Embed Size (px)
Citation preview

Current Medicinal Chemistry, 2012, 19, 1443-1474 1443
Leishmaniasis: Prevention, Parasite Detection and Treatment T. Kobets, I. Grekov and M. Lipoldová*
Laboratory of Molecular and Cellular Immunology, Institute of Molecular Genetics, Academy of Sciences of the Czech Republic, v.v.i., Prague, Czech Republic
Abstract: Leishmaniasis remains a public health problem worldwide, affecting approximately 12 million people in 88 countries; 50 000 die of it each year. The disease is caused by Leishmania, obligate intracellular vector-borne parasites. In spite of its huge health impact on the populations in vast areas, leishmaniasis is one of the most neglected diseases. No safe and effective vaccine currently exists against any form of human leishmaniasis. The spectrum and efficacy of available antileishmanial drugs are also limited. First part of this review discusses the approaches used for the vaccination against leishmaniasis that are based on the pathogen and includes virulent or attenuated parasites, parasites of related nonpathogenic species, whole killed parasites, parasites’ subunits, DNA vaccines, and vaccines based on the saliva or saliva components of transmitting phlebotomine vector. Second part describes parasite detection and quantification using microscopy assays, cell cultures, immunodetection, and DNA-based methods, and shows a progress in the development and application of these techniques. In the third part, first-line and alternative drugs used to treat leishmaniasis are characterized, and pre-clinical research of a range of natural and synthetic compounds studied for the leishmanicidal activity is described. The review also suggests that the application of novel strategies based on advances in genetics, genomics, advanced delivery systems, and high throughput screenings for leishmanicidal compounds would lead to improvement of prevention and treatment of this disease.
Keywords: Tropical disease, kala-azar, preventive medicine, animal model, therapy of visceral, cutaneous and mucocutaneous leishmaniasis, estimation of pathogen load.
1. INTRODUCTION
Leishmaniasis is a vector-borne protozoan infection with a wide clinical spectrum. About 21 Leishmania species are known to infect humans; they are transmitted by approximately 30 species of phlebotomine sand flies [1, 2]. Leishmania parasites have two basic life stages: an extracellular motile stage (promastigote) inside an invertebrate host and an intracellular non-motile stage (amastigote) inside a vertebrate host [3]. In the vertebrate host organism, Leishmania parasites infect so-called professional phagocytes (neutrophils, monocytes and macrophages) [4], as well as dendritic cells (DC) [5], immature myeloid precursor cells, sialoadhesin-positive stromal macrophages of the bone marrow, hepatocytes and fibroblasts [6]. Macrophages are supposed to be the main cellular compartment for Leishmania in the mammalian host.
It has been assumed that leishmaniasis may have been established 50 million years ago, during the Paleogene [7]. The direct evidence that people suffered from this disease came from samples 4 000 years old as DNA of L. donovani was found in Egyptian mummies from a Middle Kingdom tomb [8]. The presence of Leishmania was also detected in the facial lesions on ancient skulls from the Atacama Desert in Chile [9].
Leishmania amastigotes have been first observed by in 1885 by Cunningham in skin lesions of patients from India, but he suggested that they were members of Mycetozoa (fungi) [10]. Protozoal nature of Leishmania was first recognized in 1898 by Borovsky during his study of skin lesions in Turkmenistan [11]. Leishman in 1903 discovered similar intracellular bodies in the visceral organs of fatal cases of kala-azar from India, and established that they were morphologically related to trypanosomes [12]. Similar observation was made in the same year in India by Donovan [13]. The causal relationship between Leishmania parasites and development of cutaneous lesions was confirmed in 1908 by Martsinovsky who self infected himself with the parasite cultures [14].
Leishmaniasis includes asymptomatic infection and three main clinical syndromes. In the dermis, parasites cause the cutaneous form of the disease, which can be localized or diffuse; in the mucosa, they cause mucocutaneous leishmaniasis, and the metastatic spread of infection to the spleen and liver leads to
*Address correspondence to this author at the Institute of Molecular Genetics, Academy of Sciences of the Czech Republic, v.v.i., Vídeňská 1083, 14220 Prague 4, Czech Republic; Tel: +(420)241063243; Fax: +(420)224310955; E-mail: [email protected]
visceral leishmaniasis (also known as kala-azar or black fever). Parasites can also enter other organs; such as lymph nodes, bone marrow and lungs, and in rare cases, can even reach the brain [4].
Leishmaniasis is one of the most neglected diseases, along with other like sleeping sickness or Chagas’ disease. Such diseases are named “neglected” because they persist predominantly in marginalized and poor communities [2, 15, 16]. The type of pathology that is caused depends on the species of Leishmania, the genotype and nutritional status of the host, the transmitting vector and environmental and social factors [4]. The disease remains a public health problem worldwide, affecting approximately 12 million people in 88 countries; 50 000 die of it each year. Leishmaniasis is endemic in areas of tropics, subtropics, including southern Europe, in setting ranging from rain forests in the Americas to deserts in Asia. Leishmaniasis represents a major public health problem in the Eastern Mediterranean Region. Based on geographical distribution, the disease is divided into Old World and New World leishmaniasis [1, 2, 15, 17].
Visceral leishmaniasis is endemic in more than 60 countries, however, 90% of the 500 000 new cases that occur every year concern six countries only – India, Bangladesh, Nepal, Brazil, Ethiopia and Sudan. Visceral form of the disease is caused mainly by L. donovani in the Indian subcontinent, Asia and Africa; L. infantum in the Mediterranean basin [2], Central Asia and Transcaucasia [18], and L. chagasi in South America [2]. In Mediterranean countries, South America [2] and in Central Asia and Transcaucasia [18], the disease is zoonotic and affects mainly infants and young children. In these countries, stray and domestic dogs are the main reservoir for the infection. In the Indian subcontinent and Africa, visceral leishmaniasis is anthroponotic and affects adults and children [2].
Cutaneous leishmaniasis is endemic in more than 70 countries, with an estimation of 1.5-2 million new cases every year. Afghanistan, Syria, and Brazil are the main foci. Cutaneous form of the disease is caused mainly by L. tropica and L. major in the Old World, and by L. braziliensis, L. guyanensis, L. panamensis, L. peruviana, L. mexicana, L. amazonensis, and L. venesuelensis in the New World. Mucosal leishmaniasis develops in a small number of patients with New World cutaneous leishmaniasis, however its course is chronic and may be life-threatening [2].
The disease is spreading because of risk factors that include climate changes, population movements, long-distance tourism and trade [15]. In the last decade, leishmaniasis expanded or emerged in
1875-533X/12 $58.00+.00 © 2012 Bentham Science Publishers

1444 Current Medicinal Chemistry, 2012 Vol. 19, No. 10 Kobets et al.
several foci worldwide as the result of sand fly expansion due to natural and human factors, such as urbanization and deforestation, and global warming [2]. Cutaneous leishmaniasis is one of the top 10 diseases among tourists returning from tropical countries with skin problems. Cases of leishmaniasis were also described in organ transplant recipients [2, 16].
2. VACCINATION AGAINST LEISHMANIASIS
The aim of vaccination against an infectious agent is to provide effective immunity by induction of clonal expansion in specific memory T and/or memory B cells. Hence, a repeated encounter with the same antigen(s) will induce secondary response, which will be more rapid and more effective than the normal primary response. A good vaccine must induce a protective level of immunity at the appropriate site and this protective immune response should have an adequate duration. It must be also safe to administer, affordable by the population and suitable for the pertinent population, as different populations might differ in genetics of response. The majority of the most successful vaccines have been developed empirically; recent advances in immunology that are revealing key factors in protection against many diseases will facilitate a more rational approach to vaccines design. However, despite the considerable knowledge about the immune response against Leishmania parasites [19, 20], no safe and effective vaccine currently exists against any form of human leishmaniasis. The only long time protection against cutanenous leishmaniasis in humans is leishmanization – vaccination with the virulent parasites that had to be abandoned for safety reasons [21, 22] (see Section 2.1.1.1.), and the only successful vaccine is Leishmune®, which blocks the transmission of canine leishmaniasis [23]. Vaccine CaniLeish® was recently registered for the veterinary use in the European Union (see Section 2.1.2.1.). Probably not all responses needed for protection against leishmaniasis have been identified. Genetic analysis of susceptibility [24-30] that is hypothesis free could speed the discovery of new important defense mechanisms against leishmaniasis.
The following section and Table 1 give the examples of approaches of the development of vaccines against leishmaniasis. The comprehensive information can be found on the webpage: http://www.leishvaccines.net/
2.1. Vaccines Based on Pathogen
2.1.1. Vaccination with the Whole Organism 2.1.1.1. Leishmanization – Inoculation of Virulent Leishmania Parasites
Vaccination against cutaneous leishmaniasis using virulent parasites has been used for centuries. Bedouin or some Kurdish tribal societies traditionally exposed their babies’ bottoms to sand fly bites in order to protect them from facial lesions. Another ancient technique practicized in the Middle East was the use of a thorn to transfer infectious material from lesions to uninfected individuals [21]. Development of media for culturing Leishmania parasites enabled controlled infections with live promastigotes. Large-scale vaccination trials were carried in the past in the Soviet Union [31, 32], Israel [21], and in Iran [22]. In Iran, leishmanization of more than 160000 children was performed from 1982 to 1986; and leishmanization of 1 800 000 military personnel and soldiers, 6000 war refugees and several thousands of individuals from other population groups was carried until 1989 [22]. Several thousand persons were vaccinated during 60s in Turkmenistan [31] and in Uzbekistan [32]. These trials led to a high percentage of successful lesion development, but also faced problems with the viability and infectivity of the injected organisms, the development of large uncontrolled lesions, the enhancement of other skin diseases, and immunosuppression and
were therefore abandoned [21, 22, 33]. Establishment of a suitable condition to grow parasites and prepare stabilates [34, 35] could prevent problems with the viability and infectivity of Leishmania promastigotes. Leishmanization with well defined L. major stabilates was used for evaluation of candidate vaccines against leishmaniasis [34].
Although vaccination with live virulent parasites grown in defined conditions leads to good protection against subsequent infection [34], its use have been discontinued due to safety reasons. Leishmania parasites persist in cured animals [36, 37] and could cause serious problems in immunocompromised individuals [22]. Reactivation of persisting parasites might be especially dangerous in areas with HIV infection [38]. In addition, leishmanization cannot be applied for vaccination against visceral leishmaniasis. Due to these reasons, researchers switched to development of other types of vaccines. 2.1.1.2. Vaccination with Parasites of Related Nonpathogenic Leishmania Species
Use of vaccinia virus, a species nonpathogenic for humans in order to induce protective immunity against the related dangerous smallpox virus was the first scientifically described case of vaccination [39]. Breton and coworkers [40] applied this approach to investigate the potential of lizard parasite L. tarentolae that is nonpathogenic to humans and other mammals as a live candidate vaccine. They found that a single intraperitoneal injection of L. tarentolae elicits a protective immune response against the subsequent infectious challenge with L. donovani in susceptible BALB/c mice and led to decrease of a parasite burden in spleen and liver. L. tarentolae expresses an Amastin-like gene, cysteine protease B (cpb), lipophosphoglycan lpg3 and the leishmanolysin gp63, genes that play role in parasite virulence in Leishmania species pathogenic to humans. The degree of similarity varied from 59% and 60% for Amastin, 89% for lpg3 and 71% and 68% for cpb, in L. major and L. infantum, respectively. The amastigote A2 gene, expressed specifically by the L. donovani complex which promotes visceralization, was absent in L. tarentolae [41]. Introduction of A2 gene into L. tarentolae increased ability of this species to survive in liver of BALB/c mice [42]. The potential of L. tarentolae to protect other species than mouse from pathogenic Leishmania species has yet to be investigated. 2.1.1.3. Vaccination with Attenuated Parasites
An attenuated vaccine is a vaccine created by reducing the virulence of a pathogen in order to induce a protective immune response without causing the severe effects of the disease. Parasites can be weakened through undefined or defined genome alteration.
Treatment of Leishmania parasites by γ -irradiation [43, 44], chemical mutagenesis [45], chemical mutagenesis followed with selection for temperature-sensitivity [46], by a long term in vitro propagation [47], and by culturing in vitro under gentamicin pressure [48, 49] leads to undefined genetic alterations and parasites need to be further analyzed in order to establish the nature of introduced mutation(s). These attenuated parasites induced a protective immunity in several animal models. For example, vaccination of BALB/c mice with L. major or L. mexicana attenuated by gentamicin pressure led to decrease of lesions size after a subsequent challenge with virulent parasites [48], and vaccination with gentamicin-attenuated L. infantum protected dogs against the wild type parasite [49]. Vaccination with irradiation-attenuated L. major was shown to induce protective effect in CBA mice [44].
Defined genome alteration was achieved by targeting Leishmania genes such as dihydrofolate-reductase thymidylate synthase (dhfr-ts) [50-53], cystein-proteinase (cpa, cpb) [54], biopterin transporter (bt1) [55], lipophosphoglycan 2 (lpg2) [56], silent information regulatory 2 (sir2) [57], phosphomannomutase (pmn) [58] and centrin 1 (cen1) [59]. Vaccination of BALB/c mice
Leishmaniasis: Prevention, Parasite Detection and Treatment Current Medicinal Chemistry, 2012 Vol. 19, No. 10 1445
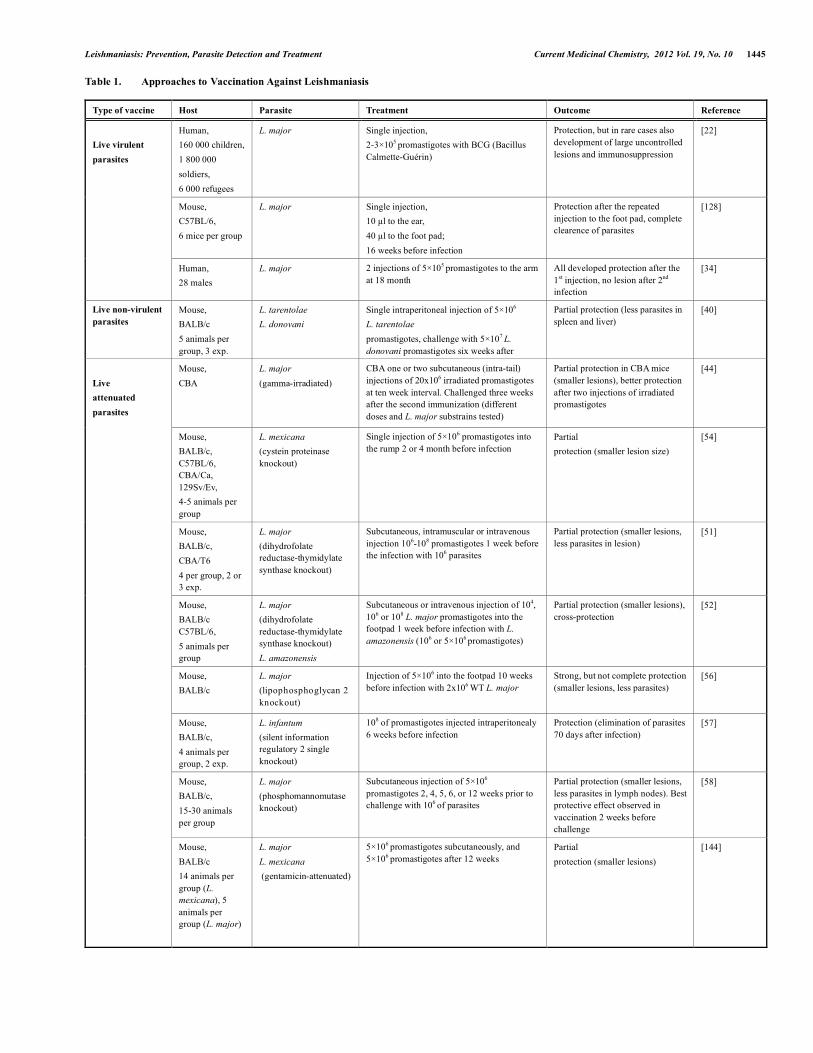
Table 1. Approaches to Vaccination Against Leishmaniasis
Type of vaccine Host Parasite Treatment Outcome Reference
Human, 160 000 children, 1 800 000 soldiers, 6 000 refugees
L. major Single injection, 2-3×105 promastigotes with BCG (Bacillus Calmette-Guérin)
Protection, but in rare cases also development of large uncontrolled lesions and immunosuppression
[22]
Mouse, C57BL/6, 6 mice per group
L. major Single injection, 10 µl to the ear, 40 µl to the foot pad; 16 weeks before infection
Protection after the repeated injection to the foot pad, complete clearence of parasites
[128]
Live virulent parasites
Human, 28 males
L. major 2 injections of 5×105 promastigotes to the arm at 18 month
All developed protection after the 1st injection, no lesion after 2nd infection
[34]
Live non-virulent parasites
Mouse, BALB/c 5 animals per group, 3 exp.
L. tarentolae L. donovani
Single intraperitoneal injection of 5×106
L. tarentolae promastigotes, challenge with 5×107 L. donovani promastigotes six weeks after
Partial protection (less parasites in spleen and liver)
[40]
Mouse, CBA
L. major (gamma-irradiated)
CBA one or two subcutaneous (intra-tail) injections of 20x106 irradiated promastigotes at ten week interval. Challenged three weeks after the second immunization (different doses and L. major substrains tested)
Partial protection in CBA mice (smaller lesions), better protection after two injections of irradiated promastigotes
[44]
Mouse, BALB/c, C57BL/6, CBA/Ca, 129Sv/Ev, 4-5 animals per group
L. mexicana (cystein proteinase knockout)
Single injection of 5×106 promastigotes into the rump 2 or 4 month before infection
Partial protection (smaller lesion size)
[54]
Mouse, BALB/c, CBA/T6 4 per group, 2 or 3 exp.
L. major (dihydrofolate reductase-thymidylate synthase knockout)
Subcutaneous, intramuscular or intravenous injection 106-108 promastigotes 1 week before the infection with 106 parasites
Partial protection (smaller lesions, less parasites in lesion)
[51]
Mouse, BALB/c C57BL/6, 5 animals per group
L. major (dihydrofolate reductase-thymidylate synthase knockout) L. amazonensis
Subcutaneous or intravenous injection of 104, 106 or 108 L. major promastigotes into the footpad 1 week before infection with L. amazonensis (106 or 5×106 promastigotes)
Partial protection (smaller lesions), cross-protection
[52]
Mouse, BALB/c
L. major (lipophosphoglycan 2 knockout)
Injection of 5×106 into the footpad 10 weeks before infection with 2x106 WT L. major
Strong, but not complete protection (smaller lesions, less parasites)
[56]
Mouse, BALB/c, 4 animals per group, 2 exp.
L. infantum (silent information regulatory 2 single knockout)
108 of promastigotes injected intraperitonealy 6 weeks before infection
Protection (elimination of parasites 70 days after infection)
[57]
Mouse, BALB/c, 15-30 animals per group
L. major (phosphomannomutase knockout)
Subcutaneous injection of 5×106 promastigotes 2, 4, 5, 6, or 12 weeks prior to challenge with 106 of parasites
Partial protection (smaller lesions, less parasites in lymph nodes). Best protective effect observed in vaccination 2 weeks before challenge
[58]
Live attenuated parasites
Mouse, BALB/c 14 animals per group (L. mexicana), 5 animals per group (L. major)
L. major L. mexicana (gentamicin-attenuated)
5×106 promastigotes subcutaneously, and 5×106 promastigotes after 12 weeks
Partial protection (smaller lesions)
[144]
1446 Current Medicinal Chemistry, 2012 Vol. 19, No. 10 Kobets et al.
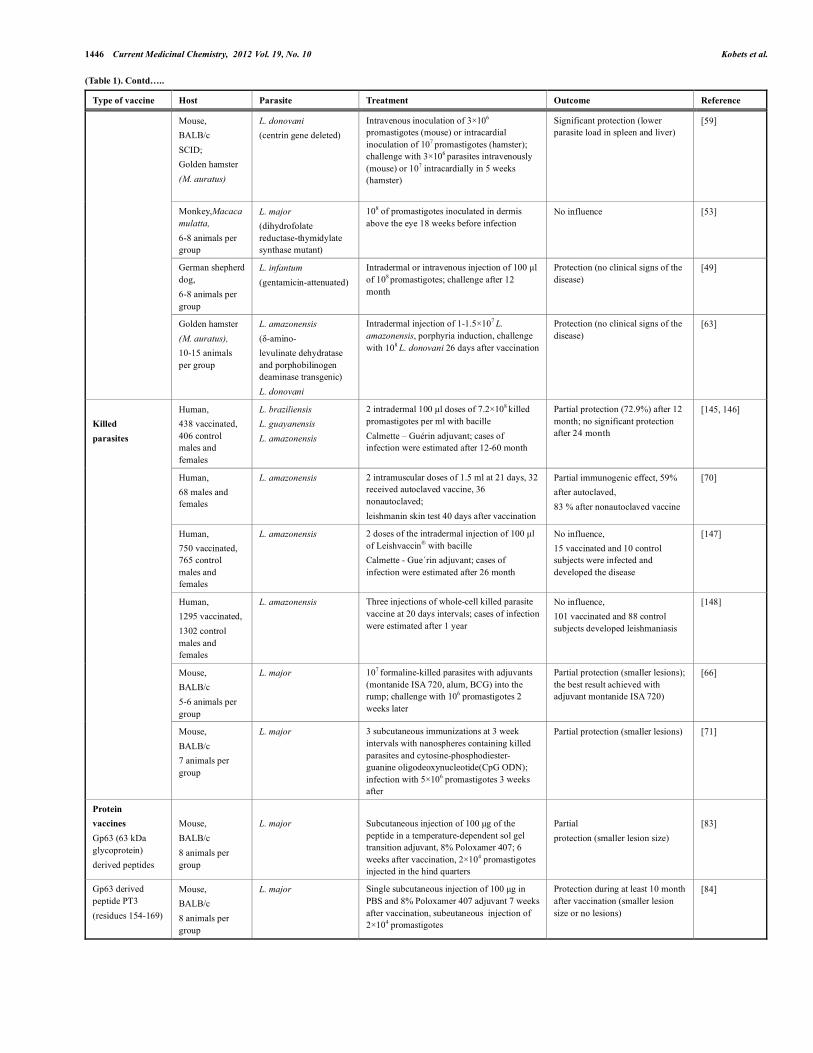
(Table 1). Contd…..
Type of vaccine Host Parasite Treatment Outcome Reference
Mouse, BALB/c SCID; Golden hamster (M. auratus)
L. donovani (centrin gene deleted)
Intravenous inoculation of 3×106
promastigotes (mouse) or intracardial inoculation of 107 promastigotes (hamster); challenge with 3×106 parasites intravenously (mouse) or 107 intracardially in 5 weeks (hamster)
Significant protection (lower parasite load in spleen and liver)
[59]
Monkey,Macaca mulatta, 6-8 animals per group
L. major (dihydrofolate reductase-thymidylate synthase mutant)
108 of promastigotes inoculated in dermis above the eye 18 weeks before infection
No influence [53]
German shepherd dog, 6-8 animals per group
L. infantum (gentamicin-attenuated)
Intradermal or intravenous injection of 100 µl of 108 promastigotes; challenge after 12 month
Protection (no clinical signs of the disease)
[49]
Golden hamster (M. auratus), 10-15 animals per group
L. amazonensis (δ-amino- levulinate dehydratase and porphobilinogen deaminase transgenic) L. donovani
Intradermal injection of 1-1.5×107 L. amazonensis, porphyria induction, challenge with 108 L. donovani 26 days after vaccination
Protection (no clinical signs of the disease)
[63]
Human, 438 vaccinated, 406 control males and females
L. braziliensis L. guayanensis L. amazonensis
2 intradermal 100 µl doses of 7.2×108 killed promastigotes per ml with bacille Calmette – Guérin adjuvant; cases of infection were estimated after 12-60 month
Partial protection (72.9%) after 12 month; no significant protection after 24 month
[145, 146]
Human, 68 males and females
L. amazonensis 2 intramuscular doses of 1.5 ml at 21 days, 32 received autoclaved vaccine, 36 nonautoclaved; leishmanin skin test 40 days after vaccination
Partial immunogenic effect, 59% after autoclaved, 83 % after nonautoclaved vaccine
[70]
Human, 750 vaccinated, 765 control males and females
L. amazonensis 2 doses of the intradermal injection of 100 µl of Leishvaccin® with bacille Calmette - Gue´rin adjuvant; cases of infection were estimated after 26 month
No influence, 15 vaccinated and 10 control subjects were infected and developed the disease
[147]
Human, 1295 vaccinated, 1302 control males and females
L. amazonensis Three injections of whole-cell killed parasite vaccine at 20 days intervals; cases of infection were estimated after 1 year
No influence, 101 vaccinated and 88 control subjects developed leishmaniasis
[148]
Mouse, BALB/c 5-6 animals per group
L. major 107 formaline-killed parasites with adjuvants (montanide ISA 720, alum, BCG) into the rump; challenge with 106 promastigotes 2 weeks later
Partial protection (smaller lesions); the best result achieved with adjuvant montanide ISA 720)
[66]
Killed parasites
Mouse, BALB/c 7 animals per group
L. major 3 subcutaneous immunizations at 3 week intervals with nanospheres containing killed parasites and cytosine-phosphodiester-guanine oligodeoxynucleotide(CpG ODN); infection with 5×106 promastigotes 3 weeks after
Partial protection (smaller lesions) [71]
Protein vaccines Gp63 (63 kDa glycoprotein) derived peptides
Mouse, BALB/c 8 animals per group
L. major
Subcutaneous injection of 100 µg of the peptide in a temperature-dependent sol gel transition adjuvant, 8% Poloxamer 407; 6 weeks after vaccination, 2×104 promastigotes injected in the hind quarters
Partial protection (smaller lesion size)
[83]
Gp63 derived peptide PT3 (residues 154-169)
Mouse, BALB/c 8 animals per group
L. major Single subcutaneous injection of 100 µg in PBS and 8% Poloxamer 407 adjuvant 7 weeks after vaccination, subeutaneous injection of 2×104 promastigotes
Protection during at least 10 month after vaccination (smaller lesion size or no lesions)
[84]
Leishmaniasis: Prevention, Parasite Detection and Treatment Current Medicinal Chemistry, 2012 Vol. 19, No. 10 1447
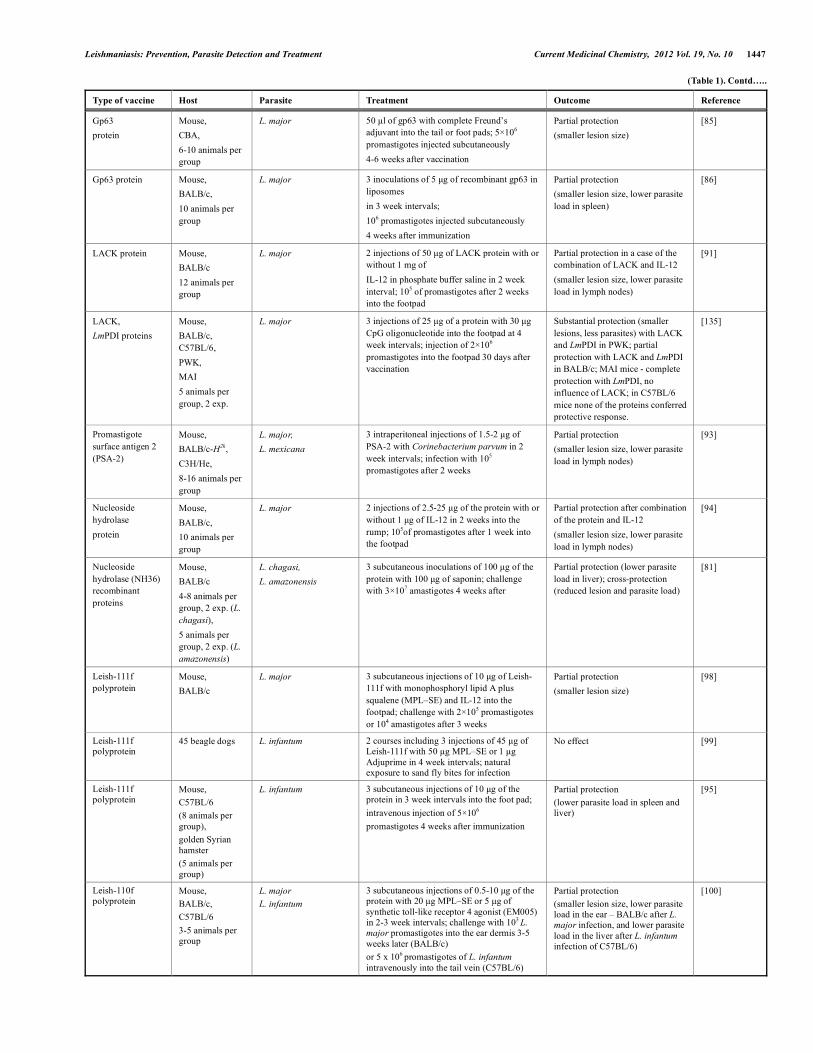
(Table 1). Contd…..
Type of vaccine Host Parasite Treatment Outcome Reference
Gp63 protein
Mouse, CBA, 6-10 animals per group
L. major 50 µl of gp63 with complete Freund’s adjuvant into the tail or foot pads; 5×106 promastigotes injected subcutaneously 4-6 weeks after vaccination
Partial protection (smaller lesion size)
[85]
Gp63 protein Mouse, BALB/c, 10 animals per group
L. major 3 inoculations of 5 µg of recombinant gp63 in liposomes in 3 week intervals; 106 promastigotes injected subcutaneously 4 weeks after immunization
Partial protection (smaller lesion size, lower parasite load in spleen)
[86]
LACK protein Mouse, BALB/c 12 animals per group
L. major 2 injections of 50 µg of LACK protein with or without 1 mg of IL-12 in phosphate buffer saline in 2 week interval; 105 of promastigotes after 2 weeks into the footpad
Partial protection in a case of the combination of LACK and IL-12 (smaller lesion size, lower parasite load in lymph nodes)
[91]
LACK, LmPDI proteins
Mouse, BALB/c, C57BL/6, PWK, MAI 5 animals per group, 2 exp.
L. major 3 injections of 25 µg of a protein with 30 µg CpG oligonucleotide into the footpad at 4 week intervals; injection of 2×106 promastigotes into the footpad 30 days after vaccination
Substantial protection (smaller lesions, less parasites) with LACK and LmPDI in PWK; partial protection with LACK and LmPDI in BALB/c; MAI mice - complete protection with LmPDI, no influence of LACK; in C57BL/6 mice none of the proteins conferred protective response.
[135]
Promastigote surface antigen 2 (PSA-2)
Mouse, BALB/c-H2k, C3H/He, 8-16 animals per group
L. major, L. mexicana
3 intraperitoneal injections of 1.5-2 µg of PSA-2 with Corinebacterium parvum in 2 week intervals; infection with 105 promastigotes after 2 weeks
Partial protection (smaller lesion size, lower parasite load in lymph nodes)
[93]
Nucleoside hydrolase protein
Mouse, BALB/c, 10 animals per group
L. major 2 injections of 2.5-25 µg of the protein with or without 1 µg of IL-12 in 2 weeks into the rump; 105of promastigotes after 1 week into the footpad
Partial protection after combination of the protein and IL-12 (smaller lesion size, lower parasite load in lymph nodes)
[94]
Nucleoside hydrolase (NH36) recombinant proteins
Mouse, BALB/c 4-8 animals per group, 2 exp. (L. chagasi), 5 animals per group, 2 exp. (L. amazonensis)
L. chagasi, L. amazonensis
3 subcutaneous inoculations of 100 µg of the protein with 100 µg of saponin; challenge with 3×107 amastigotes 4 weeks after
Partial protection (lower parasite load in liver); cross-protection (reduced lesion and parasite load)
[81]
Leish-111f polyprotein
Mouse, BALB/c
L. major 3 subcutaneous injections of 10 µg of Leish-111f with monophosphoryl lipid A plus squalene (MPL–SE) and IL-12 into the footpad; challenge with 2×105 promastigotes or 104 amastigotes after 3 weeks
Partial protection (smaller lesion size)
[98]
Leish-111f polyprotein
45 beagle dogs L. infantum 2 courses including 3 injections of 45 µg of Leish-111f with 50 µg MPL–SE or 1 µg Adjuprime in 4 week intervals; natural exposure to sand fly bites for infection
No effect [99]
Leish-111f polyprotein
Mouse, C57BL/6 (8 animals per group), golden Syrian hamster (5 animals per group)
L. infantum 3 subcutaneous injections of 10 µg of the protein in 3 week intervals into the foot pad; intravenous injection of 5×106 promastigotes 4 weeks after immunization
Partial protection (lower parasite load in spleen and liver)
[95]
Leish-110f polyprotein
Mouse, BALB/c, C57BL/6 3-5 animals per group
L. major L. infantum
3 subcutaneous injections of 0.5-10 µg of the protein with 20 µg MPL–SE or 5 µg of synthetic toll-like receptor 4 agonist (EM005) in 2-3 week intervals; challenge with 103 L. major promastigotes into the ear dermis 3-5 weeks later (BALB/c) or 5 x 106 promastigotes of L. infantum intravenously into the tail vein (C57BL/6)
Partial protection (smaller lesion size, lower parasite load in the ear – BALB/c after L. major infection, and lower parasite load in the liver after L. infantum infection of C57BL/6)
[100]
1448 Current Medicinal Chemistry, 2012 Vol. 19, No. 10 Kobets et al.
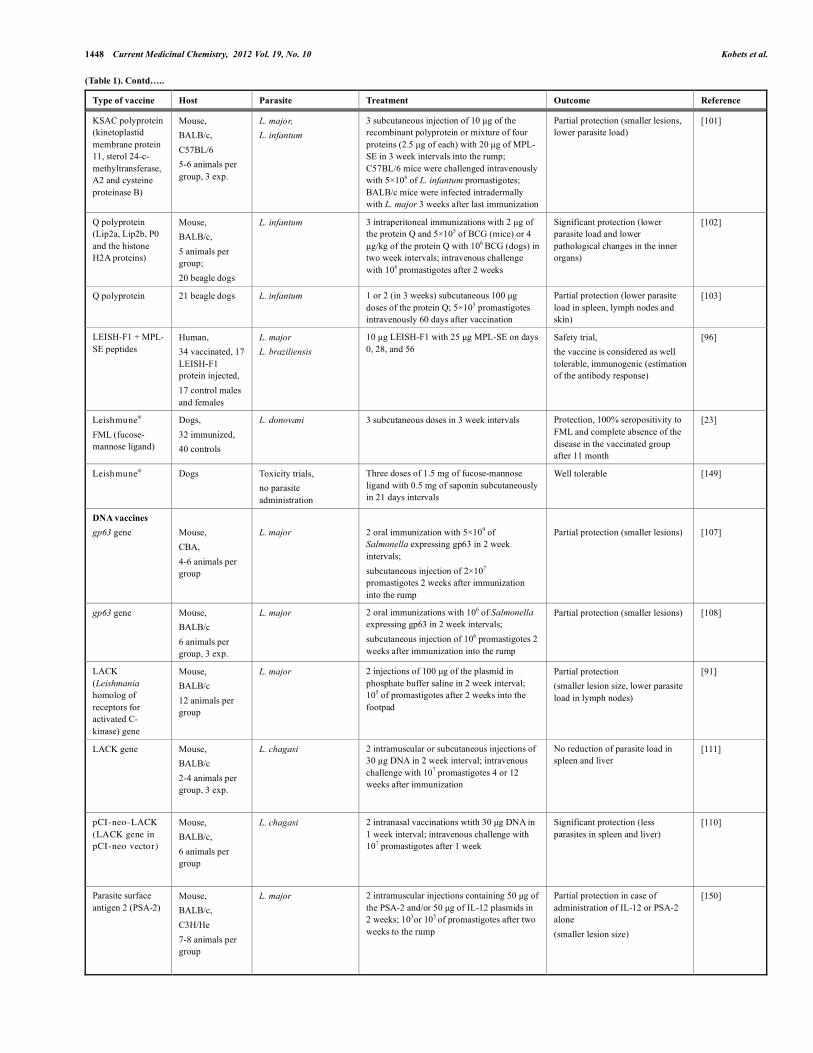
(Table 1). Contd…..
Type of vaccine Host Parasite Treatment Outcome Reference
KSAC polyprotein (kinetoplastid membrane protein 11, sterol 24-c-methyltransferase, A2 and cysteine proteinase B)
Mouse, BALB/c, C57BL/6 5-6 animals per group, 3 exp.
L. major, L. infantum
3 subcutaneous injection of 10 µg of the recombinant polyprotein or mixture of four proteins (2.5 µg of each) with 20 µg of MPL-SE in 3 week intervals into the rump; C57BL/6 mice were challenged intravenously with 5×106 of L. infantum promastigotes; BALB/c mice were infected intradermally with L. major 3 weeks after last immunization
Partial protection (smaller lesions, lower parasite load)
[101]
Q polyprotein (Lip2a, Lip2b, P0 and the histone H2A proteins)
Mouse, BALB/c, 5 animals per group; 20 beagle dogs
L. infantum 3 intraperitoneal immunizations with 2 µg of the protein Q and 5×105 of BCG (mice) or 4 µg/kg of the protein Q with 106 BCG (dogs) in two week intervals; intravenous challenge with 106 promastigotes after 2 weeks
Significant protection (lower parasite load and lower pathological changes in the inner organs)
[102]
Q polyprotein 21 beagle dogs L. infantum 1 or 2 (in 3 weeks) subcutaneous 100 µg doses of the protein Q; 5×105 promastigotes intravenously 60 days after vaccination
Partial protection (lower parasite load in spleen, lymph nodes and skin)
[103]
LEISH-F1 + MPL-SE peptides
Human, 34 vaccinated, 17 LEISH-F1 protein injected, 17 control males and females
L. major L. braziliensis
10 µg LEISH-F1 with 25 µg MPL-SE on days 0, 28, and 56
Safety trial, the vaccine is considered as well tolerable, immunogenic (estimation of the antibody response)
[96]
Leishmune® FML (fucose-mannose ligand)
Dogs, 32 immunized, 40 controls
L. donovani 3 subcutaneous doses in 3 week intervals Protection, 100% seropositivity to FML and complete absence of the disease in the vaccinated group after 11 month
[23]
Leishmune® Dogs
Toxicity trials, no parasite administration
Three doses of 1.5 mg of fucose-mannose ligand with 0.5 mg of saponin subcutaneously in 21 days intervals
Well tolerable [149]
DNA vaccines gp63 gene
Mouse, CBA, 4-6 animals per group
L. major
2 oral immunization with 5×109 of Salmonella expressing gp63 in 2 week intervals; subcutaneous injection of 2×107 promastigotes 2 weeks after immunization into the rump
Partial protection (smaller lesions)
[107]
gp63 gene Mouse, BALB/c 6 animals per group, 3 exp.
L. major 2 oral immunizations with 106 of Salmonella expressing gp63 in 2 week intervals; subcutaneous injection of 106 promastigotes 2 weeks after immunization into the rump
Partial protection (smaller lesions) [108]
LACK (Leishmania homolog of receptors for activated C- kinase) gene
Mouse, BALB/c 12 animals per group
L. major 2 injections of 100 µg of the plasmid in phosphate buffer saline in 2 week interval; 105 of promastigotes after 2 weeks into the footpad
Partial protection (smaller lesion size, lower parasite load in lymph nodes)
[91]
LACK gene Mouse, BALB/c 2-4 animals per group, 3 exp.
L. chagasi 2 intramuscular or subcutaneous injections of 30 µg DNA in 2 week interval; intravenous challenge with 107 promastigotes 4 or 12 weeks after immunization
No reduction of parasite load in spleen and liver
[111]
pCI-neo-LACK (LACK gene in pCI-neo vector)
Mouse, BALB/c, 6 animals per group
L. chagasi 2 intranasal vaccinations wtith 30 µg DNA in 1 week interval; intravenous challenge with 107 promastigotes after 1 week
Significant protection (less parasites in spleen and liver)
[110]
Parasite surface antigen 2 (PSA-2)
Mouse, BALB/c, C3H/He 7-8 animals per group
L. major 2 intramuscular injections containing 50 µg of the PSA-2 and/or 50 µg of IL-12 plasmids in 2 weeks; 103or 105 of promastigotes after two weeks to the rump
Partial protection in case of administration of IL-12 or PSA-2 alone (smaller lesion size)
[150]
Leishmaniasis: Prevention, Parasite Detection and Treatment Current Medicinal Chemistry, 2012 Vol. 19, No. 10 1449
(Table 1). Contd…..
Type of vaccine Host Parasite Treatment Outcome Reference
L. major signal peptidase
Mouse, BALB/c, 10 animals per group
L. major 2 injections of 30-100 µg of each plasmid in phosphate buffer saline in 3 week interval; 3×105of promastigotes after 3 weeks into the footpad
Partial protection (smaller lesion size, lower parasite load in lymph nodes)
[106]
Cocktail of plasmids encoding LACKp24, TSA, LmSTI1, CPa
Mouse, BALB/c 10 animals per group
L. major Intramuscular injection of 50 µg of each plasmid in phosphate buffer saline; 5×105 promastigotes after 2 weeks into the footpad or 500 promastigotes into the ear dermis
Complete protection only after low-dose infection (78% of mice with no lesions); partial protection during the standard infection
[109]
Cocktail of plasmids encoding cysteine proteinases I, II and III
Mouse, BALB/c, 13 animals per group
L. major 2 injections of 50 µg of each plasmid in solid lipid nanoparticles in phosphate buffer saline in 3 week intervals into the footpad; 3×106of promastigotes after 3 weeks into the footpad
Partial protection (smaller lesion size, lower parasite load in lymph nodes)
[112, 151]
Combination of TRYP (tryparedoxin peroxidase) plasmid with MVA (modified vaccinia virus Ankara) and TLR 1/2 agonist Pam3CSK4
Mouse, BALB/c, 8-10 animals per group
L. panamensis 2 injections, priming immunization with of 100 µg of TRYP DNA with or without 10 µg of Pam3CSK4, after 2 weeks, intraperitoneal boosting with 3×106 of TRYP-expressing MVA; 5×104 of promastigotes into the footpad 6 weeks after boosting
Partial protection, requiring TLR 1/2 activation (smaller lesion size, lower parasite load in lymph nodes)
[152]
Sand fly saliva Total saliva of Phlebotomus duboscqi
Mouse, BALB/c, 6 animals per group
L. major
Exposure to sand fly bites (30 sand flies per mouse) in 4 groups: for 15 days and for 2 days 2 weeks before infection; for 15 days and for 2 days immediately before infection; challenge with 106 promastigotes into the ear dermis
Protection only in case of short-time immunization (2 days) immediately before infection (smaller lesions, lower parasite load)
[123]
LJM11 salivary protein gene from Lutzomia longipalpis
Mouse, C57BL/6 5 animals per group, 2 exp.
L. major 3 injections with 5 µg of plasmid DNA in 15 day intervals into the ear dermis; 500 parasites after 15 days into the ear dermis
Partial protection (smaller lesion size, lower parasite load in the ear dermis)
[125]
Synthetic maxadilan (MAX) of Lutzomyia longipalpis
Mouse, CBA, 5 animals per group
L. major First injection 25 µg of MAX subcutaneously into the rump, second injection 25 µg of MAX intraperitoneally 10 days later, third – after 2 weeks; challenge with 106 L. major after 3 days
Partial protection (smaller lesions)
[124]
Protein SP15 (PpSP15) or plasmid encoding SP15 from Phlebotomus papatasi
Mouse, C57BL/6 4-5 animals per group
L. major 2 injections of 5-10 µg of plasmid DNA or 10 µl of the isolated salivary protein fraction into the ear dermis in 2 week interval; infection with 500 parasites 2 weeks later
Partial protection (smaller lesions) [126]
Plasmids encoding PpSP15, PpSP44 proteins of Phlebotomus papatasi
Mouse, C57BL/6 10 animals per group, 3 exp.
L. major Injection of 5 µg of plasmid DNA into the ear dermis; infection with 500 parasites 2 weeks later
PpSP15-immunized mice were partially protected (smaller lesions, less parasites in ear)
[127]
with L. infantum sir+/- led to complete elimination of wild type parasites after subsequent infection [57]. Complete protection against L. donovani in liver and partial protection against this parasite in spleens accompanied by a strong Th1 response was achieved after vaccination of BALB/c mice with L. donovani cen1-/- . The partially protective effect of this vaccine was observed also in hamsters [59]. Several mouse strains (Table 1) vaccinated with L. mexicana cpa/cpb-/- developed smaller lesion size after wild parasite strain challenge in comparison with naïve animals [54]. Interestingly, BALB/c vaccinated with L. major lpg2-/- were
substantially protected against virulent L. major without developing a strong Th1 response [56]. Partial protection was observed also after vaccination of BALB/c mice with L. major pmn-/- [58]. Vaccination of BALB/c and CBA/T6 mice with L. major deficient in dhfr-ts-/- led to substantial protection against L. major [51] and to partial protection against L. amazonensis in BALB/c and C57BL/6 mice [52], but had no effect in monkeys [53]. Attenuated vaccines usually have better effect when vaccination is performed shortly (1-2 weeks) before the challenge [51, 58]. Some mutants, for example L. major pmn-/- undergo a short life cycle [58], whereas L. major

1450 Current Medicinal Chemistry, 2012 Vol. 19, No. 10 Kobets et al.
lpg2-/- can persist in mouse up to whole lifetime [60] and can revert to virulence [61]. Suicidal Cassettes
Introduction into parasite genes, which will allow inducing suicide in response to external signals, such as antibiotics [62] or photoactivation [63] can prevent a long persistence and the possible reversion to virulence. A double drug sensitive strain of L. major was constructed by stably introducing into the chromosome a modified HSV-1 thymidine kinase gene and a Saccharomyces cerevisiae cytosine deaminase gene, conferring sensitivity to ganciclovir and 5-fluorocytosine, respectively, which made it possible to cure BALB/c mice infected by modified L. major using these antibiotics [62]; the protective effect of this parasite has not been tested. L. amazonensis, which episomally expressed human gene δ -aminolevulinate dehydratase and rat gene porphobilinogen deaminase, was intradermally injected into hamsters. Administration of δ -aminolevulinate led to accumulation of photosensitive uroporphyrin, which was excited by light to produce leishmanolytic oxidative species. This treatment led to protection against subsequent challenge with L. donovani [63]. Thus, these experiments show an interesting alternative for development of safe live vaccines. 2.1.1.4. Killed Parasites
A killed vaccine consists of pathogen, which is grown in culture and then killed by heat [64, 65], formaldehyde [66], by repeated cycles of freezing and thawing [67] or by autoclaving [65]. The advantage of killed vaccines is safety and low cost. These should be prepared in standardized conditions to have stable biochemical composition and immunogenicity.
Some studies with killed Leishmania were reported from 1930s and 1940s, but controlled trials were conducted only after 1970 [68, 69]. Noazin and coworkers [68] analyzed efficacy of vaccines prepared using killed whole parasites that were used in Iran, Sudan, Brazil, Colombia, and Ecuador and found that these vaccine candidates do not confer significant protection against human leishmaniasis [68]. Vaccines with killed parasites could be ineffective due to several reasons. Autoclaving of parasites, which is usually used in preparation of human vaccines lowers the immunogenicity by destroying most of the proteins [70]; in addition, inhibitory determinants might appear on surface. Replication, metabolic activity and persistence of parasites might be also necessary for the development of the protective immunity. Comparison of killed and live vaccines in a mouse experiments has shown that live parasites induce better immune response than the dead ones [57, 59, 64].
New ways to deliver vaccines, such as nanovaccines might improve the efficacy of whole-killed vaccines. Poly(D,L-lactide –co-glycoside) nanospheres as an antigen delivery system and cytosine-phosphodiester-guanine oligodeoxynucleotide (CpG ODN) as an adjuvant have been used to enhance the immune response against autoclaved L. major in BALB/c mice. Highly significant, although not complete protection against subsequent infection was observed [71].
Although an efficient killed prophylactic Leishmania vaccine has yet to be developed, killed parasites vaccines have been successfully used to improve the chemotherapy. For example, the combination of killed promastigotes of L. amazonensis with a half dose regimen of the pentavalent antimonials was highly effective for the treatment of American cutaneous leishmaniasis [72]. 2.1.2. Vaccination with Parasite Subunits
The use of parasite subunits is based on the fact that pathogen contains both the epitopes that enhance immune response against it and those that aggravate the disease [73, 74]. So, the aim is to selects proteins with epitopes enhancing host defense against the
pathogen. This selection is based on the information about the abundance and important roles of certain parasite proteins in pathogen biology, and/or the entire genome sequence is used to identify vaccine candidates. In the latter approach, which was denominated “reverse vaccinology” a large number of candidate antigens are expressed in the heterologous expression system, purified, and used to immunize mice. As a result, the proteins inducing beneficial antipathogen responses are selected [75, 76]. To date, at least 30 vaccine subunits against leishmaniasis have been tested [77, 78]. In most experimental systems that use pathogen-derived proteins, adjuvants are essential to provoke protective immunity. However, the most effective adjuvants generally cause strong inflammation, which may be essential for adjuvanticity, but may preclude their use in humans because of unacceptable side effects [21]. 2.1.2.1. Vaccination with Leishmania Fractions
Leishmune® consists of an affinity purified L. donovani promastigote glycoprotein fraction, whose composition is not completely defined, named fructose mannose ligand (FML) and the adjuvant consisting of aldehyde-containing deacylated saponins of Quillaja saponaria [79]. A fucose-mannose ligand (FML), also present on Leishmania surface, is now commercially available and was used for the treatment of dogs and proved to prevent development of canine leishmaniasis and to block its transmission [23, 79], which led to decrease of the incidence of human and visceral leishmaniasis in Brazilian endemic areas [80]. Vaccination of BALB/c mice with C-terminal domain of a secreted nucleoside hydrolase, which is the dominant antigen in the FML, has been shown to reduce parasite numbers in livers, parasite load in lesions, and lesions size after challenge with L. chagasi and L. amazonensis, respectively [81]. The first European canine Leishmania vaccine CaniLeish® (LiESAp) was released in 2011 (http://ec.europa.eu/ health/documents/community-register/html/vreg.htm.txt). It con-sists of excreted secreted proteins from Leishmania infantum (LiESAp), the dominant antigen of which is the promastigote surface antigen [82]. 2.1.2.2. Vaccination with Defined Proteins
Range of early studies showed success in the use of peptide vaccines in animal models [83, 84]. The most comprehensively studied anti-leishmanial vaccine candidate is the surface- expressed 63 kDa glycoprotein (gp63), or leishmanolysin, tested for development of vaccine against L. major [83-87]. This glycoprotein metalloprotease is a parasite receptor for host macrophages and functions in the receptor mediated uptake of promastigotes by macrophages in the mammalian host. Hence, mutant parasites lacking the protein have reduced virulence [86]. Some regimens of vaccination with gp63 led to protection against leishmaniasis [83-86], whereas others had no influence or even exagerated disease [78, 85].
The LACK antigen (Leishmania homologue of receptor for activated C kinase) 36 kDa is expressed by both promastigote and amastigote form of the parasite [88]. The LACK antigen, which is MHC class II associated, is an analogue of the mammalian receptor for activated protein kinase C (RACK) [89]. It belongs to a protein family containing the repeat motif termed WD 40 that is present in a large number of eukaryotic genes [90]. Vaccination of BALB/c mice with LACK 24kDa protein in combination with IL-12 led to partial protection against L. major [88, 91].
PSA (promastigote surface antigen) is one of the major classes of membrane-bound or secreted proteins of the parasitic protozoan Leishmania, whose main signature consists of a specific LRR (leucine rich repeats) sequence. All PSA genes found in the genomes of three sequenced Leishmania species distribute into eight subfamilies of orthologs. Seven of these subfamilies correspond to basic functions related to parasite/host interactions, the other PSA gene class, which include all so far experimentally

Leishmaniasis: Prevention, Parasite Detection and Treatment Current Medicinal Chemistry, 2012 Vol. 19, No. 10 1451
studied PSA genes, could be involved in more specialized adaptative functions [92]. Intraperitoneal vaccination of C3H/He mice with PSA-2 with Corynebacterium parvum as an adjuvant resulted in complete protection from lesion development after a challenge infection with virulent L. major. Significant protection was also obtained in the genetically susceptible BALB/c-H2k and BALB/c mice [93]. Partial protection against L. major was achieved also with vaccination of BALB/c mice with nucleoside hydrolase protein [94]. Large number of other potential candidates for vaccine design in this field is extensively studied [78]; however, this approach has brought conflicting results in many cases due to possible conformational changes and insufficient immunogenicity of individual separated peptides, which could be effective only in cocktails [21].
The LEISH-F1 + MPL®-SE vaccine is composed of the recombinant Leishmania polyprotein LEISH-F1 (formerly known as Leish-111f) antigen and the MPL®-SE adjuvant. The antigen component of the vaccine includes three proteins derived from L. major and conserved across various Leishmania species, including L. donovani and L. chagasi, which causes Old World and New World visceral leishmaniasis, respectively; and L. braziliensis, which causes both cutaneous leishmaniasis and mucosal leishmaniasis in the New World. The three proteins are: Leishmania elongation initiation factor (LeIF), thiol-specific antioxidant (TSA), and Leishmania major stress-inducible protein 1 (LmSTI1). The adjuvant component is a potent TLR4 agonist - monophosphoryl lipid A, which is derived from the lipopolysaccharide of Salmonella enterica serovar Minnesota and formulated in the adjuvant monophosphoryl lipid in a stable squalene oil-in-water emulsion (MPL®-SE) [95-97]. Immunization trials in mice demonstrated that Leish-111f was able to protect BALB/cByJ mice against L. major and L. amazonensis infection [98] and to induce partial protection against visceral leishmaniasis in C57BL/6 mice and golden Syrian hamsters [95]. However, Leish-111f failed to protect dogs against infection and did not prevent disease development in a Phase III trial in dogs [99]. The Leish-111f polyprotein was modified in order to eliminate a potential regulatory concern and an apparent proteolytic hot spot was eliminated and the new 110 kDa construct was named Leish-110f. Vaccination with this construct and MPL®-SE adjuvant completely protected BALB/c mice against lesion development after L. major infection and partially protected C57BL/6 mice against visceral leishmaniasis after infection with L. infantum [100].
KSAC is a polyprotein vaccine composed from fused proteins KMP-11 (kinetoplastid membrane protein 11), SMT (sterol 24-c-methyltransferase), A2 and CPB (cysteine proteinase B). Administration of KSAC and MPL®-SE adjuvant strongly, but not completely protected BALB/c mice against L. major infection and C57BL/6 mice partially protected against L. infantum [101].
Another multi-component antigenic protein, named Q, was formed by the genetic fusion of five fragments from the L. infantum acidic ribosomal proteins Lip2a, Lip2b, P0 and the histone H2A protein and administered with BCG adjuvant. This led to a strong protection of BALB/c mice and dogs against L. infantum infection [102]. Q protein protected dogs against L. infantum also when administered without adjuvant [103]. 2.1.3. DNA Vaccines
The concept of DNA vaccination was established by Wolff and coworkers in 1990 [104]. DNA vaccines include cloned genes of the vaccine candidates, which are injected directly into muscle tissue or skin. A plasmid is taken up and expressed by the host cells, revealing a properly folded protein of interest that causes immunization [21, 105] and usually induces more effective protective responses than vaccination with isolated protein [91, 106]. Among the first candidates for DNA vaccines was the gene for gp63 [107, 108], mentioned above (section 2.1.2.2.). Vaccines
based on gp63 induced partial protection against L. major in CBA [107] and BALB/c mice [108]. LACK (Leishmania homologue of receptors for activated C kinase) is the most extensive DNA vaccine studied so far against both cutaneous [91, 109] and visceral leishmaniasis [110, 111]; partial protection was achieved in BALB/c mice during challenge with L. major [91, 109]. Moreover, a cocktail DNA vaccine containing LACKp24 (a truncated portion of the LACK antigen), TSA (L. major homolog of the eukaryotic thiol-specific-antioxidant) and CPa (cysteine proteinase A) was able to induce complete protection after low-dose infection with L. major in BALB/c [109]. Subcutaneous or intramuscular vaccination of mice with plasmid encoding p36 L. infantum LACK protein failed to protect of BALB/c from the subsequent infection with L. chagasi although it induced strong IFN-γ production [111], whereas intranasal vaccination with the same plasmid together with the shortening the interval between last booster and infection (from 4 or 12 weeks to 1 week) led to strong, but not complete protection [110]. Immunization of BALB/c mice with a cocktail DNA vaccine, encoding cysteine proteinases type I, II, and III with solid lipid nanoparticles, potentiated protective immunity against L. major infection [112]. Knowledge of genome sequence of Leishmania [113] enabled to test 100 candidate DNA vaccines against L. major. Fourteen protective novel vaccine candidates were identified, seven vaccines exacerbated disease. Two protective antigens were identified as ribosomal proteins 60S ribosomal L22 and 40S ribosomal S19, three other had significant matches to previously identified proteins V-ATPase subunit F, dynein light chain, and amastin-like protein. Functions of remaining nine protective proteins are not known. The best novel protective antigen was an amastin-like gene that conferred significant, but not complete, decrease of footpad swelling [73].
2.2. Vaccines Based on Transmitting Phlebotomine Vector
2.2.1. Whole Saliva The possibility of vaccine development using sand fly saliva
was also considered. Sand flies are bloodsucking insects that are natural vectors of Leishmania parasites (see section 1). They deposit parasites into the host skin along with saliva, which contains immunomodulatory molecules [114, 115] that induce species-specific humoral and cellular response in the host [116-119]. Local inhabitants in endemic areas develop a specific antibody response to salivary antigens, which correlates with protection against visceral leishmaniasis [120], but not against cutaneous leishmaniasis [121, 122]. The history of immunization may also significantly change the character of anti-saliva immune response with substantial consequences for development of Leishmania. Using a mouse model, it was shown that the protection took place only in a case of short-term exposure to bites of Phlebotomus duboscqi (two days exposure to bites) right before the challenge with L. major parasites, but not in cases of long-term exposure (15 days) or short or long immunization periods followed by a significant time interval before the infection occurred (Table 1) [123]. This finding is in agreement with the field results from an endemic area of cutaneous leishmaniasis caused by L. braziliensis [122]. 2.2.2. Saliva Components
Saliva components were isolated from both New World [124, 125] and Old World [126, 127] phlebotomine sandflies and their prophylactic ability has been tested. Vaccination with Lutzomyia longipalpis salivary protein maxadilan partially protected CBA mice from L. major infection. The protective role of a LJM11 (a yellow protein from saliva of Lutzomyia longipalpis) was proved by immunization of C57BL/6 mice with peptide-encoding plasmids and subsequent infection with L. major [125]. A DNA vaccine coding Phlebotomus papatasi 15 kD salivary protein (PpSP15/SP15) from the vector of conferred partial protection

1452 Current Medicinal Chemistry, 2012 Vol. 19, No. 10 Kobets et al.
against L. major to C57BL/6 mice [126], which was accompanied by the increase of IFN-γ RNA transcripts and decrease of IL-4 transcripts in the mouse ear 2 hours after immunization [127].
2.3. Problems of Vaccination Against Leishmaniasis and Novel Vaccine Strategies
Despite the wide range of studies carried out during quite a long period of time, there is no reliable vaccine against leishmaniasis available at the present moment. The cost of developing a vaccine has been estimated to be hundreds of millions of dollars; 60–80% of this is allotted to preclinical and clinical development of the vaccine. Except for a few cases, funding for human vaccine development has been minimal until very recently. About 90% of the burden of VL is present in five countries (India, Bangladesh, Sudan, Ethiopia and Brazil). Those affected in these countries are among the poorest of the poor, and hence there is not a large enough market for pharmaceutical companies to invest a considerable amount of time and money to develop a vaccine [69]. In addition, different research groups may obtain contradictory results after testing the same candidate target for vaccine development. This might be partly due to different conditions of the experiments, as many factors influence immunogenicity and efficacy of vaccination: type of the vaccine [86], the dose, the route [128], and regimen [58, 123] of administration, type of adjuvant [66, 86] and the mode of virulent challenge [109]. The same might be true for differences in outcomes of experimental and human trials. Moreover, few of the mouse experiments are completely protective (Table 1) and usually the most efficient protection is observed when challenge with virulent parasite is performed shortly after vaccination. The short time protection might be beneficial for tourists traveling to the endemic regions, but it is not suitable for the local population.
The complexity of immune response to leishmaniasis is not yet fully explored. Some vaccines induce Th1 type of immune response, that is considered to be protective in leishmaniasis, but have no or a limited influence on organ pathology [111, 129, 130]. This is in agreement with genetic studies in mouse, where some genes influence both organ pathology and immune response including cytokine levels, whereas the others determine only cytokine levels [28, 131, 132]. Better definition of pathways that are involved in control of pathology will help to prepare more effective vaccines.
There is another important point that needs to be mentioned. Majority of vaccination experiments are performed on the mouse strain BALB/c [78] (Table 1). This is not representative of different genotypes present in the outbred human population. Moreover, it was shown that several patterns of the immune response are observed after infection with L. major, depending on the host genotype [133, 134]. Different regulatory pathways operating in different genetic backgrouds could lead to different response to vaccination. Indeed, Benhnini and coworkers [135] have shown that capacity of L. major proteins LACK (Leishmania homolog of receptor for activated C kinase) and LmPDI (L. major protein disulfide isomerase) to confer protection to L. major infection depended on the genotype of the mouse strain. In PWK and BALB/c mice vaccination with both proteins led to substantial and partial decrease of lesion size, respectively, whereas in C57BL/6 mice none of the proteins conferred protective response. Vaccination of MAI mice with LmPDI led to complete protection, while LACK had no influence on immunity. Thus, to more tightly mimic the situation in the outbred human population, larger spectrum of mouse strains needs to be tested. The antigen that confers protection in the larger range of inbred strains may have better chances to be also protective in outbred human population and should be selected for clinical trials [135]. Hence, integration of genetic studies with methods which will allow to select more
immunogenic candidate vaccine antigens by employing methods of reverse vaccinology [73, 75, 76, 136], proteomics [137] and protein engineering [138] together with the advanced delivery systems such as nanoparticles [71, 112, 139], dendritic cells [140-142], and dendritic cell-derived exosomes [141] can help to produce more efficient vaccines against leishmaniasis (Table 1).
The lack of a safe vaccine or chemoprophylaxis limits the options for prevention of leishmaniasis to elimination of reservoir populations and some form of vector control, including barriers to sand fly feeding [143]. Therefore the early pathogen detection and identification that will be described in the following section has the key importance for a timely disease treatment.
3. ASSAYS FOR PARASITE DETECTION AND QUANTI-FICATION
As with any infectious disease, spread and load of the pathogen during leishmaniasis is an important parameter, as well as the precise diagnostics of Leishmania species. Until recently, this task was a significant complication for practical doctors and researchers. The optimal methods should be fast, simple and price-worthy. Easy-to-use techniques for rapid pilot tests during field application in the absence of the equipped laboratory are also highly necessary. The main approaches for parasite detection include DNA-based techniques, microscopy assays, cell cultures, and immunodetection [153, 154].
3.1. Microscopy for Detection, Quantification and Histological Studies
3.1.1. Classical and Fluorescent Microscopy Microscopy assays were among the first methods for detection
and quantification of a wide range of pathogens, including Leishmania parasites. This kind of methods allows detecting and counting parasites directly. Histological examination of biopsies can give information about parasite load, infiltration of different cell types, formation of granulomas and other changes in Leishmania invaded tissues, reflecting the pathogenesis. Hematoxylin and eosin (H&E) or Giemsa are widely used for routine staining of tissue smears and biopsies, for example, [40, 53, 63, 102, 122, 132, 153-165], as well as various fluorescent labels, for example, [155, 157, 166-168]. Microscopy belongs to the gold standard assays, being effective both for stand-alone studies [163, 167, 169] and validation of the novel methods [132, 154, 156, 158, 159, 161, 170]. Among the most significant drawbacks of the microscopy assays is the fact that the direct detection of parasites on stained smears from aspirates of bone marrow, lymph node or spleen of patients, requires invasive procedures [171]. Microscopy analysis of large numbers of samples is usually time consuming. Moreover, quantification of parasites in certain samples may not reflect the real parasite load, because parasites are distributed in tissues unequally. 3.1.2. Electron Microscopy
Interactions between Leishmania parasites and macrophages can be studied in details using electron microscopy. Scanning electron microscopy has revealed that the phagocytosis of L. amazonensis metacyclic promastigotes begins with the engulfment of the parasite body and ends with the progressive internalization of the flagellum. Transmission electron microscopy has revealed that promastigotes are located in very long phagosomes, in which the membrane tightly follows the outline of the parasite including that of their flagellum. Immunogold cryosection electron microscopy has been useful for improving the understanding regarding the distribution and trafficking of major histocompatibility complex class II molecules in macrophages parasitized by L. amazonensis [169].
Leishmaniasis: Prevention, Parasite Detection and Treatment Current Medicinal Chemistry, 2012 Vol. 19, No. 10 1453
3.1.3. In-Vivo Imaging The data obtained from imaging fixed samples gave a very
incomplete picture of the dynamic nature of the complex processes Leishmania drives in both the insect and the mammalian host. Emerging technologies using fluorescence and bioluminescence imaging have been recently adapted for the study of host–Leishmania interactions [166] including response to therapy [166, 167]. Bioluminescence imaging of mice inoculated with transgenic Leishmania expressing the firefly luciferase provides an efficient and reliable method for delineating the various phases of infectious processes. Whole-body imaging with fluorescent parasites has also recently been developed to monitor light emission without substrate addition, making use of different colored fluorophores for multiplex imaging. Because of its nondestructive and noninvasive nature, the procedures for bioluminescent and fluorescent imaging can be performed repeatedly, allowing each animal to be used as its own control over time, overcoming the problem of animal–animal variation [166, 167].
3.2. Cell Culture Based Methods
Cell cultures include isolation and cultivation of Leishmania from cells and tissues. Viable parasites multiply and their numbers are evaluated after several days of incubation in the nutrient solution. This group of methods, along with microscopy assays,
belongs to the first techniques developed for the estimation of parasite load in the infected host organism [36, 159, 161, 172, 173]. 3.2.1. Limiting Dilution Assay
The majority of tissue culture techniques are based on limiting dilution assay [172, 173]. These methods use a range of serial dilutions to assess the ratio of cells containing viable parasites. Limiting dilution assay, originally developed for Leishmania detection and quantification, is a highly laborious and time consuming technique. Detection of viable parasites is the main advantage of this method. Since culture methods target viable cells, they should be performed immediately after isolation of the tissues and require sterile conditions. 3.2.2. Microculture and Miniculture
A microcapillary culture method (MCM) was developed for diagnosis of cutaneous leishmaniasis. In contrast to traditional culture method, MCM is more rapid, uses smaller sample, and has higher sensitivity for detection of promastigotes. In comparative studies, the average time period of incubation needed to detect promastigotes was much shorter with the microcultures than the conventional cultivation and required 2-7 days versus 2-30 days. The high sensitivity of the MCM may be explained by the use of capillary tubes, which concentrate the sample material and provide microaerophilic conditions with high CO2 that is favorable for transformation of amastigotes to promastigotes [161, 174].
Table 2. Specific Targets Used in DNA-Based Assays for Detection and Quantification of Leishmania spp.
Target Description References
kDNA Minicircle kinetoplast DNA. Each parasite contains about 10 000 of minicircles. [154, 156, 159, 180, 181]
18S rRNA / ssu-rRNA
The gene that encodes RNA included to the small ribosomal subunit. [171, 182-184]
mini-exon A very small gene of 39 nucleotides, present in Kinetoplastida. An intron, adjacent to the mini-exon, is conserved. It is followed by a sequence that varies in length considerably between species and is much less conserved.
[184, 185]
ITS1 region Internal transcribed spacer 1: the sequence located between the 18S ribosomal RNA and 5.8S ribosomal RNA genes.
[184, 186]
ITS2 region Internal transcribed spacer 2: the sequence located between the 5.8S ribosomal RNA and 28S ribosomal RNA genes.
[165]
7SL RNA
7SL RNA together with six proteins forms a signal recognition particle which mediates protein translocation across the endoplasmic reticulum in eukaryotes, including Leishmania.
[187]
gp63 The gene coding Leishmania surface protease; highly conserved. [188]
hsp70 The gene of 70kDa heat shock protein; highly conserved among eukaryotes, however, contains some variable regions; used for phylogenetic studies.
[189, 190]
DNA/RNA polymerases
Genes that encode DNA and RNA polymerases. Their sequences were used to reveal evolution of Leishmania. [191]
cyt b Cytochrome b gene is present in the mitochondrial genome, encodes the central catalytic subunit of an enzyme present in the respiratory chain of mitochondria; the gene was used for phylogenetic studies.
[192]
g6pdh The gene coding glucose-6-phosphate dehydrogenase. [193, 194]
icd The gene coding isocitrate dehydrogenase [193]
me The gene encoding cytosolic NADP-malic enzyme [193]
mpi The gene encoding mannose phosphate isomerase [193]
fh The gene encoding fumarate hydratase [193]
cpb A cluster of genes for cysteine protease B [195]
asat The gene encoding aspartate aminotransferase [176]
gpi The gene encoding glucose-6-phosphate isomerase [176]
nh1 The gene coding nucleoside hydrolase 1 [176]
nh2 The gene coding nucleoside hydrolase 2 [176]
pgd The gene coding 6-phosphogluconate dehydrogenase [176]

1454 Current Medicinal Chemistry, 2012 Vol. 19, No. 10 Kobets et al.
Miniculture method, in which parasites are cultivated inside the Eppendorf tubes, also showed better results than conventional culture methods [174]. 3.2.3. Multilocus Enzyme Electrophoresis (MLEE) for Parasite Classification
MLEE is based on the isoenzyme analysis and also requires preparation of parasite cultures. It is one of the most comprehensive methods used for identification of Leishmania, particularly the MON system, which was developed in Montpellier, France. MON system is based on 15 enzymes (malate dehydrogenase, malic enzyme, isocitrate dehydrogenase, 6-phosphogluconate dehydrogenase, glucose-6-phosphate dehydrogenase, glutamate dehydrogenase, NADH diaphorase, purine nucleoside phosphorylase, purine nucleoside phosphorylase, 2 glutamate-oxaloacetate transaminases 1 and 2, phosphoglucomutase, fumarate hydratase, mannose phosphate isomerase, glucose phosphate isomerase) [175-177]. MLEE includes Leishmania parasite isolation and cultivation, followed by preparation of enzyme extracts from the promastigotes pellets. Extracts are analyzed by electrophoresis. The distance that each reproducible enzyme band migrates from the origin (anode) is measured. The obtained set of bands defines the zymodeme, an electrophoretic profile for each extract. Attribution of the species is made by comparison of the profiles with the reference Leishmania strains [178, 179]. Isoenzymes have been used to generate phylogenetic trees and to provide a basis of the current taxonomy of L. donovani complex, which has three designated species, L. donovani, L. infantum and L. archibaldi [176]. For diagnostic purposes, lesion aspirates from patients were analyzed by MLEE [178, 179].
3.3. Assays that Detect Parasite DNA
In general, DNA-based methods were developed to detect presence of different regions of parasite DNA in experimental or clinical samples (Table 2). Techniques of this type serve two main purposes. First, they can be used for detection of Leishmania presence and measurement of parasite load in cells and tissues. Second, species- or subspecies-specific methods help to identify the parasite and make precise diagnosis. 3.3.1. Conventional Polymerase Chain Reaction (PCR), Nested PCR, and Real-Time PCR (RT-PCR) for Parasite Detection and Quantification
PCR is an extremely sensitive tool for detection of target DNA from various sources, including Leishmania. Minicircle kinetoplast DNA (kDNA) contains conserved regions that are widely used for Leishmania detection and quantification. Each parasite contains multiple copies of kinetoplast minicircles that makes kDNA a very prominent PCR target. First PCR using primers specific to conserved regions of kDNA was tested on seven Leishmania species [180] (Table 3). Amplicon can be analyzed on agarose electrophoresis or dot blot [159, 180]. Besides kDNA, a highly conserved gene gp63 (encoding the surface protease), mini-exon (a very small gene of 39 nucleotides, present in Kinetoplastida), and ITS1 region (a sequence located between the 18S ribosomal RNA and 5.8S ribosomal RNA genes) were proved to be good targets both for classical PCR combined with electrophoresis and real-time PCR, but their sensitivity was either comparable or lower than the one obtained with kDNA [184, 188, 196-198]. Small ribosomal subunit gene (ssu-rRNA) was among the first targets proposed for the Leishmania-specific PCR assays development [171, 182].
Nested PCR is based on application of two pairs of primers and includes two rounds. Additional internal primer set, specific for the particular sequences of the first amplicon, can significantly increase the sensitivity of the technique [156, 185]. For distinguishing among Leishmania species, the first round PCR using universal primers (for sequences that are conservative in all tested strains)
should be followed by the second round PCR with specific primers that amplify the sequences that are unique for each tested strain [165].
Real-time PCR (RT-PCR) allows analysis of formation of the amplicon that makes it possible to quantify the initial number of template molecules. RT-PCR assays for Leishmania detection are based on the DNA polymerase gene, kinetoplast and ribosomal DNA. Most of these tests are not species specific. In Real-time PCR, the product is detected by the fluorescent labels [194, 196, 199, 200]. In addition to the detection and quantification, RT-PCR served for Leishmania species typing. The test was developed using a polymorphism in the glucose-6-phosphate dehydrogenase locus and was performed with SYBR-green or TaqMan master mixes. This enabled identification and quantification of wide range of Leishmania species found in the Americas. Although any amplification assay based on a single-copy target is less sensitive than the one based on multi-copy targets, the method was effective for identification and quantification of parasites in human biopsy samples [194]. SYBR-green binds nonspecifically to any double-stranded DNA what comprises the major limitation of this system. Therefore, the highly specific primers and precise optimization of the synthesis conditions are necessary. In addition, background fluorescence may diminish both sensitivity and specificity [200]. The ITS1 region is commonly used target in many different eukaryotic organisms for typing purposes [184, 201-203]. It has enough conservation to serve as a PCR target but sufficient polymorphisms to facilitate species typing. The sensitivity of this RT-PCR was similar to the conventional PCR [184].
As mentioned above (Table 2), a variety of targets are used for PCR. kDNA is one of the most reliable targets for Leishmania detection and quantification since there are ~10 000 minicircles per parasite. The sensitivity and specificity of this method were compared with two other PCR assays (mini-exon and ITS1), leishmanial culture and microscopic detection in order to validate these techniques for molecular diagnosis of cutaneous leishmaniasis. The kDNA PCR was the most sensitive diagnostic assay and should be employed when species identification is not required. PCR using ssu-rRNA also showed lower sensitivity than kDNA PCR [171, 182]. However, when further parasite characterization is needed, the ITS1 PCR is both highly sensitive and specific and enables the identification Leishmania species. PCR is now the diagnostic method of choice for cutaneous and mucocutaneous leishmaniasis, and kDNA PCR is the gold standard against which all new techniques should be compared [186, 197, 204]. 3.3.2. PCR-ELISA for Parasite Detection and Quantification
Important progress was achieved with application of PCR-ELISA, which combines conventional PCR with enzyme-linked immunosorbent assay (ELISA) for the detection of labeled product. Conserved regions of kinetoplast minicircle appeared to be the preferential target used for testing [153, 154, 200, 205-207]. PCR-ELISA has sensitivity comparable to RT-PCR and allows detection of small differences in parasite load. However, some steps of this technique contain drawbacks that may influence the final result. If the product is not purified, its hybridization with the biotinylated [205, 206] or digoxigenin-labeled probes [200, 207] increases the possibility of an incomplete or nonspecific binding of the probe to the amplicon, or even DNA, and distortion of the result. Two labeled primers can be used to omit the step of hybridization. Highly optimized reaction with sensitive kDNA primers, originally labeled with biotin and digoxygenin, excludes probability of nonspecific effects, allows precise quantitative measurement of the parasite load and a fast analysis of large number of samples [154]. The improved method has been already successfully applied in two different experimental projects [123, 132].
Leishmaniasis: Prevention, Parasite Detection and Treatment Current Medicinal Chemistry, 2012 Vol. 19, No. 10 1455
Table 3. Methods for Leishmania Parasite Detection
Assay Leishmania species
Target Sensitivity (lowest detectable parasite load)
Samples Reference
Giemsa staining L. mexicana L. major
parasite cells
N/T skin biopsies [153]
Acridin orange staining L. donovani parasite cells
N/T parasite culture [155]
H&E staining L. tropica parasite cells
N/T skin biopsies [163]
Electron microscopy
L. amazonensis L. major
parasite cells
N/T macrophages [169]
L. infantum parasite cells
in vivo: 20000-40000 amastigotes/mg tissue; ex vivo: 1000-6000 amastigotes/mg tissue
whole mouse
[166]
Microscopy
In-vivo imaging
L. major parasite cells
N/T whole mouse
[167]
Limiting dilution assay L. major viable parasites
N/T skin biopsies [173]
Microcapillary culture method (MCM)
L. tropica viable parasites N/T skin biopsies [161]
Microcultures and minicultures
Leishmania spp. viable parasites N/T lesion aspirates [174]
Culture-based methods
Multilocus enzyme electrophoresis (MLEE)
L. major L. tropica
12 enzymes N/T lesion aspirates [178]
Polymerase chain reaction (PCR)
L. braziliensis L. mexicana L. major L. chagasi L. donovani L. aethiopica L. enriettii
kDNA 0.1-10 fg parasite DNA, skin biopsies
[180]
PCR + dot blot hybridization
L. braziliensis L. panamensis L. peruviania L. guyanensis
kDNA 0.1 fg parasite DNA, skin biopsies, lesion aspirates, lesion scraping
[159]
PCR L. infantum L. donovani
ssu-rRNA gene 10 parasites in 1 ml parasite DNA, blood
[171, 182]
PCR L. braziliensis L. amazonensis
kDNA N/T Giemsa stained slides
[181]
Nested PCR L. tropica kDNA 0.1 fg parasite DNA, skin biopsies
[156]
Nested PCR L. donovani mini-exon 100 fg parasite DNA, tissue aspirates
[185]
Nested PCR L. major L. gerbilii L. turanica
ITS2 N/T parasite DNA, ear biopsies
[165]
DNA-based methods
Real-time PCR (RT-PCR) L. major L. donovani L. amazonensis L. mexicana
kDNA 100 fg parasite DNA [196]
1456 Current Medicinal Chemistry, 2012 Vol. 19, No. 10 Kobets et al.
(Table 3). Contd…..
Assay Leishmania species
Target Sensitivity (lowest detectable parasite load)
Samples Reference
Real-time PCR (RT-PCR) L. braziliensis L. peruviania L. guianensis L. panamensis L. lainsoni L. utingensis L. amazonensis L. mexicana L. pifanoi L. infantum L. adleri
g6pdh 10 parasites in 0.2 ng of total DNA
parasite DNA, skin biopsies, mice footpad
[194]
Real-time PCR (RT-PCR) L. donovani L. major L. chagasi L. mexicana L. braziliensis
gp63 10 fg parasite DNA [188]
Real-time PCR (RT-PCR) L. donovani ITS1 10 fg parasite DNA [184]
PCR + enzyme-linked immunosorbent assay (PCR-ELISA)
L. braziliensis kDNA 0.5-0.6 pg parasite DNA, blood
[205]
PCR-ELISA L. infantum kDNA 1 fg parasite DNA, skin biopsies
[206]
PCR-ELISA L. major kDNA 0.3 fg parasite DNA, lymph nodes
[154]
Oligo-C-TesT L. guyanensis L. braziliensis
18S rRNA gene
N/T lesion scrapings [183]
18S rRNA gene
100 fg
Mini-exon 50 fg
Restriction fragment length polymorphism (RFLP)
L. donovani L. infantum L. tropica L. aethiopica L. braziliensis
ITS1 10 fg
parasite DNA, skin biopsies, blood, bone marrow
[184]
RFLP + reverse dot blot (RDB)
L. donovani complex L. major L. tropica L. aethiopica
7SL RNA gene 20 fg parasite DNA, skin biopsies
[187]
Reverse line blot (RLB) L. donovani L. major L. tropica L. aethiopica L. infantum
ITS1 60 fg – 5 pg, depending on probe
parasite DNA, skin biopsies
[186]
Sequencing
L. donovani L. infantum L. chagasi L. major L. tropica L. arabica L. aethiopica L. mexicana L. amazonensis L. braziliensis L. panamensis L. hertigi L. deanei L. herreri L. hoogstraali L. adleri L. gymnodactyli L. tarentolae Endotrypanum monterogeii
DNA polymerase a catalytic polypeptide and RNA polymerase II largest subunit genes
N/T
parasite DNA
[191]
Leishmaniasis: Prevention, Parasite Detection and Treatment Current Medicinal Chemistry, 2012 Vol. 19, No. 10 1457
(Table 3). Contd…..
Assay Leishmania species
Target Sensitivity (lowest detectable parasite load)
Samples Reference
Multilocus sequence typing
L. donovani L. infantum L. archibaldi
asat gpi nh1 nh2 pgd d6pdh icd me mpi fh
N/T parasite DNA [176] [193]
Sequencing L. donovani L. chagasi L. infantum L. archibaldi L. hertigi L. deanei L. equatorensis L. major L. aethiopica L. amazonesis L. garnhami L. mexicana L. tropica L. killiki L. aristidesi L. pifanoi L. enrietti L. braziliensis L. guyanensis L. panamensis L. shawi L. turanica L. arabica
cyt b N/T parasite DNA, clinical isolates
[192]
Sequencing L. donovani L. infantum L. tropica L. aethiopica L. braziliensis
mini-exon 50 fg parasite DNA, tissue samples
[184]
Sequencing L. tropica L. major L. donovani L. archibaldi L. infantum L. chagasi L. mexicana L. amazonensis L. garnhami L. braziliensis L. peruviana L. lainsoni L. guyanensis L. panamensis L. naiffi L. tarentolae Trypanosoma cruzi
hsp70 N/T parasite DNA [189]
1458 Current Medicinal Chemistry, 2012 Vol. 19, No. 10 Kobets et al.
(Table 3). Contd…..
Assay Leishmania species
Target Sensitivity (lowest detectable parasite load)
Samples Reference
Sequencing + RFLP L. guyanensis L. braziliensis L. lainsoni L. shawi L. naiffi L. amazonensis L. infantum L. utingensis L. lindenbergi
hsp70 N/T parasite DNA [190]
Polymorphism-specific PCR (PS-PSR)
L. braziliensis L. guyanensis L. panamensis L. mexicana L. amazonensis
polymorphic regions N/T parasite DNA, clinical isolates
[178]
Species-specific PCR + RFLP
L. infantum L. donovani
cpb N/T parasite DNA [195]
Polymorphism-specific PCR (PS-PCR)
L. panamensis L. braziliensis L. guyanensis L. mexicana L. amazonensis
polymorphic regions N/T parasite DNA, formalin-fixed biopsy samples
[214]
ELISA L. infantum Leishmania antigen N/T spleen, liver
[158]
ELISA L. chagasi anti-Leishmania antibodies
N/T cerebrospinal fluid
[215]
Western blot L. infantum anti-Leishmania antibodies
N/T serum [217]
Latex agglutination test (LAT)
Leishmania spp. Leishmania antigen N/T urine [219]
Recombinant K39 test (rK39)
Leishmania spp. anti-Leishmania antibodies
N/T blood [218]
Immuno-detection
Direct agglutination test (DAT)
Leishmania spp. anti-Leishmania antibodies
N/T blood, serum [218]
Delayed-type hyper-sensitivity reaction
Leishmanin skin test (LST)
L. major cell-mediated immune responce
N/T skin [221]
N/T – not tested; kDNA – kinetoplast DNA; 18S rRNA / ssu-rRNA – small ribosomal subunit RNA; ITS1 and ITS2 – internal transcribed spacers 1 snd 2; 7SL RNA – signal recognition particle RNA; gp63 – glycoprotein 63 kDa; hsp70 – heat shock protein 70 kDa; cyt B – cytochrome B; g6pdh - glucose-6-phosphate dehydrogenase; icd - isocitrate dehydrogenase; me - NADP-malic enzyme; mpi - mannose phosphate isomerase; fh - fumarate hydratase; cpb - cysteine protease B; asat - aspartate aminotransferase; gpi - glucose-6-phosphate isomerase; nh1 and nh2 - nucleoside hydrolases 1 and 2; pgd - 6-phosphogluconate dehydrogenase.
3.3.3. OligoC-TesT OligoC-TesT is a simplified and standardized PCR format for
molecular detection of Leishmania. The test is based on PCR amplification of a small sequence of the 18S rRNA gene followed by visualization of the PCR products on a dipstick by hybridization with a gold-conjugated probe. The test gives rapid result and could be useful for diagnosis of leishmaniasis, but not for parasite quantification [183, 208]. 3.3.4. Restriction Fragment Length Polymorphism (RFLP), Reverse Dot Blot (RDB), and Reverse Line Blot (RLB) for Leishmania Species Identification
A spectrum of different species can be found in patients sent to the diagnostic laboratories. Since the therapeutic strategy depends on the Leishmania species involved, it is critically important to type the infecting parasite in a sensitive way. The most commonly used
targets are 18S ribosomal RNA gene, the 39 nucleotides small mini-exon gene, and ITS1 region between the 18S ribosomal RNA and 5.8S ribosomal RNA genes. All these regions are highly repetitive and conservative, though a part of their sequences vary that allows using them for species-specific typing. Mini-exon gene is unique and tandemly repeated in Kinetoplastida, but is absent in mammalian hosts and sand fly vectors. An intron, adjacent to the mini-exon, is conserved. It is followed by a sequence that varies in length considerably between species and is much less conserved. Primers were targeted both to the conserved mini-exon and its intron in order to amplify the polymorphic intergenic region [184, 185, 198, 209-211].
The studies that compared different targets (18S ribosomal RNA gene, mini-exon and ITS1 region) revealed that the highest sensitivity was achieved with the primer for ITS1. Often the ITS1 gave clearer PCR products when compared to the mini-exon results

Leishmaniasis: Prevention, Parasite Detection and Treatment Current Medicinal Chemistry, 2012 Vol. 19, No. 10 1459
[184]. Compared to kinetoplast DNA (kDNA), which is considered to be the most sensitive PCR approach for Leishmania parasite detection, ITS1 and mini-exon PCR methods suffered from reduced sensitivity, especially when tested on clinical material collected on filter papers [197].
7SL RNA was used as a target for detection of Old World Leishmania Species. Species-specific RFLP patterns were obtained when the PCR product was digested with the restriction enzymes. Results were compared with reverse dot blot (RDB) assay, for which biotin-labeled amplicons were hybridized to specific oligonucleotide probes immobilized onto a membrane. It increased the sensitivity by tenfold compared to the detection of PCR products by gel electrophoresis [187].
Biotinylated ITS1-specific PCR primers and reverse line blot (RLB) assay were used for evaluation of 21 reference strains of several Leishmania species. Similarly to RDB, biotinylated PCR product hybridize specifically to oligonucleotide probes coupled to a membrane support. Using appropriate hybridization conditions, it is possible to differentiate between amplicons whose DNA sequences differ by only one nucleotide. The sensitivity of the RLB was comparable to that using kDNA PCR [186].
Though the gene hsp70 is conserved across prokaryotes and eukaryotes, nucleotide sequence variation is sufficient to discriminate parasite species [189]. Analysis of hsp70 gene sequence led to the identification of restriction enzymes that could also be used for PCR-RFLP based identification [190]. 3.3.5. Sequencing-Based Pathogen Classification
A single amino acid polymorphism can be responsible for change in enzyme mobility, but some distinct genotypes produce phenotypes, indistinguishable using MLEE, described above [176]. Though RFLP analysis also proved to be a reliable method for typing of Leishmania species, it was not optimal for L. Viannia subgenus (also known as L. braziliensis complex) detection. In such cases, direct sequencing of the product solved the problem. Reference strains should be used to assign the samples to the various known species [184].
Analysis of genes coding DNA and RNA polymerases has the potential for use in genetic typing and population genetics, their gene sequences were analyzed to reveal the evolution of the genus Leishmania [191]. Multilocus sequence typing, which includes amplification and sequencing of polymorphic regions (Table 3), appeared to be a very informative method for investigation of relationships between zymodemes, and more precise parasite identification and description, enhancing MLEE [176, 193].
Other DNA targets were also used for investigation of the Leishmania phylogeny. Species identification can be based on sequencing of the cytochrome b (cyt b) gene. It is contained in the mitochondrial genome of a wide variety of living forms and encodes the central catalytic subunit on an enzyme present in the respiratory chain of mitochondria. This gene has been widely used for phylogenetic studies and identification of animals and plants [212]. Using cytochrome b gene as a target allowed to explore Leishmania species/subspecies phylogenetic relations, and facilitated detection of 28 Leishmania strains in samples from patients [192, 212] (Table 3).
Besides using for RFLP, mentioned above, the gene encoding 70 kDa heat-shock protein (hsp70 and HSP70 respectively) was sequenced for studying evolutionary relationships of 16 Leishmania species from the Old and New World. This target has also been applied in phylogenetic studies of different parasites [189] (Table 3). 3.3.6. Polymorphism Specific PCR (PS-PCR)
On the basis of knowledge of the cyt b gene sequence, a polymorphism-specific PCR procedure was developed. Results
were visualized using agarose gel [179]. Discriminative PCR targeting the cbp gene (Table 2) was first used to confirm the taxonomic status of the specimens if they belong to L. donovani complex (L. infantum and L. donovani) [213]. However, for more precise detection, the product should be processed with the restriction enzymes for species specific sites [195].
The combination of PS-PCR with two subgenus-specific PCR allowed detection of major Leishmania species causing New World cutaneous leishmaniasis in formalin fixed biopsy samples. The samples are easily transported without freezing, what makes the technique beneficial for diagnostics [214].
3.4. Tests Based on Immune Response to Leishmaniasis
3.4.1. ELISA Enzyme-linked immunosorbent assay (ELISA) is based on
indirect detection of parasites in the infected host by the presence of Leishmania antigens [158] or antibodies to them [215], using anti-Leishmania antibodies and total antigen from lyzed promastigotes, respectively. Immunological cross-reaction of Leishmania with Trypanosoma cruzi, which is co-endemic in some areas, has posed a serious problem in the use of serodiagnostic tests. However, specific monoclonal antibodies against Leishmania could solve this problem [216]. One of the important advances in the indirect diagnosis of leishmaniasis was the production and identification of recombinant antigens. Specifically, the antigen rK39, a recombinant protein, resembles a part of a 230-kDa protein of the kinesin family, coded by the kinetoplast DNA of Leishmania. Reactivity with the other trypanosomatids is negative and the presence of the antibody, anti-rK39, indicates an active infection. Comparison between immunochromatographic rK39 test and ELISA exhibited no significant differences [204], nevertheless Western blot test was found to be more sensitive and suitable for the serodiagnosis of anthroponotic cutaneous leishmaniasis than conventional ELISA [217].
Estimation of the presence of Leishmania antigens and anti-Leishmania antibody levels need not reflect the real parasite load because the presence of antibodies to parasites highly depends on the individual responsiveness and general health condition of the host. Due to this fact, the diagnosis, obtained with immunodetection-based assays, should be confirmed by other methods. 3.4.2. Rapid Tests
A separate group of assays create rapid noninvasive diagnostic methods, which detect the presence of antibodies to Leishmania in the blood, serum or urine. Recombinant K39 dipstick was successfully used in the field. One-two drops of blood followed by one-two drops of buffer were placed on the pad of the dipstick and incubated for approximately 5 minutes. The test was positive when two bands, a control band and a test band, appeared within 5 minutes [218]. The direct agglutination test (DAT) can be performed either on blood or serum, measuring the titer of the immune complexes. The threshold titer chosen to define a positive test is crucial for proper analysis [218]. Latex agglutination test (LAT) is based on the detection of a leishmanial antigen in urine. In patients with visceral leishmaniasis, it was proved to be useful in the monitoring of the treatment [219]. 3.4.3. Leishmanin Skin Test (LST)
Leishmanin test was introduced in 1926 by Montenegro [220]. It includes intradermal administration of Leishmania antigen followed by the measurement of induration. It is used as an indicator of cell-mediated immunity against Leishmania parasites in vivo. Leishmanin inoculation into the skin of an individual in whom previous infection has led to a state of cell-mediated immunity will result in a delayed-type hypersensitivity reaction characterized by

1460 Current Medicinal Chemistry, 2012 Vol. 19, No. 10 Kobets et al.
erythema and induration at the site of antigen injection after a few hours. LST based on L. major antigens was used as a marker of previous exposure to L. major parasites in clinical and epidemiological studies [221], but showed lack of sensitivity in patients cured from visceral leishmaniasis [222]. Moreover, LST has significant limitations in case of visceral leishmaniasis, because patients with active disease are in a state of anergy and unresponsive to the test [222, 223].
3.5. Summary of Leishmania Detection and Quantification Assays
Taking into consideration the wide range of methods that were developed during a relatively long period of time (Table 3), the selection of the most suitable technique sometimes may become complicated.
Microscopy detection of parasites in Giemsa or hematoxylin and eosin (H&E) stained samples are considered to be the classical standard from the beginning of Leishmania studies. Though these methods are laborious, time consuming and lack sensitivity, they are highly specific, visualize the parasite directly, and remain a perfect tool for illustration of the result.
Until recently, the limiting dilution assay was commonly used for parasite quantification. This method has similar limitations as microscopy, being laborious and time consuming; moreover, all culture-based techniques are highly dependent on sterile conditions. It can take days or weeks until parasites are observed, depending on the species and number of parasites seeded. Moreover, the cultures may become contaminated. These characteristics nearly exclude the possibility of it being used for routine examinations of biopsy specimens collected under field conditions from patients [194, 197]. Nevertheless, culturing of parasites can reflect their viability and ability to multiply.
At present, the DNA-based methods create the most popular and intensively developing group of assays. They proved to be reliable, time-saving, and most suitable for the purposes of diagnostics and research, especially for analysis of large numbers of samples. These significant advantages over the other methods make them a powerful tool for Leishmania parasites detection, quantification and identification of species and subspecies.
Independently of the target, real-time PCR is the preferred method for many diagnostic laboratories because it is fast and provides the opportunity to perform a multiplex PCR. However, inhibition of the PCR can sometimes be seen, particularly with skin biopsies, and leads to a lower rise of the curve during the reaction [184]. Due to the use of fluorescent probes, the price of RT-PCR is usually a bit higher than for other PCR-based assays. Moreover, modern specialized RT-PCR machines and software required for this method are still not affordable for many clinical and scientific laboratories in developing countries.
In this aspect, PCR-ELISA is cheaper than RT-PCR and it is based on robust approaches, routinely used for a relatively long time. Common PCR machines and ELISA readers are available in laboratories all over the world, because these techniques have been widely used for research and clinical purposes. Sensitivity of PCR-ELISA is comparable to that of RT-PCR. These advantages make PCR-ELISA a preferred method in many cases.
In a number of cases, precise identification of the Leishmania species is necessary for clinicians to make correct decisions on appropriate therapy. Such information is also valuable for epidemiological studies. For a long time, the main assay for Leishmania species identification was MLEE. However, this method has some significant disadvantages. Besides the risk of a bacterial contamination during cultivation, the parasite strains with the same enzyme phenotype may in fact have distinct amino acid sequences [176, 224]. Cytochrome b gene sequencing proved to be
an effective and useful method for phylogenetic analysis of Leishmania genus [192, 212, 224]. Nevertheless, similarly to MLEE, it needs cultivation of parasites that implicates the same drawbacks as all cell culture based methods, described above.
Problems with detection of Leishmania species with MLEE can be resolved by PCR assays using primers against polymorphic regions. Combination of these techniques with RFLP or RDB/RLB significantly increases their sensitivity, helping to escape false negative diagnosis. Digestion of the PCR product with restriction enzymes allows identification of almost all pathogenic Leishmania species, thus enabling direct rapid characterization of the infecting parasite [186, 187]. Results from polymorphism specific PCR have agreed both with cyt b gene sequencing and MLEE. Therefore, all these techniques could be used to identify Leishmania species [179].
Noninvasive rapid tests (LAT, DAT, rK39 dipstick test) deserve special attention, because they are commercially available, easily applicable and give fast results. In the comparative study, rK39 dipstick test was less sensitive than DAT, but was easier to use and results could be obtained within minutes [218]. In other studies, these tests showed a comparable sensitivity and were more reliable than LAT [225]. Noninvasive rapid tests share the main disadvantage of all immunodetection assays: their results are highly dependent on the individual immune response of the organism, what may cause discrepancies in diagnostics and the presence of false negative samples. Despite of it, such tests could be especially valuable in the field conditions, for mass-surveys, as well as for confirmation of clinically suspected cases.
In summary, all the existing methods can be divided into two large groups: assays for Leishmania parasite detection and techniques suitable for both detection and quantification of the pathogen. Most of the methods indicate the presence of the pathogen in the sample without precise measurement of the level of infection. In addition, for many techniques which allow quantification of parasites, the sensitivity is either not tested or it is difficult to establish (Table 3, Sections microscopy, culture-based methods, and immunodetection). This fact restricts their use because samples with low parasite load may be errorneously classified as negative. Thus, use of these techniques alone could lead to fatal errors, especially in the clinical diagnostics of the early stages of the disease. At present, PCR-based assays (RT-PCR, PCR-ELISA) are the most suitable approaches for Leishmania parasite quantification (Table 3, Section DNA-based methods). These methods use fluorescent probes or color reaction to measure concentration of the parasite DNA according to a standard curve. Sensitivity of these techniques is very high – on the order of femtograms, so they can be successfully applied both for research and clinical purposes.
All advantages and disadvantages of the methods, described above, will determine their application both for experiments and clinical practice. Finally, it is worth to mention again that all novel methods always need standardization and quality control. For this purpose, results obtained using the newly developed techniques must be compared with results from the classical ones (microscopy, kDNA PCR, MLEE) to test the efficacy of the fresh approaches.
In conclusion, there is no one universal method that could be used in every case when the parasite load is necessary to be measured. Due to their advantages and disadvantages, all approaches have different purposes of use. PCR-based techniques are the most sensitive at the present moment, so we highly concentrated our attention on them. PCR-ELISA is robust, relatively simple and cheaper method in comparison with other quantitative assays. In addition, combination of these methods with non-PCR-based techniques, will enable an integrated view on development of leishmaniasis.
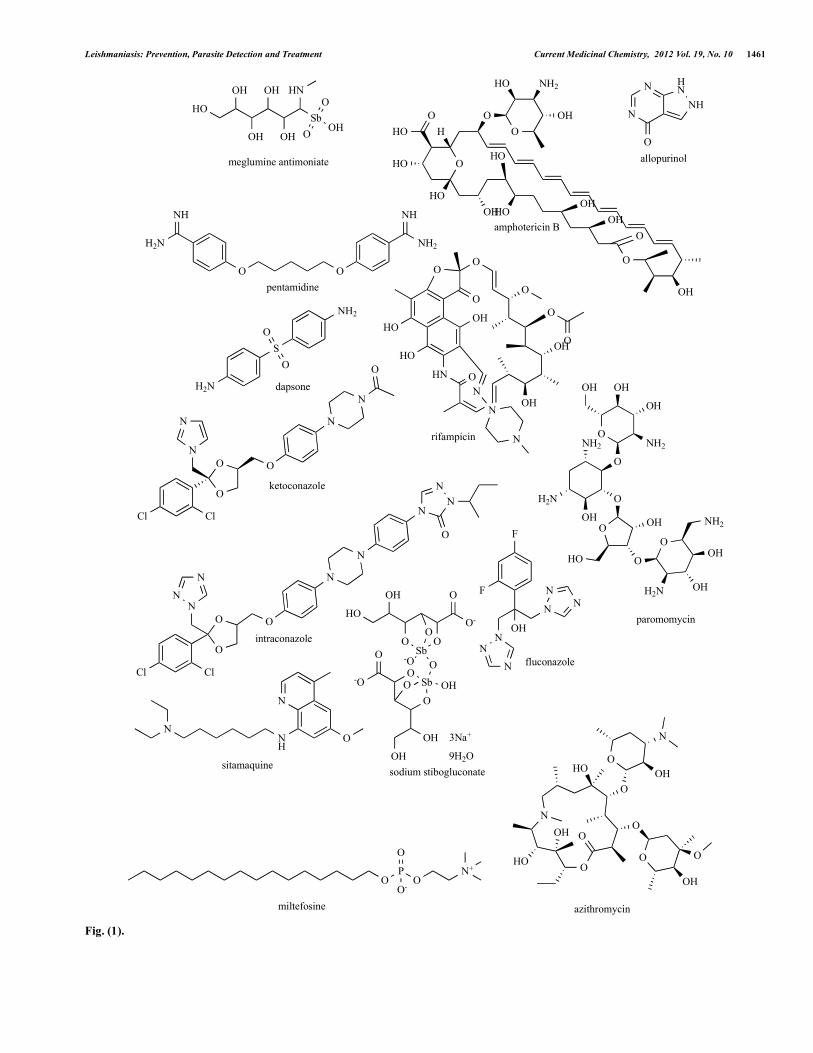
Leishmaniasis: Prevention, Parasite Detection and Treatment Current Medicinal Chemistry, 2012 Vol. 19, No. 10 1461
Sb
O
OOH
HO
OH
OH
OH
OH
HN
3Na+
9H2OOH
OH
O
Sb OH
O
Sb-O
O
OH
HO
OO
O-
O
OO
-O
O
meglumine antimoniate
sodium stibogluconate
amphotericin B
NH
NH2
OO
H2N
NH
N
NNH
HN
O
H2N
S
O
O
NH2
pentamidine
dapsone
allopurinol
N
NN
O
N
N
O
O
O
NN
N
Cl Cl
N
N
O
O
O
N
N
Cl Cl
O
ketoconazole
intraconazole
P
O
OOO-
N+
miltefosine
O
O
O
HN
O
O
O
N
N
N
O
HO
OH
OH
OH
HO
rifampicin
paromomycinOH
NN
N
N
N
N
F
F
fluconazole
O
N
OO
O O
OH
HO
HO
OH
O
O
OH
N
azithromycin
NH
N
N
O
sitamaquine
OOH
OH
O
O
HO
OH
O
OH
O
O
OH OH
OH
NH2
H2N
NH2
NH2
H2N
O
O
O
HO
HO
OH
HO
HOOH
OH
OH
O
O
OH
HO NH2
O
HO H
Fig. (1).

1462 Current Medicinal Chemistry, 2012 Vol. 19, No. 10 Kobets et al.
4. TREATMENT OF LEISHMANIASIS
Similarly to vaccination, the spectrum and efficacy of available antileishmanial drugs is also limited. Drugs that were developed to treat leishmaniasis can be divided into several groups. The first-line drugs used to treat both visceral and cutaneous leishmaniasis are pentavalent antimonials (sodium stibogluconate and meglumine antimoniate), amphothericin B and pentamidine [226-228] (Fig. 1). Amphotericin B was first introduced in early 1960s. The drug induces the formation of aqueous pores in the parasite membrane [229, 230] and has an excellent leishmanicidal activity [231]. Pentamidine is used in the treatment of cutaneous and mucocutaneous leishmaniasis in the New World [226]. A range of alternative drugs including miltefosine, paromomycin, azoles, azithromycin, allopurinol, dapsone and rifampicin have been recently introduced. However, the efficacy of the substances differs drastically depending on the clinical form of leishmaniasis, Leishmania species and geographical region [226, 232-235]. Thus, there is still a continuing need for new chemical substances with a leishmanicidal effect and for optimized delivery systems.
4.1. First-Line Drugs
4.1.1. Pentavalent Antimonials Pentavalent antimonials (SbV) have been the first-line drug for
more than 70 years. The drug is provided in two formulations: meglumine antimoniate and sodium stibogluconate. However, pentavalent forms are prodrugs [236], which are converted by gluthatione system to biologically active trivalent forms (SbIII). The latter interfere with glycolysis [237-239], fatty acid oxidative pathways [227], trypanothione balance [240-242], and type I DNA topoisomerase [243]. Pentavalent antimonials are used in the treatment of all clinical forms of leishmaniasis both in the Old and in the New World; their efficacy, however, significantly differs according to the clinical form of leishmaniasis, species of Leishmania and geografic region. In the Old World, pentavalent antimonials remain the drug of choice for the treatment of cutaneous leishmaniasis, being highly efficient against its main causative agents – L. major and L. tropica [244-246]. In the New World, treatment efficacy is about 93% in Bolivia [247], 84% in Brazil [248] and 61% in Colombia [249]. Besides, pentavalent antimonials are used for treatment of visceral leishmaniasis in most endemic areas apart from India. The cure rates are around 90-95% [250]. In India, the pentavalent antimonials were virtually abandoned due to the emergence of unresponsive L. donovani and a concomitant decrease in cure rates to 50% [251]. In addition, drug-resistant parasites were demonstrated to acquire increased in vivo survival ability [252]. Initially, the resistance of L. donovani to pentavalent antimonials was shown to be connected with high expression of γ-glutamylcysteine synthetase and high levels of parasite glutathione [253]. Recent evidence, however, implied multiple origins of drug resistance in L. donovani lines isolated from patients with visceral leishmaniasis [254]. It was proposed that the factors contributing to the occurance of L. donovani resistance to pentavalent antimonials could be the misuse of the medications [250] and a chronical exposure to arsenic in drinking water [255]. A serious disadvantage of pentavalent antimonials is their dose-limiting adverse effects. They include arthralgy, myalgy, cardiotoxicity, nephrotoxicity, hepatitis, pancreatitis, nausea and vomiting [226, 256]. Encapsulation of antimonial drugs and reduction of the dose required for effective therapy should minimize systemic toxicities and improve treatment [257]. 4.1.2. Amphotericin B
Amphotericin B was first introduced in early 1960s. It is an antifungal macrolide antibiotic isolated from Streptomyces nodosus. Antileishmanial activity of Amphotericin B is attributed to its selective affinity for ergosterol, a constitutive of Leishmania plasma
membrane, but not cholesterol, its primary sterol counterpart in mammalian cells. The drug induces the formation of aqueous pores in the parasite membrane and has an excellent leishmanicidal activity [229, 231]. The disadvantages of Amphotericin B are low water solubility, poor bioavalability, renal toxicity [230], other occasional serious toxicities [229, 231], the need of administration by slow infusion [258] and infusion based reactions like thrombophlebitis and high fever with rigor and chills [229, 231]. Toxic effects of amphotericin B deoxycholate have been largely ameliorated with the advent of lipid formulations of this drug. In these formulations, deoxycholate has been replaced by other lipids that mask amphotericin B from susceptible tissues and facilitate its preferential uptake by reticuloendothelial cells, thus achieving a targeted drug delivery to the parasite resulting in increased efficacy and reduced toxicity. Three such lipid-associated formulations of amphotericin B are commercially available: liposomal amphotericin B (L-AMB); amphotericin B lipid complex (ABLC); and amphotericin B colloidal dispersion (ABCD) [259] (Table 4). Phase II clinical trial of visceral leishmaniasis treatment with L-AMB was successfully performed in India, Kenya and Brazil (Table 5). It is interesting that the total dose of 14 mg/kg was able to cure 100% patients in India and Kenya, but only 62% patients in Brazil. With an even more striking difference, 6 mg/kg was able to cure 100% patients in India but only 20% patients in Kenya [232]. The differences in the efficacy of the treatment could be attributed either to different species of Leishmania or to a different genetics of the populations in different endemic areas. In a later study, because of its excellent safety profile, a large dose (7.5 mg/kg) of L-AMB was given to each of 203 patients with visceral leishmaniasis at 4 treatment centers, and the patients were discharged the next day: 183 patients (90%) were cured with no relapse during 6 months after treatment [260]. The effectiveness of L-AMB was worse in the cases of coinfection of HIV and visceral leishmaniasis. In Ethiopia, the total dose of 30 mg/kg in 6 doses on alternate days was able to cure only 74% of 116 patients with primary visceral leishmaniasis and only 38% of 79 patients with relapse [261]. Still, L-AMB remains the best option for the treatment of HIV and visceral leishmaniasis coinfection, which is also reflected in WHO recommendations [262].
ABLC was also highly efficient in treatment of Indian visceral leishmaniasis. Five infusions (3 mg/kg each) given to 21 patient every second day over 9 days (total dose, 15 mg/kg) resulted in a 100% apparent cure response. In 4 other patients, an apparent cure was induced by ABLC given at 3 mg/kg a day for 5 consecutive days (total dose, 15 mg/kg) [263]. Treatment of visceral leishmaniasis with ABCD was studied in India. Different regimens were studied in three groups, each including 135 patients (Table 6) [231].
Generally, the usage of lipid formulations of AmphotericinB is limited due to the high price [229, 231, 256]. In India, the cost of a single 5 mg/kg dose of AmBisome® for a 30 kg patient was about US$ 600, compared with US$ 60 for a typical treatment regimen with conventional amphotericin B [229, 231]. Although, recently, WHO concept of tiered pricing had allowed to reduce the cost of AmBisome® to US$ 252 for a typical treatment regimen in low-income countries [264]. 4.1.3. Pentamidine
Pentamidine was used to treat visceral leishmaniasis in India until it was totally abandoned due to declining efficacy and severe side effects [229] – nephrotoxicity, cardiotoxicity, diabetes mellitus-like state [230] and hypoglycemia [226]. Pentamidine is still reported to be used in the treatment of cutaneous and mucocutaneous leishmaniasis in the New World [226]. A number of clinical trials were carried out in Brazil, Colombia, Peru, Surinam and French Guyana before 2005 [265]. However, after the recommendation of WHO to use pentamidine “only when no other
Leishmaniasis: Prevention, Parasite Detection and Treatment Current Medicinal Chemistry, 2012 Vol. 19, No. 10 1463
options are available” in 2007 [262], to our knowledge, no futher clinical trials involving pentamidine were performed. Thus, the update of the information about pentamidine usage would demand a separate investigation.
4.2. Alternative Drugs
A range of alternative drugs including miltefosine, paromomycin, azoles, azithromycin, allopurinol, dapsone and rifampicin have been recently introduced [226]. From these drugs, miltefosine and paromomycin get a special attention. 4.2.1. Miltefosine
The alkylphospholipid derivative miltefosine (hexadecylphos-phocholine) is the first orally effective antileishmanial agent. In L. donovani promastigotes, miltefosine causes apoptosis-like death [266]. It was established that miltefosine 1) specifically inhibits cytochrome c oxidase, which results in significant decline in the intracellular ATP levels, reduction in the oxygen consumption rate and mitochondrial depolarization [267]; 2) inhibits alkyl-specific acyl coenzyme A acyltransferase, essential for biosynthesis of alkyl-anchored glycolipids (e.g. lipophosphoglycan) and glycoproteins (e.g. major surface glycoprotein GP63) [268]; 3) reduces the phosphatidylcholine content and enhances the phosphatidylethanolamine (PE) content in parasite membranes, suggesting a partial inactivation of PE-N-methyltransferase; 4) enhances lysophosphatidylcholine content probably due to phospholipase A2 activation; 5) strongly reduces the C24 alkylated
sterol content; 6) enhances the cholesterol content probably due to the condensation with sterols [269]. Additional antileishmanial mechanisms of action of miltefosine can be extrapolated from its effect on mitogen-stimulated fibroblasts where it inhibits phospholipase C and thus reduces both inositol 1,4,5-trisphosphate formation and Ca2+ release and diacylglycerol formation, proteinkinase C activation and proliferation [270, 271]. Furthermore, miltefosine was shown to inhibit sphingomyelin-synthase and increase the pool of its substrate, ceramide, which serves as a proapototic signal molecule [271]. Several multicenter studies in India demonstrated a high cure rate (around 94%) for miltefosine treatment of visceral leishmaniasis. Its efficacy has also been reported in SbV resistant cases [272]. The adverse effects of miltefosine are mild to moderate gastrointestinal disturbances that included vomiting and diarrhea, and renal toxicity. As miltefosine is teratogenic, it is contraindicated in pregnancy and women of child bearing age group unless contraception can be provided [229]. 4.2.2. Paromomycin
Paromomycin, obtained from cultures of Streptomyces rimosus, belongs to the class of aminoglycosides and possesses both anti-bacterial and antiprotozoal activity. Although developed in the 1960s as an anti-leishmanial agent, it remained neglected until the 1980s when topical formulations were found to be effective in cutaneous leishmaniasis and a parenteral formulation for visceral leishmaniasis was also developed [229]. It is hypothesized that paromomycin binds to a negatively charged leishmanial glycocalyx as well as to paraflagellar rod proteins and prohibitin, being
Table 4. Lipid Formulations of Amphotericin B [259]
Compound Lipid configuration Size (nm) Lipids
amphotericin B not applicable not applicable Deoxycholate
L-AMB liposome 80 Hydrogenated soy phosphatidylcholine, distearoyl phosphatidylglycerol
ABCD disklike 120-140 Cholesteryl sulfate
ABLC ribbonlike 1600-11000 Dimyristoyl phosphatidylcholine, dimyristoyl phosphatidylglycerol
Table 5. Efficacy of L-AMB Regimens in Different Endemic Areas [232]
Country Brazil Kenya India
Regimen 2 MKD* on days 1-6 and day 10 (total dose:
14mg/kg)
2 MKD on days 1-4 and day 10 (total
dose, 10mg/kg)
2 MKD on days 1-10
(total dose, 20mg/kg)
2 MKD on days 1-6 and day 10 (total
dose: 14mg/kg)
2 MKD on days 1-4 and day 10 (total
dose, 10mg/kg)
2 MKD on days 1, 5,
and 10 (total dose,
6mg/kg)
2 MKD on days 1-6 and day 10 (total
dose: 14mg/kg)
2 MKD on days 1-4 and day 10 (total
dose, 10mg/kg)
2 MKD on days 1, 5,
and 10 (total dose,
6mg/kg)
Cure 8 4 13 10 9 1 10 10 10
Failure 1 0 0 0 0 3 0 0 0
Relapse 4 0 2 0 1 1 0 0 0
% Cure 62 100 87 100 90 20 100 100 100
*MKD – mg/kg/day. Table 6. Efficacy of ABCD regimens [231]
Regimen 0.5 mg/kg on day 1, 1 mg/kg on day 2 and 1.5 mg/kg on days 3–6 (total dose 7.5 mg/kg)
0.5 mg/kg on day 1, 1.5 mg/kg on day 2 and 2 mg/kg on days 3–6 (total dose 10 mg/kg)
1 mg/kg on day 1, 2 mg/kg on day 2 and 3 mg/kg on days 3–6 (total dose 15 mg/kg)
% Cure 97% 96% 97%
Adverse effects
Chills, fever, vomiting (5.92%) Chills, fever, vomiting (8.89%); 1.48% were withdrawn
Chills, fever, vomiting (11.11%), backache (5.92%); 1.48% were withdrawn

1464 Current Medicinal Chemistry, 2012 Vol. 19, No. 10 Kobets et al.
subsequently internalized by endocytosis [273]. Inside the cell, paromomycin induces a drop of mitochondrial dehydrogenases activities, seemingly due to a shortage of respiratory substrates and the decrease of mitochondrial potential [274], promotes the association of the ribosome subunits [275], inhibits RNA and protein synthesis, affects the uptake and incorporation of macromolecular precursors and increases the amount of membranous polar lipids, leading to decreased membrane fluidity [276]. Phase 3 trial by Sundar et al. in India in 2007 showed a high six-month cure rate (94.6%) for parenteral paromomycin (dose-11 mg/kg for 21 days) which subsequently led to licensing for its use in India [277]. With different success, paromomycin in its topical formulation is used to treat cutaneous leishmaniasis in a number of countries. In the Old World, the cure rates for L. major infection ranged from 74.2% in Israel [278] to 16.6% in Iran [279]. The efficacy of topical paromomycin in treatment of L. tropica infection in Turkey was 37.5% [280]. In the New World, the cure rates ranged from 85.7% for L. braziliensis and L. mexicana in Guatemala [281] to complete clinical failure for L. mexicana and L. chagasi infection in Honduras [282]. 4.2.3. Allopurinol
Allopurinol, another alternative drug, was synthesized in the mid-1950s and later tested for leishmanicidal activity. In mammals, the compound inhibits xanthine oxidase [283]; however, Leishmania do not have this enzyme [284]. Instead, Leishmania metabolize allopurinol to an analog of adenosine nucleotides, which is incorporated into RNA. When this occurs, protein synthesis is halted, resulting in allopurinol's leishmanistatic effect [285]. Nevertheless, when used alone for treatment of cutaneous leishmaniasis caused by L. panamensis and L. braziliensis [286], allopurinol had only 33% cure rate. 4.2.4. Itraconazole, Ketokonazole and Fluconazole
Itraconazole, ketokonazole and fluconazole were initially designed for therapy of fungal infections, and are also used to treat leishmaniasis. These drugs inhibit 14-α-demethylation of lanosterol to ergosterol during cell wall synthesis and promote membrane permeability of Leishmania [226]. The success of azoles in leishmaniasis therapy varied from failure to complete release of the disease. It was reported that cutaneous lesions caused by Leishmania spp. were successfully treated with itraconazole after unresponsiveness to pentavalent antimonials [287], whereas in the case with defined L. panamensis infection such therapy was not effective [288]. High doses of fluconazole also showed promising result against L. major induced cutaneous leishmaniasis in human [289]. 4.2.5. Azithromycin
Azithromycin, a broad spectrum antibiotic used for treatment of various bacterial infections, showed certain effect against L. amazonensis, L. braziliensis and L. chagasi in vitro [290]; however the mechanism of its anti-parasitic action remains unclear [226]. As many other drugs, azithromycin was reported to have poor results (45.5% cure rate) during treatment cutaneous leishmaniasis caused by L. braziliensis [291]. 4.2.6. Sitamaquine
Sitamaquine (WR6026) is an orally administered drug tested for the treatment of visceral leishmaniasis. It causes dose-dependent inhibition of respiratory chain complex II (succinate dehydro-genase) of the respiratory chain, promotes oxidative stress and apoptosis-like death of Leishmania parasites [292]. During clinical trials, cure rates among patients with visceral L. chagasi [233] and L. donovani [234, 235] infection varied greatly from very low in Brazil to 80-90% in India and Kenya; the drug had nephrotoxic effects [233-235].
4.2.7. Rifampicin Rifampicin is a semisynthetic antibiotic derived form rifamycin,
discovered in 1957 [293]. Rifampicin blocks RNA synthesis by specially binding to and inhibiting the DNA-dependent RNA polymerase. Four weeks of daily oral administration of rifampicin (1200 mg/day) resulted in the cure of 17 of 23 (73.9%) patients with L. tropica-induced cutaneous leishmaniasis [294]. 4.2.8. Dapsone
Dapsone, a well-known anti-infectious agent, was synthesized in 1908 by Fromm and Wittmann. Dapsone inhibits the synthesis of folic acid due to structural similarity with p-aminobenzoic acid [295]. The proposed mechanism of action of dapsone as an antileishmanial agent is probably by inhibition of choline incorporation into lecithin of the cell membrane, which leads to decreasing phospholipid synthesis. Orally administrated dapsone was effective in 49 out of 60 (82%) patients with L. tropica-induced cutaneous leishmaniasis [296]. A small trial (11 patients) conducted in the New World showed a lower efficacy of the compound [297]. Finally, dapsone may cause serious adverse effects: hemolytic anemia, methemoglobinemia, hepatic involvement (hepatocellular or cholestatic disease, or both), cutaneous involvement (exanthematous eruption, Stevens-Johnson syndrome or toxic epidermal necrolysis); agranulocytosis, nephritis, pneumonitis and hypothyroidism [298].
4.3. Combinatorial Treatment
The use of combinations of different drugs may bring significant advantages and better therapeutical effects than each of the substances alone. Combinations of miltefosine and paromomycin with each other and with other antileishmanial drugs (pentavalent antimonials, amphotericin B) showed superior efficacy in treatment of visceral leishmaniasis [229, 231, 299, 300]. Pentavalent antimonials are also widely combined with other drugs. Intralesional sodium stibogluconate in combination with oral administration of ketokonazole led to the cure of 92.3% of lesions in human patients [301]. Treatment of L. braziliensis-induced mucocutaneous leishmaniasis with pentavalent antimonials in low doses combined with allopurinol showed no benefits in cure rate (63.6% vs 75% for pentavalent antimonials alone) and the rates of adverse effects [302].
4.4. Physiotherapy
Photodynamic and thermotherapy (heat therapy) were developed for local treatment of cutaneous lesions caused by Leishmania parasite; however these methods cannot affect disseminated forms of the disease.
A photodynamic effect occurs when molecules of a photosensitizer localized in the target tissue absorb light of the specific wavelength and transfer energy to biological acceptors, generating an excess of reactive oxygen species that are able to force cells into death pathways. Redox equilibrium is related to the balance between survival and death during the host-parasite interaction. Photoactivation is used to cause redox imbalance in the cell. Photosensitisers that may be used as antimicrobial agents belong to the porphyrin, phthalocyanine (Pc4), azine (methylene blue), triarylmethane (crystal violet), or cyanine families [303]. 5-aminolevulinic acid can be metabolized to the photosensitizer protoporphyrin IX and was shown to be effective against L. major with cure rates above 90% [304]; however the exact mechanism of the antiparasitic effect is not clear. It was hypothesized that 5-aminolaevulinic acid may contribute to the parasite killing indirectly via killing of macrophages [305].
Thermotherapy is based on the local application of heat using hot water soaks, circulating hot water in heating pads, alternating
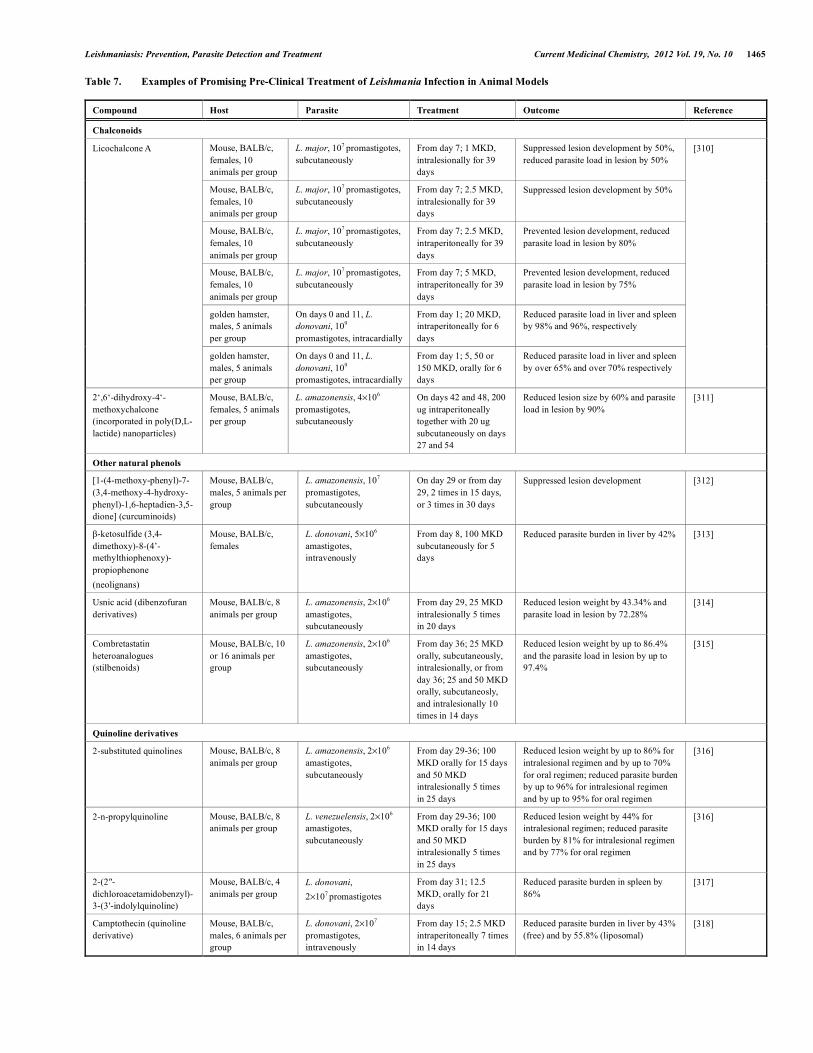
Leishmaniasis: Prevention, Parasite Detection and Treatment Current Medicinal Chemistry, 2012 Vol. 19, No. 10 1465
Table 7. Examples of Promising Pre-Clinical Treatment of Leishmania Infection in Animal Models
Compound Host Parasite Treatment Outcome Reference
Chalconoids
Mouse, BALB/c, females, 10 animals per group
L. major, 107 promastigotes, subcutaneously
From day 7; 1 MKD, intralesionally for 39 days
Suppressed lesion development by 50%, reduced parasite load in lesion by 50%
Mouse, BALB/c, females, 10 animals per group
L. major, 107 promastigotes, subcutaneously
From day 7; 2.5 MKD, intralesionally for 39 days
Suppressed lesion development by 50%
Mouse, BALB/c, females, 10 animals per group
L. major, 107 promastigotes, subcutaneously
From day 7; 2.5 MKD, intraperitoneally for 39 days
Prevented lesion development, reduced parasite load in lesion by 80%
Mouse, BALB/c, females, 10 animals per group
L. major, 107 promastigotes, subcutaneously
From day 7; 5 MKD, intraperitoneally for 39 days
Prevented lesion development, reduced parasite load in lesion by 75%
golden hamster, males, 5 animals per group
On days 0 and 11, L. donovani, 109
promastigotes, intracardially
From day 1; 20 MKD, intraperitoneally for 6 days
Reduced parasite load in liver and spleen by 98% and 96%, respectively
Licochalcone A
golden hamster, males, 5 animals per group
On days 0 and 11, L. donovani, 109
promastigotes, intracardially
From day 1; 5, 50 or 150 MKD, orally for 6 days
Reduced parasite load in liver and spleen by over 65% and over 70% respectively
[310]
2‘,6‘-dihydroxy-4‘-methoxychalcone (incorporated in poly(D,L-lactide) nanoparticles)
Mouse, BALB/c, females, 5 animals per group
L. amazonensis, 4×106 promastigotes, subcutaneously
On days 42 and 48, 200 ug intraperitoneally together with 20 ug subcutaneously on days 27 and 54
Reduced lesion size by 60% and parasite load in lesion by 90%
[311]
Other natural phenols
[1-(4-methoxy-phenyl)-7-(3,4-methoxy-4-hydroxy-phenyl)-1,6-heptadien-3,5-dione] (curcuminoids)
Mouse, BALB/c, males, 5 animals per group
L. amazonensis, 107 promastigotes, subcutaneously
On day 29 or from day 29, 2 times in 15 days, or 3 times in 30 days
Suppressed lesion development [312]
β-ketosulfide (3,4-dimethoxy)-8-(4’-methylthiophenoxy)-propiophenone (neolignans)
Mouse, BALB/c, females
L. donovani, 5×106 amastigotes, intravenously
From day 8, 100 MKD subcutaneously for 5 days
Reduced parasite burden in liver by 42% [313]
Usnic acid (dibenzofuran derivatives)
Mouse, BALB/c, 8 animals per group
L. amazonensis, 2×106 amastigotes, subcutaneously
From day 29, 25 MKD intralesionally 5 times in 20 days
Reduced lesion weight by 43.34% and parasite load in lesion by 72.28%
[314]
Combretastatin heteroanalogues (stilbenoids)
Mouse, BALB/c, 10 or 16 animals per group
L. amazonensis, 2×106
amastigotes, subcutaneously
From day 36; 25 MKD orally, subcutaneously, intralesionally, or from day 36; 25 and 50 MKD orally, subcutaneosly, and intralesionally 10 times in 14 days
Reduced lesion weight by up to 86.4% and the parasite load in lesion by up to 97.4%
[315]
Quinoline derivatives
2-substituted quinolines Mouse, BALB/c, 8 animals per group
L. amazonensis, 2×106 amastigotes, subcutaneously
From day 29-36; 100 MKD orally for 15 days and 50 MKD intralesionally 5 times in 25 days
Reduced lesion weight by up to 86% for intralesional regimen and by up to 70% for oral regimen; reduced parasite burden by up to 96% for intralesional regimen and by up to 95% for oral regimen
[316]
2-n-propylquinoline Mouse, BALB/c, 8 animals per group
L. venezuelensis, 2×106 amastigotes, subcutaneously
From day 29-36; 100 MKD orally for 15 days and 50 MKD intralesionally 5 times in 25 days
Reduced lesion weight by 44% for intralesional regimen; reduced parasite burden by 81% for intralesional regimen and by 77% for oral regimen
[316]
2-(2″-dichloroacetamidobenzyl)-3-(3′-indolylquinoline)
Mouse, BALB/c, 4 animals per group
L. donovani, 2×107 promastigotes
From day 31; 12.5 MKD, orally for 21 days
Reduced parasite burden in spleen by 86%
[317]
Camptothecin (quinoline derivative)
Mouse, BALB/c, males, 6 animals per group
L. donovani, 2×107
promastigotes, intravenously
From day 15; 2.5 MKD intraperitoneally 7 times in 14 days
Reduced parasite burden in liver by 43% (free) and by 55.8% (liposomal)
[318]
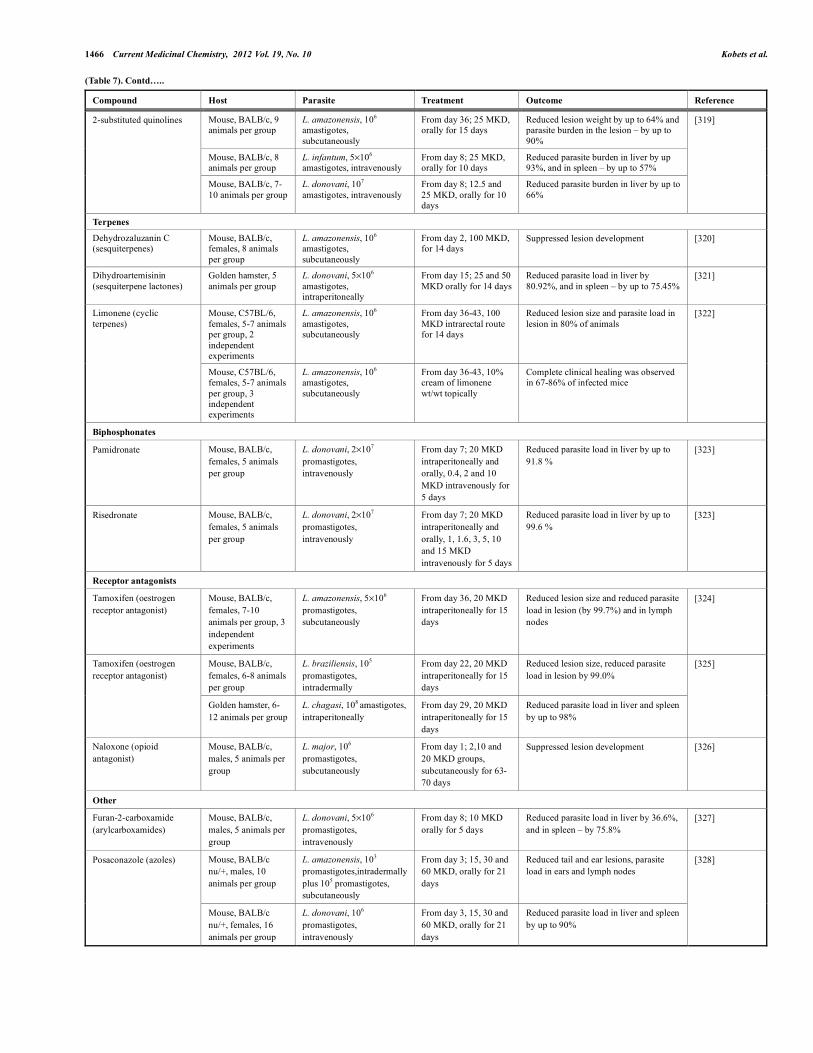
1466 Current Medicinal Chemistry, 2012 Vol. 19, No. 10 Kobets et al.
(Table 7). Contd…..
Compound Host Parasite Treatment Outcome Reference
Mouse, BALB/c, 9 animals per group
L. amazonensis, 106
amastigotes, subcutaneously
From day 36; 25 MKD, orally for 15 days
Reduced lesion weight by up to 64% and parasite burden in the lesion – by up to 90%
Mouse, BALB/c, 8 animals per group
L. infantum, 5×106
amastigotes, intravenously From day 8; 25 MKD, orally for 10 days
Reduced parasite burden in liver by up 93%, and in spleen – by up to 57%
2-substituted quinolines
Mouse, BALB/c, 7-10 animals per group
L. donovani, 107 amastigotes, intravenously
From day 8; 12.5 and 25 MKD, orally for 10 days
Reduced parasite burden in liver by up to 66%
[319]
Terpenes
Dehydrozaluzanin C (sesquiterpenes)
Mouse, BALB/c, females, 8 animals per group
L. amazonensis, 106 amastigotes, subcutaneously
From day 2, 100 MKD, for 14 days
Suppressed lesion development [320]
Dihydroartemisinin (sesquiterpene lactones)
Golden hamster, 5 animals per group
L. donovani, 5×106
amastigotes, intraperitoneally
From day 15; 25 and 50 MKD orally for 14 days
Reduced parasite load in liver by 80.92%, and in spleen – by up to 75.45%
[321]
Mouse, C57BL/6, females, 5-7 animals per group, 2 independent experiments
L. amazonensis, 106
amastigotes, subcutaneously
From day 36-43, 100 MKD intrarectal route for 14 days
Reduced lesion size and parasite load in lesion in 80% of animals
Limonene (cyclic terpenes)
Mouse, C57BL/6, females, 5-7 animals per group, 3 independent experiments
L. amazonensis, 106
amastigotes, subcutaneously
From day 36-43, 10% cream of limonene wt/wt topically
Complete clinical healing was observed in 67-86% of infected mice
[322]
Biphosphonates
Pamidronate Mouse, BALB/c, females, 5 animals per group
L. donovani, 2×107
promastigotes, intravenously
From day 7; 20 MKD intraperitoneally and orally, 0.4, 2 and 10 MKD intravenously for 5 days
Reduced parasite load in liver by up to 91.8 %
[323]
Risedronate Mouse, BALB/c, females, 5 animals per group
L. donovani, 2×107
promastigotes, intravenously
From day 7; 20 MKD intraperitoneally and orally, 1, 1.6, 3, 5, 10 and 15 MKD intravenously for 5 days
Reduced parasite load in liver by up to 99.6 %
[323]
Receptor antagonists
Tamoxifen (oestrogen receptor antagonist)
Mouse, BALB/c, females, 7-10 animals per group, 3 independent experiments
L. amazonensis, 5×106
promastigotes, subcutaneously
From day 36, 20 MKD intraperitoneally for 15 days
Reduced lesion size and reduced parasite load in lesion (by 99.7%) and in lymph nodes
[324]
Mouse, BALB/c, females, 6-8 animals per group
L. braziliensis, 105
promastigotes, intradermally
From day 22, 20 MKD intraperitoneally for 15 days
Reduced lesion size, reduced parasite load in lesion by 99.0%
Tamoxifen (oestrogen receptor antagonist)
Golden hamster, 6-12 animals per group
L. chagasi, 108 amastigotes, intraperitoneally
From day 29, 20 MKD intraperitoneally for 15 days
Reduced parasite load in liver and spleen by up to 98%
[325]
Naloxone (opioid antagonist)
Mouse, BALB/c, males, 5 animals per group
L. major, 106
promastigotes, subcutaneously
From day 1; 2,10 and 20 MKD groups, subcutaneously for 63-70 days
Suppressed lesion development [326]
Other
Furan-2-carboxamide (arylcarboxamides)
Mouse, BALB/c, males, 5 animals per group
L. donovani, 5×106
promastigotes, intravenously
From day 8; 10 MKD orally for 5 days
Reduced parasite load in liver by 36.6%, and in spleen – by 75.8%
[327]
Mouse, BALB/c nu/+, males, 10 animals per group
L. amazonensis, 103 promastigotes,intradermally plus 105 promastigotes, subcutaneously
From day 3; 15, 30 and 60 MKD, orally for 21 days
Reduced tail and ear lesions, parasite load in ears and lymph nodes
Posaconazole (azoles)
Mouse, BALB/c nu/+, females, 16 animals per group
L. donovani, 106
promastigotes, intravenously
From day 3, 15, 30 and 60 MKD, orally for 21 days
Reduced parasite load in liver and spleen by up to 90%
[328]

Leishmaniasis: Prevention, Parasite Detection and Treatment Current Medicinal Chemistry, 2012 Vol. 19, No. 10 1467
ultraviolet and infrared heat and ultrasound. Radio-frequency waves were applied in clinical trials for local heating to about 50ºC for 30 seconds with subsequent limiting of amastigote replication [306-308]. Several studies compared efficacy of pentavalent antimonials that is usually used for treatment of cutatenous leishmaniasis with thermotherapy. Healing rates for these two methods were similar both in case of skin lesions caused by L. tropica (about 70% patients were successfully cured in groups of 108 and 93 patients, for thermotherapy and intralesional stibogluconate, respectively) [306] and L. major (about 50% cured in two groups for thermotherapy and intravenous stibogluconate, 27 patients each) [308]. However, other studies show much higher cure rate for pentavalent antimonials – 97.2% for L. tropica (1030 patients, intralesional administration) [245] and 81% for L. major (207 patients, intramuscular administration) [246]. In accordance with this, the rates healing of infection with L. braziliensis were also higher for the antimonial therapy (90%) in comparison with heat application (75%) [307]. In a different study, two HIV-infected individuals with cutaneous leishmaniasis caused by L. tropica were successfully treated with heat. In one of these patients, treatment with sodium stibogluconate and rifampicin failed. Fast control on Leishmania infection is crucial for HIV-infected patients, because immunodeficiency may lead to rapid dissemination of Leishmania parasite and development of diffuse disease [309].
4.5. Perspectives
Lack of the efficiency of current antileishmanial drugs or their high price as well as the issues of adverse effects and drug resistance imply the importance of search for new leishmanicidal compounds. A range of natural and synthetic compounds is being studied for the leishmanicidal activity (Table 7). A special attention is paid to various natural phenols, quinolines and terpenes of plant origin and their analogs and derivatives [310-322]. Biphosphonates, receptor antagonists and azoles are also widely studied [323-328]. There are attempts of repurposing of antitumour [315, 318, 324, 325], antifungal [328], antimalarial [321] and anti osteoporosis [323] drugs for the treatment of leishmaniasis. Finally, the era of high throughput analysis brings new approaches, which enable the discovery of leishmanicidal compounds from new chemical classes with new modes of action [230]. On one hand, high throughput screenings for leishmanicidal compounds may be based on the decrease of Leishmania promastigote or amastigote viability, being target-independent [329, 330]. On the other hand, screenings may be performed for the inhibitors of Leishmania essential enzymes, e.g. CRK3:CYC6 cyclin-dependent kinase, being target-driven [331, 332]. Promissing drug delivery systems such as nanoparticles, liposomes, cochleates, and non-specific lipid transfer proteins could also improve treatment [333, 334]. Some of the serious adverse drug reactions might be avoided by systematically screening for pharmacogenomic risk factors. During the last few years, regulatory agencies introduced pharmacogenomics labels for several drugs, but although a priori genetic testing remains advised or recommended, it is seldom compulsory due to poor evidence-based medicine knowledge [335]. In present, none genetic data about genetic influence on response to drugs against leishmaniasis are available. Establishment of chemical-gene-disease networks is therefore of crucial importance [336].
ACKNOWLEDGEMENTS
We thank Dr. Margarita V. Strelkova, E.I. Martsinovsky Institute of Medical Parasitology and Tropical Medicine, Moscow, Russian Federation for providing us materials about leishmaniasis in the Commonwealth of Independent States, and Dr. Peter Demant, Roswell Park Cancer Institute, Buffalo, NY, USA for the critical reading of the manuscript. This study was supported by the Grant Agency of the Czech Republic (Grant Nr. 310/08/1697), and by the
Ministry of Education of the Czech Republic (Project grant LC06009). Ph.D. study of T. Kobets and I. Gregov was supported in part by the Faculty of Science and Third Faculty of Medicine, respectively, Charles University in Prague, Czech Republic.
REFERENCES
[1] Herwaldt, B.L. Leishmaniasis. Lancet, 1999, 354(9185), 1191-1199. [2] Pavli, A.; Maltezou, H.C. Leishmaniasis, an emerging infection in travelers.
Int. J. Infect. Dis., 2010, 14(12), e1032-e1039. [3] Bates, P.A. Transmission of Leishmania metacyclic promastigotes by
phlebotomine sand flies. Int. J. Parasitol., 2007, 37(10), 1097-1106. [4] Lipoldová, M.; Demant, P. Genetic susceptibility to infectious disease:
lessons from mouse models of leishmaniasis. Nat. Rev. Genet., 2006, 7(4), 294-305.
[5] Terrazas, C.A.; Terrazas, L.I.; Gomez-Garcia, L. Modulation of dendritic cell responses by parasites: a common strategy to survive. J. Biomed. Biotechnol., 2010, 2010, 357106.
[6] Bogdan, C. Mechanisms and consequences of persistence of intracellular pathogens: leishmaniasis as an example. Cell. Microbiol., 2008, 10(6), 1221-1234.
[7] Tuon, F.F.; Neto, V.A.; Amato, V.S. Leishmania: origin, evolution and future since the Precambrian. FEMS Immunol Med. Microbiol., 2008, 54(2), 158-166.
[8] Zink, A.R.; Spigelman, M.; Schraut, B.; Greenblatt, C.L.; Nerlich, A.G.; Donoghue, H.D. Leishmaniasis in ancient Egypt and Upper Nubia. Emerg. Infect. Dis., 2006, 12(10), 1616-1617.
[9] Costa, M.A.; Matheson, C.; Iachetta, L.; Llagostera, A.; Appenzeller, O. Ancient leishmaniasis in a highland desert of Northern Chile. PLoS One, 2009, 4(9), e6983.
[10] Cunningham, D. D. On the presence of peculiar parasitic organisms in the tissue of a specimen of Delhi boil. Scientific memoirs by officers of the Medical and Sanitary Departments of the Government of India; Government printing: Calcutta, India, 1885; 1, 21–31.
[11] Borovsky, P.F. [On Sart Sore.] Voenno-Medicinskij Zurnal (Military Medical Journal), 1898, 195(11), 925-941.
[12] Leishman, W.B. On the possibility of the occurence of trypanosomiasis in India. Brit. Med. J., 1903, 1,1252-1254.
[13] Donovan, C. Memoranda: on the possibility of the occurence of trypanomiasis in India. Brit. Med. J., 1903, 2, 79.
[14] Martsinovsky, E.I. [The etiology of oriental sore.] DSc. Thesis, Moscow, 1909.
[15] Dujardin, J.C.; Campino, L.; Canavate, C.; Dedet, J.P.; Gradoni, L.; Soteriadou, K.; Mazeris, A.; Ozbel, Y.; Boelaert, M. Spread of vector-borne diseases and neglect of leishmaniasis, Europe. Emerg. Infect. Dis., 2008, 14(7), 1013-1018.
[16] Machado, C.M.; Martins, T.C.; Colturato, I.; Leite, M.S.; Simione, A.J.; Souza, M.P.; Mauad, M.A.; Colturato, V.R. Epidemiology of neglected tropical diseases in transplant recipients. Review of the literature and experience of a Brazilian HSCT center. Rev. Inst. Med. Trop. Sao Paulo, 2009, 51(6), 309-324.
[17] Postigo, J.A. Leishmaniasis in the World Health Organization Eastern Mediterranean Region. Int. J. Antimicrob. Agents, 2010, 36(1), 62-65.
[18] Ponirovskii, E.N.; Strelkova, M.V.; Goncharov, D.B.; Zhirenkina, E.N.; Chernikova Iu, A. [Visceral leishmaniasis in the Commonwealth of Independent States (CIS): results and basic lines of further study]. Med. Parazitol. (Mosk.), 2006, 4, 25-31.
[19] Sacks, D.; Noben-Trauth, N. The immunology of susceptibility and resistance to Leishmania major in mice. Nat. Rev. Immunol., 2002, 2(11), 845-858.
[20] Mougneau, E.; Bihl, F.; Glaichenhaus, N. Cell biology and immunology of Leishmania. Immunol. Rev., 2011, 240(1), 286-296.
[21] Handman, E. Leishmaniasis: current status of vaccine development. Clin. Microbiol. Rev., 2001, 14(2), 229-243.
[22] Nadim, A.J.; Javadian E.; Mohebali, M. The experience of leishmanization in the Islamic Republic of Iran. East. Mediterr. Health J., 1997, 3(2), 284-289.
[23] Nogueira, F.S.; Moreira, M.A.; Borja-Cabrera, G.P.; Santos, F.N.; Menz, I.; Parra, L.E.; Xu, Z.; Chu, H.J.; Palatnik-de-Sousa, C.B.; Luvizotto, M.C. Leishmune vaccine blocks the transmission of canine visceral leishmaniasis: absence of Leishmania parasites in blood, skin and lymph nodes of vaccinated exposed dogs. Vaccine, 2005, 23(40), 4805-4810.
[24] Mock, B.A.; Fortier, A.H.; Potter, M.; Nacy, C.A. Genetic control of systemic Leishmania major infections: dissociation of intrahepatic amastigote replication from control by the Lsh gene. Infect. Immun., 1985, 50(2), 588-591.
[25] Vidal, S.M.; Malo, D.; Vogan, K.; Skamene, E.; Gros, P. Natural resistance to infection with intracellular parasites: isolation of a candidate for Bcg. Cell, 1993, 73(3), 469-485.
[26] Demant, P.; Lipoldová, M.; Svobodová, M. Resistance to Leishmania major in mice. Science, 1996, 274(5291), 1392a.
[27] Beebe, A.M.; Mauze, S.; Schork, N.J.; Coffman, R.L. Serial backcross mapping of multiple loci associated with resistance to Leishmania major in mice. Immunity, 1997, 6(5), 551-557.

1468 Current Medicinal Chemistry, 2012 Vol. 19, No. 10 Kobets et al.
[28] Lipoldová, M.; Svobodová, M.; Krulová, M.; Havelková, H.; Badalová, J.; Nohýnková, E.; Holáň, V.; Hart, A.A.; Volf, P.; Demant, P. Susceptibility to Leishmania major infection in mice: multiple loci and heterogeneity of immunopathological phenotypes. Genes Immun., 2000, 1(3), 200-206.
[29] Vladimirov, V.; Badalová, J.; Svobodová, M.; Havelková, H.; Hart, A.A.; Blažková, H.; Demant, P.; Lipoldová, M. Different genetic control of cutaneous and visceral disease after Leishmania major infection in mice. Infect. Immun., 2003, 71(4), 2041-2046.
[30] Sakthianandeswaren, A.; Curtis, J.M.; Elso, C.; Kumar, B.; Baldwin, T.M.; Lopaticki, S.; Kedzierski, L.; Smyth, G.K.; Foote, S.J.; Handman, E. Fine mapping of Leishmania major susceptibility locus lmr2 and evidence of a role for Fli1 in disease and wound healing. Infect. Immun., 2010, 78(6), 2734-2744.
[31] Kellina, O.I.; Strelkova, M.V. [Investigations on leishmaniases at the E.I. Martsinovsky Institute of Medical Parasitology and Tropical Medicine]. Med. Parazitol. (Mosk.), 2010, 4, 19-22.
[32] Gafurov, I.M. [Experience in controlling and preventing zoonotic cutaneous leishmaniasis in Uzbekistan]. Med. Parazitol. (Mosk.), 1999, 1, 58-59.
[33] Amini, H. Cutaneous lesions with very long duration as a complication of leishmanization. Iranian J. Publ. Health, 1991, 20(1-4), 43-49.
[34] Khamesipour, A.; Dowlati, Y.; Asilian, A.; Hashemi-Fesharki, R.; Javadi, A.; Noazin, S.; Modabber, F. Leishmanization: use of an old method for evaluation of candidate vaccines against leishmaniasis. Vaccine, 2005, 23(28), 3642-3648.
[35] Grekov, I.; Svobodová, M.; Nohýnková, E.; Lipoldová, M. Preparation of highly infective Leishmania promastigotes by cultivation on SNB-9 biphasic medium. J. Microbiol. Methods, 2011, 87, 273-277.
[36] Hill, J.O.; North, R.J.; Collins, F.M. Advantages of measuring changes in the number of viable parasites in murine models of experimental cutaneous leishmaniasis. Infect. Immun., 1983, 39(3), 1087-1094.
[37] Aebischer, T.; Moody, S.F.; Handman, E. Persistence of virulent Leishmania major in murine cutaneous leishmaniasis: a possible hazard for the host. Infect. Immun., 1993, 61(1), 220-226.
[38] Alvar, J.; Aparicio, P.; Aseffa, A.; Den Boer, M.; Canavate, C.; Dedet, J.P.; Gradoni, L.; Ter Horst, R.; Lopez-Velez, R.; Moreno, J. The relationship between leishmaniasis and AIDS: the second 10 years. Clin. Microbiol. Rev., 2008, 21(2), 334-359.
[39] Jenner, E. An inquiry into the causes and effects of the Variolae Vaccinae, a disease discovered in some of the western counties of England, particularly Gloucestershire, and known by the name of the cow pox; Sampson Low: London, 1798. http://www.gutenberg.org/files/29414/29414-h/29414-h.htm (accessed September 7, 2011)
[40] Breton, M.; Tremblay, M.J.; Ouellette, M.; Papadopoulou, B. Live nonpathogenic parasitic vector as a candidate vaccine against visceral leishmaniasis. Infect. Immun., 2005, 73(10), 6372-6382.
[41] Azizi, H.; Hassani, K.; Taslimi, Y.; Najafabadi, H.S.; Papadopoulou, B.; Rafati, S. Searching for virulence factors in the non-pathogenic parasite to humans Leishmania tarentolae. Parasitology, 2009, 136(7), 723-735.
[42] Mizbani, A.; Taslimi, Y.; Zahedifard, F.; Taheri, T.; Rafati, S. Effect of A2 gene on infectivity of the nonpathogenic parasite Leishmania tarentolae. Parasitol. Res., 2011, 109(3), 793-799.
[43] Alexander, J. A radioattenuated Leishmania major vaccine markedly increases the resistance of CBA mice to subsequent infection with Leishmania mexicana mexicana. Trans. R. Soc. Trop. Med. Hyg., 1982, 76(5), 646-649.
[44] Rivier, D.; Shah, R.; Bovay, P.; Mauel, J. Vaccine development against cutaneous leishmaniasis. Subcutaneous administration of radioattenuated parasites protects CBA mice against virulent Leishmania major challenge. Parasite Immunol., 1993, 15(2), 75-84.
[45] Marchand, M.; Daoud, S.; Titus, R.G.; Louis, J.; Boon, T. Variants with reduced virulence derived from Leishmania major after mutagen treatment. Parasite Immunol., 1987, 9(1), 81-92.
[46] Gorczynski, R.M. Immunization of susceptible BALB/c mice against Leishmania braziliensis. II. Use of temperature-sensitive avirulent clones of parasite for vaccination purposes. Cell Immunol., 1985, 94(1), 11-20.
[47] Mitchell, G.F.; Handman, E.; Spithill, T.W. Vaccination against cutaneous leishmaniasis in mice using nonpathogenic cloned promastigotes of Leishmania major and importance of route of injection. Aust. J. Exp. Biol. Med. Sci., 1984, 62(2), 145-53.
[48] Daneshvar, H.; Coombs, G.H.; Hagan, P.; Phillips, R.S. Leishmania mexicana and Leishmania major: attenuation of wild-type parasites and vaccination with the attenuated lines. J. Infect. Dis., 2003, 187(10), 1662-1668.
[49] Daneshvar, H.; Molaei, M.M.; Kamiabi, H.; Burchmore, R.; Hagan, P.; Phillips, R.S. Gentamicin-attenuated Leishmania infantum: cellular immunity production and protection of dogs against experimental canine leishmaniasis. Parasite Immunol., 2010, 32(11-12), 722-730.
[50] Cruz, A.; Coburn, C.M.; Beverley, S.M. Double targeted gene replacement for creating null mutants. Proc. Natl. Acad. Sci. U S A, 1991, 88(16), 7170-7174.
[51] Titus, R.G.; Gueiros-Filho, F.J.; de Freitas, L.A.; Beverley, S.M. Development of a safe live Leishmania vaccine line by gene replacement. Proc. Natl. Acad. Sci. U S A, 1995, 92(22), 10267-10271.
[52] Veras, P.; Brodskyn, C.; Balestieri, F.; Freitas, L.; Ramos, A.; Queiroz, A.; Barral, A.; Beverley, S.; Barral-Netto, M. A dhfr-ts- Leishmania major
knockout mutant cross-protects against Leishmania amazonensis. Mem. Inst. Oswaldo Cruz, 1999, 94(4), 491-496.
[53] Amaral, V.F.; Teva, A.; Oliveira-Neto, M.P.; Silva, A.J.; Pereira, M.S.; Cupolillo, E.; Porrozzi, R.; Coutinho, S.G.; Pirmez, C.; Beverley, S.M.; Grimaldi, G., Jr. Study of the safety, immunogenicity and efficacy of attenuated and killed Leishmania (Leishmania) major vaccines in a rhesus monkey (Macaca mulatta) model of the human disease. Mem. Inst. Oswaldo Cruz, 2002, 97(7), 1041-1048.
[54] Alexander, J.; Coombs, G.H.; Mottram, J.C. Leishmania mexicana cysteine proteinase-deficient mutants have attenuated virulence for mice and potentiate a Th1 response. J. Immunol., 1998, 161(12), 6794-6801.
[55] Papadopoulou, B.; Roy, G.; Breton, M.; Kundig, C.; Dumas, C.; Fillion, I.; Singh, A.K.; Olivier, M.; Ouellette, M. Reduced infectivity of a Leishmania donovani biopterin transporter genetic mutant and its use as an attenuated strain for vaccination. Infect. Immun., 2002, 70(1), 62-68.
[56] Uzonna, J.E.; Spath, G.F.; Beverley, S.M.; Scott, P. Vaccination with phosphoglycan-deficient Leishmania major protects highly susceptible mice from virulent challenge without inducing a strong Th1 response. J. Immunol., 2004, 172(6), 3793-3797.
[57] Silvestre, R.; Cordeiro-Da-Silva, A.; Santarem, N.; Vergnes, B.; Sereno, D.; Ouaissi, A. SIR2-deficient Leishmania infantum induces a defined IFN-gamma/IL-10 pattern that correlates with protection. J. Immunol., 2007, 179(5), 3161-3170.
[58] Kedzierski, L.; Curtis, J.M.; Doherty, P.C.; Handman, E.; Kedzierska, K. Decreased IL-10 and IL-13 production and increased CD44hi T cell recruitment contribute to Leishmania major immunity induced by non-persistent parasites. Eur. J. Immunol., 2008, 38(11), 3090-3100.
[59] Selvapandiyan, A.; Dey, R.; Nylen, S.; Duncan, R.; Sacks, D.; Nakhasi, H.L. Intracellular replication-deficient Leishmania donovani induces long lasting protective immunity against visceral leishmaniasis. J. Immunol., 2009, 183(3), 1813-1820.
[60] Spath, G.F.; Lye, L.F.; Segawa, H.; Sacks, D.L.; Turco, S.J.; Beverley, S.M. Persistence without pathology in phosphoglycan-deficient Leishmania major. Science, 2003, 301(5637), 1241-1243.
[61] Shankar, A.; Mitchen, T.K.; Hall, L.R.; Turco, S.J.; Titus, R.G. Reversion to virulence in Leishmania major correlates with expression of surface lipophosphoglycan. Mol. Biochem. Parasitol., 1993, 61(2), 207-216.
[62] Davoudi, N.; Tate, C.A.; Warburton, C.; Murray, A.; Mahboudi, F.; McMaster, W.R. Development of a recombinant Leishmania major strain sensitive to ganciclovir and 5-fluorocytosine for use as a live vaccine challenge in clinical trials. Vaccine, 2005, 23(9), 1170-1177.
[63] Kumari, S.; Samant, M.; Khare, P.; Misra, P.; Dutta, S.; Kolli, B.K.; Sharma, S.; Chang, K.P.; Dube, A. Photodynamic vaccination of hamsters with inducible suicidal mutants of Leishmania amazonensis elicits immunity against visceral leishmaniasis. Eur. J. Immunol., 2009, 39(1), 178-191.
[64] Nylen, S.; Mortberg, U.; Kovalenko, D.; Satti, I.; Engstrom, K.; Bakhiet, M.; Akuffo, H. Differential induction of cellular responses by live and dead Leishmania promastigotes in healthy donors. Clin. Exp. Immunol., 2001, 124(1), 43-53.
[65] Nagill, R.; Mahajan, R.; Sharma, M.; Kaur, S. Induction of cellular and humoral responses by autoclaved and heat-killed antigen of Leishmania donovani in experimental visceral leishmaniasis. Parasitol. Int.,, 2009, 58(4), 359-366.
[66] Mutiso, J.M.; Macharia, J.C.; Mutisya, R.M.; Taracha, E. Subcutaneous immunization against Leishmania major - infection in mice: efficacy of formalin-killed promastigotes combined with adjuvants. Rev. Inst. Med. Trop. Sao Paulo, 2010, 52(2), 95-100.
[67] Okwor, I.; Kuriakose, S.; Uzonna, J. Repeated inoculation of killed Leishmania major induces durable immune response that protects mice against virulent challenge. Vaccine, 2010, 28(33), 5451-5457.
[68] Noazin, S.; Khamesipour, A.; Moulton, L.H.; Tanner, M.; Nasseri, K.; Modabber, F.; Sharifi, I.; Khalil, E.A.; Bernal, I.D.; Antunes, C.M.; Smith, P.G. Efficacy of killed whole-parasite vaccines in the prevention of leishmaniasis: a meta-analysis. Vaccine, 2009, 27(35), 4747-4753.
[69] Modabber, F. Leishmaniasis vaccines: past, present and future. Int. J. Antimicrob. Agents, 2010, 36(1), 58-61.
[70] De Luca, P.M.; Mayrink, W.; Alves, C.R.; Coutinho, S.G.; Oliveira, M.P.; Bertho, A.L.; Toledo, V.P.; Costa, C.A.; Genaro, O.; Mendonca, S.C. Evaluation of the stability and immunogenicity of autoclaved and nonautoclaved preparations of a vaccine against American tegumentary leishmaniasis. Vaccine, 1999, 17(9-10), 1179-1185.
[71] Tafaghodi, M.; Eskandari, M.; Khamesipour, A.; Jaafari, M.R. Alginate microspheres encapsulated with autoclaved Leishmania major (ALM) and CpG-ODN induced partial protection and enhanced immune response against murine model of leishmaniasis. Exp. Parasitol., 2011, 129(2), 107-114.
[72] Machado-Pinto, J.; Pinto, J.; da Costa, C.A.; Genaro, O.; Marques, M.J.; Modabber, F.; Mayrink, W. Immunochemotherapy for cutaneous leishmaniasis: a controlled trial using killed Leishmania (Leishmania) amazonensis vaccine plus antimonial. Int. J. Dermatol., 2002, 41(2), 73-78.
[73] Stober, C.B.; Lange, U.G.; Roberts, M.T.; Gilmartin, B.; Francis, R.; Almeida, R.; Peacock, C.S.; McCann, S.; Blackwell, J.M. From genome to vaccines for leishmaniasis: screening 100 novel vaccine candidates against murine Leishmania major infection. Vaccine, 2006, 24(14), 2602-2616.
[74] de Matos Guedes, H.L.; Pinheiro, R.O.; Chaves, S.P.; De-Simone, S.G.; Rossi-Bergmann, B. Serine proteases of Leishmania amazonensis as

Leishmaniasis: Prevention, Parasite Detection and Treatment Current Medicinal Chemistry, 2012 Vol. 19, No. 10 1469
immunomodulatory and disease-aggravating components of the crude LaAg vaccine. Vaccine, 2010, 28(33), 5491-5496.
[75] Pizza, M.; Scarlato, V.; Masignani, V.; Giuliani, M.M.; Arico, B.; Comanducci, M.; Jennings, G.T.; Baldi, L.; Bartolini, E.; Capecchi, B.; Galeotti, C.L.; Luzzi, E.; Manetti, R.; Marchetti, E.; Mora, M.; Nuti, S.; Ratti, G.; Santini, L.; Savino, S.; Scarselli, M.; Storni, E.; Zuo, P.; Broeker, M.; Hundt, E.; Knapp, B.; Blair, E.; Mason, T.; Tettelin, H.; Hood, D.W.; Jeffries, A.C.; Saunders, N.J.; Granoff, D.M.; Venter, J.C.; Moxon, E.R.; Grandi, G.; Rappuoli, R. Identification of vaccine candidates against serogroup B meningococcus by whole-genome sequencing. Science, 2000, 287(5459), 1816-1820.
[76] Sette, A.; Rappuoli, R. Reverse vaccinology: developing vaccines in the era of genomics. Immunity, 2010, 33(4), 530-541.
[77] Working Group on Research Priorities for Development of Leishmaniasis Vaccines, Costa, C.H.; Peters, N.C.; Maruyama, S.R.; de Brito, E.C., Jr.; Santos, I.K. Vaccines for the leishmaniases: proposals for a research agenda. PLoS Negl. Trop. Dis., 2011, 5(3), e943: Available from: http://www.plosntds.org/article/info%3Adoi%2F10.1371%2Fjournal.pntd.0000943
[78] Nagill, R.; Kaur, S. Vaccine candidates for leishmaniasis: A review. Int. Immunopharmacol., 2011, 11(10), 1464-1488.
[79] Dantas-Torres, F. Leishmune vaccine: the newest tool for prevention and control of canine visceral leishmaniosis and its potential as a transmission-blocking vaccine. Vet. Parasitol., 2006, 141(1-2), 1-8.
[80] Palatnik-de-Sousa, C.B.; Silva-Antunes, I.; Morgado Ade, A.; Menz, I.; Palatnik, M.; Lavor, C. Decrease of the incidence of human and canine visceral leishmaniasis after dog vaccination with Leishmune in Brazilian endemic areas. Vaccine, 2009, 27(27), 3505-3512.
[81] Nico, D.; Claser, C.; Borja-Cabrera, G.P.; Travassos, L.R.; Palatnik, M.; Soares, I.S.; Rodrigues, M.M.; Palatnik-de-Sousa, C.B. Adaptive immunity against Leishmania nucleoside hydrolase maps its c-terminal domain as the target of the CD4+ T cell-driven protective response. PLoS Negl. Trop. Dis., 2010, 4(11), e866.
[82] Day, M.J. The immunopathology of canine vector-borne diseases. Parasit. Vectors, 2011, 4, 48.
[83] Jardim, A.; Alexander, J.; Teh, H.S.; Ou, D.; Olafson, R.W. Immunoprotective Leishmania major synthetic T cell epitopes. J. Exp. Med., 1990, 172(2), 645-648.
[84] Spitzer, N.; Jardim, A.; Lippert, D.; Olafson, R.W. Long-term protection of mice against Leishmania major with a synthetic peptide vaccine. Vaccine, 1999, 17(11-12), 1298-1300.
[85] Rivier, D.; Bovay, P.; Shah, R.; Didisheim, S.; Mauel, J. Vaccination against Leishmania major in a CBA mouse model of infection: role of adjuvants and mechanism of protection. Parasite Immunol., 1999, 21(9), 461-473.
[86] Jaafari, M.R.; Ghafarian, A.; Farrokh-Gisour, A.; Samiei, A.; Kheiri, M.T.; Mahboudi, F.; Barkhordari, F.; Khamesipour, A.; McMaster, W.R. Immune response and protection assay of recombinant major surface glycoprotein of Leishmania (rgp63) reconstituted with liposomes in BALB/c mice. Vaccine, 2006, 24(29-30), 5708-5717.
[87] Heneberg, P. Finding the smoking gun: protein tyrosine phosphatases as tools and targets of unicellular microorganisms and viruses. Curr. Med. Chem., 2012, in press.
[88] Mougneau, E.; Altare, F.; Wakil, A.E.; Zheng, S.; Coppola, T.; Wang, Z.E.; Waldmann, R.; Locksley, R.M.; Glaichenhaus, N. Expression cloning of a protective Leishmania antigen. Science, 1995, 268(5210), 563-566.
[89] Gonzalez-Aseguinolaza, G.; Taladriz, S.; Marquet, A.; Larraga, V. Molecular cloning, cell localization and binding affinity to DNA replication proteins of the p36/LACK protective antigen from Leishmania infantum. Eur. J. Biochem., 1999, 259(3), 909-916.
[90] Van der Voorn, L.; Ploegh, H.L. The WD-40 repeat. FEBS Lett., 1992, 307(2), 131-134.
[91] Gurunathan, S.; Sacks, D.L.; Brown, D.R.; Reiner, S.L.; Charest, H.; Glaichenhaus, N.; Seder, R.A. Vaccination with DNA encoding the immunodominant LACK parasite antigen confers protective immunity to mice infected with Leishmania major. J. Exp Med., 1997, 186(7), 1137-1147.
[92] Devault, A.; Banuls, A.L. The promastigote surface antigen gene family of the Leishmania parasite: differential evolution by positive selection and recombination. BMC Evol. Biol., 2008, 8, 292.
[93] Handman, E.; Symons, F.M.; Baldwin, T.M.; Curtis, J.M.; Scheerlinck, J.P. Protective vaccination with promastigote surface antigen 2 from Leishmania major is mediated by a TH1 type of immune response. Infect. Immun., 1995, 63(11), 4261-4267.
[94] Al-Wabel, M.A.; Tonui, W.K.; Cui, L.; Martin, S.K.; Titus, R.G. Protection of susceptible BALB/c mice from challenge with Leishmania major by nucleoside hydrolase, a soluble exo-antigen of Leishmania. Am. J. Trop. Med. Hyg., 2007, 77(6), 1060-1065.
[95] Coler, R.N.; Goto, Y.; Bogatzki, L.; Raman, V.; Reed, S.G. Leish-111f, a recombinant polyprotein vaccine that protects against visceral leishmaniasis by elicitation of CD4+ T cells. Infect. Immun., 2007, 75(9), 4648-4654.
[96] Velez, I.D.; Gilchrist, K.; Martinez, S.; Ramirez-Pineda, J.R.; Ashman, J.A.; Alves, F.P.; Coler, R.N.; Bogatzki, L.Y.; Kahn, S.J.; Beckmann, A.M.; Cowgill, K.D.; Reed, S.G.; Piazza, F.M. Safety and immunogenicity of a defined vaccine for the prevention of cutaneous leishmaniasis. Vaccine, 2009, 28(2), 329-337.
[97] Chakravarty, J.; Kumar, S.; Trivedi, S.; Rai, V.K.; Singh, A.; Ashman, J.A.; Laughlin, E.M.; Coler, R.N.; Kahn, S.J.; Beckmann, A.M.; Cowgill, K.D.; Reed, S.G.; Sundar, S.; Piazza, F.M. A clinical trial to evaluate the safety and immunogenicity of the LEISH-F1+MPL-SE vaccine for use in the prevention of visceral leishmaniasis. Vaccine, 2011, 29(19), 3531-3537.
[98] Skeiky, Y.A.; Coler, R.N.; Brannon, M.; Stromberg, E.; Greeson, K.; Crane, R.T.; Webb, J.R.; Campos-Neto, A.; Reed, S.G. Protective efficacy of a tandemly linked, multi-subunit recombinant leishmanial vaccine (Leish-111f) formulated in MPL adjuvant. Vaccine, 2002, 20(27-28), 3292-3303.
[99] Gradoni, L.; Foglia Manzillo, V.; Pagano, A.; Piantedosi, D.; De Luna, R.; Gramiccia, M.; Scalone, A.; Di Muccio, T.; Oliva, G. Failure of a multi-subunit recombinant leishmanial vaccine (MML) to protect dogs from Leishmania infantum infection and to prevent disease progression in infected animals. Vaccine, 2005, 23(45), 5245-5251.
[100] Bertholet, S.; Goto, Y.; Carter, L.; Bhatia, A.; Howard, R.F.; Carter, D.; Coler, R.N.; Vedvick, T.S.; Reed, S.G. Optimized subunit vaccine protects against experimental leishmaniasis. Vaccine, 2009, 27(50), 7036-7045.
[101] Goto, Y.; Bhatia, A.; Raman, V.S.; Liang, H.; Mohamath, R.; Picone, A.F.; Vidal, S.E.; Vedvick, T.S.; Howard, R.F.; Reed, S.G. KSAC, the first defined polyprotein vaccine candidate for visceral leishmaniasis. Clin. Vaccine Immunol., 2011, 18(7), 1118-1124.
[102] Molano, I.; Alonso, M.G.; Miron, C.; Redondo, E.; Requena, J.M.; Soto, M.; Nieto, C.G.; Alonso, C. A Leishmania infantum multi-component antigenic protein mixed with live BCG confers protection to dogs experimentally infected with L. infantum. Vet. Immunol. Immunop., 2003, 92(1-2), 1-13.
[103] Carcelen, J.; Iniesta, V.; Fernandez-Cotrina, J.; Serrano, F.; Parejo, J.C.; Corraliza, I.; Gallardo-Soler, A.; Maranon, F.; Soto, M.; Alonso, C.; Gomez-Nieto, C. The chimerical multi-component Q protein from Leishmania in the absence of adjuvant protects dogs against an experimental Leishmania infantum infection. Vaccine, 2009, 27(43), 5964-5973.
[104] Wolff, J.A.; Malone, R.W.; Williams, P.; Chong, W.; Acsadi, G.; Jani, A.; Felgner, P.L. Direct gene transfer into mouse muscle in vivo. Science, 1990, 247(4949 Pt 1), 1465-1468.
[105] Alarcon, J.B.; Waine, G.W.; McManus, D.P. DNA vaccines: technology and application as anti-parasite and anti-microbial agents. Adv. Parasitol., 1999, 42, 343-410.
[106] Rafati, S.; Ghaemimanesh, F.; Zahedifard, F. Comparison of potential protection induced by three vaccination strategies (DNA/DNA, Protein/Protein and DNA/Protein) against Leishmania major infection using signal peptidase type I in BALB/c mice. Vaccine, 2006, 24(16), 3290-3297.
[107] Yang, D.M.; Fairweather, N.; Button, L.L.; McMaster, W.R.; Kahl, L.P.; Liew, F.Y. Oral Salmonella typhimurium (AroA-) vaccine expressing a major leishmanial surface protein (gp63) preferentially induces T helper 1 cells and protective immunity against leishmaniasis. J. Immunol., 1990, 145(7), 2281-2285.
[108] Xu, D.; Liew, F.Y. Protection against leishmaniasis by injection of DNA encoding a major surface glycoprotein, gp63, of L. major. Immunology, 1995, 84(2), 173-176.
[109] Ahmed, S.B.; Touihri, L.; Chtourou, Y.; Dellagi, K.; Bahloul, C. DNA based vaccination with a cocktail of plasmids encoding immunodominant Leishmania (Leishmania) major antigens confers full protection in BALB/c mice. Vaccine, 2009, 27(1), 99-106.
[110] Gomes, D.C.; Pinto, E.F.; de Melo, L.D.; Lima, W.P.; Larraga, V.; Lopes, U.G.; Rossi-Bergmann, B. Intranasal delivery of naked DNA encoding the LACK antigen leads to protective immunity against visceral leishmaniasis in mice. Vaccine, 2007, 25(12), 2168-2172.
[111] Marques-da-Silva, E.A.; Coelho, E.A.; Gomes, D.C.; Vilela, M.C.; Masioli, C.Z.; Tavares, C.A.; Fernandes, A.P.; Afonso, L.C.; Rezende, S.A. Intramuscular immunization with p36 (LACK) DNA vaccine induces IFN-gamma production but does not protect BALB/c mice against Leishmania chagasi intravenous challenge. Parasitol. Res., 2005, 98(1), 67-74.
[112] Doroud, D.; Zahedifard, F.; Vatanara, A.; Najafabadi, A.R.; Taslimi, Y.; Vahabpour, R.; Torkashvand, F.; Vaziri, B.; Rafati, S. Delivery of a cocktail DNA vaccine encoding cysteine proteinases type I, II and III with solid lipid nanoparticles potentiate protective immunity against Leishmania major infection. J. Control. Release, 2011, 153(2), 154-162.
[113] Ivens, A.C.; Peacock, C.S.; Worthey, E.A.; Murphy, L.; Aggarwal, G.; Berriman, M.; Sisk, E.; Rajandream, M.A.; Adlem, E.; Aert, R.; Anupama, A.; Apostolou, Z.; Attipoe, P.; Bason, N.; Bauser, C.; Beck, A.; Beverley, S.M.; Bianchettin, G.; Borzym, K.; Bothe, G.; Bruschi, C.V.; Collins, M.; Cadag, E.; Ciarloni, L.; Clayton, C.; Coulson, R.M.; Cronin, A.; Cruz, A.K.; Davies, R.M.; De Gaudenzi, J.; Dobson, D.E.; Duesterhoeft, A.; Fazelina, G.; Fosker, N.; Frasch, A.C.; Fraser, A.; Fuchs, M.; Gabel, C.; Goble, A.; Goffeau, A.; Harris, D.; Hertz-Fowler, C.; Hilbert, H.; Horn, D.; Huang, Y.; Klages, S.; Knights, A.; Kube, M.; Larke, N.; Litvin, L.; Lord, A.; Louie, T.; Marra, M.; Masuy, D.; Matthews, K.; Michaeli, S.; Mottram, J.C.; Muller-Auer, S.; Munden, H.; Nelson, S.; Norbertczak, H.; Oliver, K.; O'Neil, S.; Pentony, M.; Pohl, T.M.; Price, C.; Purnelle, B.; Quail, M.A.; Rabbinowitsch, E.; Reinhardt, R.; Rieger, M.; Rinta, J.; Robben, J.; Robertson, L.; Ruiz, J.C.; Rutter, S.; Saunders, D.; Schafer, M.; Schein, J.; Schwartz, D.C.; Seeger, K.; Seyler, A.; Sharp, S.; Shin, H.; Sivam, D.; Squares, R.; Squares, S.; Tosato, V.; Vogt, C.; Volckaert, G.; Wambutt, R.; Warren, T.; Wedler, H.; Woodward, J.; Zhou, S.; Zimmermann, W.; Smith, D.F.; Blackwell, J.M.; Stuart, K.D.; Barrell, B.; Myler, P.J. The genome of

1470 Current Medicinal Chemistry, 2012 Vol. 19, No. 10 Kobets et al.
the kinetoplastid parasite, Leishmania major. Science, 2005, 309(5733), 436-442.
[114] Rohoušová, I.; Volf, P. Sand fly saliva: effects on host immune response and Leishmania transmission. Folia Parasitol. (Praha), 2006, 53(3), 161-171.
[115] Teixeira, C.; Gomes, R.; Collin, N.; Reynoso, D.; Jochim, R.; Oliveira, F.; Seitz, A.; Elnaiem, D.E.; Caldas, A.; de Souza, A.P.; Brodskyn, C.I.; de Oliveira, C.I.; Mendonca, I.; Costa, C.H.; Volf, P.; Barral, A.; Kamhawi, S.; Valenzuela, J.G. Discovery of markers of exposure specific to bites of Lutzomyia longipalpis, the vector of Leishmania infantum chagasi in Latin America. PLoS Negl. Trop. Dis., 2010, 4(3), e638.
[116] Theodos, C.M.; Titus, R.G. Salivary gland material from the sand fly Lutzomyia longipalpis has an inhibitory effect on macrophage function in vitro. Parasite Immunol., 1993, 15(8), 481-487.
[117] Rohoušová, I.; Ozensoy, S.; Ozbel, Y.; Volf, P. Detection of species-specific antibody response of humans and mice bitten by sand flies. Parasitology, 2005, 130(5), 493-499.
[118] Kato, H.; Anderson, J.M.; Kamhawi, S.; Oliveira, F.; Lawyer, P.G.; Pham, V.M.; Sangare, C.S.; Samake, S.; Sissoko, I.; Garfield, M.; Sigutova, L.; Volf, P.; Doumbia, S.; Valenzuela, J.G. High degree of conservancy among secreted salivary gland proteins from two geographically distant Phlebotomus duboscqi sandflies populations (Mali and Kenya). BMC Genomics, 2006, 7, 226.
[119] Drahota, J.; Lipoldová, M.; Volf, P.; Rohoušová, I. Specificity of anti-saliva immune response in mice repeatedly bitten by Phlebotomus sergenti. Parasite Immunol., 2009, 31(12), 766-770.
[120] Gomes, R.B.; Brodskyn, C.; de Oliveira, C.I.; Costa, J.; Miranda, J.C.; Caldas, A.; Valenzuela, J.G.; Barral-Netto, M.; Barral, A. Seroconversion against Lutzomyia longipalpis saliva concurrent with the development of anti-Leishmania chagasi delayed-type hypersensitivity. J. Infect. Dis., 2002, 186(10), 1530-1534.
[121] Rohoušová, I.; Volf, P.; Lipoldová, M. Modulation of murine cellular immune response and cytokine production by salivary gland lysate of three sand fly species. Parasite Immunol., 2005, 27(12), 469-473.
[122] de Moura, T.R.; Oliveira, F.; Novais, F.O.; Miranda, J.C.; Clarencio, J.; Follador, I.; Carvalho, E.M.; Valenzuela, J.G.; Barral-Netto, M.; Barral, A.; Brodskyn, C.; de Oliveira, C.I. Enhanced Leishmania braziliensis infection following pre-exposure to sandfly saliva. PLoS Negl. Trop. Dis., 2007, 1(2), e84.
[123] Rohoušová, I.; Hostomská, J.; Vlková, M.; Kobets, T.; Lipoldová, M.; Volf, P., The protective effect against Leishmania infection conferred by sand fly bites is limited to short-term exposure. Int. J. Parasitol., 2011.
[124] Morris, R.V.; Shoemaker, C.B.; David, J.R.; Lanzaro, G.C.; Titus, R.G. Sandfly maxadilan exacerbates infection with Leishmania major and vaccinating against it protects against L. major infection. J. Immunol., 2001, 167(9), 5226-5230.
[125] Xu, X.; Oliveira, F.; Chang, B.W.; Collin, N.; Gomes, R.; Teixeira, C.; Reynoso, D.; My Pham, V.; Elnaiem, D.E.; Kamhawi, S.; Ribeiro, J.M.; Valenzuela, J.G.; Andersen, J.F. Structure and function of a "Yellow" protein from saliva of the sand fly Lutzomyia longipalpis that confers protective immunity against Leishmania major infection. J. Biol. Chem., 2011, 286(37), 32383-32393.
[126] Valenzuela, J.G.; Belkaid, Y.; Garfield, M.K.; Mendez, S.; Kamhawi, S.; Rowton, E.D.; Sacks, D.L.; Ribeiro, J.M. Toward a defined anti-Leishmania vaccine targeting vector antigens: characterization of a protective salivary protein. J. Exp. Med., 2001, 194(3), 331-342.
[127] Oliveira, F.; Lawyer, P.G.; Kamhawi, S.; Valenzuela, J.G. Immunity to distinct sand fly salivary proteins primes the anti-Leishmania immune response towards protection or exacerbation of disease. PLoS Negl. Trop. Dis., 2008, 2(4), e226.
[128] Tabbara, K.S.; Peters, N.C.; Afrin, F.; Mendez, S.; Bertholet, S.; Belkaid, Y.; Sacks, D.L. Conditions influencing the efficacy of vaccination with live organisms against Leishmania major infection. Infect. Immun., 2005, 73(8), 4714-4722.
[129] Sjölander, A.; Baldwin, T.M.; Curtis, J.M.; Bengtsson, K.L.; Handman, E. Vaccination with recombinant parasite surface antigen 2 from Leishmania major induces a Th1 type of immune response but does not protect against infection. Vaccine, 1998, 16(20), 2077-2084.
[130] Campos-Neto, A. In: Leishmania. Farrel, J.P., Ed.; Kluwer Academic Publishers: Boston, 2002; Vol. 4, pp 169-190.
[131] Havelková, H.; Badalová, J.; Svobodová, M.; Vojtíšková, J.; Kurey, I.; Vladimirov, V.; Demant, P.; Lipoldová, M. Genetics of susceptibility to leishmaniasis in mice: four novel loci and functional heterogeneity of gene effects. Genes Immun., 2006, 7(3), 220-233.
[132] Kurey, I.; Kobets, T.; Havelková, H.; Slapničková, M.; Quan, L.; Trtková, K.; Grekov, I.; Svobodová, M.; Stassen, A.P.; Hutson, A.; Demant, P.; Lipoldová, M. Distinct genetic control of parasite elimination, dissemination, and disease after Leishmania major infection. Immunogenetics, 2009, 61(9), 619-633.
[133] Lipoldová, M.; Svobodová, M.; Havelková, H.; Krulová, M.; Badalová, J.; Nohýnková, E.; Hart, A.A.; Schlegel, D.; Volf, P.; Demant, P. Mouse genetic model for clinical and immunological heterogeneity of leishmaniasis. Immunogenetics, 2002, 54(3), 174-183.
[134] Babay, B.E.; Louzir, H.; Kebaier, C.; Boubaker, S.; Dellagi, K.; Cazenave, P.A. Inbred strains derived from feral mice reveal new pathogenic
mechanisms of experimental leishmaniasis due to Leishmania major. Infect. Immun., 2004, 72(8), 4603-4611.
[135] Benhnini, F.; Chenik, M.; Laouini, D.; Louzir, H.; Cazenave, P.A.; Dellagi, K. Comparative evaluation of two vaccine candidates against experimental leishmaniasis due to Leishmania major infection in four inbred mouse strains. Clin. Vaccine Immunol., 2009, 16(11), 1529-1537.
[136] Schroeder, J.; Brown, N.; Kaye, P.; Aebischer, T. Single dose novel Salmonella vaccine enhances resistance against visceralizing L. major and L. donovani infection in susceptible BALB/c mice. PLoS Negl. Trop. Dis., 2011, 5(12): e1406.
[137] Paape, D.; Aebischer, T. Contribution of proteomics of Leishmania spp. to the understanding of differentiation, drug resistance mechanisms, vaccine and drug development. J. Proteomics, 2011, 74(9), 1614-1624.
[138] Jensen, K.D.; Sercarz, E.E.; Gabaglia, C.R. Altered peptide ligands can modify the Th2 T cell response to the immunodominant 161-175 peptide of LACK (Leishmania homolog for the receptor of activated C kinase). Mol. Immunol., 2009, 46(3), 366-374.
[139] Danesh-Bahreini, M.A.; Shokri, J.; Samiei, A.; Kamali-Sarvestani, E.; Barzegar-Jalali, M.; Mohammadi-Samani, S. Nanovaccine for leishmaniasis: preparation of chitosan nanoparticles containing Leishmania superoxide dismutase and evaluation of its immunogenicity in BALB/c mice. Int. J. Nanomedicine, 2011, 6, 835-842.
[140] Remer, K.A.; Roeger, B.; Hambrecht, C.; Moll, H. Natural killer cells support the induction of protective immunity during dendritic cell-mediated vaccination against Leishmania major. Immunology, 2010, 131(4), 570-582.
[141] Schnitzer, J.K.; Berzel, S.; Fajardo-Moser, M.; Remer, K.A.; Moll, H. Fragments of antigen-loaded dendritic cells (DC) and DC-derived exosomes induce protective immunity against Leishmania major. Vaccine, 2010, 28(36), 5785-5793.
[142] Agallou, M.; Margaroni, M.; Karagouni, E. Cellular vaccination with bone marrow-derived dendritic cells pulsed with a peptide of Leishmania infantum KMP-11 and CpG oligonucleotides induces protection in a murine model of visceral leishmaniasis. Vaccine, 2011, 29(31), 5053-5064.
[143] Claborn, D.M. The biology and control of leishmaniasis vectors. J. Glob. Infect. Dis., 2010, 2(2), 127-134.
[144] Daneshvar, H.; Hagan, P.; Phillips, R.S. Leishmania mexicana H-line attenuated under pressure of gentamicin, potentiates a Th1 response and control of cutaneous leishmaniasis in BALB/c mice. Parasite Immunol., 2003, 25(11-12), 589-596.
[145] Armijos, R.X.; Weigel, M.M.; Aviles, H.; Maldonado, R.; Racines, J. Field trial of a vaccine against New World cutaneous leishmaniasis in an at-risk child population: safety, immunogenicity, and efficacy during the first 12 months of follow-up. J. Infect. Dis., 1998, 177(5), 1352-1357.
[146] Armijos, R.X.; Weigel, M.M.; Romero, L.; Garcia, V.; Salazar, J. Field trial of a vaccine against new world cutaneous leishmaniasis in an at-risk child population: how long does protection last? J. Infect. Dis., 2003, 187(12), 1959-1961.
[147] Armijos, R.X.; Weigel, M.M.; Calvopina, M.; Hidalgo, A.; Cevallos, W.; Correa, J. Safety, immunogenecity, and efficacy of an autoclaved Leishmania amazonensis vaccine plus BCG adjuvant against New World cutaneous leishmaniasis. Vaccine, 2004, 22(9-10), 1320-1326.
[148] Velez, I.D.; Gilchrist, K.; Arbelaez, M.P.; Rojas, C.A.; Puerta, J.A.; Antunes, C.M.; Zicker, F.; Modabber, F. Failure of a killed Leishmania amazonensis vaccine against American cutaneous leishmaniasis in Colombia. Trans. R. Soc. Trop. Med. Hyg., 2005, 99(8), 593-598.
[149] Parra, L.E.; Borja-Cabrera, G.P.; Santos, F.N.; Souza, L.O.; Palatnik-de-Sousa, C.B.; Menz, I. Safety trial using the Leishmune vaccine against canine visceral leishmaniasis in Brazil. Vaccine, 2007, 25(12), 2180-2186.
[150] Noormohammadi, A.H.; Hochrein, H.; Curtis, J.M.; Baldwin, T.M.; Handman, E., Paradoxical effects of IL-12 in leishmaniasis in the presence and absence of vaccinating antigen. Vaccine, 2001, 19(28-29), 4043-4052.
[151] Doroud, D.; Zahedifard, F.; Vatanara, A.; Najafabadi, A.R.; Rafati, S. Cysteine proteinase type I, encapsulated in solid lipid nanoparticles induces substantial protection against Leishmania major infection in C57BL/6 mice. Parasite Immunol., 2011, 33(6), 335-348.
[152] Jayakumar, A.; Castilho, T.M.; Park, E.; Goldsmith-Pestana, K.; Blackwell, J.M.; McMahon-Pratt, D. TLR1/2 activation during heterologous prime-boost vaccination (DNA-MVA) enhances CD8+ T Cell responses providing protection against Leishmania (Viannia). PLoS Negl. Trop. Dis., 2011, 5(6), e1204.
[153] Rodriguez, N.; Guzman, B.; Rodas, A.; Takiff, H.; Bloom, B.R.; Convit, J. Diagnosis of cutaneous leishmaniasis and species discrimination of parasites by PCR and hybridization. J. Clin. Microbiol., 1994, 32(9), 2246-2252.
[154] Kobets, T.; Badalová, J.; Grekov, I.; Havelková, H.; Svobodová, M.; Lipoldová, M. Leishmania parasite detection and quantification using PCR-ELISA. Nat. Protoc., 2010, 5(6), 1074-1080.
[155] Barreca, G.S.; Berlinghieri, M.C.; Foti, F.; Matera, G.; Foca, A. Microscopic observation of progressive immobilization of Leishmania promastigotes in acridine orange stain. J. Clin. Microbiol., 1997, 35(7), 1867-1869.
[156] Noyes, H.A.; Reyburn, H.; Bailey, J.W.; Smith, D. A nested-PCR-based schizodeme method for identifying Leishmania kinetoplast minicircle classes directly from clinical samples and its application to the study of the epidemiology of Leishmania tropica in Pakistan. J. Clin. Microbiol., 1998, 36(10), 2877-2881.

Leishmaniasis: Prevention, Parasite Detection and Treatment Current Medicinal Chemistry, 2012 Vol. 19, No. 10 1471
[157] Abdullah, S.M.; Flath, B.; Presber, H.W. Comparison of different staining procedures for the flow cytometric analysis of U-937 cells infected with different Leishmania species. J. Microbiol. Methods, 1999, 37(2), 123-138.
[158] Ferrua, B.; Le Fichoux, Y.; Suffia, I.; Rousseau, D.; Roptin, C.; Kubar, J. Quantitation of Leishmania infantum in tissues of infected BALB/c mouse by sandwich ELISA. J. Immunoassay Immunochem., 2001, 22(2), 165-181.
[159] Weigle, K.A.; Labrada, L.A.; Lozano, C.; Santrich, C.; Barker, D.C. PCR-based diagnosis of acute and chronic cutaneous leishmaniasis caused by Leishmania (Viannia). J. Clin. Microbiol., 2002, 40(2), 601-606.
[160] Royer, M.A.; Crowe, C.O. American cutaneous leishmaniasis. Arch. Pathol. Lab. Med., 2002, 126(4), 471-473.
[161] Allahverdiyev, A.M.; Uzun, S.; Bagirova, M.; Durdu, M.; Memisoglu, H.R. A sensitive new microculture method for diagnosis of cutaneous leishmaniasis. Am. J. Trop. Med. Hyg., 2004, 70(3), 294-297.
[162] Sipahi, T.; Tavil, B.; Oksal, A. Visceral leishmaniasis and Pseudomonas septicemia associated with hemophagocytic syndrome and myelodysplasia in a Turkish child. Turk. J. Pediatr., 2005, 47(2), 191-194.
[163] Saleem, K.; Ayaz, B.; Shaikh, A. Histological grading patterns in patients of cutaneous leishmaniasis. J. Coll. Physicians Surg. Pak., 2007, 17(11), 650-653.
[164] Weiss, F.; Vogenthaler, N.; Franco-Paredes, C.; Parker, S.R. Leishmania tropica-induced cutaneous and presumptive concomitant viscerotropic leishmaniasis with prolonged incubation. Arch. Dermatol., 2009, 145(9), 1023-1026.
[165] Akhavan, A.A.; Mirhendi, H.; Khamesipour, A.; Alimohammadian, M.H.; Rassi, Y.; Bates, P.; Kamhawi, S.; Valenzuela, J.G.; Arandian, M.H.; Abdoli, H.; Jalali-zand, N.; Jafari, R.; Shareghi, N.; Ghanei, M.; Yaghoobi-Ershadi, M.R. Leishmania species: detection and identification by nested PCR assay from skin samples of rodent reservoirs. Exp. Parasitol., 2010, 126(4), 552-556.
[166] Michel, G.; Ferrua, B.; Lang, T.; Maddugoda, M.P.; Munro, P.; Pomares, C.; Lemichez E.; Marty, P. Luciferase-expressing Leishmania infantum allows the monitoring of amastigote population size, in vivo, ex vivo and in vitro. PLoS Negl. Trop. Dis., 2011, 5(9), e1323. Available from: http://www.plosntds.org/article/info%3Adoi%2F10.1371%2Fjournal.pntd.0001323
[167] Lecoeur, H.; Buffet, P.; Morizot, G.; Goyard, S.; Guigon, G.; Milon, G.; Lang, T. Optimization of topical therapy for Leishmania major localized cutaneous leishmaniasis using a reliable C57BL/6 model. PLoS Negl. Trop. Dis., 2007, 1(2), e34.
[168] Filipe-Santos, O.; Pescher, P.; Breart, B.; Lippuner, C.; Aebischer, T.; Glaichenhaus, N.; Spath, G.F.; Bousso, P. A dynamic map of antigen recognition by CD4 T cells at the site of Leishmania major infection. Cell Host Microbe, 2009, 6(1), 23-33.
[169] Courret, N.; Frehel, C.; Gouhier, N.; Pouchelet, M.; Prina, E.; Roux, P.; Antoine, J.C. Biogenesis of Leishmania-harbouring parasitophorous vacuoles following phagocytosis of the metacyclic promastigote or amastigote stages of the parasites. J. Cell Sci., 2002, 115(11), 2303-2316.
[170] Shahbazi, F.; Shahabi, S.; Kazemi, B.; Mohebali, M.; Abadi, A.R.; Zare, Z. Evaluation of PCR assay in diagnosis and identification of cutaneous leishmaniasis: a comparison with the parasitological methods. Parasitol. Res., 2008, 103(5), 1159-1162.
[171] Spanakos, G.; Patsoula, E.; Kremastinou, T.; Saroglou, G.; Vakalis, N. Development of a PCR-based method for diagnosis of Leishmania in blood samples. Mol. Cell. Probes, 2002, 16(6), 415-420.
[172] Titus, R.G.; Marchand, M.; Boon, T.; Louis, J.A. A limiting dilution assay for quantifying Leishmania major in tissues of infected mice. Parasite Immunol., 1985, 7(5), 545-555.
[173] Lima, H.C.; Bleyenberg, J.A.; Titus, R.G., A simple method for quantifying Leishmania in tissues of infected animals. Parasitol. Today, 1997, 13(2), 80-82.
[174] Boggild, A.K.; Miranda-Verastegui, C.; Espinosa, D.; Arevalo, J.; Martinez-Medina, D.; Llanos-Cuentas, A.; Low, D.E. Optimization of microculture and evaluation of miniculture for the isolation of Leishmania parasites from cutaneous lesions in Peru. Am. J. Trop. Med. Hyg., 2008, 79(6), 847-852.
[175] Rioux, J.A.; Lanotte, G.; Serres, E.; Pratlong, F.; Bastien, P.; Perieres, J. Taxonomy of Leishmania. Use of isoenzymes. Suggestions for a new classification. Ann. Parasitol. Hum. Comp., 1990, 65(3), 111-125.
[176] Mauricio, I.L.; Yeo, M.; Baghaei, M.; Doto, D.; Pratlong, F.; Zemanova, E.; Dedet, J.P.; Lukes, J.; Miles, M.A. Towards multilocus sequence typing of the Leishmania donovani complex: resolving genotypes and haplotypes for five polymorphic metabolic enzymes (ASAT, GPI, NH1, NH2, PGD). Int. J. Parasitol., 2006, 36(7), 757-769.
[177] Pratlong, F.; Dereure, J.; Ravel, C.; Lami, P.; Balard, Y.; Serres, G.; Lanotte, G.; Rioux, J.A.; Dedet, J.P. Geographical distribution and epidemiological features of Old World cutaneous leishmaniasis foci, based on the isoenzyme analysis of 1048 strains. Trop. Med. Int. Health, 2009, 14(9), 1071-1085.
[178] Marco, J.D.; Bhutto, A.M.; Soomro, F.R.; Baloch, J.H.; Barroso, P.A.; Kato, H.; Uezato, H.; Katakura, K.; Korenaga, M.; Nonaka, S.; Hashiguchi, Y. Multilocus enzyme electrophoresis and cytochrome B gene sequencing-based identification of Leishmania isolates from different foci of cutaneous leishmaniasis in Pakistan. Am. J. Trop. Med. Hyg., 2006, 75(2), 261-266.
[179] Marco, J.D.; Uezato, H.; Mimori, T.; Barroso, P.A.; Korenaga, M.; Nonaka, S.; Basombrio, M.A.; Taranto, N.J.; Hashiguchi, Y. Are cytochrome B gene sequencing and polymorphism-specific polymerase chain reaction as reliable
as multilocus enzyme electrophoresis for identifying Leishmania spp. from Argentina? Am. J. Trop. Med. Hyg., 2006, 75(2), 256-260.
[180] Rodgers, M.R.; Popper, S.J.; Wirth, D.F. Amplification of kinetoplast DNA as a tool in the detection and diagnosis of Leishmania. Exp. Parasitol., 1990, 71(3), 267-275.
[181] Volpini, A.C.; Marques, M.J.; Lopes dos Santos, S.; Machado-Coelho, G.L.; Mayrink, W.; Romanha, A.J. Leishmania identification by PCR of Giemsa-stained lesion imprint slides stored for up to 36 years. Clin. Microbiol. Infect., 2006, 12(8), 815-818.
[182] Van Eys, G.J.; Schoone, G.J.; Kroon, N.C.; Ebeling, S.B. Sequence analysis of small subunit ribosomal RNA genes and its use for detection and identification of Leishmania parasites. Mol. Biochem. Parasitol., 1992, 51(1), 133-142.
[183] Espinosa, D.; Boggild, A.K.; Deborggraeve, S.; Laurent, T.; Valencia, C.; Pacheco, R.; Miranda-Verastegui, C.; Llanos-Cuentas, A.; Leclipteux, T.; Dujardin, J.C.; Buscher, P.; Arevalo, J. Leishmania OligoC-TesT as a simple, rapid, and standardized tool for molecular diagnosis of cutaneous leishmaniasis in Peru. J. Clin. Microbiol., 2009, 47(8), 2560-2563.
[184] Roelfsema, J.H.; Nozari, N.; Herremans, T.; Kortbeek, L.M.; Pinelli, E. Evaluation and improvement of two PCR targets in molecular typing of clinical samples of Leishmania patients. Exp. Parasitol., 2011, 127(1), 36-41.
[185] Katakura, K.; Kawazu, S.; Naya, T.; Nagakura, K.; Ito, M.; Aikawa, M.; Qu, J.Q.; Guan, L.R.; Zuo, X.P.; Chai, J.J.; Chang, K.P.; Matsumoto, Y. Diagnosis of kala-azar by nested PCR based on amplification of the Leishmania mini-exon gene. J. Clin. Microbiol., 1998, 36(8), 2173-2177.
[186] Nasereddin, A.; Bensoussan-Hermano, E.; Schonian, G.; Baneth, G.; Jaffe, C.L. Molecular diagnosis of Old World cutaneous leishmaniasis and species identification by use of a reverse line blot hybridization assay. J. Clin. Microbiol., 2008, 46(9), 2848-2855.
[187] Azmi, K.; Nasereddin, A.; Ereqat, S.; Schonian, G.; Abdeen, Z. Identification of Old World Leishmania species by PCR-RFLP of the 7 spliced leader RNA gene and reverse dot blot assay. Trop. Med. Int. Health, 2010, 15(8), 872-880.
[188] Tupperwar, N.; Vineeth, V.; Rath, S.; Vaidya, T. Development of a real-time polymerase chain reaction assay for the quantification of Leishmania species and the monitoring of systemic distribution of the pathogen. Diagn. Microbiol. Infect. Dis., 2008, 61(1), 23-30.
[189] Fraga, J.; Montalvo, A.M.; De Doncker, S.; Dujardin, J.C.; Van der Auwera, G. Phylogeny of Leishmania species based on the heat-shock protein 70 gene. Infect. Genet. Evol., 2010, 10(2), 238-245.
[190] Da Silva, L.A.; de Sousa Cdos, S.; da Graca, G.C.; Porrozzi, R.; Cupolillo, E. Sequence analysis and PCR-RFLP profiling of the hsp70 gene as a valuable tool for identifying Leishmania species associated with human leishmaniasis in Brazil. Infect. Genet. Evol., 2010, 10(1), 77-83.
[191] Croan, D.G.; Morrison, D.A.; Ellis, J.T. Evolution of the genus Leishmania revealed by comparison of DNA and RNA polymerase gene sequences. Mol. Biochem. Parasitol., 1997, 89(2), 149-159.
[192] Asato, Y.; Oshiro, M.; Myint, C.K.; Yamamoto, Y.; Kato, H.; Marco, J.D.; Mimori, T.; Gomez, E.A.; Hashiguchi, Y.; Uezato, H. Phylogenic analysis of the genus Leishmania by cytochrome b gene sequencing. Exp. Parasitol., 2009, 121(4), 352-361.
[193] Zemanova, E.; Jirku, M.; Mauricio, I.L.; Horak, A.; Miles, M.A.; Lukes, J. The Leishmania donovani complex: genotypes of five metabolic enzymes (ICD, ME, MPI, G6PDH, and FH), new targets for multilocus sequence typing. Int. J. Parasitol., 2007, 37(2), 149-160.
[194] Castilho, T.M.; Camargo, L.M.; McMahon-Pratt, D.; Shaw, J.J.; Floeter-Winter, L.M. A real-time polymerase chain reaction assay for the identification and quantification of American Leishmania species on the basis of glucose-6-phosphate dehydrogenase. Am. J. Trop. Med. Hyg., 2008, 78(1), 122-132.
[195] Oshaghi, M.A.; Ravasan, N.M.; Hide, M.; Javadian, E.A.; Rassi, Y.; Sedaghat, M.M.; Mohebali, M.; Hajjaran, H. Development of species-specific PCR and PCR-restriction fragment length polymorphism assays for L. infantum / L. donovani discrimination. Exp. Parasitol., 2009, 122(1), 61-65.
[196] Nicolas, L.; Prina, E.; Lang, T.; Milon, G. Real-time PCR for detection and quantitation of Leishmania in mouse tissues. J. Clin. Microbiol., 2002, 40(5), 1666-1669.
[197] Bensoussan, E.; Nasereddin, A.; Jonas, F.; Schnur, L.F.; Jaffe, C.L. Comparison of PCR assays for diagnosis of cutaneous leishmaniasis. J. Clin. Microbiol., 2006, 44(4), 1435-1439.
[198] de Toledo, J.S.; Junqueira dos Santos, A.F.; Rodrigues de Moura, T.; Antoniazi, S.A.; Brodskyn, C.; Indiani de Oliveira, C.; Barral, A.; Cruz, A.K. Leishmania (Viannia) braziliensis transfectants overexpressing the miniexon gene lose virulence in vivo. Parasitol. Int., 2009, 58(1), 45-50.
[199] Mary, C.; Faraut, F.; Lascombe, L.; Dumon, H. Quantification of Leishmania infantum DNA by a real-time PCR assay with high sensitivity. J. Clin. Microbiol., 2004, 42(11), 5249-5255.
[200] Rolao, N.; Cortes, S.; Rodrigues, O.R.; Campino, L. Quantification of Leishmania infantum parasites in tissue biopsies by real-time polymerase chain reaction and polymerase chain reaction-enzyme-linked immunosorbent assay. J. Parasitol., 2004, 90(5), 1150-1154.
[201] Parvizi, P.; Baghban, N.; Novin, E.A.; Absavaran, A. Detection, identification and molecular typing of Leishmania major in Phlebotomus

1472 Current Medicinal Chemistry, 2012 Vol. 19, No. 10 Kobets et al.
papatasi from a focus of zoonotic cutaneous leishmaniasis in central of Iran. Exp. Parasitol., 2010, 124(2), 232-237.
[202] Gausterer, C.; Stein, C.; Stimpfl, T., Application of direct PCR in a forensic case of yew poisoning. Int. J. Legal Med., 2011, in press.
[203] Jung, S.J.; Bae, M.J.; Oh, M.J.; Lee, J. Sequence conservation in the internal transcribed spacers and 5.8S ribosomal RNA of parasitic scuticociliates Miamiensis avidus (Ciliophora, Scuticociliatia). Parasitol. Int., 2011, 60(2), 216-219.
[204] de Lima, V.M.; Fattori, K.R.; Michelin Ade, F.; da Silveira Neto, L.; Vasconcelos Rde, O. Comparison between ELISA using total antigen and immunochromatography with antigen rK39 in the diagnosis of canine visceral leishmaniasis. Vet. Parasitol., 2010, 173(3-4), 330-333.
[205] Pinero, J.; Martinez, E.; Pacheco, R.; Aragon, Z.; De Armas, F.; Del Castillo, A.; Valladares, B. PCR-ELISA for diagnosis of mucocutaneous leishmaniasis. Acta Trop., 1999, 73(1), 21-29.
[206] Martin-Sanchez, J.; Lopez-Lopez, M.C.; Acedo-Sanchez, C.; Castro-Fajardo, J.J.; Pineda, J.A.; Morillas-Marquez, F. Diagnosis of infections with Leishmania infantum using PCR-ELISA. Parasitology, 2001, 122(6), 607-615.
[207] Alborzi, A.; Pourabbas, B.; Shahian, F.; Mardaneh, J.; Pouladfar, G.R.; Ziyaeyan, M. Detection of Leishmania infantum kinetoplast DNA in the whole blood of asymptomatic individuals by PCR-ELISA and comparison with other infection markers in endemic areas, southern Iran. Am. J. Trop. Med. Hyg., 2008, 79(6), 839-842.
[208] Deborggraeve, S.; Laurent, T.; Espinosa, D.; Van der Auwera, G.; Mbuchi, M.; Wasunna, M.; El-Safi, S.; Al-Basheer, A.A.; Arevalo, J.; Miranda-Verastegui, C.; Leclipteux, T.; Mertens, P.; Dujardin, J.C.; Herdewijn, P.; Buscher, P. A simplified and standardized polymerase chain reaction format for the diagnosis of leishmaniasis. J. Infect. Dis., 2008, 198(10), 1565-1572.
[209] Fernandes, O.; Murthy, V.K.; Kurath, U.; Degrave, W.M.; Campbell, D.A. Mini-exon gene variation in human pathogenic Leishmania species. Mol. Biochem. Parasitol., 1994, 66(2), 261-271.
[210] Cupolillo, E.; Grimaldi Junior, G.; Momen, H.; Beverley, S.M. Intergenic region typing (IRT): a rapid molecular approach to the characterization and evolution of Leishmania. Mol. Biochem. Parasitol., 1995, 73(1-2), 145-155.
[211] Marfurt, J.; Niederwieser, I.; Makia, N.D.; Beck, H.P.; Felger, I. Diagnostic genotyping of Old and New World Leishmania species by PCR-RFLP. Diagn. Microbiol. Infect. Dis., 2003, 46(2), 115-124.
[212] Luyo-Acero, G.E.; Uezato, H.; Oshiro, M.; Takei, K.; Kariya, K.; Katakura, K.; Gomez-Landires, E.; Hashiguchi, Y.; Nonaka, S. Sequence variation of the cytochrome b gene of various human infecting members of the genus Leishmania and their phylogeny. Parasitology, 2004, 128(5), 483-491.
[213] Hide, M.; Banuls, A.L. Species-specific PCR assay for L. infantum / L. donovani discrimination. Acta Trop., 2006, 100(3), 241-245.
[214] Mimori, T.; Sasaki, J.; Nakata, M.; Gomez, E.A.; Uezato, H.; Nonaka, S.; Hashiguchi, Y.; Furuya, M.; Saya, H. Rapid identification of Leishmania species from formalin-fixed biopsy samples by polymorphism-specific polymerase chain reaction. Gene, 1998, 210(2), 179-186.
[215] Lima, V.M.; Goncalves, M.E.; Ikeda, F.A.; Luvizotto, M.C.; Feitosa, M.M. Anti-Leishmania antibodies in cerebrospinal fluid from dogs with visceral leishmaniasis. Braz. J. Med. Biol. Res., 2003, 36(4), 485-489.
[216] McMahon Pratt, D.; David, J.R. Monoclonal antibodies that distinguish between New World species of Leishmania. Nature, 1981, 291(5816), 581-583.
[217] Zeyrek, F.Y.; Korkmaz, M.; Ozbel, Y. Serodiagnosis of anthroponotic cutaneous leishmaniasis (ACL) caused by Leishmania tropica in Sanliurfa Province, Turkey, where ACL Is highly endemic. Clin. Vaccine Immunol., 2007, 14(11), 1409-1415.
[218] Bern, C.; Jha, S.N.; Joshi, A.B.; Thakur, G.D.; Bista, M.B. Use of the recombinant K39 dipstick test and the direct agglutination test in a setting endemic for visceral leishmaniasis in Nepal. Am. J. Trop. Med. Hyg., 2000, 63(3-4), 153-157.
[219] Vilaplana, C.; Blanco, S.; Dominguez, J.; Gimenez, M.; Ausina, V.; C, T.U.; Munoz, C. Noninvasive method for diagnosis of visceral leishmaniasis by a latex agglutination test for detection of antigens in urine samples. J. Clin. Microbiol., 2004, 42(4), 1853-1854.
[220] Montenegro, J. Cutaneous reaction in leishmaniasis. Arch. Dermatol. Syphilol., 1926, 13, 187-194.
[221] Sassi, A.; Louzir, H.; Ben Salah, A.; Mokni, M.; Ben Osman, A.; Dellagi, K. Leishmanin skin test lymphoproliferative responses and cytokine production after symptomatic or asymptomatic Leishmania major infection in Tunisia. Clin. Exp. Immunol., 1999, 116(1), 127-132.
[222] Gidwani, K.; Rai, M.; Chakravarty, J.; Boelaert, M.; Sundar, S. Evaluation of leishmanin skin test in Indian visceral leishmaniasis. Am. J. Trop. Med. Hyg., 2009, 80(4), 566-567.
[223] Bern, C.; Amann, J.; Haque, R.; Chowdhury, R.; Ali, M.; Kurkjian, K.M.; Vaz, L.; Wagatsuma, Y.; Breiman, R.F.; Secor, W.E.; Maguire, J.H. Loss of leishmanin skin test antigen sensitivity and potency in a longitudinal study of visceral leishmaniasis in Bangladesh. Am. J. Trop. Med. Hyg., 2006, 75(4), 744-748.
[224] Foulet, F.; Botterel, F.; Buffet, P.; Morizot, G.; Rivollet, D.; Deniau, M.; Pratlong, F.; Costa, J.M.; Bretagne, S. Detection and identification of Leishmania species from clinical specimens by using a real-time PCR assay and sequencing of the cytochrome B gene. J. Clin. Microbiol., 2007, 45(7), 2110-2115.
[225] Singh, D.P.; Goyal, R.K.; Singh, R.K.; Sundar, S.; Mohapatra, T.M. In search of an ideal test for diagnosis and prognosis of kala-azar. J. Health Popul. Nutr., 2010, 28(3), 281-285.
[226] Goto, H.; Lindoso, J.A. Current diagnosis and treatment of cutaneous and mucocutaneous leishmaniasis. Expert Rev. Anti Infect. Ther., 2010, 8(4), 419-433.
[227] Croft, S.L.; Seifert, K.; Yardley, V. Current scenario of drug development for leishmaniasis. Indian J. Med. Res., 2006, 123(3), 399-410.
[228] Mishra, J.; Saxena, A.; Singh, S. Chemotherapy of leishmaniasis: past, present and future. Curr. Med. Chem., 2007, 14(10), 1153-1169.
[229] Sundar, S.; Chatterjee, M. Visceral leishmaniasis - current therapeutic modalities. Indian J. Med. Res., 2006, 123(3), 345-352.
[230] Cruz, A.K.; de Toledo, J.S.; Falade, M.; Terrao, M.C.; Kamchonwongpaisan, S.; Kyle, D.E.; Uthaipibull, C. Current treatment and drug discovery against Leishmania spp. and Plasmodium spp.: a review. Curr. Drug Targets, 2009, 10(3), 178-192.
[231] Sundar, S.; Mehta, H.; Chhabra, A.; Singh, V.; Chauhan, V.; Desjeux, P.; Rai, M. Amphotericin B colloidal dispersion for the treatment of Indian visceral leishmaniasis. Clin. Infect. Dis., 2006, 42(5), 608-613.
[232] Berman, J.D.; Badaro, R.; Thakur, C.P.; Wasunna, K.M.; Behbehani, K.; Davidson, R.; Kuzoe, F.; Pang, L.; Weerasuriya, K.; Bryceson, A.D. Efficacy and safety of liposomal amphotericin B (AmBisome) for visceral leishmaniasis in endemic developing countries. Bull. World Health Organ., 1998, 76(1), 25-32.
[233] Dietze, R.; Carvalho, S.F.; Valli, L.C.; Berman, J.; Brewer, T.; Milhous, W.; Sanchez, J.; Schuster, B.; Grogl, M. Phase 2 trial of WR6026, an orally administered 8-aminoquinoline, in the treatment of visceral leishmaniasis caused by Leishmania chagasi. Am. J. Trop. Med. Hyg., 2001, 65(6), 685-689.
[234] Wasunna, M.K.; Rashid, J.R.; Mbui, J.; Kirigi, G.; Kinoti, D.; Lodenyo, H.; Felton, J.M.; Sabin, A.J.; Albert, M.J.; Horton, J. A phase II dose-increasing study of sitamaquine for the treatment of visceral leishmaniasis in Kenya. Am. J. Trop. Med. Hyg., 2005, 73(5), 871-876.
[235] Jha, T.K.; Sundar, S.; Thakur, C.P.; Felton, J.M.; Sabin, A.J.; Horton, J. A phase II dose-ranging study of sitamaquine for the treatment of visceral leishmaniasis in India. Am. J. Trop. Med. Hyg., 2005, 73(6), 1005-1011.
[236] Denton, H.; McGregor, J.C.; Coombs, G.H. Reduction of anti-leishmanial pentavalent antimonial drugs by a parasite-specific thiol-dependent reductase, TDR1. Biochem. J., 2004, 381(2), 405-412.
[237] Mottram, J.C.; Coombs, G.H. Leishmania mexicana: enzyme activities of amastigotes and promastigotes and their inhibition by antimonials and arsenicals. Exp. Parasitol., 1985, 59(2), 151-160.
[238] Berman, J.D.; Waddell, D.; Hanson, B.D. Biochemical mechanisms of the antileishmanial activity of sodium stibogluconate. Antimicrob. Agents Chemother., 1985, 27(6), 916-920.
[239] Berman, J.D.; Gallalee, J.V.; Best, J.M. Sodium stibogluconate (Pentostam) inhibition of glucose catabolism via the glycolytic pathway, and fatty acid beta-oxidation in Leishmania mexicana amastigotes. Biochem. Pharmacol., 1987, 36(2), 197-201.
[240] Wyllie, S.; Cunningham, M.L.; Fairlamb, A.H. Dual action of antimonial drugs on thiol redox metabolism in the human pathogen Leishmania donovani. J. Biol. Chem., 2004, 279(38), 39925-39932.
[241] Legare, D.; Papadopoulou, B.; Roy, G.; Mukhopadhyay, R.; Haimeur, A.; Dey, S.; Grondin, K.; Brochu, C.; Rosen, B.P.; Ouellette, M. Efflux systems and increased trypanothione levels in arsenite-resistant Leishmania. Exp. Parasitol., 1997, 87(3), 275-282.
[242] Mukhopadhyay, R.; Dey, S.; Xu, N.; Gage, D.; Lightbody, J.; Ouellette, M.; Rosen, B.P. Trypanothione overproduction and resistance to antimonials and arsenicals in Leishmania. Proc. Natl. Acad. Sci. U S A, 1996, 93(19), 10383-10387.
[243] Chakraborty, A.K.; Majumder, H.K. Mode of action of pentavalent antimonials: specific inhibition of type I DNA topoisomerase of Leishmania donovani. Biochem. Biophys. Res. Commun., 1988, 152(2), 605-611.
[244] Gonzalez, U.; Pinart, M.; Reveiz, L.; Alvar, J. Interventions for Old World cutaneous leishmaniasis. Cochrane Database Syst. Rev., 2008, 4, CD005067.
[245] Uzun, S.; Durdu, M.; Culha, G.; Allahverdiyev, A.M.; Memisoglu, H.R. Clinical features, epidemiology, and efficacy and safety of intralesional antimony treatment of cutaneous leishmaniasis: recent experience in Turkey. J. Parasitol., 2004, 90(4), 853-859.
[246] Firdous, R.; Yasinzai, M.; Ranja, K. Efficacy of glucantime in the treatment of Old World cutaneous leishmaniasis. Int. J. Dermatol., 2009, 48(7), 758-762.
[247] Bermudez, H.; Rojas, E.; Garcia, L.; Desjeux, P.; Dujardin, J.C.; Boelaert, M.; Chappuis, F. Generic sodium stibogluconate is as safe and effective as branded meglumine antimoniate, for the treatment of tegumentary leishmaniasis in Isiboro Secure Park, Bolivia. Ann. Trop. Med. Parasitol., 2006, 100(7), 591-600.
[248] Oliveira-Neto, M.P.; Schubach, A.; Mattos, M.; Goncalves-Costa, S.C.; Pirmez, C. A low-dose antimony treatment in 159 patients with American cutaneous leishmaniasis: extensive follow-up studies (up to 10 years). Am. J. Trop. Med. Hyg., 1997, 57(6), 651-655.
[249] Palacios, R.; Osorio, L.E.; Grajalew, L.F.; Ochoa, M.T. Treatment failure in children in a randomized clinical trial with 10 and 20 days of meglumine antimonate for cutaneous leishmaniasis due to Leishmania viannia species. Am. J. Trop. Med. Hyg., 2001, 64(3-4), 187-193.

Leishmaniasis: Prevention, Parasite Detection and Treatment Current Medicinal Chemistry, 2012 Vol. 19, No. 10 1473
[250] Maltezou, H.C. Drug resistance in visceral leishmaniasis. J. Biomed. Biotechnol., 2010, 2010, 617521.
[251] Olliaro, P.L.; Guerin, P.J.; Gerstl, S.; Haaskjold, A.A.; Rottingen, J.A.; Sundar, S. Treatment options for visceral leishmaniasis: a systematic review of clinical studies done in India, 1980-2004. Lancet Infect. Dis., 2005, 5(12), 763-774.
[252] Vanaerschot, M.; De Doncker, S.; Rijal, S.; Maes, L.; Dujardin, J.C.; Decuypere, S. Antimonial resistance in Leishmania donovani is associated with increased in vivo parasite burden. PLoS One, 2011, 6(8), e23120.
[253] Carter, K.C.; Hutchison, S.; Henriquez, F.L.; Legare, D.; Ouellette, M.; Roberts, C.W.; Mullen, A.B. Resistance of Leishmania donovani to sodium stibogluconate is related to the expression of host and parasite gamma-glutamylcysteine synthetase. Antimicrob. Agents Chemother., 2006, 50(1), 88-95.
[254] Downing, T.; Imamura, H.; Decuypere, S.; Clark, T.G.; Coombs, G.H.; Cotton, J.A.; Hilley, J.D.; de Doncker, S.; Maes, I.; Mottram, J.C.; Quail, M.A.; Rijal, S.; Sanders, M.; Schonian, G.; Stark, O.; Sundar, S.; Vanaerschot, M.; Hertz-Fowler, C.; Dujardin, J.C.; Berriman, M. Whole genome sequencing of multiple Leishmania donovani clinical isolates provides insights into population structure and mechanisms of drug resistance. Genome Res., 2011, 21(12), 2143-2156.
[255] Perry, M.R.; Wyllie, S.; Prajapati, V.K.; Feldmann, J.; Sundar, S.; Boelaert, M.; Fairlamb, A.H. Visceral leishmaniasis and arsenic: an ancient poison contributing to antimonial treatment failure in the Indian subcontinent? PLoS Negl. Trop. Dis., 2011, 5(9), e1227.
[256] Moore, E.M.; Lockwood, D.N. Treatment of visceral leishmaniasis. J. Glob. Infect. Dis., 2010, 2(2), 151-158.
[257] Alving, C.R.; Steck, E.A.; Chapman, W.L.Jr.; Waits, V.B.; Hendricks, L.D.; Swartz, G.M.Jr.; Hanson, W.L. Therapy of leishmaniasis: superior efficacies of liposome-encapsulated drugs. Proc. Natl. Acad. Sci. U S A, 1978, 75(6), 2959-63.
[258] Eriksson, U.; Seifert, B.; Schaffner, A. Comparison of effects of amphotericin B deoxycholate infused over 4 or 24 hours: randomised controlled trial. BMJ, 2001, 322(7286), 579-582.
[259] Hiemenz, J.W.; Walsh, T.J. Lipid formulations of amphotericin B: recent progress and future directions. Clin. Infect. Dis., 1996, 22(2), S133-144.
[260] Sundar, S.; Jha, T.K.; Thakur, C.P.; Mishra, M.; Singh, V.P.; Buffels, R. Single-dose liposomal amphotericin B in the treatment of visceral leishmaniasis in India: a multicenter study. Clin. Infect. Dis., 2003, 37(6), 800-804.
[261] Ritmeijer, K.; Ter Horst, R.; Chane, S.; Aderie, E.M.; Piening, T.; Collin, S.M.; Davidson, R.N. Limited effectiveness of high-dose liposomal amphotericin B (AmBisome) for treatment of visceral leishmaniasis in an Ethiopian population with high HIV prevalence. Clin. Infect. Dis., 2011, 53(12), e152-158.
[262] Report of the Fifth Consultative Meeting on Leishmania/HIV Coinfection; WHO Document Production Services: Geneva, Switzerland, 2007; p 24.
[263] Sundar, S.; Murray, H.W. Cure of antimony-unresponsive Indian visceral leishmaniasis with amphotericin B lipid complex. J. Infect. Dis., 1996, 173(3), 762-765.
[264] Moon, S.; Jambert, E.; Childs, M.; von Schoen-Angerer, T. A win-win solution?: A critical analysis of tiered pricing to improve access to medicines in developing countries. Global Health, 2011, 7(1), 39.
[265] Tuon, F.F.; Amato, V.S.; Graf, M.E.; Siqueira, A.M.; Nicodemo, A.C.; Amato Neto, V. Treatment of New World cutaneous leishmaniasis - a systematic review with a meta-analysis. Int. J. Dermatol., 2008, 47(2), 109-124.
[266] Verma, N.K.; Dey, C.S. Possible mechanism of miltefosine-mediated death of Leishmania donovani. Antimicrob. Agents Chemother., 2004, 48(8), 3010-3015.
[267] Luque-Ortega, J.R.; Rivas, L. Miltefosine (hexadecylphosphocholine) inhibits cytochrome c oxidase in Leishmania donovani promastigotes. Antimicrob. Agents Chemother., 2007, 51(4), 1327-1332.
[268] Lux, H.; Heise, N.; Klenner, T.; Hart, D.; Opperdoes, F.R. Ether-lipid (alkyl-phospholipid) metabolism and the mechanism of action of ether-lipid analogues in Leishmania. Mol. Biochem. Parasitol., 2000, 111(1), 1-14.
[269] Rakotomanga, M.; Blanc, S.; Gaudin, K.; Chaminade, P.; Loiseau, P.M. Miltefosine affects lipid metabolism in Leishmania donovani promastigotes. Antimicrob. Agents Chemother., 2007, 51(4), 1425-1430.
[270] Maly, K.; Uberall, F.; Schubert, C.; Kindler, E.; Stekar, J.; Brachwitz, H.; Grunicke, H.H. Interference of new alkylphospholipid analogues with mitogenic signal transduction. Anticancer Drug Des., 1995, 10(5), 411-425.
[271] Wieder, T.; Reutter, W.; Orfanos, C.E.; Geilen, C.C. Mechanisms of action of phospholipid analogs as anticancer compounds. Prog. Lipid Res., 1999, 38(3), 249-259.
[272] Sundar, S.; Jha, T.K.; Thakur, C.P.; Engel, J.; Sindermann, H.; Fischer, C.; Junge, K.; Bryceson, A.; Berman, J. Oral miltefosine for Indian visceral leishmaniasis. N. Engl. J. Med., 2002, 347(22), 1739-1746.
[273] Chawla, B.; Jhingran, A.; Panigrahi, A.; Stuart, K.D.; Madhubala, R. Paromomycin affects translation and vesicle-mediated trafficking as revealed by proteomics of paromomycin-susceptible-resistant Leishmania donovani. PLoS One, 2011, 6(10), e26660.
[274] Maarouf, M.; de Kouchkovsky, Y.; Brown, S.; Petit, P.X.; Robert-Gero, M. In vivo interference of paromomycin with mitochondrial activity of Leishmania. Exp. Cell Res., 1997, 232(2), 339-348.
[275] Maarouf, M.; Lawrence, F.; Croft, S.L.; Robert-Gero, M. Ribosomes of Leishmania are a target for the aminoglycosides. Parasitol. Res., 1995, 81(5), 421-425.
[276] Maarouf, M.; Lawrence, F.; Brown, S.; Robert-Gero, M. Biochemical alterations in paromomycin-treated Leishmania donovani promastigotes. Parasitol. Res., 1997, 83(2), 198-202.
[277] Sundar, S.; Jha, T.K.; Thakur, C.P.; Sinha, P.K.; Bhattacharya, S.K. Injectable paromomycin for visceral leishmaniasis in India. N. Engl. J. Med., 2007, 356(25), 2571-2581.
[278] El-On, J.; Halevy, S.; Grunwald, M.H.; Weinrauch, L. Topical treatment of Old World cutaneous leishmaniasis caused by Leishmania major: a double-blind control study. J. Am. Acad. Dermatol., 1992, 27(2), 227-231.
[279] Faghihi, G.; Tavakoli-kia, R. Treatment of cutaneous leishmaniasis with either topical paromomycin or intralesional meglumine antimoniate. Clin. Exp. Dermatol., 2003, 28(1), 13-16.
[280] Ozgoztasi, O.; Baydar, I. A randomized clinical trial of topical paromomycin versus oral ketoconazole for treating cutaneous leishmaniasis in Turkey. Int. J. Dermatol., 1997, 36(1), 61-63.
[281] Arana, B.A.; Mendoza, C.E.; Rizzo, N.R.; Kroeger, A. Randomized, controlled, double-blind trial of topical treatment of cutaneous leishmaniasis with paromomycin plus methylbenzethonium chloride ointment in Guatemala. Am. J. Trop. Med. Hyg., 2001, 65(5), 466-470.
[282] Neva, F.A.; Ponce, C.; Ponce, E.; Kreutzer, R.; Modabber, F.; Olliaro, P. Non-ulcerative cutaneous leishmaniasis in Honduras fails to respond to topical paromomycin. Trans. R. Soc. Trop. Med. Hyg., 1997, 91(4), 473-475.
[283] Pacher, P.; Nivorozhkin, A.; Szabo, C. Therapeutic effects of xanthine oxidase inhibitors: renaissance half a century after the discovery of allopurinol. Pharmacol. Rev., 2006, 58(1), 87-114.
[284] Kelly, J.M. Molecular mechanisms of pathogenesis in Chagas disease; Landes Bioscience, Eurekah.com, Kluwer Academic, Plenum Pub.: Georgetown, New York, 2003.
[285] Martinez, S.; Gonzalez, M.; Vernaza, M.E. Treatment of cutaneous leishmaniasis with allopurinol and stibogluconate. Clin. Infect. Dis., 1997, 24(2), 165-169.
[286] Velez, I.; Agudelo, S.; Hendrickx, E.; Puerta, J.; Grogl, M.; Modabber, F.; Berman, J. Inefficacy of allopurinol as monotherapy for Colombian cutaneous leishmaniasis. A randomized, controlled trial. Ann. Intern. Med., 1997, 126(3), 232-236.
[287] Cozzani, E.; Satta, R.; Fausti, V.; Cottoni, F.; Parodi, A. Cutaneous sporotrichoid leishmaniasis resistant to pentavalent antimonial therapy: complete resolution with itraconazole. Clin. Exp. Dermatol., 2010, 36(1), 49-51.
[288] Abadir, A.; Patel, A.; Haider, S. Systemic therapy of New World cutaneous leishmaniasis: A case report and review article. Can. J. Infect. Dis. Med. Microbiol., 2010, 21(2), e79-83: Available from: http://www.ncbi.nlm.nih. gov/pmc/articles/PMC2912102/pdf/idmm21e079.pdf
[289] Emad, M.; Hayati, F.; Fallahzadeh, M.K.; Namazi, M.R. Superior efficacy of oral fluconazole 400 mg daily versus oral fluconazole 200 mg daily in the treatment of cutaneous Leishmania major infection: a randomized clinical trial. J. Am. Acad. Dermatol., 2011, 64(3), 606-608.
[290] De Oliveira-Silva, F.; de Morais-Teixeira, E.; Rabello, A. Antileishmanial activity of azithromycin against Leishmania (Leishmania) amazonensis, Leishmania (Viannia) braziliensis, and Leishmania (Leishmania) chagasi. Am. J. Trop. Med. Hyg., 2008, 78(5), 745-749.
[291] Krolewiecki, A.J.; Romero, H.D.; Cajal, S.P.; Abraham, D.; Mimori, T.; Matsumoto, T.; Juarez, M.; Taranto, N.J. A randomized clinical trial comparing oral azithromycin and meglumine antimoniate for the treatment of American cutaneous leishmaniasis caused by Leishmania (Viannia) braziliensis. Am. J. Trop. Med. Hyg., 2007, 77(4), 640-646.
[292] Carvalho, L.; Luque-Ortega, J.R.; Lopez-Martin, C.; Castanys, S.; Rivas, L.; Gamarro, F. The 8-aminoquinoline analogue sitamaquine causes oxidative stress in Leishmania donovani promastigotes by targeting succinate dehydrogenase. Antimicrob. Agents Chemother., 2011, 55(9), 4204-4210.
[293] van Ingen, J.; Aarnoutse, R.E.; Donald, P.R.; Diacon, A.H.; Dawson, R.; Plemper van Balen, G.; Gillespie, S.H.; Boeree, M.J. Why do we use 600 mg of rifampicin in tuberculosis treatment? Clin. Infect. Dis., 2011, 52(9), e194-199.
[294] Kochar, D.K.; Aseri, S.; Sharma, B.V.; Bumb, R.A.; Mehta, R.D.; Purohit, S.K. The role of rifampicin in the management of cutaneous leishmaniasis. Q. J. M., 2000, 93(11), 733-737.
[295] Curti, C.; Laget, M.; Carle, A.O.; Gellis, A.; Vanelle, P. Rapid synthesis of sulfone derivatives as potential anti-infectious agents. Eur. J. Med. Chem., 2007, 42(6), 880-884.
[296] Dogra, J. A double-blind study on the efficacy of oral dapsone in cutaneous leishmaniasis. Trans. R. Soc. Trop. Med. Hyg., 1991, 85(2), 212-213.
[297] Osorio, L.E.; Palacios, R.; Chica, M.E.; Ochoa, M.T. Treatment of cutaneous leishmaniasis in Colombia with dapsone. Lancet, 1998, 351(9101), 498-499.
[298] Bucaretchi, F.; Vicente, D.C.; Pereira, R.M.; Tresoldi, A.T. Dapsone hypersensitivity syndrome in an adolescent during treatment during of leprosy. Rev. Inst. Med. Trop. Sao Paulo, 2004, 46(6), 331-334.
[299] van Griensven, J.; Balasegaram, M.; Meheus, F.; Alvar, J.; Lynen, L.; Boelaert, M. Combination therapy for visceral leishmaniasis. Lancet Infect. Dis., 2010, 10(3), 184-194.
[300] Sundar, S.; Sinha, P.K.; Rai, M.; Verma, D.K.; Nawin, K.; Alam, S.; Chakravarty, J.; Vaillant, M.; Verma, N.; Pandey, K.; Kumari, P.; Lal, C.S.;

1474 Current Medicinal Chemistry, 2012 Vol. 19, No. 10 Kobets et al.
Arora, R.; Sharma, B.; Ellis, S.; Strub-Wourgaft, N.; Balasegaram, M.; Olliaro, P.; Das, P.; Modabber, F. Comparison of short-course multidrug treatment with standard therapy for visceral leishmaniasis in India: an open-label, non-inferiority, randomised controlled trial. Lancet, 2011, 377(9764), 477-486.
[301] El-Sayed, M.; Anwar, A.E. Intralesional sodium stibogluconate alone or its combination with either intramuscular sodium stibogluconate or oral ketoconazole in the treatment of localized cutaneous leishmaniasis: a comparative study. J. Eur. Acad. Dermatol. Venereol., 2010, 24(3), 335-340.
[302] Llanos-Cuentas, A.; Echevarria, J.; Cruz, M.; La Rosa, A.; Campos, P.; Campos, M.; Franke, E.; Berman, J.; Modabber, F.; Marr, J. Efficacy of sodium stibogluconate alone and in combination with allopurinol for treatment of mucocutaneous leishmaniasis. Clin. Infect. Dis., 1997, 25(3), 677-684.
[303] Baptista, M.S.; Wainwright, M. Photodynamic antimicrobial chemotherapy (PACT) for the treatment of malaria, leishmaniasis and trypanosomiasis. Braz. J. Med. Biol. Res., 2011, 44(1), 1-10.
[304] Asilian, A.; Davami, M. Comparison between the efficacy of photodynamic therapy and topical paromomycin in the treatment of Old World cutaneous leishmaniasis: a placebo-controlled, randomized clinical trial. Clin. Exp. Dermatol., 2006, 31(5), 634-637.
[305] Kosaka, S.; Akilov, O.E.; O'Riordan, K.; Hasan, T. A mechanistic study of delta-aminolevulinic acid-based photodynamic therapy for cutaneous leishmaniasis. J. Invest. Dermatol., 2007, 127(6), 1546-1549.
[306] Reithinger, R.; Mohsen, M.; Wahid, M.; Bismullah, M.; Quinnell, R.J.; Davies, C.R.; Kolaczinski, J.; David, J.R. Efficacy of thermotherapy to treat cutaneous leishmaniasis caused by Leishmania tropica in Kabul, Afghanistan: a randomized, controlled trial. Clin. Infect. Dis., 2005, 40(8), 1148-1155.
[307] Lobo, I.M.; Soares, M.B.; Correia, T.M.; de Freitas, L.A.; Oliveira, M.I.; Nakatani, M.; Netto, E.; Badaro, R.; David, J.R. Heat therapy for cutaneous leishmaniasis elicits a systemic cytokine response similar to that of antimonial (Glucantime) therapy. Trans. R. Soc. Trop. Med. Hyg., 2006, 100(7), 642-649.
[308] Aronson, N.E.; Wortmann, G.W.; Byrne, W.R.; Howard, R.S.; Bernstein, W.B.; Marovich, M.A.; Polhemus, M.E.; Yoon, I.K.; Hummer, K.A.; Gasser, R.A., Jr.; Oster, C.N.; Benson, P.M. A randomized controlled trial of local heat therapy versus intravenous sodium stibogluconate for the treatment of cutaneous Leishmania major infection. PLoS Negl. Trop. Dis., 2010, 4(3), e628.
[309] Prasad, N.; Ghiya, B.C.; Bumb, R.A.; Kaushal, H.; Saboskar, A.A.; Lezama-Davila, C.M.; Salotra, P.; Satoskar, A.R. Heat, Oriental sore, and HIV. Lancet, 2011, 377(9765), 610.
[310] Chen, M.; Christensen, S.B.; Theander, T.G.; Kharazmi, A. Antileishmanial activity of licochalcone A in mice infected with Leishmania major and in hamsters infected with Leishmania donovani. Antimicrob. Agents Chemother., 1994, 38(6), 1339-1344.
[311] Torres-Santos, E.C.; Rodrigues, J.M., Jr.; Moreira, D.L.; Kaplan, M.A.; Rossi-Bergmann, B. Improvement of in vitro and in vivo antileishmanial activities of 2', 6'-dihydroxy-4'-methoxychalcone by entrapment in poly(D,L-lactide) nanoparticles. Antimicrob. Agents Chemother., 1999, 43(7), 1776-1778.
[312] Alves, L.V.; Temporal, R.M.; Cysne-Finkelstein, L.; Leon, L.L. Efficacy of a diarylheptanoid derivative against Leishmania amazonensis. Mem. Inst. Oswaldo Cruz, 2003, 98(4), 553-555.
[313] Barata, L.E.; Santos, L.S.; Ferri, P.H.; Phillipson, J.D.; Paine, A.; Croft, S.L. Anti-leishmanial activity of neolignans from Virola species and synthetic analogues. Phytochemistry, 2000, 55(6), 589-595.
[314] Fournet, A.; Ferreira, M.E.; Rojas de Arias, A.; Torres de Ortiz, S.; Inchausti, A.; Yaluff, G.; Quilhot, W.; Fernandez, E.; Hidalgo, M.E. Activity of compounds isolated from Chilean lichens against experimental cutaneous leishmaniasis. Comp. Biochem. Physiol. C. Pharmacol. Toxicol. Endocrinol., 1997, 116(1), 51-54.
[315] Nakayama, H.; Caballero, E.; Maldonado, M.; Torres, S.; E., S.; Sanabria, L.; Medarde, M.; Ferreira, M.-A.; de Arias, A.; San Feliciano, A. In vivo antileishmanial efficacy of combretastatin heteroanalogues. Rev. Latinoamer. Quim., 2009, 37(1), 56-64.
[316] Fournet, A.; Ferreira, M.E.; Rojas De Arias, A.; Torres De Ortiz, S.; Fuentes, S.; Nakayama, H.; Schinini, A.; Hocquemiller, R. In vivo efficacy of oral and intralesional administration of 2-substituted quinolines in experimental treatment of new world cutaneous leishmaniasis caused by Leishmania amazonensis. Antimicrob. Agents Chemother., 1996, 40(11), 2447-2451.
[317] Chakrabarti, G.; Basu, A.; Manna, P.P.; Mahato, S.B.; Mandal, N.B.; Bandyopadhyay, S. Indolylquinoline derivatives are cytotoxic to Leishmania donovani promastigotes and amastigotes in vitro and are effective in treating murine visceral leishmaniasis. J. Antimicrob. Chemother., 1999, 43(3), 359-366.
[318] Proulx, M.E.; Desormeaux, A.; Marquis, J.F.; Olivier, M.; Bergeron, M.G. Treatment of visceral leishmaniasis with sterically stabilized liposomes containing camptothecin. Antimicrob. Agents Chemother., 2001, 45(9), 2623-2627.
[319] Nakayama, H.; Loiseau, P.M.; Bories, C.; Torres de Ortiz, S.; Schinini, A.; Serna, E.; Rojas de Arias, A.; Fakhfakh, M.A.; Franck, X.; Figadere, B.; Hocquemiller, R.; Fournet, A. Efficacy of orally administered 2-substituted quinolines in experimental murine cutaneous and visceral leishmaniases. Antimicrob. Agents Chemother., 2005, 49(12), 4950-4956.
[320] Fournet, A.; Munoz, V.; Roblot, F.; Hocquemiller, R.; Cave, A.; Gantier, J.C. Antiprotozoal activity of dehydrozaluzanin C, a sesquiterpene lactone isolated from Munnozia maronii (asteraceae). Phytother. Res., 1993, 7(2), 111-115.
[321] Ma, Y.; Lu, D.M.; Lu, X.J.; Liao, L.; Hu, X.S. Activity of dihydroartemisinin against Leishmania donovani both in vitro and in vivo. Chin. Med. J. (Engl.), 2004, 117(8), 1271-1273.
[322] Arruda, D.C.; Miguel, D.C.; Yokoyama-Yasunaka, J.K.; Katzin, A.M.; Uliana, S.R. Inhibitory activity of limonene against Leishmania parasites in vitro and in vivo. Biomed. Pharmacother., 2009, 63(9), 643-649.
[323] Yardley, V.; Khan, A.A.; Martin, M.B.; Slifer, T.R.; Araujo, F.G.; Moreno, S.N.; Docampo, R.; Croft, S.L.; Oldfield, E. In vivo activities of farnesyl pyrophosphate synthase inhibitors against Leishmania donovani and Toxoplasma gondii. Antimicrob. Agents Chemother., 2002, 46(3), 929-931.
[324] Miguel, D.C.; Yokoyama-Yasunaka, J.K.; Uliana, S.R. Tamoxifen is effective in the treatment of Leishmania amazonensis infections in mice. PLoS Negl. Trop. Dis., 2008, 2(6), e249: Available from: http://www.plosntds.org/article/info%3Adoi%2F10.1371%2Fjournal.pntd.0000249
[325] Miguel, D.C.; Zauli-Nascimento, R.C.; Yokoyama-Yasunaka, J.K.; Katz, S.; Barbieri, C.L.; Uliana, S.R. Tamoxifen as a potential antileishmanial agent: efficacy in the treatment of Leishmania braziliensis and Leishmania chagasi infections. J. Antimicrob. Chemother., 2009, 63(2), 365-368.
[326] Karaji, A.G.; Hamzavi, Y. The opioid antagonist naloxone inhibits Leishmania major infection in BALB/c mice. Exp. Parasitol., 2012, 130(1), 73-77.
[327] Le Pape, P.; Abdala, H.; Pagniez, F.; Robert, J.M.; Le Baut, G. Activity of N-lutidinylcarboxamides against Leishmania donovani and L. braziliensis. Acta Parasitologica, 1999, 44(3), 156-159.
[328] Al-Abdely, H.M.; Graybill, J.R.; Loebenberg, D.; Melby, P.C. Efficacy of the triazole SCH 56592 against Leishmania amazonensis and Leishmania donovani in experimental murine cutaneous and visceral leishmaniases. Antimicrob. Agents Chemother., 1999, 43(12), 2910-2914.
[329] Bettiol, E.; Samanovic, M.; Murkin, A.S.; Raper, J.; Buckner, F.; Rodriguez, A. Identification of three classes of heteroaromatic compounds with activity against intracellular Trypanosoma cruzi by chemical library screening. PLoS Negl. Trop. Dis., 2009, 3(2), e384: Available from: http://www.plosntds.org/article/info%3Adoi%2F10.1371%2Fjournal.pntd.0000384
[330] Sharlow, E.R.; Close, D.; Shun, T.; Leimgruber, S.; Reed, R.; Mustata, G.; Wipf, P.; Johnson, J.; O'Neil, M.; Grogl, M.; Magill, A.J.; Lazo, J.S. Identification of potent chemotypes targeting Leishmania major using a high-throughput, low-stringency, computationally enhanced, small molecule screen. PLoS Negl. Trop. Dis., 2009, 3(11), e540: Available from: http://www.plosntds.org/article/info%3Adoi%2F10.1371%2Fjournal.pntd.0000540
[331] Walker, R.G.; Thomson, G.; Malone, K.; Nowicki, M.W.; Brown, E.; Blake, D.G.; Turner, N.J.; Walkinshaw, M.D.; Grant, K.M.; Mottram, J.C. High throughput screens yield small molecule inhibitors of Leishmania CRK3:CYC6 cyclin-dependent kinase. PLoS Negl. Trop. Dis., 2011, 5(4), e1033.
[332] Cleghorn, L.A.; Woodland, A.; Collie, I.T.; Torrie, L.S.; Norcross, N.; Luksch, T.; Mpamhanga, C.; Walker, R.G.; Mottram, J.C.; Brenk, R.; Frearson, J.A.; Gilbert, I.H.; Wyatt, P.G. Identification of inhibitors of the Leishmania cdc2-related protein kinase CRK3. ChemMedChem., 2011, 6(12), 2214-2224.
[333] Immordino, M.L.; Dosio, F.; Cattel, L. Stealth liposomes: review of the basic science, rationale, and clinical applications, existing and potential. Int. J. Nanomedicine. 2006, 1(3), 297-315.
[334] Tiuman, T.S.; Santos, A.O.; Ueda-Nakamura, T.; Filho, B.P.; Nakamura, C.V. Recent advances in leishmaniasis treatment. Int. J. Infect. Dis., 2011, 15(8), e525-532.
[335] Becquemont, L. Pharmacogenomics of adverse drug reactions: practical applications and perspectives. Pharmacogenomics, 2009, 10(6), 961-969.
[336] Davis, A.P.; Murphy, C.G.; Saraceni-Richards, C.A.; Rosenstein, M.C.; Wiegers, T.C.; Mattingly, C.J. Comparative Toxicogenomics Database: a knowledgebase and discovery tool for chemical-gene-disease networks. Nucleic Acids Res., 2009, 37, D786-792.
Received: October 03, 2011 Revised: December 29, 2011 Accepted: January 02, 2012





























