Embed Size (px)
Citation preview

LEARNING AND MOTIVATION 7. 540-550 ( 1976)
Latent Inhibition: The Effects of ITI and CS Intensity during Preexposure
PAUL SCHNUR
Fordham UniversitylLincoln Center
AND
R.E. LUBOW
Tel-Aviv University, Tel-Aviv, Israel
Two experiments are reported which test the effects of intertrial interval (ITI) and stimulus intensity during nonreinforced preexposure in the latent inhibition paradigm. In Experiment 1, using mice in an escape-avoidance task, latent inhibition was a direct function of the duration of stimulus intertrial interval during preexposure. In Experiment 2, using rats in a CER task, latent inhibition was a direct function of stimulus intensity during preexposure. These findings are discussed in terms of equivalent effects which have been reported in the habituation paradigm.
A stimulus that is repeatedly presented in the absence of reinforcement eventually loses its functional impact. This effect has been observed in two related paradigms which differ in regard to conditions of response measurement. In the habituation paradigm, the probability of occurrence or the amplitude of an identifiable response is observed to decline as a function of stimulus presentation. In the latent inhibition paradigm, conditioning is retarded following nonreinforced presentations of the to-be-conditioned stimulus. Despite the difference in techniques of response measurement, the operational similarity of the two paradigms is evident, and some investigators (e.g., Ackil, Mellgren, Halgren, & Frommer, 1969; Carlton & Vogel, 1967; Lantz, 1973; Schnur, 1971) have suggested that common processes might underlie the two phenomena.
In the present experiments, the latent inhibition paradigm is used to measure the effects of varying two parameters of nonreinforced pre- exposure: stimulus intensity and the interval between successive pre- exposures, the ITI. These parameters have been investigated thoroughly
This research was supported by a National Science Foundation Grant, GB-33682, to the first author, and by an Israel Trustees Foundation Grant, 2958, to the second author. The assistance of Eli Blaustein, Jay Harburger, Dona Garrett, Molly Logan, Jill Rawnsley. and Dahlia Shabat is gratefully acknowledged. Requests for reprints should be sent to Paul Schnur, Division of Social Sciences, Fordham University at Lincoln Center. New York, New York 10023.
540 Copyright 0 1976 by Academic Press. Inc. All rights of reproductmn in any form reserved.

LATENT INHIBITION 541
within the habituation paradigm and their effects are well documented in a variety of species and response systems (Carew, Pinsker, & Kandel, 1972; Graham, 1973; Thompson, Groves, Teyler, & Roemer, 1973; Thompson & Spencer, 1966). The purpose of the present study is to determine whether the stimulus intensity and IT1 during preexposure effects, as measured in the latent inhibition paradigm, are parallel to such effects as measured in the habituation paradigm. To the extent that data parallels are found to exist between these two paradigms, the hypothesis that habituation and latent inhibition are based on common processes would be strengthened.
EXPERIMENT 1
In a review of the habituation literature, Thompson and Spencer (1966) concluded that habituation increases as an inverse function of ITI: The shorter the interval between successive stimulus presentations, the more rapid and/or more pronounced is the habituation. However, this inverse relation between IT1 and habituation is obtained only under certain conditions. That is, if the IT1 is constant over a series of habituation trials, then a group with shorter IT1 will show more habitua- tion than a group with longer ITI. Although this is the design most commonly used in habituation studies, if one considers that each stimulus presentation is both a test trial to measure habituation up to that point and a training trial for producing further habituation, this design con- founds the effects of training trial IT1 with those of test trial IT1 (Davis, 1970). To separate these effects, Davis (1970) gave two groups of rats habituation training trials with either a 2- or 16-set ITI. Subsequently, both groups were given separate test trials at ITIs of 2, 4, 8, and 16 sec. During training, startle responsiveness declined more for the 2-set IT1 group than for the 16-set IT1 group. On test trials, however, startle responsiveness was lower at each test interval for the group trained at an IT1 of 16 set than for the group trained at an IT1 of 2 sec. In short, habituation varies inversely with IT1 when the manipulation of IT1 and the measurement of its effects covary; when manipulation and measure- ment are separated, habituation is a direct function of ITI.
In the latent inhibition paradigm, habituation training trials (pre- exposure) are separated from habituation test trials (conditioning). It is possible, therefore, to evaluate the effects of preexposure IT1 independ- ently of the effects of IT1 during conditioning. In one such study, Crowell and Anderson (1972, Experiment 2) found no differences in latent inhibition as a function of IT1 with a CER procedure. In another, Lantz (1973, Experiment 2) reported a direct relation between preexposure IT1 and latent inhibition. However, the interpretation of the latter experiment is complicated by a number of factors. First, response suppression was measured after tone offset rather than during the tone

542 SCHNUR AND LUBOW
CS. Furthermore, both the amount of apparatus preexposure and the interval between preexposure and conditioning were confounded with preexposure ITI.
The purpose of the present experiment is to provide another evaluation of the effects of preexposure IT1 when IT1 during conditioning is constant. The latent inhibition paradigm is employed and two levels of ITI, 10 and 70 set, are manipulated factorially between the preexposure and conditioning phases. On the basis of the effects of IT1 in the habitua- tion paradigm (Davis, 1970), it is expected that latent inhibition will vary directly with preexposure ITI.
Method
Subjects. Seventy-two male mice of the BALB strain, obtained from the Weizman Institute, were randomly assigned to six groups of 12 mice each. The mice were 30 days of age upon arrival at the laboratory. They were maintained four to a cage with ad libitum food and water. The average age at the beginning of the experiment was 60 days.
Apparatus. The basic apparatus consisted of a commercial mouse shuttlebox 27 x 11 x 19 cm (I x w x h) with grid of 2.5mm-diameter rods, 6.5 mm apart center-to-center, and a scrambled shock source. Pro- gramming and timing of experimental events were accomplished with solid-state modules. Responses and latencies were recorded on an event recorder. The shuttlebox was housed in a sound attenuating chamber with an ambient noise level produced by an exhaust fan of approximately 60 db SPL (20 pN/m2). The CS was a tone generated by a Sonalert unit located on the ceiling of the shuttlebox, midway between the two compartments. The tone was 2.8 kHz, at 72 db SPL (20 pN/m2).
Procedure. The procedure was divided into two phases, preexposure and acquisition. During the preexposure phase, all animals were placed individually in the shuttlebox for 50 min. Twenty-four mice received 40 preexposures to the to-be-CS with a 70-set interval between exposures (Long preexposure IT1 group, L); 24 mice received 40 preexposures to the to-be-CS with a lo-set interval between exposures (Short preexposure IT1 group, S). The first preexposure for Group L began immediately after the animal was placed in the apparatus. The first preexposure for Group S began 40 min after the animal was placed in the apparatus. A third group of 24 mice received no exposure to the to-be-C& Group C. The duration of the tone in the preexposure phase was 5 sec.
The acquisition phase was begun immediately after the preexposure phase. Each of three preexposure groups. (L, S, C) was subdivided. Half of each group was tested on escape-avoidance acquisition with a 70-set IT1 (1) and half with a IO-set IT1 (s).
Thus, the experimental design was a 3 x 2 factorial, none or 40 pre- exposures, where the 40 preexposures were presented with either a

LATENT INHIBITION 543
short or long ITI, and two levels of acquisition, either short IT1 or long ITI. The six groups are designated as Ll, Ls, Sl, Ss, Cl, and Cs.
During the escape-avoidance acquisition phase, the CS was presented for 5 set without shock, after which it continued with shock (1 mA) until the animal crossed from one compartment into the other. When this occurred, tone and shock terminated together. If the mouse did not cross within 30 set from tone onset, the trial was terminated manually by turning off the tone and shock. Both for procedural purposes and data analysis, this was treated as though the response had been made by the subject. If the subject responded within 5 set of tone onset, the tone ended and there was no shock. ITI was measured from tone onset to tone onset in both phases.
All subjects were run for 60 trials. Two dependent measures were taken: latency of the crossing response and number of avoidance responses.
Results
The mean percentage of avoidance responses and the mean latency of response are shown in Table 1. It is clear that subjects receiving a long IT1 during acquisition (Groups Cl, Sl, Ll) learned more than subjects receiving a short IT1 during acquisition (Groups Cs, Ss, Ls) independently of preexposure conditions. It would appear that short acquisition ITIs produced little or no learning. This is substantiated by a 3 x 2 analysis of variance (ANOVA) on the total number of avoidance responses which revealed a significant main effect of acquisition IT1 (F(1,66) = 66.05, p < .OI) Similar analyses performed on the latency data also showed a significant effect of acquisition IT1 (F(1,66) = 26.48, p < .Ol).
Table 1 also shows that, for groups receiving the long IT1 during acquisition, the longer the IT1 during preexposure, the fewer the conditioned responses and the longer the latency of the response. The effect of preexposure IT1 was significant for total number of avoidance responses (F(2,66) = 5.83, p < .Ol) and for latency of response (F(2,66) = 5.43, p < .Ol). However, since the no preexposure condition is not on the same continuum as the two IT1 conditions, a separate 2 x 2 ANOVA was performed on the total number of avoidance responses, omitting the nonpreexposed groups. Both Preexposure IT1 and Acquisition ITI were reliable sources of differences (F(1,44) = 5.42, p < .05; F(1,44) = 33.38, p < .Ol, respectively), but the interaction between them was not significant (F(l,44) = 1.13, p > .05).
The results of this experiment relate in several ways to the empirical base of the latent inhibition phenomenon, as well as to the hypothesis that the effects of preexposure IT1 are equivalent in habituation and

544 SCHNUR AND LUBOW
TABLE 1
MEAN PERCENTAGE AND LATENCY OF AVOIDANCE RESPONSES AS A FUNCTION OF PREEXPOSURE ITI AND ACQUISITION ITI
Acquisition IT1
Preexposure ITI Short (s) Long (I)
Control (C) Mean percentage Mean latency (set)
Short (S) Mean percentage Mean latency (set)
Low 6) Mean percentage Mean latency (set)
13.06 61.39 5.32 3.16
14.31 47.36 5.54 4.17
8.20 30.97 5.86 4.94
latent inhibition paradigms. First of all, latent inhibition was demonstrated, adding to the substantial literature on the effect (Lubow, 1973). This effect is shown by the significant reduction in number of avoidance responses and increase in latencies in that group preexposed and trained with the long IT1 (Group Ll) as compared to the nonpreexposed group tested with the long ITI (Cl). That latent inhibition also was not revealed in the comparison between Group Ss and Group Cs may be attributed to the floor effect of the short IT1 acquisition schedule which precluded demonstrating learning in either group.
More importantly, the present experiment indicated that the pre- exposure IT1 affected subsequent learning. These differences were in the predicted direction, with the longer IT1 in preexposure hindering subsequent avoidance conditioning as compared to the shorter pre- exposure ITI. This effect is shown in Table 1, where Group Sl gave more avoidance responses than Group Ll and Group Ss gave more avoidance responses than Group Ls. The main effects demonstrated in the 2 x 2 ANOVA with the lack of a significant interaction substantiate the hypothesis that a longer preexposure IT1 results in poorer later learning than a shorter preexposure ITI.
EXPERIMENT 2
A second parametric characteristic of habituation is that, “The weaker the stimulus, the more rapid and/or more pronounced is habitua- tion. Strong stimuli may yield no significant habituation” (Thompson & Spencer, 1966, p. 19; Groves & Thompson, 1970; Thompson et al., 1973). This effect has also been reported in a variety of species and response systems (Thompson & Spencer, 1966). Yet, as Davis and Wagner (1968)

LATENT INHIBITION 545
point out, this finding, too, seems limited to those designs in which the manipulation of stimulus intensity during habituation and its measurement covary. When manipulation and measurement are separated, habituation is a direct function of stimulus intensity (cf. Graham, 1973).
Two previous studies have investigated the effects of preexposure intensity on latent inhibition. Lubow, Markman, and Allen (1968) failed to find a relation between preexposure intensity and latent inhibition with classical conditioning of the rabbit pinna response. On the other hand, Crowell and Anderson (1972, Experiment 1) found a direct relation between preexposure intensity and latent inhibition using rats in a CER paradigm. However, the effect was short-lived, appearing on the first test trial but not thereafter.
The purpose of the present experiment is to test for the effects of preexposure intensity independently of the effects of conditioning intensity over 16 acquisition trials. The latent inhibition paradigm is employed and two levels of CS intensity, 71 and 91 db, are manipulated factorially between the preexposure and acquisition phases. On the basis of the effects of stimulus intensity in the habituation paradigm (Davis & Wagner, 1968; Graham, 1973) it is expected that latent inhibition will vary directly with preexposure intensity.
Method
Subjects. The subjects were 24 male albino rats of the Sprague-Dawley strain. All were 70-80 days of age at the beginning of the experiment and were housed one to a cage. Upon delivery from the supplier, rats received food and water ad libitum for 6 days. During the next week, all rats were reduced to approximately 80% of their ad libitum weights by restrictive feeding and then maintained on a 24-hr feeding schedule for the duration of the experiment.
Apparatus. The apparatus consisted of two commercial rat test chambers, 24 x 30 x 24 cm, each enclosed in a light and sound attenuating box. Each chamber contained a food hopper located 2.0 cm above the grid floor and 12.5 cm from the side of the end wall. In one chamber, a manipulandum (2.6 x 3.5 cm) was located to the right of the hopper; in the other chamber, to the left of the hopper. The shock grid was made from stainless-steel bars 0.6 cm in diameter, mounted at 2.0-cm intervals. Shock was provided by a scrambled constant current source set at 2mA.
The CS was a I-kHz tone at an intensity of either 71 db SPL (20 pN/m2) or 91 db SPL (20 pN/mZ). The tones were delivered through a speaker located in the enclosure. However, the intensities were measured from a location in the center of the chamber. A ventilator fan maintained a con- stant masking noise of 75 db SPL (20 pN/m2). Experimental events were controlled and recorded automatically by solid-state equipment located in an adjacent room.

546 SCHNUR AND LUBOW
Procedure. All experimental sessions were 2-hr long and were con- ducted at the same time for each subject. In the first session, animals were magazine trained and shaped automatically. Food pellets were delivered on a variable interval (VI) l-min schedule and each bar press yielded a 45-mg pellet. All subjects began responding steadily during this sesion and hand shaping was not necessary. During the first hour of the second day, animals worked on a VI 1-min schedule of reinforcement. Starting with the second hour of this session and for all subsequent ses- sions, a VI 2-min schedule of reinforcement was in effect.
The preexposure phase of the experiment began on Day 7. Pairs of subjects were randomly assigned to one of six equal-sized groups. Group 71-71 and Group 71-91 received 24 2-min preexposures to the 71-db tone on each of Days 7- 10. Likewise, Group 91-91 and Group 91-71 received preexposures to the 91-db tone on Days 7- 10. Group O-7 1 and Group O-9 1 received no CS preexposure but spent an equivalent time in the con- ditioning chambers on Days 7- 10. All CS preexposures were presented on a fixed interval 5-min schedule while subjects bar-pressed for food.
On Days 1 l- 14, 4 days of classical conditioning were superimposed on the VI baseline. Groups 71-71, 91-71, and O-7 1 received the 71-db tone CS; Groups 91-91.71-91, and O-91 received the 91-db CS. Each 2-min CS terminated in a 2-mA, I-set footshock. The four CS-US trials per day were presented 20, 50, 70, and 100 min after the start of each daily session.
Response measure. The dependent variable was the amount of response suppression during a stimulus presentation. A suppression ratio, A/A + B was calculated: A is the number of responses during a 2-min CS and B is the number of responses during a 2-min period immediately preceding CS onset. Thus, a ratio of 0 indicates complete suppression to the CS, a ratio of .50 indicates identical response rates during CS and pre-CS intervals, and a ratio greater than .50 indicates a higher rate of responding in the presence of the CS than in its absence.
Results
Figure 1 shows mean suppression ratios for each of the six groups as a function of blocks of four acquisition trials. Each panel in Fig. 1 depicts a comparison of the three preexposure conditions for a particular acquisition condition. Looking across panels, it is clear that preexposure of the same stimulus intensity as that used in acquisition (Groups 71-71 and 91-91) retarded conditioning as compared to either nonpreexposure (Groups O-71 and O-91) or preexposure to a stimulus intensity different from that used in acquisition (Groups 91-71 and 71-91). Figure 1 also shows that preexposure of the same stimulus as that used in acquisition produced a greater decrement in conditioning relative to nonpreexposure at the higher than at the lower CS intensity. In addition, these differences
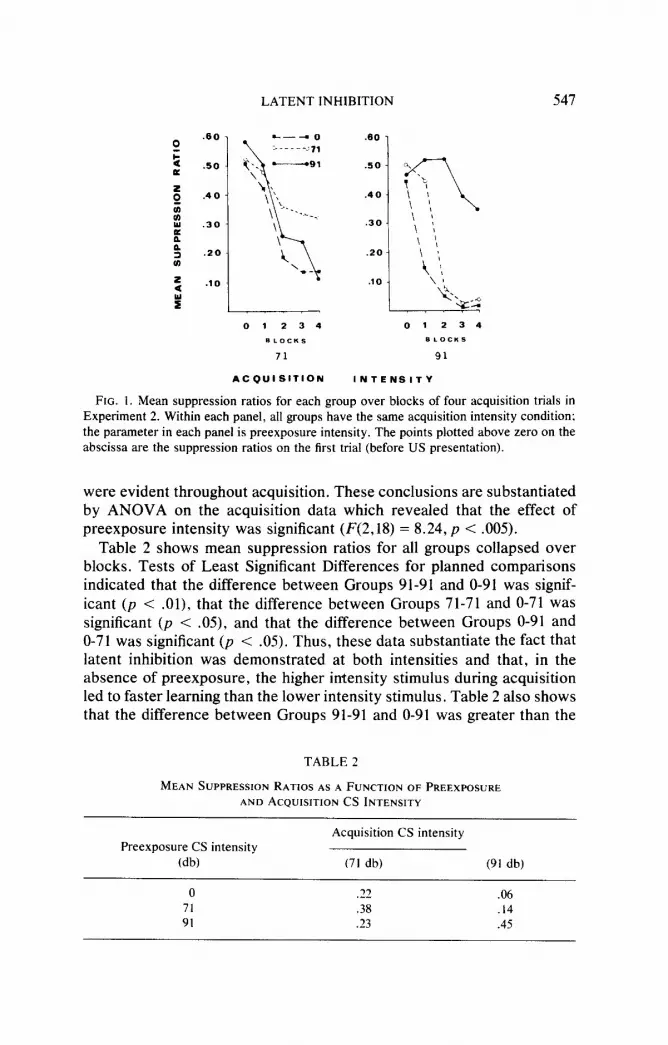
LATENT INHIBITION 547
.60 - L-4 0
j-----i71
.50 q-. .50 q-. -91 -91 \ \
\sk \sk .40 .40
\ “\,. \ “\,.
:\\; \-
\ \ --..__ --..__ .30 .30 \ \
\ \ .20 .20
L L *- *- .lO .lO
.40 \ 1 \ : \
.30 ’ : \ ! \ :
.20
.lO
I
‘i, ‘I,,
%&
0 1 234 sLOCIS
91
ACQUISITION INTENSITY
FIG. 1. Mean suppression ratios for each group over blocks of four acquisition trials in Experiment 2. Within each panel, all groups have the same acquisition intensity condition: the parameter in each panel is preexposure intensity. The points plotted above zero on the abscissa are the suppression ratios on the first trial (before US presentation).
were evident throughout acquisition. These conclusions are substantiated by ANOVA on the acquisition data which revealed that the effect of preexposure intensity was significant (F(2,lS) = 8.24, p < .005).
Table 2 shows mean suppression ratios for all groups collapsed over blocks. Tests of Least Significant Differences for planned comparisons indicated that the difference between Groups 91-91 and O-91 was signif- icant (p < .Ol), that the difference between Groups 71-71 and O-71 was significant (p < .OS), and that the difference between Groups O-91 and O-71 was significant (p < .05). Thus, these data substantiate the fact that latent inhibition was demonstrated at both intensities and that, in the absence of preexposure, the higher intensity stimulus during acquisition led to faster learning than the lower intensity stimulus. Table 2 also shows that the difference between Groups 91-91 and O-91 was greater than the
TABLE 2
MEAN SUPPRESSION RATIOS AS A FUNCTION OF PREEXPOSURE
AND ACQUISITION CS INTENSITY
Preexposure CS intensity (db)
Acquisition CS intensity
(71 db) (91 db)
0 .22 .06
71 .38 .I4
91 .23 .45

548 SCHNUR AND LUBOW
difference between Groups 71-71 and O-71. In addition, with the 91-db CS in acquisition, preexposure of the more intense CS retarded conditioning more than did preexposure of the less intense CS, the difference between Groups 91-91 and 71-91 being significant (p < .Ol). These results are consistent with the hypothesized direct relation between preexposure stimulus intensity and latent inhibition. However, with the 71-db CS in acquisition, just the opposite was true. That is, preexposure of the less intense CS produced slower conditioning (more latent inhibition) than preexposure of the more intense CS, with the difference between Groups 71-71 and 91-71 also significant (p < .05).
Discussion
The fact that CS preexposure retarded subsequent conditioned suppres- tion is consistent with other studies using the CER technique (Lubow, 1973). In addition, the latent inhibition effect obtained in the present experiment is quite durable. Figure 1 shows a difference between pre- exposed and nonpreexposed groups after4 days and 16 trials of acquisition.
With regard to the effects of preexposure intensity on latent inhibition, the results of Experiment 2 reveal an important inconsistency. If one measures the decrement in conditioning relative to control levels, it appears that latent inhibition is directly related to preexposure intensity, the difference between Groups 91-91 and O-91 being greater than the difference between Groups 71-71 and O-71 throughout acquisition (cf. Crowell & Anderson, 1972). Similarly, if one measures the rate of con- ditioning to the more intense CS, latent inhibition is again directly related to preexposure intensity. In these groups, preexposure to the more intense CS (Group 91-91) retarded conditioning more than did preexposure to the less intense CS (Group 71-91). However, if one measures the rate of conditioning to the less intense CS, latent inhibition is inversely related to preexposure intensity. In these groups, preexposure to the less intense CS (Group 71-71) retarded conditioning more than did preexposure to the more intense CS (Group 91-71).
The inconsistency in these results can be resolved if it is recognized that altering stimulus intensity between preexposure and conditioning involves a generalization test. This point has been noted also for the habituation paradigm (cf. Thompson et al. 1973). Thus, if it is assumed that in the present experiment the effects of preexposure did not generalize along the intensity dimension, the results for Groups 91-71 and 71-91 are consistent: Neither group showed latent inhibition. We are suggesting that the effects of preexposure can be highly stimulus specific (cf. Crowell & Anderson, 1972, Experiment 1). By this generalization decrement interpretation, preexposure of the more intense CS did not retard conditioning to the 91-db CS more than did preexposure of the

LATENT INHIBITION 549
less intense CS. Rather, preexposure retarded conditioning in Group 91-91, but had no effect in Group 71-91. Similarly, preexposure of the less intense CS did not retard conditioning to the 71-db CS more than did preexposure of the more intense CS. Rather, preexposure retarded conditioning in Group 71-71, but had no effect in Group 91-71. Recall, however, that Group 91-91 showed poorer conditioning than Group 71-71 even while Group O-91 showed better conditioning than Group O-71. That is, the relative decrement in conditioning produced by preexposure still warrants the conclusion that latent inhibition varies directly with preexposure intensity.
Taken together, the results of Experiments 1 and 2 demonstrate that preexposure IT1 and preexposure stimulus intensity have effects in the latent inhibition paradigm which parallel those previously reported in the habituation paradigm. This demonstration supports the hypothesis that common processes underlie the habituation and latent inhibition phenomena and is consistent with a conditioned attention theory of latent inhibition (Lubow, Alec, & Arzy, 1975: Lubow, Schnur, & Rifkin, 1976) and with a dual process theory of habituation such as that proposed by Wagner (in press).
REFERENCES
Ackil, .I. E., Mellgren, R. L., Halgren, C., & Frommer, G. P. Effects of CS preexposure on avoidance learning in rats with hippocampal lesions. Journal of Comparative and Physiological Psychology, 1%9, 69, 739-747.
Carew, T. J., Pinsker, H. M., & Kandel. E. R. Long term habituation of a defensive withdrawal reflex in aplysia. Science, 1972, 175, 451-453.
Carlton, P. L., & Vogel, J. R. Habituation and conditioning. Journal of Comparative and Physiological Psychology, 1967, 63, 348-351.
Crowell, C. R., & Anderson, D. C. Variations in intensity. interstimulus interval, and interval between preconditioning CS exposures and conditioning with rats. Journal of Comparative and Physiological Psychology, 1972, 79, 291-298.
Davis, M. Effects of interstimulus interval length and variability on startle response habituation in the rat. Journal of Comparative and Physiological Psychology, 1970, 72, 177- 192.
Davis, M., & Wagner, A. R. Startle responsiveness after habituation to different intensities of tone. Psychonomic Science, 1968, 12, 337-338.
Graham, F. K. Habituation and dishabituation of responses innervated by the automatic nervous system. In H. V. S. Peeke & M. J. Herz (Eds.), Habituation: Behavioral studies (Vol. 1). New York: Academic Press, 1973.
Groves, P. M., & Thompson, R. F. Habituation: A dual process theory. Psychological Review, 1970, 77, 419-450.
Lantz, A. E. Effect of number of trials, interstimulus interval, and dishabituation during CS habituation on subsequent conditioning in a CER paradigm. Animal Learning and Behavior, 1973, 1, 273-277.
Lubow, R. E. Latent inhibition. Psychological Bulletin, 1973, 79, 398-407. Lubow, R. E., Alec, M., & Arzy, J. Behavioral decrement following stimulus preexposure:
The effects of number of preexposures, presence of a second stimulus and

5.50 SCHNUR AND LUBOW
interstimulus interval in children and adults. Jouvnul oJ’ E.uprrimmtn/ Ps~clio/~,e~: Animal Behavior Processes, 1975. 1, 178- 188.
Lubow, R. E.. Markman, R. E.. & Allen. J. Latent inhibition and classical conditioning of the rabbit pinna response. Journal of Compurati~~r and Physiologicul P.~ychoIo~y. 1968, 66, 688-694.
Lubow. R. E., Schnur, P., & Ritkin, B. Latent inhibition and conditioned attention theory. Journal of Experimental Psychology: Animal Behurzior Processes, 1976.2, 163- 174.
Schnur, P. Selective attention: Effect of element preexposure on compound conditioning in rats. Journal of Comparative and Physiological Psychology. 1971, 76, 123- 130.
Thompson, R. F., Groves, P. M., Teyler, T. J.. & Roemer, R. A. A dual-process theory of habituation: Theory and behavior. In H. V. S. Peeke & M. J. Herz (Eds.). Habituation: Behavioral Studies (Vol. 1). New York: Academic Press, 1973.
Thompson, R. F., & Spencer, W. A. Habituation: A model phenomenon for the study of neuronal substrates of behavior. Psychological Retziew,. 1966, 73, 16-43.
Wagner, A. R. Priming in STM: An information processing mechanism for self-generated or retrieval-generated depression in performance. In T. J. Tighe & R. N. Leaton (Eds.), Habituation: Perspectives from child de~~elopment, animal brhuvior, und neurophysiology. Hillsdale. N.J.: Lawrence Erlbaum Associates, in press.
Received September 26. 1975 Revised January 10, 1976





























