Embed Size (px)
Citation preview

Linking Immune Patterns and Life History Shows Two Distinct Defense Strategies in LandSnails (Gastropoda, Pulmonata)Author(s): Jacqueline Russo and Luc MadecReviewed work(s):Source: Physiological and Biochemical Zoology, Vol. 86, No. 2 (March/April 2013), pp. 193-204Published by: The University of Chicago PressStable URL: http://www.jstor.org/stable/10.1086/669482 .
Accessed: 24/02/2013 23:14
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
The University of Chicago Press is collaborating with JSTOR to digitize, preserve and extend access toPhysiological and Biochemical Zoology.
http://www.jstor.org
This content downloaded on Sun, 24 Feb 2013 23:14:31 PMAll use subject to JSTOR Terms and Conditions

193
Linking Immune Patterns and Life History Shows Two Distinct
Defense Strategies in Land Snails (Gastropoda, Pulmonata)
* Corresponding author; e-mail: [email protected].
Physiological and Biochemical Zoology 86(2):193–204. 2013. � 2013 by TheUniversity of Chicago. All rights reserved. 1522-2152/2013/8602-2097$15.00.DOI: 10.1086/669482
Jacqueline Russo*Luc MadecUniversite de Rennes 1, Unite Mixte de Recherche 6553ECOBIO, Campus de Beaulieu, batiment 14 A, F-35042Rennes Cedex, France
Accepted 11/10/2012; Electronically Published 2/11/2013
ABSTRACT
Life history integration of the defense response was investigatedat intra- and interspecific levels in land snails of the familyHelicidae. Two hypotheses were tested: (i) fitness consequencesof defense responses are closely related to life history traits suchas size at maturity and life span; (ii) different pathways of theimmune response based on “nonspecific” versus “specific” re-sponses may reflect different defense options. Relevant immuneresponses to a challenge with E. coli were measured using thefollowing variables: blood cell density, cellular or plasma an-tibacterial activity via reactive oxygen species (ROS) level, andbacterial growth inhibition. The results revealed that the largestsnails did not exhibit the strongest immune response. Instead,body mass influenced the type of response in determining theappropriate strategy. Snails with a higher body mass at maturityhad more robust plasma immune responses than snails with alower mass, which had greater cell-mediated immune responseswith a higher hemocyte density. In addition, ROS appeared alsoto be a stress mediator as attested by differences between sitesand generations for the same species.
Introduction
Immune function depends on individual genotype as well asenvironmental heterogeneity and can be considered as a lifehistory trait that has been shown to be involved in many re-source trade-offs with other physiological functions (Stearns1989; Moret and Schmid-Hempel 2000; Wiegel and Perelson2004; Fevrier et al. 2009; Lazzaro and Little 2009). The vari-ability of the immune response is the result of not only theindividual variation in resources available for this function but
also the cost paid when several functions share the same poolof resources. Thus, a “short-lived” species should not investtoo much in an immune system when its life history strategyis based on a high demographic turnover (Rantala and Roff2005). Similar arguments can be inferred from the relationbetween the immune system and the trade-off between age andsize at maturity (Wiegel and Perelson 2004). Immune re-sponses, which often involve many interrelated components,can also result in negative correlations between the differentimmune parameters such as humoral antimicrobial peptides,phenoloxidase activity, and hemocyte number (Moret andSchmid-Hempel 2001; Freitak et al. 2007; Wilfert et al. 2007).In a theory developed for vertebrate species, “fast-living” specieswith a high reproductive rate should rely more heavily on innateinflammatory immune defenses, while “slow-living” speciesshould exhibit stronger adaptive antibody-mediated immunity(Graham 2001; Lee 2006). Invertebrates lack acquired immuneresponses homologous to those of vertebrates, but they havedeveloped an innate defense consisting of both “nonspecificresponses” always ready to act against a wide range of pathogensand “specific responses” expressed only when needed againsta narrow range of microbes (Gillespie et al. 1997; Brown et al.2003; Rolff and Siva-Jothy 2003; Hamilton et al. 2008). Thus,despite a nonacquired immunity in mollusks, they also possesselements of both nonspecific and specific immune responses.A recent transcriptomic study comparing profiles of genes dif-ferentially expressed after immune challenge in Biomphalariaglabrata would indicate that transcripts involved in a specificresponse are surprisingly overexpressed compared with thosepotentially involved in a lesser or nonspecific response (Deleuryet al. 2012). At the cellular level, the most prominent com-ponents of nonspecific innate defense involve hemocyte phag-ocytosis or encapsulation with a cascade of biochemical reac-tions such as reactive oxygen species (ROS), serum factors(agglutinins, opsonins), and lytic enzymes probably originatedfrom hemocytes (Ratanarat-Brockelman 1977; Renwrantz1979). The facet of the specific immune response depends onthe production, with different time delays, of antimicrobialpeptides that aid in recognition of various pathogens, eithergram-negative or gram-positive bacteria (Hoffmann 2003;Hamilton et al. 2008). These two modalities of defense wouldbe independent (Moret 2003).
In this study, we assessed the immune response profileswithin a taxonomic group exhibiting a great ecological andmorphological diversity—the helicid land snails. We used fivespecies that vary in body size. Within this taxon, as in manyothers, life history traits covary in a predictable way; long-livedspecies contrast with short-lived ones that are sexually matureat an early age, have a higher reproductive rate, and die sooner.
This content downloaded on Sun, 24 Feb 2013 23:14:31 PMAll use subject to JSTOR Terms and Conditions

194 J. Russo and L. Madec
Body size, which is largely involved in these combinations oflife history traits, is also known in land snails to have a crucialinfluence on many physiological processes including immuneresponse (Rantala and Roff 2005; Phillipp and Abele 2010).Thus, because an adaptive change in body size might haverepercussions on almost all fitness components, this study firstaddresses the question of a relationship between body size, lifespan, and the robustness of immune defense, which itself in-volves a balance between the immune effectors.
The second goal of this study was to examine under the sameexperimental conditions the intraspecific variation on immuneresponse. This framework was used to understand individualvariation within populations of a well-known species, namely,Cornu aspersum. This species shows a high plasticity in age andsize at maturity that is correlated with a strong intra- andinterpopulation variation in growth and reproductive com-ponents (Madec et al. 1998, 2000, 2003). Genetic parametershave been previously estimated for growth, main reproductivetraits, and adult body mass (Dupont-Nivet et al. 1997, 1998).Additional work highlighted two distinct immune strategiesbetween the subspecies C. aspersum aspersum (the garden snail)and the giant slow-living Cornu aspersum maximum (Russoand Madec 2011). Here, we analyzed snails collected from threepopulations occurring in contrasted natural environments andfrom a snail farm where animals were reared for about 20generations. In the laboratory, snails were placed under artificialconditions for two generations in order to estimate the amountof variation within and among populations and to what extentthis variation was maintained over time. In this work, twoaspects of the internal defense—that is, its nonspecific (numberof circulating hemocytes, ROS release) versus specific (plasmabacterial growth inhibition) pathways—were examined using achallenge technique in order to quantify the immune responsesin a standardized way.
Material and Methods
Chemicals and Buffer
All chemicals were obtained from Sigma Aldrich Chimie(Lyon). Snail saline buffer was prepared according to Ademaet al. (1992).
Studied Species and Populations
Three hundred wild adult specimens—that is, snails with areflected shell lip—formed the first generation of each species(F0 for the three distinct wild populations of Cornu aspersumthat have been sampled; table 1). Descriptions and geographicaland ecological distributions of the species are given in Kerneyand Cameron (1979). The five species tolerate a wide range ofnatural habitats, but the sampled populations all occurred inmore or less anthropized habitats, as shown in table 1. Snailswere collected in early spring just after hibernation ended. Theywere acclimatized to the laboratory environment for 1 wk andthen maintained under similar conditions: they were housedin polyethylene containers (10 individuals per container) at
, 16L : 8D, and relative humidity. Water20� � 1�C 80% � 5%was available from the synthetic foam covering the cage bottom,which was kept moist. They were fed ad lib. with compositesnail food (Aliments Berton, Le Boupere). Food was removed2 d before blood sampling in order to avoid an individualvariation in food intake, which could have some effects onhemocyte counts (Carballal et al. 1998).
Cornu aspersum derived from the generation of wild animalscollected from sites 1, 2, and 3 formed the F1 generation ac-cording to an experimental design for snail reproduction de-scribed in Madec et al. (2000). This part of the experiment,involving different populations reared under artificial standardconditions over two generations, was designed to clarify themagnitude and the nature of the variation of immune re-sponses. The long-term effects of these experimental conditionswere examined on sexually mature snails C. aspersum aspersum(identified as Fn) collected in a snail farm where C. aspersumwas reared for about 20 generations in the same artificial en-vironment. The snails were kept in covered plastic containersat the same density and under the same conditions as describedabove. Body mass, measured with a microbalance (SartoriusCP 224 S; �0.1 mg), showed for each population a strongpositive correlation with the linear measure of size (shell di-ameter) and therefore was used to represent adult size (seeDupont-Nivet et al. 1997). Moreover, such a similar relation-ship indicated that snail condition was, on average, comparableamong samples.
Because parasitism would cause uncontrolled modificationof the immune system, animals were dissected and checked forinfection at the end of the study (genital apparatus, digestivetract, hemolymph). Those found with pathogens were excludedfrom the analyses.
Parasitism Prevalence
Few snails were found to be parasitized. Parasite species werenot always identified: (i) flagellates (Leptomonas) in hemo-lymph of C. aspersum site 1 (2.9%) and Cepaea nemoralis(5.7%); (ii) bacteria in hemolymph of Helix pomatia (0.5%),C. aspersum site 1 (0.5%), and C. nemoralis (0.5%); and (iii)the slug mite (Acari) Riccardoella limacum in the mantle cavityof C. aspersum site 1 (2.9%) and C. nemoralis (1%).
Hemolymph Sampling
Hemolymph was collected via a capillary cardiac puncture: adrop was used immediately for cell count, and the rest wastransferred in an Eppendorf tube with 100 mL of snail salinebuffer (SSB) with anticoagulant (EDTA 0.5%). The largest spe-cies provided 1,000 mL per snail, which was kept on ice andaliquoted for the different experiments. With the smallest spe-cies, C. nemoralis and Theba pisana, a pool of four snails wasused. All snails were placed on a starvation diet 2 d beforeblood sampling in order to avoid an individual variation infood intake, which could have a significant effect on hemocytecounts (Carballal et al. 1998).
This content downloaded on Sun, 24 Feb 2013 23:14:31 PMAll use subject to JSTOR Terms and Conditions

Tabl
e1:
Geo
grap
hica
lan
dec
olog
ical
char
acte
rist
ics
ofco
llect
ing
loca
litie
san
dm
ain
life
his
tory
feat
ure
sof
the
five
hel
icid
spec
ies
Spec
ies
Loca
tion
Hab
itat
Mea
nsn
ail
mas
s(g
)A
geat
mat
uri
tyLo
nge
vity
(yr)
Shel
ldi
amet
er(m
m)
Clu
tch
size
Egg
fres
hm
ass
(mg)
Clu
tch
no.
yr�
1
Ref
eren
ces
(oth
erth
anK
ern
eyan
dC
amer
on19
79)
Cor
nuas
pers
umss
p.as
pers
um,
site
1
Mt.
St.
Mic
hel
(48�
40′ N
,01
�30′ W
)
Edg
eof
arab
lefi
elds
(pol
der
area
)F 0
9.49
�.2
8,F 1
7.44
�.2
32
3–5
29–3
410
0–15
028
–33
2–3
Mad
ecan
dD
agu
zan
1993
;M
adec
etal
.20
00C
.ss
p.as
pers
um,
site
2N
iort
(46�
19′ N
,0�
27′ W
)R
oads
ide
ditc
hF 0
12.8
7�
2.08
,F 1
8.10
�.2
2SA
SASA
120–
150
30–3
42–
3M
adec
and
Dag
uza
n19
93C
.ss
p.co
noid
eum
,si
te3
Uti
que
(37�
00′ N
,9�
56′ E
)R
oman
ruin
sF 0
20.1
0�
.83,
F 17.
96�
.30
13
16
37–4
018
0–22
038
–42
2–3
L.M
adec
etal
.,u
npu
blis
hed
data
Hel
ixpo
mat
iaSt
rasb
ourg
(48�
35′ N
,7�
45W
)
Gra
ssla
nd
23.8
8�
.75
3–5
110
35–5
030
–60
100–
150
1–2
Pol
lard
1975
;G
omot
1990
;L
igas
zew
ski
etal
.20
09O
tala
lact
eaB
ejai
a(3
6�49
′ N,
5�3′ W
)U
rban
park
(gra
ss,
flow
erbe
ds)
9.53
�.3
31
4?
25–3
640
–100
(66)
U2–
4G
amm
on19
43
The
bapi
sana
Freh
el(4
8�7′ N
,2�
31′ W
)Sa
nd
dun
e5.
29�
.19
!2
212
–20
50–8
05–
105
Sacc
hi
1971
;C
owie
1984
;B
aker
1991
Cep
aea
nem
oral
isP
aim
pon
t(4
8�01
′ N,
2�10
′ W)
Dec
idu
ous
fore
sted
ge4.
90�
.21
!2
2–4
18–2
560
–80
10–1
52–
3W
olda
and
Kre
ule
n19
73;
Car
ter
and
Ash
dow
n19
84
Not
e.SA
pse
eab
ove;
Up
un
know
n.
This content downloaded on Sun, 24 Feb 2013 23:14:31 PMAll use subject to JSTOR Terms and Conditions

196 J. Russo and L. Madec
Total Hemocyte Count (THC) and Cell Viability
The hemolymph sample (20 mL) was directly dropped onto aMalassez hemocytometer for hemocyte counts (BH2 Olympusphase contrast light microscope). The cell viability, routinelydetermined by the trypan blue exclusion test (0.02%), remainedconsistently around 95%.
Injection of Lyophilized Bacteria
After preliminary tests, a single dose of lyophilized E.73 # 10coli (ATCC 11303) per gram of fresh snail mass was used insuspension in 100 mL of SSB (Bayne 1980). This dose resultedin substantial immunomodulation (Russo and Madec 2011).Dead bacteria were used because of their lack of replication,migratory capacity, and bacterial products. Snails were injectedwith a sterile single-use syringe and needle (0.5/16 mm) intothe cephalopedal blood sinus. Control snails were injected withthe same volume (100 mL) of SSB. The snails were sampled 2h later, when the level of circulating hemocyte reached maximalrate, the bacteria growth inhibition was maximal, and the clear-ance was faster (Russo and Madec 2011).
ROS Measurement
The intracellular detection of superoxide anions released byhemocytes was performed using a colorimetric method basedon the reduction of nitroblue tetrazolium (NBT) into insolubleblue formazan, which was extracted from the cells in the pres-ence of (Pipe 1995). For NBT reduction, three aliquots of�O2
100 mL hemolymph per snail were placed into microplate wells,which were then centrifuged (1,000 g, 3 min, 0�C). Attachedcells were treated with NBT (100 mL at 0.2% in SSB) for 30min in the dark. After two washes in SSB, cells were treatedtwice with methanol (100% and 50%). The blue formazan dyewas solubilized by adding 120 mL of 2 M KOH and 140 mLdimethyl sulfoxide to each well. Snails were sampled at 2 hpostinjection of E. coli. Superoxide dismutase was used at 300units mL�1 to test the specificity of the reaction (negative con-trol). Optical density was recorded at 620 nm, and results wereexpressed per milligram of protein (Bradford method accordingto Pipe et al. 1995).
In Vitro Plasma Antibacterial Activity
The plasma antibacterial activity (PABA) was monitored byturbidometry according to Crokaert et al. (1988) against thestrain ampicillin-resistant E. coli DH5a (Department Interac-tions Biologiques et Transferts de Matieres, University ofRennes 1). Plates were read using a kinetic package (SoftMaxPro 3.1. and SpectraMax 250, Molecular Devices) with bufferserving as a blank. The snails used in this experiment werenaive, injected with either SSB (controls) or lyophilized E. coli2 h earlier. Results were expressed as bacterial growth inhibitionaccording to the formula I p (absorbance of bacterial cul-ture � absorbance of sample)/(absorbance of bacterial culture).
Protein Assay
Protein concentration was determined with the Bradford rea-gent from the same sample used to assess the immune param-eters. This colorimetric method is based on the binding of thedye Coomassie brilliant blue G to the protein. The bindingshifts the absorption of the dye from red to blue and is pro-portional to protein concentration (absorbance at 595 nm).Protein content in the extracted samples was determined fromthe standard curve of bovine serum albumin. The calibrationcurve was constructed for each set of assays, and the amountof protein was expressed in milligrams per milliliter.
Statistical Analysis
Relationships between the branches of the immune system (de-pendent matrix) and maternal body mass were first exploredby means of a redundancy analysis (RDA) with factors “species”and “challenge” (nature of injection) and “snail mass” as in-strumental variables (explanatory matrix). General linear mod-eling was then used to test for the effects of species (F0 gen-eration), “challenge,” and their interaction on the variation ofimmunity measurements. These measurements were taken on10 individuals per treatment. Moreover, a model with fixedeffects was used to determine the influence of generation (Fi),geographic origin (site), and their interaction on the intraspe-cific variation of the same measurements in C. aspersum. Beforethe results were analyzed, we checked for normality and ho-moscedasticity in the model’s error terms. A Box-Cox trans-formation was applied to any data for which observed devia-tions from group means lacked normality. We applied nocorrection for the reduction of variance in pooled samples (C.nemoralis and T. pisana) because no significant decrease wasobserved. When heterogeneity among means was detected inmultisample experiments, we used a multiple comparison ap-proach (Tukey-Kramer procedure). RDA was made with theade4TkGUI package for R environment (Thioulouse and Dray2007), and other analyses were computed using Minitab 13.3.1software.
Results
The graphical exploration of data relative to wild snails wasillustrated by the principal plane of the RDA (fig. 1). The firstaxis, which accounted for 38% of the total variance, was es-sentially based on the contribution of snail mass and thenshowed a strong opposition between small species (Theba pis-ana, Cepaea nemoralis) and larger ones (Helix pomatia, Cornuaspersum from site 3), which seemed to invest more in plasmaticantibacterial activity. Axis 2 showed that investment in he-mocyte load and, to a lesser extent, in oxidative burst was higherin snails of a small or medium size, especially in Otala lactea.
The statistical analysis of the pattern of covariation suggestedby RDA showed highly significant negative correlations betweensnail mass and THC-ROS, which were positively correlated,and positive ones between snail mass and PABA protein load,which were positively correlated (table 2). When species (in-
This content downloaded on Sun, 24 Feb 2013 23:14:31 PMAll use subject to JSTOR Terms and Conditions

Immune Patterns and Life History in Land Snails 197
Figure 1. Principal plane of the redundancy analysis (RDA; projectionof explanatory variables not shown): vectorial representation of thecorrelations between dependent variables (immune measures) andRDA factors. THC p total hemocyte count; ROS p reactive oxygenspecies; PABA p plasma antibacterial activity; PROT p total proteinload. Mean factorial scores of each species (Ca 1, 2, 3, p Cornuaspersum site 1, 2, 3; Cn p Cepaea nemoralis; Hp p Helix pomatia;Ol p Otala lactea; Tp p Theba pisana) are superimposed.
Table 2: Matrix of product-moment correlations (total below diagonal, partial abovediagonal) between variables measured
Mass THC ROS PABA PROT
THC �.411 (!.001) ... .179 (.021) .011 (.884) .183 (.018)ROS �.332 (!.001) .302 (!.001) ... �.014 (.862) �.163 (.036)PABA .642 (.001) �.286 (!.001) �.355 (!.001) ... �.064 (.411)PROT .670 (.001) �.219 (!.005) �.447 (!.001) .495 (!.001) ...
Note. “Species,” “treatment,” and “mass” effects removed. THC p total hemocyte count; ROS p reactive oxygen
species; PABA p plasma antibacterial activity; PROT p total protein load; mass p snail fresh mass. Level of
significance of r in parentheses.
cluding the five taxa with the F0 generations of C. aspersum),challenge, and snail mass (covariate) effects were removed, onlythree partial correlations remained significant (table 2, abovediagonal).
THC
The species effect was highly significant for all response vari-ables, and a big difference between treatments was also observedfor THC ( , ). Interestingly, the smallestF p 9.66 P ! 0.0012, 143
species—O. lactea, C. nemoralis, and T. pisana—were charac-terized by the highest concentration of hemocytes.
At the intraspecific level, hemocyte concentration in the wildpopulation originating from site 1 differed significantly fromthose at sites 2 (TK test, ) and 3 (TK test, ;P p 0.03 P p 0.05fig. 2). Significant differences in THC were also found betweenpopulations and generations of C. aspersum ( ,F p 13.452, 154
; , ) and treatments (P ! 0.001 F p 11.52 P ! 0.001 F p1, 154 2, 154
, ). THC was also significantly different in wild17.49 P ! 0.001versus reared (Fn) snails (TK test, ), but these0.04 ! P ! 0.001
differences decreased significantly in the F1 generation exceptfor at site 3, where the difference between the generations washighly significant (fig. 2).
ROS
The ROS production differed between wild populations of thefive species ( , ) and treatments (F p 6.94 P ! 0.001 F p6, 143 2, 143
, ; fig. 3). Intraspecific differences in C. aspersum4.82 P p 0.009were also found between treatments ( , )F p 4.5 P ! 0.0012, 154
and generations ( , ), especially betweenF p 4.9 P p 0.021, 154
generations F0 and F1 from site 3 (TK test, ).P p 0.007
PABA
Concerning PABA, the maximum inhibitory activity was ob-tained with the plasma of H. pomatia and wild C. aspersumfrom site 3, which significantly differed from all the other pop-ulations (TK test, ; fig. 4). PABA measured in0.05 ! P ! 0.001C. aspersum, which was strongly correlated with snail mass( ; ), showed significant differences betweenr p 0.601 P ! 0.001populations ( , ), especially because of theF p 7.33 P ! 0.0012, 154
distinct pattern of wild snails from site 3 in regard to all otherF0 populations (fig. 4). This pattern was not found in the F1
population from site 3, which showed no significant differencefrom the other populations of C. aspersum.
Protein Concentration
Although highly significant differences were observed betweenpopulations ( , ), the hemolymph proteinF p 16.58 P ! 0.00112, 154
load was not significantly affected by the immune stimulation(fig. 5). The most important result was the similar patternexhibited by H. pomatia and wild C. aspersum from site 3,which was significantly different from all the others (TK test,
).P ! 0.001
Discussion
This study uses immune response to address the two followingtopics: (1) If immunocompetence traded with life span, whichis known to be frequently correlated to body size in land snails(Heller 2001), the species with larger size and higher survivalshould invest more in self-defense than the smaller ones (Loch-
This content downloaded on Sun, 24 Feb 2013 23:14:31 PMAll use subject to JSTOR Terms and Conditions

198 J. Russo and L. Madec
Figure 2. Changes in the total hemocyte count in the hemolymph of naive snails (white bars), 2 h postinjection of buffer (gray bars), and 2h postinjection of lyophilized E. coli (black bars). Black stars mark significant differences between naive and challenged snails, and white starsmark significant differences between F0 and F1 generations ( ). Inset, results of post hoc tests after ANOVA (one asterisk p ;P ! 0.05 P ! 0.05two asterisks p ; three asterisks p ; means � SE, per treatment).P ! 0.01 P ! 0.001 n p 10
miller and Deerenberg 2000). (2) Assuming that environmentalconditions strongly effect immunocompetence (Schmid-Hempel 2003; Lazzaro and Little 2009; Schulenburg et al. 2009)and knowing that snail populations live in a wide range ofecological conditions, we expect that subtle differences shouldcharacterize immune functional responses at both the speciesand population levels (Sadd and Schmid-Hempel 2009).
Inter- and Intraspecific Differences in Immune Traits
Immune responses did not allow any particular species to beclearly distinguished. The possible confounding effects on var-iance components due to the experimental design (low repli-cation, sample pooling for two species) cannot be totally ruledout to explain this result, but it seems that the covariationbetween ROS and plasmatic factors appeared not to be directlytaxon specific but related to body mass.
Both wild and captive-bred snails (Russo and Madec 2011)deployed both nonspecific and specific immune response whenthey were experimentally activated with the immune elicitor E.
coli. Among the array of putative antibacterial responses usedby challenged snails, only two kinds of immune effectors weremeasured that included hemocyte density with ROS release andantibacterial activity of hemolymph. In their immune response,the small-bodied species seemed to mobilize the nonspecificdefense based on hemocyte and ROS, whereas larger snailsinvested more in a specific defense involving the lytic hemo-lymph activity. As previously suggested, these defense processeswere not mutually exclusive, the two responses acting additivelyto counter pathogens. According to “two arms being betterthan one” (Hamilton et al. 2008), snails combined both re-sponses rather than relying on one or the other.
The blood sampled from O. lactea, C. nemoralis, and T. pisanacontained as much as 1.5 times as many cells as the bloodsampled from the largest species, H. pomatia, and had twice asmany hemocytes as C. aspersum from sites 3 and 2. After thebacterial injection, these small-bodied species characterized bya higher hemocyte activity showed that the response may de-pend on the investment in hemocyte concentration. Differenceswere also found between conspecific C. aspersum: snails from
This content downloaded on Sun, 24 Feb 2013 23:14:31 PMAll use subject to JSTOR Terms and Conditions
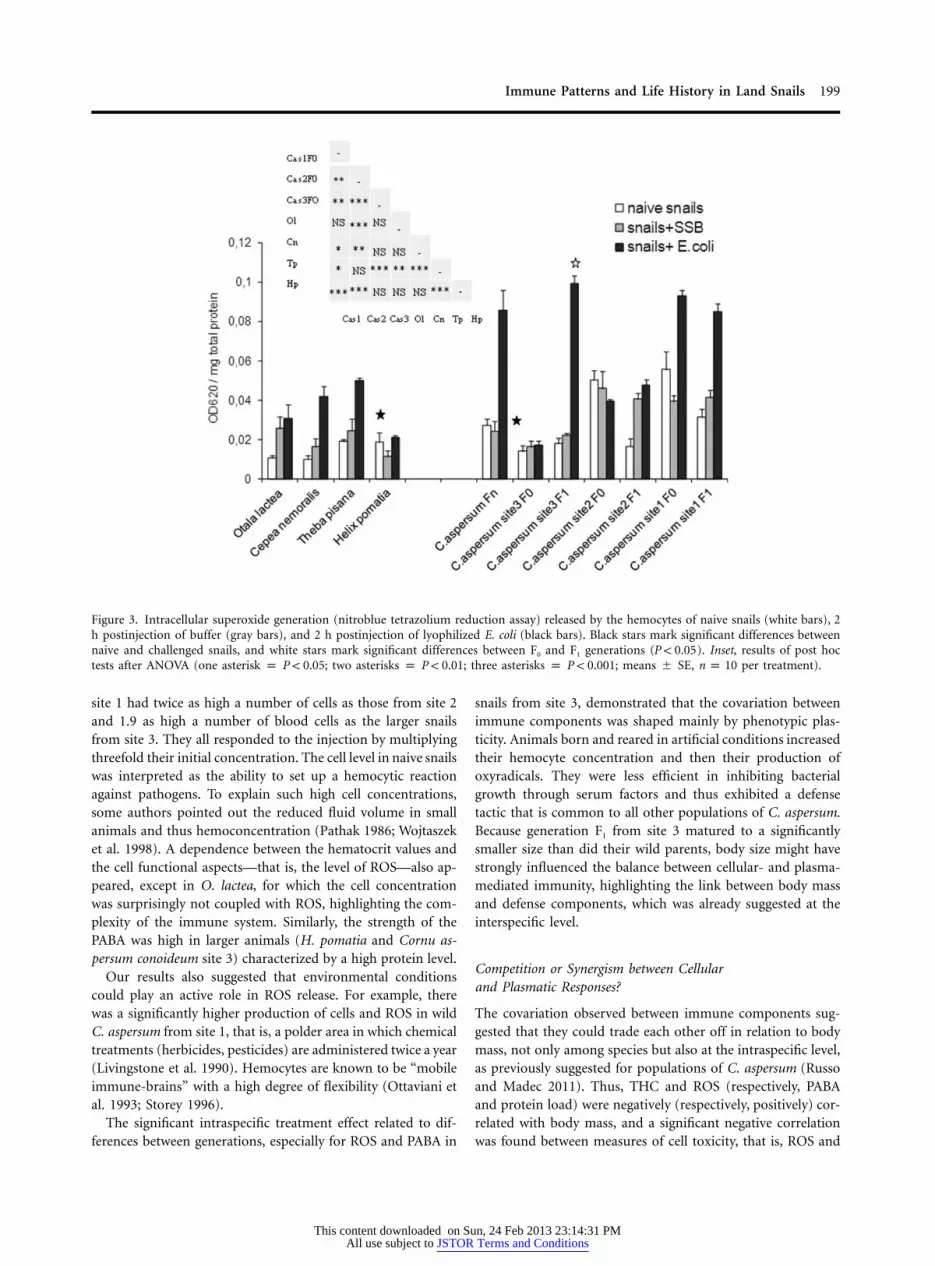
Immune Patterns and Life History in Land Snails 199
Figure 3. Intracellular superoxide generation (nitroblue tetrazolium reduction assay) released by the hemocytes of naive snails (white bars), 2h postinjection of buffer (gray bars), and 2 h postinjection of lyophilized E. coli (black bars). Black stars mark significant differences betweennaive and challenged snails, and white stars mark significant differences between F0 and F1 generations ( ). Inset, results of post hocP ! 0.05tests after ANOVA (one asterisk p ; two asterisks p ; three asterisks p ; means � SE, per treatment).P ! 0.05 P ! 0.01 P ! 0.001 n p 10
site 1 had twice as high a number of cells as those from site 2and 1.9 as high a number of blood cells as the larger snailsfrom site 3. They all responded to the injection by multiplyingthreefold their initial concentration. The cell level in naive snailswas interpreted as the ability to set up a hemocytic reactionagainst pathogens. To explain such high cell concentrations,some authors pointed out the reduced fluid volume in smallanimals and thus hemoconcentration (Pathak 1986; Wojtaszeket al. 1998). A dependence between the hematocrit values andthe cell functional aspects—that is, the level of ROS—also ap-peared, except in O. lactea, for which the cell concentrationwas surprisingly not coupled with ROS, highlighting the com-plexity of the immune system. Similarly, the strength of thePABA was high in larger animals (H. pomatia and Cornu as-persum conoideum site 3) characterized by a high protein level.
Our results also suggested that environmental conditionscould play an active role in ROS release. For example, therewas a significantly higher production of cells and ROS in wildC. aspersum from site 1, that is, a polder area in which chemicaltreatments (herbicides, pesticides) are administered twice a year(Livingstone et al. 1990). Hemocytes are known to be “mobileimmune-brains” with a high degree of flexibility (Ottaviani etal. 1993; Storey 1996).
The significant intraspecific treatment effect related to dif-ferences between generations, especially for ROS and PABA in
snails from site 3, demonstrated that the covariation betweenimmune components was shaped mainly by phenotypic plas-ticity. Animals born and reared in artificial conditions increasedtheir hemocyte concentration and then their production ofoxyradicals. They were less efficient in inhibiting bacterialgrowth through serum factors and thus exhibited a defensetactic that is common to all other populations of C. aspersum.Because generation F1 from site 3 matured to a significantlysmaller size than did their wild parents, body size might havestrongly influenced the balance between cellular- and plasma-mediated immunity, highlighting the link between body massand defense components, which was already suggested at theinterspecific level.
Competition or Synergism between Cellularand Plasmatic Responses?
The covariation observed between immune components sug-gested that they could trade each other off in relation to bodymass, not only among species but also at the intraspecific level,as previously suggested for populations of C. aspersum (Russoand Madec 2011). Thus, THC and ROS (respectively, PABAand protein load) were negatively (respectively, positively) cor-related with body mass, and a significant negative correlationwas found between measures of cell toxicity, that is, ROS and
This content downloaded on Sun, 24 Feb 2013 23:14:31 PMAll use subject to JSTOR Terms and Conditions

200 J. Russo and L. Madec
Figure 4. Escherichia coli (ampicillin-resistant DH5a) growth inhibition by snail plasma. Tests were done with naive snails, control injectedwith buffer and after receiving a single injection of lyophilized E. coli. Black stars mark significant differences between naive and challengedsnails, and white stars mark significant differences between F0 and F1 generations ( ). Inset, results of post hoc tests after ANOVA (oneP ! 0.05asterisk p ; two asterisks p ; three asterisks p ; means � SE, per treatment).P ! 0.05 P ! 0.01 P ! 0.001 n p 10
PABA. Some examples showed antagonism between hemo-lymph antibacterial activity and THC or phenol oxidase activity(Klasing and Leshchinsky 1999; Moret and Schmid-Hempel2001; Cotter et al. 2004; Wilfert et al. 2007). However, ourexperimental design was not optimal for understanding theunderlying mechanisms of the negative association observedbetween the variables, and further investigation is required tosupport such putative trade-offs (see Zera and Harshman2001).
Use of ROS or Plasmatic Factors as Killing Mechanisms:Benefits and Costs in an Evolutionary Perspective
Each component of the immune system has a priori its owncost, sometimes evaluated as costly (Sheldom and Verhulst1996) and sometimes not (Owen and Wilson 1999). Theoret-ically, a specific response should be less costly than a nonspecificone (Hoffmann et al. 1996; Moret 2003; Hamilton et al. 2008).Some authors tried to evaluate the immune cost through pro-
This content downloaded on Sun, 24 Feb 2013 23:14:31 PMAll use subject to JSTOR Terms and Conditions

Immune Patterns and Life History in Land Snails 201
Figure 5. Total protein load in hemolymph: naive snails (white bars), 2 h postinjection of buffer (gray bars), and 2 h postinjection of lyophilizedE. coli (black bars). Black stars mark significant differences between naive and challenged snails, and white stars mark significant differencesbetween F0 and F1 generations ( ). Inset, results of post hoc tests after ANOVA (one asterisk p ; two asterisks p ; threeP ! 0.05 P ! 0.05 P ! 0.01asterisks p ; means � SE, per treatment).P ! 0.001 n p 10
tein decrease (Adamo and Parsons 2006). In our study, noproteins seem to have been newly formed in response to im-mune challenge, and this has been reported in other studies(Houdijk et al. 2001; Jacot et al. 2005). However, we also haveto consider that the Bradford method just measures the con-centration of proteins with no distinction of their functionalimplications. Moreover, 2 h postexposure was perhaps too shortto evaluate the effect of inducible proteins, and immune pro-teins are not the highest part in the hemolymph. Therefore,from this experiment we are not able to estimate the real en-ergetic cost of achieving one response or the other. Perhaps thedifferent configurations represent neither an overall differencein the energy invested in immune function nor an optimalimmune response. Therefore, the immune cost may be balancedwith regard to fitness changes (Moret 2003; Rolff and Siva-Jothy 2003), and each response should reflect the effects onfitness of the host imposed by pathogens (Medley 2002; Tellaet al. 2002; Viney et al. 2005; Lee 2006; Sadd and Schmid-Hempel 2009; Graham et al. 2011). The ROS are toxic to thehost, while antimicrobial peptides had fewer negative side ef-fects, even if they can also create toxic metabolic breakdown
products from the bacteria. Thus, plasma bactericidal sub-stances can be regarded as providing selective advantages forlonger-lived species (Hoffmann et al. 1996; Tieleman and al.2005; Phillipp and Abele 2010). In trying to understand theinteractions between immune responses and other fitness com-ponents, one might ask what would be the main benefit ofROS-mediated immunity when it is obviously the more dam-aging type of defense. It is likely that snails counteract theimmunopathological effects by adjusting the antioxidant level(J. Russo and L. Madec, unpublished manuscript). Moreover,the ROS should obviously not be regarded simply as directedagainst pathogens and shaped through immune selection pres-sures. They may have been either involved in various biologicalprocesses, used as an indicator of energy consumption, or re-lated to the generally greater metabolism of small bodies(Blueweiss et al. 1978; Hennemann 1983; Dowling and Sim-mons 2009). So, for short-lived species, the oxidative rapid-acting mechanism should give a real advantage over the time-delayed production of PABA.
Until now, few studies have reported such a size-dependentcovariation between immune components (Rantala and Roff
This content downloaded on Sun, 24 Feb 2013 23:14:31 PMAll use subject to JSTOR Terms and Conditions

202 J. Russo and L. Madec
2005). The question arises as to whether this picture can begeneralized for all gastropod snails. We now have to integratethe results presented here into a life history framework to in-terpret the fitness consequences of the two strategies identified,development time and life span components, which are fre-quently correlated with body size in land snails (Heller 2001).
Acknowledgments
Dr. Annie Guiller is gratefully acknowledged for fruitful com-ments that greatly improved the manuscript.
Literature Cited
Adamo S.A. and N.M. Parsons. 2006. The emergence of life-history stage and immunity in the cricket, Gryllus texensis.Anim Behav 72:235–244.
Adema C.M., R.A. Harris, and E.C. Van Deutekom-Mulder.1992. A comparative study of haemocytes from six differentsnails: morphology and functional aspects. J Invertebr Pathol59:24–32.
Baker G.H. 1991. Production of eggs and young snails by adultTheba pisana (Muller) and Cernuella virgata (Da Costa)(Mollusca, Helicidae) in laboratory cultures and field pop-ulations. Aust J Zool 39:673–679.
Bayne C. 1980. Molluscan immunity: induction of elevatedimmunity in the land snail (Helix) by injections of bacteria(Pseudomonas aeruginosa). Dev Comp Immunol 4:43–54.
Blueweiss L., H. Fox, V. Kudzma, D. Nakashima, R. Peters, andS. Sams. 1978. Relationships between body size and somelife-history parameters. Oecologia 37:257–272.
Brown M.J.F., Y. Moret, and P. Schmid-Hempel. 2003. Acti-vation of host constitutive immune defence by an intestinaltrypanosome parasite of bumble bees. Parasitology 126:253–260.
Carballal M.J., A. Villalba, and C. Lopez. 1998. Seasonal vari-ation and effects of age, food availability, size, gonadal de-velopment, and parasitism on the hemogram of Mytilus gal-loprovincialis. J Invertebr Pathol 72:304–312.
Carter M.A. and M. Ashdown. 1984. Experimental studies onthe effects of density, size, shell colour and banding phe-notype on the fecundity of Cepaea nemoralis. Malacologia25:291–302.
Cotter S.C., L.E. Kruuk, and K. Wilson. 2004. Cost of resistance:genetic correlations and potential trade-offs in an insect im-mune system. J Evol Biol 17:421–429.
Cowie R.H. 1984. The life-cycle and productivity of the landsnail Theba pisana (Mollusca: Helicidae). J Anim Ecol 53:311–325.
Crokaert F., M.J. Lismont, M.P. Van Der Linden, and E. Your-assowsky. 1988. Determination of serum bactericidal activityagainst Escherichia coli by an automated photometricmethod. J Clin Microbiol 26:2069–2076.
Deleury E., G. Dubreuil, N. Elangovana, E. Wajnberg, J.M.Reichhart, B. Gourbal, D. Duval, O.L. Baron, J. Gouzy, and
C. Coustau. 2012. Specific versus non-specific immune re-sponses in an invertebrate species evidenced by a compar-ative de novo sequencing study. PLoS ONE 7:e32512.
Dowling D.K. and L.W. Simmons. 2009. Reactive oxygen spe-cies as universal constraints in life-history evolution. Proc RSoc B 276:1737–1745.
Dupont-Nivet M., J. Mallard, J.C. Bonnet, and J.M. Blanc. 1997.Quantitative genetics of growth traits in the edible snail Helixaspersa. Genet Sel Evol 29:571–587.
———. 1998. Quantitative genetics of reproductive traits inthe edible snail Helix aspersa Muller. J Exp Zool 281:220–227.
Fevrier Y., J. Russo, and L. Madec. 2009. Intraspecific variationin life history traits of a land snail after repeated bacterialchallenge. J Zool (Lond) 277:149–155.
Freitak D., C.W. Wheat, D.G. Heckel, and H. Vogel. 2007.Immune system responses and fitness costs associated withconsumption of bacteria in larvae of Tricoplusia ni. BMCBiol 5:56.
Gammon E.T. 1943. Helicid snails in California. Calif Dep AgricBull 32:173–187.
Gillespie J.P., M.R. Kanost, and T. Trenezek. 1997. Biologicalmediators of insect immunity. Annu Rev Entomol 42:611–643.
Gomot A. 1990. Photoperiod and temperature interaction inthe determination of reproduction of the edible snail, Helixpomatia. J Reprod Fertil 90:581–585.
Graham A.L. 2001. Use of an optimality model to solve theimmunological puzzle of concomitant infection. Q Rev Biol77:409–434.
Graham A.L., D.M. Shuker, L.C. Pollitt, K.J.R. Stuart, A.J. Wil-son, and T.J. Little. 2011. Fitness consequences of immuneresponses: strengthening the empirical framework for eco-immunology. Funct Ecol 25:5–17.
Hamilton R., M. Siva-Jothy, and M. Boots. 2008. Two arms arebetter than one: parasite variation leads to combined in-ducible and constitutive innate immune response. Proc RSoc B 275:937–945.
Heller J. 2001. Life history strategies. Pp. 417–446 in G.M.Barker, ed. The biology of terrestrial mollusks. CABI, NewYork.
Hennemann W.W. 1983. Relationship among body-mass, met-abolic-rate and the intrinsic rate of natural increase in mam-mals. Oecologia 56:104–108.
Hoffmann J.A. 2003. The immune response of Drosophila. Na-ture 426:33–38.
Hoffmann J.A., J.M. Reichhart, and C. Hetru. 1996. Innateimmunity in higher insects. Curr Opin Immunol 8:8–13.
Houdijk J.G., I. Kyriazakis, R.L. Coop, and F. Jackson. 2001.The expression of immunity to Teladorsagia circumcincta inewes and its relationship to protein nutrition depend on bodyprotein reserves. Parasitology 122:661–672.
Jacot A., H. Scheuber, J. Kurtz, and M.W.G. Brinkhof. 2005.Juvenile immune system activation induces a costly upreg-ulation of adult immunity in field crickets Gryllus campestris.Proc R Soc B 272:63–69.
This content downloaded on Sun, 24 Feb 2013 23:14:31 PMAll use subject to JSTOR Terms and Conditions

Immune Patterns and Life History in Land Snails 203
Kerney M.P. and R.A.D. Cameron. 1979. A field guide to theland snails of Britain and north-west Europe. Collins,London.
Klasing K.C. and T. Leshchinsky. 1999. Functions, costs, andbenefits of the immune system during development andgrowth. Ostrich 69:2817–2832.
Lazzaro B.P. and T.J. Little. 2009. Immunity in a variable world.Philos Trans R Soc B 364:15–26.
Lee K.A. 2006. Linking immune defenses and life history at thelevels of the individual and the species. Integr Comp Biol46:1000–1015.
Ligaszewski M., S. Krysztof, and J. Stekla. 2009. The shell fea-tures of Cornu aspersum (synonym Helix aspersa) and Helixpomatia: characteristics and comparison. Am Malacol Bull27:173–181.
Livingstone D.R., P. Garcia Martinez, X. Michel, J.F. Narbonne,S. O’Hara, D. Ribera, and G.W. Winston. 1990. Oxyradicalproduction as a pollution-mediated mechanism of toxicityin the common mussel, Mytilus edulis L., and other mollusks.Funct Ecol 4:415–424.
Lochmiller R.L. and C. Deerenberg. 2000. Trade-offs in evo-lutionary immunology: just what is the cost of immunity?Oikos 88:87–98.
Madec L., A. Bellido, and A. Guiller. 2003. Shell shape of theland snail Cornu aspersum in North Africa: unexpected evi-dence of a phylogeographical splitting. Heredity 91:224–231.
Madec L. and J. Daguzan. 1993. Geographic variation in re-productive traits of Helix aspersa Muller studied under lab-oratory conditions. Malacologia 35:99–117.
Madec L., C. Desbuquois, and M.A. Coutellec. 2000. Pheno-typic plasticity in reproductive traits: importance in the lifehistory of Helix aspersa (Mollusca: Helicidae) in a recentlycolonized habitat. Biol J Linn Soc 69:25–39.
Madec L., A. Guiller, M.A. Coutellec, and C. Desbuqois. 1998.Size fecundity relationships in the land snail Helix aspersa:preliminary results on a form outside the norm. InvertebrReprod Dev 34:83–90.
Medley G.F. 2002. The epidemiological consequences of opti-mization of the individual host immune response. Parasi-tology 125:561–570.
Moret Y. 2003. Explaining variable costs of the immune re-sponse: selection for specific versus non-specific immunityand facultative life history change. Oikos 102:213–216.
Moret Y. and P. Schmid-Hempel. 2000. Survival for immunity:the price of immune system activation for bumblebee work-ers. Science 290:1166–1168.
———. 2001. Immune defence in bumble-bee offspring. Na-ture 414:506.
Ottaviani E., E. Caselgrandi, A. Franchini, and C. Franceschi.1993. CRF provokes the release of norepinephrine by he-mocytes of Viviparus ater (Gastropoda, Prosobranchia): fur-ther evidence in favour of the evolutionary hypothesis of the“mobile immune-brain.” Biochem Biophys Res Comm 193:446–452.
Owen I.P.F. and K. Wilson. 1999. Immunocompetence: a ne-
glected life history trait or conspicuous red herring? TrendsEcol Evol 14:170–172.
Pathak J.P.N. 1986. Haemogram and its endocrine control ininsects. Pp. 49–59 in M. Brehelin, ed. Immunity in inver-tebrates. Springer, Berlin.
Phillipp E.E.R. and D. Abele. 2010. Masters of longevity: lessonsfrom long-lived bivalves: a mini-review. Gerontology 56:55–65.
Pipe R.K., J.A. Coles, and S.R. Farley. 1995. Assays for mea-suring immune response in the mussel Mytilus edulis. TechFish Immunol 1995:93–100.
Pollard E. 1975. Aspects of the ecology of Helix pomatia L. JAnim Ecol 44:305–329.
Rantala M.J. and D.A. Roff. 2005. An analysis of trade-off inimmune function, body size and development time in theMediterranean field cricket, Gryllus bimaculatus. Funct Ecol19:323–330.
Ratanarat-Brockelman C. 1977. Isolation of nematode inhibitorfrom hemolymph of the snail Helix aspersa (Muller). I. Evi-dence for opsonic factor(s) in serum. Immunology 17:437–443.
Renwrantz L. 1979. Eine Untersuchung molecular und zellularBestandteile der Hamolymphe von Helix pomatia unter be-sonderer Berucksichtigung immunobiologish aktiver Kom-ponenten. Zool Jahrb Physiol 83:283–333.
Rolff J. and M.T. Siva-Jothy. 2003. Invertebrate ecological im-munology. Science 301:472–475.
Russo J. and L. Madec. 2011. Dual strategies for immune de-fence in the land snail Cornu aspersum (Gastropoda, Pul-monata). Physiol Biochem Zool 80:212–221.
Sacchi C.F. 1971. Ecologie comparee des Gasteropodes Pul-mones des dunes mediterraneennes et atlantiques. Natura62:277–358.
Sadd B.M. and P. Schmid-Hempel. 2009. Principles of ecolog-ical immunology. Evol Appl 2:113–121.
Schmid-Hempel P. 2001. On the evolutionary ecology of host-parasite interactions: addressing the question with regard tobumblebees and their parasites. Naturwissenschaften 88:147–158.
———. 2003. Variation in immune defense as a question ofevolutionary ecology. Proc R Soc B 270:357–366.
Schulenburg H., J. Kurtz, Y. Moret, and M.T. Siva-Jothy. 2009.Introduction: ecological immunology. Philos Trans R Soc B364:3–14.
Sheldom B.C. and S. Verhulst. 1996. Ecological immunity:costly parasite defenses and trade-offs in evolutionary ecol-ogy. Trends Ecol Evol 11:317–321.
Stearns S.C. 1989. Trade-offs in life-history evolution. FunctEcol 3:259–268.
Storey K.B. 1996. Oxidative stress: animal adaptations in nature.Braz J Med Biol Res 29:1715–1733.
Tella J.L., A. Sheuerlein, and R.E. Ricklefs. 2002. Is cell-mediatedimmunity related to the evolution of life-history strategiesin birds? Proc R Soc B 269:1059–1066.
Thioulouse J. and S. Dray. 2007. Interactive multivariate data
This content downloaded on Sun, 24 Feb 2013 23:14:31 PMAll use subject to JSTOR Terms and Conditions

204 J. Russo and L. Madec
analysis in R with the ade4 and ade4TkGUI packages. J StatSoftw 22:1–14.
Tieleman B.I., J.B. Williams, R.E. Ricklefs, and K.C. Klasing.2005. Constitutive innate immunity is a component of thepace-of-life syndrome in tropical birds. Proc R Soc B 272:1715–1720.
Viney M.E., E.M. Riley, and K.L. Buchanan. 2005. Optimalimmune responses: immunocompetence revisited. TrendsEcol Evol 20:1–5.
Wiegel F.W. and A.S. Perelson. 2004. Some scaling principlesfor the immune system. Immunol Cell Biol 82:127–131.
Wilfert L., J. Gadau, and P. Schmid-Hempel. 2007. The genetic
architecture of immune defense and reproduction in maleBombus terrestris bumblebees. Evolution. 61:804–815.
Wojtaszek J., A. Poloczek-Adamowick, A. Adamowick, U. Fuks,and A. Dzugaj. 1998. Cytomorphometry and seromucoidconcentration in the hemolymph of selected snails species.Zool Pol 43:87–101.
Wolda H. and D.A. Kreulen. 1973. Ecology of some experi-mental populations of the land snail Cepaea nemoralis (L).II. Production and survival of eggs and juveniles. Neth J Zool23:168–188.
Zera A.J. and L.G. Harshman. 2001. The physiology of lifehistory trade-offs in animals. Annu Rev Ecol Syst 32:95–126.
This content downloaded on Sun, 24 Feb 2013 23:14:31 PMAll use subject to JSTOR Terms and Conditions





























