Embed Size (px)
Citation preview

Lung dendritic cells are primed by inhaled particulate antigens, and retain
MHC class II/antigenic peptide complexes in hilar lymph nodes for
a prolonged period of time
QUYNH VU, KARIN M. MCCARTHY, JOANNE M. MCCORMACK & EVELINE E. SCHNEEBERGER
Molecular Pathology Unit, Massachusetts General Hospital, Charlestown, MA, USA
SUMMARY
Intratracheal (IT) administration of heat-killed Listeria monocytogenes (HKL) results in
an influx of macrophage and dendritic cell (DC) precursors into the lung interstitium.
Low-density, FcR+, interstitial lung cells isolated from rats instilled 24 hr before with
HKL or vehicle alone, were >90% Mar1+. After culturing with granulocyte–macrophage
colony-stimulating factor (GM-CSF) for 3 days, up to 24% of the loosely adherent cells were
DC that stimulated allogeneic T-cell proliferation in an mixed lymphocyte reaction (MLR)
assay. After only an overnight incubation with GM-CSF, however, the capacity of interstitial
Mar1+ cells to stimulate HKL immune T-cell proliferation without exogenous antigen was
low. By contrast, when DC were isolated as major histocompatibility complex (MHC) class
II+ cells from rat lungs at 1, 3, 7 and 14 days after HKL instillation and cultured overnight
with GM-CSF, their antigen presentation capacity without added exogenous antigen was
robust, but declined over the 2-week period. Interestingly, hilar lymph node DC maintained
their HKL antigen-presenting capacity for up to 2 weeks after instillation of HKL. Following
IT administration of PKH-26 labelled HKL, fluorescent or immunolabelled organisms were
detected in OX62+ DC in airway epithelium, lung interstitium and hilar lymph nodes in situ
and in MHC class II+ DC isolated from these sites. We conclude that newly immigrated
Mar1+ lung DC precursors, while efficient in endocytosing particulate antigens, are incapable
of eliciting a significant proliferative response from HKL-sensitized T cells. By contrast,
MHC class II+ DC isolated from lungs and incubated overnight with GM-CSF induce
vigorous antigen-specific T-cell proliferation. Antigen-loaded lung DC in hilar lymph nodes
maintain their antigen presentation capacity for up to 2 weeks.
INTRODUCTION
Alveolar macrophage-mediated phagocytosis, combined
with the activity of the mucociliary escalator, efficiently
transports particulates out of the lung and disposes
of inhaled pathogens and airborne antigens.1 However,
if the inhaled antigenic load overwhelms this first line
of defence, immunological mechanisms including both
humoral and cell-mediated immunity serve to protect the
lung from the deleterious effects of the inhaled pathogens.
To initiate an immune response in the lung requires that
antigens traverse the epithelial barriers that line the lung
and its airways. Soluble antigens may be transported to the
lung interstitium via epithelial cell pinocytotic vesicles in
the non-perturbed lung,2 or via the paracellular pathway in
inflammatory states.3 Particulate antigens, by contrast, are
carried into the lung interstitium and from thence to local
lymph nodes via migratory cells, that include macrophages,4
and neutrophils.5 To initiate cell-mediated immunity, how-
ever, requires not only that antigens be processed and pre-
sented to T cells in the context of major histocompatibility
complex (MHC) class II antigen, but also that the necessary
co-stimulatory signals be provided,6 attributes lacking in
both neutrophils and alveolar macrophages.7,8
The accessory cell that uniquely activates naı̈ve T-cells,
is the dendritic cell (DC), which was originally characterized
Received 13 August 2001; revised 21 November 2001; accepted
12 December 2001.
Correspondence: Dr Eveline E. Schneeberger, Molecular
Pathology Unit, Massachusetts General Hospital East, 149 13th
Street, Charlestown, MA 02129, USA. E-mail: schneebe@
helix.mgh.harvard.edu
Immunology 2002 105 488–498
# 2002 Blackwell Science Ltd488

as a large motile cell endowed with abundant surface MHC
class II antigen and capable of only limited endocytosis.9
This raised the question of how cell-mediated immunity
to particulate antigens, including bacteria and myco-
bacteria, could be initiated by these cells. Data from several
studies suggested the possibility that antigenic peptides
may be released by macrophages during phagocytosis of
particulate antigens and that they then bound directly to
MHC class II on the surface of DC.10,11 Although pos-
sible, this inherently inefficient process is unlikely to be
the primary mechanism whereby peptides derived from
particulate antigens are presented to T cells.
With the development of methods for the in vitro
generation of DC from bone marrow precursors,12 it
became apparent that DC, macrophages and neutrophils
are derived from a common bone marrow progenitor cell.13
Furthermore, early in their differentiation, DC share
a number of phenotypic features with macrophages,
including the expression of FcR14 and the ability to ingest
particulates including bacteria,14,15 yeast,16 apoptotic
cells17,18 and tumour cells.19 Our recent studies have shown
that DC precursors, and not circulating mature DC, enter
the interstitial compartment of the lung,20,21 in part, by
CD18-dependent mechanisms.22 Dendritic cells isolated
from the lung and airways are phenotypically at varying
stages of differentiation and/or possibly of different
lineages.23–26
In the present study, data are presented indicating that
immature DC are among the cells that phagocytose
intratracheally (IT) administered particulate, heat-killed
Listeria monocytogenes (HKL). After engulfing the organ-
isms, the cells migrate from the lung to hilar lymph nodes,
where they adopt a mature DC phenotype. Following the
IT administration of HKL, DC isolated from airway
epithelium, the lung interstitium and from hilar lymph
nodes contain visible intracellular fragments of PKH-
26-labelled bacteria. DC isolated from the lung interstitium
and hilar lymph nodes stimulate HKL immune T-cell
proliferation in the absence of exogenous antigen for more
than a week after IT instillation. Furthermore, DC in
hilar lymph nodes retain antigenic peptides on their sur-
face for up to 2 weeks as shown by their ability to stimu-
late HKL-immune T-cell proliferation in the absence of
exogenously added bacteria.
MATERIALS AND METHODS
Animals
Pathogen-free, 6–8-week-old female Lewis rats and female
Long Evans rats (180–200 g) were obtained from Charles
River Breeding Laboratories (Kingston, NY). Animals,
housed in restricted access research animal care facilities in
Massachusetts General Hospital, were permitted free access
to food and water and underwent monthly monitoring
for viral infections. Studies were conducted in accordance
with Massachusetts General Hospital and National
Institutes of Health guidelines for the care and use of
laboratory animals.
Reagents and antibodies
Bovine serum albumin (BSA), fraction V, was obtained
from Intergen Co. (Purchase, NY). Enzymes included
collagenase (CLS 1; Worthington Biochemical Corp, Free-
hold, NJ), DNAase I (Sigma Chemical Co., St Louis, MO)
and dispase, grade II (Boehringer Mannheim Biochemicals,
Indianapolis, IN). Fetal bovine serum (FBS), PKH-26 cell
linker kit and hen egg lysozyme (HEL) were from Sigma
Chemical Co. SeaPlaque GTG agarose (low melting point
agarose) was from FMC Bioproducts (Rockland, ME).
Cytokines included recombinant interleukin-2 (rIL-2; a gift
from Hoffman-La Roche, Nutley, NJ) and murine recom-
binant granulocyte–macrophage colony-stimulating factor
(rGM-CSF; a gift from Genetics Institute, Andover, MA).
RPMI-1640 medium was from Cellgro Mediatech
(Herndon, VA). Gentamicin and sheep red blood cells
(SRBC) were from BioWhittaker (Walkersville, MD).
Monoclonal antibodies (mAb) included OX6 (anti-MHC
Class II), OX33 (anti-B cell), OX19 (anti-CD5, thymocytes
and peripheral T cells), OX52 (anti-T cell), OX62
(recognizes a subset of DC and cd T cells) (Pharmingen,
San Diego, CA) and Mar1 (antimonocyte/macrophage)
(Seikagaku, Tokyo, Japan). Rabbit anti-SRBC immuno-
globulin G (IgG) was from Diamedix Corp. (Miami, FL)
and fluorescein isothiocyanate (FITC) -labelled goat
anti-mouse IgG was from Sigma. Goat anti-mouse
IgG-coated microbeads were from Miltenyi Biotec Inc.
(Auburn, CA) and Glycergel was from Dako Corporation
(Carpinteria, CA).
Heat-killed Listeria monocytogenes (HKL).
Listeria monocytogenes was obtained from the Bacterio-
logy Laboratory of the Massachusetts General Hospital,
(Boston, MA) and heat-inactivated in a 63u water bath
for 90 min. The efficacy of heat killing was assessed
by failure of the bacteria to grow on blood agar plates.
The concentration of the organisms was determined
using McFarland nephelometric standards. Aliquots
(109 organisms/ml of saline) were stored at x20u.
Labelling of HKL
One millilitre of HKL (1r109 organisms/ml) was labelled
with 1 ml of 4r10x6M PKH-26 in diluent (supplied by
the manufacturer) and incubated for 5 min at room tem-
perature. The reaction was stopped by adding 2 ml FBS.
Labelled HKL were washed three times with PBS, utilizing
a fresh tube each time to ensure the complete removal of
free dye.
Experimental protocol
In situ immunolabelling studies. Six groups of two rats
each were anaesthetized with chloral hydrate (360 mg/kg)
and instilled IT with PKH-26-labelled HKL, 2r109
organisms in 300 ml of instillation solution (4 mg/ml Evans
blue dye, 0.4 mg/ml lyophilized rat serum protein in PBS)
to localize the injectate. Control rats were given 300 ml of
instillation solution alone. One group of rats was re-instilled
IT with 2r109 HKL 2 weeks later, as described. Rats were
489Lung dendritic cell priming by inhaled particulate antigens
# 2002 Blackwell Science Ltd, Immunology, 105, 488–498

killed 1, 3, 7, 14, or 17 days later. Frozen sections of lungs,
airways and hilar lymph nodes were immunolabelled using
OX6 mAb and FITC-tagged goat anti-mouse polyclonal
antibody and examined by fluorescence microscopy. Alter-
natively, frozen sections were immunoperoxidase labelled
using Mar1, OX62 and OX6 mAb to localize macro-
phages, DC precursors and DC, respectively, in the lungs
of experimental and control rats. In addition, double
immunolabelling with immuno-alkaline phosphatase to
detect Mar1, OX62 and OX6 mAb binding and immuno-
peroxidase to detect rabbit anti-HKL polyclonal anto-
body binding, was used to localize intracellular HKL in
macrophages, DC precursors and DC, respectively.
In vitro studies. DC and DC precursors were isolated
by two different protocols. In the first, OX6+ DC were
isolated from the lung parenchyma (exclusive of major
airways) and hilar lymph nodes from five groups of three
to four rats each that were instilled and killed at 1, 3, 7,
14 and 17 days as described above. They were tested in
antigen presentation assays without exogenously added
HKL. In further experiments OX6+ DC were isolated
separately from the airway epithelium, the lung parenchyma
and hilar lymph nodes, 24 hr after the instillation of
PKH-26-labelled HKL. They were immunolabelled for
MHC class II antigen and examined by immunofluores-
cence for the presence of HKL. In the second protocol,
experiments were designed to isolate FcR+ DC precursors
and macrophages from the lung interstitium of rats instilled
with 2r109 HKL or vehicle alone 24 hr prior to killing.
The cells were then cultured with 500 U/ml of GM-CSF for
0–3 days and their phenotype was determined by immuno-
peroxidase staining using OX6, OX62 and Mar1 mAbs
and by mixed lymphocyte reaction (MLR) assays.
Isolation of DC from airway epithelium, lung interstitium
and hilar lymph nodes
A previously described microdissection procedure was
used.24 Briefly, rats were anaesthetized with chloral hydrate
(480 mg/kg). After exsanguination via the abdominal aorta,
the thorax was opened and the hilar lymph nodes were
harvested. The lungs were perfused via the pulmonary
artery with 60–180 ml of 1.0 mM sodium ethylenediamine-
tetraacetic acid (Na2EDTA) in PBS, pH 7.4 and then
lavaged with eleven 5-ml aliquots of 1.0 mM Na2EDTA
in PBS. Ten millilitres of agarose solution (1% agarose,
2.5 U/ml dispase, 50 U/ml DNAase, 5% FBS in PBS) at
37u was infused into the airways and lungs. After tying
off the trachea, the lungs and trachea were immersed in
ice-cold PBS for 30 min to solidify the agarose. Under
sterile conditions, the heart was removed and the lung
parenchyma was dissected from the tracheo-bronchial tree.
DC were isolated separately from the dissected tracheo-
bronchial tree, the lung fragments and hilar lymph nodes
as described below.
Interstitial lung DC. The lung fragments were minced
and incubated in an enzyme solution containing 150 U/ml
collagenase, 50 U/ml DNAase in complete medium (CM)
(RPMI-1640, 5% FBS, 50 mM 2-mercaptoethanol, 1%
gentamycin) in a shaking water bath for 90 min at 37u.After passing the fragments through an 80-mesh stain-
less steel screen and filtering through four layers of gauze,
low-density cells were harvested as described below.
Airway epithelial DC. The isolated tracheo-bronchial
tree, filled with dispase/agarose, was incubated in CM in a
humidified incubator in 5% CO2/95% air for 1 hr at 37u.24
The tracheo-bronchial epithelium, with included DC, was
then flushed out with three 35-ml aliquots of 5% FBS in
PBS (FPBS). The cell suspension was passed through four
layers of gauze and washed twice with FPBS. To separate
DC from airway epithelium, the cells were resuspended
in 10 mM Na2EDTA in PBS and incubated in a shaking
water bath for 30 min at 37u. The single-cell suspension
was washed twice in FPBS and the low-density cells were
retrieved as described below.
Lymph node DC. Hilar lymph nodes were perfused with
collagenase (100 U/ml) in RPMI-1640.27 The released cells
were collected in a 50-ml tube containing 10 ml of ice-cold
RPMI. The lymph nodes were then teased with forceps
in fresh collagenase solution and the released cells were
added to the collection tube. Concentrated collagenase
solution (400 U/ml) was added to the residual lymph
node fragments and incubated for 30 min at 37u. After
vigorous pipetting, the cell suspension was transferred to the
collection tube and centrifuged at 260 g. The cells were
re-suspended in 10 ml PBS, filtered through a cell strainer
and washed twice in FPBS. Low-density cells were harvest
as described below.
Retrieval of low-density cells
BSA solutions were prepared as described,28 to yield a pH
of 7.35t0.05 and a density of 1.082 g/cm3, as determined
by refractometry (model ABBE-3L; Bausch and Lomb
Inc. Instruments and Systems Division, Rochester, NY).
Approximately 1r107x3r107 cells/ml were mixed with
the dense BSA solution and 2.5 ml aliquots were overlaid
with 1 ml of dilute BSA (dense BSA solution: PBS, 2 : 1 v/v)
and centrifuged at 12 000 g for 30 min at 4u. Low-density
cells at the interface were harvested and washed twice in
PBS. The cells were incubated overnight in CM supple-
mented with 250 U/ml GM-CSF in 5% CO2, 95% room air
at 37u. Because of the low yield, DC retrieved from the
airway epithelium were examined by immunofluorescence
only; functional studies were conducted on interstitial lung
and lymph node DC.
After overnight incubation, loosely adherent lung and
lymph node cells were harvested, and washed twice with
PBS. Lymph node DC (85% purity) were used directly in
antigen presentation assays (see below). Interstitial lung
DCs were further purified by immunomagnetic separation.
Briefly, cells were incubated with OX6 mAb (1 : 100
dilution) for 15 min at 4u. After rinsing twice with MACS
buffer (5 mM Na2EDTA, 0.5% BSA, 50 U/ml DNAse in
PBS), the cells were incubated with goat anti-mouse IgG-
coated microbeads for 15 min at 4u. OX6+ DCs were
isolated by positive selection on a MACS separator
490 Q. Vu et al.
# 2002 Blackwell Science Ltd, Immunology, 105, 488–498

(Miltenyi Biotech Inc.), yielding a DC fraction of >90%
purity.
Isolation of FcR+ immature DC from
the lung interstitium
To isolate FcR+ immature DC from the lung interstitium,
low-density cells were fractionated into FcR+ and FcRx
cells by rosetting with SRBC coated with rabbit anti-
SRBC polyclonal antibody.24 The rosetted cell pellet was
resuspended in 1 ml of CM, layered onto 2.5 ml dense
BSA solution and centrifuged at 9500 g for 15 min at 4u.The pelleted FcR+ cells were retrieved and the red blood
cells were lysed. Cell debris was removed by centrifuga-
tion with Histopaque-1077 at 800 g for 15 min at 4u.The cells were then incubated with a mixture of OX-19,
OX-52, HIS-48 mAbs, all at 20 mg/ml, followed by
incubation with goat anti-mouse IgG-coated microbeads;
each incubation was for 15 min at 4u. Macrophages and
immature DC were isolated by negative selection using
immunomagnetic separation. The harvested cells were
>90% Mar1+.
Immune T-cell isolation
HKL or HEL immune T cells were generated by immuniz-
ing Lewis rats at the base of the tail with either 2r107 HKL
or 100 mg HEL in complete Freund’s adjuvant.11 After
2 weeks, inguinal lymph nodes were harvested and the
immune T cells were isolated and cultured with rIL-2
(100 U/ml). They were re-stimulated every 3 weeks with
HKL or HEL, respectively, using irradiated, syngeneic
spleen cells as a source of APC.
MLR assay
A primary one-way MLR was conducted using isolated,
irradiated pulmonary DC (104/well) from Lewis rats and
splenic T cells (5r104/well) from Long Evans rats obtained
by separation on nylon wool columns.29 Residual MHC
class II+ cells were removed by immuno-panning with
OX-6-coated plates.
Antigen presentation assay
Lung interstitial and hilar lymph node DC from experi-
mental and control rats were irradiated (1000 rads), washed
and plated (104, 103, 102 cells/well) in triplicate in 96-well,
flat-bottom culture plates (Becton Dickinson Co., Lincoln
Park, NJ) in CM. HKL immune T cells (5r104 cells/well)
were added without antigen. To test for the specificity of
HKL antigen presentation, HEL immune T cells, without
added antigen, were substituted. Positive controls included
the addition of exogenous HKL (4r106/well) or HEL
(30 mg/well). The plates were incubated in 5% CO2, 95% O2
for 72 hr at 37u. The cells were pulsed with [3H]thymidine
(1 mCi/well, specific activity 80–90 Ci/mmol) (New England
Nuclear, Boston, MA) for 6 hr, harvested using a cell
harvester (Skatron AS, Lierbyen, Norway), and counted
in a Tri-Carb liquid scintillation spectrometer (Packard
Instrument Co. Inc., Downers Grove, IL). The per cent
relative response30 was calculated as: ({c.p.m.r[(immune
T cells+ antigen-pulsed DC)/(immune T cells+antigen-
pulsed DC+antigen)]}x{[(immune T cells+control DC)/
(immune T cells+control DC+antigen)]})r100.
Immunofluorescence
Frozen 4-mm sections of lung and lymph nodes were
prepared. These and cytocentrifuged preparations of
isolated DC were air dried and fixed in acetone. Immuno-
labelling used OX6 mAb followed by goat anti-mouse
IgG-FITC. FITC-labelled cells (green) and the engulfed
PKH-26-labelled HKL (red) were examined by fluorescence
microscopy using an Olympus epifluorescent micro-
scope and a dichroic mirror system providing the
appropriate wavelengths for fluorescein and rhodamine,
respectively.
Immunocytochemistry
Frozen sections were cut and fixed as described above.
They were immunolabelled by an indirect avidin–biotin
immunoperoxidase method (Vector Laboratories, Inc.,
Burlingame, CA) with appropriately diluted primary mAbs
(OX6, OX62, Mar1) or rabbit anti-HKL polyclonal anti-
body, post-fixed in 2% paraformaldehyde in PBS, counter-
stained with Gill no. 2 haematoxylin and the coverslips
were mounted with glycergel. For double immunolabelling,
alkaline phosphatase and peroxidase-tagged secondary
antibodies were used to detect OX6, OX62 and Mar1
(blue) and anti-HKL (red) binding.
Morphometry
Immunolabelled cells in frozen sections were counted in five
random fields of pulmonary alveoli using a 40r objective
and a 1-cm2 graticule divided into 10r10 squares.
Because of variations in the relative proportion of tissue
to alveolar space, cell counts were corrected for the fraction
of alveolar space included in the area measured, by counting
the squares of the graticule devoid of tissue and subtracting
this value from 100. The number of positively stained cells
was then divided by the number of squares that contained
tissue and the quotient was multiplied by 100. To obtain the
number of cells per cm2, the mean cell number was divided
by 0.000625 (1 cm/40).2
RESULTS
The number of OX6+ and Mar1+ cells is increased in
the lung interstitium following the intratracheal
administration of HKL
To determine the extent to which the IT administration of
HKL induced an influx of macrophage and DC precursors
into the lung interstitium, Lewis rats were instilled with
2r109 HKL and killed at 1, 7 and 14 days later. A fourth
group received a second IT dose of 2r109 HKL on day 14
and was then killed 3 days later. Sections from each of the
five lobes of the lung were immunolabelled using OX6
or Mar1 mAb. The number of immunolabelled cells in
491Lung dendritic cell priming by inhaled particulate antigens
# 2002 Blackwell Science Ltd, Immunology, 105, 488–498
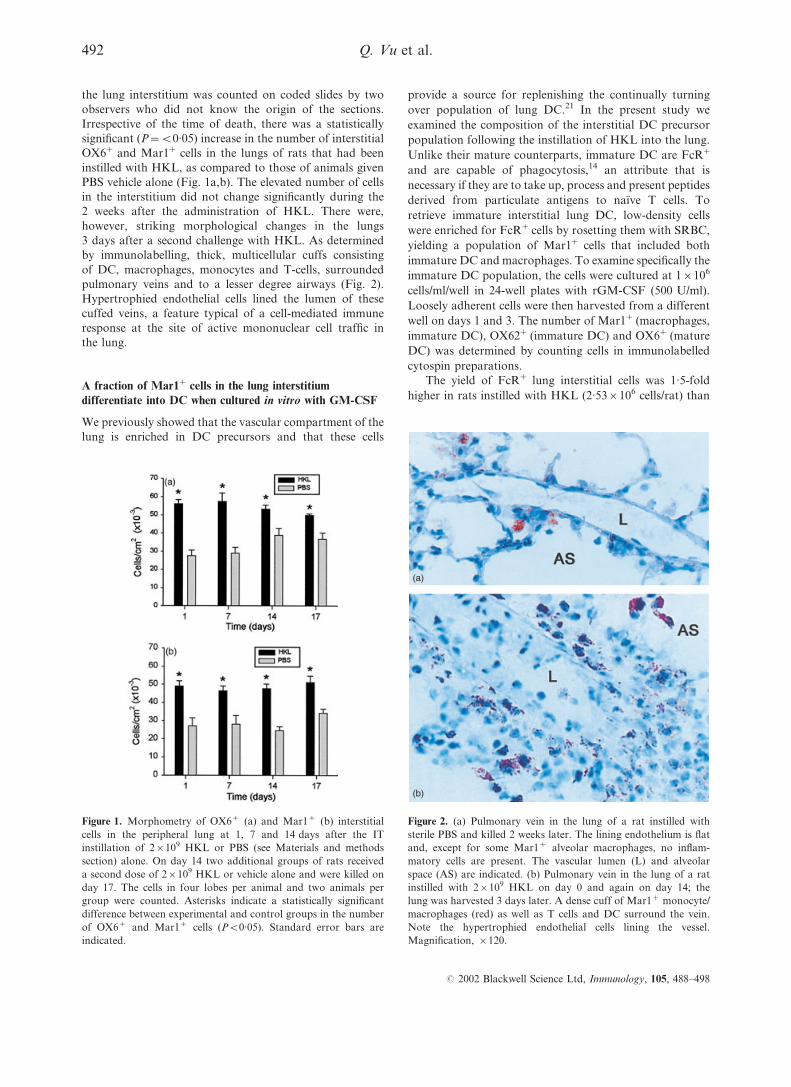
the lung interstitium was counted on coded slides by two
observers who did not know the origin of the sections.
Irrespective of the time of death, there was a statistically
significant (P=<0.05) increase in the number of interstitial
OX6+ and Mar1+ cells in the lungs of rats that had been
instilled with HKL, as compared to those of animals given
PBS vehicle alone (Fig. 1a,b). The elevated number of cells
in the interstitium did not change significantly during the
2 weeks after the administration of HKL. There were,
however, striking morphological changes in the lungs
3 days after a second challenge with HKL. As determined
by immunolabelling, thick, multicellular cuffs consisting
of DC, macrophages, monocytes and T-cells, surrounded
pulmonary veins and to a lesser degree airways (Fig. 2).
Hypertrophied endothelial cells lined the lumen of these
cuffed veins, a feature typical of a cell-mediated immune
response at the site of active mononuclear cell traffic in
the lung.
A fraction of Mar1+ cells in the lung interstitium
differentiate into DC when cultured in vitro with GM-CSF
We previously showed that the vascular compartment of the
lung is enriched in DC precursors and that these cells
provide a source for replenishing the continually turning
over population of lung DC.21 In the present study we
examined the composition of the interstitial DC precursor
population following the instillation of HKL into the lung.
Unlike their mature counterparts, immature DC are FcR+
and are capable of phagocytosis,14 an attribute that is
necessary if they are to take up, process and present peptides
derived from particulate antigens to naı̈ve T cells. To
retrieve immature interstitial lung DC, low-density cells
were enriched for FcR+ cells by rosetting them with SRBC,
yielding a population of Mar1+ cells that included both
immature DC and macrophages. To examine specifically the
immature DC population, the cells were cultured at 1r106
cells/ml/well in 24-well plates with rGM-CSF (500 U/ml).
Loosely adherent cells were then harvested from a different
well on days 1 and 3. The number of Mar1+ (macrophages,
immature DC), OX62+ (immature DC) and OX6+ (mature
DC) was determined by counting cells in immunolabelled
cytospin preparations.
The yield of FcR+ lung interstitial cells was 1.5-fold
higher in rats instilled with HKL (2.53r106 cells/rat) than
(a)
(b)
Figure 2. (a) Pulmonary vein in the lung of a rat instilled with
sterile PBS and killed 2 weeks later. The lining endothelium is flat
and, except for some Mar1+ alveolar macrophages, no inflam-
matory cells are present. The vascular lumen (L) and alveolar
space (AS) are indicated. (b) Pulmonary vein in the lung of a rat
instilled with 2r109 HKL on day 0 and again on day 14; the
lung was harvested 3 days later. A dense cuff of Mar1+ monocyte/
macrophages (red) as well as T cells and DC surround the vein.
Note the hypertrophied endothelial cells lining the vessel.
Magnification, r120.
Figure 1. Morphometry of OX6+ (a) and Mar1+ (b) interstitial
cells in the peripheral lung at 1, 7 and 14 days after the IT
instillation of 2r109 HKL or PBS (see Materials and methods
section) alone. On day 14 two additional groups of rats received
a second dose of 2r109 HKL or vehicle alone and were killed on
day 17. The cells in four lobes per animal and two animals per
group were counted. Asterisks indicate a statistically significant
difference between experimental and control groups in the number
of OX6+ and Mar1+ cells (P<0.05). Standard error bars are
indicated.
492 Q. Vu et al.
# 2002 Blackwell Science Ltd, Immunology, 105, 488–498
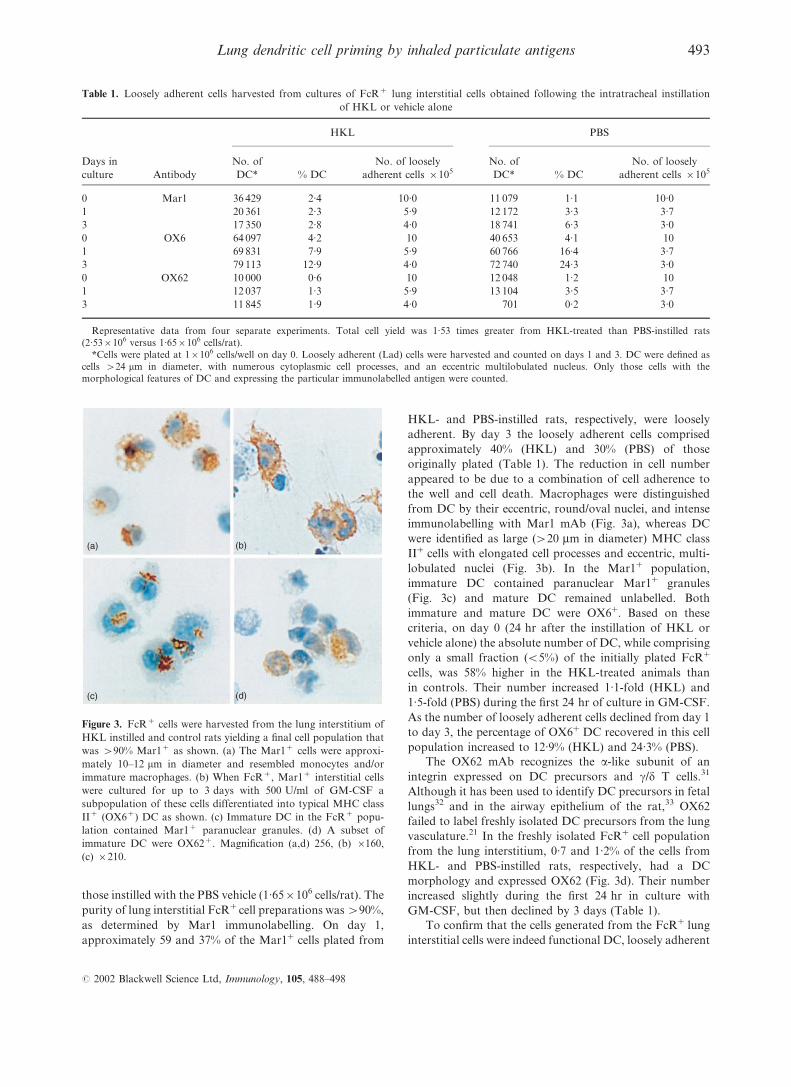
those instilled with the PBS vehicle (1.65r106 cells/rat). The
purity of lung interstitial FcR+cell preparations was >90%,
as determined by Mar1 immunolabelling. On day 1,
approximately 59 and 37% of the Mar1+ cells plated from
HKL- and PBS-instilled rats, respectively, were loosely
adherent. By day 3 the loosely adherent cells comprised
approximately 40% (HKL) and 30% (PBS) of those
originally plated (Table 1). The reduction in cell number
appeared to be due to a combination of cell adherence to
the well and cell death. Macrophages were distinguished
from DC by their eccentric, round/oval nuclei, and intense
immunolabelling with Mar1 mAb (Fig. 3a), whereas DC
were identified as large (>20 mm in diameter) MHC class
II+ cells with elongated cell processes and eccentric, multi-
lobulated nuclei (Fig. 3b). In the Mar1+ population,
immature DC contained paranuclear Mar1+ granules
(Fig. 3c) and mature DC remained unlabelled. Both
immature and mature DC were OX6+. Based on these
criteria, on day 0 (24 hr after the instillation of HKL or
vehicle alone) the absolute number of DC, while comprising
only a small fraction (<5%) of the initially plated FcR+
cells, was 58% higher in the HKL-treated animals than
in controls. Their number increased 1.1-fold (HKL) and
1.5-fold (PBS) during the first 24 hr of culture in GM-CSF.
As the number of loosely adherent cells declined from day 1
to day 3, the percentage of OX6+ DC recovered in this cell
population increased to 12.9% (HKL) and 24.3% (PBS).
The OX62 mAb recognizes the a-like subunit of an
integrin expressed on DC precursors and c/d T cells.31
Although it has been used to identify DC precursors in fetal
lungs32 and in the airway epithelium of the rat,33 OX62
failed to label freshly isolated DC precursors from the lung
vasculature.21 In the freshly isolated FcR+ cell population
from the lung interstitium, 0.7 and 1.2% of the cells from
HKL- and PBS-instilled rats, respectively, had a DC
morphology and expressed OX62 (Fig. 3d). Their number
increased slightly during the first 24 hr in culture with
GM-CSF, but then declined by 3 days (Table 1).
To confirm that the cells generated from the FcR+ lung
interstitial cells were indeed functional DC, loosely adherent
Table 1. Loosely adherent cells harvested from cultures of FcR+ lung interstitial cells obtained following the intratracheal instillation
of HKL or vehicle alone
Days in
culture Antibody
HKL PBS
No. of
DC* % DC
No. of loosely
adherent cells r105
No. of
DC* % DC
No. of loosely
adherent cells r105
0 Mar1 36 429 2.4 10.0 11 079 1.1 10.0
1 20 361 2.3 5.9 12 172 3.3 3.7
3 17 350 2.8 4.0 18 741 6.3 3.0
0 OX6 64 097 4.2 10 40 653 4.1 10
1 69 831 7.9 5.9 60 766 16.4 3.7
3 79 113 12.9 4.0 72 740 24.3 3.0
0 OX62 10 000 0.6 10 12 048 1.2 10
1 12 037 1.3 5.9 13 104 3.5 3.7
3 11 845 1.9 4.0 701 0.2 3.0
Representative data from four separate experiments. Total cell yield was 1.53 times greater from HKL-treated than PBS-instilled rats
(2.53r106 versus 1.65r106 cells/rat).
*Cells were plated at 1r106 cells/well on day 0. Loosely adherent (Lad) cells were harvested and counted on days 1 and 3. DC were defined as
cells >24 mm in diameter, with numerous cytoplasmic cell processes, and an eccentric multilobulated nucleus. Only those cells with the
morphological features of DC and expressing the particular immunolabelled antigen were counted.
(a) (b)
(c) (d)
Figure 3. FcR+ cells were harvested from the lung interstitium of
HKL instilled and control rats yielding a final cell population that
was >90% Mar1+ as shown. (a) The Mar1+ cells were approxi-
mately 10–12 mm in diameter and resembled monocytes and/or
immature macrophages. (b) When FcR+, Mar1+ interstitial cells
were cultured for up to 3 days with 500 U/ml of GM-CSF a
subpopulation of these cells differentiated into typical MHC class
II+ (OX6+) DC as shown. (c) Immature DC in the FcR+ popu-
lation contained Mar1+ paranuclear granules. (d) A subset of
immature DC were OX62+. Magnification (a,d) 256, (b) r160,
(c) r210.
493Lung dendritic cell priming by inhaled particulate antigens
# 2002 Blackwell Science Ltd, Immunology, 105, 488–498
cells were harvested following culture with 500 U/ml of
GM-CSF for 2–3 days. They were tested in an allogeneic
MLR assay using freshly isolated splenic T cells from Long
Evans rats as responder cells. A representative assay from a
total of four experiments, shows that DC in the loosely
adherent cell population induced [3H]thymidine uptake by
allogeneic T cells (Table 2).
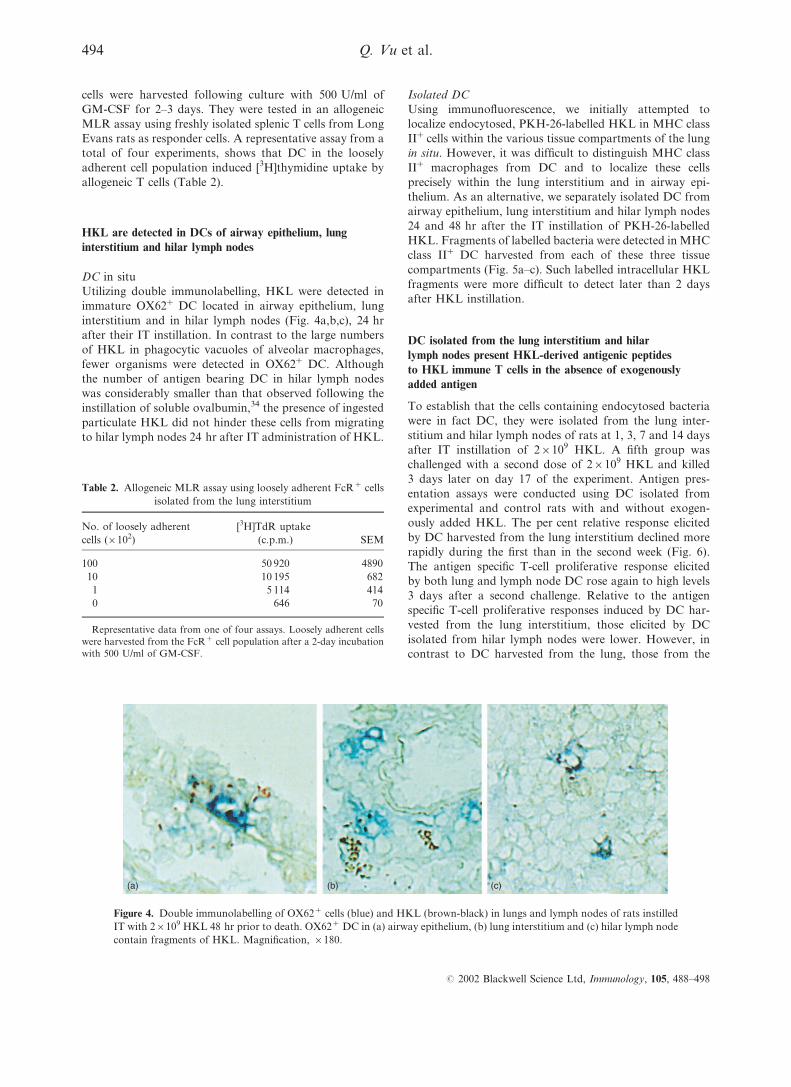
HKL are detected in DCs of airway epithelium, lung
interstitium and hilar lymph nodes
DC in situ
Utilizing double immunolabelling, HKL were detected in
immature OX62+ DC located in airway epithelium, lung
interstitium and in hilar lymph nodes (Fig. 4a,b,c), 24 hr
after their IT instillation. In contrast to the large numbers
of HKL in phagocytic vacuoles of alveolar macrophages,
fewer organisms were detected in OX62+ DC. Although
the number of antigen bearing DC in hilar lymph nodes
was considerably smaller than that observed following the
instillation of soluble ovalbumin,34 the presence of ingested
particulate HKL did not hinder these cells from migrating
to hilar lymph nodes 24 hr after IT administration of HKL.
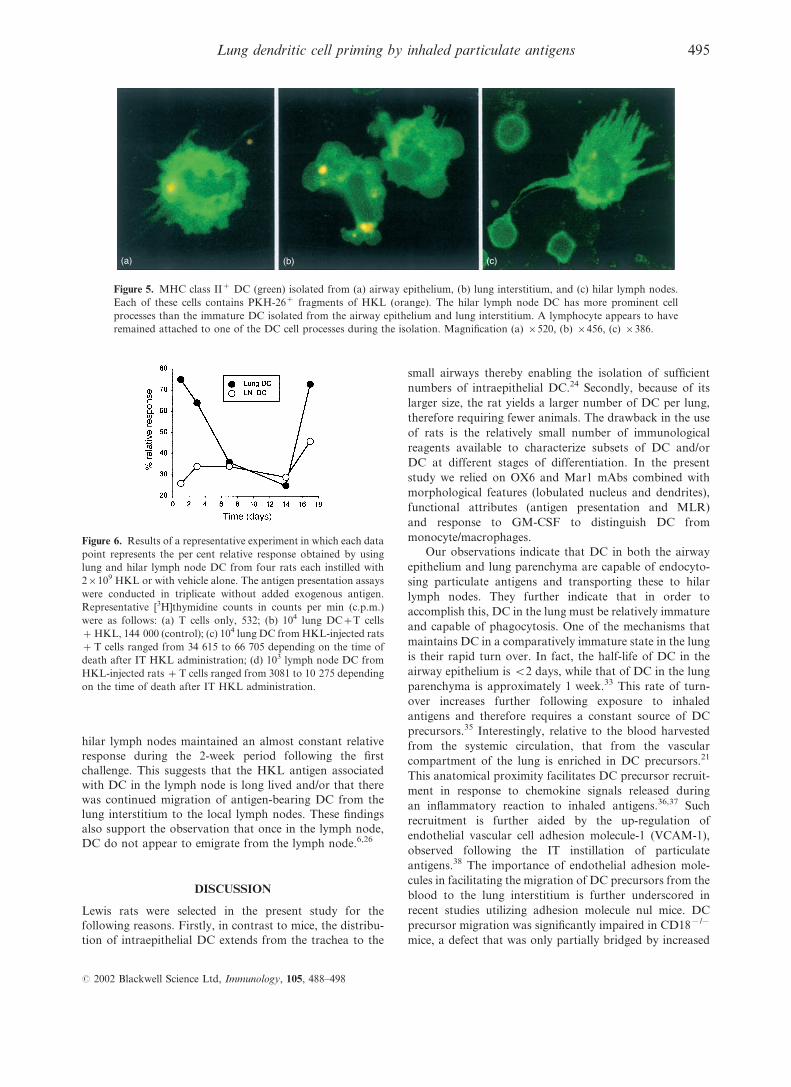
Isolated DC
Using immunofluorescence, we initially attempted to
localize endocytosed, PKH-26-labelled HKL in MHC class
II+ cells within the various tissue compartments of the lung
in situ. However, it was difficult to distinguish MHC class
II+ macrophages from DC and to localize these cells
precisely within the lung interstitium and in airway epi-
thelium. As an alternative, we separately isolated DC from
airway epithelium, lung interstitium and hilar lymph nodes
24 and 48 hr after the IT instillation of PKH-26-labelled
HKL. Fragments of labelled bacteria were detected in MHC
class II+ DC harvested from each of these three tissue
compartments (Fig. 5a–c). Such labelled intracellular HKL
fragments were more difficult to detect later than 2 days
after HKL instillation.
DC isolated from the lung interstitium and hilar
lymph nodes present HKL-derived antigenic peptides
to HKL immune T cells in the absence of exogenously
added antigen
To establish that the cells containing endocytosed bacteria
were in fact DC, they were isolated from the lung inter-
stitium and hilar lymph nodes of rats at 1, 3, 7 and 14 days
after IT instillation of 2r109 HKL. A fifth group was
challenged with a second dose of 2r109 HKL and killed
3 days later on day 17 of the experiment. Antigen pres-
entation assays were conducted using DC isolated from
experimental and control rats with and without exogen-
ously added HKL. The per cent relative response elicited
by DC harvested from the lung interstitium declined more
rapidly during the first than in the second week (Fig. 6).
The antigen specific T-cell proliferative response elicited
by both lung and lymph node DC rose again to high levels
3 days after a second challenge. Relative to the antigen
specific T-cell proliferative responses induced by DC har-
vested from the lung interstitium, those elicited by DC
isolated from hilar lymph nodes were lower. However, in
contrast to DC harvested from the lung, those from the
Table 2. Allogeneic MLR assay using loosely adherent FcR+ cells
isolated from the lung interstitium
No. of loosely adherent
cells (r102)
[3H]TdR uptake
(c.p.m.) SEM
100 50 920 4890
10 10 195 682
1 5 114 414
0 646 70
Representative data from one of four assays. Loosely adherent cells
were harvested from the FcR+ cell population after a 2-day incubation
with 500 U/ml of GM-CSF.
(a) (b) (c)
Figure 4. Double immunolabelling of OX62+ cells (blue) and HKL (brown-black) in lungs and lymph nodes of rats instilled
IT with 2r109 HKL 48 hr prior to death. OX62+ DC in (a) airway epithelium, (b) lung interstitium and (c) hilar lymph node
contain fragments of HKL. Magnification, r180.
494 Q. Vu et al.
# 2002 Blackwell Science Ltd, Immunology, 105, 488–498
hilar lymph nodes maintained an almost constant relative
response during the 2-week period following the first
challenge. This suggests that the HKL antigen associated
with DC in the lymph node is long lived and/or that there
was continued migration of antigen-bearing DC from the
lung interstitium to the local lymph nodes. These findings
also support the observation that once in the lymph node,
DC do not appear to emigrate from the lymph node.6,26
DISCUSSION
Lewis rats were selected in the present study for the
following reasons. Firstly, in contrast to mice, the distribu-
tion of intraepithelial DC extends from the trachea to the
small airways thereby enabling the isolation of sufficient
numbers of intraepithelial DC.24 Secondly, because of its
larger size, the rat yields a larger number of DC per lung,
therefore requiring fewer animals. The drawback in the use
of rats is the relatively small number of immunological
reagents available to characterize subsets of DC and/or
DC at different stages of differentiation. In the present
study we relied on OX6 and Mar1 mAbs combined with
morphological features (lobulated nucleus and dendrites),
functional attributes (antigen presentation and MLR)
and response to GM-CSF to distinguish DC from
monocyte/macrophages.
Our observations indicate that DC in both the airway
epithelium and lung parenchyma are capable of endocyto-
sing particulate antigens and transporting these to hilar
lymph nodes. They further indicate that in order to
accomplish this, DC in the lung must be relatively immature
and capable of phagocytosis. One of the mechanisms that
maintains DC in a comparatively immature state in the lung
is their rapid turn over. In fact, the half-life of DC in the
airway epithelium is <2 days, while that of DC in the lung
parenchyma is approximately 1 week.33 This rate of turn-
over increases further following exposure to inhaled
antigens and therefore requires a constant source of DC
precursors.35 Interestingly, relative to the blood harvested
from the systemic circulation, that from the vascular
compartment of the lung is enriched in DC precursors.21
This anatomical proximity facilitates DC precursor recruit-
ment in response to chemokine signals released during
an inflammatory reaction to inhaled antigens.36,37 Such
recruitment is further aided by the up-regulation of
endothelial vascular cell adhesion molecule-1 (VCAM-1),
observed following the IT instillation of particulate
antigens.38 The importance of endothelial adhesion mole-
cules in facilitating the migration of DC precursors from the
blood to the lung interstitium is further underscored in
recent studies utilizing adhesion molecule nul mice. DC
precursor migration was significantly impaired in CD18x/x
mice, a defect that was only partially bridged by increased
(a) (b) (c)
Figure 5. MHC class II+ DC (green) isolated from (a) airway epithelium, (b) lung interstitium, and (c) hilar lymph nodes.
Each of these cells contains PKH-26+ fragments of HKL (orange). The hilar lymph node DC has more prominent cell
processes than the immature DC isolated from the airway epithelium and lung interstitium. A lymphocyte appears to have
remained attached to one of the DC cell processes during the isolation. Magnification (a) r520, (b) r456, (c) r386.
Figure 6. Results of a representative experiment in which each data
point represents the per cent relative response obtained by using
lung and hilar lymph node DC from four rats each instilled with
2r109 HKL or with vehicle alone. The antigen presentation assays
were conducted in triplicate without added exogenous antigen.
Representative [3H]thymidine counts in counts per min (c.p.m.)
were as follows: (a) T cells only, 532; (b) 104 lung DC+T cells
+HKL, 144 000 (control); (c) 104 lung DC from HKL-injected rats
+T cells ranged from 34 615 to 66 705 depending on the time of
death after IT HKL administration; (d) 103 lymph node DC from
HKL-injected rats +T cells ranged from 3081 to 10 275 depending
on the time of death after IT HKL administration.
495Lung dendritic cell priming by inhaled particulate antigens
# 2002 Blackwell Science Ltd, Immunology, 105, 488–498

VCAM-1 expression.22 Recent data suggest that inter-
actions between dendritic cell-specific ligand for inter-
cellular grabbing non-integrin (DC-SIGN) and intercellular
adhesion molecule-2 may also contribute to this process.39
Following instillation of HKL into the lung there was
a significant and sustained influx of Mar1+ cells and MHC
class II+ DC precursors into the lung interstitium. In the
present study, cell counts were restricted to immunolabelled
cells in the peripheral lung interstitium, exclusive of the
connective tissue around airways and large vessels. Simi-
larly, the functional data were obtained utilizing DC
harvested from the lung parenchyma exclusive of major
airways. That DC precursors were indeed part of this newly
immigrated cell population was supported by the following
observation. Incubation of low-density, FcR+ lung
interstitial cells with GM-CSF produced a subpopulation
of functionally mature DC capable of stimulating T-cell
proliferation in an allogeneic MLR. Because their number
in the lung interstitium remained elevated for up to 2 weeks
following the instillation of HKL, it suggests that chemo-
tactic stimuli continued to be produced and/or that the
turnover rate of these precursor cells in the peripheral
lung interstitium is relatively slow. The fact that their
number in alveolar walls did not increase significantly 3 days
after a second instillation of HKL does not distinguish
between these two possibilities. However, the administra-
tion of a second dose of HKL induced the formation of
thick cuffs of cells around the pulmonary veins and airways,
indicating that substantial cell trafficking was occurring at
these anatomical sites. Further studies are needed to identify
the route by which DC precursors migrate into the lung
interstitium and to determine whether DC in the airway
epithelium enter the lung via a different route from those in
the peripheral lung interstitium.
Alveolar macrophages form the first line of defence
in the lung both by efficiently engulfing and eliminating
inhaled particulates and by producing inhibitors, including
nitric oxide, that suppress accessory cell activity in the
lung.8,40 When rats were instilled with <109 HKL/dose,
alveolar macrophages efficiently disposed of the adminis-
tered particulates and none were taken up by lung DC.41
In the present study, a dose of 2r109 HKL exceeded
the capacity of alveolar macrophages to remove all of the
administered organisms, resulting in endocytosis of HKL by
immature DC. Within 24 hr of instilling PKH-26-labelled
HKL, DC isolated separately from airway epithelium, the
lung interstitium and hilar lymph nodes contained fluores-
cent fragments of engulfed HKL. Similar fluorescent-
tagged fragments were detected in DC isolated from
hilar lymph nodes up to 48 hr after instillation of
PKH-26-labelled HKL.
Antigen presentation assays conducted, over time, in
the absence of exogenously added antigen indicated that
HKL peptides complexed to MHC class II proteins were
expressed on the surface of DC isolated from the lung
interstitium and hilar lymph nodes for prolonged periods
of time. The number of DC in the lung expressing MHC
class II/HKL peptide complexes on their surface declined
more rapidly during the first than during the second week
after the IT instillation of HKL. This was probably the
result of a combination of antigen-loaded DC emigration
from the lung and the dilutional effect of HKL-free DC
precursor migration into the lung. Similar observations
were reported in mice instilled IT with soluble, FITC-
labelled ovalbumin.34 Three days after a second IT chal-
lenge with HKL, the per cent relative response elicited by
DC isolated from the lungs and hilar lymph nodes rose to
a higher level than that observed 3 days after the first
challenge. This result is consistent with the observed
persistent elevation in the number of DC precursors in
the lungs following the first IT dose of HKL. Although the
relative response of DC harvested from local lymph nodes
was lower than that of DC isolated from the lung, it
remained elevated for up to 2 weeks after the administra-
tion of HKL. A combination of a continued migration of
antigen-loaded DC from the lung and/or a relatively slow
clearance of a particulate antigen from lymph nodes could
account for such a result. The lower relative response of
DC isolated from lymph nodes was due to a combination of
our inability to harvest all of the local lymph nodes to which
the lung DC had migrated and the relatively small number
of DC in the lymph nodes that contained engulfed HKL.
Twenty-four hours after instillation, PKH-26-labelled
HKL were predominantly found in the alveolar spaces of
the lung where they had been endocytosed by alveolar
macrophages. The question then arises by what mechanism
did the DC capture the HKL that were clearly observed
in these cells following their isolation from the airway
epithelium and lung interstitium 24 hr after instillation.
Earlier studies had shown that antigen-primed DC, but not
antigen-primed macrophages, migrate to hilar lymph nodes
following their IT instillation into the airways of rats.42
This suggests that, in contrast to alveolar macrophages,
DC express the requisite chemokine receptors that enable
them to respond to secreted chemokines and to traverse
the epithelial lining of airways and alveoli in order to
home to the lung interstitium and subsequently to hilar
lymph nodes. Among the large number of alveolar macro-
phages, a second small population of cells, comprised of
either DC precursors or immature DC, has been identified
in broncho-alveolar lavage specimens.43 These cells endo-
cytose both soluble34 and particulate antigens. They are
then thought to traverse the tight junctions of the epithelial
cell barrier and migrate to the hilar lymph nodes. Recently,
a novel alternative mechanism has been discovered whereby
DC in the intestinal epithelium extend their dendrites
through opened tight junctions into the intestinal lumen in
order to sample local, non-invasive pathogenic bacteria.44
Surprisingly, during this process DC are induced to express
the tight junction proteins occludin, claudin-1 and ZO-1,
thereby enabling them to maintain a tight junction seal
while capturing antigen in the external environment.
Whether such a mechanism also pertains to the lung
remains to be established.
ACKNOWLEDGMENTS
This study was supported by NIH grant HL36781.
496 Q. Vu et al.
# 2002 Blackwell Science Ltd, Immunology, 105, 488–498

REFERENCES
1 Bezdicek P, Crystal RG. Pulmonary Macrophages. In: Crystal
RG, West JB, Weibel ER, Barnes PJ, eds. The Lung, Scientific
Foundations 2nd edn. Philadelphia: Lippincott-Raven,
1997:859–75.
2 Schneeberger EE. The permeability of the alveolar-capillary
membrane to ultrastructural protein tracers. Ann New York
Acad Sci 1974; 221:238– 43.
3 Hulbert WC, Forster BB, Mehta JG, Man SF, Molday RS,
Walker BA et al. Study of airway epithelial permeability
with dextran. J Electron Microscopy Technique 1989;
11:137–42.
4 Harmsen AG, Muggenburg BA, Snipes MB, Bice DE. The role
of macrophages in particle translocation from lungs to lymph
node. Science 1985; 230:1277–80.
5 Harmsen AG, Mason MJ, Muggenburg BA, Gillett NA,
Jarpe MA. Migration of neutrophils from lung to tracheo-
bronchial lymph node. J Leukocyte Biol 1987; 41:95–103.
6 Banchereau J, Steinman RM. Dendritic cells and the control
of immunity. Nature 1998; 392:245–52.
7 Holt PG. Down-regulation of immune responses in the lower
respiratory tract: The role of alveolar macrophages. Clin Exp
Immunol 1986; 63:261–70.
8 Holt PG, Oliver J, Bilyk N, McMenamin C, McMenamin PG,
Kraal G et al. Downregulation of the antigen presenting
cells function(s) of pulmonary dendritic cells in vivo
by resident alveolar macrophages. J Exp Med 1993;
177:397–407.
9 Steinman RM. The dendritic cell system and its role in
immunogencity. Annu Rev Immunol 1991; 9:271–96.
10 Miyazaki H, Osawa T. Accessory functions and mutual
cooperation of murine macrophages and dendritic cells.
Europ J Immunol 1983; 13:984 –9.
11 Gong JL, McCarthy KM, Rogers RA, Schneeberger EE.
Interstitial lung macrophages interact with dendritic cells to
present antigenic peptides derived from particulate antigens
to T-cells. Immunology 1994; 81:343–51.
12 Inaba K, Inaba M, Romani N, Aya H, Deguchi M, Ikehara S
et al. Generation of large numbers of dendritic cells from
mouse bone marrow cultures supplemented with granulocyte/
macrophage colony-stimulating factor. J Exp Med 1992;
176:1693–702.
13 Inaba K, Inaba M, Deguchi M, Hagi K, Yasamizu R, Ikehara S
et al. Granulocytes, macrophages and dendritic cells arise from
a common major histocompatibility complex class II-negative
progenitor in mouse bone marrow. Proc Natl Acad Sci USA
1993; 90:3038–42.
14 Inaba K, Inaba M, Naito M, Steinman RM. Dendritic cell
progenitors phagocytose particulates, including bacillus
Calmette-Guerin organisms and sensitize mice to mycobacterial
antigens in vivo. J Exp Med 1993; 178:479–88.
15 Rescigno M, Granucci F, Citterio S, Foti M, Ricciardi-
Castagnoli P. Coordinated events during bacteria-induced
DC maturation. Immunol Today 1999; 20:200–3.
16 d’Ostiani CF, Del Sero G, Bacci A, Montagnoli C, Spreca A,
Mencacci A et al. Dendritic cells discriminate between yeasts
and hyphae of the fungus Candida albicans: Implications for
initiation of T helper cell immunity in vitro and in vivo. J Exp
Med 2000; 191:1661–73.
17 Steinman RM, Turley S, Mellman I, Inaba K. The induction
of tolerance by dendritic cells that have captured apoptotic
cells. J Exp Med 2000; 191:411–6.
18 Huang FP, Platt N, Wykes M, Major JR, Powell TJ,
Jenkins CD et al. A discrete sub-population of dendritic
cells transports apoptotic intestinal epithelial cells to T cell
areas of mesenteric lymph nodes. J Exp Medicine 2000;
191:435–43.
19 Sauter B, Albert ML, Francisco L, Larsson M, Somersan S,
Bhardwaj N. Consequences of cell death: Exposure to necrotic
tumor cells, but not primary tissue cells or apoptotic cells,
induces the maturation of immunostiuimlatory dendritic cells.
J Exp Med 2000; 191:423–33.
20 Suda T, Callahan RJ, Wilkenson RA, Van Rooijen N,
Schneeberger EE. Interferon-gamma reduces Ia+ dendritic cell
traffic to the lung. J Leukocyte Biol 1996; 60:1–10.
21 Suda T, McCarthy KM, Vu Q, McCormack J, Schneeberger EE.
Dendritic cell precursors are enriched in the vascular
compartment of the lung. Am J Resp Cell Mol Biol 1998;
19:728–37.
22 Schneeberger EE, Vu Q, LeBlanc BW, Doerschuk CM. The
accumulation of dendritic cells in the lung is impaired in
CD18x/x but not in ICAM-1x/x mutant mice. J Immunol 2000;
164:2472–8.
23 Pollard AM, Lipscomb MF. Characterization of murine lung
dendritic cells: Similarities to Langerhans cells and thymic
dendritic cells. J Exp Med 1990; 172:159–67.
24 Gong JL, McCarthy KM, Telford JR, Schneeberger EE.
Intraepithelial airway dendritic cells: a distinct subset of
pulmonary dendritic cells obtained by microdissection. J Exp
Med 1992; 175:797–807.
25 Stumbles PA, Thomas JA, Pimm CL, Lee PT, Venaille TJ,
Proksch S et al. Resting respiratory tract dendritic cells
preferentially stimulate T helper cell type 2 (Th2) responses
and require obligatory cytokine signals for induction of Th1
immunity. J Exp Med 1998; 188:2019–31.
26 Banchereau J, Briere F, Caux C, Davoust J, Lebecque S, Liu YJ
et al. Immunobiology of dendritic cells. Annu Rev Immunol
2000; 18:767–811.
27 Romani N, Bharwaj N, Pope M, Koch F, Swiggard WJ,
O’Doherty U et al. Dendritic Cells. In: Weir DM, Stewart JM,
eds. Handbook of Experimental Immunology 8th edn.
New York: Churchill Livingstone, 1997:156.1–156.4
28 Steinman RM, Cohn ZA. Identification of a novel cell type
in peripheral lymphoid organs of mice. II. functional properties
in vitro. J Exp Med 1974; 139:380–97.
29 McCarthy KM, Gong JL, Telford JR, Schneeberger EE.
Ontogeny of Ia+ accessory cells in fetal and newborn rat lung.
Am J Resp Cell Mol Biol 1992; 6:349–56.
30 Xia W, Schneeberger EE, McCarthy KM, Kradin RL.
Accessory cells of the lung: II. Ia+pulmonary dendritic cells
display surface antigen heterogeneity. Am J Resp Cell Molec
Biol 1991; 5:276–83.
31 Brenan M, Puklavec M. The MRC OX-62 antigen: a
useful marker in the purification of rat veiled cells with the
biochemical properties of an integrin. J Exp Med 1992;
175:1457–65.
32 Nelson DJ, McMenamin C, McWilliam AS, Brenan M,
Holt PG. Development of the airway intraepithelial dendritic
cell network in the rat from class II major histocompatibility
(Ia)-negative precursors: Differential regulation of Ia expression
at different levels of the respiratory tract. J Exp Med 1994;
179:203–12.
33 Holt PG, Haining S, Nelson DJ, Sedgwick JD. Origin and
steady-state turnover of class II MHC-bearing dendritic cells
in the epithelium of the conducting airways. J Immunol 1994;
153:256–61.
34 Vermaelen KY, Carro-Muino I, Lambrecht BN, Pauwels RA.
Specific migratory dendritic cells rapidly transport antigen
497Lung dendritic cell priming by inhaled particulate antigens
# 2002 Blackwell Science Ltd, Immunology, 105, 488–498

from the airways to the thoracic lymph nodes. J Exp Med 2001;
193:51–60.
35 Holt PG, Stumbles PA. Characterization of dendritic cell
populations in the respiratory tract. J Aerosol Med 2000;
13:361–7.
36 McWilliam AS, Napoli S, Marsh AM, Pemper FL, Nelson DJ,
Pimm CL et al. Dendritic cells are recruited into the
airway epithelium during the inflammatory response to a broad
spectrum of stimuli. J Exp Med 1996; 184:2429–32.
37 Stumbles PA, Strickland DH, Pimm CL, Proksch SF,
Marsh AM, McWilliam AS et al. Regulation of dendritic cell
recruitment into resting and inflamed airway epithelium: Use of
alternative chemokine receptors as a function of inducing
stimulus. J Immunol 2001; 167:228–34.
38 Wolber FM, Curtis JL, Milik AM, Fields T, Seitzman GD,
Kim KM et al. Lymphocyte recruitment and the kinetics
of adhesion receptor expression during the pulmonary
immune reponse to particulate antigen. Am J Pathol 1997;
151:1715–27.
39 Geijtenbeek TBH, Krooshoop JEB, Bleijs DA, van Vliet SJ, van
Duijnhoven GCF, Grabovsky V et al. DC-SIGN-ICAM-2
interaction mediates dendritic cell trafficking. Nature Immunol
2001; 1:353–7.
40 Thepen T, Van Rooijen N, Kraal G. Alveolar macro-
phage elimination in vivo is associated with an increase
in pulmonary immune response in mice. J Exp Med 1989;
170:499–509.
41 MacClean JA, Xia W, Pinto C, Zhao L, Liu HW, Kradin RL.
Sequestration of inhaled particulate antigens by lung
phagocytes. Am J Pathol 1996; 148:657–66.
42 Havenith CEG, Van Miert PPMC, Breedijk AJ, Beelen RHJ,
Hoefsmit ECM. Migration of dendritic cells into the draining
lymph nodes of the lung after intratracheal instillation.
Am J Resp Cell Mol Biol 1993; 9:484 –8.
43 Lambrecht BN, Carro-Muino I, Vermaelen K, Pauwels RA.
Allergen-induced changes in bone-marrow progenitor and
airway dendritic cells in sensitized rats. Am J Resp Cell Mol
Biol 1999; 20:1165–74.
44 Rescigno M, Urbano MBB, Francolini M, Rotta G, Bonasio R
et al. Dendritic cells express tight junction proteins and
penetrate gut epithelial monolayers to sample bacteria.
Nature Immunol 1901; 2:361–7.
498 Q. Vu et al.
# 2002 Blackwell Science Ltd, Immunology, 105, 488–498





























