Embed Size (px)
Citation preview

FUNDAÇÃO UNIVERSIDADE DO RIO GRANDE
MODELO ECOLÓGICO DE PREDAÇÃO EM FUNDOS VEGETADOS NO
ESTUÁRIO DA LAGOA DOS PATOS (R.S.)
JOSÉ JULIO FERRAZ DE CAMPOS JR.
Trabalho de graduação
apresentado ao curso de
Oceanologia como requisito parcial
a conclusão de curso
ORIENTADOR: Prof. Dr. Milton L. Asmus
Rio Grande
Novembro 1993

i
Além de suas mais fantásticas criações,
existem mundos de interesse para todos aqueles que
observam a natureza, tentando imitá-la
B.B. Mandelbrot

ii
RESUMO
O presente trabalho apresenta um modelo ecológico baseado na relação presa-predador
proposta para as epífitas da gramínea marinha Ruppia maritima (Potamogeatomacea) e o
gastrópode herbívoro Heleobia sp (Hidrobiidae) ,em pradarias de áreas rasas do estuário da Lagoa
dos Patos, R.S.
O modelo (versão conceitual e matemática) foi construído com base em revisão
bibliográfica, e codificado em linguagem BASIC para computador IBM-PC. Na realização das
simulações foram utilizados parâmetros com valores teóricos e calibrados devido a escassez de
dados de campo sobre a relação estudada.
Foram realizadas um total de 10 simulações plotando-se a variação de biomassas em
gramas de presas e predadores pelo tempo em dias julianos. Destas simulações, oito foram
realizadas a partir de variações dos valores dos parâmetros utilizados na primeira simulação. A
simulação final utilizou os mesmos valores da primeira, plotando-se a biomassa de presas contra a
biomassa de predadores em um gráfico denominado diagrama de curva de plano de fase. Tal
procedimento permite a observação do comportamento cíclico da variação de suas biomassas.
Através das varias simulações sugere-se a interdependência entre as presas e os predadores,
assim como o efeito dos fatores ambientais, na variação de suas biomassas.

iii
ABSTRACT
This paper presents an ecological model for the proposed predator-prey relationship
between epyfites of the seagrass Ruppia maritima (Potamogeatomacea) and the herbivorous
gastropod Heleobia sp (Hidrobiidae) in vegetated protected shallow embayment of the Patos Lagon
estuarine area, R.S.
The model ( conceptual and mathematic versions) was build based on bibliographic
information, and codified in BASIC language for IBM-PC computers. Due to the small amount of
field data about the studied relationship, theoric and calibrated values were used for the parameters
in the model.
It was carried out 10 simulations wich plotted the predator and prey biomass variations
versus time expressed in Julian days. From those simulations, eight simulations have used
variations from the parameters values utilized in the first one. The final run of the model used the
same parameters from the first simulation, but plotting prey biomass versus predator biomass, in a
graphic named phase plane curve diagram. This proceeding allows the observation of the cyclic
behavior of the predator-prey biomass variation.
The simulations suggested the interdependence between the predator and preys, and the
quantitative effect of environmental factors controlling their biomass variation.

iv
CONTEÚDO
RESUMO .............................................................................................................. 2
ABSTRACT .......................................................................................................... 3
1 INTRODUÇÃO .............................................................................................. 1
1.1 MODELOS ECOLÓGICOS. ................................................................................................................... 1
1.2 TIPOS DE MODELOS. ............................................................................................................................ 2
1.3 USOS DOS MODELOS. ........................................................................................................................... 3
1.4 MODELOS PRESA-PREDADOR.......................................................................................................... 4
2 METODOLOGIA ........................................................................................... 9
2.1 ETAPAS DA MODELAGEM ................................................................................................................. 9
2.1.1 Descrição do sistema em estudo. ............................................................................................. 9
2.1.2 Elaboração do modelo conceitual. ........................................................................................... 9
2.1.3 Elaboração do modelo matemático com base no modelo conceitual. ........................................ 9
2.1.4 Simulação do modelo. .......................................................................................................... 10
2.1.5 calibração e validação. .......................................................................................................... 10

v
2.2 ESTRUTURA DO MODELO................................................................................................................ 10
2.2.1 MODELO CONCEITUAL ................................................................................................... 10
2.2.2 DESCRIÇÃO DO MODELO ............................................................................................... 12
2.3 MODELO MATEMÁTICO .................................................................................................................. 14
2.3.2 Variação da Biomassas de Epífitas ........................................................................................ 16
2.4 MODELO DE SIMULAÇÃO ................................................................................................................ 19
3 RESULTADOS E DISCUSSÃO .................................................................. 21
3.1 SIMULAÇÕES......................................................................................................................................... 21
3.1.1 Simulação 1 .......................................................................................................................... 21
3.1.2 Simulação 2 .......................................................................................................................... 22
3.1.3 Simulação 3 .......................................................................................................................... 23
3.1.4 Simulação 4 .......................................................................................................................... 24
3.2.1 Simulação 5 .......................................................................................................................... 25
3.2.2 Simulação 6 .......................................................................................................................... 26
3.2.3 Simulação 7 .......................................................................................................................... 27
3.2.4 Simulação 8 .......................................................................................................................... 28
3.2.5 Simulação 9 .......................................................................................................................... 29
3.2.6 Simulação 10 - Diagrama de Curva de Plano de Fase ............................................................ 30

vi
4 CONCLUSÕES ........................................................................................... 32
5 REFERÊNCIAS BIBLIOGRÁFICAS ........................................................... 34
6 APÊNDICES ............................................................................................... 37
6.1 APÊNDICE I ............................................................................................................................................ 37
6.2 APÊNDICE II........................................................................................................................................... 38
6.2.1 Listagem do Modelo de Simulação ....................................................................................... 38
6.2.2 Descrição do Programa ......................................................................................................... 39
6.2.3 Funcionamento do Programa ................................................................................................ 40

i
1 INTRODUÇÃO
1.1 MODELOS ECOLÓGICOS.
Um modelo pode ser entendido como uma simplificação de um sistema real qualquer,
entendendo-se por sistema qualquer fenômeno estrutural ou funcional que tenha pelo menos dois
componentes separados com alguma interação entre eles. Por ser a simplificação do sistema que se
estuda, o modelo não pode ter todos os detalhes do sistema real, pois senão ele não seria um modelo
mas o próprio sistema (Hall & Day, 1977).
Na sua fase mais simples, os modelos tendem a terem formas verbais ou gráficas para
descrever o sistema. Com o seu desenvolvimento eles adquirem estruturas mais complexas, evoluindo
para modelos matemáticos, que se utilizam de equações para poder simular o sistema modelado
(Odum 1986, Hall & Day, 1977). Para se entender o processo da modelagem pode-se observar o
diagrama representado na figura 1, onde estão representadas as principais etapas do desenvolvimento e
uso dos modelos. Uma observação importante sobre esta representação é o fato de que um modelo é
sempre feito para ser substituído por outro melhor, na medida em que aumentam nossos
conhecimentos sobre o sistema modelado (Hall & Day, 1977; Meyer, 1992).

2
Figura 1: Diagrama representativo do processo de modelagem
1.2 TIPOS DE MODELOS.
Existem dois tipos principais do modelos, os analíticos e os de simulação. O modelo analítico é
feito geralmente com o uso de matemática relativamente complexa. Seu desenvolvimento utiliza
procedimentos matemáticos para chegar a resultados precisos de algumas equações, que descrevem
com exatidão os fenômenos estudados. Tais equações idealmente devem ser muito bem conhecidas
para que se possa chegar ao seu exato resultado. Por outro lado estas equações funcionam
satisfatoriamente apenas quando representam fenômenos lineares ou quando seu número for reduzido,
o que torna este tipo de modelo de pouca utilidade no estudo de ecossistemas completos.
Ecossistemas constantemente envolvem um grande número de componentes com
comportamento não linear, exigindo a resolução de muitas equações na sua representação.

3
O modelo de simulação possui menor exatidão no resultado de suas equações, não sendo
capaz de estudar o comportamento do sistema através de soluções matemáticas precisas. Entretanto
possibilita a resolução quase simultânea de muitas equações lineares e não lineares. Não é portanto
necessário excluir-se importantes componentes do modelo na tentativa de representar o sistema
através de um grupo de equações conhecidas (Hall & Day, 1977).
1.3 USOS DOS MODELOS.
Os modelos podem ser utilizados de várias maneiras. Entre seus objetivos destacam-se a
ajuda na conceitualização, medição e comunicação de fenômenos complicados e, algumas vezes, na
previsão da conseqüência de ações realizadas sobre o sistema modelado através do estudo deste
sistema sob condições que não podem ser observadas ou criadas na realidade ou num período de
tempo real.
Eles também ajudam a testar a validade de medições de campo e de hipóteses levantadas a
partir destes dados. Se o modelo discorda do sistema real, o modelo ou o sistema estudado, ou
ambos, são pouco conhecidos e devem ser melhor estudados. Nesse sentido a detecção dos erros
existentes no modelo auxilia no aumento do conhecimento do sistema, indicando as áreas em que é
necessário desenvolver maiores estudos para se preencher lagunas que existam nas informações
existentes sobre o sistema estudado (Hall & Day, 1977).
O fato de que a realização de um modelo envolve o uso de informações oriundas de várias
áreas de pesquisa diferentes, permite que se possam desenvolver novas linhas de estudo em áreas
que integrem diferentes campos de pesquisa em torno de um objetivo comum.

4
1.4 MODELOS PRESA-PREDADOR.
Os modelos de relações presa-predador são utilizados para o estudo de interações que
ocorrem entre duas populações nas quais uma delas depende da outra para se alimentar e sobreviver.
Tais relações dinâmicas entre presas e predadores são temas de destaque em ecologia (Berryman
1992).
Diversos modelos com diferentes características foram baseados nesta relação, para varias
populações, e interações existentes entre elas. Por volta de 1925 Lotka e Volterra desenvolveram um
dos modelos matemáticos de mais largo uso e destacada importância para este tipo de relação,
conhecido como Modelo Presa-Predador de Lotka-Volterra. Este modelo foi criado com base em
modificações observadas para populações de pescadas e tubarões no Mar Adriático por ocasião da
paralisação e posterior retomada das atividades pesqueiras como conseqüência da 1ª Guerra
Mundial. Ele funciona tendo em conta aspectos da dinâmica das populações, na qual a taxa de
variação de presa é função de seu crescimento populacional menos a predação, e a taxa de variação
do predador é função da predação menos sua morte natural. Este modelo foi posteriormente
melhorado com a introdução de dois importantes fatores:
1) A limitação de espaço disponível para o crescimento de presas e predadores em função de
suas biomassas.
2) E a relação de crescimento dos predadores não sendo mais função da densidade da
biomassa das presas disponíveis, mas da proporção entre as densidades de biomassas
de predadores e presas.

5
Este último fator gera uma resposta mais próxima da realidade quando trata da mortalidade
da população de predadores em função do alimento disponível (Meyer, 1992; Berryman, 1992;
Ginzburg & Akçakaya, 1992).
No meio marinho muitas relações do tipo presa-predador podem ser observadas e modeladas
segundo o modelo Lotka-Volterra. Exemplos vão da predação de fitoplancton por zooplancton até a
pesca industrial de peixes realizadas pelo homem.
Um exemplo destacado é a relação observada em pradarias de gramíneas submersas de
regiões estuarinas, entre epífitas assentadas sobre suas folhas e o epibentos herbívoro que alí se
alimenta destas epífitas. A população de epífitas filtra a radiação luminosa, diminuindo a qualidade
da luz que chega até a superfície foliar das gramíneas, além de aumentar essa superfície foliar,
provocando um aumento da taxa de sedimentação, assim como um aumento da resistência física da
folha as correntes, facilitando sua desfoliação (Twilley et al., 1985; Wetzel & Neckles, 1986).
Neste sistema, a presença do epibentos herbívoro é de grande importância. Tem sido
observado que declínios na abundância de fanerógamas estão ligados a um grande crescimento
excessivo de epífitas nas suas folhas.
A pastagem realizada pelos herbívoros sobre estas epífitas, controla sua abundância,
beneficiando as gramíneas. Em pradarias de Zostera marina foi constatado, que plantas sem a
presença de pastadores apresentavam até quatro vezes mais biomassa de perifiton do que as plantas
onde estes animais estavam presentes. Estes pastadores podem ainda auxiliar na reciclagem de
nutrientes através de sua produção fecal (Montfrans et al., 1984).

6
Na região do Estuário da Lagoa do Patos (Figura 2) existem várias enseadas rasas protegidas
totalmente, também denominadas de "sacos", onde se desenvolvem extensos fundos de gramíneas
marinhas representadas, principalmente por Ruppia maritima (Potamogeatomacea), e nestes fundos
há uma grande abundância e biomassa de epífitas e epibentos herbívoro associados, representado
principalmente pelo gastrópode hidrobídeo Heleobia sp. Nestas enseadas são também observadas
várias outras interações do tipo presa - predador entre a macrofauna bentônica e seus predadores
carnívoros, peixes e crustáceos, que utilizam estas áreas como zonas de criação. A importância
destas relações tem sido observada, já que a presença ou não de tais predadores provocam alterações
na estrutura da comunidade predada. Por outro lado deve-se observar que em fundos de gramíneas, a
predação por parte de peixes e outros carnívoros pode ficar prejudicada pela densa cobertura
vegetal, por vezes desenvolvida, que fornece proteção às populações associadas (Coutinho et al.,
1982; Bemvenuti, 1983; Asmus, 1984). Os trabalhos anteriormente realizados nestes fundos de
gramíneas, se caracterizam como estudos sobre a estrutura das comunidades alí existentes, com
destaque à descrição de seus organismos e suas variações no espaço e no tempo. Não existem, até o
momento, estudos voltados ao desenvolvimento de modelos que representem os aspectos dinâmicos
ou sistêmicos destas comunidades e de suas relações internas.
Neste trabalho apresenta-se um estudo de modelagem e simulação das interações do tipo
presa-predador existente entre as epífitas de Ruppia maritima e a população de Heleobia sp. Essa
modelagem enfoca um processo considerado importante nos ambientes de gramíneas submersas. Ela
é parte de um programa maior de caracterização e modelagem do ecossistema estuarino da Lagoa
dos Patos desenvolvido pelo Laboratório de Ecologia da FURG. (Mantovanelli & Asmus, 1992;

7
Teixeira da Silva & Asmus, 1992; Bonilha & Asmus, 1993). Seus objetivos mais específicos
envolveram o desenvolvimento de um modelo númerico de simulação da relação entre epífitas e
Heleobia sp., e a aplicação de exercícios teóricos de simulação que possibilitassem a criação de
hipóteses sobre os mecanismos e fatores ambientais envolvidos nessa interação.

8
Figura 2: Região do Estuário da Lagoa dos Patos.

9
2 METODOLOGIA
2.1 ETAPAS DA MODELAGEM
A modelagem de um sistema passa idealmente por quatro etapas.
2.1.1 Descrição do sistema em estudo.
Através de levantamento bibliográfico e checagem de campo determinam-se os principais
componentes, os fenômenos físicos atuantes e as interações que ocorrem entre seus componentes
bióticos e abióticos. A partir desta descrição, pode-se coletar informações ainda necessárias a
descrição do sistema e a futura elaboração do modelo. Em varios casos, como neste trabalho, o
modelo configura-se como um estudo teórico sobre um determinado aspecto do funcionamento
deste sistema, utilizando-se de informações disponíveis para avaliar o comportamento do modelo em
comparação com o comportamento do sistema real.
2.1.2 Elaboração do modelo conceitual.
Junto a descrição do sistema, representa-se o sistema de forma diagramática com o uso de
simbologia própria para representar seus componentes, fluxos e interações, fontes de energia, e
subsistemas existentes.
2.1.3 Elaboração do modelo matemático com base no modelo conceitual.
Este modelo representa na forma de equações matemáticas o sistema, ou parte do sistema,
em estudo e que se deseja posteriormente simular. Geralmente ele é composto por um conjunto de
equações representativas dos processos observados no sistema.

10
2.1.4 Simulação do modelo.
O modelo pode ser simulado de vários modos com a utilização de computadores. Usa-se
normalmente programas matemáticos que permitem a resolução em conjunto das equações
desenvolvidas, através da aplicação de rotinas apropriadas (Swartzman, 1987).
2.1.5 calibração e validação.
A calibração do modelo consiste no ajuste de suas variáveis para valores mais coerentes com
o realidade. Tenta-se produzir uma resposta mais próxima possível do comportamento real do
sistema estudado.
A validação é feita após a calibração. Ela consiste no uso de dados do sistema modelado
coletados em outras datas, ou de dados de outros sistemas similares ao estudado para testar a
capacidade do modelo em gerar para estes outros sistemas respostas compatíveis com a realidade (Hall
& Day, 1977).
2.2 ESTRUTURA DO MODELO
2.2.1 MODELO CONCEITUAL
O modelo conceitual do sistema estudado está representado na figura 3 conforme simbologia
proposta por Odum, 1988 (vide apêndice I).
Nesta representação podem ser observados seus três componentes principais que são:

11
a. Variáveis de estado representantes dos elementos estruturais internos do sistema. São
elas as epífitas, o epibentos, o compartimento de detritos, o compartimento de nutrientes
e os microorganismos decompositores de detritos.
b. Subsistemas adjacentes como as fanerógamas submersas.
c. Funções forçantes e de controle representadas pelas entradas de material ou importação
de fora do sistema para seus compartimentos internos (Hall & Day, 1977).
Figura 3: Modelo conceitual do sistema estudado.

12
2.2.2 DESCRIÇÃO DO MODELO
Na Figura 3 são representados os principais componentes e processos que atuam no sistema
da relação de predação entre o epibentos e epífitas em fundos de gramíneas marinhas.
No modelo aparecem como funções forçantes externas ao sistema a intensidade luminosa
(SOL), aporte de novos nutrientes (NUT) disponíveis às epífitas, temperatura (TEMP), salinidade
(SAL), e corrente existente no local (COR). Além das forçantes existem duas saídas e uma entrada
de matéria, são eles a exportação de detritos para fora do sistema principal, a predação e o
recrutamento de epibentos. Existe também um subsistema representando a comunidade de
fanerógamas sobre a qual a interação presa-predador ocorre (Wetzel & Neckles, 1986; Twiley et al,
1985).
Este subsistema de fanerógamas influencia diretamente na produção primária realizada pelas
epífitas pelo sombreamento que suas folhas realizam, e pela área para crescimento das epífitas que
elas proporcionam, indiretamente influenciando o bentos, que irá se alimentar de acordo com a
quantidade de biomassa de epífitas disponível.
As funções forçantes controladoras do processo de produção primária são a intensidade
luminosa, nutrientes e temperatura.
Note-se que no sistema:
- A intensidade luminosa é atenuada naturalmente pela coluna de água e pelo sombreamento
causado pela cobertura de fanerógamas, estando esta atenuação representada pela saída do sistema
de uma parte da luminosidade (Odum, 1988).

13
- Os nutrientes disponíveis para as epífitas, dentro do sistema também provêm da reciclagem
de nutrientes realizada pelo epibentos, que junto ao nutriente vindo de fora do sistema, forma o
depósito de nutrientes interno ao sistema (Montfrans et al, 1984).
- A biomassa de epífitas também influencia na produção uma vez que a quantidade
produzida depende da biomassa disponível para converter energia luminosa em matéria orgânica.
Esta biomassa também atua na atenuação da luminosidade devido ao sombreamento realizado pelo
crescimento das epífitas umas sobre as outras (Ferreira & Seeliger, 1985).
Na produção secundária, predação de bentos sobre as epífitas, ocorre influência da
temperatura, da biomassa de epífitas disponível para a predação e da biomassa de predadores
existente. A biomassa de bentos é influenciada pelo crescimento dos indivíduos alimentados pelas
epífitas, pelo recrutamento de novos predadores e pela saída de indivíduos do sistema por predação
ou outro fator de mortalidade. Sobre o bentos também ocorre uma influência da salinidade como
condicionadora de sua presença no sistema (Fenchel, 1975).
Os fluxos que representam as saídas de energia e matéria do sistema são representados por:
a. fluxo de dissipação de energia na forma de calor (segunda lei da termodinâmica)
existente em todos os componentes do sistema.
b. a perda de biomassa de bentos pela ação de predadores.
c. a exportação de detritos influenciada pela intensidade das correntes.

14
A transferência de biomassa de epífitas para o compartimento de detritos (mortalidade) está
também em parte relacionada a quebra e retirada de folhas de fanerógamas pela ação das correntes
existentes no local.
O subsistema das fanerógamas é influenciado pela biomassa de epífitas através do
sombreamento, que causam competição por CO2 e nutrientes. O crescimento das epífitas sobre as
fanerógamas também gera efeitos positivos para a gramínea. Eles reduzem a dissecação retendo a
umidade, podendo também desviar a pastagem da área fotossinteticamente ativa na base das folhas
para a parte senescente nas pontas das folhas (Montfrans et al, 1984).
2.3 MODELO MATEMÁTICO
Após concluído o modelo conceitual, fez-se um modelo matemático que representa aquele na
forma de equações serem posteriormente simuladas .
O modelo matemático não precisa necessariamente representar o mesmo modelo conceitual
feito para todo o sistema. Mais comumente se expressa apenas a parte que se tenha mais interesse em
simular. Neste caso, o modelo a ser simulado está representado na Figura 4. Matematicamente ele se
compõe de duas equações principais expostas abaixo:
EPIFt = EPIFt-1 + DE * DT Eq. 1
HELIt = HELIt-1 + DH * DT Eq. 2
Onde:
EPIF é a biomassa de epífitas.

15
HELI é a biomassa de Heleobia sp.
DE a variação da biomassa de epífitas após certo intervalo de tempo DT.
DH a variação de biomassa de Heleobia sp., após este intervalo de tempo DT.
A biomassa existente no momento t é função da biomassa existente no momento t-1 mais uma
variação desta biomassa após um período DT (Hall & Day, 1977).
Figura 4: Modelo conceitual representando a parte do sistema

16
2.3.1.1 Fatores Ambientais
A variação das biomassas de Heleobia sp. e epífitas dependem de dois fatores ambientais.
São eles a temperatura e a intensidade luminosa, representadas no modelo através de equações
teóricas do tipo cossenóide (Odum, 1989).
TEMP = TM + AMPT * COS ((2 * PI / 365) * (D + T) Eq. 3
SOL = IM + AMPI * COS ((2 * PI / 365) * (D + T) Eq. 4
Onde:
TM é a temperatura média.
AMPT é a amplitude da temperatura.
IM é a intensidade luminosa média.
AMPI é a amplitude da intensidade luminosa
T é o tempo de simulação.
D é o número de dias do calendário juliano que deve ser somado ou subtraído de T para que a
fase da curva gerada se ajuste com os valores nas datas em que foram observados.
O termo dependente do coseno é um indíce calculado para que a temperatura e a intensidade
luminosa variem sazonalmente (Kremer & Nixon, 1978; Asmus, 1990).
2.3.2 Variação da Biomassas de Epífitas
A variação de biomassa de epífitas (DE) está exposta abaixo:

17
DE = ((A'* EPIFo) - (B * EPIFo * HELIo)) Eq. 5
Sendo que:
(A' * EPIFo ) representa o crescimento da população de epífitas em função de A'.
(B * EPIFo * HELIo) representa a perda de biomassa de epífitas devido á predação.
A' = D * TEMP * LUM * SPE Eq. 6
Onde:
D é a taxa de crescimento das epífitas (Rodriquez, 1987).
TEMP é a temperatura (Montfrans et al, 1984).
SPE é a limitação de espaço disponível para o crescimento das epífitas (Berrryman, 1988).
LUM é a luminosidade real disponível (Wetzel & Neckles, 1986).
B é a taxa de predação da biomassa de epífitas existente (Odum, 1988; Meyer, 1992).
Sendo que LUM representa a luminosidade real que chega até as epifitas após a atenuação das luz
pelo sombreamento causado pelas próprias epífitas. Sua equação é:
LUM = SOL / (1 + A * EPIFo) Eq. 7
Onde:
SOL é a intensidade luminosa que chega as epífitas.
A é uma taxa de atenuação da luz devido ao sombreamento das epífitas (Odum, 1988).
E:

18
SPE = 1 - EPIFo / KE Eq. 8
Onde:
KE é a densidade de equilíbrio de epífitas ou capacidade de suporte que pode ser vista como
o valor de biomassa acima do qual não há aumento importante na população (Berryman, 1992;
Odum, 1986).
2.3.2.1 Variação da Biomassa de Heleobia sp.
A equação de variação de Heleobia sp. (DH) é:
DH = ((C' * HELIo) - (M * HELIo)) Eq. 9
Sendo que:
(C' * HELIo) representa o crescimento da biomassa de Heleobia.
(M * HELIo) representa a mortalidade natural, independente do alimento.
C' = C * TEMP * SPB * AL Eq. 10
Onde:
M é a taxa de mortalidade (Ginzburg & Akçakaya, 1992).
C é a taxa de crescimento de Heleobia (Meyer, 1992).
SPB é a limitação do espaço disponível.
AL é a limitação do alimento disponível (Montfrans, 1984; Ginzburg & Akçakaya, 1992).
Sendo que SPB é:

19
SPB = 1 - HELIo / KB Eq. 11
Onde:
KB é a densidade de equilíbrio para predadores.
AL representa a resposta do crescimento de Heleobia sp. em função da proporção existente
entre sua biomassa e a biomassa de epífitas. Ele depende da quantidade de epífitas necessárias para
manter uma Heleobia sp. (E), descrevendo a produção per-capita de Heleobia sp.. Isto faz com que
ocorra uma dependência mais direta de Heleobia sp. pela quantidade de epífitas disponível para sua
alimentação, levando em conta a interferência entre os consumidores á medida que sua população
cresce. Considera ainda a alteração da taxa de alimentação individual, a qual influencia a taxa de
alimentação total da população de predadores (Ginzburg & Akçakaya, 1992; Berryman, 1992).
AL = 1 - E * HELIo / EPIFo Eq. 12
2.4 MODELO DE SIMULAÇÃO
O modelo de simulação foi codificado em linguagem BASIC e sua descrição esta
representada no apêndice II.
Durante as simulações, os resultados obtidos foram gravados como tabelas de dados e
posteriormente lidos no programa Quattro-Pro (Borland International Inc.), o que permitiu sua
melhor apresentação na forma de gráficos.
20
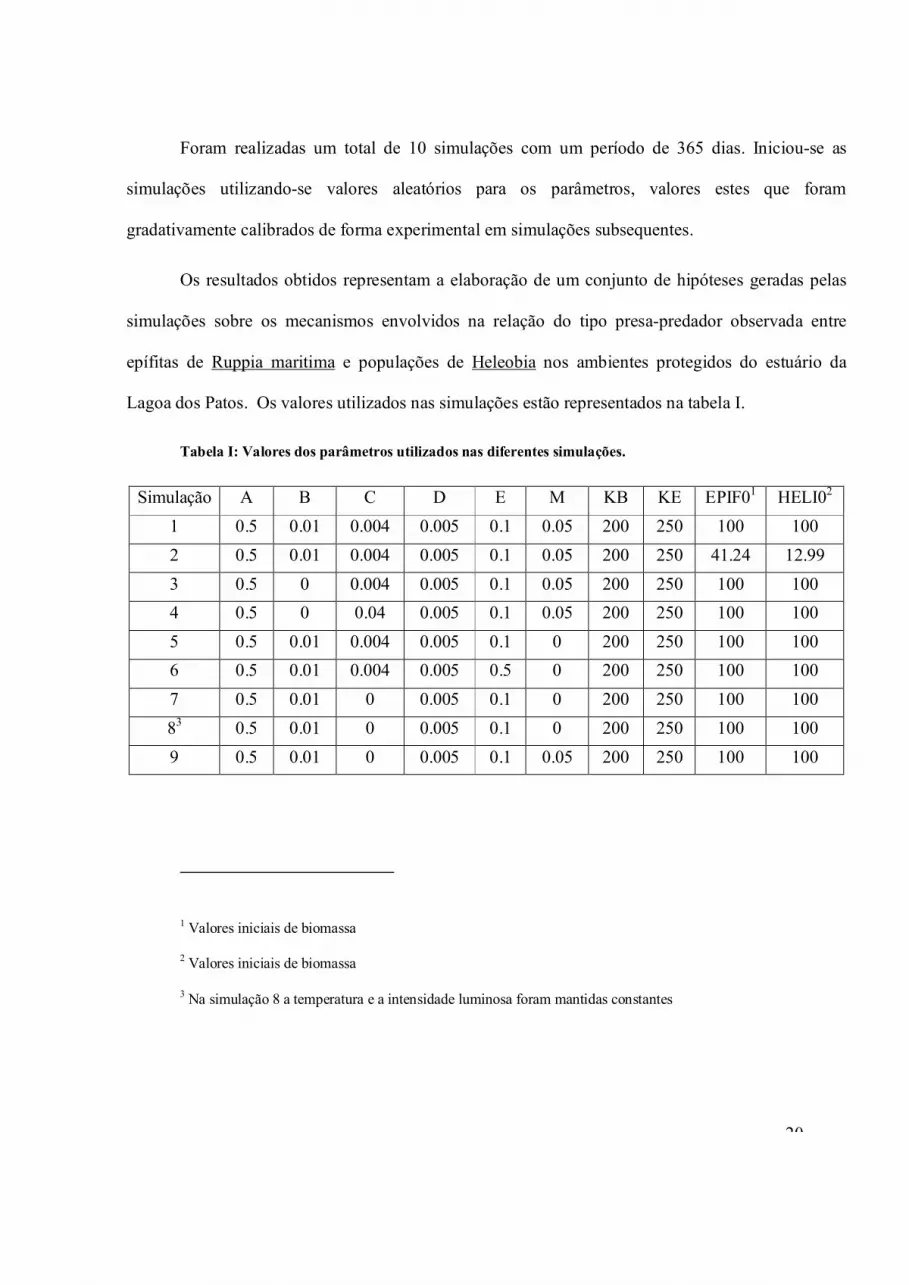
Foram realizadas um total de 10 simulações com um período de 365 dias. Iniciou-se as
simulações utilizando-se valores aleatórios para os parâmetros, valores estes que foram
gradativamente calibrados de forma experimental em simulações subsequentes.
Os resultados obtidos representam a elaboração de um conjunto de hipóteses geradas pelas
simulações sobre os mecanismos envolvidos na relação do tipo presa-predador observada entre
epífitas de Ruppia maritima e populações de Heleobia nos ambientes protegidos do estuário da
Lagoa dos Patos. Os valores utilizados nas simulações estão representados na tabela I.
Tabela I: Valores dos parâmetros utilizados nas diferentes simulações.
Simulação A B C D E M KB KE EPIF01 HELI02
1 0.5 0.01 0.004 0.005 0.1 0.05 200 250 100 100
2 0.5 0.01 0.004 0.005 0.1 0.05 200 250 41.24 12.99
3 0.5 0 0.004 0.005 0.1 0.05 200 250 100 100
4 0.5 0 0.04 0.005 0.1 0.05 200 250 100 100
5 0.5 0.01 0.004 0.005 0.1 0 200 250 100 100
6 0.5 0.01 0.004 0.005 0.5 0 200 250 100 100
7 0.5 0.01 0 0.005 0.1 0 200 250 100 100
83 0.5 0.01 0 0.005 0.1 0 200 250 100 100
9 0.5 0.01 0 0.005 0.1 0.05 200 250 100 100
1 Valores iniciais de biomassa
2 Valores iniciais de biomassa
3 Na simulação 8 a temperatura e a intensidade luminosa foram mantidas constantes

21
3 RESULTADOS E DISCUSSÃO
3.1 SIMULAÇÕES
3.1.1 Simulação 1
A simulação inicial está representada na Figura 5.
Pode-se observar a instabilidade inicial do sistema no inicio do primeiro ano estando ela
visível na forma de uma queda acentuada nas biomassa de presas e predadores. Esta instabilidade se
deve a fato de que os valores iniciais de biomassas, não estão devidamente ajustados aos valores dos
parâmetros utilizados. Isto faz com que o sistema tenda a uma grande variação inicial, até chegar a
situação de equilíbrio entre biomassas de presas e predadores ao final do primeiro ano.
Figura 5: Simulação 1

22
3.1.2 Simulação 2
Esta simulação (Figura 6) é semelhante a simulação anterior. Ela utiliza como valores iniciais
de biomassa os valores que o modelo atingiu após rodar um ano (EPIFo=41,24g e HELIo=12,99g).
Pode-se observar que não ocorre nenhum tipo de instabilidade, com o modelo desde o início da
simulação, apresentando uma oscilação constante e periódica característica de um sistema em
equilíbrio entre as populações de presas e predadores.
Figura 6: Simulação 3

23
3.1.3 Simulação 3
Nesta simulação (Figura 7), o valor do parâmetro B foi zerado de modo que não ocorre saída
de biomassa do compartimento das presas.
Observa-se que a biomassa de presas cresce até atingir a capacidade de suporte estabelecida
para o ambiente. Por outro lado a biomassa de predadores apresentou uma pequena queda. Poderia ser
esperado que a curva dos predadores também subisse até o valor de sua capacidade de suporte,
mantendo-se constante neste ponto. Dois fatores podem estar causando essa queda, assim como a
oscilação, uma baixa taxa de crescimento de Heleobia (C) gerando um desequilíbrio inicial entre um
baixo crescimento e uma maior mortalidade de predadores causando a redução da população, e a
influência da temperatura na predação que por estar também oscilando gera a variação da biomassa de
predadores.
Figura 7: Simulação 3

24
3.1.4 Simulação 4
Nesta simulação (Figura 8), o valor da taxa de crescimento de Heleobia sp. (C) foi
multiplicado por dez (C=0,04, B=0).
É possível observar que a curva de predadores não mais apresenta uma queda inicial,
crescendo como o esperado até um valor bem próximo de sua capacidade de suporte. Ela ainda
apresenta uma oscilação em menor escala devido ao fato de que a alta taxa de predação minimiza os
efeitos da queda de temperatura sobre o crescimento de predadores.
Figura 8: Simulação 4

25
3.2.1 Simulação 5
Aqui a mortalidade de predadores por outros fatores além do alimento (M) foi zerada para uma
melhor visualização da dependência dos predadores pela biomassa de presas (Figura 9).
Poderia ser esperado na ausência de mortalidade,um comportamento da biomassa de
predadores semelhante ao das presas (vide simulação 3) aumentando até o valor de sua capacidade de
suporte. Isto no entanto, não ocorre devido a mortalidade de predadores em função da disponibilidade
de alimento.
Figura 9: Simulação 5

26
3.2.2 Simulação 6
Para se testar a idéia da relação entre o valor do parâmetro E e a dependência da biomassa de
predadores, este valor foi multiplicado por cinco atingindo 0.5 (M=0) (Figura 10).
Observa-se que realmente este parâmetro tem grande influência na biomassa de predadores
pois, não só a sua oscilação ficou menor, como também a sua biomassa diminuiu, possibilitando um
pequeno aumento na biomassa de presas.
Isto deve-se ao fato de que quanto mais presas foram necessárias para alimentar um predador,
menor será a quantidade de alimento disponível para determinado número de predadores, ocasionando
uma redução de sua população até valores em que o alimento seja suficiente. Ao mesmo tempo, uma
redução da biomassa de predadores gera um aumento da biomassa de presas, até que ambos atinjam a
situação de equilíbrio.
Figura 10: Simulção 6

27
3.2.3 Simulação 7
Nesta simulação (Figura 11) a biomassa de predadores foi mantida constante zerando-se os
valores de sua taxa de crescimento (C) e de sua mortalidade (M).
Observa-se que a biomassa de presas se mantêm em valores muito baixos. Nessa condição há
uma espectativa de que ela não apresente oscilação, uma vez que depende da biomassa de predadores
para seu controle.
Deve ser observado, no entanto, que a biomassa de presas depende também da temperatura e
da intensidade luminosa que permanecem em oscilação. Elas são, nesse caso,as responsáveis pela
oscilação das presas.
Figura 11: Simulação 7

28
3.2.4 Simulação 8
Nesta simulação (Figura 12) é testada a influência da temperatura e da intensidade luminosa na
variação da biomassa de presas quando a biomassa de predadores se mantêm constante (C=0, M=0).
Para tal, os valores de temperatura e intensidade luminosa foram mantidos constantes em
17,30‘C e 39,24 E/m2/dia.
Oserva-se que o modelo tem a resposta esperada, com a biomassa de presas mantendo-se
estável junto com a de predadores.
Figura 12:Simulação 8

29
3.2.5 Simulação 9
Nesta simulação (Figura 13), a produção de biomassa de predadores foi anulada zerando-se o
valor do parâmetro C, e M volta ao seu valor original 0,05.
Observa-se que devido a mortalidade a biomassa de predadores cai a zero enquanto que a de
presas sobe até a sua capacidade suporte devido a diminuição da quantidade de predadores atuando
sobre elas.
Figura 13: Simulação 9

30
3.2.6 Simulação 10 - Diagrama de Curva de Plano de Fase
Uma outra forma de se analisar os resultados do modelo é através de uma diagrama de curva
de plano de fase que é um gráfico onde se plota as presas pelos predadores.
Com ele é possível se observar a característica cíclica do comportamento das populações e
também se elas tendem ao equilíbrio ou a instabilidade, se a curva estiver se afastando do centro o
sistema tende a instabilidade, mas se ela estiver estável ou se aproximando do seu centro o sistema
tende ao equilíbrio, isto é observado durante a simulação, ou através de setas na curva.
A próxima simulação (Figura 14) dá um exemplo deste diagrama feito para a simulação 1.
Pode-se observar também a instabilidade do inicio da simulação antes da curva adquirir o
formato cíclico ao atingir seu estado de equilíbrio.
As simulações realizadas permitiram uma clara observação do efeito da variação da
biomassa de predadores sobre a biomassa de presas, assim como a de presas sobre a de predadores
neste modelo.
A sensibilidade para o parâmetro (E), que define o quanto uma Heleobia necessita de epífitas
para o seu desenvolvimento, fica clara ao comparar-se as simulações 5 e 6 demonstrando a
importância deste parâmetro para o desenvolvimento das duas populações.

31
Figura 14: Simulação 10. Diagrama de curva de plano de fase.
Foi demonstrado também a grande importância do parâmetro (C) (taxa de predação) no
desenvolvimento das populações , chegando este parâmetro a sobrepor o efeito da temperatura
(simulações 3 e 4).Por outro lado observa-se a influencia dos parâmetros ambientais (temperatura e
intensidade luminosa), no desenvolvimento das populações. há uma clara influencia da intensidade
luminosa para as epífitas (simulações 7 e 8), e da temperatura para a Heleobia sp. Entretanto, faz-se
necessário o desenvolvimento um número maior de simulações para a obtenção de resultados mais
expressivos destes parâmetros sobre as populações estudadas.
Finalmente, este trabalho projeta para o futuro a expectativa de que, com a obtenção de um
maior número de informações sobre o sistema estudado, o modelo proposto venha a ser
completamente validado. Tal validação, por certo servir como um fator de retroalimentação,

32
devendo gerar novas versões mais refinadas e mais próximas da realidade do modelo presa-predador
aplicado a interação entre epífitas e Heleobia sp. no estuário da Lagoa do Patos.
4 CONCLUSÕES
1 - Apesar das críticas ao modelo matemático de Lotka-Volterra pelo fato de que duas
populações raramente interagem apenas entre si , a possibilidade de desenvolvimento e expansão de
suas equações utilizadas neste trabalho, com o acréscimo de novas formulações para parâmetros
como a temperatura e intensidade luminosa, permite concluir que elas podem ser adaptadas para
situações que sejam melhor compatíveis com o funcionamento real das populações em estudo.
2 - As respostas obtidas para o comportamento das variáveis a partir da terceira simulação
foram compatíveis com o esperado para um sistema em situações reais, demonstrando um
funcionamento aceitável deste modelo na sua atual fase de desenvolvimento.
3 - O modelo permitiu uma clara observação da interdependência entre predadores e presas
em várias situações. Efeitos da taxa de predação no desenvolvimento dos predadores está refletido
nas variações do parâmetro (C) nas simulações 3 e 4, assim como a dependência dos predadores pela
quantidade de epífitas predadas, expressa pelo parâmetro (E) nas simulações 5 e 6.
4 - A impossibilidade de uma completa validação deste modelo com o estudo e analise das
populações modeladas, mostra a sua utilidade na indicação de lagunas de conhecimento do
comportamento de populações na área de estudo.

33
Deve-se também ressaltar a sua utilidade como material didático para os estudos iniciais de
modelagem ecológica. Aqui são representadas de forma simplificada todas as etapas da construção e
experimentação com modelos.
5 -Obtenção de novas informações sobre as comunidades estudadas deverá futuramente
permitir uma adaptação mais perfeita deste modelo para simulações que produzam respostas
compatíveis com o comportamento real.

34
5 REFERÊNCIAS BIBLIOGRÁFICAS
ASMUS, M.L.; 1984; Estrutura da Comunidade Associada a Ruppia maritima no Estuário da Lagoa
dos Patos; Rio Grande do Sul; Brasil.; Tese de mestrado; Furg; Brasil; 154 p.
ASMUS, M.L.; 1990; Ecological Modelling of the North Inlet Marsh- Estuarine System; South
Carolina; Tese de doutorado; University of South Carolina; 179p.
BEMVENUTI, C.A.; 1983; Efeitos da Predação Sobre as Características Estruturais de uma
Comunidade Macrozoobêntonica numa Enseada Estuarina da Lagoa dos Patos, R.S. Brasil;
Tese de Mestrado; Furg; Brasil; 120 p.
BERRYMAN, ALAN A.; 1992; The Origin and Evolution of Psredator- Prey Theory; Ecology; 73(5):
1530-1535.
BONILHA, L.E. & ASMUS, M.L.; 1993; Modelo Ecológico de Fitoplancton e Zooplancton no
Estuário da Lagoa dos Patos, RS.;Anais do III Simpósio de Ecossistemas da Costa Brasileira;
Serra Negra; SP. (in press).
COUTINHO, R.; CAFRUNI, A. & SEELIGER, U.; 1982; Production Ecology of Aquatic
Macrophytes in the Patos Lagoon Estuary, Brazil; Atlantica; 5(2): 30.
FENCHEL; T.; 1975; Factors Determining the Distribution Patterns of Mud Snails (hidrobiidae);
Oecologia; 20: 1-17.
FERGUSON, R.L. & ADAMS, S.M.; 1979; A Mathematical Model of Trophyc Dynamics in
Estuarine Seagrass Communities; (in) Marsh- estuarine Systems Simulation; University of
South Carolina Press; 41-70.

35
FERREIRA, S. & SEELIGER, U.; 1985; The Colonization Process of Algal Epiphytes on Ruppia
maritima L; Bot. Marina; 28: 245-249.
GINZBURG, LEV R.; AKÇAKAYA, H.R.; 1992; Consequenses of Ratio-Dependent Predation for
Steady-State Properties of Ecosystems; Ecology; 73(5): 1536-1543.
HALL, C.A.S. & DAY Jr., J.W.; 1977; Ecosystem Modelling In Theory and Pratice: An Introduction
with case Histories; Wiley Intercience; New York; 6-35.
HYLLEBERG, J.; 1975; The Efect of Salinity and Temperature on Egestion in Mud Snais
(Gastropoda: Hydrobiidae); Oecologia; 21: 279-289.
KREMER, J.N. & NIXON, S.W.; 1978; A Coastal Marine Ecosystem: Simulation and Analysis.;
Ecological Studies; Springer-Verlag; Berlin; Vol. 24; 217p.
MANTOVANELLI, A. & ASMUS, M.L.; 1992; Caracterização e Modelagem Ecológica das
Enseadas Rasas Protegidas na Região Estuarina da Lagoa dos Patos, RS.; Simpósio sobre
Estrutura, Funcionamento e Manejo de Ecossistemas; Rio de Janeiro; Resumos pg. 141.
MEYER, J. F. C. A.; 1992; Biomatemática: Um Jeito de Ver Ecologia; V Semana Nacional de
Oceanografia; documento interno; Rio Grande; R.S.
MONTFRANS, J. van; WETZEL, R.L & ORTH, R.J.; 1984; Epiphyte- Grazer Relationships in
Seagrass Meadows: Consequences for Seagrass Growth and Production; Estuaries; 7(4a):
289-309.
ODUM, E.P.; 1986; Ecologia; Ed. Guanabara S.A.; Rio de Janeiro; 434 p.
ODUM, H.T.; BROWN, E.C.; LAHART, D.; BERSOK, C. & SENDZIMIR.J.; 1988; Environmental
Systems and Public Policy; University of Florida; Gainesville; 253p.

36
ODUM, H.T.; ODUM, E.C.; 1989; Computer Minimodels and Simulation Exercises for Science and
Social Science; University of Florida; Gainesville; 319p.
RODRIGUEZ, M.A.; 1987; Estimating Periphyton Growth Parameters using Simple Models; Limnol.
Oceanogr.; 32(2): 458-464.
SWARTZMAN, G.L.; KALUZNY, S.P.; 1987; Ecological Simulation Primer; Macmillan Pub. Co.;
NY; 370p.
TEIXEIRA DA SILVA, E. & ASMUS, M.L.; 1992; Abordagem Sistemica no Estudo de Fundos de
Fanerógamas Marinhas no Estuário da Lagoa dos Patos, RS.; V Semana Nacional de
Oceanografia; Rio Grande; RS; Resumos pg. 18.
TWILLEY, R.R.; KEMP, W.M.; STAVER, K.W.; STEVENSON, J.C. & BOYTON, W.R.; 1985;
Nutrient Enrichment of Estuarine Submersed Vascular Plant Communities. 1. Algae Growth
and Effects in Prodution of Plants and Associated Communities; Mar. Ecol. Prog. Ser.; 23:
179-191.
WETZEL, R.L. & NECKLES, H.A.; 1986; A Model of Zostera marina L. Photosynthesis and
Growth: Simulated Effects of Selected Physical-Chemical Variables and Biological
Interactions; Aquatic Botany; 26: 307-323.

37
6 APÊNDICES
6.1 APÊNDICE I
Abaixo estão representados os principais simbolos utilizados para a construção de modelos
conceituais, segundo Odum 1988.

38
6.2 APÊNDICE II
6.2.1 Listagem do Modelo de Simulação
Abaixo está representado o programa construído para realizar a simulação do modelo matemático: 10
'MODEL 5
20 CLS
30 SCREEN 1, 0, B
40 LINE (0, 0)-(639, 180), 3, B
50 OPEN "C:\SIM.PRN" FOR OUTPUT AS #1
60 T = 1
70 DT = 1
80 TF = 365
100 PI = 3.141592535898
110 EPIF = 100
120 HELI =100
130 A = .5
140 B = .01
150 C = .004
160 D = .005
170 E = .1
180 M = .05
190 KB = 200
200 KE = 250
210 TM = 17.663839#
220 AMPT = 5.485288
230 IM = 33.715658#
240 AMPI = 18.913072#
250 'EQUACOES

39
260 WRITE #1, EPIF, HELI
270 TEMP = TM + AMPT * COS((2 * PI / 365) * (270 + T))
280 SOL = IM + AMPI * COS((2 * PI / 365) * (291 + T))
290 LUM = SOL / (1 + A * EPIF)
300 PSET (T, 180 - EPIF), 3
310 PSET (T, 180 - HELI), 2
320 DE1 = B * EPIF * HELI
330 DE = (D * LUM * TEMP * EPIF * (1 - EPIF / KE)) - DE1
340 DB1 = (C * TEMP * HELI * (1 - E * HELI /EPIF)) * (1 - HELI / KB)
350 DB = DB1 - (M * HELI)
360 EPIF = EPIF + DE * DT
370 HELI = HELI + DB * DT
380 IF INKEY$ = CHR$(27) THEN STOP
390 IF EPIF <= 0 OR BENT <= 0 THEN STOP
400 T = T + (DT)
410 IF T <= TF GOTO 250
6.2.2 Descrição do Programa
Abaixo é feita a descrição do programa, estando o seu funcionamento exposto a seguir.
As linhas 10 e 250 são linhas de comentário que contém a identificação do programa e do
inicio das equações, não sendo utilizada durante o seu funcionamento.
As linhas 20 a 40 limpam a tela e definem o modo de apresentação do gráfico quando o
programa é iniciado.
A linha 50 abre um arquivo onde serão gravados os dados gerados pelo modelo para que
possam ser utilizados por outros programas.

40
As linhas 60 a 240 definem os valores iniciais das biomassas de epífitas e Heleobia e os
valores dos parâmetros utilizados pelo modelo.
A linha 260 grava no arquivo aberto na linha 50 os valores gerados pelo modelo.
Nas linhas 270 a 370 estão programadas as equações do modelo matemático exposto
anteriormente. As linhas 300 e 310 plotam a variação de biomassa de epífitas e de biomassa de
epibentos em relação ao tempo.
A linha 380 permite parar a simulação a qualquer momento apertando-se a tecla ESC, e a
linha 390 para automaticamente a simulação quando os valores de biomassa atingem 0,
especialmente no caso das epífitas para evitar erro de divisão por zero na linha 340.
A linha 400 é o contador de tempo, acrescentando um valor DT ao tempo T que é atualizado
para que sejam calculadas as biomassa no período seguinte.
A linha 410 gera um loop no o qual faz o modelo matemático ser executado novamente
enquanto o valor T for menor que TF, definido na linha 80.
6.2.3 Funcionamento do Programa
Quando for executado o programa funciona da seguinte forma.
Primeiro a tela é limpada, sendo em seguida desenhado uma moldura ao seu redor.
O programa então lê os valores iniciais dos parâmetros utilizadas.

41
Logo após é calculado os valores de TEMP, SOL, e LUM, que são utilizados do calculo do
acréscimo de biomassa DE e DH para epífitas e epibentos respectivamente. São então plotados os
valores iniciais de epífitas e de epibentos, e calculados os novos valores de biomassa de epífitas e
epibentos pela soma dos valores de DE e DH ás biomassa existentes no momento anterior, após o
qual é acrescido 1 ao valor inicial de T.
Por fim é dito ao programa que se o valor atual de T for menor ou igual a 365, definido por
TF, ele deve voltar a linha 250 executando de novo o modelo matemático, plotando os novos valores
de biomassa, e calculando os seus próximos valores até que T seja igual a 366, quando então o
programa para a sua execução.





























