Embed Size (px)
Citation preview

Litterfall, Nutrient Cycling, and Nutrient Limitation in Tropical ForestsAuthor(s): Peter M. VitousekSource: Ecology, Vol. 65, No. 1 (Feb., 1984), pp. 285-298Published by: Ecological Society of AmericaStable URL: http://www.jstor.org/stable/1939481 .
Accessed: 19/07/2013 00:11
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
Ecological Society of America is collaborating with JSTOR to digitize, preserve and extend access to Ecology.
http://www.jstor.org
This content downloaded from 206.212.0.156 on Fri, 19 Jul 2013 00:11:42 AMAll use subject to JSTOR Terms and Conditions

Ecology, 65(1), 1984, pp. 285-298 C, 1984 by the Ecological Society of Amenca
LITTERFALL, NUTRIENT CYCLING, AND NUTRIENT LIMITATION IN TROPICAL FORESTS'
PETER M. VITOUSEK2 Department of Biology, Coker Hall OJOA, University of North Carolina,
Chapel Hill, North Carolina 27514 USA
Abstract. Patterns of nitrogen, phosphorus, and calcium cycling through litterfall were evaluated using published information from 62 tropical forests. In general, lowland tropical forests have more nitrogen and lower dry mass/nitrogen ratios in litterfall than most temperate forests, while nitrogen return in montane tropical forests is comparable to that in temperate forests. Calcium return is also high in most tropical forests studied, but many tropical forests (lowland and montane) have little phosphorus return and very high dry mass/phosphorus ratios in litterfall compared to most temperate forests. Phosphorus appears to be cycled highly efficiently in such forests.
Fine litterfall in the range of tropical forests studied was predicted from climate, and the residuals of this regression were positively correlated with phosphorus but not nitrogen concentrations in litterfall. The amount of fine litterfall (uncorrected for climate) was also significantly correlated with phosphorus concentrations in moist and wet lowland tropical forests. These analyses suggest that phosphorus but not nitrogen availability limits litterfall in a substantial subset of intact tropical forests. Sites on old oxisols and ultisols, especially those in Amazonia, appear to be particularly low in available phosphorus.
Key words: calcium; montane forests; nitrogen; nutrient use efficiency; phosphorus; sesquioxide; terra firme; tropical forests.
INTRODUCTION
Nutrient cycling in tropical forests is often charac- terized as "tight" or "efficient" relative to temperate forests, but at least two distinct patterns could be im- plied by such a characterization:
1) Nutrient use within tropical trees could be "effi- cient" in that relatively large amounts of organic matter could be fixed per unit of nutrient taken up (the "re- source utility" of Hirose 1975). Such efficient use could occur either because more carbon is fixed per unit of nutrient in trees or because a larger fraction of nutrients is reabsorbed from senescing plant parts. Efficient nu- trient use of this sort (which I will call efficient within- stand cycling) would be characterized by high carbon/ nutrient ratios (low nutrient concentrations) in litter- fall, wood, and root litter (Vitousek 1982).
2) Nutrient cycling within tropical forest ecosystems could be "efficient" if most of the nutrients released from trees were rapidly taken up by roots, mycorrhizae, and decomposers, and retained within the system. Ef- ficient nutrient cycling of this sort would be charac- terized by low nutrient losses from the system as a whole despite relatively large amounts of nutrient cy- cling between trees and soil.
I will be concerned primarily with within-stand nu- trient cycling here, although the two are related in that efficient within-stand nutrient use generally results in efficient within-system cycling as well (Vitousek et al. 1982). Many, perhaps most, trees vary in the efficiency
1 Manuscript received 30 August 1982; revised 16 January 1983; accepted 21 March 1983.
2 Present address: Department of Biological Sciences, Stan- ford University, Stanford, California 94305 USA.
with which they use nutrients, depending on nutrient availability (cf. Stachurski and Zimka 1975, Miller et al. 1976, Turner 1977, Ingestad 1979). Consequently, the existence of an efficient within-stand nutrient econ- omy supports the possibility of nutrient limitation to primary production, while an inefficient within-stand nutrient economy indicates that the supply of that nu- trient to trees is adequate or better (Grubb 1977, Vi- tousek 1982). Foliar nutrient concentrations provide an alternative means of characterizing nutrient avail- ability in tropical forests (cf. Grubb 1977, Snedaker 1980, Medina et al. 1981, Peace and MacDonald 1981). As Grubb (1977) pointed out, however, foliar analyses do not include the influence of nutrient and energy reabsorption during leaf senescence, which contributes materially to the efficiency with which nutrients are used.
In this paper, I will examine the relationships among litterfall, nutrient return, and within-stand nutrient economies in a wide range of tropical forest ecosys- tems. I will emphasize litterfall and litter nutrient con- tents because there is much more information on lit- terfall and nutrient contents than on the other components of primary production in tropical forests. There is also a substantial amount of information on wood production (cf. Jordan 1982), but comparatively few useful data on the nutrient content of the annual wood increment (Bernhard-Reversat 1977). Fine root production and nutrient turnover belowground are un- derstandably rather poorly known (Klinge 1973, Jor- dan and Escalante 1980). The limited information available suggests that dry mass/nutrient ratios in lit- terfall are a good index of the nutrient economy in a stand as a whole (Vitousek 1982).
This content downloaded from 206.212.0.156 on Fri, 19 Jul 2013 00:11:42 AMAll use subject to JSTOR Terms and Conditions

286 PETER M. VITOUSEK Ecology, Vol. 65, No. 1
SOURCES AND QUALITY OF INFORMATION
I summarized the results of studies of litterfall and nutrient cycling in tropical and subtropical forests and savannas within the Tropics of Cancer and Capricorn (23027'N and S). Several reviews of litterfall in the tropics, especially Rodin and Bazilevich (1967), Klinge (1978), Brown and Lugo (1982), Vitousek (1982), Jor- dan (1983), and the complete review by Proctor (1983), were used to locate studies, and several tropical ecol- ogists generously assisted in the location of additional studies. All of the information used is summarized in Table 1; except as indicated, all of the primary studies were located and evaluated.
Methodological differences between studies are a most serious problem in any comparison of this kind. The problem may be less severe with litterfall and litter nutrient contents than with most ecosystem-level com- parisons, as the methods used are relatively simple and standardized. Different studies nonetheless use rather different techniques and report different fluxes. Some report only leaf litterfall, while others report fine lit- terfall, which is defined as leaves plus twigs plus flowers and fruit. Still others report total litterfall, which is defined as fine litterfall plus tree stems and large branches. These definitions are not always consistently applied (for example, Rodin and Bazilevich [1967] de- fine "leaf litter fall" as being identical with what I term fine litterfall), and not all studies are clear about which flux they are reporting. On the average, leaf litterfall is - 70% of fine litterfall (Bray and Gorham 1964, O'Neill
and DeAngelis 1980), and the woody component of total litterfall is too heterogeneous both spatially and temporally to allow meaningful generalizations.
The techniques used for litterfall collection can in- fluence the results obtained. Ideally, litterfall should be collected at frequent intervals from well-replicated traps located above the soil surface (Proctor 1983). Frequent collections are particularly important in low- land tropical areas where the rapid decomposition of litter in traps can lead to substantial underestimates of litterfall (Kunkel-Westphal and Kunkel 1979). Traps and frequent collections are difficult to maintain in many tropical areas, however, and collections of litter at the soil surface may be necessary (cf. Malaisse et al. 1975) despite the increased probability of contami- nation.
Most of the studies located were included in my analyses despite methodological differences. Studies which calculated litterfall from other measurements (cf. Wanner 1970, Kawanabe 1977) or which used very short collection periods (cf. Kira et al. 1967) were ex- cluded. Most of the Indian studies report litter pro- duction based on a collection of litter on the ground during the dry season. Perhaps as a consequence, their chemistry is variable and quite difficult to interpret. Finally, the results at a number of sites (particularly El Verde, Puerto Rico, and San Carlos de Rio Negro,
Venezuela) had to be collated from several publica- tions, and I was unsure that the time periods or tech- niques used were always comparable.
Information on the location and climate of each site was obtained where possible (Table 1). In some in- stances the primary studies did not report this infor- mation, which was then sought in other publications or in a climatic atlas as noted in Table 1.
EFFICIENCY OF WITHIN-STAND
NUTRIENT USE
I defined an efficient within-stand nutrient economy as one in which a relatively large amount of organic matter is produced per unit of nutrient taken up (Hirose 1975), and I used the dry mass/nutrient ratio of litter- fall as an index of that efficiency (Chapin 1980, Vitou- sek 1982). Nutrients leached from a forest canopy in throughfall or stemflow should also be included in cal- culating this efficiency, but while data on throughfall and stemflow fluxes in tropical forests exist (Nye 1961, Kenworthy 1970, Bernhard-Reversat 1977, Jordan et al. 1980, Edwards 1982), they are relatively sparse. Accordingly, I confined my analyses to nitrogen, phos- phorus, and calcium, all of which are transferred pri- marily by litterfall (Cole and Rapp 1980, Parker 1983).
My analyses were confined to sites where fine litter- fall and nutrient contents in forests at least 25 yr old were reported. The fine litter fraction was used because it is relatively well defined and because it was reported in most studies. Results from 62 of the sites listed in Table 1 met these criteria.
I used the information in Table 1 to test the hy- pothesis that within-stand nutrient use is more efficient in tropical than in temperate forests. This hypothesis would be falsified if most tropical forests failed to have dry mass/nutrient ratios in litterfall above those of most temperate forests.
Results are summarized in Fig. 1, where the dry mass/nutrient ratio (the inverse of the nutrient con- centration) in litterfall is plotted against the amount of that nutrient in litterfall for each site. Regression lines (with 95% confidence limits) for a similar analysis of temperate and boreal forests (data from Vitousek 1982) are superimposed on Fig. 1. The y axis in Fig. 1 is an index of the efficiency of within-stand cycling for that nutrient, while the x axis is the amount of that nutrient returned annually through fine litterfall. These axes are correlated, so that a spurious "pattern" could emerge if litterfall dry mass and nutrient content were unre- lated (see Vitousek 1982 for a discussion of this prob- lem).
Patterns for nitrogen
Nitrogen cycling in tropical forests follows a pattern similar to that in temperate forests, with inefficient within-stand use at high nitrogen circulation and an ascending limb of more efficient cycling at lower levels
This content downloaded from 206.212.0.156 on Fri, 19 Jul 2013 00:11:42 AMAll use subject to JSTOR Terms and Conditions
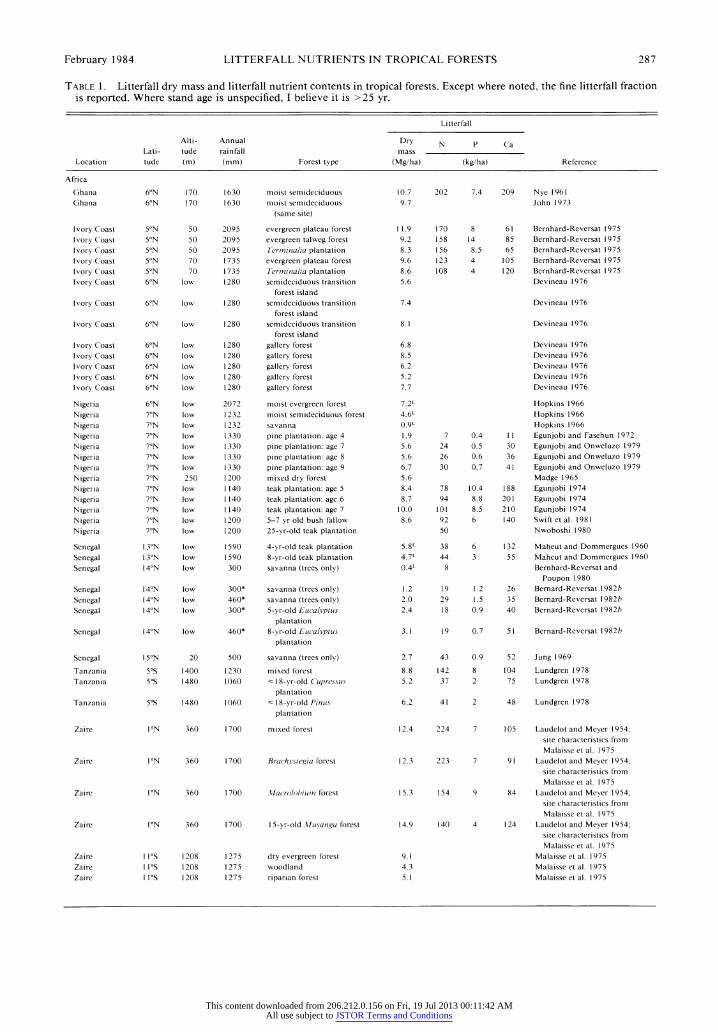
February 1984 LITTERFALL NUTRIENTS IN TROPICAL FORESTS 287
TABLE 1. Litterfall dry mass and litterfall nutrient contents in tropical forests. Except where noted, the fine litterfall fraction is reported. Where stand age is unspecified, I believe it is >25 yr.
Litterfall
Alti- Annual Dry N P Ca Lati- tude rainfall mass
Location tude (m) (mm) Forest type (Mg/ha) (kg/ha) Reference
Africa
Ghana 6?N 170 1630 moist semideciduous 10.7 202 7.4 209 Nye 1961 Ghana 6?N 170 1630 moist semideciduous 9.7 John 1973
(same site)
Ivory Coast 5?N 50 2095 evergreen plateau forest 11.9 170 8 61 Bernhard-Reversat 1975 Ivory Coast 5?N 50 2095 evergreen talweg forest 9.2 158 14 85 Bernhard-Reversat 1975 Ivory Coast 5?N 50 2095 Terminalia plantation 8.3 156 8.5 65 Bernhard-Reversat 1975 Ivory Coast 5?N 70 1735 evergreen plateau forest 9.6 123 4 105 Bernhard-Reversat 1975 Ivory Coast 5?N 70 1735 Terminalia plantation 8.6 108 4 120 Bernhard-Reversat 1975 Ivory Coast 6&N low 1280 semideciduous transition 5.6 Devineau 1976
forest island Ivory Coast 6?N low 1 280 semideciduous transition 7.4 Devineau 1976
forest island Ivory Coast 6?N low 1280 semideciduous transition 8.1 Devineau 1976
forest island Ivory Coast 6?N low 1 280 gallery forest 6.8 Devineau 1976 Ivory Coast 6?N low 1280 gallery forest 8.5 Devineau 1976 Ivory Coast 6?N low 1280 gallery forest 6.2 Devineau 1976 Ivory Coast 6?N low 1280 gallery forest 5.2 Devineau 1976
Ivory Coast 6?N low 1280 gallery forest 7.7 Devineau 1976
Nigeria 6N low 2072 moist evergreen forest 7.2L Hopkins 1966 Nigeria 7?N low 1232 moist semideciduous forest 4.6L Hopkins 1966 Nigeria 7?N low 1232 savanna 0.9L Hopkins 1966 Nigeria 7?N low 1330 pine plantation: age 4 1.9 7 0.4 11 Egunjobi and Fasehun 1972 Nigeria 7?N low 1330 pine plantation: age 7 5.6 24 0.5 30 Egunjobi and Onweluzo 1979 Nigeria 7?N low 1330 pine plantation: age 8 5.6 26 0.6 36 Egunjobi and Onweluzo 1979 Nigeria 7?N low 1330 pine plantation: age 9 6.7 30 0.7 41 Egunjobi and Onweluzo 1979 Nigeria 7?N 250 1200 mixed dry forest 5.6 Madge 1965 Nigeria 7?N low 1140 teak plantation: age 5 8.4 78 10.4 188 Egunjobi 1974 Nigeria 7?N low 1140 teak plantation: age 6 8.7 94 8.8 201 Egunjobi 1974 Nigeria 7?N low 1140 teak plantation: age 7 10.0 101 8.5 210 Egunjobi 1974 Nigeria 7?N low 1200 5-7 yr old bush fallow 8.6 92 6 140 Swift et al. 1981 Nigeria 7?N low 1200 25-yr-old teak plantation 50 Nwoboshi 1980
Senegal 13?N low 1590 4-yr-old teak plantation 5.8L 38 6 132 Maheut and Dommergues 1960
Senegal 13?N low 1590 8-yr-old teak plantation 4.7L 44 3 55 Maheut and Dommergues 1960 Senegal 14?N low 300 savanna (trees only) 0.4L 8 Bernhard-Reversat and
Poupon 1980 Senegal 14?N low 300* savanna (trees only) 1.2 19 1.2 26 Bernard-Reversat 1982h
Senegal 14?N low 460* savanna (trees only) 2.0 29 1.5 35 Bernard-Reversat 1982h
Senegal 14?N low 300* 5-yr-old Eucalvptus 2.4 18 0.9 40 Bernard-Reversat 1982h plantation
Senegal 14?N low 460* 8-yr-old Eucalyptus 3.1 19 0.7 51 Bernard-Reversat 1982h plantation
Senegal 15?N 20 500 savanna (trees only) 2.7 43 0.9 52 Jung 1969
Tanzania 5?S 1400 1230 mixed forest 8.8 142 8 104 Lundgren 1978
Tanzania 5?S 1480 1060 18-yr-old Cupressus 5.2 37 2 75 Lundgren 1978 plantation
Tanzania 5?S 1480 1060 18-yr-old Pinus 6.2 41 2 48 Lundgren 1978 plantation
Zaire 1N 360 1700 mixed forest 12.4 224 7 105 Laudelot and Meyer 1954; site characteristics from Malaisse et al. 1975
Zaire 1N 360 1700 Brachvstegia forest 12.3 223 7 91 Laudelot and Meyer 1954; site characteristics from Malaisse et al. 1975
Zaire 1N 360 1700 Macro/ohiumn forest 15.3 154 9 84 Laudelot and Meyer 1954; site characteristics from Malaisse et al. 1975
Zaire 1N 360 1700 15-yr-old Musanga forest 14.9 140 4 124 Laudelot and Meyer 1954; site characteristics from Malaisse et al. 1975
Zaire Ii'S 1208 1275 dry evergreen forest 9.1 Malaisse et al. 1975 Zaire I iS 1208 1275 woodland 4.3 Malaisse et al. 1975 Zaire I i'S 1208 1275 riparian forest 5.1 Malaisse et al. 1975
This content downloaded from 206.212.0.156 on Fri, 19 Jul 2013 00:11:42 AMAll use subject to JSTOR Terms and Conditions
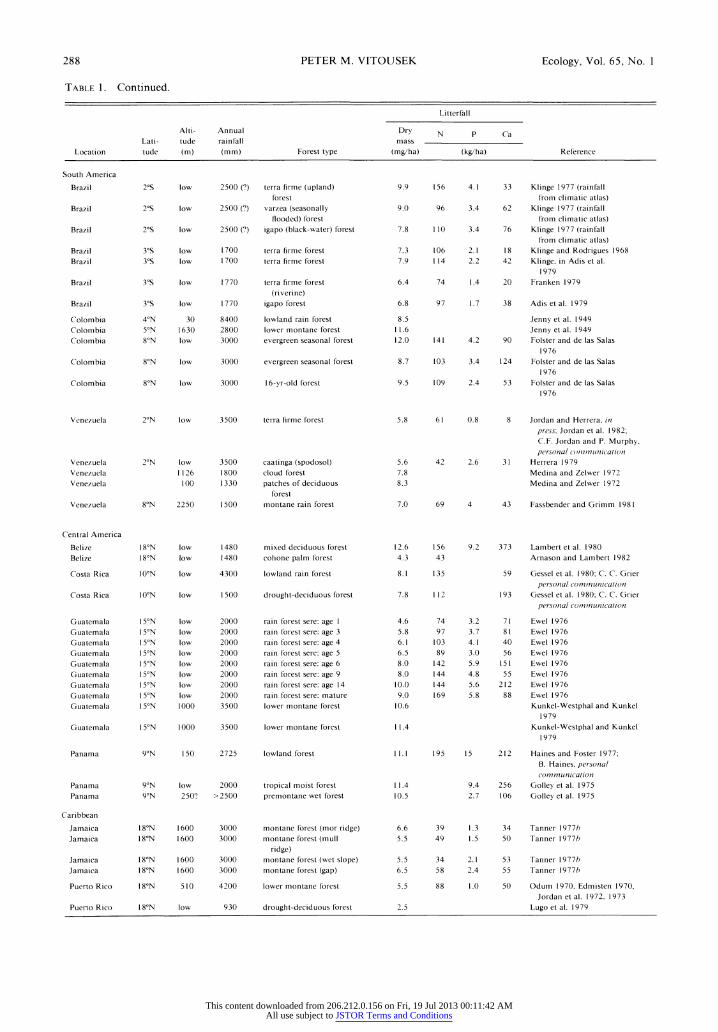
288 PETER M. VITOUSEK Ecology, Vol. 65, No. 1
TABLE 1. Continued.
Litterfall
Alti- Annual Dry N v Ca
Lati- tude rainfall mass
Location tude (m) (mm) Forest type (mg/ha) (kg/ha) Reference
South America
Brazil 2?S low 2500 (?) terra firme (upland) 9.9 156 4.1 33 Klinge 1977 (rainfall forest from climatic atlas)
Brazil 2?S low 2500 (7) varzea (seasonally 9.0 96 3.4 62 Klinge 1977 (rainfall flooded) forest from climatic atlas)
Brazil 2?S low 2500 (?) igapo (black-water) forest 7.8 110 3.4 76 Klinge 1977 (rainfall from climatic atlas)
Brazil 3YS low 1700 terra firme forest 7.3 106 2.1 18 Klinge and Rodrigues 1968 Brazil YS low 1700 terra firme forest 7.9 114 2.2 42 Klinge, in Adis et al.
1979 Brazil 3?S low 1770 terra firme forest 6.4 74 1.4 20 Franken 1979
(riverine) Brazil 3?S low 1770 igapo forest 6.8 97 1.7 38 Adis et al. 1979
Colombia 4?N 30 8400 lowland rain forest 8.5 Jenny et al. 1949 Colombia 5?N 1630 2800 lower montane forest 11.6 Jenny et al. 1949 Colombia 8?N low 3000 evergreen seasonal forest 12.0 141 4.2 90 Folster and de las Salas
1976 Colombia 8?N low 3000 evergreen seasonal forest 8.7 103 3.4 124 Folster and de las Salas
1976 Colombia 8?N low 3000 16-yr-old forest 9.5 109 2.4 53 Folster and de las Salas
1976
Venezuela 2?N low 3500 terra firme forest 5.8 61 0.8 8 Jordan and Herrera, in press: Jordan et al. 1982;
C.F. Jordan and P. Murphy, personal co(msunlnnic'ation
Venezuela 2?N low 3500 caatinga (spodosol) 5.6 42 2.6 31 Herrera 1979 Venezuela 1126 1800 cloud forest 7.8 Medina and Zelwer 1972 Venezuela 100 1330 patches of deciduous 8.3 Medina and Zelwer 1972
forest Venezuela 8?N 2250 1500 montane rain forest 7.0 69 4 43 Fassbender and Grimm 1981
Central America
Belize 18?N low 1480 mixed deciduous forest 12.6 156 9.2 373 Lambert et al. 1980 Belize 18?N low 1480 cohone palm forest 4.3 43 Arnason and Lambert 1982
Costa Rica I0?N low 4300 lowland rain forest 8.1 135 59 Gessel et al. 1980; C. C. Grier personal communication
Costa Rica I0?N low 1500 drought-deciduous forest 7.8 112 193 Gessel et al. 1980; C. C. Grier personal comnunication
Guatemala 15?N low 2000 rain forest sere: age 1 4.6 74 3.2 71 Ewel 1976 Guatemala 15?N low 2000 rain forest sere: age 3 5.8 97 3.7 81 Ewel 1976 Guatemala 15?N low 2000 rain forest sere: age 4 6.1 103 4.1 40 Ewel 1976 Guatemala 15?N low 2000 rain forest sere: age 5 6.5 89 3.0 56 Ewel 1976 Guatemala 15?N low 2000 rain forest sere: age 6 8.0 142 5.9 151 Ewel 1976 Guatemala 15?N low 2000 rain forest sere: age 9 8.0 144 4.8 55 Ewel 1976 Guatemala 15?N low 2000 rain forest sere: age 14 10.0 144 5.6 212 Ewel 1976 Guatemala I5?N low 2000 rain forest sere: mature 9.0 169 5.8 88 Ewel 1976 Guatemala 15?N 1000 3500 lower montane forest 10.6 Kunkel-Westphal and Kunkel
1979
Guatemala 15?N 1000 3500 lower montane forest 11.4 Kunkel-Westphal and Kunkel 1979
Panama 9?N 150 2725 lowland forest 11.1 195 15 212 Haines and Foster 1977; B. Haines, personal
communication
Panama 9?N low 2000 tropical moist forest 11.4 9.4 256 Golley et al. 1975 Panama 9?N 250? >2500 premontane wet forest 10.5 2.7 106 Golley et al. 1975
Caribbean
Jamaica 18?N 1600 3000 montane forest (mor ridge) 6.6 39 1.3 34 Tanner 1977b Jamaica 18?N 1600 3000 montane forest (mull 5.5 49 1.5 50 Tanner 1977b
ridge) Jamaica 18?N 1600 3000 montane forest (wet slope) 5.5 34 2.1 53 Tanner 19776 Jamaica 18?N 1600 3000 montane forest (gap) 6.5 58 2.4 55 Tanner 19776
Puerto Rico I8?N 510 4200 lower montane forest 5.5 88 1.0 50 Odum 1970. Edmisten 1970. Jordan ci at. 1972. 1973
Puerto Rico 1 8?N low 930 drought-deciduous forest 2.5 Logo ci at. 1979
This content downloaded from 206.212.0.156 on Fri, 19 Jul 2013 00:11:42 AMAll use subject to JSTOR Terms and Conditions
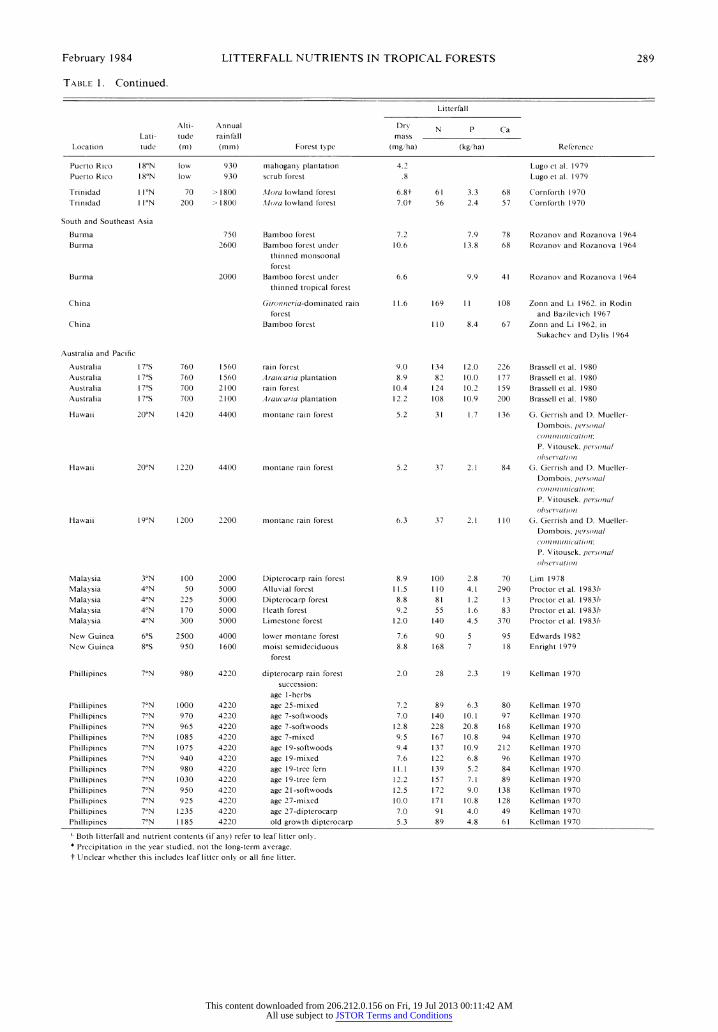
February 1984 LITTERFALL NUTRIENTS IN TROPICAL FORESTS 289
TABLE 1. Continued.
Litterfall
Alti- Annual Dry N P Ca
Lati- tude rainfall mass
Location tude (m) (mm) Forest type (mg/ha) (kg/ha) Reference
Puerto Rico 18'N low 930 mahogany plantation 4.2 Lugo et al. 1979
Puerto Rico 18'N low 930 scrub forest .8 Lugo et al. 1979
Trinidad I VN 70 > 1800 Mora lowland forest 6.8t 61 3.3 68 Cornforth 1970
Trinidad I VN 200 > 1800 MAora lowland forest 7.Ot 56 2.4 57 Cornforth 1970
South and Southeast Asia
Burma 750 Bamboo forest 7.2 7.9 78 Rozanov and Rozanova 1964
Burma 2600 Bamboo forest under 10.6 13.8 68 Rozanov and Rozanova 1964
thinned monsoonal
forest
Burma 2000 Bamboo forest under 6.6 9.9 41 Rozanov and Rozanova 1964
thinned tropical forest
China Gironneria-dominated rain 11.6 169 11 108 Zonn and Li 1962, in Rodin
forest and Bazilevich 1967
China Bamboo forest 110 8.4 67 Zonn and Li 1962. in
Sukachev and Dvlis 1964
Australia and Pacific
Australia 17?S 760 1560 rain forest 9.0 134 12.0 226 Brassell et al. 1980
Australia I7?S 760 1560 1 raucaria plantation 8.9 82 10.0 177 Brassell et al. 1980
Australia 17?S 700 2100 rain forest 10.4 124 10.2 159 Brassell et al. 1980
Australia 17?S 700 2100 .ralucaria plantation 12.2 108 10.9 200 Brassell et al. 1980
Hawaii 20'N 1420 4400 montane rain forest 5.2 31 1.7 136 G. Gerrish and D. Mueller-
Dombois, personal
c ommuzlnic ation,
P. Vitousek, pe('rsonl/
obsert ation
Hawaii 20?N 1220 4400 montane rain forest 5.2 37 2.1 84 G. Gerrish and D. Mueller-
Dombois, p)ersonal communuuficatiOn-:
P. Vitousek, personal
ohsertvatioti
Hawaii 19?N 1200 2200 montane rain forest 6.3 37 2.1 110 G. Gerrish and D. Mueller-
Dombois, personal
commzufhnic nation :
P. Vitousek, personal
obs('rtationl
Malavsia 3N 100 2000 Dipterocarp rain forest 8.9 100 2.8 70 Lim 1978
Malaysia 4?N 50 5000 Alluvial forest 11.5 110 4.1 290 Proctor et al. 19831h
Malaysia 4?N 225 5000 Dipterocarp forest 8.8 81 1.2 13 Proctor et al. 1983h
Malaysia 4?N 170 5000 Heath forest 9.2 55 1.6 83 Proctor et al. 1983h
Malaysia 4?N 300 5000 Limestone forest 12.0 140 4.5 370 Proctor et al. 19831h
New Guinea 6?S 2500 4000 lower montane forest 7.6 90 5 95 Edwards 1982
New Guinea 8?S 950 1600 moist semideciduous 8.8 168 7 18 Enright 1979
forest
Phillipines 7?N 980 4220 dipterocarp rain forest 2.0 28 2.3 19 Kellman 1970
succession:
age 1-herbs
Phillipines 7?N 1000 4220 age 25-mixed 7.2 89 6.3 80 Kellman 1970
Phillipines 7?N 970 4220 age 7-softwoods 7.0 140 10.1 97 Kellman 1970
Phillipines 7?N 965 4220 age 7-softwoods 12.8 228 20.8 168 Kellman 1970
Phillipines 7?N 1085 4220 age 7-mixed 9.5 167 10.8 94 Kellman 1970
Phillipines 7?N 1075 4220 age 19-softwoods 9.4 137 10.9 212 Kellman 1970
Phillipines 7?N 940 4220 age 19-mixed 7.6 122 6.8 96 Kellman 1970
Phillipines 7?N 980 4220 age 19-tree fern 11.1 139 5.2 84 Kellman 1970
Phillipines 7?N 1030 4220 age 19-tree fern 12.2 157 7.1 89 Kellman 1970
Phillipines 7?N 950 4220 age 21-softwoods 12.5 172 9.0 138 Kellman 1970
Phillipines 7?N 925 4220 age 27-mixed 10.0 171 10.8 128 Kellman 1970
Phillipines 7?N 1235 4220 age 27-dipterocarp 7.0 91 4.0 49 Kellman 1970
Phillipines 7?N 1185 4220 old growth dipterocarp 5.3 89 4.8 61 Kellman 1970
LBoth litterfall and nutrient contents (if any) refer to leaf litter only. * Precipitation in the year studied, not the long-term average.
t Unclear whether this includes leaf litter only or all fine litter.
This content downloaded from 206.212.0.156 on Fri, 19 Jul 2013 00:11:42 AMAll use subject to JSTOR Terms and Conditions
290 PETER M. VITOUSEK Ecology, Vol. 65, No. 1
200
\ MM W A 160 \\\MM z \M
\NW
120 - _ 8000 ~ M S
O 80 .. _. -*
S S S N *0
*I *
'I~~~~~~ 600 - . S
40
0 50 100 400 200 250
N in Litterfall (kg ha-t' yr-')
8000
600 B M
4000 - . M o S,_ M .
2000 -M M* 0M 100 1 20 0--- 0. 0 0 0 0go1
1 2 3 4 5 6 7 8 9 10 II 12 13 14
P in Litterfall (kg ha-' yr-')
800-
600 0
>400-
200 -
0 ?~~~~~~M, M 0:0 M 0 *o000
37
50 100 150 200 250 Ca in Litterfall (kg ha-'yr-')
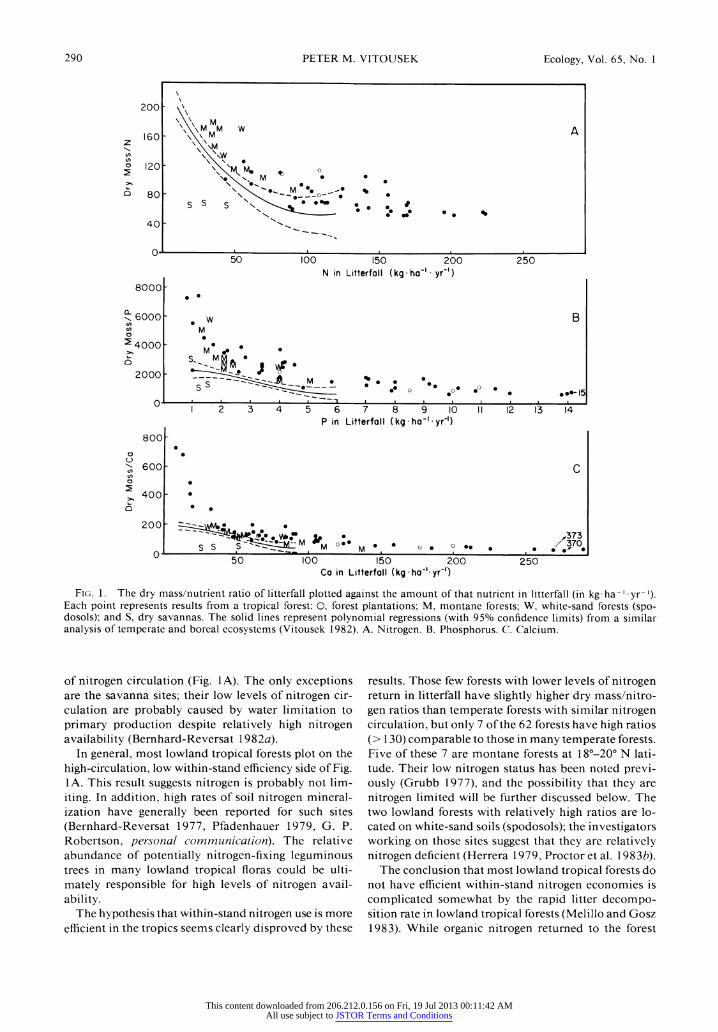
FIG. 1. The dry mass/nutrient ratio of litterfall plotted against the amount of that nutrient in litterfall (in kg ha-' yr-'). Each point represents results from a tropical forest: 0, forest plantations; M, montane forests; W, white-sand forests (spo- dosols); and S, dry savannas. The solid lines represent polynomial regressions (with 95% confidence limits) from a similar analysis of temperate and boreal ecosystems (Vitousek 1982). A. Nitrogen. B. Phosphorus. C. Calcium.
of nitrogen circulation (Fig. 1A). The only exceptions are the savanna sites; their low levels of nitrogen cir- culation are probably caused by water limitation to primary production despite relatively high nitrogen availability (Bernhard-Reversat 1 982a).
In general, most lowland tropical forests plot on the high-circulation, low within-stand efficiency side of Fig. 1A. This result suggests nitrogen is probably not lim- iting. In addition, high rates of soil nitrogen mineral- ization have generally been reported for such sites (Bernhard-Reversat 1977, Pfadenhauer 1979, G. P. Robertson, personal communication). The relative abundance of potentially nitrogen-fixing leguminous trees in many lowland tropical floras could be ulti- mately responsible for high levels of nitrogen avail- ability.
The hypothesis that within-stand nitrogen use is more efficient in the tropics seems clearly disproved by these
results. Those few forests with lower levels of nitrogen return in litterfall have slightly higher dry mass/nitro- gen ratios than temperate forests with similar nitrogen circulation, but only 7 of the 62 forests have high ratios (> 130) comparable to those in many temperate forests. Five of these 7 are montane forests at 180-200 N lati- tude. Their low nitrogen status has been noted previ- ously (Grubb 1977), and the possibility that they are nitrogen limited will be further discussed below. The two lowland forests with relatively high ratios are lo- cated on white-sand soils (spodosols); the investigators working on those sites suggest that they are relatively nitrogen deficient (Herrera 1979, Proctor et al. 1 983b).
The conclusion that most lowland tropical forests do not have efficient within-stand nitrogen economies is complicated somewhat by the rapid litter decompo- sition rate in lowland tropical forests (Melillo and Gosz 1983). While organic nitrogen returned to the forest
This content downloaded from 206.212.0.156 on Fri, 19 Jul 2013 00:11:42 AMAll use subject to JSTOR Terms and Conditions

February 1984 LITTERFALL NUTRIENTS IN TROPICAL FORESTS 291
floor in temperate-zone litterfall is unlikely to be taken up again by vegetation in that year, decomposition rates are high enough that a unit of nitrogen could cycle through a lowland tropical forest several times in a year (Dommergues 1963). The mass lost/nitrogen lost per year could thus be understated in Fig. 1A, which reports the mass lost/nitrogen lost per cycle. Nonethe- less, it seems unlikely that production could be nitrogen limited in such forests as long as the soil-plant nitrogen cycle remains intact.
Patterns for phosphorus
Within-stand phosphorus cycling in tropical forests follows a pattern like that of nitrogen in temperate forests, with low efficiency at high levels of phosphorus circulation and a sharply ascending limb of progres- sively more efficient phosphorus cycles at low levels of phosphorus circulation (Fig. 1 B). Forests on this as- cending limb have dry mass/phosphorus ratios sub- stantially above those in most temperate forests. Clear- ly phosphorus use is more efficient in these tropical forests than in most temperate forests, and phosphorus limitation to primary production appears to be a pos- sibility worth examining. Not all tropical forests use phosphorus efficiently, though; many circulate large amounts of phosphorus and have low dry mass/phos- phorus ratios (Fig. 1 B). In addition, not all forests out- side the tropics are inefficient; some eucalypts and pine plantations on extremely phosphorus-deficient soils in Australia also have efficient phosphorus economics (Melillo and Gosz 1983). However, phosphorus use is probably controlled by phosphorus availability, and as will be discussed below, phosphorus-deficient soils are much more commonly found in the tropics than the temperate zone.
The tropical forests with the most efficient within- stand phosphorus economics (generally those with lit- terfall P of <3 kg ha-' yr-') include some of the mon- tane forests. In addition, Amazonian forests generally have lower phosphorus circulation and higher dry mass/ phosphorus ratios than other lowland tropical forests. The two forests with the highest dry mass/P ratios in litterfall are an oxisol site in Venezuelan Amazonia (Jordan and Herrera 1982) and a comparable "red- yellow podzolic" (oxisol or ultisol) site in Sarawak (Proctor et al. 1983b).
Patterns for calcium
Most tropical forests have dry mass/calcium ratios in litterfall similar to those of most temperate forests (Fig. 1 C), although the maximum annual calcium cir- culation is higher in certain carbonate-rich forests (Gessel et al. 1980, Lambert et al. 1980, Proctor et al. 1983b). Six tropical forests had substantially elevated dry mass/calcium ratios, including four Amazonian forests that also had elevated dry mass/phosphorus ratios. Except for these six, tropical forests do not ap- pear to use calcium efficiently compared to temperate forests.
ARE Low NUTRIENT LEVELS CORRELATED
WITH Low FINE LITTERFALL?
Phosphorus return in litterfall varies over a 15-fold range in established (>25-yr-old) lowland tropical for- ests, while the amount of fine litterfall varies by little more than two-fold (Table 1). Clearly a consequence of efficient phosphorus economics is that relatively high levels of fine litterfall can be maintained despite low phorphorus levels. Are there limits beyond which trop- ical trees cannot compensate for low nutrient levels with increased efficiency of nutrient use, limits below which low nutrient levels are correlated with reduced fine litterfall?
My goal was to determine whether nutrient levels are correlated with fine litterfall in a way that can be distinguished from the effects of climate. On a global scale, fine litterfall is highly positively correlated (r2 =
0.77) with calculated Actual Evapotranspiration (AET), and nearly as highly correlated (inversely) with latitude (r2 = 0.67) (Meentemeyer et al. 1982). Much smaller ranges of AET, latitude, and litter production are pres- ent in the tropical forest data set (Table 1), but the presence of lowland, drought-deciduous, savanna, and montane tropical forests suggests that some correlation with climate should be expected within the tropics.
I predicted fine litterfall (Table 1) from a range of climatic parameters, then compared the residuals of this analysis with nutrient concentrations in litterfall. I took the correlation of low litter nutrient concentra- tions with strongly negative residuals from the climatic analyses as indirect evidence of nutrient limitation to fine litterfall. Finally, I examined the correlation be- tween nutrient concentrations and fine litterfall for a more homogeneous data set comprised of moist and wet lowland forests.
An edited subset of the results in Table 1 was used for these analyses. Five of the 62 sites for which nu- trient cycling information was available lacked ade- quate information on climate. Two more (1 in Zaire, 1 in Belize) had inexplicably high or low litterfall dry mass. Most of my analyses were conducted using re- sults from the remaining 55 sites.
Once again, my analyses were confined to fine lit- terfall. The results probably apply to total above-ground production as well (Vitousek 1982), but root produc- tion is relatively (and perhaps absolutely) greater in low- than in high-nutrient temperate forests (Persson 1980, Keyes and Grier 1981). Even so, where data are available, total net primary production appears to be greater in high-nutrient than low-nutrient sites in the temperate zone (J. D. Aber, personal communication).
Climate and litter production
Many studies did not report enough information to calculate AET, so I used the latitude and elevation of each site to construct a scalar (TEMP) for temperature and energy balance. This scalar was defined as:
TEMP = 26 - 0.007 x (LAT2) - 0.0045 x ELEV (1)
This content downloaded from 206.212.0.156 on Fri, 19 Jul 2013 00:11:42 AMAll use subject to JSTOR Terms and Conditions

292 PETER M. VITOUSEK Ecology, Vol. 65, No. 1
where LAT is the latitude (in degrees north or south) and ELEV is the elevation (in metres) of each site. The constant 26 is the mean annual temperature at sea level at the equator, the coefficient for latitude was selected to duplicate the mean temperature gradient at sea level between the equator and 30'N and S (Neuberger and Cahir 1969), and the coefficient for elevation was cho- sen to be intermediate between the wet and dry adi- abatic lapse rates.
Mean annual precipitation (PPT) in metres was used directly for all sites with < 2 m annual rainfall; above that level, PPT was set to 2 m. I assumed that above 2 m water availability did not affect fine litterfall; several of the highest values reported occurred just below 2 m. Very high rainfall could cause reduced production, however, especially on poorly drained sites. The sea- sonality of precipitation could also affect litter pro- duction independently of the amount of precipitation, but Holdridge et al. (1971) concluded that the amount and the seasonality of precipitation are strongly in- versely correlated in tropical environments.
I calculated higher order, logarithmic, and exponen- tial transformations and interactions of TEMP and PPT, and used a stepwise linear regression procedure (the maximum r2 procedure of SAS; Statistical Analysis System 1979) to select the best predictors of litterfall dry mass (MASS). The analyses were performed on the edited data set for all sites reporting fine litterfall dry mass and adequate climatic information (77 in all- Table 1), and were then repeated for those sites with information on nutrient cycling (55 sites).
The best single predictor of fine litterfall dry mass was a TEMP-PPT interaction term (BET) defined as:
BET = TEMP x LOG (PPT) (2)
with an r2 of 0.42 (P < .0001); the equation was:
MASS = 5.64 + 0.21 x BET (3)
The inclusion of up to four additional terms only raised the r2 to 0.47. BET was also the best single predictor of fine litterfall for the data set consisting of the 55 sites with information on nutrient return, accounting for 36% of the variation in fine litterfall. The addition of precipitation (PPT) raised the r2 to 0.46; addition of further climatic parameters did not improve the correlation.
These correlations are substantially smaller than those found by Meentemeyer et al. (1982) for a global data set including 84 sites. I believe that the difference is a consequence of the coarser climatic analysis I used and the much narrower ranges of climate and litter pro- duction found within the tropics. The correlations are similar to those observed by Brown and Lugo (1982) for tropical forest litterfall; they accounted for 44% of the variation in litterfall in their data set by using the ratio of temperature to precipitation and the logarithm of that ratio. Their predictor accounts for a somewhat smaller proportion of the variation in the data sets I
used (r2 = 0.28 for all fine litterfall, r2 = 0.39 for the subset of sites which included measurements of nu- trient return).
Nutrients and litterfall dry mass
The residuals from the climate regression were cal- culated and compared with the concentrations of in- dividual nutrients in litterfall. I assumed that the re- siduals from the climate regression represent variation that cannot be accounted for by climate; my ability to detect the possibility of nutrient limitation to fine lit- terfall is thus reduced to the extent that climate and nutrient limitation are confounded in tropical forests, and to the extent that the climate regression fails to account for all of the significant variation in litterfall due to climate. The sensitivity of this analysis is also reduced to the extent that either measurement errors or differences in methods contribute to the differences among sites in Table 1.
Results are summarized in Fig. 2. There was no sig- nificant correlation between nitrogen concentrations in litterfall and the residuals of litterfall dry mass (r2 for a second-order regression = 0.04) (Fig. 2A). Of the sev- en forests which had elevated dry mass/nitrogen ratios (Fig. 1 A), five are montane forests where the relatively low fine litterfall can be adequately predicted from cli- mate. The proximate limitation to litter production in these forests could still be nitrogen or another nutrient; that possibility cannot be tested with this analysis but will be discussed below.
The correlation between calcium concentrations and litter production is better than that for nitrogen (r2 = 0.16, P < .05), and the results for phosphorus (Fig. 2B) are still stronger. There is a highly significant second- order regression relating phosphorus concentrations in litterfall to the residuals from climate (r2 = 0.22, P < .002). As would be expected if phosphorus were lim- iting production, the relationship is strong and positive at low phosphorus concentrations (<0.04%) and ab- sent at higher concentrations (Fig. 2B).
Next, I used a stepwise multiple regression procedure (Statistical Analysis System 1979) to select the "best" combination of climate and nutrient parameters (in- cluding second- and third-order terms for each nu- trient) for predicting litter production. The best climate plus single nutrient regression was:
MASS = 9.63 + 0.42 x BET - 5.37 x PPT + 9.06 x P-4.6 x P2 (4)
where P is the phosphorus concentration (in percent) in litterfall. This regression explained 60% of the vari- ation in litterfall dry mass (P < .0001). The addition of the square of the calcium concentration raised the r2 to 0.65 (P < .0001); adding further terms did not improve the r2 value.
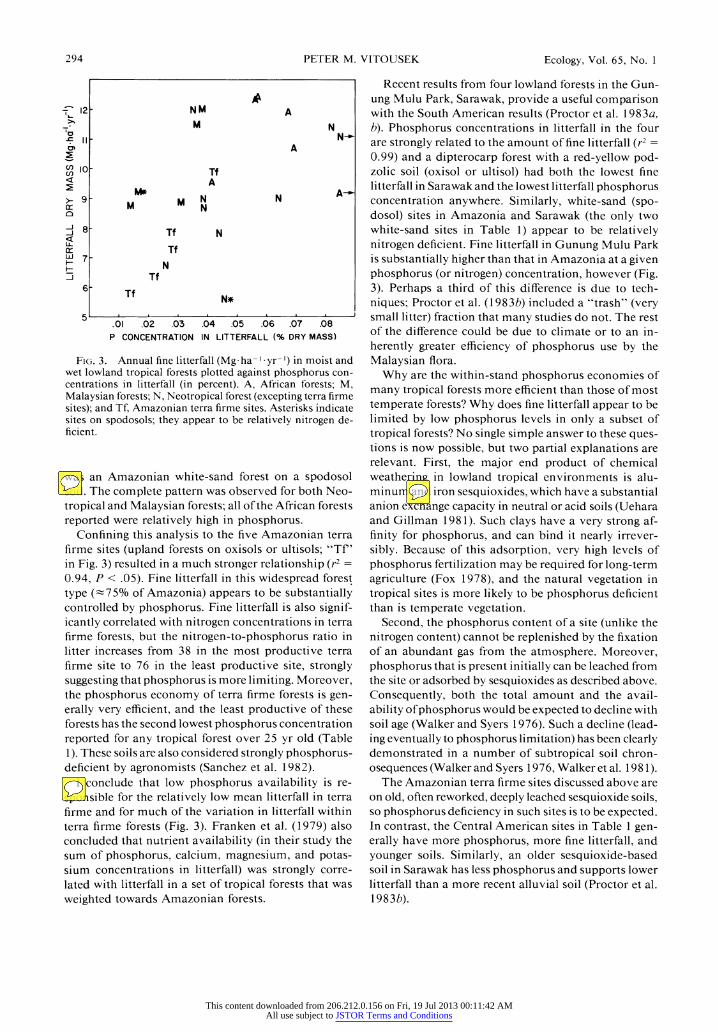
Finally, I compared fine litterfall dry mass with phos- phorus concentrations in litterfall for moist and wet lowland tropical forests (Fig. 3). This analysis was in-
This content downloaded from 206.212.0.156 on Fri, 19 Jul 2013 00:11:42 AMAll use subject to JSTOR Terms and Conditions
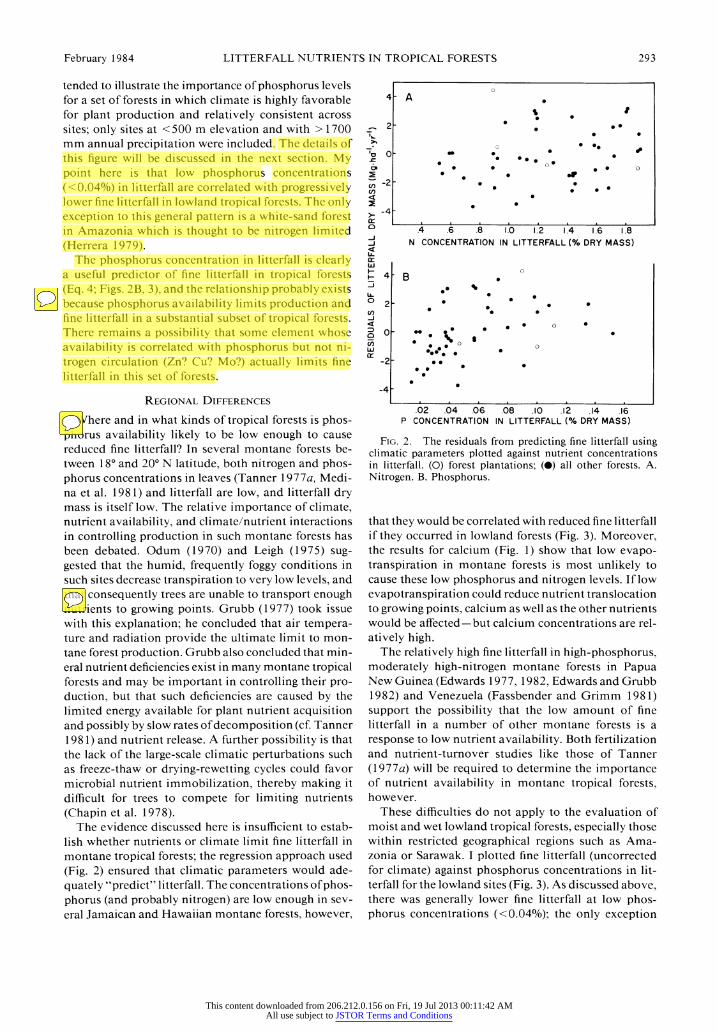
February 1984 LITTERFALL NUTRIENTS IN TROPICAL FORESTS 293
tended to illustrate the importance of phosphorus levels for a set of forests in which climate is highly favorable for plant production and relatively consistent across sites; only sites at <500 m elevation and with > 1700 mm annual precipitation were included. The details of this figure will be discussed in the next section. My point here is that low phosphorus concentrations (<0.04%) in litterfall are correlated with progressively lower fine litterfall in lowland tropical forests. The only exception to this general pattern is a white-sand forest in Amazonia which is thought to be nitrogen limited (Herrera 1979).
The phosphorus concentration in litterfall is clearly a useful predictor of fine litterfall in tropical forests (Eq. 4; Figs. 2B, 3), and the relationship probably exists because phosphorus availability limits production and fine litterfall in a substantial subset of tropical forests. There remains a possibility that some element whose availability is correlated with phosphorus but not ni- trogen circulation (Zn? Cu? Mo?) actually limits fine litterfall in this set of forests.
REGIONAL DIFFERENCES
Where and in what kinds of tropical forests is phos- phorus availability likely to be low enough to cause reduced fine litterfall? In several montane forests be- tween 180 and 20? N latitude, both nitrogen and phos- phorus concentrations in leaves (Tanner 1977a, Medi- na et al. 1981) and litterfall are low, and litterfall dry mass is itself low. The relative importance of climate, nutrient availability, and climate/nutrient interactions in controlling production in such montane forests has been debated. Odum (1970) and Leigh (1975) sug- gested that the humid, frequently foggy conditions in such sites decrease transpiration to very low levels, and that consequently trees are unable to transport enough nutrients to growing points. Grubb (1977) took issue with this explanation; he concluded that air tempera- ture and radiation provide the ultimate limit to mon- tane forest production. Grubb also concluded that min- eral nutrient deficiencies exist in many montane tropical forests and may be important in controlling their pro- duction, but that such deficiencies are caused by the limited energy available for plant nutrient acquisition and possibly by slow rates of decomposition (cf. Tanner 1981) and nutrient release. A further possibility is that the lack of the large-scale climatic perturbations such as freeze-thaw or drying-rewetting cycles could favor microbial nutrient immobilization, thereby making it difficult for trees to compete for limiting nutrients (Chapin et al. 1978).
The evidence discussed here is insufficient to estab- lish whether nutrients or climate limit fine litterfall in montane tropical forests; the regression approach used (Fig. 2) ensured that climatic parameters would ade- quately "predict" litterfall. The concentrations of phos- phorus (and probably nitrogen) are low enough in sev- eral Jamaican and Hawaiian montane forests, however,
4 A 0 % 0 0
2 *@...0
_o 0 - 00
F 0 0 @0. 0 0 0
0 0t
E(*) . .t
-4
o .4 .6 .8 1.0 1.2 1.4 1.6 1.8 j N CONCENTRATION IN LITTERFALL (% DRY MASS)
_ B -j 0 Li. 0 0 0 2 . Cl) 0 0 -j
02 04 06 0 0 0 0 ~O **'*
0 * * o
Nitron 0.00B. 0P h -2
0 0
-4
.02 .04 .06 .08 .10 .12 .14 .16 P CONCENTRATION IN LITTERFALL (% DRY MASS)
FIG. 2. The residuals from predicting fine litterfall using climatic parameters plotted against nutrient concentrations in litterfall. (0) forest plantations; (0) all other forests. A. Nitrogen. B. Phosphorus.
that they would be correlated with reduced fine litterfall if they occurred in lowland forests (Fig. 3). Moreover, the results for calcium (Fig. 1) show that low evapo- transpiration in montane forests is most unlikely to cause these low phosphorus and nitrogen levels. If low evapotranspiration could reduce nutrient translocation to growing points, calcium as well as the other nutrients would be affected-but calcium concentrations are rel- atively high.
The relatively high fine litterfall in high-phosphorus, moderately high-nitrogen montane forests in Papua New Guinea (Edwards 1977, 1982, Edwards and Grubb 1982) and Venezuela (Fassbender and Grimm 1981) support the possibility that the low amount of fine litterfall in a number of other montane forests is a response to low nutrient availability. Both fertilization and nutrient-turnover studies like those of Tanner (1977a) will be required to determine the importance of nutrient availability in montane tropical forests, however.
These difficulties do not apply to the evaluation of moist and wet lowland tropical forests, especially those within restricted geographical regions such as Ama- zonia or Sarawak. I plotted fine litterfall (uncorrected for climate) against phosphorus concentrations in lit- terfall for the lowland sites (Fig. 3). As discussed above, there was generally lower fine litterfall at low phos- phorus concentrations (<0.04%); the only exception
This content downloaded from 206.212.0.156 on Fri, 19 Jul 2013 00:11:42 AMAll use subject to JSTOR Terms and Conditions
294 PETER M. VITOUSEK Ecology, Vol. 65, No. 1
C12 NM A
M N o ~~~~~~~~~~~N-
A CX IO-_
U)O Tf E A
E 9 M M N N A 0
Tf N EL Tf
1i - H ~~~N
Ej 7Tf 6 ~ Tf N*
.01 .02 .03 .04 .05 .06 .07 .08 P CONCENTRATION IN LITTERFALL (% DRY MASS)
FIG. 3. Annual fine litterfall (Mg ha- yr-I) in moist and wet lowland tropical forests plotted against phosphorus con- centrations in litterfall (in percent). A, African forests; M, Malaysian forests; N, Neotropical forest (excepting terra firme sites); and Tf, Amazonian terra firme sites. Asterisks indicate sites on spodosols; they appear to be relatively nitrogen de- ficient.
was an Amazonian white-sand forest on a spodosol soil. The complete pattern was observed for both Neo- tropical and Malaysian forests; all of the African forests reported were relatively high in phosphorus.
Confining this analysis to the five Amazonian terra firme sites (upland forests on oxisols or ultisols; "Tf' in Fig. 3) resulted in a much stronger relationship (r2 =
0.94, P < .05). Fine litterfall in this widespread forest type (-75% of Amazonia) appears to be substantially controlled by phosphorus. Fine litterfall is also signif- icantly correlated with nitrogen concentrations in terra firme forests, but the nitrogen-to-phosphorus ratio in litter increases from 38 in the most productive terra firme site to 76 in the least productive site, strongly suggesting that phosphorus is more limiting. Moreover, the phosphorus economy of terra firme forests is gen- erally very efficient, and the least productive of these forests has the second lowest phosphorus concentration reported for any tropical forest over 25 yr old (Table 1). These soils are also considered strongly phosphorus- deficient by agronomists (Sanchez et al. 1982).
I conclude that low phosphorus availability is re- sponsible for the relatively low mean litterfall in terra firme and for much of the variation in litterfall within terra firme forests (Fig. 3). Franken et al. (1979) also concluded that nutrient availability (in their study the sum of phosphorus, calcium, magnesium, and potas- sium concentrations in litterfall) was strongly corre- lated with litterfall in a set of tropical forests that was weighted towards Amazonian forests.
Recent results from four lowland forests in the Gun- ung Mulu Park, Sarawak, provide a useful comparison with the South American results (Proctor et al. 1983a, b). Phosphorus concentrations in litterfall in the four are strongly related to the amount of fine litterfall (r2 =
0.99) and a dipterocarp forest with a red-yellow pod- zolic soil (oxisol or ultisol) had both the lowest fine litterfall in Sarawak and the lowest litterfall phosphorus concentration anywhere. Similarly, white-sand (spo- dosol) sites in Amazonia and Sarawak (the only two white-sand sites in Table 1) appear to be relatively nitrogen deficient. Fine litterfall in Gunung Mulu Park is substantially higher than that in Amazonia at a given phosphorus (or nitrogen) concentration, however (Fig. 3). Perhaps a third of this difference is due to tech- niques; Proctor et al. (1983b) included a "trash" (very small litter) fraction that many studies do not. The rest of the difference could be due to climate or to an in- herently greater efficiency of phosphorus use by the Malaysian flora.
Why are the within-stand phosphorus economies of many tropical forests more efficient than those of most temperate forests? Why does fine litterfall appear to be limited by low phosphorus levels in only a subset of tropical forests? No single simple answer to these ques- tions is now possible, but two partial explanations are relevant. First, the major end product of chemical weathering in lowland tropical environments is alu- minum and iron sesquioxides, which have a substantial anion exchange capacity in neutral or acid soils (Uehara and Gillman 1981). Such clays have a very strong af- finity for phosphorus, and can bind it nearly irrever- sibly. Because of this adsorption, very high levels of phosphorus fertilization may be required for long-term agriculture (Fox 1978), and the natural vegetation in tropical sites is more likely to be phosphorus deficient than is temperate vegetation.
Second, the phosphorus content of a site (unlike the nitrogen content) cannot be replenished by the fixation of an abundant gas from the atmosphere. Moreover, phosphorus that is present initially can be leached from the site or adsorbed by sesquioxides as described above. Consequently, both the total amount and the avail- ability of phosphorus would be expected to decline with soil age (Walker and Syers 1976). Such a decline (lead- ing eventually to phosphorus limitation) has been clearly demonstrated in a number of subtropical soil chron- osequences (Walker and Syers 1976, Walker et al. 1981).
The Amazonian terra firme sites discussed above are on old, often reworked, deeply leached sesquioxide soils, so phosphorus deficiency in such sites is to be expected. In contrast, the Central American sites in Table 1 gen- erally have more phosphorus, more fine litterfall, and younger soils. Similarly, an older sesquioxide-based soil in Sarawak has less phosphorus and supports lower litterfall than a more recent alluvial soil (Proctor et al. 1983b).
This content downloaded from 206.212.0.156 on Fri, 19 Jul 2013 00:11:42 AMAll use subject to JSTOR Terms and Conditions

February 1984 LITTERFALL NUTRIENTS IN TROPICAL FORESTS 295
CONCLUSIONS
The analyses of tropical litterfall and nutrient cycling summarized in Figs. 1-3 illustrate the far-from-origi- nal point that it is misleading to discuss mineral cycling in tropical forests as a single unified entity (cf. Grubb 1977, Ewel 1980, Jordan and Herrera 1981). These analyses also demonstrate that while many tropical forests are not characterized by "tight" within-stand nutrient economies (that in fact many forests cycle large amounts of phosphorus, nitrogen, and calcium rela- tively inefficiently) a substantial subset of tropical for- ests cycles phosphorus much more efficiently than most temperate forests. There is good, although indirect, evi- dence (Figs. 2B, 3) that litterfall in such forests is phos- phorus limited.
These conclusions differ somewhat from those of Jordan and Herrera (1981). They defined oligotrophic (infertile) and eutrophic (fertile) sites in terms of soil calcium concentrations, and they concluded that bio- mass, production, and mineral cycling do not differ substantially between intact oligotrophic and eutrophic forests. The major differences between these classes of forests that they identified are the existence of "nutrient conservation mechanisms" in intact oligotrophic for- ests and the greater ability of eutrophic forest ecosys- tems to support production following disturbance.
I suggest that soil calcium concentrations are an in- adequate index of the availability of potentially lim- iting nutrients. In fact, while Jordan and Herrera's oli- gotrophic site (an Amazonian terra firme forest) had low litterfall and among the most efficient within-stand phosphorus and calcium cycles in Table 1, their eu- trophic forest (a Puerto Rican lower montane forest) also had low litterfall and the fourth most efficient phosphorus cycle (but a calcium cycle near the median in efficiency) (Table 1). Further, results summarized here show that litter production and nutrient cycling clearly do differ between low- and high-phosphorus tropical forests (Figs. 1-3). The low phosphorus avail- ability that appears to be characteristic of geomor- phologically old tropical and subtropical soils could control both the existence of "nutrient conservation mechanisms" (sclerophylly, surface fine root mats) and the ability of sites to sustain productivity following disturbance.
ACKNOWLEDGMENTS
I thank F. Bernhard-Reversat, C. B. Davey, Y. R. Dom- mergues, P. J. Grubb, B. L. Haines, C. F. Jordan, P. A. Mat- son, D. Mueller-Dombois, J. Proctor, G. P. Robertson, and E. V. J. Tanner for directing me to information on tropical mineral cycling, for permission to quote unpublished results, and/or for critical comments on earlier drafts of this manu- script.
LITERATURE CITED
Adis, J., K. Furch, and U. Irmler. 1979. Litter production of a central Amazonian black water inundation forest. Tropical Ecology 20:236-245.
Arnason, J. T., and J. D. H. Lambert. 1982. Nitrogen cycling in the seasonally dry forest zone of Belize, Central America. In G. P. Robertson, R. Herrera, and T. H. Rosswall, editors. Nitrogen cycling in ecosystems of Latin America and the Caribbean. Plant and Soil 67:333-342.
Bernhard-Reversat, F. 1975. Recherches sur les cycles bio- geochemiques des lamentss majeurs en milieu forestier sub- equatorial (C6te d'Ivoire). Dissertation. Universit&de Paris- Sud, Paris, France.
1977. Recherches sur les variations stationelles des cycles biog&ochemiques en fort ombrophile de C6te d'I- voire. Cahiers ORSTOM (Office de la Recherche Scienti- fique et Technique Outre-mer), Serie Pedologie 15:175- 189.
1982a. Biogeochemical cycle of nitrogen in a semi- arid savanna. Oikos 38:321-332.
1982b. Les cycles biogeochemique des elements mi- neraux en plantations d'Eucalyptus camaldulensis et en for- et naturelle a Acacia seyal au Senegal. United States Agency for International Development-SEEF Project Report, Centre ORSTOM (Office de la Recherche Scientifique et Tech- nique Outre-mer) de Dakar, Senegal.
Bernhard-Reversat, F., and H. Poupon. 1980. Nitrogen cy- cling in a soil-tree system in a Sahelian savanna. Pages 363- 369 in T. Rosswall, editor. Nitrogen cycling in West African ecosystems. SCOPE-UNEP (Scientific Committee on Prob- lems of the Environment-United Nations Environmental Program) International Nitrogen Unit, Royal Swedish Academy of Sciences, Stockholm, Sweden.
Brassell, H. M., G. L. Unwin, and G. C. Stocker. 1980. The quantity, temporal distribution, and mineral element con- tent of litterfall in two forest types at two sites in tropical Australia. Journal of Ecology 68:123-139.
Bray, J. R., and E. Gorham. 1964. Litter production in forests of the world. Advances in Ecological Research 2: 101-157.
Brown, S., and A. E. Lugo. 1982. The storage and production of organic matter in tropical forests and their role in the global carbon cycle. Biotropica 14:161-187.
Chapin, F. S. 1980. The mineral nutrition of wild plants. Annual Review of Ecology and Systematics 11:233-260.
Chapin, F. S., R. J. Barsdate, and D. Bar&l. 1978. Phos- phorus cycling in Alaskan coastal tundra: a hypothesis for the regulation of nutrient cycling. Oikos 31:189-199.
Cole, D. W., and M. Rapp. 1980. Elemental cycling in for- ested ecosystems. Pages 341-409 in D. E. Reichle, editor. Dynamic properties of forest ecosystems. Cambridge Uni- versity Press, Cambridge, England.
Cornforth, I. S. 1970. Leaf-fall in a tropical rainforest. Jour- nal of Applied Ecology 7:603-608.
Devineau, J.-L. 1976. Donnees preliminaires sur la litiere et la chute des feuilles dans quelques formations foresti&res semi-d&cidueus de moyenne C6te-d'Ivoire. Oecologia Plan- tarum 11:375-395.
Dommergues, Y. 1963. Les cycles biogeochemique des ele- ments mineraux dans les formations tropicales. Bois et For- ets des Tropiques 87:19-25.
Edmisten, J. 1970. Preliminary studies of the nitrogen bud- get of a tropical forest. Pages H-21 I-H-214 in H. T. Odum and R. F. Pigeon, editors. A tropical rain forest. United States Atomic Energy Commission, Oak Ridge, Tennessee, USA.
Edwards, P. J. 1977. Studies of mineral cycling in a montane rain forest in New Guinea. II. The production and disap- pearance of litter. Journal of Ecology 65:971-992.
1982. Studies of mineral cycling in a montane rain forest in New Guinea. V. Rates of cycling in throughfall and litterfall. Journal of Ecology 70:807-827.
Edwards, P. J., and P. J. Grubb. 1982. Studies of mineral
This content downloaded from 206.212.0.156 on Fri, 19 Jul 2013 00:11:42 AMAll use subject to JSTOR Terms and Conditions

296 PETER M. VITOUSEK Ecology, Vol. 65, No. 1
cycling in a montane rain forest in New Guinea. IV. Soil characteristics and the division of mineral elements be- tween the vegetation and soil. Journal of Ecology 70:645- 666.
Egunjobi, J. K. 1974. Litterfall and mineralization in a teak (Tectona grandis) stand. Oikos 25:222-226.
Egunjobi, J. K., and F. E. Fasehun. 1972. Preliminary ob- servations on the monthly litter-fall and nutrient content of Pinus caribea L. litter. Nigerian Journal of Science 6:37- 45.
Egunjobi, J. K., and B. S. Onweluzo. 1979. Litterfall, min- eral turnover, and litter accumulation in Pinus caribea L. stands at Ibadan, Nigeria. Biotropica 11:251-255.
Enright, N. J. 1979. Litter production and nutrient parti- tioning in rainforest near Bulolo, Papua New Guinea. Ma- laysian Forester 42:202-207.
Ewel, J. J. 1976. Litterfall and leaf decomposition in a trop- ical forest succession in eastern Guatemala. Journal of Ecol- ogy 64:293-308.
1980. Tropical succession: manifold routes to ma- turity. Biotropica 12:2-7 (supplement).
Fassbender, H. W., and V. Grimm. 1981. Ciclos bioquim- icos en un ecosistema forestal de los Andes Occidentales de Venezuela. II. Producci6n y descomposicion de los re- siduos vegetales. Turrialba 31:39-47.
Folster, H., and G. de las Salas. 1976. Litter fall and min- eralization in three tropical evergreen stands, Colombia. Acta Cientifica Venezolana 27:196-202.
Fox, R. L. 1978. Studies on phosphorus nutrition in the tropics. Pages 169-187 in C. S. Andrew and E. J. Kamprath, editors. Mineral nutrition of legumes in tropical and sub- tropical soils. Commonwealth Scientific and Industrial Re- search Organization, Brisbane, Australia.
Franken, M. 1979. Major nutrient and energy contents of the litterfall of a riverine forest of central Amazonia. Trop- ical Ecology 20:211 -224.
Franken, M., V. Irmler, and H. Klinge. 1979. Litterfall in inundation, riverine, and terra firme forests of central Ama- zonia. Tropical Ecology 20:225-235.
Gessel, S. P., D. W. Cole, D. W. Johnson, and J. Turner. 1980. The nutrient cycles of two Costa Rican forests. Pages 23-44 in Progress in ecology. Volume III. Today and To- morrow's Printers and Publishers, New Delhi, India.
Golley, F. B., J. T. McGinnis, R. G. Clements, G. I. Child, and M. J. Duever. 1975. Mineral cycling in a tropical moist forest ecosystem. University of Georgia Press, Ath- ens, Georgia, USA.
Grubb, P. J. 1977. Control of forest growth and distribution on wet tropical mountains: with special reference to mineral nutrition. Annual Reviews of Ecology and Systematics 8: 83-107.
Haines, B., and R. B. Foster. 1977. Energy flow through litter in a Panamanian forest. Journal of Ecology 65:147- 155.
Herrera, R. A. 1979. Nutrient distribution and cycling in an Amazon caatinga forest on spodosols in southern Ven- ezuela. Dissertation. University of Reading, Reading, En- gland.
Hirose, T. 1975. Relations between turnover rate, resource utility and structure of some plant populations: a study of the matter budgets. Journal of the Faculty of Science, Uni- versity of Tokyo Section III, Volume XI, Part 11:355-407.
Holdridge, L. R., W. C. Grenke, W. H. Hatheway, T. Liang, and J. A. Tosi, Jr. 1971. Forest environments in tropical life zones: a pilot study. Pergamon, Oxford, England.
Hopkins, B. 1966. Vegetation of the Olokemeji Forest Re- serve, Nigeria. IV. The litter and soil with special reference to their seasonal changes. Journal of Ecology 54:687-703.
Ingestad, T. 1979. Nitrogen stress in birch seedlings. II. N, K, P, Ca, and Mg nutrition. Physiologia Plantarum 45:149- 157.
Jenny, H., S. P. Gessel, and F. T. Bingham. 1949. Com- parative study of decomposition rates of organic matter in temperate and tropical regions. Soil Science 68:419-432.
John, D. M. 1973. Accumulation and decay of litter and net production of forest in tropical West Africa. Oikos 24: 430-435.
Jordan, C. F. 1983. Productivity of tropical rain forest eco- systems and the implications for their use as future wood and energy sources. Pages 117-136 in F. B. Golley, editor. Tropical rain forest ecosystems: structure and function. Volume 14A. Ecosystems of the world. Elsevier, Amster- dam, The Netherlands.
In press. Biogeochemical cycles of an Amazonian rain forest. In N. Polunin, editor. Ecosystem theory and application. John Wiley and Sons, Chichester, England.
Jordan, C. F., and G. Escalante. 1980. Root productivity in an Amazonian rain forest. Ecology 61:14-18.
Jordan, C. F., and R. Herrera. 1981. Tropical rainforests: are nutrients really critical? American Naturalist 117:167- 180.
Jordan, C. F., W. Caskey, G. Escalante, R. Herrera, F. Mon- tagnini, R. Todd, and C. Uhl. 1982. The nitrogen cycle in a "terra firme" rain forest on oxisol in the Amazon territory of Venezuela. In G. P. Robertson, R. Herrera, and T. H. Rosswall, editors. Nitrogen cycling in ecosystems of Latin America and the Caribbean. Plant and Soil 67:325- 332.
Jordan, C. F., F. B. Golley, and J. Hall. 1980. Nutrient scavenging of rainfall by the canopy of an Amazonian rain forest. Biotropica 12:61-66.
Jordan, C. F., J. R. Kline, and D. S. Sasscer. 1972. Relative stability of mineral cycles in forest ecosystems. American Naturalist 106:237-253.
Jordan, C. F., J. R. Kline, and D. S. Sasscer. 1973. A simple model of strontium and manganese dynamics in a tropical rain forest. Health Physics 24:477-489.
Jung, G. 1969. Cycles biogeochemique dans un ecosyst&me de region tropicale s&che Acacia albida (Del.) sol ferrugi- neux tropical peu lessiv& (Dior.). Oecologia Plantarum 4: 195-210.
Kawanabe, S. 1977. A subtropical broad-leaved forest at Yona, Okinawa. Pages 268-279 in T. Shidei and T. Kira. Primary productivity of Japanese forests. University of To- kyo Press, Tokyo, Japan.
Kellman, M. C. 1970. Secondary plant succession in tropical montane Mindanao. School of Pacific Studies, Australian National University, Canberra, Australia.
Kenworthy, J. B. 1970. Water and nutrient cycling in a tropical forest. Pages 49-59 in J. R. Flenly, editor. The Water Relations of Malesian Forests, First Aberdeen-Hull Symposium on Malesian Ecology. Institute for South-East Asian Biology, University of Aberdeen, Aberdeen, Scot- land.
Keyes, M. R., and C. C. Grier. 1981. Above- and below- ground net production in 40 year old Douglas-fir stands on high and low productivity sites. Canadian Journal of Forest Research 11:599-605.
Kira, T., H. Ogawa, K. Yoda, and K. Ogino. 1967. Com- parative ecological studies on three main types of forest vegetation in Thailand. IV. Dry matter production, with special reference to the Khao Chong rain forest. Nature and Life in Southeast Asia 6:149-174.
Klinge, H. 1973. Root mass estimation in lowland tropical rain forests of central Amazonia, Brazil. I. Fine root masses of a pale yellow latosol and a giant humus podzol. Tropical Ecology 14:29-38.
1977. Fine litter production and nutrient return to the soil in three natural forest stands of eastern Amazonia. Geo-Eco-Trop 1: 159-167.
1978. Litter production in tropical ecosystems. Ma- laysian Nature Journal 30:415-422.
This content downloaded from 206.212.0.156 on Fri, 19 Jul 2013 00:11:42 AMAll use subject to JSTOR Terms and Conditions

February 1984 LITTERFALL NUTRIENTS IN TROPICAL FORESTS 297
Klinge, H., and W. A. Rodrigues. 1968. Litter production in an area of Amazonian terra firme forest. Amazoniana 1: 287-310.
Kunkel-Westphal, I., and P. Kunkel. 1979. Litter fall in a Guatemalan primary forest, with details of leaf shedding by some common tree species. Journal of Ecology 67:665- 686.
Lambert, J. D. H., J. T. Arnason, and J. L. Gale. 1980. Leaf- litter and changing nutrient levels in a seasonally dry trop- ical hardwood old forest, Belize. C.A. Plant and Soil 55: 429-443.
Laudelot, H., and J. Meyer. 1954. Les cycles d'616ments minerales et de mati&re organique en fort &quatoriale Con- golaise. Transactions of the Fifth International Congress on Soil Science 11:267-272.
Leigh, E. G. 1975. Structure and climate in tropical rain forest. Annual Review of Ecology and Systematics 6:67- 86.
Lim, M. T. 1978. Litterfall and mineral nutrient content of litter in Pasoh Forest Reserve. Malaysian Nature Journal 30:375-380.
Lugo, A. E., J. A. Gonzalez-Liboy, B. Cintr6n, and K. Dugger. 1979. Structure, productivity, and transpiration of a sub- tropical dry forest in Puerto Rico. Biotropica 10:278-291.
Lundgren, B. 1978. Soil conditions and nutrient cycling un- der natural and plantation forests in Tanzanian Highlands. Reports in Forest Ecology and Forest Soils 31, Swedish University of Agricultural Sciences, Uppsala, Sweden.
Madge, D. S. 1965. Leaf fall and litter disappearance in a tropical forest. Pedobiologia 5:273-288.
Maheut, J., and Y. R. Dommergues. 1960. Les teckeraies de Casamance: capacit& de production des peuplements caract&ristiques biologiques et maintien du potential pro- dutif des sols. Bois et Forets des Tropiques 70:25-42.
Malaisse, F., R. Freson, G. Goffinet, and M. Malaisse-Mous- set. 1975. Litter fall and litter breakdown in Miombo. Pages 137-152 in F. B. Golley and E. Medina, editors. Tropical ecological systems. Springer-Verlag, New York, New York, USA.
Medina, E., E. Cuevas, and P. L. Weaver. 1981. Compo- sicion foliar y transpiracion de especies leniosas de Pico del Este, Sierra de Luquillo, Puerto Rico. Acta Cientifica Ve- nezolana 32:159-165.
Medina, E., and M. Zelwer. 1972. Soil respiration in tropical plant communities. Pages 245-267 in P. M. Golley and F. B. Golley, editors. Tropical ecology with an emphasis on organic production. Indian National Science Academy, New Delhi, India.
Meentemeyer, V., E. 0. Box, and R. Thompson. 1982. World patterns and amounts of terrestrial plant litter production. BioScience 32:125-129.
Melillo, J. M., and J. R. Gosz. 1983. Interactions of bio- geochemical cycles in forest ecosystems. Pages 177-222 in B. Bolin and R. B. Cook, editors. The major biogeochemical cycles and their interactions. John Wiley and Sons, New York, New York, USA.
Miller, H. G., J. M. Cooper, and J. D. Miller. 1976. Effect of nitrogen supply on nutrients in litterfall and crown leach- ing in a stand of Corsican pine. Journal of Applied Ecology 13:233-248.
Neuberger, H., andJ. Cahir. 1969. Principles ofclimatology. Holt, Rinehart, and Winston, New York, New York, USA.
Nwoboshi, L. C. 1980. Nitrogen cycling in a teak plantation ecosystem in Nigeria. Pages 353-361 in T. Rosswall, editor. Nitrogen cycling in West African ecosystems. Scientific Committee on Problems of the Environment-United Na- tions Environmental Program (SCOPE-UNEP) Interna- tional Nitrogen Unit, Royal Swedish Academy of Sciences, Stockholm, Sweden.
Nye, P. H. 1961. Organic matter and nutrient cycles under moist tropical forest. Plant and Soil 13:333-345.
Odum, H. T. 1970. Summary: an emerging view of the ecological system at El Verde. Pages 1-191-1-289 in H. T. Odum and R. F. Pigeon, editors. A tropical rain forest. United States Atomic Energy Commission, Oak Ridge, Tennessee, USA.
O'Neill, R. V., and D. L. DeAngelis. 1980. Comparative productivity and biomass relations of forest ecosystems. Pages 411-449 in D. Reichle, editor. Dynamic properties of forest ecosystems. Cambridge University Press, Cam- bridge, England.
Parker, G. G. 1983. Throughfall and stemflow in the forest nutrient cycle. Advances in Ecological Research 13:57- 133.
Peace, W. J. H., and F. D. Macdonald. 1981. An investi- gation of the leaf anatomy, foliar mineral levels, and water relations of trees of a Sarawak forest. Biotropica 13: 100- 109.
Persson, H. 1980. Fine-root dynamics in a Scots Pine stand with and without near-optimum nutrient and water re- gimes. Acta Phytogeographica Suecia 68: 101 -110.
Pfadenhauer, J. 1979. Die Stickstoffmineralisation in b6den subtropischer Regenwalder in Stdbrasilien. Oecologia Plantarum 14:27-40.
Proctor, J. 1983, in press. Tropical forest litterfall. In S. Sutton, A. Chadwick, and T. C. Whitmore, editors. British Ecological Society Rain Forest Symposium. Blackwell Sci- entific, Oxford, England.
Proctor, J., J. M. Anderson, P. Chai, and H. W. Vallack. 1983a. Ecological studies in four contrasting lowland rain forests in Gunung Mulu Park, Sarawak. I. Forest environ- ment, structure, and floristics. Journal of Ecology 71:237- 260.
Proctor, J., J. M. Anderson, S. C. L. Fogden, and H. W. Vallack. 1983b. Ecological studies in four contrasting low- land rain forests in Gunung Mulu Park, Sarawak. II. Lit- terfall, litter standing crop, and preliminary observations on herbivory. Journal of Ecology 71:261-283.
Rodin, L. E., and N. I. Bazilevich. 1967. Production and mineral cycling in terrestrial vegetation. Oliver and Boyd, Edinburgh, Scotland.
Rozanov, B. G., and I. M. Rozanova. 1964. The biological cycle of nutrient elements of bamboo in the tropical forests of Burma. Botanischeskii Zhurnal 49:348-357.
Sanchez, P. A., D. E. Bandy, J. H. Villachica, and J. J. Ni- cholaides. 1982. Amazon Basin soils: management for continuous crop production. Science 216:821-827.
Snedaker, S. C. 1980. Successional immobilization of nu- trients and biologically mediated recycling in tropical for- ests. Biotropica 12:16-22 (Supplement).
Stachurski, A., and J. R. Zimka. 1975. Methods of studying forest ecosystems: leaf area, leaf production, and withdraw- al of nutrients from leaves of trees. Ekologia Polska 23:637- 648.
Statistical Analysis System. 1979. SAS User's Guide. SAS Institute, Cary, North Carolina, USA.
Sukachev, V., and N. Dylis. 1964. Fundamentals of forest biogeocoenology. Oliver and Boyd, Edinburgh, Scotland.
Swift, M. J., A. Russell-Smith, and T. J. Perfect. 1981. De- composition and mineral nutrient dynamics of plant litter in a regenerating bush-fallow in subhumid tropical Nigeria. Journal of Ecology 69:981-995.
Tanner, E. V. J. 1977a. Four montane rain forests of Ja- maica: a quantitative characterization of the floristics, the soils and the foliar mineral levels, and a discussion of the interrelations. Journal of Ecology 65:883-918.
1977b. Mineral cycling in montane rain forests in Jamaica. Dissertation. Cambridge University, Cambridge, England.
198 1. The decomposition of leaf litter in Jamaican montane rain forests. Journal of Ecology 69:263-275.
Turner, J. 1977. Effect of nitrogen availability on nitrogen cycling in a Douglas-fir stand. Forest Science 23:307-316.
This content downloaded from 206.212.0.156 on Fri, 19 Jul 2013 00:11:42 AMAll use subject to JSTOR Terms and Conditions

298 PETER M. VITOUSEK Ecology, Vol. 65, No. 1
Uehara, G., and G. Gillman. 1981. The mineralogy, chemistry, and physics of tropical soils with variable charge clays. Westview, Boulder, Colorado, USA.
Vitousek, P. M. 1982. Nutrient cycling and nutrient use efficiency. American Naturalist 119:553-572.
Vitousek, P. M., J. R. Gosz, C. C. Grier, J. M. Melillo, and W. A. Reiners. 1982. A comparative analysis of potential nitrification anitritrate mobility in forest ecosystems. Eco- logical Monographs 52:155-177.
Walker, T. W., and J. K. Syers. 1976. The fate of phosphorus during pedogenesis. Geoderma 15:1-19.
Walker, J., C. H. Thompson, I. F. Fergus, and B. R. Tunstall.
1981. Plant succession and soil development in coastal sand dunes of subtropical eastern Australia. Pages 107-131 in D. C. West, H. H. Shugart, and D. B. Botkin, editors. Forest succession: concept and application. Springer-Ver- lag, New York, New York, USA.
Wanner, H. 1970. Soil respiration, litterfall, and productiv- ity of tropical rain forests. Journal of Ecology 58:543-547.
Zonn, S. V., and C.-K. Li. 1962. Dynamics ofthe breakdown of litter and humus, and seasonal changes in their ash com- position, in two types of tropical biogeocoenoses. Soobsh- cheniya Lab. Lesoved. Moskva 6:144-152.
This content downloaded from 206.212.0.156 on Fri, 19 Jul 2013 00:11:42 AMAll use subject to JSTOR Terms and Conditions





























