Embed Size (px)
Citation preview

MAN KANN UT HI HA UN NUMARULUI MIHI US009957531B1
( 12 ) United States Patent Koepke et al .
( 10 ) Patent No . : ( 45 ) Date of Patent :
US 9 , 957 , 531 B1 May 1 , 2018
9 , 297 , 026 B2 2011 / 0151530 AL 2011 / 0165644 A1 2011 / 0201089 AL 2012 / 0252083 Al
3 / 2016 Koepke et al . 6 / 2011 Soucaille et al . 7 / 2011 Marliere 8 / 2011 Burgard et al .
10 / 2012 Koepke et al .
FOREIGN PATENT DOCUMENTS EP
( 54 ) GENETICALLY ENGINEERED BACTERIUM FOR THE PRODUCTION OF 3 - HYDROXYBUTYRATE
( 71 ) Applicant : LanzaTech New Zealand Limited , Skokie , IL ( US )
( 72 ) Inventors : Michael Koepke , Skokie , IL ( US ) ; Rasmus Overgaard Jensen , Skokie , IL ( US ) ; James Bruce Yarnton Haycock Behrendorff , Copenhagen ( DK ) ; Ryan Edward Hill , Stockholm ( SE ) ; Darmawi Juminaga , Skokie , IL ( US ) ; Alexander Paul Mueller , Skokie , IL ( US )
( 73 ) Assignee : LANZATECH NEW ZEALAND LIMITED , Auckland ( NZ )
( * ) Notice : Subject to any disclaimer , the term of this patent is extended or adjusted under 35 U . S . C . 154 ( b ) by 0 days . days .
( 21 ) Appl . No . : 15 / 658 , 668 ( 22 ) Filed : Jul . 25 , 2017
WO WO WO WO WO WO WO wo wo wo WO WO WO WO WO wo WO Wo WO WO WO
2295593 A1 3 / 2011 2007117157 Al 10 / 2007 2008028055 A2 3 / 2008 2008115080 A1 9 / 2008 2009064200 A1 5 / 2009 2009151342 Al 12 / 2009 2010001078 A2 1 / 2010 2011076691 AL 6 / 2011 2011112103 Al 9 / 2011 2012024522 A2 2 / 2012 2012026833 A1 3 / 2012 2012053905 Al 4 / 2012 2012115527 A2 8 / 2012 2013036147 A2 3 / 2013 2013180581 A1 12 / 2013 2013180584 Al 12 / 2013 2013185123 Al 12 / 2013 2013191567 Al 12 / 2013 2014036152 A13 / 2014 2015085015 A1 6 / 2015 2015101493 AL 7 / 2015 2016034691 Al 10 / 2016
Related U . S . Application Data ( 63 ) Continuation of application No . 15 / 293 , 191 , filed on
Oct . 13 , 2016 , now Pat . No . 9 , 738 , 875 . OTHER PUBLICATIONS ( 60 ) Provisional application No . 62 / 240 , 850 , filed on Oct .
13 , 2015 .
( 51 )
Abrini , Arch Microbiol , 161 : 345 - 351 , 1994 . Barker , PNAS USA , 31 : 373 - 381 , 1945 . Cheong , Nature Biotechnol , 34 : 556 - 561 , 2016 . Cho , Biotechnol Adv , 33 : 1455 - 1466 , 2015 . Clomburg , Appl Microbiol Biotechnol , 86 : 419 - 434 , 2010 . Drake , Acetogenic Prokaryotes , In : The Prokaryotes , 3rd edition , pp . 354 - 420 , New York , NY , Springer , 2006 . Heap , J Microbiol Meth , 78 : 79 - 85 , 2009 . Huang , Nature , 537 : 320 - 327 , 2016 . Hungate , Meth Microbiol , 3B : 117 - 132 , 1969 . Jones , Microbiol Rev , 50 : 484 - 524 , 1986 . Kataoka , J Biosci Bioeng , 115 : 475 - 480 , 2013 . Khoury , Trends Biotechnol , 32 : 99 - 109 , 2014 . Köpke , Appl Environ Microbiol , 80 : 3394 - 3403 , 2014 . Köpke , Curr Opin Biotechnol , 22 : 320 - 325 , 2011 . Köpke , PNAS USA , 107 : 13087 - 13092 , 2010 . Liu , Appl Microbiol Biotechnol , 53 : 545 - 552 , 2000 . Bevers , J Bacterial , 187 : 7056 - 7061 , 2005 . Makshina , Chem Soc Rev , 43 : 7917 - 7953 , 2014 . Marcellin , Green Chem , 18 : 3020 - 3028 , 2016 . Matsumoto , Appl Microbiol Biotechnol , 97 : 205 - 210 , 2013 .
( Continued )
Int . Cl . C12N 1 / 20 ( 2006 . 01 ) C12P 7 / 42 ( 2006 . 01 ) C12N 9 / 10 ( 2006 . 01 ) C12N 9 / 02 ( 2006 . 01 ) C12N 9 / 04 ( 2006 . 01 ) C12N 9 / 12 ( 2006 . 01 ) C12N 9 / 16 ( 2006 . 01 ) C12P 7 / 24 ( 2006 . 01 ) C12P 7 / 18 ( 2006 . 01 ) U . S . CI . CPC . . . . . . . . . . . . . . C12P 7 / 42 ( 2013 . 01 ) ; C12N 9 / 0006
( 2013 . 01 ) ; C12N 9 / 0008 ( 2013 . 01 ) ; C12N 9 / 1029 ( 2013 . 01 ) ; C12N 9 / 1217 ( 2013 . 01 ) ;
C12N 9 / 16 ( 2013 . 01 ) ; C12P 7 / 18 ( 2013 . 01 ) ; C12P 7 / 24 ( 2013 . 01 ) ; C12Y 101 / 01001
( 2013 . 01 ) ; C12Y 101 / 01002 ( 2013 . 01 ) ; C12Y 102 / 07005 ( 2013 . 01 ) ; C12Y 203 / 01008
( 2013 . 01 ) ; C12Y 203 / 01009 ( 2013 . 01 ) ; C12Y 207 / 02001 ( 2013 . 01 ) ; C12Y 301 / 0202
( 2013 . 01 ) Field of Classification Search CPC . . . . . . . . . . . . . . . . . . . . . . . . . . C12N 9 / 1217 ; C12N 9 / 0008 USPC . . . . . . 435 / 252 . 3 See application file for complete search history .
( 52 )
Primary Examiner — Tekchand Saidha ( 74 ) Attorney , Agent , or Firm — Andrea Schoen
( 58 ) ( 57 ) ABSTRACT The invention relates to a genetically engineered bacterium having an enzyme that converts acetyl - CoA to acetoacetyl COA , an enzyme that converts acetoacetyl - CoA to 3 - hy droxybutyryl - CoA , and an enzyme that converts 3 - hydroxy butyryl - CoA to 3 - hydroxybutyrate . The bacterium may also have enzymes to produce other downstream products , such as 3 - hydroxybutyryaldehyde , and 1 , 3 - butanediol . Typically , the bacterium is capable of producing these products from a gaseous substrate , such as syngas or an industrial waste gas .
( 56 ) References Cited U . S . PATENT DOCUMENTS
5 , 593 , 886 A 1 / 1997 Gaddy 6 , 368 , 819 B14 / 2002 Gaddy et al . 7 , 262 , 037 B2 8 / 2007 Chen et al . 9 , 096 , 860 B2 * 8 / 2015 Park . . . C12N 15 / 77 20 Claims , 43 Drawing Sheets

US 9 , 957 , 531 B1 Page 2
( 56 ) References Cited
OTHER PUBLICATIONS May , Metabol Eng , 15 : 218 - 225 , 2013 . Mock , J Bacteriol , 197 : 2965 - 2980 , 2015 . Nagarajan , Microb Cell Factories , 12 : 118 , 2013 . Packer , Nature Rev Genetics , 16 : 379 - 394 , 2015 . Peralta - Yahya , Biotechnol J , 5 : 147 - 162 , 2010 . Perez , Biotechnol Bioeng , 110 : 1066 - 1077 , 2012 . Privett PNAS USA , 109 : 3790 - 3795 , 2012 . Quan , PloS One , 4 : e6441 , 2009 . Ragsdale , Biochim Biophys Acta , 1784 : 1873 - 1898 , 2008 . Schiel - Bengelsdorf , FEBS Lett , 586 : 2191 - 2198 , 2012 . Schuchmann , Nat Rev Microbiol , 12 : 809 - 821 , 2014 . Seedorf , PNAS USA , 105 : 2128 - 2133 , 2008 . Takanashi , J Biosci Bioeng , 101 : 501 - 507 , 2006 . Tanner , Int J Syst Bacteriol , 43 : 232 - 236 , 1993 . Thauer , Bacteriol Rev , 41 : 100 - 180 , 1977 . Thompson , Appl Environ Microbiol , 56 : 607 - 613 , 1990 . Tseng , Appl Environ Microbiol , 75 : 3137 - 3145 , 2009 . van Leeuwen , Appl Microbiol Biotechnol , 93 : 1377 - 1387 , 2012 . Williams , J Gen Microbiol , 1136 : 819 - 826 , 1990 , Wolfe , Adv Microb Physiol , 6 : 107 - 146 , 1971 . Yaneva , J Biol Chem , 287 : 15502 - 15511 , 2012 . Yu , Biotechnol Bioeng , 111 : 2580 - 2586 , 2014 .
* cited by examiner

U . S . Patent May 1 , 2018 Sheet 1 of 43 US 9 , 957 , 531 B1
10 . . . . … . Kai RECRA
… … … sskate Againter Relatersivinate oney .
Art are
. . . .
1 1 - | A insite oxysianslate SAAS TEets rea
18 … … … … …
SAyer anyieR resersionate stagrate autest tae Soyoayorkissy gitake
| * * *
1
Easties RAIN Chics ARAKAMA airy trays at tyr see
* *
ii
- A . Sparted
FIG . 1

U . S . Patent May 1 , 2018 Sheet 2 of 43 US 9 , 957 , 531 B1
Damerican , 1 . SH - COA ADP fto ( 2 . 3 . 1 . 191 Buk ( 2 . 7 . 2 . 7 ) m ma Butanoyl - ping Sam as ones Butanoyl - CoA ; i Butyrate + ATP
FIG . 2

U . S . Patent May 1 , 2018 Sheet 3 of 43 US 9 , 957 , 531 B1
COA transferase ( ABE pathway ) 4
Txolage { EGZ . 3 . 3 . 93
C Isoproparol ranstorns ( 10 : 20 : 39 - *
Acetoacetate stei » lxitylxse
{ EC : 4 . 31 ) * * * * NOVOM 2x Hover his mayonnaise
Acetyl - CoA Bois Acetoncetyl - CoA uz Acetate . Acetyl - C , Acetoacetate Acetone { sohutylene
HTY
• COA kausterase rayures huts concentratin : ls of acetate
. . No direct 411 generation , ATP need to se generated via acetate biosynthesis , requiring 29ditional sect - CA
Thioesterase Sais 4 : 0 * * * *
{ soprspass } Thiolas Sextiyacetzte 890 doxylase
{ EX24 . 1 . 1 . 3 ) wa nch
Thoastarase ( EC3 . 2 . 2 . 1 w203 . 3 . 2 . 1 )
??????????????????????????????
iyu 9 { XC : 2 . 3 . 1 . 9 ) som man 1400 W
Acetyl - CoA COM Acetoacetyl - Cos
6 2x
Co . Acetoacetate : 09 . Acelone Isohutylene
Acety - Cod Acetate ATH
NOAP generation 417 need to be gesiemco via acetate Usosynthesisy 12 : { 1 } { ?? ???gk & # 1 Acctate deos sot get recycled us thus vecumintes
Pth - Buk W
Isopropano Hutyrate Kidade EC27 . 2 . 7 )
& ??˨ate secarboxylase | { { { $ 13 . 4 }
? 6 .
Thorse . 9 { EC . 2 . 3 . 1 . 9 ) sa maagannathe more
Acetyl - CoA Los Acetoacetyl - 69 2x
Proxpoxtto Suryytransferase
AC : 23 , 139 ) e On
O ! Acetoacetyt
? Pris
AYP sor Acetoacetate de Acerone Isalutylene
· Disect ATY geutration via substrate icvti kxsporio
Notenireuein for acetate formation or 31 ) xcctum acetyl
. . . . . . . . . .
FIG . 3

U . S . Patent May 1 , 2018 Sheet 4 of 43 US 9 , 957 , 531 B1
DO 0 . 2 . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , . , .
50 . 18 . . . . . . . Acetone production ( g / l 0 . 14
O 0 . 1 20 . 12 20 . 08
0 . 06 20 . 04
0 . 02
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Thi + Ptb - Buk + Adc [ E . coli BL21 ( DE3 ) + PACYC - ptb - . . .
Thi + Adc alone [ E . coli BL21 ( DE3 } + pCOLA - th1A - . .
Ptb - Buk alone [ E . coli BL21 ( DE3 ) + PACYC - ptb - buk ] No plasmid control [ E . coli BL21 ( DE3 ) ]
Media control
FIG . 4 .
0 . 2 0 . 18 0 . 16 0 . 14 0 . 12
0 . 1 0 . 08 0 . 06
Acetone ( g / L ]
0 . 04 0 . 02 . . . . . . .
10 uM IPTG 50 UM IPTG 100 UM IPTG OUM IPTG ( uninduced )
FIG . 5

US . Patent May 1 , 2018 Sheet 5 of 43 US 9 , 957 , 531 B1
?????y 93
????? : QQ 3 ???yX4XXX 949 ; .
?? ( : ?? ? 3 - hydroxybutyrate
? : ??
FIG . 6
* * * * *
| acetate O + : CH
??? % 93ye
03
C + 3 - hydroxybutyrate 3 - hydroxybutyraldehyde { , 3 823
F1G . 7

U . S . Patent May 1 , 2018 Sheet 6 of 43 US 9 , 957 , 531 B1
coa le
ÅVOR COX 40
28
{ R } Sydney
Mamos satisk * * * * * * * * * *
* * * * . 110 it
Rihydroxybutyrate { $ $ - 3 - ydroxybutyrste
keo OM O .
task ar
egokolo
FIG . 8

U . S . Patent May 1 , 2018 Sheet 7 of 43 US 9 , 957 , 531 B1
2 Aco COA
acetoacetyl - CoA
olb - out Mis acetoacetate
synthase acetone woman on the hydroxyisovalery - COA KA Serve r som * * *
3 - hydroxyisovalerate the * *
hydroxyisovalerate decarboxylase
sobutene
FIG . 9

FIG . 10
US 9 , 957 , 531 B1
Isoleucme biosynthesis
SDwKxus sn0600121au
with Bieouelvadoxo - - Xujaus
* * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * *
* * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * *
sobutane s
@
artistiningte 400
Sheet 8 of 43
2 - aceto - 2 - hydroxybutanoate
Oro
OSVOBO DejoleroSiAXOIDÁU
mini . Semik
Beoveingoxo - 2
2004 WXXN
aterialen 20 . M
distintinnan - - Shop :
pit - buk
VOYABCD 002 * fied with
May 1 , 2018
mining mo
Yoo BijenostkxOIDALE
Bejewens
2 - methylcrotony - COA Development W
40
in 2018
Yaco
????
enne testen
Superior
U . S . Patent
x 40 : 03
Voorkouengjával ?
wanne 08 091 * 200 mm
!
Alates 3 Acetyl - CoA
- 20
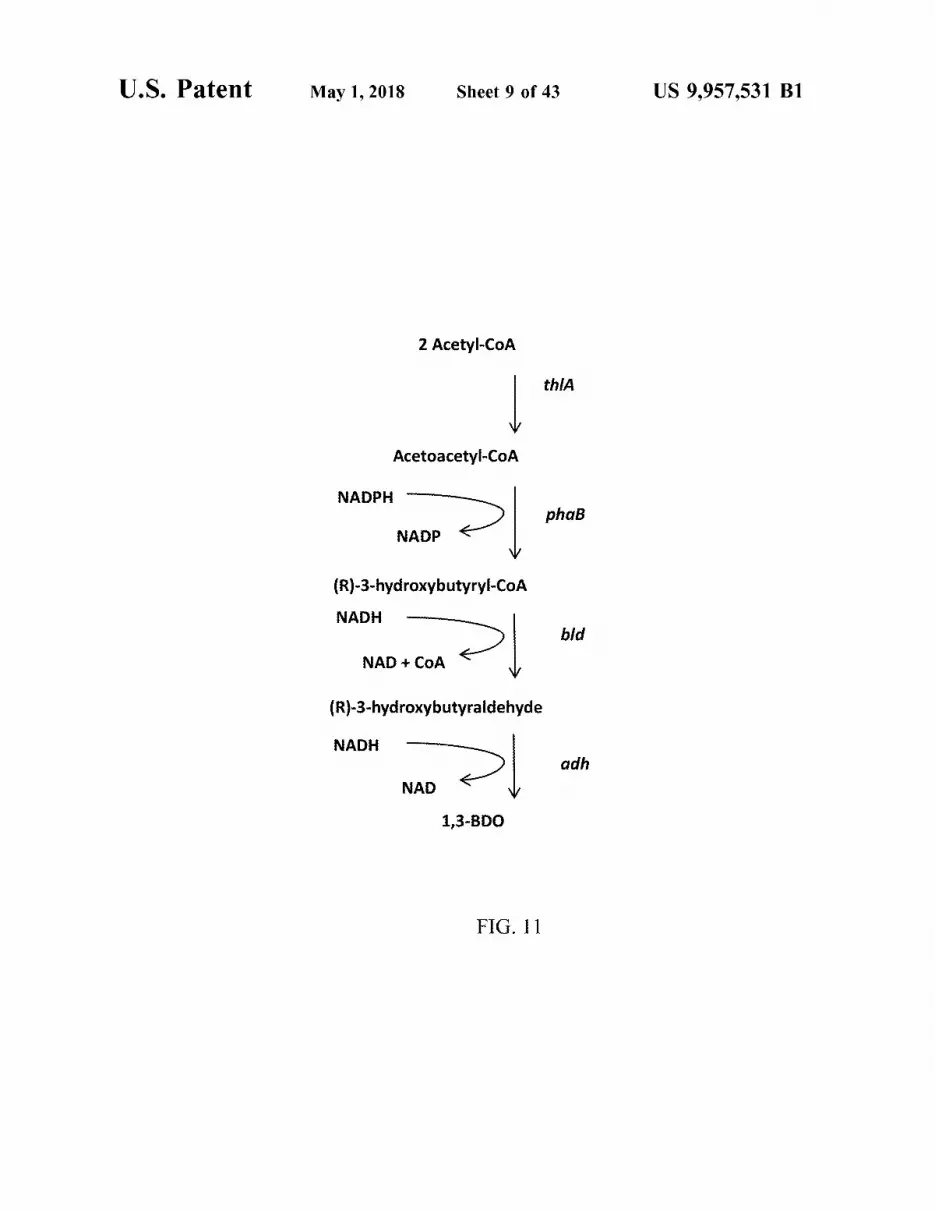
U . S . Patent May 1 , 2018 Sheet 9 of 43 US 9 , 957 , 531 B1
2 Acetyl - CoA
thia
Acetoacetyl - CoA
NADPH NADPH - and phaB
| NADP | phat phaß NADP
( R ) - 3 - hydroxybutyryl - COA NADH
bld NAD + COA
( R ) - 3 - hydroxybutyraldehyde
NADH el eeh adh NAD NAD E
1 , 3 - BDO
FIG . 11

200
US 9 , 957 , 531 B1
* * *
* * * * * * * * *
* * * * * * * * * * *
* *
* * * * * * * * *
* * * * * * * * * * * * * * * *
o 150
Sheet 10 of 43
or 100
Time ( ha )
FIG . 12
O
May 1 , 2018
0
Here
U . S . Patent
are
ca
0
( 1 / 8 ) jouedosdosi

U . S . Patent May 1 , 2018 Sheet 11 of 43 US 9 , 957 , 531 B1
thiA - adc / ptb - buk / phaB
0 * 10 50 100 2 . 50
2 . 00
1 . 50
1 . 00 - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
0 . 50 . .
0 . 00 3HB 3HB acetate ethanol acetone
FIG . 13A
thlA - adc / ptb - buk / phaB - bdh1 30 10 850 X 100
2 . 50
2 . 00
1 . 50
1 . 00
0 . 50
0 . 00 8H€ acetate acetate ethanol ethanol acetone acetone
FIG . 13B

U . S . Patent May 1 , 2018 Sheet 12 of 43 US 9 , 957 , 531 B1
thlA - adc / phaB - bdhi
0 10 050 0100
2 . 50
2 . 00
1 . 50
1 . 00
0 . 50
0 . 00 3HB acetate ethanol acetone
FIG . 13C
th : A - adc
0010 850 X 100
2 . 50
2 . 00
1 . 50 * * * * * www
1 . 00
0 . 50 0 . 00 . . . . . *
??? 3HB acetate acetate ethanol acetone * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * *
FIG . 13D

U . S . Patent May 1 , 2018 Sheet 13 of 43 US 9 , 957 , 531 B1
phab - bdh1
0 10 050 X 100
2 . 50
2 . 00
1 . 50
1 . 00 ???????????????????????
0 . 50 0 . 00 . . . . .
3HB acetate ethanol acetone
FIG . 13E
phaB 0 * 10 50 100
2 . 50
2 . 00 1 . 50 1 . 00
0 . 50 0 . 00 . . . . . . . . * * * . - -
3HB acetate ethanol acetone
FIG . 13F
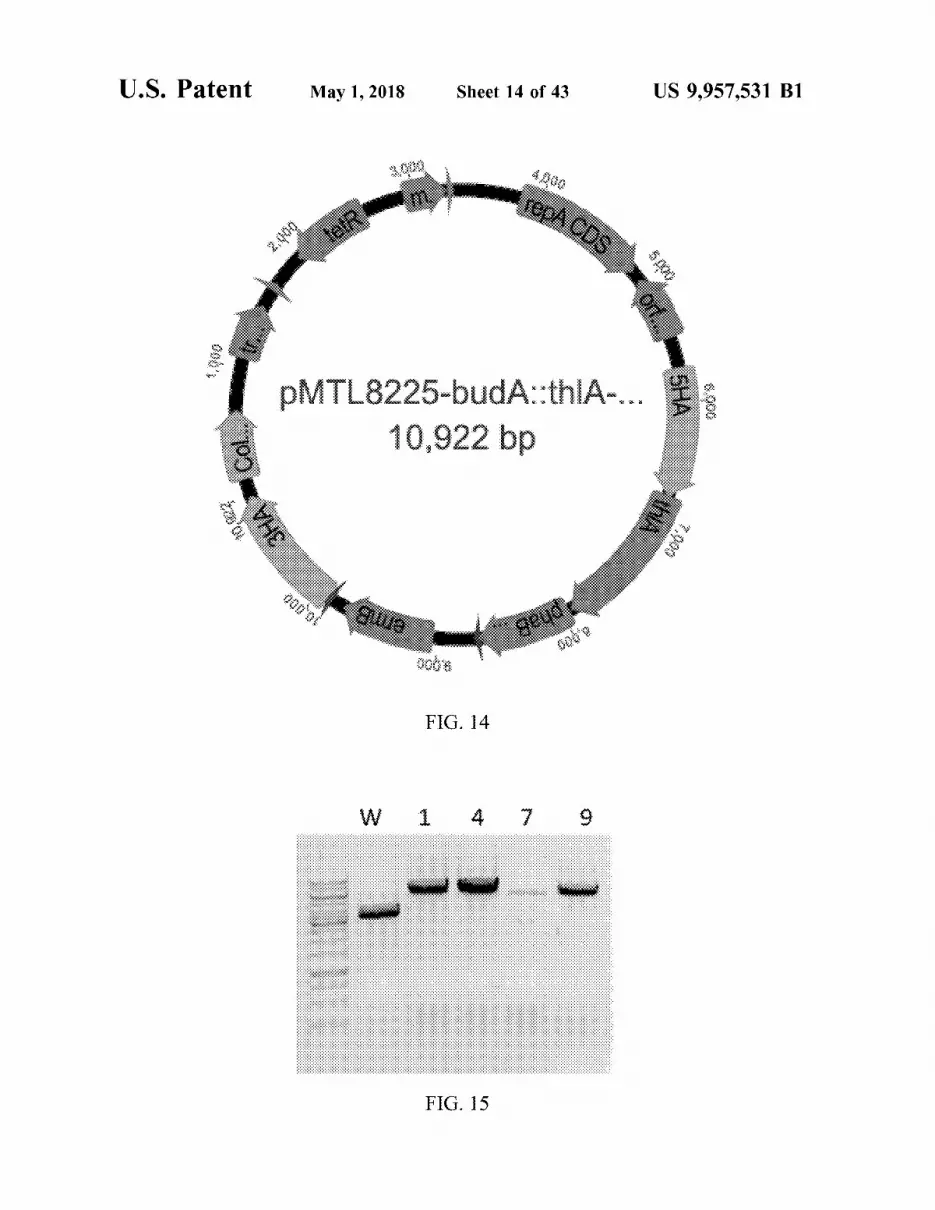
U . S . Patent May 1 , 2018 Sheet 14 of 43 US 9 , 957 , 531 B1
4 Dec
EPA LDA . Petr 5 . 920
1 . 000
PMTL8225 - budA : : thlA - . . . 10 , 922 bp
: *
2004 00010
000 0006
FIG . 14
W 1 4 9
- - -
FIG . 15

. . . . . . . . .
. . . . . . . . . . . . . .
. . . . . . . . . . . .
. .
. . . . . . .
3 - HB , 1 , 3 - BDO ( ppm )
US 9 , 957 , 531 B1
1 , 650 1 , 550 1 , 450 1 , 350
250
1 , 150 1 , 050 950 850
750
650 SSO
450
350
250
en
1 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . .
. . . . . . . . .
1 , 3 - BDO
* * * *
* * * * * * *
3HB
Sheet 15 of 43
3 encima
www . Biomass
www *
Time ( d ]
FIG . 16
May 1 , 2018
www . Ethanol wwwww Acetate
Š
Š
O
U . S . Patent
Ethanol , Biomass , Acetic acid Conc . ( g / L )
????????????????????????????????????????????????????????????

U . S . Patent May 1 , 2018 Sheet 16 of 43 US 9 , 957 , 531 B1
1 , 3 - BDC 0 . 09 0 . 08 .
. . . .
1 , 3 - BDO ( S / oooo . . . . . .
0 . 03 0 . 02 0 . 01
. . . . . . . . . . . . . . . . . . . . . . .
. . . . . . * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * *
0 02 01 09 08 001 OCT 001 091 081 Time in
LZ1561 PMTL8315 - Pfdx - th / A - phaB - bld
FIG . 17A
Biomass in
corrissenteresatare 600 151 B
noi ca in Nino insaarissasasasastana ccccc rrrrrrrrrr irror
- 9 , 50 100 OST 200 200 Time ( n )
171561 PMTL8315 - Pfdx - th / A - phaB - bld ????????????????????????
FIG . 17B

U . S . Patent May 1 , 2018 Sheet 17 of 43 US 9 , 957 , 531 B1
“ .
. .
. .
.
MMMMM . . . . . . . . . . . . . . .
" " , " i " : " : " : " AMAHAHA : :
" , " " : " ,
: : . .
…
??? ?????? * ? ? ?? ?? ?? …
… … . . MAMAHAMM …
… … … … … .
:
:
: : :
. … … … … … … . : : :
: ???HAN #
FIG . 18A
*
* . . . … . . . . . . . … . . . . . . . … . . . … . … . . . . . … . … . . … . … . . … . … . . . .
?? " " " " " " " . . . . . . , . . . . . . .
" * *
* . . . . .
. . . . . . . .
* . . .
: . .
* * * * * * * * * * * * * : : : : : : * ; * . * . * .
????? ?? rrrrrrrrrrryFFFFFF ,
FIG . 18B

3 - HB Conc . ( g / L )
A uno
frond NW . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
U . S . Patent
O
" "
" " " "
. . . . .
Continuous D = 0 . 5
2
I IT
11 11 11
D = 1 . 0
* * * * * * * * * *
? ! ? ! ?
4
KK !
1
May 1 , 2018
6
& HEY
FIG . 19
8 Time [ d ]
10
Sheet 18 of 43
1 , 3 - BDO GC - FID
12 14 - 16
US 9 , 957 , 531 B1
0 . 1
1 , 3 - BDO Conc . ( g / L )

U . S . Patent May 1 , 2018 Sheet 19 of 43 US 9 , 957 , 531 B1
1000
PMTL82256 - ptb - buk 900
800 . * * * . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4
700
600 PMTL82256 - ptb - buk 500 { { { { { { rt
400
300
100 PMTL82256 - bbb - buk WT
3 - faydroxybutyry - COA 2 - hydroxyisobutyryl - CoA Acetoacetyl - CoA
FIG . 20A
sowohl Ulmg word .
o
WT PHIB 01 PMTL82256 - ptb - buk
FIG . 20B

U . S . Patent May 1 , 2018 Sheet 20 of 43 US 9 , 957 , 531 B1
0 . 0008 0 . 0007
0 . 0008 ?r } ma 0 . 0005 € 0 . 0004
0 . 0003 } { or
0 . 0002
0 . 0001
acac - CoA LZ
acac - CoA CT2610 IZ
348 - COA
CT2640 2 - HIB - COA
2 2 HB - CoA
CT2640 WWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWWW
FIG . 21A
1 1 , 1 , 1 , 4 , 4 , 4 , 4 , , , , , , , , , , , , , , , , , , , , , , , , , , , , , * . . . . . . . * . . . * . . . * . * . . . . . . * . * . . . . . . . . * . * . . . . . * . * . .
, , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , ,
0 . 1
.
1963 CI3S24
* * * o004 * 3 Hydroxytyvo
FIG . 21B

U . S . Patent May 1 , 2018 Sheet 21 of 43 US 9 , 957 , 531 B1
Isopropanol pr OD600 ( 8 / 1 / 00 )
0 . 8
0 . 6
0 . 5
0 . 4
CAETHG _ 1780 °C . autoethanogenum
FIG . 22

U . S . Patent May 1 , 2018 Sheet 22 of 43 US 9 , 957 , 531 B1
. . . . . . . . . . . . . . . . . . .
-
00900 - - -
0 . 01 0 20 40 60 100 120 140 80
Time ( h )
A CAETHG 1280 C autoethanogenum
FIG . 23A
i
' - ' - ' - ' - '
la Isopropanol ( g / l )
enter de -
- - - - - -
- - -
-
c 0 20 40 60 80 Time in
100 120 140 160
A CAETHG 1780 oC . autoethanogenum
FIG . 23B

U . S . Patent May 1 , 2018 Sheet 23 of 43 US 9 , 957 , 531 B1
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
No n Acetate ( all
no
kansand no no ja
0 20 0 60 100 120 140 160 80 Time ( b )
Quioethanogenu . . . . . . . . . . . . . . . .
FIG . 230
. . . . . . . . . .
i
. . . . . . . . . .
0
i ( 1 / 2 ) pove433 . . . . . . . . . . . .
N
i 1 internet . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
0 . 5 000 . . . . . . - . - . - . - . - . - . . - . - . - . - . - . - - . . . . .
0 20 40 60 100 120 140 160 80 Time ( h )
FIG . 23D

U . S . Patent May 1 , 2018 Sheet 24 of 43 US 9 , 957 , 531 B1
Caus DO
a CDS * * *
www PMTL8225 - pta - ack ptb - buk 11 , 184 bp
U
a ' s
FIG . 24

U . S . Patent May 1 , 2018 Sheet 25 of 43 US 9 , 957 , 531 B1
W 1 2 3 4 5 ? 6
?????? FIG . 25

US 9 , 957 , 531 B1
?? : :
Sheet 26 of 43
:
| FIG . 26
?
May 1 , 2018
??? ? .
U . S . Patent
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .

FIG . 27 wwwwwwwwwwwwwww
03 - VAIH - 7 Yo - 24€ * 909 - 1XBeoja3y8 X338 - ki xy Tess B121
wazold Busfujw / lown
- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
US 9 , 957 , 531 B1 Sheet 27 of 43 May 1 , 2018 U . S . Patent

FIG . 28B
alaangamojpát facic acid acatio add ?????? buwwwwwwwww 1941
Product concentration ( ol 1 . 1 . 1 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
* * * . * . * . * . * . * . * . * * * . * * * . * * * . * * . * * . * * . * . * . * . * . * . * . * * * . * * * . * * * . * * * . * * * . * . * . * . * . * . * . * . * . * . * . * . * . . * . * . * . * . * . . * . * . * . . . . * . * . * . * . * . . * . * . * . * . * . * . * . . * . * * . * * . * . . . . * . * . * . . * . . * . * * . * . * . * * . * * . * . . . . . * . * . * . . * . . * . * . * . * . . . . . . . * . . * . . * . . * . . . .
50 X 100 M PTG # 0 * 10
BL21 * thIA / ptb - buk / phaB
FIG . 28A
3 - ydroxybutyrata lactic acid 008 900 jolla
4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4 . 4
Product concentration ( 9 )
* * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * * *
Old W * 001 * 09 * 0L * 0 * BL21 * thIA / ptb - buk / hbd
US 9 , 957 , 531 B1 Sheet 28 of 43 May 1 , 2018 U . S . Patent

U . S . Patent May 1 , 2018 Sheet 29 of 43 US 9 , 957 , 531 B1
LIFFFFFF *
* .
*
* *
?? ???? ???? ??
* *
? -
* - -
screat * - - - - - -
F
? F FF - 1T
?? . exe ,
????? MAN
TW / AAAAAAAyoshow ?????? ???????????????????? ??? * * * * ????????????????????????? * * * * * * , “ ? ” , ??????????????????????????? ? SEE “ ? ? ?? ? ” “ ? ” ? . tw
FIG . 29A
,
itchwestaurationstaurationstation * ??????????? :
w
:
. ??? twork . com .
* Arra - hy ” ? * * , * , HA All rs4 ? Art / That Iss = " pdam Shar m ????????????????????????????????
: ? ? ?? ? : : |
FIG . 29B

U . S . Patent May 1 , 2018 Sheet 30 of 43 US 9 , 957 , 531 B1
?? ?
???????????? ? ??
???
?????????????? ???? ? ???? ??
???? ????????????? . ? ? ?
???? ???
???? ??? ???? ?? ???? ?? ???? ????? * ????? ???
??? ???? ?????? " " "
: : " " " " " " " " " " " "
: :
FIG . 29C
* * . ?
?? ?? ???? ???
??? 333
???????????????????????????????????????????????????? ???? ??? ?? ????? ?? ?????? ?? ???? ???? ? ?????? . ? ????
*
* ??????????????????????????????????????????????????????????????????? ???? ?? ????????? ???????????????? ???????????????????? ? ?????? ???????????????? ?????????????????????????
FIG . 29D

U . S . Patent M ay1 , 2018 Sheet 31 of 43 US 9 , 957 , 531 B1
. . . . . . . . . . . . . . . . . . . . . . - - ??????? ????? ?? :
;
?????????? • ; ; ; • • • ' ' , ' ; •• • • ; $ + + + + + + : ; ; : ; ' ' t , + + + + + + + ; + - it , , , + + + + + ? ??
…
, , , , ,
abg9 / gnas # - ???? ????????
· -
-
*
* *
” - - - - * * *
. . . . . … . . . . … . . . . . . . . . . … … … . . . . . ??????????? : , : : : : . . . . . . . : . : . . . , : ? ? : ? : : : : : ? : ? : : : : : ? : ? , ;
- - ???????????????????????? ? ????????????????????????????????? ???
? : ? ?• • • • • •• • • • •?• • • •?•• •??• • • •????• • • • • • • •?• • • • • • • •?• •??• • • • • • • • •• • • •?• • • • • • •???• • • • ,
• • • • • ?????????? ???????????????????????????????? ???????????????????? ??????????????????? ?????????????? ? ????????? ?????? ? ???????????????? ???????????????????
FIG . 30

FIG . 31
US 9 , 957 , 531 B1
??????
?
??
????
???? 2
????????????????????????????????????????????????????????????????????????????????????????????????????????????? 2
?????????????????????????????????????????????????????????????????????????????????????????????????????????????? . .
ww
?
???
.
: ??? 4 ??????????????????????????????????????????????????????????????? . ????????????????????
.
?? 546492
* * * * *
????????????????????????????????????????????????????????????????????????????????? + + + ? + + + 144
? . . ????
. . . ?? - - - - - - - ?•
* *
*
*
*
. . * * *
* *
*
* *
* 4
. . .
* * * * * * :
. . . .
* + + + +
* * *
& &
, , ,
,
. . . * * * * *
. . . "
* *
+ + + *
* *
. * * * * * * * * * * * * * * * * * * * * * * = * b••A • - •??? * * * b * * * * * * * * * * * * * * * * * * * * * * *
} } }
??
& - - - - - - - - . .
& & & &
* * * * *
* *
. . . . . . . . * * *
* * .
* * *
?? ???? * * * *
* * *
??
* = * * + G . +
. . " ( : . . .
????? 22 & & & & & & &
. * * * * * * 64
& & & &
. . . .
+ + + + + A $ $ $ * * * *
. . . : - - - - - - • - 3
* * * * * * * .
???????? )
. . . . . . . . . . .
66544 444
& ?? . .
. . . ?? : . . . . . . . :
& &
?????? 36
: : : : : . . . . . . .
. . . . . . . . . . . . . . . . . . ???? . . . : 3 . . . . . . .
. .
?????????? .
. . . . . . . . . "
??????????••? . .
. .
. . . .
. . . . . . ? . . .
. . .
.
.
. ;
??
? . . . . . . . . . . . . . . . . . 4 . . . . . .
. . .
* * *
* *
. . .
. . . . . . . . . . . . ?? . . . .
??? ) 2 (
. . . . . . 4
. . . . . . . . . . . . . . . . : * * *
?? 682
28289
Sheet 32 of 43
* Cof 3346
Zi 96
3E = 0
z ?? L03 , 0
fu { padik -
??
w Al
-
3a23
????? 4
2 ?? . ????
tititititittitivil
~
??
•
4
???? *
$
??????
???? .
????
???????? ' ???? . ? . ?? ???? ' ? # ? ' ?? ' ? ' ? ' * * * * * * * * * * * *
.
1 ,
221 ,
248
= * * - - - - - 1 = - * - ' & = * - ' '
.
.
A
A
.
.
.
. - - - 3 - - 4 - - * * - ' - ' - 4 - 4 - 41
,
2214 ' ? ' ????????? '
+ ???????????? 4 ????????????????????????????????????????????????????????????????????????? )
. * * * * . . 444
S65
. .
* * * * . . . . . . . .
. . . .
. •
??????????
???? f ??
????????????
4 , 4 $ 4 . 44 ? . 4 . 44 . 4 . 64 . 2 . 4
. . .
,
May 1 , 2018
* * * *
, ,
. . .
A464444864444444 - 4 - 4444444444444244444 - 444A464944
. . . . . . . . . . . . . . . . . . . . . . . . . . . . :
* * * , ,
. .
. ,
? 2
*
. . . . . . . . . . . . . .
. . . . .
?
?????????? ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~
.
* * , ,
??
* •••••••••••• * : * : * : •••••••••••••••••••••••••••
* ' ,
??????????
*
. . 1 ? * *
??????????
. . .
*
. ' * 4 . . *
* *
* *
* *
* * * , , ,
. . .
. . ?
???? ds ? . ?? . . . . * . . . . $ . 64 . 4 . 4 . *
, ,
~
-
cccx ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; ; xxx ; ; ; xxxxxccccx ; ; ; ; ; ; ; ; ace
* *
. *
* : . . * . . . . “ “ . . + . . . .
* *
. . . . . . . . . . . . . .
, 4
. . . .
? * ; ; z :
*
??????
. . .
*
?????? .
* *
4 * *
* * 1 # * * * * * * * } * *
? *
* * *
( ????
?????? . ?? . ??
? . . . .
* * * * * ?? *
.
* * * * *
. . . . . . . . .
??????
???????????????????????? ?????? . ?? . ? .
?????????????????????????????????????????????????????????????????????????????????????????????????????????????????????? | ?? 85 ?? #
$ rg /
px
?????? : : : 20
??
Co { { 32C
_ 2 ( 56 3dcc
Z 56 376C
U . S . Patent
ttt * * * * * ttitull ,
20 -
8a2Q
| ????? i
tatitutituttiti .
. ?? " ???????????????

U . S . Patent
Buk
AOR
po * * * *
Ach
H - HS - COA
COOR
. . . . . . .
R - OH
ok , R . COOH
v
* * *
A COA
An Acid
An Aldehy An Aldehyde
word
An Alcohol / Diol
May 1 , 2018
Propanoyl - CoA
. .
Propionate
1 - Propanol
Propionaldehyde
.
.
.
.
.
. . 3 - 4 yoroxybutyryl . CA
3 . tydroxybutyrate
3 . Hydroxybutycaldersyde
1 , 3 - Butanediol
( C4 )
.
. .
. . .
.
. .
.
. .
.
Acetyl - CoA - - * *
*
V
,
.
. . . .
. .
Hexanoy . com
Captoate
exaldetide
1 - Hexaned
( C6 )
.
.
.
Sheet 33 of 43
-
Octanays - 004
Octuoate
Cateye
1 - Octanol
180 )
FIG . 32
US 9 , 957 , 531 B1

U . S . Patent May 1 , 2018 Sheet 34 of 43 US 9 , 957 , 531 B1
DTNB X - COA
pto COA Coal TNB anion – TNB - COA TNB anion + TNB - COA ADR X - P
buk Oxyluciferin / CO / AMP / PP AME D - luciferin / O , Luciferase X - acid - ATP light
FIG . 33

U . S . Patent May 1 , 2018 Sheet 35 of 43 US 9 , 957 , 531 B1
succinyl - Cod acetyl - CoA redes substrate
+ 177 muttaa 14464
22 til 1991
hati
3 - oxo - adipyl - CoA
23
3 - hydroxyadipy - C04
2 , 3 - dehydroadipyl - CoA
WWWWW adipy - COA
adipic acid
FIG . 34

U . S . Patent May 1 , 2018 Sheet 36 of 43 US 9 , 957 , 531 B1
3 - kydroxybutyry - CDA
crotonyi - C0A
31 n se inte
betyryl - CoA Crotonate
32 * * *
*
acetobutyryl - CoA crotonaldehyde
* eX93491930 *
36 33 * * *
*
ex < * * * * * * *
2 - buten - 3 - o ! 3 - hydroxyhexandyl - COA acetobutyrate
37 34
3 - hydroxyhexanoate zcelylacetone
38
1 , 3 - hexaldehyde 3 - methyl - 2 - Gutanol
39
1 , 3 - hexanediol
FIG . 35

atent May 1 , 2018 Sheet 37 of 43 US 9 , 957 , 531 B1
3 : 3y - - ? $ $ 53 ; ???
32 -
- - ?tyzy? - -
41
3 ???yxxx - 3 - XX3?? ? % 3 3
42
+ 3333 ??? XX
??? ???
xxx 47
isovaleraldehyde
45
3 3333??? ; 33
FIG . 36

U . S . Patent May 1 , 2018 Sheet 38 of 43 US 9 , 957 , 531 B1
se
se en
00 de hoy
die op
0 . 5 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . : : . ' : . : . ' : , , , , , . . .
he 4
Culture days
FIG . 37

U . S . Patent May 1 , 2018 Sheet 39 of 43 US 9 , 957 , 531 B1
* * * * * Metabolites ) ( g / L )
0 2 4 II
6 8 Time ( d )
10 12
Ethanol Butanediol Biomass
FIG . 38A
1
- - - - ( Metabolites ( g / l ) 0 . 6 0 . 5
-
0 . 4 - -
0 . 3 0 . 2 0 . 1 . . . . . . . . . . . . .
O 2 4 10 12 6 8 Time ( 0 )
Hydroxybutyrate 3 Isopropanol
FIG . 38B

U . S . Patent May 1 , 2018 Sheet 40 of 43 US 9 , 957 , 531 B1
& : AORADC * Adh
C , Acetoacyl - COA Crocetoacid Acetone / Cimethyl - 2 - 01
C3 - OH - COA woo CA 3 - OH - acid www1 , 3 - diol
C Enoxl - CoA * * C Enoate C , 2 - en - 1 - 01 900 - 28
y Acyl - Cos Coacid Calcohol Toy
Cora Acetoacyl - CoA evenementen Acetoacid 1 1 1 1 Acetone / Cua methyl - 2 - ol
( 23 - OH - COA 3 - 04 - acid , - diol
Comez Enoyl - cos word Green
C - 2 Enoate C342 2 - en - 1 - 01
1 - 2cd Cz 1 - alcohol C2 Acyl - Cos VAM
e Acetoacyl - CoA www C hcetoacid wwwC Acetone , Cox peth¥1 - 2 - 01 ????? etc etc etc etc cie
FIG . 39

US 9 , 957 , 531 B1
VVD
Sheet 41 of 43
bob 41 3w
Skep ui awe anno
FIG . 40
singing 06
FIG . 41
96
. . . . . . . . . . . . . .
009 : 21196 : 28 . 81
. . . . . . . .
03 - 10 21 . 3 - 800
15 . . .
2 . 0
?
? govori
i
seu
o o
o
May 1 , 2018
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
enes based on
atent
7 / 8
ani

U . S . Patent May 1 , 2018 Sheet 42 of 43 US 9 , 957 , 531 B1
3 - hydroxybutyrate ( g / L ] o No 0
- -
- -
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
FIG . 42A
co
. . . . . . . . . . .
o
-
-
- -
3 - hydroxybutyrate ( g / L ] TO
. . . . ' . ' . ' . ' .
0 1 2 3 4 5 6 7 8 9 10 Time [ d ]
11 12 13 14 15 16 17 18
FIG . 42B

U . S . Patent May 1 , 2018 Sheet 43 of 43 US 9 , 957 , 531 B1
. . 0 . 00 prb - buk I w / ptb - buk
II 18 18 Culture time in days
- . - . - . - . - . - . - . - . - . - . - . - . - . - . - . - . - . - . - . - , - , - , - , - , - , - , - , - , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , , . , . , . , .
FIG . 43
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
mo ) joueu i ste s 1 , 3 - BDO ( mg ) All * * * *
0 3 6 15 21 28 10 Days to culture
FIG . 44

US 9 , 957 , 531 B1
GENETICALLY ENGINEERED BACTERIUM ( Ptb ) and exogenous butyrate kinase ( Buk ) ( Ptb - Buk ) . Gen FOR THE PRODUCTION OF erally , the Ptb - Buk acts on a non - native substrate , e . g . , a
3 - HYDROXYBUTYRATE substrate other than butanoyl - CoA and / or butanoyl phos phate , and produces a non - native product , e . g . , a product
CROSS REFERENCE TO RELATED 5 other than butanoyl phosphate or butyrate . In certain APPLICATIONS embodiments , the Ptb - Buk converts acetoacetyl - CoA to This application is a continuation of U . S . patent applica acetoacetate , 3 - hydroxyisovaleryl - CoA to 3 - hydroxyisoval
tion Ser . No . 15 / 293 , 191 filed Oct . 13 , 2016 , which claims erate , 3 - hydroxybutyryl - CoA to 3 - hydroxybutyrate , or 2 - hy the benefit of U . S . Provisional Patent Application No . droxyisobutyryl - CoA to 2 - hydroxyisobutyrate .
10 62 / 240 , 850 filed Oct . 13 , 2015 , the entireties of which are The bacterium may produce one or more of an acid , an incorporated herein by reference . alkene , a ketone , an aldehyde , an alcohol , or a diol . More
specifically , the bacterium may produce one or more of BACKGROUND OF THE INVENTION acetone or a precursor thereof , isopropanol or a precursor
With recent advances in fermentation and metabolic engi - 15 thereof , isobutylene or a precursor thereof , 3 - hydroxybu neering , fermentation routes to various products have been tyrate or a precursor thereof , 1 , 3 - butanediol or a precursor identified and developed ( Clomburg , Appl Microbiol Bio thereof , 2 - hydroxyisobutyrate or a precursor thereof , adipic technol , 86 : 419 - 434 , 2010 ; Peralta - Yahya , Biotechnol J , 5 : acid or a precursor thereof , 1 , 3 - hexanediol or a precursor 147 - 162 , 2010 ; Cho , Biotechnol Adv , pii : S0734 - 9750 ( 14 ) thereof , 3 - methyl - 2 - butanol or a precursor thereof , 2 - buten 00181 - 5 , 2014 . However , all of these fermentation routes are 20 l - ol or a precursor thereof , isovalerate or a precursor thereof , energy ( ATP ) - consuming or , at best , energy ( ATP ) - neutral , or isoamyl alcohol or a precursor thereof . The bacterium which restricts product yield in energy - limited systems and does not typically produce butanol . uncouples product production from microorganism growth . The bacterium may further comprise a disruptive muta The present invention provides energy ( ATP ) - generating tion in a phosphotransacetylase ( Pta ) and an acetate kinase pathways that overcome these limitations by providing 25 ( Ack ) . The bacterium may further comprise a disruptive novel fermentation routes and pathways to a variety of mutation in a thioesterase . In another embodiment , the products , including acids , alkenes , aldehydes , alcohols , and invention provides a genetically engineered bacterium com diols . These pathways are directly coupled to microorganism prising exogenous Ptb - Buk and exogenous or endogenous growth and offer high product yields . aldehyde : ferredoxin oxidoreductase .
In particular , the invention relates to fermentation path - 30 The invention further provides a method of producing a ways involving Ptb - Buk . Phosphate butyryltransferase ( Ptb ) product comprising culturing the bacterium of any of the ( EC 2 . 3 . 1 . 19 ) natively catalyzes the reaction of butanoyl - aforementioned embodiments in the presence of a substrate . COA and phosphate to form COA and butanoyl phosphate . The product may be , for example , acetone or a precursor Butyrate kinase ( Buk ) ( EC 2 . 7 . 2 . 7 ) natively catalyzes the thereof , isopropanol or a precursor thereof , isobutylene or a reaction of butanoyl phosphate and ADP to form butyrate 35 precursor thereof , 3 - hydroxybutyrate or a precursor thereof , ( butanoate ) and ATP . Accordingly , these enzymes together 1 , 3 - butanediol or a precursor thereof , 2 - hydroxyisobutyrate ( Ptb - Buk ) natively catalyze the conversion of butanoyl - CoA or a precursor thereof , adipic acid or a precursor thereof , to butyrate and generate one ATP via substrate level phos - 1 , 3 - hexanediol or a precursor thereof , 3 - methyl - 2 - butanol or phorylation ( SLP ) . a precursor thereof , 2 - buten - 1 - ol or a precursor thereof ,
The inventors have discovered that Ptb is promiscuous 40 isovalerate or a precursor thereof , or isoamyl alcohol or a and is capable of accepting a variety of acyl - CoAs and precursor thereof . Typically , the substrate is a gaseous enoyl - CoAs as substrates , such that Ptb - Buk may be used to substrate comprising , for example , one or more of CO , CO2 , convert a number of acyl - CoAs and enoyl - CoAs to their and H , . In one embodiment , the gaseous substrate is syngas . corresponding acids or alkenates , respectively , while simul - In another embodiment , the gaseous substrate is an indus taneously generating ATP via substrate level phosphory - 45 trial waste gas . lation .
Furthermore , in combination with an aldehyde ferredoxin BRIEF DESCRIPTION OF THE DRAWINGS oxidoreductase ( AOR ) and an alcohol dehydrogenase , acids formed via the Ptb - Buk system can be further converted to FIG . 1 is a diagram of metabolic pathways for the their respective aldehydes , alcohols , or diols . AOR ( EC 50 production of various products , including acetone , isopro 1 . 2 . 7 . 5 ) catalyzes the reaction of an acid and reduced panol , isobutylene , 3 - hydroxybutyrate , 1 , 3 - butanediol , and ferredoxin ( which can , for example , be generated from 2 - hydroxyisobutyrate from acetyl - CoA . Acetyl - CoA may be oxidation of CO or hydrogen ) to form an aldehyde and generated from any suitable substrate , such as a carbohy oxidized ferredoxin . Alcohol dehydrogenase ( EC 1 . 1 . 1 . 1 drate ( e . g . , sugar ) substrate or a gaseous substrate . In the and EC 1 . 1 . 1 . 2 ) can convert an aldehyde and NAD ( P ) H to 55 present invention , acetyl - CoA is often generated from a an alcohol and NAD ( P ) . gaseous substrate . Bold arrows indicate steps that may be
Introduction of Ptb - Buk and / or AOR into a heterologous catalyzed by Ptb - Buk . species , therefore , provides a novel , alternate route to the FIG . 2 is a diagram showing the reactions natively cata formation of native and non - native products , such as as lyzed by Ptb - Buk , namely the conversion of butanoyl - CoA acids , alkenes , ketones , aldehydes , alcohols , and diols at 60 to butyrate and the generation of one ATP . high yields , thus overcoming limitations of the current state FIG . 3 is a diagram comparing the activities of COA of the art . transferase , thioesterase , and Ptb - Buk .
FIG . 4 is a graph showing average acetone production in SUMMARY OF THE INVENTION E . coli BL21 ( D3 ) modified with plasmids comprising
65 exogenous genes . This data demonstrates the ability of The invention provides a genetically engineered bacte - Ptb - Buk to convert acetoacetyl - CoA to acetoacetate in E .
rium comprising exogenous phosphate butyryltransferase coli in vivo .

US 9 , 957 , 531 B1
FIG . 5 is a graph showing the effect of induction of E . coli etoacetyl - CoA , 3 - hydroxybutyryl - CoA and 2 - hydroxy BL21 ( DE3 ) carrying both the PACYC - ptb - buk and isobutyryl - CoA ) in C . autoethanogenum expressing the Ptb PCOLA - th1A - adc plamids ( expressing thiolase , Ptb - Buk , Buk system from plasmid pMTL82256 - ptb - buk compared and acetoacate decarboxylase ) . to wild - type ( WT ) .
FIG . 6 is a diagram of a pathway designed to use Ptb - Buk 5 FIG . 21A and FIG . 21B are graphs showing reduced for acetone production , while recycling the reducing equiva acyl - CoA hydrolysis activity of C . autoethanogenum strains lents produced in the production of ( R ) - 3 - hydroxybutyryl with inactivated thioesterases ( CT2640 = thioesterase 1 , CT COA and the ATP generated by Ptb - Buk . 1524 = thioesterase 2 , CT1780 = thioesterase 3 ) compared to FIG . 7 is a diagram showing the role of aldehyde : activity found in C . autoethanogenum LZ1560 or LZ1561 . ferredoxin oxidoreductase ( AOR ) , ferredoxin , and Adh in 10 FIG . 22 is a graph showing increased specific isopropanol the production of 1 , 3 - butanediol in C . autoethanogenum . production in a C . autoethanogenum strain with disrupted More generally , AOR may be used to catalyze the conver thioesterase 3 CAETHG _ 1780 compared to wild - type C . sion of an acid to an aldehyde and Adh may be used to autoethanogenum . catalyze the conversion of the aldehyde to an alcohol / diol .
FIG . 8 is a diagram showing the stereospecificity of 15 FIGS . 23A - D are graphs showing growth ( FIG . 23A ) and Ptb - Buk for the production of ( R ) - 3 - hydroxybutyrate and isopropanol ( FIG . 23B ) , acetate ( FIG . 23C ) , and ethanol 2 - hydroxyisobutyrate . The term “ native ” in FIG . 8 refers to ( FIG . 23D ) production profiles of C . autoethanogenum native thioesterase . wild - type and strain with disrupted thioesterase 3
FIG . 9 is a diagram showing the production of isobutene ( CAETHG _ 1780 ) compared to wild - type C . autoethanoge via Ptb - Buk conversion of 3 - hydroxyisovaleryl - CoA and 20 num . 3 - hydroxyisovalerate using alternative pathway 1 . FIG . 24 is a plasmid map of pMTL8225 - pta - ack : : ptb - buk .
FIG . 10 is a diagram showing the production of isobutene FIG . 25 is a gel image indicating the replacement of pta via Ptb - Buk conversion of 3 - hydroxyisovaleryl - CoA and and ack genes replaced with ptb and buk genes and ermB 3 - hydroxyisovalerate using alternative pathway 2 . cassette .
FIG . 11 is a diagram showing the production of 1 , 3 - 25 FIG . 26 is a graph showing increased conversion 3 - hy butanediol via 3 - butyraldehyde dehydrogenase ( Bld ) . droxybutyrate to 1 , 3 - BDO by overexpression of the alde
FIG . 12 is a graph showing isopropanol production in C . hyde : ferredoxin oxidoreductase gene aorl . autoethanogenum using the Ptb - Buk system over a control . FIG . 27 is a graph showing the activity of thioesterase O PMTL85147 - th1A - adc , pMTL85147 - th1A - ptb - buk - adc . TesB , Pta - Ack , and Ptb - Buk system on CoA hydrolysis of FIGS . 13A - F are graphs showing production of 3 - hy - 30 acetoacetyl - CoA , 3 - hydroxybutyryl - CoA and 2 - hydroxy
droxybutyrate , acetate , ethanol , and acetone with modular isobutyryl - CoA compared to control ( BL21 strain ) . Ptb - Buk plasmids in E . coli with different concentrations of inducer s hows highest activity , while Pta - Ack shows no activity . IPTG ( 0 , 50 , 100 uM ) . FIG . 13A : PACYC - ptb - buk , FIGS . 28A and 28B are graphs showing production of PCOLA - th1A - adc , PCDF - phaB . FIG . 13B : PACYC - ptb - 3 - hydroxybutyrate via Ptb - Buk in combination with an buk . pCOLA - th1A - adc , PCDF - phaB - bdh1 . FIG . 13C : 35 ( S ) - specific ( Hbd ) ( FIG . 28A ) or ( R ) - specific 3 - hydroxybu PCOLA - th1A - adc , pCDF - phaB - bdh1 . FIG . 13D : PCOLA - tyrate ( PhaB ) ( FIG . 28B ) dehydrogenase . th1A - adc . FIG . 13E : PCDF - phaB - bdh1 . FIG . 13F : PCDF FIGS . 29A - D are graphs showing LC - MS / MS detection phaB . of 2 - hydroxyisobutyric acid ( 2 - HIB ) and 2 - hydroxybutyrate
FIG . 14 is a plasmid map of plasmid pMTL8225 - budA : : ( 2 - HB ) . FIG . 29A : 1 mM 2 - HIB standard . FIG . 29B : 1 mM thlA - phaB . 40 2 - HB standard .
FIG . 15 is a gel image of PCR verification of replacement FIG . 29C : 0 . 5 mM 2 - HB and 2 - HIB standard . FIG . 29D : of acetolactate synthase ( budA ) genes with thiolase ( th1A ) duplicate of C . autoethanogenum sample showing 2 - HIB and 3 - hydroxybutyryl - CoA dehydrogenase ( phaB ) genes in and 2 - HB production from gas . C . autoethanogenum for 4 clones ( 1 , 4 , 7 , 9 ) compared to FIG . 30 is a set of graphs showing GC - MS confirmation wild - type ( W ) . All clones are positive as seen by a larger 45 of 2 - hydroxyisobutyric acid ( 8 . 91 min ) production . First PCR fragment size compared to wild - type . panel : C . autoethanogenum + pMTL83155 - th1A - hbd - Pwl
FIG . 16 is a graph showing fermentation profile of a batch meaBhcmA - hcmB + pMTL82256 - tesB . Second panel : C . fermentation C . autoethanogenum budA : : thlAphaB strain autoethanogenum + pMTL83155 - thlA - hbd - Pwl - meaBh and demonstrating 3 - hydroxybutyrate and 1 , 3 - butanediol cmA - hcmB + pMTL82256 - ptb - buk ( spectrum ) . Third panel : formation from gas . 50 E . coli + pMTL83155 - th1A - hbd - Pwl - meaBhcmA - hcmB +
FIG . 17A is a graph showing production of 1 , 3 - BDO via PMTL82256 - tesB . Fourth panel : E . coli + pMTL83155 - th1A thiolase , 3 - hydroxybutyryl - CoA dehydrogenase ( Bld ) , and hbd - Pwl - meaBhcmA - hcmB + pMTL82256 - ptb - buk . butyraldehyde dehydrogenase . FIG . 17B is a graph showing FIG . 31 is a set of graphs of real time PCR showing the impact of bld expression on growth . expression of genes of the 2 - HIBA pathway ( th1A , hba ,
FIG . 18A is a graph showing the formation of 3 - hydroxy - 55 meaBhcmA , hcmB from pta - ack promoter and respectively butyrate and 1 , 3 - butanediol from gaseous substrate in C . Wood - Ljungdahl operon promoter ) in E . coli , C . autoetha autoethanogenum PMTL8315 - Pfdx - hbd1 - th1A . FIG . 18B is nogenum LZ1561 at 30° C . , and C . autoethanogenum a graph showing the reduction of acetate to ethanol in the LZ1561 at 37° C . same culture . FIG . 32 is a diagram showing the production of various
FIG . 19 is a graph showing the fermentation profile for 60 products in a microorganism comprising Ptb - Buk , AOR , and strain C . autoethanogenum PMTL8315 - Pfdx - hbd1 - th1A Adh . demonstrating formation of 3 - hydroxybutyrate and 1 , 3 - bu - FIG . 33 is a diagram showing the coupling firefly tanediol from gaseous substrate in continuous culture ( where luciferase ( Luc ) to the Ptb - Buk system to characterize indicated , media was replenished continuously with given Ptb - Buk variants . dilution rate D ) . 65 FIG . 34 is a diagram of metabolic pathways for the
FIG . 20A and FIG . 20B are graphs showing increased production of various products , including adipic acid . Bold COA hydrolysis activity on a range of acyl - CoAs ( ac arrows indicate steps that may be catalyzed by Ptb - Buk .

US 9 , 957 , 531 B1
FIG . 35 is a diagram of metabolic pathways for the ljungdahlii , and Clostridium ragsdalei do not have known production of various products , including 1 , 3 - hexanediol , native activity for this step . Escherichia coli has native 2 - methyl - 2 - butanol , and 2 - buten - 1 - ol . Bold arrows indicate activity for this step . steps that may be catalyzed by Ptb - Buk . Step 2 shows the conversion of acetoacetyl - CoA to
FIG . 36 is a diagram of metabolic pathways for the 5 acetoacetate . This step may be catalyzed by CoA - transferase production of various products , including isovalerate and ( i . e . , acetyl - CoA : acetoacetyl - CoA transferase ) ( EC 2 . 8 . 3 . 9 ) . isoamyl alcohol . Bold arrows indicate steps that may be The COA - transferase may be , for example , CtfAB , a het catalyzed by Ptb - Buk . erodimer comprising subunits CtfA and CtfB , from
FIG . 37 is a graph of 3 - HB production in C . autoetha Clostridium beijerinckii ( CtfA , WP _ 012059996 . 1 ) ( SEQ ID nogenum containing plasmid pMTL82256 - thlA - ctfAB at 10 NO : 5 ) ( CtfB , WP _ 012059997 . 1 ) ( SEO ID NO : 6 ) . This step various points of growth . may also be catalyzed by thioesterase ( EC 3 . 1 . 2 . 20 ) . The
FIG . 38A is a graph showing the growth and ethanol and thioesterase may be , for example , TesB from Escherichia 2 , 3 - butanediol production profile of strain C . autoethano coli ( NP _ 414986 . 1 ) ( SEQ ID NO : 7 ) . This step may also be genum pta - ack : : ptb - buk + pMTL85147 - th1A - ptb - buk - adc . catalyzed by a putative thioesterase , e . g . , from Clostridium FIG . 38B is a graph showing the isopropanol and 3 - HB 15 autoethanogenum or Clostridium ljungdahlii . In particular , production profile of strain C . autoethanogenum pta - ack : : three putative thioesterases have been identified in ptb - buk + pMTL85147 - th1A - ptb - buk - adc . Clostridium autoethanogenum : ( 1 ) “ thioesterase 1 ”
FIG . 39 is a diagram of a pathway scheme for producing ( AGY74947 . 1 ; annotated as palmitoyl - CoA hydrolase ; SEQ a range of C4 , C6 , C3 , C10 , C12 , C14 alcohols , ketones , enols ID NO : 8 ) , ( 2 ) “ thioesterase 2 ” ( AGY75747 . 1 ; annotated as or diols via combining known chain elongation pathway 20 4 - hydroxybenzoyl - CoA thioesterase ; SEQ ID NO : 9 ) , and ( Hbd , Crt , Bcd - EtfAB , Thl ) with Ptb - Buk + AOR / Adc - Adh . ( 3 ) “ thioesterase 3 ” ( AGY75999 . 1 ; annotated as putative
FIG . 40 is a graph showing production of 3 - HB and thioesterase ; SEQ ID NO : 10 ) . Three putative thioesterases 1 , 3 - BDO by C . autoethanogenum transformed with plasmid have also been identified in Clostridium ljungdahlii : ( 1 ) PMTL83159 - phaB - thlA at various points of growth . “ thioesterase 1 ” ( ADK15695 . 1 ; annotated as predicted acyl
FIG . 41 is a graph showing production of 3 - HB and 25 COA thioesterase 1 ; SEQ ID NO : 11 ) , ( 2 ) “ thioesterase 2 " 1 , 3 - BDO by C . autoethanogenum comprising budA knock - ( ADK16655 . 1 ; annotated as predicted thioesterase ; SEQ ID out and pMTL - HBD - ThlA at various points of growth . NO : 12 ) , and ( 3 ) “ thioesterase 3 ” ( ADK16959 . 1 ; annotated
FIG . 42A is a graph showing production of 3 - HB in a C . as predicted thioesterase ; SEQ ID NO : 13 ) . This step may autoethanogenum pMTL83159 - phaB - th1A + PMTL82256 also be catalyzed by phosphate butyryltransferase ( EC fermentation . FIG . 42B is a graph showing production of 30 2 . 3 . 1 . 19 ) + butyrate kinase ( EC 2 . 7 . 2 . 7 ) . Exemplary sources 3 - HB in a C . autoethanogenum pMTL83159 - phaB - th1A + for phosphate butyryltransferase and butyrate kinase are pMTL82256 - buk - ptb fermentation . described elsewhere in this application . Native enzymes in
FIG . 43 is a graph showing the production of 3 - HB in a Clostridium autoethanogenum , Clostridium ljungdahlii , and C . autoethanogenum strain with thioesterase knockout Clostridium ragsdalei ( or Escherichia coli ) , such as thio ( ACAETHG _ 1524 ) expressing plasmid pMTL83156 - phaB - 35 esterases from Clostridium autoethanogenum , may catalyze thlA with and without Ptb - Buk expression plasmid this step and result in the production of some amount of pMTL82256 - buk - ptb . downstream products . However , introduction of an exog
FIG . 44 is a graph showing showing ethanol and 1 , 3 - BDO enous enzyme or overexpression of an endogenous enzyme production in a C . autoethanogenum strain expressing plas may be required to produce downstream products at desir mid pMTL82256 - hbd - th1A ( 2pf ) with and without AOR 40 able levels . Additionally , in certain embodiments , a disrup overexpression plasmid pMTL83159 - aorl ( + aori ) . tive mutation may be introduced to an endogenous enzyme ,
such as an endogenous thioesterase , to reduce or eliminate DETAILED DESCRIPTION OF THE competition with introduced Ptb - Buk .
INVENTION Step 3 shows the conversion of acetoacetate to acetone . 45 This step may be catalyzed by an acetoacetate decarboxylase
Metabolic Pathways of FIGS . 1 and 34 - 36 ( EC 4 . 1 . 1 . 4 ) . The acetoacetate decarboxylase may be , for FIGS . 1 and 34 - 36 are diagrams of metabolic pathways example , Adc from Clostridium beijerinckii
for the production of various acid , alkene , ketone , aldehyde , ( WP _ 012059998 . 1 ) ( SEQ ID NO : 14 ) . This step may also be alcohol , and diol products , including acetone , isopropanol , catalyzed by an alpha - ketoisovalerate decarboxylase ( EC isobutylene , 3 - hydroxybutyrate ( R - and S - isomers ) , 1 , 3 - 50 4 . 1 . 1 . 74 ) . The alpha - ketoisovalerate decarboxylase may be , butanediol , 2 - hydroxyisobutyrate , adipic acid , 1 , 3 - hexane - for example , KivD from Lactococcus lactis ( SEQ ID NO : diol , 2 - methyl - 2 - butanol , 2 - buten - 1 - ol , isovalerate , and iso - 15 ) . Clostridium autoethanogenum , Clostridium ljungdahlii , amyl alcohol from a substrate . Bold arrows indicate steps and Clostridium ragsdalei do not have known native activity that may be catalyzed by Ptb - Buk . Exemplary enzymes are for this step . Additionally , Escherichia coli does not have provided for each of the steps and enzymatic pathways 55 known native activity for this step . Rarely , conversion of detailed in FIGS . 1 and 34 - 36 . However , additional suitable acetoacetate to acetone may occur spontaneously . However , enzymes may be known to a person of ordinary skill in the spontaneous conversion is highly inefficient and unlikely to art . result in the production of downstream products at desirable
Step 1 shows the conversion of acetyl - CoA to acetoacetyl - levels . COA . This step may be catalyzed by thiolase ( i . e . , acetyl - 60 Step 4 shows the conversion of acetone to isopropanol . COA acetyltransferase ) ( EC 2 . 3 . 1 . 9 ) . The thiolase may be , This step may be catalyzed by a primary : secondary alcohol for example , ThlA from Clostridium acetobutylicum dehydrogenase ( EC 1 . 1 . 1 . 2 ) . The primary : secondary alcohol ( WP _ 010966157 . 1 ) ( SEQ ID NO : 1 ) , PhaA from Cupriavi dehydrogenase may be , for example , SecAdh from dus necator ( WP _ 013956452 . 1 ) ( SEQ ID NO : 2 ) , BktB Clostridium autoethanogenum ( AGY74782 . 1 ) ( SEQ ID from Cupriavidus necator ( WP _ 011615089 . 1 ) ( SEQ ID 65 NO : 16 ) , SecAdh from Clostridium ljungdahlii NO : 3 ) , or AtoB from Escherichia coli ( NP _ 416728 . 1 ) ( SEQ ( ADK15544 . 1 ) ( SEQ ID NO : 17 ) , SecAdh from Clostridium ID NO : 4 ) . Clostridium autoethanogenum , Clostridium ragsdalei ( WP _ 013239134 . 1 ) ( SEQ ID NO : 18 ) , or SecAdh

US 9 , 957 , 531 B1
from Clostridium beijerinckii ( WP _ 026889046 . 1 ) ( SEQ ID NO : 12 ) , and ( 3 ) “ thioesterase 3 ” ( ADK16959 . 1 ; annotated NO : 19 ) . This step may also be catalyzed by a primary : as predicted thioesterase ; SEQ ID NO : 13 ) . This step may secondary alcohol dehydrogenase ( EC 1 . 1 . 1 . 80 ) , such as also be catalyzed by phosphate butyryltransferase ( EC SecAdh from Thermoanaerobacter brokii ( 3FSR _ A ) ( SEQ 2 . 3 . 1 . 19 ) + butyrate kinase ( EC 2 . 7 . 2 . 7 ) . Exemplary sources ID NO : 20 ) . Clostridium autoethanogenum , Clostridium 5 for phosphate butyryltransferase and butyrate kinase are ljungdahlii , and Clostridium ragsdalei have native activity described elsewhere in this application . Native enzymes in for this step ( Köpke , Appl Environ Microbiol , 80 : 3394 - Clostridium autoethanogenum , Clostridium ljungdahlii , and 3403 , 2014 ) . However , Escherichia coli does not have Clostridium ragsdalei ( or Escherichia coli ) , such as thio known native activity for this step . Knocking down or esterases from Clostridium autoethanogenum , may catalyze knocking out this enzyme in Clostridium autoethanogenum , 10 this step and result in the production of some amount of Clostridium ljungdahlii , or Clostridium ragsdalei results in downstream products . However , introduction of an exog the production and accumulation of acetone rather than enous enzyme or overexpression of an endogenous enzyme isopropanol ( WO 2015 / 085015 ) . may be required to produce downstream products at desir
Step 5 shows the conversion of acetone to 3 - hydroxyis - able levels . Additionally , in certain embodiments , a disrup ovalerate . This step may be catalyzed by a hydroxyisoval - 15 tive mutation may be introduced to an endogenous enzyme , erate synthase , such as hydroxymethylglutaryl - CoA syn - such as an endogenous thioesterase , to reduce or eliminate thase ( HMG - CoA synthase ) ( EC 2 . 3 . 3 . 10 ) from Mus competition with introduced Ptb - Buk . musculus ( SEQ ID NO : 21 ) ( US 2012 / 0110001 ) . The Step 9 shows the conversion of acetyl - CoA to 3 - methyl hydroxymethylglutaryl - CoA synthase may be engineered to 2 - oxopentanoate . This step encompasses a number of enzy improve activity . Clostridium autoethanogenum , 20 matic reactions involved in the isoleucine biosynthesis path Clostridium ljungdahlii , and Clostridium ragsdalei do not way , which is natively present in many bacteria , including have known native activity for this step . Escherichia coli Clostridium autoethanogenum , Clostridium ljungdahlii , and does not have known native activity for this step . Clostridium ragsdalei ( and Escherichia coli ) . Enzymes
Step 6 shows the conversion of 3 - hydroxyisovalerate to involved in the conversion of acetyl - CoA to 3 - methyl - 2 isobutylene ( isobutene ) . This step may be catalyzed by a 25 oxopentanoate may include citramalate synthase ( EC hydroxyisovalerate phosphorylase / decarboxylase . This step 2 . 3 . 1 . 182 ) , 3 - isopropylmalate dehydratase ( EC 4 . 2 . 1 . 35 ) , may also be catalyzed by mevalonate diphosphate decar - 3 - isopropylmalate dehydrogenase ( EC 1 . 1 . 1 . 85 ) , acetolac boxylase ( hydroxyisovalerate decarboxylase ) ( EC 4 . 1 . 1 . 33 ) . tate synthase ( EC 2 . 2 . 1 . 6 ) , ketol - acid reductoisomerase ( EC The mevalonate diphosphate decarboxylase may be , for 1 . 1 . 1 . 86 ) , and / or dihydroxyacid dehydratase ( EC 4 . 2 . 1 . 9 ) . example , Mdd from Saccharomyces cerevisiae 30 The citramalate synthase may be , for example , CimA from ( CAA96324 . 1 ) ( SEQ ID NO : 22 ) or Mdd from Picrophilus Clostridium autoethanogenum ( AGY76958 . 1 ) ( SEQ ID torridus ( WP _ 011178157 . 1 ) ( SEQ ID NO : 23 ) ( US 2011 / NO : 24 ) or CimA from Methanocaldococcus jannaschii 0165644 ; van Leeuwen , Appl Microbiol Biotechnol , 93 : ( NP _ 248395 . 1 ) ( SEQ ID NO : 25 ) . The 3 - isopropylmalate 1377 - 1387 , 2012 ) . Clostridium autoethanogenum , dehydratase may be , for example , LeuCD from Clostridium Clostridium ljungdahlii , and Clostridium ragsdalei do not 35 autoethanogenum ( WP _ 023162955 . 1 , LeuC ; AGY77204 . 1 , have known native activity for this step . Escherichia coli LeuD ) ( SEQ ID NOs : 26 and 27 , respectively ) or LeuCD does not have known native activity for this step from Escherichia coli ( NP _ 414614 . 1 , LeuC ; NP _ 414613 . 1 ,
Step 7 shows the conversion of acetone to 3 - hydroxyis - LeuD ) ( SEQ ID NOs : 28 and 29 , respectively ) . The 3 - iso ovaleryl - CoA . This step may be catalyzed by a 3 - hydroxy propylmalate dehydrogenase may be , for example , LeuB isovaleryl - CoA synthase . Clostridium autoethanogenum , 40 from Clostridium autoethanogenum ( WP _ 023162957 . 1 ) Clostridium ljungdahlii , and Clostridium ragsdalei do not ( SEQ ID NO : 30 ) or LeuB from Escherichia coli have known native activity for this step . Escherichia coli ( NP _ 414615 . 4 ) ( SEQ ID NO : 31 ) . The acetolactate synthase does not have known native activity for this step may be , for example , IlvBN from Clostridium autoethano
Step 8 shows the conversion of 3 - hydroxyisovaleryl - CoA genum ( AGY74359 . 1 , IlvB ; AGY74635 . 1 , IlvB ; to 3 - hydroxyisovalerate . This step may be catalyzed by 45 AGY74360 . 1 , IlvN ) ( SEQ ID NOs : 32 , 33 , and 34 , respec COA - transferase ( i . e . , acetyl - CoA : acetoacetyl - CoA trans - tively ) or IlvBN from Escherichia coli ( NP _ 418127 . 1 , IlvB ; ferase ) ( EC 2 . 8 . 3 . 9 ) . The COA - transferase may be , for NP 418126 . 1 , IlvN ) ( SEQ ID NOs : 35 and 36 , respectively ) . example , CtfAB , a heterodimer comprising subunits CtfA The ketol - acid reductoisomerase may be , for example , Ilvc and CtfB , from Clostridium beijerinckii ( CtfA , from Clostridium autoethanogenum ( WP _ 013238693 . 1 ) WP 012059996 . 1 ) ( SEO ID NO : 5 ) ( CtfB , 50 ( SEQ ID NO : 37 ) or IlvC from Escherichia coli ( NP WP _ 012059997 . 1 ) ( SEQ ID NO : 6 ) . This step may also be 418222 . 1 ) ( SEQ ID NO : 38 ) . The dihydroxyacid dehy catalyzed by thioesterase ( EC 3 . 1 . 2 . 20 ) . The thioesterase dratase may be , for example , IlvD from Clostridium may be , for example , TesB from Escherichia coli autoethanogenum ( WP _ 013238694 . 1 ) ( SEQ ID NO : 39 ) or ( NP _ 414986 . 1 ) ( SEQ ID NO : 7 ) . This step may also be IlvD from Escherichia coli ( YP _ 026248 . 1 ) ( SEQ ID NO : catalyzed by a putative thioesterase , e . g . , from Clostridium 55 40 ) . Clostridium autoethanogenum , Clostridium ljungdahlii , autoethanogenum or Clostridium ljungdahlii . In particular , and Clostridium ragsdalei have native activity for this step . three putative thioesterases have been identified in Step 10 shows the conversion of 3 - methyl - 2 - oxopentoate Clostridium autoethanogenum : ( 1 ) " thioesterase 1 ” to 2 - methylbutanoyl - CoA . This step may be catalyzed by ( AGY74947 . 1 ; annotated as palmitoyl - CoA hydrolase ; SEQ ketoisovalerate oxidoreductase ( EC 1 . 2 . 7 . 7 ) . The ketois ID NO : 8 ) , ( 2 ) “ thioesterase 2 " ( AGY75747 . 1 ; annotated as 60 ovalerate oxidoreductase may be , for example , the Vor 4 - hydroxybenzoyl - CoA thioesterase ; SEQ ID NO : 9 ) , and ABCD from Methanothermobacter thermautotrophicus ( 3 ) “ thioesterase 3 ” ( AGY75999 . 1 ; annotated as putative ( WP _ 010876344 . 1 , VorA ; WP _ 010876343 . 1 , VorB ; thioesterase ; SEQ ID NO : 10 ) . Three putative thioesterases WP _ 010876342 . 1 , VorC ; WP _ 010876341 . 1 , VorD ) ( SEQ have also been identified in Clostridium ljungdahlii : ( 1 ) ID NOs : 41 - 44 , respectively ) or VorABCD from Pyococcus " thioesterase 1 ” ( ADK15695 . 1 ; annotated as predicted acyl - 65 furiosus ( WP _ 011012106 . 1 , VorA ; WP 011012105 . 1 , COA thioesterase 1 ; SEQ ID NO : 11 ) , ( 2 ) “ thioesterase 2 ” VorB ; WP _ 011012108 . 1 , VorC ; WP _ 011012107 . 1 , VorD ) ( ADK16655 . 1 ; annotated as predicted thioesterase ; SEQ ID ( SEQ ID NOs : 45 - 48 , respectively ) . VorABCD is a 4 - sub

US 9 , 957 , 531 B1 10
unit enzyme . Clostridium autoethanogenum , Clostridium Step 14 shows the conversion of 3 - hydroxybutyryl - CoA ljungdahlii , and Clostridium ragsdalei do not have known to 3 - hydroxybutyrate . This step may be catalyzed by thio native activity for this step . Escherichia coli does not have esterase ( EC 3 . 1 . 2 . 20 ) . The thioesterase may be , for known native activity for this step . example , TesB from Escherichia coli ( NP _ 414986 . 1 ) ( SEQ
Step 11 shows the conversion of 2 - methylbutanoyl - CoA 5 A 5 ID NO : 7 ) . This step may also be catalyzed by a putative thioesterase , e . g . , from Clostridium autoethanogenum or to 2 - methylcrotonyl - CoA . This step may be catalyzed by Clostridium ljungdahlii . In particular , three putative thio 2 - methylbutanoyl - CoA dehydrogenase ( EC 1 . 3 . 99 . 12 ) . The esterases have been identified in Clostridium autoethanoge 2 - methylbutanoyl - CoA dehydrogenase may be , for example , num : ( 1 ) " thioesterase 1 ” ( AGY74947 . 1 ; annotated as
AcdH from Streptomyces avermitilis ( AAD44196 . 1 or palmitoyl - CoA hydrolase ; SEQ ID NO : 8 ) , ( 2 ) “ thioesterase BAB69160 . 1 ) ( SEQ ID NO : 49 ) or AcdH from Streptomyces " 2 " ( AGY75747 . 1 ; annotated as 4 - hydroxybenzoyl - COA coelicolor ( AAD44195 . 1 ) ( SEQ ID NO : 50 ) . Clostridium thioesterase ; SEQ ID NO : 9 ) , and ( 3 ) “ thioesterase 3 " autoethanogenum , Clostridium ljungdahlii , and Clostridium ( AGY75999 . 1 ; annotated as putative thioesterase ; SEQ ID ragsdalei do not have known native activity for this step . NO : 10 ) . Three putative thioesterases have also been iden Escherichia coli does not have known native activity for this 15 tified in Clostridium ljungdahlii : ( 1 ) " thioesterase 1 "
15 ( ADK15695 . 1 ; annotated as predicted acyl - CoA thio step . esterase 1 ; SEQ ID NO : 11 ) , ( 2 ) " thioesterase 2 " Step 12 shows the conversion of 2 - methylcrotonyl - CoA ( ADK16655 . 1 ; annotated as predicted thioesterase ; SEQ ID to 3 - hydroxyisovaleryl - CoA . This step may be catalyzed by NO : 12 ) , and ( 3 ) “ thioesterase 3 ” ( ADK16959 . 1 ; annotated crotonase / 3 - hydroxybutyryl - CoA dehydratase ( EC as predicted thioesterase ; SEQ ID NO : 13 ) . This step may 4 . 2 . 1 . 55 ) . The crotonase / 3 - hydroxybutyryl - CoA dehy - 20 also be catalyzed by phosphate butyryltransferase ( EC dratase may be , for example , Crt from Clostridium beijer 2 . 3 . 1 . 19 ) + butyrate kinase ( EC 2 . 7 . 2 . 7 ) . Exemplary sources inckii ( ABR34202 . 1 ) ( SEQ ID NO : 51 ) , Crt from for phosphate butyryltransferase and butyrate kinase are Clostridium acetobutylicum ( NP _ 349318 . 1 ) ( SEQ ID NO : described elsewhere in this application . Native enzymes in 52 ) , or LiuC from Myxococcus xanthus ( WP _ 011553770 . 1 ) . Clostridium autoethanogenum , Clostridium ljungdahlii , and This step may also be catalyzed by crotonyl - CoA carboxy - 25 Clostridium ragsdalei ( or Escherichia coli ) , such as thio lase - reductase ( EC 1 . 3 . 1 . 86 ) . The crotonyl - CoA carboxy - esterases from Clostridium autoethanogenum , may catalyze lase - reductase may be , for example , Cor from Treponema this step and result in the production of some amount of denticola ( NP _ 971211 . 1 ) ( SEQ ID NO : 53 ) . This step may downstream products . However , introduction of an exog also be catalyzed by crotonyl - CoA reductase ( EC 1 . 3 . 1 . 44 ) . enous enzyme or overexpression of an endogenous enzyme The crotonyl - CoA reductase may be , for example , Ter from 30 may be required to produce downstream products at desir Euglena gracilis ( AAW66853 . 1 ) ( SEQ ID NO : 54 ) . This able levels . Additionally , in certain embodiments , a disrup step may also be catalyzed by a 3 - hydroxypropionyl - CoA tive mutation may be introduced to an endogenous enzyme , dehydratase ( EC 4 . 2 . 1 . 116 ) . This 3 - hydroxypropionyl - CoA such as an endogenous thioesterase , to reduce or eliminate dehydratase may be , for example , Msed _ 2001 from Metal - competition with introduced Ptb - Buk . losphaera sedula ( WP _ 012021928 . 1 ) . This step may also be 35 Step 15 shows the conversion of 3 - hydroxybutyrate to catalyzed by a enoyl - CoA hydratase . This enoyl - CoA acetoacetate . This step may be catalyzed by 3 - hydroxybu hydratase ( 4 . 2 . 1 . 17 ) may be , for example , YngF from Bacil - tyrate dehydrogenase ( EC 1 . 1 . 1 . 30 ) . The 3 - hydroxybutyrate lus anthracis ( WP _ 000787371 . 1 ) . Clostridium autoethano - dehydrogenase may be , for example , Bdh1 from Ralstonia genum , Clostridium ljungdahlii , and Clostridium ragsdalei pickettii ( BAE72684 . 1 ) ( SEQ ID NO : 60 ) or Bdh2 from do not have known native activity for this step . Escherichia 40 Ralstonia pickettii ( BAE72685 . 1 ) ( SEQ ID NO : 61 ) . The coli does not have known native activity for this step . reverse reaction , the conversion of acetoacetate to 3 - hy
Step 13 shows the conversion of acetoacetyl - CoA to droxybutyrate , may be catalyzed by different 3 - hydroxybu 3 - hydroxybutyryl - CoA . This step may be catalyzed by 3 - hy tyrate dehydrogenase ( EC 1 . 1 . 1 . 30 ) enzymes . For example , droxybutyryl - CoA dehydrogenase ( EC 1 . 1 . 1 . 157 ) . The the conversion of acetoacetate to 3 - hydroxybutyrate may be 3 - hydroxybutyryl - CoA dehydrogenase may be , for example , 45 catalyzed by Bdh from Clostridium autoethanogenum Hbd from Clostridium beijerinckii ( WP _ 011967675 . 1 ) ( AGY75962 ) ( SEQ ID NO : 62 ) . Clostridium ljungdahlii and ( SEO ID NO : 55 ) , Hbd from Clostridium acetobutylicum Clostridium ragsdalei likely have enzymes with similar ( NP _ 349314 . 1 ) ( SEQ ID NO : 56 ) , or Hbdl from activity . Escherichia coli does not have known native activ Clostridium kluyveri ( WP _ 011989027 . 1 ) ( SEQ ID NO : 57 ) . ity for this step . This step may also be catalyzed by acetoacetyl - CoA reduc - 50 Step 16 shows the conversion of 3 - hydroxybutyrate to tase ( EC 4 . 2 . 1 . 36 ) . The acetoacetyl - CoA reductase may be , 3 - hydroxybutyrylaldehyde . This step may be catalyzed by for example , PhaB from Cupriavidus necator aldehyde : ferredoxin oxidoreductase ( EC 1 . 2 . 7 . 5 ) . The alde ( WP _ 010810131 . 1 ) ( SEQ ID NO : 58 ) . This step may also be hyde : ferredoxin oxidoreductase ( AOR ) may be , for catalyzed by acetoacetyl - CoA hydratase ( EC 4 . 2 . 1 . 119 ) . Of example , AOR from Clostridium autoethanogenum note , PhaB is R - specific and Hbd is S - specific . Additionally , 55 ( WP 013238665 . 1 ; WP 013238675 . 1 ) ( SEQ ID NOs : 63 Hbd1 from Clostridium kluyveri is NADPH - dependent and and 64 , respectively ) or AOR from Clostridium ljungdahlii Hbd from Clostridium acetobutylicum and Clostridium bei - ( ADK15073 . 1 ; ADK15083 . 1 ) ( SEQ ID NOs : 65 and 66 , jerinckii are NADH - dependent . Clostridium autoethanoge respectively ) . In further embodiments , the aldehyde : ferre num , Clostridium ljungdahlii , and Clostridium ragsdalei do doxin oxidoreductase may be or may be derived , for not have known native activity for this step . Escherichia coli 60 example , from any of the following sources , the sequences does not have known native activity for this step . of which are publically available :
Description Microrganism Accession GeneID
aldehyde : ferredoxin oxidoreductase Acidilobus saccharovorans 345 - 15 aldehyde : ferredoxin oxidoreductase Acidilobus saccharovorans 345 - 15
N C _ 014374 . 1 NC _ 014374 . 1
9498931 9499504

US 9 , 957 , 531 B1 11
- continued Description Microrganism Accession GeneID
NC _ 014374 . 1 NC 014374 . 1 NC 013926 . 1 NC _ 013926 . 1 NC 013926 . 1 NC _ 013926 . 1 NC _ 013926 . 1 NC 019942 . 1 NC 019942 . 1 NC _ 019942 . 1 NC _ 019942 . 1 NC _ 000854 . 2 NC _ 000917 . 1 NC _ 000917 . 1 NC _ 000917 . 1 NC _ 000917 . 1 NC _ 013741 . 1
9499550 9498997 8828075 8828511 8828305 8827762 8827370
14306579 14306982 14306639 14307339 1444491 1483287 1483233 1483554 1485513 8738726
NC _ 013741 . 1 8740019
NC _ 021169 . 1 15392228
NC _ 021169 . 1 15393814
NC _ 021169 . 1 15391826
NC _ 021169 . 1 15393763
NC _ 021169 . 1 15393491
NC _ 015320 . 1 NC 015320 . 1 NC _ 019791 . 1
10393142 10395048 14212403
NC _ 019791 . 1 14211524
NC _ 019791 . 1 14212092
aldehyde : ferredoxin oxidoreductase Acidilobus saccharovorans 345 - 15 aldehyde : ferredoxin oxidoreductase Acidilobus saccharovorans 345 - 15 aldehyde : ferredoxin oxidoreductase Aciduliprofundum boonei T469 aldehyde : ferredoxin oxidoreductase Aciduliprofundum boonei T469 aldehyde : ferredoxin oxidoreductase Aciduliprofundum boonei T469 aldehyde : ferredoxin oxidoreductase Aciduliprofundum boonei T469 aldehyde : ferredoxin oxidoreductase Aciduliprofundum boonei 1469 aldehyde : ferredoxin oxidoreductase Aciduliprofundum sp . MARO8 - 339 aldehyde : ferredoxin oxidoreductase Aciduliprofundum sp . MAR08 - 339 aldehyde : ferredoxin oxidoreductase Aciduliprofundum sp . MAR08 - 339 aldehyde : ferredoxin oxidoreductase Aciduliprofundum sp . MAR08 - 339 aldehyde : ferredoxin oxidoreductase Aeropyrum pernix K1 aldehyde : ferredoxin oxidoreductase Archaeoglobus fulgidus DSM 4304 aldehyde : ferredoxin oxidoreductase Archaeoglobus fulgidus DSM 4304 aldehyde : ferredoxin oxidoreductase Archaeoglobus fulgidus DSM 4304 aldehyde : ferredoxin oxidoreductase Archaeoglobus fulgidus DSM 4304 aldehyde : ferredoxin oxidoreductase Archaeoglobus profundus DSM
5631 aldehyde : ferredoxin oxidoreductase Archaeoglobus profundus DSM
5631 aldehyde : ferredoxin oxidoreductase Archaeoglobus sulfaticallidus
PM70 - 1 aldehyde : ferredoxin oxidoreductase Archaeoglobus sulfaticallidus
PM70 - 1 aldehyde : ferredoxin oxidoreductase Archaeoglobus sulfaticallidus
PM70 - 1 aldehyde : ferredoxin oxidoreductase Archaeoglobus sulfaticallidus
PM70 - 1 aldehyde : ferredoxin oxidoreductase Archaeoglobus sulfaticallidus
PM70 - 1 aldehyde : ferredoxin oxidoreductase Archaeoglobus veneficus SNP6 aldehyde : ferredoxin oxidoreductase Archaeoglobus veneficus SNP6 aldehyde : ferredoxin oxidoreductase Caldisphaera lagunensis DSM
15908 aldehyde : ferredoxin oxidoreductase Caldisphaera lagunensis DSM
15908 aldehyde : ferredoxin oxidoreductase Caldisphaera lagunensis DSM
15908 aldehyde : ferredoxin oxidoreductase Caldisphaera lagunensis DSM
15908 aldehyde : ferredoxin oxidoreductase Caldivirga maquilingensis IC - 167 aldehyde : ferredoxin oxidoreductase Caldivirga maquilingensis IC - 167 aldehyde : ferredoxin oxidoreductase Caldivirga maquilingensis IC - 167 aldehyde : ferredoxin oxidoreductase Caldivirga maquilingensis IC - 167 aldehyde : ferredoxin oxidoreductase Caldivirga maquilingensis IC - 167 aldehyde : ferredoxin oxidoreductase Caldivirga maquilingensis IC - 167 aldehyde : ferredoxin oxidoreductase Caldivirga maquilingensis IC - 167 aldehyde : ferredoxin oxidoreductase Candidatus Caldiarchaeum
sub terraneum aldehyde : ferredoxin oxidoreductase Candidatus Korarchaeum
cryptofilum OPF8 aldehyde : ferredoxin oxidoreductase Candidatus Korarchaeum
cryptofilum OPF8 aldehyde : ferredoxin oxidoreductase Candidatus Korarchaeum
cryptofilum OPF8 aldehyde : ferredoxin oxidoreductase Candidatus Korarchaeum
cryptofilum OPF8 aldehyde : ferredoxin oxidoreductase Candidatus Korarchaeum
cryptofilum OPF8 aldehyde : ferredoxin oxidoreductase Candidatus Korarchaeum
cryptofilum OPF8 aldehyde : ferredoxin oxidoreductase Chloroflexus aurantiacus 11041 aldehyde : ferredoxin oxidoreductase Clostridium acetobutylicum ATCC
824 aldehyde : ferredoxin oxidoreductase Clostridium botulinum A sfr . ATCC
3502 aldehyde : ferredoxin oxidoreductase Clostridium botulinum A str . Hall aldehyde : ferredoxin oxidoreductase Desulfovibrio vulgaris sfr .
Hildenborough aldehyde : ferredoxin oxidoreductase Desulfovibrio vulgaris sfr .
Hildenborough aldehyde : ferredoxin oxidoreductase Desulfurococcus fermentans DSM
16532 aldehyde : ferredoxin oxidoreductase Desulfurococcus fermentans DSM
16532 aldehyde : ferredoxin oxidoreductase Desulfurococcus fermentans DSM
16532
NC _ 019791 . 1 14212561
NC _ 009954 . 1 NC _ 009954 . 1 NC 009954 . 1 NC 009954 . 1 NC 009954 . 1 NC _ 009954 . 1 NC _ 009954 . 1 NC _ 022786 . 1
5710116 5710117 5709088 5708891 5710478 5710457 5709696
17602865
NC _ 010482 . 1 6094361
NC _ 010482 . 1 6094198
NC _ 010482 . 1 6093546
NC _ 010482 . 1 6093319
NC _ 010482 . 1 6094057
NC _ 010482 . 1 6093563
NC _ 010175 . 1 NC _ 003030 . 1
5828639 1118201
NC _ 009495 . 1 5187636
NC _ 009698 . 1 NC _ 002937 . 3
5400593 2796664
NC _ 002937 . 3 2795337
NC _ 018001 . 1 13061477
NC _ 018001 . 1 13061068
NC _ 018001 . 1 13062247

US 9 , 957 , 531 B1 13 14
- continued Description Microrganism Accession GeneID
NC _ 011766 . 1 7171099 NC _ 011766 . 1 7171759
NC _ 011766 . 1 7170725
NC _ 014961 . 1 10152801
NC _ 013849 . 1 NC _ 013849 . 1 NC 013849 . 1 NC 013849 . 1 NC _ 013849 . 1 NC _ 013849 . 1 NC _ 013849 . 1 NC _ 017461 . 1 NC _ 017461 . 1 NC 017461 . 1 NC _ 017461 . 1 NC _ 002939 . 5 NC _ 002939 . 5 NC 014297 . 1 NC 014297 . 1 NC 014297 . 1 NC 014297 . 1 NC _ 015948 . 1 NC _ 015948 . 1 NC _ 015948 . 1 NC _ 023013 . 1 NC _ 023013 . 1 NC _ 023013 . 1 NC _ 006396 . 1
8778536 8779007 8778940 8779639 8778820 8778745 8779874
12449263 12449994 12449294 12449682 2685730 2687039 9418623 9418760 9420819 9418748
11051410 11050783 11051433 23805333 23805138 23804665 3127969
NC _ 006396 . 1 3129232
NC _ 017941 . 2 13028168
NC _ 017941 . 2 13028399
aldehyde : ferredoxin oxidoreductase Desulfurococcus kamchatkensis 1221n
aldehyde : ferredoxin oxidoreductase Desulfurococcus kamchatkensis 1221n
aldehyde : ferredoxin oxidoreductase Desulfurococcus kamchatkensis 1221n
aldehyde : ferredoxin oxidoreductase Desulfurococcus mucosus DSM 2162
aldehyde : ferredoxin oxidoreductase Ferroglobus placidus DSM 10642 aldehyde : ferredoxin oxidoreductase Ferroglobus placidus DSM 10642 aldehyde : ferredoxin oxidoreductase Ferroglobus placidus DSM 10642 aldehyde : ferredoxin oxidoreductase Ferroglobus placidus DSM 10642 aldehyde : ferredoxin oxidoreductase Ferroglobus placidus DSM 10642 aldehyde : ferredoxin oxidoreductase Ferroglobus placidus DSM 10642 aldehyde : ferredoxin oxidoreductase Ferroglobus placidus DSM 10642 aldehyde : ferredoxin oxidoreductase Fervidicoccus fontis Kam940 aldehyde : ferredoxin oxidoreductase Fervidicoccus fontis Kam940 aldehyde : ferredoxin oxidoreductase Fervidicoccus fontis Kam94 aldehyde : ferredoxin oxidoreductase Fervidicoccus fontis Kam940 aldehyde : ferredoxin oxidoreductase Geobacter sulfurreducens PCA aldehyde : ferredoxin oxidoreductase Geobacter sulfurreducens PCA aldehyde : ferredoxin oxidoreductase Halalkalicoccus jeotgali B3 aldehyde : ferredoxin oxidoreductase Halalkalicoccus jeotgali B3 aldehyde : ferredoxin oxidoreductase Halalkalicoccus jeotgali B3 aldehyde : ferredoxin oxidoreductase Halalkalicoccus jeotgali B3 aldehyde : ferredoxin oxidoreductase Haloarcula hispanica ATCC 33960 aldehyde : ferredoxin oxidoreductase Haloarcula hispanica ATCC 33960 aldehyde : ferredoxin oxidoreductase Haloarcula hispanica ATCC 33960 aldehyde : ferredoxin oxidoreductase Haloarcula hispanica N601 aldehyde : ferredoxin oxidoreductase Haloarcula hispanica N601 aldehyde : ferredoxin oxidoreductase Haloarcula hispanica N601 aldehyde : ferredoxin oxidoreductase Haloarcula marismortui ATCC
43049 aldehyde : ferredoxin oxidoreductase Haloarcula marismortui ATCC
43049 aldehyde : ferredoxin oxidoreductase Haloferax mediterranei ATCC
33500 aldehyde : ferredoxin oxidoreductase Haloferax mediterranei ATCC
33500 aldehyde : ferredoxin oxidoreductase Haloferax volcanii DS2 aldehyde : ferredoxin oxidoreductase Haloferax volcanii DS2 aldehyde : ferredoxin oxidoreductase Haloferax volcanii DS2 aldehyde : ferredoxin oxidoreductase Halogeomefricum borinquense DSM
11551 aldehyde : ferredoxin oxidoreductase Halogeomefricum borinquense DSM
11551 aldehyde : ferredoxin oxidoreductase Halogeomefricum borinquense DSM
11551 aldehyde : ferredoxin oxidoreductase halophilic archaeon DL31 aldehyde : ferredoxin oxidoreductase halophilic archaeon DL31 aldehyde : ferredoxin oxidoreductase halophilic archaeon DL31 aldehyde : ferredoxin oxidoreductase halophilic archaeon DL31 aldehyde : ferredoxin oxidoreductase halophilic archaeon DL31 aldehyde : ferredoxin oxidoreductase halophilic archaeon DL31 aldehyde : ferredoxin oxidoreductase Halopiger xanaduensis SH - 6 aldehyde : ferredoxin oxidoreductase Halopiger xanaduensis SH - 6 aldehyde : ferredoxin oxidoreductase Halopiger xanaduensis SH - 6 aldehyde : ferredoxin oxidoreductase Halopiger xanaduensis SH - 6 aldehyde : ferredoxin oxidoreductase Halorubrum lacusprofundi ATCC
49239 aldehyde : ferredoxin oxidoreductase Halorubrum lacusprofundi ATCC
49239 aldehyde : ferredoxin oxidoreductase Halorubrum lacusprofundi ATCC
49239 aldehyde : ferredoxin oxidoreductase Haloterrigena turkmenica DSM
5511 aldehyde : ferredoxin oxidoreductase Haloterrigena turkmenica DSM
5511 aldehyde : ferredoxin oxidoreductase Haloterrigena turkmenica DSM
5511 aldehyde : ferredoxin oxidoreductase Haloterrigena turkmenica DSM
5511 aldehyde : ferredoxin oxidoreductase Haloterrigena turkmenica DSM
5511 aldehyde : ferredoxin oxidoreductase Haloterrigena turkmenica DSM
5511
NC _ 013964 . 1 NC _ 013964 . 1 NC 013967 . 1 NC _ 014735 . 1
8919329 8919033 8926544 9989054
NC _ 014729 . 1 9994424
NC _ 014729 . 1 9992444
NC _ 015954 . 1 NC _ 015954 . 1 NC 015954 . 1 NC _ 015954 . 1 NC _ 015954 . 1 NC _ 015954 . 1 NC _ 015666 . 1 NC _ 015658 . 1 NC 015666 . 1 NC _ 015666 . 1 NC _ 0 12029 . 1
11095016 11095541 11094595 11096497 11094563 11095602 10799161 10795465 10798686 10796679 7400122
NC _ 0 12029 . 1 7400291
NC _ 012029 . 1 7400689
NC _ 013744 . 1 8744461
NC _ 013744 . 1 8744695
NC _ 013743 . 1 8740954
NC _ 013745 . 1 8745418
NC _ 013743 . 1 8742968
NC _ 013743 . 1 8741246
US 9 , 957 , 531 B1 15 16
- continued Description Microrganism Accession GeneID
8741269 8745313
4781896 4782266 4782804 4781774 5562477 5562774 9716798 1452083
5142690 5143773 11972399 11971349 8680711 8680676 4795790 4847673 9347460
9347022
13845119 14408029
aldehyde : ferredoxin oxidoreductase Haloterrigena turkmenica DSM NC _ 013743 . 1 5511
aldehyde : ferredoxin oxidoreductase Haloterrigena turkmenica DSM NC _ 013745 . 1 5511
aldehyde : ferredoxin oxidoreductase Hyperthermus butylicus DSM 5456 NC _ 008818 . 1 aldehyde : ferredoxin oxidoreductase Hyperthermus butylicus DSM 5456 NC _ 008818 . 1 aldehyde : ferredoxin oxidoreductase Hyperthermus butylicus DSM 5456 NC _ 008818 . 1 aldehyde : ferredoxin oxidoreductase Hyperthermus butylicus DSM 5456 NC _ 008818 . 1 aldehyde : ferredoxin oxidoreductase Ignicoccus hospitalis KIN4 / I NC _ 009776 . 1 aldehyde : ferredoxin oxidoreductase Ignicoccus hospitalis KIN4 / I NC _ 009776 . 1 aldehyde : ferredoxin oxidoreductase Ignisphaera aggregans DSM 17230 NC _ 014471 . 1 aldehyde : ferredoxin oxidoreductase Methanocaldococcus jannaschii NC _ 000909 . 1
DSM 2661 aldehyde : ferredoxin oxidoreductase Methanocella arvoryzae MRE50 NC _ 009464 . 1 aldehyde : ferredoxin oxidoreductase Methanocella arvoryzae MRE50 NC 009464 . 1 aldehyde : ferredoxin oxidoreductase Methanocella conradii HZ254 NC _ 017034 . 1 aldehyde : ferredoxin oxidoreductase Methanocella conradii HZ254 NC _ 017034 . 1 aldehyde : ferredoxin oxidoreductase Methanocella paludicola SANAE NC _ 013665 . 1 aldehyde : ferredoxin oxidoreductase Methanocella paludicola SANAE NC _ 013665 . 1 aldehyde : ferredoxin oxidoreductase Methanocorpusculum labreanum Z NC _ 008942 . 1 aldehyde : ferredoxin oxidoreductase Methanoculleus marisnigri JR1 NC _ 009051 . 1 aldehyde : ferredoxin oxidoreductase Methanohalobium evestigatum Z NC _ 014253 . 1
7303 aldehyde : ferredoxin oxidoreductase Methanohalobium evestigatum Z NC _ 014253 . 1
7303 aldehyde : ferredoxin oxidoreductase Methanolobus psychrophilus R15 NC _ 018876 . 1 aldehyde : ferredoxin oxidoreductase Methanomethylovorans hollandica NC _ 019977 . 1
DSM 15978 aldehyde : ferredoxin oxidoreductase Methanosaeta harundinacea 6AC NC _ 017527 . 1 aldehyde : ferredoxin oxidoreductase Methanosaeta thermophila PT NC _ 008553 . 1 aldehyde : ferredoxin oxidoreductase Methanosalsum zhilinae DSM 4017 NC _ 015676 . 1 aldehyde : ferredoxin oxidoreductase Methanosarcina acetivorans C2A NC _ 003552 . 1 aldehyde : ferredoxin oxidoreductase Methanosarcina acetivorans C2A NC _ 003552 . 1 aldehyde : ferredoxin oxidoreductase Methanosarcina acetivorans C2A NC _ 003552 . 1 aldehyde : ferredoxin oxidoreductase Methanosarcina barkeri str . Fusaro NC _ 007355 . 1 aldehyde : ferredoxin oxidoreductase Methanosarcina mazei Gol NC 003901 . 1 aldehyde : ferredoxin oxidoreductase Methanosarcina mazei Gol NC 003901 . 1 aldehyde : ferredoxin oxidoreductase Methanosarcina mazei Gol NC _ 003901 . 1 aldehyde : ferredoxin oxidoreductase Methanosarcina mazei Tuco NC _ 020389 . 1 aldehyde : ferredoxin oxidoreductase Methanosarcina mazei Tuc01 NC _ 020389 . 1 aldehyde : ferredoxin oxidoreductase Methanosarcina mazei Tuc01 NC _ 020389 . 1 aldehyde : ferredoxin oxidoreductase Methanosphaerula palustris E1 - 9c NC _ 011832 . 1 aldehyde : ferredoxin oxidoreductase Methanospirillum hungatei JF - 1 NC _ 007796 . 1 aldehyde : ferredoxin oxidoreductase Methylomicrobium alcaliphilum 20Z NC _ 016112 . 1 ald ferredoxin oxidoreductase Moorella thermoacetica ATCC NC _ 007644 . 1
39073 aldehyde : ferredoxin oxidoreductase Moorella thermoacetica ATCC 7644 . 1
39073 aldehyde : ferredoxin oxidoreductase Moorella thermoacetica ATCC NC _ 007644 . 1
39073 aldehyde : ferredoxin oxidoreductase Natrialba magadii ATCC 43099 NC _ 013922 . 1 aldehyde : ferredoxin oxidoreductase Natrialba magadii ATCC 43099 NC _ 013922 . 1 aldehyde : ferredoxin oxidoreductase Natrialba magadii ATCC 43099 NC _ 013923 . 1 aldehyde : ferredoxin oxidoreductase Natrialba magadii ATCC 43099 NC _ 013922 . 1 aldehyde : ferredoxin oxidoreductase Natrinema pelli rubrum DSM 15624 NC _ 019962 . 1 aldehyde : ferredoxin oxidoreductase Natrinema pelli rubrum DSM 15624 NC _ 019962 . 1 aldehyde : ferredoxin oxidoreductase Natrinema pelli rubrum DSM 15624 NC _ 019962 . 1 aldehyde : ferredoxin oxidoreductase Natrinema sp . J7 - 2 NC 018224 . 1 aldehyde : ferredoxin oxidoreductase Natronobacterium gregoryi SP2 NC _ 019792 . 1 aldehyde : ferredoxin oxidoreductase Natronobacterium gregoryi SP2 NC _ 019792 . 1 aldehyde : ferredoxin oxidoreductase Natronobacterium gregoryi SP2 NC _ 019792 . 1 aldehyde : ferredoxin oxidoreductase Natronobacterium gregoryi SP2 NC _ 019792 . 1 aldehyde : ferredoxin oxidoreductase Natronobacterium gregoryi SP2 NC _ 019792 . 1 aldehyde : ferredoxin oxidoreductase Natronobacterium gregoryi SP2 NC _ 019792 . 1 aldehyde : ferredoxin oxidoreductase Natronococcus occultus SP4 NC _ 019974 . 1 aldehyde : ferredoxin oxidoreductase Natronococcus occultus SP4 NC _ 019974 . 1 aldehyde : ferredoxin oxidoreductase Natronococcus occultus SP NC _ 019974 . 1 aldehyde : ferredoxin oxidoreductase Natronococcus occultus SP4 NC _ 019974 . 1 aldehyde : ferredoxin oxidoreductase Natronomonas moolapensis 8 . 8 . 11 NC _ 020388 . 1 aldehyde : ferredoxin oxidoreductase Natronomonas moolapensis 8 . 8 . 11 NC _ 020388 . 1 aldehyde : ferredoxin oxidoreductase Natronomonas moolapensis 8 . 8 . 11 NC _ 020388 . 1 aldehyde : ferredoxin oxidoreductase Natronomonas pharaonis DSM 2160 NC _ 007427 . 1 aldehyde : ferredoxin oxidoreductase Natronomonas pharaonis DSM 2160 NC _ 007426 . 1 aldehyde : ferredoxin oxidoreductase Natronomonas pharaonis DSM 2160 NC _ 007426 . 1 aldehyde : ferredoxin oxidoreductase Natronomonas pharaonis DSM 2160 NC _ 007426 . 1 aldehyde : ferredoxin oxidoreductase Pyrobaculum aerophilum str . IM2 NC _ 003364 . 1 aldehyde : ferredoxin oxidoreductase Pyrobaculum aerophilum str . IM2 NC _ 003364 . 1
12511443 4462364 10822365 1475882 1474856 1473602 3625763 1479263 1481668 1480987 14656065 14656771 14654304 7271108 3924565 11361147 3831332
3830998
3831866
8824961 8823392 8826737 8825516 14335278 14333050 14333754 13349954 14210296 14207133 14209682 14207576 14206941 14206532 14403316 14405255 14403781 14402014 14651997 14652892 14651999 3694680 3702508 3702507 3702509 1464236 1464102

US 9 , 957 , 531 B1 17
- continued Description Microrganism Accession GeneID
1465126 1465445 5055904
5055700
5054881
5054644
5054547
4910224
4908822
4909927
4910099
aldehyde : ferredoxin oxidoreductase Pyrobaculum aerophilum str . IM2 NC _ 003364 . 1 aldehyde : ferredoxin oxidoreductase Pyrobaculum aerophilum str . IM2 NC _ 003364 . 1 aldehyde : ferredoxin oxidoreductase Pyrobaculum arsenaticum DSM NC _ 009376 . 1
13514 aldehyde : ferredoxin oxidoreductase Pyrobaculum arsenaticum DSM NC _ 009376 . 1
13514 aldehyde : ferredoxin oxidoreductase Pyrobaculum arsenaticum DSM NC _ 009376 . 1
13514 aldehyde : ferredoxin oxidoreductase Pyrobaculum arsenaticum DSM NC _ 009376 . 1
13514 aldehyde : ferredoxin oxidoreductase Pyrobaculum arsenaticum DSM 9376 . 1
13514 aldehyde : ferredoxin oxidoreductase Pyrobaculum calidifontis JCM NC _ 009073 . 1
11548 aldehyde : ferredoxin oxidoreductase Pyrobaculum calidifontis JCM NC _ 009073 . 1
11548 aldehyde : ferredoxin oxidoreductase Pyrobaculum calidifontis JCM NC _ 009073 . 1
11548 aldehyde : ferredoxin oxidoreductase Pyrobaculum calidifontis JCM NC _ 009073 . 1
11548 aldehyde : ferredoxin oxidoreductase Pyrobaculum islandicum DSM 4184 NC _ 008701 . 1 aldehyde : ferredoxin oxidoreductase Pyrobaculum islandicum DSM 4184 NC _ 008701 . 1 aldehyde : ferredoxin oxidoreductase Pyrobaculum islandicum DSM 4184 NC _ 008701 . 1 aldehyde : ferredoxin oxidoreductase Pyrobaculum neutrophilum V24Sta NC _ 010525 . 1 aldehyde : ferredoxin oxidoreductase Pyrobaculum neutrophilum V24Sta NC _ 010525 . 1 aldehyde : ferredoxin oxidoreductase Pyrobaculum neutrophilum V24Sta NC _ 010525 . 1 aldehyde : ferredoxin oxidoreductase Pyrobaculum oguniense TE7 NC _ 016885 . 1 aldehyde : ferredoxin oxidoreductase Pyrobaculum oguniense TE7 NC _ 016885 . 1 aldehyde : ferredoxin oxidoreductase Pyrobaculum oguniense TE7 NC _ 016885 . 1 aldehyde : ferredoxin oxidoreductase Pyrobaculum oguniense TET NC _ 016885 . 1 aldehyde : ferredoxin oxidoreductase Pyrobaculum oguniense TE7 NC _ 016885 . 1 aldehyde : ferredoxin oxidoreductase Pyrobaculum sp . 1860 NC _ 016645 . 1 aldehyde : ferredoxin oxidoreductase Pyrobaculum sp . 1860 NC _ 016645 . 1 aldehyde : ferredoxin oxidoreductase Pyrobaculum sp . 1860 NC 016645 . 1 aldehyde : ferredoxin oxidoreductase Pyrococcus abyssi GE5 NC _ 000868 . 1 aldehyde : ferredoxin oxidoreductase Pyrococcus abyssi GE5 NC _ 000868 . 1 aldehyde : ferredoxin oxidoreductase Pyrococcus abyssi GE5 NC 000868 . 1 aldehyde : ferredoxin oxidoreductase Pyrococcus abyssi GE5 NC _ 000868 . 1 aldehyde : ferredoxin oxidoreductase Pyrococcus furiosus NC _ 018092 . 1 aldehyde : ferredoxin oxidoreductase Pyrococcus furiosus COM1 NC _ 018092 . 1 aldehyde : ferredoxin oxidoreductase Pyrococcus furiosus COM1 NC _ 018092 . 1 aldehyde : ferredoxin oxidoreductase Pyrococcus furiosus COM1 NC 018092 . 1 aldehyde : ferredoxin oxidoreductase Pyrococcus furiosus DSM 3638 NC _ 003413 . 1 aldehyde : ferredoxin oxidoreductase Pyrococcus furiosus DSM 3638 NC _ 003413 . 1 aldehyde : ferredoxin oxidoreductase Pyrococcus furiosus DSM 3638 NC 003413 . 1 aldehyde : ferredoxin oxidoreductase Pyrococcus horikoshii OT3 NC _ 000961 . 1 aldehyde : ferredoxin oxidoreductase Pyrococcus horikoshii OT3 NC _ 000961 . 1 aldehyde : ferredoxin oxidoreductase Pyrococcus horikoshii OT3 NC _ 000961 . 1 aldehyde : ferredoxin oxidoreductase Pyrococcus horikoshii OT3 NC _ 000961 . 1 aldehyde : ferredoxin oxidoreductase Pyrococcus sp . NA2 NC 015474 . 1 aldehyde : ferredoxin oxidoreductase Pyrococcus sp . NA2 NC 015474 . 1 aldehyde : ferredoxin oxidoreductase Pyrococcus sp . NA2 NC 015474 . 1 aldehyde : ferredoxin oxidoreductase Pyrococcus sp . STO4 NC 017946 . 1 aldehyde : ferredoxin oxidoreductase Pyrococcus sp . STO4 NC _ 017946 . 1 aldehyde : ferredoxin oxidoreductase Pyrococcus sp . STO4 NC _ 017946 . 1 aldehyde : ferredoxin oxidoreductase Pyrococcus yayanosii CH1 NC _ 015680 . 1 aldehyde : ferredoxin oxidoreductase Pyrococcus yayanosii CH1 NC _ 015680 . 1 aldehyde : ferredoxin oxidoreductase Pyrococcus yayanosii CH1 NC 015680 . 1 aldehyde : ferredoxin oxidoreductase Pyrolobus fumarii JA NC _ 015931 . 1 aldehyde : ferredoxin oxidoreductase Pyrolobus fumarii JA NC _ 015931 . 1 aldehyde : ferredoxin oxidoreductase Pyrolobus fumarii JA NC 015931 . 1 aldehyde : ferredoxin oxidoreductase Rhodospirillum rubrum ATCC NC _ 007643 . 1
11170 aldehyde : ferredoxin oxidoreductase Staphylothermus hellenicus DSM NC _ 014205 . 1
12710 aldehyde : ferredoxin oxidoreductase Staphylothermus hellenicus DSM NC _ 014205 . 1
12710 aldehyde : ferredoxin oxidoreductase Staphylothermus hellenicus DSM NC _ 014205 . 1
12710 aldehyde : ferredoxin oxidoreductase Staphylothermus hellenicus DSM NC _ 014205 . 1
12710 aldehyde : ferredoxin oxidoreductase Staphylothermus marinus F1 NC _ 009033 . 1 aldehyde : ferredoxin oxidoreductase Staphylothermus marinus F1 NC _ 009033 . 1 aldehyde : ferredoxin oxidoreductase Thermanaerovibrio NC _ 013522 . 1
acidaminovorans DSM 6589 aldehyde : ferredoxin oxidoreductase Thermanaerovibrio NC _ 013522 . 1
acidaminovorans DSM 6589
4617364 4616724 4617494 6165427 6164958 6164976 11853778 11854024 11856490 11856176 11854908 11594868 11596631 11594049 1496313 1495669 1496580 1495287
13302148 13301806 13301219 13300785 1468181 1469073 1469843 1443218 1443341 1443932 1443598
10555029 10554020 10555341 13022107 13022436 13021314 10837518 10837112 10837264 11138144 11138776 11139127 3833668
9234557
9233414
9234134
9234110
4907444 4907343 8630284
8630027

US 9 , 957 , 531 B1 19 20
- continued Description Microrganism Accession GeneID
8630623
11082596
10041639 10041106 10042460 13037745 13038896 13037242 7988317 7987451 3233851 3233735 16550741 16548761 16550885 7018383 7016739 7017051 7017476 8096638 8096005 8096629 8095463 8096131 8096636 11015504 11015249 11015571 7419050 7418514 7420292 6941429
6943174
6941905
aldehyde : ferredoxin oxidoreductase Thermanaerovibrio NC _ 013522 . 1 acidaminovorans DSM 6589
aldehyde : ferredoxin oxidoreductase Thermoanaerobacter wiegelii NC _ 015958 . 1 Rt8 . B1
aldehyde : ferredoxin oxidoreductase Thermococcus barophilus MP NC _ 014804 . 1 aldehyde : ferredoxin oxidoreductase Thermococcus barophilus MP NC _ 014804 . 1 aldehyde : ferredoxin oxidoreductase Thermococcus barophilus MP NC _ 014804 . 1 aldehyde : ferredoxin oxidoreductase Thermococcus cleftensis NC 018015 . 1 aldehyde : ferredoxin oxidoreductase Thermococcus cleftensis NC _ 018015 . 1 aldehyde : ferredoxin oxidoreductase Thermococcus cleftensis NC _ 018015 . 1 aldehyde : ferredoxin oxidoreductase Thermococcus gammatolerans EJ3 NC _ 012804 . 1 aldehyde : ferredoxin oxidoreductase Thermococcus gammatolerans EJ3 NC _ 012804 . 1 aldehyde : ferredoxin oxidoreductase Thermococcus kodakarensis KODI NC _ 006624 . 1 aldehyde : ferredoxin oxidoreductase Thermococcus kodakarensis KODI NC _ 006624 . 1 aldehyde : ferredoxin oxidoreductase Thermococcus litoralis DSM 5473 NC _ 022084 . 1 aldehyde : ferredoxin oxidoreductase Thermococcus litoralis DSM 5473 NC _ 022084 . 1 aldehyde : ferredoxin oxidoreductase Thermococcus litoralis DSM 5473 NC _ 022084 . 1 aldehyde : ferredoxin oxidoreductase Thermococcus onnurineus NAI NC _ 011529 . 1 aldehyde : ferredoxin oxidoreductase Thermococcus onnurineus NA1 NC 011529 . 1 aldehyde : ferredoxin oxidoreductase Thermococcus onnurineus NA1 NC _ 011529 . 1 aldehyde : ferredoxin oxidoreductase Thermococcus onnurineus NA1 NC 011529 . 1 aldehyde : ferredoxin oxidoreductase Thermococcus sibiricus MM 739 NC 012883 . 1 aldehyde : ferredoxin oxidoreductase Thermococcus sibiricus MM 739 NC 012883 . 1 aldehyde : ferredoxin oxidoreductase Thermococcus sibiricus MM 739 NC 012883 . 1 aldehyde : ferredoxin oxidoreductase Thermococcus sibiricus MM 739 NC 012883 . 1 aldehyde : ferredoxin oxidoreductase Thermococcus sibiricus MM 739 NC _ 012883 . 1 aldehyde : ferredoxin oxidoreductase Thermococcus sibiricus MM 739 NC 012883 . 1 aldehyde : ferredoxin oxidoreductase Thermococcus sp . 4557 NC 015865 . 1 aldehyde : ferredoxin oxidoreductase Thermococcus sp . 4557 NC _ 015865 . 1 aldehyde : ferredoxin oxidoreductase Thermococcus sp . 4557 NC _ 015865 . 1 aldehyde : ferredoxin oxidoreductase Thermococcus sp . AM4 NC _ 016051 . 1 aldehyde : ferredoxin oxidoreductase Thermococcus sp . AM4 NC _ 016051 . 1 aldehyde : ferredoxin oxidoreductase Thermococcus sp . AM4 NC 016051 . 1 aldehyde : ferredoxin oxidoreductase Thermodesulfovibrio yellowstonii NC _ 011296 . 1
DSM 11347 aldehyde : ferredoxin oxidoreductase Thermodesulfovibrio yellowstonii NC _ 011296 . 1
DSM 11347 aldehyde : ferredoxin oxidoreductase Thermodesulfovibrio yellowstonii NC _ 011296 . 1
DSM 11347 aldehyde : ferredoxin oxidoreductase Thermofilum pendens Hrk 5 NC _ 008698 . 1 aldehyde : ferredoxin oxidoreductase Thermofilum pendens Hrk 5 NC _ 008698 . 1 aldehyde : ferredoxin oxidoreductase Thermofilum pendens Hrk 5 NC _ 008698 . 1 aldehyde : ferredoxin oxidoreductase Thermofilum pendens Hrk 5 NC _ 008698 . 1 aldehyde : ferredoxin oxidoreductase Thermofilum sp . 1910b NC 022093 . 1 aldehyde : ferredoxin oxidoreductase Thermofilum sp . 1910b NC 022093 . 1 aldehyde : ferredoxin oxidoreductase Thermofilum sp . 1910b NC 022093 . 1 aldehyde : ferredoxin oxidoreductase Thermofilum sp . 1910b NC 022093 . 1 aldehyde : ferredoxin oxidoreductase Thermogladius cellulolyticus 1633 NC 017954 . 1 aldehyde : ferredoxin oxidoreductase Thermoplasma acidophilum DSM NC _ 002578 . 1
1728 aldehyde : ferredoxin oxidoreductase Thermoplasma acidophilum DSM NC _ 002578 . 1
1728 aldehyde : ferredoxin oxidoreductase Thermoplasma vokanium GSS1 NC _ 002689 . 2 aldehyde : ferredoxin oxidoreductase Thermoplasma vokanium GSS1 NC _ 002689 . 2 aldehyde : ferredoxin oxidoreductase Thermoproteus tenax Kra 1 NC _ 016070 . 1 aldehyde : ferredoxin oxidoreductase Thermoproteus tenax Kra 1 NC 016070 . 1 aldehyde : ferredoxin oxidoreductase Thermoproteus tenax Kra 1 NC _ 016070 . 1 aldehyde : ferredoxin oxidoreductase Thermoproteus tenax Kra 1 NC 016070 . 1 aldehyde : ferredoxin oxidoreductase Thermoproteus uzoniensis 768 - 20 NC _ 015315 . 1 aldehyde : ferredoxin oxidoreductase Thermoproteus uzoniensis 768 - 20 NC _ 015315 . 1 aldehyde : ferredoxin oxidoreductase Thermoproteus uzoniensis 768 - 20 NC 015315 . 1 aldehyde : ferredoxin oxidoreductase Thermosphaera aggregans DSM NC _ 014160 . 1
11486 aldehyde : ferredoxin oxidoreductase Thermosphaera aggregans DSM NC _ 014160 . 1
11486 aldehyde : ferredoxin oxidoreductase Thermus thermophilus HB8 NC _ 006461 . 1 aldehyde : ferredoxin oxidoreductase Thermus thermophilus HB8 NC _ 006461 . 1 aldehyde : ferredoxin oxidoreductase Vulcanisaeta disfributa DSM 14429 NC _ 014537 . 1 aldehyde : ferredoxin oxidoreductase Vulcanisaeta disfributa DSM 14429NC _ 014537 . 1 aldehyde : ferredoxin oxidoreductase Vulcanisaeta disfributa DSM 14429 NC 014537 . 1 aldehyde : ferredoxin oxidoreductase Vulcanisaeta disfributa DSM 14429 NC _ 014537 . 1 aldehyde : ferredoxin oxidoreductase Vulcanisaeta disfributa DSM 14429 NC _ 014537 . 1 aldehyde : ferredoxin oxidoreductase Vulcanisaeta moutnovskia 768 - 28 NC _ 015151 . 1 aldehyde : ferredoxin oxidoreductase Vulcanisaeta moutnovskia 768 - 28 NC _ 015151 . 1 aldehyde : ferredoxin oxidoreductase Vulcanisaeta moutnovskia 768 - 28 NC _ 015151 . 1 aldehyde : ferredoxin oxidoreductase Vulcanisaeta moutnovskia 768 - 28 NC _ 015151 . 1 aldehyde : ferredoxin oxidoreductase Vulcanisaeta moutnovskia 768 - 28 NC _ 015151 . 1
4602054 4601386 4600878 4600730
16572780 16572926 16573009 16574342 13012904 1456355
1456646
1441901 1441379 11262174 11262275 11262652 11262926 10361668 10361250 10360972 9165115
9165462
3168554 3168612 9753145 9750947 9750989 9753486 9751414 10288238 10288894 10288574 10288827 10288607

US 9 , 957 , 531 B1 21 22
- continued Description Microrganism Accession GeneID
aldehyde : ferredoxin oxidoreductase Vulcanisaeta moutnovskia 768 - 28 aldehyde : ferredoxin oxidoreductase Vulcanisaeta moutnovskia 768 - 28
NC _ 015151 . 1 NC _ 015151 . 1
10288523 10288815
AOR catalyzes the reaction of an acid and reduced Step 19 shows the conversion of 3 - hydroxybutyryl - CoA ferredoxin to form an aldehyde and oxidized ferredoxin . In 10 to 2 - hydroxyisobutyryl - CoA . This step may be catalyzed by acetogens , this reaction can be coupled to oxidation CO ( via 2 - hydroxyisobutyryl - CoA mutase ( EC 5 . 4 . 99 . - ) . The 2 - hy CO dehydrogenase , EC 1 . 2 . 7 . 4 ) or hydrogen ( via ferre droxyisobutyryl - CoA mutase may be , for example , HcmAB doxin - dependent hydrogenase , EC 1 . 12 . 7 . 2 or 1 . 12 . 1 . 4 ) that from Aquincola tertiaricarbonis ( AFK77668 . 1 , large sub both yield reduced ferredoxin ( Köpke , Curr Opin Biotech unit ; AFK77665 . 1 , small subunit ) ( SEQ ID NOs : 81 and 82 , nol 22 : 320 - 325 , 2011 ; Köpke , PNAS USA , 107 : 13087 - 15 respectively ) or HemAB from Kyrpidia tusciae 13092 , 2010 ) . Clostridium autoethanogenum , Clostridium ( WP _ 013074530 . 1 , large subunit ; WP _ 013074531 . 1 , small ljungdahlii , and Clostridium ragsdalei have native activity subunit ) ( SEQ ID NOs : 83 and 84 , respectively ) . Chaperone for this step . However , overexpression of endogenous AOR MeaB ( AFK77667 . 1 , Aquincola tertiaricarbonis ; or introduction of an exogenous AOR in Clostridium WP _ 013074529 . 1 , Kyrpidia tusciae ) ( SEQ ID NOs : 85 and autoethanogenum , Clostridium ljungdahlii , or Clostridium 20 86 , respectively ) has been described to improve activity of ragsdalei may be desirable to enhance product yields . Alter HcmAB by reactivating HemAB , although MeaB is not natively , exogenous AOR may be introduced into a micro required for HcmAB function ( Yaneva , J Biol Chem , 287 : organism that does not natively comprise AOR , e . g . , E . coli . 15502 - 15511 , 2012 ) . Clostridium autoethanogenum , In particular , the co - expression of Ptb - Buk and AOR ( and , Clostridium ljungdahlii , and Clostridium ragsdalei do not optionally . Adh ) may enable such a microorganism to pro - 25 have known native activity for this step . Escherichia coli
does not have known native activity for this step . duce new non - native products . Step 20 shows the conversion of 2 - hydroxyisobutyryl Step 17 shows the conversion of 3 - hydroxybutyrylalde hyde to 1 , 3 - butanediol . This step may be catalyzed by COA to 2 - hydroxyisobutyrate . This step may be catalyzed by
phosphate butyryltransferase ( EC 2 . 3 . 1 . 19 ) + butyrate kinase alcohol dehydrogenase ( EC 1 . 1 . 1 . 1 . or 1 . 1 . 1 . 2 . ) . Alcohol 3 O ! 30 ( EC 2 . 7 . 2 . 7 ) . Exemplary sources for phosphate butyryltrans dehydrogenase can convert an aldehyde and NAD ( P ) H to an ferase and butyrate kinase are described elsewhere in this alcohol and NAD ( P ) . The alcohol dehydrogenase may be , application . Clostridium autoethanogenum , Clostridium for example , Adh from Clostridium autoethanogenum ljungdahlii , and Clostridium ragsdalei do not have known ( AGY76060 . 1 ) ( SEQ ID NO : 67 ) , Clostridium ljungdahlii native activity for this step . Escherichia coli does not have ( ADK17019 . 1 ) ( SEQ ID NO : 68 ) , or Clostridium ragsdalei , 35 known native activity for this step . Bdh? from Clostridium acetobutylicum ( NP _ 349891 . 1 ) Step 21 shows the conversion of acetyl - CoA to succinyl ( SEO ID NO : 69 ) , Bdh from Clostridium beijerinckii COA . This step encompasses a number of enzymatic reac ( WP _ 041897187 . 1 ) ( SEQ ID NO : 70 ) , Bdh1 from tions involved in the reductive TCA pathway , which is Clostridium ljungdahlii ( YP _ 003780648 . 1 ) ( SEQ ID NO : natively present in many bacteria , including Clostridium 71 ) , Bdh1 from Clostridium autoethanogenum 40 autoethanogenum , Clostridium ljungdahlii , and Clostridium ( AGY76060 . 1 ) ( SEQ ID NO : 72 ) , Bdh2 from Clostridium ragsdalei ( and Escherichia coli ) ( Brown , Biotechnol Biofu ljungdahlii ( YP 003782121 . 1 ) ( SEQ ID NO : 73 ) , Bdh2 e ls , 7 : 40 , 2014 ; U . S . Pat . No . 9 , 297 , 026 ) . Enzymes involved from Clostridium autoethanogenum ( AGY74784 . 1 ) ( SEQ in the conversion of acetyl - CoA to succinyl - CoA may ID NO : 74 ) , AdhE1 from Clostridium acetobutylicum include pyruvate : ferredoxin oxidoreductase ( PFOR ) ( EC ( NP _ 149325 . 1 ) ( SEQ ID NO : 75 ) , AdhE2 from Clostridium 45 1 . 2 . 7 . 1 ) , pyruvate carboxylase ( PYC ) ( EC 6 . 4 . 1 . 1 ) , malic acetobutylicum ( NP _ 149199 . 1 ) ( SEQ ID NO : 76 ) , AdhE enzyme / malate dehydrogenase ( EC 1 . 1 . 1 . 38 , EC 1 . 1 . 1 . 40 ) , from Clostridium beijerinckii ( WP _ 041893626 . 1 ) ( SEQ ID pyruvate phosphate dikinase ( PPDK ) ( EC : 2 . 7 . 9 . 1 ) , PEP NO : 77 ) , AdhE1 from Clostridium autoethanogenum carboxykinase ( PCK ) ( EC 4 . 1 . 1 . 49 ) , fumarate hydratase ! ( WP _ 023163372 . 1 ) ( SEQ ID NO : 78 ) , or AdhE2 from fumerase ( EC 4 . 2 . 1 . 2 ) , fumarate reductase ( EC 1 . 3 . 5 . 1 ) / Clostridium autoethanogenum ( WP _ 023163373 . 1 ) ( SEQ ID 50 succinate dehydrogenase ( EC 1 . 3 . 5 . 4 ) , and succinyl - CoA NO : 79 ) . Clostridium autoethanogenum , Clostridium ljung - synthetase ( EC 6 . 2 . 1 . 5 ) . The pyruvate : ferredoxin oxi dahlii , and Clostridium ragsdalei have native activity for doreductase may be , for example , from Clostridium this step . However , overexpression of endogenous alcohol autoethanogenum ( AGY75153 , AGY77232 ) or Escherichia dehydrogenase or introduction of an exogenous alcohol coli ( NP _ 415896 ) . The pyruvate carboxylase may be , for dehydrogenase in Clostridium autoethanogenum , 55 example , from Clostridium autoethanogenum ( AGY75817 ) . Clostridium ljungdahlii , or Clostridium ragsdalei may be The malic enzyme / malate dehydrogenase may be , for desirable to enhance product yields . Escherichia coli likely example , from Clostridium autoethanogenum ( AGY76687 ) does not have native activity for this step . or Escherichia coli ( NP _ 416714 , NP _ 417703 ) . The pyru
Step 18 shows the conversion of 3 - hydroxybutyryl - CoA vate phosphate dikinase ( PPDK ) may be , for example , from to 3 - hydroxybutyrylaldehyde . This step may be catalyzed by 60 Clostridium autoethanogenum ( AGY76274 , AGY77114 ) . butyraldehyde dehydrogenase ( EC 1 . 2 . 1 . 57 ) . The butyral - The PEP carboxykinase ( PCK ) may be , for example , from dehyde dehydrogenase may be , for example , Bld from Clostridium autoethanogenum ( AGY76928 ) or Escherichia Clostridium saccharoperbutylacetonicum ( AAP42563 . 1 ) coli ( NP _ 417862 ) . The fumarate hydratase / fumerase may ( SEQ ID NO : 80 ) . Clostridium autoethanogenum , be , for example , from Clostridium autoethanogenum Clostridium ljungdahlii , and Clostridium ragsdalei do not 65 ( AGY76121 , AGY76122 ) or Escherichia coli ( NP 416128 , have known native activity for this step . Escherichia coli NP _ 416129 , NP _ 418546 ) . The fumarate reductase / succi does not have known native activity for this step . nate dehydrogenase may be , for example , from Clostridium

23 US 9 , 957 , 531 B1
24 autoethanogenum ( AGY74573 , AGY74575 , AGY75257 , an exogenous enzyme or overexpression of an endogenous AGY77166 ) or Escherichia coli ( NP _ 415249 , NP _ 415250 , enzyme may be required to produce downstream products at NP _ 415251 , NP _ 415252 , NP _ 418575 , NP _ 418576 , desirable levels . Additionally , in certain embodiments , a NP _ 418577 , NP _ 418578 ) . The succinyl - CoA synthetase disruptive mutation may be introduced to an endogenous may be , for example , from Escherichia coli ( NP _ 415256 , 5 enzyme , such as an endogenous thioesterase , to reduce or NP _ 415257 ) . eliminate competition with introduced Ptb - Buk .
Step 22 shows shows the conversion of acetyl - CoA and Step 27 shows the conversion of shows the conversion of succinyl - CoA to 3 - oxo - adipyl - CoA . This step may be cata - 3 - hydroxbutyryl - CoA to crotonyl - CoA . This step may be lyzed by B - ketoadipyl - CoA thiolase ( EC 2 . 3 . 1 . 16 ) . The catalyzed by a crotonyl - CoA hydratase ( crotonase ) ( EC ketoisovalerate oxidoreductase may be , for example , PaaJ 10 4 . 2 . 1 . 17 ) or crotonyl - CoA reductase ( EC 1 . 3 . 1 . 38 ) . The from Escherichia coli ( WP _ 001206190 . 1 ) . Clostridium crotonyl - CoA hydratase ( crotonase ) or crotonyl - CoA reduc autoethanogenum , Clostridium ljungdahlii , and Clostridium tase may be , for example , Crt from C . acetobutylicum ragsdalei do not have known native activity for this step . ( NP _ 349318 . 1 ) ( SEQ ID NO : 52 ) or PhaJ from Aeromonas Escherichia coli does not have known native activity for this caviae ( 032472 ) . Clostridium autoethanogenum , step . 15 Clostridium Ijungdahlii , and Clostridium ragsdalei do not
Step 23 shows the conversion of 3 - oxo - adipyl - CoA to have known native activity for this step . Escherichia coli 3 - hydroxyadipyl - CoA . This step may be catalyzed by 3 - hy - does not have known native activity for this step . droxybutyryl - CoA dehydrogenase ( EC 1 . 1 . 1 . 157 ) or Step 28 shows the conversion of crotonyl - CoA to croto acetoacetyl - CoA hydratase ( EC 4 . 2 . 1 . 119 ) . The 3 - hydroxy - nate . This step may be catalyzed by phosphate butyryltrans butyryl - CoA dehydrogenase or acetoacetyl - CoA hydratase 20 ferase ( EC 2 . 3 . 1 . 19 ) + butyrate kinase ( EC 2 . 7 . 2 . 7 ) . Exem may be , for example , Hbd from Clostridium beijerinckii plary sources for phosphate butyryltransferase and butyrate ( WP 011967675 . 1 ) ( SEQ ID NO : 55 ) , Hbd from kinase are described elsewhere in this application . Native Clostridium acetobutylicum ( NP _ 349314 . 1 ) ( SEQ ID NO : enzymes in Clostridium autoethanogenum , Clostridium 56 ) , Hbdl from Clostridium kluyveri ( WP _ 011989027 . 1 ) ljungdahlii , and Clostridium ragsdalei ( or Escherichia coli ) , ( SEQ ID NO : 57 ) , or PaaH1 from Cupriavidus necator 25 such as thioesterases from Clostridium autoethanogenum , ( WP _ 010814882 . 1 ) . Of note , PhaB is R - specific and Hbd is may catalyze this step and result in the production of some S - specific . Additionally , Hbdl from Clostridium kluyveri is amount of downstream products . However , introduction of NADPH - dependent and Hbd from Clostridium acetobutyli - an exogenous enzyme or overexpression of an endogenous cum and Clostridium beijerinckii are NADH - dependent . enzyme may be required to produce downstream products at Clostridium autoethanogenum , Clostridium ljungdahlii , and 30 desirable levels . Additionally , in certain embodiments , a Clostridium ragsdalei do not have known native activity for disruptive mutation may be introduced to an endogenous this step . Escherichia coli does not have known native enzyme , such as an endogenous thioesterase , to reduce or activity for this step . eliminate competition with introduced Ptb - Buk .
Step 24 shows the conversion of 3 - hydroxyadipyl - CoA to Step 29 shows the conversion of crotonate to crotonalde 2 , 3 - dehydroadipyl - CoA . This step may be catalyzed by an 35 hyde . This step may be catalyzed by aldehyde : ferredoxin enoyl - CoA hydratase ( EC : 4 . 2 . 1 . 17 ) or enoyl - CoA reductase oxidoreductase ( EC 1 . 2 . 7 . 5 ) . Exemplary sources for alde ( EC : 1 . 3 . 1 . 38 ) . The enoyl - CoA hydratase or enoyl - CoA hyde : ferredoxin oxidoreductases are described elsewhere in reductase may be , for example , Crt from C . acetobutylicum this application . AOR catalyzes the reaction of an acid and ( NP _ 349318 . 1 ) or PhaJ from Aeromonas caviae ( 032472 ) reduced ferredoxin to form an aldehyde and oxidized ferre ( Seq . ID No . 52 ) . Clostridium autoethanogenum , 40 doxin . In acetogens , this reaction can be coupled to oxida Clostridium ljungdahlii , and Clostridium ragsdalei do not tion CO ( via CO dehydrogenase , EC 1 . 2 . 7 . 4 ) or hydrogen have known native activity for this step . Escherichia coli ( via ferredoxin - dependent hydrogenase , EC 1 . 12 . 7 . 2 or does not have known native activity for this step . 1 . 12 . 1 . 4 ) that both yield reduced ferredoxin ( Köpke , Curr
Step 25 shows the conversion of 2 , 3 - dehydroadipyl - CoA Opin Biotechnol 22 : 320 - 325 , 2011 ; Köpke , PNAS USA , to adipyl - CoA . This step may be catalyzed by trans - 2 - enoyl - 45 107 : 13087 - 13092 , 2010 ) . Clostridium autoethanogenum , CoA reductase ( EC 1 . 3 . 8 . 1 , EC 1 . 3 . 1 . 86 , EC 1 . 3 . 1 . 85 , EC Clostridium ljungdahlii , and Clostridium ragsdalei have 1 . 3 . 1 . 44 ) . The trans - 2 - enoyl - CoA reductase may be , for native activity for this step . However , overexpression of example , Bcd from C . acetobutylicum ( NP _ 349317 . 1 ) that endogenous AOR or introduction of an exogenous AOR in forms a complex with electron flavoproteins EtfAB Clostridium autoethanogenum , Clostridium ljungdahlii , or ( NP _ 349315 , NP _ 349316 ) , Cor from Streptomyces collinus 50 Clostridium ragsdalei may be desirable to enhance product ( AAA92890 ) , Cor from Rhodobacter sphaeroides yields . AOR of Pyrococcus furiosus has been demonstrated ( YP _ 354044 . 1 ) , Ter from Treponema denticola activity converting crotonaldehyde and crotonate ( Loes , J ( NP _ 971211 . 1 ) , or Ter from Euglena gracilis Bacteriol , 187 : 7056 - 7061 , 2005 ) . Alternatively , exogenous ( AY741582 . 1 ) . Clostridium autoethanogenum , Clostridium AOR may be introduced into a microorganism that does not ljungdahlii , and Clostridium ragsdalei do not have known 55 natively comprise AOR , e . g . , E . coli . In particular , the native activity for this step . Escherichia coli does not have co - expression of Ptb - Buk and AOR ( and , optionally , Adh ) known native activity for this step . may enable such a microorganism to produce new non
Step 26 shows the conversion of adipyl - CoA to adipic native products . acid . This step may be catalyzed by phosphate butyryltrans - Step 30 shows the conversion of crotonaldehyde to ferase ( EC 2 . 3 . 1 . 19 ) + butyrate kinase ( EC 2 . 7 . 2 . 7 ) . Exem - 60 2 - buten - 1 - ol . This step may be catalyzed by alcohol dehy plary sources for phosphate butyryltransferase and butyrate drogenase ( EC 1 . 1 . 1 . 1 . or 1 . 1 . 1 . 2 . ) . Alcohol dehydrogenase kinase are described elsewhere in this application . Native can convert an aldehyde and NAD ( P ) H to an alcohol and enzymes in Clostridium autoethanogenum , Clostridium NAD ( P ) . The alcohol dehydrogenase may be , for example , ljungdahlii , and Clostridium ragsdalei ( or Escherichia coli ) , Adh from Clostridium autoethanogenum ( AGY76060 . 1 ) such as thioesterases from Clostridium autoethanogenum , 65 ( SEQ ID NO : 67 ) , Clostridium ljungdahlii ( ADK17019 . 1 ) may catalyze this step and result in the production of some ( SEQ ID NO : 68 ) , or Clostridium ragsdalei , Bdh? from amount of downstream products . However , introduction of Clostridium acetobutylicum ( NP _ 349891 . 1 ) ( SEQ ID NO :

US 9 , 957 , 531 B1 25 26
vin
69 ) , Bdh from Clostridium beijerinckii ( WP _ 041897187 . 1 ) duced to an endogenous enzyme , such as an endogenous ( SEQ ID NO : 70 ) , Bdhl from Clostridium ljungdahlii thioesterase , to reduce or eliminate competition with intro ( YP _ 003780648 . 1 ) ( SEQ ID NO : 71 ) , Bdh1 from duced Ptb - Buk . Clostridium autoethanogenum ( AGY76060 . 1 ) ( SEQ ID Step 34 shows the conversion of acetobutyrate to acety NO : 72 ) . Bdh2 from Clostridium ljungdahlii 5 lacetone . This step may be catalyzed by an acetoacetate ( YP 003782121 . 1 ) ( SEO ID NO : 73 ) . Bdh2 from decarboxylase ( EC 4 . 1 . 1 . 4 ) . The acetoacetate decarboxylase Clostridium autoethanogenum ( AGY74784 . 1 ) ( SEQ ID may be , for example , Adc from Clostridium beijerinckii NO : 74 ) , AdhE1 from Clostridium acetobutylicum ( WP _ 012059998 . 1 ) ( SEQ ID NO : 14 ) . This step may also be
catalyzed by an alpha - ketoisovalerate decarboxylase ( EC ( NP _ 149325 . 1 ) ( SEQ ID NO : 75 ) , AdhE2 from Clostridium acetobutylicum ( NP _ 149199 . 1 ) ( SEQ ID NO : 76 ) , AdhE 10 4 . 1 . 1 . 74 ) . The alpha - ketoisovalerate decarboxylase may be ,
for example , KivD from Lactococcus lactis ( SEQ ID NO : from Clostridium beijerinckii ( WP _ 041893626 . 1 ) ( SEQ ID 15 ) . Clostridium autoethanogenum , Clostridium ljungdahlii , NO : 77 ) , AdhEi from Clostridium autoethanogenum and Clostridium ragsdalei do not have known native activity ( WP _ 023163372 . 1 ) ( SEQ ID NO : 78 ) , or AdhE2 from for this step . Additionally , Escherichia coli does not have Clostridium autoethanogenum ( WP _ 023163373 . 1 ) ( SEQ ID EPD 15 known native activity for this step . Rarely , conversion of NO : 79 ) . Clostridium autoethanogenum , Clostridium ljung acetoacetate to acetone may occur spontaneously . However , dahlii , and Clostridium ragsdalei have native activity for spontaneous conversion is highly inefficient and unlikely to this step . However , overexpression of endogenous alcohol result in the production of downstream products at desirable dehydrogenase or introduction of an exogenous alcohol levels . dehydrogenase in Clostridium autoethanogenum , 20 Step 35 shows the conversion of acetylacetone to Clostridium ljungdahlii , or Clostridium ragsdalei may be 3 - methyl - 2 - butanol . This step may be catalyzed by a prima desirable to enhance product yields . Escherichia coli likely ry : secondary alcohol dehydrogenase ( EC 1 . 1 . 1 . 2 ) . The pri does not have native activity for this step . mary : secondary alcohol dehydrogenase may be , for
Step 31 shows the conversion of crotonyl - CoA to butyryl - example , SecAdh from Clostridium autoethanogenum COA . This step may be catalyzed by butyryl - CoA dehydro - 25 ( AGY74782 . 1 ) ( SEQ ID NO : 16 ) , SecAdh from Clostridium genase or trans - 2 - enoyl - CoA reductase ( EC 1 . 3 . 8 . 1 , EC ljungdahlii ( ADK15544 . 1 ) ( SEQ ID NO : 17 ) , SecAdh from 1 . 3 . 1 . 86 , EC 1 . 3 . 1 . 85 , EC 1 . 3 . 1 . 44 ) . The butyryl - CoA dehy Clostridium ragsdalei ( WP _ 013239134 . 1 ) ( SEQ ID NO : drogenase or trans - 2 - enoyl - CoA reductase may be , for 18 ) , or SecAdh from Clostridium beijerinckii example , Bcd from C . acetobutylicum ( NP _ 349317 . 1 ) that ( WP _ 026889046 . 1 ) ( SEQ ID NO : 19 ) . This step may also be forms a complex with electron flavoproteins EtfAB 30 ETAB 30 catalyzed by a primary : secondary alcohol dehydrogenase
( EC 1 . 1 . 1 . 80 ) , such as SecAdh from Thermoanaerobacter ( NP _ 349315 , NP _ 349316 ) , Ccr from Streptomyces collinus brokii ( 3FSR _ A ) ( SEQ ID NO : 20 ) . Clostridium autoetha ( AAA92890 ) , Cor from Rhodobacter sphaeroides nogenum , Clostridium ljungdahlii , and Clostridium ragsda ( YP _ 354044 . 1 ) , Ter from Treponema denticola lei have native activity for this step ( Köpke , Appl Environ ( NP 971211 . 1 ) , or Ter from Euglena gracilis 35 . Microbiol . 80 : 3394 - 3403 , 2014 ) . However , Escherichia ( AY741582 . 1 ) . Clostridium autoethanogenum , Clostridium coli does not have known native activity for this step . ljungdahlii , and Clostridium ragsdalei do not have known Knocking down or knocking out this enzyme in Clostridium native activity for this step . Escherichia coli does not have autoethanogenum , Clostridium ljungdahlii , or Clostridium known native activity for this step . ragsdalei results in the production and accumulation of
Step 32 shows the conversion of butyryl - CoA to aceto - 40 acetylacetone rather than 3 - methyl - 2 - butanol ( WO 2015 / butyryl - CoA . This step may be catalyzed by thiolase or 085015 ) . acyl - CoA acetyltransferase ( EC 2 . 3 . 1 . 9 ) . The thiolase may Step 36 shows the conversion of acetobutyryl - CoA to be , for example , ThlA from Clostridium acetobutylicum 3 - hydroxyhexanoyl - CoA . This step may be catalyzed by ( WP 010966157 . 1 ) ( SEQ ID NO : 1 ) , Th1A1 from 3 - hydroxybutyryl - CoA dehydrogenase ( EC 1 . 1 . 1 . 157 ) or Clostridium kluvveri ( EDK35681 ) , Th1A2 from Clostridium 45 acetoacetyl - CoA hydratase ( EC 4 . 2 . 1 . 119 ) . The 3 - hydroxy kluyveri ( EDK35682 ) , Th1A3 from Clostridium kluyveri butyryl - CoA dehydrogenase or acetoacetyl - CoA hydratase ( EDK35683 ) , PhaA from Cupriavidus necator may be , for example , Hbd from Clostridium beijerinckii ( WP _ 013956452 . 1 ) ( SEQ ID NO : 2 ) , Bkt? from Cupriavi ( WP _ 011967675 . 1 ) ( SEQ ID NO : 55 ) , Hbd from dus necator ( WP _ 011615089 . 1 ) ( SEQ ID NO : 3 ) , or AtoB Clostridium acetobutylicum ( NP _ 349314 . 1 ) ( SEQ ID NO : from Escherichia coli ( NP _ 416728 . 1 ) ( SEQ ID NO : 4 ) . 50 56 ) , Hbdl from Clostridium kluyveri ( WP _ 011989027 . 1 ) Clostridium autoethanogenum , Clostridium ljungdahlii , and ( SEQ ID NO : 57 ) , Hbd2 from Clostridium kluyveri Clostridium ragsdalei do not have known native activity for ( EDK34807 ) , or PaaH1 from Cupriavidus necator this step . Escherichia coli has native activity for this step . ( WP _ 010814882 . 1 ) . Of note , PhaB is R - specific and Hbd is
Step 33 shows the conversion of acetobutyryl - CoA to S - specific . Additionally , Hbd1 from Clostridium kluyveri is acetobutyrate . This step may be catalyzed by phosphate 55 NADPH - dependent and Hbd from Clostridium acetobutyli butyryltransferase ( EC 2 . 3 . 1 . 19 ) + butyrate kinase ( EC cum and Clostridium beijerinckii are NADH - dependent . 2 . 7 . 2 . 7 ) . Exemplary sources for phosphate butyryltrans Clostridium autoethanogenum , Clostridium ljungdahlii , and ferase and butyrate kinase are described elsewhere in this Clostridium ragsdalei do not have known native activity for application . Native enzymes in Clostridium autoethanoge - this step . Escherichia coli does not have known native num , Clostridium ljungdahlii , and Clostridium ragsdalei ( or 60 activity for this step . Escherichia coli ) , such as thioesterases from Clostridium Step 37 shows the conversion of 3 - hydroxyhexanoyl - CoA autoethanogenum , may catalyze this step and result in the to 3 - hydroxyhexanoate . This step may be catalyzed by production of some amount of downstream products . How - phosphate butyryltransferase ( EC 2 . 3 . 1 . 19 ) + butyrate kinase ever , introduction of an exogenous enzyme or overexpres - ( EC 2 . 7 . 2 . 7 ) . Exemplary sources for phosphate butyryltrans sion of an endogenous enzyme may be required to produce 65 ferase and butyrate kinase are described elsewhere in this downstream products at desirable levels . Additionally , in application . Native enzymes in Clostridium autoethanoge certain embodiments , a disruptive mutation may be intro num , Clostridium ljungdahlii , and Clostridium ragsdalei ( or

27 US 9 , 957 , 531 B1
28 Escherichia coli ) , such as thioesterases from Clostridium widespread across many genera and kingdoms of life and autoethanogenum , may catalyze this step and result in the include , e . g . , MvaS from Staphylococcus aureus production of some amount of downstream products . How ( WP _ 053014863 . 1 ) , ERG13 from Saccharomyces cerevi ever , introduction of an exogenous enzyme or overexpres siae ( NP _ 013580 . 1 ) , HMGCS2 from Mus musculus sion of an endogenous enzyme may be required to produce 5 ( NP _ 032282 . 2 ) , and many other members of the EC downstream products at desirable levels . Additionally , in 2 . 3 . 3 . 10 group of enzymes . Clostridium autoethanogenum , certain embodiments , a disruptive mutation may be intro - Clostridium ljungdahlii , and Clostridium ragsdalei do not duced to an endogenous enzyme , such as an endogenous have known native activity for this step . Escherichia coli thioesterase , to reduce or eliminate competition with intro does not have known native activity for this step . duced Ptb - Buk . 10 Step 41 shows the conversion of 3 - hydroxy - 3 - methyl
Step 38 shows the conversion of 3 - hydroxyhexanoate to glutanoyl - CoA to 3 - methylgluconyl - CoA . This step may be 1 , 3 - hexaldehyde . This step may be catalyzed by aldehyde : catalyzed by a 3 - hydroxybutyryl - CoA dehydratase ( EC ferredoxin oxidoreductase ( EC 1 . 2 . 7 . 5 ) . Exemplary sources 4 . 2 . 1 . 55 ) . The 3 - hydroxybutyryl - CoA dehydratase may be , for aldehyde : ferredoxin oxidoreductases are described else - for example , Liuc from Myxococcus xanthus where in this application . AOR catalyzes the reaction of an 15 ( WP _ 011553770 . 1 ) . This step may also be catalyzed by a acid and reduced ferredoxin to form an aldehyde and oxi short - chain - enoyl - CoA hydratase ( EC 4 . 2 . 1 . 150 ) or an dized ferredoxin . In acetogens , this reaction can be coupled enoyl - CoA hydratase ( EC 4 . 2 . 1 . 17 ) . Clostridium autoetha to oxidation CO ( via CO dehydrogenase , EC 1 . 2 . 7 . 4 ) or nogenum , Clostridium ljungdahlii , and Clostridium ragsda hydrogen ( via ferredoxin - dependent hydrogenase , EC lei do not have known native activity for this step . Escheri 1 . 12 . 7 . 2 or 1 . 12 . 1 . 4 ) that both yield reduced ferredoxin 20 chia coli does not have known native activity for this step . ( Köpke , Curr Opin Biotechnol 22 : 320 - 325 , 2011 ; Köpke , Step 42 shows the conversion of 3 - methylgluconyl - CoA PNAS USA , 107 : 13087 - 13092 , 2010 ) . Clostridium to 2 - methylcrotonyl - CoA . This step may be catalyzed by a autoethanogenum , Clostridium ljungdahlii , and Clostridium methylcrotonyl - CoA decarboxylase ( with high structural ragsdalei have native activity for this step . However , over - similarity to glutaconate - CoA transferase ( EC 2 . 8 . 3 . 12 ) ) , expression of endogenous AOR or introduction of an exog - 25 e . g . , aibAB from Myxococcus xanthus ( WP _ 011554267 . 1 enous AOR in Clostridium autoethanogenum , Clostridium and WP _ 011554268 . 1 ) . This step may also be catalyzed by ljungdahlii , or Clostridium ragsdalei may be desirable to a methylcrotonoyl - CoA carboxylase ( EC 6 . 4 . 1 . 4 ) , e . g . , enhance product yields . Alternatively , exogenous AOR may LiuDB from Pseudomonas aeruginosa ( NP _ 250702 . 1 and be introduced into a microorganism that does not natively NP _ 250704 . 1 ) or MCCA and MCCB from Arabidopsis comprise AOR , e . g . , E . coli . In particular , the co - expression 30 thaliana ( NP _ 563674 . 1 and NP _ 567950 . 1 ) . Clostridium of Ptb - Buk and AOR ( and , optionally , Adh ) may enable such autoethanogenum , Clostridium ljungdahlii , and Clostridium a microorganism to produce new non - native products . ragsdalei do not have known native activity for this step .
Step 39 shows the conversion of 1 , 3 - hexaldehyde to Escherichia coli does not have known native activity for this 1 , 3 - hexanediol . This step may be catalyzed by alcohol step . dehydrogenase ( EC 1 . 1 . 1 . 1 . or 1 . 1 . 1 . 2 . ) . Alcohol dehydro - 35 Step 43 shows the conversion of methylcrotonyl - CoA to genase can convert an aldehyde and NAD ( P ) H to an alcohol isovaleryl - CoA . This step may be catalyzed by an oxi and NAD ( P ) . The alcohol dehydrogenase may be , for doreductase , zinc - binding dehydrogenase . This oxidoreduc example , Adh from Clostridium autoethanogenum tase , zinc - binding dehydrogenase may be , for example , ( AGY76060 . 1 ) ( SEQ ID NO : 67 ) , Clostridium ljungdahlii AibC from Myxococcus xanthus ( WP _ 011554269 . 1 ) . ( ADK17019 . 1 ) ( SEO ID NO : 68 ) , or Clostridium ragsdalei , 40 Clostridium autoethanogenum , Clostridium ljungdahlii , and Bdh? from Clostridium acetobutylicum ( NP _ 349891 . 1 ) Clostridium ragsdalei do not have known native activity for ( SEO ID NO : 69 ) , Bdh from Clostridium beijerinckii this step . Escherichia coli does not have known native ( WP _ 041897187 . 1 ) ( SEQ ID NO : 70 ) , Bdh1 from activity for this step . Clostridium ljungdahlii ( YP _ 003780648 . 1 ) ( SEQ ID NO : Step 44 shows the conversion of isovaleryl - CoA to isoval 71 ) , Bdh1 from Clostridium autoethanogenum 45 erate . This step may be catalyzed by COA - transferase ( i . e . , ( AGY76060 . 1 ) ( SEQ ID NO : 72 ) , Bdh2 from Clostridium acetyl - CoA : acetoacetyl - CoA transferase ) ( EC 2 . 8 . 3 . 9 ) . The ljungdahlii ( YP _ 003782121 . 1 ) ( SEQ ID NO : 73 ) , Bdh2 COA - transferase may be , for example , CtfAB , a heterodimer from Clostridium autoethanogenum ( AGY74784 . 1 ) ( SEQ comprising subunits CtfA and CtfB , from Clostridium bei ID NO : 74 ) , AdhEl from Clostridium acetobutylicum jerinckii ( CtfA , WP _ 012059996 . 1 ) ( SEQ ID NO : 5 ) ( CtfB , ( NP 149325 . 1 ) ( SEO ID NO : 75 ) , AdhE2 from Clostridium 50 WP _ 012059997 . 1 ) ( SEQ ID NO : 6 ) . This step may also be acetobutylicum ( NP _ 149199 . 1 ) ( SEQ ID NO : 76 ) , AdhE catalyzed by thioesterase ( EC 3 . 1 . 2 . 20 ) . The thioesterase from Clostridium beijerinckii ( WP _ 041893626 . 1 ) ( SEQ ID may be , for example , TesB from Escherichia coli NO : 77 ) , AdhEl from Clostridium autoethanogenum ( NP _ 414986 . 1 ) ( SEQ ID NO : 7 ) . This step may also be ( WP _ 023163372 . 1 ) ( SEQ ID NO : 78 ) , or AdhE2 from catalyzed by a putative thioesterase , e . g . , from Clostridium Clostridium autoethanogenum ( WP _ 023163373 . 1 ) ( SEQ ID 55 autoethanogenum or Clostridium ljungdahlii . In particular , NO : 79 ) . Clostridium autoethanogenum , Clostridium ljung - three putative thioesterases have been identified in dahlii , and Clostridium ragsdalei have native activity for Clostridium autoethanogenum : ( 1 ) “ thioesterase 1 ” this step . However , overexpression of endogenous alcohol ( AGY74947 . 1 ; annotated as palmitoyl - CoA hydrolase ; SEQ dehydrogenase or introduction of an exogenous alcohol ID NO : 8 ) , ( 2 ) “ thioesterase 2 ” ( AGY75747 . 1 ; annotated as dehydrogenase in Clostridium autoethanogenum , 60 4 - hydroxybenzoyl - CoA thioesterase ; SEQ ID NO : 9 ) , and Clostridium ljungdahlii , or Clostridium ragsdalei may be ( 3 ) “ thioesterase 3 ” ( AGY75999 . 1 ; annotated as putative desirable to enhance product yields . Escherichia coli likely thioesterase ; SEQ ID NO : 10 ) . Three putative thioesterases does not have native activity for this step . have also been identified in Clostridium ljungdahlii : ( 1 )
Step 40 shows the conversion of acetoacetyl - CoA to “ thioesterase 1 ” ( ADK15695 . 1 ; annotated as predicted acyl 3 - hydroxy - 3 - methylglutaryl - CoA . This step may be cata - 65 COA thioesterase 1 ; SEQ ID NO : 11 ) , ( 2 ) “ thioesterase 2 ” lyzed by a hydroxymethylglutaryl - CoA synthase ( HMG - ( ADK16655 . 1 ; annotated as predicted thioesterase ; SEQ ID COA synthase ) ( EC 2 . 3 . 3 . 10 ) . HMG - CoA synthases are NO : 12 ) , and ( 3 ) “ thioesterase 3 ” ( ADK16959 . 1 ; annotated

29 US 9 , 957 , 531 B1
30 as predicted thioesterase ; SEQ ID NO : 13 ) . This step may desirable to enhance product yields . Escherichia coli likely also be catalyzed by phosphate butyryltransferase ( EC does not have native activity for this step . 2 . 3 . 1 . 19 ) + butyrate kinase ( EC 2 . 7 . 2 . 7 ) . Exemplary sources Step 47 shows the conversion of isovaleryl - CoA to isoval for phosphate butyryltransferase and butyrate kinase are eraldehyde . This step may be catalyzed by butyraldehyde described elsewhere in this application . Native enzymes in 5 dehydrogenase ( EC 1 . 2 . 1 . 57 ) . The butyraldehyde dehydro Clostridium autoethanogenum . Clostridium liungdahlii , and genase may be , for example , Bld from Clostridium saccha Clostridium ragsdalei ( or Escherichia coli ) , such as thio roperbutylacetonicum ( AAP42563 . 1 ) ( SEQ ID NO : 80 ) . esterases from Clostridium autoethanogenum , may catalyze Clostridium autoethanogenum , Clostridium ljungdahlii , and
Clostridium ragsdalei likely do not have native activity for this step and result in the production of some amount of downstream products . However , introduction of an exog 10 this step . Escherichia coli does not have known native
activity for this step . enous enzyme or overexpression of an endogenous enzyme Overview of Ptb - Buk may be required to produce downstream products at desir The invention provides new pathways utilizing the Ptb able levels . Additionally , in certain embodiments , a disrup Buk enzyme system . In nature , this enzyme system is found tive mutation may be introduced to an endogenous enzyme , 15 in a range of butyrate producing microorganisms , such as such as an endogenous thioesterase , to reduce or eliminate butyrate - producing Clostridia or Butyrivibrio . In particular , competition with introduced Ptb - Buk . phosphate butyryltransferase ( Ptb ) ( EC 2 . 3 . 1 . 19 ) natively
Step 45 shows the conversion of isovalerate to isovaler catalyzes the reaction of butanoyl - CoA + phosphate to form aldehyde . This step may be catalyzed by aldehyde : ferre - COA + butanoyl phosphate and butyrate kinase ( Buk ) ( EC doxin oxidoreductase ( EC 1 . 2 . 7 . 5 ) . The aldehyde : ferredoxin 20 2 . 7 . 2 . 7 ) natively catalyzes the reaction of butanoyl phos oxidoreductase ( AOR ) may be , for example , AOR from phate and ADP to form butyrate ( butanoate ) and ATP . Clostridium autoethanogenum ( WP _ 013238665 . 1 ; Accordingly , these enzymes together ( Ptb - Buk ) natively WP _ 013238675 . 1 ) ( SEQ ID NOs : 63 and 64 , respectively ) catalyze the conversion of butanoyl - CoA to butyrate and or AOR from Clostridium ljungdahlii ( ADK15073 . 1 ; generate one ATP via substrate level - phosphorylation ( FIG . ADK15083 . 1 ) ( SEQ ID NOs : 65 and 66 , respectively ) . 25 2 ) . However , the inventors have discovered that Ptb is Further exemplary sources for aldehyde : ferredoxin oxi - promiscuous and is capable of accepting a variety of acyl doreductases are described elsewhere in this application . CoAs and enoyl - CoAs as substrates , such that Ptb - Buk may Clostridium autoethanogenum , Clostridium ljungdahlii , and be used to convert a number of acyl - CoAs and enoyl - CoAs Clostridium ragsdalei have native activity for this step to their corresponding acids or alkenates , respectively , while However , overexpression of endogenous AOR or introduc - 30 simultaneously generating ATP . It has been reported Ptb is tion of an exogenous AOR in Clostridium autoethanoge - active on a range of acyl - CoAs including acetoacetyl - CoA , num , Clostridium ljungdahlii , or Clostridium ragsdalei may in vitro ( Thompson , Appl Environ Microbiol , 56 : 607 - 613 , be desirable to enhance product yields . Alternatively , exog - 1990 ) . It has not previously been shown that acetoacetyl enous AOR may be introduced into a microorganism that phosphate could be a substrate for Buk . Although Buk is does not natively comprise AOR , e . g . , E . coli . In particular , 35 known to accept a broad substrate range ( Liu , Appl Micro the co - expression of Ptb - Buk and AOR ( and , optionally , biol Biotechnol , 53 : 545 - 552 , 2000 ) , no activity has been Adh ) may enable such a microorganism to produce new shown in vivo . non - native products . Additionally , the inventors have discovered that the intro
Step 46 shows the conversion of isovaleraldehyde to duction of exogenous Ptb - Buk enables certain microorgan isoamyl alcohol . This step may be catalyzed by alcohol 40 isms to produce useful products , including acetone , isopro dehydrogenase ( EC 1 . 1 . 1 . 1 . or 1 . 1 . 1 . 2 . ) . Alcohol dehydro - panol , isobutylene , 3 - hydroxybutyrate , 1 , 3 - butanediol , and genase can convert an aldehyde and NAD ( P ) H to an alcohol 2 - hydroxyisobutyrate , as well as other products such as and NAD ( P ) . The alcohol dehydrogenase may be , for propionate , caproate , and octonate . example , Adh from Clostridium autoethanogenum New pathways that rely on Ptb - Buk offer several major ( AGY76060 . 1 ) ( SEQ ID NO : 67 ) , Clostridium ljungdahlii 45 advantages over other known and existing pathway routes ( ADK17019 . 1 ) ( SEQ ID NO : 68 ) , or Clostridium ragsdalei , for production of products that rely on a COA - transferase Bdh? from Clostridium acetobutylicum ( NP 349891 . 1 ) as in the classic Clostridial acetone - butanol - ethanol ( ABE ) ( SEQ ID NO : 69 ) , Bdh from Clostridium beijerinckii fermentation pathway or a thioesterase ( Jones , Microbiol ( WP _ 041897187 . 1 ) ( SEQ ID NO : 70 ) , Bdh1 from Rev , 50 : 484 - 524 , 1986 ; Matsumoto , Appl Microbiol Bio Clostridium ljungdahlii ( YP _ 003780648 . 1 ) ( SEQ ID NO : 50 technol , 97 : 205 - 210 , 2013 ; May , Metabol Eng , 15 : 218 - 225 , 71 ) , Bdh1 from Clostridium autoethanogenum 2013 ) ( FIG . 3 ) . In particular , these new pathways ( 1 ) are not ( AGY76060 . 1 ) ( SEQ ID NO : 72 ) , Bdh2 from Clostridium dependent on the presence or production of particular mol ljungdahlii ( YP _ 003782121 . 1 ) ( SEQ ID NO : 73 ) , Bdh2 ecules , such as organic acids , e . g . , butyrate or acetate , from Clostridium autoethanogenum ( AGY74784 . 1 ) ( SEQ required for the CoA - transferase reaction and ( 2 ) allow for ID NO : 74 ) , AdhEl from Clostridium acetobutylicum 55 generation of ATP via substrate level phosphorylation that ( NP _ 149325 . 1 ) ( SEQ ID NO : 75 ) , AdhE2 from Clostridium would not be conserved in a thioesterase or COA - transferase acetobutylicum ( NP _ 149199 . 1 ) ( SEQ ID NO : 76 ) , AdhE reaction . The same advantages also apply when using the from Clostridium beijerinckii ( WP _ 041893626 . 1 ) ( SEQ ID Ptb - Buk system for other reactions , such as the conversion NO : 77 ) , AdhEl from Clostridium autoethanogenum of 3 - hydroxybutyryl - CoA to 3 - hydroxybutyrate . Thus , these ( WP _ 023163372 . 1 ) ( SEQ ID NO : 78 ) , or AdhE2 from 60 new pathways have the potential to yield much higher Clostridium autoethanogenum ( WP _ 023163373 . 1 ) ( SEQ ID production titers and rates by generating additional energy NO : 79 ) . Clostridium autoethanogenum , Clostridium ljung - and producing target products without co - production of dahlii , and Clostridium ragsdalei have native activity for undesired byproducts , such as acetate . this step . However , overexpression of endogenous alcohol Particularly on a commercial scale , it is not desirable for dehydrogenase or introduction of an exogenous alcohol 65 microorganisms to produce acetate ( or other organic acids dehydrogenase in Clostridium autoethanogenum , required for the CoA transferase reaction ) as byproduct , Clostridium ljungdahlii , or Clostridium ragsdalei may be since acetate diverts carbon away from target products and
state

US 9 , 957 , 531 B1 31 32
thus affects the efficiency and yield of target products . ( Schiel - Bengelsdorf , FEBS Lett , 586 : 2191 - 2198 , 2012 ) . Additionally , acetate may be toxic to microorganisms and / or The Ptb - Buk pathways described herein , however , provide may serve as a substrate for the growth of contaminating an alternative mechanism for the microorganism to generate microorganisms . Furthermore , the presence of acetate makes ATP via substrate level phosphorylation and , therefore , it more difficult to recover and separate target products and 5 avoid acetate production . In particular , acetate - forming to control fermentation conditions to favor the production of enzymes , such as Pta - Ack , that would otherwise be essential target products . ( Nagarajan , Microb Cell Factories , 12 : 118 , 2013 ) can be
ATP generation through substrate level phosphorylation replaced with Ptb - Buk as an alternative means of ATP can be used as a driving force for product synthesis , espe - generation . Since the microorganism can then rely on ATP cially in ATP - limited systems . In particular , acetogenic 10 generation via Ptb - Buk , this system provides a driving force bacteria are known to live on the thermodynamic edge of life that ensures maximum flux through the new pathways that ( Schuchmann , Nat Rev Microbiol , 12 : 809 - 821 , 2014 ) . As use Ptb - Buk . The generation of ATP may also be crucial for such , all acetogenic microorganisms isolated to date have downstream pathways that require ATP . For example , fer been described to produce acetate ( Drake , Acetogenic Pro - mentative production of isobutylene from acetone requires karyotes , In : The Prokaryotes , 3rd edition , pages 354 - 420 , 15 ATP . While the complete pathway from acetyl - CoA to New York , N . Y . , Springer , 2006 ) since the production of isobutylene is ATP - consuming when using a COA - trans acetate provides the microorganism with an option to ferase or a thioesterase , the pathway is energy neutral when directly generate ATP from substrate level phosphorylation using Ptb - Buk . via Pta ( phosphotransacetylase ) ( EC 2 . 3 . 1 . 8 ) and Ack ( ac - Exemplary sources for Ptb and Buk are provided . How etate kinase ) ( EC 2 . 7 . 2 . 1 ) . Although mechanisms such as 20 ever , it should be appreciated that other suitable sources for membrane gradients and electro bifurcation enzymes Ptb and Buk may be available . Additionally , Ptb and Buk coupled to ion or proton translocating systems , e . g . , the Rnf may be engineered to improve activity and / or genes encod complex ( Schuchmann , Nat Rev Microbiol , 12 : 809 - 821 , ing Ptb - Buk may be codon - optimized for expression in 2014 ) , conserve ATP in these microorganisms , direct ATP particular host microorganisms . generation remains critical for their survival . As a result , 25 The phosphate butyryltransferase may be or may be when introducing heterologous pathways that do not allow derived , for example , from any of the following sources , the for ATP generation , acetate is produced as a byproduct sequences of which are publically available :
Description Microorganism Accession
phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase
Clostridium sp . Clostridium sp . Clostridium saccharoperbutylacetonicum Clostridium saccharobutylicum Clostridium beijerinckii Clostridium beijerinckii Clostridium butyricum Clostridium sp . Clostridium botulinum Clostridium sp . Clostridium paraputrificum Clostridium sp . Clostridium celatum Clostridium baratii Clostridium intestinale Clostridium sp . Clostridium sp . Clostridium perfringens Clostridium botulinum Clostridium perfringens Clostridium perfringens Clostridium perfringens Clostridium perfringens Clostridium sp . Clostridium chauvoei Clostridium colicanis Clostridium cadaveris Clostridium acetobutylicum Clostridium pasteurianum Clostridium arbusti Clostridium pasteurianum Clostridium scatologenes Clostridium sp . Clostridium drakei Clostridium sp . Clostridium argentinense Clostridium akagii Clostridium sp . Clostridium carboxidivorans Clostridium sp . Clostridium sulfidigenes Clostridium tetanomorphum Clostridium hydrogeniformans Clostridium sp .
EKQ52186 WP _ 009167896 WP 015390396 WP 022743598 WP _ 026886639 WP 041893500 WP 003410761 CDB 14331 WP 049180512 CDB74819 WP _ 027098882 WP 024615655 WP 005211129 WP 039312969 WP _ 021800215 WP 042402499 WP 032117069 ABG85761 WP _ 003374233 WP 004460499 WP _ 003454254 WP 041707926 BAB82054 WP 008681116 WP 021876993 WP 002598839 WP _ 027637778 WP _ 010966357 WP 015617430 WP 010238988 WP 003445696 WP 029160341 WP _ 032120461 WP 032078800 WP _ 021281241 WP 039635970 WP _ 026883231 WP 053242611 WP 007063154 WP _ 035292411 WP _ 035133394 WP 035147564 WP _ 027633206 WP 040212965

US 9 , 957 , 531 B1 33 34
- continued Description Microorganism Accession phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase phosphate butyryltransferase
Candidatus Clostridium Clostridium sp . Clostridium sp . Clostridium lundense Clostridium algidicarnis Clostridium sp . Clostridium acetobutylicum Clostridium botulinum Clostridium botulinum Clostridium botulinum Clostridium botulinum Clostridium bornimense Clostridium botulinum Clostridium sp . Closfridiales bacterium Clostridium tetani Closfridiales bacterium Clostridium botulinum Clostridium tetani Clostridium tetani Clostridium senegalense Caloramator sp . Thermobrachium celere Clostridium cellulovorans Coprococcus comes Coprococcus comes Eubacterium sp . Clostridium sp . Caloramator australicus Clostridium sp . Lachnospiraceae bacterium Lachnospiraceae bacterium Fervidicella metallireducens Clostridium sp . Eubacterium xylanophilum Roseburia sp . Butyrivibrio crossotus Lachnospiraceae bacterium Clostridium sp . Peptostreptococcaceae bacterium Butyrivibrio crossotus Clostridium sp . Coprococcus eutactus Firmicutes bacterium Lachnospiraceae bacterium Lachnospiraceae bacterium Clostridium sp .
WP _ 040327613 WP _ 040192242 WP 050606427 WP _ 027625137 WP 029451333 WP _ 035306567 AAA75486 WP 025775938 WP _ 045541062 WP 003357252 WP 030037192 WP 044039341 WP _ 041346554 WP _ 053468896 WP _ 034572261 WP 023439553 ERI95297 WP _ 047403027 WP _ 011100667 WP _ 035111554 WP 010295062 WP _ 027307587 WP 018661036 WP 010073683 CDB84786 WP 008371924 CCZ03827 CCZ05442 WP _ 008907395 CCY59505 WP _ 035626368 WP 027440767 WP _ 035381340 CCX89274 WP _ 026834525 CDF44203 WP 005600912 WP _ 027117626 CDA68345 WP _ 026899905 CCY77124 CDE44914 WP _ 004853197 CCY23248 WP 027111007 WP _ 016293387 WP _ 046822491
- continued Microorganism Accession
In a preferred embodiment , the phosphate butyryltrans - 45 ferase is Ptb from Clostridium acetobutylicum ( WP _ 010966357 ; SEQ ID NO : 87 ) or Clostridium beijer Description inckii ( WP _ 026886639 ; SEQ ID NO : 88 ) ( WP _ 041893500 ; butyrate kinase SEQ ID NO : 89 ) . Clostridium autoethanogenum , butyrate kinase Clostridium ljungdahlii , and Clostridium ragsdalei do not so butyrate kinase natively contain phosphate butyryltransferase . butyrate kinase
The butyrate kinase may be or may be derived , for butyrate kinase butyrate kinase example , from any of the following sources , the sequences butyrate kinase
of which are publically available : butyrate kinase
WP _ 008681112 WP _ 008681114 WP _ 009167897 WP 011010889 WP 011967556 WP _ 012422882 WP _ 012450845 WP _ 015390397
Description Microorganism Accession
Clostridium sp . Clostridium sp . Clostridium sp . Clostridium perfringens Clostridium beijerinckii Clostridium botulinum Clostridium botulinum Clostridium saccharoperbutylacetonicum Clostridium beijerinckii Clostridium botulinum Clostridium chauvoei Clostridium saccharobutylicum Clostridium sp . Clostridium perfringens Clostridium beijerinckii Clostridium paraputrificum Clostridium sp . Clostridium botulinum Clostridium baratii Clostridium diolis Clostridium botulinum Clostridium beijerinckii
butyrate kinase butyrate kinase butyrate kinase butyrate kinase butyrate kinase butyrate kinase butyrate kinase butyrate kinase butyrate kinase butyrate kinase butyrate kinase
Clostridium pasteurianum Clostridium sp . Clostridium sp . Clostridium sp . Clostridium perfringens Clostridium sp . Clostridium colicanis Clostridium botulinum Clostridium perfringens Clostridium perfringens Clostridium celatum
butyrate kinase butyrate kinase butyrate kinase butyrate kinase butyrate kinase butyrate kinase
60 butyrate kinase butyrate kinase butyrate kinase butyrate kinase butyrate kinase butyrate kinase
65 butyrate kinase butyrate kinase
ALB48406 CDB14330 CDB74820 EKQ52187 QOSQKO WP _ 002582660 WP _ 002598838 WP _ 003371719 WP 003454444 WP _ 004459180 WP 005211128
WP _ 017209677 WP 017825911 WP 021876994 WP _ 022743599 WP _ 024615656 WP _ 025648345 WP _ 026886638 WP _ 027098883 WP _ 032117070 WP _ 035786166 WP _ 039312972 WP _ 039772701 WP _ 041082388 WP _ 041893502

US 9 , 957 , 531 B1 35 36
- continued design based on available crystal structures of Ptb and Buk with and without a bound substrate where the binding pocket
Description Microorganism Accession would be changed to accommodate the new substrate or through saturation mutagenesis . When activity is obtained , it butyrate kinase Clostridium sp . WP _ 042402497
butyrate kinase Clostridium baratii 5 can be further improved through iterative cycles of enzyme WP 045725505 butyrate kinase Clostridium perfringens WP 049039634 engineering . These engineering efforts would be combined butyrate kinase Clostridium botulinum WP _ 049180514 with assays to test enzyme activity . These types of strategies butyrate kinase Clostridium botulinum WP 053341511 have previously proven effective ( see , e . g . , Huang , Nature , butyrate kinase Clostridium butyricum ABU40948 butyrate kinase Clostridium sp . 537 : 320 - 327 , 2016 ; Khoury , Trends Biotechnol , 32 : 99 - 109 , CDE44915 butyrate kinase Clostridium senegalense WP _ 010295059 10 2014 ; Packer , Nature Rev Genetics , 16 : 379 - 394 , 2015 ; butyrate kinase Clostridium intestinale WP _ 021800216 Privett , PNAS USA , 109 : 3790 - 3795 , 2012 ) . butyrate kinase Eubacterium ventriosum WP _ 005363839 To improve substrate specificity of Ptb - Buk towards a butyrate kinase Closfridi ales bacterium WP 021657038 butyrate kinase Clostridium sp . specific acyl - CoA substrate , Ptb - Buk variants from public WP _ 021281242 butyrate kinase Clostridium sporogenes WP 045520059 databases or generated Ptb - Buk mutants ( for example , from butyrate kinase Clostridium sp . WP _ 050606428 15 directed evolution ) can be screened using a high throughput butyrate kinase Clostridium botulinum WP _ 012048334 assay , namely overexpressing Ptb - Buk enzyme pairs in E . butyrate kinase Clostridium botulinum WP _ 012343352 butyrate kinase Clostridium botulinum WP 003401518 coli , adding a test substrate , and screening for ATP produc butyrate kinase Clostridium argentinense WP _ 039635972 tion with a bioluminescence assay . The assay can use the butyrate kinase Clostridium botulinum WP 003357547 well - established practice of correlating ATP concentration butyrate kinase Clostridium hydrogeniformans WP _ 027633205 20 with firefly luciferase enzyme bioluminescence . The ame butyrate kinase Clostridium botulinum WP 033066487 butyrate kinase Roseburia sp . nability of this assay to multi - well plate formats would CDF44202 butyrate kinase Lachnospiraceae bacterium WP 027111008 facilitate efficient screening of substrate preference across butyrate kinase Clostridium sp . CDA68344 new Ptb - Buk combinations ( FIG . 33 ) . butyrate kinase Lachnospiraceae bacterium WP 022782491 By screening for ATP production rather than depletion of butyrate kinase Clostridium botulinum WP 012101111 25 substrate or accumulation of product , the assay avoids butyrate kinase Clostridium carboxidivorans WP 007063155 butyrate kinase Clostridium botulinum WP 041346556 measuring spontaneous hydrolysis of the CoA group . How butyrate kinase Clostridium drakei WP _ 032078801 ever , an alternative approach described in literature , is to use butyrate kinase Clostridium sp . WP _ 032120462 free CoA can be measured using the established assay using butyrate kinase Clostridium sp . WP _ 053468897
Firmicutes bacterium butyrate kinase CCZ27888 Ellman ' s reagent ( 5 , 5 ' - dithiobis - ( 2 - nitrobenzoic acid ) or butyrate kinase Clostridium sp . WP _ 035306569 30 DTNB ) ( Thompson , Appl Environ Microbiol , 56 : 607 - 613 , butyrate kinase Coprococcus comes CDB84787 1990 . ) in order to estimate the coupling efficiency of the butyrate kinase Clostridium sp . WP _ 035292410 Ptb - Buk reactions ( FIG . 33 ) . Acyl - CoAs and corresponding butyrate kinase Clostridium sp . CCX89275 free acids and phospho - intermediates can also be measured Clostridium sp . butyrate kinase WP 040212963 butyrate kinase Clostridium pasteurianum WP _ 003445697 during the validation phase using LC - MS / MS . butyrate kinase Clostridium sp . WP 053242610 35 In a high - throughput screening approach , it is difficult butyrate kinase Lachnospiraceae bacterium WP _ 016299320 gather kinetic data due to the labor involved in protein butyrate kinase Lachnospiraceae bacterium WP 022785085 butyrate kinase Lachnospiraceae bacterium WP 016281561 quantification . Instead , for each preparation of E . coli lysate butyrate kinase Eubacterium sp . CDA28786 containing Ptb - Buk enzymes , the activity against each sub butyrate kinase Clostridium scatologenes WP 029160342 strate of interest ( measured as luminescence per unit time ) butyrate kinase Lachnospiraceae bacterium WP 016228168 40 can be compared to the activity against the positive control butyrate kinase Clostridium pasteurianum WP _ 015617429 butyrate kinase Clostridium algidicarnis WP 029451332 substrate ( butyryl - CoA ) and against acetyl - CoA ( the physi butyrate kinase Lachnospiraceae bacterium WP 016293388 ological substrate that will likely provide the greatest com butyrate kinase Clostridium sulfidigenes WP _ 035133396 petition for enzyme active sites against target acyl - CoA ) . butyrate kinase Clostridium tetani WP 011100666 In order to ensure that the assay is not biased due to native butyrate kinase Clostridium tetanomorphum WP 035147567 butyrate kinase Subdoligranulum variabile wp1007045878 45 phosphotransacetylase ( Pta ) and / or acetate kinase ( Ack ) butyrate kinase Eubacterium sp . CCZ03826 activity , the assay can also be evaluated in an E . coli strain butyrate kinase Firmicutes bacterium CDF07483 where pta and / or ack genes have been knocked out . butyrate kinase Eubacterium sp . CDB13677 Production of Acetone and Isopropanol butyrate kinase Clostridium sp . WP 008400594 butyrate kinase Clostridium tetani WP _ 023439552 Acetone and isopropanol are important industrial solvents butyrate kinase Closfridiales bacterium WP 022787536 50 with a combined market size of 8 million tons and a global butyrate kinase Lachnospiraceae bacterium WP 027434709 market value of $ 8 . 5 - 11 billion . In addition , acetone and butyrate kinase Firmicutes bacterium CCY23249 isopropanol are precursors to valuable downstream prod butyrate kinase Clostridium acetobutylicum WP _ 010966356 ucts , including polymethyl methacrylate ( PMMA ) , which
has a global market value of $ 7 billion , isobutylene , which In a preferred embodiment , the butyrate kinase is Buk 55 has a global market value of $ 25 - 29 billion , and propylene ,
from Clostridium acetobutylicum ( WP _ 010966356 ; SEQ ID which has a global market value of $ 125 billion . Addition NO : 90 ) or Clostridium beijerinckii ( WP _ 011967556 ; SEQ ally , a route from acetone to jet fuel has recently been ID NO : 91 ) ( WP 017209677 ; SEQ ID NO : 92 ) reported . Currently , industrial acetone production is directly ( WP _ 026886638 ; SEQ ID NO : 93 ) ( WP _ 041893502 ; SEQ linked to petrochemical phenol production , as it is a by ID NO : 94 ) . Clostridium autoethanogenum , Clostridium 60 product of the cumene process . Around 92 % of acetone ljungdahlii , and Clostridium ragsdalei do not natively con - output by volume is a co - product of phenol production from tain butyrate kinase . cumene . This has significant implications on both environ
Since Ptb - Buk has been shown to function on a broad ment and market . In the cumene process , per mol phenol range of substrates it is reasonable to assume that if Ptb - Buk produced one mol of sodium sulfite accumulates posing a does not exhibit any activity and a desired substrate it can be 65 serious waste management problem and a challenge to engineered to achieve activity on the substrate in question . natural environments and human health . The world market A strategy could be ( but would not be limited to ) rational demand for phenol is expected to stagnate or decline , while

US 9 , 957 , 531 B1 37 38
the demand for acetone is predicted to rise . Alternative Clostridium autoethanogenum , Clostridium ljungdahlii , or phenol production routes from direct oxidation of benzene Clostridium ragsdalei ) , the microorganism may be modified are in development and expected to commercialize soon ; to knock down or knock out the expression of primary : this could result in a complete elimination of acetone secondary alcohol dehydrogenase ( e . g . , by disrupting the production . 5 gene encoding the primary : secondary alcohol dehydroge
Acetone has been produced at industrial scale for almost nase ) , such that the microorganism produces acetone with 100 years , as a by - product of butanol in ABE fermentation out converting it to isopropanol ( WO 2015 / 085015 ) . While industrial ABE fermentation declined in the second In one embodiment , the microorganism may comprise half of the 20th century due to low oil prices and high sugar more than one pathway for the production of acetone . costs , it has recently revived , with several commercial plants 10 The invention provides a microorganism capable of pro built during the last few years . Multiple groups have also ducing isopropanol or precursors thereof from a substrate . demonstrated acetone production from sugar in heterolo - The invention further provides a method of producing gous hosts that express the corresponding enzymes from isopropanol or precursors thereof by culturing such a micro ABE fermentation organisms , in particular E . coli and yeast organism in the presence of a substrate . In preferred embodi through metabolic engineering and synthetic biology 15 ments , the microorganism is derived from a parental micro approaches by several academic groups . However , low organism selected from the group consisting of Clostridium yields and high costs associated the pre - treatment needed to autoethanogenum , Clostridium ljungdahlii , or Clostridium release the polysaccharide - component of biomass make the ragsdalei . However , the microorganism may also be derived production of acetone via standard fermentation uneco - from an entirely different microorganism , e . g . , Eschericia nomic as current biochemical conversion technologies do 20 coli . The enzymatic pathways described for the production not utilize the lignin component of biomass , which can of isopropanol may comprise endogenous enzymes and , constitute up to 40 % of this material . where endogenous enzyme activity is absent or low , exog
The invention provides a microorganism capable of pro - enous enzymes . ducing acetone or precursors thereof from a substrate . The Isopropanol via steps 1 , 2 , 3 , and 4 : In one embodiment , invention further provides a method of producing acetone or 25 the invention provides a microorganism comprising precursors thereof by culturing such a microorganism in the enzymes for steps 1 , 2 , 3 , and 4 , whereby the microorganism presence of a substrate . In preferred embodiments , the is capable of producing isopropanol or precursors thereof microorganism is derived from a parental microorganism from a substrate , such as a gaseous substrate . Typically , at selected from the group consisting of Clostridium autoetha - least one of the enzymes in this pathway is exogenous to the nogenum , Clostridium ljungdahlii , or Clostridium ragsdalei . 30 microorganism . In a preferred embodiment , step 2 is cata However , the microorganism may also be derived from an lyzed by Ptb - Buk . Exemplary types and sources of enzymes entirely different microorganism , e . g . , Eschericia coli . The for steps 1 , 2 , 3 , and 4 are described elsewhere in this enzymatic pathways described for the production of acetone application . If the microorganism is derived from a parental may comprise endogenous enzymes and , where endogenous microorganism that natively contains a primary : secondary enzyme activity is absent or low , exogenous enzymes . 35 alcohol dehydrogenase capable of converting acetone to
Acetone via steps 1 , 2 , and 3 : In one embodiment , the isopropanol ( step 4 ) ( e . g . , Clostridium autoethanogenum , invention provides a microorganism comprising enzymes Clostridium ljungdahlii , or Clostridium ragsdalei ) , intro for steps 1 , 2 , and 3 , whereby the microorganism is capable duction of an exogenous enzyme for step 4 is not required of producing acetone or precursors thereof from a substrate , to produce isopropanol . However , modification of the such as a gaseous substrate . Typically , at least one of the 40 microorganism , for example , to overexpress a native prima enzymes in this pathway is exogenous to the microorganism . ry : secondary alcohol dehydrogenase may result in enhanced In a preferred embodiment , step 2 is catalyzed by Ptb - Buk . production of isopropanol . Exemplary types and sources of enzymes for steps 1 , 2 , and Isopropanol via steps 1 , 13 , 14 , 15 , 3 , and 4 : In one 3 are described elsewhere in this application . If the micro - embodiment , the invention provides a microorganism com organism is derived from a parental microorganism that 45 prising enzymes for steps 1 , 13 , 14 , 15 , 3 , and 4 , whereby the natively contains a primary : secondary alcohol dehydroge microorganism is capable of producing isopropanol or pre nase capable of converting acetone to isopropanol ( step 4 ) cursors thereof from a substrate , such as a gaseous substrate . ( e . g . , Clostridium autoethanogenum , Clostridium ljungdah - Typically , at least one of the enzymes in this pathway is lii , or Clostridium ragsdalei ) , the microorganism may be exogenous to the microorganism . In a preferred embodi modified to knock down or knock out the expression of 50 ment , step 14 is catalyzed by Ptb - Buk . Exemplary types and primary : secondary alcohol dehydrogenase ( e . g . , by disrupt - sources of enzymes for steps 1 , 13 , 14 , 15 , 3 , and 4 are ing the gene encoding the primary : secondary alcohol dehy described elsewhere in this application . If the microorgan drogenase ) , such that the microorganism produces acetone ism is derived from a parental microorganism that natively without converting it to isopropanol ( WO 2015 / 085015 ) . contains a primary : secondary alcohol dehydrogenase
Acetone via steps 1 , 13 , 14 , 15 , and 3 : In one embodiment , 55 capable of converting acetone to isopropanol ( step 4 ) ( e . g . , the invention provides a microorganism comprising exog - Clostridium autoethanogenum , Clostridium ljungdahlii , or enous enzymes for steps 1 , 13 , 14 , 15 , and 3 , whereby the Clostridium ragsdalei ) , introduction of an exogenous microorganism is capable of producing acetone or precur - enzyme for step 4 is not required to produce isopropanol . sors thereof from a substrate , such as a gaseous substrate . However , modification of the microorganism , for example , Typically , at least one of the enzymes in this pathway is 60 to overexpress a native primary : secondary alcohol dehydro exogenous to the microorganism . In a preferred embodi - genase may result in enhanced production of isopropanol . ment , step 14 is catalyzed by Ptb - Buk . Exemplary types and In one embodiment , the microorganism may comprise sources of enzymes for steps 1 , 13 , 14 , 15 , and 3 are more than one pathway for the production of isopropanol . described elsewhere in this application . If the microorgan - Production of Isobutylene ism is derived from a parental microorganism that natively 65 Isobutylene is a major chemical building block with a contains a primary : secondary alcohol dehydrogenase market size of over 15 million tons and a global market capable of converting acetone to isopropanol ( step 4 ) ( e . g . , value of $ 25 - 29 billion . Beyond its use in chemistry and as

US 9 , 957 , 531 B1 39 40
a fuel additive ( 15 Mt / yr ) , isobutylene may be converted to Production of 3 - Hydroxybutyrate isooctane , a high performance , drop - in fuel for gasoline cars . 3 - Hydroxybutyrate ( 3 - HB ) is a four carbon carboxylic Global Bioenergies has filed patent applications on the acid in the family of betahydroxy acids . 3 - hydroxybutyrate fermentative production of isobutene ( i . e . , isobutylene ) from is a cosmetic ingredient for oily skin clarification , an inter acetone , but none of the disclosed routes involve Ptb - Buk 5 mediate for anti - aging cream formulations , an intermediate ( WO 2010 / 001078 ; EP 2295593 ; WO 2011 / 076691 ; van for polyhydroxybutyrate ( PHB ) , a biodegradable polymer Leeuwen , Appl Microbiol Biotechnol , 93 : 1377 - 1387 , 2012 ) . resin , and co - monomer with other polyhydroxy acids for The invention provides a microorganism capable of pro novel bioplastics . Additionally , 3 - hydroxybutyrate has spe ducing isobutylene or precursors thereof from a substrate . cialty applications in biocompatible and biodegradable The invention further provides a method of producing 10 nanocomposites , particularly for medical implants , interme isobutylene or precursors thereof by culturing such a micro diate for C3 / C4 chemicals , chiral building blocks , and fine organism in the presence of a substrate . In preferred embodi chemicals . Although the production of ( R ) - and ( S ) - 3 ments , the microorganism is derived from a parental micro hydroxybutyrate by recombinant E . coli grown on glucose , organism selected from the group consisting of Clostridium autoethanogenum , Clostridium ljungdahlii , or Clostridium 15 the production of 3 - hydroxybutyrate has not been demon ragsdalei . However , the microorganism may also be derived strated from microorganisms grown on gaseous substrates from an entirely different microorganism , e . g . , Eschericia ( Tseng , Appl Environ Microbiol , 75 : 3137 - 3145 , 2009 ) . coli . The enzymatic pathways described for the production Notably , the system previously demonstrated in E . coli was of isobutylene may comprise endogenous enzymes and , not directly transferable to acetogens , including C . autoetha where endogenous enzyme activity is absent or low , exog - 20 nogenum , due to the presence of native thioesterases in enous enzymes . acetogens . Although E . coli also has a thioesterase TesB that
FIG . 1 shows two alternative routes to isobutylene . The can act on 3 - HB - COA , Tseng showed that background first involves the production of isobutylene via steps 1 , 2 , 3 , activity is minimal ( < 0 . 1 g / L ) . While in E . coli production 5 , and 6 . The second involves the production of isobutylene of stereopure isomers were reported , the inventors surpris via steps 1 , 2 , 3 , 7 , 8 , and 6 . Steps 2 and 8 may be catalyzed 25 ingly found that a mix of isomers were produced in C . by Ptb - Buk . Accordingly , each route may involve Ptb - Buk . autoethanogenum . Without being bound to this theory , this
Isobutylene via steps 1 , 2 , 3 , 5 , and 6 : In one embodiment , is likely a result of native isomerase activity . This enables the invention provides a microorganism comprising the combination of an ( S ) - specific 3 - hydroxybutyryl - CoA enzymes for steps 1 , 2 , 3 , 5 , and 6 , whereby the microor dehydrogenase ( Hbd ) to be combined with the ( R ) - specific ganism is capable of producing isobutylene or precursors 30 Ptb - Buk for optimized production . To produce stereopure thereof from a substrate , such as a gaseous substrate . Typi isomers , this activity can be knocked - out . Taken together , it cally , at least one of the enzymes in this pathway is exog this invention enables to produce several g / L of 3 - HB enous to the microorganism . In a preferred embodiment , compared to low production in E . coli and using Ptb - Buk step 2 is catalyzed by Ptb - Buk . Exemplary types and sources of enzymes for steps 1 , 2 , 3 , 5 , and 6 are described elsewhere 35 any 5 any combination of ( R ) - or ( S ) - specific 3 - hydroxybutyryl in this application . If the microorganism is derived from a COA dehydrogenase and native Clostridium autoethanoge parental microorganism that natively contains a primary : num thioesterase . secondary alcohol dehydrogenase capable of converting The invention provides a microorganism capable of pro acetone to isopropanol ( step 4 ) ( e . g . , Clostridium autoetha - ducing 3 - hydroxybutyrate or precursors thereof from a sub nogenum , Clostridium ljungdahlii , or Clostridium ragsda - 40 strate . The invention further provides a method of producing lei ) , the microorganism may be modified to knock down or 3 - hydroxybutyrate or precursors thereof by culturing such a knock out the expression of primary : secondary alcohol microorganism in the presence of a substrate . In preferred dehydrogenase ( e . g . , by disrupting the gene encoding the embodiments , the microorganism is derived from a parental primary : secondary alcohol dehydrogenase ) to prevent the microorganism selected from the group consisting of conversion of acetone to isopropanol and maximize the 45 Clostridium autoethanogenum , Clostridium ljungdahlii , or conversion of acetone to isobutylene . Clostridium ragsdalei . However , the microorganism may
Isobutylene via steps 1 , 2 , 3 , 7 , 8 , and 6 : In one embodi - also be derived from an entirely different microorganism , ment , the invention provides a microorganism comprising e . g . , Eschericia coli . The enzymatic pathways described for enzymes for steps 1 , 2 , 3 , 7 , 8 , and 6 , whereby the micro - the production of 3 - hydroxybutyrate may comprise endog organism is capable of producing isobutylene or precursors 50 enous enzymes and , where endogenous enzyme activity is thereof from a substrate , such as a gaseous substrate . Typi - absent or low , exogenous enzymes . cally , at least one of the enzymes in this pathway is exog - FIG . 1 shows two alternative routes to 3 - hydroxybutyrate . enous to the microorganism . In a preferred embodiment . The first involves the production of 3 - hydroxybutyrate via step 2 and / or step 8 are catalyzed by Ptb - Buk . Exemplary steps 1 , 2 , and 15 . The second involves the production of types and sources of enzymes for steps 1 , 2 , 3 , 7 , 8 , and 6 55 3 - hydroxybutyrate via steps 1 , 13 , and 14 . Steps 2 and 14 are described elsewhere in this application . If the microor - may be catalyzed by Ptb - Buk . Accordingly , each route may ganism is derived from a parental microorganism that involve Ptb - Buk . In one embodiment , the microorganism natively contains a primary : secondary alcohol dehydroge may comprise more than one pathway for the production of nase capable of converting acetone to isopropanol ( step 4 ) 3 - hydroxybutyrate , wherein Ptb - Buk may catalyze more ( e . g . , Clostridium autoethanogenum , Clostridium ljungdah - 60 than one step ( e . g . , steps 2 and 14 ) . lii , or Clostridium ragsdalei ) , the microorganism may be 3 - Hydroxybutyrate via steps 1 , 2 , and 15 : In one embodi modified to knock down or knock out the expression of ment , the invention provides a microorganism comprising primary : secondary alcohol dehydrogenase ( e . g . , by disrupt enzymes for steps 1 , 2 , and 15 , whereby the microorganism ing the gene encoding the primary : secondary alcohol dehy is capable of producing 3 - hydroxybutyrate or precursors drogenase ) to prevent the conversion of acetone to isopro - 65 thereof from a substrate , such as a gaseous substrate . Typi panol and maximize the conversion of acetone to cally , at least one of the enzymes in this pathway is exog isobutylene . enous to the microorganism . In a preferred embodiment ,

US 9 , 957 , 531 B1 42
step 2 is catalyzed by Ptb - Buk . Exemplary types and sources prising enzymes for steps 1 , 13 , 14 , 16 , and 17 , whereby the of enzymes for steps 1 , 2 , and 15 are described elsewhere in microorganism is capable of producing 1 , 3 - butanediol or this application . precursors thereof from a substrate , such as a gaseous
3 - Hydroxybutyrate via steps 1 , 13 , and 14 : In one substrate . Typically , at least one of the enzymes in this embodiment , the invention provides a microorganism com - 5 pathway is exogenous to the microorganism . In a preferred prising enzymes for steps 1 , 13 , and 14 , whereby the embodiment , step 14 is catalyzed by Ptb - Buk . Exemplary microorganism is capable of producing 3 - hydroxybutyrate types and sources of enzymes for steps 1 , 13 , 14 , 16 , and 17 or precursors thereof from substrate , such as a gaseous are described elsewhere in this application . substrate . Typically , at least one of the enzymes in this 1 , 3 - Butanediol via steps 1 , 13 , 18 , and 17 : In one embodi pathway is exogenous to the microorganism . In a preferred 10 ment , the invention provides a microorganism comprising embodiment , step 14 is catalyzed by Ptb - Buk . Exemplary enzymes for steps 1 , 13 , 18 , and 17 , whereby the microor types and sources of enzymes for steps 1 , 13 , and 14 are ganism is capable of producing 1 , 3 - butanediol or precursors described elsewhere in this application . thereof from a substrate , such as a gaseous substrate ( FIG . Production of 1 , 3 - Butanediol 11 ) . Typically , at least one of the enzymes in this pathway is
1 , 3 - Butanediol ( 1 , 3 - BDO ) is commonly used as a solvent 15 exogenous to the microorganism . Exemplary types and for food flavoring agents and is a co - monomer used in sources of enzymes for steps 1 , 13 , 18 , and 17 are described certain polyurethane and polyester resins . More importantly , elsewhere in this application . A similar route has been 1 , 3 - butanediol may be catalytically converted to 1 , 3 - buta - demonstrated in E . coli , but not in acetogens such as diene ( Makshina , Chem Soc Rev , 43 : 7917 - 7953 , 2014 ) . Clostridium autoethanogenum , Clostridium ljungdahlii , and Butadiene is used to produce rubber , plastics , lubricants , 20 Clostridium ragsdalei ( Kataoka , J Biosci Bioeng , 115 : 475 latex , and other products . While much of the butadiene 480 , 2013 ) . Although the use of Ptb - Buk results in the produced today is used for the rubber in automobile tires , it production of ( R ) - 1 , 3 - butanediol , this route , which does not can also be used to produce adiponitrile , which can be used require the use of Ptb - Buk , may result in the production of in the manufacture of nylon 6 , 6 . Global demand for buta - ( S ) - 1 , 3 - butanediol . diene is on the rise . In 2011 , there was an estimated 10 . 5 25 Production of 2 - Hydroxyisobutyrate million tons of demand , valued at $ 40 billion . 2 - Hydroxyisobutyrate ( 2 - HIB ) is a four carbon carboxylic
The invention provides a microorganism capable of pro - acid that may serve as a building block for many types of ducing 1 , 3 - butanediol or precursors thereof from a substrate . polymers . The methyl ester of methacrylic acid , which can The invention further provides a method of producing be synthesized by dehydration of 2 - hydroxyisobutyrate or 1 , 3 - butanediol or precursors thereof by culturing such a 30 via the corresponding amide , is polymerized to polymeth microorganism in the presence of substrate . In preferred ylmethacrylate ( PMMA ) for the production of acrylic glass , embodiments , the microorganism is derived from a parental durable coatings , and inks . For this compound alone , the microorganism selected from the group consisting of global market exceeds 3 million tons . Other branched C4 Clostridium autoethanogenum , Clostridium ljungdahlii , or carboxylic acids , e . g . , chloro - and amino - derivatives of Clostridium ragsdalei . However , the microorganism may 35 2 - hydroxyisobutyrate , as well as isobutylene glycol and its also be derived from an entirely different microorganism , oxide , are also used in polymers and for many other appli e . g . , Eschericia coli . The enzymatic pathways described for c ations . the production of 1 , 3 - butanediol may comprise endogenous The stereospecificity of the Ptb - Buk system is particularly enzymes and , where endogenous enzyme activity is absent useful in overcoming the limitations of the current state of or low , exogenous enzymes . 40 art with respect to the production of 2 - hydroxyisobutyrate .
In certain embodiments , the microorganism may produce Both Ptb - Buk and thioesterases are promiscuous , such that 1 , 3 - butanediol without co - production of ethanol ( or with side activity with 3 - hydroxybutyryl - CoA may divert production of only a small amount of ethanol , e . g . , less than resources away from target pathways for the production of 0 . 1 - 1 . 0 g / L ethanol or less than 1 - 10 g / L ethanol ) . 2 - hydroxyisobutyryl - CoA ( see , e . g . , FIG . 1 and FIG . 8 ) .
FIG . 1 shows three alternative routes to 1 , 3 - butanediol . 45 However , Ptb - Buk is able to distinguish between stereoiso The first involves the production of 1 , 3 - butanediol via steps mers and will act on ( R ) - 3 - hydroxybutyryl - CoA , but not on 1 , 2 , 15 , 16 , and 17 . The second involves the production of ( S ) - 3 - hydroxybutyryl - CoA . In contrast , thioesterases are not 1 , 3 - butanediol via steps 1 , 13 , 14 , 16 , and 17 . The third able to distinguish between 3 - hydroxybutyryl - CoA stereoi involves the production of 1 , 3 - butanediol via steps 1 , 13 , 18 , somers . In a preferred embodiment , an ( S ) - specific and 17 . Steps 2 and 14 may be catalyzed by Ptb - Buk . 50 acetoacetyl - CoA hydratase ( EC 4 . 2 . 1 . 119 ) ( step 13 ) is cho Accordingly , at least the first and second routes may involve sen in combination with the Ptb - Buk ( step 20 ) to avoid Ptb - Buk . In one embodiment , the microorganism may com - losses to 3 - hydroxybutyrate and maximize 2 - hydroxyisobu prise more than one pathway for the production of 1 , 3 tyrate yield ( FIG . 8 ) . The ( S ) - specific form of 3 - hydroxy butanediol . In a related embodiment , the Ptb - Buk may butyryl - CoA is also the preferred substrate for the 2 - hy catalyze more than one step ( e . g . , steps 2 and 14 ) . 55 droxyisobutyryl - CoA mutase ( EC 5 . 4 . 99 . - ) ( step 19 )
1 , 3 - Butanediol via steps 1 , 2 , 15 , 16 , and 17 : In one ( Yaneva , J Biol Chem , 287 : 15502 - 15511 , 2012 ) . embodiment , the invention provides a microorganism com - The invention provides a microorganism capable of pro prising enzymes for steps 1 , 2 , 15 , 16 , and 17 , whereby the ducing 2 - hydroxyisobutyrate or precursors thereof from a microorganism is capable of producing 1 , 3 - butanediol or substrate . The invention further provides a method of pro precursors thereof from a substrate , such as a gaseous 60 ducing 2 - hydroxyisobutyrate or precursors thereof by cul substrate . Typically , at least one of the enzymes in this turing such a microorganism in the presence of a substrate . pathway is exogenous to the microorganism . In a preferred In preferred embodiments , the microorganism is derived embodiment , step 2 is catalyzed by Ptb - Buk . Exemplary from a parental microorganism selected from the group types and sources of enzymes for steps 1 , 2 , 15 , 16 , and 17 consisting of Clostridium autoethanogenum , Clostridium are described elsewhere in this application . 65 ljungdahlii , or Clostridium ragsdalei . However , the micro
1 , 3 - Butanediol via steps 1 , 13 , 14 , 16 , and 17 : In one organism may also be derived from an entirely different embodiment , the invention provides a microorganism com - microorganism , e . g . , Eschericia coli . The enzymatic path

US 9 , 957 , 531 B1 43 44
ways described for the production of 2 - hydroxyisobutyrate In one embodiment , the microorganism may comprise may comprise endogenous enzymes and , where endogenous more than one pathway for the production of adipic acid . enzyme activity is absent or low , exogenous enzymes . Production of 1 , 3 - Hexanediol
2 - Hydroxyisobutyrate via steps 1 , 13 , 19 , and 20 : In one The invention provides a microorganism capable of pro embodiment , the invention provides a microorganism com - 5 ducing 1 , 3 - hexanediol or precursors thereof from a substrate prising enzymes for steps 1 , 13 , 19 , and 20 , whereby the ( FIG . 35 ) . The invention further provides a method of microorganism is capable of producing 2 - hydroxyisobu - producing 1 , 3 - hexanediol or precursors thereof by culturing tyrate or precursors thereof from a substrate , such as a such a microorganism in the presence of a substrate . In gaseous substrate . Typically , at least one of the enzymes in preferred embodiments , the microorganism is derived from this pathway is exogenous to the microorganism . In a 10 a parental microorganism selected from the group consisting preferred embodiment , step 20 is catalyzed by Ptb - Buk . of Clostridium autoethanogenum , Clostridium ljungdahlii , Exemplary types and sources of enzymes for steps 1 , 13 , 19 , or Clostridium ragsdalei . However , the microorganism may and 20 are described elsewhere in this application . also be derived from an entirely different microorganism ,
In certain embodiments , the invention also provides a e . g . , Eschericia coli . The enzymatic pathways described for microorganism capable of producing 2 - hydroxybutyrate 15 the production of 1 , 3 - hexanediol may comprise endogenous ( 2 - HB ) or precursors thereof from a substrate . The invention enzymes and , where endogenous enzyme activity is absent further provides a method of producing 2 - hydroxybutyrate or low , exogenous enzymes . or precursors thereof by culturing such a microorganism in The pathways depicted in FIG . 35 begin with 3 - hydroxy the presence of a substrate . Without wishing to be bound by butyryl - CoA , which may be produced via steps 1 and 13 , as any particular theory , the inventors believe the observed 20 depicted in FIG . 1 . production of 2 - hydroxybutyrate is attributable to nonspe 1 , 3 - Hexanediol via steps 1 , 13 , 27 , 31 , 32 , 36 , 37 , 38 , and cific mutase activity in microorganisms such as Clostridium 39 : In one embodiment , the invention provides a microor autoethanogenum , Clostridium ljungdahlii , and Clostridium ganism comprising enzymes for steps 1 , 13 , 27 , 31 , 32 , 36 , ragsdalei . 37 , 38 , and 39 , whereby the microorganism is capable of Production of Adipic Acid 25 producing 1 , 3 - hexanediol or precursors thereof from a sub
Adipic acid is the most important dicarboxylic acid with strate , such as a gaseous substrate . Typically , at least one of an estimated market of greater US $ 4 . 5 billion with about the enzymes in this pathway is exogenous to the microor 2 . 5 billion kgs produced annually . Over 60 % of produced ganism . In a preferred embodiment , step 37 is catalyzed by adipic acid is being used as monomer precursor for the Ptb - Buk . Exemplary types and sources of enzymes for steps production of nylon and the global market for adipic acid is 30 1 , 13 , 27 , 31 , 32 , 36 , 37 , 38 , and 39 are described elsewhere expected to reach US $ 7 . 5 billion by 2019 . Currently , adipic in this application . acid is almost exclusively produced petrochemically , e . g . by Production of 3 - Methyl - 2 - Butanol carbonylation of butadiene . The invention provides a microorganism capable of pro
The invention provides a microorganism capable of pro - ducing 3 - methyl - 2 - butanol or precursors thereof from a ducing adipic acid or precursors thereof from a substrate 35 substrate ( FIG . 35 ) . The invention further provides a method ( FIG . 34 ) . The invention further provides a method of of producing 3 - methyl - 2 - butanol or precursors thereof by producing adipic acid or precursors thereof by culturing culturing such a microorganism in the presence of a sub such a microorganism in the presence of a substrate . In strate . In preferred embodiments , the microorganism is preferred embodiments , the microorganism is derived from derived from a parental microorganism selected from the a parental microorganism selected from the group consisting 40 group consisting of Clostridium autoethanogenum , of Clostridium autoethanogenum , Clostridium ljungdahlii , Clostridium ljungdahlii , or Clostridium ragsdalei . However , or Clostridium ragsdalei . However , the microorganism may the microorganism may also be derived from an entirely also be derived from an entirely different microorganism , different microorganism , e . g . , Eschericia coli . The enzy e . g . , Eschericia coli . The enzymatic pathways described for matic pathways described for the production of 3 - methyl the production of adipic acid may comprise endogenous 45 2 - butanol may comprise endogenous enzymes and , where enzymes and , where endogenous enzyme activity is absent endogenous enzyme activity is absent or low , exogenous or low , exogenous enzymes . enzymes .
Adipic acid via steps 22 , 23 , 24 , 25 , and 26 : In one The pathways depicted in FIG . 35 begin with 3 - hydroxy embodiment , the invention provides a microorganism com - butyryl - CoA , which may be produced via steps 1 and 13 , as prising enzymes for steps 22 , 23 , 24 , 25 , and 26 , whereby the 50 depicted in FIG . 1 . microorganism is capable of producing adipic acid or pre 3 - Methyl - 2 - butanol via steps 1 , 13 , 27 , 31 , 32 , 33 , 34 , and cursors thereof from a substrate , such as a gaseous substrate . 35 : In one embodiment , the invention provides a microor Typically , at least one of the enzymes in this pathway is ganism comprising enzymes for steps 1 , 13 , 27 , 31 , 32 , 33 , exogenous to the microorganism . In a preferred embodi - 34 , and 35 , whereby the microorganism is capable of pro ment , step 26 is catalyzed by Ptb - Buk . Exemplary types and 55 ducing 3 - methyl - 2 - butanol or precursors thereof from a sources of enzymes for steps 22 , 23 , 24 , 25 , and 26 are substrate , such as a gaseous substrate . Typically , at least one described elsewhere in this application . of the enzymes in this pathway is exogenous to the micro
Adipic acid via steps 21 , 22 , 23 , 24 , 25 , and 26 : In one organism . In a preferred embodiment , step 33 is catalyzed by embodiment , the invention provides a microorganism com - Ptb - Buk . Exemplary types and sources of enzymes for steps prising enzymes for steps 21 , 22 , 23 , 24 , 25 , and 26 , 60 1 , 13 , 27 , 31 , 32 , 33 , 34 , and 35 are described elsewhere in whereby the microorganism is capable of producing adipic this application . acid or precursors thereof from a substrate , such as a gaseous Production of 2 - Buten - 1 - ol substrate . Typically , at least one of the enzymes in this The invention provides a microorganism capable of pro pathway is exogenous to the microorganism . In a preferred ducing 2 - buten - 1 - ol or precursors thereof from a substrate embodiment , step 26 is catalyzed by Ptb - Buk . Exemplary 65 ( FIG . 35 ) . The invention further provides a method of types and sources of enzymes for steps 21 , 22 , 23 , 24 , 25 , producing 2 - buten - 1 - ol or precursors thereof by culturing and 26 are described elsewhere in this application . such a microorganism in the presence of a substrate . In

45 US 9 , 957 , 531 B1
46 preferred embodiments , the microorganism is derived from as a gaseous substrate . Typically , at least one of the enzymes a parental microorganism selected from the group consisting in this pathway is exogenous to the microorganism . In a of Clostridium autoethanogenum , Clostridium ljungdahlii , preferred embodiment , step 44 is catalyzed by Ptb - Buk . or Clostridium ragsdalei . However , the microorganism may Exemplary types and sources of enzymes for steps 1 , 40 , 41 , also be derived from an entirely different microorganism , 5 42 , 43 , 44 , 45 , and 46 are described elsewhere in this e . g . , Eschericia coli . The enzymatic pathways described for application . the production of 2 - buten - 1 - ol may comprise endogenous Isoamyl alcohol via steps 1 , 40 , 41 , 42 , 43 , 47 and 46 : In enzymes and , where endogenous enzyme activity is absent one embodiment , the invention provides a microorganism or low , exogenous enzymes . comprising enzymes for steps 1 , 40 , 41 , 42 , 43 , 47 and 46 , The pathways depicted in FIG . 35 begin with 3 - hydroxy - 10 whereby the microorganism is capable of producing isoamyl butyryl - CoA , which may be produced via steps 1 and 13 , as alcohol or precursors thereof from a substrate , such as a depicted in FIG . 1 .
2 - Buten - 1 - ol via steps 1 , 13 , 27 , 28 , 29 , and 30 : In one gaseous substrate . Typically , at least one of the enzymes in embodiment , the invention provides a microorganism com this pathway is exogenous to the microorganism . Exemplary prising enzymes for steps 1 , 13 , 27 , 28 , 29 , and 30 , whereby 15 types and sources of enzymes for steps 1 , 40 , 41 , 42 , 43 , 47 the microorganism is capable of producing 2 - buten - 1 - ol or and 46 are described elsewhere in this application . precursors thereof from a substrate , such as a gaseous In one embodiment , the microorganism may comprise substrate . Typically , at least one of the enzymes in this more than one pathway for the production of isoamyl pathway is exogenous to the microorganism . In a preferred alcohol . embodiment , step 28 is catalyzed by Ptb - Buk . Exemplary 20 Production of Additional Products types and sources of enzymes for steps 1 , 13 , 27 , 28 , 29 , and The invention provides a microorganism comprising 30 are described elsewhere in this application . exogenous Ptb - Buk and exogenous or endogenous alde Production of Isovalerate hyde : ferredoxin oxidoreductase ( AOR ) . Such a microorgan
The invention provides a microorganism capable of pro - ism may produce , for example , 1 - propanol , 1 - butanol , ducing isovalerate or precursors thereof from a substrate 25 1 - hexanol , and 1 - octanol or precursors thereof from acetyl ( FIG . 36 ) . The invention further provides a method of COA generated , for example , from a gaseous substrate ( FIG . producing isovalerate or precursors thereof by culturing 32 ) . The invention further provides a method of producing such a microorganism in the presence of a substrate . In 1 - propanol , 1 - butanol , 1 - hexanol , and 1 - octanol or precur preferred embodiments , the microorganism is derived from sors thereof by culturing such a microorganism in the a parental microorganism selected from the group consisting 30 presence of a gaseous substrate . Clostridium autoethanoge of Clostridium autoethanogenum , Clostridium ljungdahlii , num , Clostridium ljungdahlii , and Clostridium ragsdalei or Clostridium ragsdalei . However , the microorganism may natively comprise AOR . However , AOR may be overex also be derived from an entirely different microorganism , pressed in such microorganisms in combination with expres e . g . , Eschericia coli . The enzymatic pathways described for sion of exogenous Ptb - Buk . Alternatively , exogenous AOR the production of isovalerate may comprise endogenous 35 and exogenous Ptb - Buk may be expressed in a microorgan enzymes and , where endogenous enzyme activity is absent ism other than Clostridium autoethanogenum , Clostridium or low , exogenous enzymes . ljungdahlii , and Clostridium ragsdalei , such as Escherichia
Isovalerate via steps 1 , 40 , 41 , 42 , 43 , and 44 : In one coli . embodiment , the invention provides a microorganism com - Production of Precursors and Intermediates prising enzymes for steps 1 , 40 , 41 , 42 , 43 , and 44 , whereby 40 The pathways depicted in FIGS . 1 , 34 , 35 , and 36 may be the microorganism is capable of producing isovalerate or modified to produce precursors or intermediates of the precursors thereof from a substrate , such as a gaseous aforementioned products . In particular , partial enzymatic substrate . Typically , at least one of the enzymes in this pathways for any of the pathways described herein may be pathway is exogenous to the microorganism . In a preferred inserted in a host microorganism to obtain production of embodiment , step 44 is catalyzed by Ptb - Buk . Exemplary 45 precursors or intermediates . types and sources of enzymes for steps 1 , 40 , 41 , 42 , 43 , and 44 are described elsewhere in this application . Definitions and Background Production of Isoamyl Alcohol
The invention provides a microorganism capable of pro - The term “ genetic modification ” or “ genetic engineering ” ducing isoamyl alcohol or precursors thereof from a sub - 50 broadly refers to manipulation of the genome or nucleic strate ( FIG . 36 ) . The invention further provides a method of acids of a microorganism . Likewise , the term " genetically producing isoamyl alcohol or precursors thereof by culturing engineered ” refers to a microorganism comprising a such a microorganism in the presence of a substrate . In manipulated genome or nucleic acids . Methods of genetic preferred embodiments , the microorganism is derived from modification of include , for example , heterologous gene a parental microorganism selected from the group consisting 55 expression , gene or promoter insertion or deletion , nucleic of Clostridium autoethanogenum , Clostridium ljungdahlii , acid mutation , altered gene expression or inactivation , or Clostridium ragsdalei . However , the microorganism may enzyme engineering , directed evolution , knowledge - based also be derived from an entirely different microorganism , design , random mutagenesis methods , gene shuffling , and e . g . , Eschericia coli . The enzymatic pathways described for codon optimization . the production of isoamyl alcohol may comprise endog - 60 “ Recombinant ” indicates that a nucleic acid , protein , or enous enzymes and , where endogenous enzyme activity is microorganism is the product of genetic modification , engi absent or low , exogenous enzymes . neering , or recombination . Generally , the term “ recombi
Isoamyl alcohol via steps 1 , 40 , 41 , 42 , 43 , 44 , 45 , and 46 : nant ” refers to a nucleic acid , protein , or microorganism that In one embodiment , the invention provides a microorganism contains or is encoded by genetic material derived from comprising enzymes for steps 1 , 40 , 41 , 42 , 43 , 44 , 45 , and 65 multiple sources , such as two or more different strains or 46 , whereby the microorganism is capable of producing species of microorganisms . As used herein , the term “ recom isoamyl alcohol or precursors thereof from a substrate , such binant ” may also be used to describe a microorganism that

US 9 , 957 , 531 B1 48
comprises a mutated nucleic acid or protein , including a The term “ energy - generating ” or the like may be used mutated form of an endogenous nucleic acid or protein . interchangeably herein with “ energy - conserving " or the like .
“ Endogenous ” refers to a nucleic acid or protein that is Both of these terms are commonly used in the literature . present or expressed in the wild - type or parental microor - “ Mutated ” refers to a nucleic acid or protein that has been ganism from which the microorganism of the invention is 5 modified in the microorganism of the invention compared to derived . For example , an endogenous gene is a gene that is the wild - type or parental microorganism from which the natively present in the wild - type or parental microorganism microorganism of the invention is derived . In one embodi from which the microorganism of the invention is derived . ment , the mutation may be a deletion , insertion , or substi In one embodiment , the expression of an endogenous gene tution in a gene encoding an enzyme . In another embodi may be controlled by an exogenous regulatory element , such 10 ment , the mutation may be a deletion , insertion , or
substitution of one or more amino acids in an enzyme . as an exogenous promoter . In particular , a " disruptive mutation ” is a mutation that “ Exogenous ” refers to a nucleic acid or protein that is not reduces or eliminates ( i . e . , " disrupts ” ) the expression or present in the wild - type or parental microorganism from activity of a gene or enzyme . The disruptive mutation may which the microorganism of the invention is derived . In one 15 partially inactivate , fully inactivate , or delete the gene or embodiment , an exogenous gene or enzyme may be derived enzyme . The disruptive mutation may be a knockout ( KO ) from a heterologous ( i . e . , different ) strain or species and mutation . The disruptive mutation may be any mutation that introduced to or expressed in the microorganism of the reduces , prevents , or blocks the biosynthesis of a product invention . In another embodiment , an exogenous gene or produced by an enzyme . The disruptive mutation may enzyme may be artificially or recombinantly created and 20 include , for example , a mutation in a gene encoding an introduced to or expressed in the microorganism of the enzyme , a mutation in a genetic regulatory element involved invention . Exogenous nucleic acids may be adapted to in the expression of a gene encoding an enzyme , the integrate into the genome of the microorganism of the introduction of a nucleic acid which produces a protein that invention or to remain in an extra - chromosomal state in the reduces or inhibits the activity of an enzyme , or the intro microorganism of the invention , for example , in a plasmid . 25 duction of a nucleic acid ( e . g . , antisense RNA , siRNA ,
“ Enzyme activity , ” or simply “ activity , ” refers broadly to CRISPR ) or protein which inhibits the expression of an enzymatic activity , including , but not limited , to the activity enzyme . The disruptive mutation may be introduced using of an enzyme , the amount of an enzyme , or the availability any method known in the art . of an enzyme to catalyze a reaction . Accordingly , “ increas Introduction of a disruptive mutation results in a micro ing ” enzyme activity includes increasing the activity of an ofan 30 organism of the invention that produces no target product or
substantially no target product or a reduced amount of target enzyme , increasing the amount of an enzyme , or increasing the availability of an enzyme to catalyze a reaction . Simi product compared to the parental microorganism from
which the microorganism of the invention is derived . For larly , “ decreasing ” enzyme activity includes decreasing the example , the microorganism of the invention may produce activity of an enzyme , decreasing the amount of an enzyme , 25 ne , 35 no target product or at least about 1 % , 3 % , 5 % , 10 % , 20 % , or decreasing the availability of an enzyme to catalyze a 30 % , 40 % , 50 % , 60 % , 70 % , 80 % , 90 % , or 95 % less target reaction . product than the parental microorganism . For example , the
With respect to enzyme activity , a “ substrate ” is a mol microorganism of the invention may produce less than about ecule upon which an enzyme acts and a “ product ” is a 0 . 001 , 0 . 01 , 0 . 10 , 0 . 30 , 0 . 50 , or 1 . 0 g / L target product . molecule produced by the action of an enzyme . A “ native 40 “ Codon optimization ” refers to the mutation of a nucleic substrate , " therefore , is a molecule upon which an enzyme acid , such as a gene , for optimized or improved translation natively acts in a wild - type microorganism and a “ native of the nucleic acid in a particular strain or species . Codon product is a molecule natively produced by the action of the optimization may result in faster translation rates or higher enzyme in the wild - type microorganism . For example , translation accuracy . In a preferred embodiment , the genes butanoyl - CoA is the native substrate of Ptb and butanoyl 45 of the invention are codon optimized for expression in phosphate and is the native substrate of Buk . Additionally , Clostridium , particularly Clostridium autoethanogenum , butanoyl phosphate is the native product of Ptb and butyrate Clostridium ljungdahlii , or Clostridium ragsdalei . In a fur ( butanoate ) is the native product of Buk . Likewise , a “ non - ther preferred embodiment , the genes of the invention are native substrate ” is a molecule upon which an enzyme does codon optimized for expression in Clostridium autoethano not natively act in a wild - type microorganism and a " non - 50 genum LZ1561 , which is deposited under DSMZ accession native product ” is a molecule not natively produced by the number DSM23693 . action of the enzyme in the wild - type microorganism . An “ Overexpressed ” refers to an increase in expression of a enzyme that is capable of acting on multiple different nucleic acid or protein in the microorganism of the invention substrates , whether native or non - native , is typically referred compared to the wild - type or parental microorganism from to as a " promiscuous ” enzyme . The inventors have discov - 55 which the microorganism of the invention is derived . Over ered that Ptb is promiscuous and is capable of accepting a expression may be achieved by any means known in the art , variety of acyl - CoAs and enoyl - CoAs as substrates , such including modifying gene copy number , gene transcription that Ptb - Buk may be used to convert a number of acyl - CoAs rate , gene translation rate , or enzyme degradation rate . and enoyl - CoAs to their corresponding acids or alkenates , The term “ variants ” includes nucleic acids and proteins respectively , while simultaneously generating ATP . Thus , in 60 whose sequence varies from the sequence of a reference preferred embodiments , the Ptb - Buk of the invention acts on nucleic acid and protein , such as a sequence of a reference non - native substrates ( i . e . , substrates other than butanoyl - nucleic acid and protein disclosed in the prior art or exem COA and / or butanoyl phosphate ) to produce non - native plified herein . The invention may be practiced using variant products ( i . e . , products other than butanoyl phosphate and / or nucleic acids or proteins that perform substantially the same butyrate ( butanoate ) ) . 65 function as the reference nucleic acid or protein . For
The term “ butyryl - COA ” may be used interchangeably example , a variant protein may perform substantially the herein with “ butanoyl - CoA . ” same function or catalyze substantially the same reaction as

US 9 , 957 , 531 B1 49 50
a reference protein . A variant gene may encode the same or promoter . Ideally , the promoter is a Wood - Ljungdahl path substantially the same protein as a reference gene . A variant way promoter , a ferredoxin promoter , a pyruvate : ferredoxin promoter may have substantially the same ability to promote oxidoreductase promoter , an Rnf complex operon promoter , the expression of one or more genes as a reference promoter . an ATP synthase operon promoter , or a phosphotransacety
Such nucleic acids or proteins may be referred to herein 5 lase / acetate kinase operon promoter . as “ functionally equivalent variants . " By way of example , A “ microorganism ” is a microscopic organism , especially functionally equivalent variants of a nucleic acid may a bacterium , archea , virus , or fungus . The microorganism of include allelic variants , fragments of a gene , mutated genes , the invention is typically a bacterium . As used herein , polymorphisms , and the like . Homologous genes from other recitation of “ microorganism ” should be taken to encompass microorganisms are also examples of functionally equiva - 10 " bacterium . ” lent variants . These include homologous genes in species A “ parental microorganism ” is a microorganism used to such as Clostridium acetobutylicum , Clostridium beijer generate a microorganism of the invention . The parental inckii , or Clostridium ljungdahlii , the details of which are publicly available on websites such as Genbank or NCBI . microorganism may be a naturally - occurring microorganism Fu equivalent variants also include nucleic acids 15 ( i . e . , a wild - type microorganism ) or a microorganism that whose sequence varies as a result of codon optimization for has been previously modified ( i . e . , a mutant or recombinant a particular microorganism . A functionally equivalent vari microorganism ) . The microorganism of the invention may ant of a nucleic acid will preferably have at least approxi be modified to express or overexpress one or more enzymes mately 70 % , approximately 80 % , approximately 85 % , that were not expressed or overexpressed in the parental approximately 90 % , approximately 95 % , approximately 20 microorganism . Similarly , the microorganism of the inven 98 % , or greater nucleic acid sequence identity ( percent tion may be modified to contain one or more genes that were homology ) with the referenced nucleic acid . A functionally not contained by the parental microorganism . The microor equivalent variant of a protein will preferably have at least ganism of the invention may also be modified to not express approximately 70 % , approximately 80 % , approximately or to express lower amounts of one or more enzymes that 85 % , approximately 90 % , approximately 95 % , approxi - 25 were expressed in the parental microorganism . In one mately 98 % , or greater amino acid identity ( percent homol - embodiment , the parental microorganism is Clostridium ogy ) with the referenced protein . The functional equivalence autoethanogenum , Clostridium ljungdahlii , or Clostridium of a variant nucleic acid or protein may be evaluated using ragsdalei . In a preferred embodiment , the parental micro any method known in the art . organism is Clostridium autoethanogenum LZ1561 , which
Nucleic acids may be delivered to a microorganism of the 30 is deposited under DSMZ accession number DSM23693 . invention using any method known in the art . For example , The term " derived from ” indicates that a nucleic acid , nucleic acids may be delivered as naked nucleic acids or protein , or microorganism is modified or adapted from a may be formulated with one or more agents , such as different ( e . g . , a parental or wild - type ) nucleic acid , protein , liposomes . The nucleic acids may be DNA , RNA , DNA , or or microorganism , so as to produce a new nucleic acid , combinations thereof , as is appropriate . Restriction inhibi - 35 protein , or microorganism . Such modifications or adapta tors may be used in certain embodiments . Additional vectors tions typically include insertion , deletion , mutation , or sub may include plasmids , viruses , bacteriophages , cosmids , and stitution of nucleic acids or genes . Generally , the microor artificial chromosomes . In a preferred embodiment , nucleic ganism of the invention is derived from a parental acids are delivered to the microorganism of the invention microorganism . In one embodiment , the microorganism of using a plasmid . By way of example , transformation ( includ - 40 the invention is derived from Clostridium autoethanogenum , ing transduction or transfection ) may be achieved by elec - Clostridium ljungdahlii , or Clostridium ragsdalei . In a pre troporation , ultrasonication , polyethylene glycol - mediated ferred embodiment , the microorganism of the invention is transformation , chemical or natural competence , protoplast derived from Clostridium autoethanogenum LZ1561 , which transformation , prophage induction , or conjugation . In cer - is deposited under DSMZ accession number DSM23693 . tain embodiments having active restriction enzyme systems , 45 The microorganism of the invention may be further clas it may be necessary to methylate a nucleic acid before sified based on functional characteristics . For example , the introduction of the nucleic acid into a microorganism . microorganism of the invention may be or may be derived
Furthermore , nucleic acids may be designed to comprise from a C1 - fixing microorganism , an anaerobe , an acetogen , a regulatory element , such as a promoter , to increase or an ethanologen , a carboxydotroph , and / or a methanotroph . otherwise control expression of a particular nucleic acid . 50 Table 1 provides a representative list of microorganisms and The promoter may be a constitutive promoter or an inducible identifies their functional characteristics .
TABLE 1 C1 - fixing Anaerobe Acetogen Ethanologen Autotroph Carboxydotroph Methanotroph
1 + + J +
+ + + +
+ + L +
+ + + +
+ + L +
+ + + +
Acetobacterium woodii Alkalibaculum bacchii Blautia producta Butyribacterium methylotrophicum Clostridium aceticum Clostridium autoethanogenum Clostridium carboxidivorans Clostridium coskatii Clostridium drakei Clostridium formicoaceticum Clostridium ljungdahlii Clostridium magnum Clostridium ragsdalei
+ + + + ILI I II III III + + + +
+ + L +
+ + +
+ + + +
N
+ + L
+ + + +

US 9 , 957 , 531 B1 51 52
TABLE 1 - continued C1 - fixing Anaerobe Acetogen Ethanologen Autotroph Carboxydotroph Methanotroph
| +
| +
+ + + +
+ +
w +
Clostridium scatologenes Eubacterium limosum Moorella thermautotrophica Moorella thermoacetica ( formerly Clostridium thermoaceticum ) Oxobacter pfennigii Sporomusa ovata Sporomusa silvacetica Sporomusa sphaeroides Thermoanaerobacter kiuvi
III III +
+ LLLLL + + III + +
+ au +
+
??
Acetobacterium woodi can produce ethanol from fructose , but not from gas . 2 It has not been investigated whether Clostridium magnum can gro One strain of Moorella thermoacetica , Moorella sp . HUC22 - 1 , has been reported to produce ethanol from gas .
4 It has not been investigated whether Sporomusa ovata can grow on CO . * It has not been investigated whether Sporomusa silvacetica can grow on CO .
It has not been investigated whether Sporomusa sphaeroides can grow on CO . ?
“ C1 ” refers to a one - carbon molecule , for example , CO , the microorganism of the invention is an autotroph . In a CO2 , CH4 , or CH2OH . “ C1 - oxygenate ” refers to a one - preferred embodiment , the microorganism of the invention carbon molecule that also comprises at least one oxygen is derived from an autotroph identified in Table 1 . atom , for example , CO , CO , , or CH2OH . “ C1 - carbon A " carboxydotroph ” is a microorganism capable of uti source ” refers a one carbon - molecule that serves as a partial lizing CO as a sole source of carbon . Typically , the micro or sole carbon source for the microorganism of the inven - 25 organism of the invention is a carboxydotroph . In a preferred tion . For example , a C1 - carbon source may comprise one or embodiment , the microorganism of the invention is derived more of CO , CO2 , CH4 , CH2OH , or CH2O2 . Preferably , the from a carboxydotroph identified in Table 1 . Cl - carbon source comprises one or both of CO and CO2 . A A “ methanotroph ” is a microorganism capable of utilizing “ C1 - fixing microorganism ” is a microorganism that has the methane as a sole source of carbon and energy . In certain ability to produce one or more products from a C1 - carbon 30 embodiments , the microorganism of the invention is derived source . Typically , the microorganism of the invention is a from a methanotroph . C1 - fixing bacterium . In a preferred embodiment , the micro - More broadly , the microorganism of the invention may be organism of the invention is derived from a C1 - fixing derived from any genus or species identified in Table 1 . microorganism identified in Table 1 . In a preferred embodiment , the microorganism of the
An " anaerobe ” is a microorganism that does not require 35 invention is derived from the cluster of Clostridia compris oxygen for growth . An anaerobe may react negatively or ing the species Clostridium autoethanogenum , Clostridium even die if oxygen is present above a certain threshold ljungdahlii , and Clostridium ragsdalei . These species were Typically , the microorganism of the invention is an anaer - first reported and characterized by Abrini , Arch Microbiol , obe . In a preferred embodiment , the microorganism of the 161 : 345 - 351 , 1994 ( Clostridium autoethanogenum ) , Tan invention is derived from an anaerobe identified in Table 1 . 40 ner , Int J System Bacteriol , 43 : 232 - 236 , 1993 ( Clostridium An " acetogen ” is a microorganism that produces or is ljungdahlii ) , and Huhnke , WO 2008 / 028055 ( Clostridium
capable of producing acetate ( or acetic acid ) as a product of ragsdalei ) . anaerobic respiration . Typically , acetogens are obligately These three species have many similarities . In particular , anaerobic bacteria that use the Wood - Ljungdahl pathway as these species are all C1 - fixing , anaerobic , acetogenic , etha their main mechanism for energy conservation and for 45 nologenic , and carboxydotrophic members of the genus synthesis of acetyl - CoA and acetyl - CoA - derived products , Clostridium . These species have similar genotypes and such as acetate ( Ragsdale , Biochim Biophys Acta , 1784 : phenotypes and modes of energy conservation and fermen 1873 - 1898 , 2008 ) . Acetogens use the acetyl - CoA pathway tative metabolism . Moreover , these species are clustered in as a ( 1 ) mechanism for the reductive synthesis of acetyl - clostridial rRNA homology group I with 16S rRNA DNA COA from CO2 , ( 2 ) terminal electron - accepting , energy 50 that is more than 99 % identical , have a DNA G + C content conserving process , ( 3 ) mechanism for the fixation ( assimi - of about 22 - 30 mol % , are gram - positive , have similar lation ) of CO2 in the synthesis of cell carbon ( Drake , morphology and size ( logarithmic growing cells between Acetogenic Prokaryotes , In : The Prokaryotes , 3rd edition , p . 0 . 5 - 0 . 7x3 - 5 um ) , are mesophilic ( grow optimally at 30 - 37° 354 , New York , N . Y . , 2006 ) . All naturally occurring aceto C . ) , have similar pH ranges of about 4 - 7 . 5 ( with an optimal gens are C1 - fixing , anaerobic , autotrophic , and non - metha - 55 pH of about 5 . 5 - 6 ) , lack cytochromes , and conserve energy notrophic . Typically , the microorganism of the invention is via an Rnf complex . Also , reduction of carboxylic acids into an acetogen . In a preferred embodiment , the microorganism their corresponding alcohols has been shown in these spe of the invention is derived from an acetogen identified in cies ( Perez , Biotechnol Bioeng , 110 : 1066 - 1077 , 2012 ) . Table 1 . Importantly , these species also all show strong autotrophic An “ ethanologen ” is a microorganism that produces or is 60 growth on CO - containing gases , produce ethanol and acetate
capable of producing ethanol . Typically , the microorganism ( or acetic acid ) as main fermentation products , and produce of the invention is an ethanologen . In a preferred embodi - small amounts of 2 , 3 - butanediol and lactic acid under cer ment , the microorganism of the invention is derived from an tain conditions . ethanologen identified in Table 1 . However , these three species also have a number of An “ autotroph ” is a microorganism capable of growing in 65 differences . These species were isolated from different
the absence of organic carbon . Instead , autotrophs use sources : Clostridium autoethanogenum from rabbit gut , inorganic carbon sources , such as CO and / or CO2 . Typically , Clostridium ljungdahlii from chicken yard waste , and

US 9 , 957 , 531 B1 53 54
Clostridium ragsdalei from freshwater sediment . These spe - Preferably , the substrate comprises about 40 - 70 mol % CO cies differ in utilization of various sugars ( e . g . , rhamnose , ( e . g . , steel mill or blast furnace gas ) , about 20 - 30 mol % CO arabinose ) , acids ( e . g . , gluconate , citrate ) , amino acids ( e . g . , ( e . g . , basic oxygen furnace gas ) , or about 15 - 45 mol % CO arginine , histidine ) , and other substrates ( e . g . , betaine , buta ( e . g . , syngas ) . In some embodiments , the substrate may nol ) . Moreover , these species differ in auxotrophy to certain 5 comprise a relatively low amount of CO , such as about 1 - 10 vitamins ( e . g . , thiamine , biotin ) . These species have differ - or 1 - 20 mol % CO . The microorganism of the invention ences in nucleic and amino acid sequences of Wood - Ljung - typically converts at least a portion of the CO in the substrate dahl pathway genes and proteins , although the general to a product . In some embodiments , the substrate comprises organization and number of these genes and proteins has no or substantially no CO . been found to be the same in all species ( Köpke , Curr Opin 10 The substrate may comprise some amount of Hy . For Biotechnol , 22 : 320 - 325 , 2011 ) . example , the substrate may comprise about 1 , 2 , 5 , 10 , 15 ,
Thus , in summary , many of the characteristics of 20 , or 30 mol % Hz . In some embodiments , the substrate Clostridium autoethanogenum , Clostridium ljungdahlii , or may comprise a relatively high amount of H2 , such as about Clostridium ragsdalei are not specific to that species , but are 60 , 70 , 80 , or 90 mol % H , . In further embodiments , the rather general characteristics for this cluster of C1 - fixing , 15 substrate comprises no or substantially no H2 . anaerobic , acetogenic , ethanologenic , and carboxydotrophic The substrate may comprise some amount of CO , . For members of the genus Clostridium . However , since these example , the substrate may comprise about 1 - 80 or 1 - 30 mol species are , in fact , distinct , the genetic modification or % CO , . In some embodiments , the substrate may comprise manipulation of one of these species may not have an less than about 20 , 15 , 10 , or 5 mol % CO , . In another identical effect in another of these species . For instance , 20 embodiment , the substrate comprises no or substantially no differences in growth , performance , or product production CO2 . may be observed . Although the substrate is typically gaseous , the substrate
The microorganism of the invention may also be derived may also be provided in alternative forms . For example , the from an isolate or mutant of Clostridium autoethanogenum , substrate may be dissolved in a liquid saturated with a Clostridium ljungdahlii , or Clostridium ragsdalei . Isolates 25 CO - containing gas using a microbubble dispersion genera and mutants of Clostridium autoethanogenum include JA1 - 1 tor . By way of further example , the substrate may be ( DSM10061 ) ( Abrini , Arch Microbiol , 161 : 345 - 351 , 1994 ) , adsorbed onto a solid support . LBS1560 ( DSM19630 ) ( WO 2009 / 064200 ) , and LZ1561 The substrate and / or C1 - carbon source may be a waste ( DSM23693 ) . Isolates and mutants of Clostridium ljung gas obtained as a byproduct of an industrial process or from dahlii include ATCC 49587 ( Tanner , Int J Syst Bacteriol , 43 : 30 some other source , such as from automobile exhaust fumes 232 - 236 , 1993 ) , PETCT ( DSM13528 , ATCC 55383 ) , ERI - 2 or biomass gasification . In certain embodiments , the indus ( ATCC 55380 ) ( U . S . Pat . No . 5 , 593 , 886 ) , C - 01 ( ATCC trial process is selected from the group consisting of ferrous 55988 ) ( U . S . Pat . No . 6 , 368 , 819 ) , 0 - 52 ( ATCC 55989 ) metal products manufacturing , such as a steel mill manu ( U . S . Pat . No . 6 , 368 , 819 ) , and OTA - 1 ( Tirado - Acevedo , facturing , non - ferrous products manufacturing , petroleum Production of bioethanol from synthesis gas using 35 refining processes , coal gasification , electric power produc Clostridium ljungdahlii , PhD thesis , North Carolina State tion , carbon black production , ammonia production , metha University , 2010 ) . Isolates and mutants of Clostridium rags nol production , and coke manufacturing . In these embodi dalei include PI 1 ( ATCC BAA - 622 , ATCC PTA - 7826 ) ( WO ments , the substrate and / or C1 - carbon source may be 2008 / 028055 ) . captured from the industrial process before it is emitted into
In some embodiments , however , the microorganism of the 40 the atmosphere , using any convenient method . invention is a microorganism other than Clostridium The substrate and / or C1 - carbon source may be syngas , autoethanogenum , Clostridium ljungdahlii , or Clostridium such as syngas obtained by gasification of coal or refinery ragsdalei . For example , the microorganism may be selected residues , gasification of biomass or lignocellulosic material , from the group consisting of Escherichia coli , Saccharomy or reforming of natural gas . In another embodiment , the ces cerevisiae , Clostridium acetobutylicum , Clostridium 45 syngas may be obtained from the gasification of municipal beijerinckii , Clostridium saccharbutyricum , Clostridium solid waste or industrial solid waste . saccharoperbutylacetonicum , Clostridium butyricum , The composition of the substrate may have a significant Clostridium diolis , Clostridium kluyveri , Clostridium pas impact on the efficiency and / or cost of the reaction . For terianium , Clostridium Novyi , Clostridium difficile , example , the presence of oxygen ( 02 ) may reduce the Clostridium thermocellum , Clostridium cellulolyticum , 50 efficiency of an anaerobic fermentation process . Depending Clostridium cellulovorans , Clostridium phytofermentans , on the composition of the substrate , it may be desirable to Lactococcus lactis , Bacillus subtilis , Bacillus licheniformis , treat , scrub , or filter the substrate to remove any undesired Zymomonas mobilis , Klebsiella oxytoca , Klebsiella pneu - impurities , such as toxins , undesired components , or dust monia , Corynebacterium glutamicum , Trichoderma reesei , particles , and / or increase the concentration of desirable Cupriavidus necator , Pseudomonas putida , Lactobacillus 55 components . plantarum , and Methylobacterium extorquens . The microorganism of the invention may be cultured to
" Substrate ” refers to a carbon and / or energy source for the produce one or more products . For instance , Clostridium microorganism of the invention . Typically , the substrate is autoethanogenum produces or can be engineered to produce gaseous and comprises a C1 - carbon source , for example , ethanol ( WO 2007 / 117157 ) , acetate ( WO 2007 / 117157 ) , CO , CO , , and / or CH . . Preferably , the substrate comprises a 60 butanol ( WO 2008 / 115080 and WO 2012 / 053905 ) , butyrate C1 - carbon source of CO or CO + CO , . The substrate may ( WO 2008 / 115080 ) , 2 , 3 - butanediol ( WO 2009 / 151342 ) , lac further comprise other non - carbon components , such as H2 , tate ( WO 2011 / 112103 ) , butene ( WO 2012 / 024522 ) , buta N2 , or electrons . diene ( WO 2012 / 024522 ) , methyl ethyl ketone ( 2 - butanone )
The substrate generally comprises at least some amount of ( WO 2012 / 024522 and WO 2013 / 185123 ) , ethylene ( WO CO , such as about 1 , 2 , 5 , 10 , 20 , 30 , 40 , 50 , 60 , 70 , 80 , 90 , 65 2012 / 026833 ) , acetone ( WO 2012 / 115527 ) , isopropanol or 100 mol % CO . The substrate may comprise a range of ( WO 2012 / 115527 ) , lipids ( WO 2013 / 036147 ) , 3 - hydroxy CO , such as about 20 - 80 , 30 - 70 , or 40 - 60 mol % CO . propionate ( 3 - HP ) ( WO 2013 / 180581 ) , isoprene ( WO 2013 /

55 US 9 , 957 , 531 B1
56 180584 ) , fatty acids ( WO 2013 / 191567 ) , 2 - butanol ( WO include pressure ( or partial pressure ) , temperature , gas flow 2013 / 185123 ) , 1 , 2 - propanediol ( WO 2014 / 0369152 ) , and rate , liquid flow rate , media pH , media redox potential , 1 - propanol ( WO 2014 / 0369152 ) . In addition to one or more agitation rate ( if using a continuous stirred tank reactor ) , target products , the microorganism of the invention may also inoculum level , maximum gas substrate concentrations to produce ethanol , acetate , and / or 2 , 3 - butanediol . In certain 5 ensure that gas in the liquid phase does not become limiting , embodiments , microbial biomass itself may be considered a and maximum product concentrations to avoid product product . inhibition . In particular , the rate of introduction of the A “ native product is a product produced by a genetically substrate may be controlled to ensure that the concentration unmodified microorganism . For example , ethanol , acetate , of gas in the liquid phase does not become limiting , since and 2 , 3 - butanediol are native products of Clostridium 10 products may be consumed by the culture under gas - limited autoethanogenum , Clostridium ljungdahlii , and Clostridium conditions . ragsdalei . A “ non - native product ” is a product that is pro duced by a genetically modified microorganism , but is not Operating a bioreactor at elevated pressures allows for an
increased rate of gas mass transfer from the gas phase to the produced by a genetically unmodified microorganism from which the genetically modified microorganism is derived 15 liquid phase . Accordingly , it is generally preferable to per The terms “ intermediate ” and “ precursor , ” which may be form the culture / fermentation at pressures higher than atmo
referred to interchangeably herein , refer to a molecular spheric pressure . Also , since a given gas conversion rate is , entity in an enzymatic pathway upstream of an observed or in part , a function of the substrate retention time and target product . retention time dictates the required volume of a bioreactor ,
“ Selectivity ” refers to the ratio of the production of a 20 the use of pressurized systems can greatly reduce the volume target product to the production of all fermentation products of the bioreactor required and , consequently , the capital cost produced by a microorganism . The microorganism of the of the culture / fermentation equipment . This , in turn , means invention may be engineered to produce products at a certain that the retention time , defined as the liquid volume in the selectivity or at a minimum selectivity . In one embodiment , bioreactor divided by the input gas flow rate , can be reduced a target product account for at least about 5 % , 10 % , 15 % , 25 when bioreactors are maintained at elevated pressure rather 20 % , 30 % , 50 % , or 75 % of all fermentation products than atmospheric pressure . The optimum reaction conditions produced by the microorganism of the invention . In one will depend partly on the particular microorganism used . embodiment , the target product accounts for at least 10 % of However , in general , it is preferable to operate the fermen all fermentation products produced by the microorganism of tation at a pressure higher than atmospheric pressure . Also , the invention , such that the microorganism of the invention 30 since a given gas conversion rate is in part a function of has a selectivity for the target product of at least 10 % . In substrate retention time and achieving a desired retention another embodiment , the target product accounts for at least time in turn dictates the required volume of a bioreactor , the 30 % of all fermentation products produced by the microor - use of pressurized systems can greatly reduce the volume of ganism of the invention , such that the microorganism of the the bioreactor required , and consequently the capital cost of invention has a selectivity for the target product of at least 35 the fermentation equipment . 30 % . Target products may be separated or purified from a
“ Increasing the efficiency , ” “ increased efficiency , " and the fermentation broth using any method or combination of like include , but are not limited to , increasing growth rate , methods known in the art , including , for example , fractional product production rate or volume , product volume per distillation , evaporation , pervaporation , gas stripping , phase volume of substrate consumed , or product selectivity . Efli - 40 separation , and extractive fermentation , including for ciency may be measured relative to the performance of example , liquid - liquid extraction . In certain embodiments , parental microorganism from which the microorganism of target products are recovered from the fermentation broth by the invention is derived . continuously removing a portion of the broth from the
Typically , the culture is performed in a bioreactor . The bioreactor , separating microbial cells from the broth ( con term “ bioreactor " includes a culture / fermentation device 45 veniently by filtration ) , and recovering one or more target consisting of one or more vessels , towers , or piping arrange - products from the broth . Alcohols and / or acetone may be ments , such as a continuous stirred tank reactor ( CSTR ) , recovered , for example , by distillation . Acids may be recov immobilized cell reactor ( ICR ) , trickle bed reactor ( TBR ) , ered , for example , by adsorption on activated charcoal . bubble column , gas lift fermenter , static mixer , or other Separated microbial cells are preferably returned to the vessel or other device suitable for gas - liquid contact . In 50 bioreactor . The cell - free permeate remaining after target some embodiments , the bioreactor may comprise a first products have been removed is also preferably returned to growth reactor and a second culture / fermentation reactor . the bioreactor . Additional nutrients ( such as B vitamins ) may The substrate may be provided to one or both of these be added to the cell - free permeate to replenish the medium reactors . As used herein , the terms “ culture ” and “ fermen - before it is returned to the bioreactor . tation " are used interchangeably . These terms encompass 55 both the growth phase and product biosynthesis phase of the EXAMPLES culture / fermentation process .
The culture is generally maintained in an aqueous culture The following examples further illustrate the invention medium that contains nutrients , vitamins , and / or minerals but , of course , should not be construed to limit its scope in sufficient to permit growth of the microorganism . Preferably 60 any way . the aqueous culture medium is an anaerobic microbial growth medium , such as a minimal anaerobic microbial Example 1 growth medium . Suitable media are well known in the art .
The culture / fermentation should desirably be carried out This example demonstrates the ability of Ptb - Buk to under appropriate conditions for production of the target 65 convert acetoacetyl - CoA to acetoacetate in E . coli in vivo product . Typically , the culture / fermentation is performed and its use in production of acetone , isopropanol , 3 - hy under anaerobic conditions . Reaction conditions to consider droxybutyrate , and isobutylene

57 US 9 , 957 , 531 B1
58 Pathways that rely on the Ptb - Buk system for acetoacetate Acetone concentrations , as well as the concentrations of
production from acetoacetyl - CoA were designed and con other metabolites such as isobutylene , were measured using structed . This was done in a modular fashion using a PDUET gas chromatography ( GC ) analysis , employing an Agilent vector system ( Novagen ) . One module contained ptb - buk 6890N headspace GC equipped with a Supelco polyethylene genes from C . beijerinckii NCIMB8052 ( GenBank 5 glycol ( PEG ) 60 - um solid - phase microextraction fiber , a NC _ 009617 , position 232027 . . . 234147 ; Cbei _ 0203 - 204 ; Restek Rtx - 1 ( 30 mx0 . 32 umx5 um ) column , and a flame NCBI - GeneID 5291437 - 38 ) on plasmid PACYC . Another ionization detector ( FID ) . Samples ( 4 ml ) were transferred module contained the thiolase gene thlA of C . acetobutyli into a 20 - ml headspace vial , upon which the fiber was cum ( Genbank NC _ 001988 , position 82040 . . . 83218 ; incubated ( exposed ) for 10 min at 50° C . The sample was CA _ P0078 ; NCBI - GenelD 1116083 ) and the acetoacetate 10 desorbed in the injector at 250° C . for 9 min . Chromatog decarboxylase gene adc of C . beijerinckii NCIMB8052 raphy was performed with an oven program of 40° C . ( 5 - min ( Genbank NC _ 009617 , position 4401916 . . . 4402656 ; hold ) and 10° C . / min to 200° C . , followed by a 5 - min hold Cbei _ 3835 ; NCBI - GeneID 5294996 ) on plasmid pCOLA . at 220° C . The column flow rate was 1 ml / min , with Ptb and buk genes were amplified from genomic DNA of C . hydrogen as the carrier gas . The FID was kept at 250° C . , beijerinckii NCIMB8052 and th1A and adc genes from an 15 with hydrogen at 40 ml / min , air at 450 ml / min , and nitrogen existing acetone plasmid PMTL85147 - thlA - ctfAB - adc ( WO at 15 ml / min as the makeup gas . 2012 / 115527 ) and cloned under control of the T7 promoter It was immediately obvious that acetone was produced in present in the pDUET vectors via restriction independent the strain carrying both the PACYC - ptb - buk and pCOLA cloning with the circular polymerase extension cloning th1A - adc plamids ( expressing thiolase , Ptb - Buk , and ( CPEC ) method ( Quan , PloS One , 4 : e6441 , 2009 ) . 20 acetoacetate decarboxylase ) . Average final acetone produc
Oligonucleotides used for amplification of ptb and buk t ion of 0 . 19 g / L was measured , whereas no acetone was genes : produced in a no plasmid control , media control , and single
SEQ ID NO : Name Sequence Direction
5
5
PACYCDuet - ptb - buk - AAGTTTTTACTCATATGTATATC reverse PACYC - ptb - R1 TCCTTCTTATACTTAAC PACYCDuet - ptb - buk - ptb - AGAAGGAGATATACATATGAGT forward PACYC - F1 AAAAACTTTGATGAGTTA PACYCDuet - ptb - buk - ACCAGACTCGAGGGTACCTAGT reverse buk - PACYC - R1 AAAccTTAGCTTGT ?c PACYCDuet - ptb - buk - TAAGGTTTACTAGGTACCCTCG forward PACYC - buk - F1 AGTCTGGTAAAGAAAC
5
Oligonucleotides used for amplification of th1A and adc genes :
plasmid controls PACYC - ptb - buk ( expressing Ptb - Buk ) or PCOLA - th1A - adc plamid ( expressing thiolase and acetoac
SEQ ID NO : Name Sequence Direction
100
101
PCOLADuet - th1A - adc - ACATATGTATATCTCCTTCTTAC reverse th1A - adc - R1 TAGCACTTTTCTAGCAATATTG pCOLADuet - thiA - adc - AGTAAGAAGGAGATATACATAT forward adc - Th1A - F1 GTTAGAAAGTGAAGTATCTAAAC pCOLADuet - th1A - adc - CAGACTCGAGGGTACCTTATTT reverse adc - PCOLA - R1 TACTGAAAGATAATCATGTAC PCOLADuet - th1A - adc - TCTTTCAGTAAAATAAGGTACC forward PCOLA - adc - F1 CTCGAGTCTGGTAAAGAAAC PCOLADuet - th1A - adc - GAAGGAGATATACATATGAAA forward thlA - PCOLA - F1 GAAGTTGTAATAGCTAGTG PCOLADuet - thiA - adc - ACAACTTCTTTCATATGTATATC reverse pCOLA - th1A - R1 TCCTTCTTATACTTAAC
102
103 104
After the plasmids PACYC - ptb - buk ( SEQ ID NO : 105 ) etate decarboxylase ) ( below reliable detection limit ) . The and pCOLA - th1A - adc ( SEQ ID NO : 106 ) were constructed , uninduced culture of the strain carrying both the PACYC they were transformed individually and together into E . coli ptb - buk and PCOLA - thlA - adc plamids ( expressing thiolase , BL21 ( DE3 ) ( Novagen ) and growth experiments carried out 55 Ptb - Buk , and acetoacetate decarboxylase ) did not produce in quadruplicates in 1 . 5 mL cultures in 12 - well plates at 28° appreciable amounts of acetone . C . with 160 rpm orbital shaking using M9 minimal medium Average Acetone Production in E . coli BL21 ( DE3 ) : ( Sambrook , Molecular Cloning : A Laboratory Manual , Vol 3 , Cold Spring Harbour Press , 1989 ) with glucose ( FIG . 4 ) . The cultures were inoculated at an OD600 nm of 0 . 1 and 60 Acetone ( g / L ) induced with different concentrations of IPTG ( 0 , 50 , 100 Thl + Ptb - Buk + Adc [ E . coil BL21 ( DE3 ) + 0 . 19 + 0 . 04 UM ) after 2 h of growth ( FIG . 5 ) . The plates were sealed PACYC - ptb - buk + pCOLA - thlA - adc ] using plate tape strips and each well was pierced with a Thl + Adc alone [ E . coil BL21 ( DE3 ) + 0 . 04 + 0 . 01
pCOLA - thlA - adc ] green tipped needle to provide micro - aerobic conditions . 65 Ptb - Buk alone [ E . coil BL21 ( DE3 ) + 0 . 03 + 0 . 01 Growth was carried out for another 64 h of induction . The PACYC - ptb - buk ] experiment was repeated in triplicate .
Strain

108 buk - Fl TTTATGTTAGAAAG
110
US 9 , 957 , 531 B1 59 60
- continued PMTL85147 - th1A - ctfAB - adc encoding thl + ctfAB + ade ( WO 2012 / 115527 ) under the control of a clostridial promoter
Strain Acetone ( g / L ) from the Wood - Ljungdahl gene cluster . In this plasmid the No plasmid control [ E . coil BL21 ( DE3 ) ] 0 . 04 + 0 . 01 ctfAB genes encoding the CoA transferase were replaced Media control 0 . 03 0 . 01 5 directly with ptb - buk genes encoding the Ptb - Buk system .
This was done as described in Example 1 using the CPEC This experiment clearly demonstrates that Ptb - Buk is able e method . The resulting plasmid is pMTL85147 - th1A - ptb
buk - adc . to perform the conversion of acetoacetyl - CoA to acetoac etate can be used in place of a COA - transferase or a 10 Oligonucleotides used for the amplification of ptb - buk
and cloning into pMTL8317 - thl - ptb - buk - adc are described thioesterase for the production of acetone , exemplified using below . a route that comprises steps 1 , 2 , and 3 of FIG . 1 .
It is well known that isopropanol can be produced from acetone by addition of a primary : secondary alcohol dehy SEQ ID NO : Name Sequence Direction drogenase ( Köpke , Appl Environ Microbiol , 80 : 3394 - 3403 , 15 - 2014 ) ( step 4 in FIG . 1 ) and that isobutylene can be 107 thiA - ATTTCCTCCCTTTCTAGCACTTT reverse
ptb - R1 TCTAGCAATATTG produced from acetone via addition of a hydroxyisovalerate adc - TAAGGTTTACTAAGGAGGTTGT forward synthase ( step 5 in FIG . 1 ) and decarboxylase ( step 6 in FIG . 1 ) ( van Leeuwen , Appl Microbiol Biotechnol , 93 : 1377 109 th1A - GCTAGAAAAGTGCTAGAAAGG forward 1387 , 2012 ) . A pathway can be constructed that includes the 20 ptb - F1 GAGGAAATGAACATG
Buk AAAACAACCTCCTTAGTAAACC reverse above - demonstrated acetone route via Ptb - Buk with the adc - R1 TTAGCTTGTTCTTC genes th1A , ptb - buk , and adc and a primary : secondary alcohol dehydrogenase gene ( e . g . , Genbank accession num ber NC _ 022592 , pos . 609711 . . . 610766 ; CAETHG 0553 ; C . autoethanogenum DSM10061 and DSM23693 ( a deri NCBI - GeneID : 17333984 ) that would allow isopropanol 25 vate of DSM10061 ) were sourced from DSMZ ( The German production via the Ptb - Buk system in E . coli comprising Collection of Microorganisms and Cell Cultures , steps 1 . 2 . 3 . and 4 of FIG . 1 . Similarly , a pathway can be Inhoffenstraße 7 B , 38124 Braunschweig , Germany ) . constructed that includes the above - demonstrated acetone Strains were grown at 37° C . in PETC medium at pH 5 . 6 route via Ptb - Buk conversion of acetoacetyl - CoA to acetoac - using standard anaerobic techniques ( Hungate , Meth Micro etate with the genes th1A , ptb - buk , and adc and genes for a 30 biol , 3B : 117 - 132 , 1969 ; Wolfe , Adv Microb Physiol , 6 : hydroxyisovalerate synthase and decarboxylase that would 107 - 146 , 1971 ) . 30 psi CO - containing steel mill gas ( col allow isobutylene production via the Ptb - Buk system in E . lected from New Zealand Steel site in Glenbrook , NZ ) or a coli comprising of steps 1 , 2 , 3 , 5 , and 6 of FIG . 1 . synthetic gas blend with same composition of 44 % CO , 32 % Acetoacetete can also be converted to 3 - hydroxybutyrate via N2 , 22 % CO . , 2 % H , was used as substrate for autotrophic a 3 - hydroxybutyrate dehydrogenase Bdh . This can be com - 35 growth . For solid media , 1 . 2 % bacto agar ( BD , Franklin bined with Ptb - Buk conversion of acetoacetyl - CoA to Lakes , N . J . 07417 , USA ) was added . acetoacetate for 3 - hydroxybutyrate production in a strain The construct was synthesized and then transformed into expressing genes th1A , ptb - buk , and bdh resulting in a C . autoethanogenum via conjugation . For this , the expres pathway comprising steps 1 , 2 , and 15 of FIG . 1 . sion vector was first introduced into the conjugative donor
40 strain E . coli HB101 + R702 ( CA434 ) ( Williams , J Gen Example 2 Microbiol , 1136 : 819 - 826 , 1990 ) ( the donor ) using standard
heat shock transformation . Donor cells were recovered in This example demonstrates the ability of Ptb - Buk to SOC medium ( Sambrook , Molecular Cloning : A Laboratory
convert acetoacetyl - CoA to acetoacetate in C . autoethano - Manual , Vol 3 , Cold Spring Harbour Press , 1989 ) at 37° C . genum in vivo and the use of Ptb - Buk in the production of 45 for 1 h before being plated on to LB medium ( Sambrook , acetone , isopropanol , 3 - hydroxybutyrate , and isobutylene Molecular Cloning : A Laboratory Manual , Vol 3 , Cold from a gaseous substrate . Spring Harbour Press , 1989 ) plates containing 100 ug / ml
To demonstrate that the Ptb - Buk system also allows spectinomycin and 25 ug / ml chloramphenicol . LB plates acetone , isopropanol , or isobutylene synthesis from gaseous were incubated at 37° C . overnight . The next day , 5 ml LB substrates , a plasmid was constructed that contains the same 50 aliquots containing 100 ug / ml spectinomycin and 25 ug / ml genes as in Example 1 , thl + ptb - buk + ade under control of a chloramphenicol were inoculated with several donor colo clostridial promoter on a shuttle vector that allows expres - nies and incubated at 37° C . , shaking for approximately 4 h , sion in acetogens such as C . autoethanogenum , C . ljung - or until the culture was visibly dense but had not yet entered dahlii or C . ragsdalei . stationary phase . 1 . 5 ml of the donor culture was harvested
The pMTL plasmid is a shuttle plasmid system for intro - 55 in a microcentrifuge tube at room temperature by centrifu ducing circular dna into Clostridia via E . coli conjugation gation at 4000 rpm for 2 min , and the supernatant was ( Heap , J Microbiol Methods , 78 : 79 - 85 , 2009 . The genes of discarded . The donor cells were gently resuspended in 500 interest ( i . e . , hbd , phaB , th1A , ptb , buk , and aorl ) were ul sterile PBS buffer ( Sambrook , Molecular Cloning : A cloned into the lacZ region of the plasmids using common Laboratory Manual , Vol 3 , Cold Spring Harbour Press , techniques in molecular biology including dna restriction 60 1989 ) and centrifuged at 4000 rpm for 2 min and the PBS digestion followed by ligation , and the golden gate dna supernatant was discarded . The pellet was introduced into an assembly technology when more than one pieces of dna anaerobic chamber and gently resuspended in 200 ul during fragments were to be cloned simultaneously into the plas - late exponential phase C . autoethanogenum culture ( the mid . The constructed plasmids are verified by DNA sequenc - recipient ) . The conjugation mixture ( the mix of donor and ing . 65 recipient cells ) was spotted onto PETC - MES + fructose agar
Production of acetone and isopropanol was previously plates and left to dry . When the spots were no longer visibly demonstrated in C . autoethanogenum using a plasmid wet , the plates were introduced into a pressure jar , pressur

62 US 9 , 957 , 531 B1
61 ized with syngas to 25 - 30 psi and incubated at 37° C . for ~ 24 ( Köpke , Appl Environ Microbiol , 80 : 3394 - 3403 , 2014 ) . It h . After 24 h incubation , the conjugation mixture was has been demonstrated that knock - out of this gene elimi removed from the plates by gently scraping it off using a 10 nates conversion of acetone to isopropanol in C . autoetha ul inoculation loop . The removed mixture was suspended in nogenum ( WO 2015 / 085015 ) . In background of this knock 200 - 300 ul PETC medium . 100 ul aliquots of the conjuga - 5 out , it becomes possible to produce acetone ( rather than tion mixture were plated on to PETC medium agar plates isopropanol ) via the Ptb - Buk system from a gaseous feed supplemented 15 ug / ml thiamphenicol to select for trans stock , using the same genes comprising steps 1 , 2 , and 3 of formants bearing the plasmid , which confers resistance to FIG . 1 . Addition of hydroxyisovalerate synthase and decar thiamphenicol via expression of chloramphenicol acetyl - boxylase genes ( van Leeuwen , Appl Microbiol Biotechnol , transferase . 10 93 : 1377 - 1387 , 2012 ) to this strain would enable isobutylene
Three distinct colonies of C . autoethanogenum bearing production from gas in C . autoethanogenum or similar the pMTL85147 - thlA - ptb - buk - adc plasmid were inoculated bacteria comprising of steps 1 , 2 , 3 , 5 , and 6 of FIG . 1 . into 2 mL of PETC - MES medium with 15 ug / ml thiam - Acetoacetate can also be converted to 3 - hydroxybutyrate phenicol and grown autotrophically at 37° C . with 100 rpm via a 3 - hydroxybutyrate dehydrogenase Bdh . A 3 - hydroxy orbital shaking for three days . Cultures were diluted to 15 butyrate dehydrogenase was identified in the genome of C . OD = 0 . 05 in 10 mL PETC - MES medium with 15 ug / ml autoethanogenum ( AGY75962 ) and other acetogens as C . thiamphenicol in serum bottles and grown autotrophically at ljungdahlii ( ADK16920 . 1 ) . This activity can be combined 37° C . with 100 rpm orbital shaking for five days , sampling with Ptb - Buk ( or COA transferase ) conversion of daily to measure biomass and metabolites . In parallel a acetoacetyl - CoA to acetoacetate for 3 - hydroxybutyrate pro control strain was examined where the expression plasmid 20 duction in a strain expressing genes th1A , ptb - buk ( or ctfAB ) encoded only thl and adc under the control of the Wood - and bdh resulting a pathway comprising steps 1 , 2 , and 15 Ljungdahl cluster promoter , with no ctfAB or ptb - buk genes of FIG . 1 . Low levels of 3 - hydroxybutyrate formation ( up to to catalyse the formation of acetoacetate from acetoacetyl 2 g / L ) via this route have been demonstrated in C . autoetha COA ( PMTL85147 - th1A - adc ) . Cultures were sampled for nogenum . These levels could be enhanced by overexpress five days in order to monitor metabolites and biomass 25 ing the Bdh gene that is only expressed in at low levels accumulation . natively .
Isopropanol concentrations as well as concentrations of In one experiment , C . autoethanogenum was transformed ethanol , acetic acid , 2 , 3 - butanediol and lactic acid were with plasmid pMTL82256 - thlA - ctfAB as described in measured by high - performance liquid chromatography Example 2 . The production was monitored for 10 days from ( HPLC ) on an Agilent LC with refractive index ( RI ) detec - 30 six biological replicates under autotrophic conditions as tion at 35° C . Samples were prepared by diluting 400 uL described in Example 2 . The average of 3 - HB after 10 days with 100 uL of 5 - sulfosalicylic acid solution ( 1 % w / v in 1 was 1 . 86 + 0 . 14 g / L . At day 10 , 1 , 3 - butanediol was produced M sulphuric acid ) , followed by a 3 minute centrifugation at ( from 3 - HB ) at an average titer of 0 . 38 + 0 . 05 g / L ( FIG . 37 ) . 14 , 000 rpm ; the supernatant was transferred to a glass vial No acetone or isopropanol was formed . This demonstrates for analysis . Separation was carried out with a 10 UL 35 that 3 - HB can be produced efficiently via acetoacetate injection on to an Alltech IOA - 2000 column ( 150 mmx6 . 5 through native enzymes . mmx8 um ) at 0 . 7 mL / min and 65° C . under isocratic In certain embodiments , it may be desirable to knock out conditions , using 5 mM sulphuric acid mobile phase . or knock down expression of 3 - hydroxybutyrate dehydro
In some instances , a longer HPLC method was used to genases , such as Bdh , to prevent carbon drain to 3 - HB and improve peak separation . In this method , isopropanol , etha - 40 therefore boost production of products such as acetone , nol , acetate , 2 , 3 - butanediol , and also 3 - hydroxybutyrate isopropanol , and isobutylene . ( which is not separated using the shorter method ) concen trations were measured by high - performance liquid chroma Example 3 tography ( HPLC ) on an Agilent 1260 Infinity LC with refractive index ( RI ) detection at 35° C . Samples were 45 This example demonstrates the ability of Ptb - Buk to prepared by diluting 400 uL with 100 uL of 5 - sulfosalicylic convert ( R ) - 3 - hydroxybutyryl - CoA to ( R ) - 3 - hydroxybu acid solution ( 1 % w / v in 1 M sulphuric acid ) , followed by tyryrate in E . coli in vivo for production of ( R ) - hydroxybu a 3 minute centrifugation at 14 , 000 rpm ; the supernatant was tyrate , acetone , isopropanol , or isobutylene . transferred to a glass vial for analysis . Separation was Pathways were designed and constructed that rely on the carried out with a 10 uL injection on to an Aminex HPX - 87H 50 Ptb - Buk system for ( R ) - 3 - hydroxybutyrate production from column ( 300 mmx7 . 8 mmx9 um ) at 0 . 6 mL / min and 35° C . ( R ) - 3 - hydroxybutyryl - CoA . Additionally , a 3 - hydroxybu under isocratic conditions , using 5 mM sulphuric acid tyrate dehydrogenase ( Bdh ) was utilized for conversion of mobile phase . ( R ) - 3 - HB to acetoacetate . It has been reported that Ralstonia
C . autoethanogenum bearing the pMTL85147 - th1A - ptb - pickettii have two 3 - hydroxybutyrate dehydrogenases Bdh1 buk - adc produced isopropanol up to 0 . 804 g IPA / g of 55 and Bdh2 that are able to convert 3 - hydroxybutyrate to biomass , whereas control strain C . autoethanogenum with acetoacetate in vitro ( Takanashi , J Biosci Bioeng , 101 : PMTL85147 - th1A - adc that does not contain Ptb - Buk pro 501 - 507 , 2006 ) . One pathway was designed making use of duced no IPA ( FIG . 12 ) . this enzyme for acetone production ( steps 1 , 13 , 14 , 15 , 3 of
This experiment clearly demonstrates that Ptb - Buk is able FIG . 1 ) , while recycling the reducing equivalents produced to perform the conversion of acetoacetyl - CoA to acetoac - 60 in the production of ( R ) - 3 - hydroxybutyryl - CoA and the ATP etate in the isopropanol pathway when using a gaseous generated by Ptb - Buk ( FIG . 6 ) . substrate . Ptb - Buk can be used in place of a COA transferase The pathways were constructed in a modular fashion or a thioesterase in a gas - fermenting acetogen such as C . using the pDUET vector system ( Novagen ) . The two mod autoethanogenum , exemplified using a route that comprises ules described in example above ( PACYC - ptb - buk for steps 1 , 2 , 3 , and 4 of FIG . 1 . 65 expression of Ptb - Buk and pCOLA - thlA - adc for expression
C . autoethanogenum contains a native primary : secondary of thiolase and acetoacetate decarboxylase ) were used alcohol dehydrogenase that converts acetone to isopropanol together with two additional modules containing either

US 9 , 957 , 531 B1 63 64
( R ) - specific 3 - hydroxybutyrate dehydrogenase phaB of 3 - hydroxybutyrate or acetone was found ( FIG . 13A - F ) . In Cupravidus necator ( WP _ 010810131 . 1 ) alone ( PCDF - these cultures , acetate levels were significantly higher . phaB ) and one with 3 - hydroxybutyrate dehydrogenase bdh1 This experiment clearly demonstrates that Ptb - Buk is able gene of Rasltonia pickettii ( BAE72684 . 1 ) ( PCDF - phaB - to perform the conversion of ( R ) - 3 - hydroxybutyrate - CoA to bdhl ) in vector pCDF . Both phaB and bdhl gene were 5 3 - hydroxybutyrate and also that Bdh1 is able in vivo to synthesized from GeneArt and cloned under control of the convert 3 - hydroxybutyrate further to acetoacetate by recy T7 promoter present in via restriction independent cloning cling the reducing equivalents produced in the production of with the circular polymerase extension cloning ( CPEC ) ( R ) - 3 - hydroxybutyryl - CoA . The experiment also highlights method ( Quan , PloS One , 4 : e6441 , 2009 ) . that Ptb - Buk is able to support growth and therefore acetate
Oligonucleotides used for amplification of bdhl gene : 10 production becomes unnecessary . Production of ( R ) - 3 - hy droxybutyrate formation was exemplified in a strain that comprises steps 1 , 13 , and 14 of FIG . 1 . Production of SEQ ID NO : Name Sequence Direction acetone was exemplified via a route that comprises steps 1 ,
111 pDuet - CATATGTATATCTCCTTCTTATA forward 13 , 14 , 15 , and 3 of FIG . 1 . insert2 - CTTAAC It is well known that isopropanol can be produced from R1 acetone by addition of a primary : secondary alcohol dehy
112 drogenase ( step 4 in FIG . 1 ) ( Köpke , Appl Environ Micro pDuet - F1 ACATATG pDuet - CCTCGAGTCTGGTAAAGAAAC forward biol , 80 : 3394 - 3403 , 2014 ) and that isobutylene can be insert2 produced from acetone via addition of a hydroxyisovalerate F1 20 synthase ( step 5 in FIG . 1 ) and decarboxylase ( step 6 in FIG .
114 insert2 - GTTTCTTTACCAGACTCGAGG forward 1 ) ( van Leeuwen , Appl Microbiol Biotechnol , 93 : 1377 pDuet - R1 1387 , 2012 ) . A pathway can be constructed that includes the Oligonucleotides used for amplification of phaB gene : above - demonstrated acetone route via Ptb - Buk with the
15
insert2 - GTTAAGTATAAGAAGGAGATAT forward
113
SEQ ID NO : Name Sequence Direction 115
116
PCDF - phaB - PACYC - CTATTCTTTGTGTCATGGTATAT forward phaB - R1 ??ccTTATTAAAG pCDF - phaB - phaB - ATAAGGAGATATACCATGACAC forward PACYC - F1 AAAGAATAGCATAC PCDF - phaB - PACYC - TGGTTTACACATGGGATAAGAT forward phaB - Fi CCGAATTCGAGCTC PCDF - phaB - phaB - AGCTCGAATTCGGATCTTATCC forward PACYC - R1 CATGTGTAAACCAC
117
118
35 After the plasmids PACYC - ptb - buk ( SEQ ID NO : 105 ) , genes th1A , ptb - buk , and adc and a primary : secondary
PCOLA - th1A - adc ( SEQ ID NO : 106 ) , PCDF - phaB ( SEQ ID alcohol dehydrogenase gene ( e . g . , Genbank NC _ 022592 , NO : 119 ) and PCDF - phaB - bdhl ( SEQ ID NO : 120 ) were pos . 609711 . . . 610766 ; CAETHG _ 0553 ; NCBI - GeneID : constructed , they were transformed individually and in com 17333984 ) that would allow isopropanol production via the binations into E . coli BL21 ( DE3 ) ( Novagen ) and growth 40 Ptb - Buk system in E . coli ( steps 1 , 13 , 14 , 15 , 3 , and 4 of experiments were carried out in quadruplicate in 1 . 5 mL FIG . 1 ) . Similarly , a pathway can be constructed that cultures in 12 - well plates at 28° C . with 160 rom orbital includes the above - demonstrated acetone route via Ptb - Buk shaking using M9 minimal medium with glucose . The with the genes th1A , ptb - buk , and adc and genes for a cultures were inoculated at an OD600 nm of 0 . 1 and after 2 hydroxyisovalerate synthase and decarboxylase that would h of growth induced with different concentrations of IPTG 45 allow isobutylene production via the Ptb - Buk system in E .
coli ( steps 1 , 13 , 14 , 15 , 3 , 5 , and 6 of FIG . 1 ) . ( 0 , 50 , 100 uM ) . The plates were sealed using BioRad plate tape strips and each well pierced with a green tipped needle Example 4 to provide micro - aerobic conditions . Growth was carried out for another 64 h of induction . The experiment was repeated 50 This example demonstrates the production of ( R ) - 3 - hy 3 times . Metabolites were measured as described in previous droxybutyrate and 1 , 3 - butanediol in C . autoethanogenum . It examples . also demonstrates production of 1 , 3 - butanediol in absence
Cultures containing a combination of plasmids PACYC - of 2 , 3 - butanediol . ptb - buk , PCOLA - thlA - adc and PCDF - phaB produced A strain of C . autoethanogenum was constructed in which between 1 . 65 - 2 . 4 g / L ( R ) - 3 - hydroxybutyrate ( depending on 55 the native pathway for 2 , 3 - butanediol production was inac level of inducer ) , with only very small amounts of byprod tivated and replaced with genes for ( R ) - 3 - hydroxybutyryl ucts ( FIGS . 13A - F ) , demonstrating the efficiency of the COA formation . This was achieved by replacing the aceto Ptb - Buk system to convert ( R ) - 3 - hydroxybutyryl - CoA to lactate decarboxylase gene ( budA ) on genome of C . ( R ) - 3 - hydroxybutyryrate and support growth ( FIG . 13A - F ) . autoethanogenum with genes for thiolase ( thlA of C . aceto In cultures that also expressed bdhl ( containing a combi - 60 butylicum ; GenBank NC _ 001988 , position 82040 . . . 83218 ; nation of plasmids PACYC - ptb - buk , pCOLA - th1A - adc , and CA _ P0078 ; NCBI - GeneID 1116083 ) and ( R ) - specific 3 - hy PCDF - phaB - bdhl ) only small amounts of ( R ) - 3 - hydroxy - droxybutyrate dehydrogenase ( phaB of Cupravidus necator ; butyryrate were found in the culture media , while between GenBank WP _ 010810131 . 1 ) resulting in strain C . autoetha 0 . 89 - 1 . 16 g / L acetone was found ( depending on level of nogenum budA : : thlAphaB . inducer ) , indicating that bdh1 gene is efficient in converting 65 To replace budA gene with thlA and phaB genes a ( R ) - 3 - hydroxybutyrate to acetoacetate and further to plasmid , pMTL8225 - budA : : th1A - phaB ( FIG . 14 ) , with E . acetone . In all plasmid combinations that lack Ptb - Buk , no coli toxin gene mazF under tet3n0 tetracycline inducible

US 9 , 957 , 531 B1 65 66
promoter ( for counter selection ) , ~ 1 kb upstream homology detector ( FID ) . Samples were prepared by diluting 400 uL of arm of budA gene , th1A , phab , erm? cassette flanked by sample with 400 uL of acetonitrile , followed by a 3 minute loxP sites and ~ 1 kb downstream homology arm of budA centrifugation at 14 , 000 rpm ; the supernatant was trans gene were assembled on plasmid pMTL - tet3no . ferred to a glass vial and the sample was dried in a Thermo
The ~ 1 kb upstream and downstream homology arms of 5 SpeedVac . Once dry , the samples were then suspended in a budA were PCR amplified from C . autoethanogenum with solution of 400 uL of N , O - Bistrifluoroacetamide ( BSTFA ) primers SN01 / SNO2 and SN07 / SN08 . th1A and phaB genes and pyridine ( 3 : 1 ratio ) and heated in a sealed glass vial for were PCR amplified from genomic DNA of Cupriavidus 60 minutes at 60° C . Samples were transferred to an necator using primers SNO3 / SN04mod . The erm? cassette autosampler for analysis using a 1 uL injection , a split ration flanked with loxP sites was PCR amplified using primers 10 of 30 to 1 , and an inlet temperature of 250° C . Chromatog SN05mod / SN06 . tet3no promoter flanked by Fsel and Pmel raphy was performed with an oven program of 70° C . ( no was synthesized and treated with restriction enzymes Fsel hold ) to a ramp of 3° C . / min to 110° C . to a ramp of 15° and Pmel and cleaned . The PCR products and digested vector were assembled using GeneArt Seamless cloning kit C . / min to 230° C . , followed by a final ramp of 40° C . / min from Life Technologies and plasmid pMTL8225 - budA : : 15 to 5 to 310° C . with a 3 - min hold . The column flow rate was 1 . 8 thlA - phaB ( SEQ ID NO : 121 ) with no mutations in the ml / min , with helium as the carrier gas . The FID was kept at inserted fragments was used to transform C . autoethanoge - 320° C . , with hydrogen at 40 ml / min , air at 400 ml / min , and num by conjugation as described in previous examples . helium at 20 ml / min as the makeup gas .
Following conjugation and selection on trimethoprim and Surprisingly , up to 1 . 55 g / L 3 - hydroxybutyrate was pro clarithromycin , 9 colonies were streaked twice on PETC - 20 duced from gas in a C . autoethanogenum budA : : th1A - phaB MES agar plates with clarithromycin and anhydrotetracy strain expressing thlA and phaB ( FIG . 16 ) . A native thio cline to induce the expression of mazF genes . The colonies esterase may convert the formed 3 - hydroxybutyryl - CoA to from clarithromycin and anhydrotetracycline should have 3 - hydroxybutyrate . In the genome sequence , three putative the budA genes replaced with thlA and phaB genes and thioesterases were identified . erm? cassette . This was verified by PCR using primers 25 Even more surprising , it was also found that , along Og31f / Og32r flanking the homology arms and KAPA poly - 3 - hydroxybutyrate formation , there was also 1 , 3 - butanediol merase ( FIG . 15 ) . formation of up to 150 mg / L ( FIG . 16 ) . This may be due to
While a band of ~ 3 . 3 kb is amplified from the wild type native aldehyde : ferredoxin oxidoreductase ( AOR ) and alco strain , bands of ~ 5 . 7 kb were amplified from colonies 1 , 4 , 7 hol dehydrogenase activity . Two AOR genes and several and 9 indicating the replacement of budA gene with th1A , 30 alcohol dehydrogenases are present in the genome of C . phaB and erm? cassette . The above event was further autoethanogenum ( Mock , J Bacteriol , 197 : 2965 - 2980 , confirmed by sequencing the PCR products of all 4 clones . 2015 ) . This reduction of 3 - hydroxybutyrate is powered by With the resulting modification the expression of th1A and reduced ferredoxin and thus can be directly coupled to CO phaB genes is driven by the promoter upstream of budA oxidation , which provides reduced ferredoxin ( CO + gene . Fdox ?CO2 + Fdred ) ( FIG . 7 ) .
SEQ ID Descrip NO : tion Sequence
PPPP 122 SN01
5NO2 5N03
125 SN04mod
ATTTACAAATTcGuccGGcc?????ccTCGTATAAATAAGATG ?TAGCTATTACAACTICTTTCATATTACATTCAccTCTATGTC GACATAGAGGTGAATGTAATATGAAAGAAGTTGTAATAGCTAG GTATAGCATACATTATACGAACGGTATTATCCCATGTGTAAACC ACCGT TTCGTATAATGTATGCTATACGAAGTTATCCTTAGAAGCAAACT TAAG GTCTAGTGTTTTTTTCTATCAATACTCTAGATACCGTTCGTATAG
126 SNO5mod
127 SN06
128 SN07
129 SNO8 130 Og3lf 131 Og32r
TGTATGCTATACGAACGGTAAGTATTGATAGAAAAAAACACTA GAC CAAAAAGGAGTTTAAACAAAAAGTCATAAACCTGGATAAC ccGTTTCTCACAACAACAATACCAG AAACCACCTTGACGATGAAACCATA
A fermentation with C . autoethanogenum budA : : th1A - 1 , 3 - BDO production was also demonstrated from gas via phaB strain was carried out . The culture was grown at 37° 53 an alternative route using a butyraldehyde dehydrogenase C . under synthetic gas ( 50 % CO , 18 % CO2 , 2 % H2 , and 30 % Bld from Clostridium saccharoperbutylacetonicum N , ) that was continuously fed into the bioreactor . The gas flow was initially set at 50 ml / min , increasing to 400 ml / min ( AAP42563 . 1 ) ( SEQ ID NO : 80 ) . The bld gene was syn over the course of the experiment , while the agitation was thesized and cloned together with the same thiolase ( thlA of increased from 200 rpm to 500 rpm . The fermentation was ou C . acetobutylicum ) and ( R ) - specific 3 - hydroxybutyrate carried out for close to 5 days . Metabolites were measured as described in examples above . dehydrogenase ( phaB of Cupravidus necator ) into a plasmid
The concentration of 1 , 3 - butanediol and other metabo - pMTL8315 - Pfdx - th1A - phaB - bld ( SEQ ID NO : 132 ) . Bld lites , such as 2 - hydroxyisobutyric acid , were measured using and phaB genes were amplified from the above plasmid via gas chromatography ( GC ) analysis , employing an Agilent 65 6890N GC equipped a Agilent CP - SIL 5CB - MS ( 50 mx0 . 25 primers in table below and cloned into existing plasmid umx0 . 25 um ) column , autosampler and a flame ionization PMTL85147 - th1A ( WO 2012 / 115527 ) .

US 9 , 957 , 531 B1 67 68
SEQ ID NO : Primer Sequence Direction 133 bid - phaB - F1 ACATGGGATAAGAAGGAGATATACATATGAT forward
AAAAG 134 bld - pMTL - R1 CGTCGACTCTAGATTAACCTGCTAAAACA forward
??????c 135 PMTL - bld - F1 GTGTTTTAGCAGGTTAATCTAGAGTC forward
GAcGTCACGC
The resulting construct was transformed into C . autoetha nogenum as described above and a growth experiment was conducted in serum bottles with 50 - mL PETC media and pressurized at 30 psi with CO - containing steel mill gas 15 ( collected from New Zealand Steel site in Glenbrook , NZ ) or a synthetic gas blend with same composition of 44 % CO , 32 % N2 , 22 % CO2 , 2 % Hz .
Scientific ) . The operon P - bbd1 - rbs2 - th1A was cloned in between restriction sites NotI and Xhol found in the multiple cloning region of the plasmid . P is the constitutive promoter which contains an intact ribosome binding site ( rbs ) . rbs2 ( SEO ID NO : 140 ) is the ribosome binding site for express ing thl A . The stepwise procedures were amplification of the P , hbd1 , and thlA from existing templates with primers listed below .
SEQ ID NO : Name Sequence Direction
141 Pfdx - F 1 AAAGGTCTCCGGCCGCGCTCACTATCTGCG forward GAACC
142 Pfdx - ri TTTGGTCTCGAATTCTGTAACACCTCCTTAA reverse TTTTTAG
143 Ppfor - F 1 AAAGGTCTCCGGCCGCAAAATAGTTGATAA forward TAATGCAGAG
144 Ppfor - ri TTTGGTCTCGAATTCCTCTC reverse CTTTTCAAGCATATA
145 hbdl - F 1 AAAGGTCTCGAATTCAAAGATCTATGTCTAT forward TAAATCAGTTGCAG
146 hbal - R1 TTTGGTCTccc?cc????? reverse ATTTCTAATATGCGAAAAATcc??????
147 th1A - F1 AAAGGTCTCAGGAGGTGTTACATATGAAAG forward AAGTTGTAATAGCTAGTGC
148 th1A - R1 TTTGGTCTCCTCGAGTATGGATCCCTAGCAC reverse TTTTCTAGCAATATTGC
1 , 3 - BDO production was demonstrated via this route The polymerase chain reactions were performed as follow from gas ( FIG . 17A ) , but production was less ( up to 67 mg / L 1 using Kapa Taq PCR Kit ( Kapa Biosystems ) . Set annealing 1 , 3 - BDO ) than via the AOR route and , in contrast to the to temperature at 56° C . , and extension for 1 minute . Repeat AOR route , growth was impacted when expressing the bld PCR reaction for 30 cycles . Afterwards . PCR products were gene comparing to the C . autoethanogenum wild - type ( FIG . desalted using the DNA Clean & Concentrator Kit ( Zymo 17B ) .
In another experiment , C . autoethanogenum transformed Research Corporation ) . with plasmid MTL83159 - phaB - th1A as described in 45 PMTL83151 plasmid backbone was prepared by carrying Example 2 produced 0 . 33 and 0 . 46 g / L of 3 - HB and 1 , 3 out the Notl / Xhol double digestion using the FastDigest BDO , respectively , in a bottle experiment under autotrophic Notl and FastDigest Xhol ( Thermo Fisher Scientific ) fol conditions as described in Example 2 ( FIG . 40 ) . lowing the protocol provided , followed by treatment with
alkaline phosphate , using the FastAP Alkaline Phosphatase Example 5 50 ( Thermo Fisher Scientific ) and the protocols provided . The
digested backbone was then desalted with the DNA Clean & This example demonstrates the production of ( S ) - 3 - hy - Concentrator Kit ( Zymo Research Corporation ) .
droxybutyrate and 1 , 3 - butanediol in C . autoethanogenum . The assembly of the PCR products and the plasmid A plasmid was constructed that expresses a thiolase ( thlA backbone was carried out using the GeneArt Type IIs Kit
from C . acetobutylicum ; SEQ ID NO : 136 ) and an ( S ) - 55 ( Thermo Fisher Scientific ) . The resulting plasmid was then specific 3 - hydroxybutyrate dehydrogenase ( hbdl from C . isolated from the E . coli plasmid expression host using the kluyveri ; SEQ ID NO : 137 ) under either a ferredoxin pro QIAprep Spin Miniprep Kit ( Qiagen ) . moter ( Pfar isolated from C . autoethanogenum ; SEQ ID NO : To introduce the assembled plasmids pMTL8315 - Pfdx 138 ) or a pyruvate - ferredoxin oxidoreductase promoter hbd1 - thlA and pMTL8315 - Ppfor - hbd1 - th1A consisting of ( Ppfor isolated from C . autoethanogenum ; SEQ ID NO : 139 ) . 60 the operons , the plasmid was first introduced into the E . coli The plasmid was constructed as follows : P - hbd1 - rbs2 - th1A CA434 strain by chemical transformation . Afterwards , con and pieced together and cloned into the pMTL83151 vector jugation was performed by mixing the transformed CA434 ( Heap , J Microbiol Meth , 78 : 79 - 85 , 2009 ) by routine strain with a C . autoethanogenum production host on a solid methods in molecular cloning , including restrictive enzyme LB - agar media , and incubation in an anaerobic environment digestion followed by ligation , overlap extension poly - 65 under pressure with a mix consisting of carbon monoxide merase chain reaction , seamless cloning ( Thermo Fisher and hydrogen as described in Example 2 . C . autoethanoge Scientific ) , and GeneArt Type IIs ( Thermo Fisher num , after conjugation , was selected by successive growth

70
3 . 7 SO
Ethanol
Biomass
US 9 , 957 , 531 B1 69
on the solid media containing the proper antibiotic and trimethroprim to remove the remaining E . coli CA434 strain , Selectivity under the anaerobic conditions . ( C - mol % )
The C . autoethanogenum strains carrying the introduced Acetate 14 . 7 Ethanol 64 . 9 PMTL8315 - Pfdx - hbdl - thlA or PMTL8315 - Ppfor - hbd1 2 , 3 - BDO 1 . 3 thlA plasmids consisting of the operon P - hbd1 - rbs2 - th1A Biomass
were grown in a 10 - mL PETC media in a 250 - ml Schott 3 - HB 10 . 4 1 , 3 - BDO bottle , sealed tight with rubber septum and cap , and pres
surized at 30 psi with CO - containing steel mill gas ( col - 10 lected from New Zealand Steel site in Glenbrook , NZ ) or a A s a comparison , in a strain expressing the same plasmid , synthetic gas blend with same composition of 44 % CO , 32 % PMTL83159 - hbd - thlA without budA knockout , the total N . , 22 % CO . , 2 % H . . Metabolites were measured as specificity for the production of 3 - HB and 1 , 3 - BDO at the described in previous examples . steady state was only 6 . 9 %
Surprisingly , there was 3 - hydroxybutyrate produced from 15 gas in C . autoethanogenum cultures expressing thlA and Selectivity hbd1 ( FIG . 18A ) . A native thioesterase may convert the ( C - mol % ) formed 3 - hydroxybutyryl - CoA to 3 - hydroxybutyrate . In the Acetate 0 . 4 genome sequence , three putative thioesterases were identi 84 . 3 fied . In the strain carrying pMTL8315 - Pfdx - hbd1 - th / A up to 2 , 3 - BDO
2 . 55 g / L 3 - hydroxybutyrate was found ( FIG . 18A ) . 3 - HB Even more surprising , it was also found that 3 - hydroxy 1 , 3 - BDO
butyrate is over time converted to 1 , 3 - butanediol , at the end of growth up to 1 . 1 g / L 1 , 3 - butanediol was produced in 25 strain carrying plasmid pMTL8315 - Pfdx - hbd1 - th1A ( FIG . Example 6 18A ) . This may be due to native aldehyde : ferredoxin oxi doreductase ( AOR ) and alcohol dehydrogenase activity . This example demonstrates that the Ptb - Buk system is Two AOR genes and several alcohol dehydrogenases are efficient in C . autoethanogenum on a range of acyl - CoAs present in the genome of C . autoethanogenum ( Mock , J 30 30 including acetoacetyl - CoA , 3 - hydroxybutyryl - CoA , and
2 - hydroxyisobutyryl - CoA Bacteriol , 197 : 2965 - 2980 , 2015 ) . This reduction of 3 - hy The Ptb - Buk system was expressed from a plasmid in C . droxybutyrate ( and reduction of acetate to ethanol ; FIG . autoethanogenum and its activity measured using a COA 18B ) is powered by reduced ferredoxin and thus can be hydrolysis assay . For this , ptb - buk genes from C . beijerinckii directly coupled to CO oxidation , which provides reduced „ NCIMB8052 ( GenBank NC 009617 . position 232027 . . . ferredoxin ( CO + Fdox ?CO2 + Fdred ) ( FIG . 7 ) . 234147 ; Cbei _ 0203 - 204 ; NCBI - GeneID 5291437 - 38 ) were
The same strain of C . autoethanogenum carrying plasmid amplified from genomic DNA of C . beijerinckii PMTL8315 - Pfdx - hbd1 - th1A was also tested in continuous NCIMB8052 and cloned under control of a pyruvate - ferre fermentation . Fermentation was carried out as described in doxin oxidoreductase promoter ( Pp for isolated from C . previous example , but the culture was turned continuous 40 an autoethanogenum ; SEQ ID NO : 139 ) into pMTL82251 with a dilution rate with fresh media of around 0 . 05 at day vector ( ( Heap , J Microbiol Meth , 78 : 79 - 85 , 2009 ) by 2 and then increased to 1 . 0 at day 3 . High 3 - hydroxybutyrate routine methods in molecular cloning , including restrictive
enzyme digestion followed by ligation , overlap extension production of up to 7 g / L was observed with 1 , 3 - BDO polymerase chain reaction , seamless cloning ( Thermo Fisher production of 0 . 5 g / L . Scientific ) , and GeneArt Type IIs ( Thermo Fisher Scientific ) To improve production of ( S ) - 3 - hydroxybutyrate and 1 , 3 as described in Example 5 . Oligonucleotides are described
butanediol and avoid synthesis of another form of butanediol below .
WWNO ? NN
SEO ID NO : Name Sequence Direction 149 Ppfor - F2 aaacagctatgaccgcGGCCGCAAAATAGTforward 150 Ppfor - R2 ttactcatTGGATTCCTCTCCITT reverse 151 Ptb - Buk - F2ggaatccaATGAGTAAAAACTTTGATGAG forward 152 Ptb - Buk - R2caggcctcgagatctcCTAGTAAACCTTAGreverse
?TTGT ?c
( 2 , 3 - butanediol ) , plasmid pMTL - HBD - ThlA was introduced The resulting plasmid pMTL82256 - ptb - buk ( SEQ ID NO : into a strain that has an inactivated 2 , 3 - butanediol pathway 153 ) was introduced into C . autoethanogenum as described where the acetolactate decarboxylase gene BudA has been 60 in previous examples . deleted ( U . S . Pat . No . 9 , 297 , 026 ) . This budA knockout Acyl - CoA hydrolysis assays were performed as follows .
C . autoethanogenum cells were harvested at OD 2 ( late eliminated the major pathway to 2 , 3 - BDO , increasing the exponential phase ) by centrifugation ( 14 , 000 rpm for 1 min specificity for 3 - HB and 1 , 3 - BDO production . When PMTL auction . When PML . at 4° C . ) . Cells were re - suspended in 500 ul lysis buffer HBD - ThlA was expressed in the budA deletion strain , a total 65 ( potassium phosphate buffer , pH 8 ) . Cells were lysed using of 15 % C - mol was achieved for both 3 - HB and 1 , 3 - BDO a freeze thaw cycle ( optional ) , sonication 6x30 s at ampli ( FIG . 41 ) . tude 20 on ice . Samples were centrifuged for 10 min at

US 9 , 957 , 531 B1 72
14 , 000 rpm at 4° C . and the supernatant with soluble described in previous example . The results of the assay show proteins was removed . The protein concentration was mea cleavage of acetoacetyl - CoA and 3 - HB - COA , but not 2 - HIB sured , e . g . , with a Bradford assay . COA , and confirm native activity is present in C . autoetha
The assay mix contained : 484 ul of potassium phosphate nogenum ( FIG . 11 ) . buffer pH 8 . 0 , 1 ul of DTNB ( final concentration of 0 . 1 An analysis of the genome of C . autoethanogenum led to mM ) , 10 ul of cell lysate , and 5 ul of COA ( final concen - identification of three putative CoA - thioesterases ( thioester tration of 500 uM ) . All the components were mixed in a hydrolases ) that could be responsible for to the cleavage of quartz cuvette ( 1 ml cuvette with a read length of 1 cm ) acetoacetyl - CoA or 3 - hydroxybutyryl - CoA thioester bond . except the protein . The assay was started by adding the cell 10 These are also present in other acetogens such as C . ljung lysate and following the reaction in a spectrophotometer at dahlii .
SEQ SEQ ID NO : C . ljungdahlii
ID Description Annotation autoethanogenum NO :
154 ADK15695 . 1 157
4 155 ADK16655 . 1 158
thioesterase 1 ( CAETHG _ 0718 ) thioesterase 2 ( CAETHG _ 1524 ) thioesterase 3 ( CAETHG _ 1780 )
Palmitoyl - CoA AGY74947 . 1 hydrolase - Hydroxybenzoyl - AGY75747 . 1 COA thioesterase Putative AGY75999 . 1 Thioesterase
156 ADK16959 . 1 159
405 nm , 30° C . for 3 min . A control without lysate was run Inactivation of these three putative CoA - thioesterases to measure autolysis of the acyl - CoA . 25 lead to higher product titers , improving efficiency of the
To determine activity , slope on the linear part of the curve Ptb - Buk system . The three putative thioesterases were inac ( usually in the first 30 s ) , was calculated . The protein amount tivated using Clos Tron technology . In brief , the targeting was normalized and slope was divided by protein amount . domain of the type II Ltr was reprogrammed using the An extinction coefficient ( 14 , 150 M - cm ) was used to Clos Tron website and the retargeted ClosTron plasmids calculate the specific activity in M / s / mg . The activity of the 30 were ordered from DNA 2 . 0 . The Clos Tron knock out negative control was subtracted . vectors PMTL007C - E2 - Cau - 2640 - 571s targeting the thio
The assay was performed with acetoacetyl - CoA , a race - esterase 1 ( CAETHG _ 0718 ) , PMTL007C - E2 - PBor3782 mic mix of 3 - hydroxybutyryl - CoA ( 3 - HB - COA ) and 2 - hy 166s targeting the thioesterase 2 ( CAETHG _ 1524 ) , and droxyisobutyryl - COA ( 2 - HIB - COA ) . The possibility of arti - PMTL007C - E2 - PBor4039 - 199s targeting the thioesterase 3 ficially low hydrolysis rates for 3 - HB - CoA and 2 - HIB - COA 35 ( CAETHG _ 1780 ) were introduced into C . autoethanoge due to potential substrate limitation was addressed by num using conjugation . repeating the hydrolysis assays for C . autoethanogenum Selection for integration was done by selecting PETC lysates using different concentrations of acyl - CoA , 500 uM supplemented with 5 ug / ml clarithromycin and successful and 200 uM . inactivation by integration of the type II intron was con
The results of the assay show significantly increased COA 40 firmed by PCR across the insertion site . hydrolysis in lysates of C . autoethanogenum carrying plas - The COA hydrolase activity on acetoacetyl - CoA of both mid pMTL82256 - ptb - buk expressing the Ptb - Buk system on wild type C . autoethanogenum and each of the C . autoetha a range of acyl - CoAs including acetoacetyl - CoA , 3 - hy - nogenum with one of the putative genes inactivated was droxybutyryl - CoA and 2 - hydroxyisobutyryl - CoA ( FIGS . measured using the assay described above . It was shown that 20A - B ) . Notably , there is also CoA hydrolysis for acyl - 45 all three strains with the inactivated putative thioesterases CoAs as 2 - hydroxyisobutyryl - CoA that are not hydrolysed showed less hydrolysis activity on acetoacetyl - CoA and by the C . autoethanogenum wild - type . With acetoacetyl - 3 - hydroxybutyryl - CoA ( FIGS . 21A - B ) . COA and 3 - hydroxybutyryl - CoA some native CoA hydroly . To demonstrate that the decreased CoA hydrolase activity , sis activity was observed . and thus an increased pool in acetoacetyl - CoA , is beneficial
50 for production of acetoacetyl - CoA derived products , the Example 7 isopropanol plasmid p MTL85147 - thlA - ctfAB - adc encoding
thl + ctfAB + adc ( WO 2012 / 115527 ) was introduced into the This example demonstrates the disruption of identified C . autoethanogenum wild - type strain and the strain with
native thioesterase genes improve efficiency of the Ptb - Buk inactivated thioesterase 1 . A growth experiment was carried and CoA transferase system by increasing the pool of 55 out 40 ml PETC medium in 1 L Schott bottles in technical available acyl - CoAs such as acetoacetyl - CoA , 3 - hydroxy - triplicates with Co gas at 37° C . at 110 rpm shaking . butyryl - CoA or 2 - hydroxyisobutyryl - CoA . Synthetic gas ( 50 % CO , 18 % CO2 , 2 % H2 , and 30 % N2 ) was
In contrast to the Ptb - Buk system , where energy is con - used as sole energy and carbon source . Headspace served in the form of ATP during conversion of acyl - CoAs exchanged once and gassed to 21 psi ( 1 . 5 bar ) at 37° C . to their respective acids , no energy is conserved if the Cons 60 under synthetic gas ( 50 % CO , 18 % CO2 , 2 % H2 , and 30 % are simply hydrolyzed . N2 ) . Samples for OD and analytics were taken twice a day .
In hydrolase assays it was found that there is native The strain with inactivated thioesterase 3 CAETHG 1780 hydrolysis activity for acetoacetyl - CoA and 3 - hydroxybu - produced significantly higher levels of isopropanol than the tyryl - CoA in C . autoethanogenum . wild - type ( FIG . 22 and FIGS . 23A - D ) .
Acyl - CoA hydrolysis assays with acetoacetyl - CoA , a 65 Similarly , knockout of thioesterases in C . autoethanoge racemic mix of 3 - hydroxybutyryl - CoA ( 3 - HB - CoA ) and num would increase the pool of 3 - hydroxybutyryl - CoA , 2 - hydroxyisobutyryl - CoA ( 2 - HIB - COA were performed as allowing more efficient utilization of 3 - hydroxybutyryl - CoA

74 US 9 , 957 , 531 B1
73 by Ptb - Buk and leading to higher production of acetone , All acetogenic microorganisms are described to produce isopropanol , isobutylene , ( R ) - 3 - hydroxybutyrate , 1 , 3 - bu acetate ( Drake , Acetogenic Prokaryotes , In : The Prokary tanediol , and / or 2 - hydroxyisobutyric acid . When plasmid otes , 3rd edition , pages 354 - 420 , New York , N . Y . , Springer , PMTL8315 - Pfdx - hbd1 - th1A of Example 5 was introduced 2006 ) as the production of acetate provides the microorgan into C . autoethanogenum strain with interrupted thioesterase 5 ism with an option to directly generate ATP from substrate 2 CAETHG 1524 , 3 - hydroxybutyrate synthesis was abol - level phosphorylation via Pta ( phosphotransacetylase ) and ished ( compared to the up to 2 . 55 g / L 3 - hydroxybutyrate that Ack ( phosphotransacetylase - acetate kinase ) . Native acetate were found when expressing this plasmid in the C . autoetha forming enzymes such as Pta - Ack are therefore considered nogenum wild type strain ) . No competing activity for 3 - hy to be essential in acetogens ( Nagarajan , Microb Cell Fac droxybutyryl - CoA is present in this strain . 10 tories , 12 : 118 , 2013 ) . Since Ptb - Buk provides an alternative These results demonstrate that by reducing thioesterase means for energy generation , it becomes possible to replace activity , a higher CoA pool for the Ptb - Buk system and the native Pta - Ack system with Ptb - Buk . product synthesis is available .
Additionally , the production of 3 - HB and 1 , 3 - BDO can be The pta and ack genes in C . autoethanogenum are in one increased by overexpression of ptb - buk . In a control experi - 15 operon . To replace pta and ack genes with ptb and buk genes ment , whereby C . autoethanogenum as described in a plasmid , pMTL8225 - pta - ack : : ptb - buk ( FIG . 24 ) , with Example 2 was transformed with plasmids pMTL83159 - mazf counter selection marker that is under tetracycline phaB - th1A from Example 4 plus pMTL82256 ( Heap , J inducible promoter , ~ 1 kb upstream homology arm , ptb , Microbiol Methods , 78 : 79 - 85 , 2009 ) , in which the latter is buk , erm? cassette flanked by loxP sites and ~ 1 kb down an empty plasmid used as a background control , the fer - 20 stream homology arm was assembled ( SEO ID NO : 160 ) . mentation of such strain resulted in a production of 3 - HB with highest titer at 1 . 68 g / L at day 10 ( FIG . 42A ) . When The ~ 1 kb upstream and downstream homology arms PMTL82256 - buk - ptb , instead of the empty plasmid were PCR amplified from C . autoethanogenum with primers PMTL82256 , was coexpressed with pMTL83159 - phaB SN22f / SN23r and SN28f / SN29r . Ptb and buk genes were th1 A in C . autoethanogenum , the fermentation resulted in a 25 PCR amplified from plPA _ 16 plasmid using primers SN2417 higher titter of 3 - HB , at 4 . 76 g / L , at an earlier time , day 4 SN25r . The erm? cassette with loxP sites was PCR ampli ( FIG . 42B ) . fied using primers SN26f / SN27r . The plasmid backbone was
Deletion of native thioesterases enhances the efficiency of PCR amplified with primers SN30f / SN31r . KAPA poly the ptb - buk system , which has preference for ( R ) - 3 - HB - merase was used for all PCR amplifications . The PCR COA . The locus of the thioesterase gene in the genome was 30 products were assembled using GeneArt Seamless cloning deleted and replaced with the buk - ptb dna fragment via the kit from Life Technologies and plasmid with no mutations in common molecular biology technique known as homolo - the insert fragments was used to transform C . autoethano gous recombination . The substitution of the thioesterase genum by conjugation as described earlier . gene by the buk - ptb was confirmed by PCR , followed by following coniugation and selection on trimethoprim and agarose gel electrophoresis and dna sequencing . clarithromycin , 7 colonies were streaked twice on PETC In a bottle experiment , when pMTL83156 - phaB - thlA was MES agar plates with clarithromycin and anhydrotetracy expressed without ptb - buk in the thioesterase deletion cline to induce the expression of mazF genes . The colonies mutant , described above , the average maximum titer of from clarithromycin and anhydrotetracycline should have 3 - HB produced was 0 . 50 + 0 . 05 g / L , similar to the titer obtained using an unmodified C . autoethanogenum strain . 40 the pta and ack genes replaced with ptb and buk genes and
erm? cassette . This was verified by PCR using primers When pMTL82256 - buk - ptb was coexpressed with the PMTL83156 - phaB - thlA plasmid in a thioesterase knockout Og29f / Og30r flanking the homology arms and KAPA poly strain , the production of 3 - HB increased to 1 . 29 + 0 . 10 g / L merase ( FIG . 25 ) . While a band of - 4 . 6 kb is amplified from ( FIG . 43 ) . the wildtype strain , bands of ~ 5 . 7 kb was amplified from
colonies 1 and 4 - 7 , indicating the replacement of pta and ack Example 8 genes replaced with ptb and buk genes and erm? cassette .
The above event was further confirmed by sequencing the This example demonstrates that it is possible to eliminate PCR products from clones 4 - 7 .
acetate production system in an acetogen C . autoethanoge With the resulting modification the expression of ptb and num with the Ptb - buk system . buk genes is driven by the promoter upstream of pta gene .
SEQ ID NO : Name Sequence
161 SN22f TTTACAAATTCGGCCGGCCAAAGATTGCTCTATGTTTAAGCT 162 SN23r CATCAAAGTTTTTACTCATCAATTTCATGTTCATTTcc?ccc? 163 SN24f AGGGAGGAAATGAACATGAAATTGATGAGTAAAAACTTTGAT
GAGT 164 SN25 r GTATAGCATACATTATACGAACGGTACTAGTAAACCTTAGCTT
GT TCTIC 165 SN26f GAAGAACAAGCTAAGGTTTACTAGTACCGTTCGTATAATGTAT
GCTATAC 166 SN27r AGAGATGAGCATTAAAAGTCAAGTCTACCGTTCGTATAGCATA
CA 167 SN28f TGTATGCTATACGAACGGTAGACTTGACTTTTAATGCTCATCTC
T 168 SN29r CATGAGATTATCAAAAAGGAGTTTAAATATCTATTTTGTCCTTA
GGA

US 9 , 957 , 531 B1 75
- continued SEO ID NO : Name Sequence
169 SN3 Of TCCTAAGGACAAAATAGATATTTAAACTCCTTTTTGATAATCTC ATG
170 SN31r AGCTTAAACATAGAGCAATCTTTGGCCGGCCGAATTTGTAAA 171 Og29f AGCCACATCCAGTAGATTGAACTTT 172 Og3 Or AATTCGCCCTACGATTAAAGTGGAA
The resulting strain C . autoethanogenum pta - ack : : ptb - buk , in which the pta - ack operon was replaced by the
the aor1 genes to form plasmid pMTL82256 - aorl . The following primers were used .
SEQ ID NO : Name Sequence Direction
173 Pfdx - Fi AAAGGTCTCCGGCCGCGCTCACTATCTGCGGAAC forward
174 Pfdx - Ri TTTGGTCTCGAATTCTGTAACACCTCCTTAATTTTreverse TAG
175 aorl - Fi AAAGGTCTCGAATTCAAAGATCTATGTATGGTTA forward TGATGGTAAAGTATTAAG
176 aorl - Ri TTTGGTCTCCTCGAGTATGGATCCCTAGAACTTACreverse CTATATATTCATCTAATCC
ptb - buk operon was transformed as described above with the After transforming the resulting plasmid pMTL82256 isopropanol production plasmid PMTL85147 - th1A - adc from aor1 into the E . coli CA434 strain , conjugation was per Example 2 . A growth study was carried out under auto - formed on the previous C . autoethanogenum 1 , 3 - BDO pro trophic conditions and analyzed for metabolic end products . 30 duction host . Thus , the resulting C . autoethanogenum strain No acetate production was observed , while isopropanol ( up carried two plasmids , one for overexpressing hbd1 and th1A , to 0 . 355 g / L ) and 3 - HB ( up to 0 . 29 g / L ) was still produced and another for aor1 , under different replication origins and alongside ethanol and 2 , 3 - butanediol ( FIGS . 39A and 39B ) . selection marker . The production for 1 , 3 - BDO was charac This demonstrates that it is possible to produce isopropanol terized and quantified following the procedures above . and 3 - HB without acetate production from gaseous sub - 35 The results clearly show that 1 , 3 - BDO production can be strates CO and / or CO2 and H , using the Ptb - Buk system .
If acetone rather than isopropanol is the target product , the improved by overexpressing aor1 . Likewise other aldehyde : ferredoxin oxidoreductase genes could be expressed in C . primary : secondary alcohol dehydrogenase gene ( SEQ ID
NO : 17 ) can be further knocked out this strain C . autoetha autoethanogenum to facilitate conversion of 3 - hydroxybu nogenum pta - ack : : ptb - buk using methods described above 40 tyrate to 1 , 3 - butanediol . and in detail in WO 2015 / 085015 . Introducing plasmid To improve of 1 , 3 - BDO production , AOR was overex pMTL85147 - th1A - adc into this strain results in production pressed to improve conversion of 3 - HB to 3 - HB - aldehyde . of acetone at similar levels as described above for isopro To do this , pMTL82256 - hbd - th1A and pMTL83159 - aor1 panol without co - production of acetate . Ethanol , 2 , 3 - butane were coexpressed in C . autoethanogenum . As compared to diol and 3 - HB may be further products . 45 the strain that carried PMTL82256 - hbd - th1A alone , the aorl
By further knock - outs it is possible to eliminate these coexpressed strain produced higher ethanol and 1 , 3 - BDO products as well , e . g . , knock - out of the acetolactate decar ( FIG . 44 ) . boxylase gene BudA results in a strain unable to produce 2 , 3 - butanediol ( U . S . Pat . No . 9 , 297 , 026 ) . 3 - HB production Example 10 may be reduced or eliminated by deletion of 3 - hydroxybu - 50 tyrate dehydrogenase gene Bdh ( SEQ ID NO : 62 ) . This example demonstrates the stereospecificity of Ptb
Buk that allows for the production of 2 - hydroxyisobutyric Example 9 acid without the production of unwanted byproducts .
2 - hydroxyisobutyic acid can be produced in E . coli and C . This example demonstrates improvement of conversion 55 autoethanogenum by introduction of a thiolase and a 3 - hy
of 3 - hydroxybutyrate to 1 , 3 - BDO by overexpression of the droxybutyryl - CoA dehydrogenase to convert acetyl - CoA to aldehyde : ferredoxin oxidoreductase gene aor1 . 3 - hydroxybutyryl - CoA , a 2 - hydroxyisobutyryl - CoA mutase
The pMTL82251 plasmid backbone was used for over enzyme for conversion of 3 - hydroxybutyryl - CoA to 2 - hy expression of the C . autoethanogenum aor1 gene . The droxyisobutyryl - CoA and an enzyme that can hydrolyse the pMTL82251 plasmid was selected since it has a different 60 COA to form 2 - hydroxyisobutyric acid . The 3 - hydroxybu replication origin and antibiotic marker , but could be co - tyryl - CoA dehydrogenase can either be ( R ) - or ( S ) - specific expressed with , the plasmid used in Example 5 that con - and the enzyme converting 2 - hydroxyisobutyryl - CoA to tained hbd1 and th1A . Preparation of the plasmid backbone 2 - hydroxybutyrate according to steps 1 , 13 , 19 , and 20 of and the assembly reaction were carried out following the FIG . 1 . This last step can either be done via a thioesterase or procedures listed above , first generating plasmid 65 the Ptb - Buk system . PMTL82256 by introducing the C . autoethanogenum ferre - Three potential candidate genes , E . coli thioesterase type doxin promoter into plasmid pMTL82251 and then adding II TesB , the C . autoethanogenum phosphate acetyltrans

US 9 , 957 , 531 B1 77 78
ferase / acetate kinase pair and the C . beijerinckii butyryl - transferase / butyrate kinase pair were cloned into E . coli PDUET T7 expression vectors via methods described above and primers below .
( SEQ ID NO : 188 ) . Hcm ) and hcmB encode a 2 - hydroxy isobutyryl - CoA mutase and meaB a chaperon from Aquin cola tertiaricarbonis , in the construct hcmA and meaB genes were fused together as one protein as described ( SEQ
SEQ ID NO : Primer Sequence
177 PETDuet - pta - ack - GGGTACCTTATTTATTTTCAACTATTTCTTTTGTATC ack - Duet12 - R1
178 PETDuet - pta - ack - TTGAAAATAAATAAGGTACCCTCGAGTC TGGTAAAG Duet12 - ack - F1
179 PETDuet - pta - ack - ??????ccATATGTAT?T??cc?????????????? Duet12 - pta - R1
180 PETDuet - pta - ack - AGGAGATATACATATGGAAAAAATTTGGAGTAAGGC pta - Duet12 - F 1
181 PETDuet - tesB - GAAATCATAATTAAGGTA CC CTCGAGTCTGGTAAAG Duet12 - tesB - F1
182 PETDuet - tesB - CCTGACTCATATGTATATCTCCTTCTTATACTTAAC Duet12 - tesB - R1
183 PETDuet - tesB - tesB - AAGAAGGAGATATACATATGAGTCAGGCACTTAAAA Duet12 - F1
184 PETDuet - tesB - AGGGTACCTTAATTATGATTTCTCATAACACCTTC testB - Duet12 - R1
The obtained plasmids PDUET - pta - ack ( SEQ ID NO : ID NO : 189 ) ( Yaneva , J Biol Chem , 287 : 15502 - 15511 , 185 ) , PDUET - ptb - buk ( SEQ ID NO : 186 ) , pDUET - tesB 25 2012 ) . The gene cassette was cloned into either a plasmid ( SEQ ID NO : 187 ) and introduced into E . coli BL21 ( DE3 ) containing thiolase ( th1A from C . acetobutylicum ; SEQ ID for expression and then assayed for their activity on NO : 136 ) and an ( S ) - specific 3 - hydroxybutyrate dehydro acetoacetyl - CoA , 3 - hydroxybutyryl - CoA and 2 - hydroxy genase ( hbd from C . acetobutylicum ; SEQ ID NO : 190 ) isobutyryl - CoA . The results are shown in FIG . 27 . E . coli ( PMTL83155 - thlA - hbd ) or an ( R ) - specific 3 - hydroxybu BL21 has a small but measurable amount of activity on all 30 tyrate dehydrogenase ( phaB from R . eutropha ) three substrates . Pta - Ack resulted in no activity above back - ( PMTL83155 - th1A - phaB ) using the restriction enzymes ground , while both thioesterase TesB and Ptb - Buk showed Kpnl and Ncol to form plasmids pMTL83155 - th1A - hbd high activity on all three substrates , including 2 - hydroxy Pwl - meaBhcmA - hcmB ( SEQ ID NO : 191 ) and isobutyryl - CoA . - PMTL83155 - th1A - phaB - Pwl - meaBhcmA - hcmB ( SEO ID
The activity of both thioesterase TesB and Ptb - Buk was NO : 192 ) , respectively . Sub - cloning of the codon optimized higher on linear acetoacetyl - CoA , 3 - hydroxybutyryl - CoA 2 - hydroxyisobutyryl - CoA mutase casette in E . coli Top - 10 than on branched 2 - hydroxyisobutyryl - CoA . This creates a was only successful after some initial cloning complications ; problem in the pathway as it results in early termination of it was found that the 2 - hydroxyisobutyryl - CoA mutase the pathway at 3 - hydroxybutyryl - CoA , in particular as 40 casette could only be cloned into the plasmid at a lower activities are higher than activities on the 2 - hydroxyisobu - temperature ( 28° C . ) . tyryl - CoA mutase enzyme . Vector pMTL83155 - th1A - hbd and pMTL83155 - th1A However , Ptb - Buk in contrast to thioesterases is able to phaB were created by first amplifying a promoter region of
distinguish between stereoisomers and will only ( or prefer the phosphate acetyltransferase of C . autoethanogenum entially ) act on ( R ) - 3 - hydroxybutyryl - CoA but not on ( S ) - 45 ( SEQ ID NO : 193 ) and cloning into vector pMTL83151 3 - hydroxybutyryl - CoA . This was demonstrated by express - ( F1797647 . 1 ; Heap , Microbiol Meth . 78 : 79 - 85 , 2009 ) ing the Ptb - Buk system either with ThlA and ( S ) - specific using Not? and Ndel restriction sites before introducing Hbd ( FIG . 28A ) or ( R ) - specific phaB ( FIG . 28B ) in the genes th1A and hbd or respectively phaB via Ndel and KpnI pDuet system in E . coli . The constructs were constructed as in a double ligation reaction . described in Examples 1 and 3 . Growth studies confirmed 50 In addition , compatible plasmid modules for expressing that appreciable amounts of 3 - hydroxybutyrate were only ptb - buk or tesB were built . For this , the respective genes formed when Ptb - Buk was expressed in combination with were amplified from genomic DNA and introduced into the ( S ) - specific Hbd but not the ( R ) - specific phaB . plasmid pMTL82256 described in Example 9 and then Therefore , a route via an ( S ) - specific 3 - hydroxybutyryl CoA dehydrogenase and the Ptb - Buk provides significant 55 ss introducing either ptb - buk or phaB using Ndel and Ncol and advantages , as the Ptb - Buk system ( unlike thioesterases ) is Seamless Cloning kit ( Life technologies ) to form plasmids not active on ( S ) - 3 - hydroxybutyryl - CoA but ( S ) - 3 - hydroxy PMTL82256 - ptb - buk ( SEQ ID NO : 194 ) and PMTL82256 butyryl - CoA is also the preferred isomer of the 2 - hydroxy - tesB ( SEQ ID NO : 195 ) . isobutyryl - CoA mutase ( Yaneva . J Biol Chem . 287 : 15502 - Plasmids pMTL83155 - thlA - hbd - Pwl - meaBhcmA - hcmB . 15511 , 2012 ) . The produced 2 - hydroxyisobutyryl - CoA can 60 PMTL83155 - thlA - phaB - Pwl - meaBhcmA - hcmB , then be used via the Ptb - Buk to produce 2 - hydroxyisobu - pMTL82256 - ptb - buk and pMTL82256 - tesB were intro tyric acid and ( unlike thioesterases ) 2 - hydroxyisobutyryl - duced into E . coli Top - 10 ( all steps at 28° C . ) and C . COA hydrolysis provides additional energy ( FIG . 8 ) . autoethanogenum by transformation as described in previ Modular constructs were designed to compare perfor - ous examples in the following combinations : PMTL83155
mance of the pathway . A gene cassette containing the 65 th1A - hbd - Pwl - meaBhcmA - hcmB + pMTL82256 - ptb - buk , Wood - Ljungdahl promoter in front of the genes meaB , PMTL83155 - th1A - hbd - Pwl - meaBhcmA - hcmB + hcmA and hcmB was codon optimized and synthesized PMTL82256 - tesB , PMTL83155 - th1A - phaB - Pwl - meaBh

79 US 9 , 957 , 531 B1
80 cmA - hcmB + pMTL82256 - ptb - buk and pMTL83155 - th1A including the control . By far the highest production occurred phaB - Pwl - meaBhcmA - hcmB + pMTL82256 - tesB . in strain carrying plasmid pMTL83155 - th1A - hbd - Pwl - me
Growth experiments were carried out with E . coli in LB aBhcmA - hcmB + pMTL82256 - ptb - buk ( 10x higher than all medium at 30° C . for 4 days and C . autoethanogenum in other routes ) , that has the optimal pathway with thiolase , PETC medium with 30 psi CO - containing steel mill gas 5 ( S ) - specific ( S ) - specific 3 - hydroxybutyryl - CoA dehydroge ( collected from New Zealand Steel site in Glenbrook , NZ ) nase , the 2 - hydroxyisobutyryl - CoA mutase , and the Ptb - Buk at 30° C . and 37° C . for 6 days . Metabolites were measured system ( FIGS . 29A - D ) . Surprisingly , also production of as described above . In addition to measurement by GC - MS , 2 - hydroxybutyrate ( 2 - HB ) ( up to 64 mg / L by LC - MS / MS 2 - Hydroxyisobutyric acid production was also confirmed and 50 mg / L by GC - MS in C . autoethanogenum ; 12 mg / L using liquid chromatography tandem mass spectrometry 10 by LC - MS / MS and 9 . 5 mg / L by GC - MS in E . coli ) was ( LC - MS / MS ) and ' H nuclear magnetic resonance ( NMR ) found in this strain , indicating unspecific mutase activity spectroscopy .
Liquid chromatography tandem mass spectrometry ( LC ( FIG . 30 ) . This was also found in the tesB strain , but again MS / MS ) data was acquired on a Dionex UltiMate 3000 at significant lower levels ( 18 mg / L in LC - MS - MS and 9 liquid chromatography system ( Dionex . California , USA ) 15 mg / L in GC - MS in C . autoethanogenum ) . Production of coupled to an ABSciex 4000 OTRAP mass spectrometer 2 - hydroxyisobutyric acid was also confirmed by NMR . ( ABSciex , Concord , Canada ) . The liquid chromatography In addition , also qRT - PCR was carried out to confirm system was controlled by Chromeleon software ( Dionex ) , expression of the genes th1A , hbd , meaBhcmA and hcmB and chromatographic separation was achieved by injecting ( FIG . 31 ) . 104 onto a Gemini - NX C18 150 mmx2 mm I . D . , 3 um 110 20 The RT - PCR graphs show that th1A gene product is A particle column ( Phenomenex , Aschaffenburg , Germany ) equipped with a pre - column Security Guard Gemini - NX expressed to slightly higher levels with the Ppta - ack promoter C18 4 mmx2 mm I . D . cartridge . The column oven tempera than hbd ( as expected with a second gene in an operon ) and ture was controlled and maintained at 55° C . throughout the that hmcB shows slightly lower expression levels than acquisition and the mobile phases were as follows : 7 . 5 mM 25 meaBhcmA . Also there is lower expression in C . autoetha aqueous tributylamine adjusted to pH 4 . 95 ( + 0 . 05 ) with nogenum at 30° C . than at 37° C . and E . coli at 30° C . For glacial acetic acid ( eluent A ) and acetonitrile ( eluent B ) . The specific cycle numbers see below . mobile phase flow rate was maintained at 300 UL / min throughout a gradient profile and was introduced directly into the mass spectrometer with no split . The mass spec - 30 Condition Target Cq Mean Cq Std Dev trometer was controlled by Analyst 1 . 5 . 2 software ( AB - E . coli 30° C . thlA thla 18 . 26 0 . 243 Sciex ) and was equipped with a TurboV electrospray source hbd 20 . 6 0 . 603 operated in negative ionisation mode . The following previ 16 . 20 0 . 108
18 . 30 0 . 666 ously optimized ( and therefore general ) parameters were C . autoethanogenum / 30° C . thlA 26 . 10 0 . 169 used to acquire scheduled Multiple Reaction Monitoring 35 Hbd 27 . 54 0 . 415 ( MRM ) data : ionspray voltage - 4500V , nebulizer ( GS1 ) , meaBhcmA 20 . 63 0 . 604
auxiliary ( GS2 ) , curtain ( CUR ) and collision ( CAD ) gases hmcB 22 . 64 0 . 697 C . autoethanogenum / 37° C . thlA 0 . 069 were 60 , 60 , 20 and medium ( arbitrary units ) , respectively , hbd 21 . 85 0 . 222 generated via a N300DR nitrogen generator ( Peak Scientific , meaBhcmA 16 . 72 0 . 119
Massachusetts , USA ) . The auxiliary gas temperature was 40 hmcB 19 . 62 0 . 173
maintained at 350° C . The entrance potential ( EP ) was - 10 volts . This method is also able to detect and separate 2 - hydroxybutyric acid . The ratio of ( S ) - 3 - hydroxybutyric acid to ( R ) - 3 - hydroxy
' H nuclear magnetic resonance ( NMR ) spectroscopy at a butyric acid was measured by high - performance liquid chro field strength of 400 MHz . Samples were prepared by 45 matography ( HPLC ) on an Agilent 1260 Infinity LC with diluting 400 uL of sample with 400 uL of 20 mM phosphate UV detection at 210 nm . Samples were prepared by cen buffer prepared with D , O and containing trimethylsilyl trifugation at 14 , 000 rpm for 3 minutes , followed by evapo proprionic acid ( TMSP ) as internal standard ( pH of 7 ) . The ration of 200 uL of supernatant to dryness . The pellet was samples were then transferred glass NMR tube ( 5 mmx8 then re - suspended in 100 % Isopropanol and sonicated under inches ) and analysed by " H NMR using presaturation for 50 heat for 1 hour . Centrifugation was repeated and the super water suppression with a 30° excitation pulse , 15 second natant transferred to an HPLC vial for analysis . Separation relaxation delay and 64 scans at a temperature of 27° C . was achieved with a 5 uL injection on to a TCI Chiral MB - S Once acquired the spectrum was transformed , flattened and column ( 250 mmx4 . 6 mmx3 um ) at 1 . 5 mL / min and 40° C . integrated using Agilent Vnmr software . The known con under isocratic conditions , using 95 - 5 hexane - isopropanol centration of TMSP was used for quantitation of 2 - hydroxy - 55 mobile phase containing 0 . 1 % trifluoracetic acid . isobutyric using the resonance at 1 . 36 ppm ( singlet ) .
In both E . coli growing heterotrophically as well as C . A stereospecific analysis of produce 3 - HB has been autoethanogenum growing autotrophically . 2 - hydroxyisobu - performed . Surprisingly it was found that in C . autoetha tyric acid could be detected in constructs PMTL83155 - thlA nogenum , a mix of isomers was produced . Enzymes Hbd bbd - Pwl - mea Bhom A - hemBAOMTL82256 - tesB ( 1 . 5 mg / L 60 and PhaB are described to be stereospecific , PhaB is R - spe in LC - MS / MS method and 8 mg / L in GC - MS in . c . cific and Hbd is S - specific and when expressing these autoethanogenum : 0 . 5 mg / L in LC - MS / MS method and 2 enzymes in E . coli a stereopure product has been observed mg / L in GC - MS in E . coli ) and pMTL83155 - th1A - phaB - ( Tseng , Appl Environ Microbiol , 75 : 3137 - 3145 , 2009 ) . Pwl - meaBhcmA - hcmB + pMTL82256 - ptb - buk ( 15 mg / L in The following table indicates the distribution of ( R ) - and LC - MS / MS method and 75 mg / L in GC - MS in C . autoetha - 65 ( S ) - form of 3 - HB at equilibrium produced via three different nogenum ; 1 . 1 mg / L in LC - MS / MS method and 8 . 5 mg / L in routes in C . autoethanogenum . These data suggest the GC - MS in E . coli ) , but not in constructs all other constructs presence of isomerase in the C . autoethanogenum .
meaBhcmA hmcB
18 . 48

81
vwa ???
15
US 9 , 957 , 531 B1 82
product production , allowing the construction of stable and Route % R - form % S - form high - yielding fermentation strains . in strains
THIA - PhaB 55 7 A genome - scale metabolic model of C . autoethanogenum 53 + 5 THIA - HBD 12 3 88 + 3 similar to the one described by Marcellin , Green Chem , 18 : ThlA - ctfAB 16 + 7 84 + 7 3020 - 3028 , 2006 was utilized . Variants of this model were
created that incorporate additional metabolic reactions , each Knockout of native isomerases may prevent interconver one representing a different genetically modified microor
sion of ( R ) and ( S ) forms of 3 - HB . Alternatively , expression ganism for non - native product formation . Three model ver or overexpression of isomerases could enable new ptb - buk 10 sions were created for each non - native product pathway , routes . For example , Hbd could be used to generate ( S ) - 3 10 incorporating either a thioesterase , acetate COA - transferase HB , isomerase could convert ( S ) - 3 - HB to ( R ) - 3 - HB , and or Ptb - Buk reaction . ptb - buk could act on ( R ) - 3 - HB to produce products of Maximum selectivities were calculated using flux balance
analysis ( FBA ) , using scripts from the COBRA Toolbox interest . V2 . 0 in MATLAB R2014a ( The Mathworks , Inc . ) with Example 11 Gurobi version 6 . 0 . 4 as the solver ( Gurobi Optimization ,
Inc . ) . Exchange reactions were constrained to represent a chemically defined minimal growth medium with CO as the This example demonstrates the production of isobutylene source of carbon and energy . An evolutionary algorithm was via Ptb - Buk conversion of 3 - hydroxyisovaleryl - CoA and used to search for the existence of strain designs incorpo 3 - hydroxyisovalerate . 20 rating up to ten gene knockouts that couple target non - native
Different routes for production of isobutylene have been chemical production with growth . described , for example the conversion of acetone to isobu - FBA predicts that pathways using Ptb - Buk or CoA trans tylene via a hydroxyisovalerate synthase and decarboxylase ferase offer the highest product selectivities due to ATP gain ( van Leeuwen , Appl Microbiol Biotechnol , 93 : 1377 - 1387 , through substrate level phosphorylation . The results are 2012 ) . However , the hydroxyisovalerate decarboxylase step 25 illustrated in Table 2 . However , it should be noted that one is an ATP requiring step and kinetics of this enzyme may not limitation of Genome - scale models and FBA analysis is that be ideal . Two alternative routes to isobutylene using the enzyme kinetics are not captured . The CoA transferase Ptb - Buk system have been identified through 3 - hydroxyis reaction requires a certain base level of acetate for function ovaleryl - CoA which has been shown in vitro to be a viable ality , therefore in reality the maximum selectivity using a substrate for the Ptb - Buk system ( Liu , Appl Microbiol 30 CoA transferase would be less than 100 % due to a base level Biotechnol , 53 : 545 - 552 , 2000 ) . of acetate required to be present .
Alternative pathway 1 consists of a synthase that converts acetone into 3 - hydroxyisovaleryl - CoA ( FIG . 9 ) .
Alternative pathway 2 proceeds via known intermediate Maximum selectivity % ( C in target 3 - methyl - 2 - oxopentanoate of the isoleucine biosynthesis 35 product / C in all fermentation products ) that is common to bacteria such as E . coli or C . autoetha Non - native product Thioesterase COA - transferase Ptb - Buk nogenum ( FIG . 10 ) .
Acetone 82 . 0 100 100 Example 12 Isopropanol 82 . 1 100 100
40 Isobutylene 55 . 9 80 . 2 3 - Hydroxybutyrate 86 . 0 100 This example describes methods for characterizing Ptb 1 , 3 - Butanediol 88 . 6
Buk variants . 2 - Hydroxyisobutyrate Given the substrate promiscuity of Ptb - Buk , it is likely
that Ptb - Buk systems of varying amino acid sequences will Table 2 . Flux balance analysis ( FBA ) showing the maxi possess varying preferences for given substrates . In order to 45 identify a Ptb - Buk system that favors a desired substrate mum possible non - native product selectivities in C . ( e . g . acetoacetyl - CoA , 3 - hydroxybutyryl - CoA , 2 - hydroxy autoethanogenum for a set of products and candidate
enzymes . isobutyryl - CoA , acetyl - CoA , and / or butyryl - CoA ) , a high It is desirable to construct strains where the target non throughput screen is desirable . Such a screen can be accom native chemical must be produced for cell growth . FBA plished by coupling firefly luciferase ( Luc ) to the Ptb - Buk 50 predicts that in most cases it would be difficult to couple system ( FIG . 33 ) . Luc reacts with D - luciferin , generating target chemical production with growth when using a thio oxyluciferin , carbon dioxide , and light . In addition to mag esterase or a CoA transferase ; instead , native products nesium and molecular oxygen , Luc requires ATP for the acetate and ethanol would be favored . However , when using reaction to proceed . ATP is a product generated by Ptb - Buk when provided an appropriate acyl - CoA or enoyl - CoA sub - 55 Ptb - Buk , many growth - coupled chemical production strain
designs exist , often incorporating a disruption of the phos strate . Therefore , Ptb - Buk reaction rates and preferences can photransacetylase - acetate kinase reactions . Table 3 summa be compared for varying substrates by quantifying the amount of light generated by a reaction containing Ptb - Buk , rizes the growth coupling ability of each strain . Luc , d - luciferin , magnesium , molecular oxygen , phosphate , ADP , and an acyl - CoA or enoyl - CoA . 60 Ability to couple non - native chemical
production with growth Example 13 Non - native product Thioesterase COA - transferase Ptb - Buk
This example uses genome - scale modeling to demon Acetone No Yes strate that high non - native product selectivities can be 65 Isopropanol No Yes achieved using Ptb - Buk . Furthermore , it shows that the use Isobutylene No No No of Ptb - Buk could permit the coupling of cellular growth with
80 . 2 100 100 100
100 100 86 . 0
No No

US 9 , 957 , 531 B1 83 84
No
No
10
- continued can be engineered to use a range of non - natural CoAs as substrates . Likewise AOR enzyme has been shown to act on
Ability to couple non - native chemical a wide range of substrates . Together these two enzymes can production with growth convert a wide range of CoAs via their acids into aldehydes , Non - native product Thioesterase COA - transferase Ptb - Buk which then can be further converted to alcohols , ketones or
enols via alcohol dehdydrogeneses , for which a wide variety 3 - Hydroxybutyrate No Yes exists in nature . While under standard conditions the reduc 1 , 3 - Butanediol No Yes Yes 2 - Hydroxyisobutyrate No Yes tion of acids with ferredoxin to aldehydes via the AOR is
endergonic ( Thauer , Bacteriol Rev , 41 : 100 - 180 , 1977 ) and as such not feasible , it surprisingly is in carboxydotrophic Table 3 . Potential to couple non - native chemical produc acetogens such as C . autoethanogenum that operate at low tion with growth in C . autoethanogenum during growth on pH and with CO or H , as substrate ( Mock , J Bacteriol , 197 : CO when reconfiguring the metabolic network with up to ten 2965 - 2980 , 2015 ) . One common limitation working with gene knockouts .
While both Ptb - Buk and CoA transferase can support high 15 15 acetogens is that they are ATP - limited , living on the ther selectivities , flux balance analysis predicts that in most modynamic edge of life ( Schuchmann , Nat Rev Microbiol , cases , only Ptb - Buk would allow the construction of stable , 12 : 809 - 821 , 2014 ) , which can be overcome by coupling this high - yielding fermentation strains that couple non - native acid reduction to ATP - linked formation of acids from CoAs chemical production with growth . via the Ptb - Buk system .
20 The Ptb - Buk system and AOR system has been demon Example 14 strated in above examples for several different products , but
can be extended to further products , for example production This example demonstrates the production of adipic acid of 2 - buten - 1 - o1 , 3 - methyl - 2 - butanol , 1 , 3 - hexanediol ( HDO ) .
via Ptb - Buk from gaseous feedstock . 2 - Buten - 1 - ol can be produced via Ptb - Buk , AOR and an Production of adipic acid in E . coli from sugar has been 25 alcohol dehydrogenase from crotonyl - CoA ( FIG . 35 ) . 1 , 3
described by a pathway utilizing Ptb - Buk ( Yu , Biotechnol Hexanediol can be produced via Ptb - Buk , AOR and an Bioeng , 111 : 2580 - 2586 , 2014 ) . However production was alcohol dehydrogenase from 3 - hydroxy - hexanoyl - CoA low , in the ug / L range . Without wishing to be bound by any ( FIG . 35 ) . By combining Ptb - Buk , Adc and an alcohol particular theory , the inventors believe that this is likely a dehydrogenase ( such as native primary : secondary alcohol function of lacking driving force in forms of reducing power 30 dehydrogenase ) , 3 - methyl - 2 - butanol can be formed from and surplus ATP . Using a reduced gaseous substrate as CO acetobutyryl - CoA . and H , and an acetogenic bacterium such as C . autoetha All of these precursors , crotonyl - CoA , 3 - hydroxy nogenum , this current limitation can be overcome . CO and hexanoyl - CoA , or acetobutyryl - CoA can be formed by H oxidation provide sufficient driving force for reduction of reduction and elongation of acetyl - CoA , acetoacetyl - CoA 3 - oxo - adipyl - CoA to 3 - hydroxyadipyl - CoA by 3 - hydroxy - 35 and 3 - HB - CoA which are described in previous examples butyryl - CoA dehydrogenase or acetoacetyl - CoA hydratase via known fermentation pathways of , for example , and 2 , 3 - dehydroadipyl - CoA to adipyl - CoA by enoyl - CoA Clostridium kluyveri ( Barker , PNAS USA , 31 : 373 - 381 , hydrolase or enoyl - CoA reductase ( FIG . 34 , steps 23 and 1945 ; Seedorf , PNAS USA , 105 : 2128 - 2133 , 2008 ) and other 25 ) , in contrast to E . coli growing heterotrophically on more Clostridia . Involved enzymes include crotonyl - CoA oxidized sugars . Acetogenic bacteria live on the energetic 40 hydratase ( crotonase ) or crotonyl - CoA reductase , butyryl limit of life and therefore ATP generating reactions like the COA dehydrogenase or trans - 2 - enoyl - CoA reductase , thio Ptb - Buk system have a strong driving force , ensuring effi - lase or acyl - CoA acetyltransferase and 3 - hydroxybutyryl cient conversion of adipyl - CoA to adipic acid ( FIG . 34 , step COA dehydrogenase or acetoacetyl - CoA hydratase ( FIG . 26 ) , in contrast to E . coli growing heterotrophically on 35 ) . Respective genes from C . kluyveri or other Clostridia sugars generating surplus ATP from glycolysis . 45 have be cloned on an expression plasmid ( U . S . 2011 /
To produce adipic acid from gas in C . autoethanogenum , 0236941 ) and and then transformed as described above in C . genes encoding a succinyl - CoA synthetase from E . coli autoethanogenum strains pta - ack : : ptb - buk or ( NP _ 415256 , NP _ 415257 ) , a ketoisovalerate oxidoreduc - CAETHG _ 1524 : : ptb - buk from previous examples for pro tase PaaJ from E . coli ( WP 001206190 . 1 ) , a 3 - hydroxybu - duction of 2 - buten - 1 - ol , 3 - methyl - 2 - butanol , 1 , 3 - hexanediol tyryl - CoA dehydrogenase Hbd from Clostridium beijer - 50 ( HDO ) . 2 - Buten - 1 - ol , 3 - methyl - 2 - butanol , and 1 , 3 - hexane inckii ( WP _ 011967675 . 1 ) , a trans - 2 - enoyl - CoA reductase diol ( HDO ) may be precursors for further downstream Crt from C . acetobutylicum ( NP _ 349318 . 1 ) , trans - 2 - enoyl - products . CoA reductase Bcd from C . acetobutylicum ( NP _ 349317 . 1 ) While these are only a few examples , it should be clear and electron flavoproteins EtfAB ( NP _ 349315 , NP _ 349316 ) that this pathway can be further extended using the same are cloned on an expression plasmid and then transformed as 55 enzymes or engineered variants thereof that have specificity described above in C . autoethanogenum strains pta - ack : : ptb - for higher chain length to produce a range of C4 , C6 , C8 , buk or CAETHG _ 1524 : : ptb - buk from previous examples . C10 , C12 , C14 alcohols , ketones , enols or diols ( FIG . 39 ) . Adipic acid is produce according to the steps depicted in Different type of molecules can be obtained also by using FIG . 34 . primer or extender units different than acetyl - CoA in the
60 thiolase step as been described elsewhere ( Cheong , Nature Example 15 Biotechnol , 34 : 556 - 561 , 2016 ) .
All references , including publications , patent applica This example demonstrates the production of various tions , and patents , cited herein are hereby incorporated by
products including 2 - buten - 1 - ol , 3 - methyl - 2 - butanol , 1 , 3 - reference to the same extent as if each reference were hexanediol ( HDO ) via Ptb - Buk and AOR . 65 individually and specifically indicated to be incorporated by As demonstrated in Example 6 , Ptb - Buk is highly pro - reference and were set forth in its entirety herein . The
miscuous and acts on a wide range of CoAs as substrates or reference to any prior art in this specification is not , and

US 9 , 957 , 531 B1 85 86
should not be taken as , an acknowledgement that that prior of any and all examples , or exemplary language ( e . g . , " such art forms part of the common general knowledge in the field as ” ) provided herein , is intended merely to better illuminate of endeavour in any country . the invention and does not pose a limitation on the scope of | The use of the terms “ a ” and " an and the and similar | the invention unless otherwise claimed . No language in the
referents in the context of describing the invention ( espe - 5 specification should be construed as indicating any non claimed element as essential to the practice of the invention . cially in the context of the following claims ) are to be Preferred embodiments of this invention are described
construed to cover both the singular and the plural , unless herein . Variations of those preferred embodiments may otherwise indicated herein or clearly contradicted by con become apparent to those of ordinary skill in the art upon text . The terms " comprising , ” having , ” “ including , ” and 10 reading the foregoing description . The inventors expect " containing ” are to be construed as open - ended terms ( i . e . , " skilled artisans to employ such variations as appropriate , and meaning “ including , but not limited to ” ) unless otherwise the inventors intend for the invention to be practiced other noted . Recitation of ranges of values herein are merely wise than as specifically described herein . Accordingly , this intended to serve as a shorthand method of referring indi - invention includes all modifications and equivalents of the vidually to each separate value falling within the range , 16 subject matter recited in the claims appended hereto as unless otherwise indicated herein , and each separate value is permitted by applicable law . Moreover , any combination of incorporated into the specification as if it were individually the above - described elements in all possible variations recited herein . All methods described herein can be per thereof is encompassed by the invention unless otherwise formed in any suitable order unless otherwise indicated indicated herein or otherwise clearly contradicted by con herein or otherwise clearly contradicted by context . The use text .
SEQUENCE LISTING
< 160 > NUMBER OF SEQ ID NOS : 195
A
< 210 > SEQ ID NO 1 1 > LENGTH : 392
< 212 > TYPE : PRT < 213 > ORGANISM : Clostridium acetobutylicum < 220 > FEATURE : ( 221 > NAME / KEY : MISC FEATURE < 223 > OTHER INFORMATION : ThiA , WP _ 010966157 . 1
< 400 > SEQUENCE : 1 Met Lys Glu Val Val Ile Ala Ser Ala Val Arq Thr Ala Ile Gly Ser |
Tyr Gly Lys Ser Leu Lys Asp Val Pro Ala Val Asp Leu Gly Ala Thr ? g 2 Ag 25 30
Ala Ile Lys Glu Ala Val Lys Lys Ala Gly Ile Lys Pro Glu Asp Val 35 40 45
?
Asn Glu Val Ile Leu Gly Asn Val Leu Gin Ala Gly Leu Gly Gin Asn 33 8 3
Pro Ala Arg Gln Ala Ser Phe Lys Ala Gly Leu Pro Val Glu Ile Pro .
2 0
& 3? g Ala Met Thr Ile Asn Lys Val Cys Gly Ser Gly Leu Arg Thr Val Ser mmmm Leu Ala Ala Gln Ile Ile Lys Ala Gly Asp Ala Asp Val Ile Ile Ala 105 110
Gly Gly Met Glu Asn Met Ser Arg Ala Pro Tyr Leu Ala Asn Asn Ala 115 120 125
5
mm mmmmmm mmmmm Arg Trp Gly Tyr Arg Met Gly Asn Ala Lys Phe Val Asp Glu Met Ile
Thr Asp Gly Leu Trp Asp Ala Phe Asn Asp Tyr His Met Gly Ile Thr 150 160
Ala Glu Asn Ile Ala Glu Arg Trp Asn Ile Ser Arg Glu Glu Gln Asp 175
Glu Phe Ala Leu Ala Ser Gln Lys Lys Ala Glu Glu Ala Ile Lys Ser 185 190 ge Ala Giu Glu ala Age Eye ser & Gly Gin Phe Lys Asp Glu Ile Val Pro Val Val Ile Lys Gly Arg Lys
195 200 205 |
Gly Glu Thr Val Val Asp Thr Asp Glu His Pro Arg Phe Gly Ser Thr ?

US 9 , 957 , 531 B1 88
- continued 215 220
Ile Glu Gly Leu Ala Lys Leu Lys Pro Ala Phe Lys Lys Asp Gly Thr 225 230 235 240
g Val Thr Ala Gly Asn Ala Ser Gly Leu Asn Asp Cys Ala Ala Val Leu 245 255
?
E EST E
1
Val Ile Met Ser Ala Glu Lys Ala Lys Glu Leu Gly Val Lys Pro Leu 265 270 RER g & 6 8
Ala Lys Ile Val Ser Tyr Gly Ser Ala Gly Val Asp Pro Ala Ile Met 275 280 mmmm mmm mmm Gly Tyr Gly Pro Phe Tyr Ala Thr Lys Ala Ala Ile Glu Lys Ala Gly
295
Trp Thr Val Asp Glu Leu Asp Leu Ile Glu Ser Asn Glu Ala Phe Ala 305 310 315 320
Ala Gin Ser Leu Ala Val Ala Lys Asp Leu Lys Phe Asp Met Asn Lys 325 335
Val Asn Val Asn Gly Gly Ala Ile Ala Leu Gly His Pro Ile Gly Ala 345 350
Ser Gly Ala Arg Ile Leu Val Thr Leu Val His Ala Met Gin Lys Arg | 355 360
Asp Ala Lys Lys Gly Leu Ala Thr Leu Cys Ile Gly Gly Gly Gin Gly 375
Thr Ala Ile Leu Leu Glu Lys Cys 385 390 ?? Lys cys
1 2
< 2102 SEO ID NO 2 < 211 > LENGTH : 393 < 212 > TYPE : PRT < 213 > ORGANISM : Cupri avidus necator < 220 > FEATURE : < 221 > NAME / KEY : MISC _ FEATURE < 223 > OTHER INFORMATION : PhaA , WP _ 013956452 . 1
1 3
2 0
2 1 2 22 2 3
< 400 > SEOUENCE : 2
Met Thr Asp Val Val Ile Val Ser Ala Ala Arg Thr Ala Val Gly Lys
Phe Gly Gly Ser Leu Ala Lys Ile Pro Ala Pro Glu Leu Gly Ala Val 25
Val Ile Lys Ala Ala Leu Glu Arg Ala Gly Val Lys Pro Glu Gin Val 35
D
? 24 & g
3? 3 40 1
Ser Glu Val Ile Met Gly Gin Val Leu Thr Ala Gly Ser Gly Gin Asn 6 ggg Pro Ala Arg Gln Ala Ala Ile Lys Ala Gly Leu Pro Ala Met Val Pro 65 70 80
Ala Met Thr Ile Asn Lys Val Cys Gly Ser Gly Leu Lys Ala Val Met mmm mmm am Leu Ala Ala Asn Ala Ile Met Ala Gly Asp Ala Glu Ile Val Val Ala 105 R
Gly Gly Gin Glu Asn Met Ser Ala Ala Pro His Val Leu Pro Gly Ser 115 120 8mm &
Arg Asp Gly Phe Arg Met Gly Asp Ala Lys Leu Val Asp Thr Met Ile ? mmmm Val Asp Gly Leu Trp Asp Val Tyr Asn Gin Tyr His Met Gly Ile Thr 145 150 160
Ala Glu Asn Val Ala Lys Glu Tyr Gly Ile Thr Arg Glu Ala Gin Asp

US 9 , 957 , 531 B1 8
- continued Glu Leu Ala Val Gly Ser Gin Asn Lys Ala Glu Ala Ala Gin Lys Ala
185 190
& Gly Lys Phe Asp Glu Glu Ile Val Pro Val Leu Ile Pro Gin Arg Lys 195 200 205
?
Gly Asp Pro Val Ala Phe Lys Thr Asp Glu Phe Val Arg Gin Gly Ala 220
A
& Thr Leu Asp Ser Met Ser Gly Leu Lys Pro Ala Phe Asp Lys Ala Gly 225 235 240
I
Thr Val Thr Ala Ala Asn Ala Ser Gly Leu Asn Asp Gly Ala Ala Ala 245 255
E g
Val Val Val Met Ser Ala Ala Lys Ala Lys Glu Leu Gly Leu Thr Pro 265 270
3 ? 3? gmmm Leu Ala Thr Ile Lys Ser Tyr Ala Asn Ala Gly Val Asp Pro Lys Val 275 280 285 mmmm Met Gly Met Gly Pro Val Pro Ala Ser Lys Arq Ala Leu Ser Ara Ala
300 mmmm mmm mmmm 3 Ep 3 Glu Trp Thr Pro Gln Asp Leu Asp Leu Met Glu Ile Asn Glu Ala Phe 305 315 320
Ala Ala Gin Ala Leu Ala Val His Gin Gin Met Gly Trp Asp Thr Ser 325 335
& Lys Val Asn Val Asn Gly Gly Ala Ile Ala Ile Gly His Pro Ile Gly 345 350
? Ala Ser Gly Cys Arg Ile Leu Val Thr Leu Leu His Glu Met Lys Arg 355 360 365
Arg Asp Ala Lys Lys Gly Leu Ala Ser Leu Cys Ile Gly Gly Gly Met 380
Gly Val Ala Leu Ala Val Glu Arg Lys 385
< 210 > SEQ ID NO 3 < 211 > LENGTH : 394 < 212 > TYPE : PRT < 213 > ORGANISM : Cupriavidus necator < 220 > FEATURE : 215 NAME / KEY : MISC FEATURE
< 223 > OTHER INFORMATION : BktB , WP _ 011615089 . 1 AAA < 400 > SEQUENCE : 3
Met Thr Arg Glu Val Val Val Val Ser Gly Val Arg Thr Ala Ile Gly ?
Thr Phe Gly Gly Ser Leu Lys Asp Val Ala Pro Ala Glu Leu Gly Ala & R
Leu Val Val Arg Glu Ala Leu Ala Arg Ala Gin Val Ser Gly Asp Asp 35
J 40
Val Gly His Val Val Phe Gly Asn Val Ile Gin Thr Glu Pro Arg Asp
Met Tyr Leu Gly Arg Val Ala Ala Val Asn Gly Gly Val Thr Ile Asn 70 ms mam &
am smmm mmmmmmm mmmm gmmmm Ala Pro Ala Leu Thr Val Asn Arg Leu Cys Gly Ser Gly Leu Gin Ala ?
Ile Val Ser Ala Ala Gln Thr Ile Leu Leu Gly Asp Thr Asp Val Ala
& Ile Gly Gly Gly Ala Glu Ser Met Ser Arg Ala Pro Tyr Leu Ala Pro 115 120
Ala Ala Arg Trp Gly Ala Arg Met Gly Asp Ala Gly Leu Val Asp Met

US 9 , 957 , 531 B1 01
- continued
Met Leu Gly Ala Leu His Asp Pro Phe His Arg Ile His Met Gly Val 145 150 155 160
Thr Ala Glu Asn Val Ala Lys Glu Tyr Asp Ile Ser Arg Ala Gin Gin 165 170 175
? Asp Glu Ala Ala Leu Glu Ser His Ara Ara Ala Ser Ala Ala Ile Lys 180 185 190
Ala Gly Tyr Phe Lys Asp Gin Ile Val Pro Val Val Ser Lys Gly Arg 200
Lys Gly Asp Val Thr Phe Asp Thr Asp Glu His Val Arg His Asp Ala
Thr Ile Asp Asp Met Thr Lys Leu Arg Pro Val Phe Val Lys Glu Asn 225 230 235 240
Gly Thr Val Thr Ala Gly Asn Ala Ser Gly Leu Asn Asp Ala Ala Ala 245 250 255
Ala Val Val Met Met Glu Arg Ala Glu Ala Glu Arg Arg Gly Leu Lys 260 265 270 mmmmmm mmmmm mmm mmm mmmm mmmmm mmmmm Pro Leu Ala Arg Leu Val Ser Tyr Gly His Ala Gly Val Asp Pro Lys
280 &
Ala Met Gly Ile Gly Pro Val Pro Ala Thr Lys Ile Ala Leu Glu Arg mmmm mmm mmmmm mmm mmm mmmm ms
Ala Gly Leu Gin Val Ser Asp Leu Asp Val Ile Glu Ala Asn Glu Ala 305 310 315 320
Phe Ala Ala Gin Ala Cys Ala Val Thr Lys Ala Leu Gly Leu Asp Pro 325 330 335
Ala Lys Val Asn Pro Asn Gly Ser Gly Ile Ser Leu Gly His Pro Ile 340 345 350
&
Gly Ala Thr Gly Ala Leu Ile Thr Val Lys Ala Leu His Glu Leu Asn 360 Ala Leu Ile Thr val Lys Ala Leu His Glu Leu Agn
Arg Val Gin Gly Arg Tyr Ala Leu Val Thr Met Cys Ile Gly Gly Gly 375 380
Gin Gly Ile Ala Ala Ile Phe Glu Arg Ile 385 390
< 210 > SEQ ID NO 4 < 211 > LENGTH : 394 < 212 > TYPE : PRT < 213 > ORGANISM : Escherichia coli < 220 > FEATURE :
21 > NAME / KEY : MISC _ FEATURE < 223 > OTHER INFORMATION : AtOB , NP _ 416728 . 1 A
< 400 > SEQUENCE : 4
Met Lys Asn cys Val Ile Val Ser Ala Val Arg Thr Ala Ile Gly Ser E
Phe Asn Gly Ser Leu Ala Ser Thr Ser Ala Ile Asp Leu Gly Ala Thr 25
o a
E Val Ile Lys Ala Ala Ile Glu Arg Ala Lys Ile Asp Ser Gln His Val 40 45 mmm ma Asp Glu Val Ile Met Gly Asn Val Leu Gin Ala Gly Leu Gly Gin Asn
55
Pro Ala Arg Gin Ala Leu Leu Lys Ser Gly Leu Ala Glu Thr Val Cys 75
Gly Phe Thr Val Asn Lys Val Cys Gly Ser Gly Leu Lys Ser Val Ala 85
Leu Ala Ala Gln Ala Ile Gin Ala Gly Gln Ala Gln Ser Ile Val Ala

US 9 , 957 , 531 B1 93 94
- continued 100 110
|
m ? $ m Gly Gly Met Glu Asn Met Ser Leu Ala Pro Tyr Leu Leu Asp Ala Lys
120 ?
& J
Ala Arg Ser Gly Tyr Arg Leu Gly Asp Gly Gin Val Tyr Asp Val Ile 135
Leu Arg Asp Gly Leu Met Cys Ala Thr His Gly Tyr His Met Gly Ile
Thr Ala Glu Asn Val Ala Lys Glu Tyr Gly Ile Thr Arg Glu Met Gin 165
Asp Glu Leu Ala Leu His Ser Gin Arg Lys Ala Ala Ala Ala Ile Glu 180 190
& Ser Gly Ala Phe Thr Ala Glu Ile Val Pro Val Asn Val Val Thr Arg 200
m
mmmm mmmm mmm mmm Lys Lys Thr Phe Val Phe Ser Gin Asp Glu Phe Pro Lys Ala Asn Ser
215
Thr Ala Glu Ala Leu Gly Ala Leu Arq Pro Ala Phe Asp Lys Ala Gly
Thr Val Thr Ala Gly Asn Ala Ser Gly Ile Asn Asp Gly Ala Ala Ala 245
Leu Val Ile Met Glu Glu Ser Ala Ala Leu Ala Ala Gly Leu Thr Pro 260 270
Leu Ala Arg Ile Lys Ser Tyr Ala Ser Gly Gly Val Pro Pro Ala Leu 280
mmmm
mmmmmm g 33 mmmmm mm Met Gly Met Gly Pro Val Pro Ala Thr Gin Lys Ala Leu Gin Leu Ala
295
Gly Leu Gin Leu Ala Asp Ile Asp Leu Ile Glu Ala Asn Glu Ala Phe
Ala Ala Gin Phe Leu Ala Val Gly Lys Asn Leu Gly Phe Asp Ser Glu 325
Lys Val Asn Val Asn Gly Gly Ala Ile Ala Leu Gly His Pro Ile Gly 340 350
Ala Ser Gly Ala Arg Ile Leu Val Thr Leu Leu His Ala Met Gin Ala 360
°
? Arg Asp Lys Thr Leu Gly Leu Ala Thr Leu Cys Ile Gly Gly Gly Gin 375
Gly Ile Ala Met Val Ile Glu Arg Leu Asn
AAAAA < 210 > SEQ ID NO 5 < 211 > LENGTH : 217 < 212 > TYPE : PRT < 213 > ORGANISM : Clostridium beijerinckii < 220 > FEATURE :
12 NAME / KEY : MISC _ FEATURE < 223 > OTHER INFORMATION : ctfA , WP _ 012059996 . 1
< 400 > SEQUENCE : 5
Met Asn Lys Leu Val Lys Leu Thr Asp Leu Lys Arg Ile Phe Lys Asp 5
Gly Met Thr Ile Met Val Gly Gly Phe Leu Asp Cys Gly Thr Pro Glu ? ? ?
Asn Ile Ile Asp Met Leu Val Asp Leu Asn Ile Lys Asn Leu Thr Ile 35
Ile Ser Asn Asp Thr Ala Phe Pro Asn Lys Gly Ile Gly Lys Leu Ile 50 55 60

US 9 , 957 , 531 B1 95 96
- continued Val Asn Gly Gin Val Ser Lys Val Ile Ala Ser His Ile Gly Thr Asn
80 Igg & ?? |
Pro Glu Thr Gly Lys Lys Met Ser Ser Gly Glu Leu Lys Val Glu Leu 95
Ser Pro Gin Gly Thr Leu Ile Glu Arg Ile Arg Ala Ala Gly Ser Gly 100 105
3 IT ? Leu Gly Gly Val Leu Thr Pro Thr Gly Leu Gly Thr Ile Val Glu Glu 120 mmm 8 mm
mmm
$ gg mmmmm Gly Lys Lys Lys Val Thr Ile Asp Gly Lys Glu Tyr Leu Leu Glu Leu 135 mmm Pro Leu Ser Ala Asp Val Ser Leu Ile Lys Gly Ser Ile Val Asp Glu
Phe Gly Asn Thr Phe Tyr Arg Ala Ala Thr Lys Asn Phe Asn Pro Tyr 175
Met Ala Met Ala Ala Lys Thr Val Ile Val Glu Ala Glu Asn Leu Val 180 185
| Lys Cys Glu Asp Leu Lys Arg Asp Ala Ile Met Thr Pro Gly Val Leu
200
Val Asp Tyr Ile Val Lys Glu Ala Ala 215
< 210 > SEQ ID No 6 < 211 > LENGTH : 221 < 212 > TYPE : PRT < 213 > ORGANISM : Clostridium beijerinckii < 220 > FEATURE : 212 NAME / KEY : MISC FEATURE 23 > OTHER INFORMATION : ctfB , WP 012059997 . 1 AA NN
< 400 > SEQUENCE : 6
Met Ile Val Asp Lys Val Leu Ala Lys Glu Ile Ile Ala Lys Arg Val ? E3 ? Ala Lys Glu Leu Lys Lys Asp Gin Leu Val Asn Leu Gly Ile Gly Leu 20 25
am & Pro Thr Leu Val Ala Asn Tyr Val Pro Lys Glu Met Asn Ile Thr Phe I ? SEE 45
Glu Ser Glu Asn Gly Met Val Gly Met Ala Gin Met Ala Ser Ser Gly
Glu Asn Asp Pro Asp Ile Ile Asn Ala Gly Gly Glu Tyr Val Thr Leu 70 &
Leu Pro Gln Gly Ser Phe Phe Asp Ser Ser Met Ser Phe Ala Leu Ile
Arg Gly Gly His Val Asp Val Ala Val Leu Gly Ala Leu Glu Val Asp 100 105 mmm gmmmmm Glu Lys Gly Asn Leu Ala Asn Trp Ile Val Pro Asn Lys Ile Val Pro
125 g mmmmm mmm Gly Met Gly Gly Ala Met Asp Leu Ala Ile Gly Ala Lys Lys Ile Ile
? Val Ala Met Gin His Thr Gly Lys Ser Lys Pro Lys Ile Val Lys Lys 150
Cys Thr Leu Pro Leu Thr Ala Lys Ala Gln Val Asp Leu Ile Val Thr ?
Glu Leu Cys Val Ile Asp Val Thr Asn Asp Gly Leu Leu Leu Lys Glu 180 185
?
|
Ile His Lys Asp Thr Thr Ile Asp Glu Ile Lys Phe Leu Thr Asp Ala 205

US 9 , 957 , 531 B1 97 98
- continued
Asp Leu Ile Ile Pro Asp Asn Leu Lys Ile Met Asp Ile 210 215 220
< 210 > SEQ ID No 7 < 211 > LENGTH : 286 < 212 > TYPE : PRT < 213 > ORGANISM : Escherichia coli 20 > FEATURE :
< 221 > NAME / KEY : MISC _ FEATURE < 223 > OTHER INFORMATION : TesB , NP _ 414986 . 1
< 400 > SEQUENCE : 7 Met Ser Gln Ala Leu Lys Asn Leu Leu Thr Leu Leu Asn Leu Glu Lys 1 Ann Len or Eye Ile Glu Glu Gly Leu Phe Arg Gly Gin Ser Glu Asp Leu Gly Leu Arg
25 30
Gin Val Phe Gly Gly Gin Val Val Gly Gin Ala Leu Tyr Ala Ala Lys 35 45 40
Glu Thr Val Pro Glu Glu Arq Leu Val His Ser Phe His Ser Tyr Phe 55 50
3 Ep 32 ° Leu Arg Pro Gly Asp Ser Lys Lys Pro Ile Ile Tyr Asp Val Glu Thr
75 65 & J
Leu Arg Asp Gly Asn Ser Phe Ser Ala Arg Arg Val Ala Ala Ile Gin
& Asn Gly Lys Pro Ile Phe Tyr Met Thr Ala Ser Phe Gin Ala Pro Glu 105 110
Ala Gly Phe Glu His Gin Lys Thr Met Pro Ser Ala Pro Ala Pro Asp 115 120 125
3
mmm mmmmmm mmmm mm
Gly Leu Pro Ser Glu Thr Gln Ile Ala Gln Ser Leu Ala His Leu Leu 130 135
Pro Pro Val Leu Lys Asp Lys Phe Ile Cys Asp Arg Pro Leu Glu Val 145 155 mmmm mmmmmm mmmm
a
mmmmmm mmmmmm g? mmm Arg Pro Val Glu Phe His Asn Pro Leu Lys Gly His Val Ala Glu Pro &
His Arg Gin Val Trp Ile Arg Ala Asn Gly Ser Val Pro Asp Asp Leu 185 190
Arg Val His Gin Tyr Leu Leu Gly Tyr Ala Ser Asp Leu Asn Phe Leu 195 200 205
Pro Val Ala Leu Gin Pro His Gly Ile Gly Phe Leu Glu Pro Gly Ile 210 215
Gln Ile Ala Thr Ile Asp His Ser Met Trp Phe His Arq Pro Phe Asn 225 235
Leu Asn Glu Trp Leu Leu Tyr Ser Val Glu Ser Thr Ser Ala Ser Ser
Ala Arg Gly Phe Val Arg Gly Glu Phe Tyr Thr Gin Asp Gly Val Leu 265 270
Val Ala Ser Thr Val Gin Glu Gly Val Met Arg Asn His Asn 275 280 285
1
< 210 > SEQ ID NO 8 < 211 > LENGTH : 436 < 212 > TYPE : PRT < 213 > ORGANISM : Clostridium autoethanogenum < 220 > FEATURE : < 221 > NAME / KEY : MISC _ FEATURE < 223 > OTHER INFORMATION : putative thioesterase 1 , AGY74947 . 1
1
< 400 > SEQUENCE : 8

US 9 , 957 , 531 B1 99 100
- continued
Met Asn Asn Asp Asn Cys Thr Ile Lys Ile Thr Pro Glu Val Ser Arq 10 15
Val Asp Glu Pro Val Asp Ile Lys Ile Asn Gly Leu Pro Lys Asn Glu
E Lys Val Ile Ile Arg Ala Val Ser Ser Asp Tyr Tyr Cys Ile Asn Ala 40
R
Ser Ile Leu Glu Ile Gly Asp Asn Thr Leu Trp Glu Ser Tyr Ala Val
Phe Glu Thr Asp Glu Cys Gly Asn Ile Asn Phe Glu Asn Ala Val Pro 65 75 80 ? 2?
S
E E & 70
Val Asp Gly Thr Tyr Ser Asn Cys Asp Lys Met Gly Leu Phe Tyr Ser 95 90
Met Arg Pro Lys Gin Ile Arg Lys Ser Lys Leu Ile Gin Lys Leu Ser
Ser Ile Asn Glu Asn Arg Lys Tyr Lys Ile Thr Phe Thr Val Glu Lys 120
Asn Gly Lys Ile Ile Gly Ser Lys Glu His Thr Arg Val Tyr Cys Asp
Asp Thr Ile Lys Ser Ile Asp Val Val Glu Lys Asn Leu Leu Ala Arg 145 150 | 155 160
Tyr Phe Thr Ser Lys Asp Asn Ile Lys His Pro Ala Ile Ile Val Leu 170 175
Ser Gly Ser Asp Gly Arg Ile Glu Lys Ala Gin Ala Ile Ala Glu Leu
Phe Ala Met Arg Gly Tyr Ser Ala Leu Ala Val Cys Tyr Phe Gly Leu 200 E e gg &
Glu Gly Thr Pro Glu Asp Leu Asn Met Ile Pro Leu Glu Tyr Val Glu
Asn Ala Val Lys Trp Leu Lys Arg Gin Asp Thr Val Asp Glu Asn Lys 225 230 235 240
Ile Ala Ile Tyr Gly Arg Ser Lys Gly Gly Glu Leu Val Leu Leu Ala 250 255
Ala Ser Met Phe Lys Asp Ile Ala Cys Val Ile Ala Asn Thr Pro Ser
Cys Tyr Val Tyr Glu Gly Ile Lys Ser Asn Lys Leu Pro Ser His His
Ser Ser Trp Met Tyr Arg Gly Arg Glu Ile Pro Tyr Leu Lys Phe Asn
Phe His Ile Ile Leu Arg Leu Ile Ile Lys Met Met Lys Lys Glu Lys 305 310 315 320
Gly Ala Leu Ala Trp Met Tyr Lys Lys Leu Ile Glu Glu Gly Asp Arg 330 335
Asp Lys Ala Thr Ile Ala Leu Asp Lys Ile Asn Gly Ser Val Leu Met
Ile Ser Ser Ala Ala Asp Glu Ile Trp Pro Ser Lys Met His Ser Glu
Thr Val Cys Ser Ile Phe Glu Lys Ser His Phe Lys His Glu Tyr Lys
& His Ile Thr Phe Ala Lys Ser Gly His Ile Leu Thr Val Pro Phe Gln 385 390 395 400
Ser Ile Tyr Pro Ser Glu Lys Tyr Pro Tyr Asp Val Glu Ser Trp Ala 410 415

US 9 , 957 , 531 B1 101 102
- continued Lys Ala Asn Met Asp Ser Trp Asn Glu Thr Ile Lys Phe Leu Glu Lys
420 425 43 0
Trp Ala Ser Lys 435
AAAAA < 210 > SEQ ID NO 9 < 211 > LENGTH : 137 < 212 > TYPE : PRT < 213 > ORGANISM : Clostridium autoethanogenum < 220 > FEATURE :
12 NAME / KEY : MISC _ FEATURE < 223 > OTHER INFORMATION : putative thioesterase 2 , AGY75747 . 1
< 400 > SEQUENCE : 9 ? Met Tyr Ile Asn Glu Thr Lys Val Val Val Arg Tyr Ala Glu Thr Asp
2 sm Lys Met Gly Ile Val His His Ser Asn Tyr Tyr Ile Tyr Phe Glu Glu 25 E ?
g Ala Arg Thr Gin Phe Ile Lys Lys Thr Gly Ile Ser Tyr Ser Gin Met 35 40 45 %
Glu Lys Asp Gly Ile Met Phe Pro Leu Val Glu Ser Asn Cys Arg Tyr 55 6 0
Leu Gin Gly Ala Lys Tyr Glu Asp Glu Leu Leu Ile Lys Thr Trp Ile 65 | 75 mm am am Lys Glu Leu Thr Pro Val Lys Ala Glu Phe Asn Tyr Ser Val Ile Arg
Glu Asn Asp Gin Lys Glu Ile Ala Lys Gly Ser Thr Leu His Ala Phe 105
I Val Asn Asn Asn Phe Lys Ile Ile Asn Leu Lys Lys Asn His Thr Glu
115 120 125
Leu Phe Lys Lys Leu Gin Ser Leu Ile 130 135
< 210 > SEQ ID NO 10 < 211 > LENGTH : 128 < 212 > TYPE : PRT
13 > ORGANISM : Clostridium autoethanogenum < 220 > FEATURE : < 221 > NAME / KEY : MISC _ FEATURE < 223 > OTHER INFORMATION : putative thioesterase 3 , AGY 75999 . 1
< 400 > SEQUENCE : 10
Met Asp Phe Ser Lys Leu Phe Lys Val Gly Ser Thr Tyr Val Ser Glu |
3
& Tyr Ile Val Lys Pro Glu Asp Thr Ala Asn Phe Ile Gly Asn Asn Gly 20 30 3? Val Val Met Leu Ser Thr Pro Ala Met Ile Lys Tyr Met Glu Tyr Thr
35 40 45 ? msm Thr Leu His Ile Val Asp Asn Val Ile Pro Lys Asn Tyr Arg Pro Val
50 55 mm mmm Gly Thr Lys Ile Asp Val Glu His Ile Lys Pro Ile Pro Ala Asn Met
S
8 3
T
Seg a ? ? 3 ?? 6
Lys Val Val Val Lys Val Thr Leu Ile Ser Ile Glu Gly Lys Lys Leu
E Arg Tyr Asn Val Glu Ala Phe Asn Glu Lys Asn cys Lys Val Gly Phe 100 110
Gly Ile Tyr Glu Gln Gln Ile Val Asn Leu Glu Gln Phe Leu Asn Arq 115 120 125
? "

T US 9 , 957 , 531 B1
103 104 - continued
< 210 > SEQ ID No 11 < 211 > LENGTH : 436 < 212 > TYPE : PRT < 213 > ORGANISM : Clostridium ljungdahlii < 220 > FEATURE : < 221 > NAME / KEY : MISC _ FEATURE < 223 > OTHER INFORMATION : putative thioesterase 1 , ADK15695 . 1
< 400 > SEQUENCE : 11 & Met Asn Asn Asp Asn Cys Thr Ile Lys Ile Thr Pro Glu Val Ser Arg
15
Val Asp Glu Pro Val Asp Ile Lys Ile Asn Gly Leu Pro Lys Asn Glu | 30 25
E Lys Val Ile Ile Arg Ala Val Ser Ser Asp Tyr Tyr Cys Ile Asn Ala 40
Ser Ile Leu Glu Ile Gly Asp Asn Thr Leu Trp Glu Ser Tyr Ala Val
Phe Glu Thr Asp Glu Cys Gly Asn Ile Asn Phe Glu Asn Ala Val Pro 65 70 75
Val Asp Gly Thr Tyr Ser Asn cys Asp Lys Met Gly Leu Phe Tyr Ser ? 95
Met Arg Pro Lys Gin Ile Arg Lys Ser Lys Leu Ile Gin Lys Leu Ser 105 110
Ser Ile Asn Glu Asn Arg Lys Tyr Lys Ile Thr Phe Thr Val Glu Lys 120
Asn Gly Lys Ile Ile Gly Ser Lys Glu His Thr Arg Val Tyr Cys Asp
Asp Thr Ile Lys Ser Ile Asp Val Val Glu Lys Asn Leu Leu Ala Arg 145 150 155
Tyr Phe Thr Ser Lys Asp Asn Ile Lys His Pro Ala Ile Ile Val Leu 175
Ser Gly Ser Asp Gly Arg Ile Glu Lys Ala Gin Ala Ile Ala Glu Leu 185 190
sms msm mmm mmmmmmmmm mmmmm mmm mmmmmm Phe Ala Met Arg Gly Tyr Ser Ala Leu Ala Val Cys Tyr Phe ly Leu 200 mmmm smmm Glu Gly Thr Pro Glu Asp Leu Asn Met Ile Pro Leu Glu Tyr Val Glu mmmm
mm
8mm ag Asn Ala Val Lys Trp Leu Lys Arg Gin Asp Thr Val Asp Glu Asn Lys 225 230 235
Ile Ala Ile Tyr Gly Arg Ser Lys Gly Gly Glu Leu Val Leu Leu Ala 255
Ala Ser Met Phe Lys Asp Ile Ala Cys Val Ile Ala Asn Thr Pro Ser 265 270
Cys Tyr Val Tyr Glu Gly Ile Lys Ser Asn Lys Leu Pro Ser His His 280
Ser Ser Trp Met Tyr Arg Gly Arg Glu Ile Pro Tyr Leu Lys Phe Asn
Phe His Ile Ile Leu Arq Leu Ile Ile Lys Met Met Lys Lys Glu Lys 305 310 315
Gly Ala Leu Ala Trp Met Tyr Lys Lys Leu Ile Glu Glu Gly Asp Arg 335
Asp Lys Ala Thr Ile Ala Leu Asp Lys Ile Asn Gly Ser Val Leu Met 345 350
Ile Ser Ser Ala Ala Asp Glu Ile Trp Pro Ser Lys Met His Ser Glu 360 8

US 9 , 957 , 531 B1 105 106
- continued
? ? Thr Val Cys Ser Ile Phe Glu Lys Ser His Phe Lys His Glu Tyr Lys 370 375 380 $ hu Lys ser His phe Lye His Glu Tyr Lys RER ? His Ile Thr Phe Ala Lys Ser Gly His Ile Leu Thr Val Pro Phe Gin
385 400 &
395
° & Ser Ile Tyr Pro Ser Glu Lys Tyr Pro Tyr Asp Val Glu Ser Trp Ala 405 415
Lys Ala Asn Met Asp Ser Trp Asn Glu Thr Ile Lys Phe Leu Glu Lys 420 425 430 ?
Trp Ala Ser Lys 435
1 0
222 1 1 1 2
< 210 > SEQ ID No 12 < 211 > LENGTH : 137 < 212 > TYPE : PRT < 213 > ORGANISM : Clostridium ljungdahliii < 220 > FEATURE : < 221 > NAME / KEY : MISC _ FEATURE < 223 > OTHER INFORMATION : putative thioesterage 2 , ADK16655 . 1
1 3
2 0
2 1
2 3
< 400 > SEQUENCE : 12 Met Tyr Ile Asn Glu Thr Lys Val Val Val Arq Tyr Ala Glu Thr Asp
15 |
Lys Met Gly Ile Val His His Ser Asn Tyr Tyr Ile Tyr Phe Glu Glu 20 25
a E | Ep 8? 30
Ala Arg Thr Gin Phe Ile Lys Lys Thr Gly Ile Ser Tyr Ser Gin Met 40
? 35
Glu Lys Asp Gly Ile Met Phe Pro Leu Val Glu Ser Asn cys Arg Tyr 3
23 3 mmmmm 55 mmm Leu Gin Gly Ala Lys Tyr Glu Asp Glu Leu Leu Ile Lys Thr Trp Ile 70 mms mam Lys Glu Leu Thr Pro Val Lys Ala Glu Phe Asn Tyr Ser Val Ile Arq
95
Glu Asn Asp Gin Lys Glu Ile Ala Lys Gly Ser Thr Leu His Ala Phe 100 105 110
Val Asn Asn Asn Phe Lys Ile Ile Asn Leu Lys Lys Asn His Thr Glu 115 120 His Thr clu
Leu Phe Lys Lys Leu Gin Ser Leu Ile 130 135
9
AAAA < 210 > SEQ ID No 13 < 211 > LENGTH : 128 | < 212 > TYPE : PRT < 213 > ORGANISM : Clostridium ljungdahlii < 220 > FEATURE : < 221 > NAME / KEY : MISC _ FEATURE < 223 > OTHER INFORMATION : putative thioesterase 3 , ADK16959 . 1
< 400 > SEQUENCE : 13
Met Asp Phe Ser Lys Leu Phe Lys Val Gly Ser Thr Tyr Val Ser Glu | 15 | 15
Tyr Ile Val Lys Pro Glu Asp Thr Ala Asn Phe Ile Gly Asn Asn Gly a 25
? Val Val Met Leu Ser Thr Pro Ala Met Ile Lys Tyr Met Glu Tyr Thr 40 45 Pro ada Met Ile ye
?g & 1 Thr Leu His Ile Val Asp Asn Val Ile Pro Lys Asn Tyr Arg Pro Val 6 0 am ?
Gly Thr Lys Ile Asp Val Glu His Ile Lys Pro Ile Pro Ala Asn Met 75 80

US 9 , 957 , 531 B1 107 108 .
- continued
Lys Val Val Val Lys Val Thr Leu Ile Ser Ile Glu Gly Lys Lys Leu 85 90 95 95
Arg Tyr Asn Val Glu Ala Phe Asn Glu Lys Asn cys Lys Val Gly Phe 100 105 110
SE 5 |
Gly Ile Tyr Glu Gln Gln Ile Val Asn Leu Glu Gin Phe Leu Asn Arq 115 120 125 Gin Gin Ile Van Aan Leu Guu Gin Phe Leu Aen Arg
1 1
2
< 210 > SEO ID NO 14 < 211 > LENGTH : 246 < 212 > TYPE : PRT < 213 > ORGANISM : Clostridium beijerinckii < 220 > FEATURE : < 221 > NAME / KEY : MISC _ FEATURE < 223 > OTHER INFORMATION : Adc , WP _ 012059998 . 1
3 0
1 22 2 22 1
< 400 > SEOUENCE : 14
Met Leu Glu Ser Glu Val Ser Lys Gln Ile Thr Thr Pro Leu Ala Ala 15
Pro Ala Phe Pro Arg Gly Pro Tyr Arg Phe His Asn Arg Glu Tyr Leu 20 25 30
E
am & Asn Ile Ile Tyr Arg Thr Asp Leu Asp Ala Leu Arq Lys Ile Val Pro 35 40
Glu Pro Leu Glu Leu Asp Arg Ala Tyr Val Arg Phe Glu Met Met Ala ARE Met Pro Asp Thr Thr ly Leu Gly Ser Tyr Thr Glu Cys Gly Gin Ala 65 70 | 75
?
&
? ? Rg d 3
Es THE & 8
&
Ile Pro Val Lys Tyr Asn Gly Val Lys Gly Asp Tyr Leu His Met Met mmm mmmmm mm 95
Tyr Leu Asp Asn Glu Pro Ala Ile Ala Val Gly Ara Glu Ser Ser Ala 100 105 110 mmmmm Tyr Pro Lys Lys Leu Gly Tyr Pro Lys Leu Phe Val Asp Ser Asp Thr
115 120
Leu Val Gly Thr Leu Lys Tyr Gly Thr Leu Pro Val Ala Thr Ala Thr
Met Gly Tyr Lys His Glu Pro Leu Asp Leu Lys Glu Ala Tyr Ala Gin 145 150 155 ER? Ile Ala Arg Pro Asn Phe Met Leu Lys Ile Ile Gin Gly Tyr Asp Gly
175
Lys Pro Arg Ile Cys Glu Leu Ile Cys Ala Glu Asn Thr Asp Ile Thr 180 185 190 RE Ile His Gly Ala Trp Thr Gly Ser Ala Arg Leu Gin Leu Phe Ser His
195 2000
9
ER & Eng g
Ala Leu Ala Pro Leu Ala Asp Leu Pro Val Leu Glu Ile Val Ser Ala
Ser His Ile Leu Thr Asp Leu Thr Leu Gly Thr Pro Lys Val Val His 225 230 235 240
Asp Tyr Leu Ser Val Lys 245
1
< 210 > SEQ ID NO 15 < 211 > LENGTH : 548 < 212 > TYPE : PRT < 213 > ORGANISM : Lactococcus lactis < 220 > FEATURE : < 221 > NAME / KEY : MISC _ FEATURE < 223 > OTHER INFORMATION : KivD
1
< 400 > SEQUENCE : 15

US 9 , 957 , 531 B1 109 , 110
- continued |
Met Tyr Thr Val Gly Asp Tyr Leu Leu Asp Arg Leu His Glu Leu Gly ? 3
10
Ile Glu Glu Ile Phe Gly Val Pro Gly Asp Tyr Asn Leu Gin Phe Leu 20 25
Asp Gin Ile Ile Ser His Lys Asp Met Lys Trp Val Gly Asn Ala Asn 40 45
Glu Leu Asn Ala Ser Tyr Met Ala Asp Gly Tyr Ala Arg Thr Lys Lys 55 &
? Ala Ala Ala Phe Leu Thr Thr Phe Gly Val Gly Glu Leu Ser Ala Val 80
Asn Gly Leu Ala Gly Ser Tyr Ala Glu Asn Leu Pro Val Val Glu Ile 90 msm Val Gly Ser Pro Thr Ser Lys Val Gin Asn Glu Gly Lys Phe Val His
100 105
His Thr Leu Ala Asp Gly Asp Phe Lys His Phe Met Lys Met His Glu 120 125
Pro Val Thr Ala Ala Arg Thr Leu Leu Thr Ala Glu Asn Ala Thr Val 135
Glu Ile Asp Arg Val Leu Ser Ala Leu Leu Lys Glu Arg Lys Pro Val 160
Tyr Ile Asn Leu Pro Val Asp Val Ala Ala Ala Lys Ala Glu Lys Pro 170 E
5
a me
mmmmm mmmmm mmm mmmmm mm Ser Leu Pro Leu Lys Lys Glu Asn Ser Thr Ser Asn Thr Ser Asp Gin
185 5mm mammmmmm mm mmmmmm 180
Glu Ile Leu Asn Lys Ile Gln Glu Ser Leu Lys Asn Ala Lys Lys Pro 200 205
Ile Val Ile Thr Gly His Glu Ile Ile Ser Phe Gly Leu Glu Lys Thr 215
Val Thr Gin Phe Ile Ser Lys Thr Lys Leu Pro Ile Thr Thr Leu Asn 240
Phe Gly Lys Ser Ser Val Asp Glu Ala Leu Pro Ser Phe Leu Gly Ile 250
3
. Tyr Asn Gly Thr Leu Ser Glu Pro Asn Leu Lys Glu Phe Val Glu Ser 260 265
Ala Asp Phe Ile Leu Met Leu Gly Val Lys Leu Thr Asp Ser Ser Thr 285
Gly Ala Phe Thr His His Leu Asn Glu Asn Lys Met Ile Ser Leu Asn 295
Ile Asp Glu Gly Lys Ile Phe Asn Glu Arg Ile Gin Asn Phe Asp Phe 320
Glu Ser Leu Ile Ser Ser Leu Leu Asp Leu Ser Glu Ile Glu Tyr Lys 330
Gly Lys Tyr Ile Asp Lys Lys Gin Glu Asp Phe Val Pro Ser Asn Ala 340 345
Leu Leu Ser Gin Asp Arg Leu Trp Gin Ala Val Glu Asn Leu Thr Gin 365
Ser Asn Glu Thr Ile Val Ala Glu Gin Gly Thr Ser Phe Phe Gly Ala 375
& Ser Ser Ile Phe Leu Lys Ser Lys Ser His Phe Ile Gly Gin Pro Leu 400
Trp Gly Ser Ile Gly Tyr Thr Phe Pro Ala Ala Leu Gly Ser Gin Ile 410
































































































































































































