Embed Size (px)
Citation preview

Metabolic adaptation in Cryptococcus neoformans during earlymurine pulmonary infection
Guanggan Hu, Po-Yan Cheng, Anita Sham, John R. Perfect1, and James W. Kronstad*
Michael Smith Laboratories, The University of British Columbia, 2185 East Mall, Vancouver, BCV6T 1Z4, Canada1 Division of Infectious Diseases, Department of Medicine, Duke University Medical Center,Durham, North Carolina
SummaryThe pathogenic fungus Cryptococcus neoformans generally initiates infection in mammalian lungtissue and subsequently disseminates to the brain. We performed serial analysis of gene expression(SAGE) on C. neoformans cells recovered from the lungs of mice and found elevated expressionof genes for central carbon metabolism including functions for acetyl-CoA production andutilization. Deletion of the highly expressed ACS1 gene encoding acetyl-CoA synthetase revealeda requirement for growth on acetate and for full virulence. Transcripts for transporters (e.g., formonosaccharides, iron, copper and acetate) and for stress-response proteins were also elevatedthus indicating a nutrient-limited and hostile host environment. The pattern of regulation wasreminiscent of the control of alternative carbon source utilization and stress response by the Snf1protein kinase in Saccharomyces cerevisiae. A snf1 mutant of C. neoformans showed defects inalternative carbon source utilization, the response to nitrosative stress, melanin production andvirulence. However, loss of Snf1 did not influence the expression of a set of genes for carbonmetabolism that were elevated upon lung infection. Taken together, the results reveal specificmetabolic adaptations of C. neoformans during pulmonary infection and indicate a role for ACS1and SNF1 in virulence.
KeywordsSAGE; acetyl-CoA; glucose; melanin; virulence
IntroductionCryptococcus neoformans causes life-threatening meningoencephalitis in patients withimmune deficiency (Casadevell and Perfect, 1998). The fungus is found in the environmentas desiccated cells and/or basidiospores produced by sexual reproduction or monokaryoticfruiting; inhalation of these cells initiates a pulmonary infection in mammals (Lin andHeitman, 2006). Human exposure to C. neoformans is thought to be common based on thepresence of antibodies in the majority of normal individuals, and many cases ofcryptococcosis in immunocompromised individuals may result from reactivation of latentasymptomatic infections (Goldman et al., 2001). C. neoformans is a facultative intracellularpathogen during early stages of murine pulmonary infection (Feldmesser et al., 2000;Feldmesser et al., 2001). In fact, Feldmesser et al., (2000) found that the percentage offungal cells internalized in alveolar macrophages reached a peak during the first 24 hours of
*Correspondence: [email protected] Tel: 604-822 4732, Fax: 604-822 2114.
NIH Public AccessAuthor ManuscriptMol Microbiol. Author manuscript; available in PMC 2009 September 1.
Published in final edited form as:Mol Microbiol. 2008 September ; 69(6): 1456–1475. doi:10.1111/j.1365-2958.2008.06374.x.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

infection. Most of the yeast cells were extracellular by 24 hours, a time associated withmacrophage cytotoxicity, and both intracellular and extracellular cryptococci were observedat 48 hours and 7 days. Fungal cells eventually disseminate from the lung via thebloodstream and reach the brain to cause meningoencephalitis.
Although the disease process is well characterized, relatively few studies have examined C.neoformans gene expression during growth in vivo (Steen et al., 2003, Rude et al., 2002, Fanet al., 2005). In an initial study, a transcriptional profile was generated by differentialdisplay RT-PCR for C. neoformans cells during meningitis in an immunosuppressed rabbitmodel of meningitis (Rude et al., 2002). This analysis revealed elevated expression for thegene ICL1 encoding isocitrate lyase, a key enzyme in the glyoxylate cycle, suggesting thatthis cycle might be important for fungal growth in the host. However, disruption of ICL1 didnot influence virulence in two animal models nor cause a growth defect in macrophages(Rude et al., 2002). Similarly, deletion of the MLS1 gene encoding malate synthase (anotherglyoxylate cycle enzyme) did not influence virulence (Idnurm et al., 2007). Subsequenttranscriptional profiling of C. neoformans from the cerebrospinal fluid (CSF) of infectedrabbits by serial analysis of gene expression (SAGE) revealed relatively high expression ofgenes involved in energy production, stress response, and small molecule transport, as wellas carbohydrate, amino acid, and lipid metabolism (Steen et al., 2003). The transcriptionalresponse of C. neoformans cells upon phagocytosis by murine macrophages has also beenexamined (Fan et al., 2005). The fungus responds to phagocytosis with elevated expressionof genes at the MAT locus and in the cAMP/protein kinase A pathway. Additionally,elevated expression was seen for genes involved in autophagy, peroxisome function,membrane transport, lipid metabolism and the response to oxidative stress (Fan et al., 2005).These studies provide the first insights into C. neoformans gene expression during infectionor upon phagocytosis in vitro. Further studies are needed, however, because gene expressionprofiles could vary substantially for chronic versus acute infections or in different hosttissues, as demonstrated for other pathogens such as Candida albicans (Barelle et al., 2006;Fradin et al., 2003; Lorenz and Fink, 2001).
Studies in other pathogenic fungi examined gene expression during infection and exploredthe roles of glycolytic functions, the glyoxylate cycle, gluconeogenesis and β-oxidation invirulence (Lorenz and Fink, 2001, Lorenz et al., 2004; Idnurm and Howlett, 2002; Solomonet al., 2004; Wang et al., 2003; Piekarska et al., 2006; Barelle et al., 2006; Schöbel et al.,2006; Ramirez and Lorenz, 2007; Thewes et al., 2007; Olivas et al, 2008). For example, thetranscriptional response of C. albicans upon phagocytosis by macrophages includes the up-regulation of functions for the glyoxylate cycle, gluconeogenesis and fatty acid degradation(Lorenz et al., 2004; Prigneau et al., 2003). In addition, phagocytosis by neutrophils inducedan amino acid deprivation response in both C. albicans and S. cerevisiae (Rubin-Bejerano etal., 2003). Mutants defective in genes encoding glyoxylate pathway functions (e.g. isocitratelyase), gluconeogenesis functions (phosphoenolpyruvate carboxykinase) and glycolyticfunctions (phosphofructokinase and pyruvate kinase) are attenuated for virulence (Lorenzand Fink, 2001; Barelle et al. 2006). For β-oxidation, deletion of the FOX2 gene encodingthe second enzyme in the pathway also resulted in attenuated virulence (Piekarska et al.,2006; Ramirez and Lorenz, 2007). However, this phenotype may result from an influence onthe glyoxylate cycle because deletion of the PEX5 gene for peroxisomal biogenesis did notcause a virulence defect (Piekarska et al., 2006). In human blood, differentially expressedgenes in C. albicans encoded functions for a general stress response, an antioxidativeresponse, the glyoxylate cycle, and virulence factors (Fradin et al., 2003). Barelle et al.(2006) used GFP fusions to examine the expression of genes in the glyoxylate cycle,gluconeogenesis and glycolysis in more detail in C. albicans. They found that the genes forthe glyoxylate pathway and gluconeogenesis were repressed by the concentration of glucosefound in blood and that the genes were induced during phagocytosis (as found by other
Hu et al. Page 2
Mol Microbiol. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

investigators). Interestingly, these genes were not expressed in fungal cells in infectedkidneys. In contrast, glycolytic genes were not induced upon phagocytosis but wereexpressed in cells in infected kidneys. Barelle et al. (2006) concluded that the glyoxylatecycle and gluconeogenesis may be important early in infection and that glycolysis isimportant during systemic disease. In light of these results, the finding that components ofthe glyoxylate cycle were not required for virulence in C. neoformans suggests that thisfungus may have different nutritional requirements during infection (Idnurm et al., 2007).Similarly, deletion of the glyoxylate cycle genes for isocitrate lyase and malate synthase didnot result in virulence defects in Aspergillus fumigatus (Schöbel et al., 2007; Olivas et al.,2008).
In this study, we examined the early transcriptome changes that occur upon C. neoformansdeposition in the murine lung. We performed SAGE analysis on C. neoformans cellsrecovered from lungs at 8 and 24 hours after infection and compared the data withpreviously described SAGE libraries from cells grown in culture, and from cells harvestedfrom the central nervous system of infected rabbits. The SAGE data suggested that murinepulmonary infection represents a nutrient-limiting environment for invading C. neoformanscells; specifically, genes in several functional categories showed elevated transcriptsincluding alternative carbon source utilization, central carbon and lipid metabolism, stressresponse and virulence. To examine carbon source utilization in more detail, wecharacterized the ACS1 gene encoding acetyl-CoA synthetase and the SNF1 gene encoding apredicted serine/threonine protein kinase in C. neoformans. Loss of ACS1 resulted in poorgrowth on acetate and a mild attenuation of virulence. The SNF1 gene was examinedbecause the pattern of gene expression resembled the regulatory influence of the well-characterized Snf1 protein kinase in S. cerevisiae; this protein mediates glucose sensing,alternative carbon source utilization and the response to stress. Deletion of the SNF1 gene inC. neoformans gene resulted in poor growth on acetate and ethanol at 37°C, reducedmelanin production and a complete loss of virulence in a murine model of cryptococcosis.
ResultsSerial analysis of gene expression during early pulmonary infection
To investigate pathogen gene expression during pulmonary cryptococcosis, we generatedSAGE libraries from fungal cells harvested by bronchoalveolar lavage (BAL) from mouselungs at 8 (21,510 tags) and 24 hours (h) (20,129 tags) after infection. The details of theSAGE analysis are presented in Experimental Procedures and in Tables S1, S2 and S3 of theSupplementary material. A comparison of the two libraries identified 382 tags withdifferential levels at the two stages of infection suggesting dynamic changes duringadaptation to the host environment. The SAGE libraries were also compared with twopreviously described libraries constructed from cells grown in low iron medium (LIM)(77,829 tags; Hu et al., 2007) or in YNB broth at 37°C (84,039 tags; Steen et al., 2002).These are referred to as in vitro libraries. In addition, the SAGE data for fungal cells fromrabbit cerebral spinal fluid (CSF) (66,217 tags; Steen et al., 2003) were compared with thepulmonary infection libraries (in vivo libraries). Overall, these libraries provided snapshotsof RNA abundance in cells from quite different conditions of temperature (i.e., 37°C forboth in vitro and the in vivo lung libraries, and 39.5°C for rabbit), nutrients (i.e, YNB broth,low iron medium (LIM) and host environment) and stress (i.e., presence or absence of hostdefense responses). The SAGE data were normalized to 20,000 tags per library to facilitatecomparisons between all five conditions. For this analysis, we focused mainly oncomparisons between in vivo (mouse lung and rabbit CSF) and in vitro (LIM and YNBbroth) conditions. Following annotation to match tags to genes, we identified severalfunctional categories containing differentially expressed genes as shown in Tables 1 and 2,and Tables S4 and S5 of the Supplementary material. A notable general observation was that
Hu et al. Page 3
Mol Microbiol. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

the patterns of expression were most similar for the libraries prepared with cells from LIMand rabbit CSF (Table S2). For the bulk of the analysis described below, we employed thedata for the 8 h lung library and the LIM library to define differential expression with theprimary goal of identifying genes that showed elevated expression during lung infection.
Adaptation during infection involves changes in central carbon metabolismInitially, we identified a number of tags for genes involved in carbon metabolism that wereexpressed at high levels during lung infection. In particular, tags for functions in theglyoxylate pathway, the TCA cycle, gluconeogenesis, lipid catabolism and two-carbonmetabolism were elevated relative to the two in vitro SAGE libraries (Table 1). Notably,these results suggested a metabolic emphasis on the generation and utilization of acetyl-CoAupon infection. The position of acetyl-CoA in central carbon metabolism is summarized inFig. 1. For the glyoxylate cycle, the tag for the gene encoding malate synthase was notablyhigher in the lung libraries (60 times higher at 8 h and 25 times at 24 h) compared with thein vitro libraries. In contrast, the tag for this gene was not present in the normalized data forthe CSF library. Disruption of the MLS1 gene for malate synthase in C. neoformanseliminates the ability of the fungus to grow on acetate but does not affect virulence (Idnurmet al., 2007). Tags for genes encoding the TCA cycle enzymes aconitase and succinatedehydrogenase were also elevated in the lung libraries. However, tags representing threeother TCA cycle enzymes were lower in the lung libraries than in one or both of the in vitrolibraries (Table 1). The SAGE analysis also revealed that a tag for the gene encodingphosphoenolpyruvate carboxykinase (PCK1), which controls the only irreversible step ingluconeogenesis, was higher in both in vivo libraries (68 and 27 times at 8 h and 24 h afterinfection, respectively, versus the in vitro libraries). This result suggests that glucose may belimiting during early infection resulting in the activation of gluconeogenesis in at least aportion of the cells in the population. Panepinto et al., (2005) found that disruption of PCK1caused poor growth on lactate and markedly reduced virulence, thus indicating theimportance of gluconeogenesis during infection.
The possibility of glucose limitation is supported in part by observations that the tags forgenes encoding glycolytic functions including glucose-6-phosphate isomerase andphosphoglycerate mutase were relatively low in the lung libraries. However, this pattern wasnot consistent for all glycolytic functions because the tags for fructose 1,6-biphosphatealdolase, hexokinase and phosphofructokinase were present at similar levels between thelibraries or elevated in one of the lung libraries. Additionally, the tag for enolase was presentat low levels in the lung libraries compared to elevated levels in the libraries from cellsgrown in low iron medium or isolated from cerebral spinal fluid. Similarly, tags for differentgenes involved in the pentose phosphate pathway (PPP) showed a mixed pattern of elevated(e.g., 6-phosphogluconate dehydrogenase), similar (e.g., phosphoketolase) or reduced (e.g.,transaldolase) expression in the in vivo versus one or both of the in vitro libraries (Table 1).
The SAGE data also indicated that functions for lipid degradation and fatty acid catabolismwere elevated during infection. Specifically, a tag for a predicted butyrylcholinesterase(triacylglycerol lipase, CGL1) was one of most abundant in the 8 h lung library, but droppedin abundance at 24 h. The level of this tag was four times higher in the 8 h library versus theLIM library and >90 times higher than in the YNB broth library; tag abundance was similarin the 24h library and the LIM library (Table 1). Lavage fluid from mammalian lungs is richin phospholipids (Rooney et al., 1994;Rose et al., 1994) and fungal lipases such as theputative Cgl1 protein may contribute to carbon acquisition during lung colonization.Interestingly, the expression of this gene was induced during phagocytosis but attempts atdisruption were not successful suggesting that the gene is essential (Fan et al, 2005). Fattyacids generated by phospholipases and triacylglycerol lipases would enter the β-oxidationpathway and eventually yield acetyl-CoA. We did identify tags for components in the β-
Hu et al. Page 4
Mol Microbiol. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

oxidation pathway in the lung libraries (e.g., enoyl-CoA hydratase/isomerase, and 2,4-dienoyl-CoA reductase (NADPH)), suggesting a role for β-oxidation upon pulmonaryinfection (Table 1). We also identified tags for functions in sterol biosynthesis andregulation, including sterol-binding proteins and a hydroxymethylglutaryl-CoA synthase(Parks and Casey, 1995;Olkkonen et al., 2006).
The ability to convert acetate to acetyl-CoA contributes to virulenceThe expression of genes encoding enzymes for the production of acetyl-CoA from pyruvateand acetate were elevated during pulmonary infection. In particular, elevated expression wasfound for the genes for acetyl-coenzyme A synthetase, (ACS1), pyruvate decarboxylase, andaldehyde dehydrogenase, as well as two acetate transporters (Table 1). Acetate utilization orproduction is potentially relevant to the pathogenesis of C. neoformans becauseHimmelreich et al. (2003) showed that it was one of the major metabolites present ininfected tissue. Acetyl-CoA is generated by fatty acid catabolism, by decarboxylation ofpyruvate via the pyruvate dehydrogenase complex or by direct activation of acetate by theAcs1 enzyme (Fig. 1). We also found tags for functions involved in the conversion ofpyruvate, acetaldehyde, and ethanol to acetate (e.g., pyruvate decarboxylase and alcoholdehydrogenase) that were elevated in the lung libraries (Table 1). A tag for a relatedfunction, succinate:fumarate antiporter, was also elevated and this protein is required forgrowth on ethanol or acetate in yeast (Table 1;Palmieri et al., 1997). Notably, a tag foraldehyde dehydrogenase was one of the most abundant in both in vivo libraries (144 copiesat 8 h and 46 at 24 h). A similar gene in Ustilago maydis is required for growth on ethanol asa sole carbon source (Basse et al. 1996). Taken together, these tags suggest that acetate andacetyl-CoA play important metabolic roles during pulmonary infection.
To examine the role of ACS1 in more detail, we first confirmed the expression of the geneduring infection by real-time quantitative PCR (Fig. 2A). In S. cerevisiae, ACS1 is expressedduring growth on non-fermentable carbon sources and under aerobic conditions, and Acs1functions in the acetate utilization pathway (Schüller, 2003). ACS1 expression is alsoregulated by the Snf1 protein kinase in yeast (Young et al. 2003) and upregulated during thegrowth of C. albicans in blood (Fradin et al., 2003). We therefore reasoned that loss of Acs1function would impair the growth of C. neoformans on acetate. Indeed, deletion of ACS1resulted in a growth defect on acetate and ethanol, and also caused poor growth on glycerol(Fig. 2B). No growth defect was observed on sucrose, arabinose, galactose or lactic acid(data not shown). In contrast, the S. cerevisiae acs1 mutant can grow on ethanol but thereare conflicting reports about growth on acetate (De Virgilio et al., 1992;Kratzer andSchüller, 1995). Complementation of the C. neoformans acs1 mutant with ACS1 generestored growth to WT (wild type) levels (Fig. 2B). The acs1 mutant did not showdifferences in capsule or melanin formation, or growth at different temperatures, comparedwith the WT strain H99 (data not shown). However, the mutant did show a delay in diseaseprogression in a mouse inhalation model of cryptococcosis (Fig. 2C). That is, mice infectedwith the mutant survived 7–10 days longer than those infected with the WT orcomplemented strains (P < 0.001). These results indicate that the ability to produce acetyl-CoA directly from acetate makes a modest contribution to growth in the host. It is possiblethat the synthesis of acetyl-coA from pyruvate, from β-oxidation or by other acetyl-CoAsynthetases may limit the impact of loss of Acs1.
We have considered the possibility that other genes encode acetyl-CoA synthetase in C.neoformans. Our BLASTp searches with Acs1 and Acs2 from S. cerevisiae identified the C.neoformans gene product that we designated Acs1 (expect value = 0.0, 56% identity).However, two genes encoding proteins of lower similarity were also found and preliminarilynamed ACS2 and ACS3. Acs2 had an expect value of E-32 (26% identity) and Acs3 had anexpect value of E-13 (21% identity). For context, a BLASTp comparison of Acs1 and Acs2
Hu et al. Page 5
Mol Microbiol. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

from S. cerevisiae yielded an expect value of 0.0 with 57% identity. Thus, for C.neoformans, there was one clear ortholog for ACS1/ACS2 and two genes with weakersimilarity. Alignments of the yeast and C. neoformans polypeptides are shown in Fig. S1(Supplementary material). Preliminary experiments indicated that disruption of the C.neoformans ACS2 gene yielded mutants that lacked notable phenotypes in terms of growthon alternative carbon sources or virulence (data not shown). The construction of double andtriple mutants will be needed to determine whether loss of these genes exacerbates thephenotypes of the acs1 mutant.
Ttransport functions show elevated expression during lung infectionThe SAGE data indicated that C. neoformans cells contained elevated transcripts for severalputative transporters upon growth in the lung environmental (Table 1). Notably, a tag for agene encoding a putative glucose transporter (designated HXT1) was the most abundanttranscript in both lung libraries, a finding suggestive of glucose limitation (Table S3,Supplementary material). This gene shared high similarity to the yeast RGT2 and SNF3genes, and was also regulated by protein kinase A (PKA) as determined by SAGE analysis(Hu et al., 2007). We confirmed the elevation of the HXT1 transcript for the lung libraries byquantitative real-time PCR and found that deletion of HXT1 resulted in early melaninproduction (Fig. S2, Supplementary material). The influence on melanin formation isinteresting because elevated glucose is known to suppress this process (Zhu and Williamson,2004). The hxt1 mutant did not show a virulence defect (data not shown).
The other transporters had predicted functions in the movement of molecules such as aminoacids, sugars (e.g., trehalose), metals (iron and copper), organic acids (e.g., acetate) andphosphate (Table 1). Trehalose transport is of interest because the trehalose pathway in C.neoformans is involved in survival in the host, the response to high-temperature stress andglycolysis (Petzold et al., 2006). The tag for a putative phosphate transporter (Pho84) wasup-regulated at both 8 and 24 h during lung infection, and this gene was also induced in C.neoformans during phagocytosis (Fan et al., 2005). In S. cerevisiae, Pho84 is involved insensing nutrient signals and activates the cAMP-PKA pathway (Giots et al., 2003;Fan et al.,2005). The candidate metal transporters included the high-affinity iron permease(CFT1,Jung et al., 2006;2008) and a copper transporter (CTR4,Waterman et al., 2007).CFT1 was previously identified as an iron-regulated gene in C. neoformans cells and thegene is also regulated by PKA in C. neoformans (Lian et al., 2005;Hu et al, 2007). CFT1encodes an iron permease and is required for the use of iron from transferrin and forvirulence in C. neoformans (Jung et al., 2008). Overall, the elevated tags for transportfunctions suggest that specific assimilation activities are required to support fungalproliferation in host tissue.
Elevated expression of virulence-associated and stress response genes indicates that thelung is a stressful environment for C. neoformans
The SAGE analysis also identified a group of elevated tags representing functions related tovirulence and stress (Table 2). For example, two tags for the flavohemoprotein gene FHB1(flavohemoglobin denitrosylase) were abundant at 8 h and 24 h after infection. FHB1 wasinduced in fungal cells during murine macrophage infection and deletion of the generesulted in hypersensitivity to nitrosative stress and attenuation of virulence in a mousemodel (de Jesus-Berrios et al., 2003;Fan et al., 2005). A tag representing the gene encodingCu, Zn superoxide dismutase (SOD1) was also elevated during lung infection, especially inthe 8 h library. Superoxide dismutase has been shown to contribute to the virulence of C.neoformans and C. gattii (Cox et al., 2003;Narasipura et al., 2003;Narasipura et al., 2005).Interestingly, a tag representing the gene for the Sch9 protein kinase was elevated at 8 h. InC. neoformans, Sch9 modulates capsule formation and thermal tolerance, and contributes to
Hu et al. Page 6
Mol Microbiol. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

virulence both independently of and in conjunction with the cAMP-PKA pathway (Wang etal., 2004). We also found that a tag for a gene for peptidyl prolyl cis-trans isomerase (Ess1),an enzyme that mediates the folding of target proteins, was elevated at 8 h. Ess1 isdispensable for growth, haploid fruiting and capsule formation, but is required for virulencein C. neoformans (Ren et al., 2005). A tag matching the gene for the FK506-resistantcalcineurin B regulatory subunit was also identified in the early infection library.Calcineurin, a serine-threonine-specific calcium-activated phosphatase, is the target of theimmunosuppressive drugs cyclosporine A and FK506, and this protein influences themating, growth at 37°C and virulence in C. neoformans (Fox et al., 2001;Fox and Heitman,2005). Finally, the in vivo SAGE libraries revealed that the expression of two genesencoding mannitol-1-phosphate dehydrogenase (Mpd1), an enzyme involved in mannitolsynthesis, were elevated during infection (Table 2). Mannitol synthesis is thought to beimportant in cryptococcosis (Chaturvedi et al., 1996), and the MPD1 gene is induced undernitric oxide stress (Chow et al., 2007). Overall, the elevated expression of genes involved instress response and functions known to contribute to virulence likely reflects C. neoformansadaptation to a hostile environment in the host.
A homolog of SNF1 influences melanin production and virulenceIn evaluating the SAGE results for pulmonary infection, we noticed that the differentialexpression of genes for carbon metabolism, transport and stress showed similarities to theregulatory pattern defined for Snf1 in S. cerevisiae (Young et al., 2003; Hong and Carlson,2007). Snf1 is a serine/threonine protein kinase that plays a major role in nutrient responseand cellular metabolism, especially in gluconeogenesis and growth on alternative carbonsources (Celenza and Carlson, 1986). We therefore identified and characterized a candidateSNF1 homolog in C. neoformans to test the hypothesis that the Snf1 regulatory pathwayinfluences the patterns of expression that we observed during infection (Fig. S3,Supplemental material). Initially, we confirmed that a cDNA of the SNF1 gene from C.neoformans was able to restore the growth of a S. cerevisiae snf1 mutant on sucrose andraffinose (data not shown). Subsequently, a SNF1 deletion mutant was constructed in C.neoformans to examine phenotypes related to carbon source utilization and virulence. Thesnf1 mutant as well as the WT and complemented strains were able to grow on eitherglucose or sucrose at 30°C and therefore did not show the growth defect on sucroseobserved in S. cerevisiae (Fig. 3, Celenza and Carlson, 1984). A similar result was obtainedfor the plant pathogenic fungus Cochliobolus carbonum in which a SNF1 mutant was able togrow on both glucose and sucrose at 30°C (Tonukari et al., 2000). At 30°C, the C.neoformans snf1 strain also grew as well as the WT and complemented strains on arabinose,fructose, raffinose, galactose, mannose, lactate, acetate, ethanol, and glycerol (Fig. 3, anddata not shown). Interestingly, different phenotypes were observed at 37°C in that the C.neoformans snf1 mutant displayed a noticeable reduction in growth on sucrose and ethanol,markedly reduced growth on acetate, and similar growth to the WT and complementedstrains on other carbon sources (glycerol, lactate, galactose, raffinose and arabinose; Fig. 3and data not shown).
Next, we examined the response of the snf1 mutant of C. neoformans to oxidative, osmotic,salt, and nitrosative stress. We did not observe significant differences between the mutantand the WT strain when cells were grown on YNB medium supplemented with NaCl, KCl,sorbitol, H2O2 or menadione, suggesting that SNF1 was not required for protection againstthese osmotic and oxidative stresses (Fig. 4 and data not shown). However, the mutantexhibited increased sensitivity to sodium nitrite at 37°C (Fig. 4). The complemented strainrestored the WT level of growth on sodium nitrite at 37°C, indicating that a functional SNF1gene is required to withstand nitrosative stress. We also tested the snf1 mutant for alteredsensitivity to several drugs known to inhibit fungal growth. We found that the mutant
Hu et al. Page 7
Mol Microbiol. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

displayed increased sensitivity to the antifungal drug amphotericin B that is commonly usedto treat cryptococcosis, but a change in sensitivity to fluconazole was not observed (Fig. 4).We also found that the snf1 mutant was more sensitive to rapamycin, perhaps indicating aninteraction with the TOR pathway. The mutant showed equal sensitivity to FK506 andcyclosporine A compared to the WT strain (data not shown). We next examined the threemain virulence factors of the fungus: the ability to grow at 37°C, capsule formation andmelanin production. We did not observe differences in growth or capsule formation betweenthe wt, snf1 mutant and complemented strains at either 30°C or 37°C (data not shown). Itwas notable, however, that the snf1 mutant produced melanin at the same level as the WTstrain at 30°C, but was unable to produce visible melanin in colonies at 37°C (Fig. 4).Complementation of the snf1 mutation with the SNF1 gene again restored the ability of cellsto produce melanin at 37°C (Fig. 4).
The motivation to examine the role of SNF1 was based in part on the pattern of geneexpression observed in the SAGE data and also on evidence that alternative carbon sourcesare important for growth in the mammalian host (Lorenz and Fink, 2001). We therefore usedquantitative real-time PCR to determine whether loss of Snf1 influenced the expression of aset of genes selected from the SAGE analysis. In parallel, we tested the influence of growthon low glucose or acetate on expression of the same genes. As shown in Fig. 5A, loss ofSnf1 did not have an appreciable effect on the expression of the metabolic genes (e.g.,ACS1, PCK1, and MLS1), or the acetate transporters ADY2 and ATO2, when cells weregrown on 2% glucose, 0.2% glucose or 2% acetate. In contrast, growth of the WT strain atthe lower glucose level resulted in a substantial elevation in the transcripts for all of thegenes. The CFT1 gene showed a different pattern in that growth on the low glucose mediumresulted in elevated expression and Snf1 was required for part of this response. Thetranscripts for a subset of the genes responded to acetate with increased levels; theseincluded ACS1 (~2.6 fold), CGL1 (~2.5 fold), CFT1 (~3.7 fold), HXT1 (~3.7 fold) andMLS1 (~2.0 fold). Of these genes, loss of Snf1 had a positive influence on the level of theHXT1 transcript during growth on acetate. Overall, we concluded that glucose levels play amajor role in regulating the genes that we observed by SAGE to have elevated transcriptsduring lung infection, and that Snf1 plays little role for these genes. These results requirecautious interpretation, however, because we have not tested whether Snf1 influences geneexpression during infection. We did find that SNF1 transcript levels were increased in thecells grown on low glucose or acetate (Fig. 5B), but they were not substantially increased incells recovered from infected lungs (Fig. S3, Supplementary material).
Given the influence of the snf1 mutation on melanin formation, the response to nitrosativestress and growth on acetate, we also examined whether Snf1 controlled the expression ofgenes related to these phenotypes. Loss of Snf1 did have an interesting influence on theexpression of the LAC1 and LAC2 genes encoding the laccase enzymes for melaninproduction. The transcript for LAC2 was slightly elevated in WT cells grown on mediumwith 0.2% glucose or with acetate, and the transcript was more markedly elevated in the snf1mutant grown on acetate. The LAC1 transcript was ~8.0 fold higher in WT cells grown on0.2% glucose compared with cells grown on acetate or 2% glucose, and Snf1 was requiredfor this enhanced transcript level. Interestingly, the LAC1 transcript levels were notinfluenced by growth on acetate in either strain. Complementation of the snf1 mutation withSNF1 restored the WT pattern of regulation (data not shown). These results suggest that partof the melanin defect in the snf1 mutant results from an influence on LAC1 expression. Snf1did not appear to appreciably influence levels for stress-related genes (e.g., FHB1, SOD1,etc.) under any of the conditions (Fig. 5C).
Finally, we hypothesized that SNF1 would also be required for virulence because of themutant phenotypes for growth on alternative carbon sources, melanin production and the
Hu et al. Page 8
Mol Microbiol. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

response to nitrosative stress at 37°C. To test this idea, we infected A/Jcr mice with the WTstrain, the snf1 mutant and the complemented strain by intranasal inoculation. All miceinfected with the WT and complemented strains succumbed to infection by day 22–24,while those infected with the snf1 mutant survived to the end of the experiment at day 60(Fig. 6). Thus the snf1 mutant was avirulent compared with the other strains (P < 0.001). Atday 60, three mice infected with snf1 mutant were sacrificed and the lung and brain tissuewere analyzed for fungal burden. Mutant cells were found at an average burden of 2.95 ×105 CFU/g (SD = 1.73 × 105) in lung tissue, but were not detected in the brain samples forany of the mice. Histopathology of brain tissue at day 60 also failed to detect cells of thesnf1 mutant thus supporting the conclusion that the mutant was unable to disseminate to orpersist in the brain (Fig. S4, Supplementary material). This observation is interesting, giventhe melanization defect of the snf1 mutant, because previous work showed that a laccase-defective, non-melanized strain was unable to escape from the lung (Noverr et al. 2004). Tospecifically test whether the snf1 mutant could disseminate beyond lung tissue, weinoculated mice by inhalation with the mutant and WT strains, and we monitored thenumbers of fungal cells in the brains and lungs at day 17. This day was selected because itimmediately precedes the time that the mice succumb to infection with the WT strain. Thisexperiment revealed that the snf1 mutant was able to reach the brain, although the fungalburden for the mutant was quite low (average of 48.50 CFUs (SD = 14.64)) compared withthe burden of WT cells (average of 1.87 × 105 (SD = 1.02 × 104)). For comparison, the lungtissue at day 17 contained an average of 1.96 × 104 CFUs (SD = 3.90 × 103) for the mutantand an average of 1.10 × 108 CFUs (SD = 2.48 × 107 CFUs) for the WT. Combined with ourprevious analysis, these results indicated that the snf1 mutant reached the brain in lownumbers but failed to persist. Histopathology also confirmed the presence of the snf1 mutantin the lung (Fig. S4, Supplementary material).
DiscussionGene expression during pulmonary cryptococcosis
In this study, we employed SAGE to compare the C. neoformans transcriptomes at 8 and 24h of murine pulmonary infection with expression patterns from in vitro culture or from CSFin a rabbit model of experimental meningitis. These comparisons provided two main insightsinto the pathogenesis of C. neoformans. First, lung infection resulted in elevated expressionof fungal genes encoding functions for the production and utilization of acetyl-CoA. Theseincluded enzymes in the glyoxylate pathway, gluconeogenesis, β-oxidation and theconversion of pyruvate, ethanol and acetate to acetyl-CoA. Two candidate acetatetransporters also showed elevated expression. This pattern was set against a background ofelevated or reduced expression of specific components of the glycolytic pathway and theTCA cycle. As proposed for C. albicans (Fradin et al., 2003), this pattern may reflectvariations in gene expression profiles in subpopulations of cells exposed to different hostenvironments (e.g., extracellular and phagocytized cells). More interestingly, it could alsoreflect adjustments to the expression levels of pathway components to enhance theproduction of specific metabolic intermediates necessary for growth in host tissue. Given theearly times of infection that we analyzed, it is also possible that some of the changes in geneexpression for metabolic functions reflect the mobilization of glycogen and/or triglyceridestores to generate glucose and acetyl-CoA. A role for acetyl-CoA as a key metabolicintermediate was tested by deleting the ACS1 gene encoding a predicted acetyl-CoAsynthase; the resulting mutant was unable to grow on acetate and showed a delayed ability tocause disease. It is likely that production of acetyl-CoA by other reactions (e.g., β-oxidation,other acetyl-CoA synthetases) partially compensated for loss of ACS1 (Fig. 1).
The second insight from this study was that the transcript profiles observed during lunginfection suggested a role for the C. neoformans homolog of the yeast Snf1 kinase, a well-
Hu et al. Page 9
Mol Microbiol. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

characterized regulator of alternative carbon source utilization and the stress response in S.cerevisiae. Deletion of SNF1 in C. neoformans revealed an involvement in carbon sourceutilization, virulence and the regulation of melanin production. Interestingly, the snf1 mutantwas able to disseminate in quite low numbers to the brain following infection but was unableto persist, perhaps due to problems with carbon source utilization and/or the response to hostdefense mechanisms. SNF1 was specifically required for the elevated transcript levels of theLAC1 gene during growth on low glucose thus revealing a new regulatory function formelanin formation in C. neoformans. However, loss of SNF1 did not influence theexpression of the genes for carbon source utilization that we identified in the SAGE analysisof lung infection. Instead, we found that the transcript levels of these genes were elevated incells grown under low glucose conditions, a result consistent with the hypothesis thatglucose is limiting during pulmonary infection.
Glucose limitation and alternative carbon sources during infectionThe availability of glucose and local microenvironments in the mammalian host can have amarked influence on the expression of functions to exploit carbon sources other than glucose(e.g., acetate, lactic acid, fatty acids). For example, Barelle et al. (2006) examined geneexpression in C. albicans during infection in light of the observation that glucose is presentat sufficiently high levels in blood (0.06–0.1%) to limit expression of functions foralternative carbon source utilization. These studies indicated that C. albicans differentiallyregulates carbon assimilation pathways depending on the stage of infection (e.g., duringinteractions with phagocytic cells versus systemic infection) and the host tissue. Similarly,our comparison of the SAGE data for infections of the lung versus the CSF revealed that thepatterns of gene expression were quite different, in support of the idea that there may betissue-specific patterns of gene expression and adaptation for C. neoformans. The patterns ofgene expression found by Thewes et al. (2007) for C. albicans during intraperitonealinfection and ex vivo liver infection also indicated that at least some of the cells experience alow glucose environment. These investigators found upregulation of the PCK1 geneencoding phosphoenolpyruvate carboxykinase for gluconeogenesis as well as upregulationof genes for the synthesis of acetyl-CoA from pyruvate. However, specific glycolyticenzymes and TCA cycle enzymes were also upregulated indicating that some of the cellswere utilizing six-carbon sugars and respiration for energy production. We found similarsets of genes to be upregulated during cryptococcal pulmonary infection, and many of thesewere also regulated by glucose in vitro. The secretions from respiratory airways generallyhave a very low concentration of glucose in healthy individuals (<0.05 mM), and acuteillness, inflammation or diabetes can elevate glucose levels (Philips et al, 2003). Forexample, bronchial aspirates from intubated patients were found to have a mean glucoseconcentration of 3.5 mM and, interestingly, glucose levels correlated with risk of infectionfor some bacterial pathogens (Philips et al., 2005). We hypothesize that many of the C.neoformans cells entering the lung experience glucose starvation. This idea is supported bythe expression patterns that we observed and, in particular, by the identification of acandidate hexose transporter (HXT1) that was the most highly expressed gene (612 tags) inthe 8 h lung library. This tag was present at much lower levels in the LIM (26 tags) and theCSF (2 tags) libraries. Our in vitro analysis indicated that the expression of the gene wasdramatically higher at 0.2% glucose compared to 2% glucose. Thus the expression of thisgene may be a useful sensor/readout of glucose levels (although other unknown factorscould also influence the expression of the gene). The hxt1 mutant also showed precociousmelanin formation and this phenotype is consistent with a role for Hxt1 in glucose sensing.
During cryptococcal meningoencephalitis, a low glucose environment is thought to occur inbrain tissue and a three to five fold drop in glucose concentrations was found in thecerebrospinal fluid of infected rabbits (Perfect et al., 1980; Kwon-Chung et al., 2000; Rude
Hu et al. Page 10
Mol Microbiol. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

et al., 2002). However, our analysis of the library from experimental meningitis in the rabbitmodel (Steen et al., 2003) did not reveal elevated expression of the glucose responsive genesthat were observed in the pulmonary infection libraries. These results indicate that a morefocused comparison of C. neoformans gene expression in different host tissues is needed.
The role of gluconeogenesis in the virulence of C. neoformans has been explored as a resultof the identification of the PCK1 gene for phosphoenolpyruvate carboxykinase as adownstream target of Vad1, a DEAD-box RNA helicase (Panepinto et al., 2005). The tag forPCK1 was elevated in our SAGE analysis of pulmonary infection, but not in the macrophageinternalized cryptococcal cells (Fan et al., 2005), suggesting a difference between thepulmonary infection and the intracellular environment. Interestingly, a pck1 mutant of C.neoformans was unable to grow on lactate or to cause disease (Panepinto et al 2005). Thisobservation led to the proposal that 3-carbon sources, rather than 2-carbon molecules suchas acetate, are preferred during infection. Our finding that a acs1 mutant did not grow onacetate or ethanol and had reduced virulence is consistent with this idea. Similarly, icl1 andmls1 mutants also can’t use acetate as a sole carbon source but retain virulence (Rude et al.,2002; Idnurm et al., 2007). Further characterization of the icl1 mutant revealed that themutant also does not grow on ethanol and shows poor growth on lactate and glycerol (J.Perfect, unpublished results).
Roles for acetyl-CoA and acetate during adaptation to the lung environmentAcetyl-CoA is a central metabolite in the balance between carbohydrate metabolism andfatty acid catabolism (Fig. 1). Regulation of the balance may be a specific adaptation to thehost environment by C. neoformans, as suggested by the abundance of transcripts forenzymes mediating the production and utilization of acetyl-CoA during pulmonaryinfection. It is possible that the expression pattern that we observed reflects the utilization ofethanol and acetate as carbon sources during infection. However, as mentioned above,mutants that cannot utilize acetate in culture retain virulence in C. neoformans. One canspeculate that the production of acetyl-CoA may be specifically important during infectionby C. neoformans because acetyl-CoA is an important precursor for the synthesis of chitin inthe cell wall and for O-acetylation of the capsule via acetyltransferase activity (in addition toits key metabolic role). β-1,4 N-acetyl glucosamine has recently been shown to play a role inattachment of the capsule to the cell wall of C. neoformans and treatment with chitinase canrelease capsule polysaccharide (Rodrigues et al., 2008). Acetylation of capsulepolysaccharide plays a role in the ability of capsule polysaccharide to inhibit neutrophilmigration and a mutant lacking capsular acetyl groups is hypervirulent (Ellerbroek et al.,2004;Janbon et al., 2001). Thus, enhanced acetyl-CoA production may meet the demands ofcapsule-related biosynthetic functions during infection.
An additional intriguing aspect of pathogen metabolism during infection is the production ofexported metabolites that may condition the host environment and contribute to virulence.The SAGE data for pulmonary infection are particularly interesting in light of thecharacterization of the metabolites produced by C. neoformans in culture and incryptococcomas from rat lung and brain (Bubb et al., 1999; Wright et al., 2002;Himmelreich et al., 2003). These studies revealed that trehalose, mannitol, glycerol, acetate,ethanol and glycerophosphorylcholine were particularly abundant among the >30metabolites that were detected. Wright et al., (2002) discussed the implications of thesemetabolites for virulence including the contribution of mannitol to defense against oxidativekilling and the possibility that acetate acidifies the extracellular environment. In fact, directmeasurement of cerebral cryptococcomas revealed a relatively low pH of 5.4 to 5.6. Wrightet al. (2002) went on to demonstrate that C. neoformans supernatants at pH 5.5 inducednecrosis in neutrophils, reduced superoxide production and influenced chemotaxis by thesecells. Our observed elevated expression of transcripts for enzymes involved in the
Hu et al. Page 11
Mol Microbiol. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

conversion of pyruvate to metabolites such as acetaldehyde, acetate and ethanol in C.neoformans during pulmonary infection is consistent with the appearance of thesemetabolites in cryptococcomas. In particular, we found tags for two candidate acetatetransporters that were elevated during infection and we hypothesize that one or both of theseproteins may contribute to acetate export. In addition to an influence on the host immuneresponse, acidification of the local environment could also contribute to the availability ofiron by triggering its release from transferrin. Indeed, Friedman et al. (2006) have shownthat Staphylococcus aureus remodels its metabolism in response to iron limitation (a keyfeature of the host environment) to produce acidic products including lactic acid. For fungalpathogens, Thewes et al. (2007) also found a pattern of expression for pH-responsive genesin C. albicans during infection, as well as upregulated expression of transporters for iron,zinc and phosphate. These authors also noted the possible connection between pHmodulation of the host environment and iron acquisition. As mentioned earlier, we foundthat the transcript for the iron permease Cft1 was elevated in pulmonary infection suggestingthat C. neoformans is experiencing iron deprivation. We recently showed that ironacquisition during infection is partially dependent on Cft1 and that this permease is requiredfor transferrin utilization (Jung et al., 2008).
Snf1 and virulence-related gene expressionIn yeast, the Snf1 protein kinase is required for the metabolic shift that occurs upon glucosedepletion, for growth on alternative carbon sources, and for the response to a variety ofenvironmental stresses such as sodium and lithium salts, alkaline pH, heat shock, andhyperosmolarity (Hardie et al., 1998; Kemp et al., 1999; Kemp et al., 2003; Vyas et al.,2001), (Alepuz et al., 1997; Hong and Carlson, 2007; Portillo et al., 2005; Ye et al., 2006).We found that Snf1 in C. neoformans plays a role in the growth response to acetate, ethanoland sucrose, and that sensitivity of the snf1 mutant was noted for nitrosative stress and forthe drugs rapamycin and amphotericin B. We found that all of the snf1 phenotypes in C.neoformans were manifested at the host temperature of 37°C, but not at 30°C. In S.cerevisiae, a snf1 mutant has reduced thermotolerance, and carbon source utilization mutantsin C. albicans have more serve defects at 37°C (Thompson-Jaeger et al., 1991; Ramirez andLorenz, 2007). In C. neoformans, snf1 mutant strains grew as well as the WT strain onmedia (either YPD or YNB) with glucose (or glycerol, galactose, raffinose, arabinose) as thesole carbon source at 37°C, indicating that loss of SNF1 did not generally reducethermotolerance, but rather influenced specific phenotypes.
SNF1 has been implicated in the virulence of plant pathogenic fungi. For example, the SNF1gene in Cochliobolus carbonum is required for the expression of cell-wall-degradingenzymes and for disease symptom formation on maize (Tonukari et al., 2000). The SNF1gene in Fusarium oxysporum, a pathogen that causes vascular wilt disease in over 100cultivated plant species, regulates the transcription of genes encoding cell wall degradingenzymes and virulence on both Arabidopsis thaliana and Brassica oleracea (Ospina-Giraldoet al., 2003). So far there has been no description of a role for SNF1 in the virulence offungal pathogens of animals. A SNF1 homologue in C. albicans is essential for viability, anddisruption of one allele resulted in morphological changes and decreased growth, but did notinfluence virulence (Petter et al., 1997). The discovery of a role for SNF1 in the virulence ofC. neoformans reveals a new regulatory connection between carbon source utilization andgrowth in mammalian hosts. Part of the virulence defect may be due to reduced melaninproduction at 37°C. Laccase expression in C. neoformans is repressed by elevatedtemperature and enhanced by glucose starvation as well as copper, iron and calcium (Zhuand Williamson, 2004). Laccase expression also occurs during early pulmonary infection(Garcia-Rivera et al., 2005). Both LAC1 and LAC2 contribute to melanin production andboth are regulated by the cAMP pathway; differences have been noted for the two genes,
Hu et al. Page 12
Mol Microbiol. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

however (Missall et al., 2005; Pukkila-Worley et al., 2005). Notably, LAC2 but not LAC1 isregulated in response to oxidative and nitrosative stresses in manner that is influenced by theTSA1 gene encoding a thiol peroxidase (Missall et al., 2004, 2005). Missall et al., (2005)presented a model for the regulation of LAC2 expression in response to nitric oxide stressand proposed that a stress-activated protein kinase might function downstream of Tsa1.
In terms of contributions to virulence, the laccases produced by the LAC1 and LAC2 havedifferent but overlapping substrate specificities that may allow the utilization of differentdiphenolic substrates in brain tissue. The LAC1 gene appears to make the larger contributionto virulence in a mouse model of cryptococcosis, although mice still succumb to infectionwith a lac1 mutant (Pukkila-Worley et al., 2005). Given these results, it is likely that theSnf1 protein influences other aspects of virulence because a snf1 mutant fails to causedisease. It is possible that the melanin defect contributes to the observed poor disseminationof the snf1 mutant from the lung to the brain because melanin formation is known toinfluence this process (Noverr et al., 2004). However, the snf1 mutant also failed to persistin brain tissue, perhaps indicating that Snf1 influences the expression of additional virulencefactors. For example, Snf1 might regulate processes that are important for virulence in C.neoformans such as gluconeogenesis (Panepinto et al. 2005), as well as control theexpression of cell surface factors that mediate interactions with the host and/or influence theresponse to host defenses. Overall, the identification of a role for Snf1 presents opportunitiesto further characterize the role of the protein in the regulation of central metabolism, theutilization of alternative carbon sources and the response of the fungal cells to the stressfulconditions of the host environment.
Experimental proceduresStrains, plasmids and media
The serotype A strain H99 (C. neoformans var. grubii) and the S. cerevisiae strainsW303-1A (MATa ade2-1 trp1-1 his3–11, 15 can1-100 ura3-1 leu2–3, 112) and MCY4908(W303-1A SNF1Δ10) were used in the study. The strains were maintained on YPD medium(1% yeast extract, 2% peptone, 2% dextrose, and 2% agar). Selectable markers for thetransformation of C. neoformans were from plasmid pCH233 (nourseothricin resistance) andpJAF1 (neomycin resistance). Plasmid BPH618 was used for yeast transformation. YPDplates containing neomycin (200 μg/ml) were used to select C. neoformans transformantsand YPD plates containing noursethricin (100 μg/ml) were used to select the SNF1-complemented transformants. S. cerevisiae transformants were selected on YNB agar (yeastnitrogen base without amino acids and uracil) supplemented with 2% glucose and the otherrequired nutrients. YNB agar (without amino acids and uracil) was supplemented with 2%glucose, 2% sucrose, 2% raffinose, or 2% maltose, and used to examine the growth of theyeast snf1 mutant. YPD and/or YNB plates (YNB with amino acids) supplemented withdifferent inhibitors or chemicals were used for phenotypic experiments. Escherichia coliwas grown on LB broth or agar supplemented with 100 μg/ml of ampicillin at 37°C.
Isolation of C. neoformans cells from mouse lungs for SAGE analysisThe C. neoformans cells for mouse inoculation were grown overnight in YPD in a 30°Cshaker, washed with phosphate-buffered saline (PBS) and resuspended in PBS at aconcentration of 8.2 × 107 cells/50 μl. A total of 20 female A/Jcr mice were anesthetizedwith ketamine and xylazine, inoculated by nasal inhalation and subjected to bronchoalveolarlavage at 8 and 24 h. At each time point, the treated mice were euthanized, a small incisionwas made in the trachea, and a capillary tube was inserted toward the lungs. The tube wassecured by silk thread and a series of 0.5 ml aliquots of ice-cold water were flushed into thelungs. A total of 10 ml of water was used per mouse and the lavage fluids from each
Hu et al. Page 13
Mol Microbiol. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

inoculation group were pooled. The cells were washed twice with ice-cold water, frozen at−80°C and lyophilized for RNA isolation. In total, 3.0 × 108 cells were obtained at 8 h and2.6 × 108 cells were collected at 24 h.
SAGE library construction, sequencing and analysisSAGE library construction, sequencing and analysis were as previously described (Steen etal., 2002, 2003; Hu et al., 2007). Briefly, RNA was isolated from lyophilized cells byvortexing with glass beads (3.0 mm, acid-washed and RNase-free) for 15 min. in 15 ml ofTRIZOL extraction buffer (Invitrogen, Carlsbad, California). The mixture was incubated for15 min at room temperature, total RNA was isolated according to the manufacturer’sinstructions and RNA quality was assessed by agarose gel electrophoresis. Total RNA wasused directly for SAGE library construction as described by Velculescu et al., (1995) usingthe I-Long SAGE kit (Invitrogen). The tagging enzyme for cDNA digestion was NlaIII and29 PCR cycles were performed to amplify the ditags during library construction. Colonieswere screened by PCR (M13F and M13R primers) to assess the average clone insert size andthe percentage of recombinants. Clones from the libraries were sequenced by BigDye primercycle sequencing on an ABI PRISM 3700 DNA analyzer. Sequence chromatograms wereprocessed using PHRED (Ewing and Green, 1998), and vector sequence was detected usingCross_match (Gordon et al., 1998). Fourteen-base-pair tags were extracted from the vector-clipped sequence, and an overall quality score for each tag was derived based on thecumulative PHRED score. Duplicate ditags and linker sequences were removed as describedpreviously (Steen et al., 2002). Only tags with a predicted accuracy of ≥99% were used, andstatistical differences between tag abundance in different libraries were determined using themethods of Audic and Claverie (1997).
The libraries yielded 21,510 (8 h) and 20,129 (24 h) tags. An overview of the abundanceclasses for both SAGE libraries is presented in Table S1, Supplementary material, with boththe number of different tag sequences and the total number of tags present in each class forthe cells from lung tissue at 8h or 24 h after infection. An overview of pair wise analyses ofdifferential expression for all of the libraries is presented in Table S2, Supplementarymaterial, and the 100 most abundant tags in each library are listed in Table 3, Supplementarymaterial. All libraries were normalized to 20,000 to allow direct comparisons, and the tagsthat appeared less than once in any given library were removed. The EST database availablefor strain H99 at the University of Okalahoma’s Advanced Center for Genome Technology(http://www.genome.ou.edu/cneo.html) was used for the preliminary assignment of tags togenes. When an EST sequence could not be identified for a particular tag, the genomicsequence for H99 at the Duke University Center for Genome Technology(http://cgt.genetics.duke.edu/data/index.html) and the Broad Institute(http://www.broad.mit.edu/cgi-bin/annotation/fungi/cryptococcus_neoformans/) was used toidentify contigs with unambiguous tag assignments. Note that a limitation of the SAGEapproach is that some transcripts are not detected because of low abundance and/or theabsence of an NlaIII site for transcript processing.
Complementation of a S. cerevisiae snf1 mutation with the SNF1 gene of C. neoformansTo obtain a cDNA for SNF1, total RNA was isolated from frozen C. neoformans cells. ThecDNA was synthesized using random hexamer priming and Superscript transcriptase II(Invitrogen Canada). A 2,270 bp PCR product was obtained using primers SNF1-cDNA-5and SNF1-cDNA-6, cloned into TOPO-TA vector (pSNF1c-topo) and then subcloned intopBPH618 to obtain pCnSNF1c. The same pair of primers was used to amplify the SNF1gene from genomic DNA of strain H99 and a PCR product of 2950 bp was obtained. Thisproduct was cloned into a TOPO-TA vector (pSNF1g-TOPO) and subcloned into pBPH618to create pCnSNF1g. Both SNF1 inserts in pCnSNF1c and pCnSNF1g were sequenced to
Hu et al. Page 14
Mol Microbiol. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

confirm the presence of the C. neoformans gene. To complement yeast snf1 mutant with C.neoformans SNF1 homologue, pCnSNF1c and pBPH618 (empty vector) were transformedinto strain MCY4908 by PEG and lithium acetate treatment. Transformants were selected onSCD (dexose)-URA and 5-FOA plates for uracil prototrophy, and on SCS (sucrose) forsucrose utilization. The sequences of the primers for this study are given in Table S6,Supplementary material.
Deletion of the SNF1 gene in C. neoformansA snf1::NEO deletion allele was constructed using a modified overlap PCR procedure(Davidson et al., 2002; Yu et al., 2004). Briefly, the primers SNF1-1/SNF1–3 and SNF1–4/SNF1–6 (Table S6, Supplementary material) were used with genomic DNA to obtain the leftand right arms for the deletion construct. The NEO selectable marker was amplified usingprimers SNF1–2/SNF1–5 and the plasmid pJAF1. The SNF1:NEO allele results in thedeletion of the complete open read frame of SNF1 (2926 bp). The resulting PCR product(3854 bp) was used to transform strain H99 by biolistic transformation (Davidson et al.,2000). Transformants were grown on YPD plates containing neomycin and screened bycolony PCR with Extaq polymerase (Takara) using primer pairs SNF1–7/SNF1–8 andSNF1–9/hug-Neo. Primer SNF1–9 was designed from the region upstream of SNF1 andhug-Neo was designed for the NEO gene. Transformants in which WT allele was replacedwere confirmed by genomic hybridization as described (Hu and Kronstad, 2006). Onemutant designated SNF1–22 contained the deletion allele and was studied further. Forcomplementation of the deletion mutation, the SNF1 gene was amplified by PCR usingprimers SNF1-rec-for and SNF1-rec-rev, and genomic DNA from strain H99. The 4627 bpproduct was digested with XbaI and cloned into the XbaI site of pCH233, creating theplasmid pSNF1rec. The strain SNF1–22 was transformed with pSNF1rec by biolistictransformation, and transformants were selected on YPD containing nourseothricin (100 ug/ml). Reintroduction of SNF1 was confirmed by colony PCR and genomic hybridization.
Quantitation of gene expression during pulmonary infection or during growth in cultureTotal RNA from frozen cells collected from the lungs of infected mice was obtained asdescribed above (from an independent experiment), and DNA was removed by treatmentwith DNase I for 30 min at 25°C. Subseqently, cDNA was synthesized using randomhexamers and Superscript transcriptase II (Invitrogen Canada). The resulting cDNA wasused for real-time PCR with primers targeted to the 3′ regions of transcripts. Primers weredesigned using PrimerExpress v3 (Applied Biosystems). The Power SYBR Green PCR mix(Applied Biosystems) was used according to the manufacture’s recommendations. AnApplied Biosystems 7500 Fast Real-time PCR system was used to detect and quantify thePCR products with the following conditions: incubation at 95°C for 10 min followed by 40cycles of 95°C for 15 sec, and 60°C for 1 min. The cDNA of the ACT1 gene was used tonormalize the data. Dissociation analysis on all PCR reactions confirmed the amplificationof a single product for each primer pair and the absence of primer dimerization. Relativegene expression was quantified using SDS software 1.3.1 (Applied Biosystems).
Growth conditions for examining the influence of reduced glucose levels, acetate and loss ofSNF1 were as follows. Cells (WT, snf1 and snf1::SNF1) were grown in YNB+2% glucose at37 °C to mid log phase, and washed with the 37 °C pre-warmed medium. Equal numbers ofcells (1.5 × 108 cells) were transferred to either YNB+2% glucose or YNB + 0.2% glucoseor YNB + 2% acetate, and cultured for 5 hours at 37 °C prior to RNA isolation. Thesequences of the primers for the PCR analysis are given in Table S7, Supplementarymaterial.
Hu et al. Page 15
Mol Microbiol. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Stress and drug response assaysTo examine the response of C. neoformans WT, snf1 and snf1::SNF1 strains to variousstress conditions, exponentially growing cultures were washed, resuspended in H2O, andadjusted to a concentration of 0.2 × 105 cells/μl. The cell suspensions were diluted 10-foldserially, and 5 μl of each dilution was spotted onto YPD and/or YNB plates supplementedwith different chemicals. Plates were incubated for two to five days (depending on theconditions) at 30°C or 37°C, and photographed. The responses of strains to oxidative,nitrosative, osmotic stress and to agents that challenged cell wall integrity were examined.The specific assays were performed on YPD and/or YNB plates supplemented with orwithout 1.2 M KCl, 1.0 M or 1.5 M NaCl, 75 mM LiCl, 0.1% SDS, 0.5 mg/ml Congo Red, 3μg/ml menadione, 0.5 mM H2O2 and 2 mM, 4 mM or 8 mM sodium nitrite (NaNO2). Forcarbon source utilization experiments, YNB agar (yeast nitrogen base, 6.7% g/liter) wassupplemented with one of the following carbon sources: 0.5% or 2% glucose, 0.5 or 2%sucrose, 2% raffinose, 2% maltose, 2% galactose, 0.5% or 2% sodium acetate, 0.5% or 2%ethanol, 0.5% or 2% glycerol, 0.5% or 2% lactic acid. Sensitivity to inhibitors of calcineurinand TOR (target of rapamycin) signalling were examined by spotting the cell dilutions onYNB plates containing 2 μg/ml or 4 μg/ml FK506, 125 μg/ml or 250 μg/ml Cyclosporin(CsA), or 1 μg/ml or 10 μg/ml rapamycin. The antifungal drugs amphotericin B (0.5 μg/ml)and fluconazole (0.5 μg/ml or 1 μg/ml) were also tested. The figures show only theconditions where the mutant gave a different response compared to the WT strain.
Capsule formation and melanin productionLIM (low-iron medium) was used to examine capsule formation. A single colony from aYPD plate for each strain was cultured overnight at 30°C in liquid YPD medium. Cells wereharvested and diluted in low iron water, and 106 cells were added into 3 ml of LIM forfurther incubation at 30°C for 48 h. After incubation, the capsule was stained by India inkand examined by differential interference microscopy (DIC). To examine melaninproduction, a single colony of each strain was incubated overnight at 30°C in liquid YPDmedium, washed and diluted to 2×104 cells per ml. Five μl of serial dilutions from this stockwere spotted onto L-DOPA plates containing 0.1% glucose. The plates were incubated forthree days at 30°C or 37°C, and melanin production was monitored and photographed daily.
Virulence assaysFor virulence assays, female A/Jcr mice (4 to 6 weeks old) were obtained from JacksonLaboratories (Bar Harbor, Maine). The C. neoformans cells for inoculation were grown inYPD medium overnight at 30°C, washed in PBS and resuspended at 1.0×106 cells/ml inPBS. Inoculation was by intranasal instillation with 50 ul of cell suspension (5.0×104). Thestatus of the mice was monitored twice per day post-inoculation. Differences in virulencewere statistically assessed with the Krustal-Wallis test. For histopathology, infected micewere euthanized by CO2 inhalation, and organs were excised and placed in 10% bufferedformalin. Fixed organs were sent to Wax-It Histology Services (Vancouver, B.C. Canada)for sectioning and staining with Mayer’s Mucicarmine. For determination of the fungal loadin organs, infected mice were euthanized by CO2 inhalation and organs were excised,weighed and homogenized in 1mL PBS using a MixerMill (Retsch). Serial dilutions of thehomogenates were plated on Sabouraud dextrose agar plates containing 35 μg/mLchloramphenicol and colony-forming units were counted after an incubation for 48 h at30°C.
Supplementary MaterialRefer to Web version on PubMed Central for supplementary material.
Hu et al. Page 16
Mol Microbiol. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

AcknowledgmentsWe thank Dr. Marian Carlson for providing yeast strains, Dr. Philip Hieter for providing plasmid pBPH618, and Dr.Joseph Heitman for providing plasmids pJAF1 and pCH233. Dr. Gary Cox provided excellent advice onperforming the pulmonary infections. We also thank the staff of Canada’s Michael Smith Genome Sciences Centrefor SAGE advice and sequencing. This work was supported by the National Institute of Allergy and InfectiousDisease (R01 AI053721), the Canadian Institutes of Health Research and the British Columbia Lung Association.J.W.K. is a Burroughs Wellcome Fund Scholar in Molecular Pathogenic Mycology.
ReferencesAlepuz PM, Cunningham KW, Estruch F. Glucose repression affects ion homeostasis in yeast through
the regulation of the stress-activated ENA1 gene. Mol Microbiol. 1997; 26:91–98. [PubMed:9383192]
Audic S, Claverie JM. The significance of digital gene expression profiles. Genome Res. 1997; 7:986–995. [PubMed: 9331369]
Barelle CJ, Priest CL, Maccallum DM, Gow NA, Odds FC, Brown AJ. Niche-specific regulation ofcentral metabolic pathways in a fungal pathogen. Cell Microbiol. 2006; 8:961–971. [PubMed:16681837]
Basse CW, Lottspeich F, Steglich W, Kahmann R. Two potential indole-3-acetaldehydedehydrogenases in the phytopathogenic fungus Ustilago maydis. Eur J Biochem. 1996; 242:648–656. [PubMed: 9022693]
Bubb WA, Wright LC, Cagney M, Santangelo RT, Sorrell TC, Kuchel PW. Heteronuclear NMRstudies of metabolites produced by Cryptococcus neoformans in culture media: identification ofpossible virulence factors. Magn Reson Med. 1999; 42:442–453. [PubMed: 10467288]
Casadevall, A.; Perfect, JR. Cryptococcus neoformans. American Society for Microbiology Press;Washington, D.C: 1998.
Celenza JL, Carlson M. Cloning and genetic mapping of SNF1, a gene required for expression ofglucose-repressible genes in Saccharomyces cerevisiae. Mol Cell Biol. 1984; 4:49–53. [PubMed:6366512]
Celenza JL, Carlson M. A yeast gene that is essential for release from glucose repression encodes aprotein kinase. Science. 1986; 233:1175–1180. [PubMed: 3526554]
Chaturvedi V, Wong B, Newman SL. Oxidative killing of Cryptococcus neoformans by humanneutrophils. Evidence that fungal mannitol protects by scavenging reactive oxygen intermediates. JImmunol. 1996; 156:3836–3840. [PubMed: 8621921]
Chow ED, Liu OW, O’Brien S, Madhani HD. Exploration of whole-genome responses of the humanAIDS-associated yeast pathogen Cryptococcus neoformans var grubii: nitric oxide stress and bodytemperature. Curr Genet. 2007; 52:137–148. [PubMed: 17661046]
Cox GM, Harrison TS, McDade HC, Taborda CP, Heinrich G, Casadevall A, Perfect JR. Superoxidedismutase influences the virulence of Cryptococcus neoformans by affecting growth withinmacrophages. Infect Immun. 2003; 71:173–180. [PubMed: 12496163]
Davidson RC, Blankenship JR, Kraus PR, de Jesus Berrios M, Hull CM, D’Souza C, Wang P,Heitman J. A PCR-based strategy to generate integrative targeting alleles with large regions ofhomology. Microbiol. 2002; 148:2607–2615.
Davidson RC, Cruz MC, Sia RA, Allen B, Alspaugh JA, Heitman J. Gene disruption by biolistictransformation in serotype D strains of Cryptococcus neoformans. Fungal Genet Biol. 2000;29:38–48. [PubMed: 10779398]
de Jesus-Berrios M, Liu L, Nussbaum JC, Cox GM, Stamler JS, Heitman J. Enzymes that counteractnitrosative stress promote fungal virulence. Curr Biol. 2003; 13:1963–1968. [PubMed: 14614821]
De Virgilio C, Burckert N, Barth G, Neuhaus JM, Boller T, Wiemken A. Cloning and disruption of agene required for growth on acetate but not on ethanol: the acetyl-coenzyme A synthetase gene ofSaccharomyces cerevisiae. Yeast. 1992; 8:1043–1051. [PubMed: 1363452]
Ellerbroek PM, Lefeber DJ, van Veghel R, Scharringa J, Brouwer E, Gerwig GJ, Janbon G,Hoepelman AI, Coenjaerts FE. O-acetylation of cryptococcal capsular glucuronoxylomannan is
Hu et al. Page 17
Mol Microbiol. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

essential for interference with neutrophil migration. J Immunol. 2004; 173:7513–7520. [PubMed:15585878]
Ewing B, Green P. Base-calling of automated sequencer traces using Phred. II. Error probabilities.Genome Res. 1998; 8:186–194. [PubMed: 9521922]
Fan W, Kraus PR, Boily MJ, Heitman J. Cryptococcus neoformans gene expression during murinemacrophage infection. Euk Cell. 2005; 4:1420–1433.
Feldmesser M, Kress Y, Novikoff P, Casadevall A. Cryptococcus neoformans is a facultativeintracellular pathogen in murine pulmonary infection. Infect Immun. 2000; 68:4225–4237.[PubMed: 10858240]
Feldmesser M, Tucker S, Casadevall A. Intracellular parasitism of macrophages by Cryptococcusneoformans. Trends Microbiol. 2001; 9:273–278. [PubMed: 11390242]
Fox DS, Cruz MC, Sia RA, Ke H, Cox GM, Cardenas ME, Heitman J. Calcineurin regulatory subunitis essential for virulence and mediates interactions with FKBP12-FK506 in Cryptococcusneoformans. Mol Microbiol. 2001; 39:835–849. [PubMed: 11251806]
Fox DS, Heitman J. Calcineurin-binding protein Cbp1 directs the specificity of calcineurin-dependenthyphal elongation during mating in Cryptococcus neoformans. Euk Cell. 2005; 4:1526–1538.
Fradin C, Kretschmar M, Nichterlein T, Gaillardin C, d’Enfert C, Hube B. Stage-specific geneexpression of Candida albicans in human blood. Mol Microbiol. 2003; 47:1523–1543. [PubMed:12622810]
Friedman DB, Stauff DL, Pishchany G, Whitwell CW, Torres VJ, Skaar EP. Staphylococcus aureusredirects central metabolism to increase iron availability. PLoS Pathog. 2006; 2:e87. [PubMed:16933993]
Garcia-Rivera J, Tucker SC, Feldmesser M, Williamson PR, Casadevall A. Laccase expression inmurine pulmonary Cryptococcus neoformans infection. Infect Immun. 2005; 73:3124–3127.[PubMed: 15845520]
Giots F, Donaton MC, Thevelein JM. Inorganic phosphate is sensed by specific phosphate carriers andacts in concert with glucose as a nutrient signal for activation of the protein kinase A pathway inthe yeast Saccharomyces cerevisiae. Mol. Microbiol. 2003; 47:1163–1181.
Goldman DL, Khine H, Abadi J, Lindenberg DJ, Pirofski LA, Niang R, Casadevall A. Serologicevidence for Cryptococcus neoformans infection in early childhood. Pediatrics. 2001; 107:E66.[PubMed: 11331716]
Gordon D, Abajian C, Green P. Consed: a graphical tool for sequence finishing. Genome Res. 1998;8:195–202. [PubMed: 9521923]
Hardie DG, Carling D, Carlson M. The AMP-activated/SNF1 protein kinase subfamily: metabolicsensors of the eukaryotic cell? Annu Rev Biochem. 1998; 67:821–855. [PubMed: 9759505]
Himmelreich U, Allen C, Dowd S, Malik R, Shehan BP, Mountford C, Sorrell TC. Identification ofmetabolites of importance in the pathogenesis of pulmonary cryptococcoma using nuclearmagnetic resonance spectroscopy. Microb Infect. 2003; 5:285–290.
Hong SP, Carlson M. Regulation of snf1 protein kinase in response to environmental stress. J BiolChem. 2007; 282:16838–16845. [PubMed: 17438333]
Hu G, Kronstad JW. Gene disruption in Cryptococcus neoformans and Cryptococcus gattii by in vitrotransposition. Curr Genet. 2006; 49:341–350. [PubMed: 16397763]
Hu G, Steen BR, Lian T, Sham AP, Tam N, Tangen KL, Kronstad JW. Transcriptional regulation byprotein kinase A in Cryptococcus neoformans. PLoS Path. 2007; 3:e42.
Idnurm A, Giles SS, Perfect JR, Heitman J. Peroxisome function regulates growth on glucose in thebasidiomycete fungus Cryptococcus neoformans. Euk Cell. 2007; 6:60–72.
Idnurm A, Howlett BJ. Isocitrate lyase is essential for pathogenicity of the fungus Leptosphaeriamaculans to canola (Brassica napus). Euk Cell. 2002; 1:719–724.
Janbon G, Himmelreich U, Moyrand F, Improvisi L, Dromer F. Cas1p is a membrane proteinnecessary for the O-acetylation of the Cryptococcus neoformans capsular polysaccharide. MolMicrobiol. 2001; 42:453–467. [PubMed: 11703667]
Jung WH, Sham AP, Lian TS, Singh A, Kosman D, Kronstad JW. Iron source preference andregulation of iron uptake in the AIDS-associated pathogen Cryptococcus neoformans. PLoS Path.2008; 4:e45.
Hu et al. Page 18
Mol Microbiol. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Jung WH, Sham A, White R, Kronstad JW. Iron regulation of the major virulence factors in the AIDS-associated pathogen Cryptococcus neoformans. PLoS Biol. 2006; 4:e410. [PubMed: 17121456]
Kemp BE, Mitchelhill KI, Stapleton D, Michell BJ, Chen ZP, Witters LA. Dealing with energydemand: the AMP-activated protein kinase. Trends Biochem Sci. 1999; 24:22–25. [PubMed:10087918]
Kemp BE, Stapleton D, Campbell DJ, Chen ZP, Murthy S, Walter M, Gupta A, Adams JJ, Katsis F,van Denderen B, Jennings IG, Iseli T, Michell BJ, Witters LA. AMP-activated protein kinase,super metabolic regulator. Biochem Soc Trans. 2003; 31:162–168. [PubMed: 12546677]
Kratzer S, Schüller HJ. Carbon source-dependent regulation of the acetyl-coenzyme A synthetase-encoding gene ACS1 from Saccharomyces cerevisiae. Gene. 1995; 161:75–79. [PubMed:7642141]
Kwon-Chung KJ, Sorrell TC, Dromer F, Fung E, Levitz SM. Cryptococcosis: clinical and biologicalaspects. Med Mycol. 2000; 38:205–213. [PubMed: 11204147]
Lian T, Simmer MI, D’Souza CA, Steen BR, Zuyderduyn SD, Jones SJ, Marra MA, Kronstad JW.Iron-regulated transcription and capsule formation in the fungal pathogen Cryptococcusneoformans. Mol Microbiol. 2005; 55:1452–1472. [PubMed: 15720553]
Lin X, Heitman J. The biology of the Cryptococcus neoformans species complex. Annu RevMicrobiol. 2006; 60:69–105. [PubMed: 16704346]
Lorenz MC, Bender JA, Fink GR. Transcriptional response of Candida albicans upon internalizationby macrophages. Euk Cell. 2004; 3:1076–1087.
Lorenz MC, Fink GR. The glyoxylate cycle is required for fungal virulence. Nature. 2001; 412:83–86.[PubMed: 11452311]
Missall TA, Cherry-Harris JF, Lodge JK. Two glutathione peroxidases in the fungal pathogenCryptococcus neoformans are expressed in the presence of specific substrates. Microbiol. 2005;151:2573–2581.
Missall TA, Pusateri ME, Lodge JK. Thiol peroxidase is critical for virulence and resistance to nitricoxide and peroxide in the fungal pathogen, Cryptococcus neoformans. Mol Microbiol. 2004;51:1447–1458. [PubMed: 14982637]
Narasipura SD, Ault JG, Behr MJ, Chaturvedi V, Chaturvedi S. Characterization of Cu,Zn superoxidedismutase (SOD1) gene knock-out mutant of Cryptococcus neoformans var. gattii: role in biologyand virulence. Mol Microbiol. 2003; 47:1681–1694. [PubMed: 12622821]
Narasipura SD, Chaturvedi V, Chaturvedi S. Characterization of Cryptococcus neoformans varietygattii SOD2 reveals distinct roles of the two superoxide dismutases in fungal biology andvirulence. Mol Microbiol. 2005; 55:1782–1800. [PubMed: 15752200]
Noverr MC, Williamson PR, Fajardo RS, Huffnagle GB. CNLAC1 is required for extrapulmonarydissemination of Cryptococcus neoformans but not pulmonary persistence. Infect Immun. 2004;72:1693–1699. [PubMed: 14977977]
Olkkonen VM, Johansson M, Suchanek M, Yan D, Hynynen R, Ehnholm C, Jauhiainen M, Thiele C,Lehto M. The OSBP-related proteins (ORPs): global sterol sensors for co-ordination of cellularlipid metabolism, membrane trafficking and signalling processes? Biochem Soc Trans. 2006;34:389–391. [PubMed: 16709169]
Ospina-Giraldo MD, Mullins E, Kang S. Loss of function of the Fusarium oxysporum SNF1 genereduces virulence on cabbage and Arabidopsis. Curr Genet. 2003; 44:49–57. [PubMed: 12845476]
Olivas I, Royuela M, Romero B, Monteiro MC, Mínguez JM, Laborda F, De Lucas JR. Ability to growon lipids accounts for the fully virulent phenotype in neutropenic mice of Aspergillus fumigatusnull mutants in the key glyoxylate cycle enzymes. Fungal Genet Biol. 2008; 45:45–60. [PubMed:17616408]
Palmieri L, Lasorsa FM, De Palma A, Palmieri F, Runswick MJ, Walker JE. Identification of the yeastACR1 gene product as a succinate-fumarate transporter essential for growth on ethanol or acetate.FEBS Lett. 1997; 417:114–118. [PubMed: 9395087]
Panepinto J, Liu L, Ramos J, Zhu X, Valyi-Nagy T, Eksi S, Fu J, Jaffe HA, Wickes B, Williamson PR.The DEAD-box RNA helicase Vad1 regulates multiple virulence-associated genes inCryptococcus neoformans. J Clin Invest. 2005; 115:632–641. [PubMed: 15765146]
Hu et al. Page 19
Mol Microbiol. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Parks LW, Casey WM. Physiological implications of sterol biosynthesis in yeast. Ann Rev Microbiol.1995; 49:95–116. [PubMed: 8561481]
Perfect JR, Lang SD, Durack DT. Chronic cryptococcal meningitis: a new experimental model inrabbits. Am J Pathol. 1980; 101:177–194. [PubMed: 7004196]
Petter R, Chang YC, Kwon-Chung KJ. A gene homologous to Saccharomyces cerevisiae SNF1appears to be essential for the viability of Candida albicans. Infect Immun. 1997; 65:4909–4917.[PubMed: 9393775]
Petzold EW, Himmelreich U, Mylonakis E, Rude T, Toffaletti D, Cox GM, Miller JL, Perfect JR.Characterization and regulation of the trehalose synthesis pathway and its importance in thepathogenicity of Cryptococcus neoformans. Infect Immun. 2006; 74:5877–5887. [PubMed:16988267]
Philips BJ, Meguer JX, Redman J, Baker EH. Factors determining the appearance of glucose in upperand lower respiratory tract secretions. Intensive Care Med. 2003; 29:2204–2210. [PubMed:14647890]
Philips BJ, Redman J, Brennan A, Wood D, Holliman R, Baines D, Baker EH. Glucose in bronchialaspirates increases the risk of respiratory MRSA in intubated patients. Thorax. 2005; 60:761–764.[PubMed: 16135681]
Piekarska K, Mol E, van den Berg M, Hardy G, van den Burg J, van Roermund C, MacCallum D,Odds F, Distel B. Peroxisomal fatty acid beta-oxidation is not essential for virulence of Candidaalbicans. Euk Cell. 2006; 5:1847–1856.
Portillo F, Mulet JM, Serrano R. A role for the non-phosphorylated form of yeast Snf1: tolerance totoxic cations and activation of potassium transport. FEBS letters. 2005; 579:512–516. [PubMed:15642368]
Prigneau O, Porta A, Poudrier JA, Colonna-Romano S, Noel T, Maresca B. Genes involved in beta-oxidation, energy metabolism and glyoxylate cycle are induced by Candida albicans duringmacrophage infection. Yeast. 2003; 20:723–730. [PubMed: 12794933]
Pukkila-Worley R, Gerrald QD, Kraus PR, Boily MJ, Davis MJ, Giles SS, Cox GM, Heitman J,Alspaugh JA. Transcriptional network of multiple capsule and melanin genes governed by theCryptococcus neoformans cyclic AMP cascade. Euk Cell. 2005; 4:190–201.
Ramirez MA, Lorenz MC. Mutations in alternative carbon utilization pathways in Candida albicansattenuate virulence and confer pleiotropic phenotypes. Euk Cell. 2007; 6:280–290.
Ren P, Rossettini A, Chaturvedi V, Hanes SD. The Ess1 prolyl isomerase is dispensable for growth butrequired for virulence in Cryptococcus neoformans. Microbiol. 2005; 151:1593–1605.
Rodrigues ML, Alvarez M, Fonseca FL, Casadevall A. Binding of the wheat germ lectin toCryptococcus neoformans suggests an association of chitin-like structures with yeast budding andcapsular glucuronoxylomannan. Euk Cell. 2008; 7:602–609.
Rooney SA, Young SL, Mendelson CR. Molecular and cellular processing of lung surfactant. FASEBJ. 1994; 8:957–967. [PubMed: 8088461]
Rose RM, Catalano PJ, Koziel H, Furlong ST. Abnormal lipid composition of bronchoalveolar lavagefluid obtained from individuals with AIDS-related lung disease. Am J Respir Crit Care Med. 1994;149:332–338. [PubMed: 8306026]
Rubin-Bejerano I, Fraser I, Grisafi P, Fink GR. Phagocytosis by neutrophils induces an amino aciddeprivation response in Saccharomyces cerevisiae and Candida albicans. Proc Natl Acad SciUSA. 2003; 100:11007–11012. [PubMed: 12958213]
Rude TH, Toffaletti DL, Cox GM, Perfect JR. Relationship of the glyoxylate pathway to thepathogenesis of Cryptococcus neoformans. Infect Immun. 2002; 70:5684–5694. [PubMed:12228298]
Schöbel F, Ibrahim-Granet O, Avé P, Latgé JP, Brakhage AA, Brock M. Aspergillus fumigatus doesnot require fatty acid metabolism via isocitrate lyase for development of invasive aspergillosis.Infect Immun. 2007; 75:1237–1244. [PubMed: 17178786]
Schüller HJ. Transcriptional control of nonfermentative metabolism in the yeast Saccharomycescerevisiae. Curr Genet. 2003; 43:139–1360. [PubMed: 12715202]
Solomon PS, Lee RC, Wilson TJ, Oliver RP. Pathogenicity of Stagonospora nodorum requires malatesynthase. Mol Microbiol. 2004; 53:1065–1073. [PubMed: 15306011]
Hu et al. Page 20
Mol Microbiol. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Steen BR, Lian T, Zuyderduyn S, MacDonald WK, Marra M, Jones SJ, Kronstad JW. Temperature-regulated transcription in the pathogenic fungus Cryptococcus neoformans. Genome Res. 2002;12:1386–1400. [PubMed: 12213776]
Steen BR, Zuyderduyn S, Toffaletti DL, Marra M, Jones SJ, Perfect JR, Kronstad J. Cryptococcusneoformans gene expression during experimental cryptococcal meningitis. Euk Cell. 2003;2:1336–1349.
Thewes S, Kretschmar M, Park H, Schaller M, Filler SG, Hube B. In vivo and ex vivo comparativetranscriptional profiling of invasive and non-invasive Candida albicans isolates identifies genesassociated with tissue invasion. Mol Microbiol. 2007; 63:1606–1628. [PubMed: 17367383]
Thompson-Jaeger S, Francois J, Gaughran JP, Tatchell K. Deletion of SNF1 affects the nutrientresponse of yeast and resembles mutations which activate the adenylate cyclase pathway.Genetics. 1991; 129:697–706. [PubMed: 1752415]
Tonukari NJ, Scott-Craig JS, Walton JD. The Cochliobolus carbonum SNF1 gene is required for cellwall-degrading enzyme expression and virulence on maize. Plant Cell. 2000; 12:237–248.[PubMed: 10662860]
Velculescu VE, Zhang L, Vogelstein B, Kinzler KW. Serial analysis of gene expression. Science.1995; 270:484–487. [PubMed: 7570003]
Vyas VK, Kuchin S, Carlson M. Interaction of the repressors Nrg1 and Nrg2 with the Snf1 proteinkinase in Saccharomyces cerevisiae. Genetics. 2001; 158:563–572. [PubMed: 11404322]
Wang P, Cox GM, Heitman J. A Sch9 protein kinase homologue controlling virulence independentlyof the cAMP pathway in Cryptococcus neoformans. Curr Genet. 2004; 46:247–255. [PubMed:15503029]
Wang ZY, Thornton CR, Kershaw MJ, Debao L, Talbot NJ. The glyoxylate cycle is required fortemporal regulation of virulence by the plant pathogenic fungus Magnaporthe grisea. MolMicrobiol. 2003; 47:1601–1612. [PubMed: 12622815]
Waterman SR, Hacham M, Hu G, Zhu X, Park YD, Shin S, Panepinto J, Valyi-Nagy T, Beam C,Husain S, Singh N, Williamson PR. Role of a CUF1/CTR4 copper regulatory axis in the virulenceof Cryptococcus neoformans. J Clin Invest. 2007; 117:794–802. [PubMed: 17290306]
Wright L, Bubb W, Davidson J, Santangelo R, Krockenberger M, Himmelreich U, Sorrell T.Metabolites released by Cryptococcus neoformans var. neoformans and var. gattii differentiallyaffect human neutrophil function. Microbes Infect. 2002; 4:1427–1438. [PubMed: 12475633]
Xue C, Bahn YS, Cox GM, Heitman J. G protein-coupled receptor Gpr4 senses amino acids andactivates the cAMP-PKA pathway in Cryptococcus neoformans. Mol Biol Cell. 2006; 17:667–679.[PubMed: 16291861]
Ye T, Garcia-Salcedo R, Ramos J, Hohmann S. Gis4, a new component of the ion homeostasis systemin the yeast Saccharomyces cerevisiae. Euk Cell. 2006; 5:1611–1621.
Yin Z, Smith RJ, Brown AJ. Multiple signalling pathways trigger the exquisite sensitivity of yeastgluconeogenic mRNAs to glucose. Mol Microbiol. 1996; 20:751–764. [PubMed: 8793872]
Young ET, Dombek KM, Tachibana C, Ideker T. Multiple pathways are co-regulated by the proteinkinase Snf1 and the transcription factors Adr1 and Cat8. J Biol Chem. 2003; 278:26146–26158.[PubMed: 12676948]
Yu JH, Hamari Z, Han KH, Seo JA, Reyes-Dominguez Y, Scazzocchio C. Double-joint PCR: a PCR-based molecular tool for gene manipulations in filamentous fungi. Fungal Genet Biol. 2004;41:973–981. [PubMed: 15465386]
Zhu X, Williamson PR. Role of laccase in the biology and virulence of Cryptococcus neoformans.FEMS Yeast Res. 2004; 5:1–10. [PubMed: 15381117]
Hu et al. Page 21
Mol Microbiol. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 1. The position of acetyl-CoA in central carbon metabolismMetabolic processes and metabolites are indicated in upper and lower case letters,respectively. Note that neither the cellular compartments for the various reactions nor theimport processes for metabolites such as ethanol and acetate are indicated.
Hu et al. Page 22
Mol Microbiol. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 2. Analysis of ACS1 expression during pulmonary infection and the role of the gene forgrowth on alternative carbon sources and virulence(A). Quantitative real time PCR to confirm the elevated expression of the ACS1 gene duringpulmonary infection relative to growth in vitro. (B) Growth of the WT strain H99, the acs1mutant and the complemented strain on the carbon sources indicated above the panels. (C)Virulence of the acs1 mutant compared with the WT strain H99 and the complementedstrain. The strains were each inoculated into 10 mice by inhalation and the mice weremonitored for signs of illness. The virulence of the acs1 mutant was statistically differentfrom that of the WT (P = 0.0001) or the complemented strain (P = 0.0008) by the Krustal-Wallis test. A representative graph is shown from a total of three experiments, and the acs1mutant displayed a similar attenuation of virulence in each experiment.
Hu et al. Page 23
Mol Microbiol. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 3. Growth of the C. neoformans snf1 mutant on alternative carbon sourcesCells of the WT strain (H99), the snf1 mutant and the complemented strain were spotted atdecreasing concentrations (from left to right) on the media indicated on the left. YPD andYNB serve as controls for growth on glucose, and YNB with sucrose, sodium acetate orethanol were used to test alternative carbon sources. The plates were incubated for two daysat 30°C or 37°C as indicated. Reintroduction of the SNF1 gene into the snf1 mutant restoredgrowth to the level of the WT strain.
Hu et al. Page 24
Mol Microbiol. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 4. Growth and melanin production by the C. neoformans snf1 mutantCells of the WT strain H99, the snf1 mutant and the complemented strain were spotinoculated in dilutions from left to right on YNB medium to test the response to salt (1.5MNaCl), nitrosative stress (8 mM NaNO2), amphotericin B (0.5 μg/ml), and rapamycin (10μg/ml). The plates were incubated at 30°C or 37°C for two days. Melanin production wastested on L-DOPA medium after three days of incubation.
Hu et al. Page 25
Mol Microbiol. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 5. Influence of glucose, acetate and deletion of SNF1 on the expression of genes identifiedby SAGE analysis(A) Expression of selected genes as measured by quantitative real-time PCR followinggrowth under the conditions indicated in the legend on the right. (B) Expression of the SNF1gene in cells grown in YNB medium with the carbon sources indicated on the X axis. (C)Expression of the genes encoding laccase or genes involved in the response to stressfollowing growth under the conditions indicated on the right. The transcript of the ACT1gene was used for normalization.
Hu et al. Page 26
Mol Microbiol. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 6.Analysis of virulence in a mouse inhalation model of cryptococcosis. (A) Ten A/Jcr micewere inoculated with the WT strain H99, the snf1 mutant or the complemented strain andmonitored for signs of illness. The virulence of the snf1 mutant was significantly differentfrom that of the WT strain (P < 0.0001) and the complemented strain (P < 0.0001) by theKrustal-Wallis test.
Hu et al. Page 27
Mol Microbiol. Author manuscript; available in PMC 2009 September 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
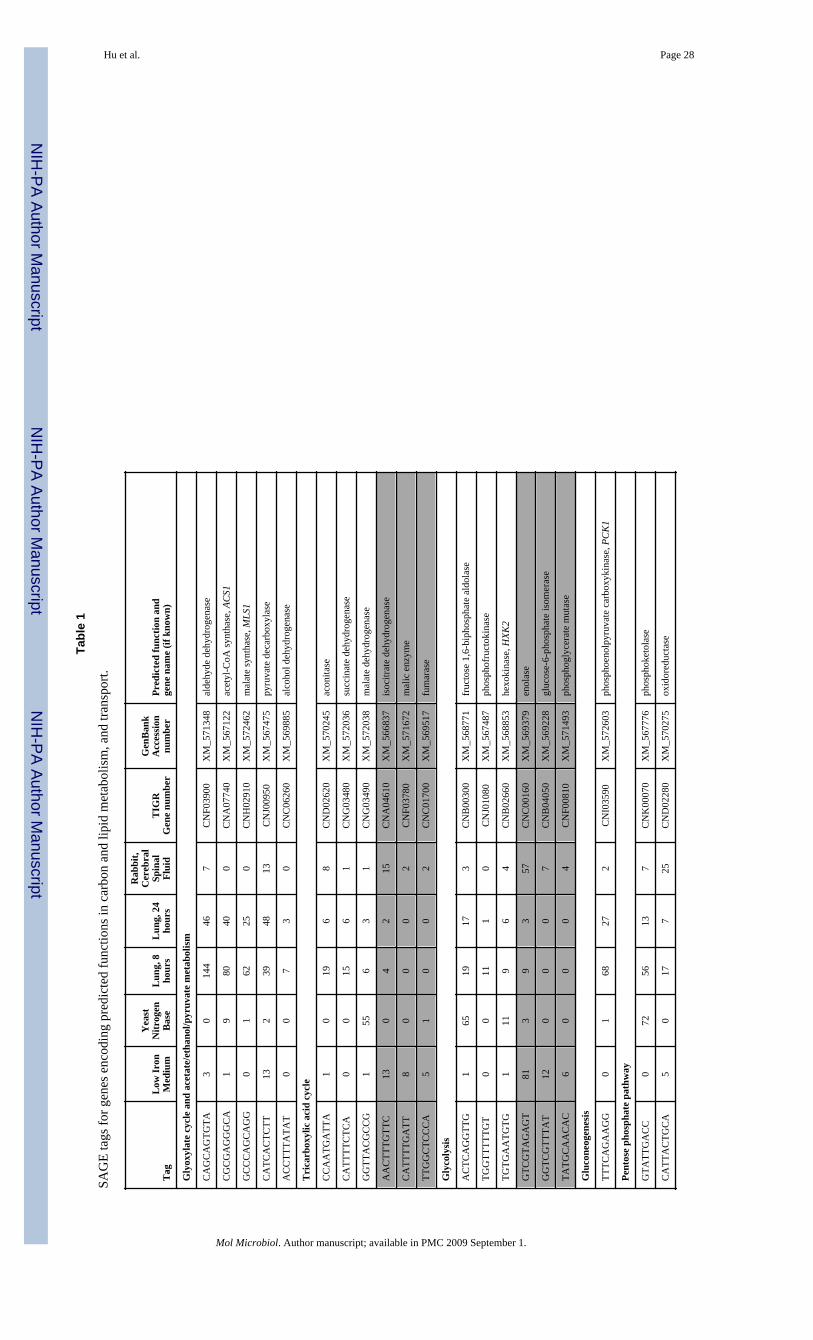
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Hu et al. Page 28
Tabl
e 1
SAG
E ta
gs fo
r gen
es e
ncod
ing
pred
icte
d fu
nctio
ns in
car
bon
and
lipid
met
abol
ism
, and
tran
spor
t.
Tag
Low
Iron
Med
ium
Yea
stN
itrog
enB
ase
Lun
g, 8
hour
sL
ung,
24
hour
s
Rab
bit,
Cer
ebra
lSp
inal
Flui
dT
IGR
Gen
e nu
mbe
r
Gen
Ban
kA
cces
sion
num
ber
Pred
icte
d fu
nctio
n an
dge
ne n
ame
(if k
now
n)
Gly
oxyl
ate
cycl
e an
d ac
etat
e/et
hano
l/pyr
uvat
e m
etab
olis
m
CA
GC
AG
TGTA
30
144
467
CN
F039
00X
M_5
7134
8al
dehy
de d
ehyd
roge
nase
CG
CG
AG
GG
CA
19
8040
0C
NA
0774
0X
M_5
6712
2ac
etyl
-CoA
synt
hase
, AC
S1
GC
CC
AG
CA
GG
01
6225
0C
NH
0291
0X
M_5
7246
2m
alat
e sy
ntha
se, M
LS1
CA
TCA
CTC
TT13
239
4813
CN
J009
50X
M_5
6747
5py
ruva
te d
ecar
boxy
lase
AC
CTT
TATA
T0
07
30
CN
C06
260
XM
_569
885
alco
hol d
ehyd
roge
nase
Tri
carb
oxyl
ic a
cid
cycl
e
CC
AA
TGA
TTA
10
196
8C
ND
0262
0X
M_5
7024
5ac
onita
se
CA
TTTT
CTC
A0
015
61
CN
G03
480
XM
_572
036
succ
inat
e de
hydr
ogen
ase
GG
TTA
CG
CC
G1
556
31
CN
G03
490
XM
_572
038
mal
ate
dehy
drog
enas
e
AA
CTT
TGTT
C13
04
215
CN
A04
610
XM
_566
837
isoc
itrat
e de
hydr
ogen
ase
CA
TTTT
GA
TT8
00
02
CN
F037
80X
M_5
7167
2m
alic
enz
yme
TTG
GC
TCC
CA
51
00
2C
NC
0170
0X
M_5
6951
7fu
mar
ase
Gly
coly
sis
AC
TCA
GG
TTG
165
1917
3C
NB
0030
0X
M_5
6877
1fr
ucto
se 1
,6-b
ipho
spha
te a
ldol
ase
TGG
TTTT
TGT
00
111
0C
NJ0
1080
XM
_567
487
phos
phof
ruct
okin
ase
TGTG
AA
TGTG
111
96
4C
NB
0266
0X
M_5
6885
3he
xoki
nase
, HXK
2
GTC
GTA
GA
GT
813
93
57C
NC
0016
0X
M_5
6937
9en
olas
e
GG
TCG
TTTA
T12
00
07
CN
B04
050
XM
_569
228
gluc
ose-
6-ph
osph
ate
isom
eras
e
TATG
CA
AC
AC
60
00
4C
NF0
0810
XM
_571
493
phos
phog
lyce
rate
mut
ase
Glu
cone
ogen
esis
TTTC
AG
AA
GG
01
6827
2C
NI0
3590
XM
_572
603
phos
phoe
nolp
yruv
ate
carb
oxyk
inas
e, P
CK
1
Pent
ose
phos
phat
e pa
thw
ay
GTA
TTG
AC
C0
7256
137
CN
K00
070
XM
_567
776
phos
phok
etol
ase
CA
TTA
CTG
CA
50
177
25C
ND
0228
0X
M_5
7027
5ox
idor
educ
tase
Mol Microbiol. Author manuscript; available in PMC 2009 September 1.

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Hu et al. Page 29
Tag
Low
Iron
Med
ium
Yea
stN
itrog
enB
ase
Lun
g, 8
hour
sL
ung,
24
hour
s
Rab
bit,
Cer
ebra
lSp
inal
Flui
dT
IGR
Gen
e nu
mbe
r
Gen
Ban
kA
cces
sion
num
ber
Pred
icte
d fu
nctio
n an
dge
ne n
ame
(if k
now
n)
TAG
TGTC
CC
G70
641
932
CN
K01
050
XM
_567
793
6-ph
osph
oglu
cona
te d
ehyd
roge
nase
AA
GTG
TGG
TA12
20
018
CN
K03
170
XM
_567
910
trans
aldo
lase
Oth
er fu
nctio
ns r
elat
ed to
car
bon
met
abol
ism
CA
TTA
TGA
CA
40
195
5C
NA
0110
0X
M_5
6708
9gl
ycer
ol 3
-pho
spha
te d
ehyd
roge
nase
ATT
CA
CG
CG
C1
017
53
CN
D02
900
XM
_570
206
glut
amin
e:fr
ucto
se-6
- pho
spha
te a
mid
otra
nsfe
rase
CC
ATA
TGTT
T3
1415
21
CN
F035
30X
M_5
7143
1gl
ycog
en p
hosp
hory
lase
- lik
e pr
otei
n
GA
CA
TTG
TTG
222
188
1C
NC
0534
0X
M_5
6972
9D
-ara
bini
tol d
ehyd
roge
nase
CA
AC
GG
GG
GG
350
71
11C
NH
0017
0X
M_5
7221
2ph
osph
oman
nom
utas
e
CTT
CC
TGTT
C23
19
620
CN
N00
430
XM
_568
570
phos
phog
luco
mut
ase
CA
TCC
CA
AA
G15
00
020
CN
C03
700
XM
_569
599
UD
P-gl
ucos
e py
roph
opho
ryla
se
TATA
AG
AC
CG
60
00
4C
NF0
3930
XM
_571
680
glyc
ogen
met
abol
ism
- rel
ated
pro
tein
Lip
id m
etab
olis
m
TAA
CC
ATA
TA19
096
177
CN
J029
70X
M_5
6757
6bu
tyry
lcho
lines
tera
se, t
riacy
lgly
cero
l lip
ase,
CG
L1
AA
CC
AC
AC
TA1
3439
1514
CN
N01
770
XM
_568
678
sim
ilar t
o B
enzo
diaz
epin
rece
ptor
TCA
TTTC
AC
T2
033
103
CN
I015
60X
M_5
7281
4st
erol
-bin
ding
pro
tein
GC
GA
AG
TAC
T0
027
30
CN
N01
550
XM
_568
632
acyl
-CoA
oxi
dase
GC
TGC
CTT
TG2
1322
61
CN
I002
40X
M_5
7273
0en
oyl-C
oA h
ydra
tase
/isom
eras
e
CA
ATG
GG
AA
T2
012
30
CN
I036
90X
M_5
7289
62,
4-di
enoy
l-CoA
redu
ctas
e (N
AD
PH)
TATG
TGA
ATC
30
93
4C
NM
0083
0X
M_5
6842
2ox
idor
educ
tase
, sho
rt ch
ain
dehy
drog
enas
e/re
duct
ase
CTA
CA
AA
TGA
10
91
1C
NH
0295
0X
M_5
7246
5ox
idor
educ
tase
, sho
rt ch
ain
dehy
drog
enas
e/re
duct
ase
GA
TAA
GG
TGT
40
86
8C
NI0
2880
XM
_572
653
oxys
tero
l-bin
ding
pro
tein
TATT
ATT
GTT
00
81
2C
NA
0332
0X
M_5
6665
8en
oyl-C
oA h
ydra
tase
AA
GC
AG
AG
GA
29
73
5C
NI0
0710
XM
_572
753
long
-cha
in-f
atty
-aci
d C
oA li
gase
TAG
TTTA
TAT
20
73
2C
NG
0268
0X
M_5
7193
1hy
drox
ymet
hylg
luta
ryl-C
oA sy
ntha
se
ATT
GA
ATG
TA0
07
225
CN
D01
120
XM
_570
351
fatty
aci
d be
ta-o
xida
tion-
rela
ted
prot
ein
Tra
nspo
rter
s
TATA
CG
ATT
T26
061
214
32
CN
B02
680
XM
_568
855
mon
osac
char
ide
trans
porte
r, H
XT1
CC
CA
TCG
TAT
820
136
7924
CN
M02
430
XM
_568
258
plas
ma
mem
bran
e iro
n pe
rmea
se, C
FT1
Mol Microbiol. Author manuscript; available in PMC 2009 September 1.

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Hu et al. Page 30
Tag
Low
Iron
Med
ium
Yea
stN
itrog
enB
ase
Lun
g, 8
hour
sL
ung,
24
hour
s
Rab
bit,
Cer
ebra
lSp
inal
Flui
dT
IGR
Gen
e nu
mbe
r
Gen
Ban
kA
cces
sion
num
ber
Pred
icte
d fu
nctio
n an
dge
ne n
ame
(if k
now
n)
CTT
GA
CA
GC
G0
542
460
CN
F029
60X
M_5
7137
1ac
etat
e tra
nspo
rter,
ADY2
TATC
TGA
TTT
191
348
10C
NG
0424
0X
M_5
7209
2he
xose
tran
spor
ter
TATT
GG
TAC
A16
028
130
CN
C03
960
XM
_569
629
phos
phat
e tra
nspo
rter
GTC
ATT
TATG
20
273
2C
NF0
2890
XM
_571
700
neut
ral a
min
o ac
id tr
ansp
orte
r
CG
GG
AA
TAC
G0
024
121
CN
H03
640
XM
_572
526
GPR
/FU
N34
fam
ily p
rote
in/a
ceta
te tr
ansp
orte
r, AT
O2
CTG
TTC
GG
CA
93
2113
11C
ND
0108
0X
M_5
7035
3hi
gh a
ffin
ity c
oppe
r tra
nspo
rter,
CTR
4
CA
CTG
TATG
C1
021
180
CN
E025
70X
M_5
7095
8su
ccin
ate:
fum
arat
e an
tipor
ter
GC
CG
GG
CTG
T1
1520
142
CN
C02
540
XM
_569
547
orga
nic
acid
tran
spor
ter
CC
GA
GA
AA
CG
20
197
2C
ND
0438
0X
M_5
7059
6m
onov
alen
t ino
rgan
ic c
atio
n tra
nspo
rter
AG
CA
CTT
ATG
10
162
0C
NI0
3490
XM
_572
882
mal
tose
per
mea
se, t
reha
lose
tran
spor
ter
CA
GTA
TCA
CT
20
152
2C
NL0
5010
XM
_568
040
gaba
per
mea
se
GTG
GTT
TTC
T1
011
92
CN
F047
60X
M_5
7144
0ne
utra
l am
ino
acid
per
mea
se
CA
TATA
TAA
C2
07
01
CN
H00
950
XM
_572
289
sper
min
e tra
nspo
rter
GG
AA
GA
GG
AA
10
63
2C
NE0
0440
XM
_571
108
Inne
r mem
bran
e so
lute
tran
spor
ter M
RS4
ATC
GG
TAC
CC
447
00
4C
NN
0103
0X
M_5
6854
4in
orga
nic
phos
phat
e tra
nspo
rter
Tags
are
list
ed b
y ab
unda
nce
base
d on
the
8 ho
ur li
brar
y; th
ese
tags
hav
e hi
gher
leve
ls in
this
libr
ary
rela
tive
to th
e LI
M li
brar
y. T
he re
mai
ning
tags
in th
e sh
aded
row
s are
list
ed b
y th
eir a
bund
ance
in th
e LI
M li
brar
y. T
he ta
g co
unts
wer
e no
rmal
ized
to 2
0,00
0 ta
gs fo
r eac
hlib
rary
.
Mol Microbiol. Author manuscript; available in PMC 2009 September 1.

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Hu et al. Page 31
Tabl
e 2
Viru
lenc
e-as
soci
ated
and
stre
ss-r
espo
nse
gene
s.
Tag
sL
ow Ir
on M
ediu
mY
east
Nitr
ogen
Bas
eL
ungs
, 8 h
ours
Lun
gs, 2
4 ho
urs
Rab
bit,
Cer
ebra
lSp
inal
Flui
dT
IGR
Gen
e nu
mbe
rG
enB
ank
Pred
icte
d Fu
nctio
n an
d ge
ne n
ame
(if k
now
n)
TATA
TGTG
TA35
024
760
262
CN
G04
220
XM
_572
088
heat
shoc
k pr
otei
n 12
CA
CG
TCC
AC
G27
6195
305
CN
D01
490
XM
_570
285
Cu/
Zn su
pero
xide
dis
mut
ase,
SO
D1
ATA
AG
CTT
TC12
074
1425
CN
G00
600
XM
_571
772
man
nito
l-1-p
hosp
hate
deh
ydro
gena
se
CA
TTTT
ATG
T14
035
1624
CN
M01
520
XM
_568
451
heat
shoc
k pr
otei
n 90
TAA
CA
TAA
TG16
030
317
CN
A01
260
XM
_567
249
glut
athi
one
S-tra
nsfe
rase
TGTT
TCTA
CA
21
243
3C
NG
0422
0X
M_5
7208
8he
at sh
ock
prot
ein
12
CTC
TTC
ATT
T8
423
1410
CN
A03
160
XM
_566
757
heat
-sho
ck p
rote
in, S
KS2
GC
GG
ATA
AA
A6
523
68
CN
B00
910
XM
_569
098
stre
ss re
spon
se R
CI p
eptid
e, c
atio
ntra
nspo
rt-re
late
d pr
otei
n
GC
TGC
CTC
TG0
421
321
CN
A01
500
XM
_566
622
alte
rnat
ive
oxid
ase
ATG
TTTT
ATT
90
207
3C
NG
0256
0X
M_5
7200
3U
DP-
gluc
uron
ic a
cid
deca
rbox
ylas
e,U
XS1
AG
ATG
GA
CG
A4
1919
84
CN
G01
060
XM
_571
865
cycl
ophi
lin-li
ke p
rote
in
AA
TTG
ATG
AG
00
1810
2C
NC
0716
0X
M_5
6984
4fla
vohe
mop
rote
in, F
HB1
TTG
ATT
TTTT
00
163
0C
NM
0186
0X
M_5
6829
4m
acro
lide-
bind
ing
prot
ein
FKB
P12
GC
GA
CA
GC
CA
01
152
1C
NC
0149
0X
M_5
6948
2pe
ptid
yl p
roly
l cis
-tran
s iso
mer
ase,
ESS1
AC
GG
AG
TGTA
10
149
2C
ND
0026
0X
M_5
7033
3FK
506-
resi
stan
t cal
cine
urin
Bre
gula
tory
subu
nit
TCG
ATG
GC
GA
20
115
14C
NE0
5040
XM
_571
033
glyo
xal o
xida
se p
recu
rsor
CA
AC
GA
TGA
T5
010
366
CN
F029
10X
M_5
7136
7LE
A d
omai
n pr
otei
n
GC
TTA
ATT
AT
01
82
1C
NN
0036
0X
M_5
6855
8pr
otei
n ki
nase
, SC
H9
GTC
TTC
ATC
A2
08
21
CN
D05
600
XM
_570
476
heat
shoc
k pr
otei
n 12
TTTC
TGA
AA
A2
07
11
CN
D05
940
XM
_570
487
caps
ule
asso
ciat
ed p
rote
in, C
AP1
TGTG
TAA
AA
G0
06
10
CN
E007
10X
M_5
7079
3m
anni
tol-1
-pho
spha
te d
ehyd
roge
nase
TCA
GA
AG
TTG
788
111
51C
NC
0420
0X
M_5
6966
7th
iore
doxi
n
TGTT
ATC
GG
T60
422
654
CN
M01
520
XM
_568
451
heat
shoc
k pr
otei
n 90
Mol Microbiol. Author manuscript; available in PMC 2009 September 1.

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Hu et al. Page 32
Tag
sL
ow Ir
on M
ediu
mY
east
Nitr
ogen
Bas
eL
ungs
, 8 h
ours
Lun
gs, 2
4 ho
urs
Rab
bit,
Cer
ebra
lSp
inal
Flui
dT
IGR
Gen
e nu
mbe
rG
enB
ank
Pred
icte
d Fu
nctio
n an
d ge
ne n
ame
(if k
now
n)
CA
TAA
TTG
GC
351
131
14C
NM
0207
0X
M_5
6828
3he
at sh
ock
prot
ein
Tags
are
list
ed b
y ab
unda
nce
base
d on
the
8 ho
ur li
brar
y an
d sh
ow h
ighe
r lev
els i
n th
is li
brar
y re
lativ
e to
the
LIM
libr
ary.
The
rem
aini
ng ta
gs in
the
shad
ed ro
ws a
re li
sted
by
thei
r abu
ndan
ce in
the
LIM
libra
ry. T
he ta
g co
unts
wer
e no
rmal
ized
to 2
0,00
0 ta
gs fo
r eac
h lib
rary
.
Mol Microbiol. Author manuscript; available in PMC 2009 September 1.





























