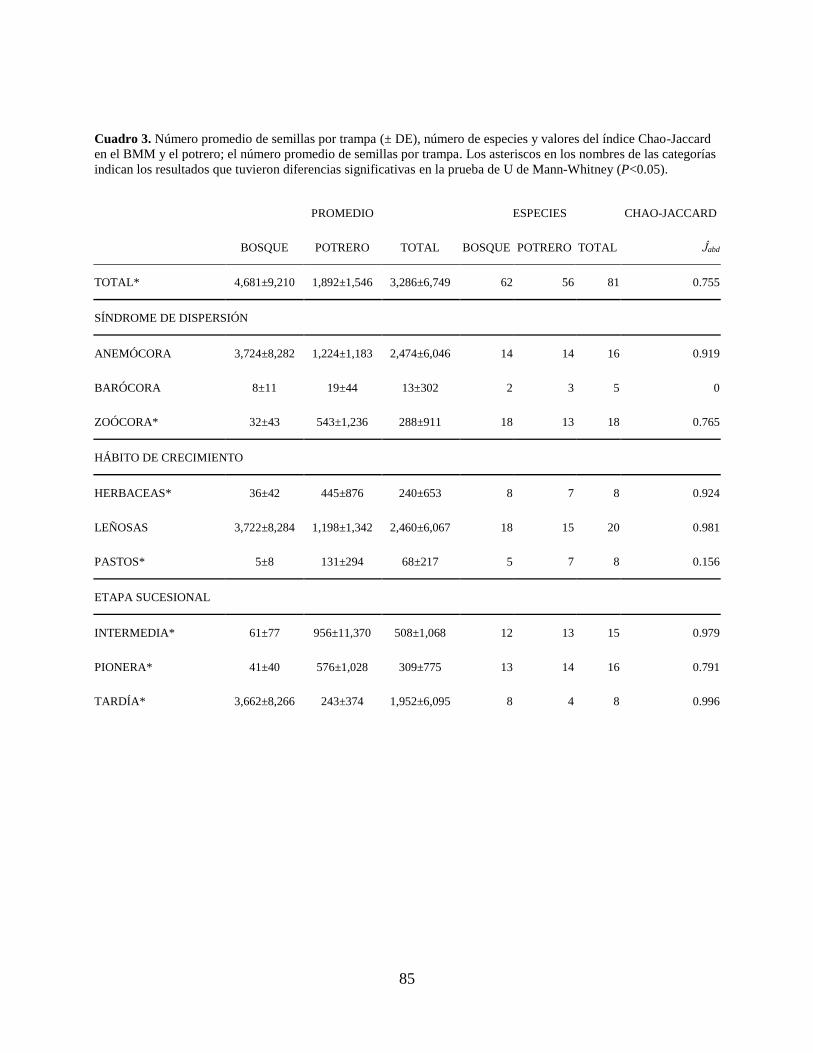
85
Cuadro 3. Número promedio de semillas por trampa (± DE), número de especies y valores del índice Chao-Jaccard
en el BMM y el potrero; el número promedio de semillas por trampa. Los asteriscos en los nombres de las categorías
indican los resultados que tuvieron diferencias significativas en la prueba de U de Mann-Whitney (P<0.05).
PROMEDIO ESPECIES CHAO-JACCARD
BOSQUE POTRERO TOTAL BOSQUE POTRERO TOTAL Ĵabd
TOTAL* 4,681±9,210 1,892±1,546 3,286±6,749 62 56 81 0.755
SÍNDROME DE DISPERSIÓN
ANEMÓCORA 3,724±8,282 1,224±1,183 2,474±6,046 14 14 16 0.919
BARÓCORA 8±11 19±44 13±302 2 3 5 0
ZOÓCORA* 32±43 543±1,236 288±911 18 13 18 0.765
HÁBITO DE CRECIMIENTO
HERBACEAS* 36±42 445±876 240±653 8 7 8 0.924
LEÑOSAS 3,722±8,284 1,198±1,342 2,460±6,067 18 15 20 0.981
PASTOS* 5±8 131±294 68±217 5 7 8 0.156
ETAPA SUCESIONAL
INTERMEDIA* 61±77 956±11,370 508±1,068 12 13 15 0.979
PIONERA* 41±40 576±1,028 309±775 13 14 16 0.791
TARDÍA* 3,662±8,266 243±374 1,952±6,095 8 4 8 0.996
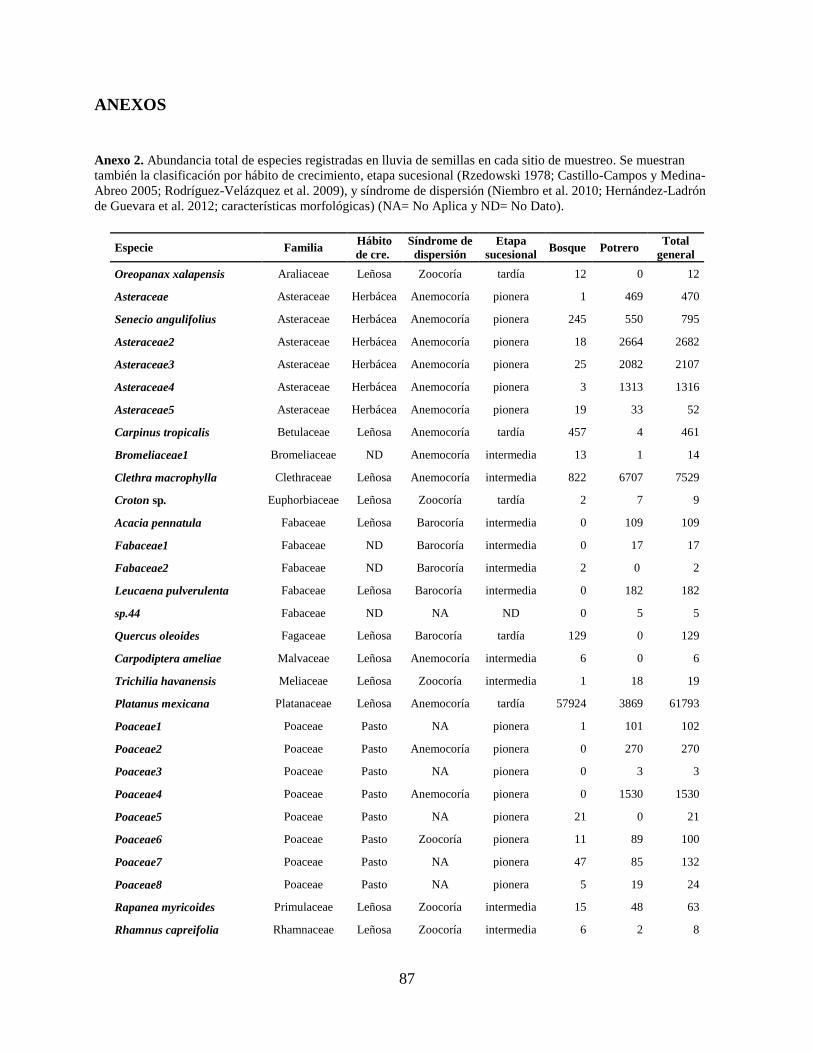
87
ANEXOS
Anexo 2. Abundancia total de especies registradas en lluvia de semillas en cada sitio de muestreo. Se muestran
también la clasificación por hábito de crecimiento, etapa sucesional (Rzedowski 1978; Castillo-Campos y Medina-
Abreo 2005; Rodríguez-Velázquez et al. 2009), y síndrome de dispersión (Niembro et al. 2010; Hernández-Ladrón
de Guevara et al. 2012; características morfológicas) (NA= No Aplica y ND= No Dato).
Especie Familia Hábito
de cre. Síndrome de
dispersión
Etapa
sucesional Bosque Potrero
Total
general
Oreopanax xalapensis Araliaceae Leñosa Zoocoría tardía 12 0 12
Asteraceae Asteraceae Herbácea Anemocoría pionera 1 469 470
Senecio angulifolius Asteraceae Herbácea Anemocoría pionera 245 550 795
Asteraceae2 Asteraceae Herbácea Anemocoría pionera 18 2664 2682
Asteraceae3 Asteraceae Herbácea Anemocoría pionera 25 2082 2107
Asteraceae4 Asteraceae Herbácea Anemocoría pionera 3 1313 1316
Asteraceae5 Asteraceae Herbácea Anemocoría pionera 19 33 52
Carpinus tropicalis Betulaceae Leñosa Anemocoría tardía 457 4 461
Bromeliaceae1 Bromeliaceae ND Anemocoría intermedia 13 1 14
Clethra macrophylla Clethraceae Leñosa Anemocoría intermedia 822 6707 7529
Croton sp. Euphorbiaceae Leñosa Zoocoría tardía 2 7 9
Acacia pennatula Fabaceae Leñosa Barocoría intermedia 0 109 109
Fabaceae1 Fabaceae ND Barocoría intermedia 0 17 17
Fabaceae2 Fabaceae ND Barocoría intermedia 2 0 2
Leucaena pulverulenta Fabaceae Leñosa Barocoría intermedia 0 182 182
sp.44 Fabaceae ND NA ND 0 5 5
Quercus oleoides Fagaceae Leñosa Barocoría tardía 129 0 129
Carpodiptera ameliae Malvaceae Leñosa Anemocoría intermedia 6 0 6
Trichilia havanensis Meliaceae Leñosa Zoocoría intermedia 1 18 19
Platanus mexicana Platanaceae Leñosa Anemocoría tardía 57924 3869 61793
Poaceae1 Poaceae Pasto NA pionera 1 101 102
Poaceae2 Poaceae Pasto Anemocoría pionera 0 270 270
Poaceae3 Poaceae Pasto NA pionera 0 3 3
Poaceae4 Poaceae Pasto Anemocoría pionera 0 1530 1530
Poaceae5 Poaceae Pasto NA pionera 21 0 21
Poaceae6 Poaceae Pasto Zoocoría pionera 11 89 100
Poaceae7 Poaceae Pasto NA pionera 47 85 132
Poaceae8 Poaceae Pasto NA pionera 5 19 24
Rapanea myricoides Primulaceae Leñosa Zoocoría intermedia 15 48 63
Rhamnus capreifolia Rhamnaceae Leñosa Zoocoría intermedia 6 2 8
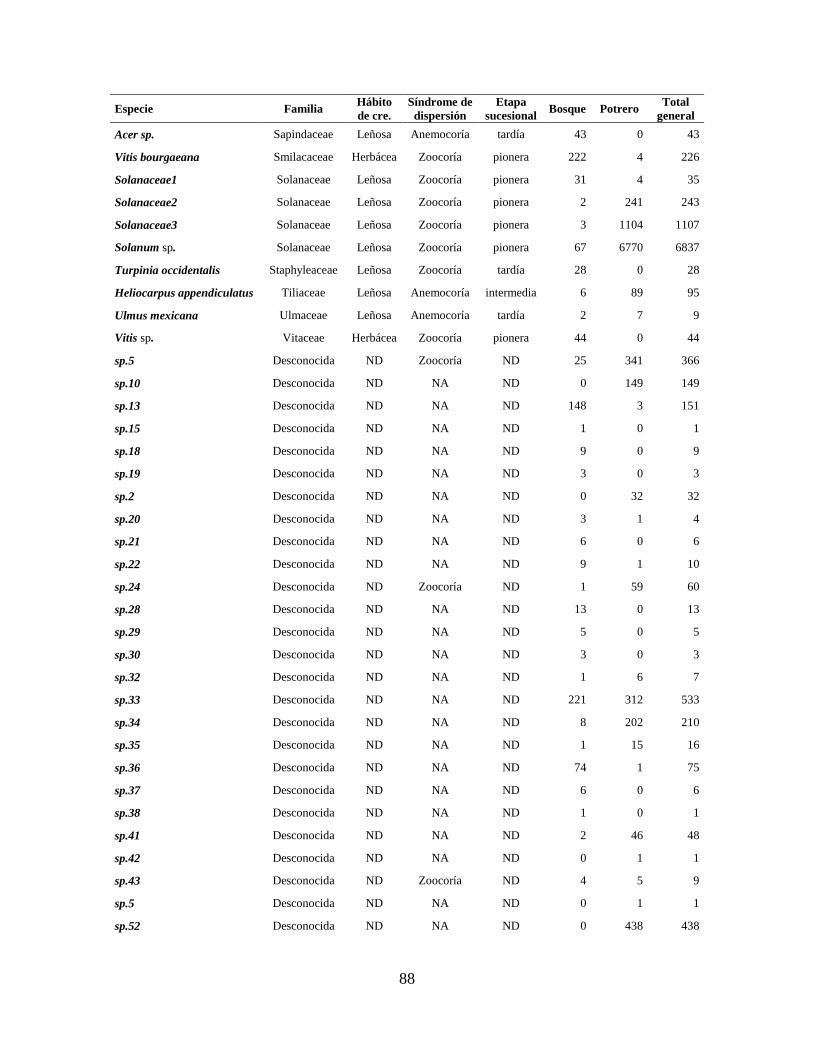
88
Especie Familia Hábito
de cre.
Síndrome de
dispersión
Etapa
sucesional Bosque Potrero
Total
general
Acer sp. Sapindaceae Leñosa Anemocoría tardía 43 0 43
Vitis bourgaeana Smilacaceae Herbácea Zoocoría pionera 222 4 226
Solanaceae1 Solanaceae Leñosa Zoocoría pionera 31 4 35
Solanaceae2 Solanaceae Leñosa Zoocoría pionera 2 241 243
Solanaceae3 Solanaceae Leñosa Zoocoría pionera 3 1104 1107
Solanum sp. Solanaceae Leñosa Zoocoría pionera 67 6770 6837
Turpinia occidentalis Staphyleaceae Leñosa Zoocoría tardía 28 0 28
Heliocarpus appendiculatus Tiliaceae Leñosa Anemocoría intermedia 6 89 95
Ulmus mexicana Ulmaceae Leñosa Anemocoría tardía 2 7 9
Vitis sp. Vitaceae Herbácea Zoocoría pionera 44 0 44
sp.5 Desconocida ND Zoocoría ND 25 341 366
sp.10 Desconocida ND NA ND 0 149 149
sp.13 Desconocida ND NA ND 148 3 151
sp.15 Desconocida ND NA ND 1 0 1
sp.18 Desconocida ND NA ND 9 0 9
sp.19 Desconocida ND NA ND 3 0 3
sp.2 Desconocida ND NA ND 0 32 32
sp.20 Desconocida ND NA ND 3 1 4
sp.21 Desconocida ND NA ND 6 0 6
sp.22 Desconocida ND NA ND 9 1 10
sp.24 Desconocida ND Zoocoría ND 1 59 60
sp.28 Desconocida ND NA ND 13 0 13
sp.29 Desconocida ND NA ND 5 0 5
sp.30 Desconocida ND NA ND 3 0 3
sp.32 Desconocida ND NA ND 1 6 7
sp.33 Desconocida ND NA ND 221 312 533
sp.34 Desconocida ND NA ND 8 202 210
sp.35 Desconocida ND NA ND 1 15 16
sp.36 Desconocida ND NA ND 74 1 75
sp.37 Desconocida ND NA ND 6 0 6
sp.38 Desconocida ND NA ND 1 0 1
sp.41 Desconocida ND NA ND 2 46 48
sp.42 Desconocida ND NA ND 0 1 1
sp.43 Desconocida ND Zoocoría ND 4 5 9
sp.5 Desconocida ND NA ND 0 1 1
sp.52 Desconocida ND NA ND 0 438 438
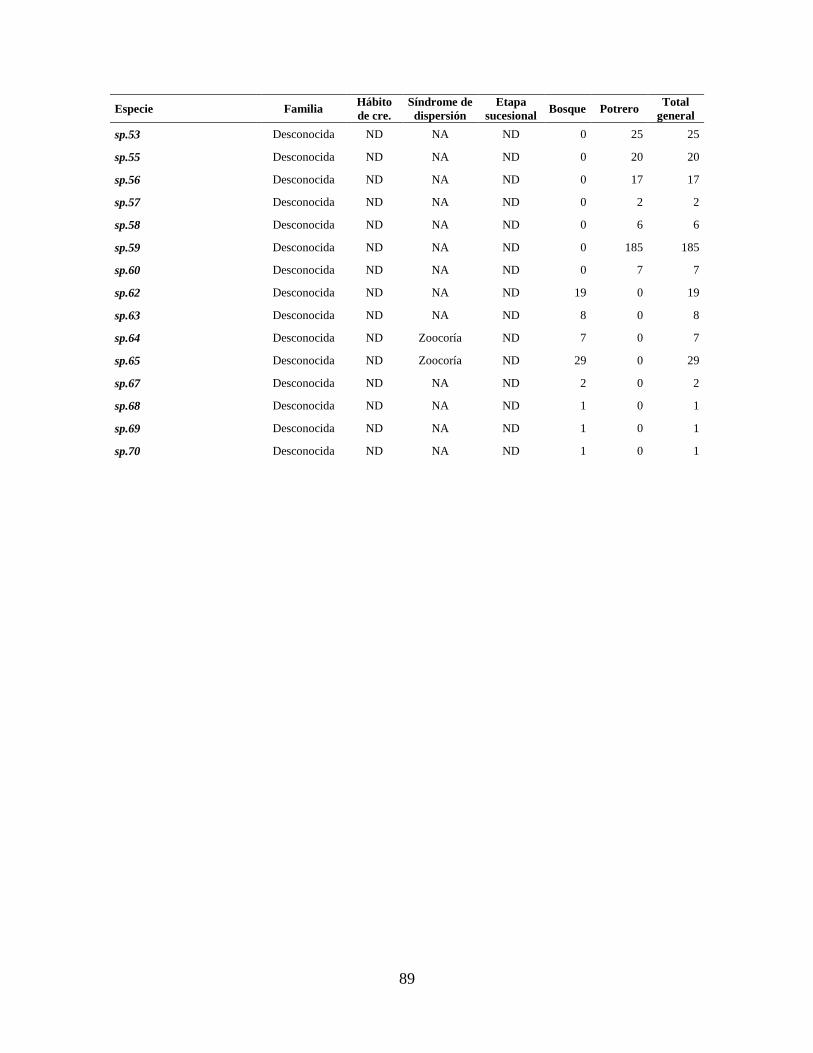
89
Especie Familia Hábito
de cre.
Síndrome de
dispersión
Etapa
sucesional Bosque Potrero
Total
general
sp.53 Desconocida ND NA ND 0 25 25
sp.55 Desconocida ND NA ND 0 20 20
sp.56 Desconocida ND NA ND 0 17 17
sp.57 Desconocida ND NA ND 0 2 2
sp.58 Desconocida ND NA ND 0 6 6
sp.59 Desconocida ND NA ND 0 185 185
sp.60 Desconocida ND NA ND 0 7 7
sp.62 Desconocida ND NA ND 19 0 19
sp.63 Desconocida ND NA ND 8 0 8
sp.64 Desconocida ND Zoocoría ND 7 0 7
sp.65 Desconocida ND Zoocoría ND 29 0 29
sp.67 Desconocida ND NA ND 2 0 2
sp.68 Desconocida ND NA ND 1 0 1
sp.69 Desconocida ND NA ND 1 0 1
sp.70 Desconocida ND NA ND 1 0 1

91
IV. CONCLUSIONES GENERALES

93
CONCLUSIONES GENERALES
1. En general, la riqueza de aves frugívoras y diversidad de plantas dispersadas en el potrero
sugieren que el sitio estudiado tiene un alto potencial de recuperación, ya que la presencia
de aves como vectores de dispersión no son una limitante. Pero de forma particular, las
aves migratorias tienen un efecto positivo en el proceso de dispersión de semillas al
incrementar la riqueza y abundancia de las semillas de especies de plantas zooócoras
durante su estancia en el sitio de estudio. Sin embargo, dada la temporalidad del
fenómeno de migración, probablemente las especies de plantas que se ven más
beneficiadas sean aquellas cuya fructificación se suscita durante este periodo.
2. Aunque la diversidad de plantas consumidas por especie de ave es relativamente baja, la
comunidad de aves frugívoras del potrero en su conjunto dispersa una diversidad
importante de semillas, comparado con lo encontrado en otros trabajos cercanos al área de
estudio. Sin embargo, la riqueza y abundancia de semillas zoócoras encontradas en las
trampas de semillas es mucho mayor a la encontrada en el análisis de las excretas, lo que
nos indica que los vertebrados voladores podrían estar dispersando una mayor diversidad
de semillas, esto es debido principalmente al sesgo que representa el método de muestreo
con redes de niebla, ya que no fueron capturadas aves de gran talla ni murciélagos.
3. Las especies de plantas encontradas en la lluvia de semillas y dispersadas por aves que
podrían catalizar la sucesión del BMM son principalmente las que pertenecen al género
Solanum ya que estas especies leñosas favorecen la misma dispersión fungiendo como
alimento, perchas y resguardo para las aves y otros dispersores. La lluvia de semillas del

94
potrero contiene otras especies de semillas abundantes que son propias del BMM, lo que
potencialmente estarían facilitando el proceso de recuperación, sobre todo por aquellas
especies anemócoras como Platanus mexicana y Clethra macrophylla. En cuanto a las
especies Conostegia xalapensis y Trichilia havanensis, estas son dispersadas de forma
importante por las aves, sin embargo, éstas no presentaron una abundancia importante en
la lluvia de semillas. A pesar de que las especies de semillas que llegan al potrero
representa un factor importante para la recuperación del sitio perturbado, su
establecimiento podría estar siendo limitado por la especies invasora de pasto Cynodon
plectostachyus que reduce el éxito de germinación de las especies importantes para la
recuperación del BMM.
4. La cercanía del potrero con un remanente del BMM tiene una importante influencia en los
procesos sucesionales, ya que la vegetación circundante determina parte de la
composición de la lluvia de semillas. El análisis de la lluvia de semillas sugiere que el
BMM tiene un importante potencial de recuperación, ya que la llegada de semillas al sitio
perturbado no es una limitante para que en el potrero se establezca vegetación propia del
BMM.

95
LITERATURA
Aide, T. M., J. K. Zimmerman, J. B. Pascarella, L. Rivera y H. Marcano-Vega. 2000. Forest
regeneration in a chronosequence of tropical abandoned pastures: Implications for
restoration ecology. Restoration Ecology. 8:328-338.
Aide, T. M., J. K. Zimmerman, M. Rosario y H. Marcano-Vega. 1996. Forest recovery in
abandoned cattle pastures along an elevational gradient in Northeastern Puerto Rico.
Biotropica. 28:537-548.
Aldrich, M., P. Bubb, S. Hostetter, y H. van de Wiel . 2000. Bosques nublados tropicales
montanos. Tiempo para la acción. (Cambridge, Ed.). Inglaterra: WWF
International/IUCN The World Conservation Union. 28 p.
Almeda, F. 1993. Familia Melastomataceae. In Flora del bajío y de regiones adyacentes. Centro
Regional del Bajío, Instituto de Ecología, A. C. México, Pátzcuaro, Mich. Fascículo 10.
Amico, G. C. y M. A. Aizen. 2005. Dispersión de semillas por aves en un bosque templado de
Sudamérica austral: ¿quién dispersa a quién? Ecología Austral. 15:89-100.
Aramburú, R. y V. Corbalán. 2000. Dieta de pichones de cotorra Myiopsitta monachus monachus
(Aves: Psittacidae) en una población Silvestre. Ornitología Neotropical. 11:241-245.
Bedoya-Patiño, J. G., J. V. Estévez-Varón y G. J. Cataño-Villa. 2010. Banco de semillas del
suelo y su papel en la recuperación de los bosques tropicales. Boletín Científico del Museo
de Historia Natural. 14:77-91.
Beltzer, A. H. 1983. Alimentación del “Benteveo” (Pitangus sulphuratus) en el valle aluvial del
río Paraná Medio (Passeriformes: Tyrannidae). Revista de la Asociación de Ciencias
Naturales del Litoral. 14:47-52.

96
Beltzer, A. H. 1985. Ecología alimentaria de Aramides ypecaha (Aves: Rallidae) en el valle
aluvial del río Paraná medio (Argentina). Revista de la Asociación de Ciencias Naturales
del Litoral. 16:73-83.
Beltzer, A. H. 1995. Biología alimentaria del Pirincho Guira guira (Aves: Cuculidae) en el valle
aluvial del río Paraná medio, Argentina. Revista de Ecología Latinoamericana. 2:13-18.
Carranza, E. 1994. Familia Platanaceae. In Flora del bajío y de regiones adyacentes. Centro
Regional del Bajío, Instituto de Ecología, A. C. México, Pátzcuaro, Mich. Fascículo 23.
Castillo-Campos, G. 1991. Vegetación y flora del municipio de Xalapa. Programa del Hombre y
la Biósfera (MAB, UNESCO) Instituto de Ecología, A.C. H. Ayuntamiento de Xalapa,
Veracruz. 148 p.
Castillo-Campos, G. y M. E. Medina-Abreo. 2005. Árboles y arbustos de la Reserva Natural de
La Mancha, Veracruz, México. 1era. Ed., 1era. reimp. Instituto de Ecología, A.C. Xalapa,
Veracruz. 143 p.
Challenger, A. 1998. Utilización y conservación de los ecosistemas terrestres de México: pasado,
presente y futuro. México, D.F. CONABIO. 847 p.
Chao, A., R. L. Chazdon, R. K. Colwell y T-J. Shen. 2005. A new statistical approach for
assessing similarity of species composition with incidence and abundance data. Ecology
Letters. 8:148-159.
Cole, R.J., K. D. Holl y R. A. Zahawi. 2010. Seed rain under tree islands planted to restore
degraded lands in a tropical agricultural landscape. Ecological Applications. 20:1255-
1269.
Colwell, R.K. 2013 EstimateS: Statistical estimation of species richness y shared species from
samples. Version 9. Persistent URL <purl.oclc.org/estimates>

97
Connell, J. H., J. G. Tracey y L. J. Webb. 1984. Compensatory recruitment, growth and mortality
as factors maintaining rain forest tree diversity. Ecological Monographs. 54:141-164.
Corlett, R.T. 1998. Frugivory and seed dispersal by vertebrates in the Oriental (Indomalayan)
Region. Biological Reviews. 73:413-448.
Cruz, A., F. Escobar, P. Gerez, M. A. Muñíz, F. Ramírez, G. Williams y CONABIO. 2010. V.
Centro de Veracruz in El Bosque Mesófilo de Montaña en México: Amenazas y
Oportunidades para su Conservación y Manejo Sostenible. 80–87p. México, D.F.,
México: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad.
Cubiña, A. y T. M. Aide. 2001. The effect of distance from forest edge on seed rain and soil seed
bank in a tropical pasture. Biotropica. 33:260-267.
Dalling, J. W., M. D. Swaine y N. C. Garwood. 1998. Dispersal patterns and seed bank dynamics
of pioneer trees in moist tropical forest. Ecology. 79:564-578.
Del Castillo, R. F. y M. A. Pérez Ríos. 2008. Changes in seed rain during secondary succession
in a tropical montane cloud forest region in Oaxaca, Mexico. Journal of Tropical Ecology.
24:433-444.
Denslow, J. S., y S. Guzmán. 2000. Variation in stand structure, light and seedling abundance
across moist forest chronosequence, Panama. Journal of Vegetation Science. 11:201-212.
Doumenge C., Gilmour D., Ruiz Perez M. y Blockhus J. 1995. Tropical montane cloud forests:
conservation status and management issues. In: Hamilton L.S., Juvik J.O. y Scatena F.N.
(eds), Tropical Montane Cloud Forests. Springer-Verlag, New York, 24-37 p.
Estrada A. y T.H. Fleming (ed.). 1986. Frugivores and seed dispersal. Tasks for Vegetation
Science 15. Dordrecht: W. Junk.
Estrada, A. y R. Coates-Estrada. 1996. Tropical rain forest fragmentation and wild populations of
primates at Los Tuxtlas, México. International Journal of Primatology. 17:759-783.

98
Estrada, A., R. Coates-Estrada y D. Meritt Jr. 1993. Bat species richness and abundance in
tropical rain forest fragments and in agricultural habitats at Los Tuxtlas, Mexico.
Ecography. 16:309-318.
Figueroa, J. A. y S. A. Castro. 2002. Effects of bird ingestion on seed germination of four woody
species of the temperate rain forest of Chiloé Island, Chile. Plant Ecology. 160:17-23.
Figueroa-Esquivel, E. M. y F. Puebla-Olivares. 2009. Importancia de un árbol tropical
(Dendropanax arboreus) para aves migratorias neárticas en México. Ornitología
Neotropical. 20:391-399.
Figueroa-Esquivel, E. M., F. Puebla-Olivares, H. Godínez-Álvarez y J. Núñez-Farfán. 2009. Seed
dispersal effectiveness by understory birds on Dendropanax arboreus in a fragmented
landscape. Biodiversity and Conservation. 18:3357–3365.
Galindo-González J., S. Guevara y V. J. Sosa. 2000. Bat- and bird-generated seed rains at
isolated trees in pastures in a tropical rainforest. Conservation Biology. 14:1693-1703.
Gálvez, J. 2002. La restauración ecológica: conceptos y aplicaciones. Universidad Rafael
Landivar- Instituto de Agricultura, Recursos Naturales y Ambiente. 8:1-23.
Gómez-Pompa, A., T. Krömer y R. Castro-Cortés. 2010. Atlas de la flora de Veracruz: Un
patrimonio natural en peligro. Comisión del Estado de Veracruz para la Conmemoración
de la Independencia Nacional y la Revolución Mexicana-Universidad Veracruzana. 530 p.
González-Espinosa, M., J.A. Meave, N. Ramírez-Marcial, T. Toledo-Aceves, F.G. Lorea-
Hernández y G. Ibarra-Manríquez. 2012. Los bosques de niebla de México: conservación
y restauración de su componente arbóreo. Ecosistemas. 21:36-54.
González-García, F., R. Straub, J. A. Lobato-García e I. MacGregor-Fors. 2014. Birds of a
neotropical green city: an up-to-date review of the avifauna of the city of Xalapa with
additional unpublished records. Urban Ecosystems. DOI 10.1007/s11252-014-0370-3.

99
González-Villareal, L. M. 1996. Familia Clethraceae. In Flora del bajío y de regiones
adyacentes. Centro Regional del Bajío, Instituto de Ecología, A. C. México, Pátzcuaro,
Mich. Fascículo 47.
Gorchov, D. L., F. Cornejo, C. F. Acorra y M. Jaramillo. 1995. Dietary overlap between
frugivorous birds and bats in the Peruvian Amazon. Oikos. 74:235-250.
Gorchov, D. L., F. Cornejo, C. F. Ascorra y M. Jaramillo. 1993. The role of seed dispersal in the
natural regeneration of rain forest after strip-cutting in the Peruvian Amazon. Vegetatio.
107/108:339:349.
Grubb, P. J. 1977. The maintenance of species-richness in plant communities: the importance of
the regeneration niche. Biological Reviews. 52:107-145.
Guevara, S. y J. Laborde. 1993. Monitoring seed dispersal at isolated standing trees in tropical
pastures: consecuences for local species availability. Vegetatio. 107:319-338.
Guevara, S., J. Meave, P. Moreno-Casasola y J. Laborde. 1992. Floristic composition and
structure of vegetation under isolated trees in neotropical pastures. Journal of Vegetation
Science. 3:655-664.
Hammer, O., D. A. T. Harper y P. D. Ryan. 2001. PAST: Paleontological Statistics software
package for education and data analysis. Paleontological Electronica. 4:9.
Henderson, C.B., K. E. Petersen y R. A. Redak. 1988. Spatial and temporal patterns in the seed
bank and vegetation of a desert grassland community. Journal of Ecology. 76:717-728.
Hernández-Ladrón de Guevara, I., O. R. Rojas-Soto, F. López-Barrera, F. Puebla-Olivares y C.
Díaz-Castelazo. 2012. Dispersión de semillas por aves en un paisaje de bosque mesófilo
en el centro de Veracruz, México: Su papel en la restauración pasiva. Revista Chilena de
Historia Natural. 85:89-100.

100
Herrera, C. 1984. A study of avian frugivores, bird-dispersed plants, and their interaction in the
Mediterranean scrublands. Ecological Monographs. 54:1-23
Herrera, C. M. 1988. Variaciones anuales en las poblaciones de pájaros frugívoros y su relación
con la abundancia de frutos. Ardeola. 35:135-142.
Herrera, C. M., P. Jordano, L. López-Soria y J. A. Amat. 1994. Recruitment of a mast-Fruiting,
bird-dispersed tree: bridging frugivore activity and seedling establishment. Ecological
Monographs. 64:315-344.
Holl, K. D. 1998. Do bird perching structures elevate seed rain and seedling establishment in
abandoned tropical pasture? Restoration Ecology. 6:253-261.
Holl, K. D. 1999. Factors limiting tropical rain forest regeneration in abandoned pasture: seed
rain, seed germination, microclimate, and soil. Biotropica. 31:229-242.
Holl, K. D. 2002. Effect of shrubs on tree seedling establishment in an abandoned tropical
pasture. Journal of Ecology. 90:179-187.
Holl, K. D., M. E. Loik, E. H. V. Lin y I. A. Samuel. 2000. Tropical montane forest restoration in
Costa Rica: overcoming barriers to dispersal and establishment. Restoration Ecology.
8:339–349.
Howe, H. F. 1977. Bird Activity and Seed dispersal of a tropical wet forest tree. Ecology. 58:539-
550.
Howe, H. F. 1984a. Implications of seed dispersal by animals for tropical reserve management.
Biological Conservation. 30:261-281.
Howe, H. F. 1984b. Constraints on the evolution of mutualisms. The American Naturalist.
123:764-777.
Howe, H. F. y J. Smallwood. 1982. Ecology of seed dispersal. Annual Review of Ecology and
Systematics. 13:201-228.

101
Howell, S. N. G., y Webb, S. 1995. The Birds of Mexico and Northern Central America. A Guide
to: The Birds of Mexico and Northern Central America. Oxford, United Kingdom: Oxford
University Press.
Hughes, J. W. y T. J. Fahey. 1988. Seed Dispersal and Colonization in a Disturbed Northern
Hardwood Forest. Bulletin of the Torrey Botanical Club. 115:89-99.
Hyatt, L. A., M. S. Rosenberg, T. G. Howard, G. Bole, W. Fang, J. Anastasia, K. Brown, R.
Grella, K. Hinman, J. P. Kurdziel, y J. Gurevitch. 2003. The distance dependence
prediction of the Janzen-Connell hypothesis: a meta-analysis. Oikos. 103:590-602
Ingle, N. R. 2003. Seed dispersal by wind, birds, and bats between Philippine montanae rainforest
and successional vegetation. Oecología. 134:251-261.
Jackson LL., N. Lopoukhine y D. Hillyard. 1995. Ecological restoration: A definition and
comments. Restoration Ecology. 3:71-75.
Janzen, D. H. 1970. Herbivores and the number of tree species in tropical forest. The American
Naturalist. 104:501-528
Jordano, P. 2000. Fruits and Frugivory. 125-166 p. in M. Fenner (ed.), editor. Seeds: the ecology
of regeneration in plant communities, 2nd edition. CABI Publ., Wallingford, UK.
Laborde, J., S. Guevara y G. Sánchez-Ríos. 2008. Tree and shrub seed dispersal in pastures: The
importance of rainforest trees outside forest fragments. Écoscience. 15:6-16.
Lamb, D. y D. Gilmour. 2003. Rehabilitation and Restoration of Degraded Forests. IUCN,
Gland, Switzerland and Cambridge, UK and WWF, Gland, Switzerland. 110 p.
Lamb, D., P. D. Erskine, y J. A. Parrotta. 2005. Restoration of degraded tropical forest
landscapes. Science. 310:1628–1632.
Levassor, C., M. Ortega y B. Peco. 1990. Seed bank dynamics of Mediterranean pastures
subjected to mechanical disturbance. Journal of Vegetation Science. 1:339-344.

102
Manson, R. H. 2004. Los servicios hidrológicos y la conservación de los bosques en México.
Madera y Bosques. 10:3-20.
Marlette, G. M. y J. E. Anderson. 1986. Seed Banks and propagule dispersal in crested-
Wheatgrass stands. Journal of Applied Ecology. 23:161-175.
Martínez, M. L., O. Pérez-Maqueo, G. Vázquez, G. Castillo-Campos, J. García-Franco, K.
Mehltreter, M. Equihua y R. Landgrave. 2009. Effects of land use change on biodiversity
and ecosystem services in tropical montane cloud forest of Mexico. Forest Ecology and
Management. 258:1856-1863.
Martínez-Garza, C., M. Osorio-Beristain, D. Valenzuela-Galván y A. Nicolás-Medina. 2011.
Intra and inter-annual variation in seed rain in a secondary dry tropical forest excluded
from chronic disturbance. Forest Ecology and Management. 262:2207-2218.
Martínez-Morales, M.A. 2007. Avifauna del bosque mesófilo de montaña del noreste de Hidalgo.
Revista Mexicana de Biodiversidad. 78:149-162.
Muñiz-Castro M. A., G. Williams-Linera y J. M. Rey-Benayas. 2006. Distance effect from cloud
forest fragments on plant community structure in abandoned pastures in Veracruz,
Mexico. Journal of Tropical Ecology. 22:431-440.
Muñiz-Castro, M. A., 2008. Sucesión secundaria y establecimiento de especies arbóreas nativas
para restauración de Bosque Mesófilo de Montaña en potreros abandonados del centro
de Veracruz. Tesis de Doctorado. Instituto de Ecología, A. C.
Nee, M. 1986. Familia Solanaceae. In Flora de Veracruz. Instituto de Ecología, A. C. México,
Xalapa, Ver. Fascículo 49
Niembro, A., M. Vázquez, y O. Sánchez. 2010. Árboles de Veracruz: 100 especies para la
reforestación estratégica. Comisión del Estado de Veracruz para la Conmemoración del

103
Bicentenario de la Independencia Nacional y el Centenario de la Revolución Mexicana,
Xalapa, México. 253 p.
Olguín P. F., A. M. Attademo, M. Quiroga, E. León y A. H. Beltzer. 2012. Biología alimentaria
de la pollona pintada Porphyriops melanops (Aves: Rallidae) en el valle de inundación
del río Paraná medio, Argentina. Revista nodo. 7:85-92.
Ortega-Pieck, A., F. López-Barrera, N. Ramírez-Marcial y J. G. García-Franco. 2011. Early
seedling establishment of two tropical montane cloud forest tree species: The role of
native and exotic grasses. Forest Ecology and Management. 261:1336–1343.
Ortiz-Pulido, R., J. Laborde y S. Guevara. 2000. Frugivoría por aves en un paisaje fragmentado:
Consecuencias en la dispersión de Semillas. Biotropica. 32:473-488.
Palmeirim, J. M., D. L. Gorchoy y S. Stoleson. 1989. Trophic structure of a neotropical frugivore
community: is there competition between birds and bats? Oecologia. 79:403-411.
Peña-Domene, M., C. Matínez-Garza y H. F. Howe. 2013. Early recruitement dynamics in
tropical restoration. Ecological Applications. 23:1124-1134.
Peterson, R. T., E. L. Chalif, M. Ramos y M. I. Castillo. 2008. Aves de México: Guía de Campo.
Distrito Federal, México: Diana. 473 p.
Pinkas, L., M. S. Oliphant y L. K. Iverson. 1971. Food habits of albacore, bluefin tuna, and
bonito in California waters. The Resources Agency Department of Fish and Game. Fish
Bulletin 152.
http://content.cdlib.org/view?docId=kt8290062w&brand=calisphere&doc.view=entire_te
xt
Pla, L. 2006. Biodiversidad: inferencia basada en el índice de Shannon y la riqueza. Interciencia.
31:583-590.

104
Posada, J. M., J. M. Aide y J. Cavelier. 2000. Cattle and weedy shrubs as restoration tools of
tropical montane rain forest. Restoration Ecology. 8:370-379.
Purata, S. 1986. Floristic and structural changes during old-field succession in the Mexican
tropics in relation to site history and species availability. Journal of Tropical Ecology.
2:257–276.
Quesada, M., G. A. Sanchez-Azofeifa, M. Alvarez-Añorve, K. E. Stoner, L. Avila-Cabadilla, J.
Calvo-Alvarado, A. Castillo, M. M. Espírito-Santo, M. Fagundes, G. W. Fernandes, J.
Gamon, M. Lopezaraiza-Mikel, D. Lawrence, L. P. C. Morellato, J. S. Powers, F. D. S.
Neves, V. Rosas-Guerrero, R. Sayago, y G. Sanchez-Montoya. 2009. Succession and
management of tropical dry forests in the Americas: Review and new perspectives. Forest
Ecology and Management. 258:1014-1024.
Ralph, J., G. Geupel, P. Pyle, T. Martin, D. DeSante, y B. Milá. 1996. Manual de métodos de
campo para el monitoreo de aves terrestres. Manual de métodos de campo para el
monitoreo de aves terrestres. USDA Forest Service Gen. Tech. Rep. PSW-GTR-159.
Ramírez-Marcial, N., M. González-Espinoza y P. F. Quintana-Ascencio. 1992. Banco y lluvia de
semillas en comunidades sucesionales de bosques de pino-encino en Los Altos de
Chiapas, México. Acta Botánica Mexicana. 20:59-75.
Reid, J. L., C. D. Mendenhall, J. A. Rosales, R. A. Zahawi y K. D. Holl. 2014. Landscape context
mediates avian habitat choise in Tropical Forest Restoration. Plos One. 9:1-8.
Ridley, H.N. 1930. The dispersal of plants throughout the world. Reeve, Ashford.
Rodríguez-Santamaría, M. F., J. M. Puentes-Aguilar y F. Cortés-Pérez. 2006. Caracterización
temporal de la lluvia de semillas en un bosque nublado del Cerro de Mamapacha (Boyacá-
Colombia). Revista de la Academia Colombiana de Ciencias. 30:619-624.

105
Rodríguez-Velázquez, J., P. Sinaca-Colin y G. Jamangapá-García. 2009. Frutos y semillas de
árboles tropicales de México. INE-SEMARNAT. 123 p.
Román-Dañobeytia, F. J., J. Castellanos-Albores, S. I. Levy-Tacher, J. Aronson, N. Ramírez-
Marcial y R. Ribeiro-Rodrigues. 2012. Responses of transplanted native tree species to
invasive alien grass removals in an abandoned cattle pasture in the Lacandon region,
Mexico. Tropical Conservation Science. 5:192–207.
Ruelas-Inzunza, E. 2006. La migración de las aves. In Moreno-Casasola, P. (ed), Entornos
Veracruzanos: La costa de La Mancha. Instituto de Ecología, A. C., Xalapa, Veracruz,
Mexico, 448-460p.
Rzedowski, G. 1993. Familia Meliaceae. In Flora del bajío y de regiones adyacentes. Centro
Regional del Bajío, Instituto de Ecología, A. C. México, Pátzcuaro, Mich. Fascículo 10.
Rzedowski, J. 1978. Vegetación de México. Limusa. México, D.F. 431 p.
Rzedowski, J., 2006.Vegetación de México. 1ra. Edición digital, Comisión Nacional para el
Conocimiento y Uso de la Biodiversidad, México, 504 p.
Salvande, M., J.A. Figueroa y J. J. Arnesto. 2011. Quantity component of the effectiveness of
seed dispersal by birds in the temperate rainforest of Chiloé, Chile. Bosque. 32:39-45.
Sibley, D. A. 2000. The Sibley Guide to Birds. New York, New York, United States: Alfred A.
Knopf, Inc. 545 p.
Snow, D. W. 1965. A possible selective factor in the evolution of fruiting seasons in tropical
forest. Oikos. 15:274-281.
Tiffney, B. H. 2004. Vertebrate dispersal of seed plants through time. Annual Review of Ecology,
Evolution, and Systematics. 35:1-29.

106
Toledo-Aceves, T. J. A. Meave, M. González-Espinoza y N. Ramírez-Marcial. 2011. Tropical
montane cloud forest: Current threats and opportunities for their conservation and
sustainable management in Mexico. Journal of Environmental Management. 92:974-981.
Van der Pijl, L. 1972. Principles of dispersal in higher plants. 1st Ed. Springer-Verlag, Berlin.
Vázquez-Balbuena, D. 2014. Deposición de semillas zoócoras en un mosaico forestal
heterogéneo del trópico seco de Veracruz y su relación con la avifauna frugívora y la
estructura de la vegetación. Tesis de Maestría. Instituto de Ecología, A. C.
Viana, D. S., L. Santamaría, T. C. Michot y J. Figuerola. 2013. Allometric scaling of long-
distance seed dispersal by migratory birds. The American Naturalist. 181:649-662.
Vieira, D. L. M., y A. Scariot. 2006. Principles of Natural Regeneration of Tropical Dry Forests
for Restoration. Restoration Ecology. 14:11-20.
Villa-Bonilla, B., O. R. Rojas-Soto, A. G. Colodner-Chamudis y C. Tejeda-Cruz. 2008.
Inventarios municipales de avifauna y su aplicación a la conservación: El caso de
Zacapoaxtla, Puebla, México. Ornitología Neotropical. 19:531-551.
Wang, B. C. and T. B. Smith. 2002. Closing the seed dispersal loop. Trends in Ecology and
Evolution. 17:379-385.
Williams-Linera, G. 2002. Tree species richness complementarity, disturbance and fragmentation
in a Mexican tropical montane cloud forest. Biodiversity and Conservation. 11:1825-
1843.
Williams-Linera, G. 2012. El bosque de niebla del centro de Veracruz: ecología, historia y
destino en tiempos de fragmentación y cambio climático. CONABIO- Instituto de
Ecología, A.C., Xalapa, Veracruz, México. 208 p.

107
Williams-Linera, G., R. H. Manson y E. Isunza. 2002. La fragmentación del bosque mesófilo de
montaña y patrones de uso del suelo en la región oeste de Xalapa. Veracruz, México.
Madera y Bosques. 8:73-89.
Wunderle, J. M. 1997. The role of animal seed dispersal in accelerating native forest regeneration
on degraded tropical lands. Forestry Ecology and Management. 99:223-235.





























