Embed Size (px)
Citation preview

Bioresource Technology xxx (2016) xxx–xxx
Contents lists available at ScienceDirect
Bioresource Technology
journal homepage: www.elsevier .com/locate /bior tech
Medium engineering for enhanced production of undecylprodigiosinantibiotic in Streptomyces coelicolor using oil palm biomass hydrolysateas a carbon source
http://dx.doi.org/10.1016/j.biortech.2016.02.0550960-8524/� 2016 Elsevier Ltd. All rights reserved.
⇑ Corresponding author at: Department of Microbial Engineering, College of Engineering, Konkuk University, Seoul 143-701, South Korea. Fax: +82 2 3437 8360E-mail address: [email protected] (Y.-H. Yang).
Please cite this article in press as: Bhatia, S.K., et al. Medium engineering for enhanced production of undecylprodigiosin antibiotic in Streptomycecolor using oil palm biomass hydrolysate as a carbon source. Bioresour. Technol. (2016), http://dx.doi.org/10.1016/j.biortech.2016.02.055
Shashi Kant Bhatia a, Bo-Rahm Lee a, Ganesan Sathiyanarayanan a, Hun-Seok Song a, Junyoung Kim a,Jong-Min Jeon a, Jung-Ho Kim a, Sung-Hee Park b, Ju-Hyun Yu c, Kyungmoon Park d, Yung-Hun Yang a,d,e,f,⇑aDepartment of Microbial Engineering, College of Engineering, Konkuk University, Seoul 143-701, South Koreab Food Ingredients Center, Foods R&D, CheilJedang, Guro-dong, Guro-Gu, Seoul 152-051, South KoreacCenter for Industrial Chemical Biotechnology, Ulsan Chemical R&BD Division, Korea Research Institute of Chemical Technology, P.O. Box 107, 141 Gajeong-ro, Yuseong-gu,Daejeon 305-600, South KoreadDepartment of Biological and Chemical Engineering, Hongik University, Sejong Ro 2639, Jochiwon, Sejong City 339-701, South KoreaeMicrobial Carbohydrate Resource Bank, Konkuk University, Seoul 143-701, South Koreaf Institute for Ubiquitous Information Technology and Applications (CBRU), Konkuk University, Seoul 143-701, South Korea
h i g h l i g h t s
� Oil palm biomass hydrolysate hasfurfural and hydroxymethylfurfuralas inhibitors.
� Furfural effects xylose utilization in S.coelicolor adversely.
� Mycelia formation is decreased withreduction in actinorhodin production.
� Furfural elicits undecylprodigiosinproduction in S. coelicolor up to 52%.
� Engineered EFB hydrolysate mediaresulted a 88% higherundecylprodigiosin production.
g r a p h i c a l a b s t r a c t
a r t i c l e i n f o
Article history:Received 15 December 2015Received in revised form 1 February 2016Accepted 16 February 2016Available online xxxx
Keywords:AntibioticBiosugarFurfuralStreptomyces coelicolorUndecylprodigiosin
a b s t r a c t
In this study, a biosugar obtained from empty fruit bunch (EFB) of oil palm by hot water treatment andsubsequent enzymatic saccharification was used for undecylprodigiosin production, using Streptomycescoelicolor. Furfural is a major inhibitor present in EFB hydrolysate (EFBH), having a minimum inhibitoryconcentration (MIC) of 1.9 mM, and it reduces utilization of glucose (27%), xylose (59%), inhibits myce-lium formation, and affects antibiotic production. Interestingly, furfural was found to be a good activatorof undecylprodigiosin production in S. coelicolor, which enhanced undecylprodigiosin production by up to52%. Optimization by mixture analysis resulted in a synthetic medium containing glucose:furfural:ACN:DMSO (1%, 2 mM, 0.2% and 0.3%, respectively). Finally, S. coelicolorwas cultured in a fermenter in minimalmedium with EFBH as a carbon source and addition of the components described above. This yielded4.2 lg/mg dcw undecylprodigiosin, which was 3.2-fold higher compared to that in un-optimizedmedium.
� 2016 Elsevier Ltd. All rights reserved.
.
s coeli-

2 S.K. Bhatia et al. / Bioresource Technology xxx (2016) xxx–xxx
1. Introduction
Utilization of biomass for economical production of commer-cialized products is a subject of debate among the global scientificcommunity (Pandey et al., 2000). Lignocellulosic raw materials areprojected to play a major role in the development of various indus-trial bioprocesses (Patel et al., 2015). These raw materials are usu-ally composed of cellulose, hemicellulose and lignin, which areassociated with each other (Ahn et al., 2012; Jonsson et al.,2013). To use lignocellulose as a rawmaterial, it must be convertedto fermentable sugars by physio-chemical pretreatment and enzy-matic hydrolysis. Lignin is a complex structure of polyalcohols, andis highly resistant to degradation by microbial enzymes (Arantesand Saddler, 2010). Pretreatment is necessary to improve enzymeaccessibility and permeability to break down the complex cell wall,in which utilizable polysaccharides are embedded (Li et al., 2013).Various pretreatment methods using acids (such as sulfuric acidand phosphoric acid) or bases (such as ammonium hydroxide,sodium hydroxide, and calcium hydroxide) have been reported topromote conversion of the complex plant cell wall to fermentablesugars by hydrolytic enzymes (Kang et al., 2013; Sindhu et al.,2015). Lignocellulose pretreatment results in release of inhibitors;i.e., aldehydes (furan aldehydes), ketones, phenolics and organicacids, which influence microbial metabolism to utilize these freesugars as a carbon source for fermentation (Allen et al., 2010). Glu-cose and xylose degradation resulted in 5-hydroxymethylfurfural(HMF) and 2-furaldehyde (furfural), respectively (Kootstra et al.,2009). Furfural induces production of reactive oxygen species(ROS) in microorganisms, which cause cellular damage and affectgrowth (Allen et al., 2010). At present, little information aboutthe microbial inhibitors released during pretreatment of lignocel-lulosic biomass, their mechanism of action, effects on carbon uti-lization and secondary metabolite production is available. Hence,the generation of inhibitors during the production of lignocellu-losic hydrolysate and their effects on microbial fermentation war-rant investigation.
Streptomyces coelicolor, an actinobacterium, produces variousindustrially important secondary metabolites (Borodina et al.,2008). S. coelicolor can synthesize twochemically distinct antibioticsas secondary metabolites—actinorhodin (a diffusible blue pigment)and the cell wall-associated, red-pigmented undecylprodigiosin(Kim et al., 2015a). Antibiotic production is controlled by many fac-tors, such as transcriptional regulators, quorum-sensing molecules(c-butyrolactone) concentrations, metabolic and nutritional status(vanWezel et al., 2000; Yang et al., 2005), and the proposed couplingof antibiotic synthesis and resistance genes (Hindra et al., 2010).These regulatory mechanisms can be altered by varying the cultureconditions and addition of various factors (Schaberle et al., 2014).Undecylprodigiosin is a focus of interest due to its immunosuppres-sive and anticancer properties (Williamson et al., 2006).
In this study, the effects of biomass-derived inhibitors (furfuraland hydroxymethylfurfural) on S. coelicolor growth, carbon sourceutilization and antibiotic production were investigated. Moreover,a minimal medium comprising EFB hydrolysate (EFBH) as a carbonsource was designed to enhance production of undecylprodigiosin.
2. Methods
2.1. Microorganisms, media and culture conditions
All reagents were purchased from Difco Laboratories (Becton–Dickinson Franklin Lakes, NJ, USA) and other chemicals; i.e., fur-fural and hydroxymethylfurfural (HMF) from Sigma–Aldrich (St.Louis, MO, USA). S. coelicolor A3 (2) M145 used for antibiotic pro-
Please cite this article in press as: Bhatia, S.K., et al. Medium engineering for ecolor using oil palm biomass hydrolysate as a carbon source. Bioresour. Techn
duction was purchased from the Korean Culture Type Collection(KCTC, South Korea). For spore production S. coelicolor was culti-vated on R5 agar plates for 7 days at 30 �C, harvested by scrapingand suspended in 20% (v/v) glycerol and stored at �80 �C. Seed cul-ture of S. coelicolor was prepared by inoculating spores in 50 ml ofLuria–Bertani (LB) liquid medium, with five 3 mm glass beads, andincubated at 30 �C with shaking at 200 rpm. The germinated sporeswere harvested by centrifugation (3200g, 4 �C, 10 min) and resus-pended in 5 ml of ion-free water. A 0.1 ml volume (2 � 106 CFU)of germinated seed culture was used as the inoculum. For produc-tion of undecylprodigiosin, S. coelicolor was cultured in minimalmedium (Difco Laboratories) containing EFB hydrolysate (EFBH)as a carbon source. EFB hydrolysate was prepared by hot watertreatment of empty fruit bunch at 190 �C for 15 min (Bench TopReactor, Model 4526, Parr Instruments), mechanical refining ofthe pretreated slurry using a valley beater (L & W, Norway), enzy-matic hydrolysis of the solid fraction of the pretreated EFB withCellic CTec2 (Novozymes, Denmark), and finally solid–liquid sepa-ration of the hydrolysate by centrifugal filtration (Eom et al., 2015).An HPLC system equipped with a Bio-Rad Aminex HPX-87H col-umn (Bio-Rad Co., Hercules, CA, USA) was used to analyze sugar,furfural and HMF contents. A mobile phase of 5 mM H2SO4 at aflow rate of 0.6 ml/min was used, and the column temperaturewas maintained at 50 �C. The EFBH concentration was adjustedto 1% glucose, and additional furfural (2 mM), DMSO (0.3%), andacetonitrile (0.2%) were added. S. coelicolor was cultured in 10 mlvolumes in the capped tubes with a capacity of 25 ml at 30 �C, withagitation at 160 rpm for 72 h, and then subjected to analysis ofantibiotic production.
2.2. Antibiotic extraction and quantification
At intervals of 24 h, 2 ml culture samples were removed, andbiomass and antibiotic production were estimated. The sampleswere divided into two aliquots. For actinorhodin estimation, anequal volume of 1 N NaOH was added to the culture aliquot. Thesample was then centrifuged at 4000g for 5 min and theactinorhodin concentration was determined by measuringthe absorbance at 633 nm (Horinouchi and Beppu, 1984). Undecyl-prodigiosin is a membrane-associated red pigment, and wasextracted from the cell pellet. An aliquot of S. coelicolor culturewas harvested by centrifugation (4000g for 5 min), suspended inmethanol and incubated at 37 �C with shaking at 200 rpm for 1 h.Cells were removed by centrifugation at 4000g for 5 min and0.1 N HCl was added to adjust pH. To quantify undecylprodigiosin,absorbance at 533 nm was measured and the concentration calcu-lated (Horinouchi and Beppu, 1984).
2.3. Targeting sub-lethal concentrations of furfural and HMF
EFB hydrolysate has furfural as the main inhibitory component,with trace amounts of HMF. The effect of these components ongrowth and antibiotic production by S. coelicolor was investigated.It is not possible to assess the effect of these inhibitors in complexEFBH, so a minimal medium containing glucose as carbon sourcewas used. S. coelicolor was cultured in minimal medium in thepresence of 1% glucose, 0.1% yeast extract and 0–4 mM inhibitorsat 30 �C for 72 h. The minimum inhibitory concentration (MICs)of the inhibitors against S. coelicolor was calculated. Antibioticextraction was also performed to estimate actinorhodin and unde-cylprodigiosin concentrations. To make stock solutions, com-pounds were dissolved in water and sterile filtered prior to theiraddition to the fermentation media.
nhanced production of undecylprodigiosin antibiotic in Streptomyces coeli-ol. (2016), http://dx.doi.org/10.1016/j.biortech.2016.02.055

Table 1Experimental design points selected by the simplex lattice methodology for the mixture analysis model using as variables glucose, furfural, acetonitrile (ACN) anddimethylsulfoxide (DMSO), and their subsequent effects on biomass, undecylprodigiosin (red) and actinorhodin (blue).
ID# Glucose Furfural ACN DMSO Biomass (mg dcw/ml) Red (lg/mg dcw) Blue (lg/mg dcw)
1 0.125 0.625 0.125 0.125 1.65 2.33 2.712 0.125 0.125 0.125 0.625 2 1.84 1.983 0.000 1.000 0.000 0.000 0.35 0.00 0.004 0.000 0.500 0.500 0.000 0.65 0.00 0.005 1.000 0.000 0.000 0.000 3.3 0.66 21.96 0.125 0.125 0.625 0.125 1.6 2.25 1.767 0.333 0.333 0.333 0.000 2.2 2.86 1.058 0.625 0.125 0.125 0.125 3.05 2.94 8.999 0.000 0.000 0.000 1.000 0.4 0.00 0.0010 0.000 0.500 0.000 0.500 0.65 0.00 0.0011 0.500 0.500 0.000 0.000 1.7 2.61 7.0512 0.333 0.000 0.333 0.333 2.8 2.60 9.7113 0.000 0.333 0.333 0.333 0.25 0.00 0.0014 0.500 0.000 0.500 0.000 1.7 2.53 5.3015 0.250 0.250 0.250 0.250 2 3.42 9.9316 0.500 0.000 0.000 0.500 2.25 1.99 25.817 0.333 0.333 0.000 0.333 2.7 2.76 18.218 0.000 0.000 0.500 0.500 0.1 0.00 0.0019 0.000 0.000 1.000 0.000 0.15 0.00 0.00
S.K. Bhatia et al. / Bioresource Technology xxx (2016) xxx–xxx 3
2.4. Effect of furfural on antibiotic production and carbon utilization
EFBH contains glucose as the major sugar together with traceamounts of xylose, so the effect of furfural on the utilization ofthese sugars was investigated. S. coelicolorwas cultured in minimalmedium, supplemented with 10 g/l glucose or 10 g/l xylose orboth, in the presence of furfural (3 mM). Samples were collectedat 24 h intervals for 72 h and biomass, antibiotic production, car-bon source and furfural utilization evaluated.
2.5. Effect of furfural
2.5.1. Field emission scanning electron microscopy (FESEM), total fattyacid profile and transcriptional analysis
To assess the effect of furfural, S. coelicolor was cultured withand without furfural and morphological changes monitored usingFESEM. Samples were prepared (Ishii et al., 2004) and FESEM wasperformed using a SUPRA 55VP FESEM (Carl Zeiss, Oberkochen,Germany) using a 15 kV accelerating voltage, and photographicimages were captured digitally. Total fatty acid of S. coelicolor, cul-tured with and without furfural was extracted and analyzed forcomposition as described already (Bhatia et al., 2015a). Transcrip-tional analysis of antibiotic regulatory genes (redD and actII-orf4)was performed using mRNA extraction and RT-PCR (Kim et al.,2015b).
2.5.2. Fluorescence spectroscopyTo assay reactive oxygen species (ROS), S. coelicolor cells grown
in the presence of furfural for 72 h were subjected to 20,70-dichlorodihydrofluorescein diacetate staining, as described previously withslight modification (Dwivedi et al., 2014). A stock solution of 20,70-dichlorodihydrofluorescein diacetate (1 mg/ml) was prepared inDMSO and stored in the dark. S. coelicolor staining was performedby incubating 200 ll of culture with 20,70-dichlorodihydrofluorescein diacetate (50 lg/ml) for 18 h at room temperature. Fluores-cence spectroscopy was performed to determine ROS using a Per-kin Elmer LS-55 fluorescence spectrometer with maximumexcitation and emission spectra at 495 and 529 nm, respectively.
2.5.3. Effect of antioxidants and organic solvents onundecylprodigiosin
Antioxidants; i.e., ascorbic acid, glycine betaine, dithiothreitol(DTT) and dimethyl sulphoxide (DMSO) were added (0.1%) to theculture medium to reduce the effects of ROS on S. coelicolor. The
Please cite this article in press as: Bhatia, S.K., et al. Medium engineering for ecolor using oil palm biomass hydrolysate as a carbon source. Bioresour. Techn
effects of acetone, acetonitrile and methanol on undecylprodi-giosin production were also investigated by adding (0.1%) in theculture media.
2.6. Mixture design for interaction effect of glucose:furfural:ACN:DMSO
To assess the interaction effect and impact of various variables;i.e., glucose:furfural:ACN:DMSO on antibiotic production by S.coelicolor, a mixture analysis using Minitab 16 was performed.Mixture analysis experiments were designed using a simplex lat-tice method and 19 sets of experiments were performed. Theexperimental design with response is shown in Table 1. Experi-ments were performed in 10 ml volumes using glucose (20%), fur-fural (80 mM), acetonitrile (ACN; 20%) and DMSO (20%), atpredetermined ratios in minimal medium. Regression analysisusing ANOVA was performed and model fitting methods wereapplied for the data analysis. Contour surface plots were createdto determine the interaction effects of the four components on bio-mass production, and actinorhodin and undecylprodigiosin syn-thesis. The combination of predictor settings that optimized thefitted response was used to verify the model.
2.7. Antibiotic production using EFB hydrolysate
EFB hydrolysate (EFBH) was used as a carbon source in minimalmedium to enhance undecylprodigiosin production by S. coelicolor.Medium composition was adjusted according to optimized syn-thetic media composition using EFBH as carbon source. S. coelicolorwas cultured for 72 h and antibiotic extraction and estimation wasperformed as described above. Scale up of the undecylprodigiosinproduction process was also performed in 100 ml and 500 ml scaleusing a 250 ml conical flask and 1 l capacity fermenter (FerementecCo. Ltd.), respectively, with the optimum parameters of 10 mlscale.
3. Results and discussion
3.1. Effects of inhibitors on growth and antibiotic production of S.coelicolor
Furfural and HMF were investigated in terms of their effects onbiomass and antibiotic production of S. coelicolor. Biomass and acti-norhodin production by S. coelicolor decreased continuously with
nhanced production of undecylprodigiosin antibiotic in Streptomyces coeli-ol. (2016), http://dx.doi.org/10.1016/j.biortech.2016.02.055

Furfural (mM)
0 1 2 3 4 5
Bio
mas
s (m
g dc
w/m
l)
1.0
1.5
2.0
2.5
3.0
3.5
4.0
Act
inor
hodi
n (µ
g/m
g dc
w)
3.0
3.5
4.0
4.5
5.0
5.5
6.0
6.5
7.0
Und
ecyl
prod
igio
sin
(µg/
mg
dcw
)
1.0
1.2
1.4
1.6
1.8
2.0
BiomassActinorhodinUndecylprodigiosin
HMF (mM)
0 1 2 3 4 5
Bio
mas
s (m
g dc
w/ m
l)
2.5
3.0
3.5
4.0
4.5
5.0
5.5
Act
inor
hodi
n (µ
g/m
g dc
w)
2
3
4
5
6
7
Und
ecyl
prod
igio
sin
(µg/
mg
dcw
)
0.4
0.6
0.8
1.0
1.2
1.4
1.6
BiomassActinorhodinUndecylprodigiosin
(a)
(b)
Fig. 1. Determination of the sub-lethal concentrations of biomass derived inhibitors, furfural and hydroxymethylfurfural (HMF). Results are means ± S.D (n = 3), P < 0.05.
4 S.K. Bhatia et al. / Bioresource Technology xxx (2016) xxx–xxx
increasing furfural concentration, while undecylprodigiosinincreased (1.87 lg/mg dcw) up to 3 mM furfural, and decreasedthereafter (Fig. 1a). The addition of 2 mM HMF resulted in anincrease in biomass (5.0 mg dcw/ml) and undecylprodigiosin pro-duction (1.4 lg/mg dcw), but reduced actinorhodin production(Fig. 1b). Actinorhodin production in the presence of furfural andHMF decreased by 43% and 50%, respectively. The MICs for furfuraland HMF were 1.9 and 2.3 mM, respectively. Thus furfural is moretoxic to microbial fermentation than HMF (Heer and Sauer, 2008).
3.2. Effect of furfural on glucose and xylose utilization and antibioticproduction
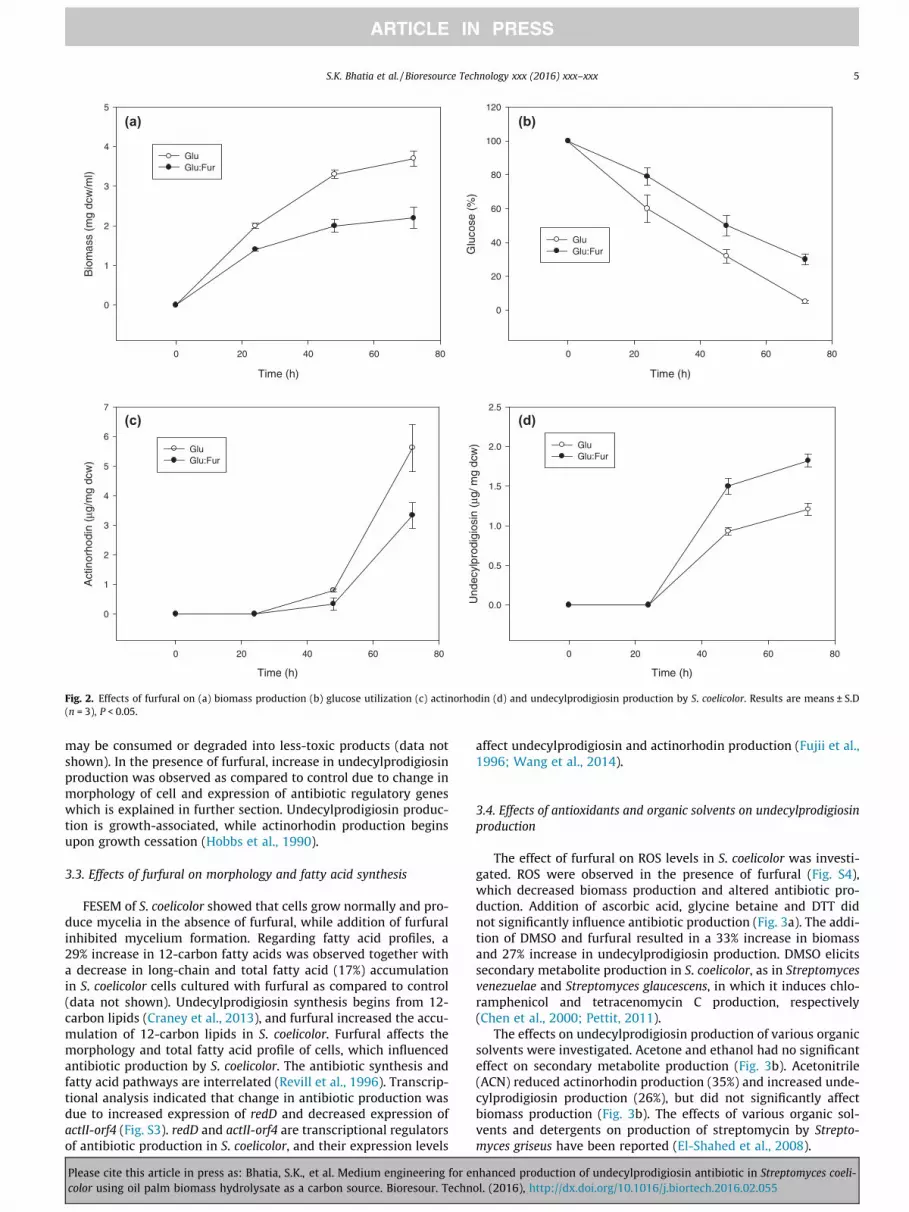
EFB hydrolysate contains glucose and xylose, so the effect offurfural on their utilization was determined. Biomass production,glucose and xylose utilization and antibiotic production were mon-itored for 72 h. Furfural reduced biomass production (41%, Fig. 2a),glucose utilization (27% decrease, Fig. 2b) and actinorhodin pro-
Please cite this article in press as: Bhatia, S.K., et al. Medium engineering for ecolor using oil palm biomass hydrolysate as a carbon source. Bioresour. Techn
duction (41%, Fig. 2c), but increased undecylprodigiosin production(52%, Fig. 2d). Xylose utilization was also reduced (59%, Fig. S1a),with a 44% reduction in biomass (Fig. S1b), and 49% reduction inactinorhodin production (Fig. S1c). Furfural enhanced undecyl-prodigiosin production by 9% (Fig. S1d), when xylose was used asthe carbon source. Furfural also effect glucose: xylose utilization,when used together as carbon source. S. coelicolor prefer to utilizeglucose as carbon source along with lower preference towardxylose (Fig. S2a). Biomass was reduced by 21%, with a 50% reduc-tion in actinorhodin and 22% in undecylprodigiosin production,respectively (Fig. S2b–S2d). Xylose utilization was affected to agreater extent than glucose utilization. Metabolism was affectedmore severely during xylose consumption than glucose consump-tion, which resulted in lower productivity when xylose was used asthe carbon source (Ask et al., 2013).
The furfural level decreased by 40%, 59% and 63% in the pres-ence of glucose, xylose and glucose: xylose, respectively, as carbonsources after 24 h. After 48 and 72 h no furfural was detected, as it
nhanced production of undecylprodigiosin antibiotic in Streptomyces coeli-ol. (2016), http://dx.doi.org/10.1016/j.biortech.2016.02.055
Time (h)
0 20 40 60 80
Bio
mas
s (m
g dc
w/m
l)
0
1
2
3
4
5
GluGlu:Fur
Time (h)
0 20 40 60 80
Glu
cose
(%
)
0
20
40
60
80
100
120
GluGlu:Fur
Time (h)
0 20 40 60 80
Act
inor
hodi
n (µ
g/m
g dc
w)
0
1
2
3
4
5
6
7
GluGlu:Fur
Time (h)
0 20 40 60 80
Und
ecyl
prod
igio
sin
(µg/
mg
dcw
)
0.0
0.5
1.0
1.5
2.0
2.5
GluGlu:Fur
(a) (b)
(c) (d)
Fig. 2. Effects of furfural on (a) biomass production (b) glucose utilization (c) actinorhodin (d) and undecylprodigiosin production by S. coelicolor. Results are means ± S.D(n = 3), P < 0.05.
S.K. Bhatia et al. / Bioresource Technology xxx (2016) xxx–xxx 5
may be consumed or degraded into less-toxic products (data notshown). In the presence of furfural, increase in undecylprodigiosinproduction was observed as compared to control due to change inmorphology of cell and expression of antibiotic regulatory geneswhich is explained in further section. Undecylprodigiosin produc-tion is growth-associated, while actinorhodin production beginsupon growth cessation (Hobbs et al., 1990).
3.3. Effects of furfural on morphology and fatty acid synthesis
FESEM of S. coelicolor showed that cells grow normally and pro-duce mycelia in the absence of furfural, while addition of furfuralinhibited mycelium formation. Regarding fatty acid profiles, a29% increase in 12-carbon fatty acids was observed together witha decrease in long-chain and total fatty acid (17%) accumulationin S. coelicolor cells cultured with furfural as compared to control(data not shown). Undecylprodigiosin synthesis begins from 12-carbon lipids (Craney et al., 2013), and furfural increased the accu-mulation of 12-carbon lipids in S. coelicolor. Furfural affects themorphology and total fatty acid profile of cells, which influencedantibiotic production by S. coelicolor. The antibiotic synthesis andfatty acid pathways are interrelated (Revill et al., 1996). Transcrip-tional analysis indicated that change in antibiotic production wasdue to increased expression of redD and decreased expression ofactII-orf4 (Fig. S3). redD and actII-orf4 are transcriptional regulatorsof antibiotic production in S. coelicolor, and their expression levels
Please cite this article in press as: Bhatia, S.K., et al. Medium engineering for ecolor using oil palm biomass hydrolysate as a carbon source. Bioresour. Techn
affect undecylprodigiosin and actinorhodin production (Fujii et al.,1996; Wang et al., 2014).
3.4. Effects of antioxidants and organic solvents on undecylprodigiosinproduction
The effect of furfural on ROS levels in S. coelicolor was investi-gated. ROS were observed in the presence of furfural (Fig. S4),which decreased biomass production and altered antibiotic pro-duction. Addition of ascorbic acid, glycine betaine and DTT didnot significantly influence antibiotic production (Fig. 3a). The addi-tion of DMSO and furfural resulted in a 33% increase in biomassand 27% increase in undecylprodigiosin production. DMSO elicitssecondary metabolite production in S. coelicolor, as in Streptomycesvenezuelae and Streptomyces glaucescens, in which it induces chlo-ramphenicol and tetracenomycin C production, respectively(Chen et al., 2000; Pettit, 2011).
The effects on undecylprodigiosin production of various organicsolvents were investigated. Acetone and ethanol had no significanteffect on secondary metabolite production (Fig. 3b). Acetonitrile(ACN) reduced actinorhodin production (35%) and increased unde-cylprodigiosin production (26%), but did not significantly affectbiomass production (Fig. 3b). The effects of various organic sol-vents and detergents on production of streptomycin by Strepto-myces griseus have been reported (El-Shahed et al., 2008).
nhanced production of undecylprodigiosin antibiotic in Streptomyces coeli-ol. (2016), http://dx.doi.org/10.1016/j.biortech.2016.02.055

Glu
Glu:Fur
Glu:Fur
:Asc
Glu:Fur
:Gly
Glu:Fur
:DTT
Glu:Fur
:DM
SO
Bio
mas
s (m
g dc
w/m
l)
0
1
2
3
4
Und
ecyl
prod
igio
sin
(µg/
mg
dcw
)
0.0
0.5
1.0
1.5
2.0
2.5
3.0
Act
inor
hodi
n (µ
g/m
g dc
w)
0
1
2
3
4
5
6
7
BiomassUndecylprodigiosinActinorhodin
Glu
Glu
:Fur
Glu
:Fur
:DM
SO
Glu
:Fur
:DM
SO
:Ace
Glu
:Fur
:DM
SO
:AC
NG
lu:F
ur:D
MS
O:E
th
Bio
mas
s (m
g dc
w m
L- )
0
1
2
3
4
Und
ecyl
prod
igio
sin
( µg
mg- d
cw)
0
1
2
3
4
5
Act
inor
hodi
n ( µ
g m
g- dcw
)
0
2
4
6
8BiomassUndecylprodigiosinActinorhodin
(a)
(b)
Fig. 3. Effects of antioxidants and organic solvents on S. coelicolor in minimal medium with glucose (Glu) as the carbon source and furfural (Fur) as an inhibitor. (a) Effects ofascorbic acid (Asc), glycine betaine (Gly), dithiothreitol (DTT) and dimethyl sulphoxide (DMSO) on antibiotic production. (b) Effects of acetone (Ace), acetonitrile (ACN) andethanol (Eth). Results are means ± S.D (n = 3), P < 0.05.
6 S.K. Bhatia et al. / Bioresource Technology xxx (2016) xxx–xxx
3.5. Mixture design for an interaction effect of glucose:furfural:ACN:DMSO
Contour surface plots for the interaction effects of glucose:fur-fural:ACN:DMSO ratios on various responses were generatedusing the Minitab 16 modeling software. ANOVA of variousresponses was performed, and p < 0.05 was taken as indicativeof statistical significance. The correlation coefficient r andadjusted coefficient R2 were both >92%, suggesting significance.The contour plot indicates that biomass production is dependentupon glucose and the roles of furfural, acetonitrile and DMSO arenegligible (Fig. 4a). All variables (glucose, furfural, ACN and
Please cite this article in press as: Bhatia, S.K., et al. Medium engineering for ecolor using oil palm biomass hydrolysate as a carbon source. Bioresour. Techn
DMSO) had a cumulative effect on undecylprodigiosin production(Fig. 4b). Glucose at higher concentrations inhibited antibioticproduction. Furfural and ACN inhibit production of actinorhodin,while DMSO has the opposite effect (Fig. 4c). The glucose:fur-fural:ACN:DMSO mixture analysis experiments suggest that fur-fural is a key player in differential production ofundecylprodigiosin, and co-feeding of ACN and DMSO furtherincreased undecylprodigiosin production. DMSO acts as anantioxidant and improves biomass production. Mixture design isan interesting approach to assessing the interaction effect of var-ious independent variables on response and their optimization formaximum production (Bhatia et al., 2015b).
nhanced production of undecylprodigiosin antibiotic in Streptomyces coeli-ol. (2016), http://dx.doi.org/10.1016/j.biortech.2016.02.055

(a)
(b)
(c)
Fig. 4. Mixture contour plots for interactive effects of glucose:furfural:DMSO:ACN(a) biomass production (mg dcw/ml) (b) undecylprodigiosin (lg/mg dcw) and (c)actinorhodin (lg/mg dcw).
S.K. Bhatia et al. / Bioresource Technology xxx (2016) xxx–xxx 7
3.6. Validation of model
A numerical optimization method was applied to predict thevalues of variables for the desired response. High and low valueswere adjusted for all responses using optimal parameter settings
Please cite this article in press as: Bhatia, S.K., et al. Medium engineering for ecolor using oil palm biomass hydrolysate as a carbon source. Bioresour. Techn
as recommended by the statistical software (Minitab 16) to obtainthe most suitable responses. D is the composite desirability and d isthe individual desirability, and their value under optimal condi-tions was close to 1.0, which confirmed the suitability of themodel. To verify the design, we chose a model to produce undecyl-prodigiosin (3.1 lg/mg dcw) with optimum biomass of2.5 mg dcw/ml and minimum actinorhodin of 10 lg/mg dcw. Theoptimum values for glucose:furfural:ACN:DMSO were predictedto be 0.484:0.252:0.151:0.111 (Fig. S5). Use of these predicted val-ues resulted in undecylprodigiosin production of 3.01 lg/mg dcwand biomass of 2.4 mg dcw/ml. The verification revealed the modelto have a high degree of accuracy (>97%), indicating its validityunder the test conditions.
3.7. Undecylprodigiosin production using EFB hydrolysate
The composition of EFB hydrolysate (EFBH) was determined.Glucose is the major carbon source (12.8%), followed by xylose/-galactose/mannose (1.58%); other sugars are present in traceamounts. Hot water treatment of EFB leads to production ofbyproducts; i.e., furfural (0.04%) and hydroxymethylfurfural(HMF), in trace amounts.
EFBH was used as a carbon source and the concentrations ofother components (furfural, ACN, DMSO) were adjusted accordingto the predicted response of synthetic media. S. coelicolor was cul-tured in minimal medium for 72 h with EFBH:furfural:ACN:DMSOcomponents, and resulted into biomass (3.1 mg dcw/ml), undecyl-prodigiosin (3.0 lg/mg dcw) and 5.0 lg/mg dcw actinorhodin pro-duction (Fig. 5). An EFBH minimal medium without furfural, ACNand DMSO resulted in production of biomass (3.4 mg dcw/ml),undecylprodigiosin (1.6 lg/mg dcw) and actinorhodin (3.4 lg/mg dcw). Engineered synthetic medium with glucose:furfural:ACN:DMSO and engineered EFBH:furfural:ACN:DMSO mediumyielded almost identical undecylprodigiosin production, 3.2 and3.0 lg/mg dcw, respectively (Fig. 5). Medium engineering to utilizeEFBH as a carbon source, with the addition of furfural, ACN andDMSO, resulted in an 88% increase in undecylprodigiosinproduction.
3.8. Scale up of undecylprodigiosin production in EFBH medium
Scale up of S. coelicolor to the 100 ml scale produced resultsidentical to those at the 10 ml scale; i.e., 3.01 lg/mg dcw undecyl-prodigiosin with 3.32 mg dcw/ml biomass, and cells exhibitedidentical morphology and growth profile. Upon further scale upto the fermenter level (500 ml), 4.2 lg/mg dcw undecylprodigiosinproduction with 4.8 mg dcw/ml biomass was recorded. Undecyl-prodigiosin production increased by 26% with 44.5% increase inbiomass in comparison to 10 ml scale. Better biomass productionlikely due to enhanced oxygen-transfer and mixing power in a fer-menter (Zhang et al., 1996). Medium engineering using EFBH ascarbon source resulted in a 3.2-fold increase in undecylprodigiosinproduction.
4. Conclusion
Furfural decrease biomass production, utilization of glucose andxylose and enhances undecylprodigiosin production by affectingcell morphology and the expression of the antibiotic regulatorygenes redD and actII-orf4. There is need to engineer S. coelicolorcells to tolerate furfural and improving their xylose utilizationcapability. Lignocellulosic biomass may be a better carbon sourceand the effects of various biomass-derived inhibitors on microbialfermentation, metabolism and carbon utilization should be inves-
nhanced production of undecylprodigiosin antibiotic in Streptomyces coeli-ol. (2016), http://dx.doi.org/10.1016/j.biortech.2016.02.055

Glu
Glu:Fur
:DM
SO:ACN
EFBH
EFBH:Fur
:DMSO:A
CN
Bio
mas
s (m
g dc
w m
L- )
0
1
2
3
4
Und
ecyl
prod
igio
sin
( µg
mg- d
cw)
0
1
2
3
4
Act
inor
hodi
n ( µ
g m
g- dcw
)
0
2
4
6
8
10
12
14BiomassUndecylprodigiosinActinorhodin
Fig. 5. Antibiotic production using EFB hydrolysate as a carbon source in minimal medium. (a) Comparison of minimal medium (Glu), engineered minimal medium (Glu:Fur:DMSO:ACN), minimal medium with EFBH as a carbon medium with EFBH as a carbon source.
8 S.K. Bhatia et al. / Bioresource Technology xxx (2016) xxx–xxx
tigated to determine the optimum growth medium formulation formaximum productivity.
Acknowledgements
This study was partially supported by the National ResearchFoundation of Korea (NRF) funded by the Ministry of Education(NRF-2011-619-E0002, NRF-2015M1A5A1037196) and AdvancedProduction Technology Development Program, Ministry of Agricul-ture, Food and Rural Affairs, Republic of Korea (1201349190011).This work was also supported by the R&D Program of MOTIE/KEIT(10049674) and the Energy Efficiency and Resources of the KoreaInstitute of Energy Technology Evaluation and Planning (KETEP)grant funded by the Korea Government Ministry of Trade, Industryand Energy (20133030000300). Also this article was also sup-ported by the KU research Professor program of Konkuk University,Seoul, South Korea.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at http://dx.doi.org/10.1016/j.biortech.2016.02.055.
References
Ahn, Y., Hu, D.H., Hong, J.H., Lee, S.H., Kim, H.J., Kim, H., 2012. Effect of co-solvent onthe spinnability and properties of electrospun cellulose nanofiber. Carbohydr.Polym. 89 (2), 340–345.
Allen, S.A., Clark, W., McCaffery, J.M., Cai, Z., Lanctot, A., Slininger, P.J., Liu, Z.L.,Gorsich, S.W., 2010. Furfural induces reactive oxygen species accumulation andcellular damage in Saccharomyces cerevisiae. Biotechnol Biofuels 3 (2), 1754–6834.
Arantes, V., Saddler, J.N., 2010. Access to cellulose limits the efficiency of enzymatichydrolysis: the role of morphogenesis. Biotechnol. Biofuels 3 (4), 1754–6834.
Ask, M., Bettiga, M., Mapelli, V., Olsson, L., 2013. The influence of HMF and furfuralon redox-balance and energy-state of xylose-utilizing Saccharomyces cerevisiae.Biotechnol. Biofuels 6 (1), 1754–6834.
Please cite this article in press as: Bhatia, S.K., et al. Medium engineering for ecolor using oil palm biomass hydrolysate as a carbon source. Bioresour. Techn
Bhatia, S.K., Yi, D.-H., Kim, Y.-H., Kim, H.-J., Seo, H.-M., Lee, J.-H., Kim, J.-H., Jeon, J.-M., Jang, K.-S., Kim, Y.-G., Yang, Y.-H., 2015a. Development of semi-syntheticmicrobial consortia of Streptomyces coelicolor for increased production ofbiodiesel (fatty acid methyl esters). Fuel 159, 189–196.
Bhatia, S.K., Yi, D.H., Kim, H.J., Jeon, J.M., Kim, Y.H., Sathiyanarayanan, G., Seo, H.M.,Lee, J.H., Kim, J.H., Park, K., Brigham, C.J., Yang, Y.H., 2015b. Overexpression ofsuccinyl-CoA synthase for poly (3-hydroxybutyrate-co-3-hydroxyvalerate)production in engineered Escherichia coli BL21(DE3). J. Appl. Microbiol. 119(3), 724–735.
Borodina, I., Siebring, J., Zhang, J., Smith, C.P., van Keulen, G., Dijkhuizen, L., Nielsen,J., 2008. Antibiotic overproduction in Streptomyces coelicolor A3 2 mediated byphosphofructokinase deletion. J. Biol. Chem. 283 (37), 25186–25199.
Chen, G., Wang, G.Y., Li, X., Waters, B., Davies, J., 2000. Enhanced production ofmicrobial metabolites in the presence of dimethyl sulfoxide. J. Antibiot. 53 (10),1145–1153.
Craney, A., Ahmed, S., Nodwell, J., 2013. Towards a new science of secondarymetabolism. J. Antibiot. 66 (7), 387–400.
Dwivedi, S., Wahab, R., Khan, F., Mishra, Y.K., Musarrat, J., Al-Khedhairy, A.A., 2014.Reactive oxygen species mediated bacterial biofilm inhibition via zinc oxidenanoparticles and their statistical determination. PLoS One 9 (11), e111289.
El-Shahed, K.Y., El-Diwany, A.I., Awad, H.M., 2008. Enhanced production ofstreptomycin and hydrolytic enzymes by Streptomyces griseus strains usingdifferent types of organic solvents and detergent compounds. Indian J.Biotechnol. 7, 341–348.
Eom, I.Y., Oh, Y.H., Park, S.J., Lee, S.H., Yu, J.H., 2015. Fermentative l-lactic acidproduction from pretreated whole slurry of oil palm trunk treated byhydrothermolysis and subsequent enzymatic hydrolysis. Bioresour. Technol.185, 143–149.
Fujii, T., Gramajo, H.C., Takano, E., Bibb, M.J., 1996. RedD and actII-ORF4, pathway-specific regulatory genes for antibiotic production in Streptomyces coelicolor A3(2), are transcribed in vitro by an RNA polymerase holoenzyme containingsigma hrdD. J. Bacteriol. 178 (11), 3402–3405.
Heer, D., Sauer, U., 2008. Identification of furfural as a key toxin in lignocellulosichydrolysates and evolution of a tolerant yeast strain. Microb. Biotechnol. 1 (6),497–506.
Hindra, Pak, P., Elliot, M.A., 2010. Regulation of a novel gene cluster involved insecondary metabolite production in Streptomyces coelicolor. J. Bacteriol. 192(19), 4973–4982.
Hobbs, G., Frazer, C.M., Gardner, D.C.J., Flett, F., Oliver, S.G., 1990. Pigmentedantibiotic production by Streptomyces coelicolor A3(2): kinetics and theinfluence of nutrients. Microbiology 136 (11), 2291–2296.
Horinouchi, S., Beppu, T., 1984. Production in large quantities of actinorhodin andundecylprodigiosin induced by afsB in Streptomyces lividans. Agric. Biol. Chem.48 (8), 2131–2133.
Ishii, S., Koki, J., Unno, H., Hori, K., 2004. Two morphological types of cellappendages on a strongly adhesive bacterium, Acinetobacter sp. strain Tol 5.Appl. Environ. Microbiol. 70 (8), 5026–5029.
nhanced production of undecylprodigiosin antibiotic in Streptomyces coeli-ol. (2016), http://dx.doi.org/10.1016/j.biortech.2016.02.055

S.K. Bhatia et al. / Bioresource Technology xxx (2016) xxx–xxx 9
Jonsson, L.J., Alriksson, B., Nilvebrant, N.O., 2013. Bioconversion of lignocellulose:inhibitors and detoxification. Biotechnol. Biofuels 6 (1), 1754–6834.
Kang, Y., Ahn, Y., Lee, S.H., Hong, J.H., Ku, M.K., Kim, H., 2013. Lignocellulosicnanofiber prepared by alkali treatment and electrospinning using ionic liquid.Fibers Polym. 14 (4), 530–536.
Kim, M., Yi, J.S., Lakshmanan, M., Lee, D.Y., Kim, B.G., 2015a. Transcriptomics-basedstrain optimization tool for designing secondary metabolite overproducingstrains of Streptomyces coelicolor. Biotechnol. Bioeng. 15 (10), 25830.
Kim, S.H., Traag, B.A., Hasan, A.H., McDowall, K.J., Kim, B.G., van Wezel, G.P., 2015b.Transcriptional analysis of the cell division-related ssg genes in Streptomycescoelicolor reveals direct control of ssgR by AtrA. Antonie van Leeuwenhoek 108(1), 201–213.
Kootstra, A.M.J., Mosier, N.S., Scott, E.L., Beeftink, H.H., Sanders, J.P.M., 2009.Differential effects of mineral and organic acids on the kinetics of arabinosedegradation under lignocellulose pretreatment conditions. Biochem. Eng. J. 43(1), 92–97.
Li, Z., Chen, C.H., Hegg, E.L., Hodge, D.B., 2013. Rapid and effective oxidativepretreatment of woody biomass at mild reaction conditions and low oxidantloadings. Biotechnol. Biofuels 6 (1), 1754–6834.
Pandey, A., Soccol, C.R., Nigam, P., Soccol, V.T., 2000. Biotechnological potential ofagro-industrial residues. I: sugarcane bagasse. Bioresour. Technol. 74 (1), 69–80.
Patel, S.K., Kumar, P., Singh, M., Lee, J.K., Kalia, V.C., 2015. Integrative approach toproduce hydrogen and polyhydroxybutyrate from biowaste using definedbacterial cultures. Bioresour. Technol. 176, 136–141.
Pettit, R.K., 2011. Small-molecule elicitation of microbial secondary metabolites.Microb. Biotechnol. 4 (4), 471–478.
Please cite this article in press as: Bhatia, S.K., et al. Medium engineering for ecolor using oil palm biomass hydrolysate as a carbon source. Bioresour. Techn
Revill, W.P., Bibb, M.J., Hopwood, D.A., 1996. Relationships between fatty acid andpolyketide synthases from Streptomyces coelicolor A3(2): characterization of thefatty acid synthase acyl carrier protein. J. Bacteriol. 178 (19), 5660–5667.
Schaberle, T.F., Orland, A., Konig, G.M., 2014. Enhanced production ofundecylprodigiosin in Streptomyces coelicolor by co-cultivation with thecorallopyronin A-producing myxobacterium, Corallococcus coralloides.Biotechnol. Lett. 36 (3), 641–648.
Sindhu, R., Binod, P., Pandey, A., 2015. Biological pretreatment of lignocellulosicbiomass – an overview. Bioresour .Technol. 24 (15), 01134–1137.
van Wezel, G.P., White, J., Hoogvliet, G., Bibb, M.J., 2000. Application of redD, thetranscriptional activator gene of the undecylprodigiosin biosynthetic pathway,as a reporter for transcriptional activity in Streptomyces coelicolor A3(2) andStreptomyces lividans. J. Mol. Microbiol. Biotechnol. 2 (4), 551–556.
Wang, W., Ji, J., Li, X., Wang, J., Li, S., Pan, G., Fan, K., Yang, K., 2014. Angucyclines assignals modulate the behaviors of Streptomyces coelicolor. Proc. Natl. Acad. Sci.USA 111 (15), 5688–5693.
Williamson, N.R., Fineran, P.C., Leeper, F.J., Salmond, G.P.C., 2006. The biosynthesisand regulation of bacterial prodiginines. Nat. Rev. Microbiol 4 (12), 887–899.
Yang, Y.H., Joo, H.S., Lee, K., Liou, K.K., Lee, H.C., Sohng, J.K., Kim, B.G., 2005. Novelmethod for detection of butanolides in Streptomyces coelicolor culture broth,using a His-tagged receptor (ScbR) and mass spectrometry. Appl. Environ.Microbiol. 71 (9), 5050–5055.
Zhang, J., Marcin, C., Shifflet, M.A., Salmon, P., Brix, T., Greasham, R., Buckland, B.,Chartrain, M., 1996. Development of a defined medium fermentation processfor physostigmine production by Streptomyces griseofuscus. Appl. Microbiol.Biotechnol. 44 (5), 568–575.
nhanced production of undecylprodigiosin antibiotic in Streptomyces coeli-ol. (2016), http://dx.doi.org/10.1016/j.biortech.2016.02.055





























