Embed Size (px)
Citation preview

GASTROENTEROLOGY 2002;122:1122-1132
Leukocyte Recruitment in Colon Cancer: Role of Cell Adhesion Molecules, Nitric Oxide, and Transforming Growth Factor 131
XAVIER BESSA,* J. IGNASI ELIZALDE,* FRANCESC MITJANS, t VIRGINIA PINOL,* ROSA MIQUEL,*
JULI~,N PANICS, * JAIME PIULATS,~ JOSEP M. PIQUIe, * and ANTONI CASTELLS* • Gastroenterology Department, Institut de Malalties Digestives, Hospital Clinic, Institut d'lnvestigacions Biomediques Agust Pi i Sunyer, University of Barcelona, Barcelona; and tLaboratorio de Bioinvestigaci6n, Merck Farina y Quimica, Barcelona, Catalonia, Spain
Background &Aims: A deficient leukocyte recruitment has been suggested in tumor vasculature, but little is known about the underlying molecular mechanism. To characterize leukocyte-endothelium interaction in experi- mental colon cancer, quantify the main endothelial cell adhesion molecules (CAMs), and evaluate the effect of tumor-derived products. Methods: Leukocyte recruitment was assessed by intravital videomicroscopy in mice bear- ing HT29-derived tumors. Endothelial CAMs were mea- sured using the dual-radiolabeled antibody technique. The role of molecules mediating leukocyte rolling (P-, E-, and L-selectin) or adhesion (intercellular adhesion molecule 1 [ICAM-1] and vascular cell adhesion molecule 1 [VCAM-1]) carcinoembryonic antigen (CEA), and transforming growth factor (TGF) I~1 was assessed through immunoblockade, whereas participation of nitric oxide (NO) and cyclooxygen- ase (COX) metabolites were evaluated by means of nonselective and selective inhibition. Results: Basal and lipopolysaccharide-stimulated leukocyte rolling and adhesion were markedly reduced in tumor vasculature. ICAM-1 immunoblockade prevented leukocyte adhesion in both tumor and nontumor microvessels. Neither baseline nor LPS-induced endothelial ICAM-1, P-, and E-selectin ex- pression in tumors were reduced with respect to nontumor vasculature. Although VCAM-1 expression was reduced in tumor endothelium, immunoneutralization of VCAM-1 failed to reverse LPS-induced leukocyte recruitment in this setting. CEA immunoblockade and COX inhibition did not modify the deficient leukocyte rolling. Nonselective NO inhibition partially reversed the defective adhesion re- sponse in tumor microvessels. Finally, TGF-I~I immuno- blockade partially and selectively restored impaired leuko- cyte rolling and adhesion in tumor microvessels. Conclusions: Impaired leukocyte recruitment in tumor vas- culature cannot be attributed to a depressed expression of the main CAMs. Selective restoration after NO inhibition and TGF-I~I immunoblockade suggests involvement of both molecules in this phenomenon.
T he prognosis of patients with colorectal cancer is determined by both local growth and metastatic
dissemination of the disease3 Among other factors, neo-
plastic progression depends on the ability of host de- fenses to reject cancer cells in an active process of immunologic surveillance2 In that regard, leukocyte- endothelium interactions in the tumor neovasculature have been proposed to play a prominent role in control- ling cancer growth because they are required for leuko- cytes to infiltrate tumor tissues, thereby exerting their antineoplastic activities)
Leukocyte recruitment in angiogenic blood microves- sels consists of a multistep cascade of finely regulated events, initiated by margination of flowing leukocytes from the central blood stream toward the vessel wall. 4 Thereafter, initial weak (low affinity) adhesive interac- tions, mediated by selectins, oq integrins, and their corresponding ligands, are established and manifested as leukocyte rolling phenomena. A proportion of rolling leukocytes become activated by diverse chemoattractants and mediators, resulting in a subsequent firm adhesion mediated by leukocyte integrins and several immuno- globulin (Ig) superfamily cell adhesion molecules (CAMs) expressed on the endothelial wall) Finally, leu- kocytes migrate to the interstitium, where they exert most of their antineoplastic actions.
This process of leukocyte recruitment has been well characterized in certain pathologic circumstances such as inflammatory bowel disease 6 and gastric Hdicobacterpylori infection, v In addition, these phenomena have been stud- ied in some experimental cancer models in which dimin- ished leukocyte rolling and adhesion in tumor vascula- ture have been observed. 8-1° Based on these studies, it has been hypothesized that the deficient leukocyte-endo- thelium interaction observed in angiogenic blood mi-
Abbreviations used in this paper: CAM, cell adhesion molecule; CEA, carcinoembryonic antigen; COX, cyclooxygenase; ICAM-1, intercellular adhesion molecule 1; L-NAME, NG-nitro-L-arginine methylester; LPS, lipopolysaccharide; mAb, monoclonal antibody; TGF-I~I, transforming growth factor I~1; VCAM-1, vascular cell adhesion molecule 1.
© 2002 by the American Gastroenterological Association 0016-5085/02/$35.00
doi:10.lO53/gast.2002.32369

April 2002 LEUKOCYTE RECRUITMENT IN COLORECTAL CANCER 1123
crovessels migh t be caused by down-regulat ion of certain
endothelial CAMs. Efforts to quantify the expression of
CAMs in t umor microvessels have ma!nly relied on im-
munosta in ing techniques, thus result ing in heteroge-
neous and sometimes conflicting results. .1-16 Technical
l imitations in quant i fying molecules wi th a high consti-
tut ive (e.g., intercellular adhesion molecule 1 [ ICAM-1])
or very low expression (e.g., vascular cell adhesion mol-
ecule 1 [VCAM-1]) may account for these discrepan-
cies. lva8 Nevertheless, these difficulties seem to have
been overcome by the development of a new technique
that uses radiolabeled monoclonal antibodies (mAbs)
against endothelial CAMs, thus allowing an accurate and
reliable quanti tat ive measurement in different vascular
beds, 6,7,19 including t umor microvessels. 2°
Unti l now, no study has been specifically addressed to
ascertain the precise molecular basis of leukocyte recruit-
ment in colon cancer. In the current investigation, leuko-
cyte-endothelium interactions in both tumor and nontumor
vasculature were analyzed by intravital fluorescence video-
microscopy in an animal model of heterotopic colon cancer,
with characterization of their molecular determinants by
means of selective immunoneutralization. Simultaneously,
expression of the main endothelial CAMs (i.e., E-selectin
and P-selectin, ICAM-1 and VCAM-1) was assessed using
the dual-radiolabeled m A b technique. Finally, the effect of
colorectal cancer-derived products involved in tumor pro-
gression (carcinoembryonic antigen [CEA], nitric oxide,
cyclooxygenase [ C O X ] metabolites, and transforming
growth factor [TGF] [31) with potential implication in
leukocyte recruitment has also been evaluated.
M a t e r i a l s a n d M e t h o d s
Animal Model of Heterotopic Colorectal Cancer
Human colon adenocarcinoma HT-29 cells (American Type Culture Collection, Manassas, VA) were cultured with RPMI 1640 medium plus 2 mmol/L Glutamax-I (Gibco BRL, Gaithersburg, MD) and 10% fetal calf serum in a humidified incubator (37°C, 6% CO2). Cells in exponential growth were detached with trypsin-EDTA and washed twice with phos- phate-buffered saline. Cell number and viability were assessed, and only cells with viability >90% were used. A total of 2 × 10 6 cells in 100 ~L phosphate-buffered saline was injected subcutaneously into the backs of 6-week-old Swiss nude mice. After 4 weeks, animals developed an evident tumor bulk. Mice were housed in the animal facility of the University of Barce- lona with a 12-hour light-dark cycle. They had free access to tap water and food but were fasted for 24 hours before the experiments. Principles of laboratory animal care (NIH pub- lication no. 86-23, revised 1985) and the procedure guidelines
for animal experiments from the Generalitat de Catalunya were followed.
mAbs
To assess the role of CAMs in leukocyte recruitment, the following mAbs were used: RMP-1, a murine IgG2a directed against mouse P-selectin21; MEL-14, a rat IgG1 di- rected against mouse L-selectin22; RME-1, a murine IgG1 directed against mouse E-selectin23; MK1.9.1, a rat lgG1 directed against mouse VCAM-124; YN-1, a rat IgG2b di- rected against mouse ICAM-125; and 3C4, a rat IgG2a directed against mouse ICAM-2. 26 MEL-14 was generously provided by Dr. P. Engel (Hospital Clfnic, Barcelona, Spain), RMP-1, RME-1, YN-1, and MK1.9.1 were scaled up and purified at Pharmacia & Upjohn Laboratories (Kalamazoo, MI), and 3C4 was purchased from PharMingen (San Diego, CA).
In Vivo Assessment of Leukocyte- Endothelium Interactions
Intravital fluorescence microscopy. Leukocyte- endothelium interactions were assessed by means of intravital fluorescence microscopy, both at baseline and 4 hours after intraperitoneal (IP) administration of lipopolysaccharide (LPS, 1 mg/kg; Salmonella abortus equi LPS; Sigma Chemical Co., St. Louis, MO) (5-6 animals per group). A detailed protocol has been published previously. 7 To assess tumor microcirculatory events, a careful skin dissection was made, creating a skin flap preserving the afferent and efferent blood supply to the tumor mass. To analyze leukocyte-endothelium interactions in co- lonic microvessels, a midline abdominal incision was made and the colon was carefully externalized. In both types of experi- ments, mice were positioned on a 20 × 30-cm Plexiglas board in a manner that allowed the inner surface of the skin or the bowel wall to be placed over a glass slide covering a 3.5 × 3.5-cm hole centered in the Plexiglas. This technical approx- imation allowed us to clearly differentiate tumor from peritu- mor and colonic microvessels. A schematic of the intravital microscopy system used in these experiments is shown in Figure 1.
After surgery, mice were allowed to stabilize for 10 minutes and then 0.05 mL of fluorescent dye acridine orange solution was administered. Images from 2-3 different venules with internal diameters of 30-35 gtm in each localization were selected and recorded for 1 minute each. Because of potential hazards of prolonged fluorescence emission on oxyradical gen- eration, illumination periods were restricted to 1 minute. 27 Venular diameter was measured online using a video caliper (Microcirculation Research Institute, Texas A&M University, College Station, TX). Rolling leukocytes were defined as those white blood cells that moved at a velocity less than that of free-flowing leukocytes in the same vessel. The flux of rolling leukocytes was measured as the number of rolling leukocytes that passed a fixed point within a small (10 ~m) viewing area of the vessel in a 30-second period. Leukocytes were considered adherent to venular endothelium when stationary for 30 sec- onds or longer and expressed as number per 100-1~m length of

1124 BESSAETAL. GASTROENTEROLOGY Vol. 122, No. 4
Acr~e °range s°7°n
// + ,. ~" " Externalized Intestine
Fluorescence Filter I I
FI~
EX. 420-490 nm ~ . . . . . . . . . Em. 520 nrn
High Resolution Monllor
" . . . . . . . . . . -2> Epl fluorescence
Figure 1. Schematic of the intravital microscopy system. In an anes- thetized animal, a portion of the intestine is exteriorized through a laparotomy (for tumor examination, a skin flap is created and the tumor placed over a glass-covered window in the microscopy stage). Light produced by a mercury lamp is projected to the excitation filter ( 420 -490 nm) and through the objective lens to the tissue surface. Reflected light is filtered by the emission filter (520 nm), and the image is digitized by the CCD camera. The video signal is recorded and later analyzed.
venule. Venular blood flow (Vbf) was estimated from the mean of the velocity of 3 free-flowing leukocytes (ffl), using the empirical relationship of Vbf = ffl/1.628 Venular wall shear rate (',/) was calculated assuming cylindrical geometry, using the Newtonian definition ~/ = 8(Vbf/diameter).29
Role of selectins in leukocyte ro l l i ng . Molecules involved in leukocyte roiling were assessed in additional groups of tumor-bearing mice (4 animals per group). After performing a baseline examination of leukocyte-endothelium interactions, blocking doses of anti-P-selectin (RMP-1, 100 Dg), anti-E-selectin (RME-1, 100 b~g), or anti-L-selectin (MEL-14, 80 b~g) alone or in combination were administered intravenously. Ten minutes later, leukocyte rolling was reas- sessed in the same venules.
Role of ICAM-1 and VCAM-1 in leukocyte adhe- sion. The contribution of ICAM-1 and VCAM-1 as mediators of leukocyte adhesion in angiogenic blood microvessels was analyzed by administration of the appropriate mAbs at block- ing doses in additional groups of tumor-bearing mice (4 animals per group). Fifteen minutes before the administration of LPS (1 mg/kg IP), a blocking dose of an anti-ICAM-1 (YN-1, 50 ~g) or anti-VCAM-1 (MK 1.9.1, 30 ~g) mAb was administered through the lateral tail vein. Four hours later, animals were anesthetized and a second dose of the correspond- ing mAb was injected intravenously before assessing leuko- cyte-endothelium interactions using the intravital microscopy technique.
Expression of Endothelial CAMs
Level of expression of the main endothelial CAMs (P-selectin, E-selectin, ICAM-1, and VCAM-1) was assessed using the dual-radiolabeled mAb technique) ° At baseline and after administration of LPS (4 hours), 5-7 animals per group were studied. Details of this technique have been described previously. 6,7,25,3° In brief, a mixture of binding mAbs (10 ~g
125I-YN-1 and 40 I-~g unlabeled YN-1 for ICAM-1, 10 b~g 125I-MK1.9.1 and 30 I*g unlabeled MK1.9.1 for VCAM-1, and a unique labeled dose of 10 b~g 125I-RMP-1 or 125I-RME-1 for P-selectin and E-selectin, respectively) with a nonbinding mAb to correct for the nonspecific accumulation (5 I*g 131I UPC-10; mouse IgG, Sigma Qufmica, Madrid, Spain) was administered through the femoral vein catheter. Antibodies were labeled using the iodogen method. A blood sample was obtained through the carotid artery catheter 5 minutes after injection of the mAb mixture. The animals were then hepa- rinized (1 mg/kg sodium heparin) and rapidly exsanguinated. Entire organs were harvested and weighed. 125I (binding mAb) and 1311 (nonbinding mAb) activities in different organs and in 50-1*L aliquots of cell-free plasma were counted in a gamma counter (Packard, Meridien, Australia) with automatic correc- tion for background activity and spillover. The accumulated activity of each mAb in an organ was expressed as nanograms of binding antibody per gram of tissue. 6
Because the expression of different CAMs is calculated as nanograms of mAb binding per gram of tissue and mAb accumulation is influenced by changes in endothelial surface area, we estimated the total endothelial surface area using the dual radiolabeled mAb technique for ICAM-2. ICAM-2 is constitutively expressed on the endothelial surface and, con- trary to ICAM-1, is not up-regulated in activated endothelial ceils; therefore, it is a reliable marker for endothelial surface area) 9,2° For ICAM-2, 10 I~g 125I-3C4 and 60 Dg unlabeled 3C4 were administered to each animal along with 5 I-~g 131I UPC-10 as a nonbinding mAb. For interorgan comparisons, values for endothelial CAMs were adjusted for ICAM-2 ex- pression according to the following formula: corrected mAb accumulation = ([ng Specific mAb/g Tissue] / [ n g ICAM-2/g Tissue]).
Effect of Colorectal Cancer Products Involved in Tumor Progression on Leukocyte Recruitment
CEA. Modulatory effects of CEA on leukocyte recruit- ment were established by immunoblockade using a polyclonal rabbit antibody against CEA (DAKO, Glostrup, Denmark). Previous in vitro experiments have shown the blocking ability of this antibody) 1 A pharmacokinetic study of this antibody was performed after IP administration of 2 different doses (5 and 10 mg/kg) of 125I-labeled antibody. Based on plasma concentrations, the dose selected for treatment was 5 mg/kg (calculated half-life, 15-18 hours; circulating levels, 6.5 ~g/mL at 6 hours). Accordingly, a blocking dose of 5 mg/kg was administered IP 24 and 4 hours before the experiment.
COX metabolites. The influence of COX metabolites on leukocyte recruitment was evaluated by both nonselective and selective COX-2 inhibition. For this purpose, a nonselec- tive COX inhibitor (indomethacin; Sigma Qufmica) (7.5 mg/kg subcutaneously) and a selective COX-2 inhibitor (SC- 236; Searle-Pharmacia, Skokie, IL) (2 mg/kg intravenously) were administered 1 hour before the experiment. The in vivo inhibitory effect of these doses was evaluated by measuring
April 2002 LEUKOCYTE RECRUITMENT IN COLORECTAL CANCER 1125
serum prostaglandin (PG) E 2 concentration in tumor-bearing mice (6 animals per group) treated with indomethacin and SC-236. The PGE2 concentration was measured by enzyme immunoassay (Amersham, Uppsala, Sweden) after extraction of PGE2 on Sep Pak C18 cartridges (Waters Associates, Milford, MA). From 0.5 to 1 mL of conditioned medium was acidified with 50% acetic acid (50 IxL) and applied to cartridges pre- activated with ethanol and distilled water. Cartridges were then washed with distilled water and petroleum benzine, and PGE2 was eluted with 4 mL methylformate. The eluted PGE2 was then dried and reconstituted for enzyme immunoassay. The recovery rate for the extraction procedure was 85%, as determined by the addition of synthetic PGE 2 to conditioned medium. The amounts of PGE2 were corrected by the total amount of protein in the corresponding samples. Whereas administration of indomethacin significantly decreased PGE2 production (baseline: 1171 + 267 pg/mL vs. indomethacin, 853 -+ 208 pg/mL, P = 0.02; LPS: 1461 + 312 pg/mL vs. indomethacin, 853 + 208 pg/mL, P = 0.001), no change was observed after administration of SC-236 (data not shown).
NO. NG-nitro-L-arginine methylester (L-NAME; Sigma Qufmica), a nonselective NO synthase inhibitor, was admin- istered through the tail vein 1 hour before the experiment. A dose-response curve was constructed with increasing doses of L-NAME (from 0.5 to 10 mg/kg). In a separate group of animals, involvement of the inducible NO synthase isoform was assessed by its selective inhibition with administration of aminoguanidine (Sigma Qufmica) (100 mg/kg intravenously)
1 hour before the experiment. LPS-induced adhesion was evaluated after NO inhibition.
For that purpose, 1 hour after administration of L-NAME (10 mg/kg intravenously), LPS (1 mg/kg IP) was injected. Leuko- cyte-endothelium interactions were assessed by intravital videomicroscopy 4 hours after administration of LPS.
T G F - ~ I . A blocking dose of an mAb against TGF-]31 (IgG1 mouse anti-human TGF-~I; Sigma Qufmica) (1 mg/kg IP) was administered 24 and 4 hours before the experiment. LPS-induced adhesion was evaluated after administration of TGF-[31. For that purpose, LPS (1 mg/kg IP) was injected 1 hour after administration of TGF-]31. Leukocyte-endothelium interactions were assessed by intravital videomicroscopy 4 hours after administration of LPS.
Immunohistochemistry for TGF-[31 was performed auto- matically using the TechMate 500 and EnVision System (DAKO, Carpinteria, CA). Briefly, 4-b~m dewaxed sections were treated with 0.05% saponin for 30 minutes at room temperature. After blocking endogenous peroxidase, sections were incubated with the primary antibody (TGF-[31; Santa Cruz Biotechnology, Santa Cruz, CA; 1:70 dilution) for 30 minutes at room temperature and washed in ChemMate buffer solution (DAKO). A peroxidase-labeled anti-rabbit polyclonal antibody was then applied for 30 minutes. Slides were incu- bated with diaminobenzidine substrate chromogen solution, washed in water, and counterstained with hematoxylin.
Statistical Methods
Data were analyzed by means of the Student t test and analysis of variance with the Bonferroni (post-hoc) test for parametric variables or the Mann-Whitney U test and the Kruskal-Wallis test for nonparametric variables. All values are reported as mean + standard deviation. Significance was con- sidered with an ot risk of 0.05.
R e s u l t s
Leukocyte-Endothelium Interactions in Tumor and Nontumor Vasculature
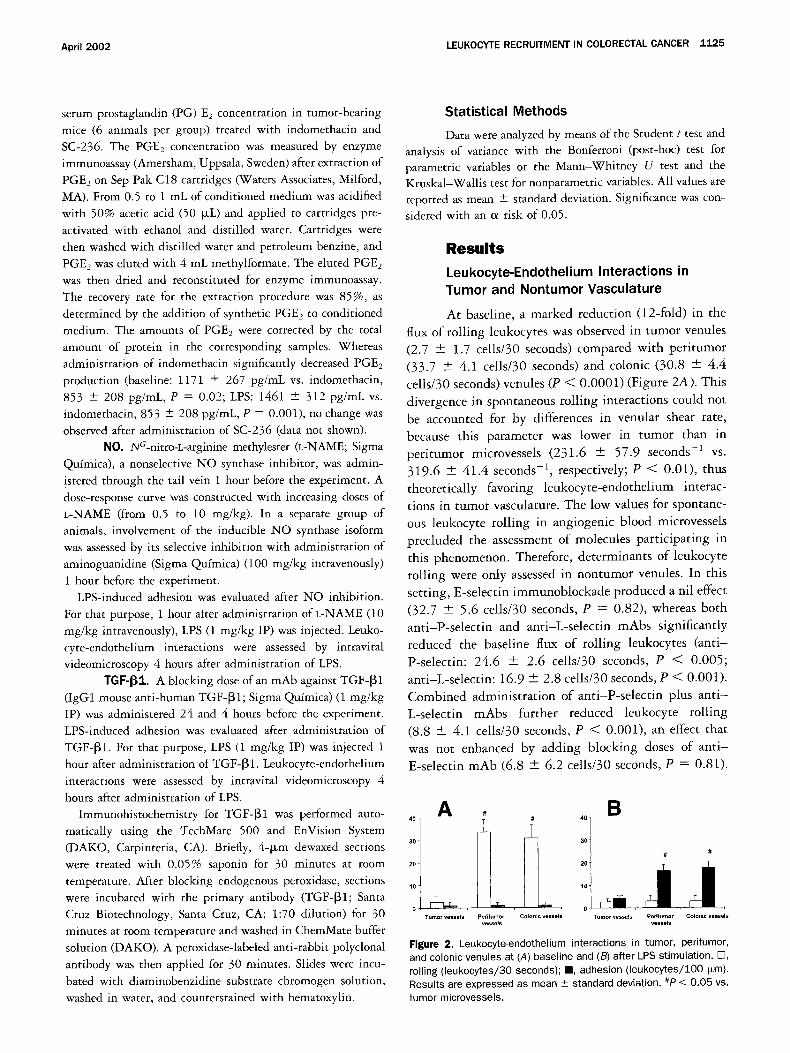
At baseline, a marked reduction (12-fold) in the
flux of roiling leukocytes was observed in tumor venules
(2.7 + 1.7 cells/30 seconds) compared with peritumor
(33.7 + 4.1 cells/30 seconds) and colonic (30.8 --- 4.4
cells/30 seconds)venules (P < 0.0001) (Figure 2A). This
divergence in spontaneous roiling interactions could not
be accounted for by differences in venular shear rate,
because this parameter was lower in tumor than in
peritumor microvessels (231.6 + 57.9 seconds -1 vs.
319.6 + 41.4 seconds -1, respectively; P < 0.01), thus
theoretically favoring leukocyte-endothelium interac-
tions in tumor vasculature. The low values for spontane- ous leukocyte rolling in angiogenic blood microvessels
precluded the assessment of molecules participating in
this phenomenon. Therefore, determinants of leukocyte
rolling were only assessed in nontumor venules. In this
setting, E-selectin immunoblockade produced a nil effect
(32.7 + 5.6 cells/30 seconds, P = 0.82), whereas both
anti-P-selectin and anti-L-selectin mAbs significantly
reduced the baseline flux of roiling leukocytes (anti-
P-selectin: 24.6 + 2.6 cells/30 seconds, P < 0.005;
anti-L-selectin: 16.9 + 2.8 cells/30 seconds, P < 0.001).
Combined administration of anti-P-selectin plus ant i -
L-selectin mAbs further reduced leukocyte rolling
(8.8 --- 4.1 cells/30 seconds, P < 0.001), an effect that
was not enhanced by adding blocking doses of ant i -
E-selectin mAb (6.8 + 6.2 cells/30 seconds, P = 0.81).
0 ~ o Tumor vessels Periturnor Colonic vessels Tumor vessels PerRumor Colonic vessels
v~sels vessels
Figure 2. Leukocyte-endothelium interactions in tumor, peritumor, and colonic venules at (A) baseline and (B) after LPS stimulation. E3, rolling (leukocytes/30 seconds); II, adhesion ( leukocytes/ lO0 i~m). Results are expressed as mean _+ standard deviation. #P < 0.05 vs. tumor microvessels.
1126 BESSA ET AL. GASTROENTEROLOGY Vol. 122, No. 4
A B
t 5
E
~5 ==
Basal LPS LPS LPS Basal LPS LPS LPS + + + +
ICAM*I VCAM- I fCAM-I VCAM-1
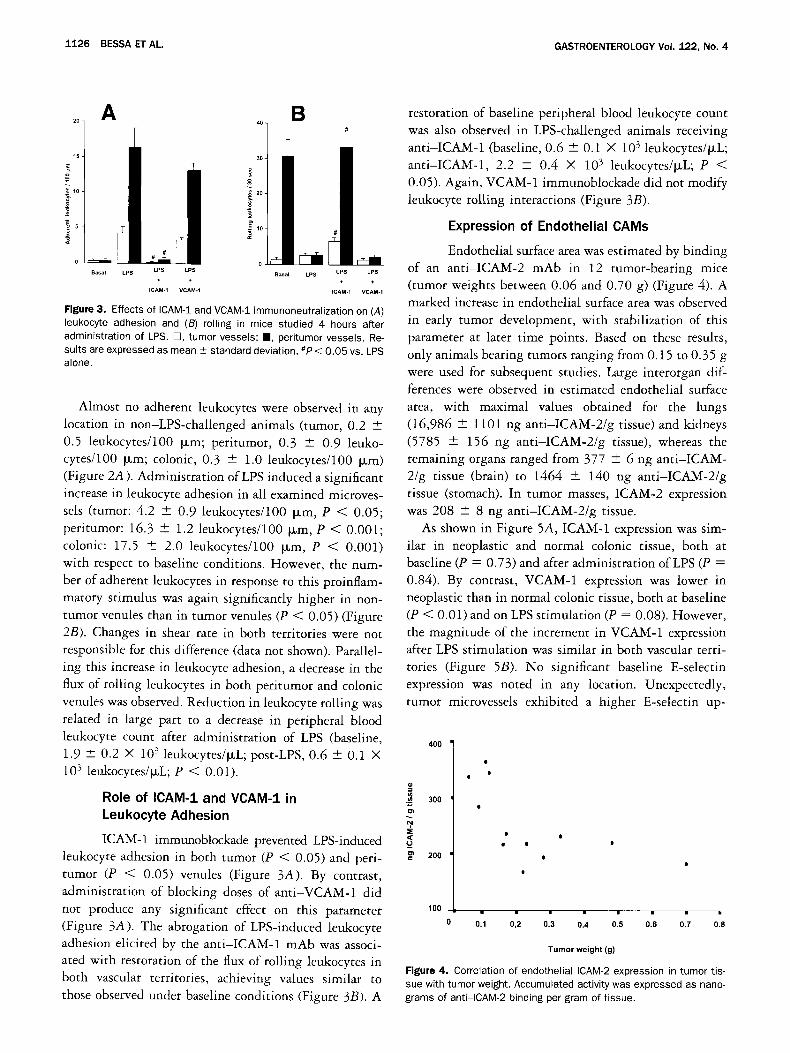
Figure 3. Effects of ICAM-1 and VCAM-1 immunoneutralization on (A) leukocyte adhesion and (B) rolling in mice studied 4 hours after administration of LPS. E3, tumor vessels; B, peritumor vessels. Re- suits are expressed as mean _+ standard deviation. #P < 0.05 vs. LPS alone.
Almost no adherent leukocytes were observed in any location in non-LPS-challenged animals (tumor, 0.2 ---
0.5 leukocytes/100 Dm; peritumor, 0.3 + 0.9 leuko- cytes/100 I~m; colonic, 0.3 -4- 1.0 leukocytes/100 b~m) (Figure 2A). Administration of LPS induced a significant increase in leukocyte adhesion in all examined microves- sels (tumor: 4.2 _+ 0.9 leukocytes/100 ~m, P < 0.05; peritumor: 16.3 + 1.2 leukocytes/100 b~m, P < 0.001; colonic: 17.5 + 2.0 leukocytes/lO0 Fum, P < 0.001) with respect to baseline conditions. However, the num- ber of adherent leukocytes in response to this proinflam- matory stimulus was again significantly higher in non- tumor venules than in tumor venules (P < 0.05) (Figure 2B). Changes in shear rate in both territories were not responsible for this difference (data not shown). Parallel- ing this increase in leukocyte adhesion, a decrease in the flux of rolling leukocytes in both peritumor and colonic venules was observed. Reduction in leukocyte roiling was related in large part to a decrease in peripheral blood leukocyte count after administration of LPS (baseline, 1.9 + 0.2 × 103 leukocytes/p~L; post-LPS, 0.6 -+ 0.1 × 103 leukocytes/btL; P < 0.01).
Role of ICAM-1 and VCAM-1 in Leukocyte Adhesion
ICAM-1 immunoblockade prevented LPS-induced leukocyte adhesion in both tumor (P < 0.05) and peri- tumor (P < 0.05) venules (Figure 3A). By contrast, administration of blocking doses of anti-VCAM-1 did
not produce any significant effect on this parameter (Figure 3A). The abrogation of LPS-induced leukocyte adhesion elicited by the anti-ICAM-1 mAb was associ- ated with restoration of the flux of rolling leukocytes in both vascular territories, achieving values similar to those observed under baseline conditions (Figure 3B). A
restoration of baseline peripheral blood leukocyte count was also observed in LPS-challenged animals receiving anti-ICAM-1 (baseline, 0.6 - 0.1 × 103 leukocytes/~L; anti-ICAM-1, 2.2 + 0.4 × 103 leukocytes/~L; P < 0.05). Again, VCAM-1 immunoblockade did not modify leukocyte rolling interactions (Figure 3B).
Expression of Endothelial CAMs
Endothelial surface area was estimated by binding of an anti-ICAM-2 mAb in 12 rumor-bearing mice (tumor weights between 0.06 and 0.70 g) (Figure 4). A marked increase in endothelial surface area was observed in early tumor development, with stabilization of this parameter at later time points. Based on these results, only animals bearing tumors ranging from 0.15 to 0.35 g were used for subsequent studies. Large interorgan dif- ferences were observed in estimated endothelial surface area, with maximal values obtained for the lungs (16,986 + 1101 ng anti-ICAM-2/g tissue) and kidneys (5785 -+ 156 ng anti-ICAM-2/g tissue), whereas the remaining organs ranged from 377 --- 6 ng anti-ICAM- 2/g tissue (brain) to 1464 + 140 ng anti-ICAM-2/g tissue (stomach). In tumor masses, ICAM-2 expression was 208 -+ 8 ng anti-ICAM-2/g tissue.
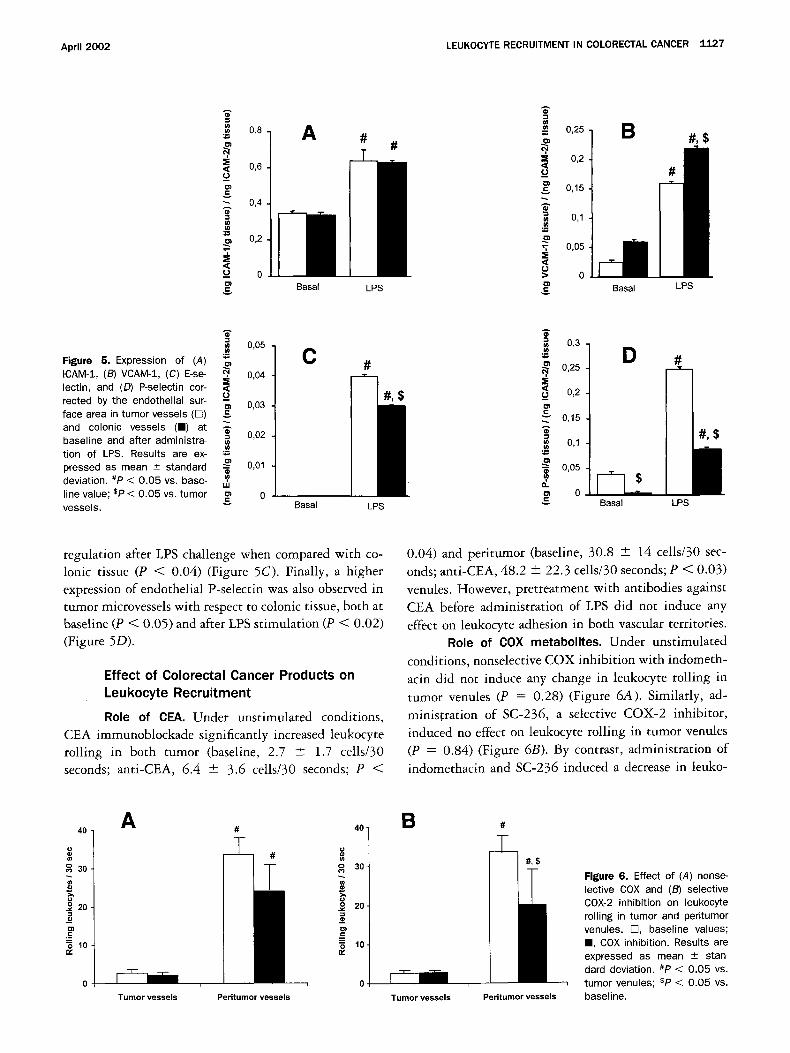
As shown in Figure 5A, ICAM-1 expression was sim- ilar in neoplastic and normal colonic tissue, both at baseline (P = 0.73) and after administration ofLPS (P = 0.84). By contrast, VCAM-1 expression was lower in neoplastic than in normal colonic tissue, both at baseline (P < 0.01) and on LPS stimulation (P = 0.08). However, the magnitude of the increment in VCAM-1 expression after LPS stimulation was similar in both vascular terri- tories (Figure 5B). No significant baseline E-selectin expression was noted in any location. Unexpectedly, tumor microvessels exhibited a higher E-selectin up-
400
== 300
:~ 200
100
• • II
0.1 0,2 0.3 0.4 0.5 0.6 0.7 0.8
Tumor weight ( g )
Figure 4. Correlation of endothelial ICAM-2 expression in tumor tis- sue with tumor weight. Accumulated activity was expressed as nano- grams of anti-ICAM-2 binding per gram of tissue.
April 2002 LEUKOCYTE RECRUITMENT IN COLORECTAL CANCER 1127
== 0,8 o~
< 0,6 _o
t ~ 0,4 g
m 0,2 "T =E
---- 0
A
Basal LPS
0,25
=i 0,2
0,15
0,1
"T 0,05
L~ > 0
Basal LPS
0,05 Figure 5. Expression of (A) ICAM-1, (B) VCAM-1, (C) E-se- ~, 0,04 lectin, and (D) P-selectin cor- rected by the endothelial sur- - 0,O3 face area in tumor vessels (rT) and colonic vessels (m) at ~.
0,02 baseline and after administra- tion of LPS. Results are ex-
0,01 pressed as mean _+ standard deviation. #P < 0.05 vs. base- ,,,, line value; sP < 0.05 vs. tumor ~ 0 vessels.
C
Basal LPS
"~ 0,3 u~
0,25
< __. 0,2
0,15
~ 0,1
0,05
a .
~ o Basal LPS
regulation after LPS challenge when compared with co- Ionic tissue (P < 0.04) (Figure 5C). Finally, a higher expression of endothelial P-selectin was also observed in tumor microvessels with respect to colonic tissue, both at baseline (P < 0.05) and after LPS stimulation (P < 0.02) (Figure 5D).
Ef fec t of C o l o r e c t a l C a n c e r P r o d u c t s on
L e u k o c y t e R e c r u i t m e n t
R o l e o f CEA. Under unstimulated conditions, CEA immunoblockade significantly increased leukocyte rolling in both tumor (baseline, 2.7 4- 1.7 cells/30 seconds; anti-CEA, 6.4 + 3.6 cells/30 seconds; P <
0.04) and peritumor (baseline, 30.8 + 14 cells/30 sec- onds; anti-CEA, 48.2 4- 22.3 cells/30 seconds; P < 0.03) venules. However, pretreatment with antibodies against CEA before administration of LPS did not induce any effect on leukocyte adhesion in both vascular territories.
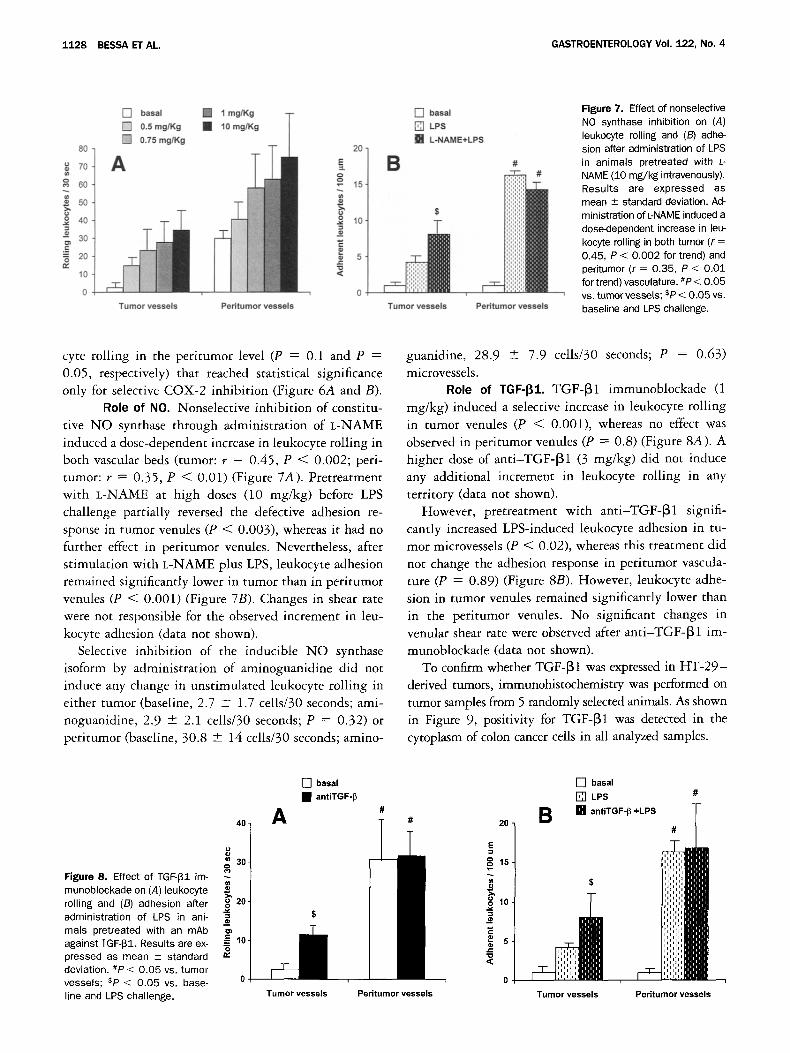
R o l e o f COX metabol i tes . Under unstimulated conditions, nonselective COX inhibition with indometh- acin did not induce any change in leukocyte rolling in tumor venules (P = 0.28) (Figure 6A). Similarly, ad- ministration of SC-236, a selective COX-2 inhibitor, induced no effect on leukocyte rolling in tumor venules (P = 0.84) (Figure 6B). By contrast, administration of indomethacin and SC-236 induced a decrease in leuko-
#
30
I 9 20 _== _== o 10 n/
A
S - T T T u m o r vessels
40
Peritumor vessels
40-
30
I o 20-
== o 10. n,
O.
a #
Tumor vessels
#,$
Peritumor vessels
Figure 6. Effect of (A) nonse- lective COX and (B) selective COX-2 inhibition on leukocyte rolling in tumor and peritumor venules. C], baseline values; B, COX inhibition. Results are expressed as mean --- stan- dard deviation. #P < 0.05 vs. tumor venules; sP < 0.05 vs. baseline.
1 1 2 8 BESSA ET AL. GASTROENTEROLOGY Vol. 122, No. 4
80
o 7o
60 (#
50 o~ 9 4o
- 30 == E 20 o
lO
[ ] basal [ ] 0.5 mg/Kg [ ] 0.75 mg/Kg
A
A ~ - t l . , _
Tumor vessels Peritumor vessels
20-
o o 15-
°~ 10- ] C
s
[ ] basal [ ] LPS [ ] L-NAME+LPS
B
$
Tumor vessels Peritumor vessels
Figure 7, Effect of nonselective NO synthase inhibition on (A) leukocyte rolling and (B) adhe- sion after administration of LPS in animals pretreated with L-
NAME (10 mg/kg intravenously). Results are expressed as mean +- standard deviation. Ad- ministration of L-NAME induced a dose-dependent increase in leu- kocyte rolling in both tumor (r = 0.45, P < 0.002 for trend) and peritumor (r = 0.35, P < 0.01 for trend) vasculature. #P < 0.05 vs. tumor vessels; sP< 0.05 vs. baseline and LPS challenge.
cyte rolling in the peritumor level (P = 0.1 and P = 0.05, respectively) that reached statistical significance only for selective COX-2 inhibition (Figure 6A and B).
Role of NO. Nonselective inhibition of constitu- tive NO synthase through administration of L-NAME induced a close-dependent increase in leukocyte rolling in both vascular beds (tumor: r = 0.45, P < 0.002; peri- tumor: r = 0.35, P < 0.01) (Figure 7A). Pretreatment with L-NAME at high doses (10 mg/kg) before LPS challenge partially reversed the defective adhesion re- sponse in tumor venules (P < 0.003), whereas it had no further effect in peritumor venules. Nevertheless, after stimulation with L-NAME plus LPS, leukocyte adhesion remained significantly lower in tumor than in peritumor venules (P < 0.001) (Figure 7B). Changes in shear rate were not responsible for the observed increment in leu- kocyte adhesion (data not shown).
Selective inhibition of the inducible NO synthase isoform by administration of aminoguanidine did not induce any change in unstimulated leukocyte rolling in either tumor (baseline, 2.7 + 1.7 cells/30 seconds; ami- noguanidine, 2.9 + 2.1 cells/30 seconds; P = 0.32) or peritumor (baseline, 30.8 + 14 cells/30 seconds; amino-
guanidine, 28.9 + 7.9 cells/30 seconds; P = 0.63) microvessels.
Role of TGF-[31. TGF-[31 immunoblockade (1 mg/kg) induced a selective increase in leukocyte rolling in tumor venules (P < 0.001), whereas no effect was observed in peritumor venules (P = 0.8) (Figure 8A). A higher dose of anti-TGF-[31 (3 mg/kg) did not induce any additional increment in leukocyte rolling in any territory (data not shown).
However, pretreatment with anti-TGF-[31 signifi- cantly increased LPS-induced leukocyte adhesion in tu- mor microvessels (P < 0.02), whereas this treatment did not change the adhesion response in peritumor vascula- ture (P = 0.89) (Figure 8B). However, leukocyte adhe- sion in tumor venules remained significantly lower than in the peritumor venules. No significant changes in venular shear rate were observed after anti-TGF-I31 im- munoblockade (data not shown).
To confirm whether TGF-[31 was expressed in HT-29- derived tumors, immunohistochemistry was performed on tumor samples from 5 randomly selected animals. As shown in Figure 9, positivity for TGF-[31 was detected in the cytoplasm of colon cancer cells in all analyzed samples.
Figure 8. Effect of TGF-I~I im- munoblockade on (A) leukocyte rolling and (B) adhesion after administration of LPS in ani- mals pretreated with an mAb against TGF-I~I. Results are ex- pressed as mean _+ standard deviation. #P < 0.05 vs. tumor vessels; *P < 0.05 vs, base- line and LPS challenge.
40
30- g
==
20 o _== == .__- 10 o
[ ] basal • antiTGF-~
A
Tumor vessels
#
i
Peritumor vessels
20
E
o ° 15 x--
C
8 5 "O ,<
[ ] basal
[ ] LPS #
Tumor vessels Peritumor vessels
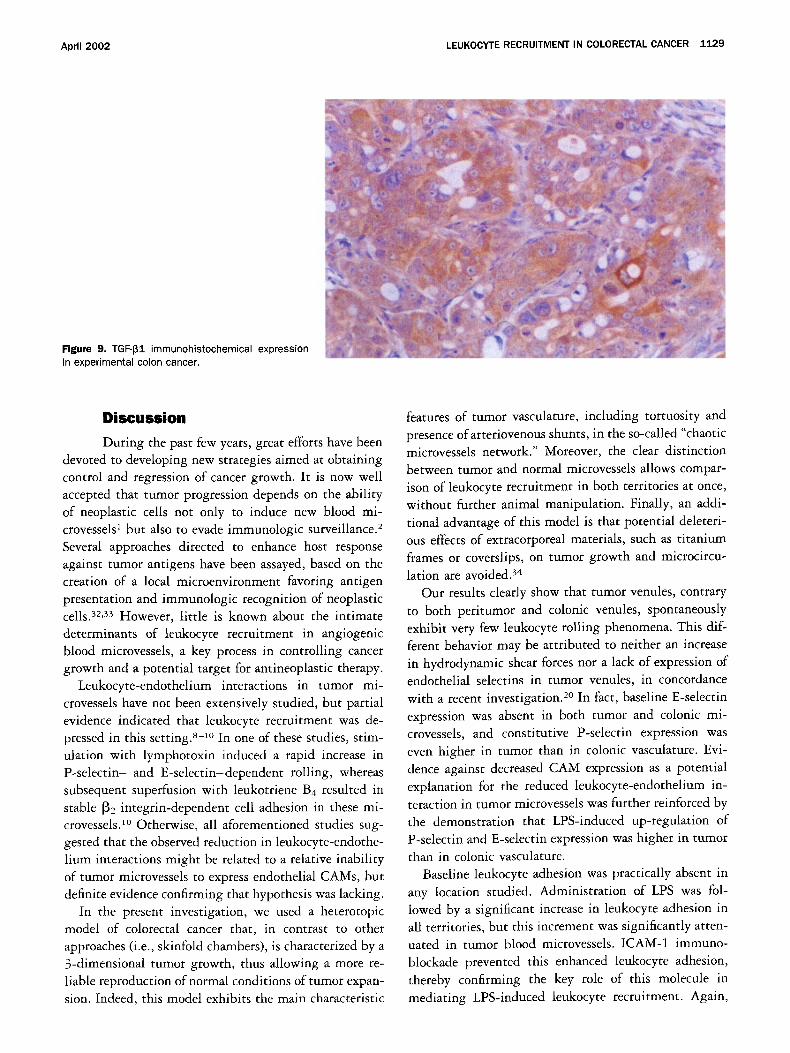
April 2002 LEUKOCYTE RECRUITMENT IN COLORECTAL CANCER 1129
Figure 9. TGF-I31 immunohistochemical expression in experimental colon cancer.
Discussion
During the past few years, great efforts have been devoted to developing new strategies aimed at obtaining control and regression of cancer growth. It is now well accepted that tumor progression depends on the ability of neoplastic cells not only to induce new blood mi- crovessels 1 but also to evade immunologic surveillance. 2 Several approaches directed to enhance host response against tumor antigens have been assayed, based on the creation of a local microenvironment favoring antigen presentation and immunologic recognition of neoplastic cells. 32,33 However, little is known about the intimate determinants of leukocyte recruitment in angiogenic blood microvessels, a key process in controlling cancer growth and a potential target for antineoplastic therapy.
Leukocyte-endothelium interactions in tumor mi- crovessels have not been extensively studied, but partial evidence indicated that leukocyte recruitment was de- pressed in this setting. *-1° In one of these studies, stim- ulation with lymphotoxin induced a rapid increase in P-selectin- and E-selectin-dependent rolling, whereas subsequent superfusion with leukotriene B4 resulted in stable ~2 integrin-dependent cell adhesion in these mi- crovessels) ° Otherwise, all aforementioned studies sug- gested that the observed reduction in leukocyte-endothe- lium interactions might be related to a relative inability of tumor microvessels to express endothelial CAMs, but definite evidence confirming that hypothesis was lacking.
In the present investigation, we used a heterotopic model of colorectal cancer that, in contrast to other approaches (i.e., skinfold chambers), is characterized by a 3-dimensional tumor growth, thus allowing a more re- liable reproduction of normal conditions of tumor expan- sion. Indeed, this model exhibits the main characteristic
features of tumor vasculature, including tortuosity and presence of arteriovenous shunts, in the so-called "chaotic microvessels network." Moreover, the clear distinction between tumor and normal microvessels allows compar- ison of leukocyte recruitment in both territories at once, without further animal manipulation. Finally, an addi- tional advantage of this model is that potential deleteri- ous effects of extracorporeal materials, such as titanium frames or coverslips, on tumor growth and microcircu- lation are avoided. 34
Our results clearly show that tumor venules, contrary to both peritumor and colonic venules, spontaneously exhibit very few leukocyte rolling phenomena. This dif- ferent behavior may be attributed to neither an increase in hydrodynamic shear forces nor a lack of expression of endothelial selectins in tumor venules, in concordance with a recent investigation. 2° In fact, baseline E-selectin expression was absent in both tumor and colonic mi- crovessels, and constitutive P-selectin expression was even higher in tumor than in colonic vasculature. Evi- dence against decreased CAM expression as a potential explanation for the reduced leukocyte-endothelium in- teraction in tumor microvessels was further reinforced by the demonstration that LPS-induced up-regulation of P-selectin and E-selectin expression was higher in tumor than in colonic vasculature.
Baseline leukocyte adhesion was practically absent in any location studied. Administration of LPS was fol- lowed by a significant increase in leukocyte adhesion in all territories, but this increment was significantly atten- uated in tumor blood microvessels. ICAM-1 immuno- blockade prevented this enhanced leukocyte adhesion, thereby confirming the key role of this molecule in mediating LPS-induced leukocyte recruitment. Again,

1130 BESSAETAL. GASTROENTEROLOGY Vol. 122, No. 4
demonstration of a similar expression of ICAM-1 in both
tumor and normal blood microvessels ruled out the possibility of a reduced CAM expression as a potential
determinant for the reduced leukocyte-endothelium in-
teractions existing in this experimental model of colon cancer.
VCAM-1 expression was slightly lower in tumor than in colonic microvessels, both at baseline and after LPS
stimulation. However, two observations suggest that
decreased expression of VCAM-1 is not the clue to explain the reduced leukocyte-endothelium interactions observed in tumor venules. First, VCAM-1 up-regulation
in tumor microvessels in response to LPS stimulation did not restore leukocyte adhesion to the levels observed in
colonic venules. Second and more important, VCAM-1 immunoneutralization had a null effect on leukocyte
recruitment after endoroxin challenge in both tumor and nontumor vessels.
Together, these results suggest that alternative mech- anisms should account for the impaired leukocyte-endo-
thelium interactions in the heterotopic model of colorec- tal cancer. In that sense, it was reasonable to hypothesize that tumor-derived products could be responsible for this
event. Among others, CEA, COX metabolites, NO, and
TGF-~I were potential candidates because they are in-
volved in both tumor progression and leukocyte recruit- ment.
CEA, an Ig family member, is constitutively expressed in intestinal epithelium and overexpressed in most colo- rectal cancers. Recent reports suggested that CEA-pro-
ducing cells may be selectively spared by cellular im-
mune defense mechanisms in tumor cell~lymphocyte interactions. Soluble CEA drastically inhibits the cyto- toxic activity of LAK cells against colon cancer cells in a dose-dependent manner. 35,36 Moreover, the use of CEA-
blocking antibodies stimulates lymphocyte adhesion to
HT-29 cells in v i t ro) * However, in our study, CEA immunoblockade induced a parallel, nonselective in- crease in leukocyte rolling interactions in both peritumor
and tumor microvessels, thus arguing against the partic- ipation of this molecule in the impaired tumor leukocyte
recruitment observed in colon cancer. Several lines of evidence support the involvement of
COX in colorectal cancer growth. 3v In addition, it has
been suggested that PGE2, a metabolite of this enzyme,
down-regulates HLA-DR expression, which may allow malignant cells to escape immunosurveillance) 8 Further-
more, intravital videomicroscopy studies have shown that indomethacin and aspirin promote leukocyte adhe- sion in postcapillary mesenteric venules) 9,4° However, both nonselective and selective COX-2 inhibition pro-
duced a nil effect on leukocyte rolling at tumor vascu- lature in the present investigation. It is important to
point out that, whereas administration of indomethacin
decreased serum PGE2 levels, treatment with SC-236 did not modify production of PGE> These results are in concordance with recent investigations showing that HT-29 cells express a low level of COX-2 protein. 41,42 In
that sense, it seems that the experimental model used in
the current study may not be adequate to evaluate the effect of COX-2 metabolites and, consequently, involve- ment of these products in the impaired leukocyte recruit-
ment observed in tumor vasculature cannot be defini-
tively ruled out. NO, a critical mediator in many biological processes
including cancer, 43-45 was also a potential candidate. In
a previous study, endogenous N O was found to be re- sponsible for a diminished leukocyte-endothelium inter- action. 46 However, high doses of a nonselective NO
synthase inhibitor were used in this investigation; there-
fore, changes in leukocyte recruitment may be caused by its hemodynamic effects at the systemic level. In the current study, nonselective N O inhibition before LPS
challenge partially reversed the defective adhesion re-
sponse in tumor microvessels without causing significant changes in shear rate, whereas it did not further increase
leukocyte adhesion in nontumor vasculature. Selective
inhibition of the inducible isoform did not induce any changes in any vascular territory. These facts suggest that N O coming from constitutive N O synthase participates
in leukocyte-endothelium interaction in several vascular
territories and is one of the mediators of the impaired
leukocyte recruitment observed in colon cancer. TGF-~I is a multifunctional polypeptide that pro-
motes angiogenesis and inhibits epithelial cell prolifera-
tion. Increased transcription of the TGF-~I gene was
shown in colorectal cancer-derived cell lines, and high levels of TGF-[31 have been observed in patients with this neoplasm. 47,48 This molecule has also been impli-
cated in tumor progression and metastasis in many ex- perimental cancer models. 49,5° However, TGF-[3 has
been reported to inhibit a variety of immune responses,
such as the ability of T lymphocytes to proliferate in response to cytokines, generation of tumor-specific cyto- toxic T lymphocytes, expression of interferon gamma
messenger RNA induced by interleukin 2, and tumor necrosis factor ot secretion. 49,51,52 Furthermore, TGF-[31
has been shown to decrease leukocyte adhesion both in vitro in cultured endothelial cells 53,54 and in vivo in an experimental model of gallbladder t umor ) 5 In the present study, TGF-[31 immunoblockade partially but selectively restores leukocyte rolling and adhesion in

April 2002 LEUKOCYTE RECRUITMENT IN COLORECTAL CANCER 1131
tumor vasculature. This observation suggests that TGF-~I is one of the factors that allow tumor cells to avoid immunosurveillance by circulating lymphocytes. The molecular mechanism mediating this phenomenon is unknown, in that sense, it has been suggested that TGF-~I inhibits E-selectin expression in cultured hu- man umbilical vein endothelial cells. 53 However, in our model, up-regulation of E-selectin expression in tumor microvessels after administration of LPS was identical to that observed in nontumor vasculature.
In summary, the results of the present study confirm a deficient leukocyte-endothelium interaction in colorectal cancer. This alteration could not be attributed to a depressed expression of the main endothelial CAMs in- volved in leukocyte recruitment, but rather to a func- tional blocking effect mediated at least in part by N O and TGF-~I . This observation opens attractive new ap- proaches to control tumor growth by favoring the inter- action of immune effector ceils with tumor endothelium.
References 1. Liota LA, Stetler-Stevenson W. Tumor invasion and metastasis:
an imbalance of positive and negative regulation. Cancer Res 1991;51:5054s-5059s.
2. Ferrara N. Natural killer cells, adhesion and tumor angiogenesis. Nature Med 1996;2:971-972.
3. Yu WG, Ogawa M, Mu J, Umehara K, Tsujimura T, Fujiwara H, Hamaoka T. IL-12 induced tumor regression correlates with in situ activity of IFN-~ produced by tumor infiltrating cells and its secondary induction of antitumor pathways. J Leuk Bio11997;62: 4 5 0 - 4 5 7 .
4. Pan6s J, Granger DN. Leukocyte-endothelial cell interactions: molecular mechanisms and implications in gastrointestinal dis- ease. Gastroenterology 1998;114 :1066-1090.
5. Albelda SM, Smith CW, Ward PA. Adhesion molecules and inflam- matory injury. FASEB J 1994;8 :504-512.
6. Sans M, Pan6s J, Ardite E, Elizalde JI, Arce Y, Flena M, Palacin A, Fern~ndez-Checa JC, Anderson DC, Lobb R, Piqu6 JM. VCAM-1 and ICAM-1 mediate leukocyte-endothelial cell adhesion in rat experimental colitis. Gastroenterology 1999 ;116 :874-883 .
7. Elizalde JI, G6mez J, Pan~s J, Lozano M, Casadevall M, Ramirez J, Pizcueta P, Marco F, Diaz de Rojas F, Granger DN, Piqu6 JM. Platelet activation in mice and human Helicobacter pylori infec- tion. J Clin Invest 1997;10 :996-1005.
8. Wu NZ, Klitzman B, Dodge R, Dewhirst MW. Diminished leuko- cyte-endothelial interaction in tumor microvessels. Cancer Res 1992;52 :4265-4628.
9. Fukumara D, Saleshi HA, Witwer B, Tuma RF, Melder R J, Jain RK. Tumor necrosis factor e-induced leukocyte adhesion in normal and tumor vessels: effect of tumor type, transplantation site, and host strain. Cancer Res 1995 ;55 :4824-4829 .
10. BorgstrSm P, Hughes GK, Hansell P, Wolitzky BA, Sriramarao P. Leukocyte adhesion in angiogenic blood vessels. Role of E-selec- tin, P-selectin, and [32 integrin in lymphotoxin-mediated leukocyte recruitment in tumor microvessels. J Clin Invest 1997 ;99 :2246- 2253.
11. Fox SB, Turner GDH, Gatter KC, Harris AL. The increased expres- sion of adhesion molecules ICAM-2, E- and P-selectins on breast cancer endothelium. J Pathol 1995;177:369-376.
12. Nelson H, Ramsey PS, Donohue JH, Wold LE. Cell adhesion molecule expression within the microvasculature of human colo-
rectal malignancies. Clin Immunol Immunopathol 1994 ;72 :129- 136.
13. Maurer CA, Friess H, Kretschmann B, Wildi S, M(~ller C, Graber H, Schilling M, B(Jchler MW. Over-expression of ICAM-1, VCAM-1 and ELAM-1 might influence tumor progression in colorectal cancer. Int J Cancer 1998 ;79 :76 -81 .
14. Piali L, Fichtel A, Terpe H J, Imhof BA, Gisler RH. Endothelial vascular cell adhesion molecule 1 expression is suppressed by melanoma and carcinoma. J Exp Med 1995;181:811-816.
15. Griffioen AW, Damen CA, Martinotti S, Blihjam GH, Groenewegen GC. Endothelial intercellular adhesion molecule-1 expression is suppressed in human malignancies: the role of angiogenic fac- tors. Cancer Res 1996;56:1111-1117.
16. Hellwig S, Damen C, van Adrichem N, Blijham G, Groenewegen G, Griffioen AW. Endothelial CD34 is suppressed in human malig- nancies: role of angiogenic factors. Cancer Lett 1997;120:203- 211.
17. Kuzu I, Bicknell R, Fletcher CD, Gatter KC. Expression of adhe- sion molecules on the endothelium of normal tissue vessels and vascular tumors. Lab Invest 1993;69:322-328.
18. Zocchi MR, Poggi A. Lymphocyte-endothelial cell adhesion mole- cules at the primary tumor site in human lung and renal cell carcinomas. J Natl Cancer Inst 1993;85 :246-247.
19. Vallien G, Langley R, Jennings S, Specian R, Granger DN. Expres- sion of endothelial cell adhesion molecules in neovascularized tissue. Microcirculation 2000;7 :249-258.
20. Langley RR, Russell J, Eppihimer M J, Alexander S J, Gerritsen M, Specian RD, Granger DN. Quantification of murine endothelial cell adhesion molecules in solid tumors. Am J Physiol 1999;277: 1156-1166 .
21. Walter UM, Ayer LM, Wolitzky BA, Wagner DD, Hynes RO, Man- ning AM, Issekutz AC. Characterization of a novel adhesion func- tion blocking monoclonal antibody to rat/mouse P-selectln gen- erated in the P-selectin-deficient mouse. Hybridoma 1997;16: 249 -257.
22. Ley K, Bullard DC, Arbones ML, Bosse R, Vestweber D, Tedder TF, Beaudet AL. Sequential contribution of L- and P-selectin to leukocyte rolling in vivo. J Exp Med 1995 ;181 :669-675 .
23. Walter UM, Ayer LM, Manning AM, Frenette PS, Wagner DD, Hynes RO, Wolitzky BA, Issekutz AC. Generation and character- ization of a novel adhesion function blocking monoclonal anti- body recognizing both rat and mouse E-selectin. Hybridoma 1997;16:355-361.
24. Henninger DD, Pan6s J, Russell J, Gerritsen M, Anderson DC, Granger DN. Cytokine-induced VCAM-1 and ICAM-1 expression in different organs in the mouse. J Immunol 1997;158: 1825-1832.
25. Komatsu S, Pan6s J, Grisham MB, Rusell JM, Mori N, Granger DN. Effects of intestinal stasis on intercellular cell adhesion molecule 1 expression in the rat: role of enteric bacteria. Gas- troenterology 1997;112:1971-1978.
26. Xu H, Bickford JK, Luther E, Carpenito C, Takei F, Springer TA. Characterization of murine intercellular adhesion molecule-2. J Immunol 1996 ;156 :4909-4914 .
27. Reed MWR, Miller FN. Importance of light dose in fluorescent microscopy. Microvasc Res 1988;36 :104-107.
28. Davis MJ. Determination of volumetric flow in capillary tubes using an optical doppler velocimeter. Microvasc Res 1987;34: 223-230.
29. Lipowsky HH, Kowalcheck S, Zweifach BW. The distribution of blood rheological parameters in the microvasculature of cat mes- entery. Circ Res 1978;43 :738-749.
30. Pan6s J, Perry MA, Anderson DC, Manning A, Leone B, Cepinskas G, Rosenbloom CL, Miyasaka M, Kvietys PR, Granger DN. Re- gional differences in constitutive and induced ICAM-1 expression in vivo. Am .I Physiol 1998;269:1955-1964.
31. Gr(Jngberg E, Eckert K, Maurer HR. Docetaxel treatment of HT-29

1132 BESSA ET AL. GASTROENTEROLOGY Vol. 122, No. 4
colon carcinoma cells reinforces the adhesion and immunocyto- toxicity of peripheral blood lymphocytes in vitro. Int J Onco11998; 12:957-963.
32. Ogawa M, Yu WG, Umehara K, Iwasaki M, Wijesuriya R, Tsujimura T, Kubo T, Fujiwara H, Hamaoka T. Multiple roles of interferon~ in the mediation of interleukin 12-induced tumor regression. Cancer Res 1998;58:2426-2432.
33. Coughlin CA, Salhany EH, Wysocka M, Aruga E, Kurzawa H, Ghang AE, Hunter CA, Fox JC, Trinchieri G, Lee W. Interleukin 12 and interleukin 18 synergistically induce murine tumor regression which involves inhibition of angiogenesis. J Clin Invest 1998; 101:1441-1452.
34. Lehr HA, Leuning M, Menger MD, Nolte D, Messmer K. Dorsal skinfold chamber technique for intravital microscopy in nude mice. Am J Pathol 1993;143:1055-1062.
35. Rivoltini L, Cattoretti G, Arienti F, Mastroianni A, Parmiani G. CEA and NCA expressed by colon carcinoma cells affect their interac- tion with and lisability by activated lymphocytes. Int J Biol Mark- ers 1992;7:143-147.
36. Kammerer R, Von Kleist S. CEA expression of colorectal adeno- carcinoma is correlated with their resistance against LAK-cell lysis. Int J Cancer 1994;57:341-347.
37. Taketo M. Cyclooxygenase-2 inhibitors in tumorigenesis. J Natl Cancer Inst 1998;90:1609-1620.
38. Arvind P, Papavassiliou ED, Tsioulais G J, Qiao L, Lovelace CIP, Duceman B, Rigas B. Prostaglandin E 2 down-regulates the ex- pression of HLA-DR antigen in human adenocarcinoma cell lines. Biochemistry 1995;34:5604-5609.
39. Asako H, Kubes P, Wallace J, Gaginella T, Wolf RE, Granger DN. Indomethacin-induced leukocyte adhesion in mesenteric venules: role of lipoxygenase products. Am J Physiol 1992;262:903-908.
40. Asako H, Kubes P, Wallace J, Wolf RE, Granger DN. Modulation of leukocyte adhesion in rat mesenteric venules by aspirin and salicylate. Gastroenterology 1992;103:146-152.
41. Shao J, Sheng H, Inoue H, Morrow JD, DuBois RN. Regulation of constitutive cyclooxygenase-2 expression in colon carcinoma cells. J Biol Chem 2000;275:33951-33956.
42. Hsi LC, Baek S J, Eling TE. Lack of cyclooxygenase-2 activity in HT-29 human colorectal carcinoma cells. Exp Cell Res 2000; 256:563-570.
43. Wink DA, Vodovotz Y, Laval J, Laval F, Dewhirst MW, Mitchell J. The multifaceted roles of nitric oxide in cancer. Carcinogenesis 1998;19:711-721.
44. Takahashi M, Fukuda K, Ohata T, Sugimura T, Wakabayashi K. Increased expression of inducible and endothelial constitutive nitric oxide synthases in rat colon colon tumors induced by azoxymethane. Cancer Res 1997;57:1233-1237.
45. Ambs S, Merriam WG, Bennett WP, Felley-Bosco E, Ogonfusika MO, Oser SM, Klein S, Shields PG, Billiar TR, Harris C. Frequent nitric oxide synthase-2 expression in human colon adenomas: implication for tumor angiogenesis and colon cancer progres- sion. Cancer Res 1998;58:334-341.
46. Fukumara D, Yuan F, Endo M, Jain RK. Role of nitric oxide in tumor microcirculation. Am J Pathol 1997;150:713-725.
47. Anzano MA, Rieman D, Prichett W, Bowen-Pope DF, Greig R. Growth factor production by human colon carcinoma cell lines. Cancer Res 1989;49:2898-2904.
48. Tsushima H, Kawata S, Tamura S, Ito N, Shirai Y, Kiso S, Imai Y, Shimomukai H, Nomura Y, Matsuda Y, Matsuzawa Y. High levels of transforming growth factor 131 in patients with colorectal can- cer: association with disease progression. Gastroenterology 1996;110:375-382.
49. Lagadec P, Reveneau S, Lejeune P, Pinard D, Borman T, Bauer J, Jeanin JF. Immunomodulator OM163-induced reversal of tumor- mediated immunosuppression and downregulation of TGF-~I in vivo. J Pharmacol Exp Ther 1996;278:926-933.
50. Mooradian DL, McCarthy JB, Komanduri KV, Furcht LT. Effects of transforming growth factor-l~l on human pulmonary adenocarci- noma cell adhesion, motility, and invasion in vitro. J Natl Cancer Inst 1992;84:523--527.
51. Fox FE, Ford HC, Douglas R, Cherian S, Nowell PC. Evidence that TGF-beta can inhibit human T-lymphocyte proliferation thorough paracrine and autocrine mechanisms. Cell Immunol 1993;150: 45-58.
52. Tada T, Ohzeki S, Utsumi K, Takiuchi H, Muramatsu M, Li XF, Shimizu J, Fujiwara H, Mamaoka T. Transforming growth factor- 13-induced inhibition of T cell function. J Immunol 1991;146: 1077-1082.
53. Gamble JR, Khew-Goodall Y, Vadas MA. Transforming growth factor-~ inhibits E-selectin expression on human endothelial cells. J Immunol 1993;150:4494-4503.
54. Gamble JR, Vadas MA. Endothelial adhesiveness for blood neu- trophiles is inhibited by transforming growth factor-~l. Science 1998;242:97-99.
55. Hojo M, Morimoto T, Maluccio M, Asanos T, Morimoto K, Lagman M, Shimbo T, Suthanthiran M. Cyclosporine induces cancer pro- gression by a cell-autonomous mechanism. Nature Med 1999; 5:1203-1208.
Received May 8, 2001. Accepted December 19, 2001. Address requests for reprints to: Antoni Castells, M.D., Gastroenter-
ology Department, Hospital Clinic, Villarroel 170, 08036 Barcelona, Catalonia, Spain. e-mail: [email protected]; fax: (34) 93-227- 93-87.
Supported by grants from the Ministerio de Ciencia y Tecnologia (SAFO0-O038), the Fondo de Investigaci6n Sanitaria (FIS 0 0 / 0 5 7 4 and 01/0104-02) , and from the Ag~ncia d'Avaluaci6 de Tecnologia M~dica of the Generalitat de Catalunya ( 2 / 6 / 9 6 and 026 /16 /00 ) . X.B. and V.P. received a research grant from the Hospital Clinic and from Institut d'lnvestigacions Biom~diques Agust Pi i Sunyer (IDIBAPS).
Presented in part at the 1999 and 2000 annual meetings of the American Gastroenterological Association in Orlando, Florida, and San Diego, California, respectively.
The authors thank Dr. Wladimiro Jim~nez, Dr. Antonio Soriano, Margarita Maynar, and Elena Gonzalvo for their technical support.





























