Embed Size (px)
Citation preview

0014-2980/98/1212-4257$17.50+.50/0© WILEY-VCH Verlag GmbH, D-69451 Weinheim, 1998
Linkage of a major quantitative trait locus to Yaagene-induced lupus-like nephritis in(NZW × C57BL/6)F1 mice
Marie-Laure Santiago1, Charles Mary1, Daniel Parzy2, Chantal Jacquet3, XavierMontagutelli4, R. Michael E. Parkhouse5, Robert Lemoine6, Shozo Izui7 and LucReininger1
1 INSERM U399, Universite de la Mediterranee, Marseille, France2 Institut de Medecine Tropicale du Service de Sante des Armees, Marseille, France3 INSERM U475, Montpellier, France4 Unite de Genetique des Mammiferes, Institut Pasteur, Paris, France5 Institute for Animal Health, Pirbright, GB6 Institut de Pathologie, Universite de Lausanne, Lausanne, Switzerland7 Department of Pathology, CMU, Geneva, Switzerland
In the present study, we mapped the major quantitative trait loci (QTL) differing between theNZW and C57BL/6 inbred strains of mice by making use of (NZW × C57BL/6.Yaa)F1 mice,a model in which the lupus-like autoimmune syndrome observed in male mice is associatedwith the presence of an as yet unidentified Y chromosome-linked autoimmune accelerationgene, Yaa. Linkage analysis of 126 C57BL/6 × (NZW × C57BL/6.Yaa)F1 backcross malesprovided evidence for a major QTL on chromosome 7 controlling both the severity of glomer-ulonephritis and the production of IgG anti-DNA autoantibody and retroviral gp70-anti-gp70immune complexes. Two additional QTL of C57BL/6 origin on chromosome 17 had noapparent individual effects, but showed strong epistatic interaction with chromosome 7 QTLfor disease severity and anti-DNA autoantibody production. Our data also identified on chro-mosome 13 a QTL of NZW origin with a major effect on the level of gp70, and showing anadditive effect with the chromosome 7 QTL on the level of gp70 immune complexes. Ourstudy thus provides a model to dissect the complex genetic interactions that result in mani-festations of murine lupus-like disease.
Key words: Systemic lupus erythematosus / Genetic / Autoantibody / Retroviral gp70
Received 3/8/98Revised 22/9/98Accepted 24/9/98
[I 18705]
Abbreviations: BC1: Backcross B6: C57BL/6 GN: Glo-merulonephritis IC: Immune complexes NZB: New Zea-land Black NZW: New Zealand White QTL: Quantitativetrait locus (loci) Yaa: Y-linked autoimmune acceleration
1 Introduction
Identification of specific genetic loci that contribute tosusceptibility to SLE in human has been hampered byseveral factors, including disease heterogeneity andcomplex interactions of environmental and genetic fac-tors [1, 2]. Murine SLE can be instrumental to studies inhuman [3]. Linkage analyses using microsatellites (orsimple sequence repeats) as markers recently performedin crosses involving the lupus-prone New Zealand Black(NZB) and New Zealand White (NZW) mice identified atleast 12 loci in suggestive or significant linkage with
lupus traits [4–8]. To date, narrowing the interval contain-ing NZB and NZW lupus susceptibility loci represents amajor goal toward identification of the genes them-selves. The incomplete association of lupus susceptibil-ity loci with disease traits and their interactions in athreshold manner add, however, to the complexity of thepolygenic inheritance of murine SLE [4, 6, 7, 9], andremain a major difficulty for the identification of genesresponsible for lupus susceptibility.
Single gene defects that lead to or accelerate autoim-mune disease include an as yet unidentified autoimmuneacceleration gene present on the Y chromosome derivedfrom BXSB mice, Yaa (Y-linked autoimmune accelera-tion) [10]. The Yaa gene has been shown to be responsi-ble for the acceleration of the lupus-like autoimmunesyndrome in inbred BXSB mice and in their F1 hybridswith NZB or NZW mice (for review see reference [11]).Noteworthy, F1 hybrids of NZW and C57BL/6 (B6) mice
Eur. J. Immunol. 1998. 28: 4257–4267 QTL linked with Yaa gene-induced lupus-like nephritis 4257

carrying the Yaa gene (B6.Yaa) have been shown todevelop severe lupus-like nephritis closely associatedwith elevated serum levels of IgG anti-DNA antibodiesand circulating retroviral gp70-anti-gp70 immune com-plexes (gp70 IC) [12]. In contrast to the lymphoprolifera-tion (lpr) mutation, which can induce autoantibodies in avariety of genetic backgrounds [13], the Yaa gene doesnot affect immunologically normal B6 mice. Since(NZW × B6)F1 males lacking the Yaa gene, like females,are essentially normal, the Yaa gene appears capable ofpotentiating an autosomal genome which is by itself notsufficient, so that mice develop a lupus-like syndrome.This strongly suggests that relatively small numbers ofcritical genes, such as NZW dominant susceptibilityalleles, may be sufficient to cause full expression of SLEin presence of the Yaa gene. Defining the effects andchromosomal position of NZW background genes con-tributing to lupus-like disease in (NZW × B6.Yaa)F1 micerepresents a model for dissecting the complex geneticinteractions that result in manifestations of murine SLE.We have used this approach in a B6 × (NZW × B6.Yaa)F1backcross study and report the mapping of a major QTLon chromosome 7 inherited from the NZW strain predis-posing to both glomerulonephritis (GN) and autoanti-body production. Results are discussed, along with theresults of other linkage studies involving the NZW strain,as evidence for interaction of the Yaa gene with a gene orgroup of closely linked genes that may play a key role incontrolling B cell abnormalities and mediating lupus sus-ceptibility.
2 Results
2.1 Inheritance of GN in a backcross of B6females with (NZW × B6.Yaa)F1 males
To determine the NZW genetic contribution to the Yaagene-induced acceleration of autoimmune disease in(NZW × B6.Yaa)F1 mice, a total of 126 B6 × (NZW ×B6.Yaa)F1 male backcross (BC1) mice were bred andexamined histologically for indications of renal diseasewhen animals were killed either moribund or at the end ofa 15-month observation period. The distribution of histo-logical grade of the 126 BC1 mice was clearly bimodal(Fig. 1 A), with 63 mice showing evidence of the severelethal form of GN (grade n 3). The 50 % disease inci-dence observed in these backcross mice is compatiblewith the hypothesis that a dominant allele of NZW originat a single locus is controlling the severity of GN in pres-ence of the Yaa gene, although the data may also reflectthe interaction of multiple threshold genetic influencesthat combine to produce a polygenic trait [4, 6, 7].
2.2 Distribution of serum levels of IgG anti-DNAantibodies, retroviral gp70 IC and gp70antigen
Beginning at 2 months of age, the 126 BC1 mice wereserially bled and the sera were analyzed for IgG anti-DNAantibodies and retroviral gp70 IC. Results presented inFig. 1B and C show that among 6–8-month-old parentalcontrol mice, none of B6.Yaa, but nearly all (NZW ×B6.Yaa)F1 male mice developed markedly elevated IgGanti-DNA antibodies (19 ± 8 U/ml vs. 125 ± 45 U/ml,respectively) or retroviral gp70 IC (0.3 ± 0.2 ? g/ml vs.4.7 ± 3.5 ? g/ml, respectively), indicating that these auto-antibodies could be used to discriminate parental phe-notypes among BC1 mice. The cumulative incidence ofBC1 mice producing high titers of anti-DNA antibodiesgreater than 3 SD above the mean of B6.Yaa males(43 U/ml) increased from 16 % (20/126) at 2 months to50 % (63/126) at 8 months of age, and did not changethereafter (data not shown). For retroviral gp70 IC, againapproximately half of the BC1 mice (66/126), determinedat 6 months of age, were found to develop high levels ofgp70 IC greater than 3 SD above the mean of B6.Yaamales (0.9 ? g/ml).
It has been previously shown that in contrast to NZWmice which produce large amounts of serum retroviralgp70 antigen, B6 mice produce low levels of serum gp70[14]. This raised a possibility that some genetic factorsinfluencing the production of gp70 may implicate gp70IC formation. We therefore performed an analysis of thedistribution of serum gp70 in BC1 mice. Since after 2months of age circulating gp70 does not change greatlythroughout life, levels of gp70 in BC1 and control micewere determined at 6 months. Results of this analysisindicated that there was a wide spread of gp70 levelsamong BC1 mice with values overlapping B6.Yaa (2.3 ±0.7 ? g/ml) and (NZW × B6.Yaa)F1 (9.2 ± 3.3 ? g/ml)parental phenotypes (Fig. 1D). Of the 126 BC1 mice, 65(52 %) developed serum levels of gp70 greater than 3 SDabove the mean of B6.Yaa males (4.4 ? g/ml).
2.3 Association of serum levels of IgG anti-DNAantibodies or retroviral gp70 IC with severeGN
The development of severe GN among BC1 progeny wasnext investigated in relation to the production of anti-DNA antibodies or gp70 IC (Fig. 1 and Table 1). Compari-sons show that there was a significant associationbetween severe nephritis and high titers of IgG anti-DNAantibodies (p = 4.0 × 10–4). A much stronger associationwas, however, detected by the analysis of gp70 IC distri-bution, showing a remarkable bias toward high levels of
4258 M.-L. Santiago et al. Eur. J. Immunol. 1998. 28: 4257–4267

Figure 1. Inheritance of lupus-like autoimmune disease in 126 B6 × (NZW × B6.Yaa)F1 males: (A) Incidence of glomerular lesionsin backross mice. Black bars represent grades 3 and 4 GN considered significant contributors to clinical disease and/or death.Serum levels of (B) anti-DNA IgG autoantibodies (U/ml), (C) retroviral gp70 IC ( ? g/ml), and (D) total gp70 antigen ( ? g/ml) in indi-vidual backcross (BC1) mice as compared to parental B6.Yaa and (NZW × B6.Yaa)F1 (F1) mice. Anti-DNA autoantibody levels areshown for peak levels before 8 months of age, and gp70 IC and gp70 antigen are shown at 6 months of age. The horizontal linesrepresent the mean plus 3 SD for autoantibody and antigen levels in age-matched B6.Yaa males. Backcross mice with autoanti-body and antigen levels below/above the horizontal line were assigned a low/high autoantibody phenotype, respectively. Back-cross mice that developed severe nephritis (grade n 3) are shown as solid circles, and mice that developed no abnormality tomoderate nephritis (grade X 3) are depicted as open circles.
gp70 IC in mice with severe GN (p = 8.9 × 10–8), whichsuggests that autoantibodies to gp70 are the major path-ogenic antibodies in the (NZW × B6.Yaa)F1 model oflupus nephritis as shown in other lupus mice [8, 14, 15].
A comparison of IgG anti-DNA antibodies and gp70 ICrevealed a significant correlation between both autoim-mune responses (rs = 0.32; p = 2.0 × 10–4), which mightreflect a common mechanism regulating their produc-tion. When the serum levels of retroviral gp70 antigenwere plotted against the levels of gp70 IC, a strongercorrelation was observed (rs = 0.53; p X 1.0 × 10–4), sug-gesting the possibility that the levels of gp70 influencethe formation of gp70 IC.
2.4 Linkage of marker loci to GN
The comparison of genotypes of BC1 mice classified aspositive or negative for severe GN was conducted byChi-square analysis. The only significant linkage wasobserved with chromosome 7 loci, with the strongesteffect for D7Mit79 locus ( £ 2 = 21.5; p X 4.0 × 10–6). Miceheterozygous at this locus developed a significantlymore severe GN than homozygous mice. The intervalwhere the GN-controlling locus lies is limited byD7Mit114 and D7Nds5 marker loci. No other region ofthe genome was found to be associated with the severityof GN. In particular, no association was found with chro-mosome 17 loci. However, when BC1 mice were classi-
Eur. J. Immunol. 1998. 28: 4257–4267 QTL linked with Yaa gene-induced lupus-like nephritis 4259
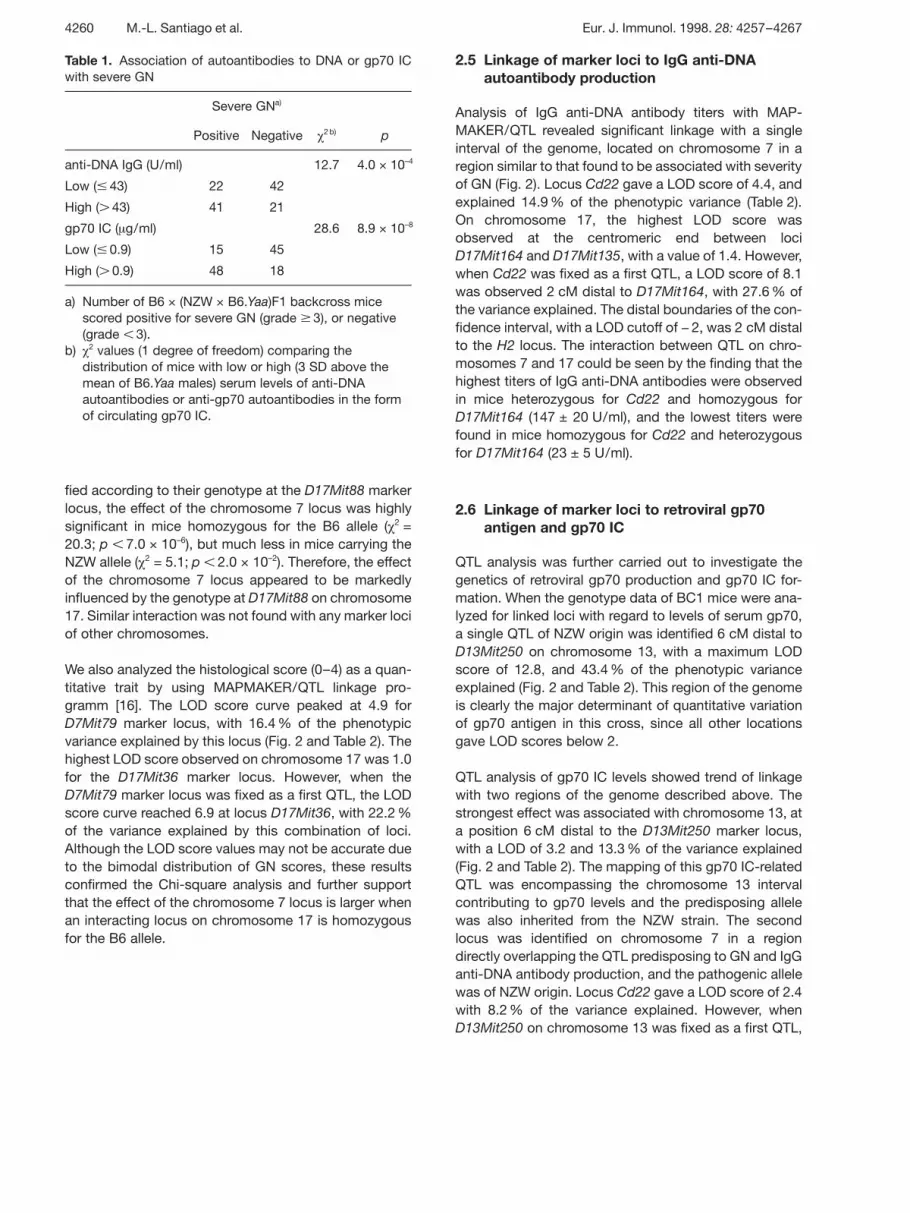
Table 1. Association of autoantibodies to DNA or gp70 ICwith severe GN
Severe GNa)
Positive Negative £2 b) p
anti-DNA IgG (U/ml) 12.7 4.0 × 10–4
Low ( p 43) 22 42
High ( G 43) 41 21
gp70 IC ( ? g/ml) 28.6 8.9 × 10–8
Low ( p 0.9) 15 45
High ( G 0.9) 48 18
a) Number of B6 × (NZW × B6.Yaa)F1 backcross micescored positive for severe GN (grade n 3), or negative(grade X 3).
b) £ 2 values (1 degree of freedom) comparing thedistribution of mice with low or high (3 SD above themean of B6.Yaa males) serum levels of anti-DNAautoantibodies or anti-gp70 autoantibodies in the formof circulating gp70 IC.
fied according to their genotype at the D17Mit88 markerlocus, the effect of the chromosome 7 locus was highlysignificant in mice homozygous for the B6 allele ( £ 2 =20.3; p X 7.0 × 10–6), but much less in mice carrying theNZW allele ( £ 2 = 5.1; p X 2.0 × 10–2). Therefore, the effectof the chromosome 7 locus appeared to be markedlyinfluenced by the genotype at D17Mit88 on chromosome17. Similar interaction was not found with any marker lociof other chromosomes.
We also analyzed the histological score (0–4) as a quan-titative trait by using MAPMAKER/QTL linkage pro-gramm [16]. The LOD score curve peaked at 4.9 forD7Mit79 marker locus, with 16.4 % of the phenotypicvariance explained by this locus (Fig. 2 and Table 2). Thehighest LOD score observed on chromosome 17 was 1.0for the D17Mit36 marker locus. However, when theD7Mit79 marker locus was fixed as a first QTL, the LODscore curve reached 6.9 at locus D17Mit36, with 22.2 %of the variance explained by this combination of loci.Although the LOD score values may not be accurate dueto the bimodal distribution of GN scores, these resultsconfirmed the Chi-square analysis and further supportthat the effect of the chromosome 7 locus is larger whenan interacting locus on chromosome 17 is homozygousfor the B6 allele.
2.5 Linkage of marker loci to IgG anti-DNAautoantibody production
Analysis of IgG anti-DNA antibody titers with MAP-MAKER/QTL revealed significant linkage with a singleinterval of the genome, located on chromosome 7 in aregion similar to that found to be associated with severityof GN (Fig. 2). Locus Cd22 gave a LOD score of 4.4, andexplained 14.9 % of the phenotypic variance (Table 2).On chromosome 17, the highest LOD score wasobserved at the centromeric end between lociD17Mit164 and D17Mit135, with a value of 1.4. However,when Cd22 was fixed as a first QTL, a LOD score of 8.1was observed 2 cM distal to D17Mit164, with 27.6 % ofthe variance explained. The distal boundaries of the con-fidence interval, with a LOD cutoff of − 2, was 2 cM distalto the H2 locus. The interaction between QTL on chro-mosomes 7 and 17 could be seen by the finding that thehighest titers of IgG anti-DNA antibodies were observedin mice heterozygous for Cd22 and homozygous forD17Mit164 (147 ± 20 U/ml), and the lowest titers werefound in mice homozygous for Cd22 and heterozygousfor D17Mit164 (23 ± 5 U/ml).
2.6 Linkage of marker loci to retroviral gp70antigen and gp70 IC
QTL analysis was further carried out to investigate thegenetics of retroviral gp70 production and gp70 IC for-mation. When the genotype data of BC1 mice were ana-lyzed for linked loci with regard to levels of serum gp70,a single QTL of NZW origin was identified 6 cM distal toD13Mit250 on chromosome 13, with a maximum LODscore of 12.8, and 43.4 % of the phenotypic varianceexplained (Fig. 2 and Table 2). This region of the genomeis clearly the major determinant of quantitative variationof gp70 antigen in this cross, since all other locationsgave LOD scores below 2.
QTL analysis of gp70 IC levels showed trend of linkagewith two regions of the genome described above. Thestrongest effect was associated with chromosome 13, ata position 6 cM distal to the D13Mit250 marker locus,with a LOD of 3.2 and 13.3 % of the variance explained(Fig. 2 and Table 2). The mapping of this gp70 IC-relatedQTL was encompassing the chromosome 13 intervalcontributing to gp70 levels and the predisposing allelewas also inherited from the NZW strain. The secondlocus was identified on chromosome 7 in a regiondirectly overlapping the QTL predisposing to GN and IgGanti-DNA antibody production, and the pathogenic allelewas of NZW origin. Locus Cd22 gave a LOD score of 2.4with 8.2 % of the variance explained. However, whenD13Mit250 on chromosome 13 was fixed as a first QTL,
4260 M.-L. Santiago et al. Eur. J. Immunol. 1998. 28: 4257–4267

Figure 2. Interval mapping scans showing QTL on chromosomes 7, 13 and 17. Scans for GN (−), anti-DNA IgG (--), gp70 IC (...)and total gp70 (−) are shown. The positions of marker loci are indicated on the x axis. The distances between each marker indi-cated in cM were generated by MAPMAKER/QTL [16] using the Kosambi function, whereas the position from the centromere ofproximal locus markers are those indicated in the MGD map. Log-likelihood (LOD) values for the presence of a QTL were calcu-lated at 2-cM intervals by MAPMAKER/QTL. The most likely position (95 % confidence intervals) for each QTL, determined by its2-LOD support interval, is indicated by a horizontal bar below the plot. As discussed in the text, analysis of genotypic combina-tions (indicated in parentheses as fixed chromosomes) revealed interaction of chromosome 7 and 13 QTL for gp70 IC formation,and of chromosome 7 and 17 QTL for GN severity and anti-DNA autoantibody production.
a LOD score of 5.4 was observed at the Cd22 locus with19.9 % of the variance explained (Fig. 2 and Table 2),indicating an additive effect of chromosomes 7 and 13QTL to increase gp70 IC formation.
3 Discussion
We used a panel of B6 × (NZW × B6.Yaa)F1 backcrossmales to define susceptibility loci linked with the Yaagene-induced lupus-like autoimmune syndrome. Agenomic segment of NZW origin on chromosome 7 wasfound to exhibit significant linkage to several SLE sever-ity traits, namely GN, IgG anti-DNA antibody production,and, due to its combined genotype with an additional
QTL linked to the production of retroviral gp70 antigenon chromosome 13, to gp70 IC formation, suggesting asingle locus with pleiotropic effects. Our study revealedalso two QTL of B6 origin on chromosome 17 showingno individual effects but strongly interacting with thechromosome 7 locus to enhance GN severity and levelsof serum IgG anti-DNA autoantibodies. Thus, the datafrom this backcross demonstrate that one critical geneor a closely linked group of genes of NZW origin on chro-mosome 7 play a key role in the predisposition of (NZW ×B6)F1 mice to Yaa gene-induced lupus-like autoimmunesyndrome.
In this backcross study, significant QTL for severe GNand anti-DNA autoantibody production clearly overlap
Eur. J. Immunol. 1998. 28: 4257–4267 QTL linked with Yaa gene-induced lupus-like nephritis 4261

Table 2. Estimated relative contribution to SLE-related traits of the genotypic combinations at chromosome 7, 13, and 17 QTLa)
Trait Peak marker Chromosome Position (cM) SLE traitseverityb)
LOD score Phenotypicvariance (%)
GN D7Mit79 7 25 W G B 4.9 16.4
D17Mit36 17 24 B G W 1.0 −
D7Mit79 + D17Mit36 7 and 17 − − 6.9 22.2
Anti-DNA antibodies Cd22 7 24 W G B 4.4 14.9
D17Mit164 [+ 2]c) 17 6 B G W 1.4 −
Cd22 + D17Mit164 [+ 2]c) 7 and 17 − − 8.1 27.6
gp70 IC Cd22 7 24 W G B 2.4 8.2
D13Mit250 [+ 6]c) 13 29 W G B 3.2 13.3
CD22 + D13Mit250 [+ 6]c) 7 and 13 − W G B 5.4 19.9
gp70 antigen D13Mit250 [+ 6]c) 13 29 W G B 12.8 43.4
a) Analysis of data from 126 B6 × (NZW × B6.Yaa)F1 backcross mice studied histologically for renal pathology andserologically for circulating autoantibodies and gp70 antigen.
b) Parental alleles are ordered such that SLE trait severity is associated with heterozygosity NZW/B6 (W) or homozygosity B6/B6 (B) at the position considered.
c) Peak marker positioned 2 cM distal to D17Mit164, or 6 cM distal to D13Mit250.
on chromosome 7 within a region centromeric toD7Nds5 of NZW mice. Other linkage analyses that havemapped NZW lupus susceptibility loci on the centro-meric portion of chromosome 7 include a study by Morelet al. [4]. Using (NZM × B6)F1 × NZM backcross mice,these authors mapped a locus, designated Sle3, contrib-uting to the incidence and severity of lupus-like renal dis-ease. This lupus susceptibility locus was most closelylinked to the pink-eyed dilution (p) locus, located approx-imately 10 cM distal to the D7Nds5 marker locus. Inaddition, a second peak for GN was identified 4 cM cen-tromeric of D7Nds5, suggesting the possible existenceof a second lupus susceptibility gene within Sle3. Inter-estingly, the study of Morel et al. [4] also showed that amarker close to the centromere on chromosome 7 was intrend of linkage for anti-DNA antibody production. Thepresent mapping of a locus centromeric of D7Nds5which is essential for the SLE phenotype in B6 × (NZW ×B6.Yaa)F1 mice emphasizes the presence of a secondlupus susceptibility allele at the centromeric end of Sle3.
In another study using (NZB × NZW)F2 (BWF2) inter-crosses, Kono et al. [6] identified a dominant NZW sus-ceptibility locus, designated Lbw5, linked to early mor-tality that also mapped on chromosome 7 near theD7Nds5 marker locus. However, the linkage analysis ofBWF2 mice to antichromatin antibody production orsevere GN did not reveal linkage to the Lbw5 locus.Although this study may indicate the potential involve-ment of a third NZW lupus susceptibility allele within the
Sl3/Lbw5 region, it should be emphasized that back-ground genes may modify the phenotype linked with aparticular locus and that linkage analysis reveals onlythose trait-causing genes that differ between parentalstrains used [17, 18]. The apparent contribution of theLbw5 locus to a distinct phenotype in BWF2 mice, thus,may reflect variations of the strain combinations andcrosses used among the different studies, and conse-quently, this locus could also define the NZW allele thatleads to a susceptible phenotype in presence of the Yaagene.
The present study identified a genomic segment of NZWorigin on chromosome 13 exhibiting highly significantlinkage for retroviral gp70 antigen and explaining 43 % ofthe variance of BC1 mice; it is the major locus whichaccounts for the production of elevated serum levels ofgp70. Although it has been suggested that multiplegenes are involved in the control of serum levels of gp70,the locus identified in the present study maps to a chro-mosomal location different from that of previously identi-fied loci: Gv2 and Sgp2 located at the telomeric end ofchromosome 7 [19, 20], and Sgp1 linked to the H2 locuson chromosome 17 [21]. Since the locus on chromo-some 13 showed an additive effect on gp70 IC formationbut had no apparent influence on the development ofIgG anti-DNA autoantibody responses and severe GN, itmay be that this NZW locus regulates the production ofgp70 antigen but not the antibody response to gp70. Infact, the weak linkage with gp70 IC probably reveals that
4262 M.-L. Santiago et al. Eur. J. Immunol. 1998. 28: 4257–4267

the gene located in this region is controlling the produc-tion of gp70 IC only indirectly. Accordingly, higher levelsof gp70 IC could be a consequence of increased con-centrations of circulating gp70.
It should be stressed that our backcross study revealedtwo QTL of B6 origin on chromosome 17 which stronglyinteract with the chromosome 7 locus to enhance theseverity of the renal disease and anti-DNA antibody pro-duction, although these two QTL remained undetectedwhen single-locus interval mapping was applied. Thecombinations of NZW alleles at chromosome 7 and 17QTL resulted in reduced incidence of severe GN andlower titers of anti-DNA autoantibodies, compared withcombinations of NZW allele at chromosome 7 and B6allele at chromosome 17 QTL. Although this may sug-gest the presence of one or two NZW dominant alleleson chromosome 17 that moderate the aggravating effectof the chromosome 7 locus, this conclusion is not com-patible with the high incidence of severe lethal lupus-likedisease in (NZW × B6.Yaa)F1 mice. Alternatively, ourdata may argue for the presence in the B6 background ofsusceptibility loci modifying expression of lupus-like dis-ease, but which by themselves are not sufficient to pre-dispose to the Yaa gene effect. This is supported bystudies showing linkage of background B6 or BALB/cloci to autoantibody production in crosses with the MRLstrain in presence of the Faslpr mutation or with the NZWstrain, respectively [22, 23]. It is of interest that confi-dence intervals for both chromosome 17 loci overlappedthe MHC locus. Several investigators have previouslyshown the importance of the H2 locus, particularly het-erozygosity, in the (NZB × NZW)F1 murine model of SLE[4, 6, 8, 24, 25]. In fact, the H2b haplotype (B6 origin)expressed in the present backcross mice has beenshown to exhibit a dominant effect on autoimmuneresponses in lupus-prone BXSB and (NZB × BXSB)F1mice [26, 27]. This could explain an increased suscepti-bility associated with the H2b/z heterozygosity (vs. theH2z/z) observed in (NZM × B6) × NZM backcross mice, inwhich the H2z/z is likely a less susceptible haplotype [4].Since some gene dosage effect of the MHC class II mol-ecules on the development of autoimmune responseshas been previously described [28, 29], our presentresults could be explained by the fact that the MHC genedosage effect may be more apparent in mice whosegenetic background is less predisposed to autoimmunedisease.
The overlapping QTL for GN severity, IgG anti-DNA anti-body production and gp70 IC formation on chromo-some 7 suggest that the basic defect produced by thislocus is B lymphocyte hyperactivity, which in presence ofthe Yaa gene gives rise to accelerated autoantibody pro-duction, immune complex formation and renal disease.
In fact, the possible presence within this region of anNZW allele that mediates polyclonal B cell activation isconsistent with the spontaneous production of IgM andIgG antibodies against both nuclear and non-nuclearantigens observed in B6 mice congenic for the NZWallele at the Sle3 locus [30]. In a previous study, we haveshown that genetic defects associated with the develop-ment of lupus-like disease in (NZB × NZW)F1 mice, suchas hypergammaglobulinemia and antinuclear antibodyproduction, are likely to be expressed at the level of Bcells in NZW mice [31, 32]. Since the Yaa gene abnormal-ity is likely to be expressed at the level of B cells [33], onecan speculate that the NZW susceptibility allele on thechromosome 7 locus defined in our present study couldcontribute to the development of SLE in(NZW × B6.Yaa)F1 mice as a result of its combinedexpression with the Yaa gene in the B lineage. Thismodel would tie together the findings that the Yaa geneaccelerates the progression of SLE and enhances theoverall antibody responses against self antigens as wellas foreign antigens only in mice that are predisposed toautoimmune disease, but not in immunologically normalCBA/J and B6 mice which no not manifest B cell abnor-malities [11].
The chromosome 7 region flanked by D7Mit76 andD7Nds5 marker loci is rich in genes that have an impor-tant role in the control of B cell differentiation and activa-tion, and could be candidate genes for the genetic pre-disposition to the Yaa gene effect. Among them, the sur-face glycoprotein of B lymphocytes CD22 is a negativeregulator of antigen receptor signaling. Mice deficient inCD22 exhibit augmented intracellular calcium responsesfollowing BCR stimulation and spontaneously produceanti-DNA antibodies [34–36]. Interestingly, the LODscore curves supporting a proximal chromosome 7 locuscontrolling the Yaa gene-induced lupus-like disease in(NZW × B6.Yaa)F1 mice peaked in the vicinity of theCd22 locus. Other B lineage cell-related genes withpotential relevance to the Yaa gene effect include Cd79a,which encodes the Ig § component of the B cell antigenreceptor [37], bcl3, a member of the I O B family, whichencodes proteins that regulate the NF- O B family of tran-scription factors [38], and Otf2, an octamer binding tran-scription factor which mediates transcription of Ig genes[39].
Clearly, our present study provides a model in which thegenetic complexity of the susceptibility to lupus-like dis-ease is markedly reduced, indicating that the Yaa genemay help to dissect the genetic predisposition to murinelupus-like disease in other strain combinations. In the(NZW × B6.Yaa)F1 model, the construction of congenicand recombinant strains in which the effects of the chro-mosome 7 locus can be evaluated, should allow fine
Eur. J. Immunol. 1998. 28: 4257–4267 QTL linked with Yaa gene-induced lupus-like nephritis 4263

mapping and ultimately permit positional cloning of thegene(s) underlying the NZW predisposition to murineSLE. Finally, without determining the exact nature of thegenes corresponding to this susceptibility locus, throughthe use of conserved synteny between proximal mousechromosome 7 and human chromosome 19q13 region[40], our study may help define the genetic basis ofhuman SLE and related autoimmune diseases [41].
4 Materials and methods
4.1 Mice
Parental NZW and B6 female mice were purchased fromHarlan Olac Ltd. (Oxon, GB) and IFFA-CREDO (Lyon,France), respectively. B6.Yaa males were established byrepeated backcrosses (n G 20), as described previously [12].The (NZW × B6.Yaa) and B6 × (NZW × B6.Yaa)F1 backcrossmice were obtained by local breeding in our own animalfacility. Blood samples were collected every 2 months fromthe age of 2 months by orbital sinus puncture, and the serawere stored at − 20 °C until use. Only male animals were uti-lized in the present study.
4.2 Serological assays
The presence of serum IgG anti-DNA antibodies wasassessed by ELISA in which heat-denatured calf thymusDNA (Sigma Chemical Co., St. Louis, MO) was used as coat-ing antigen, as previously described [42]. Results areexpressed in titration units (U/ml) in reference to a standardcurve obtained from a pool serum of 3–4-month-old MRL-lpr/lpr mice.
Serum levels of retroviral gp70 IC were quantified by ELISAcombined with the precipitation of the serum with PEG(average molecular weight 6000), which precipitates only theantibody-bound gp70, and not free gp70, as described [43].Results are expressed as ? g/ml of gp70 complexed withanti-gp70 autoantibodies by referring to a standard curveobtained from NZB sera with known amounts of gp70. Theconcentrations of total gp70 (free and complexed forms) inserum samples were determined by the same ELISA.
4.3 Histopathology
Kidney samples were collected at the time of killing whenmice were moribund or at the end of the experiment, at 15months of age. Histological sections were stained with peri-odic acid-Schiff reagent or hematoxylin and eosin. Theextent of GN was graded on a 0 to 4 scale based on theintensity and extent of histopathological changes, where agrade of 0 was given to kidneys without glomerular lesions,1 corresponded to minimal thickening of the mesangium, 2
to noticeable increases in both mesangial and glomerularcellularity, 3 were characterized by the preceding featureswith superimposed inflammatory exudates and capsularadhesions, and in 4, the glomerular architecture was obliter-ated in G 70 % of glomeruli, and tubular cast formation wasextensive. GN with grade 3 or 4 was considered significantcontributor to clinical disease and/or death.
4.4 Genetic mapping using simple sequence lengthpolymorphism
Genotypes were determined by PCR using 95 selected sim-ple length polymorphism markers over the autosomalgenome. Oligonucleotides flanking simple sequence repeatswere purchased from Research Genetics (Huntsville, AL).DNA from NZW, B6 and backcross mice was extracted fromtail biopsies kept frozen at − 70 °C before use. Amplificationof the simple sequence repeats was achieved by PCR with athermal cycler (HyBaid, GB). The PCR products were loadedonto a 3.5 % Nusieve-1.5 % agarose gel, electrophoresed at8 V/cm for 2–3 h and visualized by ethidium bromide stain-ing and UV transillumination (254 nm). Coverage of thegenome was estimated by taking the position of the markerloci on the Mouse Genome Database (MGD) mouse geneticmap obtained via the internet at http://www.infor-matics.jax.org (The Jackson Laboratory, Bar Harbor, ME)and applying a swept radius of 20 cM, unless a chromo-some end was within 20 cM. We thus obtained a geneticlinkage map in which the longest predicted interval of a genelocus to a chromosomal marker was p 20 cM, except for thecentromeric portions of chromosomes 6 and 18, where themost proximal markers were 26 and 22 cM distant from thecentromeric ends, and the telomeric portions of chromo-somes 3 and 8, where the most distal markers were 34 and27 cM distant from the telomeric ends, respectively. Thegenotypic data of this cross have been deposited at theMGD (The Jackson Laboratory, Bar Harbor, ME). It should benoted that we increased the density of markers in certainregions reported to show linkage to autoimmune traits incrosses involving the NZW strain [4–8, 22], namely:D1Mit268, 15, 36, 17, 155 (for Sle1/Lbw7); D4Mit17, 9, 36(for Sle2/Lbw2); D5Mit99 (for Lbw3); D6Mit10 (for Lbw4);D7Mit178, 76, 114, 246, 79, 230, 38, D7Nds5 (for Sle3/Lbw5); D11Mit164 (for Lbw8); D14Mit257 (for Nwa);D16Mit12 (for Nwa1); D17Mit135, 16, 36 (for Sle4/Lbw1);D18Mit186, 188, 49 (for Lbw6); and D19Mit11 (for Nwa2).The Cd22 locus was analyzed by PCR using a 5' primerCATCCTCCTTTTTCAGAA and a 3' primer AGAT-TCCCACTGTCATTG hybridizing to exon 4 which contains a18-bp deletion in the NZW strain but not in the B6 strain [44].
4.5 Statistical analysis
Comparisons of the distribution of high and low levels ofanti-DNA IgG antibodies or retroviral gp70 IC among miceeither positive (grade 3 or 4) or negative (grade 0 to 2) for
4264 M.-L. Santiago et al. Eur. J. Immunol. 1998. 28: 4257–4267

severe GN were tested by Chi-square analysis, using a stan-dard (2 × 2) contingency matrix. Correlations betweenserum levels of anti-DNA IgG antibodies and retroviral gp70IC and gp70 antigen were tested using the non-parametricSpearman rank correlation coefficient, rs.
The association of GN with marker loci was either tested bya Chi-square test for goodness-of-fit against an expected50:50 distribution using a standard (2 × 2) contingencymatrix, or by using the linkage program, MAPMAKER/QTL[16], using the 0–4 GN score. Logarithmic transformation ofserum levels of anti-DNA IgG antibodies, retroviral gp70antigen and gp70 IC resulted in normalized distributions andwere used as quantitative traits for MAPMAKER/QTL analy-sis. The LOD threshold is set at 1.9 for suggestive linkage,and 3.3 for significant linkage, based on the recommenda-tion of Lander and Kruglyak [45]. However, since four pre-sumably independent phenotypic characters were analyzed,a Bonferroni correction factor of 1/4 was applied and theLOD threshold was therefore set at 2.5 and 3.9, respectively.
Acknowledgments: L. Reininger was supported by a grantfrom the Association de Recherche sur la Polyarthrite and S.Izui was supported by the Swiss National Foundation forScientific Research.
5 References
1 Klinman, D. M. and Steinberg, A. D., Inquiry intomurine and human lupus. Immunol. Rev. 1995. 144:157–193.
2 Kotzin, B. L., Systemic lupus erythematosus. Cell 1996.85: 303–306.
3 Andrews, B. S., Eisenberg, R. A., Theofilopoulos, A.N., Izui, S., Wilson, C. B., McConahey, P. J., Murphy,E. D., Roths, J. B. and Dixon, F. J., Spontaneousmurine lupus-like syndromes. Clinical and immunopath-ological manifestations in several strains. J. Exp. Med.1978. 148: 1198–1215.
4 Morel, L., Rudofsky, U. H., Longmate, J. A., Schiffen-bauer, J. and Wakeland, E. K., Polygenic control of sus-ceptibility to murine systemic lupus erythematosus.Immunity 1994. 1: 219–229.
5 Drake, C. G., Babcock, S. K., Palmer, E. and Kotzin, B.L., Genetic analysis of the NZB contribution to lupus-likeautoimmune disase in (NZB × NZW)F1 mice. Proc. Natl.Acad. Sci. USA 1994. 91: 4062–4066.
6 Kono, D. H., Burlingame, R. W., Owens, D. G., Kura-mochi, A., Balderas, R. S., Balomenos, D. and Theofi-lopoulos, A. N., Lupus susceptibility loci in New Zealandmice. Proc. Natl. Acad. Sci. USA 1994. 91:10168–10172.
7 Drake, C. G., Rozzo, S. J., Hirschfeld, H. F., Smarnwo-rawong, N. P., Palmer, E. and Kotzin, B. L., Analysis ofthe New Zealand Black contribution to lupus-like renaldisease. Multiple genes that operate in a threshold man-ner. J. Immunol. 1995. 154: 2441–2447.
8 Vyse, T. J., Drake, C. G., Rozzo, S. J., Roper, E., Izui,S. and Kotzin, B. L., Genetic linkage of IgG autoanti-body production in relation to lupus nephritis in NewZealand hybrid mice. J. Clin. Invest. 1996. 98:1762–1772.
9 Theofilopoulos, A. N., The basis of autoimmunity: PartII. Genetic predisposition. Immunol. Today 1995. 16:150–159.
10 Murphy, E. D. and Roths, J. B., A Y chromosome asso-ciated factor in strain BXSB producing accelerated auto-immunity and lymphoproliferation. Arthritis Rheum.1979. 22: 1188–1194.
11 Izui, S., Iwamoto, M., Fossati, L., Merino, R., Takaha-shi, S. and Ibnou-Zekri, N., The Yaa gene model of sys-temic lupus erythematosus. Immunol. Rev. 1995. 144:137–156.
12 Izui, S., Higaki, M., Morrow, D. and Merino, R., The Ychromosome from autoimmune BXSB/MpJ miceinduces a lupus-like syndrome in (NZW × C57BL/6)F1male mice, but not in C57BL/6 male mice. Eur. J. Immu-nol. 1988. 18: 911–915.
13 Izui, S., Kelley, V. E., Masuda, K., Yoshida, H., Roths,J. B. and Murphy, E. D., Induction of various autoanti-bodies by mutant gene lpr in several strains of mice. J.Immunol. 1984. 133: 227–233.
14 Izui, S., McConahey, P. J., Theofilopoulos, A. N. andDixon, F. J., Association of circulating retroviral gp70-anti-gp70 immune complexes with murine systemiclupus erythematosus. J. Exp. Med. 1979. 149:1099–1116.
15 Maruyama, N., Furukawa, F., Nakai, Y., Sasaki, Y.,Ohta, K., Ozaki, S., Hirose, S. and Shirai, T., Geneticstudies of autoimmunity in New Zealand mice. IV. Contri-bution of NZB and NZW genes to the spontaneousoccurrence of retroviral gp70 immune complexes in(NZB × NZW)F1 hybrid and the correlation to renal dis-ease. J. Immunol. 1983. 130: 740–746.
16 Lander, E. S. and Botstein, D., Mapping mendelian fac-tors underlying quantitative traits using RFLP linkagemaps. Genetics 1989. 121: 185–199.
17 Rozzo, S. J., Vyse, T. J., Drake, C. G. and Kotzin, B. L.,Effect of genetic background on the contribution of NewZealand Black loci to autoimmune lupus nephritis. Proc.Natl. Acad. Sci. USA 1996. 93: 15164–15168.
18 Lander, E. S. and Schork, N. J., Genetic dissection ofcomplex traits. Science 1994. 265: 2037–2048.
Eur. J. Immunol. 1998. 28: 4257–4267 QTL linked with Yaa gene-induced lupus-like nephritis 4265

19 Stockert, E., Sato, H., Itakura, K., Boyse, E. A. andOld, L. J., Location of the second gene required forexpression of the leukemia-associated mouse antigenGIX. Science 1972. 178: 862–863.
20 Maruyama, N., Lindstrom, C. O., Sato, H. and Dixon,F. J., Serum gp70 production regulated by a gene onmurine chromosome 7. Immunogenetics 1983. 18:365–371.
21 Maruyama, N. and Lindstrom, C. O., H-2-linked regula-tion of serum gp70 production in mice. Immunogenetics1983. 17: 507–521.
22 Vyse, T. J., Morel, L., Tanner, F. J., Wakeland, E. K. andKotzin, B. L., Backcross analysis of genes linked toautoantibody production in New Zealand White mice. J.Immunol. 1996. 157: 2719–2727.
23 Vidal, S., Kono, D. H. and Theofilopoulos, A. N., Locipredisposing to autoimmunity in MRL-Faslpr and C57BL/6-Faslpr mice. J. Clin. Invest. 1998. 101: 696–702.
24 Hirose, S., Nagasawa, R., Sekikawa, I., Hamaoki, M.,Ishida, Y., Sato, H. and Shirai, T., Enhancing effect ofH-2-linked NZW gene(s) on the autoimmune traits of(NZB × NZW)F1 mice. J. Exp. Med. 1983. 158: 228–233.
25 Kotzin, B. L. and Palmer, E., The contribution of NZWgenes to lupus-like disease in (NZB × NZW)F1 mice. J.Exp. Med. 1987. 165: 1237–1251.
26 Merino, R., Fossati, L., Lacour, M., Lemoine, R.,Higaki, M. and Izui, S., H-2-linked control of the Yaagene-induced acceleration of lupus-like autoimmunedisease in BXSB mice. Eur. J. Immunol. 1992. 22:295–299.
27 Merino, R., Iwamoto, M., Gershwin, M. E. and Izui, S.,The Yaa gene abrogates the major histocompatibilitycomplex association of murine lupus in (NZB × BXSB)F1hybrid mice. J. Clin. Invest. 1994. 94: 521–525.
28 Cohen, P. L., Creech, E., Nakul-Aquaronne, D., McDa-niel, R., Ackler, S., Rapoport, R., Sobel, E. S. andEisenberg, R. A., Antigen nonspecific effect of majorhistocompatibility complex haplotype on autoantibodylevels in systemic lupus erythematosus-prone lpr mice.J. Clin. Invest. 1993. 91: 2761–2768.
29 Ibnou-Zekri, N., Iwamoto, M., Fossati, L., McCo-nahey, P. J. and Izui, S., Role of the major histocompati-bility complex class II E § gene in lupus susceptibility inmice. Proc. Natl. Acad. Sci. USA 1997. 94:14654–14659.
30 Morel, L., Mohan, C., Yu, Y., Croker, B. P., Tian, N.,Deng, A. and Wakeland, E. K., Functional dissection ofsystemic lupus erythematosus using congenic mousestrains. J. Immunol. 1997. 158: 6019–6028.
31 Reininger, L., Radaszkiewicz, T., Kosco, M., Mel-chers, F. and Rolink, A. G., Development of autoim-
mune disease in SCID mice populated with long-term «invitro» proliferating (NZB × NZW)F1 pre-B cells. J. Exp.Med. 1992. 176: 1343–1353.
32 Reininger, L., Winkler, T. H., Kalberer, C., Jourdan, M.,Melchers, F. and Rolink, A. G., Intrinsic B cell defects inNZB and NZW mice contribute to systemic lupus erythe-matosus in (NZB × NZW)F1 mice. J. Exp. Med. 1996.184: 853–861.
33 Merino, R., Fossati, L., Lacour, M. and Izui, S., Selec-tive autoantibody production by Yaa+ B cells in autoim-mune Yaa+-Yaa− bone marrow chimeric mice. J. Exp.Med. 1991. 174: 1023–1029.
34 O’Keefe, T. L., Williams, G. T., Davies, S. L. and Neu-berger, M. S., Hyperresponsive B cells in CD22-deficient mice. Science 1996. 274: 798–801.
35 Otipoby, K. L., Andersson, K. B., Draves, K. E., Klaus,S. J., Farr, A. G., Kerner, J. D., Perlmutter, R. M., Law,C.-L. and Clark, E. A., CD22 regulates thymus-independent responses and the lifespan of B cells.Nature 1996. 384: 634–637.
36 Sato, S., Miller, A. S., Inaoki, M., Bock, C. B., Jansen,P. J., Tang, M. L. K. and Tedder, T. F., CD22 is both apositive and negative regulator of B lymphocyte antigenreceptor signal transduction: altered signaling in CD22-deficient mice. Immunity 1996. 5: 551–562.
37 Sakaguchi, N., Kashiwamura, S.-I., Kimoto, M., Thal-mann, P. and Melchers, F., B lymphocyte lineage-restricted expression of mb-1, a gene with CD3-likestructural properties. EMBO J. 1988. 7: 3457–3464.
38 Kerr, L. D., Duckett, C. S., Wamsley, P., Zhang, Q.,Chiao, P., Nabel, G., Baeurle, T. W. and Verma, I. M.,The proto-oncogene bcl-3 encodes an I O B protein.Genes Dev. 1992. 6: 2352–2363.
39 Nelsen, B. and Sen, R., Regulation of immunoglobulingenes transcription. Int. Rev. Cytol. 1992. 133: 365–371.
40 Stubbs, L., Carver, E. A., Shannon, M. E., Kim, J.,Geisler, J., Generoso, E. E., Stanford, B. G., Dunn, W.C., Mohrenweiser, H., Zimmermann, W., Watt, S. M.and Ashworth, L. K., Detailed comparative map ofhuman chromosome 19q and related regions of themouse genome. Genomics 1996. 35: 499–508.
41 Tsao, B. P., Cantor, R. M., Kalunian, K. C., Chen, C.-J.,Badsha, H., Singh, R., Wallace, D. J., Kitridou, R. C.,Chen, S.-L., Shen, N., Song, Y. W., Isenberg, D. A., Yu,C.-L., Hahn, B. H. and Rotter, J. I., Evidence for linkageof a candidate chromosome 1 region to human systemiclupus erythematosus. J. Clin. Invest. 1997. 99: 725–731.
42 Rolink, A. G., Radaszkiewicz, T. and Melchers, F., Theautoantigen-binding B cell repertoires of normal and ofchronically graft-versus-host-diseased mice. J. Exp.Med. 1987. 165: 1675–1687.
4266 M.-L. Santiago et al. Eur. J. Immunol. 1998. 28: 4257–4267

43 Merino, R., Fossati, L., Lacour, M., Lemoine, R.,Higaki, M. and Izui, S., H-2 linked control of the Yaagene induced acceleration of lupus-like autoimmunedisease in BXSB mice. Eur. J. Immunol. 1992. 22:295–299.
44 Law, C.-L., Torres, R. M., Sundberg, H. A., Parkhouse,R. M. E., Brannan, C. I., Copeland, N. G., Jenkins, N.A. and Clark, E. A., Organisation of the murine cd22locus. Mapping to chromosome 7 and characterizationof two alleles. J. Immunol. 1993. 151: 175–187.
45 Lander, E. S. and Kruglyak, L., Genetic dissection ofcomplex traits: guidelines for interpreting and reportinglinkage results. Nat. Genet. 1995. 11: 241–247.
Correspondence: Luc Reininger, INSERM U399, Universitede la Mediterranee, 27 bd Jean Moulin, F-13385 Marseillecedex 5, FranceFax: +33-491 79 60 63e-mail: luc.reininger — medecine.univ-mrs.fr
Eur. J. Immunol. 1998. 28: 4257–4267 QTL linked with Yaa gene-induced lupus-like nephritis 4267





























