Embed Size (px)
Citation preview

Biochemical and Molecular Actions of Nutrients
Meal Feeding Alters Translational Control of Gene Expression in Rat Liver1
Ali K. Reiter, Stephen J. Crozier, Scot R. Kimball, and Leonard S. Jefferson2
Department of Cellular and Molecular Physiology, The Pennsylvania State University College of Medicine,Hershey, PA 17033
ABSTRACT Meal feeding after a period of food deprivation results in a subsequent increase in the protein andRNA content of the liver. To gain insight into the mechanisms involved in the response to food intake, changes inthe association of selected mRNAs with polysomes were examined. On the day of the study, rat livers werecollected at 0, 15, 60, and 180 min after the start of feeding and analyzed for biomarkers of the translational controlof protein synthesis. Protein synthesis was increased within 60 min and was sustained for 180 min. Assembly ofthe active eukaryotic initiation factor (eIF) 4F complex was elevated within 15 min, as indicated by the relativeassociation of eIF4E � eIF4G, but returned to the basal value within 180 min. Phosphorylation of the ribosomalprotein (rp) S6 kinase S6K1 and its substrate rpS6 was increased within 15 min and was sustained for at least 180min. Both eIF4F assembly and activation of S6K1 have been linked to upregulated translation of a subset ofmRNAs. To identify translationally regulated mRNAs, polysomal (i.e., actively translated) and nonpolysomal(nontranslated) fractions were isolated and subjected to microarray analysis. The mRNAs encoding 78 proteins,including 42 proteins involved in protein synthesis, exhibited increased abundance in polysomes in response tofeeding. Overall, the results demonstrate that protein synthesis as well as ribosomal protein mRNA translationundergo rapid and sustained stimulation in the liver after meal feeding and thus contribute to the previouslyobserved increases in protein and RNA content. J. Nutr. 135: 367–375, 2005.
KEY WORDS: ● translation initiation ● eukaryotic initiation factors 4E and 4G ● mTOR● microarray analysis
The protein and RNA content of the liver decline duringfood deprivation, and both macromolecules are replenishedupon commencement of feeding (1,2). In rat liver, the fastingto feeding transition results in a rapid aggregation of ribosomeson mRNA, an effect that is sustained for an extended periodof time (2,3). The observed accumulation of polysomes sug-gests a stimulation of the process involving the initiation ofmRNA translation, which would explain in part the subse-quent increase in protein and RNA content.
Further evidence for the involvement of translation initia-tion in the feeding-induced increase in the protein and RNAcontent of the liver is provided by the reported increase inphosphorylation of eukaryotic initiation factor (eIF)3 4E-bind-ing protein 1 (4E-BP1) and ribosomal protein (rp) S6 kinase 1(S6K1) (4,5), 2 biomarkers of signaling through the mamma-lian target of rapamycin (mTOR), 1 h after food intake (4,6).Increased phosphorylation of 4E-BP1 and S6K1 in rat liver wasalso reported to occur in response to oral administration of
leucine (7). Phosphorylation of 4E-BP1 on multiple residuespromotes its dissociation from the 5�-7-methyl(m7)-GTP-capbinding protein (7,8), eIF4E, allowing eIF4E to associate witheIF4G to form the active eIF4F complex that mediates thebinding of mRNA to the 40S ribosomal subunit (9). Thus, inresponse to food intake, increased 4E-BP1 phosphorylationtypically enhances eIF4F complex assembly and thereby stim-ulates translation initiation (4,5).
Increased phosphorylation and the resulting activation ofS6K1 enhance the phosphorylation of rpS6, which is impli-cated in mediating translational control of mRNAs containinga 5�-terminal oligopyrimidine tract (TOP) sequence (10). ThemRNAs encoding most ribosomal proteins as well as severalelongation factors contain a 5�-TOP sequence, suggesting thatthe 5�-TOP sequence permits translational regulation of pro-teins involved in the translation process itself. Therefore,translation of mRNAs containing a 5�-TOP sequence couldcontribute to the increase in protein and RNA content of theliver in response to food intake (11). An increase in S6K1 andrpS6 phosphorylation in rat liver parallels the increased poly-some association of mRNAs containing a 5�-TOP sequenceafter oral administration of leucine (12,13). The effect of oralleucine administration is prevented by treatment with rapa-mycin, demonstrating a dependence on mTOR-signaling forthe stimulation of 5�-TOP translation in rat liver (13). How-ever, rapamycin does not alter the global rate of proteinsynthesis in the liver. Thus, mTOR signaling is activated inresponse to feeding or leucine administration; however, it is
1 Supported by National Institutes of Health grant DK-13499. A.K.R. wassupported by training grant GM08619 from the National Institutes of Health.S.J.C. was supported by the American Heart Association.
2 To whom correspondence should be addressed.E-mail: [email protected].
3 Abbreviations used: 4E-BP, eukaryotic initiation factor 4E binding protein;eIF, eukaryotic initiation factor; eEF, eukaryotic elongation factor; GAPDH, glyc-eraldehyde-3-phosphate dehydrogenase; Hsp8a, heat shock protein 8a; m7GTP,7-methyl-GTP; mTOR, mammalian target of rapamycin; QRT-PCR, quantitativereal-time PCR; rp, ribosomal protein; S6K1, ribosomal protein S6 kinase 1; TOP,terminal oligopyrimidine tract.
0022-3166/05 $8.00 © 2005 American Society for Nutritional Sciences.Manuscript received 29 June 2004. Initial review completed 9 August 2004. Revision accepted 22 December 2004.
367
by guest on January 22, 2015jn.nutrition.org
Dow
nloaded from

unlikely that activation of the pathway accounts for thestimulation of global rates of protein synthesis (12,13). Onthe other hand, stimulation of mTOR signaling may play animportant role in the feeding-induced increase in RNAcontent of the liver through activation of ribosome biosyn-thesis (13).
The overall objective of the present study was to gaininsight into the mechanism(s) involved in the regulation ofprotein synthesis and the translation of selected mRNAs inthe liver in response to feeding. To maximize the chance ofseeing novel changes, a time course analysis was performed toinvestigate both rapid and delayed changes in protein synthe-sis and mRNA translation.
MATERIALS AND METHODS
Animals. Male Sprague-Dawley rats (age 21 d) weighing 45–60g (Charles River Laboratory) were maintained on a reverse 12-hlight:dark cycle with free access to water. The day after arrival, therats were subjected to meal training as described previously (5).Briefly, rats were allowed access to food (Harlan-Teklad RodentChow) for 3 h/d 2 h after commencement of the dark cycle for 14 d.During the first 3–4 d after the start of the meal-training regimen,daily food consumption gradually increased and body weight wasmaintained or decreased slightly (10–15%). Thereafter, daily foodconsumption stabilized and body weight increased steadily throughthe end of the study. On the day of the experiment, rats wererandomly assigned to 4 treatment groups including a food-deprivedcontrol group and groups that were permitted to feed for 15, 60, or180 min. At 10 min before removal of the liver, a flooding dose (1.0mL/100 g body weight) of L-[2,3,4,5,6-3H]phenylalanine (150mmol/L containing 3.70 GBq/L) was administered via tail vein in-jection. After administration of [3H]phenylalanine, rats were re-turned to their cages and had free access to food and water. Rats werekilled by decapitation, trunk blood was collected for measurement ofamino acid specific radioactivity, and the liver was excised for analysisof protein synthesis and biomarkers of mRNA translation. The ex-perimental protocol for the study was reviewed and approved by theInstitutional Animal Care and Use Committee of The PennsylvaniaState University College of Medicine.
Sample preparation. One portion (�0.5 g) of the liver wasweighed and homogenized in 7 volumes of Buffer A consisting of (inmmol/L) 20 HEPES (pH 7.4), 100 KCl, 0.2 EDTA, 2 EGTA, 50 NaF,50 �-glycerophosphate, 0.1 phenylmethylsulfonyl fluoride, 1 benza-midine, and 0.5 sodium vanadate using a Polytron homogenizer. Analiquot of the homogenate was used for measurement of proteinsynthesis, as described below. The remaining homogenate was cen-trifuged at 10,000 � g for 10 min at 4°C. The resulting supernatantwas used for analysis of the phosphorylation status of S6K1, rpS6,eIF4G, and 4E-BP1 as well as the association of eIF4G, 4E-BP1, and4E-BP2 with eIF4E as described below. A second portion (�1 g) ofthe liver was weighed and homogenized in 3 volumes of Buffer Bconsisting of (in mmol/L) 40 HEPES (pH 7.5), 100 KCl, and 5magnesium chloride. The homogenate was centrifuged at 3000 � gfor 15 min, and the resulting supernatant was used for analysis ofpolysome aggregation as described below. A third portion of liver(�0.3 g) was homogenized in 5 volumes of Buffer C consisting of (inmmol/L) 45 HEPES (pH 7.4), 0.375 magnesium acetate, 0.075EDTA, 95 potassium acetate, 2.03 digitonin, 10% (v:v) glycerol, and0.003 microcystin. The homogenate was centrifuged at 10,000 � g for10 min at 4°C, and the resulting supernatant was analyzed for eIF2Bactivity as described below.
Measurement of protein synthesis. The rate of synthesis of totalmixed liver protein was measured as previously described (14).Briefly, the fractional rate of protein synthesis was estimated from therate of incorporation of radioactive phenylalanine into total protein.The fractional rate of protein synthesis is the rate of incorporation of[3H]phenylalanine into protein using the serum phenylalanine spe-cific radioactivity as representative of the precursor pool during theincorporation period, i.e., the time elapsed from injection of [3H]phe-nylalanine until tissue homogenization (5).
Polysome profiles. One volume of detergent [10% (v:v) TritonX-100, 0.24 mol/L deoxycholate sodium salt] was added to 9 volumesof the supernatant of livers homogenized in Buffer B; an aliquot of thesample was layered over a 10–70% linear sucrose density gradient.The gradients were centrifuged at 90,000 � g for 3 h at 4°C in aBeckman SW28 rotor. After centrifugation, the gradients were frac-tionated on an Isco gradient fractionator while the UV absorption at254 nm was recorded continuously. Five fractions (5.0 mL each) anda final 2.5-mL fraction were collected for extraction of total RNA asdescribed below.
Measurement of protein phosphorylation status. Phosphoryla-tion of eIF4G was assessed by Western blot analysis using an anti-phospho-eIF4G (Ser 1108) antibody (Cell Signaling Technology)and normalized to total eIF4G using a polyclonal anti-eIF4G anti-body. Phosphorylation of 4E-BP1 was assessed as a change in elec-trophoretic mobility during SDS-PAGE. The association of eIF4G,4E-BP1, or 4E-BP2 with eIF4E was evaluated as described previously(15). eIF4E phosphorylation was evaluated using a polyclonal anti-body to eIF4E (Ser 209) (Cell Signaling Technology) and normalizedto total eIF4E. Hyperphosphorylation of S6K1 was assessed as de-creased migration during SDS-PAGE using a polyclonal anti-S6K1antibody; S6K1 phosphorylation was also analyzed using a polyclonalantibody that recognizes only the Thr 389 phosphorylated form ofS6K1 (Cell Signaling Technology). Finally, rpS6 phosphorylationwas evaluated using a 1:1 mixture of polyclonal antibodies thatrecognize either the Ser 235/236 or Ser 240/244 phosphorylated formsof rpS6 (Cell Signaling Technology).
Polysomal RNA extraction, purification, and microarray analy-sis. Total RNA was isolated from sucrose density gradient fractionsby performing 2 phenol:chloroform (5:1) (Ambion) extractions. The2nd phenol:chloroform extraction was followed by an RNA precip-itation step using 0.1 volume 5 mol/L ammonium acetate (Ambion)and 2 volumes 100% ethanol. The precipitate was washed with 100%ethanol and dissolved in RNA Storage Solution (Ambion) contain-ing Anti-RNase (Ambion) to prevent degradation. RNA from eachcondition was then pooled into 1 of 2 fractions termed subpolysomal,consisting of nonribosome-associated mRNA, 40S and 60S ribosomalsubunits, and mRNA associated with a single 80S ribosome, orpolysomal, consisting of mRNA associated with multiple ribosomes.Subpolysomal RNA from each condition was pooled with subpoly-somal RNA from the same treatment group; polysomal RNA fromeach condition was pooled with polysomal RNA from the sametreatment group. The RNA was further purified using the RNeasy Kit(Qiagen) and eluted in RNase free water (Ambion). DNase I treat-ment was then performed on the samples using the DNase Free Kit(Ambion). The quality, purity, and concentration of the RNA wasassessed using a LabChip analyzed on an Agilent 2100 Bioanalyzer.Microarray analysis was performed in the Juvenile Diabetes ResearchFoundation Functional Genomics Core Facility at The PennsylvaniaState University College of Medicine. Briefly, total RNA (50 �g)from each treatment group was indirectly labeled with CY5; totalRNA (50 �g) from a rat reference library (Stratagene) was indirectlylabeled with CY3. Then, the labeled cDNA was hybridized to a RatLiver Array (MWG Biotech). The microarrays were analyzed on aPerkin Elmer ScanArray 4000XL, and the data were analyzed usingGeneSpring version 6.0.
Total RNA extraction from liver. Total RNA was extractedfrom the liver using an RNeasy Kit (Qiagen). The quality andquantity of the RNA was assessed by measuring the A260 and A280with a Beckman Coulter spectrophotometer.
Quantitative real-time PCR. Quantitative real-time PCR (QRT-PCR) was performed as previously described (16). Briefly, 1 �g totalRNA was incubated with 1 �L Oligo (dT) (Invitrogen) for 10 min at70°C, and the sample was then placed on ice. Next, 5 �L 2X reactionbuffer, 2 �L 0.1 mmol/L dithiothreitol (Invitrogen), and 1 �L Super-script Platinum Taq Polymerase were added to the reaction mixture.The samples were then incubated at 42°C for 65 min and stored at�20°C until QRT-PCR was performed. Serial dilutions were per-formed on the samples to a final dilution of 1:16. QRT-PCR wasperformed using QuantiTect SYBR Green PCR following the man-ufacturer’s instructions. The primers used for each mRNA analyzed wereas follows: albumin-upper-5�-TGGGCAGTAGCTCGTATGA-3� and
REITER ET AL.368
by guest on January 22, 2015jn.nutrition.org
Dow
nloaded from
lower-5�-CAACAGGTCGCCGTGACA-3�; eEF1A-upper-5�-CTA-ATATGCCGTGGTTCAAG-3� and lower-5�-CGCAGAGGC-TTGTCAGTTG-3�; glyceraldehyde-3-phosphate dehydrogenase(GAPDH)-upper-5�-GGGCTGCCTTCTCTTGTGA-3� and lower-5�-TGAACTTGCCGTGGGTAGA-3�; Hsp8a-upper-5�-TGTCCT-CATCAAGCGCAATA-3� and lower-5�-GGCCCTTTCACCT-TCATAC-3�; rpS6-upper-5�-ACTGGCTGTCAGAAACTCAT-3�and lower-5�-CCACATAACCCTTCCACTCT-3�; rpS8 upper-5�-CGTGCTCTGAGATTGGATGT-3� and lower- 5�-CGGAC-AAGCTCGTTATTGG-3�; rpL26 upper-5�-TCTCACATTCGGAG-GAAGAT-3� and lower-5�-TGTCCTCGAACAACCTGAAC-3�.
Statistical analysis. All data were analyzed by InStat 3.0 (Graph-Pad Software) using an ANOVA multiple comparisons test andfurther analyzed using Dunnet’s Multiple Comparison Test when theP-value for the ANOVA was �0.05. The level of significance was setat P � 0.05 for all statistical tests. All data are presented as means� SEM.
RESULTS
Fractional rate of protein synthesis in response to feeding.The rate of synthesis of total mixed liver protein, as analyzedusing the flooding dose technique (14), did not differ from thefood-deprived control value 15 min after commencement offeeding (Table 1). However, by 60 and 180 min, the syntheticrate increased to 118 and 128% of the food-deprived controlvalue, respectively.
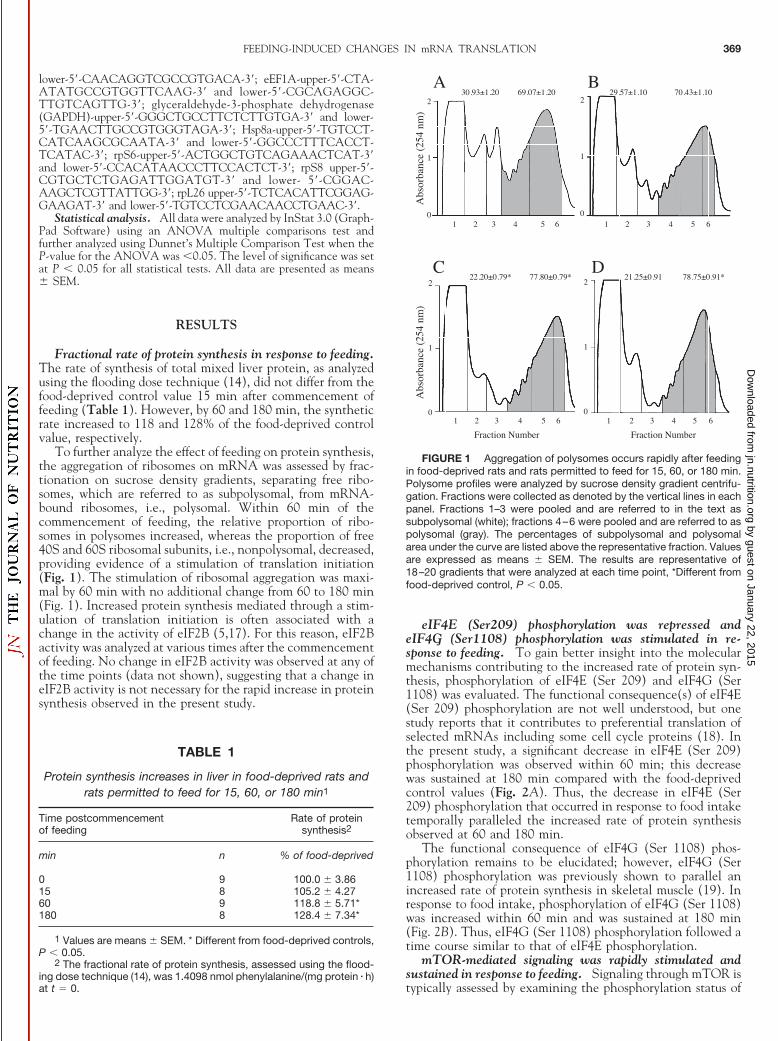
To further analyze the effect of feeding on protein synthesis,the aggregation of ribosomes on mRNA was assessed by frac-tionation on sucrose density gradients, separating free ribo-somes, which are referred to as subpolysomal, from mRNA-bound ribosomes, i.e., polysomal. Within 60 min of thecommencement of feeding, the relative proportion of ribo-somes in polysomes increased, whereas the proportion of free40S and 60S ribosomal subunits, i.e., nonpolysomal, decreased,providing evidence of a stimulation of translation initiation(Fig. 1). The stimulation of ribosomal aggregation was maxi-mal by 60 min with no additional change from 60 to 180 min(Fig. 1). Increased protein synthesis mediated through a stim-ulation of translation initiation is often associated with achange in the activity of eIF2B (5,17). For this reason, eIF2Bactivity was analyzed at various times after the commencementof feeding. No change in eIF2B activity was observed at any ofthe time points (data not shown), suggesting that a change ineIF2B activity is not necessary for the rapid increase in proteinsynthesis observed in the present study.
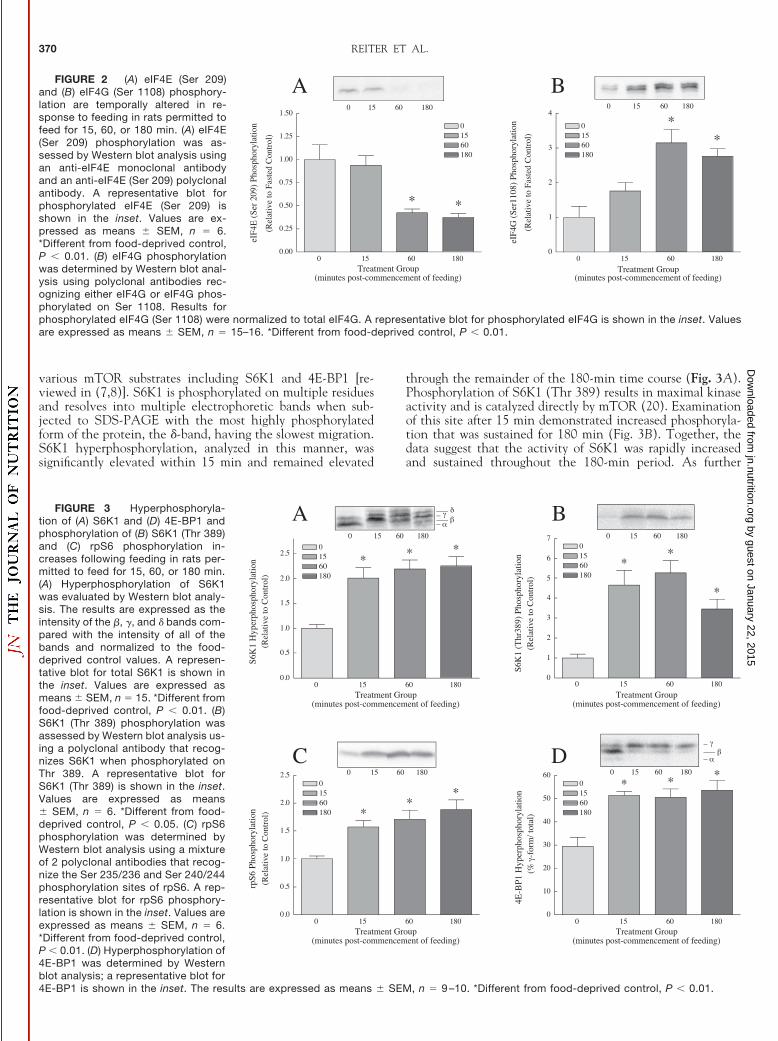
eIF4E (Ser209) phosphorylation was repressed andeIF4G (Ser1108) phosphorylation was stimulated in re-sponse to feeding. To gain better insight into the molecularmechanisms contributing to the increased rate of protein syn-thesis, phosphorylation of eIF4E (Ser 209) and eIF4G (Ser1108) was evaluated. The functional consequence(s) of eIF4E(Ser 209) phosphorylation are not well understood, but onestudy reports that it contributes to preferential translation ofselected mRNAs including some cell cycle proteins (18). Inthe present study, a significant decrease in eIF4E (Ser 209)phosphorylation was observed within 60 min; this decreasewas sustained at 180 min compared with the food-deprivedcontrol values (Fig. 2A). Thus, the decrease in eIF4E (Ser209) phosphorylation that occurred in response to food intaketemporally paralleled the increased rate of protein synthesisobserved at 60 and 180 min.
The functional consequence of eIF4G (Ser 1108) phos-phorylation remains to be elucidated; however, eIF4G (Ser1108) phosphorylation was previously shown to parallel anincreased rate of protein synthesis in skeletal muscle (19). Inresponse to food intake, phosphorylation of eIF4G (Ser 1108)was increased within 60 min and was sustained at 180 min(Fig. 2B). Thus, eIF4G (Ser 1108) phosphorylation followed atime course similar to that of eIF4E phosphorylation.
mTOR-mediated signaling was rapidly stimulated andsustained in response to feeding. Signaling through mTOR istypically assessed by examining the phosphorylation status of
TABLE 1
Protein synthesis increases in liver in food-deprived rats andrats permitted to feed for 15, 60, or 180 min1
Time postcommencementof feeding
Rate of proteinsynthesis2
min n % of food-deprived
0 9 100.0 � 3.8615 8 105.2 � 4.2760 9 118.8 � 5.71*180 8 128.4 � 7.34*
1 Values are means � SEM. * Different from food-deprived controls,P � 0.05.
2 The fractional rate of protein synthesis, assessed using the flood-ing dose technique (14), was 1.4098 nmol phenylalanine/(mg protein � h)at t � 0.
FIGURE 1 Aggregation of polysomes occurs rapidly after feedingin food-deprived rats and rats permitted to feed for 15, 60, or 180 min.Polysome profiles were analyzed by sucrose density gradient centrifu-gation. Fractions were collected as denoted by the vertical lines in eachpanel. Fractions 1–3 were pooled and are referred to in the text assubpolysomal (white); fractions 4–6 were pooled and are referred to aspolysomal (gray). The percentages of subpolysomal and polysomalarea under the curve are listed above the representative fraction. Valuesare expressed as means � SEM. The results are representative of18–20 gradients that were analyzed at each time point, *Different fromfood-deprived control, P � 0.05.
FEEDING-INDUCED CHANGES IN mRNA TRANSLATION 369
by guest on January 22, 2015jn.nutrition.org
Dow
nloaded from
various mTOR substrates including S6K1 and 4E-BP1 [re-viewed in (7,8)]. S6K1 is phosphorylated on multiple residuesand resolves into multiple electrophoretic bands when sub-jected to SDS-PAGE with the most highly phosphorylatedform of the protein, the �-band, having the slowest migration.S6K1 hyperphosphorylation, analyzed in this manner, wassignificantly elevated within 15 min and remained elevated
through the remainder of the 180-min time course (Fig. 3A).Phosphorylation of S6K1 (Thr 389) results in maximal kinaseactivity and is catalyzed directly by mTOR (20). Examinationof this site after 15 min demonstrated increased phosphoryla-tion that was sustained for 180 min (Fig. 3B). Together, thedata suggest that the activity of S6K1 was rapidly increasedand sustained throughout the 180-min period. As further
FIGURE 2 (A) eIF4E (Ser 209)and (B) eIF4G (Ser 1108) phosphory-lation are temporally altered in re-sponse to feeding in rats permitted tofeed for 15, 60, or 180 min. (A) eIF4E(Ser 209) phosphorylation was as-sessed by Western blot analysis usingan anti-eIF4E monoclonal antibodyand an anti-eIF4E (Ser 209) polyclonalantibody. A representative blot forphosphorylated eIF4E (Ser 209) isshown in the inset. Values are ex-pressed as means � SEM, n � 6.*Different from food-deprived control,P � 0.01. (B) eIF4G phosphorylationwas determined by Western blot anal-ysis using polyclonal antibodies rec-ognizing either eIF4G or eIF4G phos-phorylated on Ser 1108. Results forphosphorylated eIF4G (Ser 1108) were normalized to total eIF4G. A representative blot for phosphorylated eIF4G is shown in the inset. Valuesare expressed as means � SEM, n � 15–16. *Different from food-deprived control, P � 0.01.
FIGURE 3 Hyperphosphoryla-tion of (A) S6K1 and (D) 4E-BP1 andphosphorylation of (B) S6K1 (Thr 389)and (C) rpS6 phosphorylation in-creases following feeding in rats per-mitted to feed for 15, 60, or 180 min.(A) Hyperphosphorylation of S6K1was evaluated by Western blot analy-sis. The results are expressed as theintensity of the �, �, and � bands com-pared with the intensity of all of thebands and normalized to the food-deprived control values. A represen-tative blot for total S6K1 is shown inthe inset. Values are expressed asmeans � SEM, n � 15. *Different fromfood-deprived control, P � 0.01. (B)S6K1 (Thr 389) phosphorylation wasassessed by Western blot analysis us-ing a polyclonal antibody that recog-nizes S6K1 when phosphorylated onThr 389. A representative blot forS6K1 (Thr 389) is shown in the inset.Values are expressed as means� SEM, n � 6. *Different from food-deprived control, P � 0.05. (C) rpS6phosphorylation was determined byWestern blot analysis using a mixtureof 2 polyclonal antibodies that recog-nize the Ser 235/236 and Ser 240/244phosphorylation sites of rpS6. A rep-resentative blot for rpS6 phosphory-lation is shown in the inset. Values areexpressed as means � SEM, n � 6.*Different from food-deprived control,P � 0.01. (D) Hyperphosphorylation of4E-BP1 was determined by Westernblot analysis; a representative blot for4E-BP1 is shown in the inset. The results are expressed as means � SEM, n � 9 –10. *Different from food-deprived control, P � 0.01.
REITER ET AL.370
by guest on January 22, 2015jn.nutrition.org
Dow
nloaded from
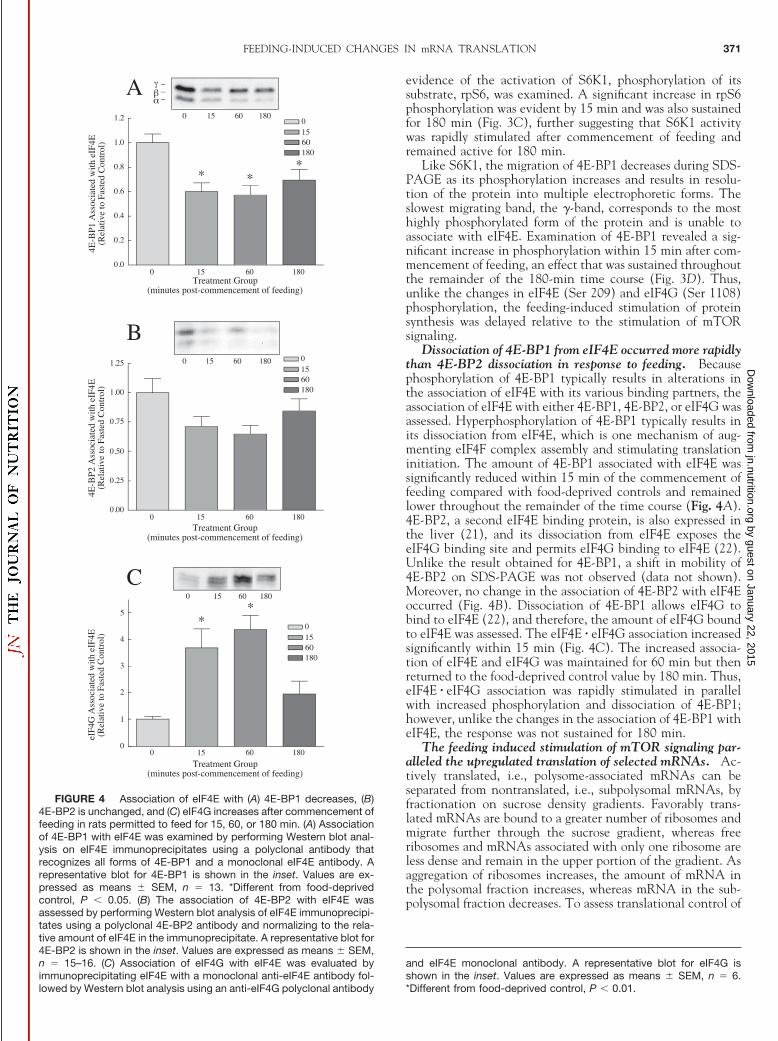
evidence of the activation of S6K1, phosphorylation of itssubstrate, rpS6, was examined. A significant increase in rpS6phosphorylation was evident by 15 min and was also sustainedfor 180 min (Fig. 3C), further suggesting that S6K1 activitywas rapidly stimulated after commencement of feeding andremained active for 180 min.
Like S6K1, the migration of 4E-BP1 decreases during SDS-PAGE as its phosphorylation increases and results in resolu-tion of the protein into multiple electrophoretic forms. Theslowest migrating band, the �-band, corresponds to the mosthighly phosphorylated form of the protein and is unable toassociate with eIF4E. Examination of 4E-BP1 revealed a sig-nificant increase in phosphorylation within 15 min after com-mencement of feeding, an effect that was sustained throughoutthe remainder of the 180-min time course (Fig. 3D). Thus,unlike the changes in eIF4E (Ser 209) and eIF4G (Ser 1108)phosphorylation, the feeding-induced stimulation of proteinsynthesis was delayed relative to the stimulation of mTORsignaling.
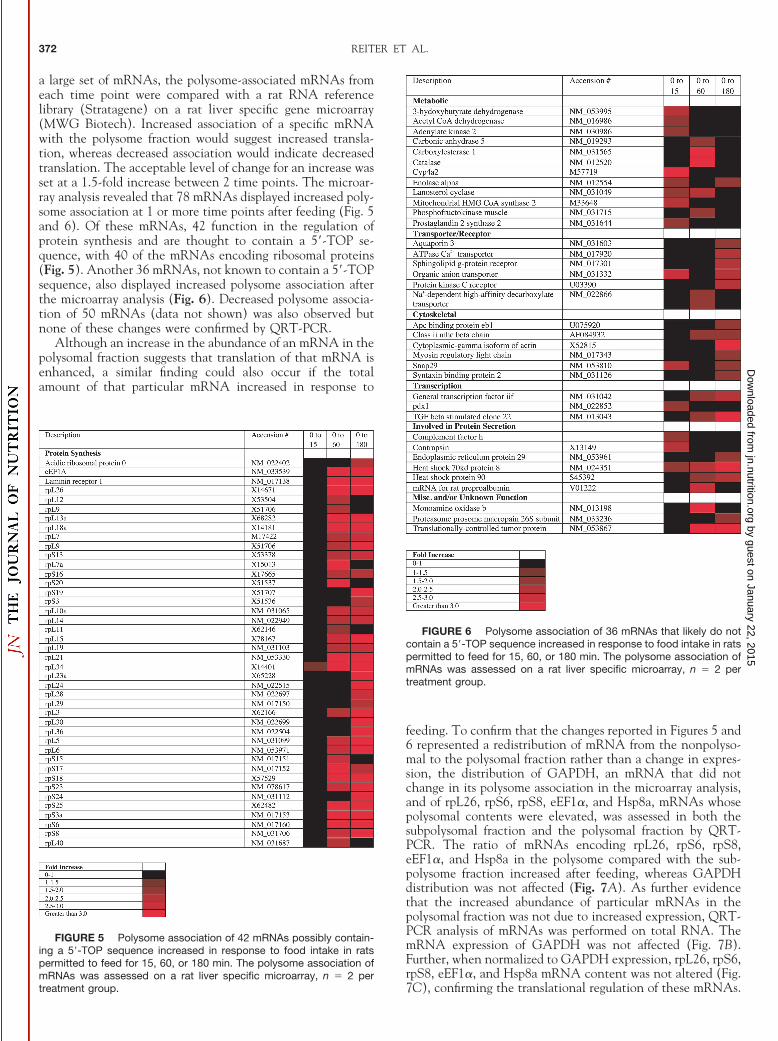
Dissociation of 4E-BP1 from eIF4E occurred more rapidlythan 4E-BP2 dissociation in response to feeding. Becausephosphorylation of 4E-BP1 typically results in alterations inthe association of eIF4E with its various binding partners, theassociation of eIF4E with either 4E-BP1, 4E-BP2, or eIF4G wasassessed. Hyperphosphorylation of 4E-BP1 typically results inits dissociation from eIF4E, which is one mechanism of aug-menting eIF4F complex assembly and stimulating translationinitiation. The amount of 4E-BP1 associated with eIF4E wassignificantly reduced within 15 min of the commencement offeeding compared with food-deprived controls and remainedlower throughout the remainder of the time course (Fig. 4A).4E-BP2, a second eIF4E binding protein, is also expressed inthe liver (21), and its dissociation from eIF4E exposes theeIF4G binding site and permits eIF4G binding to eIF4E (22).Unlike the result obtained for 4E-BP1, a shift in mobility of4E-BP2 on SDS-PAGE was not observed (data not shown).Moreover, no change in the association of 4E-BP2 with eIF4Eoccurred (Fig. 4B). Dissociation of 4E-BP1 allows eIF4G tobind to eIF4E (22), and therefore, the amount of eIF4G boundto eIF4E was assessed. The eIF4E � eIF4G association increasedsignificantly within 15 min (Fig. 4C). The increased associa-tion of eIF4E and eIF4G was maintained for 60 min but thenreturned to the food-deprived control value by 180 min. Thus,eIF4E � eIF4G association was rapidly stimulated in parallelwith increased phosphorylation and dissociation of 4E-BP1;however, unlike the changes in the association of 4E-BP1 witheIF4E, the response was not sustained for 180 min.
The feeding induced stimulation of mTOR signaling par-alleled the upregulated translation of selected mRNAs. Ac-tively translated, i.e., polysome-associated mRNAs can beseparated from nontranslated, i.e., subpolysomal mRNAs, byfractionation on sucrose density gradients. Favorably trans-lated mRNAs are bound to a greater number of ribosomes andmigrate further through the sucrose gradient, whereas freeribosomes and mRNAs associated with only one ribosome areless dense and remain in the upper portion of the gradient. Asaggregation of ribosomes increases, the amount of mRNA inthe polysomal fraction increases, whereas mRNA in the sub-polysomal fraction decreases. To assess translational control of
FIGURE 4 Association of eIF4E with (A) 4E-BP1 decreases, (B)4E-BP2 is unchanged, and (C) eIF4G increases after commencement offeeding in rats permitted to feed for 15, 60, or 180 min. (A) Associationof 4E-BP1 with eIF4E was examined by performing Western blot anal-ysis on eIF4E immunoprecipitates using a polyclonal antibody thatrecognizes all forms of 4E-BP1 and a monoclonal eIF4E antibody. Arepresentative blot for 4E-BP1 is shown in the inset. Values are ex-pressed as means � SEM, n � 13. *Different from food-deprivedcontrol, P � 0.05. (B) The association of 4E-BP2 with eIF4E wasassessed by performing Western blot analysis of eIF4E immunoprecipi-tates using a polyclonal 4E-BP2 antibody and normalizing to the rela-tive amount of eIF4E in the immunoprecipitate. A representative blot for4E-BP2 is shown in the inset. Values are expressed as means � SEM,n � 15–16. (C) Association of eIF4G with eIF4E was evaluated byimmunoprecipitating eIF4E with a monoclonal anti-eIF4E antibody fol-lowed by Western blot analysis using an anti-eIF4G polyclonal antibody
and eIF4E monoclonal antibody. A representative blot for eIF4G isshown in the inset. Values are expressed as means � SEM, n � 6.*Different from food-deprived control, P � 0.01.
FEEDING-INDUCED CHANGES IN mRNA TRANSLATION 371
by guest on January 22, 2015jn.nutrition.org
Dow
nloaded from
a large set of mRNAs, the polysome-associated mRNAs fromeach time point were compared with a rat RNA referencelibrary (Stratagene) on a rat liver specific gene microarray(MWG Biotech). Increased association of a specific mRNAwith the polysome fraction would suggest increased transla-tion, whereas decreased association would indicate decreasedtranslation. The acceptable level of change for an increase wasset at a 1.5-fold increase between 2 time points. The microar-ray analysis revealed that 78 mRNAs displayed increased poly-some association at 1 or more time points after feeding (Fig. 5and 6). Of these mRNAs, 42 function in the regulation ofprotein synthesis and are thought to contain a 5�-TOP se-quence, with 40 of the mRNAs encoding ribosomal proteins(Fig. 5). Another 36 mRNAs, not known to contain a 5�-TOPsequence, also displayed increased polysome association afterthe microarray analysis (Fig. 6). Decreased polysome associa-tion of 50 mRNAs (data not shown) was also observed butnone of these changes were confirmed by QRT-PCR.
Although an increase in the abundance of an mRNA in thepolysomal fraction suggests that translation of that mRNA isenhanced, a similar finding could also occur if the totalamount of that particular mRNA increased in response to
feeding. To confirm that the changes reported in Figures 5 and6 represented a redistribution of mRNA from the nonpolyso-mal to the polysomal fraction rather than a change in expres-sion, the distribution of GAPDH, an mRNA that did notchange in its polysome association in the microarray analysis,and of rpL26, rpS6, rpS8, eEF1�, and Hsp8a, mRNAs whosepolysomal contents were elevated, was assessed in both thesubpolysomal fraction and the polysomal fraction by QRT-PCR. The ratio of mRNAs encoding rpL26, rpS6, rpS8,eEF1�, and Hsp8a in the polysome compared with the sub-polysome fraction increased after feeding, whereas GAPDHdistribution was not affected (Fig. 7A). As further evidencethat the increased abundance of particular mRNAs in thepolysomal fraction was not due to increased expression, QRT-PCR analysis of mRNAs was performed on total RNA. ThemRNA expression of GAPDH was not affected (Fig. 7B).Further, when normalized to GAPDH expression, rpL26, rpS6,rpS8, eEF1�, and Hsp8a mRNA content was not altered (Fig.7C), confirming the translational regulation of these mRNAs.
FIGURE 5 Polysome association of 42 mRNAs possibly contain-ing a 5�-TOP sequence increased in response to food intake in ratspermitted to feed for 15, 60, or 180 min. The polysome association ofmRNAs was assessed on a rat liver specific microarray, n � 2 pertreatment group.
FIGURE 6 Polysome association of 36 mRNAs that likely do notcontain a 5�-TOP sequence increased in response to food intake in ratspermitted to feed for 15, 60, or 180 min. The polysome association ofmRNAs was assessed on a rat liver specific microarray, n � 2 pertreatment group.
REITER ET AL.372
by guest on January 22, 2015jn.nutrition.org
Dow
nloaded from
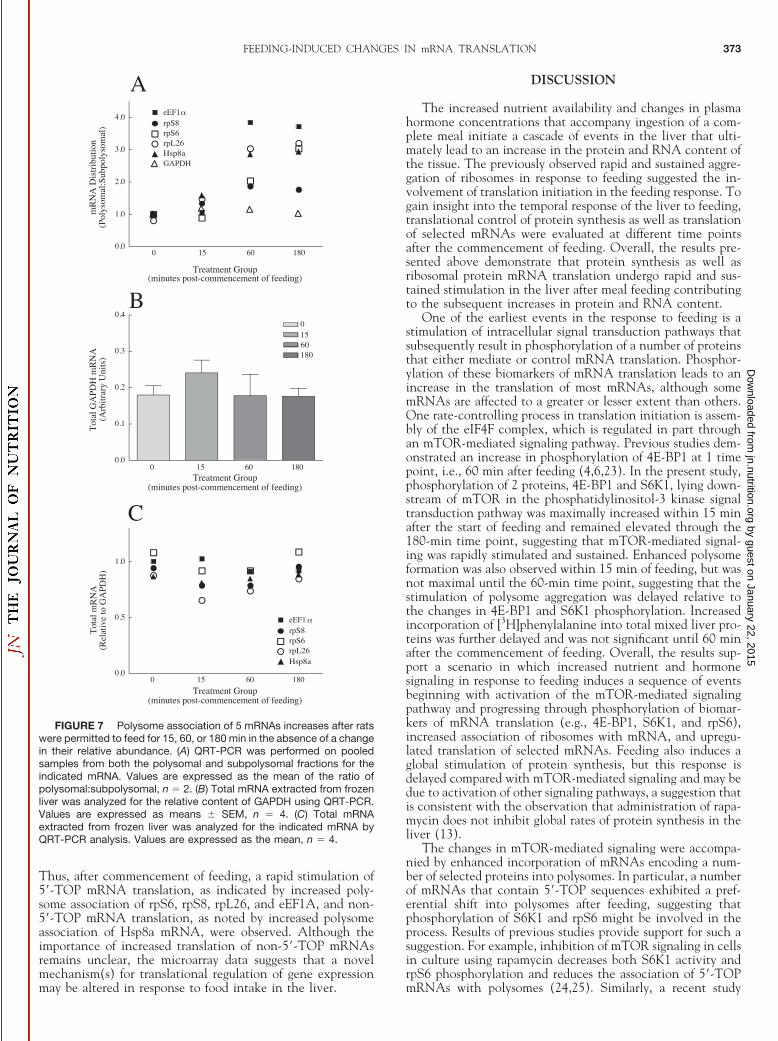
Thus, after commencement of feeding, a rapid stimulation of5�-TOP mRNA translation, as indicated by increased poly-some association of rpS6, rpS8, rpL26, and eEF1A, and non-5�-TOP mRNA translation, as noted by increased polysomeassociation of Hsp8a mRNA, were observed. Although theimportance of increased translation of non-5�-TOP mRNAsremains unclear, the microarray data suggests that a novelmechanism(s) for translational regulation of gene expressionmay be altered in response to food intake in the liver.
DISCUSSION
The increased nutrient availability and changes in plasmahormone concentrations that accompany ingestion of a com-plete meal initiate a cascade of events in the liver that ulti-mately lead to an increase in the protein and RNA content ofthe tissue. The previously observed rapid and sustained aggre-gation of ribosomes in response to feeding suggested the in-volvement of translation initiation in the feeding response. Togain insight into the temporal response of the liver to feeding,translational control of protein synthesis as well as translationof selected mRNAs were evaluated at different time pointsafter the commencement of feeding. Overall, the results pre-sented above demonstrate that protein synthesis as well asribosomal protein mRNA translation undergo rapid and sus-tained stimulation in the liver after meal feeding contributingto the subsequent increases in protein and RNA content.
One of the earliest events in the response to feeding is astimulation of intracellular signal transduction pathways thatsubsequently result in phosphorylation of a number of proteinsthat either mediate or control mRNA translation. Phosphor-ylation of these biomarkers of mRNA translation leads to anincrease in the translation of most mRNAs, although somemRNAs are affected to a greater or lesser extent than others.One rate-controlling process in translation initiation is assem-bly of the eIF4F complex, which is regulated in part throughan mTOR-mediated signaling pathway. Previous studies dem-onstrated an increase in phosphorylation of 4E-BP1 at 1 timepoint, i.e., 60 min after feeding (4,6,23). In the present study,phosphorylation of 2 proteins, 4E-BP1 and S6K1, lying down-stream of mTOR in the phosphatidylinositol-3 kinase signaltransduction pathway was maximally increased within 15 minafter the start of feeding and remained elevated through the180-min time point, suggesting that mTOR-mediated signal-ing was rapidly stimulated and sustained. Enhanced polysomeformation was also observed within 15 min of feeding, but wasnot maximal until the 60-min time point, suggesting that thestimulation of polysome aggregation was delayed relative tothe changes in 4E-BP1 and S6K1 phosphorylation. Increasedincorporation of [3H]phenylalanine into total mixed liver pro-teins was further delayed and was not significant until 60 minafter the commencement of feeding. Overall, the results sup-port a scenario in which increased nutrient and hormonesignaling in response to feeding induces a sequence of eventsbeginning with activation of the mTOR-mediated signalingpathway and progressing through phosphorylation of biomar-kers of mRNA translation (e.g., 4E-BP1, S6K1, and rpS6),increased association of ribosomes with mRNA, and upregu-lated translation of selected mRNAs. Feeding also induces aglobal stimulation of protein synthesis, but this response isdelayed compared with mTOR-mediated signaling and may bedue to activation of other signaling pathways, a suggestion thatis consistent with the observation that administration of rapa-mycin does not inhibit global rates of protein synthesis in theliver (13).
The changes in mTOR-mediated signaling were accompa-nied by enhanced incorporation of mRNAs encoding a num-ber of selected proteins into polysomes. In particular, a numberof mRNAs that contain 5�-TOP sequences exhibited a pref-erential shift into polysomes after feeding, suggesting thatphosphorylation of S6K1 and rpS6 might be involved in theprocess. Results of previous studies provide support for such asuggestion. For example, inhibition of mTOR signaling in cellsin culture using rapamycin decreases both S6K1 activity andrpS6 phosphorylation and reduces the association of 5�-TOPmRNAs with polysomes (24,25). Similarly, a recent study
FIGURE 7 Polysome association of 5 mRNAs increases after ratswere permitted to feed for 15, 60, or 180 min in the absence of a changein their relative abundance. (A) QRT-PCR was performed on pooledsamples from both the polysomal and subpolysomal fractions for theindicated mRNA. Values are expressed as the mean of the ratio ofpolysomal:subpolysomal, n � 2. (B) Total mRNA extracted from frozenliver was analyzed for the relative content of GAPDH using QRT-PCR.Values are expressed as means � SEM, n � 4. (C) Total mRNAextracted from frozen liver was analyzed for the indicated mRNA byQRT-PCR analysis. Values are expressed as the mean, n � 4.
FEEDING-INDUCED CHANGES IN mRNA TRANSLATION 373
by guest on January 22, 2015jn.nutrition.org
Dow
nloaded from

reported that rapamycin prevents the leucine-induced redis-tribution of ribosomal protein mRNAs into polysomes in ratliver (13), although it does not prevent its shift into mono-somes (i.e., mRNA associated with a single ribosome). Thelatter result agrees with reports suggesting that S6K1 activa-tion and rpS6 phosphorylation may not be sufficient for en-hanced translation of 5�-TOP mRNAs (26,27). Despite theuncertainty of the dependence of 5�-TOP mRNA translationon S6K1 activation or rpS6 phosphorylation, it is clear thatfeeding rapidly promotes an increase in the polysome associ-ation of ribosomal protein mRNAs in the liver. Moreover, theenhanced association of ribosomal protein mRNAs withpolysomes observed in the present study, in combinationwith previous studies reporting an mTOR-mediated in-crease in rDNA transcription (28), provides a likely expla-nation for the subsequent increase in ribosome content inresponse to feeding (1,2).
Additionally, the results of the microarray analysis providedinsight into the translational upregulation of 78 differentmRNAs (Figs. 5 and 6) and downregulation of 50 mRNAs(data not shown). Although 42 of the 78 mRNAs did containor were likely to contain such a sequence, 36 mRNAs wereidentified that are not known to contain a 5�-TOP sequence.Of the non-5�-TOP mRNAs, Hsp8a was confirmed to shiftfrom the subpolysomal fraction to the polysomal fraction byQRT-PCR. Thus, it is possible that some of the 36 non-5�-TOP mRNAs may be regulated by a novel mechanism(s).Interestingly, albumin mRNA incorporation into polysomeswas observed to increase by microarray analysis. Because therat albumin mRNA lacks a 5�-TOP sequence (accession#M16825), the results suggest that its translation as well asother mRNAs may be regulated through unique mechanismsthat have not yet been identified.
As with S6K1 phosphorylation, 4E-BP1 hyperphosphoryla-tion was rapidly stimulated and sustained for an extendedperiod of time. The increase in phosphorylation of 4E-BP1corresponded to a decrease in 4E-BP1 � eIF4E association.Thus, like S6K1 and rpS6 phosphorylation, the changes in4E-BP1 paralleled the observed stimulation of 5�-TOP trans-lation. In contrast, the association of 4E-BP2 with eIF4E wasnot altered at any time up to 180 min after feeding. 4E-BP2has not been studied extensively although it shares 60% se-quence identity with 4E-BP1, and the residues phosphorylatedon 4E-BP1 are fairly well conserved (21). The conclusion fromprevious studies is that 4E-BP1 and 4E-BP2 are regulated inthe same manner and serve the same function, i.e., to sequestereIF4E. However, the data presented in the present work andelsewhere (29) suggest that 4E-BP1 and 4E-BP2 are differen-tially regulated, raising the possibility that they may servedifferent functions in the regulation of the translational con-trol of protein synthesis. It is interesting to speculate that4E-BP1 may be involved primarily in the regulation of 5�-TOPtranslation, whereas 4E-BP2 may have other functions inregulating translation initiation. Future studies using micelacking either 4E-BP1 or 4E-BP2 could address this issue.
Unlike the increased phosphorylation of mTOR substratesin response to feeding, a decrease in eIF4E (Ser 209) phos-phorylation was observed within 60 min and maintainedthrough the remainder of the time course. The result was notentirely unexpected because oral administration of leucinesignificantly reduces eIF4E (Ser 209) phosphorylation in skel-etal muscle, despite a stimulation of protein synthesis andmTOR-mediated signaling (30). However, the results of thepresent work demonstrate once again an association betweena decrease in eIF4E (Ser 209) phosphorylation and an increasein the global rate of total protein synthesis in rat liver, al-
though the functional consequences of decreased eIF4E (Ser209) phosphorylation are unclear.
eIF4G (Ser 1108) phosphorylation increases in cells inculture after serum stimulation (31) and parallels an increasedrate of protein synthesis (32), but the functional consequenceof the increase in phosphorylation at Ser1108 or a subsequentresidue has not been elucidated. Phosphorylation of eIF4G(Ser1108) corresponded to the aggregation of ribosomes asindicated by the polysome profile analysis, and the observedmaximal elevation in phosphorylation at 60 min paralleled anincrease in the global rate of protein synthesis. Thus, althoughlittle is known about the function of eIF4G (Ser 1108) phos-phorylation, the data support a model in which eIF4G (Ser1108) phosphorylation functions in the regulation of theglobal rate of total protein synthesis. One possible function ofeIF4G (Ser 1108) phosphorylation that has not been exploredis that it may increase the affinity of eIF4G for other compo-nents of the eIF4F complex such as eIF4A. Another possibilitymight be to increase the association of eIF4E with eIF4G.However, in the present study eIF4E � eIF4G association de-creased by 180 min, a time at which eIF4G (Ser 1108) phos-phorylation was still elevated. The basis for the maintenanceof protein synthesis at a time when the eIF4G � eIF4E associ-ation had returned to the food-deprived control value is un-clear; however, it is possible that once mRNA is associatedwith polysomes, the requirement for the eIF4G � eIF4E com-plex in maintaining mRNA in polysomes is reduced, resultingin a sustained elevation of protein synthesis. Thus, furtherstudy of the functional consequence of eIF4G phosphorylationis required to gain insight into its importance in the regulationof protein synthesis. However, because a definitive role foreIF4G phosphorylation in modulating the function of theprotein has not been delineated, the possibility that thechanges in eIF4G phosphorylation observed in the presentstudy represent a nonfunctional modification cannot be elim-inated.
Another rate-controlling process in translation initiation isthe assembly of the 43S preinitiation complex consisting ofthe eIF2 � GDP � initiator methionyl tRNAi complex bound tothe 40S ribosomal complex. The rate-limiting enzyme for thisprocess is eIF2B, which catalyzes the exchange of GDP boundto eIF2 for GTP. The activity of eIF2B is regulated by phos-phorylation of eIF2� (Ser 51), which competitively inhibitsthe guanine nucleotide exchange activity of eIF2B. Previouswork demonstrated a correlation between eIF2B activity andchanges in the global rate of protein synthesis. Feeding ani-mals a diet devoid of 1 or more essential amino acids stimu-lated an increase in eIF2� (Ser 51) phosphorylation, a de-crease in eIF2B activity, and a decrease in the rate of globalprotein synthesis (12). However, 1 h after the feeding of acomplete meal, Yoshizawa et al. (6) did not observe anysignificant change in eIF2B activity. The possibility remainedthat an increase in eIF2B activity occurred during the first 60min of feeding. For this reason, eIF2B activity was assessed inthe present work, but no change in its activity, as measured byan in vitro activity assay, or in eIF2� (Ser51) phosphorylationwas observed at any time (data not shown). The results suggestthat the changes in protein synthesis and polysome aggrega-tion were not regulated through modulation of eIF2B activity.
Overall, a role for changes in phosphorylation of eIF4E andeIF4G must be considered because the time course for changesin these biomarkers is the same as that observed for alterationsin the global rate of protein synthesis. Additionally, the resultsillustrate a rapid and sustained stimulation of mTOR-mediatedsignaling that parallels an increase in polysomal association ofmany ribosomal protein mRNAs, as well as other mRNAs that
REITER ET AL.374
by guest on January 22, 2015jn.nutrition.org
Dow
nloaded from

are not known to possess a 5�-TOP sequence, and emphasizethe coordinated response to increase the rate of protein syn-thesis and ribosome biogenesis. Finally, after feeding, bothhepatic protein synthesis and ribosomal protein mRNA trans-lation undergo rapid and sustained stimulation, which is likelymediated by both mTOR-dependent and -independent signal-ing events.
ACKNOWLEDGMENTS
We thank Lynne Hugendubler and Sharon Rannels for technicalassistance and Samer Al-Murrani and Terrence Rager for assistancein the QRT-PCR work performed in the JDRF Functional GenomicsCore Facility.
LITERATURE CITED1. Munro, H. N. (1968) Role of amino acid supply in regulating ribosome
function. Fed. Proc. 27: 1231–1237.2. Kosterlitz, H. W. (1947) The effects of changes in dietary protein on the
composition and structure of the liver cell. J. Physiol. 106: 194–210.3. Yokogoshi, H. & Yoshida, A. (1986) Time-dependent changes in ag-
gregation of hepatic ribosomes after meal feeding of rats. J. Nutr. 116: 472–474.4. Yoshizawa, F., Kimball, S. R. & Jefferson, L. S. (1997) Modulation of
translation initiation in rat skeletal muscle and liver in response to food intake.Biochem. Biophys. Res. Commun. 240: 825–831.
5. Anthony, T. G., Reiter, A. K., Anthony, J. C., Kimball, S. R. & Jefferson,L. S. (2001) Deficiency of dietary EAA preferentially inhibits mRNA translationof ribosomal proteins in liver of meal-fed rats. Am. J. Physiol. 281: E430–E439.
6. Yoshizawa, F., Kimball, S. R., Vary, T. C. & Jefferson, L. S. (1998)Effect of dietary protein on translation initiation in rat skeletal muscle and liver.Am. J. Physiol. 275: E814–E820.
7. Gingras, A. C., Raught, B. & Sonenberg, N. (2001) Regulation oftranslation initiation by FRAP/mTOR. Genes Dev. 15: 807–826.
8. Wang, X., Li, W., Parra, J. L., Beugnet, A. & Proud, C. G. (2003) The Cterminus of initiation factor 4E-binding protein 1 contains multiple regulatoryfeatures that influence its function and phosphorylation. Mol. Cell Biol. 23: 1546–1557.
9. Pain, V. M. (1996) Initiation of protein synthesis in eukaryotic cells. Eur.J. Biochem. 236: 747–771.
10. Meyuhas, O. & Hornstein, E. (2000) Translational Control of TOPmRNAs. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
11. Burnett, P. E., Barrow, R. K., Cohen, N. A., Snyder, S. H. & Sabatini, D. M.(1998) RAFT1 phosphorylation of the translational regulators p70 S6 kinase and4E-BP1. Proc. Natl. Acad. Sci. U.S.A. 95: 1432–1437.
12. Anthony, T. G., Anthony, J. C., Yoshizawa, F., Kimball, S. R. & Jefferson,L. S. (2001) Oral administration of leucine stimulates ribosomal protein mRNAtranslation but not global rates of protein synthesis in the liver of rats. J. Nutr. 131:1171–1176.
13. Reiter, A. K., Anthony, T. G., Anthony, J. C., Jefferson, L. S. & Kimball,S. R. (2004) The mTOR signaling pathway mediates control of ribosomalprotein mRNA translation in rat liver. Int. J. Biochem. Cell Biol. 36: 2169–2179.
14. Garlick, P. J., McNurlan, M. A. & Preedy, V. R. (1980) A rapid andconvenient technique for measuring the rate of protein synthesis in tissues byinjection of [3H]phenylalanine. Biochem. J. 192: 719–723.
15. Kimball, S. R., Jurasinski, C. V., Lawrence, J. C., Jr. & Jefferson, L. S.(1997) Insulin stimulates protein synthesis in skeletal muscle by enhancing theassociation of eIF-4E and eIF-4G. Am. J. Physiol. 272: C754–C759.
16. Kubica, N., Kimball, S. R., Jefferson, L. S. & Farrell, P. A. (2004)Alterations in the expression of mRNAs and proteins that code for species
relevant to eIF2B activity after an acute bout of resistance exercise. J. Appl.Physiol. 96: 679–687.
17. Shah, O. J., Antonetti, D. A., Kimball, S. R. & Jefferson, L. S. (1999)Leucine, glutamine, and tyrosine reciprocally modulate the translation initiationfactors eIF4F and eIF2B in perfused rat liver. J. Biol. Chem. 274: 36168–36175.
18. Scheper, G. C., van Kollenburg, B., Hu, J., Luo, Y., Goss, D. J. & Proud,C. G. (2002) Phosphorylation of eukaryotic initiation factor 4E markedly re-duces its affinity for capped mRNA. J. Biol. Chem. 277: 3303–3309.
19. Lang, C. H., Wu, D., Frost, R. A., Jefferson, L. S., Kimball, S. R. & Vary,T. C. (1999) Inhibition of muscle protein synthesis by alcohol is associated withmodulation of eIF2B and eIF4E. Am. J. Physiol. 277: E268–E276.
20. Pullen, N. & Thomas, G. (1997) The modular phosphorylation andactivation of p70s6k. FEBS Lett. 410: 78–82.
21. Tsukiyama-Kohara, K., Vidal, S. M., Gingras, A. C., Glover, T. W., Hanash,S. M., Heng, H. & Sonenberg, N. (1996) Tissue distribution, genomic structure,and chromosome mapping of mouse and human eukaryotic initiation factor4E-binding proteins 1 and 2. Genomics 38: 353–363.
22. Mader, S., Lee, H., Pause, A. & Sonenberg, N. (1995) The translationinitiation factor eIF-4E binds to a common motif shared by the translation factoreIF-4 gamma and the translational repressors 4E-binding proteins. Mol. Cell Biol.15: 4990–4997.
23. Kimball, S. R., Jefferson, L. S., Nguyen, H. V., Suryawan, A., Bush, J. A.& Davis, T. A. (2000) Feeding stimulates protein synthesis in muscle and liverof neonatal pigs through an mTOR-dependent process. Am. J. Physiol. 279:E1080–E1087.
24. Jefferies, H. B., Reinhard, C., Kozma, S. C. & Thomas, G. (1994)Rapamycin selectively represses translation of the “polypyrimidine tract” mRNAfamily. Proc. Natl. Acad. Sci. U.S.A. 91: 4441–4445.
25. Jefferies, H. B., Fumagalli, S., Dennis, P. B., Reinhard, C., Pearson, R. B.& Thomas, G. (1997) Rapamycin suppresses 5�TOP mRNA translation throughinhibition of p70s6k. EMBO J. 16: 3693–3704.
26. Stolovich, M., Tang, H., Hornstein, E., Levy, G., Cohen, R., Bae, S. S.,Birnbaum, M. J. & Meyuhas, O. (2002) Transduction of growth or mitogenicsignals into translational activation of TOP mRNAs is fully reliant on the phos-phatidylinositol 3-kinase-mediated pathway but requires neither S6K1 nor rpS6phosphorylation. Mol. Cell Biol. 22: 8101–8113.
27. Tang, H., Hornstein, E., Stolovich, M., Levy, G., Livingstone, M., Temple-ton, D., Avruch, J. & Meyuhas, O. (2001) Amino acid-induced translation ofTOP mRNAs is fully dependent on phosphatidylinositol 3-kinase-mediated sig-naling, is partially inhibited by rapamycin, and is independent of S6K1 and rpS6phosphorylation. Mol. Cell Biol. 21: 8671–8683.
28. Hannan, K. M., Brandenburger, Y., Jenkins, A., Sharkey, K., Cavanaugh,A., Rothblum, L., Moss, T., Poortinga, G., McArthur, G. A., et al. (2003) mTOR-dependent regulation of ribosomal gene transcription requires S6K1 and is me-diated by phosphorylation of the carboxy-terminal activation domain of the nu-cleolar transcription factor UBF. Mol. Cell Biol. 23: 8862–8877.
29. von Manteuffel, S. R., Dennis, P. B., Pullen, N., Gingras, A. C., Sonenberg,N. & Thomas, G. (1997) The insulin-induced signalling pathway leading to S6and initiation factor 4E binding protein 1 phosphorylation bifurcates at a rapa-mycin-sensitive point immediately upstream of p70s6k. Mol. Cell Biol. 17: 5426–5436.
30. Anthony, J. C., Yoshizawa, F., Anthony, T. G., Vary, T. C., Jefferson, L. S.& Kimball, S. R. (2000) Leucine stimulates translation initiation in skeletalmuscle of postabsorptive rats via a rapamycin-sensitive pathway. J. Nutr. 130:2413–2419.
31. Raught, B., Gingras, A. C., Gygi, S. P., Imataka, H., Morino, S., Gradi, A.,Aebersold, R. & Sonenberg, N. (2000) Serum-stimulated, rapamycin-sensitivephosphorylation sites in the eukaryotic translation initiation factor 4GI. EMBO J.19: 434–444.
32. Lang, C. H., Frost, R. A., Deshpande, N., Kumar, V., Vary, T. C., Jefferson,L. S. & Kimball, S. R. (2003) Alcohol impairs leucine-mediated phosphorylationof 4E-BP1, S6K1, eIF4G, and mTOR in skeletal muscle. Am. J. Physiol. 285:E1205–E1215.
FEEDING-INDUCED CHANGES IN mRNA TRANSLATION 375
by guest on January 22, 2015jn.nutrition.org
Dow
nloaded from





























