Embed Size (px)
Citation preview

Lipopeptide structure determines TLR2 dependent cellactivation levelUte Buwitt-Beckmann1, Holger Heine1, Karl-Heinz Wiesmuller2, Gunther Jung3, Roland Brock4
and Artur J. Ulmer1
1 Department of Immunology and Cell Biology, Research Center Borstel, Borstel, Germany
2 EMC microcollections GmbH, Tubingen, Germany
3 Institute of Organic Chemistry, University of Tubingen, Tubingen, Germany
4 Department of Molecular Biology, Institute for Cell Biology, University of Tubingen, Tubingen, Germany
Lipoproteins ⁄peptides (LP) are major constituents of
the cell wall of bacteria. Bacterial LP are di-O-acylated-
S-(2,3-dihydroxypropyl)-cysteinyl residues N-terminally
coupled to distinct polypeptides, as found in the
macrophage-activating lipopeptide from Mycoplasma
fermentans (MALP2) [1] and in LP from the N termi-
nus of the cytochrome subunit of the photoreaction
center of Rhodopseudomonas viridis [2]. The S-(2,3-di-
hydroxypropyl)-cysteine may be N-acylated with a
third fatty acid via an amide-linkage as is the case for
the LP from the cell wall of Escherichia coli [3] and
numerous other Gram-negative bacteria [4]. These LP
activate the innate immune system and promote the
formation of adaptive immunity as an adjuvant during
stimulation with specific antigens.
The receptor responsible for a functional recognition
of LP by cells is the Toll-like receptor 2 (TLR2) [5,6].
TLRs are prominent pattern recognition receptors
of the innate immune system recognizing various
invading microorganisms by conserved molecular
Keywords
TLR2; lipopeptides; ligand recognition;
structure–activity relationship
Correspondence
A. J. Ulmer, Research Center Borstel,
Parkallee 22, 23845 Borstel, Germany
Tel: +49 4537 188448
Fax: +49 4537 188435
E-mail: [email protected]
(Received 19 August 2005, revised 13
October 2005, accepted 20 October 2005)
doi:10.1111/j.1742-4658.2005.05029.x
Bacterial lipoproteins ⁄peptides are composed of di-O-acylated-S-(2,3-dihyd-
roxypropyl)-cysteinyl residues N-terminally coupled to distinct polypep-
tides, which can be N-acylated with a third fatty acid. Using a synthetic
lipopeptide library we characterized the contribution of the lipid portion to
the TLR2 dependent pattern recognition. We found that the two ester
bound fatty acid length threshold is beyond eight C atoms because almost
no response was elicited by cellular challenge with analogues carrying shor-
ter acyl chains in HEK293 cells expressing recombinant human TLR2. In
contrast, the amide bound fatty acid is of lesser importance. While two
ester-bound palmitic acids mediate a high stimulatory activity of the
respective analogue, a lipopeptide carrying one amide-bound and another
ester-bound palmitic acid molecule was inactive. In addition, species speci-
fic LP recognition through murine and human TLR2 depended on the
length of the two ester bound fatty acid chains. In conclusion, our results
indicate the responsibility of both ester bound acyl chains but not of the
amide bound fatty acid molecule for the TLR dependent cellular recogni-
tion of canonical triacylated LP, as well as a requirement for a minimal
acyl chain length. Thus they might support the explanation of specific
immuno-stimulatory potentials of different microorganisms and provide a
basis for rational design of TLR2 specific adjuvants mediating immune
activation to distinct levels.
Abbreviations
BbMALP2, macrophage-activating lipopeptide from Mycoplasma fermentans; LP, lipoproteins ⁄ peptides; huTLR2, human TLR2; muTLR2,
murine TLR2; IL, interleukin.
6354 FEBS Journal 272 (2005) 6354–6364 ª 2005 The Authors Journal compilation ª 2005 FEBS

structures, so called pathogen-associated molecular
patterns [7]. To date 11 TLRs have been described.
The receptors differ not only in ligand specificity but
also in their expression pattern on different cells of the
innate immune system. In addition, distinct TLRs are
able to induce the expression of different sets of
inflammatory target genes [8]. TLR2 recognizes the
most diverse set of molecular structures, including,
lipoteichoic acid, lipoarabinomanan, bacterial LP, as
well as molecules from yeast, spirochetes and fungi [8].
Unlike other TLRs, which are functionally active as
homomers [9], TLR2 forms heteromers either with
TLR1 or TLR6 [9] to attain specificity for a given sti-
mulus [10–13]. The previous concept, which has stated
that di-acylated LP like MALP2 signals through
TLR2 ⁄ 6, whereas tri-acylated LP, as realized in tri-
palmitoyl-S-(2,3-dihydroxypropyl)-cysteinyl-seryl-tetra-
lysine (Pam3C-SK4), signals through TLR2 ⁄ 1, has
recently been challenged by the finding that Pam2C-
SK4 as well as MALP2-SK4 is recognized by TLR2
in a TLR6-independent manner [14]. The molecular
mechanism of the recognition of LP by TLR2 is not
clear yet. Recently it was demonstrated that binding
of Pam3C-SK4 to the LPS-binding protein, and
soluble- or membrane-bound CD14 results in spatial
proximity of LP, CD14, TLR2 and TLR1 [15–17] and
subsequent TLR2 signalling.
Among the TLR2-dependent bacterial ligands, LP
are primary candidates for analysing the structural
requirements for TLR2 activation. Firstly, these
molecules are available from various bacterial species
expressing different molecular structures. Secondly,
chemical synthesis provides a vast variety of analytic-
ally well defined LP analogues [18,19]. Although the
structural requirements for optimal adjuvant activity
of synthetic LP have been studied in detail [20], the
structure–activity relationships for the recognition of
LP by TLR2 are still poorly defined. Recently we
have systematically investigated the contribution of
all proteinogenic amino acids except cysteine in the
peptide moiety of Pam3Cys-lipohexapeptides. Ninety-
five LP amide subcollections were synthesized by
combinatorial peptide chemistry. None of these lipo-
hexapeptides subcollections substantially exceeded the
biological activity of Pam3C-SK4 indicating that the
biological activity is more or less independent of
peptide sequences in this LP library [19]. However,
some exceptions from these general results have been
found (Wiesmuller et al. unpublished data). Concern-
ing the contribution of the fatty acids in TLR2-
dependent signalling evidence that the fatty acid
composition has an impact on the biological activity
has been presented [21]. For this reason we have
now systematically investigated the structure-activity
relationships for the lipid moiety of LP, tightly
focusing on recognition by TLR2. LP libraries were
synthesized having different fatty acid compositions
at the three reactive groups of the LP-scaffold,
namely the ester-bound fatty acids and the amide-
bound fatty acids. We could demonstrate that the
amide-bound fatty acids contribute, if at all, only
minutely to the recognition by TLR2. In contrast,
long chain ester-bound fatty acids are essential for
the induction of signalling through TLR2. Interest-
ingly, a longer chain-length of the ester-bound fatty
acids is necessary for the activation of huTLR2 in
comparison to muTLR2, revealing a species specificity
of LP recognition.
Results and discussion
The structure-activity relationships in the recognition
of TLR2 by various bacterial LP have been poorly
defined so far. In particular, little information exists
on the relevance of the fatty acid composition and dis-
tribution within the LP head group for TLR2 activa-
tion. In order to define the structural requirements for
the lipid moiety of LP during TLR2 activation, inde-
pendent from a coreceptor bias, HEK293 cells trans-
fected with either the huTLR2 or muTLR2 were
selected as a cellular test system. These cells express
both TLR1 and TLR6. At first it should be noted that
all bioactive LP used in our study were found to be
TLR2 dependent: There was no response to these LP
in nontransfected HEK293 cells, cells transfected with
an empty vector, nor in TLR2-deficient mice (data not
shown).
In a first set of experiments a LP collection was tes-
ted that was composed of synthetic LP analogues of
Pam3C-SK4 in which the amide-bound as well as the
two ester-bound fatty acids were modified. The amide-
bound fatty acids in our collection were Hex, Pe, Dec,
Myr, and Pam and the ester- bound fatty acids Ac,
Hex, Oct, or Pam (for abbreviations of the fatty acids
see Table 1). Due to the synthesis protocol for this
lipopeptide collection [22], both ester-bound fatty acids
were identical. HEK293 cells were transfected with
huTLR2, stimulated with 1000 nm of LP and the
release of interleukin (IL)-8 in the culture supernatant
was determined after 24 h of culture. Our results
clearly indicate that the amide-bound fatty acids have,
if at all, only a minute effect on the TLR2-mediated
IL-8 release: no significant differences in the response
of cells stimulated with LP containing N-bound Hex,
Pe, Dec, Myr, or Pam acyl residues were observed
(Table 2).
U. Buwitt-Beckmann et al. TLR2 recognition by lipopeptides
FEBS Journal 272 (2005) 6354–6364 ª 2005 The Authors Journal compilation ª 2005 FEBS 6355

Previously it had been postulated that the presence
or absence of an N-bound fatty acid is responsible
for converting a TLR2 ⁄TLR6 dependent LP into a
TLR2 ⁄TLR1 dependent LP. However, our recent
results, showing that Pam2C-SK4 and also MALP2-
SK4 (the elongated MALP2 analogue Pam2C-
GNNDESNISFKEKSK4) are TLR6-independent LP,
required the rejection of this hypothesis. At least all
LP with a SK4-peptid tail might be recognized by
TLR2 in a TLR6 independent manner. It therefore
seems reasonable to assume that all the LP analogues
of this collection are TLR6 independent. Whether
these TLR2 agonists signal in a TLR1-dependent man-
ner remains to be investigated. It should be noted,
however, that even the well-studied reference lipohexa-
peptide Pam3C-SK4 exerts not only TLR1-dependent
bioactivity, but also, at least in part, a TLR1-inde-
pendent activity [12,14].
In contrast to the amide-bound fatty acids, the iden-
tity of the ester-bound fatty acids is of great relevance
for the TLR2-dependent response of the cells. For
fatty acids with a short chain length (Ac or Hex) only
a minute or no response could be observed. The
response was slightly enhanced after stimulation with a
LP having ester-bound octanoic acids [at least with
respect to nuclear factor jB (NFjB) translocation] andwas maximal after stimulation with palmitoylated LP
analogues. The same structure-activity relationship for
the stimulation of HEK293-huTLR2 cells was also
observed, when the activation of NFjB, as determined
in a luciferase-reporter assay, instead of IL-8 release
was estimated (Table 2). This indicates that the nature
of the ester-bound but not the amide-bound fatty acids
of LP is of great relevance throughout from TLR2-
induced signal transduction up to cytokine release.
To obtain more detailed information on the dose–
response function of lipopeptides, for selected synthetic
LP analogues the IL-8 release by HEK293-huTLR2
cells was determined over a concentration range of
1-1000 nm. As shown in Fig. 2A, all four tested lipo-
hexapeptide analogues with two ester-bound Pam
moieties exhibited nearly the same dose–response
characteristic, independently from the nature of the
N-bound acyl moiety. On the other hand, HEK293-
huTLR2 were unresponsive to all lipohexapeptide
Table 1. Abbreviation, structure and name of fatty acids used in
this study.
AC C2H4O2 Acetic acid
Hex C6 H12 O2 Hexanoic acid
BA C7 H6 O2 Benzoic acid
Pe C8 H8 O2 Phenylacetic acid
Oct C8 H16 O2 Octanoic acid
Pel C9 H18 O2 Nonanoic acid
Dec C10 H20 O2 Decanoic acid
Dod C12 H24 O2 Dodecanoic acid
Myr C14 H28 O2 Tetradecanoic acid ⁄myristic acid
Pam C16 H32 O2 Hexadecanoic acid ⁄ palmitic acid
Lin C18 H32 O2 Octadeca-9,12E-dienoic acid
Ole C18 H34 O2 Octadeca-9E-enoic acid
Ara C20 H40 O2 Eicosanoic acid
Table 2. Stimulation of HEK-huTLR2 by various synthetic lipohexapeptide analogues having different O- and N-acylated head groups. The LP
analogues were used at a concentration of 1000 nM. IL-8 release was measured after a culture period of 24 h, the Luciferase-reporter assay
for NFjB was performed after 6 h of stimulation. The results are expressed as percentage response in relation to the response to
Pam3CSK4. Each value represents the mean of triplicates.
IL-8 release
HexAc2C-SK4 0% PeAc2C-SK4 0% DecAc2C-SK4 1% PamAc2C-SK4 2% MyrAc2C-SK4 1%
HexHex2C-SK4 0% PeHex2C-SK4 0% DecHex2C-SK4 0% PamHex2C-SK4 1% MyrHex2C-SK4 1%
HexOct2C-SK4 0% PeOct2C-SK4 0% DecOct2C-SK4 1% PamOct2C-SK4 2% MyrOct2C-SK4 3%
HexPam2C-SK4 79% PePam2C-SK4 91% DecPam2C-SK4 97% PamPam2C-SK4 100% MyrPam2C-SK4 156%
Luciferase-reporter assay for NFjB
HexAc2C-SK4 14% PeAc2C-SK4 1% DecAc2C-SK4 24% PamAc2C-SK4 22% MyrAc2C-SK4 26%
HexHex2C-SK4 2% PeHex2C-SK4 2% DecHex2C-SK4 3% PamHex2C-SK4 10% MyrHex2C-SK4 2%
HexOct2C-SK4 14% PeOct2C-SK4 17% DecOct2C-SK4 20% PamOct2C-SK4 2% MyrOct2C-SK4 51%
HexPam2C-SK4 111% PePam2C-SK4 103% DecPam2C-SK4 119% PamPam2C-SK4 100% MyrPam2C-SK4 145%
Fig. 1. Structures and denotations of some synthetic LP analogues
used in this study. To illustrate the structure denotation of LP ana-
logues used in this paper, the structures of three synthetic LP ana-
logues are given. The chemical structure of the scaffold with the
four reactive groups is shown. R1 represents the amide-bound fatty
acid, R2 and R3 represent the ester-bound fatty acids, and R4 repre-
sents the peptide moiety.
TLR2 recognition by lipopeptides U. Buwitt-Beckmann et al.
6356 FEBS Journal 272 (2005) 6354–6364 ª 2005 The Authors Journal compilation ª 2005 FEBS

analogues carrying two ester bound hexanoic acids.
This lack of activity was independent from the nature
of the acyl moiety, i.e. short or long chain length
(Fig. 2B). In addition, for a series of LP analogues
with an increasing chain length of the two ester bound
fatty acids, no response in HEK293-huTLR2 cells was
detected for Ac, Hex, or Oct but only for Pam moiet-
ies. This structure–activity relationship again was the
same for a series of LP with a short (Hex) or a long
(Myr) amide-bound fatty acid (Fig. 3A and B). In
summary, all LP analogues with short O-acylated fatty
acids (Ac, Hex, or Oct) were found to be inactive for
HEK293-huTLR2. Moreover, the amide-bound fatty
acid is of low or even no relevance for the TLR2-medi-
ated stimulation of HEK-huTLR2 cells.
The results, presented so far, indicate that LP ana-
logues with two ester-bound fatty acids having a short
chain length of eight carbons or less are incapable of
activating cells through TLR2, whereas a chain length
of 16 carbons, such as that present in palmitic acid is
optimal. Therefore, we decided to analyse the impact
of the acyl chain length as a specific aspect of TLR2
dependent LP recognition. For these investigations a
further collection of LP with a SSNASK4-peptide moi-
ety was synthesized and characterized by electro-MS.
It should be noted that the dose kinetic of the
huTLR2 dependent cellular response, revealed upon
application of lipohexapeptide Pam3C-SK4, largely
resembled that obtained upon challenge with the lipo-
decapeptide Pam3C-SSNASK4 (data not shown). For
these analogues with a longer peptide moiety the stim-
ulatory activity was a function of the chain length of
the two ester-bound fatty acids as well. Only a margi-
nal response was found for the LP analogues having
ester-bound decanoyl (10 carbon atoms) moieties or
acyl residues with less numbers of carbons. Ester-
bound fatty acids with at least 14 carbon atoms (Myr)
were necessary to obtain an optimal response
(Table 3). Activation of IL-8 release in HEK293-huT-
LR2 cells again showed the same structure–activity
relationships as activation of the translocation of
NFjB (Table 3). These results obtained with LP at a
fixed concentration of 1000 nm were again confirmed
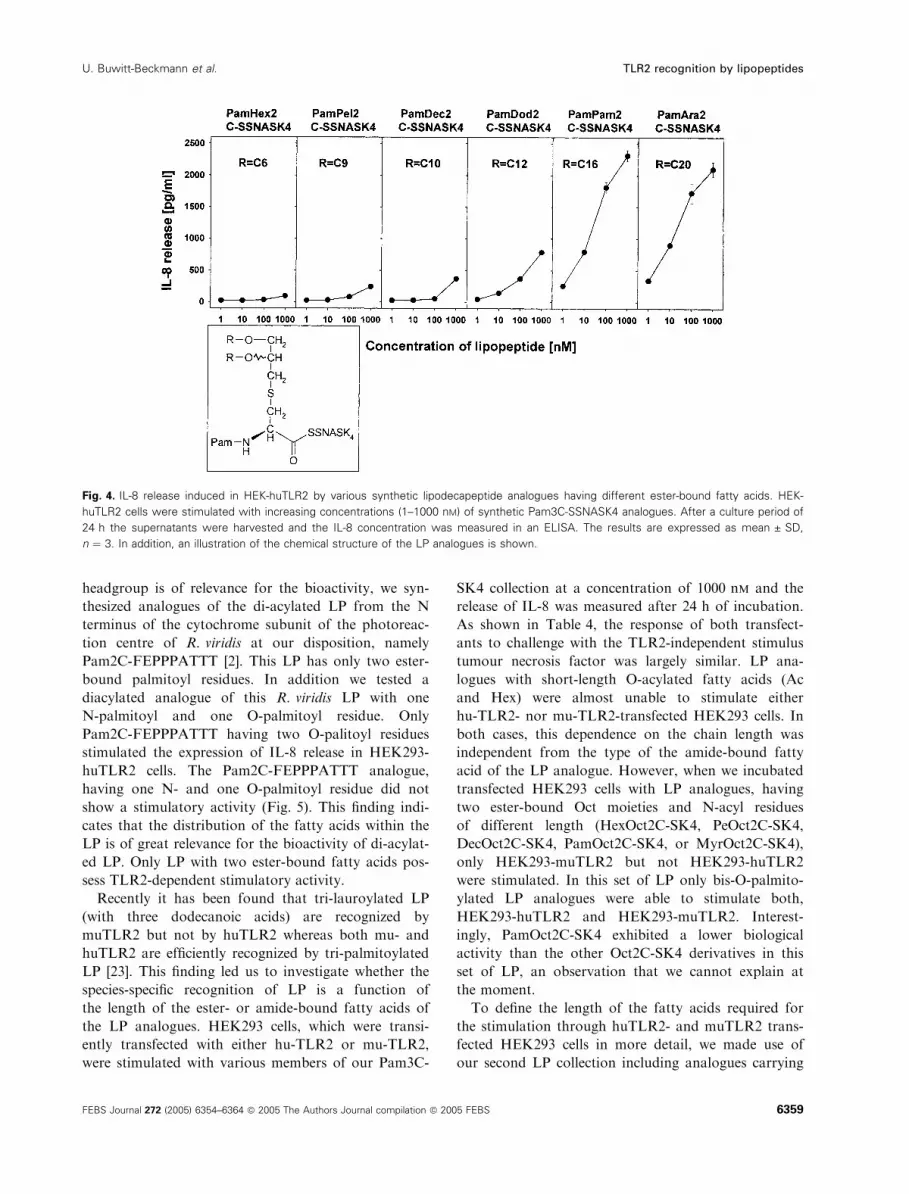
by the acquisition of dose–response curves (Fig. 4). It
should be noted that a LP with ester-bound Ara moi-
eties had nearly the same agonistic activity as a LP
Fig. 2. IL-8 release induced in HEK-huTLR2 by various synthetic LP analogues having differently N-acylated cysteine residues. HEK-huTLR2
cells were stimulated with increasing concentrations (1–1000 nM) of synthetic LP analogues. Results obtained for (A) bis-O-palmitoylated
lipohexapeptide analogues and (B) bis-O-hexanoylated lipohexapeptide analogues having different N-acylated head groups. After a culture
period of 24 h the supernatants were harvested and the IL-8 concentration was measured by an ELISA. The results are expressed as
mean ± SD, n ¼ 3. The chemical structures of the LP analogues are included.
U. Buwitt-Beckmann et al. TLR2 recognition by lipopeptides
FEBS Journal 272 (2005) 6354–6364 ª 2005 The Authors Journal compilation ª 2005 FEBS 6357

with ester-bound Pam moieties. Therefore, it can be
concluded that a LP needs a minimum chain length of
the ester-bound fatty acids for optimal response. How-
ever, this response cannot be increased further by an
elongation of the ester-bound fatty acids.
Hitherto we could conclude that only LP with at
least two long-chain ester-bound fatty acids, such as
Myr or Pam, are able to signal through huTLR2. To
scrutinize whether the positional distribution of the two
fatty acids within the S-(2,3-dihydroxypropyl)-cysteinyl
Table 3. Stimulation of HEK-huTLR2 by various synthetic lpodecapeptide analogs having different ester-bound fatty acids. The LP analogues
were used at a concentration of 1000 nM. IL-8 release was measured after a culture period of 24 h, the Luciferase-reporter assay for NFjB
was performed after 6 h of stimulation. Each value represents the mean of triplicates.
IL-8 release (% in relation to Pam3C-SSNASK4) Luciferase-reporter assay for NFjB
PamHex2C-SSNASK4 2% C6a) PamHex2C-SSNASK4 13%
PamBa2C-SSNASK4 4% C7 PamBa2C-SSNASK4 4%
PamPe2C-SSNASK4 0% C8 PamPe2C-SSNASK4 8%
PamOct2C-SSNASK4 6% C8 PamOct2C-SSNASK4 15%
PamPel2C-SSNASK4 8% C9 PamPel2C-SSNASK4 15%
PamDec2C-SSNASK4 3% C10 PamDec2C-SSNASK4 29%
PamDod2C-SSNASK4 37% C12 PamDod2C-SSNASK4 69%
Pam Myr2C-SSNASK4 88% C14 Pam Myr2C-SSNASK4 126%
PamPam2C-SSNASK4 100% C16 PamPam2C-SSNASK4 100%
PamOle2C-SSNASK4 119% C18 PamOle2C-SSNASK4 125%
PamLin2C-SSNASK4 101% C18 PamLin2C-SSNASK4 165%
PamAra2C-SSNASK4 91% C20 PamAra2C-SSNASK4 186%
aChain length of O-acylated fatty acids.
Fig. 3. IL-8 release induced in HEK-huTLR2 by various synthetic lipohexapeptide analogues having different O-acylated head groups.
HEK-huTLR2 cells were stimulated with increasing concentrations (1–1000 nM) of synthetic LP analogues. Results obtained for (A)
HexPam2C-SK4 and (B) MyrPam2C-SK4 analogues having different ester-bound fatty acids. After a culture period of 24 h the supernatants
were harvested and the IL-8 concentration was measured in an ELISA. The results are expressed as mean ± SD, n ¼ 3. The chemical struc-
tures of the LP analogs are included.
TLR2 recognition by lipopeptides U. Buwitt-Beckmann et al.
6358 FEBS Journal 272 (2005) 6354–6364 ª 2005 The Authors Journal compilation ª 2005 FEBS
headgroup is of relevance for the bioactivity, we syn-
thesized analogues of the di-acylated LP from the N
terminus of the cytochrome subunit of the photoreac-
tion centre of R. viridis at our disposition, namely
Pam2C-FEPPPATTT [2]. This LP has only two ester-
bound palmitoyl residues. In addition we tested a
diacylated analogue of this R. viridis LP with one
N-palmitoyl and one O-palmitoyl residue. Only
Pam2C-FEPPPATTT having two O-palitoyl residues
stimulated the expression of IL-8 release in HEK293-
huTLR2 cells. The Pam2C-FEPPPATTT analogue,
having one N- and one O-palmitoyl residue did not
show a stimulatory activity (Fig. 5). This finding indi-
cates that the distribution of the fatty acids within the
LP is of great relevance for the bioactivity of di-acylat-
ed LP. Only LP with two ester-bound fatty acids pos-
sess TLR2-dependent stimulatory activity.
Recently it has been found that tri-lauroylated LP
(with three dodecanoic acids) are recognized by
muTLR2 but not by huTLR2 whereas both mu- and
huTLR2 are efficiently recognized by tri-palmitoylated
LP [23]. This finding led us to investigate whether the
species-specific recognition of LP is a function of
the length of the ester- or amide-bound fatty acids of
the LP analogues. HEK293 cells, which were transi-
ently transfected with either hu-TLR2 or mu-TLR2,
were stimulated with various members of our Pam3C-
SK4 collection at a concentration of 1000 nm and the
release of IL-8 was measured after 24 h of incubation.
As shown in Table 4, the response of both transfect-
ants to challenge with the TLR2-independent stimulus
tumour necrosis factor was largely similar. LP ana-
logues with short-length O-acylated fatty acids (Ac
and Hex) were almost unable to stimulate either
hu-TLR2- nor mu-TLR2-transfected HEK293 cells. In
both cases, this dependence on the chain length was
independent from the type of the amide-bound fatty
acid of the LP analogue. However, when we incubated
transfected HEK293 cells with LP analogues, having
two ester-bound Oct moieties and N-acyl residues
of different length (HexOct2C-SK4, PeOct2C-SK4,
DecOct2C-SK4, PamOct2C-SK4, or MyrOct2C-SK4),
only HEK293-muTLR2 but not HEK293-huTLR2
were stimulated. In this set of LP only bis-O-palmito-
ylated LP analogues were able to stimulate both,
HEK293-huTLR2 and HEK293-muTLR2. Interest-
ingly, PamOct2C-SK4 exhibited a lower biological
activity than the other Oct2C-SK4 derivatives in this
set of LP, an observation that we cannot explain at
the moment.
To define the length of the fatty acids required for
the stimulation through huTLR2- and muTLR2 trans-
fected HEK293 cells in more detail, we made use of
our second LP collection including analogues carrying
Fig. 4. IL-8 release induced in HEK-huTLR2 by various synthetic lipodecapeptide analogues having different ester-bound fatty acids. HEK-
huTLR2 cells were stimulated with increasing concentrations (1–1000 nM) of synthetic Pam3C-SSNASK4 analogues. After a culture period of
24 h the supernatants were harvested and the IL-8 concentration was measured in an ELISA. The results are expressed as mean ± SD,
n ¼ 3. In addition, an illustration of the chemical structure of the LP analogues is shown.
U. Buwitt-Beckmann et al. TLR2 recognition by lipopeptides
FEBS Journal 272 (2005) 6354–6364 ª 2005 The Authors Journal compilation ª 2005 FEBS 6359

the longer SSNASK4 peptide chain. N-Palmitoylated
lipodecapeptides with short length bis-O-acylated head
groups (PamHex2C-SSNASK4, PamBa2C-SSNASK4,
and PamPe2C-SSNASK4) did not stimulate HEK293-
huTLR2 and HEK293-muTLR2 cells (Table 5). The
stimulatory activity of PamOct2C-SSNASK4, Pam-
Pel2C-SSNASK4, and PamDec2C-SSNASK4 was low
or moderate in HEK293-muTLR2 cells but was absent
in HEK293-huTLR2 cells. A strong response of
HEK293-muTLR2 cells was observed upon stimula-
tion with PamDod2C-SSNASK4, whereas responsive-
ness of HEK293-huTLR2 to this LP was low.
N-palmitoylated LP carrying long length ester-bound
carboxylic acids (PamMyr2C-SSNASK4, PamOle2C-
SSNASK4, PamLin2C-SSNASK4, and PamAra-
Hex2C-SSNASK4), all exhibited an almost similar
high stimulatory activity in both, HEK293-huTLR2
and HEK293-muTLR2 cells. Taken together, from
these results we conclude that the species-specific
recognition of LP is not a function of the length of
the N-fatty acyl, but of the O-fatty acyl groups of LP
analogues.
Table 4. The response of HEK-huTLR2 and HEK-muTLR2 to lipo-
hexapeptide analogues with various amide- and ester-bound fatty
acids. The lipopeptides were used at a concentration of 1000 nM,
tumour necrosis factor was used at a concentration of 10 ngÆmL)1.
The results are expressed as IL-8 release (pgÆmL)1), mean ± SD,
n ¼ 3.
Control 0 ± 0 11 ± 11
Tumour necrosis factor 1494 ± 120 1216 ± 9
HexAc2C-SK4 1 ± 1 68 ± 16
HexHex2C-SK4 0 ± 0 47 ± 14
HexOct2C-SK4 0 ± 0 1404 ± 167
HexPam2C-SK4 1689 ± 298 4281 ± 152
PeAc2C-SK4 0 ± 0 0 ± 0
PeHex2C-SK4 9 ± 9 0 ± 0
PeOct2C-SK4 2 ± 2 1744 ± 193
PePam2C-SK4 1940 ± 108 3418 ± 193
DecAc2C-SK4 27 ± 10 290 ± 1
DecHex2C-SK4 0 ± 0 83 ± 8
DecOct2C-SK4 23 ± 6 2705 ± 41
DecPam2C-SK4 2072 ± 137 3321 ± 269
PamAc2C-SK4 32 ± 1 76 ± 2
PamHex2C-SK4 14 ± 3 146 ± 11
PamOct2C-SK4 34 ± 7 564 ± 15
PamPam2C-SK4 2126 ± 378 2876 ± 99
MyrAc2C-SK4 21 ± 2 19 ± 19
MyrHex2C-SK4 15 ± 5 0 ± 0
MyrOct2C-SK4 62 ± 8 2111 ± 160
MyrPam2C-SK4 3320 ± 256 2551 ± 340
Table 5. The response of HEK-huTLR2 and HEK-muTLR2 to lipo-
decapeptide analogues with various ester-bound fatty acids. The LP
analogs were used at a concentration of 1000 nM. IL-8 release was
measured after a culture period of 24 h. Each value represents the
mean ± SD of triplicates.
Structure
IL-8 release (pgÆmL)1)
hu-TLR2 mu-TLR2
PamHex2C-SSNASK4 0 ± 0 10 ± 10
PamBa2C-SSNASK4 0 ± 0 3 ± 3
PamPe2C-SSNASK4 0 ± 0 10 ± 3
PamOct2C-SSNASK4 16 ± 2 331 ± 35
PamPel2C-SSNASK4 4 ± 4 133 ± 4
PamDec2C-SSNASK4 31 ± 5 1031 ± 40
PamDod2C-SSNASK4 164 ± 11 2632 ± 431
Pam Myr2C-SSNASK4 1286 ± 341 2224 ± 147
PamOle2C-SSNASK4 1319 ± 104 1398 ± 108
PamLin2C-SSNASK4 2086 ± 279 2689 ± 75
PamAra2C-SSNASK4 1948 ± 57 3340 ± 13
Fig. 5. Response of HEK-huTLR2 cells to doubly palmitoylated synthetic LP analogues from R. viridis having a different positional distribution
of the two fatty acids. HEK-huTLR2 cells were stimulated with rising concentrations (1–1000 nM) of two synthetic Pam2C-FEPPPATTT ana-
logues, which only differ in the positions of the two Pam. There are either two ester-bound Pam or one esterfied Pam and one amidated
Pam as indicated. After a culture period of 24 h the supernatants were harvested and the IL-8 concentration was measured in an ELISA.
The results are expressed as mean ± SD, n ¼ 3. The chemical structures of the LP analogues are included.
TLR2 recognition by lipopeptides U. Buwitt-Beckmann et al.
6360 FEBS Journal 272 (2005) 6354–6364 ª 2005 The Authors Journal compilation ª 2005 FEBS

This paper presents a systematic investigation of the
structure–activity relationships between the fatty acid
patterns in LP and the huTLR2, as well as the
muTLR2-dependent pattern recognition. The analyses
revealed that the chain length of the two ester-bound
fatty acids is an essential determinant for the biological
activity of LP in HEK293-huTLR2 and in HEK293-
muTLR2 cells. Only fatty acids, having a chain length
of 12 or more carbons were able to substantially sti-
mulate the cells. In contrast, the amide-bound fatty
acids of the LP analogues had no remarkable effect on
the recognition by the TLR2 transfected HEK293 cells.
LP analogues without amide-bound fatty acid were
nearly as active as LP with this substitution (data not
shown). However, it should be kept in mind that the
amide-bound fatty acid of LP beside the peptide moi-
eties determines the coreceptor usage (TLR2 ⁄TLR1 vs.
TLR2 ⁄TLR6 [13,24–26]. Whereas the di-palmitoylated
LP from mycoplasma and the di-palmitoylated LP
form the R. viridis are TLR2 ⁄TRL6 dependent, their
tri-palmitoylated analogues are TLR6-independent. In
addition we could demonstrate that the chain length
of the two ester-bound fatty acids is a determinant
for the species-specific recognition by huTLR2 vs.
muTLR2. Whereas LP having at least two ester-bound
fatty acids with 12 carbon atoms are required for acti-
vation of huTLR2, muTLR2 is already activated by
LP having fatty acids with only eight carbon atoms.
Finally, for HEK293-huTLR2 cells, the distribution of
fatty acids for diacylated LP is also of great relevance
for the bioactivity of LP. In comparison to LP with
two ester-bound fatty acids, LP with one amide-bound
and one ester-bound fatty acid show a markedly
reduced bioactivity in HEK293-huTLR2 cells.
Our investigations have been run in a cell model
using HEK293 cells transfected with human or murine
TLR2. This model has the advantage of being a con-
trolled in vitro system, in which the receptor, which is
responsible for the recognition of the ligand, is well
defined. Preliminary results indicate that a analogous
structure–activity relationship exists also in native cells
(e.g., human and murine macrophages, unpublished
data). Furthermore, using a limited number of Pam3C-
SK4 analogues it has been shown that they induce
specific patterns of cytokines, chemokines and costimu-
latory membrane molecules in human dendritic cells,
depending on the source of amide-bound and ester-
bound fatty acids [27]. Therefore, we conclude that
our results obtained in HEK293-TLR2 cells reflect the
structural relationship of LP-recognition also in native
cells and the native immune system. and indeed, also
in vivo the structure of LP determines the adjuvant
activity [28]. LP with a N-acyl-S-diacylglyceryl cysteine
backbone has been found in all bacteria. During bac-
terial infection these LP may be involved in the initi-
ation of the response of the innate immune system and
activation ⁄modulation of antigen-presenting cells inclu-
ding dendritic cells, thereby modulating the pattern of
the response of the adaptive immune system. There-
fore, it is reasonable to conclude that the structure of
bacterial LP may play a role for the outcome of a bac-
terial infection. However, at present it is still poorly
understood to which extent bacterial LP contribute to
the extent and pattern of the immune response during
bacterial infection.
Taken together, our results show the substantial
importance of the two ester-bound acyl residues within
LP for the induction of signalling through TLR2. Very
similar to the TLR4 ligand LPS [29], the number, the
chain length, and the distribution of the fatty acids is of
great consequence for the stimulatory activity of LP.
LP are amphiphilic molecules and have been described
to form, like LPS, supramolecular structures in aqueous
solution air–water interfaces, and lipid bilayer mem-
branes [30–33]. As has been determined with LPS, the
supramolecular conformation and the molecular shape
are important for its biological activity and the recogni-
tion by LPS binding molecules, e.g. TLT4 [34]. In con-
trast to LPS, however, the supramolecular structure
and molecular conformation of LP, which is optimal
for the recognition by TLR2 remains to be investigated.
Experimental procedures
Materials
All lipopeptide collections and single lipohexa- and lipo-
decapeptides were synthesized and analysed according to
published procedures [35] by EMC microcollections GmbH
(Tubingen, Germany, E-mail: [email protected]).
The abbreviations, structures and names of the fatty acids
used in this paper are given in Table 1. The abbreviations
of the amino acids relate to the single letter code. The
structure denotation of LP are explained in Fig. 1. All
SSNASK4- and SK4- LP analogues were solubilized in
water at a concentration of 1 mm, resulting in a clear solu-
tion. Pam2C-FEPPPATTT analogues were solubilized in
dimethyl sulfoxind at 1 mm and diluted in culture medium
prior stimulation of the cells. All synthetic LP used in this
paper stimulate cells in a strictly TLR2-dependent manner
as determined by the use of TLR2-deficient mice and
untransfected HEK293 cells. Unless otherwise indicated, all
fine chemicals were from Sigma-Aldrich (Deisenhofen, Ger-
many), Serva (Heidelberg, Germany), Merck Biosiences
(Darmstadt, Germany) or Roche Diagnostics (Mannheim,
Germany).
U. Buwitt-Beckmann et al. TLR2 recognition by lipopeptides
FEBS Journal 272 (2005) 6354–6364 ª 2005 The Authors Journal compilation ª 2005 FEBS 6361

Stable transfected HEK293 cells
Expression plasmid containing a Flag-tagged version of
hu-TLR2 was a kind gift from P. Nelson, Seattle, USA and
was subcloned into pREP9 (Invitrogen, Karlsruhe,
Germany). Stable transfection of HEK293 cells with
pREP9-Flag-huTLR2 was performed using Superfect
Transfection Reagent (Quiagen, Hilden, Germany) accord-
ing to the manufacturer’s recommendations. Positive cells
were selected by fluorescence-activated cell sorting. Clonal
cell lines were obtained by limiting dilution. Stably trans-
fected cells were maintained in DMEM supplemented with
10% FCS, 0.5 unitsÆmL)1 penicillin, 0.5 lgÆmL)1 strepto-
mycin, and 400 lgÆmL)1 G418 (Biochrom, Berlin,
Germany). Stable transfected HEK293-huTLR2 cells were
plated on 48-well dishes (Greiner, Frickenhausen,
Germany) at a concentration of 3 · 105ÆmL)1 in 400 lLcomplete medium without G418. The following day cells
were stimulated with the indicated LP for 24 h. Superna-
tants were collected and analysed for an IL-8 content with
a commercial ELISA (Biosource, Solingen, Germany).
Transient transfected HEK293 cells
HEK293 cells were plated at a density of 1.5 · 105ÆmL)1 in
96-well plates in DMEM supplemented with 10% FBS,
0.5 unitsÆmL)1 penicillin, 0.5 lgÆmL)1 streptomycin. The fol-
lowing day, cells were transiently transfected using Polyfect
(Quiagen, Hilden, Germany) according to the manufactures’
protocol. Expression plasmid containing the Flag-tagged ver-
sion of mu-TLR2 was a kind gift from E. Lien, MA, USA
and was subcloned into pREP9 (Invitrogen, Karlsruhe, Ger-
many). pREP9-Flag-mu-TLR2 and pREP9-Flag-huTLR2
plasmids were used at 200 ng per transfection. The efficiency
of the transient transfection was 50–70% as determined by
FACS analysis using a anti-TLR2 mAb. After 6 h of trans-
fection cells were washed and stimulated for further 24 h.
Interleukin-8 content in the culture supernatants was quanti-
fied using a commercial ELISA (Biosource, Camarillo, CA,
USA). The kinetics of the response are shown in Fig. 6.
NF-jB reporter assay
HEK293 cells were cotransfected with 150 ng per transfec-
tion NF-jB firefly luciferase reporter plasmid and 50 ng per
transfection control Renilla luciferase plasmid (both were
kindly provided by Dr D.T. Golenbock, Worcester, USA).
After transfection the cells were stimulated with LP analogs
for 6 h and lysed by passive lysis buffer (Promega, Mann-
heim, Germany). Luciferase activity was measured by using
a luminometer (Berthold, Bad Wildbad, Germany) and was
calculated in relative light units as a ratio of NF-jB-dependent firefly luciferase activity to NF-jB-independentRenilla luciferase activity. The kinetics of the response are
shown in Fig. 6.
Statistics
The results are given in mean of three cultures run in a
given experiment. Standard deviation was less than 15% or
as given in the figures and tables. Each experiment was
repeated at least three times. One of these three experi-
ments, giving representative results, is shown in each figure.
Acknowledgements
The skillful technical assistance by P. Prilla and
C. Schneider (Research Center Borstel) and by
M. Wacker and B. Patzold (EMC microcollections
GmbH) is acknowledged. We thank A. Wallisch for
final reading of this manuscript. This work was sup-
ported by the Deutsche Forschungsgemeinschaft (Ul
68 ⁄ 3-1) and the Bundesministerium fur Forschung und
Technologie (project Biochance Nr. 0312662).
References
1 Muhlradt PF, Kiess M, Meyer H, Sussmuth R & Jung
G (1997) Isolation, structure elucidation, and synthesis
of a macrophage stimulatory lipopeptide from Myco-
plasma fermentans acting at picomolar concentration.
J Exp Med 185, 1951–1958.
2 Metzger JW, Beck-Sickinger AG, Loleit M, Eckert M,
Bessler WG & Jung G (1995) Synthetic S-(2,3-dihydroxy-
propyl)-cysteinyl peptides derived from the N-terminus of
the cytochrome subunit of the photoreaction centre of
Rhodopseudomonas viridis enhance murine splenocyte
proliferation. J Pept Sci 1, 184–190.
Fig. 6. Kinetics of the IL-8 release and NFjB activation after stimu-
lation of HEK-huTLR2 with Pam3-CSK4. HEK-huTLR2 cells were sti-
mulated with Pam3C-SK4 at a concentration of 1000 nM. After
various culture periods (4 h, 8 h, 16 h, 24 h, 48 h) the supernatants
were harvested and the IL-8 concentration was measured in an
ELISA. The Luciferase-reporter assay for NFjB was performed
after 2 h, 4 h, 6 h, and 8 h of stimulation. The results are
expressed as mean ± SD, n ¼ 3.
TLR2 recognition by lipopeptides U. Buwitt-Beckmann et al.
6362 FEBS Journal 272 (2005) 6354–6364 ª 2005 The Authors Journal compilation ª 2005 FEBS

3 Braun V (1975) Covalent lipoprotein from the outer
membrane of Escherichia coli. Biochim Biophys Acta
415, 335–377.
4 Madan BM & Sankaran K (2002) DOLOP – database
of bacterial lipoproteins. Bioinformatics 18, 641–643.
5 Takeuchi O, Kaufmann A, Grote K, Kawai T, Hoshino
K, Morr M, Muhlradt PF & Akira S (2000) Cutting
edge: preferentially the R-stereoisomer of the mycoplas-
mal lipopeptide macrophage-activating lipopeptide-2
activates immune cells through a toll-like receptor
2- and MyD88-dependent signaling pathway. J Immunol
164, 554–557.
6 Lien E, Sellati TJ, Yoshimura A, Flo TH, Rawadi G,
Finberg RW, Carroll JD, Espevik T, Ingalls RR, Radolf
JD & Golenbock DT (1999) Toll-like receptor 2 func-
tions as a pattern recognition receptor for diverse bac-
terial products. J Biol Chem 274, 33419–33425.
7 Janeway CA Jr & Medzhitov R (2002) Innate immune
recognition. Annu Rev Immunol 20, 197–216.
8 Heine H & Lien E (2003) Toll-like receptors and their
function in innate and adaptive immunity. Int Arch
Allergy Immunol 130, 180–192.
9 Ozinsky A, Underhill DM, Fontenot JD, Hajjar AM,
Smith KD, Wilson CB, Schroeder L & Aderem A
(2000) The repertoire for pattern recognition of patho-
gens by the innate immune system is defined by coop-
eration between toll-like receptors. Proc Natl Acad Sci
USA 97, 13766–13771.
10 Alexopoulou L, Thomas V, Schnare M, Lobet Y,
Anguita J, Schoen RT, Medzhitov R, Fikrig E & Flavell
RA (2002) Hyporesponsiveness to vaccination with Bor-
relia burgdorferi OspA in humans and in TLR1- and
TLR2-deficient mice. Nat Med 8, 878–884.
11 Morr M, Takeuchi O, Akira S, Simon MM & Muhlradt
PF (2002) Differential recognition of structural details
of bacterial lipopeptides by toll-like receptors. Eur J
Immunol 32, 3337–3347.
12 Takeuchi O, Sato S, Horiuchi T, Hoshino K, Takeda
K, Dong Z, Modlin RL & Akira S (2002) Cutting edge:
role of Toll-like receptor 1 in mediating immune
response to microbial lipoproteins. J Immunol 169,
10–14.
13 Takeuchi O, Kawai T, Muhlradt PF, Morr M, Radolf
JD, Zychlinsky A, Takeda K & Akira S (2001) Discri-
mination of bacterial lipoproteins by Toll-like receptor
6. Int Immunol 13, 933–940.
14 Buwitt-Beckmann U, Heine H, Wiesmuller KH, Jung
G, Brock R, Akira S & Ulmer AJ (2005) Toll-like
receptor 6-independent signaling by diacylated lipopep-
tides. Eur J Immunol 35, 282–289.
15 Manukyan M, Triantafilou K, Triantafilou M, Mackie
A, Nilsen N, Espevik T, Wiesmuller KH, Ulmer AJ &
Heine H (2005) Binding of lipopeptide to CD14 induces
physical proximity of CD14, TLR2 and TLR1. Eur J
Immunol 35, 911–921.
16 Vasselon T, Detmers PA, Charron D & Haziot A
(2004) TLR2 recognizes a bacterial lipopeptide through
direct binding. J Immunol 173, 7401–7405.
17 Schroder NW, Heine H, Alexander C, Manukyan M,
Eckert J, Hamann L, Gobel UB & Schumann RR
(2004) Lipopolysaccharide binding protein binds to tri-
acylated and diacylated lipopeptides and mediates
innate immune responses. J Immunol 173, 2683–2691.
18 Wiesmuller KH, Bessler WG & Jung G (1992) Solid
phase peptide synthesis of lipopeptide vaccines eliciting
epitope-specific B-, T-helper and T-killer cell response.
Int J Pept Protein Res 40, 255–260.
19 Spohn R, Buwitt-Beckmann U, Brock R, Jung G,
Ulmer AJ & Wiesmuller KH (2004) Synthetic lipopep-
tide adjuvants and Toll-like receptor 2–structure-activity
relationships. Vaccine 22, 2494–2499.
20 Bessler WG, Mittenbuhler K, Esche U & Huber M
(2003) Lipopeptide adjuvants in combination treatment.
Int Immunopharmacol 3, 1217–1224.
21 Muller SD, Muller MR, Huber M, Esche UU, Kirsch-
ning CJ, Wagner H, Bessler WG & Mittenbuhler K
(2004) Triacyl-lipopentapeptide adjuvants: TLR2-depen-
dent activation of macrophages and modulation of
receptor-mediated cell activation by altering acyl-moi-
eties. Int Immunopharmacol 4, 1287–1300.
22 Reutter F, Jung G, Baier W, Treyer B, Bessler WG &
Wiesmuller KH (2005) Immunostimulants and Toll-like
receptor ligands obtained by screening combinatorial
lipopeptide collections. J Pept Res 65, 375–383.
23 Grabiec A, Meng G, Fichte S, Bessler W, Wagner H &
Kirschning CJ (2004) Human but not murine toll-like
receptor 2 discriminates between tri-palmitoylated and
tri-lauroylated peptides. J Biol Chem 279, 48004–48012.
24 Takeuchi O, Sato S, Horiuchi T, Hoshino K, Takeda
K, Dong Z, Modlin RL & Akira S (2002) Cutting edge:
role of Toll-like receptor 1 in mediating immune
response to microbial lipoproteins. J Immunol 169,
10–14.
25 Akira S (2003) Mammalian Toll-like receptors. Curr
Opin Immunol 15, 5–11.
26 Buwitt-Beckmann U, Heine H, Wiesmuller KH, Jung
G, Brock R, Akira S & Ulmer AJ (2005) Toll-like
receptor 6-independent signaling by diacylated lipopep-
tides. Eur J Immunol 35, 282–289.
27 Reschner A, Moretta A, Landmann R, Heberer M,
Spagnoli GC & Padovan E (2003) The ester-bonded
palmitoyl side chains of Pam3CysSerLys4 lipopeptide
account for its powerful adjuvanticity to HLA class
I-restricted CD8+ T lymphocytes. Eur J Immunol 33,
2044–2052.
28 Bessler WG, Baier W, vd EU, Hoffmann P, Heinevetter
L, Wiesmuller KH & Jung G (1997) Bacterial Lipopep-
tides Constitute Efficient Novel Immunogens and Adju-
vants in Parenteral and Oral Immunization. Behring
Institute Mitt, 390–399.
U. Buwitt-Beckmann et al. TLR2 recognition by lipopeptides
FEBS Journal 272 (2005) 6354–6364 ª 2005 The Authors Journal compilation ª 2005 FEBS 6363

29 Heine H, Rietschel ET & Ulmer AJ (2001) The biology
of endotoxin. Mol Biotechnol 19, 279–296.
30 Reichel F, Roelofsen AM, Geurts HPM, Feiters MC &
Boons G-J (2003) Stereochemical dependence of the
self-assembly of the immunoadjuvants Pam3Cys and
Pam3Cys Series. J Am Chem Soc 121, 7989–7997.
31 Prass W, Ringsdorf H, Bessler W, Wiesmuller KH &
Jung G (1987) Lipopeptides of the N-terminus of
Escherichia coli lipoprotein: synthesis, mitogenicity and
properties in monolayer experiments. Biochim Biophys
Acta 900, 116–128.
32 Metzger JW, Sawyer WH, Wille B, Biesert L, Bessler
WG & Jung G (1993) Interaction of immunologically-
active lipopeptides with membranes. Biochim Biophys
Acta 1149, 29–39.
33 Boncheva M, Duschl C, Beck W, Jung G & Vogel H
(2003) Formation and characterization of lipopeptide
layers at interfaces for the molecular recognition of anti-
bodies. Langmuir 12, 5636–5642.
34 Seydel U, Hawkins L, Schromm AB, Heine H, Scheel
O, Koch MH & Brandenburg K (2003) The generalized
endotoxic principle. Eur J Immunol 33, 1586–1581.
35 Metzger JW, Kempter C, Wiesmuller KH & Jung G
(1994) Electrospray mass spectrometry and tandem
mass spectrometry of synthetic multicomponent peptide
mixtures: determination of composition and purity. Anal
Biochem 219, 261–277.
TLR2 recognition by lipopeptides U. Buwitt-Beckmann et al.
6364 FEBS Journal 272 (2005) 6354–6364 ª 2005 The Authors Journal compilation ª 2005 FEBS





























