Embed Size (px)
Citation preview

Recent advances of research in antinutritional factorsin legume seeds andoilseeds
EAAP publication No. 110, 2004Toledo, Spain
8-10 March 2004

Recent advances of research in antinutritional factorsin legume seeds and oilseeds

The EAAP series is published under the direction of Dr. P. Rafai
EAAP – European Association for Animal Production
INIA – Instituto Nacional de Investigación y Tecnología Agraria yAlimentaria
Ministerio de Ciencia y Tecnología
Junta de Comunidades de Castilla-La Mancha
The European Association for Animal Production wishes to express its appreciation to theMinistero per le Politiche Agricole e Forestali and the Associazione Italiana Allevatori for theirvaluable support of its activities

Recent advances ofresearch in antinutritionalfactors in legume seedsand oilseeds
Proceedings of the fourth internationalworkshop on antinutritional factors inlegume seeds and oilseeds
EAAP publication No. 110Toledo, Spain8-10 March 2004
Editors:
M. Muzquiz, G.D. Hill, C. Cuadrado, M.M. Pedrosa and C. Burbano
Wageningen AcademicWageningen AcademicP u b l i s h e r ssseessbP u b l i s h e r sP u b l i s h e r sP u b l i s h e r s

ISBN: 978-90-76998-39-8e-ISBN: 978-90-8686-524-6
DOI: 10.3920/978-90-8686-524-6
ISSN 0071-2477
Subject headings:animal and human nutrition
feed and food technologyplant breeding
First published, 2004
© Wageningen Academic Publishers The Netherlands, 2004
This work is subject to copyright. All rights are reserved, whether the whole or part of the material is concerned. Nothing from this publication may be translated, reproduced, stored in a computerised system or published in any form or in any manner, including electronic, mechanical, reprographic or photographic, without prior written permission from the publisher, Wageningen Academic Publishers, P.O. Box 220, 6700 AE Wageningen, the Netherlands, www.WageningenAcademic.com
The individual contributions in this publication and any liabilities arising from them remain the responsibility of the authors.
The designations employed and the presentation of material in this publication do not imply the expression of any opinion whatsoever on the part of the European Association for Animal Production concerning the legal status of any country, territory, city or area or of its authorities, or concerning the delimitation of its frontiers or boundaries.
The publisher is not responsible for possible damages, which could be a result of content derived from this publication.

Other sponsors Asociación Española de Leguminosas Provital Carburos Metálicos Organising committee A. Brenes, CSIC, Instituto del Frío, Dpto. de Metabolismo y Nutrición, José Antonio
Novais, 10, Ciudad Universitaria, 28040 Madrid, Spain C. Burbano, INIA, SGIT, Dpto. de Tecnología de Alimentos, Carretera de La Coruña,
Km 7,5, Aptdo. 8111, 28040 Madrid, Spain C. Cuadrado, INIA, SGIT, Dpto. de Tecnología de Alimentos, Carretera de La
Coruña, Km 7,5, Aptdo. 8111, 28040 Madrid, Spain M.M. Pedrosa, INIA, SGIT, Dpto. de Tecnología de Alimentos, Carretera de La
Coruña, Km 7,5, Aptdo. 8111, 28040 Madrid, Spain M. Muzquiz, INIA, SGIT, Dpto. de Tecnología de Alimentos, Carretera de La Coruña,
Km 7,5, Aptdo. 8111, 28040 Madrid, Spain A. Varela, INIA, SGIT, Dpto. de Tecnología de Alimentos, Carretera de La Coruña,
Km 7,5, Aptdo. 8111, 28040 Madrid, Spain Scientific committee A. Brenes, CSIC, Instituto del Frío, Departamento de Metabolismo y Nutrición. José
Antonio Novais, 10. Ciudad Universitaria. 28040 Madrid, Spain C. Burbano, INIA, SGIT, Dpto. de Tecnología de Alimentos, Carretera de La Coruña,
Km 7,5, Aptdo. 8111, 28040 Madrid, Spain C. Cuadrado, INIA, SGIT, Dpto. de Tecnología de Alimentos, Carretera de La
Coruña, Km 7,5, Aptdo. 8111, 28040 Madrid, Spain G.D. Hill, Plant Sciences Group, PO Box 84, Lincoln University Canterbury, New
Zealand A.J.M. Jansman, Animal Sciences Group, Division of Nutrition and Food,
Edelhertweg 15, PO Box 65, 8200 AB Lelystad, The Netherlands M. Muzquiz, INIA, SGIT, Dpto. de Tecnología de Alimentos, Carretera de La Coruña,
Km 7,5, Aptdo. 8111, 28040 Madrid, Spain A.F.B. van der Poel, Animal Nutrition Group, Wageningen University & Research
Centre, Marijkeweg 40, 6709 PG Wageningen, The Netherlands L.A. Rubio, CSIC, Estación Experimental del Zaidín, Unidad de Nutrición, Profesor
Albareda 1, 18008 Granada, Spain


Contents Preface 1 Opening session Opening address
A. Cazorla 5Words of welcome
M. Muzquiz 7
Session I. Analysis of antinutritional factors in legume seeds and oilseeds
Recent advances in analytical methods for identification and quantification of
phenolic compounds. M. Stobiecki and H.P.S. Makkar 11
Phenolic compounds and pyrimidine glycoside determination in Vicia narbonensis seed. M. Arias, L.T. Ortiz and M. de los Mozos 29
Evaluation of antinutritional factors and nitrogen compounds of selected pea cultivars (Pisum sativum L.). N. Ariza, M.A. Martín-Cabrejas, R.M. Esteban, E. Mollá and F.J. López-Andréu 35
The effect of cyanogenic glucosides of seed of Stizolobium deeringianum from Tabasco, Mexico. L. Barrientos, J. Vargas, F. Navarro, M. Ruíz, F. López-Dellamary, A. Rodríguez and J. Pineda 39
Determination of the antinutritional factors in the seed of tropical grain legumes with potential for human and animal feeding. M.F. Díaz, M.A. Martín–Cabrejas, N. Ariza, F.J. López–Andreu, L. Jaime and C. Vidal–Valverde 43
Using near infrared reflectance spectroscopy (NIRS) for ANFs analysis in oilseed Brassicas. R. Font, M. del Río and A. de Haro-Bailón 49
Phytate and α-galactoside content of wild lupins from Jalisco, México. P.M. García-López, C.M. Gurrola Díaz, P. Garzón de la Mora, M.L. Isaac Virgen, M.A. Ruiz Lopez, C. Cuadrado, C. Burbano, M.M. Pedrosa and M. Muzquiz 53
Variation in alkaloid type and content during Erythrina americana seed development. R. García-Mateos, M. Soto-Hernández, M. Martínez 57
Antinutritional factors in different oilseeds usable as renewable resources compared with rapeseed. B. Matthäus 63

Flatulence-producing galactosyl cyclitols D-chiro-inositol fraction in maturing yellow lupin seed. A.I. Piotrowicz-Cieślak 69
Flatulence-producing oligosaccharides and galactosyl cyclitols in maturing Andean lupin (Lupinus mutabilis) seed. A.I. Piotrowicz-Cieślak, D.J. Michalczyk and R.J. Górecki 73
A modified trypsin inhibitor affinity chromatography assay for industrial application. I. Wolters, J. de Groot, H. Boer and A.F.B. van der Poel 77
The correlation between taste and structure of lupin alkaloids. W. Wysocka and J. Jasiczak 81
Session II. Mechanism of action of antinutritional factors in
legume seeds and oilseeds The mode of action of ANFs on the gastrointestinal tract and its microflora.
A. Pusztai, S. Bardocz and M.A. Martín-Cabrejas 87Lupinus campestris seed as a source of compounds with antimutagenic activity.
C. Jiménez, G. Loarca-Piña and G. Dávila 101Decreased levels of heat shock protein 70 and 90 in gut epithelial cells after
exposure to plant lectins. J. Koninkx, H. Ovelgönne, A. Pusztai, S. Bardocz, S. Ewen, H. Hendriks and J. van Dijk 107
Physiological mechanisms in the impairment of nutrient digestion and transport capacity in the salmon intestine by soybean antinutrients. Å. Krogdahl and A.M. Bakke-McKellep 111
Influence of lectins isolated from soybean seed and evening primrose seed and cake on rat metabolism. H. Leontowicz, M. Leontowicz, J.J. Czerwiński, H. Kostyra and R. Krzemiński 115
Session III A. Beneficial effects of antinutritional factors in
human nutrition Beneficial (antiproliferative) effects of different substances.
F.M. Lajolo, M.I. Genovese, I.F. Pryme and T.M. Dale 123Investigation of legume seed protease inhibitors as potential anti-carcinogenic
proteins. A. Clemente, D.A. MacKenzie, I.T. Johnson and C. Domoney 137
Assessment of the potential of legume lectins to act as a mucosal adjuvant. E. Gelencser, G. Grant and D. Kelly 143
A modelling approach to develop an antinutrients food databank with human nutrition implications. Mª Joyanes 149
Beneficial outcome in hypercholesterolemia after intake of heat treated chickpea in rats. M.A. Zulet, M.T. Macarulla, M.P. Portillo and J.A. Martínez 153

Session III B. Negative effects of antinutritional factors in human nutrition
Storage proteins: physiological and antigenic effects.
L.A. Rubio, J. Rodríguez, C. Fernández and J.F. Crespo 159 Morphological changes of rat brain neurons after intracranial administration of
the alkaloid gramine. A. Hernández-Torres, J. Bañuelos-Pineda P.M. García-López, M.A. Ruiz-López and P. Garzón de la Mora 177
Plasma antibody responses to grain legume proteins in weaned piglets. J.P. Lallès, P. Salgado and J.P.B. Freire 181
The nutritional potential of Mexican piñon (Jatropha curcas). Toxic and antinutritional factors. J. Martínez-Herrera, L. Chel-Guerrero and A.L. Martínez-Ayala 185
Immunodetection of legume proteins resistant to digestion in weaned piglets. P. Salgado, J.P.B. Freire and J.P. Lallès 189
Session IV. Effects of antinutritional factors on monogastric
animals Recent progress on research on the effects of antinutritional factors in legume
and oil seeds in monogastric animals. A. Brenes, A.J.M. Jansman and R.R. Marquardt 195
The cholesterol lowering effect of lupin meal in chicken diets. A. Brenes, I. Arija, C. Centeno and A. Viveros 219
Thermal processing of rapeseed meal: nutritional evaluation in digestibility experiments with pigs. R. de Schrijver, S. de Vos and J. Vande Ginste 225
Microbial protease addition to a soybean meal diet for weaned piglets: effects on performance, digestion, gut flora and gut function. N. Dierick, J. Decuypere, K. Molly and E. Vanderbeke 229
Rate of muscle protein synthesis in rats fed raw and extruded faba bean diets. F. Marzo, E. Urdaneta, J. Barrenetxe, F. Ibañez and S. Santidrian 235
Effect of carob (Ceratonia siliqua L.) seed in broiler chick diets on nutrient digestibility and intestinal viscosity. L.T. Ortiz, M.L. Rodríguez, C. Alzueta, A. Rebolé, C. Centeno and J. Treviño 239
In vivo and in vitro ileal digestibility of protein and amino acids of peas containing different tannin levels. E. Święch, L. Buraczewska and M.Taciak 243
The effect of trypsin inhibitor level in soy products on in vitro and in vivo (pigs and rats) protein and amino acid digestibility. E. Święch, L. Buraczewska and M.Taciak 247
Nutritional value and physiological effects of industrial soybean products differing in protein solubility and trypsin inhibitor content. M. Taciak, E. Święch and B. Pastuszewska 251

Intestinal enzymatic activity and amino acid uptake in brush border membrane vesicles of rats fed extruded kidney bean (Phaseolus vulgaris). E. Urdaneta, J. Barrenetxe, P. Aranguren, S. Santidrian and F. Marzo 255
Session V. Application of biotechnology and processing to
reduce ANFs and to increase nutritional quality. Potential use of GMOs
A. The potential of genetically modified legume and oilseed crops for food and
non-food use. R. Greiner and C. Domoney 261
α-Galactosides in Catalan landraces of beans (Phaseolus vulgaris L.) and location, candidates for protected designation of origin. L. Bosch, F. Casañas, M. Pujolà, E. Centelles, P. Beltran, A. Florez and F. Nuez 277
Facile breeding markers to lower contents of vicine and convicine in faba bean seeds and trypsin inhibitors in pea seeds. G. Duc, P. Marget, D. Page and C. Domoney 281
Identification of molecular markers tightly linked to low tannin and vicine-convicine content in faba beans. N. Gutierrez, G. Duc, P. Marget, C.M. Avila, M.J. Suso, J.I. Cubero, M.T. Moreno and A.M. Torres 287
B. Technical and biotechnological modifications of antinutritional factors in
legume and oilseeds. Gy. Hajos and A.U. Osagie 293
Modifications of seed storage proteins during germination and seedling growth of faba bean cotyledons. C. Cuadrado, E. Guillamón, C. Goyoaga, M.M. Pedrosa, P. Altares, C. Burbano, M. Muzquiz and C. Romero 307
Effects of exogenous enzymes on the content of bioactive compounds in lentils and peas. M. Dueñas, T. Hernández & I. Estrella 311
Content and distribution of vicine, convicine and L-DOPA throughout the germination and seedling growth of Vicia faba L. seed. C. Goyoaga, C. Cuadrado, M.M. Pedrosa, E. Guillamón, P. Altares, M. Muzquiz and C. Burbano. 317
Detoxification of rapeseed meal by controlled instantaneous pressure drop. J. Haddad and K. Allaf 323
The inactivation of soybean trypsin inhibitors by hydro-thermo-mechanical treatment. J. Haddad and K. Allaf 329
Changes of phytate content of Lupinus albus and L. mutabilis seed during controlled instantaneous pressure drop treatment. J. Haddad, R. Greiner and K. Allaf 333

Effect of aqueous, acid and alkaline thermal treatments on protein quality of Lupinus campestris seed. C. Jiménez-Martínez, R. Mora-Escobedo and G. Dávila-Ortíz 337
Elimination of quinolizidine alkaloids, α-galactosides and phenolic compounds from Lupinus campestris seed via aqueous, acid and alkaline thermal treatment. C. Jiménez-Martínez, M.M. Pedrosa, M. Muzquiz and G. Dávila-Ortíz 343
Limited phytate hydrolysis by endogenous phytase of sesame seeds during germination. K. Żyła, T. Fortuna, M. Mika and M. Czubak 347
Closing session Grain legumes and oilseeds – the way ahead.
G.D. Hill 353Closing comments
A. Brenes 365 Author index 367 Keyword index 369


1
Preface This book contains the proceedings of the fourth International Workshop on Antinutritional Factors in Legumes and Oilseeds held from 8-10 March 2004 in Toledo Spain. Again the workshop has focused on aspects related to the presence, effects and inactivation, by processing, or by plant breeding, of the antinutritional factors (ANFs) in legume seed (particularly soy, lupin, pea, bean, faba bean and lentil) and in other oilseeds. Presentations from Cuba and Mexico have extended both the range of legumes considered and have introduced new genera for our consideration.
As in the past the papers presented cover methods of chemical analysis of the ANFs, their content in seed and their mode of action in animals and in humans. From there the papers consider their inactivation by processing, plant breeding and biotechnology. At this meeting we have for the first time a specific section on the potential beneficial effect of the consumption of ANFs, something that was hinted at at our last meeting in Wageningen in 1998.
Each session was opened by a major review paper, which covered developments since the last workshop. Although previous workshops had considered allergic reactions in domestic livestock this time there is a major review on the role of legumes in the development of allergies in humans. The inclusion of all oilseeds has also provided an opportunity to review crops such as cottonseed and linseed. We also have a contribution that considers the effects of ANFs in fish digestion, in the area of aquaculture.
More than fifty papers were submitted and have been included in these proceedings. On behalf of the organising committee I wish to acknowledge the financial support of
the organisations and companies listed in the proceedings. Also on behalf of the other members of the committee I would like to give special
thanks to George Hill of Lincoln University, New Zealand for his work in reviewing all of the papers, his final editing of the proceedings and his concluding summary and recommendations for the future.
On behalf of the organising and scientific committees, M. Muzquiz Madrid, March 2004


Opening session


5
Opening address A. Cazorla Director General, Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria, Carretera de La Coruña Km 7,5, 28040, Madrid, Spain On behalf of INIA it gives me great pleasure to welcome you all to this 4th International Workshop on Antinutritional Factors in Legumes and Oilseeds, which is being hosted by the first time in Spain in this beautiful and historic city of Toledo.
In Spain we have a major interest in the nutritional quality of legume seed in human diet. Dry legume seed plays a major part in Spanish cuisine and chickpea, faba bean, lentil, peas and beans in many coloured forms are still regularly eaten despite the inroads of ventures like McDonalds and Pizza Hut. The grain legumes are important in Spain and in other Mediterranean countries. However, there is a considerable part of the population of the world that depend on the grain legumes as a major source of their daily dietary protein intake. To all of these people legumes are of vital interest.
Besides the contribution legume seed makes to human diet they are also a major part of animal rations for both monogastrics and ruminants. Following the problems experienced in Europe with ‘Mad Cow Disease’ and BSE the use of animal meals for animal feeding in the EU has been banned. In Europe, even before the ban, there was a major shortfall in high protein animal feeds. The ban has made matters worse and as a result we need to look at new potential sources of vegetable protein. It is interesting to note that the proceedings of this workshop includes papers from Cuba and Mexico which look at legume and other plant species that have not been widely used as animal feeds.
At INIA we have a major interest in the nutritional value of legumes, particularly in the area of human nutrition because of their major importance in the Spanish diet. Besides our direct research in this area we have co-ordinated a network, which involves a number of other research organisations, universities, hospitals and commercial enterprises to integrate work in Spain on these valuable protein sources. We welcome the representatives of many different countries of the world to this fourth meeting on this topic. At INIA we are pleased to note that the holding of this meeting in Spain has attracted a number of presentations from different part of Iberoamerica a region which we also have research collaboration. We also have close linkages with parts of eastern Europe such as the Polish Academy of Science and there are a number of papers from Polish research organisations, which we work with.
In conclusion I again welcome you to this Workshop. I hope that the next two and a half days are both pleasant and scientifically fruitful. That you renew old friendships and that new associations are made and finally that you take away fond memories of your time in Toledo.


7
Words of welcome M. Muzquiz INIA, SGIT, Dpto. de Tecnología de Alimentos, Carretera de La Coruña, Km 7,5, Aptdo. 8111, 28040 Madrid, Spain On behalf of the organising committee I am pleased to welcome you to the 4th International Workshop on Antinutritional Factors in Legume Seeds and Oilseeds held in Toledo, Spain.
After the workshops in 1988, 1993 and 1998 in Wageningen, The Netherlands, which were very successful, the Organizing Committee of the previous Workshops proposed that I organise the present workshop in Spain.
The average Spanish diet is considered as a model, which is being followed by other countries, mainly because of the great variety of products in our regions, which are consistent with the good nutrition. Today, Spanish gastronomy and nutrition can be included in what is known as the ‘Mediterranean Diet’, which means something more than a healthy and well balanced diet. The grain legumes are one of the most relevant components of this diet and their consumption has always had a very important socio-economical role in Spain.
From a nutritional point of view the presence of both protein and starch in adequate proportions, along with fibre, vitamins, and microelements, has made the legumes a focus of justified nutritional interest. However, in these rich sources of vegetable protein, there are a series of compounds, generally known as antinutrients, that impede the digestion of some of their most interesting components, and in some cases they are simply toxic or cause undesirable physiological side effects (e.g. flatulence).
However, it has recently been discovered that, in small quantities, they can have a beneficial effect on health, aiding the prevention of illnesses such as cancer and coronary disease. As a result they are now being called non-nutritional compounds because although they have no direct nutritional value they are not always harmful. From a biochemical point of view the non-nutritional compounds are very varied, they do not appear equally in all pulses and their physiological effects differ.
As can be seen from the program this workshop is somewhat different from that of the previous workshops. We, as organizing committee, thought that emphasis in the program on the ANFs in oilseed as well as those in legume seed, would benefit the scope and interest in the workshop. Also, more attention is given to the mode of action and beneficial and negative effects of ANFs in human nutrition.
As in the past, each session of the program is introduced by an invited review paper, which gives an overview of a particular aspect of ANFs, and mainly emphasises recent developments in the topic since the last ANF workshop in 1998. These main papers are followed by short papers presented orally and by contributions presented as posters.
The program contains seven main papers and about fifty short papers and posters. An essential part of the program is the group discussions on the last day of the workshop. This will provide an excellent opportunity to discuss further new issues, which will arise during the workshop and to bring forward our own ideas and new items, which can provide guidelines and directions for future research in this area.
As the organising committee we are proud to have the book of the proceedings ready for the start of the workshop. This would not have been possible without your assistance in meeting our deadlines and we thank you for your cooperation.
Finally, we hope the Workshop is of benefit to you, both from a scientific point of view and the personal experience of being in Toledo, one of the oldest cities in the Iberian

8
Peninsula with a history of more than two thousand years. Please, enjoy yourselves during your time in our country.

Session I Analysis of antinutritional factors in legume seeds and oilseeds


11
Recent advances in analytical methods for identification and quantification of phenolic compounds M. Stobiecki1 & H.P.S. Makkar2 1Institute of Bioorganic Chemistry PAS, Noskowskiego 12/14, 61-704 Poznań, Poland 2Animal Production and Health Section, Joint FAO/IAEA Division of Nuclear Techniques in Food and Agriculture, International Atomic Energy Agency, Vienna, Austria Summary Phenolic compounds are one of the largest groups of secondary metabolites present in the Plant Kingdom. This class of natural products has a wide range of biological activity, ranging from beneficial to adverse, in both humans and animals. Recent advances made in the analysis of simple phenolics and polyphenolics (tannins) are reviewed.
For simple phenolic compound mixtures, choice of a separation and identification method is dependent on factors such as the physico-chemical properties (polarity, thermal stability and molecular weight) and the number of components in the mixtures. Gas chromatography is the method of choice for low molecular weight aromatic acids, alcohols and simple flavonoid aglycones. More polar and thermally unstable compounds, mainly glycosidic conjugates of phenolic acids and flavonoids, are separated well on reversed phase liquid chromatography (LC) columns. The most flexible separation method is capillary electrophoresis, where the highest sensitivity and separation can be achieved. Many different detectors [ultra violet (UV), mass spectrometry (MS), nuclear magnetic resonance (NMR), electrochemical, laser induced fluorescence (LIF)] are used for the identification of compounds eluted from the chromatographic columns. The UV and MS detectors are used widely due to their efficient provision of information, their high sensitivity and ease of use.
Tannin analysis remains highly problematic. Reversed-phase high-pressure liquid chromatography (HPLC) has been used for separation of low molecular weight polymers. However, this technique has not been particularly useful for complex hydrolysable tannins (HT) and condensed tannin (CT) polymers beyond tetramers. The UV detection method is widely used because of its simplicity. However, it is not specific due to interference by other moieties present in plant extracts. Fluorescence and electrochemical detection systems can also be used. Structural information can be generated using MS and NMR approaches. Most tannin-containing plants contain both HT and CT, but unfortunately usually only CT (proanthocyanidins), are analysed. Furthermore, the biological effects are ascribed mostly to the CT, which might be misleading. There are two new assays for HT. These are based on their hydrolysis to gallic acid and then its determination, using HPLC. The second is based on the methanolysis of HT to methyl gallate and then its determination using potassium iodate; protein precipitation assays, both isotopic (based on 125I-labelled bovine serum albumin) and non-isotopic, representing the operational property of both HT and CT. There is a tannin bioassay based on an in vitro simulation of the rumen and measurement of tannin activity for both free and bound tannins in terms of rumen fermentation parameters; a 14C-polyethylene glycol binding assay; and a 13C-NMR-based assay will also be presented.
Keywords: chromatography, mass spectrometry, ionisation, phenolic compounds, tannins

12
Introduction The composition of plant secondary metabolites in tissues of agriculturally important crop plants is influenced by both biological (interaction with symbiotic and pathogenic microorganisms or insects) and physical (temperature, draught, UV light irradiation) stimuli (Dixon & Paiva 1995; Bednarek et al., 2003). In the last decade, the development and use of genetically modified plants has received considerable attention. Changes in the genomes may also influence the profiles of different secondary metabolites due to the introduction of new genes and the over expression or suppression of existing ones (Fiehn, 2001; Bovy et al., 2002; Sumner et al., 2003).
Various classes of phenolic plant secondary metabolites are of increasing concern because of their diverse biological roles. These plant metabolites, when consumed, may affect the health of individuals, due to their phytoestrogenic (Beck et al., 2003; Boue et al., 2003), anti-oxidant (Shirley, 1996; Fritz et al., 2003) and anticarcinogenic (Dixon & Steel, 1999; Gerhauser et al., 2003) activity.
The chemical structure of polyphenols is diverse, due to: (1) the pattern of hydroxyl groups substitution, (2) the placement of the flavonoid B phenolic ring and (3) sugar substitution on the aglycones, and different possibilities of isomerization in the aglycone and glycosidic moieties (Fig. 1). At present, over flavonoid 4,000 structures and their conjugates are known. These diverse structural differences affect physico-chemical properties like polarity, thermal stability and molecular mass of these substances. These parameters influence the choice of the analytical methods used for the identification and/or structural analysis of this class of natural products.
During the last decade, exciting developments have occurred in the field of instrumentation. These have made methods for the separation of a chemical compound from a mixture and its identification, structural characterization and quantification highly efficient and sensitive. This has widened and strengthened the analytical capabilities of research and industrial laboratories. The methods can be classified as direct that do not require any modification of the compound; and indirect, for which chemical treatment of the material is often necessary. The former methods are easier and require fewer resources. However, unfortunately, not all classes of compounds can be analysed using this approach because of their complicated chemical structures. Hydrolysable and condensed tannins, due to their high degree of polymerisation, are in this category.
Three aspects have to be taken into account when a choice of instrumental methods is being considered for the identification and characterisation of compounds present in a complicated mixture. These are: speed of analysis, selectivity and sensitivity. At the present stage of analytical method development it is not possible to achieve all determinations with one set of instruments. However, by using different kinds of detectors on line (UV diode array, MS and/or NMR) with chromatographic methods, the selectivity and in some cases the sensitivity of the analysis can be improved (Niessen, 1999; Gelpi, 2002).
For the identification and characterization of natural phenolic products in mixtures, the most suitable detector is the mass spectrometer linked to chromatographic instrumentation (gas and liquid chromatography or capillary electrophoresis). Application of different kinds of ionisation methods and analysers is possible (Fig. 2) with mass spectrometers joined to chromatographic systems.
In these instruments, different physico-chemical effects are utilized both for the ionisation and separation of the ions created. On one hand, the amount of internal energy in ionised molecules plays an important role due to their strong influence on the degree of fragmentation; and on the other hand the resolution achieved during separation of the ions

13
created also has impact on the amount of information obtainable from the resultant mass spectra. The proper choice of chromatographic method and mass spectrometer type is very important for the efficient detection of target compounds in a mixture. In some cases, when gas chromatography is applied, chemical modification of the compounds is necessary to enable their evaporation and separation. However, some rearrangement reactions during derivation may cause structural changes of the compounds being investigated.
O
OH
HO
OH
R
O
O
OH
HO
R
OHO
O
OH
HO
OH
OH
R+ O
OH
HOR
OH
OH
O
OH
HOR
OH
OH
OH
OH
RHO
OH
O
HO O
OHRR
OH
R
O
HO
OH
O
O
R
OH
OH
HO
OH
OA C
B
- possible glycosilation side
R = H, OH
(3)(2)
(5) (6)
(8)(7)
(4)
(1)
Figure 1. Chemical structures of main classes of flavonoids present in plant material:
(1) flavan, (2) flavone, (3) flavonol (places of possible glycosilation indicated), (4) isoflavone, (5) anthocyanidin, (6) flavan-3-ol (7) phenylpropanoid acid and (8) proanthocyanidin type B (condensed tannin - CT).
The amount of structural information obtained from a mass spectrum depends on the
ionisation method used. The highest energy transfer occurs during electron ionisation (EI) and in these cases fragmentation of molecular ions may be seen. When other ionisation methods: chemical ionisation (CI), electrospray (ESI) and atmospheric pressure ionisation (APCI) (also called “soft ionisation” methods) are applied in GC/MS and/or LC/MS and EC/MS systems, the fragmentation of compounds is not common. An important achievement in mass spectrometric technology is collision induced dissociation tandem mass spectrometry (CID MS/MS). During CID MS/MS analysis, precursor ions (with a defined m/z value)

14
extracted in the first analyser collide with atoms of an inert gas in the collision cell, and the ionised fragments created (product ions) in this process are separated in the second analyser (Jennings, 1996). In the last couple of years a multistage tandem mass spectrometry (MSn) instrument with an ion trap analyser has been developed. In this analyser the fragment ions created can be further studied using an additional MSn stage. However, the fragmentation mechanisms are different during high- and low-energy collisions used in electro-magnetic or quadrupole and ion trap analysers. It has been shown that low-energy CID MS/MS spectra of C-glycosidic flavonoids differ from those obtained when high-energy measurements are performed (Waridel et al., 2001).
Control and datacollection
Sample introduction
Scheme of mass spectrometer
Ionization Analyzer Detector
directGCLCEC
EICIES
APCIMaldiFAB
LSIMS
B/E or E/BQ
Ion trapTOF
FT ICR
LOQ – 10-9 - 10 –12 mol, LOD – 10 –18 mol
Figure 2. Block diagram of mass spectrometer with description of the consecutive parts.
LOQ = limit of quantification, LOD = limit of detection. Mass spectrometric approach for the identification and structural analysis of phenolic natural products Mass spectrometry, due to its sensitivity, is the method of choice for the identification and structural analysis of phenolic conjugates isolated from plant materials. However, various kinds of biological activity of different classes of natural products makes it necessary to monitor these compounds and their metabolites in food and feed products or in organisms. Below are some examples, where mass spectrometric techniques have been used for the analysis of a single compound after its isolation from biological materials of different origins. Mass spectrometric analysis of single compounds The structural characterization of flavonoid glycosides, isolated as pure compounds from plant material, is possible using different mass spectrometric strategies (Claeys et al., 1996; Frański et al., 1999). The molecular mass of the compound under investigation can be estimated using soft ionisation methods (fast atom bombardment - FAB, ESI, APCI). In most cases some fragment ions are observed in the mass spectra, enabling the estimation of the mass of aglycone and the size of sugar rings (Stobiecki, 2000). Additional information on the substitution pattern of sugars on the aglycone can be obtained using CID MS/MS.

15
Differentiation of sugar ring substitution on the aglycone and the placement of interglycosidic linkage in diglycosides (1-6 or 1-2) is also possible (Cuyckens, et al., 2000,2001, 2002; Ma, et al., 2001; Bylka at al., 2002; Sikorska et al., 2003). In some cases, analysis of the MS/MS spectra also enables the structure of the aglycone to be determined (Claeys et al., 1996; Huck et al., 2001; Vanhoenacker et al., 2001). Structural information about the compounds may also be available when simple chemical modifications (methylation followed by methanolysis in 1N HCl and a second derivation step is made with different groups (deuterated methyl, acetyl or trimethylsilyl)) are applied and a GC/MS analysis of the products produced is performed. From the MS spectra and the retention times, it is possible to identify sugars and linkages between sugars (placement of the glycosidic bond), the aglycone and the substitution of sugar on the phenolic moiety (Frański et al., 1999, 2002) (Fig. 3).
- identification ofaglicone and sugar
- placement of sugar onaglicone
- aglycone and sugaridentification
Flavonoid glycosides
CIDMS/MS
CIDMS/MS
Methanolysis
Acetylation
MethylationCH3/CD3
Hydrolysis TFA
Reduction MetylationCH3/CD3
MethylationCH3/CD3
GC/MSGC/MS
GC/MSGC/MS
Acetylation
- placement of sugar onaglicone
- glycosidic linkage position
- sugaridentification
- confirmation of molecular mass
- differentiation O-di-glicosides from di-O-glycosides- confirmation of native methyl grouppresence
- molecular mass- mass of aglycone and sugar
- sugar sequence-differentiation C-and O- glycosides
Figure 3. Mass spectrometric approach for the structural analysis of flavonoid conjugates. Gas chromatography - mass spectrometry Gas chromatography - mass spectrometry is a well established method. For more than two decades it has been possible to couple a capillary GC column directly to the MS ion sources. This arrangement achieves a high sensitivity analysis. Electron and chemical ionisation (EI and CI respectively) methods are applied in the GC/MS systems for routine studies. An important limitation of this method is the volatility of the target compounds. Most of phenolic acid derivatives and flavonoids have to be substituted on their polar groups (hydroxyl and/or carboxyl) prior to injection into GC/MS systems to increase their volatility. Trimethylsilyl (TMS) and methyl groups (CH3) are often used for the determination. In some cases it is necessary to introduce deuterium atoms into the methyl groups (CD3) in order to distinguish between native methyl groups, present in the molecule being studied, and those, which were introduced during chemical treatment. Chemical treatment may cause some rearrangements in the molecules produced, especially during methylation of different classes of flavanones. However, EI mass spectra for the methyl derivatives of flavonoids provide

16
more structural data than for the trimethylsilyl derivatives (Stobiecki & Wojtaszek, 1993; Bednarek et al., 2001).
In the case of phenolic glycoconjugates, especially glucoronides of simple phenols, for example, catechols conjugates present in physiological fluids, utilization of less energetic chemical ionisation allows molecular mass information to be obtained from mass spectra from GC/MS runs (Stobiecki et al., 1984). This class of compounds may often be identified in physiological fluids during studies on the metabolism of natural products in the human and animal gut. Additional chemical treatment (hydrolysis or methanolysis) may be necessary to obtain information on the aglycone and sugar parts of the compounds under investigation.
Different analysers may be utilized in GC/MS systems, the most popular being the low-resolution quadrupols and ion traps. In the last few years, the application of time of flight (Tof) analysers has received considerable attention. Hyphenation of a gas chromatograph with Tof MS analyser assures the collection of high-resolution data for the ions produced, which allows the elucidation of the elemental composition of the molecular and fragment ions. Fiehn (2001) demonstrated the utility of GC/MS Tof instrumentation for plant metabolism studies. This instrumental approach can provide additional structural information about isomeric natural products under investigation. Liquid chromatography - mass spectrometry Introduction of new “soft ionisation” methods revolutionised the application of mass spectrometric analysis of the compounds present in the eluent from a column mounted on a liquid chromatograph. A liquid chromatograph coupled to a mass spectrometer with electrospray (ES) or atmospheric pressure chemical ionisation (APCI), because of easiness of its utilization, has been widely used for qualitative and later quantitative analysis of different classes of natural products in samples extracted from plant tissues. Both positive and/or negative ions mass spectra are registered during the analysis of flavonoids and their conjugates. However, the composition of the mobile phase used for separation of the components on the LC column influences response linearity, detection limits and precision (Careri et al., 1999; Zhao et al., 2002).
When an LC/MS system is used, a few important chromatographic parameters, which need to be considered, are: column diameter and flow rate, composition of the liquid phase and its gradient. The first two parameters influence sensitivity and the third affects the resolution achieved on the column and performances of the mass analyser. The application of solid components in the liquid phase will cause rapid contamination of the ion source and only the volatile components of the liquid phase can be utilized in LC/MS systems. At high flow rates it is sometime necessary to split the column eluate prior to its introduction into the capillary in the ESI source. In some cases, to obtain additional structural information, hydrolysis of the plant extracts followed by GC/MS analysis of the products obtained is necessary (Fig. 4). The approach based on the EI mass spectra of the methylated derivatives permits differentiation of the isomeric structures of the aglycones present in the samples. For example, kaemferol - flavonol and orobol - isoflavon have been characterized in extracts of white lupin (Lupinus albus) after methanolysis and methylation of the extract sample (Bednarek et al., 2001).
Many papers describe applications of LC/MS systems with different ionisation methods (CF FAB – continuous flow fast atom bombardment, ESI and APCI). Papers about LC/MS systems which possess the ability for tandem mass spectrometric analysis for the study of free flavonoids and their glycosides in the extracts, obtained directly from different plant tissue, have been published in the last decade. These classes of metabolites were studied in fruit,

17
seed and roots or green parts of plants (Sumner et al., 1996; Justesen et al., 1998; Lin et al., 2000; Klejdus et al., 2001; Bednarek et al., 2001; de Rijke et al., 2001; Gu & Gu, 2001; Tomas-Barberan et al., 2001; Frański et al., 2003). Other papers report research on plant based food components (Mellenthin & Galensa, 1999; Friedrich et al., 2000; Vanhoenacker et al., 2001; Sanchez-Rabaneda et al., 2003) and the presence of flavonoid metabolites in physiological fluids (Li et al., 2001; Blaut et al., 2003). In most of these publications the application of a double detection system where UV and MS detectors are coupled on line to an LC column, has been used (Fig. 5). An important aspect of LC/MS analysis is sample preparation. In some of the above studies, plant tissue extracts were subjected to direct analysis. In others solid phase extraction (SPE), mainly on reversed phase silica gel, preceded LC/MS analysis. In some cases, preconcentration of the group of target metabolites may improve analysis sensitivity. Utilization of an UV diode array together with a MS detector increases the structural information available. On the basis of the UV spectra, it is possible to differentiate many classes of flavonoids; for example, isoflavones are distinguishable from flavonols and flavones. Use of MS detection also permits evaluation of the degree aglycone hydroxylation (Fig. 5). It is also possible to arrange a system, where an NMR instrument is coupled to a LC column. The application of this triple detector system has been demonstrated by Wolfender et al. (1997).
Sample
HPLC/DAD
Deacetylation
Retention time UV Spectra
LC/MS
Methanolysis,Methylation
MethylationMethanolysis,
Acetylation
GC/MS
ESI/MS EI/MS
MW of glycosidesand aglycones.
Differentiationof flavonoid
classes.
Comparisonwith standard
compounds
Identification of aglyconesand placement
of glycosidic bond.
Target compounds identification
Figure 4. Structural information obtainable from LC/MS and/or GC/MS analyses of extracts
from plant tissues. Capillary electrophoresis - mass spectrometry Applications of different techniques of capillary electrophoresis combined with a mass spectrometer CE/MS have been described by Niessen (1999) in his book. Three CE techniques were given for the separation of natural products in plant extracts. These are: capillary zone electrophoresis (CZE), isotachophoresis, and micellar electrokinetic chromatography (MEKC). Three different coupling systems have been applied in CE/MS instruments: direct CE-MS coupling, coaxial coupling, and liquid junction. There are some advantages of capillary electrophoresis over liquid chromatography. These are the shorter

18
time necessary for a single analysis and better resolution of sample components. However, the efficient utilization of CE combined to a mass spectrometer needs a highly experienced operator because of the very low flow in the capillary. Additional problems can arise due to the presence of non-volatile buffer components, such as additives like cyclodextrines and surfactants, which can lead to an intolerable decrease of sensitivity and background. Their presence in the buffer does not allow the production of high quality mass spectra and a rapid contamination of ion source can occur. Over the past few years, capillary electrophoresis has been extensively used for analysis of polyphenols, but in most of these applications detectors other than MS detectors have been used due to problems with non-volatile components in the buffers. The use of CE/MS analysis has been limited and so far; only a small number of publications have appeared using this methodology (Tomas-Barberan & Garcia-Viguera, 1997; Vanhoenacker et al., 2001; Urbanek et al., 2002). Tannin assays A number of different tannin assays are available. The methods generally used are categorized into three groups: chemical, protein precipitation, and other. Chemical methods The most commonly used procedures in this category are redox reactions (Folin-Ciocalteu, Folin-Denis or Prussian blue methods), the vanillin assay, the metal complexing assay, and the acid butanol assay with and without addition of iron (Hagerman et al., 1997). Each method measures different types of tannins based on the chemistry of the reaction between phenols and the reagents used. Total phenols: The Folin-Denis, Folin-Ciocalteu or Prussian blue methods are used to measure total phenols. These assays are based on the oxidation of the phenolic analyte and reduction of the reagent to form a chromophore. The presence of reducing agents can interfere with this assay. Another method, commonly known as the ferric chloride assay is based on the formation of coloured phenolic-metal ion complexes. It is useful for measuring total phenols. Interference from non-phenolics is unlikely, however, non-tannin phenolics cannot be distinguished from tannins by this method. In the Folin-Denis, Folin-Ciocalteu or the Prussian blue methods, the results are generally expressed as tannic or gallic acid equivalents (Makkar et al., 1993). Tannins: In the Makkar et al. (1993) method, tannins are distinguished from non-tannins by using a solid matrix, polyvinylpolypyrrolidone (PVPP). This method assumes that the phenolics, which bind to proteins, also bind to PVPP. Total phenols are measured in a plant extract using the Folin-Ciocalteu method before and after treatment with PVPP. The PVPP has a high affinity for tannins and therefore removal of the PVPP following treatment, using centrifugation removes tannins from the extract. The difference between total phenol values, in the extract, before and after PVPP treatment is a measure of tannins. The ferric chloride assay could also be used in place of the Folin-Ciocalteu assay for the measurement of total phenols before and after PVPP treatment. The former method is preferred because of its higher sensitivity. The method based on PVPP treatment (Makkar et al., 1993) measures total tannins. However, it provides no information on the presence or absence of condensed or hydrolysable tannins.

19
Figure 5. Ultra violet (UV) and single ion chromatograms of extracts of white lupin (Lupinus
albus) leaves. Numbers on the UV and single ions chromatograms correspond to flavonoid glycosides identified in the lupin leaf extract (Bednarek et al. Phytochemistry 56: 77-85, with permission).
Condensed tannins: The vanillin assay in methanol (Broadhurst & Jones, 1978; Price et al., 1978) has been widely used for measuring condensed tannins in sorghum (Sorghum sp.) and in beans (Phaseolus sp.). This assay measures flavan-3-ols and dihydrochalcones, which are non-tannin in addition to condensed tannins. Another disadvantage of this method is that monomeric flavans give a higher colour yield than condensed tannins. Further, proanthocyanidins based on 5-deoxyflavanols, such as profistinidin in quebracho tannin do not react at all. The presence of acetone interferes with this assay by forming a chromogen with acidified vanillin. The chromogen formed has an absorption peak at 548 nm, which produces a substantial error in the determination of condensed tannins. In addition the reproducibility of this method is not good (Makkar & Becker, 1993, 1994).
The butanol-HCl method originally proposed by Bate-Smith (1973) for the measurement of condensed tannins is simple and more specific than the vanillin assay. Later, this method was modified by the inclusion of iron in the butanol-HCl reagent, which was claimed to increase sensitivity and reproducibility of the assay (Porter et al., 1986). This method is based on the oxidative cleavage of the interflavan bonds in the presence of mineral acids in alcoholic solutions at about 95 °C to yield pink-coloured anthocyanidins, which are measured at 550 nm. The method is sensitive to the presence of water. An increase in the water content decreases the colour yield of the anthocyanidins. Tannins besides being

20
extractable exist in the bound form, as contaminants in plant fibre or protein fractions. The bound proanthocyanidins or condensed tannins are generally measured by butanol-HCl-iron reagent. The hydrolysis of bound proanthocyanidins to anthocyanidins is a prerequisite for the determination of these condensed tannins.
For accurate determination of bound tannins, samples should be freeze-dried because drying, even at temperatures as low as 50 °C, can decrease the measured values (Makkar & Singh, 1995). Even in freeze-dried samples, not all bound condensed tannins react quantitatively in the butanol-HCl reagent. This leads to an underestimation of bound condensed tannins (Makkar et al., 1999). Thus the butanol-HCl method should be used with caution as a quantitative assay. Values obtained using this method do not correlate well with the biological values of tannin-containing feeds. However, the method is a simple way of showing the presence of condensed tannins in feedstuffs.
Terrill et al. (1992) also proposed a method, based on butanol-HCl, for the determination of extractable, protein- and fibre-bound condensed tannins. The classification of condensed tannins as bound to protein and fibre by this method has not been validated and could be arbitrary and loosely categorized. In addition the nutritional significance of the values obtained for fibre and protein bound tannins has not been evaluated. The presence of hydrolysable tannins in the bound form in feedstuffs has not yet been shown, although these are present in the heartwood of Castanea sativa and in Quercus petraea (Peng et al., 1991). Other methods for the analysis of insoluble tannins include the use of 13C-NMR (Makkar et al., 1999) and thiolysis and phloroglucinol degradation (Matthews et al., 1992). These methods, because of their complexity, have not been used much.
The relative degree of polymerisation of the proanthocyanidins can be measured from the ratio of the results from the vanillin assay in glacial acetic acid (in which only terminal units react to form a chromophore) and those from the butanol-HCl assay (in which only extender units react to form a chromophore) (Butler et al., 1982). A simple method for the characterization of proanthocyanidins and the determination of the degree of their polymerisation, based on acidic degradation of proanthocyanidins with phenylmethanethiol (thiolysis) followed by HPLC (Labarbe et al., 1999; Guyot et al., 2001) could offer valuable information on structure-activity relationship of proanthocyanidins in foods and feeds. Hydrolysable tannins: An approach employed in some methods for determination of hydrolysable tannins is the conversion of the hydrolysable tannins to a common unit and determining the common units by spectrophotometric or HPLC methods. These methods are useful for simple hydrolysable tannins but may provide limited information for complex oligomeric hydrolysable tannins.
The rhodanine method (Inoue & Hagerman, 1988) determines gallotannins as gallic acid equivalent. In this method gallotannins are acid hydrolysed under anaerobic conditions to gallic acid, which is reacted with rhodanine to give a pink chromophore (measured at 520 nm). Free gallic acid, before acid hydrolysis, is also measured. This is subtracted from the total gallic acid (gallotannins plus free gallic acid) measured after hydrolysis to obtain gallotannins as gallic acid equivalents. The number of gallic acid units differs between different gallotannins, and therefore this method does not provide an absolute quantification of gallotannins. In addition, some ellagitannins are known to contain gallic acid. Measuring gallic acid before and after acid hydrolysis by HPLC could increase the specificity and sensitivity of the assays.
The sodium nitrite method (Bate-Smith, 1977) for the determination of ellagitannins, based on determining the product of hydrolysis of hexahydroxydiphenic acid esters

21
(ellagitannins) requires large quantities of pyridine as a solvent. This introduces a significant toxicity risk when the method is used for routine analyses of large sets of sample.
The potassium iodate method (Bate-Smith, 1977) measures both gallotannins and ellagitannins and is used occasionally but neither the reaction chemistry nor the specificity of the method has been established. Practical limitations of the method include an inconvenient reaction temperature (the method requires a refrigerated water bath); the need to run each reaction for a different time period to achieve maximum colour yield; the occurrence of interfering yellow oxidation products; formation of several chromophores with different lambda maxes for some plant extracts comprised of mixtures of tannins and the formation of precipitates when extracts from some plants are analysed.
Recently, Hartzfeld et al. (2002) modified the potassium iodate method to include a first step in which all of the hydrolysable tannins in the sample are converted to a single chemical species, methyl gallate. Differences in the reactivity of the parent compounds are thus overcome. Results from various laboratories can be compared directly since they are reported in terms of a commercially available standard compound, methyl gallate. The conditions for methanolysis have been established to assess the temperature and time required to maximise the yield of methyl gallate. The temperature for methanolysis has been fixed at 85 oC to avoid the disappearance of some methyl gallate and the formation of unidentified phenolic products. For routine analysis, 20 h has been suggested as a convenient time for methanolysis. In addition, the reaction conditions between methyl gallate and potassium iodate have been optimised to form a stable chromogen. A similar approach has been used (Lei et al., 2001) in which anhydrous methanolic HCl has been used to release ellagic acid and gallic acid (as methyl gallate) followed by the determination of individual moieties using HPLC. This method could be adapted to measure soluble and insoluble hydrolysable tannins. Protein precipitation methods The quantification of tannins based on their operational properties i.e., their capacity for complexing with proteins (protein precipitation assays) (Makkar, 1989) are considered to provide better information on the biological value of feeds and fodders containing tannins. The method (Hagerman & Butler, 1980) based on binding of 125I-labelled bovine serum albumin (BSA) for the determination of the protein precipitation capacity of tannins is accurate and sensitive. Recently, this method has been simplified (Henson et al, 2003). However, the method requires special equipment and some degree of expertise, because it is a radio-assay.
In the protein precipitation method for the determination of tannins (Makkar et al., 1987), the protein in the tannin-protein complexes has been measured using the ninhydrin assay of amino acids released by alkaline hydrolysis of the complex. Subsequently, this method has been modified to enable measurement of both protein and phenolics in a tannin-protein complex (Makkar et al., 1988). This enables the measurement of the ratio of protein to tannin in the tannin-protein complex, which can be defined as the specific activity of tannins. This represents the amount of protein, which is bound by a unit of phenolics/tannins in the complex. This could provide valuable information regarding changes in the nature of tannins, vis-à-vis their protein binding capacity associated with, for example, development, maturity, post harvest storage and detanninification of foods and feeds. In this method (Dawra et al., 1988), a plant extract containing these moieties is applied to chromatography paper and reacted with protein bovine serum albumin. Unbound protein is washed off, and the protein in the tannin-protein complex is stained with Ponceau S, a dye specific for proteins. Protein-bound dye is eluted and the optical absorbance of the eluate is measured at

22
525 nm and converted to protein using a calibration curve. The dye bound can also be measured using an image analyser (Hoffman et al., 2002). The sensitivity of the chromatography paper-protein-Ponceau S method is about 20 times higher than that of other methods (Makkar et al., 1987, 1988). A further advantage is that, acetone containing plant extracts can be used in the chromatography paper-protein-Ponceau S method whereas the presence of acetone interferes with other protein precipitation assays. The preparation of plant extract in aqueous acetone (generally 70 %) is desirable because the solubility of tannins and phenolics from various plant sources is higher in aqueous acetone (Makkar et al., 1988; 1990), and acetone prevents the oxidation of phenols. Another protein precipitation method, which is insensitive to acetone, is the radial diffusion assay (Hagerman, 1987), in which tannin molecules migrate through agarose gel, which is impregnated with the protein, BSA.
In most protein-binding assays the protein used is BSA, because it has the advantage of being inexpensive, easily available and soluble. However, these methods can be used with any other protein. Tannin bioassay: All available protein precipitation assays measure tannins under conditions (temperature, pH, ionic strength, nature of protein), which are different from those in the rumen. Therefore the results obtained have limited applicability for predicting the nutritive value of tannin-containing feedstuffs. Polyvinylpyrrolidone and polyethylene glycol bind to tannins and make them inert (Makkar et al., 1995). This property has been exploited together with the in vitro gas method to quantify the effects of tannins on rumen fermentation (Makkar & Becker, 1996). Incubation of polyethylene glycol 6,000 with a tannin-containing feedstuff in the in vitro system increases gas production. The percentage increase in gas production represents the effects of the tannins; the higher the percentage increase in gas production, the greater the effect. Later studies (Makkar et al., 1998) showed that measurement of gas production alone could be misleading as tannins can affect nutrient partitioning. Measurement of microbial mass production should be combined with gas measurement to give a better insight into the effects of the tannins. This is achieved using purine and
15N
measurements. The full potential of this bioassay can be exploited if both the gas production and the microbial mass/protein production are measured.
Increase in nitrogen degradability of a feed when incubated in an in vitro rumen fermentation system in the presence of PEG is also a measure of tannin activity (Getachew et al., 2000; Jones et al., 2000) and this increase in nitrogen degradability has also been found to predict the effects of tannins in ruminants (Jones et al., 2000). 14C-labelled polyethylene glycol binding assay In this method the feed samples (without tannin extraction) are kept in contact with PEG spiked with 14C-labelled PEG, and the radioactivity bound to the feed sample is a measure of tannins; the higher the activity, the higher the tannin content (Silanikove et al., 2001). The method is reportedly useful since there is no need to extract tannins, and it is considered to be a measure of both bound and extractable tannins (Silanikove et al., 2001). High correlations between the PEG-binding assay and protein precipitation capacity method (Makkar et al., 1993; 1998) have been obtained and the values are useful in predicting the negative effects of tannins on ruminal degradation (Silanikove et al., 2001). A major source of error in this method could arise from the formation of tannin-PEG complexes, which will not be recovered in the feed sample and hence will introduce underestimated tannin values. There is a need to study the extent of formation of soluble versus precipitable PEG-tannin complexes.

23
Near infrared based method: Tannins in legume forages were quantified using near infrared reflectance spectroscopy. The wavelength, 2.150 µm was found to be an indispensable wavelength for the prediction of condensed tannins (Goodchild et al., 1997). This method can be used to determine tannins in large sample sets of homogenous feeds such as forage legumes.
None of the above methods provide unambiguous information on the composition or size of tannins. Although NMR and FAB mass spectroscopy are powerful tools for the analysis of proanthocyanidins and of their structure it is not currently possible to obtain useful mass spectra for polymeric condensed tannins. Furthermore the spectra are difficult to interpret. Gamble et al. (1996) used solid-state NMR spectroscopy. The results from this technique agree well with those from the butanol-HCl method. The use of NMR spectroscopy is expensive and it requires special expertise. Therefore, it cannot be used as a routine technique. Although oligomers and flavonoid monomers can be separated by HPLC, condensed tannins cannot be resolved by HPLC. No method based on HPLC is available for reliably quantifying condensed tannins in feedstuffs. For ecological and nutritional studies that may include hundreds of samples it is impractical to attempt to characterize and measure each individual compound. Instead, broad screening methods based on the similar chemistries of groups of related compounds could provide quantitative information about the classes of compounds. This strategy has been employed in many studies of condensed tannins (under appropriate conditions, degradation of condensed tannins to anthocyanidins and then its determination spectrophotometrically). The same approach could be employed for hydrolysable tannins. The leaves from most browses and trees contain both condensed and hydrolysable tannins but the determination of hydrolysable tannins has been neglected in the past, and without quantifying hydrolysable tannins or at least screening for their absence the adverse effects of tannins have been attributed to the condensed tannins.
From the above discussion, it is evident that each type of tannin responds differently in each of these assays. This variability in response makes it impossible to use any single method. To qualitatively and quantitatively describe the tannins in an extract, a set of methods, based on the objective of the study, is recommended. References Bate-Smith, E.C. 1973. Tannins of herbaceous leguminosae. Phytochemistry 12, 1809–1812. Bate-Smith, E.C. 1977. Astringent tannins of Acer species. Phytochemistry 16, 1421–1427. Beck, V., Unterrieder, E., Krenn, L., Kubelka, W. & Jungbauer A. 2003. Comparison of
hormonal activity (estrogen, androgen and progestin) of standardized plant extracts for large scale use in hormone replacement therapy. Journal of Steroid Biochemistry and Molecular Biology 84, 259-268.
Bednarek, P., Frański, R., Kerhoas, L., Einhorn, J., Wojtaszek, P. & Stobiecki, M. 2001. Profiling changes in metabolism of isoflavonoids and their conjugates in Lupinus albus treated with biotic elicitor. Phytochemistry 56, 77-85.
Bednarek, P., Wojtaszek, P., Kerhoas L., Einhorn, J., Frański, R. & Stobiecki, M. 2003. Profiling of flavonoid conjugates in Lupinus albus and Lupinus angustifolius responding to abiotic and biotic stimuli. Journal of Chemical Ecology, 29, 1127-1142.
Blaut, M., Schoefer, L. & Braune, A. 2003. Transformation of flavonoids by intestinal microorganisms. International Journal Vitamin and Nutrition Research 73, 79-87.
Boue, S.M., Wiese, T.E., Nehls, S., Burow, M.E., Elliott, S., Carter-Wientjes, C.H., Shih, B.Y., Mclachlan, J.A. & Cleveland, T.E. 2003. Evaluation of the estrogenic effects of legume extracts containing phytoestrogens. Journal of Agricultural and Food Chemistry 51, 2193-2199.

24
Bovy, A., Vos, R. de, Kemper, M., Schijlen, E., Pertejo, M.A., Muir, S., Collins G., Robinson, S., Verhoyen, M., Hughes, S., Santos-Buelga, C. & Tunen, A. van 2002. High-flavonol tomatoes resulting from from the heterologous expression of the maize transcription factors genes LC and C1. The Plant Cell 14, 2509-2526.
Broadhurst, R.B. & Jones, W.T. 1978. Analysis of condensed tannins using acidified vanillin. Journal of Science of Food and Agriculture 29, 788–794.
Butler, L.G., Price, M.L. & Brotherton, J.E. 1982. Vanillin assay for proanthocyanidins (condensed tannins), modification of the solvent for estimation of the degree of polymerisation. Journal of Agricultural and Food Chemistry 30, 1087–1089.
Bylka, W., Frański, R. & Stobiecki, M. 2002. Differentiation between isomeric acacetin-6-C-(6"-O-malonyl)-glucoside and acacetin-8-C-(6"-O-malonyl)-glucoside by using low-energy CID mass spectra. Journal Mass Spectrometry 37, 648-650.
Careri, M., Elviri, L. & Mangia, A. 1999. Validation of liquid chromatography ionspray mass spectrometry method for analysis of flavanones, flavones and flavonols. Rapid Communication in Mass Spectrometry 13, 2399-2405.
Claeys, M., Li, Q., Heuvel, H. van den, & Dillen, L. 1996. Mass spectrometric studies on flavonoid glucosides. In Applications of Modern Mass Spectrometry in Plant Sciences. R.P Newton & T.J. Walton (Eds.) Clarendon Press, Oxford. 182-194.
Cuyckens, F., Ma, Yl., Pocsfalvi, G. & Claeys M. 2000. Tandem mass spectral strategies for the structural characterization of flavonoid glycosides. Analysis 28, 888A-895A.
Cuyckens, F., Rozenberg, R., Hoffman, E. de, & Claeys M. 2001. Structure characterization of flavonoid O-glycosides by positive and negative nano-electrospray ionisation ion trap mass spectrometry. Journal Mass Spectrometry 36, 1203-1210.
Cuyckens, F., Shahat, A.A., Pieters, L. & Claeys M. 2002. Direct stereochemical assignment of hexose and pentose residues in flavonoid O-glycosides by fast atom bombardment and electrospray ionisation mass spectrometry. Journal Mass Spectrometry 37, 1272-1279.
Dawra, R.K., Makkar, H.P.S. & Singh, B. 1988. Protein binding capacity of microquantities of tannins. Analytical Biochemistry 170, 50–53.
De Rijke, E., Zafra-Gomez, A., Ariese, F., Brinkman, U.A.Th. & Gooijer, C. 2001. Determination of isoflavone glucoside malonates in Trifolium pratense L. (red clover) extracts: Quantification and stability studies. Journal of Chromatography A 932, 55-64.
Dixon, R.A. & Paiva, N.L. 1995. Stress-induced phenylpropanoid metabolism. The Plant Cell 7, 1085-1097.
Dixon, R.A. & Steel, C.L. 1999. Flavonoids and isoflavonoids – a gold mine for metabolic engineering. Trends Plant Sciences 4, 394-400.
Fiehn, O. 2001. Combining genomics, metabolome analysis, and biochemical modelling to understand metabolic networks. Comparative and Functional Genomics 2, 155-168.
Frański, R., Bednarek, P., Wojtaszek, P., & Stobiecki, M. 1999. Identification of flavonoid diglycosides in yellow lupin (Lupinus luteus L.) with mass spectrometric techniques. Journal Mass Spectrometry 34, 486-495.
Frański, R., Eitner, K., Sikorska, M., Matławska, I. & Stobiecki, M. 2003. Electrospray mass spectrometric decomposition of some glucuronic acid containing flavonoid diglycosides. Phytochemical Anaysis 14, 170-175.
Frański, R., Matławska, I., Bylka, W., Sikorska, M., Fiedorow, P. & Stobiecki, M. 2002. Differentiation of interglycosidic linkages in permethylated flavonoid glycosides from linked scan mass spectra (B/E). Journal of Agricultural and Food Chemistry 50, 976-982.

25
Friedrich, W., Eberhardt, A. & Galensa, R. 2000. Investigation of proanthocyanidins by HPLC with electrospray ionization mass spectrometry. European Food Research and Technology, 211, 56-64.
Fritz, K.L., Seppanen, C.M., Kurzer, M.S. & Csallany, A.S. 2003. The in vivo antioxidant activity of soybean isoflavones in human subjects. Nutrition Research 23, 479-487.
Gamble, G.R., Akin, D.E., Makkar, H.P.S. & Becker, K. 1996. Biological degradation of tannins in Sericea lespedeza by the white rot fungi Ceriporiopsis subvermispora and Cyathus stercoreus analysed by solid state 13C NMR spectroscopy. Applied and Environmental Microbiology 62, 3600–3604.
Gelpi E. 2002. Interfaces for coupled liquid-phase separation/mass spectrometry techniques. An update on recent developments. Journal of Mass Spectrometry 37, 241-253.
Gerhauser, C., Klimo, K., Heiss, E., Neumann, I., Gamal-Eldeen, A., Knauft, J., Liu, G.Y., Sitthimonchai, S. & Frank, N. 2003. Mechanism-based in vitro screening of potential cancer chemopreventive agents. Mutation Research-Fundamental Molecular Mechanisms of Mutagenesis 523, 163-172
Getachew, G., Makkar, H.P.S. & Becker, K. 2000. Effect of polyethylene glycol on in vitro degradability of nitrogen and microbial protein synthesis from tannin-rich browse and herbaceous legumes. British Journal of Nutrition 84, 73–83.
Goodchild, A.V., El Haramein, F.J., Makkar, H.P.S., El Monein, A.A. & Williams, P.C. 1997. 18th International Conference on Near-Infrared Spectroscopy, 15-19 September 199. Haus der Technik, Essen, Abstracts 37.
Gu, L., & Gu, W. 2001. Characterisation of soy isoflavones and screening for novel malonyl glycosides using high performance liquid chromatography electrospray ionisation-mass spectrometry. Phytochemical Anaysis 12, 377-382.
Guyot, S., Marnet, N. & Drilleau, J-M. 2001. Thiolysis-HPLC characterization of apple proanthocyanidins covering a large range of polymerisation states, Journal of Agricultural and Food Chemistry 49, 14–20.
Hagerman, A.E. 1987. Radial diffusion method for determining tannin in plant extracts. Journal of Chemical Ecology 13, 437–449.
Hagerman, A.E. & Butler, L.G. 1980. Determination of protein in tannin-protein precipitates. Journal of Agricultural and Food Chemistry 28, 944–947.
Hagerman, A.E., Zhao, Y. & Johnson, S. 1997. Methods for determination of condensed and hydrolysable tannins. In Antinutrients and Phytochemicals in Food. F. Shahidi (Ed.). ACS Symposium Series No. 662, American Chemical Society. 209–222.
Hartzfeld, P.W., Forkner, R., Hunter, M.D. & Hagerman, A.E. 2002. Determination of hydrolyzable tannins (gallotannins and ellagitannins) after reaction with potassium iodate. Journal of Agricultural and Food Chemistry 50, 1785–1790.
Henson, G.L., Niemeyer, L., Ansong, G., Forkner, R., Makkar, H.P.S. & Hagerman, A.E. 2003. Modified method for determining protein binding capacity of plant polyphenolics using radiolabeled protein. Phytochemical Analysis, (In press).
Hoffmann, E.M., Muetzel S. & Becker, K. 2002. A modified dot-blot method of protein determination applied in the tannin-protein precipitation assay to facilitate the evaluation of tannin activity in animal feeds. British Journal of Nutrition 87, 421–426.
Huck, C.W., Buchmeister, M.R. & Bonn, G.K. 2002. Fast analysis of flavonoids in plant extracts by liquid chromatography – ultraviolet absorbance detection on poly(carboxylic acid)–coated silica and electrospray ionization tandem mass spectrometric detection. Journal of Chromatography A 943, 33-38.
Inoue, K.H. & Hagerman, A.E. 1988. Determination of gallotannins with rhodanine. Analytical Biochemistry 169, 363–369.

26
Jennings, K.R. 1996. MS/MS instrumentation. In Applications of Modern Mass Spectrometry in Plant Sciences. R.P. Newton & T.J. Walton (Eds.). Clarendon Press, Oxford. 25-43.
Jones, R.J., Meyer, J.H.F., Bechaz, M. & Stoltz, M.A. 2000. An approach to screening potential pasture species for condensed tannin activity. Animal Feed Science and Technology 85, 269–277.
Justesen, U., Knuthesen, P. & Leth, T. 1998. Quantitative analysis of flavonols, flavones and flavanones in fruits vegetables and beverages by high-performance liquid chromatography with photodiode array and mass spectrometric detection. Journal Chromatography A 799, 101-110.
Klejdus, B., Vitamvasova-Sterbova, D. & Kuban, V. 2001. Identification of isoflavone conjugates in red clover (Trifolium pratense) by liquid chromatography-mass spectrometry after two-dimensional solid phase extraction. Analitica Chimica Acta 450, 81–97.
Labarbe, B., Cheynier V., Brossaud, F., Souquet, J-M. & Moutounet, M. 1999. Quantitative fractionation of grape proanthocyanidins according to their degree of polymerisation. Journal of Agricultural and Food Chemistry 47, 2719–2723.
Lei, Z., Jervis, J. & Helm, R.F. 2001. Use of methanolysis for the determination of total ellagic and gallic acid contents of wood and food products. Journal of Agricultural and Food Chemistry 49, 1165–1168.
Li, C.A., Meng, X.F., Winnik, B., Lee, M.J., Lu, H.; Sheng S.Q., Buckley, B. & Yang, C.S. 2001. Analysis of urinary metabolites of tea catechins by liquid chromatography/electrospray ionization mass spectrometry. Chemical Research in Toxicology 14, 702-707.
Lin, L.Z., He, X.G., Lindenmaier, M., Yang, J., Cleary, M., Qiu, S.X. & Cordell, G.A. 2000. LC/ESI/MS study of the flavonoid glycoside malonates of red clover (Trifolium pratense). Journal of Agricultural and Food Chemistry 48, 354-356.
Ma, Y., Cuyckens, F., Heuvel, H. van den & Claeys, M. 2001. Mass spectrometric methods for the characterisation and differentiation of isomeric O-diglycosyl flavonoids. Phytochemical Anaysis 12, 159-165
Makkar, H.P.S. 1989. Protein precipitation methods for quantitation of tannins: a review. Journal of Agricultural and Food Chemistry 37, 1197–1202.
Makkar, H.P.S. & Becker, K. 1993. Vanillin-HCl method for condensed tannins: effect of organic solvents used for extraction of tannins. Journal of Chemical Ecology 19, 613–621.
Makkar, H.P.S. & Becker, K. 1994. Some problems in the determination of tannins and possible solutions. Acta Horticulturae 381, 782–788.
Makkar, H.P.S. & Becker, K. 1996. A bioassay for tannins. The XVIIIth International Conference on Polyphenols, Bordeaux, 15-18 July 1996. Polyphenols Communications 96, 197–198.
Makkar, H.P.S., Blümmel, M. & Becker, K. 1998. Potential and limitations of in vitro gas method for studying the effects of plant defensive components on rumen fermentation. In Recent Advances of Research in Antinutritional Factors in Legume Seeds and Rapeseed. A.J.M Jansman, G.D. Hill, J. Huisman & A.F.B. van der Poel (Eds). Proceedings of the 3rd International Workshop on Antinutritional Factors in Legume Seeds and Rapeseed, 8–10 July 1998, Wageningen, 173-177.
Makkar, H.P.S., Blümmel, M. & Becker, K. 1995. Formation of complexes between polyvinyl pyrrolidone and polethylene glycol with tannins and their implications in gas production and true digestibility in in vitro techniques. British Journal of Nutrition 73, 897–913.

27
Makkar, H.P.S., Blümmel, M., Borowy, N.K. & Becker, K. 1993. Gravimetric determination of tannins and their correlations with chemical and protein precipitation methods. Journal of Science of Food and Agriculture 61, 161–165.
Makkar, H.P.S., Dawra, R.K. &, Singh, B. 1987. Protein precipitation assay for quantitation of tannins: Determination of protein in tannin-protein complex. Analytical Biochemistry 166, 435–439.
Makkar, H.P.S., Dawra, R.K. &, Singh, B. 1988. Determination of both tannin and protein in a tannin-protein complex. Journal of Agricultural and Food Chemistry 36, 523–525.
Makkar, H.P.S., Gamble, G. & Becker, K. 1999. Limitation of the butanol-hydrochloric acid-iron assay for bound condensed tannins. Food Chemistry 66, 129–133.
Makkar, H.P.S. & Singh, B. 1995. Determination of condensed tannins in complexes with fibre and proteins. Journal of Science of Food and Agriculture 69, 129–132.
Makkar, H.P.S., Singh, B., Negi, S., 1990. Tannin levels and their degree of polymerisation and specific activity in some agroindustrial byproducts. Biological Wastes 31, 137-144.
Matthews, S., Mila, I., Scalbert, A., Pollet, B., Lapierre, C., Herve du Terrill, T.H., Rowan, A.M., Doughlas, G.B. & Barry, T.N. 1992. Determination of extractable and bound condensed tannin concentrations in forage plants, protein concentrate meals and cereal grains. Journal of Science of Food and Agriculture 58, 321–329.
Mellenthin, O. & Galensa, R. 1999. Analysis of polyphenols using capillary zone electrophoresis and HPLC: Detection of soy, lupin, and pea protein in meat products. Journal of Agricultural and Food Chemistry 47, 594-602.
Niessen, W.M.A. 1999. Liquid Chromatography - Mass Spectrometry. Second Edition, Marcel Dekker Inc., New York.
Peng, S., Scalbert, A. & Monties, B. 1991. Insoluble ellagitannins in Castanea sativa and Quercus petraea woods. Phytochemistry 30, 775–778.
Price, M.L., Scoyoc, S. van & Butler, L.G. 1978. A critical evaluation of the vanillin reaction as an assay for tannin in sorghum grain. Journal of Agricultural and Food Chemistry 26, 1214–1218.
Porter, L.J., Hrstich, L.N. & Chan, B.G. 1986. The conversion of procyanidin and prodelphinidins to cyaniding and delphinidin. Phytochemistry 25, 223–230.
Sanchez-Rabaneda, F., Jauregui, O., Casals, I., Andres-Lacueva, C., Izquierdo-Pulido, M. & Lamuela-Raventos, R.M. 2003. Liquid chromatographic/electrospray ionization tandem mass spectrometric study of the phenolic composition of cocoa (Theobroma cacao). Journal Mass Spectrometry 38, 35-42.
Shirley, B.W. 1996. Flavonoids biosynthesis ”new” function for an ”old” pathway. Trends Plant Sciences 1, 377-382.
Sikorska, M., Matławska, I., Frański, R. & Stobiecki, M. 2003. Application of mass spectrometric techniques for structural analysis of apigenin 8-C-(6-O-glucopyranosyl) glucopyranoside - a novel flavonoid C-diglycoside. Rapid Communications Mass Spectrometry 17, 1380-1382.
Silanikove, N., Perevolotsky, A. & Provenza, F.D. 2001. Use of tannin-binding chemicals to assay for tannins and their negative postingestive effects in ruminants. Animal Feed Science and Technology 91, 69–81.
Stobiecki, M. 2000. Review - Application of mass spectrometry for identification and structural studies of flavonoid glycosides. Phytochemistry 54,,237-256.
Stobiecki, M., Carr, S.A., Seviert, H-J., Reinhold, V.N. & Goldman, P. 1984. The contribution of bacterial microflora in metabolic profiling. 32nd ASMS Annual Conference on Mass Spectrometry and Allied Topics. San Antonio, Texas 27 May-1 June 1984. Abstracts, 580-581.

28
Stobiecki, M. & Wojtaszek, P. 1993. Application of gas chromatography-mass spectrometry for identification of isoflavonoids in lupin root extracts. Journal Chromatography 509, 391-398.
Sumner, L.W., Mendes, P. & Dixon, R.A. 2003. Plant metabolomics: large-scale phytochemistry in the functional genomics era. Phytochemistry 62, 817-836.
Sumner, L.W., Paiva, N.L., Dixon, R.A. & Geno, P.W. 1996. High performance liquid chromatography/continuous-flow liquid secondary ions mass spectrometry of flavonoid glycosides in leguminous plant extracts. Journal Mass Spectrometry 31, 472-485.
Terrill, T.H., Rowan, A.M., Doughlas, G.B. & Barry, T.N. 1992. Determination of extractable and bound condensed tannin concentrations in forage plants, protein concentrate meals and cereal grains. Journal of Science of Food and Agriculture 58, 321-329.
Tomas-Barberan, F.A., Gil, M.I., Cremin, P., Waterhouse, A.L., Hess-Pierce, B. & Kader A.A. 2001. HPLC-DAD-ESIMS analysis of phenolic compounds in nectarines, peaches, and plums. Journal of Agricultural and Food Chemistry 49, 4748-4760
Tomas-Barberan, F.A. & Garcia-Viguera, C. 1997. Capillary electrophoresis versus HPLC in plant polyphenol analysis. Analysis 25, M23-M25.
Urbanek, M., Blechtova, L., Pospisilova, M. & Polasek, M. 2002. On-line coupling of capillary isotachophoresis and capillary zone electrophoresis for the determination of flavonoids in methanolic extracts of Hypericum perforatum leaves or flowers. Journal Chromatography A 958, 261-271.
Vanhoenacker, G., de Villiers, A., Lazou, K., de Keukeleire, D. & Sandra, P. 2001. Comparison of high-performance liquid chromatography - Mass spectroscopy and capillary electrophoresis - Mass spectroscopy for the analysis of phenolic compounds in diethyl ether extracts of red wines. Chromatographia 54, 309-315.
Waridel, P., Wolfender, J-L., Ndjoko, K., Hobby, K.R., Major, H.J. & Hostettmann, K. 2001. Evaluation of quadrupole time of flight tandem mass spectrometry and ion trap multiple-stage mass spectrometry for differentiation of C-glycosidic flavonoid isomers. Journal Chromatography A 926, 29-41.
Wolfender, J-L., Rodriguez, S. & Hostettmann, K. 1997. Liquid chromatography/ultra violet/mass spectrometric and liquid chromatography/nuclear magnetic resonance spectroscopic analysis of crude extracts Gentinaceae species. Phytochemical Analysis 8, 97-104.
Zhao, J.J., Yang, A.Y. & Rogers D. 2002 Effects of liquid chromatography mobile phase buffer contents on the ionisation and fragmentation of analytes in liquid chromatography/ion spray tandem mass spectrometric determination. Journal of Mass Spectrometry 37, 421-433.

29
Phenolic compounds and pyrimidine glycoside determination in Vicia narbonensis seed M. Arias1, L.T. Ortiz2 & M. de los Mozos1
1Centro de Investigación Agraria de Albaladejito, Carretera Toledo-Cuenca, Km 174, 16194 Cuenca, Spain 2Universidad Complutense de Madrid, Facultad de Veterinaria, Departamento de Producción Animal, Ciudad Universitaria, 28040, Madrid, Spain Summary The total phenolics, condensed tannins, vicine, convicine and L-DOPA content in the seed of 21 Vicia narbonensis accessions were measured. The results varied between 2.01 - 5.03 % for total phenolics, 0.22 - 0.94 % for condensed tannins, 0.012 - 0.067 % for vicine, 0.000 - 0.017 % for convicine and 0.000 – 0.011 % for L-DOPA. The results for total phenolics and condensed tannins were higher than those previously reported for this species. On the hand contrary, the vicine and convicine content was lower than the previously published value. The L-DOPA content was also very low. Keywords: Vicia narbonensis, polyphenolic compounds, pyrimidine glycosides,
antinutritional factors Introduction: Vicia narbonensis (narbon bean) is a grain legume from the Mediterranean region. It is related to faba bean (Vicia faba) and is postulated to be a wild ancestor of the crop. It is well adapted to medium to low rainfall areas and to neutral to alkaline soils (Castleman et al., 1999). It shows some characteristics of agronomic value such as erect growth habit, good nitrogen fixation and low to no attack by diseases and pests.
The seed contains up to 28 % protein in the dry matter (DM) (Aletor et al., 1994). The straw contains up to 9 % protein (Abd El-Moneim, 1992). However, it is not usually used for human or animal feeding due to a high content of antinutritional factors (ANFs), which include several phenolic compounds (polyphenols and tannins), pyrimidine glucosides (vicine, convicine and L-DOPA) and γ-glutamyl-S-ethenyl-cysteine (GEC).
Tannins are phenolic plant secondary compounds and are widely distributed in the plant kingdom, especially in pulses. Tannins exist primarily in condensed (CT) and hydrolysable forms (HT) (Haslam, 1989). The HT molecule contains a carbohydrate (generally D-glucose) as a central core. The hydroxyl groups of these carbohydrates are esterified with phenolic groups such as gallic acid or ellagic acid (Haslam, 1989). Structurally, CT are complexes of oligomers and polymers of flavonoid units linked by carbon-carbon bonds (Hangerman & Bluter, 1991). Tannins complex with proteins and other macromolecules and convert them into inert compounds with negative effects on animal nutrition (Reed, 1995).
Vicine and convicine are glucosides that occur mainly in Vicia faba seed. They are hydrolysed by the gut microflora and produce reactive compounds, which generate the free radicals divicine and isouramil. They are responsible for the disease favism (Marquardt et al., 1983). The compound L-DOPA (3,4, dihydroxifenilalanine) is a precursor of dopaquinone, which is a factor in haemolysis when Vicia faba is eaten (de Haro, 1983).

30
This species can store, a significant amount of the dipeptide GEC. Seed content varies from 0.41 to 3.77 % (Castleman, 2000). It reduces palatability and causes damage in monogastric animals (Enneking, 1995).
Despite the ANFs, which are present, sheep and cattle can graze V. narbonensis. It can also be used for grain and high quality hay production and is an acceptable feed supplement for sheep for short periods. It’s use, as long term feed has not been established. Its inclusion in monogastric diets is not recommended (Castleman, 2000).
This work presents an initial analysis of the level of ANFs in samples of Spanish V. narbonensis seed. Materials and methods Plant material This work analysed 20 germplasm accessions of V. narbonensis (Table 1) from the Centro de Recursos Fitogenéticos del Instituto Nacional de Investigación y Tecnología Agraria y Alimentaría (CRF-INIA). Seed was multiplied in the 2001 - 2002 season and was stored at the Banco Regional de Germoplasma de Albaladejito (Cuenca, Spain). The cultivar Altair was also analysed. Table 1. Origin of the Vicia narbonensis samples from Spain analysed in this work.
CRF number Province CRF number Province BG-005512 Jaen BG-019585 Cádiz BG-031093 Jaen BG-001894 Cuenca BG-013234 Ciudad Real BG-023509 Madrid BG-009987 Jaen BG-022759 Sicilia BG-019584 Cádiz BG-013235 Granada BG-029694 Madrid BG-009982 Toledo BG-029056 Granada BG-018826 Toledo BG-013237 Toledo BG-013236 Granada BG-025291 Jaen BG-022216 Baleares BG-001571 Cádiz BG-011729 Córdoba
Methods Flour samples were used for all analyses. Flour was obtained by milling entire seeds. The seeds were milled twice, initially without a sieve and then through a 1 mm screen. The seed dry matter (DM) was estimated. The mean flour moisture content was 9.39 %.
Total polyphenols were determined by the Folin-Denis method (Burns, 1963). One g of sample was boiled in 150 ml of distilled water for 2 h. It was diluted to 250 ml, cooled and filtered to obtain 2 ml of extract. This was mixed with 2 ml of Folin Denis reagent and 5 ml of saturated sodium carbonate solution. The volume was made up to 100 ml with distilled water. After 40 min absorbency was read, at 725 nm. Total polyphenols in the sample were calculated by a standard curve obtained from a standard tannic acid solution. Results are given as the percent of tannic acid equivalents in DM.
The vanillin colorimetric method (Hagerman & Butler, 1978) was used, to measure condensed tannins using a catequine solution as a standard. In an acid environment this reacts with vanillin to give a pink colour that was measured by spectrophotometrically. About 700 -

31
800 mg of flour was stirred with 50 ml of HCl solution 1 % in methanol for 20 h. The extract filtrate was transferred to a dark test tube to which vanillin (4 % in methanol) and concentrated HCl were added. The mixture was stirred and left to stand for 20 min in the dark and the absorbency was then read at 500 nm. For the blank a double quantity was prepared. The condensed tannin content of the samples was calculated from a standard curve (0.0, 0.1, 0.2, 0.3, 0.4 mg/ml of catequine). The results are given as percent of catequine units in the DM.
Vicine, convicine and L-DOPA analyses were by High Performance Liquid Chromatography (HPLC) (Marquardt & Frolich, 1981). Sample were extracted with 5 % perchloric acid and stirred with an Ultra Turrax (T25 basic, IKA, Germany) for 1 min. The extract was centrifuged for 10 min (Sorvall Superspeed RC2-B, EEUU) at 20,200 g to obtain a supernatant. It was then filtered through a 0.45 µm Millipore membrane and injected into a Beckman System Gold chromatograph with 20 µl/vial. Vials were kept on ice until injected to avoid glycoside hydrolysis. The compounds were measured with reverse phase HPLC using Beckman System Gold chromatography equipment. The moving phase was 0.05 M ammonic phosphate buffer at pH 2. It was prepared from concentrated phosphoric acid and double distilled, deionised water, and everything was filtered through a 0.45 µm Millipore membrane. Sample analysis was under isocratic conditions with a 1.2 ml/min flux and an injection volume of 20 µl. The wavelength used was 280 nm. Every sample was analysed twice. For the stationary phase a C18 reverse phase column was used. A stock standard solution was obtained by dissolving 34.4 mg of vicine, 15 mg of convicine and 189 mg of L-DOPA in 100 ml of deionised double distilled water. From this solution other dilutions were made with 5 % perchloric acid (400, 600, 800, 900 and 950 µl) to obtain a standard curve (these concentrations are usually used to determine these compounds in Vicia faba). In both cases diluted standards were filtered through a 0.45 µm Millipore membrane. All samples were frozen until injected to minimize possible glycoside hydrolysis.
Due to the low vicine, convicine and L-DOPA concentration in the samples, a second standard curve with new dilutions was made by taking 100, 200 and 300 µl of the standard No 1 (0.05 ml of the mother solution with 1 ml of 5 % perchloric acid) to give a standard curve that was linear between concentrations of 0 and 0.02 mg/ml for vicine, 0 to 0.001 mg/ml for convicine and 0.04 mg/ml for L-DOPA). Results and discussion The seed content of the different ANFs is shown in Table 2. There was high variation in the total polyphenol content of the samples (range: 2.02 – 5.03 %). The average was 2.78 % of tannic acid equivalents, which was higher than reported by Berger et al. (1999). There was a mean value of 0.57 % tannic acid equivalents (range: 0.24 – 1.12 % tannic acid equivalents). In V. Faba seed the mean value was 1.36 % tannic acid equivalents (range: 1.05 to 1.89 %).
In line with the high polyphenol content the condensed tannin content was also high with a range of 0.23 % to of 0.94 % of catequine equivalents. The mean value was 0.43 %. This was higher than reported by Aletor et al. (1994), at 0.21 % (range: 0.14 – 0.33 %). Previous V. faba data is not comparable with ours, because the units used were different.
The vicine and convicine concentration in all samples was very low. Mean values were 0.025 % for vicine and 0.005 % for convicine. These values are lower than those of Pitz et al. (1980) who reported a mean of 0.75 % for vicine and 0.08 % for convicine in V. narbonensis seed. These concentrations are lower than those usually found in V. faba (means: 0.72 % for vicine and 0.27 % for convicine). From the results the amount of L-DOPA in V. narbonensis seed is negligible.

32
Table 2. The antinutritional factor content of Spanish Vicia narbonensis seed. Percent CRF code Total polyphenols
(Tannic acid equivalents)
Condensed tannins(Catequine equivalents) Vicine Convicine L-DOPA
BG-005512 2.23 0.57 0.018 0.002 0.008 BG-031093 4.59 0.65 0.022 0 0 BG-013234 2.07 0.52 0.016 0.001 0.009 BG-009987 2.54 0.34 0.023 0.008 0.007 BG-019584 2.75 0.23 0.067 0.017 0.007 BG-029694 5.03 0.94 0.025 0.003 0.006 BG-029056 2.23 0.59 0.019 0.001 0.008 BG-013237 2.36 0.55 0.015 0.001 0.01 BG-025291 2.51 0.38 0.022 0.006 0.008 BG-001571 2.48 0.34 0.034 0.011 0.007 BG-019585 3.03 0.30 0.027 0.006 0.008 BG-001894 2.64 0.31 0.034 0.012 0.007 BG-023509 3.77 0.50 0.028 0.001 0.008 BG-022759 2.73 0.54 0.023 0.005 0.011 BG-013235 2.15 0.35 0.012 0.005 0.006 BG-009982 2.01 0.57 0.016 0.001 0.008 BG-018826 2.49 0.26 0.045 0 0 BG-013236 2.09 0.22 0.014 0.003 0.008 BG-022216 2.76 0.45 0.023 0.007 0.008 BG-011729 2.02 0.35 0.017 0.006 0.008 Altair 2.44 0.56 0.015 0.004 0.007 Average 2.59 0.43 0.025 0.005 0.007 S.E. 0.176 0.037 0.003 0.001 0.001 C.V. (%) 29.8 37.62 51.3 92.95 37
As could be expected from their chemical composition there was a moderate positive
correlation between the tannin and the polyphenol content (r = 0.58; α = 0.015) and between vicine and convicine (r = 0.62; α = 0.003). There was a negative correlation between tannins and convicine (r = -0.53; α = 0.029). Acknowledgements We thank the research team of the Food and Technology Department, INIA, especially C. Burbano and M. Muzquiz, for their technical and personal assistance and for supply of vicine, convicine and L-DOPA standards. We also thank the research team Animal Production Department, Veterinary Faculty, Universidad Complutense de Madrid and AGROSA, Semillas Selectas, S.A. for the supply of Altair seeds. This work was developed from a pre-doctoral scholarship sponsored by INIA (B.O.E. No 62, 13 March 2001). References: Abd El-Moneim, A.M. 1992. Narbon vetch (Vicia narbonensis): A potential feed legume
crop for the dry areas in West Asia. Journal of Agronomy and Crop Science 169, 347-353.

33
Aletor, V.A., Goodchild, A.V. & Abd El-Moneim, A.M. 1994. Nutritional and antinutritional characteristics of selected Vicia genotypes. Animal Feed Science and Technology 47, 125-139.
Berger, J.D., Siddique, K.H.M. & Loss, S.P. 1999. Cool season grain legumes for Mediterranean environments: species × environment interaction in seed quality traits and anti-nutritional factors in the genus Vicia. Australian Journal of Agricultural Research 50, 389-401.
Burns, R.E. 1963. Methods of tannin analysis for forage crop evaluation. Georgia Agricultural Experiment Station Technical Bulletin No. 32.
Castleman, G. 2000. A guide for Feeding Narbon Beans to Animals. Department of Natural Resources and Environment. Victorian Institute for Dryland Agriculture, Australia. 4 pp.
Castleman, G.H., Mock, L. & Heazlewood, C. 1999. New narbon bean (Vicia narbonensis) N9035*002. Technical Information Dossier. pp. 1-13.
De Haro, A. 1983. La calidad nutritiva de las leguminosas-grano y su control genético. In Leguminosas de Grano. J.I. Cubero & M.T. Moreno (Eds). Mundi-Prensa, Madrid. pp. 211-247.
Enneking, D. 1995. The toxicity of Vicia species and their utilisation as grain legumes. Centre for Legumes in Mediterranean Agriculture Occasional Publication Nº 6. University of Western Australia, Nedlands. (1st edition, Enneking, D., 1994. PhD thesis, University of Adelaide, South Australia. 119 pp.
Hagerman, A. & Butler, L. 1978. Protein precipitation method for the quantitative determination of tannins. Journal of Agricultural and Food Chemistry 26, 809-812.
Hagerman, A.E. & Butler, L.G. 1991. The specificity of proanthocyanidin-protein interactions. Journal of Biological Chemistry 256, 4494-4497.
Haslam, E. 1989. Plant Polyphenols. Vegetable Tannins Revisited. Cambridge University Press, Cambridge.
Marquardt R.R. & Frolich A. 1981. Rapid reversed-phase high-performance liquid chromatographic method for the quantitation of vicine, convicine and related compounds. Journal of Chromatography 208, 373-379.
Marquardt R.R., Muduuli, D.S. & Frohlich A.A. 1983. Purification and some properties of vicine and convicine isolated from faba bean (Vicia faba L.) protein concentrate. Journal of Agricultural and Food Chemistry, 31,839-844.
Pitz, W.J., Sosulski, F.W. & Hogge, L.R. 1980. Occurrence of vicine and convicine in seeds of some Vicia species and other pulses. Canadian Institute of Food Science and Technology Journal 13, 35.
Reed, J.D. 1995. Nutritional toxicology of tannins and related polyphenols in forage legumes. Journal of Animal Science 73, 1516-1528.


35
Evaluation of antinutritional factors and nitrogen compounds of selected pea cultivars (Pisum sativum L.) N. Ariza, M.A. Martín-Cabrejas, R.M. Esteban, E. Mollá & F.J. López-Andréu Departamento Química Agrícola, Facultad de Ciencias, Universidad Autónoma de Madrid, 28049 Madrid, Spain Summary The nutritional characteristics of eight different pea cultivars (Pisum sativum L.) from several European countries were studied. The nutrient content, particularly of protein, showed considerable variability, which depended on genetics, climate, soil and cultural conditions. In contrast with other grain legumes such beans (Phaseolus vulgaris) and soybean (Glycine max), the results of the analysis showed that most of the pea cultivars did not contain significant amounts of lectin, chymotrypsin and α-amylase inhibitor. Thus, no particular processing was needed to reduce their levels of antinutritional factors (ANFs) for human nutrition.
This work also studied the pea seed protein solubility as the nutritive value of peas depends on the availability of their seed proteins for digestion and absorption in the digestive tract. The pea cultivars contained higher levels of extractable nitrogen (N) compounds than other legume seeds. Exhaustive fractionation studies with sodium dodecylsulphate (SDS) have provided valuable information. The general trend with SDS was that a high percent of the protein was soluble. The final insoluble residue contained a very low level of N. These results potentially provide useful guidance on the utilisation of these seeds and their products. Keywords: peas, antinutritional factors, nitrogen compounds, varieties Introduction The grain legumes include peas, beans, lentils (Lens culinaris), peanuts (Arachis hypogaea), and other plants with pods. Legumes are an important food source and are widely consumed worldwide. Legumes have been cultivated for thousands of years, although many of the cultivars of beans and peas that are common today were unknown until relatively recently. Legumes play an important role in traditional diets of many regions of the world, and in many western countries peas have started to play a significant dietary role because of their beneficial nutritional effects (Messina, 1999). Thus, it is now recommended to increase the amount of legume in the diet of diabetics because of their apparent beneficial effect in reducing postprandial glycemia (Thorne et al., 1983). Recently, attention has also focused on the growing evidence that legume seed can play an important role in the prevention of chronic diseases (Kushi et al., 1999).
However, in some countries legume seed is not utilised sufficiently in human diets because of their ANFs such as lectins and enzyme inhibitors. Among the grain legumes, dry peas provide a good source of protein, vitamins, calories, and structural carbohydrates mainly due to their high soluble dietary fibre content (Kosson et al., 1994). In Western countries peas are used in a variety of day-to-day food dishes.
The aim of the present work was to determine the variation in some of those compounds of interest from a nutritional point of view for some selected pea varieties grown in Europe.

36
The results should permit the selection of a number of pea cultivars for large-scale cultivation in Europe. Material and methods Plant material Eight varieties of dry peas from different European countries were used in this study (Table 1). Pea seeds were ground and freeze-dried for analysis. Table 1. The cultivar, type and origin of the peas studied.
Pea cultivar Type Origin Deso Commercial Holland Solara Commercial Holland Frisson Commercial France Amino Commercial France Progress-9 Commercial Great Britain Ballet Commercial Great Britain RV-8 Commercial Valladolid (Spain) Cea Commercial Valladolid (Spain)
Methods Antinutritional factors: Pea flours were extracted (1:10, w/v) by stirring overnight with a 0.02 M sodium phosphate buffer at pH 7.0 containing NaCl (8 g/l) at ≈ 1 °C followed by centrifuging at 50,000 g for 25 min. The resultant clear supernatants were used for ANF evaluations. Chymotrypsin inhibitor activity was determined by the method of Grant et al. (1986). The α-amylase inhibitor content was measured by the starch/iodine procedure of Piergiovanni (1992).
Triplicate assays were conducted for inhibitor determination. Haemagglutinating activity was estimated in sodium phosphate extracts by a serial dilution procedure using rat blood cells (Grant et al., 1986). The assays were reproducible to ± 1 dilution and values presented are the mean of four separate measurements. Seed protein solubilisation: Seed meals were extracted by stirring overnight with 20 g/l of sodium dodecylsulphate (SDS) (1:10, w/v) at room temperature centrifuging at 50,000 g for 20 min. and decanting the supernatant. The residue was re-extracted and the two extracts were combined. Proteins in the extracts were precipitated with trichloroacetic acid solution (final concentration 50 g/l) at 1ºC followed by centrifuging at 50,000 g for 20 min. (Martín-Cabrejas et al., 1995). The N content of the pellets (protein-N) and the supernatants (non-protein-N) was estimated using the Kjeldahl procedure (AOAC, 1995). Results and discussion None of the pea varieties contained any α-amylase or chymotrypsin inhibitors. They had very low lectin levels (Table 2). Nutritionally these results are interesting because these ANFs have deleterious or toxic effects on animals and humans (Liener, 1989, 1994). Generally, the major ANF limiting legume seed use is lectin. Although, lectins are generally more resistant

37
to heat-denaturation than other plant proteins, pea lectins can be inactivated by a short cooking time because of their very low content. Therefore, these pea varieties should not need pre-treatment before they can be safely used as a food (Ariza, 2000). Table 2. Antinutritional factor content of dry pea seed (activity) (g/100 g DM).
Pea cultivar α-amylase inhibitor Chymotrypsin inhibitor Haemagglutinating activityDeso n.d1. n.d. 0.06 Solara n.d. n.d. 0.06 Frisson n.d. n.d. 0.06 Amino n.d. n.d. 0.12 Progress-9 n.d. n.d. 0.06 Ballet n.d. n.d. 0.06 RV-8 n.d. n.d. 0.06 Cea n.d. n.d. 0.06
1n.d. = not detected
The nutritive value of peas depends on seed protein availability for digestion and absorption in the digestive tract. These pea varieties contained significant levels of N (Table 3) compared to some other legumes. The highest N levels were in cvs Frisson, Deso and Progress-9. The SDS N solubility study has also provided valuable information. Table 3. Pea seed nitrogen (N) solubility in sodium dodecylsulphate (SDS) (mg N/g DM). N-Soluble in SDS
Pea cultivar Total N Total soluble N Protein N Non-Protein N Deso 44.2 ± 0.2 18.8 15.1 ± 1.9 3.7 ± 0.7 Solara 38.8 ± 0.1 16.5 12.6 ± 1.4 3.9 ± 0.3 Frisson 47.8 ± 0.4 17.4 13.5 ± 2.6 3.8 ± 0.3 Amino 38.5 ± 0.1 11.1 7.5 ± 0.8 3.5 ± 0.0 Progress-9 42.2 ± 0.3 22.8 15.4 ± 1.6 7.4 ± 0.5 Ballet 39.2 ± 0.2 11.8 6.1 ± 0.0 5.7 ± 1.2 RV-8 38.8 ± 0.2 14.4 11.7 ± 2.7 2.6 ± 0.6 Cea 38.4 ± 0.1 13.4 10.9 ± 5.6 2.5 ± 0.5
Extraction with SDS promoted a high degree of N solubilisation in most of the peas;
their content varied from 28 to 36 % of total N, except in cvs Ballet and Amino, which had lower levels. The SDS soluble fraction was divided into protein-N and non-protein-N. The main component of the supernatants was protein-N (52 % to 80 % of the SDS-fraction). The pea cvs Progress-9, Deso and Frisson had the highest content. This corroborates that peas have high levels of digestible proteins. The non-protein-N comprised peptides, free amino acids, and non-protein nitrogen compounds.
These results show that the main components were easily extractable proteins, which were not linked to other cell compounds such as starch, complex carbohydrates or dietary fibre. These proteins are highly digestible and would be the main reserve proteins (globulins and albumins) of the pea seed (Periago et al., 1994).
This data gives a clear indication that genetic and environmental factors can cause significant changes in the solubility of various nitrogen compounds in pea seeds. The British (Progress-9), French (Frisson) and Dutch (Deso) cultivars were the most suitable from a nutritional point of view and could selected for use for large-scale cultivation in Europe.

38
References AOAC 1995. Official Methods of Analysis, 16th Edition. Association of Official Analytical Chemists, Washington, DC. Ariza, N. 2000. Estudio de componentes nutricionales y antinutricionales en guisante (Pisum
sativum, L.). Influencia de la variedad y del proceso de germinación. Tesis, Universidad Autónoma de Madrid.
Grant, G., McKenzie, N.H., Watt, W., Stewart, J.C., Dorward, P.M. & Pusztai, A. 1986. Nutritional evaluation of soybeans (Glycine max): Nitrogen balance and fractionation studies. Journal of the Science of Food and Agriculture 37, 1001-1010.
Kosson, R., Czuchajowska, Z. & Pomeranz, Y. 1994. Smooth and wrinkled peas. 1. General physical and chemical characteristics. Journal of Agricultural and Food Chemistry 42, 91-95.
Kushi, L.H., Meyer, K.A. & Jacobs, D.R. 1999. Cereals, legumes, and chronic disease risk reduction: evidence from epidemiologic studies. American Journal of Clinical Nutrition 70, 451S-458S.
Liener, I.E. 1989. Control of antinutritional and toxic factors in oil-seeds and legumes. In Food Uses of Whole Oil and Protein Seeds. E.W. Lusas, D.R. Erickson & W. Nip (Eds). AOCS: Champaign, IL. pp. 344-371.
Liener, I.E. 1994. Implications of antinutritional components in soybean foods. Critical Review of Food Science and Nutrition 34, 31-67.
Martín-Cabrejas, M.A., Esteban, R.M., Waldron, K., Grant, G., Bardocz, S. & Pusztai, A. 1995. Hard-to-cook phenomenon in beans: Changes in antinutrient factors and nitrogenous compounds during storage. Journal of the Science of Food and Agriculture 69, 429-435.
Messina, M.J. 1999. Legumes and soybeans: overview of their nutritional profiles and health effects. American Journal of Clinical Nutrition 70, 439-450.
Periago, M.J., Ros, G., Englyst, H.N. & Rincón, F. 1994. Estudio del guisante (Pisum sativum) en función de la variedad y tamaño. Revista Española de Ciencia y Tecnología de Alimentos 34, 565-575.
Piergiovanni, A.R. 1992. Effect of some experimental parameters on activity of cowpea α-amylase inhibitors. Lebensmittel-Wissenschaft und-Technologie 25, 321-324.
Thorne, MJ., Jenkins, D.J., Wolever, T.M. & Jenkins, A.L. 1983. Factors affecting starch digestibility and the glycemic response with special reference to legumes. American Journal of Clinical Nutrition 38, 481-488.

39
The effect of cyanogenic glucosides of seed of Stizolobium deeringianum from Tabasco, Mexico L. Barrientos1, J. Vargas1, F. Navarro1, M. Ruíz2, F. López-Dellamary1, A. Rodríguez1 & J. Pineda3 1Wood, Cellulose, and Paper Research Department, University of Guadalajara, México 2Botany and Zoology Institute, University of Guadalajara, México 3Neurobiology Laboratory, University of Guadalajara, México Summary Antinutritional factors (ANFs) in velvet bean seed (Stizolobium deeringianum) were studied. Chemical analysis of the seeds showed that the total phenol content was 3.45 % (by spectrophotometer), condensable tannins were about 0.03 % (by Skin’s power method) and cyanogenic glucosides (CG) were 0.42 mg of HCN/100 g dry mater (DM).
A study with Wistar rats from the same litter and of about the same weight was also conducted. Each rat was given a dose of 4.2 and 5.4 ml of glucosides (obtained as HCN) mixed in water. The feeding experiment lasted for 10 and 21 d. Each day the rats were weighed and food consumption was recorded. At the end of the experiment, rat liver damage was analysed.
Results of the chemical analysis and the rat study showed that the presence of ANFs in velvet bean seed, do not present a problem for human and animal consumption, provided beans are processed before consumption. In conclusion this wild legume species, can be proposed as an alternative non-traditional food source.
Keywords: anti-nutritional factors, cyanogenic glucosides, phenolic compounds, Stizolobium
deeringianum Introduction In nature, there are a great many wild plants. Many of these, have a high seed protein content and could be used as alternative food sources. Among such plants is velvet bean (Stizolobium deeringianum), an Indian native legume.
Velvet bean is characterised by fast growth, high seed production and it does not demand high soil fertility. In Mexico, velvet bean, known as Nescafé, is grown in the Tamulté de las Sabanas region of Tabasco where the native people use it as a food (toasted) in April and May. This legume is potentially an important protein source (25.5 %) for human and animal consumption. However, the presence ANFs limits the direct consumption of seed (Casares & López 1959). The aim of this work was to determine some of the ANFs present in velvet bean seed. Materials and methods Sample preparation Velvet bean seed was collected from the Mexican state of Tabasco. Seed was air-dried and milled using a 0.5 mm Wiley mill. The flour was oven dried at 56 °C.

40
Seed chemical analysis Total phenols were measured by spectrophotometry with a 565 nm Perkins Elmer Lambda 2. Condensable tannins were measured using the TAPPI technique T-207-OM-89. Cyanogenic glucosides were measured using a UV-visible spectrophotometer (Perkin Elmer Lambda 2) at 565 nm (AOAC, 1989). Biological test A biological test used Swiss Wistar rats. Male rats from the same litter weighing between 200 and 300 g were used. There were total of twenty rats, one group of 4 rats for control, two groups of same number of rats (8) for raw seed and two groups of 4 rats for the cooked seed treatments.
For the untreated and treated rat groups 200 ml of water and 150 g of food was supplied. For the treatments a dose of HCN, obtained from cyanogenic glucosides in the ratio of 5.4 ml/100 g for raw seed and 4.2 ml/100 g for cooked seed, was mixed into the water. Each day’s consumption was measured. The experiment was conducted for two time periods of 10 and 21 d. After 10 d two rats from each group were sacrificed for histological damage studies. The remaining rats were kept until 21 d.
To determine histopathological damage to the rats, at the end of each time period rats were killed by gassing. The kidney, pancreas, liver and lymph nodes were sampled. All slices were examined histologically to determine any morphological changes (Buxk, et al., 1991; Gapalan et al., 1992).
A further experiment was carried out with lambs to determine in situ digestibility. Triplicate nylon bags were used for 72 h. Protein, crude fibre, dry matter (DM) and organic matter (OM) were measured (Buck et al., 1999). Results and discussion The total phenol content was 3.45 % and condensed tannins were 0.03 % (Tappy, 1999). Cyanogenic glucosides were 0.42 mg of HCN/100 g DM. Plant products that contain more than 20 mg HCN/100 ml are consider to have high HCN levels. The amount found in this plant suggest it could be used in diets, without causing severe damage (Montgomery, 1964).
Rat weight and liver studies showed that in most of the 5 rat groups there was no significant difference in food consumption. Rats in cage 2 fed cooked seed + 5.8 ml of HCN consumed the most water. Comparing rat water intake between animals on cooked and raw seed, the latter animals drank the most water. There was no significant difference in rat weights among five cages (Buck et al., 1991).
Table 1 shows the damage caused by the consumption of raw and cooked seed after 10 d of treatment. Liver damage was minor and showed degeneration in response to both raw and cooked seed. The kidneys showed minor amiloidosis, which should not be significant for consumption. The most significant damage was in the pancreas, where moderate vacuolation was observed after ingestion of raw seed. This was not seen with cooked seed (Kamalu, 1993a,b).
Table 2 shows the damage caused by seed consumption after 21 d. There was severe and moderate hepatitis after ingestion of both raw and cooked seed. In the kidney there was marked pigmentation and moderate nephrosis with both raw and cooked seed.

41
Table 1. Histopathological changes in Wistar rat organs after 10 d of consuming raw and cooked velvet bean seed.
Organ Seed treatment Liver Kidney Pancreas Raw Discrete blurred
deterioration Discreet amyloidosis in tubular lumen
Disassociation of acinis with moderate vacuole formation
Cooked Discrete blurred deterioration
Discreet amyloidosis in the tubular lumen
No change
Kamalu et al. (1995) reported that linamarin, a type of cyanogenic glucoside in cassava,
(Manihot escuelenta), hydrolyses in the gastrointestinal tract causing electrolyte imbalance in cells by inhibition of Na+ K+ ATPase. Table 2. Histopathological changes in Wistar rat organs after 21 d of consuming raw and
cooked velvet bean seed. Organ Seed treatment HCN (%) Liver Kidney Lymph nodes
Raw 5.4 Severe blurred deterioration
Discrete blurred necrosis with the presence of a haemosidersine-like pigment
Presence of a haemosidersine-like pigment
Cooked 4.2 Moderate hepatitis Moderate unpigmented necrosis
Presence of a small quantity of pigment
The in situ digestibility trial showed a DM digestibility of 89 and 84 % for raw and
cooked seed respectively. There was no difference in OM digestibility between raw and cooked seed (mean 83 %). These results confirm that the digestibility of velvet bean seed is high (Tejeda, 1985). Conclusions Chemical analysis of cyanogenic glucosides in velvet bean seed showed there was a low concentration in the seed (0.42 mg/100 g DM). At that level it is probably not harmful for human and animal consumption. An in situ digestibility study showed that velvet bean seed was highly digestible at about 80 %. This is higher than the 60 % that can be considered normal. A HCN toxicity study, in rats, showed severe organ damage when raw seed was fed but cooked seed was not harmful. Thus, we recommend that seed is cooked, to release the toxic compounds, before consumption. References AOAC. 1989. Official Methods of Analysis (14th Edition). Association of Official Analytical
Chemists, Arlington. Chapter XIII. Buck, W.B., Osweiler, G.D. & Gelder, G.A.V. 1991. Veterinary toxicology: Clinic and
Diagnostics (Spanish) Modern Manual. pp. 127-131.

42
Casares, R. & Lopez H.C. 1959. Bromatology studies of the seeds of Stizolobium deeringianum, acclimatized in Spain. Archivos Instituto de Aclimatación 8, 19-22..
Gopalan, V., Pastuszyn, A., Galey, W.R. Jr & Glew, R.H. 1992. Exolytic hydrolysis of toxic plant glucosides by guinea pig liver cytosolic β-glucosidase. Journal of Biological Chemistry 267, 14027-14032.
Kamalu, B.P. 1993a. Pathological changes in growing dogs fed on a balanced cassava (Manihot escuelenta Crantz) diet. British Journal of Nutrition 69, 921-934.
Kamalu, B.P. 1993b. The adverse effects of long–term cassava (Manihot escuelenta Crantz) consumption. Journal of Food Science and Nutrition 46, 65-93.
Kamalu, B.P. 1995. The adverse effects of long-term cassava (Manihot escuelenta Crantz) consumption. International Journal of Food Science and Nutrition 46, 65-93.
Montgomery, R.D. 1964. Observations on the cyanide content and toxicity of tropical pulses. West Indian Medical Journal 13, 1-11.
Tappi Test Methods 1998. Technique T 207 OM-93 Tappi Press, Atlanta Ga. Tejada, H.I. 1985. Manual de Laboratorio Para Análisis de Ingredientes Utilizados en la
Alimentación Animal. Apoya a la Investigación y Experimentación Pecuaria México, México City. pp. 353-354.

43
Determination of the antinutritional factors in the seed of tropical grain legumes with potential for human and animal feeding M.F. Díaz1, M.A. Martín–Cabrejas2, N. Ariza2, F.J. López–Andreu2, L. Jaime3 & C. Vidal–Valverde3 1Instituto de Ciencia Animal, Carretera Central, Km 47, 1/2, San José de las Lajas, La Habana, Cuba 2Departamento de Química Agrícola, Facultad de Ciencias, Universidad Autónoma de Madrid, 28049 Madrid, Spain 3Instituto de Fermentaciones Industriales, CSIC, Juan de la Cierva 3, E - 28006, Madrid, Spain Summary This study aimed at evaluating the content of anti-nutritional factors (ANFs) such as protease inhibitors, condensed tannins and inositol phosphate in meals from the seed of eight cultivars from the tropical grain legume species Vigna unguiculata (cowpea), Glycine max (soybean), Canavalia ensiformis (jackbean), Lablab purpureus (dolichos) and Stizolobium niveum (mucuna). The soybeans contained the highest trypsin (15.86 g/100 g for Incasoy 27 and 13.29 g/100 g in Duocrop) and chymotrypsin inhibitor levels (6.40 g/100 g in Duocrop), followed by dolichos. Mucuna had the highest amount of inositol phosphate (16.36 µmol/g) and condensed tannins (0.38 %), together with the cowpea variety INIFAT 93 (0.43 %, for tannins). This study showed the variability in ANF concentrations among legume species and varieties. This indicates the need to determine the ANF content of all legume species or cultivars used for human and animal feeding. Generally, cowpea was the legume with the lowest ANF content specially the cv. Trópico 782. The presence of more than one ANF in all of the seeds evaluated, and the different chemical composition of their seeds indicates the need for processing to increasing their nutritive value. Keywords: tropical legumes, anti-nutritional factors Introduction Legume seeds are an important source of protein, energy, vitamin and minerals for human and animal consumption. In Cuba, legume species like Glycine max, Vigna unguiculata, Lablab purpureus, Canavalia ensiformis and Stizolobium niveum have been introduced and genetically improved for tropical conditions. Studies have been carried out at the Instituto de Ciencia Animal that have proved the agronomic and nutritional potential of these legume crops as non-conventional feed sources for monogastric animals (Díaz et al., 2001).
The presence in these legume seeds of toxic and ANFs has limited their use in human and animal feeding. In general, their consumption has been associated with pancreatic hypertrophy or hyperplasia, decrease in digestibility and amino acid absorption, and reduced essential mineral bioavailability (Martin-Cabrejas et al., 1995). The aim of this study was to evaluate the ANF content in eight legumes with agronomic and nutritional potential for human and animal feeding in the tropics.

44
Materials and methods Eight seed samples from the tropical grain legumes Vigna unguiculata cvs White, Inifat 93 and Trópico 782; Glycine max cvs Incasoy 27 and Duocrop; Canavalia ensiformis; Lablab purpureus cv. Rongai and Stizolobium niveum were analysed.
The legumes were sown in the rainy season on a typical red ferrallitic soil and were hand harvested when the seed was ripe. The seeds were sun-dried to reduce their moisture content (12 - 14 %) and were stored at 6 - 10 ºC. The seed moisture was < 15 %. Three representative samples of 200 g were taken using the method of Lees (1969). The design was completely randomised. Chemical analysis Trypsin inhibitors were determined according to Grant et al. (1986). Chymotrypsin inhibitors were estimated by the method of Martin-Cabrejas et al. (1998), α-amylase inhibitors as per Piergiovanni (1992) and condensed tannins by the Porter et al. (1986) method. Estimation of inositol-phosphate was according to Honke et al. (1998). Calculations The amount of inhibitor in the different enzymatic assays was calculated comparing the amount of sample or inhibitor required to give a 50 % inhibition of enzyme activity. It was expressed as grams of commercial inhibitor equivalent to 100 g of legume meal. The assays were in triplicate. Statistical analysis The data were statistically analysed and Duncan's (1955) test was used to determine differences among means. Results and discussion Trypsin inhibitors The soybeans had the highest levels of trypsin inhibitors. Dolichos was intermediate and cowpea, mucuna and jackbean had the lowest concentrations (Table 1). Other authors have reported the high trypsin inhibitor content of soybean seed, compared to other grain legumes (e.g. Armour et al., 1998). They noted that more than 6 % of soybean protein comprised two powerful protease inhibitors (Kunitz and Bowman Birk Inhibitor). These inhibitors decreased growth, caused pancreatic hypertrophy and hyperplasia and adenomatose lesions in the pancreas of monogastric animals fed raw soybean seed. The effect was directly related to animal age and the time of exposure to the diet (Martín-Cabrejas et al., 1998). Chymotrypsin inhibitors The soybean cv. Duocrop had the highest chymotrypsin inhibitor content. However, it was not significantly different from Incasoy 27, mucuna or dolichos. Cowpeas and jackbean had the lowest content (Table 1).

45
Table 1. Protease inhibitors (g/100 g) and condensed tannins (%)in legume seeds.
Legume Trypsin inhibitor
Chymotrypsin inhibitor
α-amylase inhibitor
Condensed tannins
V. unguiculata Trópico 782 3.08d* 2.78cd 0.011b 0.0330ef Inifat 93 2.83de 2.20d 0.020a 0.4300a V. white 2.71de 4.35bc 0.011b 0.2600c
Glycine max Incasoy 27 15.86a 5.10ab 0.004e 0.0070f Duocrop 13.29b 6.40a 0.009d 0.1040d
S. niveum 1.61f 5.08ab 0.012bc 0.3800b C. ensiformis 1.86ef 2.16d 0.010d 0.0460e L. purpureus 7.19c 5.52ab 0.012bc 0.2700c SE ± 0.30*** 0.476** 0.0003** 0.0098*** *Within columns means followed by a different letter are significantly different (P < 0.05) (Duncan 1955). **P < 0.01, ***P < 0.001.
αααα-amylase inhibitors All the legumes had very low concentrations of this inhibitor. The cowpea cv. Inifat 93 had the highest amount. The soybean cultivars had the least and the other legumes were intermediate (Table 1). Condensed tannins The cowpea cv. Inifat 93 and mucuna had the highest condensed tannin content. The soybean cv. Incasoy 27, the cowpea cv. Trópico 782 and the jackbean had the least (Table 1). There were differences in tannin levels among cultivars from the same legume species. Yoshida et al. (1996) showed that although there was variability in the tannin content of varieties of the same species, the type of anthocyanidin was the same for all varieties, irrespective of environmental conditions. Inositol phosphates Mucuna had highest concentration of total inositol phosphates and IP5 + IP6 and dolichos the lowest. In the cowpeas cv. Trópico 782 contained the lowest amount of this ANF. Between the two soybean cultivars Duocrop contained less than Incasoy 27 (Table 2). The results show that the relative proportion of each inositol phosphate analysed (IP3, IP4, IP5 and IP6) varied among the different legume species. Further, inositol hexaphosphate (IP6) was the compound that was most common in the samples.
This study showed the presence of more than one ANF in all of the tropical grain legume seeds evaluated, as well as variable concentration in the all legumes. Generally, Vigna unguiculata had the lowest ANF content, especially cv. Trópico 782.
Kakade et al. (1972) in soybean and Oluwatosin (1999) in cowpea demonstrated that the environment primarily affected the level of tannins, trypsin inhibitors and phytates in these legumes. They found significant genotype by environment interactions. Honke et al. (1998), in a study of seed inositol phosphates, in Pisum sativum, Vicia faba and Lupinus luteus, in consecutive years, found that although the environment (temperature, rainfall and

46
humidity) did not affect the total inositol phosphate content of the seed it did affect the form in which they accumulated during seed maturation.
Table 2. Inositol phosphates (IP3, IP4, IP5, IP6) (µmol/g) in legume seed. Legume Total IP IP5 + IP6 IP6 IP5 IP4 IP3 V. unguiculata
Trópico 782 9.38c* 7.64d 5.04c 2.61d 0.96c 0.78d Inifat 93 12.56b 10.24b 5.89b 4.35b 1.34b 0.98cd V. white 13.02b 10.35b 6.20ab 4.15bc 1.42b 1.26bc
Glycine max Incasoy 27 13.04b 10.41ab 6.68a 3.70c 1.10bc 1.54ab Duocrop 10.50c 8.76c 6.00b 2.66d 0.93c 0.90cd
S. niveum 16.36a 11.30a 5.81b 5.49a 3.33a 1.72a C. ensiformis 11.80b 9.52bc 4.98c 4.55b 1.34b 0.93cd L. purpureus 7.72d 6.97d 5.28c 1.69e 0.44d 0.30e SE ± 0.37*** 0.27*** 0.15** 0.16*** 0.10*** 0.10*** *Within columns means followed by a different letter are significantly different (P < 0.05) (Duncan 1955). **P < 0.01, ***P < 0.001.
The presence of more than one ANF, with differing chemical composition, and sites in
the seed leads to a search for processes to produce biochemical and structural modifications to reduce the level of these substances with an expected increase in the nutritive value of the final product.
Germination and fermentation produce positives changes in the nutritional value of legume seed such as increases in essential amino acids, soluble proteins and in vitro protein digestibility (Kiers et al., 2000); significant decreases in α-galactosides and penta and hexa-phosphate inositols (Uwaegbute et al., 2000), a reduction in proteases and the degree of tannin polymerisation (Nnam, 1999). These processes also increase the levels of vitamin B1, B2 and C in legume seed (Blázquez, 1999). References Armour, J.C., Chanaka Perera, R.L., Buchan, W.C. & Grant, G. 1998. Protease inhibitors
and lectins in soya beans and effect of aqueous heat - treatment. Journal of the Science of Food and Agriculture 78, 225-231.
Blázquez, I. 1999. Contenido de vitamina B1 y vitamina B2 en guisantes y lentejas. Efecto de los procesos de germinación y extracción alcohólico. Tesis de Licenciatura en Ciencias Química. Universidad Autónoma de Madrid, Facultad de Ciencias, Departamento de Química, Agrícola, Geología y Geoquímica. Madrid, 75 p.
Díaz M.F., Padilla, C., Aguirre, L., Lon-Wo, E., Castro, M. & Cino, D.M. 2001. Utilización de las leguminosas como alternativa en la alimentación de animales monogástricos. Informe final de proyecto. PNCT No 008. "Producción de alimento animal por vías biotecnológicas". Instituto de Ciencia Animal, La Habana, Cuba.
Duncan, D.B. 1955. Multiple range and multiple F test. Biometrics 11, 1-14. Grant, G., McKenzie, N.H., Watt, W., Stewart, J.C., Dorward, P.M. & Pusztai, A. 1986.
Nutritional evaluation of soybeans (Glycine max). Nitrogen balance and fractionation studies. Journal of the Science of Food and Agriculture 37, 1001-1010.

47
Honke, J., Kozlowska, H., Vidal-Valverde, C., Frías, J. & Górecki, R. 1998. Changes in quantities of inositol phosphate during maturation and germination of legume seeds. Lebensmittel Untersuchung und - Forschung 206, 279-283.
Kakade, M.L., Simons, N.R., Liener, I.E. & Lambert, J.W. 1972. Biochemical and nutritional assessment of different varieties of soybean. Journal of Agricultural and Food Chemistry 20, 87-90.
Kiers, J.L., Nout, R.M.J. & Rombouts, F.M. 2000. In vitro digestibility of processed and fermented soya bean, cowpea and maize. Journal of the Science of Food and Agriculture 80, 1325-1331.
Lees, R. 1969. Manual de Análisis de Alimento. Edición Acribia, Zaragoza. pp. 231. Martín-Cabrejas, M.A., Ariza N., López Amorós, M.L., Mollá, E. & López-Andreus, F.J.
1998. Determinación del contenido de inhibidores de quimotripsina en leguminosas. Alimentaria 98/99, 99–102.
Martín-Cabrejas, M.A., Esteban, R.M., Waldron, K.W., Maina, G., Grant, G., Bardocz, S. & Pusztai, A. 1995. Hard-to-cook phenomenon in beans: Changes in antinutrient factors and nitrogenous compounds during storage. Journal of the Science of Food and Agriculture 69, 429-435.
Nnam, N.M. 1999. Nitrogen and mineral utilization of young children fed blends of fermented or unfermented corn (Zea mays L.) African yam bean (Sphenostylis stenocarpa) and cowpea (Vigna unguiculata). Ecology of Food and Nutrition 38, 21-34.
Oluwatosin, O.B. 1999. Genotype x environment influence on cowpea (Vigna unguiculata (L) Walp) antinutritional factors: 1. Trypsin inhibitors, tannins, phytic acid and haemagglutinin. Journal of the Science of Food and Agriculture 79, 265-272.
Piergiovanni, A.R. 1992. Effect of some experimental parameters on activity of cowpea α-amylase inhibitors. Lebensmittel-Wissenschaft und-Technologie 25, 231-234.
Porter, L.J., Hritisch, L.N. & Chen, B.G. 1986. The conversion of procyanidins and prodelphinidins to cyanidin and delphinidin. Phytochemistry 25, 223–230.
Uwaegbute, A.C., Iroegbu, C.U. & Eke, O. 2000. Chemical and sensory evaluation of germinated cowpeas (Vigna unguiculata) and their products. Food Chemistry 68, 141-146.
Yoshida, K., Sato, Y., Okuno, R., Kameda, K., Isobe, M. & Kondo, T. 1996. Structural analysis and measurement on anthocyanins from colored seed coats of Vigna, Phaseolus, and Glycine legumes. Bioscience, Biotechnology and Biochemistry 60, 589 -593.


49
Using near infrared reflectance spectroscopy (NIRS) for ANFs analysis in oilseed Brassicas R. Font, M. del Río & A. de Haro-Bailón Instituto de Agricultura Sostenible, Departamento de Agronomía y Mejora Genética Vegetal, Finca Alameda del Obispo s/n, 14080 Córdoba, Spain Summary Brassica oilseeds are grown primarily for the oil and protein contained in seed making them of great value for human and animal nutrition, and also for industry. However, the antinutritional factors (ANFs) contained in the oil (erucic acid) and meal (glucosinolates and fibre) makes Brassica seed unacceptable for human and animal nutrition. Standard methods for ANFs analysis are expensive and time-consuming. In contrast, Near Infrared Reflectance Spectroscopy (NIRS) is a rapid, non-destructive and economical method of analysis that works without the use of hazardous chemicals. In this paper, we test the possible use of NIRS for determining ANFs in intact Brassica sp. seed. The r2 and RPD of the equations for erucic acid (EA), glucosinolates (GSL) and acid detergent fibre (ADF) in the external validation were, respectively, 0.91 and 3.28 (% oil); 0.82 and 2.18 µmol/g DW and 0.83 and 2.40 (% DW). Our results indicate that NIRS can predict ANFs in the Brassica seed with sufficient accuracy for screening. Keywords: Brassica, NIRS, erucic acid, ADF, glucosinolates Introduction The germination capacity of Brassica seed and plant resistance to low temperatures, have made Brassica oilseeds one of the few edible oil crops that can be cultivated in the temperate agricultural zones of the world, at high elevations and, as winter crops, under relatively cool growing conditions (Kimber & McGregor, 1995). Species such as Ethiopian mustard (Brassica carinata A. Braun) and Indian mustard (B. juncea L. Czern. & Coss.) can be grown under environmental stress conditions in hot climates, and can be cultivated as oilseed crops under Mediterranean conditions (Fereres et al., 1983).
Brassica oilseeds are mainly used for their high oil and meal contents. Among the different uses of the oil are edible oils used to produce margarine for human consumption, and industrial use, mainly as engine lubricants and additives, as well as their use in the plastic and nylon industries. On the other hand, the residual meal is a good source of protein for human and animal nutrition.
However, Brassica oil and meal contain antinutritional factors (ANFs) that can make them unacceptable for monogastric nutrition. From a plant breeding point of view the main ANFs are the erucic acid (EA) in the oil for B. carinata, glucosinolates (GSL) in B. juncea, and fibre in all three allotetraploid species.
The standard methods of analysis used to determining these ANFs are expensive, time-consuming and requires specialised personnel. This makes the analysis of large numbers of seed samples difficult for monitoring ANF levels, as is required in plant breeding programs.
In the last 30 years, NIRS has been widely used as a fast and accurate method of qualitative and quantitative analysis in many fields (Williams & Norris, 1987). The

50
Department of Agronomy and Plant Breeding (DAPB) of the Institute for Sustainable Agriculture (IAS, CSIC, Córdoba), has been using NIRS for fifteen years to determine seed quality components in different plant species (De Haro et al., 1989; Font et al., 1998, 2002). The most attractive features of NIRS analysis are its speed, minimal sample preparation and it is non-destructive nature. It is thus possible to analyse large numbers of samples quickly.
The objective of this work was to test the potential of NIRS for determining EA, GSL and acid detergent fibre (ADF) levels in intact Brassica seed. Materials and methods Plant material Plant material used in this work was form the oilseed species B. juncea, B. carinata and B. napus, from the germplasm collection at the DAPB. This collection comprises accessions of different geographical origin and represents most of the genetic variability among these species. The plants were grown over different years, in Córdoba (Spain). They were harvested individually and their seed was stored for NIRS analysis. Reference analysis methods Seed EA content was determined by gas liquid chromatography (Garcés & Mancha, 1993). The AOAC method (AOAC, 1990 method 989.03) was used to determine ADF and GSL content determined as per Quinsac & Ribaillier (1985). NIRS analysis
The NIRS analysis consisted in the following steps: 1. Seed samples of each Brassica species were scanned with a NIRS spectrometer model 6500 (Foss-NIRSystems, Inc., Silver Spring, Md) in reflectance mode, equipped with a transport module, and their NIR spectra recorder as log (1/Reflectance) in the range 400 to 2500 nm, as independent files.
The EA level was analysed in B. carinata samples, and the GSL content in B. juncea samples. For ADF analysis the seed samples were from B. juncea, B. carinata and B. napus. For EA and GSL analysis, representative samples of B. carinata and B. juncea files, respectively, containing the whole spectral variability in the germplasm collection, were selected for calibration (66 % of the whole set). The rest were used for validation. Spectral selection was done by determining the Mahalanobis (H) distance of each spectrum from the mean population spectrum. The selected samples were then analysed by the reference methods for these parameters. For the ADF analysis, the spectra files of all three Brassica species were considered together and the calibration and validation groups were made, following the same procedure as in EA and GSL determination.
Calibration equations for the different components were developed on the calibration sets using Modified Partial Least Square (MPLS) regression (GLOBAL v. 1.50 program, WINISI II, Infrasoft International, LLC, Port Matilda, Pa), with different mathematical pre-treatments [0,0,1,1 (derivative, gap, first smooth, second smooth); 1,4,4,1; 2,5,5,2] of the original spectra. The equations obtained by the calibration process were then validated on the validation sets, to test the prediction ability of each equation over samples of the same characteristics. The prediction ability of the different equations for each parameter was evaluated attending their coefficients of determination in the external validation (r2) and ratio

51
of the standard deviation (SD) to standard error of prediction (SEP), which is known as RPD (Williams & Sobering, 1996). Results and discussion Second derivative transformation of the raw optical data (2, 5, 5, 2) gave the best combination of r2 and RPD for EA and ADF; while the first derivative transformation (1, 4, 4, 1) of the original spectra, showed a high r2 and RPD for GSL. The selected calibration equations for EA, GSL and ADF resulted in standard errors of calibration (SECs) of 3.11 (% oil, DW), 17.00 (µmol/g, DW) and 0.60 (% DW). The coefficients of determination in the calibration (R2) were 0.96, 0.80 and 0.82, respectively (Table 1). This indicates equations, which are of good (GSL) to excellent (EA and ADF) precision (Shenk & Westerhaus, 1996).
The external validation indicated that the three seed components were predicted with sufficient accuracy for screening purposes, as their RPD values were > 3 or close to it (Williams & Sobering, 1996). The level of precision shown by the equations in the calibration was then validated. The external validation confirmed the predictive ability shown by the three components in the calibration.
Biston et al. (1988), Davis et al. (1991) and Font et al. (2003) have extensively used NIRS as an alternative method for ANF analysis in Brassica seed. (The predictive abilities of the equations for ANFs reported by these authors varied widely depending on factors such as species analysed, sample pre-treatment (intact or ground seed) the spectroscopic technique used (reflectance or transmittance), and by the ranges and SDs of the sample groups used in their studies. Table 1. Calibration and external validation statistics for erucic acid (EA) (n = 67),
glucosinolates (GSL) (n = 208), and acid detergent fibre ADF (n = 150) for the selected equations.
Calibration External validation ANF Range Mean SD SEC R2 Range Mean SD RPD R2 EA1 0.01 - 50.6 34.0 15.3 3.1 0.96 0.03 - 49.7 33.7 15.1 3.3 0.91
GSL2 16.1 -196.7 123.7 37.6 17.0 0.80 21.9 - 187.3 127.6 34.2 2.2 0.82ADF3 5.3 - 16.3 11.0 2.2 0.6 0.92 6.7 -15.5 11.0 2.0 2.4 0.831B. carinata. 2B. juncea. 3B. carinata, B. juncea and B. napus. Conclusions The results of this work show that NIRS can determine the EA, GSL and ADF content of intact Brassica seed samples with sufficient accuracy for screening and plant breeding purposes. This non-destructive technique reduces the time required for analysis, is low cost, does not use of hazardous chemicals and leaves the seed undamaged. References AOAC 1990. AOAC method 989.03. In Official Methods of Analysis, 15th Edition. K.
Helrich (Ed.). Association of Official Analytical Chemists, Arlington, Va. Volume 2, 781-782.
Biston, R., Dardenne, P., Cwikowski, M., Marlier, M., Severin, M. & Wathelet, J.P. 1988. Fast analysis of rapeseed glucosinolates by near infrared reflectance spectroscopy. Journal of the American Oil Chemists Society 65, 1599-1600.

52
Davis, J.B., Hall, M.H., Eckert, J.W., Corsini, J.A. & Auld, D.L. 1991. Comparison of near-infrared reflectance analyzes with GC analyzes of glucosinolate concentration in rapeseed. EUCARPIA Cruciferae Newsletter 14-15, 120-121.
De Haro, A., López-Medina, J., Cabrera, A. & Martín, A. 1989. Determination of tannin in the seeds of Vicia faba by NIR. In Recent Advances of Research in Antinutritional Factors in Legume Seeds. J. Huisman, T.F.B. van der Poel & I.E. Liener. Proceedings 1st International Workshop on Antinutritional Factors in Legume Seeds, Wageningen 23–25 November 1988, 297-300.
Fereres, E., Fernández-Martínez, J.M., Minguez, Y. & Domínguez, J. 1983. Productivity of Brassica juncea and Brassica carinata in relation to rapeseed, B. napus. I. Agronomic studies. In Proceedings 6th International Rapeseed Congress, Paris, 17-19 May, 1983, 293-298.
Font, R, Del Río, M, Fernández, J, Bancroft, A.E., Chinoy, C., Morgan, C. & De Haro, A. 2002. Seed oil content analysis of Ethiopian mustard (Brassica carinata A. Braun) by near infrared spectroscopy. EUCARPIA Cruciferae Newsletter 24, 5-6.
Font, R, Del Río, M., Fernández-Martínez, J.M. & De Haro, A. 1998. Determining quality components in Indian mustard by NIRS. EUCARPIA Cruciferae Newsletter 20, 67-68.
Font, R., Del Río, M., Fernández-Martínez, J.M. & De Haro, A. 2003. Acid detergent fiber analysis in oilseed brassicas by near infrared spectroscopy. Journal of Agricultural and Food Chemistry 51, 2917-2922.
Garcés, R. & Mancha, M. 1993. One step lipid extraction and fatty acid methyl esters preparation from fresh plant tissues. Analytical Biochemistry 211, 139-143.
Kimber, D.S. & McGregor, D.I. 1995. The species and their origin, cultivation and world production. In Brassica Oilseeds. Production and Utilization. D.S. Kimber & D.I. McGregor (Eds). CAB International, Wallingford. pp. 1-7.
Quinsac, A. & Ribaillier, D. 1985. Quantitative analysis of glucosinolates in rapeseed seeds. Optimization of desulphatation. In Advances in the Production and Utilization of Cruciferous with Special Emphasis to Oilseed Rape. World crops: Production, Utilization, Description. H. Sorensen (Ed.). Kluwer Academic Publishers, Dordrecht. Volume 11. pp. 85-96.
Shenk, J.S. & Westerhaus, M.O. 1996. Calibration the ISI way. In Near Infrared Spectroscopy: The Future Waves. A.M.C. Davies & P.C. Williams (Eds.). NIR Publications, Chichester. Pp. 198-202.
Williams, P.C. & Norris, K.H. 1987. Near-infrared Technology in the Agricultural and Food Industries. P.C Williams & K.H. Norris (Eds.). American Association of Cereal Chemists, St. Paul, Mn.
Williams, P.C. & Sobering, D.C. 1996. How do we do it: a brief summary of the methods we use in developing near infrared calibrations. In Near Infrared Spectroscopy: The Future Waves. A.M.C. Davies & P.C. Williams (Eds). NIR Publications, Chichester. pp. 185-188.

53
Phytate and αααα-galactoside content of wild lupins from Jalisco, México P.M. García-López1, C.M. Gurrola Díaz2, P. Garzón de la Mora2, M.L. Isaac Virgen2, M.A. Ruiz Lopez1, C. Cuadrado3, C. Burbano3, M.M. Pedrosa3 & M. Muzquiz3 1Laboratorio de Biotecnología, Departamento de Botánica y Zoología, Universidad de Guadalajara, Aptdo. 139, Zapopan, Jalisco, 45100 México 2Instituto de Enfermedades Crónico Degenerativas, Universidad de Guadalajara, CUCS, Guadalajara, Jalisco, México 3Departamento Tecnología de Alimentos, SIGT-INIA, Aptdo 8111, 20080, Madrid, Spain Summary The phytate and α-galactoside content of Lupinus montanus and L. stipulatus, seed was measured. The phytate content of these lupin seeds ranged from 2.82 to 3.66 mg/g. The main
α-galactosides in both lupin seeds were raffinose (4.8 - 9.3 mg/g), stachyose (33.0 - 43.1 mg/g), and verbascose (8.0 - 15.1 mg/g). The level and type of phytates and α-galactosides in these wild lupins are similar to those already reported for other sweet or bitter lupins. Keywords: Lupinus montanus, L. stipulatus, phytates, α-galactosides Introduction Almost 100 wild species of the genus Lupinus have been reported in Mexico. Lupinus montanus, and L. stipulatus, are two of 15 species described from the Mexican state of Jalisco (McVaugh, 1987). These species also grow in other Mexican states like Estado de México and Guanajuato (Dunn, 1979; McVaugh, 1987).
Like other wild lupins from South America, Europe and the Mediterranean region, the seed of these Mexican lupins have high protein, dietary fibre and alkaloid contents (Ruiz et al, 2000). Alkaloids are the main constraint to the consumption of both wild and domesticated lupins. However, the presence of oligosaccharides and phytates could be additional antinutritional factors (ANFs) to their use in human and animal nutrition (Trugo & von Baer, 1998).
The aim of this study was to determine, the oligosaccharide and phytate content of seed from populations of L. montanus and L. stipulatus collected from different locations in Mexico in 1996/97.
Materials and methods Experimental Seed of bitter L. montanus and L. stipulatus, for this study, was collected from several populations in 1996/97 from different locations in the Mexican state of Jalisco. Lupin seed was ground to pass through a 1mm sieve (Tecator, Cyclotec 1093).

54
αααα-Galactosides The α-Galactosides were extracted using ion exchange columns (Muzquiz et al., 1992). Samples (20 µl) were analysed using a Beckman HPLC chromatograph fitted with a 156 refractive index detector. A Spherisorb-5-NH2 column (250 × 4.6 mm id) was used with acetonitrile/water (1 ml/min), 65:35 (v/v) as the mobile phase. Individual sugars were quantified by comparison with external standards of sucrose, raffinose, stachyose and verbascose. Calibration curves were drawn for all four sugars. There was a linear response in the range 0 - 5 mg/ml and the determination coefficient was 0.99. Inositol phosphates The individual inositol phosphates (IP3-IP6) were extracted according Muzquiz et al. (1999) and measured by HPLC (Lehrfeld, 1994). Analysis was with a Beckman System Gold HPLC equipped with a refractive index. The column was a macrosporous polymer PRP-1 (150 × 4.1 mm, 5µm) heated at 45 °C and was equilibrated with the mobile phase for 1 h. The mobile phase was 515 ml of methanol with 485 ml of water. Eight ml of tetrabutylammonium hydroxide (40 % in water), 1 ml 5M sulphuric acid, 0.5 ml 91 % formic acid and 100 µl of a phytic acid hydrolysate (6 mg/ml) were added sequentially. The pH was adjusted to 4.3 with 9 M sulphuric acid. The mobile phase was filtered through a Millipore filter (0.45 µm) and degassed under a vacuum. The flow rate was 1.2 ml/min and the injection volume was 20 µl. The standard used was sodium phytate (Sigma Chemicals, USA). There was a linear response in the range 0.36 - 7.3 mg/ml. Results and discussion αααα-Galactosides Table 1 shows the oligosaccharide composition of L. montanus, and L. stipulatus seed collected from several locations in the state of Jalisco. Total α-galactosides were in the range of 53.8 to 60.0 mg/g DM. This was lower than values found in domesticated lupins such as L. albus, L. mutabilis, L. angustifolius but was similar to levels in L. hispanicus and L. consentinii (Trugo& Almeida, 1988; Zdunczyk et al, 1998).
Table 1. The α-galactoside content (mg/g) of L. montanus and L. stipulatus collected at
several locations in Jalisco, Mexico in 1996/97. Lupin species L. montanus L. stipulatus Sugar CV1 CV2 B B
Sucrose 6.4 ± 0.25 3.1 ± 0.01 4.6 ± 0.05 5.3 ± 0.05 Raffinose 5.9 ± 0.21 9.3 ± 0.08 4.8 ± 0.02 6.8 ± 0.27 Stachyose 43.1 ± 1.09 38.7 ± 0.02 36.5 ± 0.09 33.0 ± 0.45 Verbascose 8.0 ± 0.02 12.0 ± 0.35 15.1 ± 0.01 13.9 ± 0.50
Total α-galactosides 57.0 ± 1.09 60.0 ± 0.25 56.4 ± 0.23 53.8 ± 0.68 1Colima' Volcano, 1996; 2Colima' Volcano: El Refugio, 1997; B, Bolaños, 1997.
Stachyose was the main oligosaccharide present in the lupin seed. Its concentration at
33.0 to 43.1 mg/g was higher than reported for L. hispanicus, L. luteus, L. angustifolius,

55
L. albus and L. mutabilis, but lower than in the Polish cultivars of L. albus, Bardo and Bac (Zdunczyk et al, 1998). The amount of this sugar in L. montanus, and L. stipulatus seed was 60 % of the total seed sugar content. This is slightly higher than the 53 % reported by Trugo & Almeida (1988) in domesticated and wild lupins.
The highest raffinose and verbascose concentration in total oligosaccharides of the two species was in L. montanus seed from Colima' Volcano: El Refugio (1997) at 9.3 mg/g and 12.0 mg/g, respectively. The raffinose concentration in L. montanus from Colima' Volcano (1996), and Bolaños (1997) and in L. stipulatus from Bolaños, (1997), was lower than in L. hispanicus. However, the amount in L. montanus from Colima' Volcano: El Refugio (1997) was higher (Muzquiz et al., 1989). The verbascose level in both species was lower than in L. luteus (Saini, 1989), unlike the Polish L. albus cultivars Bardo and Bac (Zdunczyk et al, 1998). Lupin seed sucrose content ranged from 3.1 mg/g in L. montanus from Colima' Volcano: El Refugio (1997) to 6.4 mg/g in L. montanus from Colima' Volcano (1996). These values are, lower than the in L. hispanicus, hispanicus, L. hispanicus bicolor, and the L. albus cultivars Bardo and Bac (Muzquiz, et al, 1989, Zdunczyk et al, 1998). Although it is known that genotype and environment can modify the proportions and total oligosaccharide content in lupin seed it was not possible to ascertain if these factors had any effect on the proportion and types of sugars found in L. montanus and L. stipulatus seed. Inositol phosphates Table 2 shows the inositol phosphate composition of the L. montanus, and L. stipulatus seed. Total phytates ranged from 2.82 to 3.97 mg/g. These values are lower than those reported by Burbano et al (1995). On the other hand the total inositol phosphates in L. stipulatus from Bolaños (1997) at 2.82 mg/g was lower than levels in L. albus (3.00 mg/g) and L. luteus (3.25 mg/g) (Muzquiz, et al 1998).
Table 2. The Inositol phosphate content (mg/g) of L. montanus and L. stipulatus seed
collected at several locations in Jalisco, Mexico 1996/97. Lupin species L. montanus L. stipulatusInositol phosphate CV1 CV2 B B
IP4 0.03 ± 0.0 0.05 ± 0.02 0.10 ± 0.02 0.11 ± 0.00 IP5 0.39 ± 0.0 0.55 ± 0.19 0.73 ± 0.09 0.65 ± 0.03 IP6 3.23 ± 0.0 3.37 ± 0.66 2.60 ± 0.11 2.06 ± 0.13
Total 3.66 ± 0.0 3.97 ± 0.87 3.42 ± 0.22 2.82 ± 0.16 1CV= Colima' Volcano 1996. 2CV= Colima' Volcano: El Refugio, 1997, B, Bolaños 1997
The amount of inositol phosphate IP6 in all the samples was lower than in L. luteus
(7.17 mg/g), Phaseolus vulgaris, and Vicia faba, but was similar to the level in Lupinus albus, Lens culinaris, and Cicer arietinum (Burbano et al, 1995). On the other hand the levels of inositol phosphate, in the form IP4 and IP5, in L. montanus from Colima' Volcano (1996), Colima' Volcano: El Refugio (1997), and Bolaños (1997) and in L. stipulatus were lower and higher, respectively than in L. luteus and L. albus. The form IP3, which is present in L. luteus and L. albus, was not found in any of lupin samples (Burbano et al 1995).

56
References Burbano, C., Muzquiz, M., Osagie, A., Ayet, G. & Cuadrado, C. 1995. Determination of
phytate and lower inositol phosphates in Spanish legumes by HPLC methodology. Food Chemistry 52, 321-325.
Dunn, D.B. 1979. Lupinus. In Flora Fanerogamica del Valle de México. J. Rzedowski & G.C. Rzedowski (Eds). Instituto de Ecología, México, DF. pp. 326-338.
Lehrfeld, J. 1994. Separation and quantification of phytic acid and some inositol phosphates in foods: problems and solutions. Journal of Agricultural and Food Chemistry 42, 2726-2731.
McVaugh, R. 1987. Flora Novo-Galiciana. A Descriptive Account of Vascular Plants of Western Mexico. Vol. V. Leguminosae. University of Michigan Press, Ann Arbor.
Muzquiz, M., Burbano, C., Ayet, C., Pedrosa, M.M. & Cuadrado, C. 1999. The investigation of antinutritional factors in Phaseolus vulgaris. Environmental and varietal differences. Biotechnologie Agronomie Société et Environnement 3, 210-216.
Muzquiz, M., Burbano, C., Gorospe, M.J. & Ródenas, I. 1989. A Chemical study of Lupinus hispanicus seed-toxic and antinutritional components. Journal of the Science of Food Agriculture 47, 205-214.
Muzquiz, M., Pedrosa, M.M., Cuadrado, C., Ayet, G., Burbano, C., & Brenes, A. 1998. Variation of alkaloids, alkaloids esters, phytic acid and phytase activity in germinated seed of Lupinus albus and Lupinus luteus. In Recent Advances of Research in Antinutritional Factors in Legume Seeds and Rapeseed. A.J.M Jansman, G.D. Hill, J. Huisman & A.F.B. van der Poel (Eds). Proceedings of the 3rd International Workshop on Antinutritional Factors in Legume Seeds and Rapeseed, Wageningen, 8–10 July 1998, 387-390.
Muzquiz, M., Rey, C., Cuadrado, C. & Fenwick, G.R. 1992. Effect of germination on the oligosaccharides content of lupin species. Journal of Chromatography 607, 349-352.
Ruiz López, M.A., García López, P.M., Castañeda Vázquez, H., Zamora, N.J.F., Garzón de la Mora, P., Bañuelos Pineda, J., Burbano, M., Pedrosa, M., Cuadrado, C. & Muzquiz, M. 2000. Chemical composition and antinutrient content of three Lupinus species from Jalisco, Mexico. Journal of Food Composition and Analysis 13, 193-199.
Saini, H.S. 1989. Legume seed oligosaccharides. In Recent Advances of Research in Antinutritional Factors in Legume Seeds. J. Huisman, T.F.B. van der Poel & I.E. Liener, (Eds). Proceedings of the 1st International Workshop on Antinutritional Factors in Legume Seeds, Wageningen 23–25 November 1988, 329-341.
Trugo, L.C. & Almeida, D.C.F. 1988. Oligosaccharide contents in the seeds of cultivated lupins. Journal of the Science of Food Agriculture 45, 21-24.
Trugo, L.C. & Baer, D. von, 1998. Analytical methods for the analysis of antinutritional factors in legume seeds. In Recent Advances of Research in Antinutritional Factors in Legume Seeds and Rapeseed. A.J.M Jansman, G.D. Hill, J. Huisman & A.F.B. van der Poel (Eds). Proceedings of the 3rd International Workshop on Antinutritional Factors in Legume Seeds and Rapeseed, Wageningen 8–10 July 1998, 11-28.
Zdunczyk, Z., Juskiewicz, J., Frejnagel, S. & Gulewicz, K. 1998. Influence of alkaloids and oligosaccharides from white lupin seeds on utilization of diets by rats and adsorption of nutrients in the small intestine. Animal Feed Science Technology 72, 143-154.

57
Variation in alkaloid type and content during Erythrina americana seed development R. García-Mateos1, M. Soto-Hernández2 & M. Martínez3
1Universidad Autónoma Chapingo, Departamento de Preparatoria Agrícola, Carretera México-Texcoco Km 38.5, 56230 Chapingo, México 2Colegio de Postgraduados, Programa de Botánica, Carretera. México-Texcoco, Km 35.5, 56230 Montecillo, México 3Instituto de Química, Universidad Nacional Autónoma de México, Ciudad Universitaria, Coyoacán, Distrito Federal 04510, México Summary The flowers, green beans, seeds and pod valves of Erythrina americana were studied to determine the level and type of alkaloid present at different stages of pod development. The moisture content of the mature and dry seeds was 78.9 and 5.7 % respectively. The alkaloids were present in young tissues and at seed maturity. On a dry matter basis there was a high alkaloid content in flowers and dry seed and low levels in dry pods. The lowest alkaloid concentration was in mature seeds. A gas chromatography-mass spectrometry (GC/MS) analysis showed β-erythroidine was present. It was the major alkaloid in all the tissues examined. Erysodine and erysovine were found in the mature and dry tissues but not in flowers. The N-oxide alkaloids were only found in mature tissues. Keywords: alkaloids, Erythrina americana, gas chromatography-mass spectrometry,
physiology Introduction Erythrina is a large genus in the Legume family. Plants in it display a wide range of morphological variation and ecological diversity. It is prominent among trees and is attracting increased research and development attention. It is distributed throughout the tropics. The greatest concentration of Erythrina spp. is found in southern México (27 species) and in Central America (Neill, 1993). Many of the approximately 115 Erythrina spp. are used in agroforestry systems as fences, windbreaks, shade or support for other plants (Neill, 1993; Russo, 1993). As a typical legume it improve the soil and provides animal fodder, human food, medicines and wood products. Their unique flowers and seeds also make them popular as ornamental handcrafts (Musálem, 1993). These attributes are diminished by the toxicity of the plants because they accumulate alkaloids in all plant parts but particularly in the seed. Erythrina alkaloids can be considered typical for this genus; they have unusual structures and have a restricted distribution in the Leguminosae. Flowers and seed of E. americana are rich in alkaloids (Aguilar et al., 1981), this suggest that these are possible accumulation sites.
During pod development there is translocation of nitrogen compounds such as amino acids, alkaloids and amides, which accounts for these compounds being the major nitrogen compounds in the mature seed and in the pods. The free amino acids concentration decreases when storage protein and other nitrogen compounds such as alkaloids are formed (Salisbury and Ross, 1994).

58
Sotelo et al. (1993) and Robinson (1979) suggested that the development and maturation process in a plant can affect alkaloid synthesis, according to their relative concentration and structural diversity. Waller & Nowacki (1978) assumed that during plant morphological and physiological processes, the alkaloid profile can be modified.
Few studies have been done on changes of alkaloid content and type during pod development and nothing has been reported in the literature on these changes in Erythrina spp. The aim of this work was to determine changes in the different alkaloid types during of pod development from flowering to dry seed. Materials and methods Plant material Flowers, green beans, seed and pods at different development stages were collected. They were collected at the following intervals: flower → green bean (whole pods present) (0 - 30 d); green bean →→→→ mature pod (without seed) and seed (31 - 45 d); mature pods and seed →→→→ dry pods and seeds (46 - 96 d). The maturation process lasted approximately 96 d. Samples were collected from plants growing in the gardens of the Chemistry Faculty at the University Campus in México City. A botanist from the Herbarium of the Colegio de Postgraduados, Montecillo, Estado de México certified their authenticity. Alkaloid extraction All plant samples were air-dried and milled separately. They were extracted with hexane by Soxhlet extraction for 48 h to remove fats. Alkaloids were extracted by the Games et al (1974) method. This method gives, through partitioning two fractions: i) a free alkaloid fraction (hexane and methanol soluble) which are non-conjugated and ii) a liberated alkaloid fraction obtained after acid hydrolysis because in it the alkaloids are conjugated with sugars. Gas chromatography-Mass spectrometry (GC/MS) The total alkaloid mixture (free and liberated alkaloids) was analysed as trimethylsilyl (TMSi) derivatives according to Games et al. (1974). Alkaloid identification was by comparison with samples previously purified and characterized in our laboratory (erysodine, erysovine, erythraline, α- and β-erythroidines, erysopine), interpretation of mass spectra characteristics and use of reference data. Results and Discussion In our study free alkaloids included the hexane and methanol soluble fraction, as the hexane fraction contains significant amounts of alkaloids (Hargreaves et al., 1974; Sotelo et al., 1993).
The alkaloid yield of each of the plant parts examined is shown in Table 1. Free alkaloids were most common in all the tissues analysed except in dry pods where liberated alkaloids were at the same concentration as free alkaloids.
The quantitative analysis of total alkaloids (free and liberated alkaloids) showed interesting trends in variation of alkaloid concentrations during pod maturation. For example flowers synthesized moderate levels of alkaloids mainly as free alkaloids, synthesis declined at the green bean stage and attained their highest accumulation in dry seed. This means that

59
the alkaloid levels increased during seed maturation. This conforms to Waller & Nowacki (1978). Table 1. The alkaloid content (g/100 g DM) during pod development of Erythrina americana.
Development stage
Component Flowers Green beans
Mature seed
Mature podsa Dry seed Dry pods
Total alkaloids 1.10 0.39 0.17 0.64 1.05 0.27 Free alkaloidsb 1.02 0.21 0.15 0.58 0.75 0.13 Liberated alkaloids 0.08 0.18 0.02 0.06 0.30 0.14
a = mature pods (without seeds); b = free alkaloids (hexane + methanol fraction)
The methanol soluble alkaloids of E. americana were identified by GC/MS. In the analysis it was convenient to divide the alkaloids into two groups, those that contain a conjugated 1,6 diene system (dienoid alkaloids) and those, which contain an isolated 1(6)-double bond (alkenoid alkaloid). The structures of alkaloids identified are shown in Fig. 1-4.
The mass spectra of all of the dienoid alkaloids showed essentially the same fragmentation pattern. The major peaks were at M+, M+-15 (m/z 356), M+-31 (m/z 340), M+-58 (m/z313), M+-72 (m/z 299) and M+-85 (m/z 286) (Boar & Widdowson, 1970).
Figure 1. Structure of dienoid alkaloids. Figure 2. Structure of alkenoid alkaloids.
Dienoid Alkaloid R1 R2 R3 R4 Alkenoid Alkaloid R1 R2 R3 R4
Erysodine H Me Me H Erythratidine Me Me Me α-OH Erysovine Me H Me H Erysotrine Me Me Me H 11β-Methoxyerythraline -CH2 Me MeO 11β-Hydroxyerysovine Me H Me OH
The fragmentation of the second group of Erythrina alkaloids (e.g. erythratidine), those having a 1(6)-double bond, was more complex and varied more than those of the above group. The ions M+-15 (m/z 388) and M+-31 (m/z 372) were of relatively minor importance, but the ion M+-58 (m/z 345) that correspond to a retro Diels-Alder reaction in ring A was the major peak in this group.
The rare alkaloids erythartine-N-oxide and erythristemine-N-oxide were also observed. They showed the typical fragmentation pattern of the diene aromatic alkaloids besides the fragment of M+-16 that supported the evidence of the N-O residue. The non-aromatic α- and β-erythroidines (Fig. 4), respectively, belong to the diene group and fragment
R1
R2
R3
123
45
6 7
8
1011
14
15
16
17O
O
O
R4
N
R1
R2
R3
R4
O
O
O
N

60
by loss of the methoxyl substituent at C-3. α-erythroidine showed a M+ m/z 273 and a major fragment ion at m/z 242 (M+-31). The derivative of β-erythroidine-TMS showed a similar fragmentation pattern except for the presence of an intense ion at m/z 73, M+ at m/z 345 and an intense ion at m/z 130 due to fragment C2HO-TMS. These results confirm that the alkaloids α-erythroidine, and β-erythroidine are found in all stages of fruit maturation including in flowers (Aguilar et al., 1981; Sotelo et al., 1993). The explanation of this pattern can be inferred through their biosynthesis because, as Barton et al. (1974) pointed out, erysodine is one of the main alkaloids that remains until the end of biosynthesis. Only alkylation or dealkylation of the phenolic group or decarboxylation of ring D determine the transformation of erysodine as precursor of the lactonic alkaloid β-erythroidine through several intermediate stages (Dyke & Quessy, 1981), so this compound is the final product of the biosynthetic pathway in the plant material that was examined. It is possible that of all the possible biosynthetic precursors, mainly the aromatic Erythrina alkaloids, are shunted into this pathway and are converted mainly into β-erythroidine. Thus the aromatic Erythrina alkaloids precursors may not be present, or detected in significant amounts, because are rapidly converted to β-erythroidine.
Figure 3. Structure of the N-oxide alkaloids N-oxide Alkaloid R1 R2 R3 R4
Erythrartine-N-oxide Me Me Me OH Erythristemine-N-oxide Me Me Me MeO
α-erythroidine β-erythroidine
Figure 4. Structure of the non-aromatic alkaloids. Conclusions Alkaloids accumulation was not only observed at the end of seed maturation but also in young tissues like flowers, mature pods and seed. The alkaloid level increased during pod maturation. These results were supported with a useful structural probe with mass spectrometry that allowed the dienoid alkaloids to be distinguished from alkenoid or the non-aromatic alkaloids.
O+
-R1
R2
R3
R4
O
O
O
N
CH3
O
NO
O
O
O
CH3O
N

61
References Aguilar, M.I., Giral, F. & Espejo, O. 1981. Alkaloids from the flowers of Erythrina
americana. Phytochemistry 20, 2061-2062. Barton, D.H.R., Potter, C.H. & Widdowson, D.A. 1974. Phenol oxidation and biosynthesis,
Part XXIII. On the benzyltetrahydroisoquinoline origins of the Erythrina alkaloids. Journal of the Chemical Society Perkin I, 346-348.
Boar, R.B., Widdowson, D.A. 1970. Mass spectra of the Erythrina alkaloids: a novel fragmentation of the spiran system. Journal of the Chemical Society B, 1591-1595.
Dyke, S.F. & Quessy, S.N. 1981. Erythrina and related alkaloids. In The Alkaloids, Vol. 18. R.F.H. Manske (Ed.). Academic Press, New York. pp. 1-98.
Games, D.E., Jackson, A.H., Khan, N.A. & Millington, D.S. 1974. Alkaloids of some African, Asian, Polynesian and Australian species of Erythrina. Lloydia 37, 581-588.
Hargreaves, R.T., Johnson, R.D., Millington, D.S., Mondal, M.H., Beavers, W., Becker, L., Young, C., Rinehart K.L. 1974. Alkaloids of American species of Erythrina. Lloydia 37, 569-580.
Musálem, M.A. 1993. Erythrina en México: distribución, uso e investigación. In Erythrina in the New and Old Worlds. S.B. Westley & M.H. Powell (Eds). Nitrogen Fixing Tree Association, Hawaii. pp. 46-48.
Neill, D.A. 1993. The genus Erythrina: taxonomy, distribution and ecological differentiation. In Erythrina in the New and Old Worlds. S.B. Westley & M.H. Powell (Eds). Nitrogen Fixing Tree Association, Hawaii. pp. 15-27.
Robinson, T. 1979. The evolutionary ecology of alkaloid. In Herbivores: Their Interactions with Secondary Plant Metabolites. G.A. Rosenthal & D.H. Janzen (Eds). Academic Press, New York. pp. 413-448.
Russo, R.O. 1993. The use of Erythrina species in the Americas. In Erythrina in the New and Old Worlds. S.B. Westley & M.H. Powell (Eds). Nitrogen Fixing Tree Association, Hawaii. pp. 28-45.
Salisbury, F.B. & Ross, C.W. 1994. Crecimiento y Desarrollo. In Fisiología Vegetal. N.G. Philp (Ed.). Grupo Editorial Ibero América, México City. pp. 388-390
Sotelo, A., Soto, M., Lucas, B., Giral, F. 1993. Comparative studies of the alkaloidal composition of two Mexican Erythrina species and nutritive value of the detoxified seeds. Journal of Agricultural and Food Chemistry 41, 2340-2343.
Waller, G.R. & Nowacki, E.K. 1978. Metabolic (catabolic) modifications of alkaloids by plants. In Alkaloid Biology and Metabolism in Plants. G.R. Waller & E.K. Nowacki (Eds). Plenum Press, New York. pp 183-249.


63
Antinutritional factors in different oilseeds usable as renewable resources compared with rapeseed B. Matthäus Institute for Lipid Research, Federal Centre for Cereal, Potato and Lipid Research, Post Box 1705, D-48006 Münster, Germany. Summary Twelve different oilseeds, usable as renewable resources for the production of oil, were investigated for their composition and content of glucosinolates, inositol phosphates, sinapine and condensed tannins. The results were compared with the composition of Brassica napus (rapeseed). All of the seeds contained one or more antinutritional factors (ANFs), some in higher amounts than B. napus. Nevertheless, the use of most of the seeds in animal nutrition may be possible. Only seed of Lepidium sativum, Sinapis alba (high amounts of glucosinolates and sinapine) and Lesquerella fendleri (a high amount of condensed tannins) seemed to be unsuitable. Key words: antinutritional factors, glucosinolates, inositol phosphates, oilseeds, sinapine Introduction An important marketing point for oilseeds is the possibility of complete exploitation of the agricultural product after oil extraction. For a useful product it is necessary not only to exploit the oil, but also the resulting press cake. From an economic point of view the use of the press cake in animal rations gives the highest added value to the product. Most commercial oilseed press cakes are rich in protein and have an interesting amino acid composition. However, they also contain at least one ANF, which lowers their nutritionla value (Bell, 1989; Griffiths et al, 1998). Therefore the current use of press cake in human or animal nutrition is limited.
To assess the potential of oilseed residues for animal rations information on the nature and quantity of the ANFs in the seed is important. Only then is it possible to assign effects on animals to the composition of the oilseed. For many years different plants have been investigated in breeding experiments regarding their potential for oil production. Some of these oilseeds are used as renewable resources, but less is known about their ANF composition.
The aim of this work was to investigate different oilseeds, usable as renewable resources, with regard to their ANFs such as glucosinolates, tannins, sinapine and inositol phosphates. A major objective was to compare the composition of ANFs with the amounts found in Brassica napus (rapeseed), currently one of the most important oilseeds grown in Europe. Materials and methods Seed of Brassica carinata (BC), Camelina sativa (CAM), Coriandrum sativum (COR), Euphorbia lagascae (EU), Lepidium sativum (LE), Lesquerella fendleri (FE), Madia sativa (MAD), Vernonia galamensis (VE), Sinapis alba (SA), Cannabis sativa (HS), Lallemantia

64
iberica (LI), Crambe abyssinica (CA), and Brassica napus (RS) was obtained from different plant breeding programs.
Inositol phosphate determination was by HPLC after HCl extraction and purification with Dowex 1 × 2 (Matthäus et al, 1995). Glucosinolates were determined by HPLC after purification with anion-exchange columns (EC, 1990) and sinapine was extracted from the seed by methanol/water (70:30) and injected into an HPLC (Bjerg et al, 1984). Condensed tannins were determined photometrically using a modified vanillin assay (Butler et al, 1982). Results and discussion Glucosinolates were only found in seed from the family Brassicaceae. Their amount and composition differed among genera (Fig. 1). The amounts in B. napus, Camelina sativa and Lesquerella fendleri seed were 15 µmol/g, 24 µmol/g and 27 µmol/g, respectively. In the other seeds their content was significant higher (71 µmol/g in Crambe abyssinica; 77 µmol/g in Brassica carinata; 138 µmol/g in Sinapis alba and 160 µmol/g in Lepidium sativum). In most of the seeds the glucosinolate composition consisted of one main glucosinolate, which accounted for 92 to 100 % of total glucosinolates. Only in Brassica napus and Camelina sativa was the distribution of the glucosinolates more diversified. The glucosinolate content of Crambe abyssinica, Sinapis alba, Brassica carinata and Lepidium sativum suggested that the use of residues of these seeds would be strongly limited by their high concentration of different glucosinolates. The glucosinolate concentration was at least five times more than in 00-rapeseed.
Figure 1. Composition of glucosinolates from different oilseeds
The main inositol phosphate present in the seeds was inositol hexaphosphate (IP6). The
amount ranged from 6.6 mg/g (Euphorbia lagascae) to 23.1 mg/g (Brassica carinata). Small amounts of inositol pentaphosphate (IP5), as a degradation product were detected in all samples. The content ranged from 0.6 mg/g (Euphorbia lagascae) to 3.8 mg/g (Brassica carinata) (Fig. 2), no other degradation products of IP6 were found. Compared with B. napus only seed of Euphorbia lagascae had significant lower levels of inositol phosphates. Phytic acid, and its degradation products, can cause poor mineral and protein bioavailability but some protective effects are described. Thus by the use of added minerals possible negative effects of this group of compounds could be avoided.
020406080
100120140160180
Glu
cotro
paeo
lin
Prog
oitri
n
Sini
grin
Glu
cona
pin
4-H
ydro
.glu
cobr
.
Glu
cobr
assi
cin
MSG
-9
MSG
-10
MSG
-11
Glu
cotro
paeo
lin
Glu
coib
erin
Sina
lbin
4-H
ydro
.glu
cobr
.
Glu
cona
stur
iin
Prog
oitri
n
Epi-P
rogo
itrin
Glu
cona
pin
4-H
ydro
.glu
cobr
.
PRO
Glu
cona
pin
4-H
ydro
.glu
cobr
.
Glu
cobr
assi
cana
pin
Glu
cobr
assi
cin
Glu
cona
stur
iin
Con
tent
of g
luco
sino
late
s [µ
mol
/g]
BC CS LEFE
SA CA RS

65
Figure 2. Concentration of inositol penta- and –hexaphosphate of different oilseeds.
Sinapine was found in seeds of the family Brassicaceae. The least was in Lesquerella fendleri (0.2 mg/g), while Lepidium sativum contained the most (13 mg/g) (Figure. 3). High amounts of sinapine were also found in Sinapis alba (11.5 mg/g), varying between 0.2 mg/g and nearly 18 mg/g. The variation in seeds of other genera was smaller. Only in seeds of Lesquerella fendleri and C. abyssinica was the amount of sinapine significant lower than in Brassica napus. The other seeds contained comparable or higher amounts. Therefore it could be expected that eggs from hens fed a diet containing these residues could have a crabby or fishy taint (Pearson et al, 1980).
Figure 3. Concentration of sinapine in different oilseeds.
The average condensed tannins content of the seed varied from 0.04 mg gallic acid equivalents (GAE)/g seed (Euphorbia lagascae) to 19.6 mg/g (Lepidium sativum) (Fig. 4). There were high amounts of condensed tannin in seed of Brassica carinata, B. napus, Lepidium sativum and Lesquerella fendleri. In the other seeds the amounts were significantly lower (≤ 2.1 mg/g). A negative effect of the use of such residues as a feed in animal nutrition as a result of the amount of condensed tannins is unlikely. This is because tannins are only seriously toxic when consumed in large amounts at more than 1 % of the diet. However, the
0
2
4
6
8
10
12
14
16
18
20
BC CAM LE FE SA CA RS
Con
tent
of s
inap
ine
[mg/
g] (c
alcu
late
d as
sin
apin
eth
iocy
anat
e)
0
5
10
15
20
25
30
35
40
BC
CAM
CO
R
EU
LE
FE
MAD
VE
SA
CA LI
RS
HS Con
cent
ratio
n of
inos
itol p
enta
- and
-hex
apho
spha
te
(phy
tic a
cid)
[mg/
g]
Inositol pentaphosphate
Inositol hexaphosphate

66
high amount in Lesquerella fendleri seed could result in the residue being rejected. A annin level of about 20 mg/g DM leads to rejection by grazing animals (Kumar et al, 1984).
Figure 4. Concentration of condensed tannins in different oilseeds.
Conclusions All of the seeds contained one or more ANFs, some in higher amounts than in Brassica napus. Nevertheless, based on their composition and the amounts of ANFs ith should be possible to use most of these seeds in animal nutrition. Only seed of Lepidium sativum, Sinapis alba (high glucosinolates and sinapine) and Lesquerella fendleri (high condensed tannins) seem to be unsuitable. However, some of these seeds may contain other ANFs, which could impede the use of their residues in animal rations. References Bell, J.M. 1989. Nutritional characteristics and protein uses of oilseed meals. In Oil Crops
of the World. G. Röbbelen, R.K. Downey & A. Achri (Eds). McGraw-Hill Publishing Company, New York. pp. 192-207.
Bjerg, B., Olsen, O., Rassmussen, K.W. & Sorensen, H. 1984. New principles of ion-exchange techniques suitable to sample preparation and group separation of natural products prior to liquid chromatography. Journal of Liquid Chromatography 7, 691-707.
Butler, L.G., Price, M.L. & Brotherton, J.E. 1982. Vanillin assay for proanthocyanidins (condensed tannins): Modification of the solvent for estimation of the degree of polymerization. Journal of Agricultural and Food Chemistry 30, 1087-1089.
EC. 1990. Determination of the oilseed glucosinolate content by HPLC. Journal of the European Communities L170, 03.07.27-34.
Griffiths, D.W., Birch, A.N.E. & Hillmann, J.R. 1998. Antinutritional compounds in the Brassicaceae: Analysis, biosynthesis, chemistry and dietary effects. Journal of Horticultural Science & Biotechnology 73, 1-18.
Kumar, R. & Singh, M. 1984. Tannins: Their adverse role in ruminant nutrition. Journal of Agricultural and Food Chemistry 32, 447-453.
0
5
10
15
20
25
BC
CAM
CO
R
EU
LE
FE
MAD
VE
SA
CA LI
RS
HS
Con
tent
of c
onde
nsed
tann
ins
[mg/
g]
(cal
cula
ted
as c
atec
hin)

67
Matthäus, B., Lösing, R. & Fiebig, H.-J. 1995. Determination of inositol phosphates IP3-IP6 in rapeseed and rapeseed meal by an HPLC method. 1. Method. Fat Science Technology 97, 289-291.
Pearson, A.W., Butler, E.J. & Fenwick, G.R. 1980. Rapeseed meal and egg taint: The role of sinapine. Journal of the Science of Food and Agriculture 31, 898-904.


69
Flatulence-producing galactosyl cyclitols D-chiro-inositol fraction in maturing yellow lupin seed A.I. Piotrowicz-Cieślak Department of Plant Physiology and Biotechnology, Faculty of Biology, University of Warmia and Mazuria in Olsztyn, 10-718 Olsztyn-Kortowo, Poland Summary
Embryos from yellow lupin (Lupinus luteus) seed (cv. Juno), were placed at 20 and 25 d after flowering on MS agar medium supplemented which D-chiro-inositol, galactinol and galactinol + D-chiro-inositol. Trigalacto-chiro-inositol was synthesized most intensely. The levels of galacto-chiro-inositol and digalacto-chiro-inositol 3, 5 and 10 d after placing the embryos on the medium were increased. Key words: Lupinus luteus, yellow lupin seed, galactosyl cyclitols, D-chiro-inositol Introduction Galactosyl cyclitols accumulate in developing or mature legume seeds, especially in lupin seed (Piotrowicz-Cieślak et al., 2003). Galactosyl cyclitols probably play an important role in the acquisition of plant desiccation tolerance and tolerance to biotic and abiotic stresses.
The galactosyl cyclitols in yellow lupin seed were represented by derivatives of the following compounds: myo-inositol, D-pinitol A, D-pinitol B and D-chiro-inositol (Piotrowicz-Cieślak et al., 2003). The α-(1→6) bonds of the galactosyl cyclitols are not hydrolysed by intestinal mucosal enzymes, and the α-galactosides pass into the colon where they are digested by intestinal bacteria. This results in the production of flatulence. Because of this the galactosyl cyclitols are considered to be antinutritional factors (ANFs), which induce flatulence.
The total amount of galactosyl cyclitols in yellow lupin seed ranged from 11.8 to 23.8 mg/g in the cotyledon and axis, respectively (Piotrowicz-Cieślak et al., 1999). The D-chiro-inositol fraction contained galacto-chiro-inositol (fagopyritol B1, 0-α-D-galactopyranosyl-(1→2)- D-chiro-inositol), digalacto-chiro-inositol (fagopyritol B2, 0-α-D-galactopyranosyl-(1→6)-0-α-D-galactopyranosyl-(1→2)-D-chiro-inositol) and trigalactopinitol B (0-α-D-galactopyranosyl-(1→6)-0-α-D-galactopyranosyl-(1→6)-0-α-D-galactopyranosyl-(1→2)-D-chiro-inositol) (Obendorf, 1997).
The object of this work was to determine the accumulation patterns of cyclitols in the embryos of maturing yellow lupin seed.
Materials and methods
Embryos of yellow lupin (cv. Juno), were taken 20 and 25 d after flowering and placed on a MS agar medium supplemented which D-chiro-inositol, galactinol and galactinol + D-chiro-inositol. The level of galactosyl cyclitols of the D-chiro-inosytol series was determined at the time of inoculation (0 d) and after 1, 3, 5 and 10 d in culture. The embryos were analysed for their content of D-chiro-inositol derivatives: galacto-chiro-inositol, digalacto-chiro-inositol

70
and trigalacto-chiro-inositol by gas chromatography. The level of D-chiro-inositol derivatives was determined separately in the embryonic axes and cotyledons.
Results and discussion
Studies on carbon transformation in plant micro-reproduction indicate that sucrose is the main carbon source (Copeland, 1990). Therefore, it is the most common component in microbiological media. Sucrose is an extra-cellular carbon source, however, its production by photosynthesis in cultured cells cannot be ruled out. Frequent studies have tested numerous sugars and have shown that sucrose was the most essential sugar and it was added to the medium at 2 or 3 % (Hdider & Desjardins, 1994.). This experiment used a 3 % sucrose concentration.
Sugar intake by the developing embryos was gradual. A gradual embryo sugar was accompanied by a gradual sugar decrease in the medium. This gradual sugar assimilation was observed many times (Piotrowicz-Cieślak et al., 2000). The D-chiro-inositol derivative accumulation profile in 20 and 25 d old embryos was similar (Fig. 1, 2).
Figure 1. Composition of D-chiro-inositol derivatives: galacto-chiro-inositol (•), digalacto-
chiro-inositol (□), trigalacto-chiro-inositol (o) 20 days after flowering.
Trigalacto-chiro-inositol was synthesized most intensely. Three to 5 d after placing the embryos on the medium the galacto-chiro-inositol and digalacto-chiro-inositol content increased. However, at 10 d the level of galacto-chiro-inositol and digalacto-chiro-inositol fell. The most intense synthesis of trigalacto-chiro-inositol was in cotyledons cultured on the medium, which was supplemented with both galactinol and D-chiro-inositol. It was at 4.5 and 5.6 mg/g of DM, respectively in 20 and 25 d old embryos. Based on these results synthesis intensity was determined by the availability and amount of D-chiro-inositol and galactinol. The amount of trigalacto-chiro-inositol produced by the seeds was four times higher than in

71
yellow lupin seed. The increase in trigalacto-chiro-inositol content most likely results from a decrease in the amount of galacto-chiro-inositol and digalacto-chiro-inositol. Galactinol is commonly recognised as a donor of galactosyl groups. It possibly carries galactosyl residues onto D-chiro-inositol thus extending the galactosyl chain. Galactosyl cyclitol synthesis is not limited by the amount of substrate therefore it proceeds very rapidly. In naturally maturing seeds, trigalacto-chiro-inositol synthesis is quite active and the highest intensities are found 40 to 45 d after flowering. It can be assumed that the availability of galactinol and D-chiro-inositol is the principal factor affecting the level of D-chiro-inositol galactosyl cyclitol synthesis.
Figure 2. The composition of D-chiro-inositol derivatives: galacto-chiro-inositol (•),
digalacto-chiro-inositol (□), trigalacto-chiro-inositol (o) 25 days after flowering.
Acknowledgements This work was funded by grants from the Polish Committee for Scientific Research; project No. 3 PO6A 005 23.
References Copeland, L. 1990. Enzymes of sucrose metabolism. In Methods in Plant Biochemistry.
Vol. 3. Enzymes of The Primary Metabolism. P.J. Lea (Ed.). Academic Press, London. pp. 73-85.
Hdider, C. & Desjardins, Y. 1994. Effects of sucrose on photosynthesis and phosphoenolopuryvate carboxylase activity of in vitro cultured strawberry plantlets. Plant Cell, Tissue and Organ Culture 36, 27-33.

72
Obendorf, R.L. 1997. Oligosaccharides and galactosyl cyclitols in seed desiccation tolerance (Review Update). Seed Science Research 7, 63-74.
Piotrowicz-Cieślak, A.I., Górecki, R.J. & Adomas, B. 1999. The content and composition of soluble carbohydrates in lupin seeds of different species and cultivars. Plant Breeding and Seed Science 43 (2), 25-34.
Piotrowicz-Cieślak, A.I., Ryszard, J., Górecki, R.J. & Rejowski, A. (2000). Soluble oligosaccharides and galactosyl cyclitols in maturing lupin seeds. Horticulture and Vegetable Growing 19 (3)-2, 273-280.
Piotrowicz-Cieślak, A.I., Gracia-Lopez, P.M. & Gulewicz, K. 2003. Cyclitols, galactosyl cyclitols and raffinose family oligosaccharides in Mexican wild lupin seeds. Acta Societatis Botanicorum Poloniae 72, 109-114.

73
Flatulence-producing oligosaccharides and galactosyl cyclitols in maturing Andean lupin (Lupinus mutabilis) seed A.I. Piotrowicz-Cieślak, D.J. Michalczyk & R.J. Górecki Department of Plant Physiology and Biotechnology, Faculty of Biology, University of Warmia and Masuria in Olsztyn, 10-718 Olsztyn-Kortowo, Poland Summary The flatulence-producing oligosaccharides are important components of Lupinus mutabilis seed. Maturing lupin seeds were harvested at 10 d intervals from 10 to 50 d after flowering. Mature dry seed contained 11.5 % oligosaccharides, 2.0 % sucrose, 0.6 % galactosyl cyclitols, 0.4 % cyclitols and 0.01 % fructose. Key words: Lupinus mutabilis, Andean lupin, maturing, oligosaccharides, raffinose family,
galactosyl cyclitols Introduction The Andean lupin (Lupinus mutabilis) is the only lupin crop from the New Word. A high seed protein (the highest among lupins at > 44 % DM) and a high fat (≈25 % DM) content are characteristics of L. mutabilis seed (Hatzold et al., 1983). Lupin seed contain a range of antinutritional factors (ANFs). In the major domesticated lupin species these are alkaloids, saponins, tannins, lectins, trypsin inhibitors, phytates and α-galactosides (raffinose family oligosaccharides and galactosyl cyclitols) (Góórecki et al., 1997; Gladstones, 1998). These ANFs can make lupin seed unsuitable for the feeding of monogastric animals (Price et al., 1988). Oligosaccharides of the raffinose family and galactosyl cyclitols have been identified as a major cause of flatulence in animals and in humans (Dey, 1997). The objective of this work was to examine raffinose family oligosaccharides and galactosyl cyclitols in maturing L. mutabilis seed. Materials and methods Lupinus mutabilis plants were grown in a greenhouse at 25 oC day and 17 oC night temperature. Seeds were harvested at 10 d intervals from 10 to 50 d after flowering. Their water content, germination characteristics and soluble carbohydrates were measured. Soluble sugars were analysed according to Horbowicz & Obendorf (1994) with slight modification. Results and discussion Fourteen soluble carbohydrates were detected in L. mutabilis seed: fructose, sucrose, cyclitols, galactosyl cyclitols and raffinose family oligosaccharides (Fig. 1, Table 1). Fructose, sucrose and cyclitols (D-pinitol, D-chiro-inositol, and myo-inositol) were dominant at early stages of seed growth. Mature Andean lupin seed accumulated 11.5 % of its DM as stachyose, raffinose and verbascose. The soluble sugars profile of the maturing seed showed that the concentration of raffinose, stachyose and verbascose decreased during seed maturation (Table 1).

74
Sucrose was present throughout seed development. Lupinus mutabilis accumulated galactosyl cyclitols in the seed, among the galactosyl cyclitols, galactinol was dominant. Galactosyl cyclitols have been found in the seed of various plant species (Horbowicz & Obendorf 1994). They are formed after a cyclitol binds with one or two galactose molecules with an α-(1→6) and α-(1→2) or α-(1→3) galactosyl bond. These compounds may form higher order homological sequences. Among the galactosyl cyclitols, the greatest amount of data has been collected on the galactinol (0-α-D-galactopyranosyl-(1→3)-D-myo-inositol) that is the carrier of the D-myo-inositol galactosyl residues to sucrose and higher homologous sequences such as the raffinose family sugars (Day, 1997). Galactosyl cyclitols, like the raffinose family oligosaccharides, belong to the basic group of storage compounds present in lupin seed (Piotrowicz-Cieślak et al., 1999). The L. mutabilis seeds contained 12 unidentified carbohydrates. Table 1. Carbohydrate compositiona of seed of Lupinus mutabilis (mg/g seed DM).
Days after flowering Soluble carbohydrates 10 20 30 40 50 Fructose 10.5 ± 2.3 7.2 ± 1.2 9.5 ± 1.1 1.2 ± 0.4 0.2 ± 0.1 Sucrose 85.0 ± 7.2 37.7 ± 4.2 24.5 ± 2.3 24.2 ± 4.2 20.4 ± 2.2
Cyclitols D-pinitol 2.2 ± 0.2 3.7 ± 0.7 4.3 ± 1.5 2.1 ± 0.4 1.3 ± 0.2 D-chiro-inosytol 1.1 ± 0.1 1.1 ± 0.3 0.7 ± 0.6 0.2 ± 0.2 0.20 ± 0.04myo-inosytol 5.1 ±0.2 8.2 ± 0.1 5.7 ± 0.3 3.2 ± 0.8 2.7 ± 0.1
Galactosyl cyclitols Galactopinitol A 0 0 0 0.1 ± 0.05 0.2 ±0.04 Galactopinitol B 0 0 0 0 Trace Galactinol 0.5 ± 0.02 8.7 ± 1.2 7.8 ± 0.1 4.5 ± 0.1 2.0 ± 0.2 Ciceritol 0 0 0.3 ± 0.02 0.7 ± 0.1 1.2 ± 0.1 Digalacto-myo-inosytol 0 0 0 0.2 ± 0.01 1.0 ± 0.2 Trigalactopinitol A 0 0 0 0.8 ± 0.2 1.6 ± 0.2
Raffinose family oligosaccharides (RFO) Raffinose 0 0 5.6 ± 0.7 18.3 ± 3.2 24.3 ± 3.3 Stachyose 0 1.2 ± 0.1 26.1 ± 2.3 46.2 ± 6.3 60.6 ± 5.2 Verbascose 0 0 8.5 ± 2.2 21.3 ± 2.1 30.4 ± 1.2 Sum of RFO 0 1.2 40.3 85.8 115.2 Sucrose/RFO 0 30.9 0.6 0.3 0.2 Total galactosyl cyclitols 0.5 8.7 8.2 6.4 6.0 Total sol. carbohydrate 104.5 67.8 93.4 123.2 125.2 aData are the means of 4 replicates/treatment ± standard deviation.
The sucrose to RFO ratio, specifically, reflects seed storage capacity (Horbowicz & Obendorf, 1994). Seeds with a ratio value below 1 can be stored for more than 10 years. The greatest storage capacity in this seed was 50 days after flowering (0.09). Acknowledgements This work was funded by grants from the Polish Committee for Scientific Research. (Project No. 3 PO6A 005 23)

75
Figure 1. The chromatogram profile of the soluble carbohydrates of Lupinus mutabilis seed.
(1-D-pinitol, 2-fructose, 3-D-chiro-inositol, 4-myo-inositol, 5, 6, 7, 8, 9, 10-unknown, 11-internal standard, 13, 14 -unknown, 15 sucrose, 16, galactopinitol A, 17- galactopinitol B, 18-unknown, 19- galactinol, 20-unknown, 21-raffinose, 22-ciceritol, 23- digalacto-myo-inositol, 24-stachyose 25- trigalactopinitol A, 26- verbascose).
References Dey, P.M. 1997. Carbohydrate metabolism. In Plant Physiology. P.M. Dey & J.B.
Harbourne. (Eds). Academic Press, London. pp. 150-204 Gladstones, J.S. 1998. Distribution, origin, taxonomy, history and importance. In Lupins as
Crop Plants: Biology, Production and Utilization. J.S. Gladstones, C. Atkins & J. Hamblin (Eds). CAB International, Wallingford. pp. 335-376.
Górecki, R.J., Piotrowicz-Cieślak, A.I., Lahuta L.B. & Obendorf R.L. 1997. Soluble carbohydrates in desiccation tolerance of yellow lupin seeds during maturation and germination. Seed Science Research 7, 107-115.
Hatzold, T., Elmadfa, I., Gross, R., Wink, M., Hartmann, T. & Witte, L. 1983. Qunolizidine alkaloids in seed of Lupinus mutabilis. Journal of Agricultural and Food Chemistry 31, 934-938.
Horbowicz, M. & Obendorf, R.L. 1994. Seed desiccation tolerance and storability: dependence on flatulence-producing oligosaccharides and cyclitols – review and survey. Seed Science Research 4, 385-405.
Piotrowicz-Cieślak, A.I., Górecki, R.J. & Adomas, B. 1999. The content and composition of soluble carbohydrates in lupin seeds of different species and cultivars. Plant Breeding and Seed Science 43 (2), 25-34.
Price, K.R., Lewis, J., Wyatt, G.M. & Fenwick, G.R. 1988. Flatulence - causes, relation to diet and remedies. Nahrung 32, 609-623.


77
A modified trypsin inhibitor affinity chromatography assay for industrial application I. Wolters1, J. de Groot2, H. Boer1 and A.F.B. van der Poel1
1Animal Nutrition Group, Wageningen University & Research Centre, Marijkeweg 40, 6709 PG Wageningen, The Netherlands 2Wageningen Centre for Food Sciences, Wageningen University & Research Centre, Food Chemistry Group, Bomenweg 2, 6703 HD Wageningen, The Netherlands Introduction For the trypsin inhibitor analysis (TIs) in e.g. soybeans (Glycine max), peas (Pisum sativum) and white kidney beans (Phaseolus vulgaris), Roozen & de Groot (1987, 1991) developed a method based on trypsin-sepharose affinity chromatography. The advantages of this method are increased assay sensitivity by concentrating the TIs on the column, and an increased specificity compared with earlier assay methods (Kakade et al., 1974). However, industrial laboratories still hesitate to use affinity chromatography since special and expensive equipment is needed for the extraction procedure used prior to the affinity chromatography.
This study describes a new extraction and filtration method for the assay. The procedure has been modified by reducing the time for the procedure and by using simple laboratory equipment. Further, the extraction procedure is more controlled and reproducible. The method is more applicable for use in industrial laboratories. Materials and methods Plant material Full fat soybeans, grown in Argentina, were obtained from a commercial supplier. They were steam treated at 118 °C for 2.5, 5, 10 and 20 min, respectively using a pressurized toaster (van der Poel et al., 1990). The processed soybeans were dried (16 h at 35 °C). Processed and unprocessed beans were successively ground in two steps, through a 6 mm and a 1 mm sieve.
Trypsin and Kunitz trypsin-inhibitor were obtained from Merck (No. 1.08350) and Fluka (No. 93619) respectively. The CNBr-sepharose 4B was purchased from Amersham Biosciences (No. 17-0430-01). All other reagents were analytical grade.
Defatting Samples were defatted at room temperature by a modified Soxhlet extraction (Rettberg 150 ml). The cooling coil was 30 cm. Vaporised petroleum-ether (40 - 60 °C) entered the cooling coil in the middle. The temperature of the condensed petroleum-ether was therefore no higher then 25 ° C ± 1 °C, measured with a 1 mm thermocouple on the bottom of the filled thimble. The extraction thimble with 25 g of sample was placed on a 2 cm high stainless steel sieve to prevent mixing of the petroleum-ether fractions of successive extraction cycles. After 1 h of extraction the thimble filled with the defatted sample was placed in a tight-fitting glass tube and the sample was dried with a gentle air stream through the thimble.

78
Extraction The defatted samples were extracted with 0.2 M di-sodiumhydrogen phosphate-0.5 M sodium chloride solution (pH 8.2) (ratio sample:extraction liquid = 1:15). The suspension was stirred at room temperature, under vacuum, which was maintained with a vacuum pump, for 60 min. During extraction the pH of the suspension was kept at pH 8.2 by adjusting the pH three times in the first 30 min with 3.5 M sodium hydroxide.
The suspension was subsequently centrifuged at a minimum of 1,840 g. The supernatant was saved and the pellet was re-extracted with 0.2 M di-sodium hydrogen phosphate-0.5 M sodium chloride solution (pH 8.2). The re-extraction procedure was as above. Ultra filtration The combined supernatants were ultra filtered, through a 0.2-µm membrane (S&S NC 20, type 401314, 50 mm) at a pressure of 1.8 bar in a stirred ultra filtration cell (70 ml). Affinity chromatography The coupling of trypsin to CNBr-activated Sepharose 4B was performed according to the supplier’s instructions (Amersham Biosciences). To obtain optimal trypsin coupling to the sepharose particles, the sepharose particles should be kept suspended in the trypsin solution, while mixing the suspension “end over end“ during the coupling procedure (6 – 8 rpm). Coupling was performed at ± 4°C.
A maximum binding-capacity of 3.5 – 4.0 mg of Kunitz trypsin-inhibitor protein/trypsin-sepharose column (1.8 × 1.5 cm) was obtained.
After equilibration of the trypsin-sepharose column with 0.2 M di-sodium hydrogen phosphate-0.5 M sodium chloride solution (pH 8.2) at a flow rate of 1.8 ml/min, x ml of sample was applied to the column. The volume applied, depends on the amount of TIs expected in the sample and the maximum binding capacity of the column. The column should not be overloaded (Roozen & de Groot, 1991). The column should be washed with 15 ml of the phosphate-sodium chloride solution (pH 8.2) and subsequently with 15 ml of 0.05 M sodium acetate-0.5 M NaCl (pH 5.2). Elute the bound trypsin inhibitor with 15 ml of 0.1 M Glycine HCl-0.5 M NaCl (pH 3.0). Protein and dry matter determination Protein concentration of the collected eluate was determined as per Lowry et al. (1951). Calculation of the amount of trypsin inhibitor (TI)/g of dry sample was as follows.
mg TI/g dry sample = gVVa
×××
2
115
where a = the concentration of TIs in the eluate (mg/ml); 15 = volume of the eluate (ml); V1 = volume phosphate/chloride solution (pH 8.2) used to extract the sample (ml); V2 = volume of the extract applied to the affinity column (ml); g = amount of defatted dry sample used for the extraction (g).
The dry matter content of the soybean samples was determined by oven drying at 103 °C for 4 h.

79
Results and discussion Data pertaining to the analysed criteria of the soybeans steamed for different times are shown in Table 1. The moisture content, remaining fat content of the sample after defatting, TIs (mg/g crude protein). The volume of extract used in the trypsin-sepharose column varied with cooking time. It was 1 – 2 ml for raw beans, 15 – 25 ml after 2.5 min 40 – 60 ml after 5 min, 75 ml after 10 min and 445 ml after 20 min cooking. Table 1. Trypsin inhibitor (TI) content (mg/g dry matter (DM)) in defatted soybean samples
determined by affinity chromatography
Soybean sample TIs (mg/g DM)* TIs (mg/g CP)* Moisture (g/kg) Remaining oil (g/kg)
Untreated 25.6 ± 1.1 (n=2) 48.0 ± 2.1 (n=2) 125 26
2.5 min. 3.1 ± 0.2 (n=3) 5.93 ± 0.30 (n=3) 85 15
5 min. 0.66 ± 0.01 (n=2) 1.33 ± 0.01 (n=2) 61 18 10 min 0.26 0.48 127 19 20 min 0.15 0.27 138 18
*Mean ± SD
From this validation experiment it can be seen that the levels of trypsin inhibitor are in range with the time of steam treatment. Moreover, low levels of trypsin inhibitor can be estimated as low levels of TIs can be concentrated by the elution of a large volume of extract through the column (Table 1).
The extraction of soy samples with this new extraction and filtration method gave a very clear extract, necessary for affinity chromatography, without the need to use an expensive centrifuge (Roozen & de Groot, 1991). A further advantage is that the defatting, extraction and filtration can be done in 6 - 7 h. Together with the affinity chromatography it will give a result in one-day for untreated samples or samples treated for a short time.
In Table 2, levels of trypsin inhibitors (TIs) are compared with values of trypsin inhibitor activity (TIA) in heat processed full fat soybeans from Argentina and China (Qin et al., 1998), determined by the method of Kakade et al. (1974). For comparison the TIA values of Qin et al. (1988) have been converted to give values in mg of Kunitz inhibitor/soybean sample. The unprocessed soybean sample gave similar TI values by the different methods. Further validation experiments with the TI-assay will be carried out and the inactivation pattern for TIs can be judged to be similar for soybeans of the different origin. Table 2. Comparison of trypsin inhibitor (TI) contents and trypsin inhibitor activity (TIA) in
samples of full fat soybeans after toasting at 118 °C. TIs
(mg/g soybean) TIA
(mg/g soybean) TIA
(mg/g soybean) Origin Argentina China
Unprocessed 17.91 17.02 22.32 Toasted at 118 oC
2.5 min 2.2 3.9 9.4 5 min 0.5 2.1 4.6 10 min 0.2 1.3 2.3 20 min 0.1 0.4 0.8
1Wolters et al. ((This manuscript). 2Qin et al. (1998).

80
References Kakade, M.L., Rackis, J.J., McGhee, J.E. & Puski, G. 1974. Determination of trypsin
inhibitor activity of soy products: a collaborative analysis of an improved procedure. Cereal Chemistry 51, 376-382.
Lowry, O.H., Rosebrough, N.J., Farr, A.L. & Randell, R.J. 1951. Protein measurement with the Folin phenol reagent. Journal of Biological Chemistry 193, 265–275.
Qin, G.X., Verstegen, M.W.A. & Poel, A.F.B. van der 1998. Effect of temperature and time during steam treatment on the protein quality of full-fat soybeans from different origin. Journal of the Science of Food and Agriculture 77, 393-398.
Roozen, J.P. & Groot, J. de 1987. Analysis of low levels of trypsin inhibitor activity in food. Lebensmittel-Wissenschaft und Technologie 20, 305–308.
Roozen, J.P., Groot, J. de, 1991. Analysis of trypsin inhibitors and lectins in white kidney beans (Phaseolus vulgaris, var. Processor) in a combined method. Journal of the American Organisation of Agricultural Chemists 74, 940-943.
Van der Poel, A.F.B., Blonk, J., Zuilichem, D.J. van & Oort, M. van, 1990. Thermal inactivation of lectins and trypsin inhibitor activity during steam processing of dry beans (Phaseolus vulgaris L.) and effects on protein quality. Journal of the Science of Food and Agriculture 53, 215-228.

81
The correlation between taste and structure of lupin alkaloids W. Wysocka1 & J. Jasiczak2
1Faculty of Chemistry, Adam Mickiewicz University, Grunwaldzka 6, 60 780 Poznań, Poland 2Faculty of Commodity Science, Poznań University of Economics, Al. Niepodleglosci 10, 60 967 Poznań, Poland Summary A topological molecular matrix of the bitter taste receptor has been constructed by a superposition of model bitter compounds. The matrix allows the analysis of correlations between the taste of individual compounds and their spatial structure. The main important factor in the chemoreception of taste is the conformation of the alkaloid molecules. Results of the computer analysis were supported by sensory tests. Among the alkaloids investigated, sparteine was the most bitter. Keywords: theory of taste, chemoreception of taste, alkaloids, conformation of bis-
quinolizidine alkaloids Introduction Lupins (Lupinus spp) and other grain legumes have a high nutritive value due to their high seed protein content. However, the main antinutritional factors in lupins are the quinolizidine alkaloids, which are bitter tasting toxic compounds. The range of bitterness of particular alkaloids differs depending on their chemical structure. This problem has posed a challenge to workers who have tried to study the correlation between the structure and the taste of lupin alkaloids. Recently, significant effort has been devoted to obtain information from the interaction of flavour molecules with taste receptors. The most extensive literature on this topic is available for sweet compounds. Thus, most chemoreception theories have been formulated for sweetness (Zalewski & Jasiczak, 1994).
In initial attempts to describe the correlation between the flavour of bis-quinolizidine alkaloids and their spatial structure, we (Jasiczak & Wysocka, 1996; Jasiczak et al., 1999) provided a preliminary topological model of a bitter taste receptor. The models adopted by the authors for mapping the bitter taste receptor were based on such agonists (bitter-flavoured compounds) as methylurea, tetraiodosaccharine and quinine. Modelling of the molecular matrix (mould) of bitter taste receptors On the basis of literature descriptions of the electric charge in electrophilic and nucleophilic centres for sweet-flavoured compounds (glucophores) (Jasiczak et al., 2000) we modified our earlier topological matrix of the bitter taste receptor. The matrix shows the receptor’s critical size (which is its minimal size) in the Y,Z plane. The receptor’s depth (shown in the X,Y plane) is 3 Å, which meets the criterion of Shellenberger’s spatial barrier (Zalewski & Jasiczak, 1994). The receptor is fully open towards the X axis forming a half-open cavity.
The topological matrix that takes into account the above criteria can be formulated by superimposing model bitter compounds. The model compounds selected were quinine, tetraiodosaccharine, khellin and methylurea. The electro-static potential of these compounds facilitated their superposition. The spatial orientation of the compounds was determined by the

82
alignment of the proper hydrophilic (electrophilic and nucleophilic) centres with the complementary centres in the receptors.
The total area of van der Waals forces interactions (VdW net) was determined for a super-molecule created in the above manner. The area may be taken as the critical area of the receptor. In the diagram below the critical area is marked by a solid line (Fig. 1). The line delimits the scope of impact of model agonists in the Van der Waals (VdW) radius inside the walls of the hypothetical receptor. It is proposed that the resulting topographic form be called the molecular matrix (mould) of bitter taste. Figure 1 shows the matrix in the critical plane Y,Z and in the side view (plane X,Y) that describes the Shellenberger barrier. The molecular matrix has been additionally divided into sectors (A to D) to facilitate description of the interrelations between the agonists and the matrix.
Figure 1. The molecular matrix of bitter taste in the Y,Z plane and its side view. On the basis of the information in the literature and our research, we propose the following regions in the molecular matrix:
- A region of strongly active nucleo-electrophilic centres (on the Y axis), which represents the hydrophilic part of the matrix. In this part, the polar groups of the agonist interact with the matrix, which anchors the antagonist in the receptor and the steroidal alignment of the hydrophobic part of the compound.
- A, B, C sectors where hydrophobic forces make the antagonists interact with the walls of the molecular matrix describing the bitter taste receptor and makes the nucleophilic centre of the antagonist, i.e. the electrophilic centre of the receptor (of the ns/er type), be engaged in a complementary interaction with the electrophilic centre of the antagonist, i.e. the nucleophilic centre of the receptor (of the es/nr type),
- The interface between sectors B and C, which includes a well-defined region of hydrophobic, impact that comprises, among others, a sub-region of the interaction of π electrons (the π-Stacking Region or the Aryl Region). In this region, the aromatic rings of the agonists, substituted with electro-acceptor groups, affect those areas in the receptor that abound with π electrons,

83
- Matrix walls in sector A which, stimulated by the interaction with the agonists, significantly affect the intensity of bitter sensations (i.e. the profile of intense bitter taste),
- Open space in sector D of the molecular matrix, which, devoid of the critical dimension, allows very large molecule antagonists to easily penetrate the receptor.
Computer fitting of lupin alkaloid in the molecular matrix of the bitter taste receptors The molecular matrix, as designed, allows satisfactory computer analyses of various classes of compounds by fitting them into the matrix taking into regard their expected taste qualities. The matrix also allows the analysis of correlations between the taste of individual compounds and their spatial structure.
Lupin alkaloids having a sparteine skeleton can occur in two conformations: the full chair conformation for all rings or with the boat ring C. Figure 2, shows a projection of two sparteine conformers in the molecular matrix in the plane Y,Z and side views (in the aspect of Shallenberger’s spatial barrier).
X
Z
Y
Figure 2. Projections of sparteine conformers into the molecular matrix of bitter taste.
As shown in Figure 2, the boat-shaped conformer of bis-quinolizidine alkaloids with its VdW net fits well into the molecular matrix of the bitter taste receptor (Fig. 2, left side), especially in sectors A and B. A strong stimulation of sector A results in sensations of bitterness that are likely to be highly intense. On the other hand, the VdW net for quinine shows that the model compound is unlikely to engage in any interaction with sector A walls but strongly affects sector B and, particularly sector C, as determined by the use of the molecule’s topology. The observations suggest that the boat-shaped sparteine conformer should stimulate a bitter taste at a level of intensity that is very close to that observed in the case of quinine. The results of sensory tests support this conclusion.
Regardless of which active centre (N16 or N1) is captured in the matrix’s hydrophilic centre, the full-chair conformer of bis-quinolizidine alkaloid will not fit into its hydrophobic

84
part. This means it will not stimulate the receptor (Fig. 2, right side). Such conformers of bis-quinolizidine alkaloids will remain tasteless. This conclusion has also been supported by sensory tests.
This hypothesis suggests that bitter taste is stimulated exclusively by bis-quinolizidine alkaloids occurring in boat-shaped conformations, which is confirmed by the results of computer analyses and is also supported by the results of sensory tests (Jasiczak et al., 1999).
References
Jasiczak, J., Jonska-Muteba E. & Zalewski R.I. 2000. Structure-activity relationship of
sweet molecules: phenylurea derivatives. Polish Journal of Chemistry 74, 1259-1273. Jasiczak, J. & Wysocka, W. 1996. Korelacja pomiedzy struktura a smakiem gorzkim
alkaloidow lubinowych. In Lubin: Kierunki Badan i Perspektywy Uzytkowe. I. Frencel & K. Gulewicz (Eds). Polish Lupin Association, Poznań. pp. 138-152.
Jasiczak J., Wysocka W. & Skolik A. 1999. Matryca molekularna receptorow smaku gorzkiego gorzkiego w badaniach struktury alkaloidow bis-chinolizydynowych. In Na Pograniczu Chemii i Biologii, Vol. III. H. Koroniak & J. Barciszewski (Eds). Wyd. Naukowe UAM, Poznań. pp. 503-529.
Zalewski R.I. & Jasiczak J. 1994. Structure-property relationship in sweeteners. Journal of Chemical Information and Computer Sciences 34, 179-183.

Session II Mechanism of action of antinutritional factors in legume seeds and oilseed


87
The mode of action of ANFs on the gastrointestinal tract and its microflora A. Pusztai1, S. Bardocz1 & M.A. Martín-Cabrejas2
1*Independent Consultants - Aberdeen, AB10 6SF, Scotland, UK 2Universidad Autónoma de Madrid, Facultad de Ciencias, Sección de Química Agrícola, CVII, 28049 Madrid, Spain Summary New directions in antinutritional factor (ANF) research in the last decade have led to major developments in our understanding of their role in nutrition. In these, instead of carrying out simple mechanistic studies and measurements of the negative effects of the various ANFs on the nutritional performance of laboratory and farm animals as done in the past, their mode of action in the gastrointestinal tract and particularly their effects on metabolism and the gut microflora has been investigated. The scientific interest in ANFs is now also turning to studies of their possible useful and beneficial applications as gut, metabolic and hormonal regulators and as probiotic/prebiotic agents, even though there is still much to learn about the role of ANFs in these applications. With the development of genetically modified (GM) plants by transferring the genes of some of the natural insecticidal protein ANFs to major crop plants, a major new avenue in plant protection applications has opened up. Keywords: protein, low molecular weight ANFs, harmful effects, beneficial applications Introduction Plants contain constituents, which are nutritionally deleterious to humans and animals, which ingest them. These so-called antinutritional factors, ANFs, make no positive contribution to the nutritional value of the diet but rather depress it. Legumes are particularly rich in these phytochemicals. Some of these ANFs are proteins, such as the lectins, enzyme inhibitors, or other proteins with antinutritional effects. However, the most numerous ANFs are relatively low molecular weight phytochemicals, such as the polyphenols, tannins, phytates, saponins and some oligosaccharides. It is therefore not surprising that for such chemically different classes of compounds their modes of action are very diverse. These effects can vary between the hardly noticeable (subclinical) to the overtly toxic or indeed poisonous. Although early studies on ANFs were rather simplistic and mechanistic in nature they have still given useful qualitative, and sometimes quantitative, indications about these harmful effects. Moreover, based on these several methods have been invented and tried in nutritional practice for the reduction or possible elimination of the negative effects of ANFs.
High temperature heating denatures almost all proteins. Therefore the most convenient and frequently used methods for the elimination of the harmful effects of protein ANFs are based on various forms of heat treatment. As proteins are direct gene products, it has more recently also become possible to use genetic antisense techniques for the elimination of protein ANFs. Unfortunately, neither of these two methods can be directly applied to remove the antinutritional effects of the low molecular weight ANFs.
Because chemically ANFs are a diverse class of compounds, and their mode of action and the methods used for their removal are different and because their beneficial and harmful *Formerly: The Rowett Research Institute, Bucksburn, Aberdeen AB21 9SB, Scotland, UK

88
effects are also different it is practical to describe the progress in ANF studies separately for protein ANFs and low molecular weight ANFs, respectively. In this short review the objective was not to comprehensively cover developments in ANF research but rather to highlight those results, which appeared to be the most significant to the authors. Size limitation precluded the possibility of a more comprehensive review. Advances in ANF research - Proteins Lectins Practically all lectins are resistant to gut proteolysis (Pusztai et al., 1990). Therefore they remain biologically active during their passage through the gastrointestinal tract. Accordingly, their effects on the gut, body metabolism, immune and endocrine systems and the bacterial flora are the direct consequence of their interaction with the highly glycosylated receptors of the surface cells of the gut. This occurs not only in mammals and birds but also in fish, such as the Atlantic salmon (Salmo salar) and the rainbow trout (Oncorhynchus mykiss) (Buttle et al., 2001). Due to differences in glycosylation along the gastrointestinal tract the effect of the same lectin can be different in different parts of the gut. Lectins can also modify the glycosylation of gut surface cells. Thus, from understanding the molecular and cellular mechanism of their interactions with the gut, it may be possible to utilise lectins as reagents for various nutritional or medical applications (see van Damme et al., 1997).
As lectins given orally induce the release of gut peptide hormones that are needed for the normal functioning of the gastrointestinal tract, a potentially important clinical application of lectins is to use them to reverse the grave condition of small bowel atrophy in patients who need to be fed parenterally (Jordinson et al., 1999). Similar, though not the same, effects were seen when peanut (Arachis hypogaea) agglutinin was intravenously infused (Jordinson et al., 2000). The kidney bean (Phaseolus vulgaris) lectin (PHA) can be used to suppress gastric acid output while stimulating the release of cholecystokinin (CCK), and consequently pancreatic enzyme secretion into the duodenum (Pusztai, 1999; Kordás et al., 2000, 2001;). Some lectins, such as PHA can also be used to modulate food consumption and gastric emptying in experimental animals (Baintner et al., 2003). Wheat germ (Triticum spp) agglutinin and PHA have also been shown to reduce the levels of heat shock proteins in gut epithelial cells (Ovelgönne et al., 2000). It may also be possible to use PHA as a dietary adjunct or a therapeutic agent to stimulate gut function and ameliorate obesity not only in animals but also in humans if a safe and effective dose-range can be established in clinical trials (Pusztai et al., 1998). Similar observations have been made with soybean (Glycine max) whey of high lectin content (Pusztai et al., 1997a). Friedman & Brandon (2001) and Lajolo & Genovese (2002) have recently published useful reviews on the effects of lectins and trypsin inhibitors.
Insecticidal effects of lectins: Lectins have been found to be one of the most potent naturally occurring insecticidal proteins. It is therefore not surprising that many transgenic plants have been developed using lectin genes. As the topic of genetically modified plants is outside the scope of this review they will not be reviewed here. However, some key papers describing the interaction between lectins and insects will be briefly mentioned. Thus, some lectins, particularly the Listera ovata (LOA) and Galanthus nivalis agglutinin (GNA) blocked larval development of the legume pod borer, Maruca vitrata (Machuka et al., 1999). In contrast, the African yam bean (Sphenostylis stenocarpa) seed lectin inhibited the development of the cowpea weevil (Callosobruchus maculatus) but it did not affect the larvae of the legume pod

89
borer (Maruca testualis) (Machuka et al., 2000). In a well-researched study it was shown that PHA was lethal to the western tarnished plant bug (Lygus hesperus) but that this was due to the strong binding of the lectin to the cells of the midgut region of the bug, which became swollen to such an extent that the gut lumen was completely blocked and finally the toxic lectin was endocytosed (Habibi et al., 2000). It was also shown that PHA is toxic to the tomato moth (Lacanobia oleracea) as apparently the lectin was extensively bound to gut glycoproteins of the moth (Fitches et al., 2001). Studies of the mechanism of lectin action showed that the binding of the garlic (Allium sativum) leaf lectin to homopteran pests was correlated with its insecticidal activity (Bandyopadhyay et al., 2001). Even aphid parasitoids were affected by lectins, such as GNA (Couty et al., 2001a; Couty et al., 2001b; Couty & Poppy, 2001; Sétamou, et al., 2002). Galanthus nivalis agglutinin was also effective on stalkborers (Busseola spp) (Sétamou et al., 2002). Wheat germ agglutinin showed insecticidal effects against some insects, such as the European corn borer (Ostrinia nubilalis) because this N-acetylglucosamine-specific lectin interferes with the formation of the peritrophic membrane in the insect larvae (Hopkins & Harper, 2001). Conclusive experimental evidence was put forward supporting the idea that the insecticidal activity of lectins is truly dependent on lectin function because in the presence of the haptenic sugar the insecticidal activity of the lectin was significantly reduced or even abolished (Triguéros et al., 2000). A review of the role of lectins and protease inhibitors in plant defence has been recently published by Murdock & Shade (2002). It has also been shown that even lectin-like proteins, such as arcelin-1 in bean seed can have significant insecticidal effects against a number of insects, such as some species of Bruchids (Paes et al., 2000). Effects on tumours: Lectins given orally can have diverse effects on different tumours. It has been shown that peanut agglutinin (PNA), stimulates the proliferation in colonic explants in vitro (Ryder et al., 1994) and eating peanuts increases rectal proliferation in individuals with mucosal expression of peanut lectin receptor (Ryder et al., 1998). In contrast, orally given PHA, or mistletoe (Viscum album) lectin, ML-1, significantly reduced, and in some cases eliminated, the development of tumours in NMR and Balb/c mice into which Krebs II ascites tumour cells had been intraperitoneally or subcutaneously injected before the lectin treatment (Pryme et al., 1999a; Pryme et al., 1999b; Pryme et al., 2002). The beneficial lectin effect was complex and due to several factors. These included a deprivation of nutrients of the growing tumour by the high nutrient requirements of the lectin-induced compulsory opposing gut growth, inhibition by ML-1 of angiogenesis in the growing tumour and stimulation of the immune system combating tumour growth. The capacity of various lectins, including Bacillus thuringiensis Cry1Ac protoxin to modulate both the systemic and mucosal immune systems is now firmly established (Haas et al., 1999; Vázquez et al., 1999; Lavelle et al., 2000; Lavelle et al., 2001). Lectins and bacteria: Lectins are known to directly interact with gut bacteria (Kellens et al., 1995; Porter et al., 1998). A good example for this is the agglutination of different strains of Salmonella typhimurium by Concanavalin A (Con A) (Naughton et al., 2000). Furthermore, the indirect effects of lectins on the composition of the gut flora have also been studied. By modifying the glycosylation of epithelial surface cells and thus changing the sites to which bacteria can bind, lectins can induce shifts in the gut bacterial population, with important nutritional and physiological consequences (Beuth et al., 1995). For references also see van Damme et al. (1997). The bacterial flora in the gut can modulate the glycosylation patterns of the intestinal mucosa (Freitas et al., 2002). An early example of lectin-induced change in the bacterial population is PHA-induced coliform overgrowth in the rat small intestine (Pusztai et

90
al., 1993). The PHA induces an increase in crypt cell proliferation rate that leads to the flooding of the small intestinal villi with polymannosylated juvenile enterocytes, which in turn can form attachment sites to type-1-fimbriated Escherichia coli. As this binding of E. coli is mannose-dependent, the effects of the PHA-induced coliform overgrowth can be partially reversed by GNA, which is a mannose-specific lectin from snowdrop bulbs. This blocking of the attachment site of a bacterium by a lectin with similar sugar binding-specificity is called chemical probiosis (Pusztai et al., 1993).
A good example for this type of probiosis is the inhibition of Salmonella-binding to Caco-2 cells by PNA because one of the receptors recognised by Salmonella is galactosyl-N-acetylgalactosamine and this sugar structure is also recognised by PNA (Poschet & Fairclough, 1999). Orally given GNA significantly reduced the numbers of S. typhimurium S986 in the lower part of the small intestine and the large intestine of rats infected with this pathogen. As a result it also significantly improved rat growth (Naughton et al., 2000). However, GNA had much less effect on infection with S. enteritidis 857 and with Concanavalin A the infection became worse, particularly in the case of S. typhimurium. Protease inhibitors Pancreatic serine-proteases inhibitors are the best-known and most studied ANFs. Legumes, and particularly soybean and winged bean (Psophocarpus tetragonobolus) (Nishino et al., 20001) are major sources of these inhibitors. Traditionally they belong to two major classes: the Kunitz trypsin inhibitor or the family of Bowman-Birk trypsin/chymotrypsin inhibitors. This latter family occurs widely in legume seeds while Kunitz trypsin inhibitor is mainly found in soybeans. In most instances inclusion of protease inhibitors in the diet reduces the growth rate and feed-utilisation of animals. However, this does not appear to hold true for Atlantic salmon whose growth rate is stimulated by the presence of small amounts of potato (Solanum tuberosum) protease inhibitors in their diet (Sveier et al., 2001). References in previous reviews can be traced from the reviews of Friedman & Brandon (2001) and Lajolo & Genovese (2002) in which their occurrence and removal methods are described. This short review will emphasise possible useful applications.
Trypsin inhibitors are employed as tools in studies of diet-induced stimulation of pancreatic enzyme secretion. When trypsin/chymotrypsin inhibitors from the diet reach the duodenum they neutralise the proteases present. Therefore food digestion would stop without further influx of proteases from the pancreas. The reduction in duodenal protease level is a signal for the release of cholecystokinin from the duodenal epithelial endocrine cells that, in turn, after reaching the exocrine pancreas, stimulate the secretion of more serine proteases into the duodenum. Thus, in a negative feedback loop protease levels in the small bowel are regulated by the protease inhibitors consumed in the diet. Despite the attractiveness of this theory it is clear that this is not the only mechanism by which pancreatic protease secretion is controlled. Recent studies have shown that protease inhibitors pre-complexed with trypsin or chymotrypsin are just as effective in releasing serine-proteases from the pancreas as free, uncomplexed inhibitors (Pusztai et al., 1997b).
Similar to lectins, some of these inhibitors have insecticidal properties and their genes are frequently used in plant genetic modification. Even though this is outside the scope of this review, some of the well-studied inhibitor - insect interactions will be given. Thus, winged bean trypsin inhibitor extracts from rice (Oryza sativa) plants genetically modified with the gene mwti1b inhibited growth of the rice stem borer (Chilo suppressalis) (Mochizuki et al., 1999). The protease inhibitor accumulated in maize (Zea mays) plants in response to wounding or insect feeding is highly active against midgut proteases of the larvae of

91
Spodoptera littoralis (Tamayo et al., 2000). Thus, the gene of this trypsin inhibitor is a good candidate for use in plant protection against lepidopteran pests by genetic engineering. Other studies have confirmed the importance of protease inhibitors in plant protection. (Ortego et al., 2001) showed that when the level of wound-inducible proteinase inhibitors in potato was reduced by transgenic techniques the weight gain of Colorado potato beetle (Leptinotarsa decemlineata) larvae and beet armyworm (Spodoptera exigua) fed on genetically modified potatoes was significantly higher than that on conventional potatoes. Similarly, in the presence of soybean trypsin inhibitor the antilarval effectiveness of Bacillus thuringiensis endotoxin was increased, probably by increasing toxin retention time in the larval midgut (Zhang et al., 2000). Potato proteases were found to be good inhibitors of midgut proteases in the honeybee (Apis mellifera). When they were fed to bees at moderate concentrations, the bee lifespan was reduced (Malone et al., 1998). (Burgess et al., 2002) reported on prey-mediated effects of the protease inhibitor aprotinin on the predatory carabid beetle (Nebria brevicollis). With these beetles were fed on Helicoverpa armigera larvae that had originated from field-collected adults fed on an artificial diets with or without aprotinin, initially the beetles grew better in the absence of aprotinin in the prey than in its presence. However, at the last stage of feeding this growth pattern was reversed. This was found despite the fact that the trypsin level but not the chymotrypsin and elastase level, was lower in the aprotinin-containing prey (Burgess et al., 2002).
Trypsin inhibitors of the Kunitz type from soybean are human allergens (See Sicherer et al., 2000). Similarly, many of the trypsin inhibitors from potato are allergenic and cause wheal-and-flare responses in skin prick tests (Seppala et al., 2001).
Other useful and possible therapeutic applications are known. The most important of these is the potentially beneficial anticarcinogenic effect of soybean Bowman-Birk inhibitor for humans (Hawrylewicz et al., 1995; Kennedy, 1998), even though long-term use of this inhibitor in the diet of small mammals increases the chances of neoplastic pancreatic changes (Wormsley, 1988). It appears that used with soybean phytoestrogens, such as diadzein and genistein (McMichael-Phillips et al., 1998) and/or phospholipids (Foehr, et al., 1999) Bowman-Birk inhibitor may be a particularly effective anticarcinogen. Although human clinical trials have started to assess the effectiveness of this inhibitor in human cancer therapy, it is important to exercise caution. Most of the work to date suggesting a role for this inhibitor in cancer treatment has been done with in vitro tumour cells, (Friedman & Brandon, 2001).
Other possible beneficial effects of soybean trypsin inhibitors have been reported, such as the protection of mice against gentamycin-induced nephrotoxicity (Smetana et al., 1992) and a reduction in colon inflammation in mice with dextran sulphate sodium-induced ulcerative colitis (Ware et al., 1999). α-Amylase inhibitors α-Amylase inhibitors are widely distributed in plants (Buonocore & Silano, 1986; Grant et al., 1995), particularly in legumes and in some cereals. Some of these inhibitors in vitro were not found to be fully active against porcine pancreatic amylase unless the inhibitor and the enzyme were pre-incubated in the absence of starch. This was suggested as an explanation of the apparent lack of inhibitor effectiveness in starch degradation in humans (Koukiekolo et al., 1999). However, in other clinical studies purified α-amylase inhibitor preparations effectively inhibited intraduodenal amylase (Layer et al., 1985) and in normal or diabetic rats starch digestion was also inhibited, although not fully stopped, by intragastric administration of the inhibitor (Menezes & Lajolo 1987; Lajolo et al., 1991). In another study starch

92
digestion in the rat small intestine was inhibited by a preparation of pure kidney bean α-amylase inhibitor, leading to occasional blocking of the caecum by accumulated undigested starch, particularly at high inhibitor intake. Growth depression occurred due to losses of body nitrogen, lipids and carbohydrates (Pusztai et al., 1995). However, when peas (Pisum sativum) were genetically modified with the gene for kidney bean α-amylase inhibitor, neither the genetically engineered peas nor the inhibitor purified from them inhibited starch digestion in the rat intestine in vivo (Pusztai et al., 1999). Advances in ANF research - low molecular weight ANFs Polyphenols Polyphenols are one of the most numerous and widely distributed groups of substances in the plant kingdom. However, reliable data on polyphenol intake from food is scarce, because flavonoid intake is only calculated from the intake of flavonols, flavonons and flavanons (Bravo, 1998). To be accurate, these estimates ought to also include other compounds (catechins, anthocyanidins, isoflavones and chalcones).
Tannins (tannic acid) are water-soluble polyphenols present in many plant foods. They are considered nutritionally undesirable because they form complexes with proteins, starch and digestive enzymes and reduce the nutritional value of foods. However, it is now accepted that the most important dietary effect of condensed tannins in the digestive tract is due to their formation of less digestible complexes with dietary proteins, rather than inhibition of digestive enzymes (Chung et al., 1998a).
The bioavailability, absorption and metabolism of dietary polyphenols is primarily determined by such factors as their chemical structure, degree of glycosylation/acylation, conjugation with other molecules, molecular size, solubility. The metabolism and excretion of absorbed polyphenols is well known in experimental animals. However, such data are scarce for humans. Hydroxyl groups of the polyphenols are usually conjugated with glucuronic acid or sulphate in the liver, and methylation may also occur (Hollman & Katan, 1999). These groups are excreted in the urine and bile. In addition, these conjugates can be hydrolysed by microorganisms in the colon, and thus, can be reabsorbed and enter the enterohepatic cycle. The microorganisms cleave the heterocyclic ring, producing different phenolic acids, which are then absorbed and excreted in the urine (Heilman & Merfort, 1998). Different secondary metabolites have been detected in most mammalian species (Cook & Samman, 1996, Hollman, 1997).
Polyphenols are metabolised and absorbed in the gut, but information is still scarce about the uptake efficiency and the permanence of phenolic compounds, or their conjugates or derivatives, in the body. This knowledge is of great importance, because some of the physiologic effects of food polyphenols depend on their circulating levels (e.g. their antioxidant capacity (Rios et al., 2002). Recent studies have found that monomers, dimers and trimers of catechin are passively taken up through tight junctions, although this is more limited for the trimers. Higher oligomers and polymers might also enter the body by transcytosis as is known to happen for other large molecules, such as carrageens, β-lactoglobulin, and lactalbumin (Clifford, 1999).
Epidemiological studies on flavonoids and their beneficial health effects remain inconclusive. There is an inverse relationship between dietary flavonoid intake and mortality from coronary heart disease (CHD). This may be partially due to oxidation inhibition of low density lipoprotein and reduced platelet aggregation (Cook & Samman, 1996; Lampe, 1999; Sesso et al., 2003). Soybeans are unique among legumes because of their isoflavone (a

93
subclass of flavonoids) content. They have received considerable attention in recent years to establish a potential role in the prevention and treatment of a number of chronic diseases including certain forms of cancer (Kushi et al., 1999), osteoporosis, heart disease, and for their ability to relieve menopausal symptoms (Nijveldt et al., 2001).
The presence of tannins in legumes appears to be a double-edged sword. On one hand they may be beneficial to health due to their preventive potential in carcinogenesis and mutagenesis. However, on the other hand, they may be harmful because of their involvement in cancer-initiation, hepatoxicity, and antinutritional activity. Many tannins have been shown to reduce the mutagenicity of mutagens. These compounds may therefore be anticarcinogens. Thus, quercetin shows potent anticarcinogenic activity against cancers of the skin, colon and mammary gland in rodents. Gallic-, caffeic- and chlorogenic acid also reduced the effectiveness of some mutagens. Ellagic acid has been reported to significantly inhibit tumour formation in the colon, oesophagus, liver, lung, tongue and skin of rats and mice both in vitro and in vivo. Recently, this compound has also been shown to be a possible chemopreventive agent in human carcinogenesis. Several oligomers of hydrolysable tannins, including agrimoniin, oenothein B and coriariin A, exhibited antitumour activity against sarcomas (Chung et al., 1998a).
Tannins have also been shown to be harmful. They can produce hepatic necrosis in humans and grazing animals. By binding and precipitating the epithelial proteins tannins can penetrate the superficial cells and induce liver damage (Chung et al., 1998b). Tannin-related compounds also caused tumours in experimental animals. It was suggested that this may not have been due to DNA mutation but because of their ability to cause cellular irritation and damage (Ramanathan et al., 1992; Onodera et al., 1994; Jeng et al., 2001). Tannins, can act as co-carcinogens or promoters in inducing skin carcinomas in the presence of other carcinogens. However, further research is needed to identify the role of all the components in the cancer-initiating process. The immunomodulation activity of tannic acid has also been investigated (Feldman et al., 1999).
Tannins can theoretically serve as natural regulators of microbial populations in different habitats including the human gastrointestinal tract (Chung et al., 1998b). However, before tannins could be used in medical applications the effect of these compounds on the intestinal microflora and the biotransformation of other chemicals in the gastrointestinal tract need to be studied in detail (Haslam, 1996). Although ingestion of large quantities of tannins may result in adverse health effects, the intake of small quantities of some tannins could protect the gastrointestinal tract against ulcers and gastric or colon carcinomas and thus may be beneficial to human health. However, for this it will be necessary to determine the right dose of the appropriate tannin.
Phytate Phytate is considered to be an antinutrient because its contribution to poor mineral bioavailability of legumes (Harland & Morris, 1995, Adams et al., 2002). Suboptimal mineral status has been shown to cause increased morbidity, poor pregnancy outcomes, impairment of growth, immune competence and cognitive function. All this underlines the need to optimise Fe and Zn bioavailability (Lönnerdal, 2002). Phytic acid is also known to be an antioxidant.
Recent studies in pigs, as models for humans, have tried to elucidate the pathway of inositol hydrolysis in the gut (Schlemmer et al., 2001). Phytate degradation in the gut was nearly complete and similar for diets, which were very low in intrinsic feed phytases (phytase-inactivated diet) and the control diet. However, the apparent availability of total phosphorus

94
was significantly higher in pigs fed the control than the phytase-inactivated diet. Inositol hexaphosphate is absorbed through the stomach and upper small intestine, is quickly dephosphorylated in the mucosal cells and is distributed to different tissues. The dephosphorylation of inositol hexaphosphate in the gastrointestinal tract in humans and animals could have a possible role in the chemoprevention of pulmonary carcinogenesis (Wattenberg, 1999). It has also been postulated that it may play a role in reducing the risk of colon and possibly breast cancer (Vucenik et al., 1997). Phytic acid forms chelates with various metals, which can suppress damaging iron-catalysed redox reactions (Graf & Eaton, 1993). By suppressing the cell damage of the intestinal epithelium, the harmful effects of the oxygen radicals produced by colonic bacteria may be reduced by dietary phytic acid (Messina, 1999). Oligosaccharides Oligosaccharides commonly present in legumes are held responsible for gas production in the gut, because there is no α-galactosidase in the human small bowel mucosa to hydrolyse the α-(1-6) galactose linkage present in the galactose-containing oligosaccharides (raffinose, stachyose and verbascose) consumed in the diet. These sugars therefore pass unchanged into the large intestine where the colon bacteria metabolise them forming large amounts of CO2, H2, and sometimes methane. It is possible to remove a part of the oligosaccharides, and thus reduce flatulence by processing. However, nowadays this is not always done because they may also have beneficial effects for humans. One of these is their promotion of the growth of bifidobacteria in the colon. Thus, oligosaccharides are now considered as prebiotics, improving colon health, increasing longevity, and decreasing the risk of colon cancer (Messina, 1999). Saponins Legumes are a major source of saponins. They are poorly absorbed from the gut, because they form insoluble complexes with 3-β-hydroxysteroids and interact and form large, mixed micelles with bile acids and cholesterol. Saponins are therefore antinutrients. Although saponins have been shown to lower cholesterol levels in some animals, their hypocholesterolemic effects in humans is more speculative. They may have anticancer properties, as suggested by a recent rodent study in which feeding a saponin-containing diet inhibited the development of preneoplastic lesions in the colon (Koratkar & Rao, 1997), but these results may be irrelevant to humans (Ridout et al., 1988). References Adams, C.L., Hambidge, M., Raboy, V., Dorsch, J.A., Sian, L., Westcott, J.L. & Krebs, N.F.
2002. Zinc absorption from a low-phytic acid maize. American Journal of Clinical Nutrition 76, 556-559.
Baintner, K., Kiss, P., Pfüller, U., Bardocz, S. & Pusztai, A. 2003. Effect of orally and intraperitoneally administered plant lectins on food consumption of rats. Acta Physiologica Hungarica 90, 97-107.
Bandyopadhyay, S., Roy, A. & Das, S. 2001. Binding of garlic (Allium sativum) leaf lectin to the gut receptors of homopteran pests is correlated to its insecticidal activity. Plant Science 161, 1025-1033.

95
Beuth, J., Ko, H.L., Pulverer, G., Uhlenbruck, G. & Pichlmaier, H. 1995. Importance of lectins for the prevention of bacterial infections and cancer metastases. Glycoconjugate Journal 12, 1-6.
Bravo, L. 1998. Polyphenols: chemistry, dietary sources, metabolism and nutritional significance. Nutritional Reviews 56, 317-333.
Buonocore, V. & Silano, V. 1986. Biochemical, nutritional and toxicological aspects of alpha-amylase inhibitors from plant foods. Advances in Experimental and Medical Biology 199, 483-507.
Burgess, E.P.J., Lövei, G.L., Malone, L.A., Nielsen, I.W., Gatehouse, H.S. & Christeller, J.T. 2002. Prey-mediated effects of the protease inhibitor aprotinin on the predatory beetle Nebria brevicollis. Journal of Insect Physiology 48, 1093-1101.
Buttle, L.G., Burrells, A.C., Good, J.E., Williams, P.D. Southgate, P.J. & Burrells, C. 2001. The binding of soybean agglutinin (SBA) to the intestinal epithelium of Atlantic salmon, Salmo salar and Rainbow trout, Oncorhynchus mykiss, fed high levels of soybean meal. Veterinary Immunology and Immunopathology 80, 237-244.
Chung, K.T., Wei, C.I. & Johnson M.G. 1998b. Are tannins a double-edged sword in biology and health? Trends in Food Science & Technology 9, 168-175.
Chung, K.T., Wong, T.Y., Wei, C.I., Huang, Y.W. & Lin, Y. 1998a. Tannins and human health: a review. Critical Reviews in Food Science and Nutrition 38, 421-464.
Clifford, M.N. 1999. CPT and Health. Intermediate report, FAIR Project Contract No PL95-0653.
Cook, N.C. & Samman, S. 1996. Flavonoids-chemistry, metabolism, cardioprotective effects, and dietary sources. Journal of Nutritional Biochemistry 7, 66-76.
Couty, A., Down, R.E., Gatehouse, A.M.R., Kaiser, L., Pham-Delégue, M.H. & Poppy, G.M. 2001b. Effects of artificial diet containing GNA and GNA-expressing potatoes on the development of the aphid parasitoid Aphidius ervi Haliday (Hymenoptera: Aphidiidae). Journal of Insect Physiology 47, 1357-1366.
Couty, A. & Poppy, G.M. 2001. Does host-feeding on GNA-intoxicated aphids by Aphelinus abdominalis affect their longevity and/or fecundity? Entomologia Experimentalis et Applicata 100, 331-337.
Couty, A., Vina, G. de la, Clark, S.J., Kaiser, L., Pham-Delégue, M.H. & Poppy, G.M. 2001a. Direct and indirect sublethal effects of Galanthus nivalis agglutinin (GNA) on the development of a potato-aphid parasitoid, Aphelinus abdominalis (Hymenoptera: Aphelinidae). Journal of Insect Physiology 47, 553-561.
Feldman, K.S., Sahasrabudhe, K., Smith, R.S. & Scheuchenzuber, J. 1999. Immunostimulation by plant polyphenols: a relationship between tumor necrosis factor-α production and tannin structure. Bioorganic & Medical Chemistry Letters 9, 985-990.
Fitches, E., Ilett, C., Gatehouse, A.M.R., Gatehouse, L.N., Greene, R., Edwards, J.P. & Gatehouse, J.A. 2001. The effects of Phaseolus vulgaris erythro- and leucoagglutinating isolectins (PHA-E and PHA-L) delivered via artificial diet and transgenic plants on the growth and development of tomato moth (Lacanobia oleracea) larvae lectin binding to gut glcoproteins in vitro and in vivo. Journal of Insect Physiology 47, 1389-1398.
Foehr, M.W., Tomei, L.D., Goddard, J.G., Pemberton, P.A. & Bothurst, I.C. 1999. Antiapoptotic activity of Bowman-Birk inhibitor can be attributed to copurified phospholipids. Nutrition and Cancer 34, 199-205.
Freitas, M., Axelsson, L.G., Cayuela, C., Mitvedt, T. & Trugnan, G. 2002. Microbial-host interactions specifically control the glycosylation pattern in the intestinal mouse mucosa. Histochemistry Cell Biology 118, 149-161.

96
Friedman, M. & Brandon, D.L. 2001. Nutritional and health benefits of soy proteins. Journal of Agricultural and Food Chemistry 49 1069-1086).
Graf, E. & Eaton, J.W. 1993. Suppression of colon cancer by dietary phytic acid. Nutrition and Cancer 19, 11-19.
Grant, G., Edwards, J.E. & Pusztai, A. 1995. α-Amylase inhibitor levels in seeds generally available in Europe. Journal of the Science of Food and Agriculture 67, 235-238.
Haas, H., Falcone, F.H., Schramm, G., Haisch, K., Gibbs, B.F., Klaucke, J., Pöppelmann, M., Becker, W.M., Gabius, H.J. & Schlaak, M. 1999. Dietary lectins can induce in vitro release of IL-4 and IL-13 from human basophils. European Journal of Immunology 29, 918-927.
Habibi, J., Backus, E.A. & Huesing, J.E. 2000. Effects of phytohemagglutinin (PHA) on the structure of midgut epithelial cells and localization of its binding sites in western tarnished plant bug, Lygus hesperus Knight. Journal of Insect Physiology 46, 611-619.
Harland, B.F. & Morris, E.R. 1995. Phytate: a good or a bad food component? Nutrition Research 15, 733-754.
Haslam, E. 1996. Natural polyphenols (vegetable tannins) as drugs: Possible modes of action. Journal of Natural Products 59, 205-215.
Hawrylewicz, E.J., Zapata, J.J. & Blair, W.H. 1995. Soy and experimental cancer: animal studies. Journal of Nutrition 125, 698S-708S.
Heilmann, J. & Merfort, Y. 1998. Aktueller Kenntnisstand zum metabolismus von flavonoiden-. II. Resorption und metabolismus von flavonen, flavononen, flavanen, proantocyanidinen und isoflavonoiden. Pharmacology in unserer Zeit 27, 173-183.
Hollman, P.C.H. 1997. Bioavailability of flavonoids. European Journal of Clinical Nutrition 51, Supplement 1, S66-S69.
Hollman, P.C.H. & Katan, M.B. 1999. Dietary flavonoids: intake, health effects and bioavailability. Food Chemistry Toxicology 37, 937-942.
Hopkins, T.L. & Harper, M.S. 2001. Lepidopteran peritrohic membranes and effects of dietary wheat germ agglutinin on their formation and structure. Archives of Insect Biochemistry and Physiology 47, 100-109.
Jeng, J.H., Chang, M.C. & Hahn, L.J. 2001. Role of areca nut in betel quid-associated chemical carcinogenesis: current awareness. Oral Oncology 37, 477-492.
Jordinson, M., Fitzgerald, A.J., Goodlad, R.A., Brynes, A., Grant, G., Pignatelli, M. & Calam, J. 2000. Systemic effect of peanut agglutinin following intravenous infusion in to rats. Alimentary Pharmacology Therapy 14, 835-840
Jordinson, M., Goodlad, R.A., Brynes, A., Bliss, P., Ghatei, M.A., Bloom, S.R., Fitzgerald, A., Grant, G., Bardocz, S., Pusztai, A., Pignatelli, M. & Calam, J. 1999. Gastrointestinal responses to a panel of lectins in rats maintained on total parenteral nutrition. American Journal of Physiology, Gastrointestinal Liver Physiology 39, G1235-G1242.
Kellens, J.T., Jacobs, J.A., Peumans, W.J. & Stobberingh, E.E. 1995. Agglutination of Streptococcus milleri by lectins.. Journal of Medical Microbiology 41, 14-19.
Kennedy, A.R. 1998. The Bowman-Birk inhibitor from soybeans as an anticarcinogenic agent. American Journal of Clinical Nutrition 68, 1406S-1412S.
Koratkar, R. & Rao, A.V. 1997. Effect of soya bean saponins on azoxymethane-induced preneoplastic lesions in the colon of mice. Nutrition and Cancer 27, 206.209.
Kordás, K., Burghardt, B., Kisfalvi, K., Bardocz, S., Pusztai, A. & Varga, G. 2000. Diverse effects of phytohaemagglutinin on gastrointestinal secretions in rats. Journal of Physiology (Paris) 94, 31-36.

97
Kordás, K., Szalmay, G., Bardocz, S., Pusztai, A. & Varga, G. 2001. Phytohaemagglutinin inhibits gastric acid but not pepsin secretion in conscious rats. Journal of Physiology (Paris) 95, 1-6.
Koukiekolo, R., Le Berre-Anton, V., Desseaux, V., Moreau, Y., Rougé, P., Marchis-Mouren, G. & Santimone, M. 1999. Mechanism of porcine pancreatic α-amylase inhibition of amylose and maltopentaose hydrolysis by kidney bean (Phaseolus vulgaris) inhibitor and comparison with that by acarbose. European Journal of Biochemistry 265, 20-26.
Kushi, L.H., Meyer, K.A. & Jacobs, D.R. 1999. Cereals, legumes, and chronic disease risk reduction: evidence from epidemiologic studies. American Journal of Clinical Nutrition 70, 451S-458S.
Lajolo, F.M., Finardi-Filho, F. & Menezes, E.W. 1991. Amylase inhibitors in Phaseolus vulgaris beans. Food Technology 45, 119-121.
Lajolo, F.M. & Genovese, M.I. 2002. Nutritional significance of lectins and enzyme inhibitors from legumes. Journal of Agricultural and Food Chemistry 50, 6592-6598.
Lampe, J.W. 1999. Health effects of vegetables and fruit: assessing mechanisms of action in human experimental studies. American Journal of Clinical Nutrition 70, 475S-490S.
Lavelle, E.C., Grant, G., Pusztai, A., Pfüller, U. & O'Hagan, D.T. 2000. Mucosal immunogenecity of plant lectins in mice. Immunology 90, 30-37.
Lavelle, E.C., Grant, G., Pusztai, A., Pfüller, U. & O'Hagan, D.T. 2001. The identification of plant lectins with mucosal adjuvant activity. Immunology 102, 77-86.
Layer, P., Carlson, G.L. & DiMagno, E.P. 1985. Partially purified white bean amylase inhibitor reduces starch digestion in vitro and inactivates intraduodenal amylase in humans. Gastroenterology 88, 1985-1992.
Lönnerdal, B. 2002. Phytic acid-trace element (Zn, Cu, Mn) interactions. International Journal of Food Science and Technology 37, 749-758.
Machuka, J., Damme, E.J.M. van, Peumans, W.J. & Jackai, L.E.N. 1999. Effect of plant lectins on larval development of the legume pod borer; Maruca vitrata. Entomologia Experimentalis et Applicata 93, 179-187.
Machuka, J.S., Okeola, O.G., Chrispeels, M.J. & Jackai, L.E.N. 2000. The African yam bean seed lectin affects the development of the cowpea weevil but does not affect the development of larvae of the legume pod borer. Phytochemistry 53, 667-674.
Malone, L.A., Burgess, E.P.J., Christeller, J.T. & Gatehouse, H.S. 1998. In vivo responses of honey midgut proteases to two protease inhibitors from potato. Journal of Insect Physiology 44, 141-147.
McMichael-Phillips, D.F., Harding, C., Morton, M., Roberts, S.A., Howell, A., Potten, C.S. & Bundred, N.J. 1998. Effects of soy protein supplementation on epithelial proliferation in the histologically normal human breast. American Journal of Clinical Nutrition 68, 1431S-1435S.
Menezes, E.W. & Lajolo, F.M. 1987. Inhibition of starch digestion by black bean α-amylase inhibitor, in normal and diabetic rats. Nutrition Reports International 36, 1185-1189.
Messina, M.J. 1999. Legumes and soybeans: overview of their nutritional profiles and health effects. American Journal of Clinical Nutrition 70, 439S-450S.
Mochizuki, A., Nishizawa, Y., Onodera, H., Tabei, Y., Toki, S., Habu, Y., Ugaki, M. & Ohashi, Y. 1999. Transgenic rice plants expressing trypsin inhibitor are resistant against rice stem borers, Chilo suppressalis. Entomologia Experimentalis et Applicata 93, 173-178.
Murdock, L.L. & Shade, R.E. 2002. Lectins and protease inhibitors as plant defenses against insects. Journal of Agricultural and Food Chemistry 50, 6605-6611.

98
Naughton, P.J., Grant, G., Bardocz, S. & Pusztai, A. 2000. Modulation of Salmonella infection by the lectins of Canavalia ensiformis (Con A) Galanthus nivalis (GNA) in a rat model in vivo. Journal of Applied Microbiology 88, 720-727.
Nijveldt, R.J., Nood, E., Hoorn, D.E.C., Boelens, P.G., Norren, K. & Leeuwen, P.A.M. 2001. Flavonoids: a review of probable mechanisms of action and potential applications. American Journal of Clinical Nutrition 74, 418-425.
Nishino, N., Taniguchi, M. & Sakaguchi, E. 2001. Response of intestinal proteinase activities to the feeding of isolated winged bean (Psophocarpus tetragonobolus) and soya bean (Glycine max) trypsin inhibitors in rats. Journal of the Science of Food and Agriculture 81, 1055-1059.
Onodera, H., Kitaura, K., Mitsumori, K., Yoshida, L., Yasuhara, K., Shimo, T., Takahashi, M. & Hayashi, T. 1994. Study on the carcinogenicity of tannic acid in F344 rats. Food Chemistry Toxicology 32, 1101-1106.
Ortego, F., Novillo, C., Sánchez-Serrano, J.J. & Castañera, P. 2001. Physiological response of Colorado potato beetle and beet armyworm larvae to depletion of wound-inducible proteinase inhibitors in transgenic potato plants. Journal of Insect Physiology 47, 1291-1300.
Ovelgönne, J.H., Koninkx, J.F.J.G., Pusztai, A., Bardocz, S., Kok, W., Ewen, S.W.B., Hendriks, H.G.C.J.M. & Dijk, J.E. van, 2000. Decreased levels of heat shock proteins in gut epithelial cells after exposure to plant lectins. Gut 46, 679-687.
Paes, N.S., Gerhardt, I.R., Coutinho, M.V., Yokoyama, M., Santana, E., Harris, N., Chrispeels, M.J. & Grossi de Sa, M.F. 2000. The effect of arcelin-1 on the structure of the midgut of bruchid larvae and immunolocalization of the arcelin protein. Journal of Insect Physiology 46, 393-402.
Poschet, J.F. & Fairclough, P.D. 1999. Effect of lectins on Salmoinella tphimurium invasion in cultured human intestinal cells. Gut 44(S1), W252.
Porter, J. & Pickup, R.N. 1998. Separation of natural populations of coliform bacteria from freshwater and sewage by magnetic bead sorting. Journal of Microbiology Methods 33, 221-226.
Pryme, I.F., Pusztai, A., Bardocz, S. & Ewen, S.W.B. 1999a. A combination of dietary protein depletion and PHA-induced gut growth reduce the mass of a murine non-Hodgkin lymphoma. Cancer Letters 139, 145-152.
Pryme, I.F., Bardocz, S., Pusztai, A. & Ewen, S.W.B. 1999b. The growth of an established murine non-Hodgkin lymphoma tumour is limited by switching to a phytohaemagglutinin-containing diet. Cancer Letters 146, 87-91.
Pryme, I.F., Bardocz, S. Pusztai, A. & Ewen, S.W.B. 2002. Dietary mistletoe supplementation and reduced growth of murine non-Hodgkin lymphoma. Histology and Histopathology 17, 261-271.
Pusztai, A. 1999. Phytohaemagglutinin stimulates pancreatic enzyme secretion in rats by a combination of cholecystokinin- and noncholecystokinin-linked pathways. In Biology of the Pancreas in Growing Animals. S.G. Pierzynowski & R. Zabielski (Eds). Elsevier Science B.V., Amsterdam, 273-286.
Pusztai, A., Ewen, S.W.B., Grant, G., Peumans, W.J., Damme, E.J.M. van, Rubio, L. & Bardocz, S. 1990. Relationship between survival and binding of plant lectins during small intestinal passage and their effectiveness as growth factors. Digestion 46, 308-316.
Pusztai, A., Grant, G., Bardocz, S., Alonso, R., Chrispeels, M.J., Schroeder, H.E., Tabe, L.M. & Higgins, T.J.V. 1999. Expression of the insecticidal bean α-amylase inhibitor transgene has minimal detrimental effect on the nutritional value of peas fed to rats at 30 % of the diet. Journal of Nutrition 129, 1597-1603.

99
Pusztai, A., Grant, G., Bardocz, S., Baintner, K., Gelencsér, E. & Ewen, S.W.B. 1997b. Both free and complexed trypsin inhibitors stimulate pancreatic secretion and change duodenal enzyme levels. American Journal of Physiology, Gastrointestinal Liver Physiology 35, G340-G350.
Pusztai, A., Grant, G., Bardocz, S., Gelencsér, E. & Hajós, G. 1997a. Novel dietary strategy for overcoming the antinutritional effects of soybean whey of high agglutinin content. British Journal of Nutrition 77, 933-945.
Pusztai, A., Grant, G., Buchan, W.C., Bardocz, S., Carvalho de, A.F.F.U. & Ewen, S.W.B. 1998. Lipid accumulation in obese Zucker rats is reduced by inclusion of raw kidney bean (Phaseolus vulgaris) in the diet. British Journal of Nutrition 79, 213-221.
Pusztai, A., Grant, G., Duguid, T., Brown, D.S., Peumans, W.J., van Damme, E.J.M. & Bardocz, S. 1995. Inhibition of starch digestion by α-amylase inhibitor reduces the efficiency of utilization of dietary proteins and lipids and retards the growth of rats. Journal of Nutrition 125, 1554-1562.
Pusztai, A., Grant, G., Spencer, R.J., Duguid, T.J., Brown, D.S., Ewen, S.W.B., Peumans, W.J., van Damme, E.J.M. & Bardocz, S. 1993. Kidney bean lectin-induced Escherichia coli overgrowth in the small intestine is blocked by GNA, a mannose-specific lectin. Journal of Applied Bacteriology 75, 360-368.
Ramanathan, R., Tan, C.H. & Das, N.P. 1992. Tannic acid promotes benzo(a)pyrene-induced mouse skin carcinogenesis at low concentrations. Medical Science Research 20, 711-712.
Ridout, C.L., Wharf, G., Price, L.R., Johnson, L.T. & Fenwick, G.R. 1988. UK mean daily intakes of saponins-intestine-permeabilizing factors in legumes. Food Science and Nutrition 42, 111-116.
Rios, L.Y., Bennett, R.N., Lazarus, S.A., Rémésy, C., Scalbert, A. & Williamson, G. 2002. Cocoa procyanidins are stable during gastric transit in humans. American Journal of Clinical Nutrition 76, 1106-1110.
Ryder, S.D., Jacyna, M.R., Levi, A.J. Rizzi, P.M. & Rhodes, J.M. 1998. Eating peanuts increases rectal proliferation in individuals with mucosal expression of peanut lectin receptor. Gastroenterology 114, 44-49.
Ryder, S.D., Parker, N., Eccleston, D., Haqqani, M.T. & Rhodes, J.M. 1994. Peanut lectin (PNA) stimulates the proliferation of colonic explants from patients with ulcerative colitis, Crohn's disease, and colonic polyps. Gastroenterology 106, 117-124.
Schlemmer, U., Jany, K.D., Berk, A., Schulz, E. & Rechkemmer, G. 2001. Degradation of phytate in the gut of pigs- Pathway of gastrointestinal inositol phosphate hydrolysis and enzymes involved.. Archives of Animal Nutrition 55, 255-280.
Seppälä, U., Majmaa, H., Turjanmaa, K., Helin, J., Reunala, T., Kalkkinen, N. & Paloosuo, T. 2001. Identification of four novel potato (Solanum tuberosum) allergens belonging to the family of soybean trypsin inhibitors. Allergy 56, 619-626.
Sesso, H.D., Gaziano, J.M., Liu, S. & Buring, J.E. 2003. Flavonoid intake and the risk of cardiovascular disease in women. American Journal of Clinical Nutrition 77, 1400-1408.
Sétamou, M., Bernal, J.S., Legaspi, J.C. & Mirkov, T.E. 2002a. Effects of snowdrop lectin (Galanthus nivalis agglutinin) expressed in transgenic sugarcane on fitness of Cotesia flavipes (Hymenoptera: Braconidae), a parasitoid of the nontarget pest Diatraea saccharalis (Lepidoptera: Crambidae). Annals of the Entomology Society of America 95, 75-83.
Sétamou, M., Bernal, J.S., Legaspi, J.C., Mirkov, T.E. & Legaspi, B.C. Jr. 2002b. Evaluation of lectin-expressing sugarcane against stalkborers (Lepidoptera: Pyralidae): Effects on life history parameters.. Journal of Economic Entomology 95, 469-477.

100
Sicherer, S.H., Sampson, H.A. & Burks, A.W. 2000. Peanut and soy allergy, a clinical and therapeutical dilemma. Allergy 55, 515-521.
Smetana, S., Khalef, S., Bar-Khayim, Y. & Birk, Y. 1992. Antimicrobial gentamycin activity in the presence of exogenous protease inhibitor (Bowman-Birk inhibitor) in gentamycin-induced nephrotoxicity in rats. Nephron 61, 68-72.
Sveier, H., Kvamme, B.O. & Raae, A.J. 2001. Growth and protein utilization in Atlantic salmon (Salmo salar L.) given a protease inhibitor in the diet. Aquaculture Nutrition 7, 255-264.
Tamayo, M.C., Rufat, M., Bravo, J.M. & San Segundo, B. 2000. Accumulation of a maize proteinase inhibitor in response to wounding and insect feeding, and characterization of its activity toward digestive proteinases of Spodoptera littoralis larvae. Planta 211, 62-71.
Triguéros, V., Wang, M., Pére, D., Paquereau, L., Chavant, L. & Fournier, D. 2000. Modulation of a lectin insecticidal activity by carbohydrates. Archives of Insect Biochemistry and Physiology 45, 175-179.
Van Damme, E.J.M., Peumans, W.J., Pusztai, A. & Bardocz, S. 1997. Plant lectins in mammalian nutrition, immunology, metabolism and as oral therapeutic and immune agents. In Handbook of Plant Lectins: Properties and Biomedical Applications. E.J.M van Damme, , W.J. Peumans, A. Pusztai & S. Bardocz (Eds). John Wiley & Sons, Chichester, 31-50.
Vázquez, R.I., Moreno-Fierros, L., Neri-Bazán, L., de la Riva, G.A. & López-Revilla, R. 1999. Bacillus thuringiensis Cry1Ac protoxin is a potent systemic and mucosal adjuvant. Scandinavian Journal of Immunology 49, 578-584.
Vucenik, Y., Yang, G.Y. & Shamsuddin, A.M. 1997. Comparison of pure inositol hexaphosphate and high-bran diet in the prevention of DMBA-induced rat mammary carcinogenesis. Nutrition and Cancer 28, 7-13.
Ware, J.H., Wan, X.S., Newbern, P. & Kennedy, A.R. 1999. Bowman-Birk inhibitor reduces colon inflammation in mice with dextran sulphate sodium-induced ulcerative colitis. Digestive Diseases and Sciences 44, 986-990.
Wattenberg, L.W. 1999. Chemoprevention of pulmonary carcinogenesis by myo-inositol. Anticancer Research 19, 3659-3661.
Wormsley, K.G. 1988. Trypsin inhibitors: potential concern for humans. Journal of Nutrition 118, 134-136.
Zhang, J.H., Wang, C.Z. & Qin, J.D. 2000. The interactions between soybean trypsin inhibitor and delta-endotoxin of Bacillus thuringiensis in Helicoverpa armigera larva. Journal of Invertebrate Pathology 75, 259-266.

101
Lupinus campestris seed as a source of compounds with antimutagenic activity C. Jiménez1, G. Loarca-Piña2 & G. Dávila1 1Departamento de Graduados e Investigación en Alimentos, Escuela Nacional de Ciencias Biológicas, Instituto Politécnico Nacional, México D.F. 11340 México 2PROPAC, Universidad Autónoma de Querétaro, Querétaro, Qro., 76010 México Summary Lupin (Lupinus spp) seed can contain high amounts of protein (40 %) and oil (14 %). About 90 species of lupin have been reported in Mexico. The use of this genus as a source of food has been limited by the presence of antinutritional factors (ANFs), such as phenolic compounds (PC), carbohydrates (CH) and quinolizidine alkaloids (QAs). However, it has also been suggested that consumption of these compounds can affect human health and may reduce the risk of disease. The aim of this work was to determine the effect of Lupinus campestris PC, CH and QAs, on the mutagenicity of 1-nitropyrene (1-NP) using Salmonella typhimurium tester strain YG1024. The results indicated that Lupinus campestris seed has 11 mg (+) catechin eq/g seed coat; 120.3 and 2.13 mg/g of seed of CH and QAs, respectively. The 1-NP mutagenicity was inhibited in 86, 76 and 75 % for PC, CH and QAs at concentrations of 200, 512 and 13.6 µg/tube, respectively. Keywords: Lupinus campestris phenolic compounds, quinolizidine alkaloids, carbohydrates,
antimutagenicity Introduction Recently emphasis has been placed on dietary habits as significant factors affecting human health. Epidemiological studies have shown a good relationship between good dietary habits and a low cancer incidence (Stravic, 1994).
Lupins are cultivated on approximately 106 ha in the world. In Mexico there are about 90 wild species. Lupinus campestris seed contains about 44 % protein (Jiménez et al, 2001). Lupins contain PC, CH and QAs that may affect human health or reduce disease risk (Stravic, 1994; Tzyh et al., 1999).
Nitroarenes are potent mutagens for Salmonella typhimurium. The compound 1-NP is a mutagen that requires metabolic activation towards arylhydroxilamines by acetyl-CoA:N-Hydroxyarylamine o-acetyltransfere, which is present in bacterial cells (Mermelstein et al., 1981).
In this study a microsuspension assay was used to examine the effects of the PC, CH, and QAs from Lupinus campestris seed against the mutagenicity of 1-NP. Materials and methods Wild L. campestris seed was collected over a range of 50 km in the Mexican State of Morelos. Seed was stored at 4°C, until used. Phenolic compounds, CH and QAs were extracted and measured using the methods of Deshpande & Cheryan (1987), Muzquiz et al. (1992) and Muzquiz et al. (1994) respectively. To test mutagenicity we used Salmonella typhimurium tester strain YG 1024. The mutagenic activity of 1-NP and the antimutagenic activity of PC,

102
CH and QAs, isolated from Lupinus campestris seed, were determined using the Kado microsuspension assay (Kado et al., 1983, 1986). In each experiment samples were tested in triplicate. Antimutagenicity was expressed as the percent reduction of 1-NP mutagenicity according to the formula:
% Inhibition = 100-[x1/x2(100)] where x1 is the number of revertants/plate in the presence of the extract, expressed as mg (+) catechin equivalent/g seed, CH or QAs mg/g of seed and x2 is the number of revertants/plate in the absence of extract. Statistical analysis Mutagenicity results represent the average and standard deviation (SD) of triplicate plates/dose for the two experiments in each test series. Statistical differences between the control and treatments were analysed by Dunnett’s method. Results and discussion Measurement of phenolic compounds, carbohydrates and quinolizidine alkaloids The testa of L campestris was 20 % of the total seed. From the testa 543 mg/100 g of methanol extract of PC was obtained with a concentration of 11 mg (+) catechin eq/g of seed. This was lower than in other legumes. The CH content of L. campestris seed was 120 mg/g. This included the di- and oligosaccharides sucrose, melibiose, raffinose, ciceritol, stachyose and verbascose. The QAs concentration was 2.13 mg/g, lower than reported by Hatzold et al. (1983) in L. mutabilis (3.1 mg/g). The major alkaloids were hydroxyaphyllidine and hydroxyaphylline with concentrations of 1.47 and 0.49 mg/g respectively. They represented 92.4 % of the total QAs present. The alkaloids (eight components) were identified by their mass fragmentation. Antimutagenic activity of the phenolic compounds The dose-response curve of 1-NP mutagenicity in tester strain YG-1024 is shown in Fig. 1. The highest dose tested (1.6 µg/tube) was toxic to the bacteria. There was a good dose-response relationship up to approximately 0.5 µg/tube, so 0.4 µg/tube was chosen for all the antimutagenicity assays.
The PC from L. campestris seed were not toxic to the bacteria at the concentrations tested as the number of revertant colonies was similar to the control. The inhibitory effect of PC from L. campestris seeds on 1-NP mutagenicity in tester strain YG 1024 is shown in Table 1. Inhibition on 1-NP was dose-dependent between 0 - 200 µg phenolic extract/tube and gave a maximum inhibition of 86.25 %.
The results show that the PC present in L. campestris seed are similar to those in other legume seeds and they inhibit damage caused by toxic substances like nitroarenes. González de Mejía et al. (1999) and Cardador et al. (2002) showed that PC from common bean (Phaseolus vulgaris) cv. Flor de Mayo FM-38 inhibited benzo[a]pyrene and aflatoxin B1, mutagenicity with a dose related response with Salmonella tester strains YG1024, TA98 and TA100 in a microsuspension assay (50 - 85 %).

103
0
300
600
900
1200
0 0,5 1 1,5 2
1-NP (µg/plate)
Reve
rtant
s /pl
ate
Figure 1. The dose response curve of 1-NP mutagenicity in Salmonella typhimurium
YG1024. The spontaneous mutation rate was 43 ± 13.4 revertants/plate. Table 1. The antimutagenic activity of phenolic compounds extracted from L. campestris
testas against 1-NP2 in YG1024. Phenolic extract (µg/tube)1 Revertants/plate Inhibition (%)
0.0 1,144 ± 51a 0.0 0.025 1,055 ± 53b 8.1 0.05 1,006 ± 26b 12.1 0.10 870 ± 25b 24.0 0.20 763 ± 08b 33.3
25.00 604 ± 22b 47.2 50.00 481 ± 19b 57.9
100.00 223 ± 20b 80.5 200.00 157 ± 21b 86.3
1Concentration of PC expressed as µg equivalent of (+) catechin per tube. 21-NP -Nitropirene (400 ng/tube). The spontaneous mutation rate was 64 ± 3 revertants/plate. Different letters indicate a statistically significant difference from the control (Dunnett α= 0.05).
Antimutagenic activity of the carbohydrates The CH concentrations tested were in the range of 8 - 512 µg/plate. They were neither toxic nor mutagenic to the bacteria (Table 2). Both water and dimethyl sulphoxide (DMSO) were used as solvents to prepare the CH solutions. The maximum inhibition (32 to 76 %) was obtained for both solvents at the highest CH concentration (512 µg/tube). The higher inhibition from the DMSO extract could be due to the capacity of DMSO to increase CH solubility. Sreekumar & Hozono (1998) and Wang et al. (2002) reported similar results with an inhibition of 40 - 63 %. Antimutagenic activity of the quinolizidine alkaloids The inhibitory effect of the QAs is shown in Table 3. The QAs doses tested were in the range of 13.6 – 435.0 µg/tube. In contrast to the antimutagenic effects shown by PC and CH, the lowest dose of QAs had the highest inhibition of 1-NP mutagenicity (75 % reduction). However as the concentration of QAs increased their inhibition of mutagenicity decreased.

104
Table 2. The antimutagenic activity of carbohydrates (CH) extracted from L. campestris seed against 1-NP2 in YG1024.
Dissolved in water Dissolved in DMSO CH (µg/tube)1 Revertants/plate Inhibition (%) Revertants/plate Inhibition (%)
0 1,348 ± 120a 0.0 1,344 ± 31a 0.0 8 1,275 ± 16b 5.4 1,246 ± 61a 7.3
16 1,214 ± 71b 9.9 1,023 ± 37b 23.9 32 1,185 ± 35b 12.1 915 ± 55b 31.9 64 1,163 ± 66b 13.7 844 ± 46b 37.2
128 1,106 ± 40b 17.9 788 ± 50b 41.4 256 959 ± 65b 28.9 537 ± 55b 60.0 512 912 ± 20b 32.3 318 ± 8b 76.3
1Concentration of CH expressed as µg per tube. 21-NP -Nitropirene (400 ng/tube). Spontaneous mutation rate was 41 ± 2.5 revertants/plate. Different letters indicate a statistically significant difference from the control (Dunnett α= 0.05).
Table 3. The antimutagenic activity of quinolizidine alkaloids (QAs) extracted from
L. campestris seed against 1-NP2 in YG1024. QAs (µg/tube)1 Revertants/plate Inhibition (%)
0.0 1,450 ± 102a 0.0 13.6 357 ± 27b 75.4 27.2 859 ± 25b 40.8 54.4 1,332 ± 13b 8.2
108.8 1,327 ± 16a 8.4 217.5 1,470 ± 13a -1.4 435.0 1,849 ± 10b -26.6
1Concentration of QAs expressed as µg per tube. 21-NP -Nitropirene (400 ng/tube). Spontaneous mutation rate was 38 ± 2 revertants/plate. Different letters indicate a statistically significant difference from the control (Dunnett α= 0.05)
A concentration of QAs of 217.5 µg/tube did not show any antimutagenic effect. At a
concentration of 435 µg/tube the extract was mutagenic as the number of revertants was higher than in the control (1,849 and 1,450 respectively). Culvenor & Petterson (1986) reported that Lupinus angustifolius alkaloids were neither toxic nor mutagenic for tester strains TA98, TA100 and TA1538 using an incorporation and incubation assay. They also reported there was no effect in the chromosome aberration and point mutation test using a culture of Chinese hamster ovary cells. Acknowledgements
The authors thank the Instituto Politécnico Nacional and Consejo Nacional de Ciencia y Tecnología (CONACyT) through 33995 projects, for financial support for this research. Cristian Jiménez Martínez acknowledges a study grant from CONACyT.

105
References Cardador, M.A., Castaño, T.E. & Loarca, P.F.G. 2002. Antimutagenic activity of natural
phenolic compounds present in common beans against aflatoxin B1. Food Additives and Contaminants 19, 62-65.
Culvenor, C.C. & Petterson, D.S. 1986. Lupin toxins - alkaloids and phomopsis. Proceeding 4th International Lupin Conference, Geraldton, 15 – 22 August 1986. pp. 188-198.
Deshpande, S.S. & Cheryan, M. 1987. Determination of phenolic compounds of dry beans using vanillin, redox and precipitation assays. Journal of Food Science 52, 332-334.
González de Mejía, E., Castaño, T.E. & Loarca-Piña, G. 1999. Antimutagenic effect of natural phenolic compounds in beans. Mutation Research. Genetic Toxicology and Environmental Mutagenesis 44, 1-9
Hatzold, T., Ibraim, E., Gross, R., Wink M., Harmannn, H. & Witte, L. 1983. Quinolizidine alkaloids in seed of Lupinus mutabilis. Journal of Agricultural and Food Chemistry 31, 934-938
Jiménez, M.C., Hernández, S.H., Alvarez, M.G., Robledo, Q.N., Martínez, H.J. & Dávila, O.G. 2001. Effect of aqueous and alkaline thermal treatment on chemical composition and oligosaccharides, alkaloid and tannin content of Lupinus campestris seeds. Journal of The Science of Food and Agriculture 81, 421-428.
Kado, N., Langley, D. & Eisenstadt, E. 1983. A simple modification of the Salmonella liquid -incubation assay. Increased sensitivity for detecting mutagens in human urine. Mutation Research, 121, 25-32.
Kado, N.Y., Guirguis, G.N., Flessel, C.P., Chan, R.C., Chang, K. & Wesolowski, J.J. 1986. Mutagenicity of fine (<2.5 µm) airborne particles: diurnal variation in community air determinate by a Salmonella micro preincubation (microsuspension) procedure. Environmental Mutagenesis 8, 53-66.
Mermelstein R., Kiriazides D. K., Butler M., McCoy E. C. & Rosenkranz H. S., 1981. The extraordinary mutagenicity of nytropirenes in bacteria. Mutation Research 89,187-196.
Muzquiz, M., Cuadrado, C., Ayet, G., Cuadra, C. de la, Burbano, C. & Osagie, A. 1994. Variation of Alkaloid components of lupin seeds in 49 genotypes of Lupinus albus L. from different countries and locations. Journal of Agricultural and Food Chemistry 42, 1447-1450.
Muzquiz M., Rey, C. & Cuadrado, C. 1992. Effect of germination on the oligosaccharides content of lupin species. Journal of Chromatography 607, 349-352.
Sreekumar, O. & Hozono, A. 1998. The antimutagenic properties of a polysaccharide produced by Bifidobacterium longum and its cultured milk against some heterocyclic amine. Journal of Microbiology 44,1029-1036.
Stavric B., 1994, Antimutagens and anticarcinogens in foods. Food and Chemical Toxicology 32, 79-90.
Tzyh, C.H, Yu, C.L., Iou, S.C. & Jen. K.L. 1999. Inhibition of eleven mutagens by various tea extracts, (-)Epilogallocatechin-3-gallate, Gallic Acid and caffeine. Food and Chemical Toxicology 37, 569-579.
Wang, H.X., Andrade, L. & Engeseth, N.J. 2002. Antimutagenic effect of various honeys and sugar against Trp-p-1. Journal of Agricultural and Food Chemistry 50, 6923-6928.


107
Decreased levels of heat shock protein 70 and 90 in gut epithelial cells after exposure to plant lectins J. Koninkx1, H. Ovelgönne1, A. Pusztai2, S. Bardocz2, S. Ewen3, H. Hendriks1 & J. van Dijk1 1Department of Pathobiology, Division of Pathology, Faculty of Veterinary Medicine, Utrecht University, Yalelaan 1, Postbus 80.158, 3508 TD Utrecht, The Netherlands 2The Rowett Research Institute, Greenburn Road, Bucksburn, Aberdeen AB21 9SB, United Kingdom 3Department of Pathology, University of Aberdeen Medical School, Aberdeen AB1 2ZX, United Kingdom Summary To investigate whether PHA, a lectin from kidney bean (Phaseolus vulgaris) and WGA, a lectin from wheat germ (Triticum aestivum) were able to modify the heat shock response in gut epithelial cells, jejunal tissue sections from PHA and WGA-fed rats were screened for the expression of heat shock proteins (HSP) 70 and 90 using monoclonal antibodies. In addition, the levels of HSP70 and 90 in differentiated lectin-exposed Caco-2 cells were analysed by polyacrylamide gel electrophoresis and immunoblotting. Compared to the levels of HSP70 and 90 in the intestinal cells of lactalbumin-fed (LA) rats, the PHA and WGA-fed rats showed a severe decline in HSP levels. These results were confirmed by in vitro experiments using differentiated Caco-2 cells exposed to PHA-E4 and WGA. Keywords: heat shock proteins, lectins, rat small intestine, cell line Introduction Living cells have a highly conserved heat shock or stress response to adverse changes in their environmental conditions (Hightower, 1991). As a consequence of dietary intake, gut epithelial cells are regularly exposed to high levels of potentially harmful substances. At relatively high dietary intake lectins such as PHA and WGA induce hyperplastic growth of the rat gut (Pusztai et al., 1993) and alterations of the activities of brush border enzymes (Pusztai et al., 1996). Both in vitro (Koninkx et al., 1992) and in vivo (Pusztai et al., 1993) these lectins cause structural lesions in gut epithelial cells leading to severe disruptions in the integrity of the epithelial layer.
To assess the stress caused by exposure to these lectins we used differentiated Caco-2 cells to investigate if the lectins induce changes in the heat shock response of these cells. In particular, we examined the effect of both lectins on the expression of HSP90 and HSP70. We also screened slides made from the jejunum of rats, which had been experimentally exposed to either PHA or WGA, for the expression of HSPs in their gut epithelium. Materials and methods Animals, diets, tissue processing for histology and HSP immunostaining Six male, conventionally grown specific pathogen free inbred Hooded-Lister rats of the Rowett colony, kept single in metabolism cages, were prefed for 3 d (6 g/rat/d) on a semisynthetic, good quality diet consisting mainly of maize starch, potato starch and glucose.

108
This diet contained 10 % (w/w) lactalbumin (LA) as the sole protein. The rats weighing 80-90 g were divided into three groups of 2 rats each and pair-fed on different experimental diets for 10 d. The control group was kept on the same LA diet as used for pre-feeding. The WGA and PHA groups were fed diets based on the LA control diet in which 7 % (w/w) of the LA was replaced by an equal amount of either WGA or PHA (93 g LA + 7 g WGA or PHA/kg of diet).
The food intake of all three groups was restricted to the voluntary intake of the PHA-group (6 g diet/rat/d). This amount of diet contained 42 mg/d of WGA or PHA. On the morning of the tenth day, after overnight fasting, rats were offered 2 g of their respective diets and killed under ether anaesthesia 2 h later. The abdomen was cut open and 2 gut sections each of 2 cm cut 7 cm from the pylorus were taken for histology and heat shock protein immunostaining. After deparaffination sections were incubated with monoclonal antibodies against HSPs70 and 90 and subsequently processed with an avidin/biotin/peroxidase staining kit. Endogenous peroxidase activity was blocked by exposure to methanol-hydrogen peroxidase. The specificity of the immunostaining was controlled by omitting the primary antibody, the secondary antibody or the substrate. Cell culture Caco-2 cells were routinely grown in supplemented DMEM (Dulbecco’s modified Eagle’s medium) as described previously (Koninkx et al., 1992). Cells were exposed to lectins in plain DMEM in order to be free from potentially reactive carbohydrates. Cell incubation with 100 µg/ml of lectin was performed for 8, 24, 32, 56, and 80 h. Western blot analysis Proteins separation was by polyacrylamide gel electrophoresis (Laemmli, 1970). Subsequently proteins were transferred to ImmobilonTM-P PVDF (polyvinylidene difluoride) transfer membrane and HSPs were detected using monoclonal antibodies and a peroxidase coupled detection system. Results HSP content of rat gut tissue A semiquantitative evaluation of the staining intensities is shown in Table 1. Compared to the jejunal crypts of LA-fed rats the staining intensity of the intestinal cells in the crypts of PHA-fed and WGA-fed rats was decreased considerably. Also the villi of these gut sections showed a weaker intensity of anti-HSP immunostaining. The greatest reduction in staining intensity was in the villi of rats fed PHA or WGA after immunostaining with anti-HSP90. A decrease in immunostaining intensity was not found when rats were fed heat inactivated (60 min. at 100 oC) lectins (data not shown). HSP content of differentiated Caco-2 cells during lectin exposure The lectins PHA-E4 and WGA were used to determine the effect of lectin exposure on HSP synthesis in differentiated Caco-2 cells. The cellular content of HSP70 and HSP90 is given in Figure 1. The results obtained from control Caco-2 cells clearly showed that constitutive levels of HSPs are present in these enterocyte-like cells.

109
Table 1. The intensity of immunostaining of the jejunal crypts and villi of rats fed on diets containing LA, PHA, or WGA.
HSPs Lectin Crypt staining intensity Villus staining intensity HSP70 LA PHA WGA ++++ ++ + ++++ +++ ++ HSP90 LA PHA WGA ++++ ++ + ++++ ++ + The sections of the jejunum were stained with monoclonal antibodies to HSP70 and HSP90. The intensity of the immunostaining of PHA and WGA fed rats was matched with the staining intensity of the corresponding LA fed rats. Based on this comparison the intensity of the staining was scored using an arbitrary scale (strong staining ++++, moderate staining +++, weak staining ++, very weak staining +).
Generally, the content of HSPs of non-proliferating differentiated Caco-2 cells decreased over time during exposure to 100 µg/ml of PHA-E4 or WGA. The Western blot (Fig. 1) clearly demonstrated an inhibitory effect of PHA-E4 on both HSP70 and HSP90. In contrast, WGA interfered only with the HSP70 content. This lectin has little or no effect on the HSP90 content. Lectin exposure did not inhibit the cellular protein synthesis machinery (data not shown). Incubation of Caco-2 cells with 100 µg of PHA-E4/ml in the presence of 1 mg of fetuin/ml or 100 µg of WGA in the presence of 0.1 M GlcNAc failed to induce the observed changes in HSP synthesis (data not shown).
Figure 1. Inhibition of HSP70 and HSP90 synthesis in differentiated Caco-2 cells during
incubation with lectins. Discussion Cells have a highly conserved set of stress proteins (HSPs), which are involved in coping with chemical or physical damage (Hightower, 1991). Our data clearly demonstrate that lectins, which are known to cause structural lesions in gut epithelial cells on exposure, interfere with the levels of HSPs in these cells. The intensity of the immunostaining in the crypt cells of the jejunum of PHA and WGA-fed rats was dramatically decreased compared with the jejunum of LA-fed rats (Table 1). In differentiated Caco-2 cells the levels of HSPs decreased over time during exposure to lectins (Fig. 1). Lectins are known to interfere with the cytoskeleton of gut cells. Cytoskeletal lesions in rat enterocytes have been observed (Bardocz et al., 1991) and a shift in the ratio of filamentous to globular actin could be measured in differentiated Caco-2 cells (Draaijer et al., 1989). Studies on the function of HSP70 (Majecak & Luftig,

110
1991) and HSP90 (Koyasu et al., 1986) demonstrated the actin-binding activity by these proteins, which stabilise the actin filaments by cross-linking. After lectin exposure, HSPs might be directed to stabilising the cytoskeleton as well as chaperoning of cellular proteins. If the resulting demand for HSPs is not met by adequate synthesis, the level of soluble HSPs decreases.
Considering the evidence linking HSP levels to increased stress resistance it is likely that the lectins PHA-E4 and WGA are not only detrimental to gut intestinal cells by damaging their apical membranes. Depression of the amount of stress proteins in enterocytes by these lectins may leave the cells more vulnerable to exposure to agents produced by the digestive tract, as well as to harmful agents in the diet. Considering that pathogenic bacteria induce stress proteins in intestinal cells, lectin induced downregulation of these proteins may also give increased cellular susceptibility to bacterial invasion. References Bardocz, S., Grant, G., Brown, D., Ewen, S., Stewart, J. & Pusztai, A. 1991. Effect of
fasting and refeeding on basolateral polyamine uptake and metabolism by the rat small bowel. Digestion 50, 28-35.
Draaijer, M., Koninkx, J., Hendriks, H., Kik, M., Dijk J. van, & Mouwen J. 1989. Actin cytoskeletal lesions in differentiated human colon carcinoma Caco-2 cells after exposure to soybean agglutinin. Biology of the Cell 65, 29-35.
Hightower, I. 1991. Heat shock, stress proteins, chaperones and proteotoxicity. Cell 66, 191-197.
Koninkx, J., Hendriks, H., Rossum J. van, Ingh T. van den, Mouwen, J. 1992. Interaction of legume lectins with the cellular metabolism of differentiated Caco-2 cells. Gastroenterology 102, 1516-1523.
Koyasu, S., Nishida, E., Kadowaki, T., Matsuzaki, F., Iida, K., Harada, F., Kasuga, M., Sakai, H. & Yahara, I. 1986. Two mammalian heat shock proteins, Hsp90 and Hsp100, are actin binding proteins. Proceedings of the National Academy of Science USA 83, 8054-9058.
Laemmli, U. 1970. Cleavage of the structural proteins during the assembly of the head of bacteriophage T4. Nature 277, 680-685.
Majecak, D. & Luftig, R. 1991. Stabilisation of actin filaments at early times after adenovirus infection and in heat-shocked cells. Virus Research 19, 31-45.
Pusztai, A., Ewen, S., Grant, G., Brown, D., Stewart, J., Peumans, W., Damme E. van, Bardocz, S. 1993. Antinutritive effects of wheat germ agglutinin and other N-acetylglucosamine-specific lectins. British Journal of Nutrition 70, 313-321.
Pusztai, A., Koninkx, J., Hendriks, H., Kok, W., Hulscher, S., Damme E. van, Peumans, W., Grant, G. & Bardocz, S. 1996. Effect of the insecticidal Galanthus nivalis agglutinin on metabolism and the activities of brush border enzymes in the rat small intestine. Journal of Nutritional Biochemistry 7, 677-682.

111
Physiological mechanisms in the impairment of nutrient digestion and transport capacity in the salmon intestine by soybean antinutrients Å. Krogdahl & A.M. Bakke-McKellep Norwegian School of Veterinary Science, PO Box 8146 Dep., N-0033 Oslo, Norway Summary This presentation summarises the results of three experiments. These experiments demonstrate the negative effects of soybean (Glycine max) meal on various aspects of the digestive functions of the fish (Salmo salar). The main effects were found in the distal intestine and involved luminal digestion, brush border hydrolysis, nutrient transport across the mucosa as well as immunological and morphological processes. Keywords: Salmo salar, soybean, salmonid digestion, nutrient transport, immune functions Introduction The use of soybean meal as an alternative protein source to fishmeal for salmonids in aquaculture has met with limited success. High inclusion levels of full-fat and solvent-extracted soybean meal reduce weight gain and feed efficiency (Olli et al., 1994b; Olli & Krogdahl, 1994; Rumsey et al., 1994a; Olli & Krogdahl, 1995; Refstie et al., 1998), and cause morphological changes in the distal intestinal epithelium (Ingh et al., 1991; Rumsey et al., 1994a,b; Baeverfjord & Krogdahl, 1996; Bakke-McKellep et al., 2000). The precise causative agent of the inflammatory response and/or the reduced digestive function is not known. Some efforts have been made to identify the causal agent. However, its exact nature is still unknown. However, it is known that the compound is alcohol-soluble (Olli & Krogdahl, 1995; Ingh et al., 1996; Bureau et al., 1998). This paper summarizes the results of three experiments with Atlantic salmon (Salmo salar L.), which were conducted to shed light on the physiological mechanisms, which underlie the negative effects of soybean-meal-induced enteritis in salmonids. Experiment 1. Effects of graded levels of standard soybean meal on intestinal structure, mucosal enzyme activities, and pancreatic response in Atlantic salmon (Krogdahl et al., 2004) In this experiment the levels of dietary soybean meal inclusion provided 0, 10, 15, 20, 25 or 35 % of the protein from soybeans. The fish weighed 280 g and were kept in salt water. The results of a comparative slaughter procedure showed that soybean inclusion reduced nutrient utilisation, particularly energy. The effects were mainly due to effects on nutrient digestibility. The effect on energy utilization was apparent even at a 10 % soybean inclusion. Histological examination (Fig. 1) of the distal intestines also showed that their morphology was affected at the 10 % soybean inclusion level. Higher inclusion levels affected the distal mucosa more severely giving characteristics, which are typical in soybean fed salmonids: a widening of the central area (lamina propria) which is infiltrated by a mixed population of inflammatory cells identified as lymphocytes, macrophages, eosinophilic and neutrophilic granular cells, and diffuse IgM (Baeverfjord & Krogdahl 1996; Bakke-McKellep et al., 2000), and concurrent changes in enterocyte structure, including a decrease, or even a loss, of the
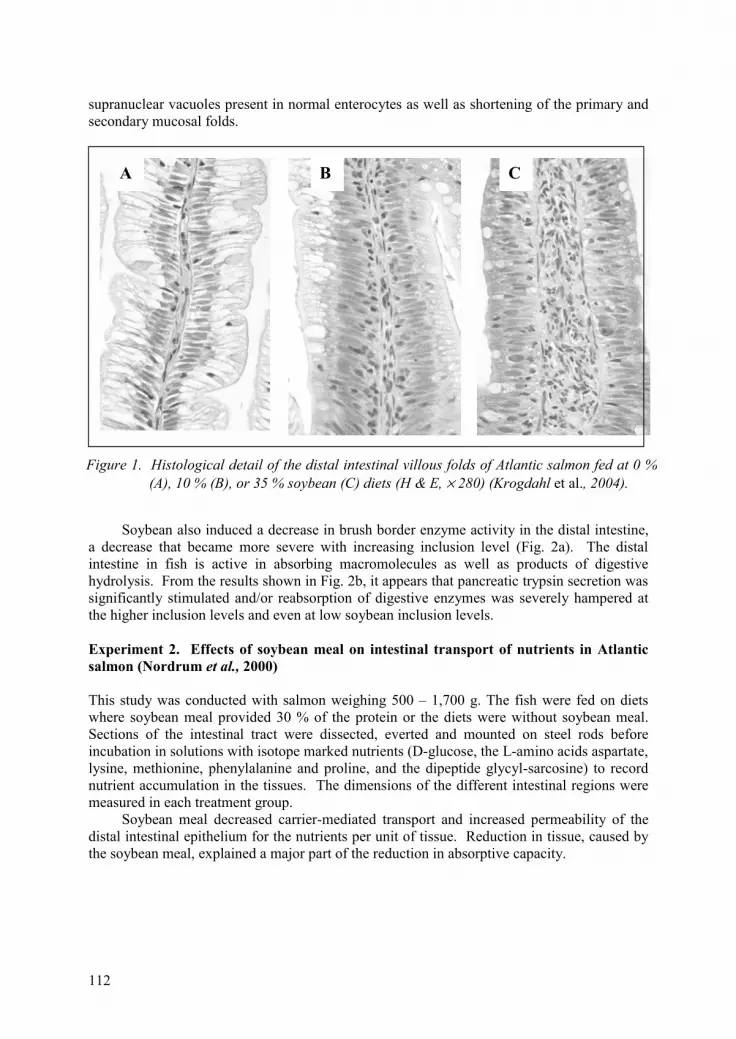
112
supranuclear vacuoles present in normal enterocytes as well as shortening of the primary and secondary mucosal folds.
Soybean also induced a decrease in brush border enzyme activity in the distal intestine, a decrease that became more severe with increasing inclusion level (Fig. 2a). The distal intestine in fish is active in absorbing macromolecules as well as products of digestive hydrolysis. From the results shown in Fig. 2b, it appears that pancreatic trypsin secretion was significantly stimulated and/or reabsorption of digestive enzymes was severely hampered at the higher inclusion levels and even at low soybean inclusion levels. Experiment 2. Effects of soybean meal on intestinal transport of nutrients in Atlantic salmon (Nordrum et al., 2000) This study was conducted with salmon weighing 500 – 1,700 g. The fish were fed on diets where soybean meal provided 30 % of the protein or the diets were without soybean meal. Sections of the intestinal tract were dissected, everted and mounted on steel rods before incubation in solutions with isotope marked nutrients (D-glucose, the L-amino acids aspartate, lysine, methionine, phenylalanine and proline, and the dipeptide glycyl-sarcosine) to record nutrient accumulation in the tissues. The dimensions of the different intestinal regions were measured in each treatment group.
Soybean meal decreased carrier-mediated transport and increased permeability of the distal intestinal epithelium for the nutrients per unit of tissue. Reduction in tissue, caused by the soybean meal, explained a major part of the reduction in absorptive capacity.
Figure 1. Histological detail of the distal intestinal villous folds of Atlantic salmon fed at 0 %(A), 10 % (B), or 35 % soybean (C) diets (H & E, × 280) (Krogdahl et al., 2004).
A B C

113
Figure 2. Effects of graded levels of soybean meal on (A) mucosal enzyme activity (AP =
alkaline phosphatase; LAP = Leucine aminopeptidase) (Units: U/mg protein) and (B) on faecal trypsin activity)(Unit: U/mg DM) (Krogdahl et al., 2004).
Experiment 3. Changes in immune and enzyme histochemical phenotypes of cells in the intestinal mucosa of Atlantic salmon with soybean meal-induced enteritis (Bakke-McKellep et al., 2000) Salmon with an average body weight of 927 g were used in this study. They were fed a fishmeal-based diet (control) or a diet in which 30 % of the fishmeal protein was replaced with soybean meal protein (SBM diet). In salmon fed SBM, there was markedly reduced enzyme reactivity in distal intestinal epithelial cells, both in the brush border (5'-nucleotidase, Mg2+-ATPase, alkaline phosphatase, and leucine aminopeptidase) and the intracellular structures (alkaline and acid phosphatase, non-specific esterase, and alanine aminopeptidase). There appeared to be an increased presence of monocytes, including macrophages, neutrophilic granulocytes and IgM in the lamina propria cells of the SBM-fed fish. The mid intestine showed little response to the diet. The results suggest that the toxic/antigenic component(s) of SBM affect the differentiation of distal intestinal epithelial cells and may help explain the reduced nutrient digestibility, previously reported, in salmonids fed on extracted SBM. Conclusions Standard soybean meal appears to reduce nutrient utilisation in salmonids by affecting the distal part of the intestine in several ways. The reduction in nutrient hydrolysis at the brush border may limit nutrient absorption. Impairment of transport capacity across the intestinal mucosa may add another dimension to the effect of soybean meal on salmonid digestion. Finally, the excessive loss of pancreatic enzymes in the faeces, most likely due to reduced protein reabsorption, may further diminish the capacity for luminal digestion in salmon.
AP LAP Maltase Isomaltase Lactase Sucrase
Enzy
me
activ
ities
0
10
20
30
40
F0F10F15F20F25F35
Soybean inclusion level, % of protein0 5 10 15 20 25 30 35
Faec
al tr
ypsin
, mg
mg-1
faec
es, d
ry m
atte
r
0
2
4
6
8
A B

114
References Baeverfjord, G. & Krogdahl, A. (1996). Development and regression of soybean meal
induced enteritis in Atlantic salmon, Salmo salar L., distal intestine: a comparison with the intestines of fasted fish. Journal of Fish Diseases 19, 375-387.
Bakke-McKellep, A.M., Press, C.M., Baeverfjord, G., Krogdahl, A. & Landsverk, T. (2000). Changes in immune and enzyme histochemical phenotypes of cells in the intestinal mucosa of Atlantic salmon, Salmo salar L., with soybean meal-induced enteritis. . Journal of Fish Diseases 23, 115-127.
Bureau, D.P., Harris, A.M. & Cho, C.Y. (1998). The effects of purified alcohol extracts from soy products on feed intake and growth of Chinook salmon (Oncorhynchus tshawytscha) and rainbow trout (Oncorhynchus mykiss). Aquaculture 161, 27-43.
Ingh, T.S.G.A.M. van den, Krogdahl, Å., Olli, J.J., Hendricks, H.G.C.J.M. & Koninkx, J.F.J.G. (1991). Effects of soybean-containing diets on the proximal and distal intestine in Atlantic salmon (Salmo salar): a morphological study. Aquaculture 94, 297-305.
Ingh, T.S.G.A.M. van den, Olli, J.J. & Krogdahl, Å. (1996). Alcohol-soluble components in soybeans cause morphological changes in the distal intestine of Atlantic salmon, Salmo salar L. Journal of Fish Diseases 19, 47-53.
Nordrum, S., Bakke McKellep, A.M., Krogdahl, Å. & Buddington, R. 2000. Effects of soybean meal and salinity on intestinal transport of nutrients in Atlantic salmon (Salmo salar, L) and rainbow trout (Oncorhynchus mykiss). Comparative Biochemistry and Physiology 125B, 317-335.
Olli, J. & Krogdahl, Å. (1994). Nutritive value of four soybean products as protein sources in diets for rainbow trout (Oncorhynchus mykiss, Walbaum) reared in fresh water. Acta Agricultura Scandinavia, Section A 44, 185-192.
Olli, J. & Krogdahl, Å. (1995). Alcohol soluble components of soybeans seem to reduce lipid digestibility in fish-meal-based diets for Atlantic salmon, Salmo salar L. Aquaculture Research 26, 831-835.
Olli, J., Krogdahl, Å., Ingh, T.S.G.A.M. van den & Brattås, L.E. (1994b). Nutritive value of four soybean products in diets for Atlantic salmon (Salmo salar, L.). Acta Agricultura Scandinavia, Section A 44, 50-60.
Krogdahl, A., Bakke-McKellep, A.M. & Baeverfjord, G. 2004. Effects of graded levels of standard soybean meal on intestinal structure, mucosal enzyme activities, and pancreatic response in Atlantic salmon (Salmo salar L.). Aquaculture Nutrition. (In press).
Refstie, S., Storebakken, T. & Roem, A.J. (1998). Feed consumption and conversion in Atlantic salmon (Salmo salar) fed diets with fish meal, extracted soybean meal or soybean meal with reduced content of oligosaccharides, trypsin inhibitors, lectins and soya antigens. Aquaculture 162, 301-312.
Rumsey, G.L., Endres, J.G., Bowser, P.R., Earnest-Koons, K.A., Anderson, D.P. & Siwicki, A.K. (1994a). Soy protein in diets of rainbow trout: Effects on growth, protein absorption, gastrointestinal histology, and non-specific serologic and immune response. In Nutrition and Utilization Technology in Aquaculture. C. Lim & D.J. Sessa (Eds). AOAC Press, Champaign, Il. pp. 166-188.
Rumsey, G.L., Siwicki, A.K., Anderson, D.P. & Bowser, P.R. (1994b). Effect of soybean protein on serological response, non-specific defense mechanisms, growth, and protein utilization in rainbow trout. Veterinary Immunology and Immunopathology 41, 323-339.

115
Influence of lectins isolated from soybean seed and evening primrose seed and cake on rat metabolism H. Leontowicz1, M. Leontowicz1, J.J. Czerwiński1, H. Kostyra2 & R. Krzemiński1 1Department of Physiological Sciences, Faculty of Veterinary Medicine, Warsaw Agricultural University, Nowoursynowska 159, 02-787 Warsaw, Poland 2Division of Food Science, Institute of Animal Reproduction and Food Research, Polish Academy of Sciences, Tuwima 10, 10-747 Olsztyn, Poland Summary Agglutination of human red blood cells (RBC) by lectins isolated from soybean (Glycine max L.) seed and evening primrose (Oenothera paradoxa, Hudziok L.) seed and cake and their dietary influence on metabolic processes in rats was studied. Lectin extracts agglutinated human RBC in different way and at different times. Lectins offered for 14 d at 0.1 g/d/kg BW reduced trypsin and α-amylase activity in the pancreas, enhanced α-amylase activity in pancreatic juice and improved the plasma lipid profile. Keywords: Glycine max, Oenothera paradoxa, lectins, rat, pancreatic enzyme activity Introduction Plant lectins (Liener, 1994) are mainly proteins or glycoproteins with a specificity for carbohydrate structures (Pusztai, 1993). Most of them can agglutinate in vitro RBC of animals and humans (Jaffe & Seldl, 1992). However some bind to the plasma proteins and thus do not agglutinate RBC (Kelsall et al., 2002).
At high doses, lectins can seriously damage the small intestine (Pusztai, 1996), impair absorption and nutrient transport through the intestinal wall (Bardocz et al., 1995), stimulate intestinal and pancreatic growth (Kelsall et al., 2002), diminish enzyme activity (protease, disaccharidase), disturb lipid absorption in the small intestine and modulate hormone secretion (insulin, CCK, gastrin) (Liener, 1994).
Lectins (and other antinutritional factors (ANFs) are also present in evening primrose seed (Leontowicz et al., 1998b; Kostyra et al., 1999) which is widely used as a source of oil rich in γ–linolenic acid (Kulasek et al., 1997). There is no published data on changes in lectin activity of evening primrose seed after oil extraction.
The aim of this study was to determine the activity of lectin (L), isolated from soybean seed, evening primrose seed or cake, and the effect of these lectins, when fed for 2 weeks in a standard rat diet at 15 mg d/rat on the enzyme activity of the pancreas and pancreatic juice, and on some plasma metabolic parameters. Materials and methods Protein extracts of soybean seed (S) and evening primrose seed (EPS) or cake (EPC) were obtained (Santiago et al., 1989). Lectin activity was estimated by the agglutination test (Liener, 1989) of human RBC on blood groups O, A, B and AB.
An in vivo experiment used male Wistar rats (145 ± 8 g). All experimental rats, were fed ad libitum, with a standard diet (17 % protein) for 14 d. There were 8 control (C) rats and
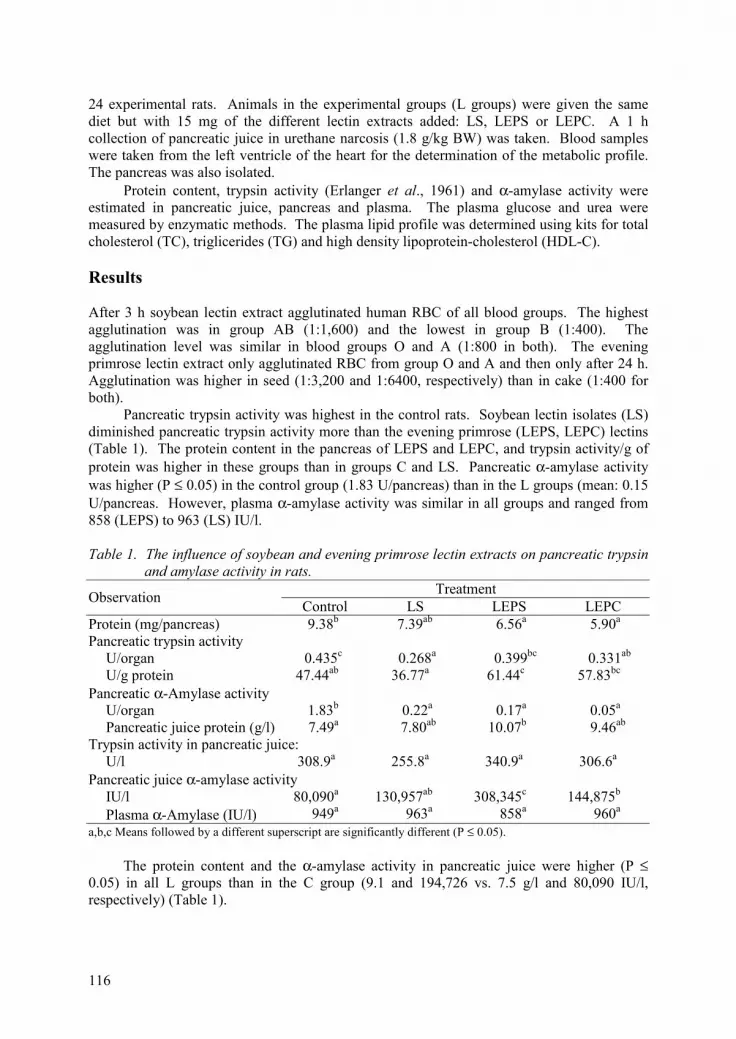
116
24 experimental rats. Animals in the experimental groups (L groups) were given the same diet but with 15 mg of the different lectin extracts added: LS, LEPS or LEPC. A 1 h collection of pancreatic juice in urethane narcosis (1.8 g/kg BW) was taken. Blood samples were taken from the left ventricle of the heart for the determination of the metabolic profile. The pancreas was also isolated.
Protein content, trypsin activity (Erlanger et al., 1961) and α-amylase activity were estimated in pancreatic juice, pancreas and plasma. The plasma glucose and urea were measured by enzymatic methods. The plasma lipid profile was determined using kits for total cholesterol (TC), triglicerides (TG) and high density lipoprotein-cholesterol (HDL-C).
Results After 3 h soybean lectin extract agglutinated human RBC of all blood groups. The highest agglutination was in group AB (1:1,600) and the lowest in group B (1:400). The agglutination level was similar in blood groups O and A (1:800 in both). The evening primrose lectin extract only agglutinated RBC from group O and A and then only after 24 h. Agglutination was higher in seed (1:3,200 and 1:6400, respectively) than in cake (1:400 for both).
Pancreatic trypsin activity was highest in the control rats. Soybean lectin isolates (LS) diminished pancreatic trypsin activity more than the evening primrose (LEPS, LEPC) lectins (Table 1). The protein content in the pancreas of LEPS and LEPC, and trypsin activity/g of protein was higher in these groups than in groups C and LS. Pancreatic α-amylase activity was higher (P ≤ 0.05) in the control group (1.83 U/pancreas) than in the L groups (mean: 0.15 U/pancreas. However, plasma α-amylase activity was similar in all groups and ranged from 858 (LEPS) to 963 (LS) IU/l.
Table 1. The influence of soybean and evening primrose lectin extracts on pancreatic trypsin
and amylase activity in rats. Treatment Observation Control LS LEPS LEPC
Protein (mg/pancreas) 9.38b 7.39ab 6.56a 5.90a Pancreatic trypsin activity
U/organ 0.435c 0.268a 0.399bc 0.331ab U/g protein 47.44ab 36.77a 61.44c 57.83bc
Pancreatic α-Amylase activity U/organ 1.83b 0.22a 0.17a 0.05a Pancreatic juice protein (g/l) 7.49a 7.80ab 10.07b 9.46ab
Trypsin activity in pancreatic juice: U/l 308.9a 255.8a 340.9a 306.6a
Pancreatic juice α-amylase activity IU/l 80,090a 130,957ab 308,345c 144,875b Plasma α-Amylase (IU/l) 949a 963a 858a 960a
a,b,c Means followed by a different superscript are significantly different (P ≤ 0.05).
The protein content and the α-amylase activity in pancreatic juice were higher (P ≤ 0.05) in all L groups than in the C group (9.1 and 194,726 vs. 7.5 g/l and 80,090 IU/l, respectively) (Table 1).

117
Plasma glucose level was similar in all groups and ranged from 10.36 (LEPC) to 12.36 (C) mmol/l. Lectin extracts, especially those from evening primrose, diminished plasma urea concentration to 5.45 (LEPS) and 5.70 (LEPC) compared with 7.19 mmol/l in the C group (P ≤ 0.05). They also enhanced plasma protein concentration (Table 2).
Table 2. The influence of lectin extracts from soybean and evening primrose on some plasma
metabolic parameters. Experimental Observation Control
LS LEPS LEPC Glucose (mmol/l) 12.36a 11.66a 10.80a 10.36a Total protein (g/l) 5.54a 6.07b 6.16b 6.12b Urea, mmol/l 7.19b 7.41b 5.45a 5.70a TC, mmol/l 1.64ab 2.00c 1.50a 1.91bc TG, mmol/l 1.48b 1.04a 0.94a 1.19a HDL-C, mmol/l 0.86a 1.09bc 0.93ab 1.21c TC/HDL 1.91b 1.83ab 1.61a 1.58a a,b,c Means followed by a different superscript are significantly different (P ≤ 0.05).
Lectin extracts modified the plasma lipid profile (P ≤ 0.05) in the L groups compared
with the control rata. The mean HDL-C level was 1.08 vs. 0.86 mmol/l, TG - 1.06 vs. 1.48 mmol/l and the proportion of TC/HDL-C- was 1.67 vs. 1.91 in the L and C groups, respectively. Discussion Significant amount of lectins, which are consumed can reach the circulation in their functional form and bind to blood glycoproteins. Lectin extract from soybean seeds agglutinated human RBC of all blood groups after 3 h. Lectins from evening primrose only agglutinated group A and O cells and then only after a much longer period. Similar effects of lectin agglutination of RBC were observed in soybean (Leontowicz et al., 2001) and from evening primrose cake (Kostyra et al., 1999).
In contrast with this work Kostyra et al. (1999) reported agglutination of blood cells from groups B and AB (the highest). High lectin activity of the lectin from EP seed (and other ANFs) can restrict utilisation of this seed, and its cake, in human and animal nutrition. The soybean and evening primrose lectin extracts at the doses fed did not significantly affect food intake, body weight gain or food efficiency ratio during this 14 d feeding experiment with rats.
All lectin extracts added to standard rat diet (agglutination 0) at 0.1 g/kg BW decreased (P ≤ 0.05) pancreatic trypsin and α-amylase activity expressed as U/organ. Diminished pancreatic trypsin activity was observed after feeding rats a semi-purified diet, which contained 5 or 15 mg of pea lectin (Leontowicz et al., 2000). Grant et al. (1997), in in vitro experiments found, that soybean lectins (also from kidney bean (Phaseolus vulgaris)) could induce the release of α-amylase from pancreatic acini by binding lectins to the surface carbohydrates on the acini. In this work the α-amylase activity of the pancreatic juice was higher (P ≤ 0.05) in all lectin groups. It is possible some lectins may have reached the pancreas and stimulated α-amylase secretion. However, trypsin activity was not significantly changed.
The lectin extracts also changed plasma lipid parameters in. There was an increase in HDL-C but reduced TG and the proportion of TC/HDL-C was observed. These changes were

118
more favourable in the case of the LEPS than with the other lectin extracts. Similar changes in plasma lipid profile after feeding 15 mg LS/d/rat with atherogenic diet (1 % cholesterol) were obtain by Leontowicz et al. (unpublished).
It can be concluded that lectin extracts isolated from oil seeds (soybean), evening primrose seed or cake agglutinated human RBC in different ways and after different times. Lectin extracts fed for 2 weeks at 15 mg/d/rat decreased trypsin and amylase activity in the pancreas and increased α-amylase activity in pancreatic juice. Lectin extracts (mainly from evening primrose seed) improved the plasma lipid profile. References Bardocz, S., Grant, G., Ewen, B., Duguid, B.J., Brown, D.S., Englyst, K. & Pusztai, A.
1995. Reversible effect of phytohaemagglutinin on the growth and metabolism of rat gastrointestinal tract. Gut 37, 353-360.
Erlanger, B.F., Kokowski, N. & Cohen, W. 1961. The preparation end properties of two new chromogenic substrates of trypsin. Archives of Biochemistry and Biophysics 95, 271-278.
Grant, G., Henderson, L.T., Edwards, J.E., Ewan, E.C., Bardocz, S. & Pusztai, A. 1997. Kidney bean and soybean lectins cause enzyme secretion by pancreatic acini in vitro. Life Sciences 60, 1589-1595.
Jaffe W.G. & Seldl, D.S. 1992. Toxicology of plant lectin. In Food Poisoning. Handbook of Natural Toxins. A.T. Tu (Ed.). 7, 264-290.
Kelsall, A., Fitzgerald, A.J., Howard, C.V., Ewans, R.C., Singh, R., Rhodes, J.M. & Goodlad, R.A. 2002. Dietary lectins can stimulate pancreatic growth in the rat. International Journal of Experimental Pathology 83, 203-208.
Kostyra, H., Kostyra, E., Krawczuk, S. & Leontowicz, H. 1999. Influence of extrusion of seed lectins on their haemagglutination properties. Cost 98. European Commission 6, 55-59.
Kulasek G., H. Leontowicz, M. Leontowicz & B. Bałasińska, 1997. Comparison of diet utilisation with raw and extruded primrose cake by rats. 16th International Congress of Nutrition. Montreal, 27 July – 1 August 1997.
Leontowicz, H., Leontowicz, M., Biernat, M. Gralak, M.A., Krzemiński, R. & Czerwiński, J. 2000. The effect of pea lectins on pancreas and jejunum in rats fed semipurified diet. COST 98. European Commission. 9, 111-115.
Leontowicz, H., Leontowicz, M., Kostyra, H., Kulasek, G., Krzemiński, R. & Podgurniak, M. 2001. Effects or raw or extruded legume seeds on some functional and morphological gut parameters in rats. Journal of Animal and Feed Sciences 10, 169-183.
Leontowicz, M, Leontowicz, H., Kostyra, H. Kulasek, G. & Podgurniak, M. 1998. The influence of extrusion on Evening primrose cake on inactivations of antinutritional factors and metabolic response in rats. IIIrd Symposium: Oil from evening primrose seeds and others oils containing acids n-6 or n-3 in prophylactic and therapy, Sulejów, 15 - 16 May 1989, 189-194.
Liener, I.E. 1989. Control of antinutritional and toxic factors in oil seeds and legumes. In Food Uses of Whole Oil and Protein Seeds. W.E.W. Lusas, D.R. Erikson & W. Nip (Eds). American Oil Chemists Association, Champaign Il, 344-371.
Liener, I.E. 1994. Implications of antinutritional components in soybean foods. Critical. Reviews in Food Science and Nutrition 34, 31-67.
Pusztai, A. 1993. Dietary lectins are metabolic signal for the gut and modulate immune and hormone functions. European Journal of Clinical Nutrition 47, 691-699.

119
Pusztai, A. 1996. Chemical probiosis: Blockage by food (plant) lectins of infection of the gut with pathogenic bacteria. In COST 98. Effects of Antinutrients on the Nutritional Value of Legume Diets. S. Bardocz, F.V. Nekrep & A. Pusztai (Eds). European Commission 3, 1-6.
Santiago, J.G., Levy-Benshimol, A. & Carmona, A. (1993). Effect of Phaseolus vulgaris on glucose absorption, transport and metabolism in rat everted intestinal sacs. Journal of Nutritional Biochemistry 4, 426-430.


Session III A Beneficial effects of antinutritional factors in human nutrition


123
Beneficial (antiproliferative) effects of different substances F.M. Lajolo1, M.I. Genovese1, I.F. Pryme2 & T.M. Dale2
1Departamento de Alimentos e Nutrição Experimental, Universidade de São Paulo, Av. Prof. Lineu Prestes, 580, CEP 05508-900, São Paulo, Brazil 2Department of Biochemistry and Molecular Biology, University of Bergen, Jonas Lies vei 91, NO-5009 Bergen, Norway Summary Legume seed contains antinutritional factors (ANFs) such as lectins and trypsin inhibitors that may adversely affect their nutritional properties. However, processing easily inactivates them and chronic ingestion of residual levels is unlikely to pose risks to human health. On the other hand, recent research has shown potential beneficial effects of some of the ANFs. Among them, there is substantial evidence for a role of trypsin inhibitors, phytic acid, saponins and lectins as cancer risk reducing factors. A brief summary of some of these results is presented. Keywords: lectins, trypsin inhibitors, phytic acid, saponins, tumour growth Lectins In 1888 Stillmark reported that castor bean (Ricinus communis) extracts agglutinated red blood cells from different animal species. The seed extracts were found to contain haemagglutinating proteins, defined as agglutinins. Lectins are a very heterogeneous group of proteins of non-immune origin and have gone through several definition steps resulting in less strictness over the years. Some years ago van Damme et al. (1997) defined lectins as: ‘carbohydrate-binding proteins possessing at least one non-catalytic domain, which bind reversibly to a specific mono- or oligosaccharide’. The continuous discovery of new and diverse lectins with similarities found in several different classes of proteins seems to call for even less strict definitions in the future. Maybe the most accurate definition of a lectin would simply be: ‘proteins that specifically bind or cross-bind carbohydrates’.
Classically lectins were divided into 4 groups based on their sugar specificity, such as glucose/mannose, galactose/N-acetylgalactose, N-acetylglucosamine, fucose or sialic acid. Now myriads of plant lectins, animal lectins and even microbiological lectins have been isolated (Prokop et al., 1968; Ashwell & Morell, 1974; Barondes, 1981; Liener et al., 1986; Razin & Mirelman, 1986), and a new and unambiguous classification system has been introduced (van Damme et al., 1997).
Lectins are widespread in the plant kingdom, occurring in seeds and all kinds of vegetative tissues. They often form part of the plant defence system against predators such as insects, worms. In many cases lectins severely interfere with the digestive processes of the insects and worms. Lectins and many other low molecular weight components in foodstuffs are thus called antinutrients because they often have a negative impact on the nutritional value of diets, which contain them. However, most of the antinutritional effects can be eliminated, or substantially reduced, with proper storage, heat treatment, soaking, germinating and cooking. Plant lectins are not always harmful, as biologically active proteins. Some can modify and sometimes improve gut function and body metabolism. They can also alter health

124
status as well. Animal lectins are usually secreted from cells where they may participate in cell-cell-interactions, recognition of immune defence systems, immunoregulation and the prohibition of autoimmunity (Petersen et al., 2001; Wallis, 2002; Holmskov et al., 2003). Due to the homology between plant- and animal-lectins, it is likely that dietary plant lectins can mimic, and to some extent amplify, the effects normally exerted by the lectin counterparts produced by the animal.
Orally administered lectins
Many lectins taken orally are resistant to low pH and proteolytic breakdown due to their carbohydrate shell and their complex structure. They thus remain fully active during their passage through the entire alimentary canal (Pusztai et al., 1990). The surface of the mammalian gut is highly glycosylated and the epithelium is covered by glycolipids and glycoproteins. Through their saccharide binding, lectins bind to epithelial membrane glycoproteins, including hormone receptors, growth factors and cytokines, transport proteins and brush-border membrane enzymes, glycolipids, gangliosides and secreted mucins (Pusztai et al., 1990, 1995). Although lectin binding is most extensive in the small intestine, similar binding occurs throughout the entire digestive system from the oral cavity to the distal colon. The lectin can send messages into the cell via secondary messengers or by the lectin itself entering the cell by endocytosis.
The major effects mediated by lectin binding in the digestive tract are: (a) hyperplasia, increase in crypt cell production rate (Bardócz et al., 1994a, b; Pryme et al., 1998a), (b) overstimulation of secretory cells and exhaustion of their secretory capacity, (c) replacement of appropriate endogenous ligands bound to their receptors, and (d) immunostimulation (Lavelle et al., 2000). Both (a) hyperplasia and (d) immunostimulation are interesting aspects in cancer research.
Oral immunogenicity
The function of the gut associated lymphoid tissue is to absorb minute samples of oral antigens via Peyers patches M-cells and activate the gut immune system. As lectins can bind to M-cells (Sharma et al., 1996), their endocytosis is more extensive, which makes them more powerful immunogens than other antigens. Lectins influence the immune system by evoking local and systemic immune responses (Bardócz et al., 1999). They can induce strong immunogenic reactions when presented both orally or intranasally (Lavelle, 1999; Lavelle et al., 2000). Mitogenic lectins induce lymphocyte proliferation. Several components of the immune system, such as mast cells, basophils, eosinophils and others produce or release cytokines and chemokines (Ebisawa et al., 1997) when exposed to appropriate lectins.
Kidney bean lectin – phytohaemagglutinin (PHA)
Legumes are generally a rich source of lectins. Phytohaemagglutinin (PHA) is the lectin present in the kidney bean (Phaseolus vulgaris). Kidney bean seeds contain large quantities of PHA, which is a complex mixture of isolectins with molecular weights of 118 kDa (Pusztai & Stewart, 1978). All isolectins are tetramers composed of two different subunits, namely an erythroagglutinating E-type subunit, and a mitogenic leucoagglutinating L-type subunit. Five different isolectins are found in most bean varieties - E4, E3L, E2L2, EL3 and L4 (Leavitt et al., 1977). The lectin PHA is only inhibited by complex oligosaccharides (Kaifu & Osawa, 1976; Irimura et al., 1981; Hammarstrom et al., 1982). The sugar specificity for PHA-E is

125
the complex of (Galβ(1,4)GlcNAcβ (1,2)Man) while PHA-L has its specificity for the complex of (Gal β (1,4)GlcNAcβ (1,2)[Gal β (1,4) GlcNAc β (1,6)]Man) (van Damme et al., 1997).
Phytohaemagglutinin binds extremely efficiently to the brush border epithelium of the digestive tract and is partially endocytosed. As mentioned above PHA is known to mediate gut hyperplasia. However, it also has many other biological activities, like the agglutination of human and animal erythrocytes. It is also a potent mitogen, and is capable of producing suppressor cells by reacting with splenic cells. These cells then suppress lymphocyte transformation in mixed human lymphocyte cultures (Sampson et al., 1975). Phytohaemagglutinin reacts with mast cells causing an IgE-independent histamine release and degranulation in vitro (Hook et al., 1974). In fat cell membranes PHA reacts with the insulin receptor, and mimics most of the biological effects of insulin (Pusztai & Watt, 1974). A lipolytic effect has been observed in both rats (Bardócz et al., 1996) and mice (Bardócz et al., 1994a,b), where PHA fed animals showed a major loss of body lipids (Pryme et al., 1996a). As a typical lectin, orally administered PHA is a powerful antinutrient (Pusztai & Bardócz, 1996).
The gut
Based on the difficulties on reducing the polyamine body pool, a novel lectin-approach has been adopted to manipulate the levels of polyamines in the body using the special properties of lectins, and especially PHA. The induction of hyperplastic growth in the gut causes extensive accumulation of polyamines in gut tissues. The aim is to restrict polyamine availability for tumour cells, since a competitor for the growing tumour would reduce the body pool of polyamines, and thus reduce the rate of tumour growth (Pryme et al., 1999a; Pryme & Bardócz, 2001).
The gut mucosa is one of the most active tissues in the body. This is illustrated by the fact that an estimated 40 % of total body protein synthesis per day occurs in this organ (Bardócz et al., 1990). The gut is the first line of defence in the body, and it is therefore crucial to keep the gut structure intact for survival of the species. If gut growth cannot be satisfied by the diet, then the body will inevitably draw on its own reserves to be able to support compensatory gut growth. In the small intestine the villus cells proliferate in the crypts, emerge up the villus while differentiating, and finally the fully differentiated villus cells reach their destination, the villus apex (Bardócz et al., 1995). Here the cells participate in nutrient uptake, and ligand binding. After a transit time of 36 h the cells die due to the strain caused by their enormous activity, and are sloughed off from the gut surface and ultimately emerge in the faeces.
There is a continuous production of villi cells in the crypts and the rate of proliferation is adjusted according to needs. The villi contain rapidly growing cells and therefore require a range of important nutrients. Furthermore, they need a large supply of polyamines to sustain their growth (Bardócz, 1989; Seiler et al., 1996). When feeding on a lectin diet, and especially PHA, lectin-induced hyperplasia further increases demand for extraneous polyamines.
Gut hyperplasia
Hyperplasia is defined as: ‘increased cell production in normal tissue; an excess of normal tissue’. Phytohaemagglutinin affects growth and metabolism of the digestive tract of mice, and therefore seems to be a mitogen for the villus (Pusztai, 1991; Bardócz et al., 1994a, b).

126
Phytohaemagglutinin affects the entire digestive system of mice and rats, inducing a large increase in the weight of the small intestine. The small intestine of LA-fed mice has been shown weigh of 334.4 mg + 54.2 mg, (on average) while its weight in mice fed PHA for 12 d increased to 580.6 mg + 130.3 mg (Pryme et al., 1998a). Although the wet weight of the stomach decreases with PHA feeding, that of the small intestine, caecum and colon increases significantly. Increased turnover of epithelial cells leads to a speeding up of the loss and degradation of villus cells in the lumen. The released lectin is then free to react with the epithelium in more distal parts of the gut, and the physiological stimulus of PHA, therefore, in due course, affects the entire alimentary canal.
In mice given PHA in the diet the structure of the jejunum changes considerably and its morphology becomes characteristic for that of a rapidly growing tissue. Furthermore, the increase in the protein, DNA and RNA content in a 20 cm section of the jejunum has confirmed that the lectin is a growth factor for the small intestine (Bardócz et al., 1994a). Due to the hyperplastic response, the length of the crypts in PHA-fed animals shows a marked elongation, whereas the villi only show slight signs of elongation (Pryme et al., 1998a).
Immunocytochemical visualisation has shown the localisation of PHA in the mouse jejunum. Intense binding of the lectin to the microvillus border of the villi and evidence of endocytosis has also been observed (Ewen et al., 1998).
Phytohaemagglutinin and tumour growth Instead of limiting the supply of dietary polyamines Bardócz, Pryme and their coworkers (Bardócz et al., 1997; Pryme et al., 1999a; Pryme & Bardócz, 2001) used a novel approach to manipulate the body polyamine pools. When PHA was included in the diet (up to 7 mg/g diet), a fully reversible, dose-dependent growth of the small intestine was observed in mice (Bardócz et al., 1994a; Pryme et al., 1996a, b; Pryme et al., 1998a). As shown in Table 1, 12 days after feeding mice a PHA-containing diet (7 mg PHA/g diet), the dry weight of the small intestine had increased by about 230 % while the corresponding tumour weight was reduced by more than 30 %. These results demonstrate clear competition between the hyperplastic growth of the small intestine induced by the lectin and tumour growth.
Table 1. The dry weight (mg) of the small intestine and non-Hodgkin lymphoma tumours
obtained from mice 12 d after subcutaneous injection of 2 x 106 tumour cells when fed on a control diet or a phytohaemagglutinin (PHA) containing diet.
Treatment group Tissue Control fed PHA fed Small intestine 219.2 ± 43.4* 509.0 ± 102.7 Tumour 410.6 ± 198.2 280.5 ± 82.6 *The values represent means (+ SEM) for five animals in each treatment group.
Prior to the onset of hyperplastic growth an accumulation of polyamines occurs in the intestinal epithelia (Bardócz et al., 1990). The levels of putrescine, spermidine and spermine in non-Hodgkin lymphoma (NHL) tumour cells obtained from the control group (non-lectin fed), and mice fed PHA indicated a build up of polyamines in the cells in the latter group. This occurred before the NHL tumour entered a proliferative growth phase (Pryme et al., 1998b). These results show that when the animals are fed PHA, and gut hyperplasia is induced, then tumour growth is slow during the initial stages of epithelial proliferation. There is thus a correlation in time with the polyamine requirement of the gut and the limited tumour growth. It appears that gut cells are more efficient in procuring polyamines from the

127
body pool than the tumour cells, further illustrating the point that the tumour is unable to produce sufficient polyamines by the de novo route to support its growth.
It seems, therefore, that the two different growth signals i.e. the induction of tumour growth and the PHA-induced gut hyperplasia appear to compete with one another for important exogenous nutrients such as the polyamines. The data shows that growth of an NHL tumour was initially slowed as a result of simultaneously occurring gut hyperplasia. Tumour growth, however, was not completely arrested.
Since it has been established that extraneous polyamines from the diet are required to sustain tumour growth, and the hyperplastic growth of the gut that occurs in response to ingesting the lectin is a polyamine-dependent process, it has become apparent that this induction of gut hyperplastic growth can be used to effectively compete with tumour proliferation. Results obtained so far have clearly shown that dietary inclusion of the plant lectin PHA results in a low initial rate of proliferation of NHL tumours in mice (Pryme et al., 1994a, b; 1995; 1996b; 1998a, b; 1999b; 2000). Results showing changes in the weight and tissue polyamine content indicate that interorgan competition between the tumour and vital organs can be used to manipulate the metabolism of tumour-bearing mice. The promising results, obtained with PHA, indicate that lectins, which demonstrate such gut growth-promoting properties, may be extremely valuable with respect to the development of new avenues of anti-cancer strategy. These effects are now known not be limited to PHA alone since mistletoe lectins have recently been shown to have similar biological properties (Pryme et al., 2002). The potential use of lectins as dietary supplements needs further evaluation.
Protease inhibitors
Protease inhibitors are widely distributed in plants, animals and microorganisms. Protease inhibitors in legume seed can have an important impact on their nutritional value as they inhibit pancreatic serine proteases, impairing protein digestion.
Protease inhibitors in legume seeds contain no carbohydrates and belong to two different families, referred to as Kunitz and Bowman-Birk. Kunitz type inhibitors have a molecular weight of about 20 kDa, with two disulphide bridges, and act specifically against trypsin. Bowman-Birk type inhibitors have a molecular weight in the range of 8 to 10 kDa, with seven disulphide bridges, and inhibit trypsin and chymotrypsin simultaneously at independent binding sites. Both protease inhibitors are found in soybean (Glycine max). In common bean (Phaseolus vulgaris), lima bean (P. lunatus), cowpea (Vigna unguiculata) and lentil (Lens culinaris), the protease inhibitors have been characterized as members of the Bowman-Birk family (Lajolo & Genovese, 2002).
The effect of trypsin inhibitors on animal growth is not only a consequence of inhibition intestinal protein digestion, since when inhibitors are present in diets consisting of free amino acids decreased growth was also observed. Kunitz and Bowman-Birk inhibitors cause an enlargement of the pancreas (hypertrophy and hyperplasia) in rodents and birds, and hypersecretion of digestive enzymes. This loss of the sulphur-rich endogenous proteins trypsin and chymotrypsin would cause growth depression, as legume seed proteins are generally deficient in the sulphur amino acids (Lajolo & Genovese, 2002). The mechanism of action of both trypsin inhibitors, Kunitz and Bowman-Birk, is suppression of the negative feedback regulation of pancreatic secretion through increased release of the hormone cholecystokinin from the intestinal mucosa, as reviewed by Liener (1994).
Nutritionally they do not represent a problem since the most common grain legume processing methods, such as cooking at atmospheric pressure and cooking under pressure, are efficient in inactivating most or even all of the inhibitory protease activity. Due to their being

128
proteins, trypsin inhibitors are inactivated under conditions that lead to irreversible protein denaturation. As all human food legumes are heated before consumption by humans, it can be expected that these ANFs will be destroyed (Liener, 1994).
On the other hand, Bowman-Birk inhibitor (BBI) has been shown to be effective in preventing or suppressing carcinogen-induced transformation in vitro and carcinogenesis in animal carcinogenesis assays, and achieved Investigational New Drug status from the FDA in 1992 (Kennedy, 1995, 1998a, b). The BBI showed inhibited the growth and survival of human prostate cancer cells (Kennedy & Wan, 2002). It also reduced the incidence and frequency of colon tumours in dimethylhydrazine-treated rats. This effect was not observed with autoclaved BBI, suggesting that protease inhibitor activity was necessary for anticarcinogenic activity (Kennedy et al., 2002). Recently, studies with humans showed no toxic effects of BBI (Wan et al., 1999; Armstrong et al., 2000). Bowman-Birk inhibitor was effective when administered to animals by different routes, including the diet. The amount of residual Bowman-Birk inhibitor in autoclaved soybeans would be enough to suppress carcinogenesis. Half of an oral dose given to animals was absorbed, distributed throughout the body and excreted in the urine (Kennedy, 1998a). Urinary excretion of BBI in humans after soy consumption has recently been demonstrated (Wan et al., 2000). According to Clawson (1996), the effect of protease inhibitors would be indirect and the most attractive hypothesis would be that dietary protease inhibitors induce synthesis and distribution of endogenous protease inhibitors, which have widespread effects on cell growth and behaviour.
Phytic acid
Phytic acid (myo-inositol hexakisphosphate, IP6) is abundant in cereal and legume seed, contributing about 1 to 7 % of their dry weight. It acts as a phosphate storage depot (Zou & Erdman, 1995). Phytic acid (or its salts, the phytates) represents, on average, 70 % of the total phosphorus content of dry beans, which have a phytic acid concentration of 0.3 to 0.6 % (Lajolo et al., 1996). In the pH range of 0.5 - 9.0 it presents a sterically stable form with one phosphate at carbon position 2 in the axial position and five phosphates in the equatorial position. At pH values over 9.5 it adopts a sterically hindered conformation with five phosphates in the axial position and one phosphate in the equatorial position (Shamsuddin, 2002). The anionic nature of the six phosphate groups imparts a high cation binding property to the molecule, and the low solubility of some of the salts formed gives poor absorption from the gastrointestinal tract. Thus, consumption of foods rich in phytates has been associated with a reduced mineral bioavailability. Processing of foods leads to hydrolysis to tri, tetra, and penta inositol phosphate, which have a lower mineral chelating activity (Harland & Narula, 1999). Phytic acid also decreased in vitro protein digestibility. This was associated with the formation of macrocomplexes through interaction with positively charged groups (like ε-NH3 of lysine) in the proteins. It also affects starch digestion and the glycemic index. This effect was the result of a direct interaction with starch, or with enzymes involved in starch digestion, or the chelation of the ion calcium required for α-amylase activity (Lajolo et al., 1996).
In vivo, phytic acid (IP6) is rapidly absorbed by rats and inositol and IP1-6 were present in gastric epithelial cells after 1 h of IP6 intragastric administration. The IP6 metabolites, inositol and IP1, are transported via plasma, reaching distant organs, and are eliminated in the urine. In humans, unmetabolised IP6 was excreted in the urine in doses, which corresponded to 1 - 3 % of the total administered. Being a highly charged molecule, IP6 seems to be transported across cell membranes by binding proteins such as clathrin adaptor complex AP2, AP180, and coatomer of COP I coat (Shamsuddin et al., 1997; Shamsuddin 1999).

129
Phytic acid and its lower phosphorylated forms, IP 1-5, are present in most mammalian cells at much lower concentrations than in plants (15 - 100 µM range), and are important for regulation of vital cellular functions. Inositol is the precursor of phospholipids and inositol phosphates in cells and has a regulatory effect on the biosynthetic enzymes involved. In animal cells, IP3 is central in cellular signal transduction and intracellular function. Its release from phosphatidylinositol bisphosphate, present in the plasma membrane, causes its rapid diffusion through the cytosol to bind to IP3-gated Ca2+-release channels in the membrane of the endoplasmic reticulum and release Ca2+. This causes its rapid dephosphorylation and consequent inactivation, although a part is further phosphorylated to IP4 which inhibits IP3 and promotes refilling of the intracellular Ca2+ store from the extracellular fluid, acting as an ‘off-switch’ which maintains calcium homeostasis (Shamsuddin et al., 1997; Harland & Narula, 1999).
Although there are no studies with humans, a reasonable amount of evidence points to the anticarcinogenic activity of phytic acid. According to a review by Fox & Eberl (2002) of the literature from 1966 to 2002 (Medline search selecting 28 studies), a large number of animal studies have shown that phytic acid exhibits anti-neoplastic properties in breast, liver, colon, prostate, sarcomas, leukaemia and skin cancer. The proposed mechanisms of action, not yet elucidated, include antioxidant activity (inhibition of iron mediated oxidative reactions and also tumour progression. Magnesium and zinc are critical for cell proliferation - by metal chelation), enhanced immunity (increase in natural killer [NK] cell activity and the antagonism of fibroblast growth factors), and altered gene expression, causing greater differentiation of malignant cells and even a complete reversion to normal phenotypes. The IP6 would interfere with signal transduction by blocking phosphatidyl inositol 3 kinase, stimulate p53 and p21 WAF1/Cip1 tumour suppressor genes, and decrease mitosis by arresting proliferation in the G0/G1 phase. An increase in papilloma formation was observed as a side effect of the sodium salt of phytic acid but not of the potassium or magnesium salts.
Many of the studies were performed in vitro with cancer cell lines. They almost instantaneously absorbed IP6 and the rate and pattern of IP6 metabolism varied depending on cell type. The IP6 treatment was cytostatic, not cytotoxic. Intracellular inositols accumulated mostly in the cytosol as inositol and IP 1-6 (Shamsuddin, 1999). Inositol is also an anti-neoplastic agent but to a lesser extent than IP6. It can act synergistically with it (Shamsuddin, 2002).
In rats, 1 % phytic acid in the diet greatly increased (more than 100-fold) its physiological levels in brain and plasma, without affecting inositol triphosphate levels. In malignant cells treated with phytic acid, the intracellular concentration of inositol triphosphate doubled without changes in the phytic acid level (Grases et al., 2002).
In cultured cells treated with H2O2 and Cu(II), phytic acid (but not myo-inositol) inhibited the formation of 8-oxo-7,8-dihydro-2'-deoxyguanosine but did not scavenge H2O2, indicating that it acts as an antioxidant inhibiting the generation of reactive oxygen species from H2O2 by chelating metals (Midorikawa et al., 2001). This was also the mechanism suggested for the decrease in iron-mediated colon cancer risk (Zhou & Erdman, 1995). Ferry et al. (2002), on the other hand, reported that HeLa cells treated with phytic acid induced apoptosis by inhibiting the Akt-nuclear factor kappaB pathway (a cell surviving signal) and also by causing mitochondrial permeabilization, followed by cytochrome c release and activation of the apoptotic machinery, caspase 9, caspase 3 and poly (ADP-ribose) polymerase.
Phytic acid significantly decreased the number of S-phase cells in human breast and colon cancer cell lines, arresting cells in the G0/G1-phase, and decreased Ki-67 and PCNA expression levels, which act as proliferation markers (El-Sherbiny et al., 2001).

130
In advanced human prostate carcinoma cells, phytic acid gave significant dose and time-dependent growth inhibition associated with G1 arrest and apoptosis induction. It was shown to modulate the cyclin-dependent kinase (CDK) inhibitors–CDK–cyclin complex and decrease CDK-cyclin kinase activity and, at higher doses, to activate the caspase cascade (Singh et al., 2003).
Phytic acid was also anti-proliferative with human leukaemic cell lines via down-modulation of genes involved in transcription and cell cycle regulation and upregulation of cell cycle inhibitors (Deliliers et al., 2002).
Although all these studies were conducted with IP6, it is not generally accepted as the biological effector. There are some indications that the physiologically active metabolites are the diphosphorylated inositol phosphates produced by further phosphorylation of IP6 (Shears, 2001). Contrary to this, there is a hypothesis that the anti-neoplastic activity of IP6 is mediated via lower IPs (Shamsuddin, 2002).
There are no studies, in humans or animals, showing the possibility of anticarcinogenic effects resulting from the ingestion of IP6 which is naturally present in the diet, e.g. in soy or dry beans. Future studies should consider the presence of lectins and isoflavones along with IP6 as biologically active agents and their potential for synergistic action.
Saponins
Saponins are amphiphilic compounds present in foods of plant origin, particularly in legume seed. Besides their cholesterol-lowering effect, soy saponins have been reported to inhibit tumour development both in vivo and in vitro (Rao & Sung, 1995). In human colon cancer cells, soy saponins decreased cell growth in a concentration-dependent manner, suppressing protein kinase C activation and inducing differentiation, although they did not induce apoptosis (Oh & Sung, 2001). Soyasapogenol B glycosides presented anti-invasive activity and induced apoptosis in SNB19 human glioblastoma cells, probably by stimulating cytochrome-c release and subsequent activation of a caspase cascade (Yanamandra et al., 2003). Soyasapogenol B also showed a growth inhibitory effect in human breast cancer cells (Rowlands et al., 2002) and antigenotoxic activity against 2-acetoxyacetylaminofluorene-induced DNA damage in Chinese hamster ovary cells (Berhow et al., 2000). Soyasaponin I was also reported to be a potent and specific inhibitor of sialyltransferase activity (Wu et al., 2001).
Dietary intake of soysaponins also reduced azoxymethane-induced preneoplastic lesions in the colon of mice, without showing any adverse effects on their growth and overall health (Koratkar & Rao, 1997). Other substances with promising anticarcinogenic properties are tannins (Santos-Buelga & Scalbert, 2000), polyphenolic compounds that are naturally present in legume seeds such as pea (Pisum sativum), lentil and bean. The tannin content of dry beans (Phaseolus vulgaris) varies according to seed coat colour, where the tannins are concentrated. During cooking these thermostable water-soluble substances migrate into both the cooking water and the cotyledons, where they interact with storage proteins, mainly phaseolin, decreasing its digestibility. Tannins also form complexes with digestive enzymes and impair dietary protein and carbohydrate digestion (Lajolo et al., 1996). A high amount of tannins (27 % of DM) has been shown to be strongly associated with dietary fibre and is thus partially responsible for its beneficial effects (Saura-Calixto, 1998).

131
Conclusions There is increasing evidence, which strongly suggesting that diet and nutrients play a causative role in the development of many disease states, including cancer. Some data has indicated that overnutrition can significantly promote the development of certain cancers, including those of the colon, pancreas, kidney, breast, ovary endometrium and prostate. It is important, however, that nutrition should not be regarded as a sole means of cancer treatment/prevention but rather as a necessary component to be taken into consideration when planning an anti-cancer strategy. Individual dietary factors may be useful in the design of active methods of preventing the development of disease or in the active management of tumour growth. It seems likely that legume ANFs may have an important future role as dietary supplements in disease management. References Armstrong, W.B., Kennedy, A.R., Wan, X.S., Atiba, J., McLaren, E. & Meyskens, F.L.
2000. Single-dose administration of Bowman-Birk inhibitor concentrate in patients with oral leukoplakia. Cancer Epidemiology Biomarkers & Prevention 9, 43-47.
Ashwell, G. & Morell, A.G. 1974. The role of surface carbohydrates in the hepatic recognition and transport of circulating glycoproteins. Advances in Enzymology & Related Areas of Molecular Biology 41, 99-128.
Bardócz, S. 1989. Polyamines in tissue regeneration. In Physiology of Polyamines, Volume 1. U. Bachrach & Y.M. Heimer (Eds.). CRC Press, Boca Raton, Fl. pp. 96-106.
Bardócz, S., Brown, D.S., Grant, G. & Pusztai, A. 1990. Luminal and basolateral polyamine uptake by rat small intestine stimulated to grow by Phaseolus vulgaris lectin phytohaemagglutinin in vivo. Biochimica Biophysica Acta 1034, 46-52.
Bardócz, S., Ewen, S.W.B., Grant, G. & Pusztai, A. 1995. Lectins: Biomedical aspects. A Pusztai & S. Bardócz (Eds.). Taylor & Francis, London. pp. 103-116.
Bardócz, S., Grant, G., Duguid, T.J., Brown, D.S., Pusztai, A. & Pryme, I.F. 1994b. Effect of phytohaemagglutinin on the growth of Krebs II tumour cells, body metabolism and internal organs of mice. International Journal of Oncology 5, 1369-1374.
Bardócz, S., Grant, G., Duguid, T.J., Brown, D.S., Pusztai, A. & Pryme, I.F. 1997. Intracellular levels of polyamines in Krebs II lymphosarcoma cells in mice fed phytohaemagglutinin-containing diets are coupled with altered tumour growth. Cancer Letters 121, 25-29.
Bardócz, S., Grant, G., Duguid, T.J., Brown, D.S., Sakhri, M., Pusztai, A., Pryme, I.F., Mayer, D. & Way�, K. 1994a. Phytohaemagglutinin in the diet induces growth of the gut and modifies some organ weights in mice. Medical Science Research 22, 101-103.
Bardócz, S., Grant, G. & Pusztai, A. 1996. The effect of phytohaemagglutinin at different dietary concentrations on the growth, body composition and plasma insulin of the rat. British Journal of Nutrition 76, 613-626.
Bardócz, S., Grant, G., Pusztai, A. & Ewen, S.W. B. 1999. Lectins as gut growth factors - a summary. In COST 98, Effects of antinutrients on the nutritional value of legume diets. S. Bardócz, G. Hajós & A. Pusztai (Eds.). EU, Luxembourg, 6. pp. 61-64.
Barondes, S.H. 1981. Lectins: their multiple endogenous cellular functions. Annual Review of Biochemistry 50, 207-231.
Berhow, M.A., Wagner, E.D., Vaughn, S.F. & Plewa, M.J. 2000. Characterization and antimutagenic activity of soybean saponins. Mutation Research-Fundamental and Molecular Mechanisms of Mutagenesis 448, 11-22.

132
Clawson, G.A. 1996. Protease inhibitors and carcinogenesis: a review. Cancer Investigation 14, 597-608.
Deliliers, G.L., Servida, F., Fracchiolla, N.S., Ricci, C., Borsotti, C., Colombo, G. & Soligo, D. 2002. Effect of inositol hexaphosphate (IP(6)) on human normal and leukaemic haematopoietic cells. British Journal of Haematology 117, 577-587.
Ebisawa, M., Tachimoto, H., Iikura, Y., Akiyami, K. & Saito, H. 1997. Progress in Allergy and Clinical Immunology, Volume 4. Hogrefe & Huber Publishers, Seattle.
El-Sherbiny, Y.M., Cox, M.C., Ismail, C.A., Shamsuddin, A.M. & Vucenik, I. 2001. G0/G1 arrest and S phase inhibition of human cancer cell lines by inositol hexaphosphate (IP6). Anticancer Research 21 (4A), 2393-2403.
Ewen, S.W.B., Bardócz, S., Grant, G., Pryme, I.F. & Pusztai, A. 1998. The effect of PHA and mistletoe lectin binding to epithelium of rat and mouse gut. In COST 98, Effects of antinutrients on the nutritional value of legume diets. S. Bardócz, U. Pfüller & A. Pusztai (Eds.). EU, Luxembourg, 5. pp. 221-225.
Ferry, S., Matsuda, M., Yoshida, H. & Hirata, M. 2002. Inositol hexakisphosphate blocks tumor cell growth by activating apoptotic machinery as well as by inhibiting the Akt/NF kappa B-mediated cell survival pathway. Carcinogenesis 23, 2031-2041.
Fox, C.H. & Eberl, M. 2002. Phytic acid (IP6), novel broad spectrum anti-neoplastic agent: a systematic review. Complementary Therapies in Medicine 10, 229-234.
Grases, F., Simonet, B.M., Vucenik, I., Perello, J., Prieto, R.M. & Shamsuddin, A.M. 2002. Effects of exogenous inositol hexakisphosphate (InsP (6)) on the levels of InsP (6) and of inositol trisphosphate (InsP (3)) in malignant cells, tissues and biological fluids. Life Science 71, 1535-1546.
Hammarstrom, S., Hammarstrom, M.L., Sundblad, G., Arnarp, J. & Lonngren, J. 1982. Mitogenic leukoagglutinin from Phaseolus vulgaris binds to a pentasaccharide unit in N-acetyllactosamine-type glycoprotein glycans. Proceedings National Academy of Science USA 79, 1611-1615.
Harland, B.F. & Narula, G. 1999. Food phytate and its hydrolysis products. Nutrition Research 19, 947-961.
Holmskov, U., Thiel, S. & Jensenius, J.C. 2003. Collectins and ficolins: Humoral lectins of the innate immune defense. Annual Reviews of Immunology 21, 547-578.
Hook, W.A., Dougherty, S.F. & Oppenheim, J.J. 1974. Release of histamine from hamster mast cells by concanavalin A and phytohemagglutinin. Infection and Immunity 9, 903-908.
Irimura, T., Tsuji, T., Tagami, S., Yamamoto, K. & Osawa, T. 1981. Structure of a complex-type sugar chain of human glycophorin A. Biochemistry 20, 560-566.
Kaifu, R. & Osawa, T. 1976. Synthesis of O-beta-D-galactopyranosyl-(1 linked to 4)-O-(2acetamido-2-deoxy-beta-D-glucopyranosyl)-(1 linked to 2)-D-mannose and its interaction with various lectins. Carbohydrate Research 52, 179-185.
Kennedy, A.R. 1998a. The Bowman-Birk inhibitor from soybeans as an anticarcinogenic agent. American Journal of Clinical Nutrition 68, 1406S-1412S.
Kennedy, A.R. 1998b. Chemopreventive agents: protease inhibitors. Pharmacology & Therapeutics 78, 167-209.
Kennedy, A.R. 1995. The evidence for soybean products as cancer preventive agents. Journal of Nutrition 125, 733S-743S.
Kennedy, A.R., Billings, P.C., Wan, X.S. & Newberne, P.M. 2002. Effects of Bowman-Birk inhibitor on rat colon carcinogenesis. Nutrition and Cancer 43, 174-186.

133
Kennedy, A.R. & Wan, X.S. 2002. Effects of the Bowman-Birk inhibitor on growth, invasion, and clonogenic survival of human prostate epithelial cells and prostate cancer cells. Prostate 50, 125-133.
Koratkar, R. & Rao, A.V. 1997. Effect of soya bean saponins on azoxymethane-induced preneoplastic lesions in the colon of mice. Nutrition and Cancer 27, 206-209.
Lajolo, F.M. & Genovese, M.I. 2002. Nutritional significance of lectins and enzyme inhibitors from legumes. Journal of Agricultural and Food Chemistry 50, 6592-6598.
Lajolo, F.M., Genovese, M.I. & Menezes, E.W. 1996. Qualidade nutricional do feijão. In Cultura do Feijoeiro Comum no Brasil. R.S. Araújo, C.A. Rava, L.F. Stone & M.J. de O Zimmermann (Eds). Potafos, Piracicaba. pp. 23-56.
Lavelle, E.C. 1999. Lectins in oral vaccination. In COST 98, Effects of antinutrients on the nutritional value of legume diets. S. Bardócz, G. Hajós, & A. Pusztai (Eds.). EU. Luxembourg, 6. pp. 148-159.
Lavelle, E.C., Grant, G., Pusztai, A., Pfüller, U. & O'Hagan, D.T. 2000. Mucosal immunogenicity of plant lectins in mice. Immunology 99, 30-37.
Leavitt, R.D., Felsted, R.L. & Bachur, N.R. 1977. Biological and biochemical properties of Phaseolus vulgaris isolectins. Journal of Biological Chemistry 252, 2961-2966.
Liener, I.E. 1994. Implications of antinutritional components in soybean foods. Critical Reviews of Food Science & Nutrition 34, 31-67.
Liener, I.E., Sharon, N. & Goldstein, I.J. 1986. The Lectins, Properties, Functions and Application in Biology and Medicine. Academic Press, New York.
Midorikawa, K., Murata, M., Oikawa, S., Hiraku, Y. & Kawanishi, S. 2001. Protective effect of phytic acid on oxidative damage with reference to cancer chemoprevention. Biochemical and Biophysical Research Communications 288, 552-557.
Oh, Y.J. & Sung, M.K. 2001. Soybeans saponins inhibit cell proliferation by suppressing PKC activation and induce differentiation of HT-29 human colon adenocarcinoma cells. Nutrition and Cancer 39, 132-138.
Petersen, S.V., Thiel, S., Jensen, L., Steffensen, R. & Jensenius, J.C. 2001. An assay for the mannan-binding lectin pathway of complement activation. Journal of Immunological Methods 257, 107-116.
Prokop, O., Uhlenbruck, G. & Kohler, W. 1968. A new source of antibody-like substances having anti-blood group specificity. A discussion on the specificity of Helix agglutinins. Vox Sanguinis 14, 321-333.
Pryme, I.F. & Bardócz, S. 2001. Anti-cancer therapy: diversion of polyamines in the gut. European Journal of Gastroenterology & Hepatology 13, 1-6.
Pryme, I.F., Bardócz, S., Grant, G., Duguid, T.J., Brown, D.S. & Pusztai, A. 1995. Switching between control and phytohaemagglutinin-containing diets affects growth of Krebs II ascites cells and produces differences in the levels of putrescine, spermidine and spermine. Cancer Letters 93, 233-237.
Pryme, I.F., Bardócz, S., Grant, G., Duguid, T.J., Brown, D.S. & Pusztai, A. 1998b. The effect of PHA on the growth and polyamine content of Krebs II non-Hodgkin lymphoma tumour cells in mice. In COST 917, Biogenically active amines in food. S. Bardócz, A. White & G. Hajós (Eds). EU, Luxembourg, 2. pp. 123-129.
Pryme, I.F., Bardócz, S., Pusztai, A. & Ewen, S.W.B. 2000. The extent of PHA-induced gut hyperplasia is reduced by a developing tumour. In COST 98, Effects of Antinutrients on the Nutritional Value of Legume Diets. S. Bardócz, E. Gelencsér & A. Pusztai (Eds). EU, Luxembourg, 8. pp. 65-69.

134
Pryme, I.F., Bardócz, S., Pusztai, A. & Ewen, S.W.B. 2002. Dietary mistletoe lectin supplementation and reduced growth of a murine non-Hodgkin lymphoma. Histology & Histopathology 17, 261-271.
Pryme, I.F., Grant, G., Pusztai, A. & Bardócz, S. 1999a. Limiting the availability of polyamines for a developing tumour. In Polyamines in Health and Nutrition. S. Bardócz & A. White (Eds). Kluwer Academic Publishers, Norwell, Mass. pp. 283-291.
Pryme, I.F., Pusztai, A. & Bardócz, S. 1994a. A diet containing the lectin phytohaemagglutinin (PHA) slows down the proliferation of Krebs II cell tumours in mice. Cancer Letters 76, 133-137.
Pryme, I.F., Pusztai, A. & Bardócz, S. 1994b. The initial growth rate of Krebs II ascites cell tumours in mice is slowed down by the inclusion of phytohaemagglutinin in the diet. International Journal of Oncology 5, 1105 - 1107.
Pryme, I.F., Pusztai, A., Bardócz, S. & Ewen, S.W.B. 1998a. The induction of gut hyperplasia by phytohaemagglutinin in the diet and limitation of tumour growth. Histology & Histopathology 13, 575-583.
Pryme, I.F., Pusztai, A., Bardócz, S. & Ewen, S.W.B. 1999b. A combination of dietary protein depletion and PHA-induced gut growth reduce the mass of a murine non-Hodgkin lymphoma. Cancer Letters 139, 145-152.
Pryme, I.F., Pusztai, A., Grant, G. & Bardócz, S. 1996a. The effect of switching between a phytohemagglutinin-containing and a control diet on the growth and lipid content of a Krebs II lymphosarcoma tumor. Journal of Experimental Therapeutics & Oncology 1, 273-277.
Pryme, I. F., Pusztai, A., Grant, G. & Bardócz, S. 1996b. Phytohemagglutinin-induced gut hyperplasia and the growth of a mouse lymphosarcoma tumor. Journal of Experimental Therapeutics & Oncology 1, 171-176.
Pusztai, A. 1991. Plant lectins. Cambridge University Press, Cambridge. Pusztai, A. & Bardócz, S. 1996. Biological effects of plant lectins on the gastrointestinal
tract: metabolic consequences and applications. Trends in Glycoscience and Glycotechnology 8, 149-165.
Pusztai, A., Ewen, S.W.B., Grant, G., Peumans, W.J., Damme, E.J. van, Coates, M.E. & Bardócz, S. 1995. Lectins and also bacteria modify the glycosylation of gut surface receptors in the rat. Glycoconjugate Journal 12, 22-35.
Pusztai, A., Ewen, S.W.B, Grant, G., Peumans, W. J., Damme, E.J. van, Rubio, L. & Bardócz, S. 1990. Relationship between survival and binding of plant lectins during small intestinal passage and their effectiveness as growth factors. Digestion 46, 308-316.
Pusztai, A. & Stewart, J.C. 1978. Isolectins of Phaseolus vulgaris. Physicochemical studies. Biochimica Biophysica Acta 536, 38-49.
Pusztai, A. & Watt, W.B. 1974. Isolectins of Phaseolus vulgaris. A comprehensive study of fractionation. Biochimica Biophysica Acta 365, 57-71.
Rao, A.V. & Sung, M.K. 1995. Saponins as anticarcinogens. Journal of Nutrition 125, S717-S724.
Razin, S. & Mirelman, D. 1986. Microbial Lectins and Agglutinins, Wiley & Sons, New York.
Rowlands, J.C., Berhow, M.A. & Badger, T.M. 2002. Estrogenic and antiproliferative properties of soy sapogenols in human breast cancer cells in vitro. Food and Chemical Toxicology 40, 1767-1774.
Sampson, D., Grotelueschen, C. & Kauffman, H.M. 1975. The human splenic suppressor cell. Transplantation 20, 362-367.

135
Santos-Buelga, C. & Scalbert, A. 2000. Proanthocyanidins and tannin-like compounds – nature, occurrence, dietary intake and effects on nutrition and health. Journal of the Science of Food and Agriculture 80, 1094-1117.
Saura-Calixto, F. 1998. Antioxidant dietary fibre product: A new concept and a potential food ingredient. Journal of Agricultural and Food Chemistry 46, 4303-4306.
Seiler, N., Delcros, J.G. & Moulinoux, J.P. 1996. Polyamine transport in mammalian cells. An update. International Journal of Biochemistry & Cell Biology 28, 843-861.
Shamsuddin, A.M. 1999. Metabolism and cellular functions of IP6: a review. Anticancer Research 19 (5A), 3733-3736.
Shamsuddin, A.M. 2002. Anti-cancer function of phytic acid. International Journal of Food Science and Technology 37, 769-782.
Shamsuddin, A.M., Vucenik, I. & Cole, K.E. 1997. IP6: a novel anti-cancer agent. Life Sciences 61, 343-354.
Sharma, R., Damme, E.J. van, Peumans, W.J., Sarsfield, P. & Schumacher, U. 1996. Lectin binding reveals divergent carbohydrate expression in human and mouse Peyer's patches. Histochemistry & Cell Biology 105, 459-465.
Shears, S.B. 2001. Assessing the omnipotence of inositol hexakisphosphate. Cellular Signalling 13, 151-158.
Singh, R.P., Agarwal, C. & Agarwal, R. 2003. Inositol hexaphosphate inhibits growth, and induces G1 arrest and apoptotic death of prostate carcinoma DU145 cells: modulation of CDKI-CDK-cyclin and pRb-related protein-E2F complexes. Carcinogenesis 24, 555-563.
Stillmark, H. 1888. Über Ricin, ein giftiges Ferment aus dem Samen von Ricinus comm. und einigen anderen Euphorbiaceen. Thesis, University of Dorpat, Dorpat (Turtu).
Van Damme, E.J.M., Peumans, W.J., Pusztai, A. & Bardócz, S. 1997. Handbook of Plant Lectins: Properties and Biomedical Applications. Wiley & Sons, Chichester.
Wallis, R. 2002. Structural and functional aspects of complement activation by mannose-binding protein. Immunobiology 205, 433-445.
Wan, X.S., Lu, L.J.W., Anderson, K.E., Ware, J.H. & Kennedy, A.R. 2000. Urinary excretion of Bowman-Birk inhibitor in humans after soy consumption as determined by a monoclonal antibody-based immunoassay. Cancer Epidemiology Biomarkers & Prevention 9, 741-747.
Wan, X.S., Meyskens, F.L., Armstrong, W.B., Taylor, T.H. & Kennedy, A.R. 1999. Relationship between protease activity & neu oncogene expression in patients with oral leukoplakia treated with the Bowman-Birk inhibitor. Cancer Epidemiology Biomarkers & Prevention 8, 601-608.
Wu, C.Y., Hus, C.C., Chen, S.T. & Tsai, Y.C. 2001. Soyasaponin I, a potent and specific sialyltransferase inhibitor. Biochemical and Biophysical Research Communications 284, 466-469.
Yanamandra, N., Berhow, M.A., Konduri, S., Dinh, D.H., Olivero, W.C., Nicolson, G.L. & Rao, J.S. 2003. Triterpenoids from Glycine max decrease invasiveness and induce caspase-mediated cell death in human SNB19 glioma cells. Clinical & Experimental Metastasis 20, 375-383.
Zhou, J.R. & Erdman, J.W. 1995. Phytic acid in health and disease. Critical Reviews in Food Science and Nutrition 35, 495-508.


137
Investigation of legume seed protease inhibitors as potential anti-carcinogenic proteins A. Clemente1, D.A. MacKenzie2, I.T. Johnson3 & C. Domoney1 1John Innes Centre, Department of Metabolic Biology, Norwich Research Park, Norwich NR4 7UH, United Kingdom 2Institute of Food Research, Department of Food Safety Science, Norwich Research Park, Norwich NR4 7UA, United Kingdom
3Institute of Food Research, Department of Nutrition, Health and Consumer Science, Norwich Research Park, Norwich NR4 7UA, United Kingdom Summary Most studies on the health-promoting properties of plant protease inhibitors have been carried out with the Bowman-Birk inhibitor (BBI) from soybean (Glycine max) but other legume seeds are also rich sources of protease inhibitors. We investigated two pea (Pisum sativum) protease inhibitors (TI1B and TI2B), homologous to BBI, but differing most significantly in amino acids at the two sites of enzyme inhibition. We over-expressed the pea protease inhibitor variants, using an expression system capable of efficiently folding extensively disulphide-bonded proteins and examined their effect, compared with BBI, on the proliferation of HT29 human colon cancer cells in vitro. Significant decreases in proliferation rates were observed using both pea and soybean protease inhibitors, with the pea proteins showing the largest reduction. Keywords: Aspergillus niger, inhibitory domains, Pisum sativum, anti-proliferation, protease inhibitors Introduction Proteases are key players in a wide range of biological processes and aberrant functioning of certain proteases has been linked to disease states, including cancer progression, in mammals. An understanding of the role played by proteases in these processes provides opportunities for therapeutic intervention. The role of diet in the prevention of human disease and, in particular, the positive contribution made by plant foods is widely publicised. However, the evidence for this is mainly epidemiological with little direct evidence for the benefits of individual plant components.
Naturally occurring protease inhibitors are abundant in many plant foods; some have the potential to inhibit digestive enzymes but can survive digestion and be transported as biologically active peptides. These dietary proteins are candidates for the modulation of protease activity in relation to disease. Studies with soybean inhibitors provide evidence that the inhibition of serine (chymotrypsin-like) proteases is relevant to the arrest of certain mammalian tumours (Kennedy, 1998). A class of serine protease inhibitors from pea has been characterised extensively at the biochemical and genetic levels (Domoney et al., 2002; Page et al., 2002). We have investigated the functional properties of two pea protease inhibitors that are homologous to BBI and have evaluated the anti-proliferative effects of these proteins on human colon cancer cells using an in vitro cell assay designed for high-throughput and rapid screening.

138
Materials and methods Expression and purification of pea protease inhibitors in Aspergillus niger Two pea protease inhibitor variants were expressed as recombinant proteins, using Aspergillus niger as an expression system capable of efficiently re-folding disulphide-bonded proteins. Sequences corresponding to mature TI1B and TI2B proteins were amplified from cDNAs (accession numbers X83211 and X83210, respectively), using primers that included an XbaI cleavage site to allow cloning into the glucoamylase fusion vector (pIGF) (Archer et al., 1994). The N-terminal primer was designed for in-frame fusion to the glucoamylase carrier protein and included a dibasic amino acid KEX2 processing site (KR). The KEX2 was expected to process the fusion protein C-terminal to the KR sequence just before the N-terminus of TI (GDD) in vivo. Glucoamylase-TI fusion expression plasmids were co-transformed with the pyrG+ selection plasmid, pAB4-1, into protoplasts of A. niger. Transformants were screened either by PCR, for the presence of the target gene, or directly for functional protein expression by trypsin inhibition activity assays (Domoney & Welham, 1992), prior to analysis of growth media by SDS-PAGE. Spent culture media were filtered, dialysed against distilled water and freeze-dried. Purified recombinant pea protease inhibitors were recovered by FPLC on MonoQ and MonoS columns. Functional properties of pea protease inhibitors Trypsin inhibitor activity was quantified, based on the hydrolysis of N-α-benzoyl-DL-arginine-p-nitroanilide (BAPNA) (Domoney & Welham, 1992). Chymotrypsin inhibitor activity measurements utilised N-benzoyl-L-tyrosine ethyl ester (BTEE) as a specific substrate, monitored in reactions at A256nm. The inhibition constants (Ki) of the recombinant pea inhibitors and BBI were determined for trypsin and chymotrypsin by competitive binding studies, using the substrates BAPNA and BTEE, in assays at pH 7.5 and pH 7.8, respectively. Cell proliferation Ninety-six well microtitre plates were inoculated at a density of 2,000 HT29 human colon adenocarcinoma cells per well in 200 µl of media that gave optimal growth. Cells were incubated under 5 % CO2 in humidified air for 24 h. After coating the wells, BBI or recombinant pea protease inhibitors were added at a range of concentrations, defined by chymotrypsin inhibitor units (CIU). Control wells received no inhibitors. Cells were harvested after 4 d without changing the media. Proliferation was assessed by vital staining, followed by cell lysis. The optical densities of the resulting solutions at 550 nm were measured using an automatic plate reader. The IC50 values were determined, based on the concentration of protease inhibitor that decreased cell proliferation by 50 % compared with the controls. Results and discussion Expression of recombinant pea protease inhibitors We used a protein fusion strategy to express individual pea protease inhibitor isoforms as recombinant proteins. In each case, the inhibitor protein was linked to part of the A. niger

139
glucoamylase protein, designed to target the fusion protein to the secretory pathway, and the inclusion of a processing site at the fusion junction allowed the release of the target protein.
The yield of recombinant pea protease inhibitors, determined from assays of A. niger growth media, was in the range of 17 to 24 mg/l. This was theoretically two orders of magnitude higher than previously obtained in Escheria coli (data not shown). The N-terminal sequences of the recombinant proteins produced by Aspergillus niger showed a consistent loss of three (GDD) or six (GDDVKS) amino acids compared with that predicted from the sequences. Molecular mass determination by mass spectrometry indicated further processing at the C-terminal ends with a loss of eight (EVEEVIKN) or nine (SEVEEVIKN) amino acids. The N-terminal processing of the recombinant protease inhibitors could be due to aberrant processing by KEX2 in the Golgi system or by limited later proteolysis. Processing at N- and C-terminal ends of pea protease inhibitors has been described and explains the derivation of multiple isoforms in vivo from relatively few genes (Domoney, 1999); isoforms lacking nine C-terminal amino acids have been identified in vivo. The expression of recombinant full-length protease inhibitors may be achieved, using A. niger mutants deficient in proteases. Functional properties of recombinant pea protease inhibitors Recombinant TI1B had a much higher specific chymotrypsin inhibitory activity compared with TI2B (Table 1). This is likely to reflect sequence differences at positions P1 and P2 of the inhibitory domains in these proteins. Both TI1B and TI2B showed a higher binding affinity (lower Ki) for chymotrypsin compared with trypsin. The strongest trypsin inhibitor was BBI but it had a value intermediate between TI1B and TI2B for chymotrypsin inhibition. These differences cannot readily be correlated with specific amino acid differences at or around the active sites (Table 1). More fundamental knowledge of the contribution of individual amino acids is necessary, in addition to an understanding of the role of amino-terminal sequences in stabilising protease-protease inhibitor complexes. Anti-proliferative effects of pea protease inhibitors on human colon carcinoma cells A statistically significant (P < 0.05) and dose-dependent decrease in the growth of HT29 human colon adenocarcinoma cells was observed after treatment with BBI, recombinant TI1B or TI2B. In this in vitro system, the recombinant pea proteins, TI1B and TI2B, were significantly more effective in inhibiting cell proliferation compared with BBI (Fig. 1), with an IC50 value of less than 50µM in the case of TI1B. These results may be explained following the identification of candidate proteases and through dissection of the apoptotic and mitotic processes that are affected in the HT29 cells. Such knowledge will be necessary to assess the therapeutic use of protease inhibitors as dietary proteins capable of inhibiting or preventing carcinogenesis in the gastrointestinal tract. Acknowledgements AC is a recipient of an EU Marie Curie fellowship (MCFI-1999-01516). Mass spectrometry was carried out by Dr Andrew R. Bottrill and Edman sequencing by Dr Mike J. Naldrett, both of the Joint IFR-JIC Proteomics Facility.

140
Ta
ble
1. A
min
o ac
id s
eque
nce,
con
stan
t of i
nhib
ition
(Ki)
and
spec
ific
activ
ity o
f try
psin
(T) a
nd c
hym
otry
psin
(C) i
nhib
itory
do
mai
ns o
f the
reco
mbi
nant
pea
pro
teas
e in
hibi
tors
, TI1
B an
d TI
2B, c
ompa
red
with
BBI
. Th
e re
activ
e pe
ptid
e bo
nd
site
P1-
P 1’ i
s sho
wn
in b
old
for e
ach
dom
ain.
A
min
o ac
id se
quen
ce o
f inh
ibito
ry
site
s K
i (n
M)
Spec
ific
inhi
bito
ry a
ctiv
ity
(U/m
g pr
otei
n)
T
C
T
C
T C
TI
1B
CLC
TKSN
PPTC
C
ICA
YSN
PPK
C
16.3
5 ±
8.20
7.
38 ±
0.9
2 2,
926
± 28
9 4,
973
± 29
0 TI
2B
CLC
TKSD
PPTC
C
ICA
LSY
PPQ
C
19.5
4 ±
9.07
13
.16
± 1.
34
2,59
6 ±
132
3,26
0 ±
270
BB
I C
AC
TKSN
PPQ
C
CIC
AL
SYPA
QC
1.
47 ±
1.1
6 10
.10
± 2.
25
4,39
6 ±
159
3,82
7 ±
336

141
Figure 1. Effect of BBI and the pea protease inhibitors, TI1B and TI2B (300 CIU added in
each case), on the in vitro growth of HT29 human adenocarcinoma colon cancer cells.
References Archer, D.B., Jeenes, D.J. & MacKenzie, D.A. 1994. Strategies for improving heterologous
protein production from filamentous fungi. Antonie van Leeuwenhoek 65, 245-250. Domoney, C. 1999. Inhibitors of legume seeds. In Seed Proteins. P.R. Shewry & R. Casey
(Eds). Kluwer Academic Publishers, The Netherlands. pp. 635-655. Domoney, C. & Welham, T. 1992. Trypsin inhibitors in Pisum: variation in amount and
pattern of accumulation in developing seeds. Seed Science Research 2, 147-154. Domoney, C., Welham, T., Ellis, N., Mozzanega, P. & Turner, L. 2002. Three classes of
proteinase inhibitor gene have distinct but overlapping patterns of expression in Pisum sativum plants. Plant Molecular Biology 48, 319-329.
Kennedy, A.R. 1998. Chemopreventive agents: protease inhibitors. Pharmacology and Therapeutics 78, 167-209.
Page, D., Aubert, G., Duc, G., Welham, T. & Domoney, C. 2002. Combinatorial variation in coding and promoter sequences of genes at the Tri locus in Pisum sativum accounts for variation in trypsin inhibitor activity in seeds. Molecular Genetics and Genomics 267, 359-369.
TI1B TI2B BBI
0102030405060708090
100
protease inhibitor
gro
wth
as
perc
ent o
f con
trol


143
Assessment of the potential of legume lectins to act as a mucosal adjuvant E. Gelencser1, G. Grant2 & D. Kelly2
1CFRI, Central Food Research Institute, Herman Ottó út 15, 1022 Budapest, Hungary 2Rowett Research Institute, Greenburn Road Bucksburn, AB21 9SB Aberdeen, Scotland, United Kingdom
Summary Mucosal vaccines can be used to treat systemic inflammatory diseases through the induction of antigen-specific mucosal tolerance. An in vivo mouse model was developed to demonstrate the beneficial adjuvant effect of Phaseolus vulgaris lectin (PHA) using a well-studied T-dependent dietary antigen ovalbumin (OVA). A preliminary experiment was conducted to determine the adjuvant dose. Antigen uptake increased in the circulation after the oral administration of a single feed of 25 mg OVA co-administered with 10 µg or 100 µg PHA-L adjuvant. For the assessment of tolerance induction mice were sensitised with a single feed of 2 mg of OVA and immunised subcutaneously with 100 µg OVA and 10 µg of complete Freud adjuvant 7 d later. Peripheral tolerance was assessed one week later. Our studies with the legume agglutinin PHA clearly demonstrated that orally administered lectin can facilitate oral tolerance induction to an unrelated, but anatomically co-localized marker antigen (OVA). Keywords: mucosal adjuvant, legume agglutinin, mucosal antigen uptake, oral tolerance Introduction Studies have shown that oral treatment with low doses of a soluble antigen, conjugated to the non-toxic lectinic B subunit of the cholera toxin (CTB) markedly suppressed IgE antibody responses and allergic reactions in sensitised mice. The authors suggested that the specific actions in the strong enhancement of oral tolerance by CTB-antigen conjugates are probably complex and include increased uptake of the antigen and CTB to the relevant mucosal surface and to the relevant mucosal antigen presenting cells (APC). The CTB bound on APC has a direct immune-modulatory effect which is associated with the development of an actively tolerogenic APC signal as well as the TGF-β-secreting suppressive-regulatory T-cells in mucosal tissues and draining lymph nodes (Borrebaeck & Carlsson, 1989).
It is postulated that plant lectins could have a similar significant adjuvant effect (Holmgren et al., 2003). Phaseolus vulgaris lectin (PHA) is not only mitogenic but it can also produce suppressor T cells by reacting with spleen cells. These cells can suppress lymphocyte transformation in mixed lymphocyte culture systems. Although PHA-binding may cause some microvillar damage of enterocytes, this is a fully reversible and is rapidly repaired by endogenous mechanisms and reorganisation of epithelial structures (Pusztai et al., 1997).
The main objective of our research was to demonstrate the beneficial adjuvant effect of PHA in the induction of peripheral tolerance to a well-studied T-dependent dietary antigen, ovalbumin (OVA) using an in vivo mouse model.

144
Materials and methods Preliminary animal experiment to determine adjuvant dose Female BALB/c mice were maintained on a standard diet under standard animal house conditions until use at 6 - 8 weeks of age. They had free access to water and food prior to the experiments. On the day of experiment mice were randomly selected into four groups of four animals in each test groups and one animal in the control group. Mice were starved for 3 h before the experiment started. To determine antigen uptake, mice were given a single feed of 25 mg OVA with or without 10 or 100 µg PHA-L adjuvant dissolved in 0.2 ml of saline via a stainless steel gavage needle. Control mice were not treated. To determine the kinetics of antigen uptake control animals were sacrificed prior to the experiment and a group of 3 test animals were killed either, 15 min, 30 min, 60 min and 120 min after the initial gavages. For all mice, 1 % heparin blood samples were obtained by cardiac aortic puncture under halothane anaesthesia. Plasma samples were stored at -20 °C until required for detection of serum antigen and antibody levels. Mouse model experiment to assess tolerance to induction Groups of BALB/c female mice (10 mice/group) fed on an OVA-free diet were given, by gastric intubation, a single dose of 2 mg OVA dissolved in 0.2 ml of saline with or without 10 or 100 µg PHA adjuvant. Control animals were given 0.2 ml of saline. Seven days later the animals were subcutaneously immunised with 100 µg OVA and 10 µg complete Freud adjuvant. To determine serum IgG levels 5 animals from each group were killed before immunization. Peripheral tolerance was assessed one week later. Plasma samples were stored at -20 °C until required for detection of serum antigen and antibody levels. Intestinal ovalbumin uptake Sandwich ELISA was used to measure OVA in mouse sera. A microtiter plate (Dynex) was coated with 10 µg/ml of monoclonal anti-chicken egg albumin-mouse IgG1 (Sigma) diluted (1:800) in carbonate-bicarbonate buffer at pH 9.6 and incubated for 1 h at 37 °C. Plates were washed five times with phosphate buffered saline containing 0.1 % tween (PBS–T, pH 7.4) before adding each successive reagent. Unoccupied binding sites on the plate were blocked with 2 % gelatine in PBS-T for 30 min at 37 °C. The assay was carried out on frozen samples. Two fold dilutions of samples from individual mice or standard OVA (20-0.6 µg/ml), prepared in duplicate, were assayed. After incubation for 30 min at 37 °C the specific primary antibody (anti-chicken egg albumin rabbit serum, Sigma) was added at working dilution (1:4000). After incubation for 30 min at 37 °C biotinylated secondary antibody (biotinylated anti-rabbit IgG, Sigma) was added at working dilution (1:1500). Following the next incubation step of 30 min at 37 °C ExtAvidin Peroxidase conjugate (Sigma) was added at working dilution (1:2000). Plates were incubated for 30 min at room temperature. This was followed by colour development using hydrogen peroxide and ortho-phenylendiamine containing Sigma Fast (Sigma) substrate reaction. The reaction was inhibited with 3 M sulphuric acid. Optical densities were measured at 492 nm against a reference filter at 630 nm by an ELISA reader (Denley Wellscan). For calibration of the results the linear regression (Y = Abs, X = LnConc) mode axes set up was used. Serum OVA levels were expressed in µg/ml.

145
Determination of antigen-specific antibody levels Antigen-specific antibody (OVA IgG) levels in the serum samples were monitored with ELISA assays in the samples from the mouse model experiments. Microtiter plates (Dynex) were coated with OVA (Sigma) in PBS, 7.4 at 5 µg/ml. After incubation (37 °C, 3 h) the plates were blocked with PBS containing 5 % foetal calf serum by incubation (37 °C, 1 h). Blocking buffer was replaced by diluted serum samples (1:10). Determinations were in duplicate for each animal. Diluted pulled mouse serum was added as an inside standard and PBS was used as a blank. After incubation overnight at 4 °C the antibodies were detected with biotinylated anti-mouse IgG (Sigma) diluted to 1:10000. The immune-complex was identified with diluted ExtAvidin Peroxidase conjugate (Sigma, 1:1000) by incubation (1 h, room temperature). Antigen-specific antibodies were visualised 20 min later using Sigma Fast (Sigma) substrate reaction. There were washings between the reactions. The reaction was stopped by 3 M sulphuric acid. Absorbance was read at 492 nm against a 630 nm reference filter with an ELISA reader. The results were evaluated in relation to the internal standard and are expressed in arbitrary units. Results and discussion Effect of PHA on uptake of a soluble marker antigen (OVA) via the intestine The first property of PHA, which we considered might be important in their mucosal adjuvancy, was an increase in the uptake of a soluble marker antigen (OVA) via the intestine. To test this idea, mice were fed 25 mg of OVA after feeding 10 or 100 µg PHA-L adjuvant and the level of immunogenic OVA was measured in the serum. As shown in Figure 1, control mice fed OVA had detectable amounts of OVA within 15 min Serum OVA levels rose quickly in mice fed 10 µg or 100 µg PHA-L co-administered to the test dose of OVA and these values peaked at 15 min and 120 min or 30 min respectively. Role of PHA-L in the induction of peripheral tolerance To demonstrate the beneficial adjuvant effect of PHA an in vivo mouse model was developed using the T-dependent dietary antigen ovalbumin (OVA). For the assessment of tolerance induction mice were sensitised with a single feed of 2 mg of OVA and immunised subcutaneously with 100 µg OVA and 10 µg complete PHA-L adjuvant 7 d later. Peripheral tolerance was assessed one week later (Fig. 2). As Figure 2 shows the serum OVA-IgG levels were suppressed more in mice given 100 µg PHA-L adjuvant co-administered to OVA (A) on the first day of the challenge compared with mice given only OVA (B). The serum OVA-IgG levels were enhanced in control mice, which were only given saline (C).
Our studies, with a legume agglutinin (PHA), have clearly shown that an orally administered lectin can facilitate oral tolerance induction to an unrelated, but anatomically co-localized marker antigen (OVA).

146
Figure 1. The serum OVA concentration at various times after feeding 25 mg OVA to BALB/c
mice with or without 10 µg or 100 µg PHA adjuvant. *P < 0.001. Mean ± s.d. (n = 12 at each time). 100 µg of PHA-L enhanced the uptake of orally administered protein into the serum. Serum levels in mice fed 25 mg OVA rose rapidly (A) and to more if the mice had been given 10 µg (B) or 100 µg of orally co-administered PHA-L (C) during the challenge.
Figure 2. The effect of PHA adjuvant on systemic immune responses. Serum OVA-IgG
responses in mice at various times after feeding 2 mg of OVA to BALB/c mice co-administered with or without 100 µg PHA adjuvant then immunized subcutaneously with 100 µg OVA and 10 µg complete Freud adjuvant 7 days later (day 7). Control mice fed saline. Peripheral tolerance assessed 1 week later (day 14). Results are pooled from 5 mice/group for each time and expressed as the mean of antibody units. *P < 0.001. Mean ± s.d. (n = 5 at each time). 100 µg PHA-L adjuvant co-administered to OVA (A) on the first day of the challenge suppressed the levels of serum OVA-IgG compared to mice which were only given OVA (B). Serum levels of OVA-IgG were enhanced in control mice given only saline (C).
References Borrebaeck, C.A.K. & Carlsson, R. 1989. Lectins as mitogens. In Advances in Lectin
Research. H. Franz (Ed). VEB Verlag und Gesundheit, Berlin. pp. 11-27.
0
5
10
15
20
25
0 15 30 60 120
Time after antigen feeding (min)
Seru
m O
VA
, ug/
ml
A B C* *
0
50
100
150
200
250
300
A B CAnt
i OV
A se
um Ig
G a
ntib
ody
units
day 7day 14
*

147
Holmgren, J., Czerkinsky, C., Eriksson, K. & Mharandi, A. (2003). Mucosal immunization and adjuvants: a brief overview of recent advances and challenges. Vaccine 21, Suppl. 2., S89-S95.
Pusztai, A., Gelencser, E., Grant, G. & Bardocz, S. (1997). Nutritional manipulation of immune competence in young non-ruminant animals. In Recent Advances in Animal Nutrition. P.C. Garnsworthy & J. Wiseman (Eds). Nottingham University Press, Nottingham. pp. 29-43.


149
A modelling approach to develop an antinutrient food databank with human nutrition implications Mª Joyanes Agencia Española de Seguridad Alimentaría, Carretera de Majadahonda a Pozuelo, Km 2, 28220, Madrid, Spain Summary Advances in the field of nutrition over the last decade have led to an understanding of the importance of non-nutrient food compounds. Beyond their interactions with nutrients, their functions in biological processes, toxicity, and some beneficial effects have been reported. Information regarding their modulating effects permits a wider perspective of these compounds than a simple positive/negative effect dichotomy. A knowledge of non-nutrients in food plays an important role in nutritional assessment, in the choice of the best varieties or conditions to obtain optimal nutritional values, in the application of industrial processes that avoid or minimise negative effects and in the development of functional foods.
A number of models to develop databanks of bioactive substances have been proposed. This extensive field needs to be divided according to previously established criteria.
Despite the large number of bioactive substances, including anti-nutrients, that have been identified, there is currently no database, which contains information on their presence and quantity in different foods. Different methods used, in compiling the tables makes it difficult to include all of the information, which is available in scientific papers into a single databank. For these reasons, it is necessary to compile a databank of these compounds using appropriate units. Keywords: non-nutrients, databank Introduction Although the so-called anti-nutrients have now been studied for over a century, their biological role is still not completely understood. Several hypotheses concerning their physiological functions have been suggested. Historically, it was assumed that these compounds reduced nutritional utilisation and food value because they inhibited or blocked important metabolic pathways. Legumes and oilseeds, important sources of protein and energy in several countries, are rich in these compounds (Ali & Muzquiz, 1998). The concept of antinutritional factors must be revised as a result of new data regarding their properties and recent interest in the beneficial or adverse effects on health of new non-nutrient substances. The term ‘bioactive substance’, used in the broad sense, includes all non-nutrient food components that interact in biological processes (Watzl & Leitzmann, 1995). It is surprising that food composition tables do not include either toxins, which are commonly present in plants or anti-nutrients, even in genetically modified foods (Novak & Haslberger, 2000).
The development of a database of non-nutritive food components is necessary to estimate their intake in epidemiological nutritional studies and to facilitate advice with regard to their intake. Recent dietary reference intakes have included some non-nutrient compounds. All food composition databanks need to be revised and updated continuously (Joyanes et al., 2001) due to changes in foods and new findings regarding the properties of non-nutrients and the way in which they affect biological processes. There is little information on some of these

150
food constituents. Nevertheless, the interest in bioactive substances is reflected in the growing number of publications that have appeared in this area over the last three decades (Pennington, 2002). However, these compounds and their classification remain controversial. The scientific community does not fully agree with the definition of a new nutrient offered by Hendrich et al. (1994), who wrote, ‘when obtained from the usual diet, a food component that sustains or enhances physiological functions and/or prevents diseases is a nutrient’.
In recent years great advances have been made in obtaining and compiling specific data on non-nutrient components, such as carotenoids (Granado et al., 1997) or flavonoids (Peterson et al., 1998). Ali and Muzquiz (1998) compiled data on anti-nutrients in legume seeds used in human nutrition. Materials and methods Scientific papers on antinutrients and computerised scientific literature searches using the terms ‘antinutrient’, and ‘bioactive substances’ were reviewed after our work on the criteria for optimising food composition tables in the databank. This focuses on external factors and specific problems affecting the non-nutrient content of food. The identification of certain information gaps e.g. units and expressions, equivalents or modulating effects to give a model for these kinds of food components. Results External factor affecting non-nutrient food content Environmental and genetics factors clearly influence the antinutrient content of certain plants, e.g. Phaseolus vulgaris, but not in the same way or to the same degree. Within a species, variety can influences the lectin and saponin content. Geographical location especially affects phytates and α-galactosides (Muzquiz et al, 1998). The high polyphenol and flavone content of olive (Olea europea) oil is associated with high antioxidant activity. Olives differ in their total biophenols content due, not only to their geographical origin, but also to the processing that they undergo (Blekas et al., 2002). Units and equivalents The results expressed as units of weight refer to the same units as the whole sample, percent should be used where possible, for comparative proposes. Equivalents are indirect and do not necessarily measure the same property. Consequently this limits their value and could be a source of error. Depending on the method used, e.g., in antioxidant capacity measurements the method plays a role and must be identified, as it may not reflect the same potential or site of action (Dae-Ok et al., 2002). Some non-nutrient compounds modulating dietary effects The protein antinutrients, as biologically active proteins, can modify or improve gut function, alter the gastrointestinal flora, body metabolism and health status. Bean (Phaseolus vulgaris) lectin can stimulate gut function and could ameliorate obesity if a safe, and effective, dose-range could be established for humans. This is due to its lipase inhibitory effects and lipolytic activity (Pusztai et al., 1998). Flavonoids are phytochemicals with potentially beneficial biological effects that are poorly characterized in existing composition tables. The flavonoid

151
content, in seed of the family Fabaceae, depends on plant species. The most common are flavonols and isoflavonoids, which are associated with hormone effects. Inulin and oligofructose are indigestible compounds that selectively stimulate growth and/or activity of potentially health-enhancing intestinal bacteria as prebiotics (Flickinger et al., 2003). Other compounds have pro-vitamin action, because they can be biologically transformed into vitamins, or anti-vitamins. Legumes and oilseeds have several compounds with antioxidant properties (Gorinstein et al., 2003). Model for a non-nutrient databank Common databank problems Many of the problems of a non-nutrient food composition databank are the same as those that occur with food composition tables for nutrients. The evaluation of existing data for scientific quality, collection and representative sample size used, the quality of the analysis, the correct taxonomical classification of the plant food, botanical varieties, processes, and the development a coding system, must all be used to obtain accurate food quality information. There are also generalised statistical conditions, which must be used in all cases. Specific problems in the development of bioactive substances databanks
• To define bioactive substances: this field is very broad and presumably activity depends on the specific structure of the compounds involved.
• More precise definitions are required of individual bioactive components and the selection, development and evaluation of their functions and interactions.
• Identification, definition and filling gaps. • Development of standards. • The use of common units of expressions, to obtain a coherent comparable database. Proposal
1. Determination in foods, which are most frequently consumed in each country, must be the
first step. 2. Identify, classify and quantify non-nutrient compounds, by their chemical family and
mode of action. 3. Data must be expressed in a form related to the edible part of the food by weight: units
measured must be those needed by food analysts. 4. The most active component (e.g. isomer) must be identified. Their principal and
functional nature; after which it is necessary to develop biomarkers, or equivalent-factors according to their function. Determine the safe upper intake level.
5. Improve information on the relationship between non-nutrients and nutrients where this is not known.
Conclusions Non-nutrient food component databases need to divide up the huge number of substances by selection of the most commonly consumed foods and then to identify the more important components according to their physiological or pathological activity; the development of standards; the use units of measurement which permit the use of the corrected data; improve

152
knowledge of relationships between non-nutrients and nutrients and to determine upper intake levels. References Ali, R. & Muzquiz, M. 1998. ANFs in tropical legume seeds for human nutrition. In Recent
Advances of Research in Antinutritional Factors in Legume Seeds and Rapeseed. . A.J.M. Jansman, G.D. Hill, J. Huisman & A.F.B. van der Poel (Eds). Proceedings 3rd International Workshop on Antinutritional Factors in Legume Seeds and Rapeseed, Wageningen 8-10 July 1998, 107-122.
Blekas, G., Vassilakis, C., Harizanis, C., Tsimidou, M. & Boskou, D.G. 2002. Biophenols in table olives. Journal of Agricultural and Food Chemistry 50, 3688-3692.
Dae-Ok, K., Won Lee, K., Joo Lee, H. & Yong Lee, C. 2002. Vitamin C equivalent antioxidant capacity (VCEAC) of phenolic phytochemicals. Journal of Agricultural and Food Chemistry 50, 3713-3717.
Flickinger, E.A., Loo J van & Fahey Jr, G.C. 2003. Nutritional responses to the presence of inulin and oligofructose in the diets of domesticated animals: A review. Critical Review Food Science and Nutrition 43, 19-60.
Gorinstein, S., Martin-Belloso, O., Katrich, E., Lojek, A., Ciz, M., Gligelmo-Miguel, N., Haruenkit, R., Park, Y.S., Jung, S.T. & Trakhtenberg, S. 2003. Comparison of the contents of the main biochemical compounds and the antioxidant activity of some Spanish olive oils as determined by four different radical scavenging tests. Journal of Nutritional Biochemistry 14, 154-159.
Granado, F., Olmedilla, B., Blanco, I., Gil-Martinez, E. & Rojas-Hidalgo, E. 1997. Variability in the intercomparison of food content data: A user’s point of view. Critical Review Food Science and Nutrition, 37, 621-633.
Hendrick, S., Kwang-Won, L., Xia, X., Huei-Ju & Murphy, P.A. 1994. Defining food components as new nutrients. Journal of Nutrition 24 (9 Suppl), 1789S-1792S.
Joyanes, Mª., Lema, L. & Outschoorn, I. 2001. Criteria for optimising food composition tables in relation to studies of habitual food intake. Congress Book 4th International Food Data Conference, Bratislava, 24-26 August 2001, 5-6.
Muzquiz, M., Burbano, C., Ayet, G., Pedrosa, M.P. & Cuadrado, C. 1999. The investigation of antinutritional factors in Phaseolus vulgaris. Environmental and varietal differences. Biotechnology, Agronomy Society and Environment 3, 210-216.
Novak, W.K. & Haslberger, A.G. 2002. Substantial equivalence of antinutrients and inherent plant toxins in genetically modified novel foods. Food Chemical Toxicology 38, 473-83.
Pennington, J.A.T. 2002. Composition databases for bioactive food components. Journal of Food Composition and Analysis 15, 419-434.
Peterson, J. & Dwyer, J. 1998. Taxonomic classification helps identity flavonoids-containing foods on a semiquantitative food frequency questionnaire. Journal of American Dietetic Association 6, 677-685.
Pusztai, A., Grant, G., Buchan, W.C., Bardocz, S., Carvalho, A.F. de & Ewen, S.W. 1998. Lipid accumulation in obese Zucker rats is reduced by inclusion of raw kidney bean (Phaseolus vulgaris) in the diet. British Journal of Nutrition 79, 213-21.
Watzl, B. & Leitzmann C. 1995. Bioaktive Substanzen in Lebensmitteln. Hippokrates Verlag, Stuttgart.

153
Beneficial outcome in hypercholesterolemia after intake of heat treated chickpea in rats M.A. Zulet1, M.T. Macarulla2, M.P. Portillo2 & J.A. Martínez1 1Department of Physiology and Nutrition, University of Navarra, 31008 Pamplona, Spain 2Department of Food Science and Nutrition, University of País Vasco, 01006 Vitoria, Spain Summary Legume seed consumption, particularly soybean (Glycine max), has been associated with improvements in the lipid profile in hypercholesterolemic individuals. Inclusion of chickpea (Cicer arietinum) in the diet gave beneficial responses in hypercholesterolemic rats, particularly in relation to cholesterol metabolism pathways. Because of these effects, it seems that the inclusion of chickpea in the diet could be recommended to lower saturated fats and cholesterol and to promote cardio vascular health. Keywords: Cicer arietinum, legume, cholesterol, fat, hypercholesterolemia Introduction Legume seeds have been associated with preventive or therapeutic properties in processes like hypercholesterolemia, diabetes and cancer (Messina, 1999). Although most studies have been on soybean, other legumes such as beans (Phaseolus vulgaris), peas (Pisum sativum) and chickpeas have also been claimed to have hypocholesterolaemic properties when included in the diet (Zulet & Martínez, 1995; Macarulla et al., 2001). This effect has not only been associated with the legume protein but with the presence of other components in the seed such as saponins, isoflavones, phytic acid, trypsin inhibitors, soluble dietary fibre and the lipid composition of the seeds (Erdman, 2000). The aim of this study was to assess possible therapeutic effects on lipid metabolism of a diet containing heated chickpea seed in a diet-induced model of hypercholesterolemia. Materials and methods Animals and diets Twenty four male Wistar rats weighing about 200 g were assigned to two groups. Control rats (n = 8) were fed a semi-purified diet containing casein as a protein source and olive oil (5 %) as fat and rats (n = 16), which received casein and fat enriched in saturated fatty acids (coconut oil 25 %) and cholesterol (1 %) as previously described (Zulet et al., 1999a). After 26 d, animals on the high-fat diet were hypercholesterolemic. They were then assigned to two new subgroups which received the following diets for 16 d: one hypercholesterolemic group (n = 8) continued on the same high-fat diet (HH) and the remaining hypercholesterolemic rats (n = 8) were fed on a diet containing chickpea as a protein source and olive oil (5 %) as fat (HL). Whole chickpea seed was ground into a fine powder and autoclaved at 112 ºC for 12 min prior to incorporation into the diet. The control group continued on the same initial diet (CC). The composition of the different diets and the chickpea can be obtained from Zulet et al. (1999a, b).

154
Procedures At the end of the experimental period (42 d) the rats were euthanized. Serum was separated from blood samples and the abdomen and liver were removed, weighed and frozen at –20 ºC. Total Serum cholesterol (CHOL); high density lipoproteins (HDL-cholesterol), low density lipoproteins (LDL-cholesterol), and triacylglycerols (TG) were measured using commercial kits (Boehringer Mannheim). Liver cholesterol was analysed with o-phataldehyde reagent (Zulet & Martínez, 1995). The enzyme 3-Hydroxy-3-methylglutaryl-CoA (HMGCoA) reductase was isolated from liver microsomes and assayed through the rate of NADP+ release (Macarulla et al, 2001). Lipoprotein lipase (LPL) activity was measured in adipose tissue by a fluorometric assay (Zulet et al, 1999a). Statistical analysis The results are presented as the mean ± SEM. The data was statistically analysed using a one-way ANOVA followed by a Fisher PLSD test, or by the Kruskal-Wallis and Mann-Whitney-U tests as appropriate. All the statistical analyses were performed using the Statview� package. Results Animals fed on the high-fat diet enriched with saturated fat and cholesterol (HH group) showed a significant increase in total cholesterol (+362 %; P < 0.01) and LDL-cholesterol (+2,660 %; P < 0.001). There was also a lower HDL-cholesterol level (-62 %; P < 0.01) compared to rats fed the control diet (CC group). Moreover, these disturbances were linked with increased liver cholesterol (+1,583 %; P < 0.01). However, in the HL group there were significant decreases in total cholesterol (-70 %; P < 0.01), LDL-cholesterol (-88 %; P < 0.01) and the total cholesterol/HDL ratio (-86 %; P < 0.01) as well as liver cholesterol (-47 %; P < 0.01) compared to the HH group. Liver HMGCoA reductase activity increased significantly (+127 %; P < 0.05). There was a significant decrease in LPL activity (-55 %; P < 0.01) in the chickpea fed rats compared to the HH group (Fig. 1) Discussion The importance of dietary intake in the management of lipid status is well established. Because of this there has been considerable interest in soy based foods among both consumers and health care professionals. This is due to their potential benefits in a variety of chronic diseases, especially cardiovascular illnesses (Messina et al, 1999). However, only a few experiments have been done with other grain legumes such as chickpea. This species is a leading food staple in developing countries on account of its low price and high nutritive value. In contrast, animal and human nutrition studies have confirmed the hypercholesterolemic effects of consuming saturated fatty acids and cholesterol by increasing serum cholesterol and altering the lipoprotein pattern (Zulet et al, 1999a).
The health benefits observed after legume consumption have been attributed to their fibre content, the vegetable protein, fatty acid content, trypsin inhibitors, phytic acid, saponins and isoflavones (Anderson & Major, 2002). In this trial the chickpea was autoclaved, prior to feeding, to avoid undesirable responses such as reduced growth, low nitrogen retention and the inhibition of intestinal functions from eating raw chickpea seed. Previous studies have shown that methods such as wet and dry-heating, germination and fermentation reduce the concentration of several antinutritional factors such as phytic acid, trypsin inhibitors,

155
polyphenols and lectins, and improve in vitro protein digestibility (Larralde & Martínez, 1989). As thermolabile factors are reduced or eliminated by autoclaving (Zulet & Martinez, 1995), the hypocholesterolemic effects of the chickpea diet were probably due either to amino acid, carbohydrate and/or fat composition, or to thermostable factors in the chickpea seed such as isoflavones and saponins.
Figure 1. Liver HMGCoA activity and adipose tissue LPL activity in rats fed a control diet for
42 d (CC); rats fed a high-fat diet for 42 d (HH) and rats fed a high-fat diet for 26 d and a chickpea based diet for the following 16 d (HL). (*P < 0.05; ** P < 0.01; *** P < 0.001).
Attempts have been made to explain the mechanisms by which legume seed
constituents exert a beneficial effect on plasma lipids and fatty liver content (Messina, 1999; Erdman, 2000). These have included LDL receptor upregulation, inhibitory effects on intestinal cholesterol absorption, increased faecal bile acid excretion, bile acid synthesis from cholesterol in the liver accompanied by increased LDL removal from the blood and cholesterol loss from the body. Other legume seed components such as isoflavones improve the lipid profile and act as antioxidants, which might inhibit LDL oxidation (Jenkins et al., 2002). Further, some beneficial effects of legume seed intake may involve plasma concentration changes in insulin, glucagon, or thyroid hormones (Frühbeck et al., 1997). Several studies have shown a relative depletion of tissue cholesterol and an increase in hepatic cholesterogenesis following legume seed intake (Macarulla et al., 2001). These results are in keeping with the increase in HMGCoA reductase activity found in this study, which was probably due to suppression of cholesterol intake and reductions in total cholesterol and LDL cholesterol and liver cholesterol. Additionally, the decrease in LPL activity after legume intake might be associated with the observed rise in TG levels, since it has been reported that LPL may play an important role in mediating triacylglycerol uptake.
The results of this study suggest that the regular consumption of chickpea seed might exert a protective and corrective role in some alterations of lipid metabolism, which may provide cardiovascular and other health benefits similar to those described after soybean consumption. References
Anderson, J.W. & Major, A.W. 2002. Pulses and lipaemia: effect: potential in the
prevention of cardiovascular disease. British Journal of Nutrition 88, S263-271.
0 2 4 6 8
10 12 14 16
HMGCoA LPL
nmol/min x mg prot
CC HH HL
*
*** ** *
*

156
Erdman, J.W. 2000. Soy protein and cardiovascular disease. Circulation 102, 2555-2563. Frühbeck, G., Monreal, I. & Santidrian, S. 1997. Hormonal implications of the
hypocholesterolemic effect of intake of field beans (Vicia faba L.) by young men with hypercholesterolemia. American Journal of Clinical Nutrition 66, 1452-1460.
Jenkins, D.J.A., Kendall, C.W.C., Jackson, C.J.C., Connelly, P.W., Parker, T., Faulkner, D., Vidgen, E., Cunnane, S.C., Leiter, L.A. & Josse, R.G. 2002. Effects of high- and low-isoflavone soyfoods on blood lipids, oxidized LDL, homocysteine, and blood pressure in hyperlipidemic men and women. American Journal of Clinical Nutrition 76, 365-372.
Larralde, J. & Martínez, J.A. 1989. A reappraisal of the nutritional utilization of legumes. Journal of Physiology and Biochemistry 45, 220-227.
Macarulla, M.T., Medina, C., Diego, M.A. de, Chavarri, M., Zulet, M.A., Martínez, J.A., Nöel-Suberville, C., Higueret, P. & Portillo, M.P. 2001. Effects of the whole seed and a protein isolate of faba bean (Vicia faba) on the cholesterol metabolism of hypercholesterolaemic rats. British Journal of Nutrition 85, 607-614.
Messina, M.J. 1999. Legumes and soybeans: overview of their nutritional profiles and health effects. American Journal of Clinical Nutrition 70, 439S-450S.
Zulet, M.A., Barber, A., Garcin, H., Higueret, P. & Martínez, J.A. 1999a. Alterations in carbohydrate and lipid metabolism induced by a diet rich in coconut oil and cholesterol in a rat model. Journal of the American College of Nutrition 18, 36-42.
Zulet, M.A., Macarulla, M.T., Portillo M.P., Nöel-Suberville, C., Higueret, P. & Martínez, J.A. 1999b. International Journal of Vitamin and Nutrition Research 69, 403-411.
Zulet, M.A. & Martínez, J.A. 1995. Corrective role of chickpea intake on a dietary-induced model of hypercholesterolemia. Plant Food for Human Nutrition 48, 269-277.

Session III B Negative effects of antinutritional factors in human nutrition


159
Storage proteins: physiological and antigenic effects L.A. Rubio1, J. Rodríguez2, C. Fernández2 & J.F. Crespo2 1Unidad de Nutrición, Estación Experimental del Zaidín (CSIC), Profesor Albareda 1, 18008 Granada, Spain 2Servicio de Alergia, Hospital Universitario 12 de Octubre, Avenida de Córdoba s/n, 28041 Madrid, Spain Summary Increased intestinal excretion of endogenous proteins and urinary N loss, changes in plasma amino acid concentration, modulation of plasma lipids and cholesterolemia, and adverse immune reactions have been described in animals and humans consuming legume- or legume protein-based diets. Most allergens described, so far, as causing allergic reactions to legume seeds in humans are storage proteins. The chemical structure of these particular groups of proteins is probably the ultimate cause of their nutritional and/or physiological behaviour. Keywords: legumes, storage proteins, protein and lipid metabolism, allergy Introduction Legume seeds are important sources of nutrients such as minerals, vitamins and proteins for both animals and humans. However, they also contain significant amounts of natural toxins or antinutritional factors (ANFs). The deleterious effects of ANFs have been extensively investigated in animal and human nutrition. Increased intestinal excretion of endogenous proteins and loss of N in the urine, changes in plasma amino acid concentrations, modulation of plasma lipids and cholesterolemia, and adverse immune reactions have been described in animals and humans consuming legumes- or legume protein-based diets. In particular, legumes have been implicated in adverse reactions characterized by abnormal or exaggerated immunological responses to specific food proteins that result in a variety of symptoms (food allergy or hypersensitivity). The most important proteins in legume seeds from both the nutritional and allergenic points of view are the storage proteins.
The protein content of legume seed is between 20 and 50 % of the DM. Most of this protein (about 80 %) is in the form of storage proteins. These are those laid down in the protein bodies during seed development for future use at a metabolically more active stage. Storage proteins are mainly salt soluble globulins, which can be further separated into two major fractions, vicilin and legumin, by changing the solution pH. For example, the soybean (Glycine max) vicilin and legumin isoelectric points are about 4.8 and 6.6 respectively (Thanh et al., 1975). They are generally glycoproteins (1 - 5 % carbohydrate), rich in amides (glutamic acid-glutamine, aspartic acid-asparagine and arginine) and poor in sulphur amino acids (methionine, cystine).
Until recently, feed and food proteins were supposed to be degraded in the gastrointestinal tract to amino acids, which were then absorbed and utilised primarily by the liver to synthesise proteins or they entered the general amino acid pool of the organism. This simplistic approach has changed recently as a number of observations have indicated that dietary proteins entering the organism can significantly influence a number of physiological functions. The best-known examples of this are probably proteins like some lectins and the protease inhibitors. However, the effects observed in humans and animals after the ingestion

160
of other dietary proteins have also attracted recent attention. In this review, a number of observations are reported which suggest that closer attention should be paid to traditional dietary proteins to better understand feed- or food-host interactions. On the basis of that understanding, proteins and/or foodstuffs could be manipulated to increase their potentially beneficial effects in both animal production and human health. I - Physiological effects Digestibility and N metabolism. Feeding growing animals with diets based on legume seed meal as the main protein source results in poorer performance than expected based on the chemical composition of the diet. It is generally thought that the presence of ANFs together with the slower digestion rate of legume, compared to animal protein, could explain their lower nutritional efficiency. However, this conclusion is mostly based on experiments in which whole seed meal was used in the diet. When purified proteins were tested usually only in vitro values were reported. One of the few reports on in vivo digestibility with purified proteins is that of Aubry & Boucrot (1986). They showed that after 2 h gastric emptying the intestinal absorption of pea (Pisum sativum) vicilin and legumin in rats was as high as casein. However, the nutritional value, measured as protein efficiency ratio and biological value, of diets based on purified legume proteins or seed meals containing low or no ANFs is lower than control diets even though both the faecal and ileal digestibility of globulins purified from legume seeds such as soybean, faba bean (Vicia faba), and narrow-leafed lupin (Lupinus angustifolius) was no different from control values in the rat (Rubio et al., 1991, 1994). Furthermore, the ileal and faecal nitrogen (N) digestibility of whole legume seed meal low in ANFs are usually similar or close to control values (Rubio et al., 1995, 1998, van der Poel et al., 1992). These results suggest that undenatured legume globulins are highly digestible in the small intestine, and therefore the lower legume protein digestibility when the whole meal is used in the diet is likely to be due to other factors such as lectins, tannins and/or trypsin inhibitors.
Santoro et al. (1997) found that in rats fed a diet containing highly purified phaseolin, the main globulin from the Phaseolus vulgaris seed, the faecal N digestibility, based on a conventional estimation method, was only 37.5 %. However, using immunological techniques, it was shown that the bulk of faecal N was not chemically related to phaseolin. After correction for the non-phaseolin N, the true digestibility of the phaseolin was estimated to be 74.3 %. It was suggested that phaseolin and/or undigested fragments derived from the native molecule may, as reported for the lectins, stimulate secretion of endogenous N, possibly mucins. Similarly, Huisman et al. (1992) observed in pigs that the ileal true protein digestibility of peas was between 93 and 95 % using the 15N-dilution technique. This indicated that the pea protein was almost completely digested by enzymes in the small intestine. However, at the ileal level apparent protein digestibility was between 74 and 79 %. Thus, the lower apparent ileal protein digestibility of pea proteins can be attributed to excretion of endogenous protein. The apparent ileal protein digestibility of toasted common beans (P. vulgaris) was about zero (-4 %), while the true ileal protein digestibility was about 66 %. Although the reasons for these effects are at present unclear, they might be related to abnormal immune reactions (see below) against the proteins, or their fractions produced after enzymatic hydrolysis in the small intestine. Whatever the reason for the effect, it suggests that some dietary legume storage proteins can induce changes in the digestive process and are not simply degraded to a higher or lower extent.

161
The inclusion of narrow-leafed lupin, faba bean, chickpea (Cicer arietinum) or soybean purified globulins in rat diets had a profound effect on the nutritional performance of the animals, with lower gain:feed ratios leading to low net weight gains compared with control animals (Rubio et al., 1995, 1998). Except for soybean globulins, this lower performance was accompanied by a significantly increased urea excretion in the urine. High urea excretion is a well-known effect associated with legume feeding, and an inverse correlation between the biological value of different proteins and blood urea concentration has been reported (Eggum, 1970). As urea is the main end product of protein catabolism in mammals, high plasma urea values are associated with disturbances in protein metabolism and increased protein degradation, which finally result in a loss of N in the urine. The final result is lower N retention and net protein utilisation values. However, in the above experiments the diets were fully supplemented with essential amino acids to give the same values as in the control diets, and total N digestibility in the small intestine was high. Therefore, the nutritional value of the diets based on control (lactalbumin) or legume proteins should, theoretically be very similar. Nevertheless, it is possible that the rate of protein digestion and/or the absorption of amino acids/peptides were different from those required by the animal for adequate protein synthesis. The absorption of small peptides and free amino acids in the small intestine occurs by independent mechanisms and their rate probably differs (Webb, 1990). As a result, the mix of amino acids available for protein synthesis in the tissues at a given moment could be imbalanced. This imbalance can lower the rate of protein synthesis, resulting in increased amino acid catabolism (Benevenga et al., 1993). In this respect, we recently determined in Caco-2 cells that amino acids from legume proteins are probably absorbed at different rates from those of proteins of animal origin such as casein (Rubio & Seiquer, 2002).
A number of papers have reported on the plasma amino acid profiles of animals fed legume-based diets. Thus, free plasma concentrations of glycine, histidine and arginine were higher than controls in rats fed diets based on faba bean, lupin, chickpea and soybean meals or their respective globulin proteins. On the other hand, threonine, leucine and lysine levels were lower in animals fed the same diets. Other amino acids (tyrosine, asparagine, glutamine, serine) were not significantly affected, or were only affected in some cases (alanine, methionine, isoleucine, phenilalanine and ornithine) (Rubio et al., 1999). The higher glycine and arginine content in the legume proteins than in lactalbumin might explain the higher plasma values found in the rats fed legume-based diets. However, the same does not apply to histidine. The amount of threonine, leucine and lysine, whose plasma values were substantially lower in rats fed legume proteins, were similar or higher in the legume proteins compared with the lactalbumin. Furthermore, in all cases the legume-based diets were supplemented with amino acids to match the control values. Lower lysine values were found in the rats after just 9 h of feeding with supplemented lupin-based diets compared with lactalbumin (Rubio et al., 1995). Batterham et al. (1986) reported low lysine availability in rats and pigs fed white lupin (Lupinus albus) meal. The cause was unknown and lysine supplementation did not overcome the poorer weight gain of pigs fed on lupin seed meal compared with soybean meal. Therefore, effects on plasma amino acid concentrations, at least for lysine, are fast and are probably independent of supplementation with the deficient amino acids in the diet.
Other effects on physiological parameters in animals fed diets based in soybean protein have been reported which also suggest a physiological effect on protein metabolism. Thus, Nielsen et al. (1994) found that, compared with hydrolysed soybean protein, intact casein gave 2.2 to 2.8 times higher rates of protein synthesis and degradation respectively at the same N balance. They suggested that the effect was related to differences in plasma amino acid concentrations observed with the two proteins. Chiji et al. (1990) and Hara & Kiriyama (1991) studied the absorption of free methionine or oligo-L-methionine (mixture of synthetic

162
hexa- and heptapeptides of methionine) added to diets containing casein or soybean protein in rats. Apart from the fact that the methionine and oligo-L-methionine differed in their nutritional effects, there were different rates of oligo-L-methionine digestion due to the presence of casein or soybean protein in the diet. The amino acids from casein were absorbed faster than those from the soy protein isolate. Also, Hajós et al. (1996) recently showed that enzyme methionine enrichment (chemically incorporated into the protein structure) of soy proteins significantly improved their nutritional value compared with methionine supplemented soy proteins. On the other hand, over the last 15 years evidence has accumulated for the absorption of significant amounts of peptides and even intact proteins during normal digestion (Webb 1990). Furthermore, Friedrich et al. (1984) observed that, for similar total absorption values, the dynamics of the digestive-absorptive process in rats are different for proteins of both animal and plant origin when treated with different enzymes. Therefore, as also suggested by Sholz-Ahrens et al. (1990), it is possible that some absorbed peptide(s) arising from protein hydrolysis in the intestine are biologically active and can produce a metabolic effect. As a result there may be altered activity of amino acid degrading enzymes in the liver (Cenarruzabeitia et al., 1979; Iritani et al., 1986) with increased urea production and loss of amino acids for protein synthesis. Liver relative weights were lower in rats fed whole faba bean, lupin or defatted soy seed meals or their globulin proteins. This lower liver weight is likely to be the result of changes in tissue composition. In fact, both glycogen concentration and total content in the liver were lower in rats fed lupin meal diets (Rubio et al., 1995) compared with lactalbumin or casein controls. Total protein and RNA levels were also lower. Total DNA did not differ from the controls. Lipid metabolism A number of mechanisms have been claimed to explain the effects of proteins on lipid metabolism. However, two circumstances should be pointed out. Firstly, there is uncertainty about the effects of potentially active components in the protein preparations used which may contain up to 20 % of non-protein material (soluble fibre, phytosterols, saponins, phytates etc.). Secondly, most of the work has compared soybean proteins with casein, and there is much less published information on other plant and animal proteins. According to Beynen (1990) the mechanisms proposed to explain the effects of legume proteins on lipid metabolism can be classified into two major groups: a) those with primary features at the intestinal level, and b) those with primary features at the postabsoptive level. For the first group, two concepts have been proposed to explain intestinal steroid absorption inhibition in animals fed on soy protein. (1) Undigested protein binds bile acids and neutral steroids and reduces steroid re-absorption (Sugano et al., 1988). However, soy protein is highly digestible in vivo (see above) and less digestible treated casein does not lower serum cholesterol concentration (West et al., 1989). Dietary casein (a highly phosphorylated protein compared to soybean protein) and phosphopeptides derived from it remove calcium from the calcium phosphate sediment in the intestine, dissolve the sediment and solubilise phosphate which leads to a reduction of binding sites for bile acids and bile acid absorption is increased (van der Meer, 1983). However, the phosphorylation concept may not extend to other animal proteins that are not highly phosphorylated but do increase serum cholesterol concentration. Also, milk-whey protein lowered plasma and liver cholesterol with respect to casein while reducing faecal bile acid excretion. This led Zhang & Beynen (1993) to conclude that the effect probably resides in different protein structure or the non-protein components rather than in different amino acid composition per se.

163
The mechanisms proposed at the postabsoptive level involve: (1) the absorption of potentially active oligopeptides derived from soybean hydrolysis, and (2) effects of single amino acids (lysine, arginine). Both the potential absorption of peptides and the effects of legume proteins on plasma amino acids profile have been discussed above. More specifically, Lovati et al. (1992) found that in vitro soybean 7S and 11S globulins increased uptake and/or degradation of 125I-LDL in Hep G2 and HSF cell lines. Furthermore, rats fed a casein-cholesterol diet treated daily with 7S or 11S globulins for 2 weeks had reduced cholesterolemia and their liver membrane preparations showed a non-significant increase in the maximal binding of labelled cholesterol-rich lipoprotein fraction (β-VLDL) to high affinity receptors. Bloch et al. (1988) suggested that peptides reported to cross the intestinal barrier and reach the blood stream might be implicated in these effects. Further, Sirtori et al. (1999) showed, in a double-blind study with humans, there was a significant cholesterol-lowering effect of soybean protein, even when it only partly replaced animal protein in the diet.
Regarding mechanism (2), although numerous studies have dealt with the influence of dietary amino acids on plasma lipids and cholesterol, the results are far from unequivocal. Kingman et al. (1993), using diets based on beans (Phaseolus vulgaris and P. lunatus) peas or lentils (Lens culinaris) with pigs, concluded that the mechanism for the hypocholesterolaemic effect of legume feeding does not involve increased cholesterol clearance via the intestinal route, but rather has effects on plasma amino acids. Rubio et al. (1995) also found plasma triglyceride concentration in rats fed diets containing whole Lupinus angustifolius meal; legume globulin proteins or lupin fibre was significantly lower than in control animals, while plasma cholesterol levels were not affected. The addition of arginine to a control lactalbumin-based diet led to a decrease in plasma triglyceride concentration. These results agree with those of Sugano et al. (1982) who reported that the arginine:lysine ratio is more effective in regulating triglycerides than serum cholesterol levels. Horigome & Cho (1992) determined that soybean protein affects the concentration of serum cholesterol, triglycerides and free amino acids in rats. With the current available information, it is not possible to definitely conclude whether specific amino acids are responsible for the cholesterol-modulating effect. However, it seems likely that differences in hormones are involved in the underlying mechanism. According to this hypothesis, legume proteins would affect either the plasma insulin/glucagon ratio (Noseda & Fragiacomo, 1980) or thyroid hormones (Sholz-Ahrens et al., 1990) through their effects on plasma amino acid concentrations. The immune system Over the last 10-15 years a number of articles and reviews (Sissons, 1989; Lallès et al., 1993; Lallès & Peltre, 1996) have dealt with the mode of action and effects of legume proteins acting as antigens/allergens in farm animals. Early work with calves first indicated the possibility that immune responses to dietary soybean protein might be linked with disorders in gastrointestinal function as high titres of circulatory anti-soy antibodies (IgG type against glycinin and β-conglycinin, the major storage proteins of soybean) no abnormalities in gut microflora were observed in calves suffering from diarrhoea after receiving a soy-based diet. Loss of normal villous architecture, oedema, increased numbers of intra-epithelial lymphocytes and crypt hyperplasia were observed in calves, which had been orally sensitised to soybean flour (Pedersen et al., 1984). Intestinal hypersensitivity to dietary antigens has also been postulated to produce a predisposition to diarrhoea, which sometimes occurs when piglets are weaned onto soy based diets (Miller et al., 1983). Consequently, when milk replacers containing legume proteins are fed to calves, growth is depressed by 20 - 30 % of

164
that of control animals fed skim milk protein (Lallès, 1993). In piglets legume-based starter diets usually give a transient reduction in performance.
The relative spleen and thymus weights are lower than in control animals in rats fed diets based on lupin, chickpea or soybean meals or their globulins (Wang & MacIntosh, 1996; Rubio et al., 1999). These results are consistent with those of others on immune system parameters of rats and mice fed legume based diets, although the reasons for them are not clear. Thus, Tarwid et al. (1985) found that liver, kidney and spleen relative weights and total white blood cell counts and lymphocyte counts were significantly depressed in rats fed lentil-based diets. Esparza et al. (1996) reported lower liver and spleen relative weights in pea-fed mice compared to casein fed controls. There were also significant increases in immunoglobulins and IgG after legume feeding. The percentage of splenic T cells and helper T cells was significantly increased by the legume diet. They suggested that feeding mice on a diet containing peas as protein source reduced nutrients utilisation and impaired nutritional performance that may be associated with an immune mechanism.
The mechanisms involved in these immunological effects are unclear. Most of the work has been on calves. This is due to the fact that in pigs or even rats natural transient hypersensitivity precedes the induction of oral tolerance after weaning, with increased numbers of T lymphocytes in the intestinal lamina propria and epithelium. However, the actual dietary allergens involved are still not identified (Dréau et al., 1994). With preruminant calves direct skin tests and in vitro lympho-proliferation assays show that β-conglycinin is probably the most allergenic protein in soybeans. Other proteins like α-conglycinin, glycinin, Bowman-Birk trypsin inhibitor and lectins also seem to be involved to a lesser extent. The increased densities of T lymphocytes in the mucosa together with a lack of detectable antisoy IgE antibodies (implicated in Type I immediate hypersensitivity) led Lallès et al. (1996) to conclude that in chronic situations the intestinal disorders were caused by specific T-cell activation. It is important to note that the proteins implicated in immune reactions are those that survive undigested for the longest time in both the small intestine and in in vitro assays (Sissons et al., 1984; Li et al., 1991; Lallès et al., 1993). However, the cellular and molecular basis of intestinal villus atrophy and crypt hyperplasia, probably involving the release of cytokines by activated intestinal T lymphocytes and macrophages, is still unexplored. II - Antigenic effects in humans Legumes represent an example of the effect of the staple diet on the development of food allergies through ingestion. Thus, high consumption of peanuts (Arachis hypogaea) and soybeans in North America correlates well with the importance of these foods in allergic reactions. However, green bean, chickpea, and lentil allergy are found in Mediterranean countries and India where these foods are an important source of fibre and inexpensive protein. In fact, lentil, chickpea, and beans still provide 4.9 % of the protein in the Spanish diet. Prevalence Peanut allergy occupies a special place in the spectrum of food allergies, because of its frequency and severity. It is responsible for the greatest number of deaths, particularly in the United States and the United Kingdom. An increase in the frequency of this allergy has also been noted in France (Moneret-Vautrin et al., 1998). Several population-based studies have estimated the prevalence of peanut allergy. Tariq et al. (1996) followed a cohort on the Isle of

165
Wight (in the United Kingdom) from birth until the age of 4 years. At 4 years, 0.5 % of the children had had an allergic reaction to peanuts. On the basis of this and other studies, the estimated prevalence of peanut allergy in these countries is between 0.6 % and 1.0 % (Kanny et al., 2001). A recent follow-up study found that the prevalence of peanut allergy had increased up to 1.5 % on the Isle of Wight. This suggests that the problem is increasing (Grundy et al., 2002). Several factors may account for the apparent increase in prevalence; these include the early age of exposure to peanuts, and the way peanuts are cooked, roasted versus boiled or fried (Sampson, 2002). The clinical expression of peanut allergy ranges from mild to severe anaphylactic reactions. Peanuts are responsible for the majority of fatal and near-fatal anaphylactic food-induced reactions. Sometimes symptoms are developed after ingestion of less than 2 mg of peanut protein: one peanut contains about 200 mg (Al-Muhsen et al., 2003). Soybean is another important legume seed, which causes anaphylactic reactions. In Sweden from 1993 to 1996 there were 61 cases of severe food reactions reported in children. Five of them were fatal. Peanut, soy and tree nuts caused 45 of the 61 reactions and 4 of the fatalities were caused by soy (Foucard & Malmheden, 1999).
There is little published information on the role of other temperate legumes including lentil, chickpea, pea, green beans, and sweet lupin species. Most reports are from Spain and Greece, where green beans, lentils, and chickpeas are included in the Mediterranean diet. Our previous studies have shown that in Spain, with a typical Mediterranean diet, lentils are the legumes most commonly implicated in IgE-mediated food hypersensitivity. In 355 children who were diagnosed as having food hypersensitivity, lentil allergy occurred in 10.1 % of patients, while peanuts caused allergic reactions in 6.7 % (Crespo et al., 1995). We addressed the clinical and immunologic features in 22 lentil-allergic children (Pascual et al., 1999). The study showed that allergic reactions to lentil usually start before 4 years of age. The most common symptoms are oropharyngeal and acute urticaria through ingestion. Symptomatic reactivity to chickpea is frequently present. Kalogeromitros et al. (1996) described a case related to the ingestion of cooked lentil and chickpea and with inhalation exposure to cooking lentil soup. Recently, Patil et al. (2001) verified, with double blind, placebo-controlled food challenges the diagnosis of chickpea allergy in 31 patients from India.
Legume allergens can elicit respiratory symptoms by inhalation, sometimes skin symptoms are also observed. Besides anecdotal reports of allergic respiratory reactions from exposure to cooking vapours of some legumes (peas, chickpeas, beans, and lentils) (Martin et al., 1992; Garcia Ortiz et al., 1995) and to peanuts in commercial flights (Sicherer et al., 1999), white lupin (Lupinus albus) flour seems to be an important cause of allergic sensitisation in exposed workers. Thus, it might give rise to occupational asthma and food allergy (Crespo et al., 2001). Allergenic proteins of legume seeds The majority of allergenic proteins from vegetable foods are either pathogenesis-related proteins or seed storage proteins. Most allergens described so far as causing allergic reactions to legume seeds in humans are storage proteins. For practical purposes, food allergy is generally subdivided into disorders mediated by IgE antibodies and those resulting from non-IgE-mediated mechanisms. However, IgE-mediated reactions are the most common immunologic disorders following legume ingestion. Several factors may account for the appearance of allergic reactions to one or another legume, such as the amount of legume consumed in the area, the intrinsic ability to produce IgE-mediated reactions and the genetic background.

166
Cupin superfamily Allergenic vicilins: The most relevant among the allergenic vicilins in legumes is the major peanut allergen Ara h 1. Its linear IgE-binding epitopes have been identified and mutational analysis has shown that a single amino acid substitution can abolish their IgE binding properties (Burks et al., 1997). Another relevant vicilin is the soybean alpha subunit of the globulin β-conglycinin designated as Gly m Bd 60 K (76 kD). The compound Gly m Bd 28K is a vicilin-like glycoprotein of MW 26 kD, a minor component fractionated from the 7S globulin fraction (Ogawa et al., 2000). On the other hand, Len c 1 is an allergen isolated and characterized from boiled lentils with a MW from 12 to 16 kD and corresponds to a gamma-vicilin subunit (Sánchez-Monge et al., 2000). Vicilins with allergenic properties have been identified in seeds such as walnut (Juglans regia) (Teuber et al., 1999), cashew (Anacardium occidentale) (Wang et al., 2002) and sesame (Sesamum indicum) (Beyer et al., 2002). Allergenic legumins: Ara h 4 from peanut is the N-terminal fraction of a 57 kD MW glycinin subunit, with a MW of 14 kD (Kleber-Hanke et al., 1999). Linear IgE epitopes have been mapped and mutational analysis has decreased IgE binding by between 35 and 85 %, retaining its ability to stimulate T cell activation. The cDNA clone of Ara h 4 encodes a protein with 70 % homology to glycinins. Soybean glycinin possesses five subunits, all of which react with IgE from soybean-allergic patients (Helm et al., 2000). Other allergenic globulins are found in hazelnut (Corylus avellana), coconut (Cocos nucifera) walnut (Teuber & Peterson, 1999) and almond (Prunus amygdalus) (Pasini et al., 2000). Prolamin superfamily 2S-albumin allergens: Three relevant allergens from peanut belong to the conglutin family, related to 2S albumins. The compound Ara h 2 is recognized by serum IgE from more than 90 % of peanut-allergic patients, and ten linear IgE-binding epitopes have been identified (Stanley et al., 1997). Ara h 2 also has trypsin inhibitor functions, enhanced by roasting. Mutational analysis shows a loss of IgE binding with single amino acid substitutions (Stanley et al., 1997). Two isoforms of Ara h 2 have been identified, the second being the most potent allergen (Chatel et al., 2003). The other two allergens from peanut are Ara h 6 of 14.5 kD MW, which is recognized by 38 % of sera from peanut-allergic subjects and Ara h 7, 15.8 kD MW recognised by 43 % (Kleber-Hanke et al., 1999). Other 2S albumin allergens are found in mustard (Brassica juncea) (Monsalve et al., 1993), oilseed rape (Brassica napus) (Monsalve et al., 1997), Brazil nut (Bertholletia excelsa) (Nordlee et al., 1996), walnut (Teuber et al., 1999), sunflower (Helianthus annus) seed (Kelly et al., 2000), cashew nut (Teuber et al., 2002) and sesame seed (Pastorello et al., 2001). Lipid transfer proteins: Among this group of proteins major allergens are found in several Rosaceae fruits, particularly from the genus Prunus (Pastorello et al., 2001). An allergen of 8 kD MW from soybean hulls was also identified (Gly m 1). This allergen is identical with the hydrophobic protein of soybean (HPS), a member of a cystine-rich protein family with the same pattern of cystine residues as LTP (Baud et al., 1993). Other allergenic proteins Several allergenic proteins not included in the seed-storage group have been identified. They are not as numerous as in other plant species, but several have been identified in legumes.

167
PR-10 proteins: The allergen of this plant-defence family, birch (Betula spp) pollen, Bet v 1 was the first allergenic PR-10 homologue identified. After that, many allergens, which are homologous to Bet v 1 have been described in several fruits of the Rosaceae and in other vegetables and nuts. Recently, SAM22, a Bet v 1 homologue, has been characterized in soybean (Kleine-Tebbe et al., 2002). Significant IgE binding to SAM22 occurred in 17 out of 20 patients allergic to birch pollen, apple (Malus sylvestris) or hazelnut. They also reacted clinically to soy oral challenges for the first time. Profilins: These are actin-binding proteins, with a MW 12-15 kD, of which the first allergen, Bet v 2 was identified in birch pollen. They are ubiquitous. Profilin allergens are found in many fruits and in vegetables. Recombinant allergens Ara h 5 (Kleber-Hanke et al., 1999) and Gly m 3 (Rihs et al., 1999) have been produced from cDNAs coding for profilins from peanut and soybean respectively. Recombinant Gly m 3 is recognized by IgE in 69 % of sera, but not its fragments. This indicates that the integrity of the molecule conformation is needed to retain its antigenic properties. Clinical allergy to legumes Several studies with children have shown a broad immunologic reactivity (IgE antibodies) to several legumes, positive results of skin testing and determination of legume-specific IgE, in allergic patients, which correlate poorly with the outcome of oral challenges (Bernhisel-Broadbent & Sampson, 1989). This was particularly so when an objective procedure such as the double blind, placebo-controlled food challenge method (DBPCFC) was used. Since most information came from paediatric studies performed in North America, which were focused on children allergic to peanut and/or soybean, we sought to evaluate legume allergy in adults, considering a scenario where legumes other than peanut may be relevant.
We analysed a case-series of 34 adult patients diagnosed in the Hospital Universitario 12 de Octubre, in Madrid who showed clinical allergy to one or more members of the legume family. Their median age was 29.5 years (range 16 - 71 years) with a male/female ratio of 0.4. The diagnostic procedure included a clinical questionnaire, skin testing, specific serum IgE assessment by the CAP-FEIA System (Pharmacia, Uppsala, Sweden), and challenge testing. Oral tolerance to legumes and other foods was evaluated by the DBPCFC procedure (Bock et al., 1988). Reactions reported to occur on inhalation in occupational or domestic settings were documented by inhalation specific challenges. Contact urticaria was documented by epicutaneous testing.
Overall, 45 reactions were considered positive in the 34 patients allergic to legumes (Table 1). The majority of reactions were in response to peanut (20 patients), followed by chickpea (5 patients), lentil (4 patients), and green bean (4 patients). Table 2 lists the symptoms of the reactions to the most common legumes. Interestingly, 7 reactions were caused by inhalation of the aerosolised legume (raw green bean, 3; grasspea (Lathyrus sativus) flour, 2; and lupin flour 2). The grass pea and lupin-reactive patients also had clinical symptoms on ingestion of the implicated legumes. The raw green bean-reactive patients tolerated ingestion of boiled green beans.
Peanut-reactive patients showed limited clinical reactivity to other members of the legume family, only 2 out of the 20 (10 %) peanut-allergic subjects reacted to other legumes (Table 3). Conversely, most patients allergic to chickpeas (80 %) were clinically reactive to other legumes, particularly lentil. No patients reacted exclusively to lentils, pea or broad bean (Vicia faba). Overall, the clinical cross-reactivity among members of the legume family seems to be lower than in other botanical families. In this study, only 6 out of 32 legume-

168
Table 1. Relative frequency of different legumes causing 45 reactions in 34 patients. Legume Number of reactions Proportion (%)
Broad bean 1 2 Chickpea 5 11 Grass pea 33 6 Green beans 41,2 9 Lentil 4 9 Lupin (Lupinus albus) 24 4 Pea 1 2 Peanut 20 45 Soybean 2 4 White bean 3 6 1Three reactions on inhalation; 2One reaction on contact; 3One reaction only on ingestion, one reaction only on inhalation, one reaction both on ingestion and inhalation; 4Both reactions on inhalation and ingestion. Table 2. Symptoms of reactions to the most common legumes causing food allergy in 34
patients. Bean Symptoms Peanut Chickpea Lentil Green White
Grass Pea
Oral 13 5 2 2 1 Cutaneous 2 1 1 Gastrointestinal 1 1 1 Respiratory 1 3 1 Generalised anaphylaxis 6 1 2 2 Table 3. Associated legume allergies in patients allergic to peanut and chickpea.
Legume Peanut-allergic patients (n = 20)
Chickpea-allergic patients (n = 5)
Peanut - 0 Chickpea 0 - Lentil 0 4 Green beans 0 0 White bean 0 2 Grass pea 1 0 Soybean 1 0 Lupin 0 0 Pea 0 1 Broad bean 0 1
Total 2 8 allergic patients reacted to more than one legume species, i.e. clinical cross-reactions were only present in 18.8 % of the patients. This contrasts with previous studies, which evaluated interspecies clinical cross-reactivity in legumes. Bernhisel-Broadbent & Sampson (1989) evaluated the cross-allergenicity in legumes in children with food hypersensitivity. In this study only 2 out of 41 (5 %) children were allergic to more than one legume (peanut and soybean). Bock & Atkins (1989) studied 32 children with positive DBPCFC to peanut. Each patient was skin tested with pea and soybean. Twenty-two patients had a positive skin test to soybean or pea, but only two (6 %) patients had positive double-blind challenges (one to pea and one to soybean). The reason for these differences could be related to the study design,

169
selection criteria, and the legumes evaluated, as chickpea and lentil were not included in previous studies, which were performed in the USA. Generally, the rate of cross-reactions in legume-allergic patients seems to be lower than in other botanical families. Rodriguez et al. (2000) evaluated clinical cross-reactivity among foods of the family Rosaceae in 34 adult patients complaining of adverse reactions. Peach (Prunus persica) was the most common offender and clinical cross-reactivity of peach with other Rosaceae was 46 %.
In summary, IgE-mediated reactions are the most common immunologic disorders following legume ingestion. These foods represent an eloquent example of the effect of a staple diet on the development of food allergies through ingestion. Most allergens so far described as causing allergic reactions to legume seeds in humans are storage proteins. In Spain, lentil is the legume most commonly implicated in IgE-mediated food hypersensitivity in children. However, in adult patients the majority of reactions are observed in response to peanut ingestion. In addition, occupational exposure to some airborne legume allergens could be relevant in eliciting IgE-mediated respiratory allergy through inhalation. Peanut-reactive adult patients have limited clinical reactivity to other members of the legume family, while most patients allergic to chickpea are also clinically reactive to other legumes, particularly lentil. Overall, the clinical cross-reactivity among members of legume family in adults seems to be lower than in other botanical families. However, adult patients allergic to peanut show extensive clinical reactivity to other foods, particularly peaches and tree nuts (Table 4). Further studies are needed to determine the immunologic basis of this multiple reactivity and to developed clinical guidelines to provide, with proper elimination, therapeutic diets in these patients. Table 4. Associated non- legume food allergies in 20 peanut-allergic patients.
Food Number of reactions Proportion (%) Peach 8 40 Almond 6 30 Walnut 6 30 Hazelnut 5 25 Chestnut 3 15 Apple 2 10 Grape 2 10 Tomato 2 10 Honey 2 10 Melon, Kiwi, Cherry Apricot, Banana, Pear, Barley 1 each 5 each
In conclusion, increased intestinal excretion of endogenous proteins and loss of N
through the urine, changes in plasma amino acids concentration, modulation of plasma lipids and cholesterolemia, and adverse immune reactions have been described in animals and humans consuming legume- or legume protein-based diets. The chemical structure of these particular groups of proteins is probably the ultimate cause of their nutritional and/or physiological behaviour. Therefore, the study of the mechanisms involved in the effects will hopefully open new ways to the elimination of undesirable effects and/or the utilisation of new biologically useful dietary components.

170
References Al-Muhsen, S., Clarke, A., & Kagan, R. 2003. Peanut allergy: an overview. Canadian
Medical Association Journal 168, 1279-1282. Aubry, M. & Boucrot, P. 1986. Etude comparée de la digestion des viciline et lectine
radiomarquées de Pisum sativum chez le rat. Annals in Nutrition and Metabolism 30, 175-182.
Batterham, E.S., Andersen, L.M., Lowe, R.F. & Darnell, R.E. 1986. Nutritional value of lupin (Lupinus albus)-seed meal for growing pigs: availability of lysine, effect of autoclaving and net energy content. British Journal of Nutrition 56, 645-659.
Baud, F., Pebay-Peyroula, E., Cohen-Addad, C., Odani, S., & Lehmann, M.S. 1993. Crystal structure of hydrophobic protein from soybean; a member of a new cystine-rich family. Journal of Molecular Biology 231, 877-887.
Bernhisel-Broadbent, J., & Sampson, H.A. 1989. Cross-allergenicity in the legume botanical family in children with food hypersensitivity. Journal of Allergy and Clinical Immunology 83, 435-440.
Beyer, K., Bardina, L., Grishina, G. & Sampson, H.A. 2002. Identification of sesame seed allergens by 2-dimensional proteomics and Edman sequencing: seed storage proteins as common food allergens. Journal of Allergy and Clinical Immunology 110, 154-159.
Beynen, A.C. 1990. Comparison of the mechanisms proposed to explain the hypocholesterolemic effect of soybean protein versus casein in experimental animals. Journal of Nutritional Science and Vitaminology 36, S87-S93.
Bloch, K.J., Wright, J.A., Bishara, S.M. & Bloch, M.B. 1988. Uptake of polypeptide fragments of proteins by rat intestine in vitro and in vivo. Gastroenterology 95, 1272-1278.
Bock, S.A. & Atkins, F.M. 1989. The natural history of peanut allergy. Journal of Allergy and Clinical Immunology 83, 900-904
Bock, S.A., Sampson, H.A., Atkins, F.M., Zeiger, R.S., Lehrer, S., Sachs, M., Bush, R.K. & Metcalfe D.D. 1988. Double-blind, placebo-controlled food challenge (DBPCFC) as an office procedure: a manual. Journal of Allergy and Clinical Immunology 82, 986-997.
Burks, A.W., Shin, D., Cockrell, G., Stanley, J.S., Helm, R.M. & Bannon, G.A. 1997. Mapping and mutational analysis of the IgE-binding epitopes on Ara h 1, a legume vicilin protein and a major allergen in peanut hypersensitivity. European Journal of Biochemistry 245, 334-339.
Cenarruzabeitia, M.N., Santidrián, S., Bello, J. & Larralde, J. 1979. Effect of raw field bean (Vicia faba) on amino-acid-degrading enzymes in rats and chicks. Nutrition and Metabolism 23, 203-210.
Chatel, J.M., Bernard, H. & Orson, F.M. 2003. Isolation and characterization of two complete Ara h 2 isoforms cDNA. International Archives of Allergy and Immunology 131, 14-18.
Chiji, H., Harayama, K. & Kiriyama, S. 1990. Effects of feeding rats low protein diets containing casein or soy protein isolate supplemented with methionine or oligo-L-methionine. Journal of Nutrition 120, 166-171.
Crespo, J.F., Pascual, C., Burks, A.W., Helm, R.M. & Esteban, M.M. 1995. Frequency of food allergy in a pediatric population from Spain. Pediatric Allergy and Immunology 6, 39-43.
Crespo, J.F., Rodriguez, J., Vives, R., James, J.M., Reano, M., Daroca, P., Burbano, C., & Muzquiz, M. 2001. Occupational IgE-mediated allergy after exposure to lupin seed flour. Journal of Allergy and Clinical Immunology 108, 295-297.

171
Dréau, D., Lallès, J.P., Philouze-Rome, V., Toullec, R. & Salmon, H. 1994. Local and systemic immune responses to soybean protein ingestion in early-weaned pigs. Journal of Animal Science 72, 2090-2098.
Eggum, B.O. 1970. Blood urea measurement as a technique for assessing protein quality. British Journal of Nutrition 24, 983-988.
Esparza, M.L., Larralde, J. & Martinez, J.A. 1996. Development-related nutritional utilization and immune function in mice fed on two different protein sources. Nutrition Research 16, 829-839.
Foucard, T. & Malmheden, Y.I. 1999. A study on severe food reactions in Sweden-- is soy protein an underestimated cause of anaphylaxis? Allergy 54, 261-265.
Friedrich, M., Noack, J., Proll, J. & Noack, R. 1984. Absorption of enzymatic protein hydrolysates and equimolar amino acid mixtures in the perfused small intestine of the rat. Biomed Biochimica Acta 43, 117-130.
Garcia Ortiz, J.C., Lopez-Asunsolo, A., Cosmes, P. & Duran, A.M. 1995. Bronchial asthma induced by hypersensitivity to legumes. Alergologia et Inmunopatologia (Madrid) 23, 38-40.
Grundy, J., Matttews, S., Bateman, B., Dean, T. & Arshad, S. 2002. Rising prevalence of allergy to peanut in children: data from 2 sequential cohorts. Journal of Allergy and Clinical Immunology 110, 784-789.
Hajós, G., Gelencsér, E., Grant, G., Bardocz, S., Sakhri, M., Duguid, T., Newman, A.M. & Pusztai, A. 1996. Effect of proteolyic modification and methionine enrichment on the nutritional value of soy albumins for rats. Journal of Nutritional Biochemistry 7, 481-487.
Hara, H. & Kiriyama, S. 1991. Absorptive behaviour of oligo-L-methionine and dietary proteins in a casein or soybean protein diet: porto-venous differences in amino acid concentrations in unrestrained rats. Journal of Nutrition 121, 638-645.
Helm, R.M., Cockrell, G., Connaughton, C., Sampson, H.A., Bannon, G.A., Beilinson, V., Nielsen, N.C. & Burks, A.W. 2000. A soybean G2 glycinin allergen. 2. Epitope mapping and three-dimensional modelling. International Archives of Allergy and Immunology 123, 213-219.
Horigome, T. & Cho, Y.S. 1992. Dietary casein and soybean protein affect the concentrations of serum cholesterol, triglycerides and free amino acids in rats. Journal of Nutrition 122, 2273-2282.
Huisman, J., Heinz, Th., Poel, A.F.B. van-der, Leeuwen, P. van, Souffrant, W.B. & Verstegen, M.W.A. 1992. True protein digestibility and amounts of endogenous protein measured with the 15N-dilution techniques in piglets fed on peas (Pisum sativum) and common beans (Phaseolus vulgaris). British Journal of Nutrition 68, 101-110.
Iritani, N., Nagashima, K., Fukuda, H., Katsurada, A. & Tanaka, T. 1986. Effects of dietary proteins on lipogenic enzymes in rat liver. Journal of Nutrition 116, 190-197.
Kalogeromitros, D., Armenaka, M., Galatas, I., Capellou, O. & Katsarou, A. 1996. Anaphylaxis induced by lentils. Annals of Allergy Asthma and Immunology 77, 480-482.
Kanny, G., Moneret Vautrin, D.A., Flabbeed, J., Beaudouin, E., Morisset, M. & Thevenin, F. 2001. Population study of food allergy in France. Journal of Allergy and Clinical Immunology 108, 133-140.
Kelly, J.D., Hlywka, J.J. & Hefle, S.L. 2000. Identification of sunflower seed IgE-binding proteins. International Archives of Allergy and Immunology 121, 19-24.

172
Kingman, S.M., Walker, A.F., Low, A.G., Sambrook, I.E., Owen, R.W. & Cole, T.J. 1993. Comparative effect of four legume species on plasma lipids and faecal steroid excretion in hypercholesterolemic pigs. British Journal of Nutrition 69, 409-421.
Kleber-Hanke, T., Crameri, R., Appenzeller, U., Schlaak, M. & Becker, W.M. 1999. Selective cloning of peanut allergens, including profilin and 2S albumins, by phage display technology. International Archives of Allergy and Immunology 119, 265-274.
Kleine-Tebbe, J., Vogel, L., Crowel, D.N., Haustein, U.F. & Vieths, S. 2002. Severe oral allergy syndrome and anaphylactic reactions caused by a Bet v 1-related PR-10 protein in soybean, SAM22. Journal of Allergy and Clinical Immunology 110, 797-804.
Lallès, J.P. 1993. Nutritional and antinutritional aspects of soybean and field pea proteins used in veal calf production, a review. Livestock Production Science 34, 181-202.
Lallès, J.P., Dréau, D., Féménia, F., Parodi, A.L. & Toulec, R. 1996. Feeding heated soyabean flour increases the density of B and T lymphocytes in the small intestine of calves. Veterinary Immunological Immunopathology 52, 105-115.
Lallès, J.P. & Peltre, G. 1996. Biochemical features of grain legume allergens in humans and animals. Nutrition Reviews 54, 101-107.
Lallès, J.P., Salmon, H., Bakker, N.P.M. & Tolman, G.H. 1993. Effects of dietary antigens on health, performance and immune system of calves and piglets. In Recent Advances of Research in Antinutritional Factors in Legume Seeds. A.F.B. van der Poel, J. Huisman & H.S. Saini (Eds). Proceedings 2nd International Workshop on Antinutritional Factors in Legume Seeds, Wageningen, 1-3 December 1993, 253-270.
Li, D.F., Nelssen, J.L., Reddy, P.G., Blecha, F., Klemm, R.D., Giesting, D.W., Hancock, J.D., Allee, G.L. & Goodband, R.D. 1991. Measuring suitability of soybean products for early-weaned pigs with immunological criteria. Journal of Animal Science 69, 3299-3307.
Lovati, M.R., Manzoni, C., Corsini, A., Granata, A., Frattini, R., Fumagalli, R. & Sirtori, C.R. 1992. Low density lipoprotein receptor activity is modulated by soybean globulins in cell culture. Journal of Nutrition 122, 1971-1978.
Martin, J.A., Compaired, J.A., Hoz, de la, B., Quirce, S., Alonso, M.D., Igea, J.M. & Losada, E., 1992. Bronchial asthma induced by chickpea and lentil. Allergy 47, 185-187.
Miller, B.G., Newby, T.J., Stokes, C.R., Hampton, D. & Bourne, F.J. 1983. The role of dietary antigen in the aetiology of post weaning diarrhoea. Annales de Recherche Veterinaire 14, 487-492.
Nordlee, J.A., Taylor, S.L., Townsend, J.A., Thomas, L.A. & Bush, R.K. 1996. Identification of a Brazil-nut allergen in transgenic soybeans. New England Journal of Medicine 14, 688-692.
Moneret-Vautrin, D.A., Rance, F., Kanny, G., Olsewski, A., Gueant, J.L., Dutau, G. & Guerin L. 1998. Food allergy to peanuts in France--evaluation of 142 observations. Clinical Experimental Allergy 28, 1113-1119.
Monsalve, R.I., Gonzalez de la Pena, M.A., Lopez-Otin, C., Fiandor, A., Fernandez, C.,Villalba, M. & Rodriguez, R. 1997. Detection, isolation and complete amino acid sequence of an aeroallergenic protein from rapeseed flour. Clinical Experimental Allergy 27, 833-841.
Monsalve, R.I., Gonzalez de la Pena, M.A., Menendez-Arias, L., Lopez-Otin, C.,Villalba, M. & Rodriguez, R. 1993. Characterization of a new oriental-mustard (Brassica juncea) allergen, Bra j IE: detection of an allergenic epitope. The Biochemical Journal 293,625-632.

173
Nielsen, K., Kondrup, J., Elsner, P., Juul, A. & Jensen, E.S. 1994. Casein and bean-bean protein have different effects on whole body protein turnover at the same nitrogen balance. British Journal of Nutrition 72, 69-81.
Noseda, G. & Fragiacomo, C. 1980. Effects of soybean protein diet on serum lipids, plasma glucagon and insulin. Diet and drugs in Atherosclerosis 61-65.
Ogawa, A., Samoto, M. & Takahashi, K. 2000. Soybean allergens and hypoallergenic soybean products. Journal of Nutritional Science and Vitaminology (Tokyo) 46, 271-279.
Pascual, C.Y., Fernandez Crespo, J., Sanchez Pastor, S., Padial, M.A., Díaz Pena, J.M., Martín Muñoz, F. & Martín Esteban, M. 1999. Allergy to lentils in Mediterranean pediatric patients. Journal of Allergy and Clinical Immunology 103, 154-158.
Pasini, G., Simonato, B., Giannattasio, M., Gemignani, C. & Curioni, A. 2000. IgE binding to almond proteins in two CAP-FEIA-negative patients with allergic symptoms to almond as compared to three CAP-FEIA-false-positive subjects. Allergy 55, 955-958.
Pastorello, E.A., Pompei, C., Pravettoni, V., Brenna, O., Farioli, L., Trambaioli, C. & Conti, A. 2001. Lipid transfer proteins and 2S albumins as allergens. Allergy 56, Supplement 67,45-47.
Patil, S.P., Niphadka, P.V. & Bapat, M.M. 2001. Chickpea: a major food allergen in the Indian subcontinent and its clinical and immunochemical correlation. Annals of Allergy Asthma and Immunology 87, 140-145.
Pedersen, H.E., Sissons, J.W., Turvey, A. & Sondergaard, I. 1984. Histologic and metabolic responses to bean-bean antigens in the gut of the preruminant calf. Proceedings of the Nutrition Society 43, 114A.
Rihs, H.P., Chen, Z., Rueff, F., Petersen, A., Rozynek, P., Heimann, H. & Baur, X. 1999. IgE binding of the recombinant allergen soybean profilin (rGly m 3) is mediated by conformational epitopes. Journal of Allergy and Clinical Immunology 104, 1293-1301.
Rodriguez, J., Crespo, J.F., Lopez-Rubio, A., Cruz-Bertolo, J., de la, Ferrando-Vivas, P., Vives, R., & Daroca, P. 2000. Clinical cross-reactivity among foods of the Rosaceae family. Journal of Allergy and Clinical Immunology 106, 183-189.
Rubio, L.A., Grant, G., Bardocz, S., Dewey, P. & Pusztai, A. 1991. Nutritional response of growing rats to faba beans (Vicia faba) and faba bean fractions. British Journal of Nutrition 66, 533-542.
Rubio, L.A., Grant, G., Cavallé, C., Martinez-Aragón, A. & Pusztai, A. 1994. High in vivo (rat) digestibility of faba bean (V. faba), lupin (L. angustifolius) and soybean (G. max) globulins. Journal of the Science of Food and Agriculture 66, 289-292
Rubio, L.A., Grant, G., Daguid, T., Brown, D., Bardocz, S. & Pusztai, A. 1998. The nutritional utilization by rats of chickpea (Cicer arietinum) meal and its isolated globulin proteins is poorer than that of defatted soybean or lactalbumin. Journal of Nutrition 128, 1042-1047.
Rubio, L.A., Grant, G., Daguid, T., Brown, D. & Pusztai, A. 1999. Organ relative weights and plasma amino acid concentrations in rats fed diets based in legume (faba bean, lupin, chickpea, soybean) seed meals or their fractions. Journal of the Science of Food and Agriculture 79, 187-194.
Rubio, L.A., Grant, G., Scislowsky, P., Brown, D., Annand, M. & Pusztai, A. 1995. The utilization of lupin (L. angustifolius) and faba bean globulins by rats is poorer than of soybean globulins or lactalbumin but the nutritional value of lupin seed meal is lower only than that of lactalbumin. Journal of Nutrition 125, 2145-2155.

174
Rubio, L.A. & Seiquer, I. 2002. Transport of amino acids from in vitro digested legume proteins or casein in Caco-2 cell cultures. Journal of Agricultural and Food Chemistry 50, 5202-5207.
Sanchez-Monge, R., Pascual, C.Y., Diaz-Perales, A., Fernandez-Crespo, J., Martin-Esteban, M. & Salcedo, G. 2000. Isolation and characterization of relevant allergens from boiled lentils. Journal of Allergy and Clinical Immunology 106, 955-961.
Sampson, H.A. 2002. Peanut Allergy. The New England Journal of Medicine 346, 1294-1299. Santoro, L.G., Grant, G. & Pusztai, A. 1997. Effects of short-term feeding of rats with a
highly purified phaseolin preparation. Plant Foods in Human Nutrition 51, 61-70. Sholz-Ahrens, K., Hagemeister, H., Unshelm, J., Agergaard, N. & Barth, C.A. 1990. Response
of hormones modulating plasma cholesterol to dietary casein or soy protein in minipigs. Journal of Nutrition 120, 1387-1392.
Sicherer, S.A., Furlong, T.J., Simone, J. de, & Sampson, H.A. 1999. Self-reported allergy to peanut in commercial airliners. Journal of Allergy and Clinical Immunology 104, 186-189.
Sirtori, C.R., Pazzucconi, F., Colombo, L., Battistin, P., Bondioli, A. & Descheemaeker, K. 1999. Double-blind study of he addition of high-protein soy milk v. cow´s milk to the diet of patients with severe hypercholesterolaemia and resistance to or intolerance of statins. British Journal of Nutrition 82, 91-96.
Sissons, J.W. 1989. Aetiology of diarrea in pigs and pre-ruminant calves. In Recent Advances in Animal Nutrition,. W. Haresign W & D.J.A. Cole (Eds). Butterwoths, London. pp 261-282
Sissons, J.W., Pedersen, H.E. & Thurston, S.M. 1984. Soya-bean antigen survival in the digestive tract of the preruminant calf. Proceedings of the Nutrition Society 43, 115A.
Stanley, J.S., King, N., Burks, A.W., Huang, S.K., Sampson, H., Cockrell, G., Helm, R.M., West, C.M. & Bannon, G.A. 1997. Identification and mutational analysis of the immunodominant IgE binding epitopes of the major peanut allergen Ara h 2. Archives of Biochemistry and Biophysics 342, 244-253.
Sugano, M., Ishiwaki, N., Nagata, Y. & Imaizumi, K. 1982. Effects of arginine and lysine addition to casein and bean-bean protein on serum lipids, apolipoproteins, insulin and glucagon in rats. British Journal of Nutrition 48, 211-221.
Sugano, M., Yamada, Y., Yoshida, K., Hashimoto, Y., Matsuo, T. & Kimoto, M. 1988. The hypocholesterolemic action of the undigested fraction of soybean protein in rats. Atherosclerosis 72, 115-122.
Tariq, S.M., Stevens, M., Matthews, S., Ridout, S., Twiselton, R. & Hide, D.V. 1996. Cohort study of peanut and tree nut sensitization by age of 4 years. British Medical Journal 313, 514-517.
Tarwid, J.N., Morrall, R.A. & Mills, J.H. 1985. Effects of feeding Ascochyta-infected and normal lentils to rats (short-term study). Canadian Journal of Compared Medicine 49, 409-413.
Teuber, S.S. & Peterson, W.R. 1999. Systemic allergic reaction to coconut (Cocos nucifera) in 2 subjects with hypersensitivity to tree nut and demonstration of cross-reactivity to legumin-like seed storage proteins: new coconut and walnut food allergens. Journal of Allergy and Clinical Immunology 103, 1180-1185.
Teuber, S.S., Sathe, S.K., Peterson, W.R. & Roux, K.H. 2002. Characterization of the soluble allergenic proteins of cashew nut (Anacardium occidentale L.). Journal of Agricultural and Food Chemistry 50, 6543-6549.
Thanh, V.H., Okubo, K. & Shibasaki, K. 1975. Isolation and characterization of the multiple 7S globulins of soybean proteins. Plant Physiology 56, 19-22.

175
Van der Meer, R. 1983. Is the hypocholesterolemic effect of dietary casein related to its phosphorylation state? Atherosclerosis 49, 339-341.
Van der Poel, A.F.B., Dellaert, L.M., Norel, A. van & Helpster, J.P.F.G. 1992. The digestibility in piglets of faba bean (Vicia faba L.) as affected by breeding towards the absence of condensed tannins. British Journal of Nutrition 68, 793-800.
Wang, Y.H. & McIntosh, G.H. 1996. Extrusion and boiling improve rat body weight gain and plasma cholesterol lowering ability of peas and chickpeas. Journal of Nutrition 126, 3054-3062
Wang, F., Robotham, J.M., Teuber, S.S., Tawde, P., Sathe, S.K. & Roux, K.H. 2002. Ana o 1, a cashew (Anacardium occidentale) allergen of the vicilin seed storage protein family. Journal of Allergy and Clinical Immunology 110, 160-166.
Webb, K.E. 1990. Intestinal absorption of protein hydrolysis products: a review. Journal of Animal Science 68, 3011-3022.
West, C.E., Spaaij, C., Clous, W.M., Twisk, H.P., Goertz, M.P.H., Hubbard, R.W., Kuyvenhoven, M.W., Meer, R. van der, Roszkowski, W.F., Sanchez. A. & Beynen, A.C. 1989. Comparison of the hypocholesterolemic effects of dietary soybean protein with those of formaldehyde-treated casein in rabbits. Journal of Nutrition 119, 843-856.
Zhang, X. & Beynen, A.C. 1993. Lowering effect of dietary milk-whey protein v. casein on plasma and liver cholesterol concentrations in rats. British Journal of Nutrition 70, 139-146.


177
Morphological changes of rat brain neurons after intracranial administration of the alkaloid gramine A. Hernández-Torres1, J. Bañuelos-Pineda2 P.M. García-López3, M.A. Ruiz-López3 & P. Garzón de la Mora1 1Instituto de Enfermedades Crónico-Degenerativas, Departamento de Fisiología, Centro Universitario de Ciencias de la Salud, Sierra mojada No. 950, Colonia Independencia C.P. 44340, Guadalajara, Jalisco, México 2Laboratorio de Neuromorfología, Departamento de Medicina Veterinaria, Centro Universitario de Ciencias Biológicas y Agropecuarias, Universidad de Guadalajara, Carretera Guadalajara-Nogales Km 15.5, Zapopan, Jalisco, México 3Laboratorio de Biotecnología, Departamento de Botánica y Zoología, Centro Universitario de Ciencias Biológicas y Agropecuarias, Universidad de Guadalajara, Carretera Guadalajara-Nogales Km 15.5, A.P. 1-139 Zapopan, Jalisco, México Summary The effect of a daily intracranial dose of the alkaloid gramine (3-dimethyl-aminomethyl indole) on brain neuron morphology was evaluated in Swiss Wistar rats. Rats were fitted with a cranial osteal cannula by stereotaxic surgery and treated for five consecutive days with 2, 3, 5 or 10 µg/d of gramine. The intracranial administration of gramine 2 to 5 µg/d had no apparent toxic or behavioural effect. However, animals treated with 10 µg of gramine/d became agitated, developed hirsutism and caudal erection.
Histopathological analysis of brains from experimental animals showed an increase in the number of neurons with eosinophilia. The numbers of neurons with eosinophilia increased as the gramine dose increased from 2 to 10 µg/d. Neurons with eosinophilia and cavitation were observed in the deep stratus of the frontal and frontoparietal cortex, deep stratus of the posterior cingular cortex, several thalamic nuclei, putamen, caudate nucleus, lateral and medial hypothalamus. From these results it can be concluded that intracranial administration of gramine caused histopathological changes in rat brain neurons. Keywords: morphological changes, neurons, gramine, indole alkaloids, lupin Introduction Although the quinolizidine alkaloids are the main alkaloids present in the genus Lupinus the tryptophan derived alkaloid gramine (3-dimethyl-aminomethyl indole) has recently been reported in Lupinus luteus and L. hartwegii (Schmeller & Wink, 1998; Pastuszewska et al., 2001).
Quinolizidine alkaloids exert physiological effects on the central nervous, circulatory and respiratory systems including hypotension, hypoglycemia, diuretic and anti-inflammatory responses. Gramine has also been reported to affect the central nervous, circulatory and respiratory systems (Schmeller & Wink, 1998; Pastuszewska et al., 2001). Gramine has also been shown to be a serotonin antagonist and an acetylcholinesterase and butyrilcholinesterase inhibitor (Avery & Horvitz, 1990; Katz & Frost, 1995).
Dietary administration of gramine to experimental animals has given inconsistent effects on body composition and feed intake. Feeding 0.67 g/kg body weight and 0.38 g/kg

178
body weight decreased weight gains in broiler chickens and rats, respectively (Pastuszewska et al., 2001). However, feeding pigs with a yellow lupin (L. luteus) based diet with a high gramine content had no adverse effects on feed intake, growth or performance (Wasilewko et al., 1997).
Since 1999 gramine has been used in Europe, as dietary supplement, to treat depression, epilepsy and Alzheimer disease. It is also used as a sedative, blood pressure modulator and for the treatment of nicotine withdrawal symptoms.
Oral administration of gramine to pigs, rats and broiler chickens did not cause any histopathological changes to the heart, kidneys, liver, or testicles. However, reductions in haemoglobin concentration and in white blood cells numbers were observed (Pastuszewska et al., 2001). Even though gramine appears to be a safe dietary supplement for the treatment of several diseases of the central nervous system, it is important to determine if this alkaloid causes morphological changes to brain neurons. The aim of this study was to determine possible histological changes in the rat brain after intraventricular cerebral administration of gramine. Materials and methods The effect of daily intracranial administration of the alkaloid gramine on the neuron morphology of the brain was evaluated with Swiss Wistar rats weighing 250 - 350 g. Forty-eight rats were divided into six groups of 8 animals each (4 male and 4 female) and randomly assigned to a control group (T0) or to one of 5 experimental groups (T1 to T5). During the experiment the rats were kept in a 12 h dark-light cycle (08:00 to 20:00 h), at 25 ± 1 °C and a relative humidity of 65 - 70 %. They had free access to food and water. Rats were housed in individual cages and were handled in accordance with the National Institute of Health guidelines for experimental animals (NIH Animal Care and Use Committee, 1992).
Gramine, or distilled water, was administered via a cannula inserted through the cranial osteal plates during stereotaxic surgery. Animals in the experimental groups were treated for five consecutive days with a daily dose (10 µl) of distilled water (T1), 2 (T2), 3 (T3), 5 (T4) or 10 (T5) µg of gramine in 10 µl of distilled water. Rats were placed in an open area (60 × 60 cm) for 30 min after gramine or water was administered and any behavioural changes were recorded. Two days after the last treatment, control and experimental animals were injected with 50 mg/kg of pentobarbital and perfused with a 0.9 % NaCl (2 min) and a Bowin (15 min) solution.
Brains were removed by craniotomy and were fixed by immersing them for 12 hr at 4 °C in a fixative solution. The brains were dehydrated in ethanol solutions of increasing concentrations (40 to 100 %) and set in a paraffin block. Brain coronal slices of 15 µm were taken from 4 brains in each group (2 of each sex), mounted in tissue slices and stained with haematoxylin-eosin. Sagital slices were taken of the remaining brains and they were stained in the same way. All brain slices were examined histologically to determine if changes in brain morphology had occurred. Results There were no apparent signs of toxicity or behavioural changes after the intracranial administration of distilled water (10 µl) or 2, 3 or 5 µg of gramine. Animals treated with 10 µg of gramine exhibited anxiety, excessive restlessness, grooming, as well as unsynchronised exploratory behaviour, hirsute hair and caudal erection.

179
Histological examination of the brain tissues of the experimental animals showed that administration of 10 µl of distilled water resulted in a small number of eosinophilic neurons in the deep cerebellar nuclei, cerebellum, dentate gyrus, hippocampus CA4 zone, diverse nuclei of the lateral cerebral stem, deep and superior layers of the frontal, frontoparietal cortex and in Purkinje cell regions (Fig. 1). The intracranial administration of gramine significantly increased the number of eosinophilic neurons. Eosinophilic neuron number increased with gramine dose.
A B C Figure 1. Midsagital brain tissue slides of animals treated with 10 µl distilled water.
(A - cerebellum Purkinje cell; B - dentate girus neurons; C - deep cerebellar nuclei neurons. Brain neurons shown in this slide display eosinophilia. Brain tissue slices were stained with haematoxylin-eosin and were viewed at 20X.
Histopathological evaluation of brain tissues from animals treated for 5 consecutive
days with 10 µg gramine in 10 µl of distilled water (T5) showed neuronal cavitation and neuronal necrosis along the thickness of the frontoparietal and frontal cortex, deep layers of the lateral cingular cortex, thalamicus nuclei, caudate-putamen, lateral and medial hypothalamus, septohippocampal nucleus, deep cerebellum nuclei and diverse nuclei of the dorsal and ventral stem (Fig. 2). Based on these results it can be concluded that daily intracranial administration of 10 µg of gramine causes severe damage in diverse neurons of the rat brain.
A B C Figure 2. Midsagital brain tissue slides of animal treated with 10 µg gramine in 10 µl of
distilled water. (A - Neurons of deep layers of the frontoparietal cortex; B - ventral stem nucleus neurons; C - Deep cerebellar nuclei. All neurons showed eosinophilia and cavitation. Brain tissue slices were stained with eosin-eosin (HE) and viewed at 20X.
Discussion The brain histopathological changes observed in this study are contrary to an earlier report, which indicated that the dietary admistration of gramine has no detrimental effect, other than a reduction in, feed intake (Pastuszewska et al., 2001). Long term studies are needed to establish how gramine damages brain neurons.

180
References Avery, L. & Horwitz, H.R. 1990. Effects of starvation and neuroactive drugs on feeding in
Caemorhabditis elegans. Journal of Experimental Zoology 253, 263-270. Goelz, M.F., Rothenbacher, H., Wiggins, J.P., Kendall, W.A. & Hershberg, T.V. 1980.
Some histopathological effects of the alkaloids gramine and hordenine on meadow voles (Microtus pennsylvanucus). Toxicology 18, 125-131.
Katz, P.S. & Frost W.N. 1995. Intrinsic nueromodulation in the Tritonia swim SPJ: serotonin mediated both neuromodulation and neurotransmission by the dorsal swim interneurons. Journal of Neurophysiology 74, 2281-2294.
NIH Animal Care and Use Committee. 1992. Using animals in intramural research: guidelines for investigators. NIH Animal Care and Use Committee (Eds). NIH Training Center, Md.
Pastuszewska, B., Smulikowska, S., Wasilewko, J., Buraczewska, L., Ochtabinska, A., Mieczkowska, A., Lechowskai, R. & Bielecki, W. 2001. Response of animal to gramine. I. Performance and selected hematological, biochemical and histological parameters in growing chicken rats and pigs. Archives of Animal Nutrition 55, 1-16.
Schmeller, T. & Wink, M. 1998. Utilization of alkaloids in modern medicine. In Alkaloids: Biochemistry, Ecology and Medicinal Applications. M.F. Roberts and M. Wink (Eds). Plenum Press, New York. pp. 435-459.
Wasilewko, J., Burraczewska, L., Lechowski, R. & Wysocka, W. 1997. Toxicological effects of lupin alkaloids in pigs. In Proceedings of the Conference Lupin in Modern Agriculture. Polish Lupin Society, Agricultural University Olsztyn, Olsztyn-Kortowo, 223-227.

181
Plasma antibody responses to grain legume proteins in weaned piglets J.P. Lallès1, P. Salgado1,2 & J.P.B. Freire2 1Institut National de la Recherche Agronomique INRA-UMRVP, 35590 Saint-Gilles, France 2Instituto Superior de Agronomia, Tapada de Ajuda, 1349-017 Lisboa, Portugal Summary Experiments were conducted to investigate legume immunogenicity in weaned piglets. Diets contained either casein (control) or an equal protein mixture of casein and raw pea (Pisum sativum), faba bean (Vicia faba), narrow-leafed lupin (Lupinus angustifolius), Kabuli and desi chickpea (Cicer arietinum) or soybean (Glycine max) meal as their protein source. After four weeks of feeding, jugular blood was collected and plasma analysed for anti-legume protein IgG antibodies. The ELISA antibody titres to crude legume protein extracts were higher in the legume-fed piglets than in control animals, except for pea. This was accounted for by higher titres to 7S, but not to 11S, legume seed storage proteins. Western blotting revealed many immunogenic proteins corresponding mostly to the 7S protein family. Minor chickpea proteins were found to be highly immunogenic. Overall faba bean and chickpea seed were the most immunogenic. Keywords: piglet, weaning, legume proteins, plasma IgG Introduction Soybean is a major protein source for animal feeding and is usually well digested. However, it is also known for its allergenicity in humans and for the immune-mediated gut disorders it induces in piglets and calves (Lallès & Peltre, 1996). Specific plasma antibodies were found in early-weaned piglets after feeding so-called antigenic soybean products (Lallès, 1999). These responses may be detrimental to the small intestine and affect pig performance in the first weeks after weaning.
Data on immune responses, in weaned piglet, to legume seeds, other than soybean, are scarce. Therefore, we conducted this study to investigate the plasma antibody responses to pea, faba bean, narrow-leafed lupin and chickpea seed using specific ELISA and western blotting. Materials and methods Animals, diets and blood sampling The experiments used 30 (8.7 ± 0.3 kg body weight (BW), Exp. 1) and 18 (8.2 ± 0.3 kg BW, Exp. 2) male piglets (Duroc × Landrace) weaned at 28 d. They were divided into 5 (Exp. 1) and 3 (Exp. 2) groups of 6 pigs on the basis of litter origin and average weaning BW. Piglets were fed (40 g/kg BW/d) one of 5 (Exp. 1) or 3 (Exp. 2) experimental diets from weaning. A control diet based on casein (C) as the protein source (Exp. 1 and 2), and diets in which 50 % of the crude protein (N × 6.25) was supplied by pea (P), faba bean (FB), narrow-leafed lupin (L) seed and soybean defatted meal (SBM) in Experiment 1, and Kabuli (KCP) or desi (DCP) chickpea seed in Experiment 2, were formulated. The legume protein sources differed in their chemical composition and their antinutritional factor content including trypsin inhibitors,

182
oligosaccharides, alkaloids and tannins. The diets contained 25 and 12.5 % (as fed) of casein in the control and legume-based diets, respectively, and 47.5, 39.6, 29.3, 49.1, 47.8 and 23.7 % of P, FB, L, KCP, DCP and SBM, respectively. After four weeks of feeding, blood samples were taken from the jugular vein and plasma was prepared and frozen (–20 ºC). Anti-legume protein antibody determination by ELISA A crude protein extract (CPE) was obtained from ground legume seeds by extraction in Tris-HCl pH 7.5 and centrifuging. The globulin fraction then served for the purifying of the legumin and vicilin protein families using FPLC (Melo et al., 1994).
Titres of plasma IgG specific for CPEs and purified globulins were determined using ELISA. This assay included plate coating with CPEs (1 mg/ml) or purified globulins (1 µg/ml) in carbonate buffer at pH 9.6, saturation with skim-milk powder, incubation of serial dilutions of pig plasma, incubation with a rabbit anti-pig IgG and then a goat anti-rabbit IgG peroxidase-labelled, and peroxidase activity measurement using ABTS 2-2’-azinobis 3-ethylbenz-thiazoline sulphonic acid as the substrate. Titres were expressed as the log of the highest doubling dilutions (initial dilution 1:20) showing an optical density twice that of the background. The ELISA antibody titres were compared by the Mann-Whitney’s test and differences were taken as significant at P < 0.10. Legume protein antigen repertoires studied by western blotting Crude protein extracts (40 µg) and purified globulins (10 µg) were separated by SDS-PAGE under reducing conditions. Molecular weight (MW) standards were also run. Separated proteins were transferred from the gels to nitrocellulose membranes. They were saturated with skim-milk powder, and incubated with pig plasmas (dilution 1:160), then a rabbit anti-pig IgG and finally a goat anti-rabbit IgG labelled with peroxidase. The IgG-antigen binding was revealed using diaminobenzidine and H2O2. Results Plasma IgG anti-legume protein antibody titres are shown in Table 1. Antibody titres to CPEs were significantly higher in legume-fed piglets than in the control animals for FB, L, KCP and DCP. These differences were accounted for by the antibody responses to vicilin (7S) proteins. Antibody titres to pea vicilin were significantly higher in pea-fed piglets than in control animals.
Plasmas from the legume-fed piglets with the highest ELISA titres were analysed by western blotting. Many protein bands of MW 94 to 14.4 kDa were identified (Fig. 1). In the case of pea, stained bands from the CPE did not appear to correspond with pea legumin polypeptides (Lane 11S, Fig. 1). In contrast, binding to pea vicilin proteins was shown at MWs of 72, 67 and 46 kDa (Lane 7S). In faba bean there was major staining between 75 and 43 kDa, with strong bands at 65 and 47 kDa (Lanes CPE and 7S). There were fainter bands between 37 and 20 kDa. Bands at 60, 37 and 24 kDa corresponded to faba bean legumin proteins (lane 11S). There was strong staining of four proteins of the faba bean vicilin family (Lane 7S). For narrow-leafed lupin, different bands between 72 and 45 kDa plus two bands at 29 and 27 kDa stained clearly (Lane CPE). There was faint staining of lupin vicilin family proteins (Lane 7S). There were many stained chickpea bands between 77 and 16 kDa, among which were seven strong bands in the Kabuli chickpea CPE. Proteins from KCP legumin did not appear to be stained (Lane 11S) and only one strong 7S band was observed. A staining

183
pattern close to that with KCP was observed in the plasma of piglets fed DCP (not shown). No staining was observed for SBM (not shown). Table 1. Median plasma antibody titres against CPEs, 11S and 7S proteins from different
legumes as determined by ELISA in controls (C) and legume-fed (Legume) piglets.
CPE 11S 7S Legume species C Legume C Legume C LegumePea 61 7ns 5 6ns 5 8* Faba bean 5 9* 6 9ns 5 8# Blue lupin 5 7# 6 8ns 6 9* Chickpea - Kabuli 6 9* 5 7ns 5 8* Chickpea - desi 6 8* 5 7ns 5 8# Soybean meal 5 5ns 6 6ns 5 6ns 1Titre expressed as log2 dilution from initial dilution 1:20. *, #, ns Difference from the corresponding control *P < 0.05, #P < 0.10, ns not significant.
Figure 1. Western blots of representative plasmas from piglets fed pea (P), faba bean (FB),
blue lupin (L) and Kabuli chickpea (KCP). Discussion Our ELISA data shows that piglets fed different raw legume seeds raised systemic antibodies consistently directed against the vicilin (or 7S) protein fraction. This is in line with the results of Seabra et al. (2001) with piglets fed on other legume seeds. The biochemical basis of such a biased antibody response towards 7S proteins is unclear because 7S and 11S globulins are structurally related. However, legumin and vicilin proteins have structural differences, which account for their specific biochemical, rheological, and probably immunological properties.
MW (kDa) 94 ▬ 67 ▬ 43 ▬ 30 ▬ 20.1 ▬
CPE 11S 7S
P FB WCPL
CPE 11S 7S CPE 11S 7S CPE 11S 7S

184
Western blotting confirmed proteins of the vicilin family were more immunogenic than the legumins. This was particularly true for faba bean, pea and narrow-leafed lupin. In contrast chickpeas exhibited a complex CPE staining pattern with only a few bands corresponding to 7S proteins. In our results bands stained against proteins of the legumin family were rare, or absent. Finally, western blotting showed immunogenicity of additional, non-storage globulins, especially in the chickpeas. Although some bands may be related to protease inhibitors, we did not attempt to identify them further. Seabra et al. (2001) concluded that piglets displayed antibodies against β-conglutin of Lupinus luteus and against vicilin of Vicia sativa and Lathyrus cicera. Apart from their study, and except for soybean and peanut (Arachis hypogaea), data on legume seed antigenic protein repertoires in animals are scarce. In humans, allergies were reported to pea proteins (many bands > 25 kDa; 20, 14 and 2kDa), lupin (55 to 35 kDa, 21 kDa) and chickpea (78 to 20 kDa and low MW proteins) (Lallès & Peltre, 1996; San Ireneo et al., 2000).
In conclusion, the proteins of pea, faba bean, narrow-leafed lupin, and chickpea induced systemic antibody responses in weaned piglets. Faba bean and chickpea were the most immunogenic species. Vicilin proteins were much more immunogenic than the legumin proteins. Finally the non-storage globulin proteins of chickpea were noticeable immunogens. Acknowledgement Financial support of a PRAXIS XXI scholarship of the Fundação para a Ciência e a Tecnologia and Instituto de Ciências Agrárias Mediterrânicas, Portugal is acknowledged. References Lallès, J.P. 1999. Soy products as protein sources for preruminants and young pigs. In Soy
in Animal Nutrition. J.K. Drakley (Ed.). Federation of Animal Science Societies, Savoy (USA). pp. 106-126.
Lallès, J.P. & Peltre, G. 1996. Biochemical features of grain legume allergens in humans and animals. Nutrition Reviews 54, 101-107.
Melo, T.S., Ferreira, R.B. & Teixeira, A.N. 1994. The seed storage proteins from Lupinus albus. Phytochemistry 37, 641-648.
San Ireneo, M.M., Sandin, M.D.I., Fernandez-Caaldas, E., Lizana, F.M., Fletes M.J.R. & Borrego, M.T.L. 2000. Specific IgE levels to Cicer arietinum (chickpea) in tolerant and nontolerant children: Evaluation of boiled and raw extracts. International Archives of Allergy and Immunology 121, 137-143.
Seabra, M., Carvalho, S., Freire, J., Ferreira, J., Mourato, M., Cunha, L., Cabral, F., Teixeira, A. & Aumaitre, A. 2001. Lupinus luteus, Vicia sativa and Lathyrus cicera as protein sources for piglets: ileal and total tract apparent digestibility of amino acids and antigenic effects. Animal Feed Science and Technology 89, 1-16.

185
The nutritional potential of Mexican piñon (Jatropha curcas). Toxic and antinutritional factors J. Martínez-Herrera1, L. Chel-Guerrero2 & A.L. Martínez-Ayala1 1Departamento de Biotecnología, Centro de Desarrollo de Productos Bióticos-IPN, AP. 24, Yautepec, Mor. 62731, México 2Facultad de Ingeniería Química, Universidad Autónoma de Yucatán, Mérida, Yucatán, México. Summary Jatropha curcas or Mexican piñon is a native Mexican plant from the family Euphorbiaceae. The seed has a high protein (27 - 32 %) and oil (58 - 60 %) content. Although the seed cake meal is rich in protein (50 %), it is toxic to rats, mice and ruminants and therefore, cannot be used as animal feed. Several cases of human poisoning have been recorded after accidental consumption of seed. However, in Mexico non-toxic lines of J. curcas have been found. No phorbol esters were found but trypsin inhibitor, lectin and phytate were present. With the aim of selecting non-toxic genotypes of this species in this work we studied the nutritional potential and antinutritional factors (ANFs) in J. curcas seed from different states of Mexico. Defatted Jatropha curcas meal contained 43 – 50 % protein.
The seed storage proteins i.e. the albumins, globulins, glutelins and prolamins were sequentially extracted according to their solubility. The major protein fraction was globulin (44 %), followed by glutelin and albumin (40 and 12 %), respectively. The prolamins were a minor fraction (3.4 %). The digestibility (86 – 90 %) was slightly higher than reported values for some legume seeds. Both defatted and non-defatted meal tested positively for alkaloids, trypsin inhibitor activity (28 - 35 UTI/g) and haemagglutination. Keywords: Jatropha curcas, piñoncillo, trypsin inhibitor, haemagglutination Introduction The search for new protein sources has become important over the past few decades. This is not only to meet the increased demand for protein but also to seek alternative crops that can be cultivated in marginal soils and to seek plant species capable of producing high quality protein. Jatropha curcas commonly, referred to as physic nut, purging nut, Piñon and Piñoncillo is native to Central America and Mexico where it occurs naturally in coastal forests. Jatropha curcas is a drought resistant shrub or tree belonging to the family Euphorbiaceae. It is cultivated in Central and South America, South East Asia, India and Africa (Schmook & Serralta, 1997).
The oilseed cake, left after oil extraction, is currently used as a fertiliser. However, it may have potential as a livestock feed, as it is rich in crude protein (50 - 58 %) depending on the residual oil level. The essential amino acid levels, except for lysine, in J. curcas meal protein are higher than those in the FAO reference protein for a growing child of 2 - 5 years (Makkar & Becker, 1997). However, both seed and oil have are toxic. A mixed seed sample from different trees in the Papantla region of Veracruz State, Mexico was found to be non-toxic. These kernels after roasting are consumed by humans (Makkar et al., 1998). There is little information on the proteins in J. curcas seed. The only member of this family that has been studied is the castor bean (Ricinus communis) about 44 % of total seed protein is type

186
2S, 14 % is 7S, and 42 % is 11S (Shewry & Pandya, 1999). In this work, we studied the protein composition and ANFs in Jatropha curcas seed from different states of Mexico to obtain information for plant selection purposes. Materials and methods Jatropha meal samples The Jatropha curcas seed was collected in the states of Veracruz and Morelos, Mexico. Dry seeds were dehulled by hand and ground to flour, which was defatted with hexane. Protein fractions Albumins, globulins, prolamins, and glutelins from J. curcas seed were sequentially extracted using appropriate solutions (Osborne, 1924). Chemical analyses The seed samples were analysed for protein (N × 6.25), fat, ash and fibre using AOAC (1990) procedures. Carbohydrates were determined by difference. In vitro digestibility A multienzyme system of trypsin, chymotrypsin and peptidase was used (Hsu et al., 1977). Five ml of the multienzyme solution was added to a protein suspension that was being stirred at 37°C. There was an immediate rapid decline in pH. This was caused by the freeing of amino acid carboxyl groups from the protein chain by the proteolytic enzymes. The drop in pH was recorded automatically over a 10-minute period with a recording pH meter. Trypsin inhibitor and haemagglutinin activity Trypsin inhibitor activity was determined essentially according to Smith et al. (1980) except that the enzyme was added last, after the inhibitor was mixed with the substrate. The results are expressed as trypsin inhibited/g of sample (UTI/g).
Lectin analysis was by haemagglutination assay in round-bottomed wells of microtiter plates with a 1 % (v/v) trypsinised erythrocytes (from cows blood) suspension in phosphate buffered saline (PBS). The sedimentation pattern of the erythrocyte suspensions was read after 2 h at room temperature. A positive pattern which indicated agglutination was a uniform covering of the bottom of the well by erythrocytes while a negative pattern (indicating no agglutination) was a circular clump of erythrocytes surrounded by a concentric, clear zone of equal size to the blank (which contained PBS instead of sample). Haemagglutination activity was defined as the minimum amount of J. curcas meal in mg/ml of assay medium that produced agglutination.

187
Results and discussion Nutritional potential of Mexican piñon Mexican piñon seed has a high fat and protein content. The chemical composition of the J. curcas meals is shown in Table 1. The J. curcas seed contained 26.3 – 28.1 % protein, which is in the range of values reported for other legumes and 60 % fat. The protein level was higher in the defatted meal to 48.3 – 49.9 %. Thus Mexican piñon meal from non-toxic varieties could be a good protein source for both animals and humans. Table 1. Proximate analysis of entire and defatted Jatropha curcas seed meals of seed from
Morelos and Veracruz. Morelos Veracruz
Component (%)a Meal Defatted meal Meal Defatted meal Protein 26.3 ± 1.1 48.3 ± 2.0 28.1 ± 1.2 49.9 ± 0.9 Fat 59.9 ± 0.9 2.9 ± 0.5 57.1 ± 1.0 3.5 ± 0.5 Ash 4.7 ± 0.2 8.6 ± 0.2 4.1 ± 0.1 8.3 ± 0.3 Fibre 4.8 ± 0.2 8.1 ± 0.4 4.7 ± 0.2 8.5 ± 0.2 Carbohydrateb 4.3 32.0 4.0 29.8 aDry basis, mean of three determinations. bBy difference.
Seed storage proteins (albumins, globulins, glutelins and prolamins), which had been sequentially extracted, according to their solubility, had the same distribution in seed from Morelos and Veracruz. The major protein fraction was globulin (40.8 - 44.4 %), followed by glutelin and albumin (38 - 39.8 and 12.3 - 13.8 % respectively). Prolamin was a minor fraction at only 3.4 - 7.4 %. Table 2. Protein fractions in Mexican piñon seed from Morelos and Veracruz.
Percentage of total proteina Protein fraction Morelos Veracruz Albumin 12.3 ± 1.5 13.8 ± 1.8 Globulin 44.4 ± 2.3 40.8 ± 2.0 Prolamin 3.4 ± 0.8 7.4 ± 3.1 Glutelin 39.8 ± 2.5 38.0 ± 1.1 aMean of three determinations
The protein digestibility was 88 and 90 % in the lines from Morelos and Veracruz respectively. This is slightly higher than reported values for some legume seeds. Toxic/antinutritional components Jatropha curcas seed has been reported to be toxic to humans, rodents and livestock but in Mexico, some lines are non-toxic (Makkar et al., 1997). The results for trypsin inhibitor activity and haemagglutination of defatted Mexican piñon meal from lines from Morelos and Veracruz are shown in Table 3. Trypsin inhibitor activity in the defatted meals from Morelos and Veracruz seed varied from 28 to 35 UTI/g.
Lectin activity was the same in the Morelos and Veracruz seed lines. Similar lectin values, in toxic and non-toxic lines were reported by Makkar et al. (1997). This suggests that lectin is not the main toxic principle in J. curcas seed.

188
Table 3. Trypsin inhibitor activity and lectin content of meal of Mexican piñon varieties from Morelos and Veracruz.
Origin Trypsin inhibitor (UTI/g) Lectin activity* Morelos 28 51 Veracruz 35 51 *Minimum amount of meal (mg/ml) which produced haemagglutination
Trypsin inhibitors and lectins are heat labile, and their adverse effects could be mitigated by heat treatment before meal or seeds are consumed. References AOAC. 1990. Official Methods of Analysis. 15th Edition. Association of Official
Analytical Chemist, Washington, DC. Hsu, H.W. Vavak, D.L. Satterlee, L.D. & Miller, G.A. 1977. A multienzyme technique for
estimating protein digestibility. Journal of Food Science 42, 1269-1273. Makkar, H.P.S., Aderibigbe, A.O. & Becker K. 1998. Comparative evaluation of non-toxic
and toxic varieties of Jatropha curcas for chemical composition, digestibility, protein degradability and toxic factors. Food Chemistry 62, 207-215.
Makkar, H.P.S. & Becker, K. 1997. Potential of Jatropha seed cake as a protein supplement in livestock feed and constraints to its utilization. In G.M. Gübitz, M. Mittelbach & M. Trabi (Eds). Symposium “Jatropha 97:” Biofuels and Industrial Products from Jatropha curcas, Managua, Nicaragua, 23-27 February 1977, 190-206.
Makkar, H.P.S.; Becker, K., Sporer, F. & Wink M. 1997. Studies on nutritive potential and toxic constituents of different provenances of Jatropha curcas. Journal of Agricultural and Food Chemistry 45, 3152-3127.
Osborne, T.B. 1924. The Vegetable Proteins. Longmans-Green, London. Schmook, B. & Serralta-Peraza, L. 1997. Jatropha curcas: Distribution and uses in the
Yucatán Peninsula of México. . In G.M. Gübitz, M. Mittelbach & M. Trabi (Eds). Symposium “Jatropha 97:” Biofuels and Industrial Products from Jatropha curcas, Managua, Nicaragua, 23-27 February 1977, 53-57.
Shewry, P. & Pandya, R. 1999. Seeds Proteins. P.R. Shewry & R. Casey (Eds). Kluwer Academic Publishers, Norwell. Ma.
Smith, C., Megen, W. van, Twaalfhoven, L. & Hitchcock, C. 1980. The determination of trypsin inhibitor levels in foodstuffs. Journal of the Science of Food and Agriculture 31, 341-350.

189
Immunodetection of legume proteins resistant to digestion in weaned piglets P. Salgado1,2, J.P.B. Freire2 & J.P. Lallès1 1Institut National de la Recherche Agronomique INRA-UMRVP, 35590 Saint-Gilles, France 2Instituto Superior de Agronomia, Tapada de Ajuda, 1349-017 Lisboa, Portugal Summary Experiments were conducted to investigate ileal digestion of legume proteins in piglets weaned onto diets whose protein was provided by casein (control) or an equal protein level mixture of casein and raw pea (Pisum sativum), faba bean (Vicia faba), narrow-leafed lupin (Lupinus angustifolius), Kabuli and desi chickpea (Cicer arietinum) or soybean (Glycine max) meal. Ileal digesta were analysed by western blotting with antibodies raised against the main 11S and 7S legume proteins. Only a few immunoreactive legume protein bands were detected by immunoblotting. They mainly belonged to the 7S family in the case of pea, faba bean and chickpea. Nearly intact narrow-leafed lupin proteins of the 11S family were identified. Keywords: piglet, weaning, legume proteins, ileal digestion Introduction Grain legume seeds are rich in protein and can be incorporated into weaning diets for piglets as alternatives to soybean meal. Legume seeds are of nutritional interest, but their protein digestibility is variable and is generally lower than casein based control diets. Different factors, including antinutritional factors (ANFs) and the fibre content, but also the natural resistance of legume storage proteins to digestion may explain these differences (Lallès & Jansman, 1998). As a consequence there are usually increased ileal losses of both exogenous and endogenous nitrogen (N) and amino acid (AA).
Storage globulins are approximately 70 % of legume seed proteins and are composed of two major families, 11S or legumin and 7S or vicilin (Casey, 1999). Pea legumin is more resistant to hydrolysis by trypsin than by pepsin, and the β−polypeptides of legumin are more resistant than the α−polypeptides (Plumb & Lambert, 1990; Perrot et al., 1999). In contrast, vicilin is hydrolysed better by trypsin than by pepsin. The 7S globulin of kidney bean (Phaseolus vulgaris), phaseolin, is highly resistant to hydrolysis by both pepsin and trypsin (Begbie & Ross, 1993). Despite high homology, globulins of various legume seeds exhibit marked differences in their susceptibility to hydrolysis (Nielsen et al., 1988).
Little is known on the biochemistry of legume protein ileal digestion in young pigs. The aim of this study was to investigate it using western blotting. Materials and methods Animals, diets and digesta collection The experiments used 30 (8.7 ± 0.3 kg body weight (BW), Exp. 1) and 18 (8.2 ± 0.3 kg, Exp. 2) male piglets (Duroc × Landrace) weaned at 4 weeks. They were divided into 5 (Exp. 1) and 3 (Exp. 2) groups of 6 pigs on the basis of litter origin and average weaning BW. Piglets were individually fed (40 g/kg BW/d) one of the 5 (Exp. 1) or 3 (Exp. 2) experimental diets from

190
weaning. After 12 d of adaptation to the diets, piglets were fitted with an ileo-rectal anastomosis.
A control casein (C) based diet (Exp. 1 and 2), and diets in which 50 % of the crude protein (CP) (N × 6.25) was supplied by pea (P), faba bean (FB), narrow-leafed lupin, seed and defatted soybean meal (SBM) in Experiment 1, and by Kabuli (KCP) or desi (DCP) chickpea seed in Experiment 2, were formulated. The proteins differed in their chemical composition and ANF content. The diets contained 25 and 12.5 % (as fed) of casein for the control and legume-based diets, respectively, and 47.5, 39.6, 29.3, 49.1, 47.8 and 23.7 % of P, FB, L, KCP, DCP and SBM, respectively. Other ingredients were maize starch, wheat straw, synthetic AAs, soybean oil, sucrose and minerals. The CP and energy content of the diets varied between 220 - 240 g and 16 - 18 MJ/kg DM, respectively.
Ileal digesta were collected totally from anastomosed pigs over 7 d of the fourth week of the experiment, weighed, mixed with preservatives and frozen at –20 °C. They were then freeze-dried and ground (1 mm screen). Soluble protein in the digesta was extracted in borate buffer pH 8.0. Supernatants were prepared by centrifuging and were frozen (–20 °C). Legume protein digestion studied by western blotting Crude protein extracts (CPE) were obtained from ground legume seed by extraction in Tris-HCl at pH 7.5. The globulin fraction of the legumes was isolated and fractionated into the legumin and vicilin protein families by FPLC (Melo et al., 1994). Hyperimmune plasmas were prepared in rabbits. The SDS-PAGE determinations were carried out on polyacrylamide gels. Samples run on a given gel included molecular weight (MW) standards, casein, control digesta (a pooled sample) from casein-fed piglets, CPE and the 11S and 7S preparations for a given legume species and the ileal digesta of 5 (out of 6) piglets fed the corresponding legume diet. Samples were run under reducing conditions. Separated proteins were transferred from the gel to nitrocellulose membranes that were saturated using skim-milk powder. The membranes were incubated with the prepared hyperimmune plasmas, and then with an anti-rabbit IgG labelled with peroxidase. Protein binding was revealed using diaminobenzidine and H2O2. The MWs were calculated from the MW standards by linear regression. Results Western blotting allowed our anti-legume antibodies to recognise protein bands with most MW, as expected, being between 40 - 20 and 70 - 40 kDa, respectively (Table 1). Higher MW protein bands were observed for lupin 11S, and lower for faba bean and lupin 7S. Some of our antibody reagents were not totally specific and cross-reactions between globulin families were observed, as mentioned by Perrot et al. (1999).
The major protein bands, stained on the western blots, carried out for each CPE extract or purified globulin are summarised in Table 2. They were usually of low intensity. One to four protein bands with MWs between 62 and < 14 kDa, depending on legume species, were detected with the anti-CPE reagents. In contrast, no protein bands were stained with the anti-11S reagents, except in lupin and Kabuli chickpea (one band each). A few protein bands (2 - 3) stained with the anti-7S reagents, except in lupin and soybean meal (no staining).

191
Discussion Our anti-legume protein plasmas did not recognise proteins from casein and control digesta or endogenous proteins, identified in the ileal digesta, of the pigs (Salgado, 2001). This shows that the protein bands detected in the ileal digesta of legume-fed piglets were of legume origin. However, the staining was usually faint and this supports the fairly high ileal protein digestibility of these legumes (Salgado, 2001). Table 1. Summary of the molecular weight (kDa) of the major protein bands recognised by
hyperimmune plasmas prepared against 11S and 7S proteins of legume seeds and used in SDS-PAGE.
Legume species Anti-11S plasma Anti-7S plasma Pea 43, 26, 24, 20 70, 50, 38 Faba bean 41, 39, 33, 25 70, 55, 50, 44, 42, 33, 27 Narrow-leafed lupin 63, 55, 43, 25, 19 73, 64, 57, 40, 38, 20 Chickpea1 42, 38, 24, 22, 20 75, 58, 38 Soybean 40, 20 76, 72, 53 1hyperimmune plasmas prepared to 11S and 7S proteins from Kabuli chickpea only. Table 2. Molecular weights (kDa) of the major protein bands recognised by SDS-PAGE in
the ileal digesta of piglets using anti-crude protein extracts (CPE), 11S and 7S protein antibodies.
Legume species Anti-CPE Anti-11S Anti-7S Pea 50 (4)1, 40 (4), 33 (4), 19 (3) none (5) 50 (5), 22 (4), 20 (4) Faba bean 25 (5) none (5) 70 (4), 50 (5) Narrow-leafed lupin
50 (5), 36 (4), 20 (3) 20 (4) none (5)
Chickpea Kabuli 62 (2), 27-24 (5), <14 (5) 25 (3) 35 (3), 32 (1) desi 62 (3), 29-25 (4), <14 (4) none (5) 73 (1), 62 (2), 53 (5), 34 (4), 28 (1)Soybean 44 (4), 40-39 (5), 24 (2) none (5) none (5) 1number of individual digesta in parentheses stained for each band (total number/treatment = 5).
Legume proteins are partially resistant to in vitro (Nielsen et al., 1988, Plumb &
Lambert, 1990, Perrot et al., 1999) and in vivo digestion (Begbie & Ross, 1993; Crévieu et al., 1997; Lallès et al., 1999; Carbonaro et al., 2000). Less immuno-recognition was observed with anti-11S plasmas (Table 2). This data, with piglets, indicates that the 11S proteins were digested more than 7S proteins, except in lupin and soybean. However, basic polypeptides of pea legumin and soybean have been detected in ileal digesta of chicken (Crévieu et al., 1997) and veal calves (Lallès et al., 1999), respectively. In the case of faba bean the proteins of MW 70 and 50 kDa are probably from vicilin (Carbonaro et al., 2000). Only one band at 20 kDa, from lupin, was found in common between the CPE and the 11S preparations, suggesting a legumin origin. Two bands of 36 and 20 kDa, observed here, were still present after fermenting lupin with rumen contents (Spencer et al., 1988). This emphasises their high resistance to breakdown. We observed a number of protein bands in chickpea, some of which were apparently from vicilin. Kabuli and desi chickpeas had different protein staining patterns. Discrepancies among the protein bands revealed using anti-CPE plasmas on one hand and those stained by anti-11S and anti-7S reagents on the other hand also suggest that partially digested globulins and other, non-globulin proteins would

192
escape digestion. This was the case for low MW (<14 kDa) proteins from the chickpeas. Although some of the bands might be related to protease inhibitors, we did not attempt to identify them further. In conclusion, a few faintly stained bands were identified in the ileal digesta of legume-fed piglets as being of legume origin. There were less 11S (legumin) proteins present than those from the 7S (vicilin) family. Acknowledgement Financial support of a PRAXIS XXI program of the Fundação para a Ciência e a Tecnologia and Instituto de Ciências Agrárias Mediterrânicas, Portugal is acknowledged. References Begbie, R. & Ross, A.W. 1993. Resistance of the kidney bean reserve protein, phaseolin, to
proteolysis in the porcine digestive tract. Journal of the Science of Food and Agriculture 61, 301-307.
Carbonaro, M., Grant, G., Cappelloni, M. & Pusztai, A. 2000. Perspectives into factors limiting in vivo digestion of legume proteins: Antinutritional compounds or storage proteins? Journal of Agricultural and Food Chemistry 48, 742-749.
Casey, R. 1999. Distribution and some properties of seed globulins. In Seed Proteins. P.R. Shewry & R. Casey (Eds). Kluwer Academic Publications, Dordrecht. pp. 159-169.
Crévieu, I., Carré, B., Chagneau, A.M., Quillien, L., Guéguen, J. & Bérot, S. 1997. Identification of resistant pea (Pisum sativum L) proteins in the digestive tract of chickens. Journal of Agricultural and Food Chemistry 45, 1295-1300.
Lallès, J.P. & Jansman, A.J.M. 1998. Recent progress in the understanding of the mode of action and effects of antinutritional factors from legume seeds in non-ruminant farm animals. In Recent Advances of research in antinutritional factors in legume seeds and rapeseed. Proceedings 3rd International Workshop on Antinutritional Factors in Legume Seeds and Rapeseed, Wageningen 8-10 July 1998, 219-232.
Lallès, J.P., Tukur, H.M., Salgado, P., Mills E.N.C., Morgan, M.R.A., Quillien, L., Levieux, D. & Toullec, R. 1999. Immunochemical studies on gastric and intestinal digestion of soybean glycinin and β -conglycinin in vivo. Journal of Agricultural and Food Chemistry 47, 2797-2806.
Melo, T.S., Ferreira R.B. & Teixeira, A.N. 1994. The seed storage proteins from Lupinus albus. Phytochemistry 37, 641-648.
Nielsen, S.S., Deshpande, S.S., Hermodson, M.A. & Scott, M.P. 1988. Comparative digestibility of legume storage proteins. Journal of Agricultural and Food Chemistry 36, 896-902.
Perrot, C., Quillien, L. & Guéguen, J. 1999. Identification by immunoblotting of pea (Pisum sativum L) proteins resistant to in vitro enzymatic hydrolysis. Science des Aliments 19, 377-390.
Plumb, G.W. & Lambert, N.A. 1990. Comparison of the trypsinolysis products of nine 11S globulin species. Food Hydrocolloids 3, 465-473.
Salgado, P. 2001. Digestion des protéines de légumineuses et réponses immunitaires chez le porcelet et le porc en croissance. Thèse de Doctorat, Ecole Nationale Supérieure Agronomique de Rennes, France.
Spencer, D., Higgins, T.J.V., Freer, M., Dove, H. & Coombe, J.B. 1988. Monitoring the fate of dietary proteins in rumen fluid using gel electrophoresis. British Journal of Nutrition 60, 241-247.

Session IV Effects of antinutritional factors on monogastric animals


195
Recent progress on research on the effects of antinutritional factors in legume and oil seeds in monogastric animals A. Brenes1, A.J.M. Jansman2 and R.R. Marquardt3 1CSIC, Instituto del Frío, Departamento de Metabolismo y Nutrición, José Antonio Novais, 10. 28040 Madrid, Spain 2Animal Sciences Group, Division of Nutrition and Food, Edelhertweg 15, PO Box 65, 8200 AB Lelystad, The Netherlands 3University of Manitoba, Department of Animal Science, Winnipeg, Manitoba R3T 2N2, Canada Introduction In recent years the interest in Europe in the production of legume and oilseeds as alternative protein sources replacing the more traditional ingredients in diets for monogastric animal species has grown considerably. The recent ban on the use of animal protein lead to an increased demand for alternative protein sources in Europe. The most commonly used legumes are peas (Pisum sativum), faba beans (Vicia faba), and lupins (Lupinus spp). They contain several natural constituents, which can interfere with nutrient digestion and utilization or interfere with specific physiological processes within the animal: trypsin inhibitors (TI), lectins, tannins, α-galactosides and alkaloids.
Some oil seeds e.g. sunflower (Helianthus annuus), cottonseed (Gossypium spp) and rapeseed-canola (Brassica spp.) are primarily grown for oil production for human consumption. They are suitable protein sources in the diets of monogastric animals, despite the presence of antinutritional factors (ANFs) (e.g. mucilage, chlorogenic acid, gossypol, erucic acid). Their oil contains large proportion of highly unsaturated fatty acids and it may, therefore, also contribute to animal essential acid fatty acid requirements. The composition of oilseed meals is influenced by the processing conditions during oil extraction. Hull content, preconditioning, cooking and the solvent extraction procedure determine the nutritive value of the meal.
This paper reviews information published since the third ANF Workshop in 1998 on legume seeds and oilseeds in animal feeds for non-ruminant with special emphasis on ANFs in the seed. Legume seed Peas Compared with other legumes, currently grown pea varieties have a relatively ANF low content and a relatively high protein and energy content. As a result, they are increasingly used as a feed ingredient in monogastric diets. However, Gdala et al. (1992) reported that pig diets containing pea seed had lower protein digestibility values than soybean (Glycine max) meal-based diets. This can partly be explained by the presence of ANFs, including protease inhibitors, oligosaccharides and/or fibrous material. All these factors limit accessibility of legume seed protein to digestive enzymes and increase the amount of endogenous nitrogen (N) present at the terminal ileum (Lallès & Jansman, 1998).
Although considerable information on the nutritional value of peas for pigs is available, in practice, peas as an ingredient in animal feeds sometimes have a relatively poor image

196
(Jansman & Verstegen, 2002). The limited supply of peas and their nutritional problems are factors that limit their use in different countries.
In earlier studies, a negative effect of trypsin inhibitor activity (TIA) on ileal protein or amino acid digestibility has been observed. However, variability in protein or amino acid ileal digestibilities might not only be associated with TIA, but also with fibre content or other unidentified factors. In most of these studies, a low TIA was associated with large seeds with a low fibre content, whereas, a high TIA was associated with small seeds with a high fibre content. For this reason, most of research in the last few years with peas has studied factors that affect nutrient utilisation, particularly protein and amino acids digestibility. Grala et al. (1999) showed that differences in apparent ileal digestibility of protein and amino acids observed in piglets fed diets containing peas were related to differences in ileal recovery of both endogenous and dietary N. Moreover, the presence of protease inhibitors and the extremely high water-holding capacity of the fibre fraction of peas, may be factors which negatively affect the apparent ileal digestibility of crude protein and amino acids (AAs). Their effect is to increase the rate of passage of digesta in the small intestine. Differences in the neutral detergent fibre (NDF) content were, in part, responsible for the variation of apparent ileal digestibility the majority of the AAs in pea seed used in growing-finishing pig diets. However, with the exception of tryptophan, TIA was not related to the variation in ileal AA digestibility values (Fan & Sauer, 1999). These observations are contrary to the results of Grosjean et al. (2000). The explanation may lie in the small variation in the fibre content of the peas used in this study. The authors suggest that a high TIA is the main factor explaining the low digestibility, rather than the time of cultivation (winter versus spring).
Canibe & Bach-Knudsen (2001) and Bach-Knudsen et al. (2001) investigated the degradation of pea fibre along the gastrointestinal tract of pigs and its relation to changes in some of their physico-chemical properties (water-holding capacity and extent of fermentation). Ileal and caecal digesta containing pea cotyledon fibre showed the highest swelling and water retention properties, and a higher digestibility of non-starch polysaccharides (NSP) compared to pea hull fibre. Moreover, the particle size of pea cotyledons and hulls was drastically reduced in the faeces compared with the ileum and the caecum. The digestibility of the different carbohydrate fractions in peas are compared with barley (Hordeum vulgare) in Table 1.
Salgado et al. (2002a, b) compared the effect of pea and narrow-leafed lupin seed (Lupinus angustifolius) on gut morphology and enzyme activity of the intestinal mucosa of weaned and growing pigs. The relatively longer jejunal villi in pigs fed a pea based diet compared to lupin diets reflected the high level of energy absorbed from the soluble sugars and volatile fatty acids formed in the large intestine by the fermentative degradation of fermentable fibre present in the peas. In pigs fed lupin-based diets, the amount of insoluble fibre ingested and the low amino acid digestibility of this fraction could have increased the rate of mucosal cell death and consequently decreased the villus height. Higher maltase, sucrase, and particularly, peptide hydrolase activities may have been stimulated by the structural complexity of the legume proteins and their need for extensive degradation to amino acids and oligopeptides prior to absorption.
Recent research on peas has also focused on the effects of processing and the addition of enzymes on apparent seed digestibility. Extrusion and micronisation of raw peas improved their AA digestibility and the efficiency of feed utilisation when fed to early weaned pigs (Owusu-Asiedu et al., 2002). However, there was an inconsistency between improved pig performance and nutrient digestibility. Similarly, O‘Doherty & Keady (2001) using expansion processing, Thacker & Racz (2001) using enzymes in hulled and dehulled pea diets in growing and finishing pigs, and Nonn et al. (1999) studying the efficacy of two enzyme

197
preparations in weaned piglets were unable to improve the nutritional value of the diet. Alonso et al. (2000) using extruded peas in rat diets did not appear to improve the nutritional quality of the diet. However, the supplementation of raw and extruded peas with free amino acids up to the required values for rats greatly improved the nutritional quality of extruded supplemented peas. Associated with the reduction in ANFs in peas, their extrusion prior to mixing consistently increased the apparent nutrient digestibility in diets containing peas (O’Doherty & Keady, 2001). Protein and AA digestibility can be improved significantly more by extrusion than by dehulling (Mariscal-Landin et al., 2002). Abd El-Haddy & Habiba (2003) found that extrusion of different legume seeds, including peas, that had been soaked in water for 16 h improved their in vitro protein digestibility, enhanced phosphorus availability, reduced tannin and polyphenol concentration, and eliminated trypsin and α-amylase inhibitors when present.
Table 1. Ileal and total tract digestibility of different carbohydrate fractions of peas and
barley (Bach Knudsen et al., 2001). Digestibility Content (g/kg DM) Ileum Total tract Peas Barley Peas Barley Peas Barley Sugars 39 16 0.95 - 1.00 Non-dig. oligosacchar. 50 10 0.54 - 1.00 1.00 Starch 454 587 0.91 0.96 1.00 1.00 NSP 180 187 0.18 0.17 0.88 0.52 Total carbohydrates 723 800 0.70 0.78 0.97 0.89
The use of near-isogenic lines of peas with high or low levels of TIA will permit a more
meaningful assessment of the effects of pea TI proteins in animals. Hedemann et al. (1999) indicated that TIA content was correlated with a significant negative effect on protein digestibility and biological value. The TIA content of the pea seed was reflected in the relative activity of pancreatic chymotrypsin. Likewise, Wiseman et al. (2003) showed that the digestibility of AAs by young poultry was significantly improved using lines of pea with a low TIA in the seed compared with near isogenic counterparts having a high TIA.
The nutritional value of 25 white flowered pea lines, 12 coloured peas and 5 wrinkled peas in mash or pelleted diets was evaluated using adult cockerels (Grosjean et al. 1999). Feed peas had a higher feeding value than coloured-flowered and wrinkled peas. The difference can mainly be explained by the presence of tannins in the testas of the coloured-flowered peas, the proportion of amylose and amylopectin in the starch and a different fibre content. Pelleting the diet improved the feeding value of the peas. Grinding to a small particle size can greatly improve pea starch digestibility or metabolisable energy in poultry (Carré, 2002). However, the digestibility of peas with a small mean particle size was still inferior to that of maize (Zea mays). Small particles may also increase water losses with a subsequent reduction in the litter quality (Carré, 2000).
Recent data on the ANFs of 18 pea lines showed that peas with yellow cotyledons had the highest TIA (Vidal et al., 2003). Peas with brown testas had the lowest verbascose and sucrose content and were richest in inositol hexaphosphate.
The presence of TIA and the fibre properties are the main factors affecting ileal digestibility of peas in pigs. Solubility, water retention capacity and particle size of pea fibre all affect the fermentation process. Pigs fed legume containing diets showed significant differences in intestinal enzyme activity. Further research is necessary to study the effect of specific seed fractions on gastrointestinal health and microbial activity. Plant breeding for

198
low TIA in the seed offers the potential to improve the nutritional value of peas. There is also a need for the development of molecular marker techniques for the selection of protein quality determinants in plant breeding programs. Faba bean Faba beans have been studied extensively during the past 20 years as a source of protein-rich supplement in the diet of monogastric animals (Marquardt et al., 1989). Flis et al. (1999) studied the nutritional value of pig diets containing 300 g/kg of diet of field bean (Vicia faba) with various levels of condensed tannins. Despite a slightly lower nutritional value, as expressed by metabolisable energy and digestible protein, the high-tannin diet (1 g/kg phenols and 0.6 g/kg proanthocyanidins) did not lower pig growth rate (25 - 63 kg) when compared with animals fed a low tannin or dehulled faba bean diet. In other studies faba bean tannins have been shown to effect growth and nutrient utilisation in poultry (Marquardt et al., 1989). Higher digestibility of amino acids in faba bean seeds were reported from white flowered (tannin-free) than from coloured flowered cultivars (tannin containing) (Jansman et al., 1993) in pigs. Mariscal-Landin et al. (2002) corroborated these results. However, according to the latter authors, dehulling also significantly improved the arginine and tryptophan digestibility but not the other AAs. Salgado et al. (2002c) studied the nature of endogenous protein losses at the terminal ileum produced by feeding legume seed to weaned pigs. These proteins shared N-terminal amino acid sequences with enzymes of the serine protease family.
The biological activity of small amounts of faba bean proanthocyanidins has been determined in rats (Juskiewicz et al., 2001). Proanthocyanidins extracted from faba bean testas (1 and 3 g/kg) when included in the diet had no effect on crude protein digestibility, N retention or the biological value of dietary protein. A lower activity of β-glucuronidase activity was found in the digesta compared to the control group. This produced an effect on the large bowel microflora. This data is not in agreement with other studies where it was shown that faba beans condensed tannins reduce AA digestibility and retention (Marquardt et al., 1989).
The effect of different (bio)technological processes on the nutritional value of faba bean has been studied. The feeding of germinated faba beans to rats gave a significant improvement in growth performance, N retention, apparent ileal digestibility of some AAs, and faecal starch and NSP digestibility compared to feeding non-germinated faba bean (Rubio et al., 2002a; Rubio, 2003). Similarly, faba beans extrusion improved their nutritive value (Mariscal-Landin et al., 2002; Abd-El-Hady & Habiba, 2003).
Joa et al. (2002) studied systemic and local immune responses to legume proteins in piglets weaned at 28 days of age. Piglets fed legume-containing diets had higher plasma immunoglobulin G titres to legume proteins than piglets fed casein-based diets. The most immunoreactive proteins belonged to the vicilin family.
Other studies with faba beans suggest that their relatively low nutritional value is partially due to the poor nutritive quality of the major reserve proteins (vicilin and convicilin) rather than the presence of any known ANFs (Rubio, 2000). Characterization of the gastrointestinal digesta for globulin content and AA analysis of undigested protein of whole cooked faba beans indicated that it was mainly its structural properties and not the binding of polyphenols, which determined the extent of protein aggregation on autoclaving. Therefore, this may be responsible for its low digestibility (Carbonaro et al., 2000). Purified legume storage proteins (faba bean, lupin and chickpea (Cicer arietinum) globulins) and casein were subjected to in vitro digestion followed by the use of a bicameral Caco-cell culture system to determine AA transport across the cell layer (Rubio & Seiquer, 2002b). The results

199
confirmed previous in vivo observations that legume seed proteins are probably digested and absorbed at rates different from those of animal proteins.
The low nutritional value of faba beans and other legumes is due in part, to the structural properties of their storage proteins. They are difficult to digest and be absorbed in the intestinal tract, and by their immunogenicity. Further work is needed to highlight the nature of this undigested dietary proteins or its fragments and their effect on the secretion and digestion of endogenous constituents, and their impact on protein metabolism.
Lupins Low-alkaloid lupin seed, of several species are a potential source of good quality protein supplements for monogastric animals such as poultry and pigs. They are free of antinutritional compounds such as trypsin inhibitors and haemagluttinins (van Barneveld, 1999). However, there are limitations to the inclusion of lupin seed in the diet due to the presence of NSP and oligosaccharides of the raffinose family. Research over the last five years has mainly concentrated on the adverse effects of lupin seed and its fractions on nutrient digestibility of nutrients, and on gastrointestinal tract morphology. The possibile use of enzymes (proteases, galactosidases and carbohydrases) to improve the nutritional value and to study the mechanism of action of these enzymes in lupin seed has also been investigated.
The carbohydrates of lupins are different, as is common with other legume seeds, they contains negligible levels of starch, and high levels of soluble and insoluble NSP and oligosacharides. Detailed information on the polysaccharides compositions of whole lupin, lupin kernel and lupin hull was reported by van Barneveld (1997) and van Laar et al. (2000). Lupin kernels contain the raffinose series of oligosaccharides (74 to 80 g/kg) in addition to high concentrations of total NSPs (350 g/kg) of which 15 % are soluble.
On the basis of recent analysis (Wasilewko & Buracewska, 1999), the average alkaloid content in Lupinus albus cultivars was around 0.8 g/kg dry matter (DM), which is twice the amount found in L. angustifolius. These values may be much higher depending on weather conditions during flowering. The tolerance of animals to the lupin alkaloid appears to vary among different species. In pigs, the toxicity of sweet varieties of lupin is reported to be at 0.33 g/kg total alkaloid in the diet (Allen, 1998). Fast growing chickens may be more sensitive to the toxic challenge because of the high metabolic rate associated with rapid growth. Skeletal deformity, mortality and increased content of cytochrome P-450 in the liver microsomes of birds has been observed by Olkowski et al. (2001) in some broiler chickens fed raw narrow-leafed lupin based diets. Feed refusal associated with the consumption of sweet lupin has not been associated with the presence of alkaloids.
Performance responses to the inclusion of lupin in pig and poultry diets can be impaired if the overall NSP content of the diet is high (van Barneveld, 1997; Brenes et al., 2002). A reduction in voluntary feed intake, an increase in the feed to gain ratio and the size of the gastrointestinal tract, and a decrease in nutrient digestibility may be, in part, attributed to the indigestible fibre content. Maximum performance in chicks fed a lupin diet was achieved by the removal of hulls from lupin seed. This was because dehulled seed contains less NSP and their nutritive value was higher than that of whole seed (Brenes et al., 2002; Rubio et al., 2003). The strong inverse relationship shown by Dunshea et al. (2001) in pigs between feed intake and retention time in the GI tract suggest that delayed digestion could results in a lower feed intake and in a substantial increase in hindgut fermentation of pigs fed diets containing a high proportion of lupin seed.
Cell wall material isolated from the endosperm of lupin seeds was studied to relate cell wall composition and structure with fermentation characteristics (van Laar et al., 2000).

200
Faecal microbes from pigs appeared to degrade pectins or pectin-related sugars (galactose, arabinose, uronic acid) faster than cellulose. Probably, the pectin-rich cell wall is composed of a relatively “open” structure in which a cellulose-xyloglucan network forms the backbone of the cell wall. Table 2 shows the differences in volatile fatty acid (VFA) production profiles obtained for the cell wall substrates from lupin, soybean, pea and faba bean.
Table 2. Percentage of residues of cell walls after 144 h in vitro fermentation, VFA
production pattern (% of total production) (acetic, Ac; propionic, Pr, butyric acid, Bu, and Ac/Pr ratio) and gas production yield (GPY; ml/g) for the fermentation of legume seed endosperm cell wall material (Van Laar et al., 2000).
Legume seed Residue (%) Ac Pr Bu Ac/Pr GPY
Faba bean 14.4ab 60.2a 31.4b 8.4b 1.9a 428a L. albus 12.8b 47.8d 43.4a 8.8b 1.1c 420ab Pea 15.7ab 57.8b 32.8b 9.4b 1.8ab 433a Soy 17.1a 54.3c 33.6b 12.1a 1.6b 391b abc P < 0.05 for data in the same column.
Morphological and functional changes of the intestinal mucosa have also been observed
in growing pigs. Salgado et al. (2002b) showed that the duodenal and jejunal crypt width was lower in pigs fed lupin-containing diets because the amount of insoluble fibre ingested and the apparent ileal digestibility could have increased the rate of cell death and consequently decreased the villus height. Significant differences in intestinal enzyme activity were also observed. In poultry, increasing the lupin content in the diet enlarged the relative size of several sections of the gastrointestinal tract (Brenes et al., 2002) related to the high level of non-enzymatically digestible constituents.
Lupin seed contains significant levels of raffinose family oligosaccharides that appear to be indigestible in the stomach and small intestine. The role of dietary oligosaccharides in poultry is not clear. Rubio et al. (1998) in chickens and Nemcova et al. (1999) in weanling pigs reported a prebiotic effect of oligosaccharides. The results suggest that the effect on the Lactobacilli count is related to the presence of the lupin meal or fructooligosaccharides in the diet. Oligosaccharides and NSP should be regarded as being potentially responsible for the changes described in the microbial ecology of the chicken and pig gut. Recent studies using ethanol fractions showed that the addition to the diet of an oligosaccharide fraction from lupin seed improved the performance of chickens and confirmed that oligosaccharides in lupins should not be considered as an anti-nutrient (Hughes et al., 1998; Brenes et al., 2003). Unlike chickens, in pigs, van Barneveld et al. (1997) indicated that ethanol extraction of lupin, to reduce its oligosaccharide content, improved energy and AA digestibility.
Studies using multi-activity enzymes in poultry diets were conducted with different concentration of lupin seeds in the diet. Kocher et al. (2000a) investigated the effect of enzymes on changes in the composition of NSP along the digestive tract. Enzyme addition increased NSP digestibility in the ileum but had no effects on protein digestibility and fermentation in the ileum and ceca. Similarly, Hughes et al. (2000) showed that enzyme addition caused a small improvement in apparent metabolisable energy (AME) of the diet and had a beneficial effect on excreta moisture when the content of lupin kernel isolate in the diet was low (50 g/kg). At higher levels (100 and 150 g/kg) the enzyme partially depolymerised some insoluble NSPs, which resulted in increased ileal viscosity and depressed the AME and growth performance of chickens. Brenes et al. (2002, 2003) using a multienzymatic preparation (proteases, galactosidases and carbohydrases) in a poultry diet improved chick

201
performance and counteracted the negative effect caused by the fibrous components in lupin seed. Enzymes reduced the size of the gastrointestinal tract in birds fed whole, dehulled lupin and lupin hull diets.
The nutritive value of transgenic lupin seed of narrow-leafed lupin seed having a higher methionine content was evaluated using broiler chickens (Ravindran et al., 2002). Because of their methionine content, transgenic lupins have the potential to reduce the amount of supplemental methionine needed in lupin-based poultry diets (saving of 0.6 g/kg in poultry diets containing 250 g lupin/kg). The higher AME value obtained in chicks fed transgenic lupins may be related to its lower content of soluble NSP.
In conclusion, detailed information on the polysaccharide components and the fermentation characteristic of the kernel cell walls of lupin seed have been reported. The presence of soluble sugars and the physico-chemical properties of the fibre fraction in lupin seed may be factors that negatively affect nutrient digestibility and may induce morphological and functional changes in the intestinal tract. Beneficial effects on growth performance and intestinal microflora (Lactobacilli) of lupin oligosaccharides in chicken diets confirm that this fraction must not be considered as an anti-nutrient. More studies are needed to define the prebiotic effect of legume oligosaccharides. Transgenic lupins have the potential to lower the amount of supplemental methionine needed in lupin-based diets. Further studies on the significance of a lower NSP content in transgenic lupins are needed. Chickpeas Chickpea seed is usually grown for human consumption but approximately 20 % of the production is damaged during harvesting and processing. They are sold as a by-product at low prices for livestock feeding. Based on seed colour and site of origin, chickpeas are generally classified as either being Kabuli or desi types. The two chickpea races differ in their nutrient composition Kabuli types have a lower fibre, higher starch and higher fat content than desi types (Gil et al., 1996). To date the animal feed industry has been reluctant to use chickpea seed in pig and poultry diets due to the suspected presence of ANFs such protease and amylase inhibitors, lectins, polyphenols and oligosaccharides (Cerioli et al., 1998) and the lack of information on possible inclusion levels. Recently, Viveros et al. (2001) showed that the inclusion of up to 450 g/kg of Kabuli and up to 150 g/kg of desi chickpea seed in the diet had a negative effect on the performance of chickens, including weight gain. They also affected the length of the digestive tract. In addition incorporation of Kabuli chickpeas into the diet reduced the digestibility of dietary protein and starch, α-amylase and trypsin activities, and its AME. Autoclave treatment of desi chickpeas improved bird performance. This was probably due to a reduction or inactivation of the trypsin, chymotrypsin and α-amylase inhibitor or structural modifications of fibre, which increased the ratio between soluble and insoluble fibre. Farrell et al. (1999) also demonstrated that birds fed steam-pelleted chickpea diets gained more weight and consumed more food than those fed cold-pelleted diets.
Salgado et al. (2001), investigated the digestibility of Kabuli (cream) and desi (brown) chickpeas in the young pig, and evaluated their impact on intestinal morphology and function. Differences in digestibility values were moderate as well their effects on the morphology (villus atrophy and crypt hyperplasia) and function of intestinal tissue (duodenal specific activities of aminopeptidases, lactase, sucrase and alkaline phosphatase). Thacker et al. (2002) compared the nutrient digestibility of 16 varieties of Kabuli and desi chickpeas in swine using the mobile bag technique. They showed that Kabuli chickpeas appeared to have a greater potential as a protein and energy source for use in swine diets than desi chickpeas.

202
Mustafa et al. (2000a) demonstrated that the inclusion up to 300 g/kg of both types of chickpea in pig finishing diets had no detrimental effects on pig performance.
Rubio et al. (2002) measured the effect of a germinated Kabuli chickpea based diet on performance, N utilisation and AA digestibility of the rat. Germination had no detectable effect on these parameters. Rubio et al. (1998) also demonstrated that inclusion of globulins extracted from chickpeas into rat diets increased the relative weight of the gastrointestinal tract, reduced the relative weight of the spleen and thymus, indicating changes in the immune system, and lowered the threonine, leucine and lysine concentration of blood plasma.
In conclusion, the nutritional value of Kabuli and desi chickpeas and their effect on the morphology and the enzymes of the gastrointestinal tract in chickens and pigs have been studied. Biotechnological processes (autoclaving, pelleting and germination) have been used to improve the nutritional value of this seed. Further work is needed to obtain more information on the lower utilisation of chickpea protein and the physico-chemical properties of the fibre fraction, and the possible immunomodulatory or immunogenic effects of chickpea. Oilseeds Linseed Linseed (Linum usitatissimum) or flax, is a potentially valuable protein (200 - 230 g/kg) and energy (400 - 430 g crude fat/kg) source for animal diets (Lee et al., 1995). The majority of the oil is linolenic acid (Panford & Deman, 1990). However, most reports indicate that the addition of linseed to broiler chick diets in amounts above 100 g/kg has adverse effects on performance and nutrient utilisation (Roth-Maier & Kirchgessner, 1995; Treviño et al., 2000; Ortiz et al., 2001; Rodríguez et al., 2001). This is attributed to the presence ANFs and toxic factors in this seed.
Linseed hulls contain substantial amounts of mucilage; about 80 g/kg seed (Bhatty & Cherdkiatgumchai, 1990). It has a high water-absorption capacity with functional properties similar to those of gum Arabic and can increase the viscosity of digesta leading to reduced nutrient availability (Bhatty, 1995; Rodríguez et al., 2001). Mucilage consists of neutral and acidic polysaccharides. The neutral fraction is composed of branched arabinoxylan with a (1-4)-β-D-xylan backbone to which arabinose and galactose side-chains are attached, and an acidic fraction that has a backbone of (1-2)-linked α-L-rhamnopyranosyl and (1-4)-linked D-galactopyranosyluronic acid residues, with side-chains of fucose and galactose residues (Fedeniuk & Biliaderis, 1994).
Recentl work of Rebolé et al. (2002) and Alzueta et al. (2002) confirms that a linseed mucilage extract, when incorporated into a chicken diet, increased the digesta viscosity and negatively influenced fat digestion and N-corrected apparent metabolisable energy (AMEn). Table 3 shows that the substitution of a demucilaged-linseed diet for linseed in the basal diet significantly improved fat digestibility including the major fatty acids and the AMEn content, and reduced jejunal digesta viscosity.
Studies with ground full-fat linseed fed in mash, pellet and crumbled diets when assayed with birds of different ages showed that pelleting or crumbling of the diet significantly increased the AMEn value of linseed and that the difference observed in young versus mature birds was possibly due to a greater tolerance of the latter to linseed. Likewise, the nutritive value of a maize-soybean-linseed diet was not affected by the addition of sepiolite or a multienzyme preparation with high xylanase and/or pectinase activity (Alzueta et al., 2002).

203
Table 3. Effects of diets containing linseed or demucilaged linseed on performance, nutrient digestibility, energy values and jejunal digesta viscosity of broilers chickens (1 to 23 d of age) (Alzueta et al., 2003).
Diets Control Linseed-80 Linseed-160 Dem. Linseed-160Body weight (g) 908a 801b 716c 820b Gain:food ratio 0.712a 0.647b 0.581c 0.678ab Crude fat 0.846a 0.683c 0.536d 0.773b C16:0 0.846a 0.694c 0.553d 0.770b C18:0 0.868a 0.699c 0.555d 0.801b C18:1n-9 0.896a 0.748c 0.609d 0.804b C18:2n-6 0.902a 0.818c 0.718d 0.855b C18:3n-3 0.743a 0.492c 0.433d 0.730b Total fatty acids 0.890a 0.733c 0.598d 0.799b AMEn (MJ/kg) 14.75a 13.58b 12.33c 14.90a Viscosity (cP) 1.9a 8.7b 137.1c 5.4b abc P < 0.05 for data in the same row.
Linseed also contains a dipeptide of glutamic acid and proline (linatine), which is
present in the cotyledons (100 mg/kg) that adversely affects growth and food efficiency in birds because it acts as a pyridoxine antagonist (Mandokhot & Singh, 1983). Its effects can be completely alleviated by administering vitamin B6 to chicks. Finally, cyanogenic glycosides, mainly linustatin and neolinustatin, have also been reported to be involved in reduced chick growth (Mazza & Oomah, 1995).
Organoleptic evaluation of eggs produced by laying hens fed diets containing graded levels of linseed and vitamin E suggest that up to 100 g/kg of the seed used in the birds diet decreases the acceptability of the eggs as a result of changed aroma and flavour. These effects seemed to be accentuated by using high levels of vitamin E in the diets (Leeson et al., 1998). Novak & Scheideler (2001) showed that the addition of linseed to the diet of laying hens did not have any adverse effects on egg production. However, linseed supplementation can significantly alter weight of yolk solids and yolk, and albumen percentages. A reduction in hepatic fat content without a dramatic rise in lipid peroxidation in the liver of laying hens was also produced by inclusion of linseed in the diet (Schumann et al., 2000).
An alternative means of increasing the long-chain n-3 polyunsaturated fatty acid (PUFA) content of pig tissues would be to include α-linolenic acid (18:3n-3), the precursor to eicocosapentaenoic and docosahexaenoic acids, into the feed. A potential commercial source of 18:3n-3 is whole linseed which is about one-third oil, of which just over 50 % is 18:3n-3. Increasing levels of ground linseed in a pig diet for the final 25 days before slaughter increased the content of healthy omega-3 fatty acid in the carcass with no detrimental effects on animal production (Cunnane et al., 1990; Romans et al., 1995a,b; Matthews et al., 2000).
The presence of substantial amounts of mucilage from linseed hulls influences nutrient availability and viscosity of the digesta in chickens fed this product. Enzyme and sepiolite addition do not appear to improve the nutritive value of linseed. Inclusion of linseed in the diet of laying hens causes a reduction in hepatic fat content and significantly alters the yolk and albumen egg percentage. Increasing the linseed content of pig diets up to 100 g/kg has no adverse effect on carcass or meat quality whilst enhancing the level of n-3 fatty acids, which have a potential positive effect in humans. More research is required to evaluate the possible prebiotic effects of mucilage on intestinal microflora and to estimate the impact on human health of the incorporation of linseed oil in monogastric diets.

204
Cottonseed Cottonseed meal (CSM), a by-product of the cotton industry after extraction of oil from cottonseed, is fed to livestock as a valuable protein source. It is used primarily in ruminant diets because of their tolerance to gossypol. Its use in poultry diets has been restricted due to fears of gossypol toxicity and concerns about protein quality (low lysine levels) and high fibre. Gossypol is toxic and is approximately 4 - 17 g/kg by weight of glanded cottonseed, depending of the variety and environmental conditions during growth (Tchiegang & Bourely, 1997). Gossypol has two naphthalene rings with identical constituents that can partially rotate around the bond connecting the rings, forming two identical structures that have no plane of symmetry and differing optical properties [(+) and (-) enantiomers] (Huang et al., 1987). Animal studies, in which the pure enantiomers of gossypol have been used, indicated that the (-) enantiomer may be the most toxic of the two (Chen et al., 1987; Gamboa et al., 2001). Bayley et al. (2000) and Gamboa et al. (2001) conclusively showed a link between the ratio of the two enantiomers and toxicity. Histopathological liver changes in broilers due to gossypol feeding occurs at levels lower than those that affect body weight (Henry et al., 2001a).
Total gossypol in CSM is determined by its concentration in the kernel. The amount of free gossypol is related to temperature, pressure, cooking time and moisture conditions during processing (Forster & Calhoun, 1995). Considerable effort has been devoted to devising practical methods to reduce and detoxify the free gossypol in order to utilize CSM in poultry diets. The expander-solvent extraction process, used by most companies to extract oil from cottonseed, results in CSM with 0.3 to 1.4 g/kg of free gossypol and 10.9 to 11.6 g/kg of bound gossypol. Another technique used to inactivate free gossypol in pigs and poultry diets is to add iron at a 1:1 ratio in relation to the concentration of free gossypol (Boling et al., 1998).
There are several reports showing differences in the tolerance level that may be due to factors such as age and strain of birds, protein and lysine level in the diet. Broilers diets formulated with CSM require higher lysine levels to obtain performance comparable to soybean meal but with slightly higher body fat. The higher level of lysine required is probably due primarily to the unavailability of the lysine bound to gossypol during oil extraction and heating of CSM. Extrusion of CSM is not necessary for use in broiler diets (Henry et al., 2001b). It has been suggested that a plasma total gossypol concentration of 10µg/ml could be used as an upper limit for establishing safe levels of dietary CSM (Gamboa et al., 2001).
Laying hens fed diets containing CSM can produce eggs that have pink albumen discolorations and brown yolks due to the presence of cyclopropenoid fatty acids and free gossypol from the CSM. The beneficial effects obtained by the use of expanded CSM can be lost if soapstock is added back to the meal (Davis et al., 2002) as it is a rich source of free gossypol. The meal can be fed to laying hens at a higher percentage (200-300 g/kg) without causing egg yolk discoloration when this fraction is not added to the CSM. However, there is dramatic variability within a population of laying hens in their susceptibility to gossypol-related egg discoloration due to genetic differences.
High fibre oilseed cakes are generally not used in Europe for pig feeding. However, digestibility experiments using several commercial cotton seed meals in growing pigs diets were performed by Fevrier et al. (2001) to establish the relationship between the ileal digestibility of AAs and the chemical composition of CSM. Their results showed a curvilinear relationship between ileal digestibility of AAs and NSP. Thus the problems

205
arising from ANFs, like gossypol, are probably not the main factors limiting the nutritive value of CSM for pig feeding.
In conclusion, gossypol is the main toxin in cottonseed and forms two identical structures differing in their optical properties (+ and – enantiomers). The use of expander-solvent extracted CSM and the addition of iron to the diet reduces and inactivates the toxic effects of gossypol in broilers. Pink albumen discoloration and brown yolks is caused by addition of CSM to laying hens diets. The NSP composition of CSM appreciably modifies the true digestibility of AAs in pigs. The development of cotton cultivars with a lower total gossypol content or a higher (+)- to (-)-gossypol ratio would be of benefit to the animal feed industry. Sunflower Sunflower (Helianthus annuus L.) is the second largest oilseed crop as a world source of vegetable oil. The by-product from the oil industry is used as an alternative protein source in animal feeds. Its crude protein content depends on the dehulling and oil extraction process and ranges from 290 to 450 g/kg in an inverse relationship to the fibre content (320 to 140 g/kg crude fibre). The use of whole full fat seed, in animal feeds, is limited for technical and nutritional reasons. For example, sunflower seed cannot be ground alone as its preservation after grinding is difficult. Also, its relatively high oil content makes it an unbalanced feedstuff with a very low digestible carbohydrate content and an AA profile which is lysine deficient (Alzueta et al., 1999; San Juan & Villamide, 2001,).
Like other oilseeds, sunflower contains several undesirable ANFs. Chlorogenic acid (CGA), caffeic acid (CA) and quinic acid (QA) are the main acids present in sunflower seed. The CGA and CA represent 700 g/kg of the total acids and are mainly located in the kernel (0.7-2.8 mg/g) (Canibe et al., 1999; Pedrosa et al., 2000). The major phenolic compound in sunflower seed is CGA; its presence has been reported to affect protein digestibility by inhibiting proteolytic enzyme activity (Milic et al., 1968). Interactions between CGA and sunflower proteins may also lower the nutritive value of sunflower protein, since the condensation products are resistant to proteolytic enzymes. Polyphenol oxidase oxidises CGA in the seed to a highly reactive o-quinone intermediate. This can interact with the free NH2 group of lysine (Pedrosa et al., 2000). This interaction could further lower seed nutritive value since sunflower protein is already lysine deficient. However, in experiments with rats and chickens it was found that up to 6 g/kg of CGA did not interfere with AA digestibility and nutritive value. There was no correlation between CGA concentration and protein quality (Eklund et al., 1975; Treviño et al., 1998; Canibe et al., 1999). Perhaps, higher concentrations of dietary chlorogenic acid than those used in these experiments would be necessary to cause detrimental nutritional effects.
Nearly all the studies on sunflower seed in poultry diets have been done with the meal that remains after decortication and oil extraction (Zatari & Sell, 1990; Musharaf, 1991). Many of the studies indicate that sunflower protein could partially replace soybean protein in the diet at an inclusion level in the diet ranging from 120 to 200 g/kg without any adverse effect on performance. Villamide & San Juan (1998) showed that an excreta collection period of 48 h was suitable for determining the true metabolisable energy of sunflower seed (high fibre content) meal based on prediction equations in chickens and that its true AA digestibility was directly related to its crude protein content. Recombined products (a mix of meal and oil to match the original composition of seed) have been compared with full-fat seed when fed to pigs and poultry (Lee et al., 1995; Arija et al., 2000a; San Juan & Villamide, 2001). The recombined material had a better utilisation (energy value and oil digestibility) than full fat

206
seed. This reflected a positive effect of the oil extraction process on substrate accessibility for enzymatic and/or fermentative digestion.
The feeding of full-fat sunflower (FFSS), and their fractions, to livestock has not been extensively investigated as a protein source for broiler chickens. The addition of up 50 g/kg full fat sunflower kernel (FFSK) adversely affected chicken performance and fat digestibility. However, the inclusion of sunflower hulls up to 50 g/kg had no adverse effects (Arija et al., 1998, 1999). The nutritive quality of sunflower protein (AA profile) is poorer than that of soybean but the two were equal when the diet was supplemented with lysine (Alzueta et al., 1999). Likewise, the apparent metabolisable energy content increased significantly as the inclusion level of FFSK was increased. The addition of an enzyme complex to the diet had no effect on the apparent digestion of fat, fatty acids and crude protein (Rodríguez et al., 1998; Rebolé et al., 1999). The addition of 150 g/kg of FFSK to broiler chicken diets also caused alterations to the jejunal mucosa with a shortening and thickening of the villi, hyperplasia and vacuolar degeneration of enterocytes, and hypertrophy and hyperplasia of goblet cells (Arija et al., 2000b).
In conclusion, chlorogenic acid is the major phenolic compound considered as an ANF in sunflower seed. Recent studies indicate that dietary chlorogenic acid at a concentration lower than 6 g/kg did not affect the nutritional quality of the diet. Nutritional evaluation of full-fat sunflower as affected by the oil extraction process has also been evaluated. More research is required to evaluate the effect in animals of feeding higher concentrations of dietary chlorogenic acid and to study new varieties of high oleic acid sunflower seed, which contains much lower levels of linoleic acid than that of ordinary sunflower. This seed can serve as a concentrated source of monounsaturated fatty acids and lead to its incorporation into muscular tissue. The potential health benefit of these fatty acids in the human diet is well known (Pérez-Jiménez, 1995; Roche, 2001). Canola Canola is a variety of rapeseed (Brassica napus) specifically bred to have low concentrations of total glucosinolates (< 30 mmol/kg oil free DM) and erucic acid (< 20 g/kg of total fatty acids). The development of new cultivars of rapeseed that have low levels of these compounds has been the major reason for the renewed interest in its meal, specifically canola meal, as feedstuffs for monogastric animals.
Although canola meal contains significantly less glucosinolates than older seed varieties, there may still be sufficient quantities remaining after processing to cause reduced performance or undesired physiological effects when it is fed to animals for a relatively long period of time or at a relatively high concentration. The presence of ANFs in this seed has been a major quality factor influencing the use of the by-product meal. Glucosinolates per se are not toxic, however, endogenous myrosinase present in the seeds and present in the indigenous gastrointestinal tract flora of the animal can hydrolyse them to yield toxic breakdown products (van Kempen & Jansman, 1994). Depending on the nature of the glucosinolates and the reaction conditions isothiocianates, thiocianates, or nitriles may be formed. These compounds can impair feed intake and growth, and interfere with thyroid and liver function (Campbell & Schöne, 1998). In addition to glucosinolates, others factors such as dietary fibre content (140 g/kg of crude fibre), and the level of tannins, phytic acid and sinapine (ester of sinapic acid and choline) may contribute to decreased animal performance.
Mullan et al. (2000) evaluated 20 samples of canola meal produced in southwestern Australia. The average oil and total glucosinolate levels were 378 g/kg and 14.4 mmol/kg (oil free DM). The study showed that single-press canola meal was a suitable alternative to other

207
protein sources. However, for growing-finishing pigs levels greater than 150 g/kg tended to decrease growth performance and produced thyroid hypertrophy.
There may be scope to include solvent-extracted canola meal into pig diets at greater levels than possible by the use of expeller-extracted meal since some ANFs may be reduced by a solvent extraction procedure including a toasting step. King et al. (2001) examined whether higher levels of solvent-extracted canola could be used compared to expeller-extracted meal in diets for different classes of pigs. Their results with weaner pigs, grower-finisher pigs, and lactating sows, respectively, indicated that low glucosinolate (<5 µmol/g), solvent extracted canola meal could be included at levels up to 250, 300, and 200 g/kg, respectively, without adversely affecting pig performance.
Opalka et al. (2001) investigated the effect of long-term feeding with graded levels of low glucosinolate rapeseed on the reproductive performance and endocrine status of gilts and their piglets. Long-term feeding with graded levels of glucosinolates (growth and lactation, 0.45 or 0.78 and pregnancy, 0.16 and 0.32 mM glucosinolates/kg diet) did not affect reproductive performance of gilts and primiparous sows. The rapeseed (RSM) diets, however, did affect the mean concentrations of plasma free thyroxine, triiodothyronine, prolactin, progesterone and androstenedione in gilts during the midluteal phase of the first oestrus cycle. In contrast, there was no significant difference in serum concentrations of hormones examined in piglets originating from the control and the RSM groups. Schöne et al. (2001) also showed that glucosinolates and their degradation compounds may affect the thyroid and the mammary glands resulting in a lower iodine (I) milk transfer and higher renal and intestinal I excretion.
The ileal digestibility of N and AA in canola meal for pigs has been widely studied (Grala et al., 1998; de Lange et al., 1998). Special efforts have recently been made to distinguish between endogenous and dietary N losses at the end of the ileum for pigs (Grala et al., 1998, 1999). Jondreville et al. (2000) showed that the true digestibility (TD) of N and AA was lower in canola meal compared with sunflower meal (SFM) and soybean meal (SBM). The high fibre concentration in RSM and SFM, as well as the large amount of N bound to the fibre in pigs fed RSM, probably explains these results. Differences in TD of AA between canola meal and soybean meal were related to different recoveries of both endogenous and dietary AAs.
The inclusion of enzymes (Thacker, 2001) and extrusion processing of canola meal based diets did not improve their nutritional value for growing-finishing pigs (Keady & O´Doherty, 2000).
Full-fat canola seed was evaluated as an alternative source in diets fed to weaner and grower-finisher pigs (Brandt et al., 1999). There was no significant difference in the intake, growth rate or feed conversion of weaner pigs using 240 g/kg of full-fat rapeseed meal compared to a control diet based on full-fat soybean. However, an inclusion level of 160 g/kg in grower-finisher pig diets was recommended. The back fat of pigs consuming diets with 160 and 240 g/kg of full-fat canola meal had 13 % higher iodine numbers than pigs that received 0 and 80 g/kg of full-fat canola in their diets.
Based of concerns over possible accumulation of triglycerides in the heart as a result of small amounts of erucic acid in the oil the effect of dietary canola oil (low erucic acid rapeseed) was studied by Innis & Dyer (1999) and Green & Innis (2000). The modest accumulation of erucic acid associated with feeding canola oil was not associated with any biochemical evidence of heart triglyceride accumulation at 10 and 18 days of age in piglets.
The nutritive value of the meal derived from recently developed varieties of canola has been evaluated in broilers (Slominski et al., 1999). Their results showed yellow-seeded B. napus was superior to its black-seeded counterpart or other yellow-seeded species

208
(B. juncea, B. rapa) with regard to protein, fibre, and true metabolisable energy content and overall nutrient utilisation as determined by a superior feed to gain ratio in broiler chickens.
Janjecic et al. (2002) showed that inclusion of 300 g/kg of rapeseed in the diet affected chicken performance, increased liver size and caused slight to medium hypertrophy of thyroid epithelial tissue as well as slight thymus and cloacal bursa follicle hypertrophy.
Pre-press-solvent extraction is currently the most effective method of extracting oil from canola meal. Recently, Mustafa et al. (2000b) reported that processing, especially at the desolventing/toasting stage, had a significant impact on protein degradation rates in the rumen. It was hypothesized that the moisture incorporated into canola meal during desolventing, as spare steam, promotes toasting (Maillard reactions) and may contribute to losses in AA content and reduce digestibility. Newkirk & Classen (2002) showed that desolventing, without spare steam, produced a non-toasted meal that gave improved broiler growth and feed efficiency compared to toasted canola meal. Newkirk et al. (2003a) also showed that the desolventing/toasting process reduced lysine content, apparent ileal digestibility of most AAs and the metabolisable energy in broilers fed canola meal products, and also increased meal variability. Finally, a study conducted by Newkirk et al. (2003b) confirmed that non-toasted canola meals contains higher levels of digestible AAs than the conventionally toasted canola meal that is commonly produced by commercial crushing plants in Western Canada.
A second important factor in determining the suitability of canola meal for poultry diets, after considering processing condition, is the concentration of indigestible oligosaccharides (25 g/kg) and NSP (180 g/kg) of which 15 g/kg is soluble (Bell, 1993). The addition of enzymes to canola-based diets had no significant effect on growth performance and AME. However, the addition of enzymes (cellulase, β-glucanase, xylanase) significantly reduced the concentration of soluble NSP in the jejunum (Kocher et al., 2000b). Studies with a similar multiactivity enzyme preparation showed the potential of these products to enhance the digestion of canola meal polysaccharides in vitro (Slominski & Campbell, 1990; Simbaya et al., 1996). Newkirk & Classen (2001) designed experiments to investigate the effects of prior in vitro hydrolysis of phytate on the nutritional value of canola meal for broiler chickens. There was no significant effect on the performance of broilers fed pre-treated meal with purified or crude phytase.
In conclusion, significant quantities of glucosinolates still remain in canola meal after processing. The desolventing/toasting process has a significant negative impact on AA digestibility in chickens. Modification of the method by the elimination of spare steam and reducing the level of browning and toasting of the meal results in higher levels of digestible AAs and improved broiler performance over that obtained with conventional toasted CM. Before a general recommendation to eliminate CM toasting can be made, the impact of this process must be studied on other animal species such as pigs and cattle. Further research is also needed to study the effect of ANFs in canola meal in pig diets.

209
Conclusions and future prospects Legume seeds The nutritional value of different legume seeds has been improved by the development of varieties that have reduced levels of ANFs. There are, however, still indications that low levels of ANFs may be present in some legume seed and that they interfere with protein digestibility and metabolism including their effects on the loss of endogenous protein in the digestive tract and consequences for N metabolism. The immunogenic effects of some legumes protein, especially in young animals, also requires more attention.
The potential use of some ANFs and legume proteins as functional foods should be considered. The effect of legume seed or specific legume seed constituents on microbial activity in different parts of the digestive tract and it’s consequences for gastrointestinal health and function is also an area, which needs exploration. Oilseeds There is limited data on the physiological effects of ANFs present in rapeseed on pig reproduction. This seems to justify the initiation of more detailed studies on this seed. Strategies for the economic extraction, modification and clinical evaluation of different natural compounds in linseed need to be developed to further evaluate the functional properties of the seed. Whole seed is a potential commercial source of linoleic acid for pigs and poultry. More detailed studies on the influence of the fatty acid composition of oilseeds on that of the various tissues in chickens and pigs also need to be considered.
In conclusion, not much new information on the mode of action of the various (classical) ANFs has been published over the last five years. Further development of the functional properties of various legume and oil seeds is an interesting new area, especially in relation to the nutrition and (intestinal) health of the target animal species and their consequences for human health. References Abd El-Haddy, E.A. & Habiba, R.A. 2003. Effect of soaking and extrusion conditions on
antinutrients and protein digestibility of legume seeds. Lebensmittel-Wissenschaft und-Technologie 36, 285-293.
Allen, J.G. 1998. Toxins and lupinosis. In Lupins as Crop Plants. Biology, Production and Utilization. J.S. Gladstones, C. Atkins & J. Hamblin (Eds). CAB International, Wallingford. pp. 411-435
Alzueta, C., Ortiz, L., Rebolé, A., Rodríguez, M.L., Centeno, C. & Treviño, J. 2002. Effects of removal of mucilage and enzyme or sepiolite supplement on the nutrient digestibility and metabolisable energy of a diet containing linseed in broiler chickens. Animal Feed Science and Technology 97, 169-181.
Alzueta, C., Treviño, J., Rebolé, A., Ortiz, L.T., Rodríguez, M.L. & Centeno, C., 1999. Full-fat sunflower seed as a protein source for broiler chicks. Journal of the Science of Food and Agriculture 79, 1681-1686.
Arija, I., Brenes, A., Viveros, A. & Elices, R. 1998. Effects of inclusion of full-fat sunflower kernels and hulls in diets for growing broiler chickens. Animal Feed Science and Technology 70, 137-149.

210
Arija, I., Viveros, A., Brenes, A. & Canales, R. 1999. Estudio del valor nutritivo de la semilla de girasol entera descascarillada en raciones de pollos broiler y su efecto sobre la concentración de ácidos grasos en la grasa abdominal. Archivos de Zootecnia 48, 249-259.
Arija, I., Viveros, A., Brenes, A., Canales, R., Pizarro, M. & Castaño, M. 2000b. Histological alterations in the intestinal epithelium caused by the inclusion of full-fat sunflower kernels in broiler chicken diets. Poultry Science 79, 1332-1334.
Arija, I., Viveros, A., Canales, R. & Brenes, A. 2000a. Efecto de la inclusión de la semilla de girasol entera y reconstituida en raciones de pollos broiler. Investigación Agraria: Producción y Sanidad Animales 15, 81-94
Bach Knudsen, K.E., Canibe, N. & Sorensen, H. 2001. Limitations in the use of peas (Pisum sativum L) in diets for pigs and possible ways to overcome some of these problems. In Towards the Sustainable Production of Healthy Food, Feed and Novel Products. AEP (Ed). Proceedings 4th European Conference on Grain Legumes, 8-12 July 2001, Cracow, 122-123.
Bailey, C.A., Stipanovic, R.D., Ziehr, M.S., Haq, A.U., Sattar, M., Kubena, L.F., Kim, H.L. & Vieira, R.M. 2000. Cottonseed with a high (+)- to (-)-gossypol enantiomer ratio favourable to broiler production. Journal of Agricultural and Food Chemistry 48, 5692-5695.
Bell, J.M. 1993. Factors affecting the nutritional value of canola meal: A review. Canadian Journal of Animal Science 73, 679-697.
Bhatty, R.S. 1995. Nutrient composition of whole seed and flaxseed meal. In Flax Seed in Human Nutrition. C.S. Cunnane & L.U. Thompson (Eds). AOCS Press, Champaign, IL, 2242.
Bhatty, R.S. & Cherdkiatgumchai, P. 1990. Compositional analysis of laboratory prepared and commercial samples of linseed meal and of hull isolated from flax. Journal of the American Oil Chemists’ Society 67, 79-84.
Boling, S.D., Edwards, H.M., Emmert, J.L., Biehl, R.R. & Baker, D.H. 1998. Bioavailability of iron in cottonseed meal, ferric sulphate, and two ferrous sulphate by-products of the galvanizing industry. Poultry Science 77, 1388-1392.
Brandt, D.A., Merwe, J.P. van der & Brand, T.S. 1999. Full-fat canola meal as a protein source for weaner and grower-finisher pig. Australian Journal of Experimental Agriculture 39, 21-28.
Brenes, A., Marquardt, R.R., Guenter, W. & Viveros, A. 2002. Effect of enzyme addition on the performance and gastrointestinal tract size of chicks fed lupin seed and their fractions. Poultry Science 81, 670-678.
Brenes, A., Slominski, B., Marquardt, R.R., Guenter, W. & Viveros, A. 2003. Effect of enzyme addition on the digestibilities of cell wall polysaccharides and oligosaccharides from whole, dehulled and ethanol extractions of white lupin in chickens. Poultry Science 82, 1716-1725.
Campbell, L.D. & Shöne, F., 1998. Effects of antinutritional factors in rapeseed. In Recent Advances of Research in Antinutritional Factors in Legume Seeds and Rapeseed. A.J.M. Jansman, G.D. Hill, J. Huisman, A.F.B. van der Poel (Eds). Proceedings 3rd International Workshop on Antinutritional Factors in Legume Seeds and Rapeseed, Wageningen 8–10 July 1998, 185-198.
Canibe, N. & Bach-Knudsen, K.E. 2001. Degradation and physicochemical changes of barley and pea fibre along the gastrointestinal tract of pigs. Journal of the Science of Food and Agriculture 82, 27-39.

211
Canibe, N., Pedrosa, M.M., Robredo, L.M. & Bach-Knudsen, K.E. 1999. Chemical composition, digestibility and protein quality of 12 sunflower (Helianthus annuus L) cultivars. Journal of the Science of Food and Agriculture 79, 775-1782.
Carbonaro, M., Grant, G., Cappelloni, M. & Pusztai, A. 2000. Perspective into factors limiting in vivo digestion of legume proteins: antinutritional compounds or storage proteins. Journal of Agricultural and Food Chemistry 48, 742-749.
Carré, B. 2000. Effets de la taille des particules alimentaires sur les processus digestifs chez les oiseaux d’élevage. Productions Animales 12, 131-136.
Carré, B. 2002. Processing of peas for poultry. Saskatchewan Pulse Growers Symposium. Cerioli, C., Fiorentini, L., Prandini, A. & Piva, G. 1998. Antinutritional factors and nutritive
value of different cultivars of pea, chickpea and faba bean. In Recent Advances of Research in Antinutritional Factors in Legume Seeds and Rapeseeds. A.J.M. Jansman, G.D. Hill, J. Huisman & A.F.B. van der Poel (Eds). Proceedings 3rd International Workshop on Antinutritional Factors in Legume Seeds and Rapeseed, Wageningen 8–10 July 1998, 43-46.
Chen, Q.Q., Chen, H. & Lei, H.P. 1987. Comparative study on the metabolism of optical gossypol in rats. Journal Ethnopharmacology 20, 31-37.
Cunnane, S.C., Stitt, P.A., Ganguli, S. & Armstrong, J.K. 1990. Raised omega-3 fatty acid levels in pigs fed flax. Canadian Journal of Animal Nutrition 70, 251-254.
Davis, A.J. & Dale, N. 2002. The use of cottonseed meal with or without added soapstock in laying hen diets. Journal of Applied Poultry Research 11, 127-133.
De Lange, C.F.M., Gabert, V.M., Gillis, D. & Patience, J.F. 1998. Digestible energy and apparent ileal amino acid digestibilities in regular or partial mechanically dehulled canola meal samples fed to growing pigs. Canadian Journal of Animal Science 78, 641-648.
Dunshea, F.R., Gannon, J.N., Barneveld, R.J. van, Mullan, B.P., Campbell, R.G. & King, R.H. 2001. Dietary lupins (Lupinus angustifolius and Lupinus albus) can increase digesta retention in the gastrointestinal tract of pigs. Australian Journal of Agricultural Research 52, 593-602.
Fan, M.Z. & Sauer, W.C. 1999. Variability of apparent ileal amino acid digestibility in different pea samples for growing-finishing pigs. Canadian Journal of Animal Science 79, 467-475.
Farrell, D.J., Perez-Maldonado, R.A. & Mannion, P.F. 1999. Optimum inclusion of field beans, chick peas and sweet lupins in poultry diets. II. Broiler experiment. British Poultry Science 40, 674-680.
Fedeniuk, R.W. & Biliaderis, C.G. 1994. Composition and physicochemical properties of linseed (Linum usitatissimum L.). Journal of Agricultural and Food Chemistry 42, 40-247.
Fevrier, C., Lechevestrier, Y., Lebreton, Y. & Jaguelin-Peyraud, Y. 2001. Prediction of standardized ileal true digestibility of amino acids from the chemical composition of oilseeds meals in the growing pig. Animal Feed Science and Technology 90, 103-115.
Flis, M., Sobotka, W., Purwin, C. & Zdunczyk, Z. 1999. Nutritional value of diets containing filed beans (Vicia faba L.) seeds with high or low proanthocyanidin levels for pig. Journal of Animal and Feed Sciences 8, 171-180.
Forster, L.A. Jr. & Calhoun, M.C. 1995. Nutrient values for cottonseed products deserve new look. Feedstuffs 67 (44), October, 23.
Gamboa, D.A., Calhoun, M.C., Kuhlman, S.W., Haq A.U. & Bailey, C.A. 2001. Tissue distribution of gossypol enantomers in broilers fed various cottonseed meals. Poultry Science 80, 920-925.

212
Gil, J., Nadaj, S., Moreno, M.T. & Haro, A. 1996. Variability of some physico-chemical characters in desi and Kabuli chickpea types. Journal of the Science of Food and Agriculture 71, 179-184.
Gdala, J., Buracewska, L. & Grala, W. 1992. The chemical composition of different types and varieties of pea and the digestion of their protein in pigs. Journal of Animal and Feed Sciences 1, 71-79.
Grala, W., Verstegen, M.W.A., Jansman, A., Huisman, J. & Leeuwen, P. van, 1998. Ileal apparent protein and amino acid digestibilities and endogenous nitrogen losses in pigs fed soy-bean and rapeseed products. Journal of Animal Science 76, 557-568.
Grala, W., Verstegen, M.W.A., Jansman, A., Huisman, J. & Leeuwen, P. van, 1999. Apparent protein digestibility and recovery of endogenous nitrogen at the terminal ileum of pigs fed diets containing various soybean products, pea or rapeseed hulls. Animal Feed Science and Technology 80, 231-245.
Green, T.J. & Innis, S.M. 2000. Low erucic acid canola oil does not induce heart triglyceride accumulation in neonatal pigs fed formula. Lipids 35, 607-612.
Grosjean, F., Barrier-Guillot, B., Bastianelli, D., Rudeaux, F., Bourdillon, A. & Peyronnet, C. 1999. Feeding value of three categories of pea (Pisum sativum, L.) for poultry. Animal Science 69, 591-599.
Grosjean, F., Jondreville, C., Williatte-Hazouard, I., Skiba, F., Cacarrouée, B. & Gatel, F. 2000. Ileal digestibility of protein and amino acids of feed peas with different trypsin inhibitor activity in pigs. Canadian Journal of Animal Science 80, 643-652.
Hedemann, M.S., Welham, T., Boisen, S., Canibe, N., Bilham, L. & Domoney, C. 1999. Studies on the biological responses of rats to seed trypsin inhibitors using near-isogenic lines of Pisum sativum L (pea). Journal of the Science of Food and Agriculture 79, 1647-1653.
Henry, M.H., Pesti, G.M., Bakall, R., Lee, J., Toledo, R.T., Eitenmiller, R.R. & Phillips, R.D. 2001. The performance of broilers chicks fed diets containing extruded cottonseed meal supplemented with lysine. Poultry Science 80, 762-768.
Henry, M.H., Pesti, G.M. & Brown, T.P. 2001. Pathology and histopathology of purified gossypol toxicity in broiler chicks. Avian Diseases 45, 598-604.
Huang, L.D., Zheng, D.K. & Si, Y.K. 1987. Resolution of racemic gossypol. Journal Ethnopharmacology 20, 13-20.
Hughes, R.J., Barneveld, R.J. van & Choct, M. 1998. Factors influencing the nutritive value of lupin for broiler chickens. Chicken Meat Research Committee Final Report No. DAS10CM. Rural Industries Research and Development Corporation, Canberra, Australia.
Hughes, R.J., Choct, M., Kocher A. & Barneveld, R.J. van, 2000. Effect of food enzymes on AME and composition of digesta from broiler chickens fed on diets containing non-starch polysaccharides isolated from lupin kernel. British Poultry Science 41, 318-323.
Innis, S.M. & Dyer, R.A. 1999. Dietary canola oil alters haematological indices and blood lipids in neonatal piglets fed formula. Journal of Nutrition 129, 1261-1268.
Janjecic, Z., Grbesa, D., Muzic, S., Curic, S., Rupic, V., Liker, B., Dikic, M., Antunovic, B. & Zupanic, D. 2002. Influence of rapeseed on productivity and health of broiler chicks. Acta Veterinaria Hungarica 50, 37-50.
Jansman, A.J.M., Huisman, J. & Poel, A.F.B. van der, 1993. Ileal and faecal digestibility in piglets of faba beans (Vicia faba L.) varying in tannin content. Animal Feed Science and Technology 42, 83-96.

213
Jansman A.J.M. & Verstegen, M.W.A., 2002. Potential of peas as alternative for other protein sources in diets for pigs. Proceeding Eastern Conference. Guelph, Canada. 29-44.
Joa, P.S., Freire, J.B., Ferreira, R.B., Seabra, M., Teixeira, A.R., Toullec, R. & Lallés, J.P. 2002. Legume proteins of the vicilin family are more immunnogenic than those of legumin family in weaned piglets. Food and Agricultural Immunology 14, 51-63.
Jondreville, C., Broecke, J. van den, Gatel, F., Grosjean, F., Cauwenberghe, S. van, & Seve, B. 2000. Ileal amino acid digestibility and estimates of endogenous amino acid losses in pigs fed rapeseed meal, sunflower meal and soybean meal. Canadian Journal of Animal Science 80, 495-506.
Juskiewicz, J., Wroblewska, M., Zhaki, K., Zdunczyk, Z. & Hussein, L. 2001. Biological activities of faba beans proanthocyanidins. Acta Alimentaria 30, 63-69.
Keady, U. & O´Doherty, J.V. 2000. The effect of extrusion on the nutritive value of rapeseed meal for growing and finishing pigs. Irish Journal of Agricultural and Food Research 39, 419-431.
King, R.H., Eason, P.E., Kerton, D.K. & Dunshea, F.R. 2001. Evaluation of solvent-extracted canola meal for growing pigs and lactating sows. Australian Journal of Agricultural Research 52, 1033-1041.
Kocher, A., Choct, M., Hughes, R.J. & Broz, J. 2000a. Effect of food enzymes on utilisation of lupin carbohydrates by broilers. British Poultry Science 41, 75-82.
Kocher, A., Choct, M., Porter, M.D. & Broz, J. 2000b. The effects of enzyme addition to broiler diets containing high concentration of canola or sunflower meal. Poultry Science 79, 1767-1774.
Lallés, J.P. & Jansman, A.J.M. 1998. Recent progress in the understanding of the mode of action and effects of antinutritional factors from legume seeds in non-ruminants farm animals. In Recent Advances of Research in Antinutritional Factors in Legume Seeds. A.J.M. Jansman, G.D. Hill, J. Huisman & A.F.B. van der Poel, (Eds). Proceedings 3rd International Workshop on Antinutritional Factors in Legume Seeds and Rapeseed, Wageningen 8–10 July 1998, 219-232.
Lee, K., Qi, G.H. & Sim, J.S. 1995. Metabolisable energy and amino acid availability of full-fat seeds, meals and oils of flax and canola. Canadian Journal of Animal Science 71, 897-903.
Leeson, S., Caston, L. & MacLaurin, T. 1998. Organoleptic evaluation of eggs produced by laying hens fed diets containing graded levels of flaxseed and vitamin E. Poultry Science 77, 1436-1440.
Mandokhot, V.M. & Singh, N. 1983. Studies on linseed (Linum usitatissimum) as a protein source. 2. Evidence of toxicity and treatments to improve quality. Journal of the Science of Food and Agriculture 20, 291-295.
Mariscal-Landín, G., Lebreton, Y. & Seve, B. 2002. Apparent and standardised true ileal digestibility of protein and amino acids from faba bean, lupin and pea, provided as whole seeds, dehulled or extruded in pig diets. Animal Feed Science and Technology 97, 183-198.
Marquardt, R.R. 1989. Dietary effects of tannins, vicine and convicine. In Recent Advances of Research in Antinutritional Factors in Legume Seeds. J Huisman, T.F.B. van der Poel & I.E. Liener (Eds). Proceedings 1st International Workshop on Antinutritional Factors in Legume Seeds, Wageningen, 23–25 November 1988, 141-155.
Matthews, K.R., Homer, D.B., Thies, F. & Calder, P.C. 2000. Effect of whole seed (Linum usitatissimum) in the diet of finishing pigs on growth performance and on the quality and acid composition of various tissues. British Journal of Nutrition 83, 637-643.

214
Mazza, G. & Oomah, B.D. 1995. Flaxseed, dietary fibre and cyanogens. In Flax Seed in Human Nutrition. S.C. Cunnane, & L.U. Thompson (Eds). AOCS Press, Champaign, IL, .2242.
Milic, B., Stojanovic, S., Vucurevic, N. & Turcic, M. 1968. Chlorogenic and quinic acids in sunflower meal. Journal of the Science of Food and Agriculture 19, 108-113.
Mullan, B.P., Pluske, J.R., Allen, J. & Harris, D.J. 2000. Evaluation of Western Australian canola meal for growing pigs. Australian Journal of Agricultural Research 51, 547-553.
Musharaf, N.A. 1991. Effect of graded levels of sunflower seed meal in broiler diets. Animal Feed Science and Technology 33, 129-137.
Mustafa, A.F., Christensen, D.A., Mckinnon, J.J. & Newkirk, R. 2000b. Effects of stage of processing of canola seed on chemical composition and in vitro protein degradability of canola meal and intermediate products. Canadian Journal of Animal Science 80, 211-214.
Mustafa, A.F., Thacker, P.A., Mckinnon, J.J., Christensen, D.A. & Racz, V.J., 2000a. Nutritional value of feed grade chickpeas for ruminants and pigs. Journal of the Science of Food and Agriculture 80, 1581-1588.
Nemcova, R., Bomba, A., Gancarcicova, S., Eric, R. & Guba, P. 1999. Study of the effect of Lactobacillus paracasei and fructooligosaccharides on the faecal microflora in weanling pigs. Berliner und Munchener Tierarzliche Wochenscherift 112, 225-228.
Newkirk, R.W. & Classen, H.L. 2001. The non-mineral nutritional impact of phytate in canola meal fed to broiler chicks. Animal Feed Science and Technology 91, 115-128.
Newkirk, R.W. & Classen, H.L. 2002. The effects of toasting canola meal on body weight, feed conversion efficiency, and mortality in broiler chickens. Poultry Science 81, 815-825.
Newkirk, R.W., Classen, H.L. & Edney, M.J. 2003a. Effects of prepress-solvent extraction on the nutritional value of canola meal for broiler chickens. Animal Feed Science and Technology 104, 111-119.
Newkirk, R.W., Classen, H.L., Scott, T.A. & Edney, M.J. 2003b. The digestibility and content of amino acids in toasted and non-toasted canola meals. Canadian Journal of Animal Science 83, 131-139.
Novak, C. & Scheideler, S.E. 2001. Long term effects of feeding flaxseed-based diets. 1. egg production parameters, components, and eggshell quality in two strains of laying hens. Poultry Science 80, 1480-1489.
O’Doherty, J.V. & Keady, U. 2001. The effect of expander processing and extrusion on the nutritive value of peas for pigs. Animal Science 72:43-53.
Olkowski, A.A., Olkowski, B.I., Amarowicz, R. & Classen, H.L. 2001. Adverse effect of dietary lupine in broiler chickens. Poultry Science 80, 621-625.
Opalka, M., Dusza, L., Koziorowski, M., Staszkiewicz, J., Lipiñski, K. & Tywoñczuk, J. 2001. Effect of long-term feeding with graded levels of low glucosinolate rapeseed meal on endocrine status of gilts and their piglets. Livestock Production Science 69, 233-243.
Ortiz, L., Rebolé, A., Alzueta, C., Rodríguez, M.L. & Treviño, J., 2001. Metabolisable energy value and digestibility of fat and fatty acids in linseed determined with growing broiler chickens. British Poultry Science 42, 57-63.
Owusu-Asiedu, A., Baidoo, S.K. & Nyachoti, C.M. 2002. Effect of heat processing on nutrient digestibility in pea and supplementing amylase and xylanase to raw, extruded or micronized pea-based diets on performance of early-weaned pig. Canadian Journal of Animal Science 82, 367-374.

215
Panford, J.A. & Deman, J.M. 1990. Determination of oil content of seeds by NIR: influence of fatty acid composition on wavelength selection. Journal of American Oil Chemists’ Society 67, 473-482.
Pedrosa, M.M., Muzquiz, M., García-Vallejo, C., Burbano, C., Cuadrado, C., Ayet, G. & Robredo, L.M., 2000. Determination of caffeic and chlorogenic acids and their derivatives in different sunflower seeds. Journal of the Science of Food and Agriculture 80, 459-464.
Pérez-Jiménez, F., Espino, A., López-Segura, F., Blanco, J., Ruiz, V., Prada, J.L., López-Miranda, J., Jiménez, J. & Ordovas, J.M., 1995. Lipoprotein concentration in normo-lipidemic male consuming olive oil and oleic acid-rich sunflower oil. American Journal of Clinical Nutrition 62, 769-775.
Ravindran, V., Tabe, L.M., Molvig, L., Higgins, T.V.J. & Bryden, W.L. 2002. Nutritional evaluation of transgenic high-methionine lupins (Lupinus angustifolius L) with broiler chickens. Journal of the Science of Food and Agriculture 82, 280-285.
Rebolé, A., Rodríguez, M.L., Alzueta, C., Ortiz, L.T. & Treviño, J. 1999. A short note on effect of enzyme supplement on the nutritive value of broiler chick diets containing maize, soybean meal and full-fat sunflower seed. Animal Feed Science and Technology 78, 153-158.
Rebolé, A., Rodríguez, M.L., Ortiz, L., Alzueta, C., Centeno, C. & Treviño, J. 2002. Mucilage in linseed: effects on the intestinal viscosity and nutrient digestion in broiler chicks. Journal of the Science of Food and Agriculture 82, 1171-1176.
Roche, H.M., 2001. Olive oil, high-oleic acid sunflower oil and coronary heart disease. British Journal of Nutrition 85: 3-4.
Rodríguez, M.L., Alzueta, C., Rebole, A., Ortiz, L., Centeno, C. & Treviño, J., 2001. Effect of inclusion level of linseed on the nutrient utilisation of diets for growing broiler chickens. British Poultry Science 42, 368-375.
Rodríguez, M.L., Ortiz, L.T., Treviño, J., Rebole, A., Alzueta, C. & Centeno, C. 1998. Studies on the nutritive value of full-fat sunflower seed in broiler chicks diets. Animal Feed Science and Technology 71, 341-349.
Romans, J.R., Johnson, R.C., Wulf, D.M., Libal, G.W. & Costello, W.J. 1995a. Effects of ground flaxseed in swine diets on pig performance and on physical and sensory characteristics and omega-3 fatty acid content of pork: I. Dietary level of flax seed. Journal of Animal Science 73, 1982-1986.
Romans, J.R., Wulf, D.M., Johnson, R.C., Libal G.W. & Costello, W.J. 1995b. Effects of ground flaxseed in swine diets on pig performance and on physical and sensory characteristics and omega-3 fatty acid content of pork: II. Duration of 15 % dietary flaxseed. Journal of Animal Science 73, 1987-1999.
Roth-Maier, D.A. & Kirchgessner, M. 1995. Feeding of full-fat flax seed and sunflower seeds to fattening. Archiv für Geflügelkunde 59, 319-322.
Rubio, L.A. 2000. Physiological effects of legume storage proteins. Nutrition Abstracts and Reviews A 70, 197-204.
Rubio, L.A. 2003. Carbohydrates digestibility and faecal nitrogen excretion in rats fed raw or germinated faba bean (Vicia faba)- and chickpea (Cicer arietinum)-based diets. British Journal of Nutrition 90, 301-310.
Rubio, L.A., Brenes, A. & Centeno, C. 2003. Effect of feeding growing broiler chickens with practical diets containing sweet lupins (Lupinus angustifolius) seed meal. British Poultry Science 44: 391-397.

216
Rubio, L.A., Grant, G., Daguid, T., Brown, D., Bardocz, S. & Pusztai, A. 1998. The nutritional utilization by rat of chickpea (Cicer arietinum) meal and its isolated globulin proteins is poorer than that of defatted soybean or lactalbumin. Journal of Nutrition 128, 1042-1047.
Rubio, L.A., Muzquiz, M., Burbano, C., Cuadrado, C. & Pedrosa, M. 2002a. High apparent ileal digestibility of amino acids in raw and germinated faba bean (Vicia faba) and chickpea (Cicer arietinum)-based diets for rats. Journal of the Science of Food and Agriculture 82, 1710-1717.
Rubio, L.A. & Seiquer, I. 2002. Transport of amino acids from in vitro digested legume proteins or casein in Caco-2 cell cultures. Journal of Agricultural and Food Chemistry 50, 5002-5005.
Salgado, P., Lallès, J.P., Toullec, R., Mourato, M., Cabral, F. & Freire, J.P.B. 2001. Nutrient digestibility of chickpea (Cicer arietinum) seeds and effects on the small intestine of weaned piglets. Animal Feed Science and Technology 91, 197-212.
Salgado, P., Freire, J.P.B., Mourato, M., Cabral, F., Toullec, R. & Lallés, J.P. 2002a. Comparative effects of different legume protein sources in weaned piglets: nutrient digestibility, intestinal morphology and digestive enzymes. Livestock Production Science 74, 191-202.
Salgado,P., Martins, J.M., Carvalho, F., Abreu, M., Freire, J.P.B., Toullec, R., Lallès, J.P. & Bento, O., 2002b. Component digestibility of lupin (Lupinus angustifolius) and pea (Pisum sativum) seeds and effects on the small intestine and body organs in anastomosed and intact growing pigs. Animal Feed Science and Technology 98, 187-201.
Salgado, P., Montagne, L., Freire, J.P.B., Ferreira, R.B., Texeira, A., Bento, O., Abreu, M.C., Toullec, R. & Lallès, J.P. 2002c. Legume grains ileal losses of specific endogenous serine-protease proteins in weaned pigs. Journal of Nutrition 132, 1913-1920.
San Juan, L.D. & Villamide, M.J. 2001. Nutritional evaluation of sunflower products for poultry as affected by the oil extraction process. Poultry Science 80, 431-437.
Schöne, F., Leitere, M., Hartung, H., Jahreis, G. & Tischendorf, F. 2001. Rapeseed glucosinolates and iodine in sows affect the milk iodine concentration and the iodine status of piglets. British Journal of Nutrition 85, 659-670.
Schumann, B.E., Squires, E.J. & Leeson, S. 2000. Effect of dietary flaxseed, flax oil and n-3 fatty acid supplement on hepatic and plasma characteristics relevant to fatty liver haemorragic syndrome in laying hens. British Poultry Science 41, 465-472.
Simbaya, J., Slominski, B.A., Guenter, W., Morgan, A. & Campbell, L.D. 1996. The effects of protease and carbohydrase supplementation on the nutritive value of canola meal for poultry-in vitro and in vivo studies. Animal Feed Science and Technology 61, 219-234.
Slominski, B.A. & Campbell, L.D. 1990. Non-starch polysaccharides of canola meal: Quantification, digestibility in poultry and potential benefit of dietary enzyme supplementation. Journal of the Science of Food and Agriculture 53, 175-184.
Slominski, B.A., Simbaya, J., Campbell, L.D., Rakow, G. & Guenter, W. 1999. Nutritive value for broilers of meals derived from newly developed varieties of yellow-seeded canola. Animal Feed Science and Technology 78, 249-262.
Tchiegang, C. & Bourely, J. 1997. Critical studies on the conventional official colorimetric method of determination of free gossypol in cottonseed. Rivista Italiana delle Soslanze Grasse 74, 24 7-251.
Thacker, P.A. 2001. Effect of enzyme supplementation on the performance of growing-finishing pigs fed barley-based diets supplemented with soybean meal or canola meal. Asian-Australian Journal of Animal Science 14, 1008-1013.

217
Thacker, P.A., Qiao, S. & Racz, V.J. 2002. A comparison of the nutrient digestibility of desi and Kabuli chickpeas fed to swine. Journal of the Science of Food and Agriculture 82, 1312-1318.
Thacker, P.A. & Racz, V.J. 2001. Performance of growing/finishing pigs fed hulled and dehulled peas with and without dietary enzymes. Asian-Australian Journal of Animal Science 14, 1434-1439.
Treviño, J., Rebolé, A., Rodríguez, M.L., Ortiz, L.T., Centeno, C. & Alzueta, C. 1998. Nutritional effect of chlorogenic acid fed to growing broiler chicks. Journal of the Science of Food and Agriculture 76, 156-160.
Treviño, J., Rodríguez, M.L., Ortiz, L.T., Rebolé, A. & Alzueta, C. 2000. Protein quality of linseed for growing broiler chicks. Animal Feed Science and Technology 84, 155-166.
Van Barneveld, R.J. 1999. Understanding the nutritional chemistry of lupin (Lupinus spp.) seed to improve livestock production efficiency. Nutrition Research Reviews 12, 203-230.
Van Kempen, G.J.M. & Jansman, A.J.M. 1994. Use of EC produced oil seeds in animal feeds. In Recent Advances in Animal Nutrition. P.C. Garnsworthy & D.J.A. Cole (Eds). Nottingham University Press, Nottingham, 31-56.
Van Laar, H., Tamminga, S., Williams, B.A. & Verstegen, M.W.A. 2000. Fermentation of the endosperm cell walls of monocotyledon and dicotyledon plant species by faecal microbes from pigs. The relationship between cell wall characteristics & fermentability, 2000. Animal Feed Science and Technology 88, 13-30.
Vidal, C., Frias, J., Hernández, A., Martín, P.J., Sierra, I., Rodríguez, C., Blázquez, I. & Vicente, G. 2003. Assessment of nutritional compounds and antinutritional factors in pea (Pisum sativum) seeds. Journal of the Science of Food and Agriculture 83, 298-306.
Villamide, M.J. & San Juan, L.D. 1998. Effect of chemical composition of sunflower seed meal on its true metabolisable energy and amino acid digestibility. Poultry Science 77, 1884-1892.
Viveros, A., Brenes, A., Elices, R., Arija, I. & Canales, R. 2001. Nutritional value of raw and autoclaved Kabuli and desi chickpeas (Cicer arietinum) for growing chickens. British Poultry Science 42, 242-251.
Wasilewko, K. & Buracewska, L. 1999. Chemical composition including content of amino acids, minerals and alkaloids in seeds of three lupin species cultivated in Poland. Journal of Animal and Feed Science 8, 1-12.
Wiseman, J., Al-Mazzoqi, W., Welham, T. & Domoney, C. 2003. The apparent ileal digestibility, determined with young broilers, of amino acids in near-isogenic lines of peas (Pisum sativum L) differing in trypsin inhibitor activity. Journal of the Science of Food and Agriculture 83, 644-655.
Zatari, I.M. & Sell, J.L. 1990. Sunflower meal as a component of fat-supplemented diets for broiler chickens. Poultry Science 69, 1503-1507.


219
The cholesterol lowering effect of lupin meal in chicken diets A. Brenes1, I. Arija2, C. Centeno1 & A. Viveros2 1Instituto del Frío, Departamento Metabolismo y Nutrición. José Antonio Novias, 10. Ciudad Universitaria. 28040 Madrid, Spain 2Departamento de Producción Animal, Facultad de Veterinaria, Ciudad Universitaria, 28040 Madrid, Spain Summary An experiment studied the effect of different concentrations of white lupin (Lupinus albus) seed (0, 20 and 40 %), with and without added cholesterol (1 %), in chicken diets on their performance, relative liver weight, liver fat, intestinal pH and viscosity, and different blood serum parameters. Lupin seed depressed bird performance. There were progressive decreases in liver fat (17 %), cecal pH (6 %), serum glucose (20 %), cholesterol (27 %), total biliary salts (46 %) and total protein (14 %), and increased jejunum viscosity (14 %) with increased amounts of lupin. Cholesterol supplementation had no effect on performance, cecal pH and plasma triglycerides. Relative liver weight (10 %), liver fat (46 %), jejunum viscosity (5 %), cholesterol (60 %), total biliary salts (60 %) and total protein (39 %) were increased, and serum glucose (14 %) was reduced by the addition of cholesterol. These results indicate that inclusion of lupin seed in chicken diets decreased serum cholesterol and affected other related parameters. Cholesterol addition increased different blood serum parameters and liver fat. Keywords: Lupinus albus, cholesterol, chickens Introduction Evidence for a hypocholesterolemic effect of grain legumes on raised cholesterol level in animal models and man has accumulated in recent years (Kingman, 1991). Soy (Glycine max) protein was effective in lowering the level of serum triglycerides and cholesterol in humans and animals (Sirtori et al., 1995). This hypocholesterolemic effect is thought to be at least partially attributable to the amino acid profile of the plant proteins (Carroll, 1991) or to a complex system inducing enhanced expression of LDL receptors (Lovati et al., 1987). On the other hand, in soybean, and in some other legume seeds, the dietary fibre fraction could be an important component, which reduces plasma cholesterol levels (Uberoi et al., 1992). Likewise, soluble and insoluble non-starch polysaccharides (NSP) in the hull and cotyledons of legume seeds have also been reported to be effective in lowering serum cholesterol (Lairon, 1996). White lupin seed protein content is comparable to that of soybean (32 - 45 %).
The carbohydrate chemistry of lupins is different from most other legumes. They contain negligible amounts of starch and high levels (up to 500 g/kg) of soluble and insoluble NSP, and oligosaccharides (van Barneveld, 1999). Lupin kernels contain pectic substances and the major polysaccharide is β-(1-4)-galactan (van Barneveld, 1997). Currently, there is limited information on the physiological effects of lupin seed, particularly on lipid metabolism, compared with other legume seeds. Therefore the purpose of this study was to determine the effect of lupin seed in normal or hypercholesterolaemic birds on different physiological parameters.

220
Materials and methods One hundred and eight, day old male broiler chicks, were housed in an environmentally controlled room. Chicks were allocated to 18 pens, each pen containing six chicks, to receive six diets. There were three replicates. Diets and water were provided ad libitum. The lupin diets (0, 20 and 40 %), with and without cholesterol (1 %) were fed for 3 weeks. Lupins were included in the diets at the expense of soybean and maize (Zea mays). Amino acids levels were balanced in the diets. At the end of the experiment birds were weighed and feed consumption was recorded. Ten birds were randomly selected from each treatment, and blood samples were taken to determine blood glucose, cholesterol, triglycerides, total biliary salts and total serum protein. The liver, jejunum and ceacum were removed for liver fat, viscosity and pH determination, using 8 chicks per treatment.
The data were subjected to analysis of variance using the General Linear Model (SAS Institute, 1989). The data was analysed factorially by ANOVA with 3 lupin concentration × 2 cholesterol concentrations. Results Increased amounts of lupin seed in the diet significantly depressed body weight (12 %), feed efficiency (7 %), liver fat (17 %) and cecal pH (6 %) and increased jejunum viscosity (14 %). Similarly, increased dietary lupin concentration significantly reduced plasma glucose (20 %), cholesterol (27 %), total biliary salts (47 %) and total protein (14 %) compared with control fed chicks. However, relative liver weight and triglycerides were not affected (Table 1).
The addition of 1 % of cholesterol did not affect performance, cecal pH or triglycerides. However, relative liver weight (10 %), liver fat (46 %), jejunum viscosity (5 %), plasma cholesterol (60 %), total biliary salts (60 %) and total protein (39 %) were significantly reduced. Plasma glucose was significantly reduced (14 %). The statistical analysis showed there were significant interactions between lupin concentration and cholesterol content for jejunum viscosity, plasma glucose and triglycerides. Discussion This work shows that raw, low alkaloid, white lupin seed at up to 20 % in diets for broilers reduced growth. Centeno et al. (1990) and Brenes et al. (2002) obtained similar results. Part of the effect may have been due to the high hull fibre content or the level of NSPs in the cotyledons. Gdala et al. (1998) and Brenes et al. (1993, 2002) demonstrated the detrimental effect of fibre by adding hulls to a dehulled lupin seed diet. The white lupin seed meal used here contained up to 316 g NSP/kg. The main monosaccharides were galactose, glucose, arabinose and xylose. There was 80 g oligosaccharides/kg, most of which was in the cotyledons. In birds, degradation of lupin fibre in the small intestine is very limited (Brenes et al., 2003). However, a significant fraction of total NSP is pectin-like substances composed of branched β-(1 to 4>4) galactans, which are quite labile and probably highly susceptible to fermentative breakdown. Considerable amounts of NSP become soluble when ingested by chickens, as shown by increased ileal-soluble NSP levels (Kocher et al., 2000) and the ileal viscosity. The increase in intestinal viscosity and the reduction in cecal pH suggest that the carbohydrates in the lupin seeds are potentially responsible for these effects. The adverse effects of soluble NSPs, especially in poultry, are due to their ability to increase the digesta viscosity (Annison et al., 1996), to modify gastrointestinal tract physiology and change the gut ecosystem (Rubio et al., 1998).

221
Tabl
e 1.
Effe
ct o
f die
tary
whi
te lu
pin
mea
l con
cent
ratio
n w
ith a
nd w
ithou
t cho
lest
erol
on:
bod
y w
eigh
t (BW
), fe
ed c
onsu
mpt
ion
(FC
), fe
edef
ficie
ncy
(FE)
, re
lativ
e liv
er w
eigh
t (R
LW),
liver
fat
(LF
), ce
cal
pH (
pH),
inte
stin
al v
isco
sity
(IV),
and
the
gluc
ose
(G),
chol
este
rol (
Ch)
, tri
glyc
erid
e (T
), to
tal b
iliar
y sa
lts (B
S) a
nd to
tal p
rote
in (T
P) c
onte
nts
in p
lasm
a of
bro
iler
chic
ks a
t 3 w
eeks
of
age
Trea
tmen
t Lu
pin
Ch
BW
FC
FE
R
LW
LF
pH
IV
G
Ch
T B
S TP
1
0 0
624a
826
1.30
c 2.
90bc
5.34
d 6.
25a
2.19
d 25
4a 10
3bc
33b
12bc
2.
66cd
2 20
0
580bc
748
1.37
ab
3.08
abc
6.56
c 6.
25a
2.33
b 24
9a 10
9bc
26c
9bc
2.
87c
3 40
0
543d
761
1.40
a 2.
83c
4.57
e 6.
07a
2.32
bc
199c
74c
15d
7c
2.44
d
4 0
1 62
1a 81
3 1.
33bc
3.
13ab
8.74
a 6.
15a
2.20
cd
220b
182a
23c
22a
3.90
a
5 20
1
607ab
810
1.33
bc
3.17
ab8.
09ab
6.28
a 2.
31bc
d20
7bc
134b
20cd
15
ab
3.87
a
6 40
1
555cd
782
1.41
a 3.
36a
7.09
bc5.
61b
2.68
a 18
3d 13
7b 42
a 11
bc
3.28
b
Pool
ed S
EM
48.0
37
.0
0.02
0.
29
1.22
0.
31
0.12
14
.1
35.6
6.
4 6.
6 0.
28
Lupi
n (L
) 0
62
2a 81
9 1.
31c
3.12
7.
04a
6.20
a 2.
19c
239a
142a
28a
17a
3.28
a
20
594b
779
1.35
b 3.
11
7.32
a 6.
26a
2.32
b 22
8b 12
2ab
23b
12ab
3.
34a
40
549c
771
1.40
a 3.
01
5.83
b 5.
84b
2.50
a 19
1c 10
4b 29
a 9
b 2.
83b
Cho
lest
erol
(Ch)
0 58
2 80
2 1.
35
2.94
b 5.
49b
6.19
2.
28b
236a
95b
25
10b
2.66
b
1 59
5 77
8 1.
35
3.22
a 8.
03a
6.01
2.
50a
203b
152a
28
16a
3.69
a
Sour
ce o
f var
iatio
n
L
0.01
0.
09
0.01
0.
48
0.01
0.
01
0.01
0.
01
0.02
0.
03
0.01
0.
01
Ch
0.20
0.
20
0.93
0.
05
0.01
0.
05
0.01
0.
01
0.01
0.
06
0.01
0.
01
L ×
Ch
1.41
0.
20
0.14
0.
07
0.06
0.
08
0.01
0.
04
0.10
0.
01
0.57
0.
18

222
The results also show that white lupin seed in the diet was beneficial to hypercholesterolemic chicks. It decreased plasma cholesterol, glucose and total biliary salt concentration and reduced liver fat. These results are similar to those of Eder et al. (1996), Chango et al. (1998) and Rubio et al. (2003) with chicks and rats fed white lupin seeds. The results suggest that viscous NSPs can enhance bile secretion and cause a significant loss of these acids in the faeces (Ikegami et al., 1990). The net effect may be an altered intestinal lipid metabolism in resulting in increased hepatic synthesis of bile acids from cholesterol to re-establish the composite pool of these metabolites in the enterohepatic circulation. Soluble fibres also may be useful modulators of post-prandial glycaemic and insulinaemic responses. Diets containing legume seeds may have special advantages, as legume carbohydrates may be digested and absorbed more slowly than those from other foods.
The low concentration of glucose, cholesterol and total biliary salts in plasma, caused by lupin feeding, may have some physiological importance and may be able to be extrapolated to humans. The avian liver is a major site of lipid biosynthesis similar to the human liver. Thus, it provides a useful model for studying lipid metabolism regulation in humans. In conclusion, overall the results suggest that white lupin seed in the diet may be effective in lowering cholesterol absorption as well as the plasma glucose level in chickens. References Annison, G., Hughes, R.J. & Choct, M. 1986. Effects of enzyme supplementation on the
nutritive value of dehulled lupins. British Poultry Science 37, 152-172. Brenes, A., Marquardt, R.R., Guenter, W. & Rotter, B.A. 1993. Effect of enzyme
supplementation on the nutritional value of raw, autoclaved, and dehulled lupins (Lupinus albus) in chicken diets. Poultry Science 72, 2281-2293.
Brenes, A., Marquardt, R.R., Guenter, W. & Viveros, A. 2002. Effect of enzyme addition on the performance and gastrointestinal tract size of chicks fed lupin seed and their fractions. Poultry Science 81, 670-678.
Brenes, A., Slominski, B., Marquardt, R.R., Guenter, W. & Viveros, A. 2003. Effect of enzyme addition on the digestibilities of cell wall polysaccharides and oligosaccharides from whole, dehulled, and ethanol extracted fractions of white lupin in chickens. Poultry Science (In press).
Carroll, K.K. 1991. Review of clinical studies on cholesterol-lowering response to soy protein. Journal of the American Dietetic Association 91, 820-827.
Centeno, C., Yuste, P., Rubio, L.A., Treviño, J. & Brenes, A. 1990. Influence of lupin (Lupinus albus) and flavomycin supplementation in broiler diets. Archivos de Zootecnia 39, 15-24.
Chango, A., Villaume, C., Bau, H.M., Schwertz, A., Nicolas, J.P. & Mejean, L. 1998. Effects of casein, sweet white lupin and sweet yellow lupin diet on cholesterol metabolism. Journal of the Science of Food and Agriculture 76, 303-309.
Eder, K., Roth-Maier, D. & Kirchgessner, M. 1996. The effect of enzyme supplements and high amounts of white lupins on concentrations of lipids in serum and meat in fattening chickens. Archives of Animal Nutrition 49, 221-228.
Gdala, J. 1998. Composition, properties, and nutritive value of dietary fibre of legume seeds. A review. Journal of Animal and Feed Science 7, 131-149.
Ikegami, S., Tshuchishasi, F., Harada, H., Tshuchishasi, N., Nishide, E. & Innami, S. 1990. Effect of viscous indigestible polysaccharides on pancreatic-biliary secretion and digestive organs in rats. Journal of Nutrition 120, 353-360.

223
Kingman, S.M. 1991. The influence of legume seeds on human plasma lipid concentrations. Nutrition Research Reviews 4, 97-123.
Kocher, A., Choct, M., Hughes, R.J. & Broz, J. 2000. Effect of food enzymes on utilisation of lupin carbohydrates by broilers. British Poultry Science 41, 75-82.
Lairon, D. 1996. Dietary fibres: effects on lipid metabolism and mechanisms of action. European Journal of Clinical Nutrition 50, 125-133.
Lovati, M.R., Manzoni, C., Canavesi, A., Sirtori, M., Vaccarino, V., Marchi, M., Gaddi, G. & Sirtori, C.R. 1987. Soybean protein diet increases low density lipoprotein receptor activity in mononuclear cells from hypercholesterolemic patients. Journal of Clinical Investigations 80, 125-130.
Rubio, L.A., Brenes, A. & Centeno, C. 2003. Effects of feeding growing broiler chickens with practical diets containing sweet lupins (Lupinus angustifolius). British Poultry Science 44,391-397.
Rubio, L.A., Brenes, A., Setien, I., Asunción, G. de la, Duran, N. & Cutuli, M.T. 1998. Lactobacilli counts in crop, ileum and caecum of growing broiler chickens fed on practical diets containing whole or dehulled sweet lupin (Lupinus angustifolius) seed meal. British Poultry Science 39, 354-359.
Sirtori, C.R., Lovati, M.R., Manzoni, C., Gianazza, E., Bondioli, A., Staels, B. & Auwers, J. 1995. Reduction of serum cholesterol by soy proteins. Nutrition, Metabolism and Cardiovascular Diseases 8, 334-340.
SAS Institute, 1989. SAS User’ Guide. SAS Institute Inc., Cary, Nc. Uberoi, S.K., Vadhera, S. & Soni, G.L. 1992. Role of dietary fibre from pulses and cereals
as hypocholesterolemic and hypolipidemic agent. Journal of Food Science and Technology 29, 281-283.
Van Barneveld, R.J. 1997. Chemical composition of the ground whole seed kernels and hulls of L. angustifolius cv Gungurru and L. albus cv Kiev mutant. In Understanding the Nutritional Value of Lupins. Pig Research and Development Corporation Final Report DAS 33P. pp. 10-18.
Van Barneveld, R.J. 1999. Understanding the nutritional chemistry of lupin (Lupinus spp) seed to improve livestock production efficiency. Nutrition Research Reviews 12, 203-230.


225
Thermal processing of rapeseed meal: nutritional evaluation in digestibility experiments with pigs R. de Schrijver, S. de Vos & J. Vande Ginste Nutrition Laboratory, Catholic University of Leuven, Kardinaal Mercierlaan 92, B-3001 Leuven, Belgium Summary The nutritional value of heat-processed diets containing low glucosinolate rapeseed meal (Brassica spp) was investigated in growth trials and ileal digestibility experiments carried out with barrows (20 to 100 kg). Starter, grower and finisher diets were either prepared as meal, were pelleting at 80 °C or expanded at 100 °C and then pelleted. During the complete fattening period, the pelleted diet significantly (P < 0.05) improved the growth rate and the ileal digestibility of dry matter (DM), protein, phosphorus and calcium. However the feed:gain ratio and nitrogen (N) retention were not affected by pelleting (NS). When pelleting was preceded by expansion there was no additional improvement in nutrition. Generally, heat-treated diets contained significantly (P < 0.05) more soluble N. However, soluble dietary fibre and available lysine levels were not affected (NS). Keywords: rapeseed meal, pigs, expansion, pelleting Introduction Expansion processing has been introduced in the feed industry as a pre-pelleting conditioning step in order to improve the physical quality of the pellets and to decrease energy expenditure during subsequent pelleting. During the expansion process the feed is subjected to mild heating (90 - 130 °C) and pressure (38 - 40 bar). The question is whether the processing conditions also have nutritional consequences, in particular with regard to digestibility and nutrient availability of nutrients in feedstuffs of lower quality such as rapeseed meal. The use of this feedstuff in pig diets is limited because of its high crude fibre content giving lower protein and amino acid digestibility (Bell, 1984), and the negative effect of glucosinolates on metabolic processes and feed intake (Schöne et al., 1997).
Previous experiments with pigs showed that pelleting highly digestible meal diets containing barley (Hordeum vulgare), wheat (Triticum aestivum), soybean (Glycine max) meal and cassava (Manihot escuelenta), favourably effected crude protein (CP) and DM utilisation. Vande Ginste and de Schrijver (1997) found these effects were not improved by an additional expansion step prior to pelleting. However, it is possible that expansion processing of diets with lower nutritional quality, such as rapeseed meal, may give positive effects on nutrient utilisation.
The objective of this study was to investigate if expansion and pelleting of pig diets, which contained substantial amounts of rapeseed meal, could affect animal performance and ileal digestibility of CP, DM, phosphorus and calcium. Furthermore, in vitro measurements were carried out to determine whether thermal feed processing affected the amounts of soluble protein, available lysine, free amino acids and soluble dietary fibre and whether these effects may be related to animal performance.

226
Materials and methods Growth and ileal digestibility experiments used starting (20 - 40 kg), growing (40 - 70 kg) and finishing barrows (70 - 100 kg). The starter, grower and finisher diets contained 350 g/kg of low glucosinolate rapeseed meal and were balanced for ileal digestible amino acids and net energy. Other major dietary ingredients were wheat, soybean meal, barley and cassava. The diets were prepared either as meal (M), were pelleted (P) or expanded and then pelleted (EP). The maximum temperature during the expanding and pelleting process was 100 °C and 80 °C, respectively.
The growth trial used 72 barrows (Belgian Landrace × Piétrain), which were randomly divided, into 12 pens of 6 animals. Each of the experimental diets (M, P or EP) was fed to 4 pens. During the whole experiment, each pen received starter, grower and finisher diets, which were processed in the same way. Animals had free access to feed and water. Feed intake, body weight gain and the feed conversion ratio were measured.
The ileal digestibility trials used 12 individually housed barrows (Belgian Landrace × Piétrain) that had been surgically fitted with a post-valve T-caecum cannula. The animals were divided in 3 groups of 4 pigs and fed one of the 3 experimental diets (M, P or EP). Feed and water were supplied ad libitum. With each pig, two 5-day digestibility trials were carried out during the starting, growing and finishing phase, respectively. The apparent ileal digestibility of DM, P, Ca and N was measured, and N retention. The acid insoluble ash (AIA) content of feed, faeces and chyme samples was used as an indigestible marker to determine ileal and faecal output. Ileal digesta were collected daily for 3 h. To prevent further enzymatic activity, chyme samples were frozen (-20 °C) every hour. Faeces were collected and weighed twice daily, homogenised and frozen.
At the end of the experiment, chyme and faeces samples were pooled per pig and per digestibility trial and were analysed for AIA, DM, P, Ca and N. Daily total urine volume was recorded and 1 % was kept at -20 °C. At the end of the trial the urine samples were filtered and pooled per pig and per digestibility trial pending N analysis. Feed intake was recorded daily and was corrected for refusal and spillage. Results for apparent digestibility and retention were expressed as a percentage of intake.
Feed samples were analysed through in vitro laboratory tests for available lysine (Carpenter, 1960), N solubility (de Schrijver, 1976), soluble and insoluble fibre (AOAC, 1993) and free amino acid content. The free amino acids were measured by extracting the feed samples for 1 h with 0.1 N HCl containing 2% thiodiglycol. The extracted amino acids were analysed using cation-exchange chromatography combined with a lithium-citrate buffer system.
The results were statistically analysed by analysis of variance using the General Linear Model procedure of SAS (1988). The LSD-test was used to compare differences between treatment means. Probability levels of P < 0.05 were considered significant. Results and discussion Compared to the M diets, feeding the P diets to pigs from 20 to 100 kg significantly (P < 0.05) increased body weight gain and tended to reduce the feed:gain ratio (P < 0.1; Table 1). However, pig performance was not further improved when the feed was expanded prior to pelleting.

227
Table 1. The effect of feed processing on pig performance (20 - 100 kg of body weight)1. Diet M P EP Daily feed intake (kg) 2.12a ± 0.08 2.33a ± 0.14 2.26a ± 0.10 Daily weight gain (kg) 0.71b ± 0.09 0.79a ± 0.08 0,77a ± 0.11 Feed conversion ratio 2.98a ± 0.07 2.95a ± 0.05 2.94a ± 0.07 1Values are means ± standard deviation (n = 24). a,bMeans in a row with a different superscript are statistically different (P < 0.05). M = meal diet; P = pelleted diet; EP = expanded and pelleted diet.
The improved performance of pigs fed the P diets versus the M diets was related to significantly (P < 0.05) higher ileal digestibility of CP, DM, P and Ca (Table 2). There was no difference between the P and EP groups in nutrient digestibility (NS) and N retention was not affected by feed processing (NS).
Table 2. The effect of diet processing on apparent ileal digestibility of nitrogen, dry matter,
phosphorus and calcium, and nitrogen retention in pigs (20 - 100 kg of body weight)1.
Diet M P EP Ileal N-digestibility (%) 68.2b ± 2.7 75.1a ± 2.3 73.5a ± 3.1 N-retention (%) 50.5a ± 2.4 50.9a ± 3.1 49.7a ± 2.6 Ileal DM-digestibility (%) 65.0b ± 2.8 70.5a ± 3.0 71.7a ± 2.5 Ileal P-digestibility (%) 38.2b ± 2.2 43.7a ± 3.4 44.9ab ± 4.4 Ileal Ca-digestibility (%) 50.3b ± 2.9 56.1a ± 3.1 54.8a ± 3.7 1Values are means ± standard deviation (n=4). a,b Means in a row with a different superscript are statistically different (P < 0.05). M = meal; P = pelleted diet; EP = expanded and pelleted diet.
Compared with the M diets, the P and EP diets had higher dietary protein solubility (P < 0.05; Table 3). This suggests the transformation of cell wall structures, caused by shear forces, during expansion and pelleting, or increased protein release due to cell rupture (Marty et al., 1994). Available lysine and soluble dietary fibre levels were generally not affected (NS) by thermal processing, although the soluble fibre content did tend to increase. The combination of expansion and pelleting often gave the highest level of soluble fibre. This is in line with our previous results (de Schrijver & Vande Ginste, 1998). There was no effect of heat processing on either free amino acids or total glucosinolates (< 5 µmol/g DM) in the diet.
In conclusion, expansion and subsequent pelleting of rapeseed meal containing pig diets did not give any supplementary nutritional advantage over pelleting alone. Generally, this work confirms our earlier results from pig experiments using diets containing 28.5 % rapeseed meal (de Schrijver & Vande Ginste, 1998). References AOAC (1993). Methods of Analysis for Nutrition Labelling. Association of Official
Analytical Chemists, Arlington, Va. Bell, J.M. (1984). Nutrients and toxicants in rapeseed meal: a review. Journal of Animal
Science 58, 996-1010. Carpenter, K.J. (1960). The estimation of the available lysine in animal-protein foods.
Biochemical Journal 77, 604-610.

228
Table 3. The effect of diet processing on lysine availability, protein solubility and dietary fibre solubility.
Diet M P EP
Available lysine1 Starter diet 93.3a ± 0.4 92.6a ± 0.2
Grower diet 91.1a ± 0.5 92.0a ± 0.3 92.6a ± 0.4 Finisher diet 90.3a ± 0.3 91.7a ± 0.3 92.0a ± 0.4
Protein solubility2 Starter diet 55.1c ± 0.6 63.8a ± 0.3 64.8a ± 0.7 Grower diet 57.0b ± 0.7 64.1ab ± 0.6 68.1a ± 0.5 Finisher diet 59.4b ± 0.7 66.3a ± 0.8 64.9a ± 0.6
Soluble dietary fibre3 Starter diet 13.2a ± 1.9 14.2a ± 1.6 15.3a ± 2.3 Grower diet 12.9 a ± 2.0 16.4a ± 1.9 17.8a ± 1.8 Finisher diet 11.2a ± 2.6 14.1a ± 2.1 15.5a ± 2.5
1Expressed as percentage of total lysine content (n = 3). 2Expressed as percentage of total protein content (n = 3). 3Expressed as percentage of total dietary fibre (n = 4). a,b,cMeans in a row with a different superscript are statistically different (P < 0.05). M = meal; P = pelleted diet; EP = expanded and pelleted diet.
De Schrijver, R. 1976. Comparative study of analytical methods to predict soybean oil meal
quality. Mededelingen Faculteit Landbouwwetenschappen Gent 41, 1777-1783. De Schrijver, R. & Vande Ginste, J. 1998. Heat processing of rapeseed meal containing
diets: effects on growth performance and ileal nutrient digestibility in pigs. In Recent Advances of Research in Antinutritional Factors in Legume Seeds and Rapeseed. A.J.M. Jansman, G.D. Hill, J. Huisman & A.F.B. van der Poel (Eds). Proceedings 3rd International Workshop on Antinutritional Factors in Legume Seeds and Rapeseed, Wageningen, 8-10 July 1998, 405-408.
Marty, B.J, Chavez, E.R. & Lange, C.F.M. de 1994. Recovery of amino acids at the distal ileum for determining apparent and true ileal amino acid digestibilities in growing pigs fed various heat-processed full-fat soybean products. Journal of Animal Science 72, 2029-2037.
Schöne, F., Groppel, B., Hennig, A. & Jahreis, G. 1997. Rapeseed meals, methimazole, thiocyanate and iodine affect growth and thyroid. Investigations into glucosinolate tolerance in the pig. Journal of the Science of Food and Agriculture 74, 69-80.
Statistical Analysis Systems Institute Inc. 1988. SASSTAT User’s Guide. Release 6.03, Carr, Nc.
Vande Ginste, J. & Schrijver, R. de 1997. Heat processing of pig diets: effects on faecal and ileal nutrient digestibility and in vitro protein characteristics. In Digestive Physiology in Pigs. J.-P Laplace, C. Février, and A. Barbeau (Eds). Proceedings 7th International Symposium on Digestive Physiology in Pigs, INRA. EAAP Publication No. 88, 634-638.

229
Microbial protease addition to a soybean meal diet for weaned piglets: effects on performance, digestion, gut flora and gut function N. Dierick1, J. Decuypere1, K. Molly2 & E. Vanderbeke3 1Faculty of Agricultural and Applied Biological Sciences, Department of Animal Production, Ghent University, Proefhoevestraat, 10, B-9090 Melle, Belgium 2Vitamex N.V., B-9031 Drongen, Belgium 3Aveve N.V., B-2170 Merksem, Belgium Summary Transient hypersensitivity to soy (Glycine max) proteins, resulting in postweaning lag, is well known in pig nutrition. This hypersensitivity precedes the induction of oral tolerance, fully established by the age of 12 weeks. As antigenic soy proteins are not much reduced by heat and show resistance to mammalian proteases, the alleviation of antigenic effects by using a microbial exogenous protease in a soybean meal (SBM) based weaner diet, was studied in a combined growth trial and slaughter experiment (1 h, 2 h, 3 h post-prandial). There was a non-significant trend for piglets to perform better (growth, F:G ratio) on the protease containing diets. Also on the protease diets, ileal digestibility, and to a lesser extent faecal digestibility, of dry matter (DM), organic matter (OM), crude protein (CP), HCl fat and energy were increased, although again not significantly. Electrophoretic profiles of SDS-PAGE and ELISA and immunoblotting tests showed less residual antigenicity against the soy protein in the stomach and foregut content of piglets on the protease diets. Also the IgG titers to soy proteins in the sera of protease fed piglets were lower. Furthermore, protease addition significantly reduced total anaerobic flora in the stomach and the foregut in a dose/time dependent manner. The number of intra-epithelial lymphocytes (IELs) and the crypt depth was reduced giving a higher villus height:crypt depth (V:C) ratio. Both phenomena (V:C ratio; IELs) indicate a healthier and better functioning mucosa when protease was used. For nearly all parameters studied, the effects were more pronounced at 2 h post-prandial at the lower enzyme dose. There is little published data on the potential use of proteases in animal diets. Our data shows that the addition of protease may be a potential way of increasing the amounts of SBM in piglet starter diets. Keywords: soybean meal, proteases, antibodies Introduction The recent European ban on the use of antibiotic growth promoters and the ban on the use of animal protein in pig nutrition may result in decreased animal performance. Moreover, there is evidence that > 80 % of piglet deaths in the post-weaning month seem to involve digestive disorders either alone or associated with other disorders. Plant globulins are less digestible than animal proteins resulting in poor performance when they are fed to early-weaned piglets (Dréau et al., 1995). Vegetable protein sources also increase fibre, phytate, complex storage proteins and antinutritional proteins in the diet. To date most work on the mode of action and effect of legume proteins acting as antigens/allergens in farm animals has been carried out in calves (Dréau & Lallès, 1999).
Addition of exogenous enzymes could help alleviate this disadvantage and maintain economic production. For preruminant calf feeding, the hydrolysis of soy proteins by acid or

230
proteolytic treatments, gives products with low antigen levels based on ELISA tests. However, protein digestibility was still not as high as in milk protein (0.82 versus 0.95). The few trials conducted to date on the effects of protease (all or not in combination with NSPase) supplementation to plant based diets or protease pre-treatment of plant proteins for pigs have given variable result (Thorpe & Beal, 2001). Possible explanations may be: genotype, age, health status and housing of the pigs, the target substrate is not well defined, enzyme type and activity, inclusion level, method of application (addition versus pre-treatment), different methods of determining enzyme activity, diet formulations exceeding nutrient recommendations and the presence or absence of antimicrobial growth promoters in the growth trials. Nevertheless the greatest effect has been observed in early-weaned piglets in the 7 d immediately post-weaning, helping to overcome the transient deleterious effect of feeding SBM.
In this study, the addition of a protease to soybean meal diets as a potential alternative to increasing the amount of SBM in weaned piglet starter diets was investigated. The effect of feeding protease at two dose levels in SBM based diets on the performance, faecal score, survival of globulins in the stomach and small intestine using ELISA and immunoblotting methods, the nutrient digestibility, gut flora, small intestine morphology and histology and on serum specific-specific antibodies in piglets was investigated in a slaughter experiment. The question is whether a change in the antigen content, due to exogenous enzymic action, has practical meaning in the feeding of weaned pigs. To date the suitability of simple addition of proteases to SBM diets in pig nutrition for upgrading the (legume) protein fraction has not been investigated. Materials and methods Thirty-six newly weaned piglets (21 d of age; 6.5 kg) were divided according to litter, sex and weight in 3 homogenous groups of 12 piglets and fed either a SBM diet (D1) or were supplemented with either 0.05 % (D2) or 0.1 % protease (D3) (Biofeed Pro, Novozymes, DK).
The feeds contained 11.2 g/kg apparent ileal digestible lysine, 7.1 g/kg apparent ileal digestible methionine + cystine, 7.3 g/kg apparent ileal digestible threonine, 2.2 g/kg apparent ileal digestible tryptophan and had a net energy content (NE) of 9.45 MJ/kg. The diets were free of antibiotic, copper sulphate and zinc oxide beyond animal requirements and were offered dry and ad libitum. Water was continuously available via nipples.
Piglets were weighted individually on days 0 and at slaughter. Feed intake was recorded per pen for the total period. On days 10, 11 and 12 postweaning, 4 piglets of each experimental group, were euthanized at 1 h, 2 h and 3 h post-prandial for sampling of blood and digesta. It has been reported that changes in gut flora and in villus structure peak at 7 - 10 d post weaning. The gastro-intestinal tract (GIT) was immediately removed and dissected. Content samples were taken from the stomach, the upper small intestine (first 3 m) and the lower small intestine (last 3 m), for bacteriological and chemical analyses. Gut tissues for histomorphometry were sampled and processed at 2 sites: 3 m distal from the pylorus and 3 m proximal to the caecum, using standard procedures. For digestibility determination Celite 545 was added to the diets as an inert marker. The Ghent University Animal Ethics Committee approved the experimental protocol.
Feeds and digesta (stomach and small intestinal content, faeces) were analysed for macronutrients using EU standard methods except for total dietary fibre (TDF), which was determined by the Prosky method. The bacteriological counts were on fresh serially diluted ten-fold samples. The medium and culture conditions used were Reinforced Clostridial agar + 0.001 % hemin (48 h, 37 °C, anaerobically, total colony count).

231
Electrophoretic separations (Laemmli method) of the proteins from feed and digesta were carried out under reducing conditions. Immunoblotting was carried out using pig antisoy antiserum. Pig polyclonal antibodies were prepared as follows: 4 pigs (age 11 weeks; fed a diet completely free of legume proteins) received an inter muscular injection of soy proteins (2 ml), prepared from raw untoasted soybeans and mixed in Freund's complete adjuvant. Two months after the initial injection a booster protein injection was given as before but using Freund's incomplete adjuvant. Pigs were bled 10 d after the booster injections and serum was prepared.
Assessment of polyclonal antibodies in the serum was by non-competitive indirect inhibition ELISA, using rabbit anti-pig immunoglobulin IgG HRP conjugate. For analysis of total soy antigenicity in feed and digesta, pig antisoy antiserum was used. Results and discussion The piglets on the protease containing diets showed a 5 to 8 % improvement in growth rate and a better feed conversion ratio (NS). Feed intake was the same among diets. The apparent small intestinal and overall digestibilities of the macronutrients are shown in Table 1, which, shows that protease addition improved digestion. Table 1. Small intestinal and overall digestibility of nutrients and energy (%, mean ± SD) Diet DM OM CP HCl fat TDF Energy Small intestine D1 45.4 ± 0.8 48.7 ± 1.4 56.2 ± 2.7 47.8 ± 6.8 13.8 ± 11.5 51.6 ± 1.1D2 50.1 ± 8.4 52.1 ± 6.9 63.4 ± 8.0 57.7 ± 8.7 21.8 ± 14.6 58.6 ± 6.7D3 47.6 ± 2.5 51.3 ± 2.4 59.6 ± 3.9 57.8 ± 1.7 18.3 ± 11.4 55.3 ± 2.4Overall D1 73.4 ± 2.3 76.3 ± 2.2 69.5 ± 3.8 30.5 ± 5.3 88.5 ± 0.5 72.7 ± 2.8 D2 74.6 ± 2.0 77.2 ± 1.9 72.4 ± 2.1 44.6 ± 3.9 89.0 ± 0.8 74.2 ± 1.5 D3 74.1 77.2 72.3 47.3 87.4 74.8 Ileal samples: pooled per diet (3 periods with 4 pigs each). No significant differences. Faecal samples: 2 mixed per diet (pens), except for D3 (n=1)
Table 2 gives the effect of protease on gut flora and morphology. The total bacterial load in the gut was significantly lowered by the protease, in a dose/time dependent manner. Some evidence suggests that proteases reduce bacterial attachment in the GIT. Further in vitro studies show that the inhibitory factors (antimicrobial biopeptides?) might not only originate from SBM, but also from whey powder (accelerated lactoferricin production via exogenous protease). The V:C ratio was improved and the number of IELs, which are believed to be the first line of defence against dietary and pathogenic insult, were significantly lowered. This indicates a more healthy and better functioning mucosal status. There was a significant positive relationship between animal growth and villus height (r = +0.63**). There was also a significant negative relationship piglet growth the IEL number (r = -0.45*). The IgG levels (Fig. 1) against soybean proteins were lower in the serum of piglets fed the proteases in agreement with the lower levels of residual antigenicity (Fig. 2) in the gut content of piglets on the protease diets and with the altered SDS-PAGE and immunoblotting profiles of the immunogenic soy proteins (data not shown). Our results suggest that protease addition may be a potential way of increasing the amount of SBM in weaner piglet starter diets without lowering animal performances and gut health.
232
Table 2. Effect of diet and post-prandial time (PPT) on total anaerobic count, gut morphology and the number of intra-epithelial lymphocytes in the proximal small intestine
PPT Diet St1 SI11 SI2
1 V2 C2 V:C2 IELs3 1 h D1 7.51a 6.70a 7.54a 408a 294a 1.39a 43a D2 6.71b 6.22b 7.42a 405a 261b 1.55b 38b D3 6.04c 5.59c 7.39a 436b 274b 1.59b 45a 2 h D1 7.53a 7.05a 7.78a 432a 300a 1.44a 41a D2 6.73b 6.09b 7.77a 473b 326b 1.45a 33b D3 6.47b 5.84b 7.83a 413a 278a 1.49b 41a 3 h D1 7.20a 6.36ab 7.77a 451a 310a 1.45a 39a D2 7.23a 6.65a 7.39a 423b 280b 1.51b 36a D3 7.12a 6.09b 7.85a 410b 304a 1.35c 39a 1 – 3 h D1 7.41a 6.70a 7.70a 429a 301a 1.43a 41a D2 6.89b 6.32ab 7.52a 428a 287b 1.49b 35b D3 6.54b 5.84b 7.69a 422a 284b 1.49b 42a
1Counts (CFU, log10/g) in stomach (St), proximal (SI1) and distal (SI2) small intestine, respectively. 2V = villus height; C = crypt depth; V:C = villus height:crypt depth ratio (in µ). 3IELs = IELs/100 enterocytes; a,b,c: P<0.05 (per post-prandial period)
Figure 1. Serum antisoy IgG levels Figure 2. Residual stomach antigenicity. Acknowledgements The Belgian Ministry of Agriculture (R & D Direction) and N.V. Aveve (B), N.V. Kemin Europa (B) and N.V. Vitamex (B) supported this research financially. The authors thank Dr C. van Nevel for his help in morphological and histochemical measurements. References Dréau, D. & Lallès, J. 1999. Contribution to the study of gut hypersensitivity reactions to
soybean proteins in preruminant calves and early-weaned piglets. Livestock Production Science 60, 209-218.
0.150
0.200
0.250
0.300
0.350
D11h
D12h
D13h
D21h
D2 2h D2
3h D3 1h D3
2h D33h
Diet and post-prandial time
Absorption (450 nm)
0 0.1 0.2 0.3 0.4 0.5 0.6
D1 1h D1
2h D1 3h D2
1hD2 2h D2
3h D3 1h D3
2hD33h
Diet and post-prandial time
Absorption (450 nm)

233
Dréau, D., Lallès, J., Toullec, R. & Salmon, H. 1995. B and T lymphocytes are enhanced in the gut of piglets fed heat-treated soybean proteins. Veterinary Immunology and Immunopathology 60, 69-79.
Thorpe, J. & Beal, J. 2001. Vegetable protein meals and the effects of enzymes. In Enzymes in Farm Animal Nutrition. M. Bedford & G. Partridge (Eds). CABI, Wallingford. pp. 125-143.


235
Rate of muscle protein synthesis in rats fed raw and extruded faba bean diets F. Marzo1, E. Urdaneta1, J. Barrenetxe1, F. Ibañez1 & S. Santidrian2 1Animal Physiology & Nutrition Laboratory, Universidad Pública de Navarra, 31006 Pamplona Spain 2Department of Human Physiology, Universidad de Navarra, 31008 Pamplona, Spain Summary A feeding experiment was conducted with male Wistar rats to determine the effects of extruded faba bean (Vicia faba L.) on gastrocnemius (GM) muscle protein synthesis rate and DNA and RNA content. Extrusion abolished proteolytic enzyme inhibitor activity. Processing also significantly (P < 0.01) reduced the tannin and phytate content. Over 15 d, animals were fed, ad libitum, with diets containing either casein (Control, C), raw (RFB) or extruded faba bean (EFB) as the sole protein source (11 %) supplemented with methionine (0.25 %). The energy content was 15.6 MJ/kg diet. There was a significant (P < 0.01) reduction in weight gain and increase in food intake in rats fed RFB compared with control animals. Extrusion significantly improved both weight gain and food intake. There was no difference in muscle weight, protein, RNA and DNA content between rats fed control and extruded faba bean diets. However, GM weight and total RNA content were significantly increased (P < 0.01) in EFB group compared with the RFB group. Protein synthesis was significantly (P < 0.01) decreased in rats fed RFB compared to those fed control and EFB diets. The possible implications of these findings are discussed. Keywords: extrusion cooking, Vicia faba, gastrocnemius muscle, rat Introduction Faba beans (Vicia faba L.) are commonly used as a protein source for different kind of animals (Messina, 1999). Many legumes, when raw, contain antinutritional factors (ANFs), such as proteolytic enzyme inhibitors, tannins and phytates. They are also generally deficient in sulphur amino acids. The ANFs have a negative effect on protein digestibility and metabolism in monogastrics, including humans (Marzo et al., 1998; Grant, 1999b). Processing methods like extrusion have been developed to inactivate these ANFs and have been used on raw legume seed (Harper, 1995; Alonso et al., 2000a). Sulphur amino acid deficiency can be corrected by supplementing the legume protein with methionine.
The aim of this work was to report on the protein synthesis rate of skeletal muscle, protein turnover and nucleic acid composition in rats fed an extruded faba bean diet. Materials and methods Male Wistar rats weighing about 80 - 90 g were distributed into 3 dietary groups of 6 animals each and housed in individual metabolism cages. Over 15 d, animals were fed ad libitum diets, which contained casein (Control, C), raw or extruded faba bean (RFB and EFB groups, respectively) as the sole protein source. Methionine (0.25 %) was added to the three diets, which contained 11.0 % protein, and 15.6 MJ/kg diet. The faba beans were ground to 0.5

236
mm. Extrusion cooking used a Clextral X-5 model BC 45 twin-screw extruder (F-42100 Firminy, France).
The rats were housed at controlled temperature (22 ± 1 °C), relative humidity (50 ± 5 %), ventilation (at least 15 complete changes of air/h), and with an artificial light-dark cycle (12 h light/dark period). Body weight change and food intake were recorded daily. At the end of the experiment the rats were killed and the gastrocnemius (GM) muscle from the hind legs were excised and weighed. Protein synthesis was measured in the GM administering a flooding dose of [3H] phenylalanine (Garlick, 1980). Muscle protein, DNA and RNA were also measured (Giles & Myer, 1965; Munro & Fleck, 1966; Bradford, 1976).
One-way analysis of variance (ANOVA), followed by Fisher's least significant difference (LSD) test, was used to determine differences among treatments. Differences were considered significant with a P value of 1 %. Comparisons were performed using the StatView/Apple Macintosh version 4.01 non-FPU (Abacus Concepts, 1992-1993) statistical package. Results and discussion Raw faba bean contained significant amounts of trypsin (1.7 ± 0.2 g/kg) and chymotrypsin (0.6 ± 0.1 g/kg) inhibitor and lectin (32 x 104 HU/kg) but no detectable α-amylase inhibitor. The protease inhibitors and the lectin were completely inactivated or eliminated by extrusion cooking. Extrusion processing also significantly reduced (P < 0.01) seed tannin and phytate content from 2.6 ± 0.2 to 1.0 ± 0.1 g/kg and 1.9 ± 0.2 to 1.5 ± 0.1 g/kg respectively.
The experimental results are summarised in Table 1. There was a significant (P < 0.01) reduction in weight gain and an increase in food intake in RFB rats compared with the control group. The results of Wang & McIntosh (1996) support these findings. They showed that extrusion significantly improved both body weight gain and food intake. There was no difference in muscle weight, protein, RNA and DNA content between rats fed the control and the EFB diets. However, GM weight and total RNA content significantly increased (P < 0.01) in EFB rats compared with RFB animals. Protein synthesis rate (Ks) was significantly (P < 0.01) decreased in rats fed RFB compared with those fed the control and EFB diets. Raw faba bean protein was poorly utilised by rats. This appeared to be due to a combination of factors including amino acid deficiency, the presence of ANFs and possibly, the refractory nature of the reserve proteins and other seed constituents such as starch (Grant et al., 1995). Heat-treatment of legumes inactivates or eliminates the major ANFs (lectins and enzyme inhibitors) from the seed and greatly increases protein and starch digestibility (Alonso et al., 2000b). Table 1. Weight gain, food intake, gastrocnemius muscle weight, protein content, total DNA
and RNA, and protein synthesis rate (ks) of male growing rats fed diets containing raw (RFB) and extruded Vicia faba (EFB) as the sole protein source over 15 d.
Diet C RFB EFB Weight gain (g) 126 ± 6a 101 ± 3b 116 ± 4a Food intake (g/100 g BW) 138 ± 3b 152 ± 3a 143 ± 2b Gastrocnemius (mg/100 g BW) 518 ± 18a 504 ± 14b 512 ± 12a Protein content (mg) 176 ± 8.2a 171 ± 9.2a 174 ± 7.5a DNA (mg) 0.98 ± 0.06a 0.90 ± 0.06a 0.96 ± 0.04a RNA (mg) 1.33 ± 0.12a 0.94 ± 0.09b 1.29 ± 0.10a ks ( %/d) 13.2 ± 0.3a 10.8 ± 0.3b 11.8 ± 0.4a Ks, protein synthesis rate is the percent of newly synthesized protein/d. Values are mean ± SEM of 6animals/group. Means with different superscripts are significantly different (P≤ 0.01).

237
In conclusion, extrusion processing of faba bean seed improved muscle protein synthesis rate and growth in legume-fed rats. The efficiency of this thermal technology in inactivating proteolitic enzyme inhibitors and others ANFs may explain the observed improvements. Acknowledgments This work was supported by Spanish DGICYT 97-0675.
References Alonso, R., Aguirre, A. & Marzo, F. 2000a. Effects of extrusion and traditional processing
methods on antinutrients and in vitro digestibility of protein and starch in faba bean and kidney beans. Food Chemistry 68, 159-165.
Alonso, R., Grant, G., Dewey, P. & Marzo, F. 2000b. Nutritional assessment in vitro and in vivo of raw and extruded peas (Pisum sativum L.). Journal of Agricultural and Food Chemistry 48, 2286-2290.
Bradford, M.M. 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Annals of Biochemistry 72, 248-254.
Garlick, P.J., McNurlan, M.A. & Preedy, V.R. 1980. A rapid and convenient technique for measuring the rate of protein synthesis in tissues by injection of [3H]phenylalanine. Biochemistry Journal 192, 719-723.
Giles, K.W. & Myer, A. 1965. An improved diphenilamine method for the estimation of deoxiribonucleic acid. Nature 206, 93.
Grant, G. 1999a. Protein protease inhibitors from plants. In Secondary Plant Products: Antinutritional and Beneficial Actions in Animal Feeding. J.C. Caygill & I. Mueller-Harvey (Eds). Nottingham University Press, Nottingham. pp. 71-86.
Grant, G. 1999b. Plant lectins. In Secondary Plant Products: Antinutritional and Beneficial Actions in Animal Feeding. J.C. Caygill & I. Mueller-Harvey (Eds). Nottingham University Press, Nottingham. pp. 87-110.
Grant, G., Dorward, P.M., Buchan, W.C., Armour, J.C. & Pusztai, A. 1995. Consumption of diets containing raw soyabean (Glycine max) or kidney bean (Phaseolus vulgaris), cowpeas (Vigna unguiculata) or lupin seed (Lupinus angustifolius) by rats for up to 700 days. Effects upon growth, body composition and organ weights. British Journal of Nutrition 73, 17-29.
Harper, J.M. 1995. Extrusion technology: Current status and future potential. South African Journal of Food Science and Nutrition 7, 135-141.
Marzo, F., Alonso, R., Aguirre, A. & Santidrian, S. 1998. The effects of extruded kidney beans (EKB) (Phaseolus vulgaris L. var. Pinto) on the growth, organ weight and hormonal levels of the rat. FASEB Journal 4836, A835.
Messina, M.J. 1999. Legumes and soybeans: overview of their nutritional profiles and health effects. American Journal of Clinical Nutrition 70, 4398-4508.
Munro, H,N. & Fleck, A. 1966. Recent developments in the measurements of the nucleic acids in biological materials. Analyst 91, 78-88.
Wang, Y.H.A. & McIntosh, GH. 1996. Extrusion and boiling improve rat body weight gain and plasma cholesterol lowering ability of peas and chickpeas. Journal of Nutrition 126, 3054-3062.


239
Effect of carob (Ceratonia siliqua L.) seed in broiler chick diets on nutrient digestibility and intestinal viscosity L.T. Ortiz1, M.L. Rodríguez1, C. Alzueta1, A. Rebolé1, C. Centeno2 & J. Treviño1 1Departamento de Producción Animal, Facultad de Veterinaria, Universidad Complutense, Ciudad Universitaria, 28040 Madrid, Spain 2Instituto del Frío, CSIC, Ciudad Universitaria, 28040 Madrid, Spain Summary Inclusion of carob Ceratonia siliqua L seed at 60 (CS-60 diet) and 90 g/kg (CS-90 diet) in a reference diet markedly (P < 0.01) depressed nutrient utilisation in broiler chicks. The reduction in apparent digestibility of crude protein (CP) and crude fat was 12.7 and 20.9 % for the CS-60 diet, and 29.5 and 23.8 % for the CS-90 diet, respectively. The apparent metabolisable energy value followed a similar trend to crude fat digestibility. The viscosity of the jejunal digesta was drastically increased by inclusion of carob seed in the diet, and this might explain most of the results observed in this study. Keywords: Ceratonia siliqua, digestibility, intestinal viscosity, chicks Introduction Carob bean is the seed of Ceratonia siliqua a perennial legume tree native to the Mediterranean basin and southwestern Asia. The seed contain a very high level of gum (42 – 46 % by weight; Saura-Calixto, 1987), which is basically a neutral galactomannan polymer.
Numerous studies have shown that soluble polysaccharides, such as β-glucans and arabinoxylans, can depress the digestion and absorption of nutrients in monogastrics (Campbell & Bedford, 1992; Choct et al., 1996). On the other hand, it is also known that certain types of soluble fibre can have beneficial effects on human and animal health, such as reducing the risk of coronary heart disease by lowering blood pressure and serum cholesterol (Williams et al., 1995; Beagger et al., 1996).
The aim of this study was to evaluate the effect of inclusion of carob seed into the diet on apparent protein and fat digestibility, apparent metabolisable energy (AME) and intestinal viscosity in broiler chicks. Materials and methods The carob seed used in this study was obtained commercially from Valencia, Spain. Seed was ground and analysed as described below.
Five-day-old male broiler chicks (Cobb) were randomly distributed into 3 experimental groups of 10 chicks each. Chicks were housed in wire metal cages, 2 birds/cage, and fed for 16 d (5 to 21 d of age) on a reference diet (RD) or one of two test diets containing carob seed at 60 (CS-60) and 90 (CS-90) g/kg (Table 1). Titanium dioxide was added to the diets as an indigestible marker. All diets were formulated to meet National Research Council (1994) requirements for broiler chicks.
Excreta samples were collected from each cage during the last three days of the experiment and used to estimate crude fat digestibility. On day 22 all birds were slaughtered

240
and the jejunum and ileum from each bird was dissected out. Their contents were flushed into plastic containers. Jejunal content was used for viscosity measurements, and ileal content for pH measurement, dry matter, nitrogen and titanium analyses to estimate CP digestibility. The analyses were performed on pooled samples from the two birds in the same cage. Table 1. Composition of experimental diets (g/kg, as fed basis). Experimental diets Ingredients RD CS-60 CS-90 Maize 479 397 356 Soybean meal 410 404 400 Sunflower oil 66 94 109 Carob seed 0 60 90 Methionine 2 2 2 Titanium dioxide 5 5 5 Basal mixture1 38 38 38 1Basal mixture provided, as g/kg: sodium chloride, 3.0; calcium carbonate, 10.0; dicalcium phosphate, 19.0; BHT, 1.0; vitamin and mineral premix, 5.0. For composition of vitamin/mineral premix see (Alzueta et al., 2003).
Dry matter (DM), CP, soluble and insoluble dietary fibre and ash were analysed by standard AOAC methods (AOAC, 1995). Crude fat was determined using petroleum ether following acidification of the samples with 4N HCl (Wiseman et al., 1992). Neutral and acid detergent fibre and lignin was determined as described by Robertson & van Soest (1981). Gross energy was measured in an adiabatic bomb calorimeter. Titanium was determined colourimetrically (Short et al., 1996). While total phenol and condensed tannin content were determined by the Folin-Denis (Burns, 1963) and vanillin-hydrochloric acid (Broadhurst & Jones, 1978) methods respectively. Jejunal digesta viscosity and ileal digesta pH were measured with a digital viscometer and a micro pH-electrode, respectively. Results were analysed by analysis of variance using the Statgraphics software package (Version 5.0, Statistical Graphics, Rockville, Md). Results and discussion The carob seed composition is shown in Table 2. The seed contained 681 g/kg DM of dietary fibre, approximately half of which was soluble fibre. Table 2. Analysed composition of carob seed (g/kg DM). Crude protein 183.0 Neutral detergent fibre 300.1 Crude fat 18.8 Acid detergent fibre 123.3 Ash 45.1 Lignin 9.8 Total phenolics1 32.1 Soluble fibre 346.8 Condensed tannins2 8.0 Insoluble fibre 334.4 1Expressed as tannic acid equivalents (g/kg DM). 2Expressed as catechin equivalents (g/kg DM).
The results of this experiment (Table 3) show that inclusion of carob seed in the diet depressed nutrient digestion (P < 0.01). The effect was more marked as the inclusion rate increased from 60 to 90 g/kg of the ration.

241
Table 3. Effect of dietary inclusion of carob seed in the diet on the apparent digestibility of crude protein and crude fat, AME content (MJ/kg DM), jejunal digesta viscosity (cP) and of ileal digesta pH measured in 22-d-old broiler chicks.
Carob inclusion (g/kg) 0 60 90
Pooled SEM
ANOVA P-value
Crude protein 0.800a 0.698b 0.564c 0.026 < 0.01 Crude fat 0.870a 0.688b 0.663b 0.011 < 0.01 AME 14.22a 11.49b 11.41b 0.111 < 0.01 Viscosity 1.2a 115.2b nm 7.00 < 0.01 pH 6.8a 6.4ab 5.7b 0.27 < 0.05 nm = not measurable. a,b,c Means within a row with a different superscript are significantly different (P < 0.05).
The apparent digestibility coefficients of CP in the CS-60 and CS-90 diets were reduced
by 12.7 and 29.5 %, respectively, compared with the reference diet. For crude fat, the corresponding digestibility coefficients for the carob diets decreased by 20.9 and 23.8 %, respectively. Carob seed in the diet had a negative effect on the AME content and values decreased in a similar way to crude fat digestibility. These findings agree with the results of Vohra & Kratzer (1964), who found chicken growth was inhibited by about 26 % by the dietary inclusion of carob gum at 20 g/kg. Harmuth-Hoene & Schwerdtfeger (1979) also found a marked reduction of protein digestibility when rats were fed on a diet with 100 g/kg of carob gum.
Feeding chicks diets containing carob seed drastically increased digesta viscosity and caused sticky droppings. In chicks on the CS-90 diet, the jejunal content was gel-like. The viscosity could not be measured because no supernatant was obtained after centrifugation. Viscosity is considered to be the mechanism by which soluble fibre can depress nutrient digestion by limiting the diffusion of digestive enzymes and nutrients and by stimulating microbial activity in the small intestine resulting in greater competition with the host animal for nutrients (Bedford, 1995; Choct et al., 1996). Considering this hypothesis, it is possible that the lower ileal pH found in the carob seed-fed chicks than in chicks on the reference diet might indicate microbial degradation of carob gum, yielding lactic acid and short chain fatty acids, since this neutral galactomannan polymer is highly susceptible to fermentation breakdown (Bravo, 1999). In addition to the effect of intestinal viscosity, the presence of tannins in carob seed was probably another factor that contributed to depressed nutrient digestion, particularly protein. It is well known that tannins, specially condensed tannins, can inhibit the activity of some digestive enzymes and form tannin-protein complexes, which are resistant to enzymic hydrolysis (Marquardt, 1989). This may be the reason why the decreased protein digestibility values were not in line with those for fat digestibility and AME content.
In conclusion, the results of this study show that inclusion of carob seed in chick diets impaired their nutritive value and was associated with a drastic increase in intestinal viscosity. The reduced ileal pH found is possibly a reflection of microbial activity in the small intestine. This aspect of our results requires further research employing microbial methods. References Alzueta, C., Rodríguez, M.L., Cutuli, M.T., Rebolé, A., Ortiz, L.T., Centeno, C. & Treviño,
J. 2003. Effect of whole and demucilaged linseed in broiler chicken diets on digesta viscosity, nutrient utilisation and intestinal microflora. British Poultry Science 44, 67-74.

242
Association of Official Analytical Chemists International, 1995. Official Methods of Analysis. AOAC, Arlington, Va.
Beagger, M., Andersen, O., Nielsen, J.D. & Rytting, K.L. 1996. Dietary fibres reduce blood pressure, serum total cholesterol and platelet aggregation in rats. British Journal of Nutrition 75, 483-493.
Bedford, M.R. 1995. Mechanism of action and potential environmental benefits from the use of feed enzymes. Animal Feed Science and Technology 53, 145-155.
Bravo, L. 1999. Propiedades y aplicaciones de la fibra de la algarroba (Prosopis pallida L.). Alimentaria 3, 67-73.
Broadhurst, R.B. & Jones, W.T. 1978. Analysis of condensed tannins using acidified vanillin. Journal of the Science of Food and Agriculture 29, 788-794.
Burns, R.E., 1963. Methods of tannin analysis for forage crop evaluation. Georgia Agricultural Experiment Station Technical Bulletin No. 32.
Campbell, G.L. & Bedford, M.R. 1992. Enzyme applications for monogastric feeds: a review. Canadian Journal of Animal Science 72, 449-456.
Choct, M., Hughes, R.J., Wang, J., Bedford, M.R., Morgan, A.J. & Annison, G. 1996. Increased small intestinal fermentation is partly responsible for the antinutritive activity of non-starch polysaccharides in chickens. British Poultry Science 37, 609-621.
Harmuth-Hoene, A. & Schwerdtfeger, E. 1979. Effect of indigestible polysaccharides on protein digestible and nitrogen retention in growing rats. Nutrition and Metabolism 23, 399-407.
Marquardt, R.R., 1989. Dietary effects of tannins, vicine and convicine. In Recent Advances of Research in Antinutritional Factors in Legume Seeds. J. Huisman, T.F.B. van der Poel & I.E. Liener (Eds). Proceedings 1st International Workshop on Antinutritional Factors in Legume Seeds, Wageningen, 23-25 November 1988, 141-155.
National Research Council, 1994. Nutrient Requirements of Poultry, 9th Edition. Academic Press, Washington, DC.
Robertson, J.B. & Soest, P.J. van, 1981. The detergent system of analysis and its application to human food. In The Analysis of Dietary Fibre in Foods. W.P.T. James & O. Theander (Eds). Marcel Dekker, New York. pp. 123-158.
Saura-Calixto, F. 1987. Determinación de la composición química de algarroba (Ceratonia siliqua). Azúcares, taninos, pectinas y aminoácidos. Anales de Bromatología XXXIX, 81-93.
Short, F.J., Gorton, P., Wiseman, J. & Boorman, K.N. 1996. Determination of titanium dioxide added as inert marker in chicken digestibility studies. Animal Feed Science and Technology 59, 215-221.
Vohra, P. & Kratzer, F.M. 1964. Growth inhibitory effect of certain polysaccharides for chickens. Poultry Science 43, 1164-1169.
Williams, C.L., Bollella, M., Spack, A. & Puder, D. 1995. Soluble fiber enhances the hypocholesterolemic effect and the step I diet in childhood. Journal of the American College of Nutrition 14, 251-257.
Wiseman, J., Edmunds, B.K. & Shepperson, N. 1992. The apparent metabolisable energy of sunflower oil and sunflower acid oil for broiler chickens. Animal Feed Science and Technology 36, 41-51.

243
In vivo and in vitro ileal digestibility of protein and amino acids of peas containing different tannin levels E. Święch, L. Buraczewska & M. Taciak The Kielanowski Institute of Animal Physiology and Nutrition, Polish Academy of Sciences, Department of Monogastric Nutrition, PL 05-110 Jabłonna, Poland Summary The standardized, true ileal digestibility of protein and amino acids was determined in cannulated growing pigs and in adult rats fed diets which including pea (Pisum sativum) cvs Agra and Grapis which respectively contained 0.4 and 5.2 g tannin/kg of seed dry matter (DM). In vitro digestibility was determined in two low-tannin and four high-tannin pea cultivars by the two-step pepsin/pancreatin method of Boisen & Fernández (1995). The standardised ileal protein and amino acids digestibility was significantly higher in pigs fed the low-tannin pea cv. Agra than with the high-tannin cv. Grapis. True amino acid ileal digestibility in rats was also higher in animals fed the low-tannin cv Agra. However, values were only significantly different for methionine, cystine, isoleucine, histidine, phenylalanine and valine. Standardized ileal amino acids digestibility was determined in vitro in six pea cultivars. It decreased linearly with increased tannin content. In conclusion, the in vitro method seems to be a useful way to rank peas, according to their amino acid digestibility, for pigs. Keywords: Pisum sativum, tannins, digestibility, amino acids, pigs, rats, in vitro Introduction Compared with other grain legumes, peas have a relatively low antinutritional factor (ANF) content. They have a high protein and energy content and are increasingly used as a feed ingredient for pigs (Castell et al., 1996). Their lysine content is relatively high, but the level of sulphur-amino acids is relatively low.
Two sub-species of peas are cultivated in Poland, P. sativum arvense, which has coloured flowers and seed and is mainly used in animal feeds and P. sativum hortense, which is white-flowered and is used in human nutrition and for monogastric animals, especially pigs. Coloured-flowered pea seed contains more tannins than white-flowered peas. Tannins are known to negatively affect nutrient digestibility.
The aim of the study was to evaluate an in vitro assay as an alternative way of estimating pea protein value for pigs. Standardised true ileal protein and amino acid digestibility of peas with different tannin levels was determined in pigs, rats and in vitro. Materials and methods Two cultivars of white flowered (Agra, Albatros) and four cultivars of coloured flowered (Grapis, Selga, Retro, Almara) peas were used in this study. For the in vivo trials, two diets were prepared; one used the pea cv. Agra and the other cv. Grapis. Based on chemical analysis both diets contained the same amount of crude protein and metabolisable energy, 158 g/kg and 13.4 MJ/kg, respectively. The pig experiment used 12 male castrates (55 - 65
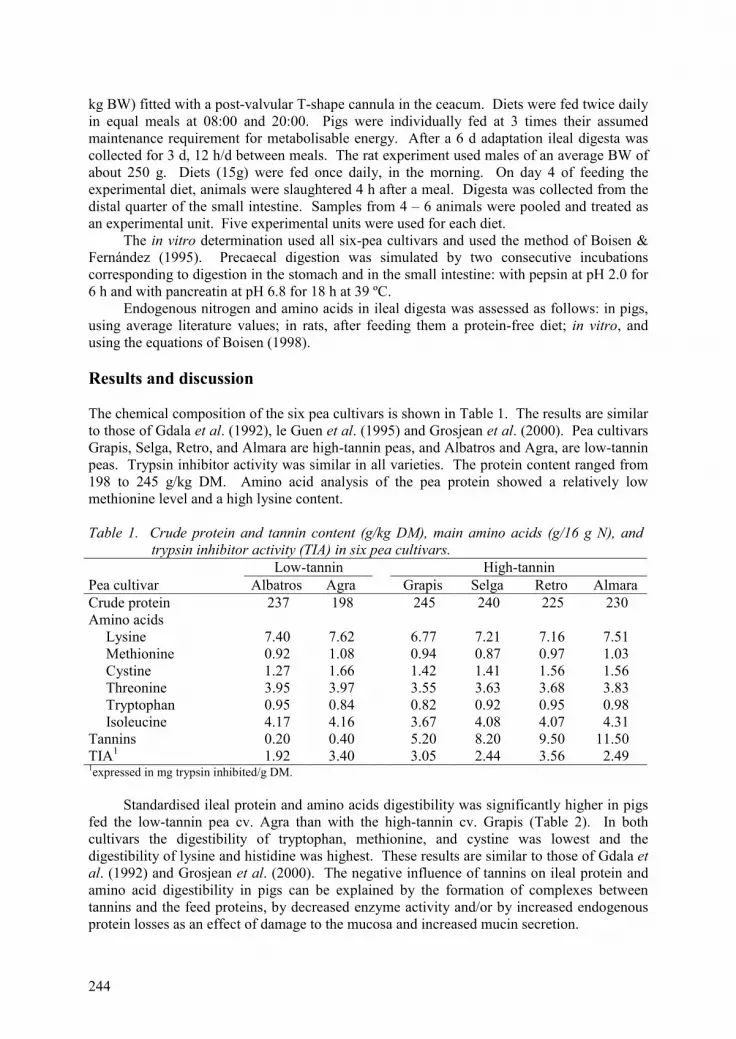
244
kg BW) fitted with a post-valvular T-shape cannula in the ceacum. Diets were fed twice daily in equal meals at 08:00 and 20:00. Pigs were individually fed at 3 times their assumed maintenance requirement for metabolisable energy. After a 6 d adaptation ileal digesta was collected for 3 d, 12 h/d between meals. The rat experiment used males of an average BW of about 250 g. Diets (15g) were fed once daily, in the morning. On day 4 of feeding the experimental diet, animals were slaughtered 4 h after a meal. Digesta was collected from the distal quarter of the small intestine. Samples from 4 – 6 animals were pooled and treated as an experimental unit. Five experimental units were used for each diet.
The in vitro determination used all six-pea cultivars and used the method of Boisen & Fernández (1995). Precaecal digestion was simulated by two consecutive incubations corresponding to digestion in the stomach and in the small intestine: with pepsin at pH 2.0 for 6 h and with pancreatin at pH 6.8 for 18 h at 39 ºC.
Endogenous nitrogen and amino acids in ileal digesta was assessed as follows: in pigs, using average literature values; in rats, after feeding them a protein-free diet; in vitro, and using the equations of Boisen (1998). Results and discussion The chemical composition of the six pea cultivars is shown in Table 1. The results are similar to those of Gdala et al. (1992), le Guen et al. (1995) and Grosjean et al. (2000). Pea cultivars Grapis, Selga, Retro, and Almara are high-tannin peas, and Albatros and Agra, are low-tannin peas. Trypsin inhibitor activity was similar in all varieties. The protein content ranged from 198 to 245 g/kg DM. Amino acid analysis of the pea protein showed a relatively low methionine level and a high lysine content. Table 1. Crude protein and tannin content (g/kg DM), main amino acids (g/16 g N), and
trypsin inhibitor activity (TIA) in six pea cultivars. Low-tannin High-tannin Pea cultivar Albatros Agra Grapis Selga Retro Almara Crude protein 237 198 245 240 225 230 Amino acids
Lysine 7.40 7.62 6.77 7.21 7.16 7.51 Methionine 0.92 1.08 0.94 0.87 0.97 1.03 Cystine 1.27 1.66 1.42 1.41 1.56 1.56 Threonine 3.95 3.97 3.55 3.63 3.68 3.83 Tryptophan 0.95 0.84 0.82 0.92 0.95 0.98 Isoleucine 4.17 4.16 3.67 4.08 4.07 4.31
Tannins 0.20 0.40 5.20 8.20 9.50 11.50 TIA1 1.92 3.40 3.05 2.44 3.56 2.49 1expressed in mg trypsin inhibited/g DM.
Standardised ileal protein and amino acids digestibility was significantly higher in pigs fed the low-tannin pea cv. Agra than with the high-tannin cv. Grapis (Table 2). In both cultivars the digestibility of tryptophan, methionine, and cystine was lowest and the digestibility of lysine and histidine was highest. These results are similar to those of Gdala et al. (1992) and Grosjean et al. (2000). The negative influence of tannins on ileal protein and amino acid digestibility in pigs can be explained by the formation of complexes between tannins and the feed proteins, by decreased enzyme activity and/or by increased endogenous protein losses as an effect of damage to the mucosa and increased mucin secretion.

245
Table 2. Standardized (pig), true (rat), ileal protein (CP) and amino acid (%) digestibility of two pea cultivars determined in pigs and rats (mean ± SD; n = 6).
Animal Pig Rat Pea cv. Agra (LT)1 Grapis (HT) P ≤ Agra (LT) Grapis (HT) P ≤ CP 75.3 ± 1.6 70.6 ± 1.7 0.001 77.0 ± 1.8 78.2 ± 0.6 NS Lys 85.4 ± 1.7 82.2 ± 2.6 0.05 87.8 ± 1.7 86.9 ± 0.6 NS Met 75.5 ± 2.4 63.3 ± 3.1 0.001 89.5 ±1.9 83.1 ± 1.6 0.001Cys 76.0 ± 3.2 65.8 ± 3.5 0.001 80.0 ± 1.5 74.3 ± 0.7 0.001Thr 78.3 ± 3.2 73.3 ± 1.1 0.01 82.9 ± 1.7 81.8 ± 1.2 NS Trp 71.4 ± 2.3 59.3 ± 3.1 0.001 64.9 ± 6.0 65.3 ± 1.4 NS Ile 78.5 ± 2.8 73.7 ±1.4 0.01 80.5 ±1.8 77.6 ± 1.4 0.05 His 84.3 ± 0.9 77.5 ± 1.5 0.001 84.5 ± 2.4 81.4 ± 1.1 0.01 Leu 75.6 ± 2.5 71.2 ± 1.9 0.01 84.2 ± 2.2 80.6 ± 1.0 NS Phe 81.5 ± 1.8 74.2 ± 2.7 0.001 84.6 ± 1.8 82.4 ± 0.9 0.05 Val 77.6 ± 2.1 71.4 ± 1.6 0.001 82.1 ± 1.8 79.4 ± 1.4 0.05 1LT, HT – low- and high-tannin pea cultivars containing 0.4 and 5.2 g tannin/kg DM, respectively.
True ileal amino acids digestibility was higher in rats fed the diet with cv. Agra than with cv. Grapis (Table 2). However, values were only significantly different for methionine, cystine, isoleucine, histidine, phenylalanine, and valine. In contrast to the pigs, rats fed the high-tannin diet showed hypertrophy of the parotid glands and a high secretion of proline-rich protein (Jansman et al., 1994). This proline-rich protein forms complexes with tannins and the tannins do not interact with the feed proteins or other endogenous proteins such as the digestive enzymes. Table 3. In vitro standardized ileal protein (CP) and amino acid (%) digestibility of six pea
cultivars containing different tannin levels (g/kg DM). Low-tannin High-tannin Pea cv. Albatros Agra Grapis Selga Retro Almara Tannins 0.2 0.4 5.2 8.2 9.5 11.5 CP 95.9 86.3 82.6 87.3 85.0 85.6 Lys 94.8 90.2 87.0 86.0 83.5 84.1 Met 91.6 88.3 84.0 82.0 79.9 80.5 Cys 90.6 84.7 81.1 82.2 80.0 80.2 Thr 91.4 85.2 81.7 81.6 78.9 79.2 Ile 93.9 88.3 83.8 85.0 82.4 83.0 His 93.9 89.3 84.7 84.9 82.3 82.9 Leu 94.2 88.4 85.0 83.9 82.5 83.2 Phe 93.8 88.5 84.0 85.0 82.2 83.0 Val 93.3 87.0 83.2 83.9 81.5 82.2
The standardised in vitro ileal protein digestibility was highest in the cv. Albatros and lowest in cvs Retro and Almara (Table 3) after the exclusion of cv. Grapis, which was the lowest for unknown reasons. The values of standardized the in vitro ileal protein and amino acid digestibilities were higher than the corresponding in vivo values for cvs Agra and Grapis. The standardised in vitro ileal amino acid digestibility in the six pea cultivars decreased linearly with increased tannin content (Table 4). The highest correlations were obtained for methionine, lysine, histidine, and lower ones for phenylalanine, valine, and cystine. Garrido

246
et al. (1991) found a close negative (r = -0.88) relationship between the tannin content of faba bean (Vicia faba) seed and their in vitro protein digestibility. Table 4. Relationship between tannin content (x) and standardized ileal in vitro protein and
amino acids digestibility (Y) (n = 6). Equation: Y = b + aX r P ≤ CP Y = 90.1 – 5.13x -0.532 NS Lys Y = 92.4 – 8.23x -0.920 0.01 Met Y = 89.9 – 9.39x -0.957 0.01 Cys Y = 87.3 – 7.08x -0.831 0.05 Thr Y = 88.1 – 8.72x -0.880 0.05 Ile Y = 90.6 – 7.84x -0.851 0.05 His Y = 91.3 – 8.45x -0.903 0.05 Leu Y = 90.9 – 8.14x -0.879 0.05 Phe Y = 90.7 – 7.95x -0.865 0.05 Val Y = 89.7 – 7.81x -0.840 0.05
In conclusion, the in vitro method seems to be useful way of ranking pea cultivars for pig feeding according to their amino acid digestibility. References Boisen, S. 1998. A new protein evaluation system for pig feeds and its practical application.
Acta Agriciculturæ Scandinavica Section A, Animal Science 48, 1-11. Boisen, S. & Fernández, J.A. 1995. Prediction of the apparent ileal digestibility of protein
and amino acids in feedstuffs and feed mixtures for pigs by in vitro analyses. Animal Feed Science and Technology 51, 29–43.
Castell, A.G., Guenter, W. & Igbasan, F.A. 1996. Nutritive value of peas for nonruminant diets. Animal Feed Science and Technology 60, 209–227.
Garrido, A., Gómez-Cabrera, A., Guerrerro, J.E. & Marquardt R.R. 1991. Chemical composition and digestibility in vitro of Vicia faba L. cultivars varying in tannin content. Animal Feed Science and Technology 35, 205-211.
Gdala, J., Buraczewska, L. & Grala, W. 1992. The chemical composition of different types and varieties of pea and the digestion of their protein in pigs. Journal of Animal and Feed Sciences 1, 71–79.
Grosjean, F., Jondreville, C., Williatte-Hazouard, I., Skiba, F., Carrouèe, B. & Gatel F. 2000. Ileal digestibility of protein and amino acids of feed peas with different trypsin inhibitor activity in pigs. Canadian Journal of Animal Science 80, 643–652.
Jansman, A.J.M., Frohlich, A.A. & Marquardt R.R. 1994. Production of proline-rich proteins by the parotid glands of rats in enhanced by feeding diets containing tannins from faba beans (Vicia faba L.). Journal of Nutrition 124, 249-258.
Le Guen, M.P., Huisman, J., Guèguen, J., Beelen, G. & Verstegen M.W.A. 1995. Effects of a concentrate of pea antinutritional factors on the pea protein digestibility in piglets. Livestock Production Science 44, 157–167.

247
The effect of trypsin inhibitor level in soy products on in vitro and in vivo (pigs and rats) protein and amino acid digestibility E. Święch, L. Buraczewska & M. Taciak The Kielanowski Institute of Animal Physiology and Nutrition, Polish Academy of Sciences, Department of Monogastric Nutrition, PL 05 – 110 Jabłonna, Poland Summary Standardized true ileal protein and amino acid digestibility of soybean (Glycine max) diets containing different levels of trypsin inhibitor activity (TIA) were determined in cannulated growing pigs and adult rats and using an in vitro method. Three diets based on soybean meal (SBM) were used. In two diets part of the SBM was replaced with raw (RSB) or extruded (ESB) soybean. Inclusion of RSB or ESB in the diets increased their TIA from 0.90 (in the control diet containing only SBM) to 3.94 and 2.23 units, respectively. Raw soybean, in the diet, significantly decreased standardised true ileal protein digestibility in both animal species. Standardised ileal protein and amino acid digestibility of the ESB diet compared with the SBM diet, was also lower in rats. However, there was no significant difference for all of the amino acids. The standardised in vitro protein and amino acid ileal digestibility was not affected by TIA. Keywords: trypsin inhibitor activity, digestibility, amino acids, pigs, rats, in vitro Introduction Raw soybean is a high-protein feedstuff, but it contains various antinutritional factors (ANFs) including lectins and protease inhibitors (Huisman & Jansman, 1991). Soybean can be processed in several ways, e.g. toasting, roasting or by extrusion, to partially inactivate trypsin inhibitors. However, during heating, amino acids can react with reducing sugars, fat oxidation products, or with phenolic acids, which can reduce their digestibility.
Trypsin inhibitors can cause serious disturbances to physiological processes, including digestion and absorption. Most research on the mode of action of ANFs in animals and studies on reducing their activity by processing have been carried out on small animals such as rats, mice, and chickens.
The aim of this study was to evaluate the effect of different levels of trypsin inhibitor activity (TIA) in soy products on their standardised true ileal protein and amino acid digestibility determined in pigs, rats, and in vitro. Materials and methods The soybean meal (SBM), raw soybean (RSB), and extruded soybean (ESB) used in the study were chemically analysed. Three diets were prepared for the pigs and rats: one diet with SBM (235g/kg) and two diets with mixtures of SBM (150g/kg) and RSB or ESB (100g/kg). The ESB diet was not used in the pig experiment. All diets contained similar amounts of crude protein and calculated metabolisable energy.
The pig experiment used 12 male castrates (65 - 75 kg BW) fitted with a post-valvular T-shape cannula in the ceacum. Diets were fed twice daily in equal meals at 08:00 and 20:00.

248
Pigs were individually fed at 3 times their assumed maintenance requirement for metabolisable energy. After a 6-day adaptation period, ileal digesta was collected for 3 d, 12 h/d between meals.
The rat experiment used male rats of average BW about 250 g. Diets (15 g) were fed once daily, in the morning. On the fourth day of feeding the experimental diet animals were slaughtered 4 h after a meal. Digesta was collected from the distal quarter of the small intestine. Samples from 4 – 6 animals were pooled and treated as an experimental unit. There were 5 experimental units per diet.
The in vitro determination of the soy products used the method of Boisen & Fernández (1995). Precaecal digestion was simulated by two consecutive incubations corresponding to digestion in the stomach and in the small intestine with pepsin at pH 2.0 for 6 h and with pancreatin at pH 6.8 for 18 h at 39 ºC.
The level of endogenous nitrogen and amino acids in the ileal digesta was assessed for the pigs, from average literature values, in rats, after feeding a protein-free diet and in vitro, using the equations of Boisen (1998). Results and discussion Raw soybean and ESB contained less crude protein (387 and 362 g/kg DM, respectively) than SBM (519 g/kg DM) due to the higher fat content of the RSB and the ESB (220 and 240 g/kg DM). Trypsin inhibitor activity, expressed in mg inhibited trypsin/g DM, was 3.8, 32.6 and 15.5 for the SBM, RSB, and ESB, respectively (Table 1). Heating raw soybean can reduce TIA by 57 – 90 %, sometimes down to the level found in SBM (Myer & Froseth, 1989; Herkelman et al., 1992). Inclusion of the RSB or ESB in the diets increased the TIA from 0.90 (in the control diet containing only SBM) to 3.94 and 2.23 units, respectively. Table 1. Crude protein and ether extract (g/kg DM), major amino acids (g/16 g N), and
trypsin inhibitor activity (TIA) in soy products (SBM, RSB, and ESB). SBM RSB ESB Crude protein 518.9 386.9 361.5 Ether extract 24.8 220 240 Amino acids
Lysine 6.29 6.32 6.33 Methionine 1.40 1.44 1.46 Cystine 1.53 1.60 1.59 Threonine 4.07 4.22 4.06 Tryptophan 1.35 1.38 1.31 Isoleucine 4.62 4.63 4.58
TIA1 3.8 32.6 15.5 1expressed in mg trypsin inhibited/g DM.
Inclusion of RSB in the diet significantly decreased (P ≤ 0.001) standardized ileal protein and amino acid digestibility in pigs (Table 2). Similarly, true ileal protein and amino acid digestibility of RSB diet was lower in rats than in the SBM diet (P ≤ 0.001) (Table 3). Inclusion of ESB in rat diets also decreased true ileal protein and amino acids digestibility. However, the values were only significantly different for lysine, tryptophan, histidine, leucine, phenylalanine, and valine (P ≤ 0.001).
Protein digestibility in rats was low compared with amino acid digestibility in rats and protein digestibility in pigs (Tables 2 and 3).

249
Table 2. Standardised ileal protein (CP) and amino acid digestibility (%) of soy products determined in pigs and in vitro.
Pigs (n = 6) In vitro SBM SBM + RSB P ≤ SBM SBM + RSB SBM+ ESBCP 84.5 ± 2.3b 61.9 ± 3.0a 0.001 90.4 93.5 94.4 Lys 89.9 ± 2.2b 72.7 ± 1.8a 0.001 91.7 93.9 94.6 Met 91.0 ± 2.5b 79.1 ± 2.4a 0.001 92.6 95.4 96.2 Cys 82.8 ± 3.7b 67.6 ± 3.1a 0.001 88.4 90.6 91.3 Thr 86.7 ± 2.7b 62.4 ± 3.6a 0.001 89.8 92.0 92.6 Trp 91.7 ± 2.2b 62.5 ± 3.8a 0.001 Ile 88.6 ± 1.6b 64.3 ± 2.0a 0.001 90.9 94.0 94.9 His 90.1 ± 1.5b 72.6 ± 1.9a 0.001 81.8 95.1 96.0 Leu 86.7 ± 1.7b 60.3 ± 2.4a 0.001 90.9 92.5 93.2 Phe 89.3 ± 1.3b 66.5 ± 1.9a 0.001 91.2 94.2 95.1 Val 87.3 ± 1.9b 63.1 ± 2.5a 0.001 89.9 92.4 93.1
Table 3. True ileal protein (CP) and amino acid digestibility of soy products determined in
rats (mean ± SD, n = 5). SBM SBM + RSB SBM + ESB P ≤ CP 76.4 ± 1.9b 64.0 ± 1.5a 76.2 ± 2.1b 0.001 Lys 86.8 ± 1.1c 72.2 ± 1.4a 83.9 ± 1.8b 0.001 Met 89.9 ± 1.2b 73.2 ± 0.9a 90.0 ± 1.6b 0.001 Cys 79.3 ± 3.0b 54.1 ± 2.4a 77.4 ± 2.8b 0.001 Thr 80.6 ± 2.0b 61.9 ± 1.7a 77.9 ± 2.7b 0.001 Trp 77.1 ± 1.9c 42.5 ± 2.0a 68.3 ± 3.6b 0.001 Ile 82.3 ± 1.4b 63.1 ± 2.4a 80.3 ± 2.0b 0.001 His 84.6 ± 1.8c 68.2 ± 1.6a 82.1 ± 2.4b 0.001 Leu 83.9 ± 1.4c 66.4 ± 2.1a 80.2 ± 2.4b 0.001 Phe 84.3 ± 1.4c 68.7 ± 2.2a 80.6 ± 2.2b 0.001 Val 83.3 ± 1.9c 63.7 ± 2.1a 78.6 ± 2.9b 0.001
The negative effect of trypsin inhibitor on protein and amino acid digestibility is related
to the formation of complexes between trypsin inhibitor and trypsin. This decreases trypsin activity and increases protein losses. The protein and amino acid digestibility of the differently processed soy products may be also related to changes in their protein structure and damage to amino acids during heating.
In contrast with the in vivo results, standardised ileal protein and amino acid digestibility determined in vitro was not negatively affected by the inclusion of RSB or ESB in the soybean mixtures (Table 2). It is possible that soy TIA did not influence digestion. This result is probably due to the excess of enzyme used for the in vitro digestion. These results are similar to those of Periago et al. (1996) and Clemente et al. (2000), who also found no effect of TIA on the in vitro protein digestibility of pea (Pisum sativum). Capetillo et al. (2001), however found a significantly (P ≤ 0.05) lower in vitro protein digestibility of raw soybean compared with soybean meal (52 and 87 %, respectively) and Martinez et al. (1998) found a negative relationship (r = -0.65, P ≤ 0.05) between TIA and in vitro protein digestibility in green beans (Phaseolus vulgaris).

250
In conclusion, rats seem to be a useful model to rank soy products according to their amino acid digestibility for pigs because rats, like pigs, are sensitive to TIA in feeds. References Boisen, S. 1998. A new protein evaluation system for pig feeds and its practical application.
Acta Agriculturæ Scandinavica Section A, Animal Science 48, 1-11. Boisen, S. & Fernández, J.A. 1995. Prediction of the apparent ileal digestibility of protein
and amino acids in feedstuffs and feed mixtures for pigs by in vitro analyses. Animal Feed Science and Technology 51, 29–43.
Capetillo, C.M., Abreu, J.E. & Belmar, R. 2001. Standarization of an in vitro method for predicting N digestibility in pig diets. Cuban Journal of Agricultural Science 35, 349–353.
Clemente, A., Vioque, J., Sánchez-Vioque, R., Pedroche, J., Bautista, J. & Millán, F. 2000. Factors affecting the in vitro protein digestibility of chickpea albumins. Journal of the Science of Food and Agriculture 80, 79 – 84.
Herkelman, K.L., Cromwell, G.L., Stahly, T.S., Pfeiffer. T.W. & Knabe, D. 1992. Apparent digestibility of protein and amino acids in raw and heated conventional and low-trypsin-inhibitor soybeans for pigs. Journal of Animal Science 70, 818–826.
Huisman, J. & Jansman A.J.M. 1991. Dietary effects and some analytical aspects of antinutritional factors in peas (Pisum sativum), common beans (Phaseolus vulgaris) and soybeans (Glycine max L.) in monogastric farm animals. A literature review. Nutrition Abstract and Reviews, Series B 61, 901–921.
Martinez, C., Ros, G., Periago, M.J., Ortuňo, J., López, G. & Rincón, F. 1998. In vitro protein digestibility and mineral availability of green beans (Phaseolus vulgaris L.) as influenced by variety and pod size. Journal of the Science of Food and Agriculture 77, 414-420.
Myer, R.O. & Froseth, J.A. 1989. Extruded mixtures of beans (Phaseolus vulgaris) and soyabeans as protein sources in swine and chick diets. In Recent Advances of Research in Antinutritional Factors in Legume Seeds. J. Huisman, T.F.B van der Poel & I.E. Liener (Eds). Proceedings 1st International Workshop on Antinutritional Factors in Legume Seeds, Wageningen 22-25 November 1988, 254–258.
Periago, M.J., Ros, G., Martinez, M.C., Rincón, F., López, G., Ortuňo, J. & Ros, F. 1996. In vitro estimation of protein and mineral availability in green peas affected by antinutritional factors and maturity. Lebensmittel Wissenschaft und Technologie 29, 481-488.

251
Nutritional value and physiological effects of industrial soybean products differing in protein solubility and trypsin inhibitor content M. Taciak, E. Święch & B. Pastuszewska The Kielanowski Institute of Animal Physiology and Nutrition, Polish Academy of Sciences Instytucka 3, 05-110 Jabłonna, Poland Summary The chemical and amino acid composition, trypsin inhibitor activity (TIA), protein solubility in sodium borate and in potassium hydroxide, protein dispersibility index (PDI) were determined in different soybean protein products and casein. The in vitro protein digestibility was also measured. The true digestibility (TD) and biological value (BV) were determined in rats in a 10 d N-balance test. Growth, weight of internal organs and selected gut parameters were assayed in a 4 week experiment.
Trypsin inhibitor activity was highest in extruded soybean (16.85 mg/g). In the other soybean products it was low. The TD and BV of the soybean products were generally lower than in casein except for the TD of soybean isolate and the BV of soybean oil meal (SBM). The TD and BV of extruded soybean did not reflect its high TIA. The pancreas weight either did not differ or was significantly lower in Provasoy 45 and SBM fed rats than on casein fed animals. It was not correlated with TIA. In this study the in vitro ileal protein digestibility values reflected the TD determined in rats. There were correlations between protein solubility in KOH and TD, and in vitro protein digestibility. Keywords: soybean proteins, protein solubility, trypsin inhibitor activity, nutritional value Introduction Recently, many new high protein plant products based, mainly, on soybean are on sale. However, their technology and nutritional value for milk protein replacement in the nutrition of young monogastric animals is not known. They may differ in both the temperature of heat treatment, which affects TIA, protein digestibility, and the concentration or removal of fibre from the bean. There is therefore a need to develop methods to determine the quality and nutritional value of these different protein products. Dudley-Cash (1999) and Batal et al. (2000) proposed protein solubility in KOH and the PDI for monitoring soybean meal quality. Lee & Garlich (1992) proposed solubility in sodium borate. In the case of nutritional value Boisen & Fernandez (1995) developed a method for measuring in vitro ileal protein digestibility.
The objective of this work was to compare the quality and nutritional value of different soybean protein products and casein by in vitro tests and experiments with rats designed to measure protein utilisation (balance test) and their effect on growth rate and selected gut parameters. Materials and methods The following soybean products, SBM, extruded soybean (ESB), soybean protein isolate (SBI), Hamlet Protein-100 (HP-100), Hamlet Protein-300 (HP-300), Soycomil K (SK), Provasoy 45

252
(P45), and casein (CAS), as a control, were compared. Amino acid composition was determined (Buraczewska & Buraczewski, 1984), TIA according to Kakade et al. (1974) and the NDF and ADF fractions according to van Soest (1973). Three measures of protein solubility were used. Solubility in 0.5 % KOH was determined as per Pastuszewska et al. (1998), in sodium borate according to Lee & Garlich (1992) and the PDI that measures protein solubility in water after homogenisation at 8,500 RPM for 10 min. was determined according to Dudley-Cash (1999). All protein products were analysed, in vitro, for ileal protein digestibility (Boisen & Fernandez, 1995).
Two experiments were carried out with male rats (Ifz:BOA strain). Diets contained the same amount of crude protein (9.5 %, N × 6.25), crude fibre (4 %) and fat (4.3 %). True digestibility and BV were determined in a nitrogen balance experiment with 64 rats aged 29 ± 1 d with a mean body weight of 78 g. A growth experiment was performed for 4 weeks on 64 rats, 24 ± 1 d. old and mean body weight 54 g. After euthanasia the small intestine, caecum, liver, pancreas, spleen, thymus gland and kidneys were removed and weighed. The mass and pH of caecal digesta were recorded. Results and discussion The main differences in chemical composition were between CAS, SBI, which had the highest crude protein (> 800 g/kg DM) and the lowest fat contents (≈ 0 g/kg DM), and ESB, which had the lowest crude protein (425 g/kg DM) and the highest fat (186 g/kg DM) content. The TIA was the highest in ESB (16.85 mg/g) but was low in other soybean products (Table 1.). The amino acid composition of all the soybean products was similar. Casein had higher levels of lysine, cystine + methionine, valine, isoleucine and leucine than the soybean proteins. The TD and BV of the soybean products were generally lower than that of casein except for the TD of SBI (Table 2.), this is similar to the results of Mariotti et al. (1999). Among all the soybean products, SBM followed by P45 had the highest while SBI had the lowest BV. The BVs of other products were similar. The high TIA of ESB had no effect on TD and BV values. Total weight gain was greater on casein than on soybean proteins, except for P45 (Table 3.). This in accordance with York & Clemens (1998) and can be explained by the essential amino acid content. In rats fed soybean products the greatest weight gain and the lowest feed to gain ratio was in rats fed SBM and P45. The lowest weight gain and the highest feed to gain ratio was in rats fed SBI. These results are similar to those of Emmert & Baker (1995), and can be explained by different treatments applied during the manufacture of the protein products. Table 1. Proximate analysis of soybean protein products (g/kg DM) and trypsin inhibitor
activity (TIA, mg/g)
Product Crude protein
Ether extract Ash Crude fibre NDF1 ADF2 TIA
SBM 517 27.2 75.1 48.6 86.4 64.4 3.40 ESB 425 186.7 49.0 59.4 91.0 62.6 16.85 SBI 804 2.6 122.7 1.9 nd nd 4.77 HP-100 586 8.6 75.4 32.1 68.4 41.5 2.16 HP-300 592 7.6 70.1 35.8 77.8 45.1 1.84 SK 696 4.3 66.9 32.0 69.9 41.7 3.16 P45 541 15.2 65.3 33.5 80.0 44.2 5.57 CAS 832 nd3 99.5 nd nd nd nd 1neutral detergent fibre. 2acid detergent fibre. 3not determined.

253
Table 2. The true digestibility (TD), biological value (BV), in vitro apparent and standardised protein digestibility and protein solubility (%) in KOH and sodium borate and the protein dispersibility index (PDI).
In vitro digestibility Protein solubility Product TD BV Apparent Standardised KOH Borate PDI SBM 91.4a 80.8d 88.6 94.8 75.2 26.8 14.3 ESB 91.9a 74.4bc 88.1 94.6 72.3 14.9 8.1 SBI 98.1c 69.5a 90.4 96.6 96.8 55.9 63.5 HP-100 92.0a 74.2bc 88.7 95.2 79.7 20.5 11.6 HP-300 92.4a 72.1ab 87.8 94.5 79.2 18.3 10.3 SK 94.7b 72.8b 89.0 95.8 81.1 10.2 5.1 P45 91.5a 77.2c 88.2 94.9 86.1 55.7 25.5 CAS 96.9c 82.3d 92.5 98.6 nd 5.4 0.8
Differences in the weights of the small intestine, pancreas, testes, spleen and ceacum indicate the physiological or metabolic effects of the soy proteins when fed to rats. The highest small intestine weight was in rats fed SBI. It is difficult to explain this without further histological examination. Pancreas weight was not increased by TIA, and, contrary to expectations, it was low in rats fed on the ESB diet, with the highest TIA. Fermentable material in the diet can influence the weight of caecal tissue and digesta. In the case of CAS and SBI there were low caecal tissue and digesta weights. There were higher values in rats fed the other soybean proteins. This may be due to different amounts of fermentable carbohydrates reaching caecum. The caecal digesta pH value supports this interpretation. Table 3. The total weight gain (TWG, g), feed to gain ratio (FGR), organ mass (g/100 g),
mass (g/100 g) and pH of caecal digesta in rats. Caecum Product TWG FCR Small
intestine Pancreas Testes Tissue Digesta pH Spleen
SBM 115cd 3.37b 2.76ab 0.246a 1.365bc 0.379cd 1.27bc 6.42a 0.195b
ESB 109bcd 3.54bc 2.83b 0.267abc 1.298abc 0.358abc 1.05abc 6.54abc 0.200bc
SBI 79a 4.31d 3.12c 0.320cd 1.688d 0.295ab 1.10abc 6.95d 0.215cd
HP-100 114cd 3.45b 2.89b 0.313bcd 1.384bc 0.349abc 1.17bc 6.49ab 0.190ab
HP-300 95b 3.74c 2.84b 0.278abc 1.484c 0.380cd 1.01ab 6.47ab 0.183ab
SK 105bc 3.58bc 2.69ab 0.276abc 1.425bc 0.369bcd 1.07abc 6.60bc 0.174a
P45 123d 3.39b 2.75ab 0.254ab 1.130a 0.445d 1.33c 6.57abc 0.200bc
CAS 123d 3.02a 2.71ab 0.310bcd 1.251ab 0.288a 0.83a 6.98d 0.189ab
In this study in vitro the ileal protein digestibility values reflected the TD determined on
rats. There were correlations between TD and in vitro protein digestibility, both apparent and standardised. Variation in protein solubility varied with the soy protein product and the solvent used. Among soy products protein solubility was the highest in SBI in all solvents (Table 2), the lowest was ESB in KOH and SK in sodium borate and PDI. According to Batal et al. (2000), the PDI is a more consistent and sensitive indicator of heat processing of soybean flakes, than KOH solubility. In their experiment there were regular changes in heating time while in our work evaluated different soy products of unknown technology. There were correlations between protein solubility in KOH and TD, KOH solubility and in vitro apparent and standardised protein digestibility.
In conclusion, the commercial soybean products tested had variable protein values. This can be probably explained by the different treatments that had been used during manufacture. In vitro protein digestibility and protein solubility in KOH can partly replace N-balance tests on

254
rats, but these tests are not sufficient to determine the nutritional value of the products. To do this a long duration growth experiment is required. References Batal, A.B., Douglas, M.W., Engram, A.E. & Parsons, C.M. 2000. Protein dispersibility
index as an indicator of adequately processed soybean meal. Poultry Science 79, 1592-1596.
Boisen, S. & Fernandez, J.A. 1995. Prediction of the apparent ileal digestibility of protein and amino acids in feedstuffs and feed mixtures for pigs by in vitro analyses. Animal Feed Science Technology 51, 29-43.
Buraczewska, L. & Buraczewski, S. 1984. A note on determination of methionine and tryptophan. In T. Zebrowska, L. Buraczewska, S. Buraczewski, J. Kowalczyk & B. Pastuszewska (Eds.) Proceedings 6th International Symposium on Amino Acids, Serock, June 1981, 47-50.
Dudley-Cash, W.A. 1999. Methods for determining quality of soybean meal protein important. Feedstuffs 71, 10-11.
Emmert, J.K & Baker, D.H. 1995. Protein quality assessment of soy products. Nutrition Research 15, 1647-1656.
Kakade, M.L., Rackis, J.J., McGhee, J.E. & Puski, G. 1974. Determination of trypsin inhibitor activity of soy products: collaborative analysis of an improved procedure. Cereal Chemistry 52, 376-382.
Lee, H. & Garlich, J.D. 1992. Effect of overcooked soybean meal on chicken performance and amino acid availability. Poultry Science 71, 499-508.
Mariotti, F., Mahe, S., Benamouzig, R., Luengo, C., Dare, S., Gudichon, C. & Tome, D. 1999. Nutritional value of [15N]-soy protein isolate assessed from ileal digestibility and postprandial protein utilization in humans. Journal of Nutrition 129, 1992-1997.
Pastuszewska, B., Buraczewska, L., Ochtabińska, A. & Buraczewski, S. 1998. Protein solubility as an indicator of overheating rapeseed oilmeal and cake. Journal of Animal Feed Science 7, 73-82.
Van Soest, P.J. 1973. Collaborative study of acid detergent fibre and lignin. Journal Association Official Agricultural Chemists 56, 513-530.
York, J.J. & Clemens, E.T. 1998. Casein, red meat and soyprotein effects on nutrient digestibility and the colonic microstructure of the laboratory rat. Nutrition Research 18, 1057-1066.

255
Intestinal enzymatic activity and amino acid uptake in brush border membrane vesicles of rats fed extruded kidney bean (Phaseolus vulgaris) E. Urdaneta1, J. Barrenetxe1, P. Aranguren1, S. Santidrian2 & F. Marzo1 1Animal Physiology & Nutrition Laboratory, Public University of Navarra, Campus Arrosadia, 31006 Pamplona, Spain 2Department of Human Physiology, University of Navvarra, Irunlarrea s/n, 31008 Pamplona, Spain Summary Legume seeds are an important protein, vitamin and mineral source for humans. However, the nutritional value of legume seed is limited by antinutritional factors (ANFs). Extrusion is a technological process that can reduce these factors without affecting the legume nutritional value. In this work we analysed the effect of raw and extruded beans (Phaseolus vulgaris L.) on intestinal enzymatic activity, amino acid absorption, and their effect on rat growth rate. Male Wistar rats were fed ad libitum for 15 d on three semi-synthetic diets: control (C, 11 % protein from casein), raw and extruded kidney bean (6 % protein from each treated legume + 5 % protein from casein). Rats fed extruded kidney bean had an improved growth rate (P < 0.001), increased intestinal L-leucine absorption (P < 0.001), and higher intestinal enzymatic activity (P < 0.001) compared with the rats fed raw kidney bean. Extrusion seemed to improve animal growth and intestinal physiology.
Keywords: legumes, extrusion, intestinal absorption, enzymes, Phaseolus vulgaris Introduction Legumes are an important source of dietary protein, vitamins, complex carbohydrate, fibre and minerals for animal and human nutrition. Recently their consumption has greatly increased due to nutritional claims that beans have beneficial effects for human health, preventing cardiovascular disease (Friedman & Brandon, 2001), diabetes (Bardocz et al., 1996) and other disorders. However, legumes have limited nutritional value due to the presence of ANFs that have undesirable physiological and biochemical effects. In particular the nutritional value of raw kidney bean is very low due to the presence of a high concentration of lectins that are detrimental (Bardocz et al., 1996). Work in our laboratory has demonstrated these negative effects in laboratory rats (Alonso et al., 2001; Cavallé de Moya et al., 2003;). On the other hand, recently, studies have shown that intake of these compounds at the correct dose may be beneficial by acting as an antioxidant (Friedman & Brandon, 2001), hypoglycaemic (Bardocz et al., 1996) and anticarcinogenic compound (Gerber et al., 2002).
Different technological processes can modify the composition and nutrient availability in raw seed (Della Valle et al., 1994). Extrusion is an economical industrial procedure that is now widely used to improve the nutritive value of legume seed. It can totally or partially reduce ANFs present in legume seed (Alonso et al., 2000). However, chemical alteration of nutrients caused by the process could also decrease nutrient assimilation from heat-treated foods. The aim of the present study was to analyse the effect of an extruded kidney bean diet on intestinal enzymatic activity and leucine absorption of growing rats.

256
Materials and methods Male Wistar rats (80 g; CRIFFA, Barcelona) were fed ad libitum for 15 d. Three semi-synthetic diets: control (C, 11 % protein from casein), raw (RKB) and extruded (EKB) kidney bean (6 % protein content from each legume treatment + 5 % protein from casein) were formulated and body weight and food intake were recorded daily. On day 15, animals were sacrificed and a part of the jejunum and other organs were removed immediately. The jejunum part was processed for isolation of brush border membrane vesicles (BBMV) using the method of Shirazy-Beechey et al. (1990), to undertake absorption studies and determine enzyme activity.
Sucrase (E.C. 3.2.1.48) and maltase activity were determined using the method of Dahlqvist (1964). Alkaline phosphatase (E.C. 3.1.3.1) was determined by measuring the rate of hydrolisis of p-nitrophenilphosphate to p-nitrophenol (Uzeato & Fujita, 1983). The activity of two dipeptidases anchored at the brush border membrane of the enterocytes was also determined (Andria et al., 1980). Finally, amino acid uptake by BBMV was measured using the rapid filtration technique of Hopfer et al. (1975) with slight modifications. Statistical analysis of the data used SPSS software using a one way analysis of the variance test (ANOVA, Kruskall-Wallis). Post-hoc comparisons were done using Least Significance Difference procedures. Differences among groups were considered significant when P < 0.05. Results and discussion The presence of raw bean in the diet decreased final animal body weight (P < 0.001). The same effect was not seen in animals fed an extruded bean diet (Table 1). Intestinal weight was significantly increased (P < 0.01) by raw kidney beans in the diet compared with those fed on extruded kidney beans (Table 1). Table 1. Body weight gain (BWG), food intake and intestinal weight of the different
experimental groups. Results expressed as mean ± SD (n = 5). a: P < 0.05 vs control group; c: P < 0.001 vs control group; f: P < 0.001 vs RKB group.
Parameters C RKB EKB BWG (g) 95.0 ± 2.9 10.0 ± 3.2c 87.0 ± 4.0f Food Intake (g) 232.0 ± 0.0 87.0 ± 5.0a 232.0 ± 0.0f Intestinal Weight (g/100g BW) 4.0 ± 0.4 11.8 ± 1.3c 4.5 ± 0.4f
Raw bean significantly inhibited (P < 0.001) L-leucine uptake by the BBMV compared with the control (Fig. 1). The same effect was observed for intestinal enzyme activity of sucrase (P < 0.001), maltase (P < 0.001) and dipeptidilpeptidase IV (P < 0.001) in the BBMV (Table 2). For both parameters extrusion significantly (P < 0.001) modified the effect. The effects of lectin, on intestinal enzyme activity, has been described in the literature with similar results to those reported here (Ayyagari et al., 1993).
In conclusion, extrusion of Phaseolus vulgaris ameliorates intestinal nutrient uptake and the growth of animals fed on a diet containing the legume. This technique could be useful for the development of different processes and for the development of new animal feds.

257
Table 2. Intestinal enzymatic (µmol/mg protien) activity in BBMV from animals fed control, raw or extruded kidney bean diets. Results expressed as mean ± SD (n = 3). C, P <0.001 vs control group; f, P < 0.001 vs RKB group.
Enzyme C RKB EKB Sucrase 362.0 ± 2.8 196.1 ± 1.4c 265.3 ± 3.1c,f Maltase 479.6 ± 4.5 297.9 ± 4.5c 399.4 ± 6.1c,f Dip. Peptidase IV 0.8 ± 0.01 0.6 ± 0.01c 0.8 ± 0.02c,f
Figure 1. The 0.1 mM L-leucine uptake by BBMV obtained from animals from the different
experimental groups. Results expressed as mean ± SD (n = 4). *** P < 0.001 vs control group; ** P<0.01 vs control group.
References Alonso, R., Aguirre, A. & F. Marzo, F. 2000 Effects of extrusion and traditional processing
methods on antinutrients and in vitro digestibility of protein and starch in faba bean and kidney beans. Food Chemistry 68, 159-165.
Alonso, R., Grant, G. & Marzo, F. 2001. Thermal treatment improves nutritional quality of pea seeds (Pisum sativum L.) without reducing their hypocholesterolemic properties. Nutrition Research 21, 1067-1077.
Andria, G., Cucchiara, S., Vizia, B. de, Ritis, G. de, Mazzacca, G. & Auricchio, S. 1980. Brush border and cytosol peptidase activities of human small intestine in normal subjects and celiac patients. Pediatric Research 14, 812-818.
Ayyagari, R., Raghunath, M. & Rao, B.S. 1993. Early effects and the possible mechanism of the effect of concanavalin A (con A) and Phaseolus vulgaris lectin (PHA-P) on intestinal absorption of calcium and sucrose. Plant Foods for Human Nutrition 43, 63-70.
***
***
***
0 4000 5000 6000
pmol
L-L
euci
ne/m
g pr
ot
0
100
200
300
400
500
seconds
***
***
***
Control RKBEKB
***
***
**

258
Bardocz, S., Grant, G. & Pusztai, A. 1996. The effect of phytohaemagglutinin at different dietary concentrations on the growth, body composition and plasma insulin of the rat. British Journal of Nutrition 76, 613-626.
Cavallé de Moya, C., Grant, G., Frühbeck, G., Urdaneta, E., García, M., Marzo, F. & Santidrián, S. 2003. Local (gut) and systemic metabolism of rats is altered by consumption of raw beans (Phaseolus vulgaris L. var. arthropurpurea). British Journal of Nutrition 89, 311-318.
Dahlqvist, A. 1964. Method for the assay of intestinal disaccharidases. Annals of Biochemistry 7, 18-25.
Della Valle, G., Quillien, L. & Gueguen, J. 1994. Relationships between processing conditions and starch and protein modifications during extrusion-cooking of pea flour. Journal of the Science of Food and Agriculture 64, 509-517.
Friedman, M. & Brandon, D.L. 2001. Nutritional and health benefits of soy proteins. Journal of Agricultural and Food Chemistry 49, 1069-1086.
Gerber, M., Boutron-Ruault, M.C., Hercberg, S., Riboli, E., Scalbert, A. & Siess, M.H. 2002. Food and cancer: state of the art about the protective effect of fruits and vegetables. Bulletin du Cancer 89, 293-312.
Hopfer, U., Sigrist-Nelson, K., Perotto, J. & Murer, H. 1975. Intestinal sugar transport: studies with isolated plasma membranes. Annals New York Academy of Science 264, 414-427.
Shirazy-Beechey, S.P., Davies, A.G., Tebbut, K., Dyer, J., Ellis, A., C.J. Taylor, C.J., Fairclough, P. & Beechey, R.B. 1990. Preparation and properties of brush border membrane vesicles from human small intestine. Gastroenterology 98, 676-685.
Uezato, T. & Fujita, M. 1983. Developmental transition of alkaline phosphatase from suckling to adult type in rat small intestine: molecular species and effect of injected cortisone and thyroxine. Journal of Biochemistry 94, 1483-1488.

Session V A Application of biotechnology and processing to reduce ANFs and to increase nutritional quality. Potential use of GMOs.


261
The potential of genetically modified legume and oilseed crops for food and non-food use R. Greiner1 & C. Domoney2
1Federal Research Centre for Nutrition, Centre for Molecular Biology, Haid-und-Neu-Strasse 9, 76131 Karlsruhe, Germany 2John Innes Centre, Norwich Research Park, Colney, Norwich NR4 7UH, United Kingdom Summary Gene technology provides a substantial opportunity to alter the composition of crops to match current and emerging food and industrial uses. Many of the genetic improvements in legumes and oilseeds in the foreseeable future will result from the use of molecular genetic techniques to introduce new genes, modify existing ones and to provide more efficient means to identify specific combinations of genes. World-wide, there are currently hundreds of field trials of novel transgenic legume and oilseed lines, evaluating traits such as herbicide resistance, resistance to insects, fungi and viruses, male sterility and restoration, modified oil, protein and starch composition, pharmaceutical production, production of biodegradable plastics, and stress tolerance. The enormous value of genetic modification as a laboratory tool should not be under-estimated. Such an approach allows the function of genes and the role of enzymes to be defined rapidly and target genes thus identified may be manipulated subsequently, using either transgenic or conventional breeding. Keywords: crop plants, genetic modification, input traits, nutritional quality, output traits Introduction Legumes and oilseeds are of great global importance due to their significance in human and animal nutrition. Although the primary marketplace for legumes and oilseeds is the food and feed market, these crops are being used additionally for a variety of non-food purposes. Gene technology provides a substantial opportunity to tailor the composition of grains to current and emerging food and industrial uses. A well-defined, preferably simple, shoot regeneration protocol is a pre-requisite for the production of transgenic plants. Grain legumes are one of the groups of dicotyledonous crops least amenable to transformation, although they are usually susceptible to Agrobacterium infection. Particle bombardment is an alternative procedure for those legumes that fail to respond to Agrobacterium-mediated gene transfer. Over the last few years, transgenic crops have moved from being a laboratory curiosity to providing new varieties grown in large areas throughout the world. To date, the predominant transgenic crops are maize (Zea mays), soybean (Glycine max), oilseed rape (Brassica spp) and cotton (Gossipium spp). It is not surprising that oilseed rape and soybean were the first genetically modified crops to be commercialised, since Brassica species, particularly Brassica napus, and soybean have been the focus of much of the fundamental research conducted on plant genetic engineering techniques. Gene transfer protocols and regeneration systems have now been developed for the following target legume and oilseed species (Babaoglu et al., 2000; Kuchuk et al., 2001): oilseed rape (Brassica napus), soybean (Glycine max), brown mustard (Brassica juncea), cotton (Gossypium hirsutum), pea (Pisum sativum), chickpea (Cicer arietinum), lentil (Lens culinaris), lupin (Lupinus angustifolius, L. mutabilis), common

262
bean (Phaseolus vulgaris), narbon bean (Vicia narbonensis), faba bean (V. faba), moth bean (Vigna aconitifolia), black gram (V. mungo), mungbean (V. radiata), cowpea (V. unguiculata), alfalfa (Medicago sativa), subterranean clover (Trifolium subterranean), white clover (Trifolium repens), peanut (Arachis hypogaea), sunflower (Helianthus annuus), linseed (Linum usitatissimum), white mustard (Sinapis alba), and poppy (Papaver somniferum). Input traits Genetically modified crop production characteristics, such as herbicide tolerance and insect resistance, have become known as "input traits", and constitute the first wave of GM products to reach the market. The benefits from input traits are confined mainly to crop production systems and are therefore captured principally by growers and agribusiness. However, many input traits also have substantial benefits to agro-ecosystems, such as reduced chemical usage, as well as having the potential to reduce food costs through improvements in production efficiency. Some of the key examples of input traits that have been developed in legumes and oilseeds are outlined below. Herbicide tolerance Resistance to a number of different herbicides has been incorporated into legumes and oilseeds, using genes isolated mainly from microorganisms (Babaoglu et al., 2000; Kuchuk et al., 2001). Many of these, such as tolerance to Roundup Ready®, Liberty Link®, and Bromoxynil®, are already being used commercially in several oilseed crops, notably soybeans, canola and cotton. Insect resistance Control of insect pests represents one of the major input costs of world agricultural production. The genetic engineering of insect resistance has been challenged by the great adaptability of insects and by the problem of non-target impacts. In general, resistance based on multiple genes is less easily broken than a monogenically based resistance.
Crops expressing insecticidal crystal proteins from Bacillus thuringiensis (Bt) were among the first transgenic products approved for commercial use (Hegedus et al., 2002). These proteins are valuable in the control of lepidopteran and, to a lesser extent, coleopteran insects. However, there is still a need to develop alternative strategies, particularly for control of homopteran insects, many of which transmit agronomically important diseases and are not targeted by existing Bt toxins. Vegetative insecticidal genes from Bacillus species may provide a useful alternative or supplementary source of resistance in the future, but first these have to be expressed effectively in transgenic plants (Hegedus et al., 2002). The insect gut is the obvious target for the pore-forming or lytic activity of both the Bt and the vegetative insecticidal proteins.
Nutrient uptake by the insect presents another target. In this regard, inhibitors of digestion have proved effective against diverse lepidopterans and/or sucking pests. Trypsin-like enzymes are the dominant proteases in lepidopteran larvae and genes encoding trypsin-chymotrypsin inhibitors have been expressed in plants (Murdock & Shade, 2002). These transgenic plants demonstrated successful anti-feedant activity against lepitopteran larvae. Unfortunately, the utility of protease inhibitors is complicated by the observation that insects compensate for the loss of proteolytic activity by increasing the synthesis of the affected

263
protease and by expressing novel proteases that are insensitive to the inhibitor (Gatehouse et al., 1997). Some insects feed almost exclusively on starch and these are particularly susceptible to amylase inhibitors. Therefore, the expression of α-amylase inhibitors in some legumes may confer natural resistance to such insects. Field trials of transgenic peas expressing an α-amylase inhibitor gene from the common bean, Phaseolus vulgaris, showed a high resistance to attack by pea weevils (Bruchus pisorum) (Morton et al., 2000). This technology holds promise, therefore, for protecting grain legumes against coleopteran pests. Few insecticidal proteins have been found that are active against sap-sucking insects, such as plant hoppers, leaf hoppers, and aphids, although, surprisingly, inhibitors may prove useful also for some of these (Wang et al., 2003). Sugar-binding proteins, or lectins, have been purified from several plant species, and many lectins have been reported to have toxic effects on sap-sucking insects that derive most of their energy reserves from the sugars being transported in the phloem (Murdock & Shade, 2002). Furthermore, chitinase, lipoxygenase, cholesterol oxidase and the biotin-binding proteins, avidin and strepavidin, have also been shown to have strong insecticidal activity, but few of these genes have been tested in transgenic crop plants (Hegedus et al., 2002). Disease resistance The majority of virus-resistant transgenic plants result from the application of the concept of pathogen-derived resistance, based on the use of virus-derived genes and gene segments as the source of resistance. Many viral gene products, for example, coat proteins, replicase, movement proteins, and proteinases, can trigger a host response. A host response induced by many of these, that leads to inhibition of viral multiplication and death of infected parts, represents the best known barrier for viral infection (Fuchs & Gonsalves, 2002). Others are being investigated to provide additional mechanisms in the near future. In particular, apoptosis, the programmed cell death of single cells thereby preventing necrosis of tissues, might provide an interesting candidate process to develop for improved resistance. It has been shown additionally that RNA alone can mediate protection. In order to block translation of viral RNA by means of hybridisation, antisense RNA has been expressed in various plants. Surprisingly, in some cases, sense RNA also provided protective effects by co-suppression (Fuchs & Gonsalves, 2002).
Potential anti-fungal genes include hydrolases of fungal cell walls, namely β-1,3-glucanase and chitinase. For example, an enhanced tolerance against several fungi was shown in chitinase-expressing rape (Brassica napus) under natural field conditions. Furthermore, genes encoding plant protease inhibitors, polygalacturonic acid inhibitors, and osmotin have been used to increase fungal tolerance of plants (Tenhagen, 2002).
Several components of the general plant response system have been manipulated in order to obtain protection against pathogens, such as viruses and fungi, but so far only with limited success. Pathogens encounter an oxidative burst early, followed by the transcriptional activation of pathogenesis-related proteins, accumulation of phytoalexins, and a systemic signal mediated by salicylic acid, leading to systemically acquired resistance in non-inoculated tissues or organs (Tenhagen, 2002). A further general response has been attributed to ribosome-inactivating proteins, which accumulate in certain plants to high levels in an inactive pre-form that is rendered active during wounding to inactivate ribosomes of distantly related, but not the same, species (Tenhagen, 2002).
Plants have also developed an efficient mechanism to combat pathogens by synthesising small anti-microbial peptides, collectively termed defensins (Tenhagen, 2002). Defence peptides exhibit a broad spectrum of anti-microbial activity against bacteria, fungi, and even

264
enveloped viruses. The large diversity of these peptides provides a huge reservoir of genes that can be expressed in plants to enhance tolerance against pathogens. Until now, the over-expression of plant defence peptide genes to achieve higher tolerance against a broad range of pathogens has only been partly successful.
The efficient protection of many wild-type plants against microbial pathogens appears to be mediated at least in part by plant secondary metabolites that are toxic to microbes. Before we can engineer pathogen tolerance routinely by manipulating secondary metabolism in plants, we must develop a better understanding of the complex interactions between different metabolic pathways and their regulation. Current strategies are based on the expression of a single gene or a few genes to equip a plant with a novel secondary metabolite. A successful example is the expression of a grapevine stilbene synthase gene in alfalfa under the control of its native pathogen-inducible promoter. The enzyme generates the stilbene resveratrol, which has anti-microbial properties in the transgenic plants (Hipskind & Paiva, 2000).
It is evident that single target mechanisms have a high chance of being overcome by pathogens. From this viewpoint, more complex defence strategies should be beneficial for enhanced long-term pathogen tolerance.
Nematode resistance Nematodes are very difficult to control chemically, and the nematocides currently available have highly undesirable environmental characteristics. Transgenic approaches offer the opportunity to pursue alternative strategies for establishing nematode resistance in plant species (Cai et al., 2002). Natural nematode tolerance plant genes have been identified and introduced into elite breeding lines. The combination of these resistance genes with artificial resistance mechanisms, such as proteinase inhibitors, lectins or monoclonal antibodies, offers the opportunity to breed varieties with improved and broad resistance. Hybrid pollination systems Hybrid vigour has been described in many plant species and has formed the basis for producing high-yielding cultivars in several crops, including oilseeds. However, the high cost of developing conventional hybrid systems, coupled with the fact that suitable systems do not exist for some important crops, has prompted several efforts to develop genetic methods of preventing pollen development. The most advanced of these involves the introduction of the barnase gene that encodes a ribonuclease enzyme that destroys mRNA specifically in the developing pollen to generate male-sterility in the female parent. The barstar gene is incorporated in the male parent to inhibit the action of the ribonuclease and restore normal fertility in the resulting hybrid plants. This system is already in commercial use for production of hybrid canola varieties and could also be used in other crops (Fobert, 2002; Palmer & Keller, 2002). Output traits The modification of product quality characteristics using gene technology depends on a well-established understanding of the pathways for biosynthesis of plant products, a rapidly expanding knowledge of the genetic control of these pathways, and an increasing availability of cloned genes for key enzymatic steps.

265
Improved protein quality Legume seeds and oilseeds are deficient in both cysteine and methionine; animals are capable of converting methionine to cysteine but not vice versa. Consequently, there is an interest in increasing the methionine content of legume seeds. The main interest in improving the sulphur amino acid content of legume seeds has focused on the methionine-rich 2S albumins from Brazil nut (Bertholletia excelsa) and sunflower. Expression of 2S albumin genes has resulted in significant increases in the methionine content of oilseed rape and narbon bean (Müntz et al., 1998). However, the Brazil nut protein can be allergenic to some humans and the commercial development of seeds expressing this protein has therefore been suspended. It is clear that, in some species, amino acid reserves may limit the plant's ability to respond to the added demand imposed by the transgene and the limited sulphur reserves may simply be re-allocated from endogenous proteins to the new sulphur sink (Demidov et al., 2003: Hagan et al., 2003). Attempts have been made also, using in vitro gene mutation, to alter appropriate amino acid codons into methionine and/or lysine codons, or to insert stretches of additional codons for these amino acids, into individual genes for legume and oilseed storage proteins (Müntz et al., 1998). In addition, amino acid metabolism within the seed has been modified in order to increase the free amount of the respective essential amino acid by engineering regulatory key enzymes in the biosynthetic pathway (Galili & Höfgen, 2002). High-lysine soybean and rapeseed have been produced by this approach, since synthetic lysine is used to supplement soy- and rape-based animal feed (Falco et al., 1995). It has been shown recently that expression of both a methionine-rich albumin and a feed-back insensitive bacterial aspartate kinase, to stimulate methionine biosynthesis, can result in narbon bean protein with 2 - 2.4 times higher methionine content than the wild-type (Demidov et al., 2003).
Forage crops are excellent sources of protein for grazing animals, but significant protein value is lost to bacteria during digestion in the rumen. To improve forage protein quality, lucerne (Medicago sativa), lupin and subterranean clover have been modified to produce a protein rich in sulphur-containing amino acids, but also resistant to rumen degradation (Tabe et al., 1993). Feeding trials using modified lupins resulted in an 8 % increase in wool growth and a 7 % increase in live weight gain in sheep; in addition, other aspects of ruminant animal production may be improved through increasing the supply of sulphur amino acids absorbed into the bloodstream (Molvig et al., 1997). In general, protein quantity, as well as quality, may be improved indirectly as a pleiotropic consequence of manipulating pathways leading to other components; an increase in protein content has been demonstrated following antisense inhibition of the starch pathway in narbon bean (Weber et al., 2000). Improved nutritional quality of plant oils The manipulation of seed oil content via transgene insertion was an early goal of the application of modern biotechnology to agriculture and has led to the production of crops with modified oils that are under trial, principally in the USA and Canada. There are three obvious targets for the modification of edible oils in seeds. First, to increase the total 18:0 and 18:1 content of the plant oil so that it has an acceptable solid fat functionality for use in margarine and other confectionery applications but without the deleterious health effects of partially hydrogenated oils rich in trans-fatty acids (Thelen & Ohlrogge, 2002). The oleic acid levels have been raised (using transgenic approaches) to 88 % in soybean oil, to 89 % and 75 % in canola oil from Brassica napus and Brassica juncea respectively, and to 77 % in cottonseed oil (Liu et al., 2002; Thelen & Ohlrogge, 2002). In addition, transgenic soybeans, sunflower, cotton and canola produce an oil enriched in stearic acid (35-40 %) (Thelen & Ohlrogge,

266
2002). By inter-crossing a high-oleic and a high-stearic genotype of cotton, it has been possible to obtain a cottonseed oil with intermediate levels of oleic and stearic acid (Liu et al., 2002). Secondly, work is ongoing to increase monounsaturates (18:1) with a concomitant decrease in polyunsaturates (18:2, 18:3), combined with a reduction in total saturates (16:0, 18:0) (Coughlan & Kinney, 2002). This would yield oil that is more chemically stable and having a reduced total saturated fat content. High-oleic/low-linolenic and high-lauric oilseed rape is being grown commercially in Canada. Furthermore, a low-saturated (3 %) low-linolenic (3 %) soybean oil is on the market in the USA. Thirdly, there is increasing interest in producing very long-chain polyunsaturates, such as linolenic acid (18:3ω6, GLA), docosahexenoic acid (22:6ω3, DHA) and eicosapentenoic acid (20:5ω3, EPA), which are nutritionally beneficial as precursors for certain prostaglandins and as cholesterol-lowering agents (Huang et al., 2001). The accumulation of up to 68 % linolenic and up to 17 % stearidonic acid (18:4ω3, SDA) in oilseed rape has been reported (Ursin et al., 2000). It seems likely, therefore, that genetically modified oilseeds having nutritionally effective levels of long-chain polyunsaturates will become a reality but there is still much progress needed, in particular for some fatty acids (e.g. fish oil-types). Altered carbohydrate composition Carbohydrates provide clothing and shelter as the cellulose of cotton and wood, a means of communication as the cellulose of paper, dietary fibre as starch and β-glucans, the basis of most beverages ranging from fruit juice and soft drinks to cognac, and dietary nutrients. So far, the main efforts in carbohydrate engineering in plants have been directed to altering starch yield and type, and to changing the degree of branching in amylopectin (Schulman, 2002). The utilisation of starch and other plant carbohydrates as replacements for petrochemicals offers great potential for the farming of crops containing specialised storage products. In addition to non-food uses, applications ranging from fat substitutes to fibre as resistant starch in novel foods promise to create new markets for plant carbohydrates. Additional objectives include the conversion of starch to simple sugars (fructose or trehalose) or the production of novel sugars (mannitol or sorbitol), to confer osmoregulatory and stress-ameliorating benefits to plants, and the production of fructans, as potential pro- or pre-biotic anti-tumorigenic components of the human diet (Schulman, 2002).
Cellulose is of high value for the pulp, paper and textile industries. Genes have been targeted to improve the properties of the cotton fibre (Chapple & Carpita, 1998; Delmer & Haigler, 2002); these include increased length, strength, thermal properties, improved dye-binding, and resistance to shrinkage and wrinkling. These properties have been improved through classical breeding efforts but additional improvement is possible through genetic engineering. For example, 'naturally blue' cotton would provide a product of enhanced value and would give considerable environmental benefit. Genes from bacteria have been isolated that generate indigo, or indigoidine, from common plant chemicals and it is hoped to divert some of these precursors into the production of blue pigments, initially in model plants.
Cellulose is also an important carbon source for ruminants and the value of forage crops could be improved greatly if the ratio of cellulose to other wall polymers was increased. Secondary metabolite production The biosynthetic pathways for the synthesis of secondary metabolites are rapidly being elucidated using biochemistry, gene technology and genomics in model systems such as Arabidopsis. The increasing availability of key genes controlling these pathways is now

267
enabling the content of these valuable micro-nutrients to be enhanced in plant foods. It is possible to use gene technology to increase greatly the naturally low levels of phytosterols in oils up to levels that have a nutritional benefit. Seed oils also contain low levels of β-carotene, which the body converts into vitamin A. Research has shown that β-carotene levels in canola (Brassica napus) seed can be raised up to 50-fold by the introduction of just one gene targeted to the plastid (Verporte & Memelink, 2002). As β-carotene is lipid-soluble, the majority of it will be extracted in the seed oil. The entire pathway to β-carotene has been engineered into rice ('golden rice') for the future relief of blindness in millions of children, following development and trials of appropriate rice cultivars (Potrykus, 2001). The levels of antioxidants in seed oils may also be enhanced, once genes in the tocopherol and tocotrienol pathways are expressed transgenically. So far, it has been shown that the relative proportions of α-tocopherol (vitamin E) and ß-tocopherol, which differ in their effectiveness as antioxidants, can be modified by gene technology (Yan & Kerr, 2002). There has been one successful attempt to increase plant L-ascorbic acid content through genetic engineering (Hancock & Viola, 2002).
Gene technology is being used in an attempt to increase the levels of condensed tannins in pasture legumes (white clover and lucerne) in order to reduce the risk of bloat in ruminants (Xie et al., 2003). Such tannin-accumulating transgenic lines would also have better protein quality, which could boost dairy and beef productivity. Other approaches to increase plant nutritive value Lignin is the major structural component of secondary thickened plant cell walls. Both lignin concentration and lignin methoxyl content have been negatively correlated with forage digestibility for ruminant animals. Therefore, there is considerable interest in the potential for genetic manipulation of lignin to improve digestibility of forages (Chapple & Carpita, 1998; Dixon et al., 1996). Several reports have been published on alterations in lignin content or composition as a result of genetic manipulation but, due to pleiotropic effects on, for example, increased disease susceptibility, there is much to be achieved before practical application (Dixon et al., 1996). Furthermore, genetic engineering is being used to reduce the content of anti-nutrients such as phytate (Lucca et al., 2001; Raboy, 2001), protease inhibitors (Welham & Domoney, 2000) and glucosinolates, as well as oestrogen-like compounds, which may lower animal fertility. Industrial crops Most output traits are being genetically modified initially for improved food uses. However, in the future, there is likely to be a "third wave" of GM crops where the product modifications will be designed for specific industrial use. Metabolic engineering research is currently at an advanced stage in the process of re-directing the biosynthetic capabilities of seeds towards the production of novel compounds, such as industrial raw materials and pharmaceuticals. The development of GM crops containing industrial compounds could contribute to converting traditional agricultural crops into efficient producers of more valuable chemical commodities and thereby provide renewable plant sources of raw materials currently obtained from non-renewable petroleum.

268
Industrial plant fatty acids In addition to their importance in human nutrition, plant fatty acids are also major ingredients of non-food products, such as soaps, detergents, lubricants, biofuels, cosmetics, and paints. In principle, it is now possible to envisage the production on an agricultural scale of a vegetable oil containing any fatty acid chain length ranging from C6 to C24 with varying degrees of unsaturation, epoxidation, branching or hydroxylation, depending on the intended application (Cahoon et al., 2002). Oils have been obtained containing 8:0, 10:0, 12:0 and 14:0, and various combinations of these fatty acids, in significant amounts (Thelen & Ohlrogge, 2002). For example, genetic modification of oilseed rape resulted in oil enriched in 8:0 (10 mol %) and 10:0 (25 mol %) or containing 14:0 at up to 40 mol %. These oils have diverse applications in the food, pharmaceutical, and lubricant industries. The first commercial oil in this respect was derived from transgenic oilseed rape in 1995, contained 40 mol % laurate (12:0), and found applications in the manufacture of soaps and detergents. A wide range of oils have been produced through genetic engineering, including those enriched in fatty acids with epoxy and hydroxyl groups and conjugated double bonds (Cahoon et al., 2002). These types of oils can be used in industrial applications such as plasticizers, drying oils, adhesives, and nylon and composite precursors. A more ambitious plan is the production of long-chain wax esters (C20, C22, C24) in oilseeds (Thelen & Ohlrogge, 2002). Many of the waxes found in nature have commercial uses in the lubricant, food, and cosmetic industries. Limitations on supply has led to interest in manipulating crops to produce wax at reasonable cost. Considerable progress has been made in identifying the key enzyme involved in wax production in jojoba (Simmondsia chinensis) seeds and co-expression of three enzymes on the pathway has led to the accumulation of waxes at 50-70 % of oil molecules in Arabidopsis seeds (Lardizabal et al., 2000). This approach suggests that commercial oilseed crops may be manipulated in a similar fashion. Biodegradable plastics Polyhydroxyalkanoates (PHAs) are bacterial polyesters having properties of biodegradable thermoplastics and elastomers and their synthesis in crop plants is seen as an attractive system for the sustained production of large amounts of polymers at low cost. Since the first demonstration of the synthesis of polyhydroxybutyrate in plants in 1992, the range of PHAs, which have been synthesised in plants, has increased to include a number of co-polymers (Poirier, 2002). Even though steady progress has been made towards creating transgenic crops for commercialisation of PHAs, the production of PHAs in high quantity in plants, without affecting the yield of other plant products, remains a problem. It has been demonstrated, by expressing two enzymes of the polyhydroxybutyrate pathway under the control of fibre-specific promoters in cotton, that improvement of insulating properties of fibres is feasible (John & Keller, 1996). Pharmaceuticals, peptides and industrial enzymes Recent work suggests that plants provide economic bio-reactors for large-scale production of industrial and pharmaceutical proteins. The most widely studied therapeutic proteins produced in plants have been monoclonal antibodies for passive immunotherapy or diagnostics, and antigens for use in oral vaccines (Daniell et al., 2001; Larrick & Thomas 2001). Although several such proteins have been expressed successfully in transgenic plants, very few have made their way into clinical trials. The most advanced product in human

269
clinical trials is an anti-Streptococcus mutans secretory immunoglobulin A plantibody, currently in phase II clinical trials for prevention of dental caries (Larrick & Thomas, 2001). The recent availability of large amounts of secretory immunoglobulin A plantibodies opens up a number of novel therapeutic opportunities for disorders of the mucosal immune system. Furthermore, transgenic plants have been constructed that express proteins such as enkephalins, α-interferon, human serum albumin, hirudin, α-1-antitrypsin, avidin, β-glucuronidase, lactoferrin, collagen, and two of the most expensive drugs: glucocerebrosidase and granulocyte-macrophage colony-stimulating factor. Recombinant bioactive avidin and β-glucuronidase were the first recombinant plant-derived proteins to be produced commercially (Larrick & Thomas, 2001). The low yield of recombinant protein and its subsequent purification represent the major limiting factors in the use of plants to express foreign proteins. Consequently, the therapeutic anticoagulant hirudin and the human neuropeptide leu-enkephalin have been produced using oleosin-fusion technology, which allows extraction using canola oil bodies (Parmenter et al., 1995).
Gene technology is being exploited also to modify the content or profiles of morphinan alkaloids for pharmaceutical use. Oilseed poppies remain the exclusive source of several alkaloids of pharmaceutical importance. Modifying the activities of particular enzymes involved in alkaloid synthesis can result in either elevated or reduced concentrations of alkaloids in transgenic plants (Hughes & Shanks, 2002).
Edible vaccines produced in transgenic plants have the potential to revolutionise vaccination. Several vaccines for human and animal use, including vaccines for hepatitis B (lupin, lettuce (Latuca sativa)) (Kapusta et al., 1999), foot-and-mouth disease (lucerne) (Wigdorovitz et al., 1999) and bovine pneumonia pasteurellosis (white clover) (Lee et al., 2001), have been produced in transgenic plants and demonstrated to be both immunogenic and protective.
Many industrial processes involve the degradation of plant products and much research on recombinant enzyme production in plants has focused on the production of enzymes, such as amylases, phytases, cellulases, proteases and lipases (Giddings, 2001). Phytoremediation The concept of phytoremediation to clean up metal-contaminated soil has invoked considerable interest (Krämer & Chardonnens, 2001). This technology relies on metal-accumulating plants to transport and concentrate polluting metals from the soil into harvestable shoots. The introduction of metal-binding peptides and proteins, such as metallothioneins, metal transporters or phytochelatins, into high biomass plants is an attractive strategy. Numerous attempts to engineer the production of metallothioneins in plants have been reported (Hasegawa et al., 1997; Mejáre & Bülow, 2001; Pan et al., 1994). So far, however, it is mainly model plants that have been tested only on a laboratory scale. Thus, it remains to be seen whether phytoremediation will be an industrially viable concept. Further activities Photosynthetic enhancement and yield increase Clearly, the first priority for plant breeders is yield and it is assumed generally that improved energy capture can be translated into higher yields of harvestable commodities. Although there is still little reliable information that relates transgenic modification of specific photosynthetic genes to performance under agricultural conditions, results from many

270
preliminary tests have been published. One approach has been to introduce key biochemical features of the C4 photosynthetic pathway into C3 plants (Jeanneau et al., 2002). The justification for this approach is based on the fact that C3 photosynthesis suffers from O2 inhibition due to the oxygenase reaction of ribulose 1,5-bisphosphate carboxylase/oxygenase, and the subsequent loss of CO2 from photorespiration. The first transgenic plants exhibited reduced O2 inhibition of photosynthesis but photosynthetic rates were comparable to those of untransformed plants.
Nitrogen and phosphorus are the most important nutrients that limit agricultural production. Research is ongoing to improve ammonium assimilation and phosphorus uptake by means of genetic engineering. It has been demonstrated that constitutive over-expression of cytosolic glutamine synthetase increases photosynthetic productivity under conditions of N-deficiency (Fuentes et al., 2001). Modification of the nitrogen-fixing symbiosis between legume roots and bacteria of the species Rhizobium and Bradyrhizobium is also a strategy to improve ammonium assimilation. Recent progress in our knowledge of nodule organogenesis and function has enabled various metabolic pathways to be targeted for genetic manipulation (Dixon et al., 1996). Since nodules of tropical legumes export fixed nitrogen in the form of ureides, allantoin, and allantoic acid, which are synthesised by the oxidation of purines, de novo purine synthesis is a critically important pathway in these nodules (Atkins, 1991). Over-expressing the rate-limiting enzyme to increase purine synthesis in such nodules may be a way to improve nitrogen fixation, resulting in enhancement of overall plant productivity.
When plants are grown under phosphate-deficient conditions, increased production and secretion of organic acids make phosphate available for uptake by high-affinity phosphate transporters in the plasma membrane, due to the liberation of phosphate from poorly soluble forms in the soil. Transgenic plants over-expressing specialised phosphatases, such as apyrase or phytases, have shown enhanced growth coupled with augmented phosphate transport (Thomas et al., 1999; Richardson et al., 2001).
These preliminary data suggest that transgenic technology offers a promising approach to reducing agricultural nitrogen and phosphorus fertilisation and the concurrent deleterious impacts on the environment. Abiotic stress Abiotic stresses such as drought, low temperatures, heat, high salinity and pollution are environmental factors that dramatically limit plant growth and crop productivity. Molecular studies have focused on the identification of genes that are activated by different forms of stress. However, the function of few of these genes is known, which makes it difficult to design approaches to improve stress tolerance. Since active oxygen species are involved in a wide variety of environmental stresses, antioxidant enzymes are obvious molecular targets for the production of stress-tolerant transgenic plant varieties (van Breusegem & Inzé, 2002). Several antioxidative stress enzymes such as superoxide dismutase, ascorbate peroxidases, and catalases have been genetically engineered into plants. In general, transgenic plants expressing an additional superoxide dismutase enzyme have improved stress tolerance and vigour (Samis et al., 2002). Furthermore, small heat shock proteins have the potential to protect cells from stress damage (Sun et al., 2002). Accumulated data from the genetic modification of such heat shock proteins suggest that increased production correlates with better plant stress tolerance. A further strategy to obtain very drought- and salt-tolerant transgenic plants is to over-produce osmolytes, such as mannitol, fructans, proline, glycine-betaine, trehalose and polyols (Rontein et al., 2002). The feasibility of using antibody-expressing transgenic plants to neutralise toxic metals was shown recently (Drake et al.,

271
2002). Furthermore, through a biotechnological approach, certain crops have been shown to be more aluminium tolerant due to an increase in citric acid synthesis (Clemens et al., 2002). All in all, the studies indicate that higher stress tolerance can be achieved by genetic engineering, but that multiple mechanisms to engineer stress tolerance must be utilised for optimal effects. One approach may be to use a single gene encoding a stress-inducible transcription factor to up-regulate simultaneously many genes involved in stress response. This may be possible in the future through the design of specific synthetic transcription factors (Guan et al., 2002).
A completely different approach to avoid yield losses due to abiotic stress is to ensure that seeds develop during the most favourable environmental conditions, while avoiding unfavourable risk factors such as drought and frost. It has been demonstrated that transgenic manipulation of genes that control flowering time is possible (Fobert, 2002). By using chemically-inducible promoters, flowering could be triggered at any point during development by spraying fields with the appropriate inducers. The decision on if and when to spray can be based on environmental conditions and crop performance during the growing season. In certain circumstances, reducing flowering time by as little as a day may have a significant impact on yield. Future aspects and perspective The continued development of legume and oilseed crops remains a great challenge. Most transgenic varieties developed initially carry only a single modified trait, but genetic engineering is now moving to the introduction of multigenic or stacked traits. Such lines can be developed either by multiple or sequential transformation, or by conventional crossing of single trait lines. Plant biotechnology is now at the threshold of an exciting new area in which emphasis is on the manipulation of metabolic pathways. Over-expression of single biosynthetic genes in plants may yield enhanced amounts of the desired metabolites within a pathway or even novel metabolites not normally produced by the plant. Inhibition of the activity of genes for biosynthetic enzymes may be utilised to knock out pathway side-branches or catabolism of a particular secondary metabolite, thus enhancing product yield. Over-expression of regulatory genes of biosynthetic pathways represents an elegant way to manipulate several pathway enzymes more easily. Regulatory genes are also implicated in the responses of a plant to environmental challenges, the control of flowering, seed development, fibre development, partitioning of photo-assimilate into different storage compounds and a multitude of other important metabolic pathways. In order to manipulate plant productivity and product quality effectively, it is critical, therefore, to understand these metabolic networks and their regulation. The value of transgenic technologies as laboratory tools cannot be over-emphasised. In the future, these technologies will become more refined and more predictable, with respect to promoter behaviour, genome integration sites for transgenes, control of downstream pleiotropic effects of novel genes and regulation of transgenes by nuclear matrix attachment regions.
In the longer term, commercial uptake and exploitation of many transgenic lines being developed currently will not take place for many reasons. The majority will serve as laboratory tools to unravel aspects of the function of genes that may be exploited later for crop improvement using conventional breeding. Although transgenesis may represent the only means of achieving some desirable traits, it must be viewed alongside traditional breeding methods as part of a suite of techniques available to modern plant breeding. Besides fundamental considerations of public acceptance and gene product safety, there are other issues that may prevent particular crops or crop products from reaching the marketplace.

272
These include the consequences of gene flow, problems associated with traceability and segregation of food and non-food plants, and methods for maintaining strict control over access to plants producing, for example, pharmaceutical compounds. These are all issues that are being considered and addressed currently in various nations by appropriate regulatory authorities; further discussion of these issues falls outside the scope of this paper. Additionally, commercial viability will weigh against the further development of some transgenic products when extraction/production costs and competitiveness against 'natural' products are considered. Limitations to achieving commercial concentrations of products exist in many cases. The reasons for particular limitations, for example a lack of specific acyltransferases in the case of some fatty acids (Larson et al., 2002), are beginning to be understood as solutions to these and many other problems continue to be sought. Acknowledgements Research on legumes at JIC is supported in part by DEFRA, UK. The JIC is supported by a competitive strategic grant from the Biotechnology and Biological Sciences Research Council, UK. We are grateful to Professor Alison Smith and Dr Matthew Hills, JIC, for their constructive comments on parts of this paper. References Atkins, C.A. 1991. Ammonia assimilation and export of nitrogen from the legume nodule.
In Biology and Biochemistry of Nitrogen Fixation. M. Dilworth & A. Glenn (Eds). Elsevier, Amsterdam. pp. 293-319.
Babaoglu, M., Davey, M.R. & Power, J.B. 2000. Genetic engineering of grain legumes: key transformation events. AgBiotechNet 2, 1-14.
Cahoon, E.B., Ripp, K.G. Hall, S.E. & McGonigle, B. 2002. Transgenic production of epoxy fatty acids by expression of a cytochrome P450 enzyme from Euphorbia lagascae seed. Plant Physiology 128, 615-624.
Cai, D., Wyss, U., Jung, C. & Kleine, M. 2002. Genetic engineering for resistance to nematodes. In Transgenic Plants and Crops. G.G. Khachatourians, A. McHughen, R. Scorza, W.-K. Nip & Y.H. Hui (Eds). Marcel Dekker, New York. pp. 233-248.
Chapple, C. & Carpita, N. 1998. Plant cell walls as targets for biotechnology. Current Opinion in Plant Biology 1, 179-185.
Clemens, S., Palmgren, M.G. & Krämer, U. 2002. A long way ahead: understanding and engineering plant metal accumulation. Trends in Plant Science 7, 309-315.
Coughlan, S.J. & Kinney, A.J. 2002. Transgenic plants as sources of modified oil. In Plant Biotechnology and Transgenic Plants. K.-M. Oksman-Caldentey & W.H. Barz (Eds.). Marcel Dekker, New York. pp. 305-321.
Daniell, H., Streatfield, S.J. & Wycoff, K. 2001. Medical molecular farming: production of antibodies, biopharmaceuticals and edible vaccines in plants. Trends in Plant Science 6, 219-226.
Delmer, D.P. & Haigler, C.H. 2002. The regulation of metabolic flux to cellulose, a major sink for carbon in plants. Metabolic Engineering 4, 22-28.
Demidov, D., Horstmann, C., Meixner, M., Pickardt, T., Saalbach, I., Galili, G. & Müntz, K. 2003. Additive effects of the feed-back insensitive bacterial aspartate kinase and the Brazil nut 2S albumin on the methionine content of transgenic narbon bean (Vicia narbonensis L.). Molecular Breeding 11, 187-201.

273
Dixon, R.A., Lamb, C.J., Masoud, S., Sewalt, V.J.H. & Paiva, N.L. 1996. Metabolic engineering: prospects for crop improvement through the genetic manipulation of phenylpropanoid biosynthesis and defense responses - a review. Gene 179, 61-71.
Drake, P.M.W., Chargelegue, D., Vine, N.D., Dolleweerd, C.J. van, Obregon, P. & Ma, A.K.-C. 2002. Transgenic plants expressing antibodies: a model for phytoremediation. The FASEB Journal 16, 1855-1860.
Falco, S.C., Guida, T., Locke, M., Mauvais, J., Sanders, C., Ward, R.T. & Webber, P. 1995. Transgenic canola and soybean seeds with increased lysine. Biotechnology 13, 577-582.
Fobert, P. 2002. Improving crop performance through transgenic modification of flowering. In Transgenic Plants and Crops. G.G. Khachatourians, A. McHughen, R. Scorza, W.-K. Nip & Y.H. Hui (Eds). Marcel Dekker, New York. pp. 183-196.
Fuchs, M. & Gonsalves, D. 2002. Genetic engineering and resistance to viruses. In Transgenic Plants and Crops. G.G. Khachatourians, A. McHughen, R. Scorza, W.-K. Nip & Y.H. Hui (Eds). Marcel Dekker, New York. pp. 217-231.
Fuentes, S.I., Allen, D.J., Ortiz-Lopez, A. & Hernández, G. 2001. Over-expression of cytosolic glutamine synthase increases photosynthesis and growth at low nitrogen concentrations. Journal of Experimental Botany 52, 1071-1081.
Galili, G. & Höfgen, R. 2002. Metabolic engineering of amino acids and storage proteins in plants. Metabolic Engineering 4, 3-11.
Gatehouse, J.A., Shannon, A.L., Burgess, E.P.J. & Christeller, J.T. 1997. Characterization of major midgut proteinase cDNAs from Helicoverpa armigera larvae and changes in gene expression in response to four proteinase inhibitors in the diet. Insect Biochemistry and Molecular Biology 27, 929-944.
Giddings, G. 2001. Transgenic plants as protein factories. Current Opinion in Biotechnology 12, 450-454.
Guan, X., Ho, T., Beachy, R.N. & C.F. Barbas. 2002. Controlling gene expression in plants using synthetic zinc finger transcription factors. The Plant Journal 32, 1077-1086.
Hagan, N.D., Upadhyaya, N., Tabe, L.M. & Higgins, T.J.V.. 2003. The redistribution of protein sulfur in transgenic rice expressing a gene for a foreign, sulfur-rich protein. The Plant Journal 34, 1-11.
Hancock, R.D. & Viola, R. 2002. Biotechnological approaches for L-ascorbic acid production. Trends in Biotechnology 20, 299-305.
Hasegawa, I., Terada, E., Sunairi, M., Wakita, H., Shinmachi, F., Noguchi, A., Nakajima, M. & Yakazi, J. 1997. Genetic improvement of heavy metal tolerance in plants by transfer of the yeast metallothionein gene (CUP1). Plant and Soil 196, 277-281.
Hegedus, D.D., Gruber, M.Y., Braun, L. & Khachatourians, G.G. 2002. Genetic engineering and resistance to insects. In Transgenic Plants and Crops. G.G. Khachatourians, A. McHughen, R. Scorza, W.-K. Nip & Y.H. Hui (Eds). Marcel Dekker, New York. pp. 249-278.
Hipskind, J.D. & Paiva, N.L. 2000. Constitutive accumulation of a resveratrol-glucoside in transgenic alfalfa increases resistance to Phoma medicaginis. Molecular Plant-Microbe Interactions 13, 551-562.
Huang, Y.-S., Mukerji, P., Das, T. & Knutzon, D.S. 2001. Transgenic production of long-chain polyunsaturated fatty acids. World Review of Nutrition and Dietetics 88, 243-248.
Hughes, E.H. & Shanks, J.V. 2002. Metabolic engineering of plants for alkaloid production. Metabolic Engineering 4, 41-48.
Jeanneau, M., Vidal, J., Gousset-Dupont, A., Lebouteiller, B., Hodges, M., Gerentes, D. & Perez, P. 2002. Manipulating PEPC levels in plants. Journal of Experimental Botany 53, 1837-1845.

274
John, M.E. & Keller, G. 1996. Metabolic pathway engineering in cotton: Biosynthesis of polyhydroxybutyrate in fiber cells. Proceedings of the National Academy of Sciences USA 93, 12768-12773.
Kapusta, J., Modelska, A., Figlerowicz, M., Pniewski, T., Letellier, M., Lisowa, O. Yusibov, V., Koprowski, H., Plucienniczak, A. & Legocki, A.B. 1999. A plant-derived edible vaccine against hepatitis B virus. The FASEB Journal 13, 1796-1799.
Krämer, U. & A.N. Chardonnens, A.N. 2001. The use of transgenic plants in the bioremediation of soils contaminated with trace elements. Applied Microbiology and Biotechnology 55, 661-672.
Kuchuk, N., Griga, M., Kusturkova, G. & Ilieva-Stoilova, M. 2001. Biotechnology. In Carbohydrates in Grain Legume Seeds. C.L. Hedley (Ed.). CAB International, Wallingford. pp. 145-226.
Lardizabal, K.D., Metz, J.G., Sakamoto, T., Hutton, W.C., Pollard, M.R. & Lassner, M.W. 2000. Purification of a jojoba embryo wax synthase, cloning of its cDNA, and production of high levels of wax in seeds of transgenic Arabidopsis. Plant Physiology 122, 645-655.
Larrick, J.W. & Thomas, D.W. 2001. Producing proteins in transgenic plants and animals. Current Opinion in Biotechnology 12, 411-418.
Larson, T.R., Edgell, T., Byrne, J., Dehesh, K. & Graham, I.A. 2002. Acyl CoA profiles of transgenic plants that accumulate medium-chain fatty acids indicate inefficient storage lipid synthesis in developing oilseeds. The Plant Journal 32, 519-527.
Lee, R.W.H., Strommer, J., Hodgins, D., Shewen, P.E., Niu, Y. & Lo, R.Y.C. 2001. Towards development of an edible vaccine against bovine pneumonic pasteurellosis using transgenic white clover expressing a Mannheimia haemolytica A1 leukotoxin 50 fusion protein. Infection and Immunity 69, 5786-5793.
Liu, Q., Singh, S.P. & A.G. Green, A.G. 2002. High-stearic and high-oleic cottonseed oils produced by hairpin RNA-mediated post-transcriptional gene silencing. Plant Physiology 129, 1732-1743.
Lucca, P., Hurrell, R. & Potrykus, I. 2001. Genetic engineering approaches to improve the bioavailability and the level of iron in rice grains. Theoretical and Applied Genetics 102, 392-397.
Mejáre, M. & Bülow, L. 2001. Metal-binding proteins and peptides in bioremediation and phytomediation of heavy metals. Trends in Biotechnology 19, 67-73.
Morton, R.L., Schroeder, H.E., Bateman, K.S., Chrispeels, M.J., Armstrong, E. & Higgins, T.J.V. 2000. Bean α-amylase inhibitor 1 in transgenic peas (Pisum sativum) provides complete protection from pea weevil (Bruchus pisorum) under field conditions. Proceedings of the National Academy of Sciences USA 97, 3820-3825.
Molvig, L., Tabe, L., Eggum, B.O., Moore, A., Craig, S., Spencer, D. & Higgins T.J.V. 1997. Enhanced methionine levels and increased nutritive value of seeds of transgenic lupins (Lupinus angustifolius L.) expressing a sunflower seed albumin gene. Proceedings of the National Academy of Sciences USA 94, 8393-8398.
Müntz, K., Christov, V., Saalbach, G.,. Saalbach, I., Waddell, D., Pickardt, T., Schieder, O. & Wüstenhagen, T. 1998. Genetic engineering for high methionine grain legumes. Nahrung 42, 125-127.
Murdock, L.L. & Shade, R.E. 2002. Lectins and protease inhibitors as plant defenses against insects. Journal of Agricultural and Food Chemistry 50, 6605-6611.
Palmer, C.E. & Keller, W.A.. 2002. Transgenic oilseed brassicas. In Transgenic Plants and Crops. G.G. Khachatourians, A. McHughen, R. Scorza, W.-K. Nip & Y.H. Hui (Eds). Marcel Dekker, New York. pp. 773-792.

275
Pan, A., Tie, F., Duau, Z., Yang, M., Wang, Z., Li, L., Chen, Z. & Ru, B. 1994. α-domain of human metallothionein IA can bind to metals in transgenic tobacco plants. Molecular and General Genetics 242, 666-674.
Parmenter, D.L., Boothe, J.G., Rooijen, G.J. van, Yeung, E.C. & Moloney, M.M. 1995. Production of biologically active hirudin in plant seeds using oleosin partitioning. Plant Molecular Biology 29, 1167-1180.
Poirier, Y. 2002. Polyhydroxyalkanoate synthesis in plants as a tool for biotechnology and basic studies of lipid metabolism. Progress in Lipid Research 41, 131-155.
Potrykus, I. 2001. Golden rice and beyond. Plant Physiology 125, 1157-1161. Raboy, V. 2001. Seeds for a better future: 'low phytate' grains help to overcome malnutrition
and reduce pollution. Trends in Plant Science 6, 458-462. Richardson, A.E., Hadobas, P.A. & Hayes, J.E. 2001. Extracellular secretion of Aspergillus
phytase from Arabidopsis roots enables plants to obtain phosphorus from phytate. The Plant Journal 25, 641-649.
Rontein, D., Basset, G. & Hanson, A.D. 2002. Metabolic engineering of osmoprotectant accumulation in plants. Metabolic Engineering 4, 49-56.
Samis K., Bowley, S. & McKersie, B. 2002. Pyramiding Mn-superoxide dismutase transgenes to improve persistence and biomass production in alfalfa. Journal of Experimental Botany 53, 1343-1350.
Schulman, A. 2002. Transgenic plants as producers of modified starch and other carbohydrates. In Plant Biotechnology and Transgenic Plants. K.-M. Oksman-Caldentey & W.H. Barz (Eds). Marcel Dekker, New York. pp. 255-282.
Sun, W., Montague, M. van, & N. Verbruggen, N. 2002. Small heat shock proteins and stress tolerance in plants. Biochimica et Biophysica Acta 1577, 1-9.
Tabe, L.M., Higgins, C.M., McNabb, W.C. & Higgins, T.J.V. 1993. Genetic engineering of grains and pasture legumes for improved nutritive value. Genetica 90, 181-200.
Tenhagen, R. 2002. Transgenic plants with enhanced tolerance against microbial pathogens. In Plant Biotechnology and Transgenic Plants. K.-M. Oksman-Caldentey & W.H. Barz (Eds). Marcel Dekker, New York. pp. 549-569.
Thelen, J.J. & Ohlrogge, J.B. 2002. Metabolic engineering of fatty acid biosynthesis in plants. Metabolic Engineering 4, 12-21.
Thomas, C., Sue, Y., Naus, K., Lloyd, A. & Roux, S. 1999. Apyrase functions in plant phosphate nutrition and mobilizes phosphate from extracelullar ATP. Plant Physiology 119, 543-55.
Ursin,V., Knutzon, D., Radke, S., Thornton, J. & Knauf, V. 2000. Production of beneficial dietary omega-3 and omega-6 fatty acids in transgenic canola. In Abstracts 14th
International Symposium on Plant Lipids, 13. Van Breusegem, F. & Inzé, D. 2002. Transgenic plants expressing tolerance toward
oxidative stress. In Plant Biotechnology and Transgenic Plants. K.-M. Oksman-Caldentey & W.H. Barz (Eds). Marcel Dekker, New York. pp. 497-516.
Verporte, R. & Memelink, J. 2002. Engineering secondary metabolite production in plants. Current Opinion in Biotechnology 13, 181-187.
Wang, T.L., Domoney, C., Hedley, C.L. Casey, R. & Grusak, M.A. 2003. Can we improve the nutritional quality of legume seeds? Plant Physiology 131, 886-891.
Weber, H., Rolletschek, H., Heim, U., Golombek, S., Gubatz. S. & U. Wobus. 2000. Antisense-inhibition of ADP-glucose pyrophosphorylase in developing seeds of Vicia narbonensis moderately decreases starch but increases protein content and affects seed maturation. The Plant Journal 24, 33-43.

276
Welham, T. & Domoney, C. 2000. Temporal and spatial activity of a promoter from a pea enzyme inhibitor gene and its exploitation for seed quality improvement. Plant Science 159, 289-299.
Wigdorovitz, A., Carrilho, C., Dus Santos, M.J., Trono, K., Peralta, A., Gómez, M.C, Rios, R.D., Franzone, P.M., Sadir, A.M., Escribano, J.M. & Borca, M.V. 1999. Induction of a protective antibody response to foot and mouth disease virus in mice following oral or parenteral immunization with alfalfa transgenic plants expressing the viral structural protein VP1. Virology 255, 247-353.
Xie, D.-Y., Sharma, S.B., Paiva, N.L. Ferreira, D. & Dixon, R.A. 2003. Role of anthocyanidin reductase, encoded by BANYULS in plant flavonoid biosynthesis. Science 299, 396-399.
Yan, L. & Kerr, P.S. 2002. Genetically engineered crops: their potential use for improvement of human nutrition. Nutrition Reviews 60, 135-141.

277
α-Galactosides in Catalan landraces of beans (Phaseolus vulgaris L.) and location, candidates for protected designation of origin L. Bosch1, F. Casañas1, M. Pujolà1, E. Centelles1, P. Beltran1, A. Florez1 & F. Nuez2 1ESAB, Universidad Politécnica de Catalunya, Urgell, 187, 08036 Barcelona, Spain 2COMAV, Universidad Politécnica de Valencia, Camino de Vera 14, 46022 Valencia, Spain Summary Traditional landraces of dry beans (Phaseolus vulgaris) are still cultivated in many places in Europe due to their organoleptic quality. These landraces, which are generally of regional interest, deserve attention from plant breeders because of their social and economic importance. Three landraces, which are still cultivated in Catalonia were assessed for raffinose and stachyose content (both accounting for flatulence) after being cultivated in three locations that are candidates for Protected Designation of Origin. The location effect was significant (P < 0.05) for both stachyose and raffinose, while the variety effect only reached the same level of significance for raffinose. Environmental correlations were greater than genetic ones, either between stachyose and raffinose or between these two traits and other chemical compounds of the seed. From a practical point of view (flatulence production), the differences found in raffinose and stachyose content, although significant, does not seem large enough to justify the inclusion of these two compounds in the definition of Catalan Protected Designations of Origin. Keywords: Phaseolus vulgaris, landraces, α-galactosides, genetic correlations, environmental
correlations Introduction Breeding of commercial varieties of dry beans (Phaseolus vulgaris L.) has focused on yield and nutritive value. Little attention has been paid to organoleptic quality. In Spain, beans are a minor crop, but some traditional landraces are still cultivated due to their gastronomic prestige, sometimes associated with a typical culinary speciality. Such landraces have a poor yield performance and other agronomic traits but the high price they obtain, in local markets, makes their cultivation worthwhile.
To protect valuable landraces associated with certain regions and local dishes, the European Community promotes the Protected Designation of Origin (PDO). To contribute to this task regarding traditional beans of Catalonia, our laboratory has launched a program to ascertain the chemical basis of the gastronomic differences and to determine the effect of genotype and environment on the final product.
In addition to gastronomic traits we are also monitored some chemical compounds with undesirable side effects or antinutritional properties. Among others, we checked the α-galactosides of sucrose, which are partially responsible for flatulence (Alles et al., 1996). Significant differences in raffinose and stachyose content have been found among varieties (Drum et al., 1990; Muzquiz et al., 1999) and locations (Muzquiz et al., 1999). Thus, information on these compounds might be useful for the definition of PDOs. The objectives of this experiment were i) to describe the genetic and environmental variation in raffinose and stachyose in Catalan landraces, cultivated in areas that are candidates for PDO, and ii) to evaluate whether these traits would be of use in the definition of PDOs.

278
Materials and methods Field trials included the Catalan landraces Castellfollit del Boix, Genoll de Crist, and Montcau (pure line derived from the Ganxet landrace), and the checks Canela (from Vegas Bañezanas Company, La Bañeza, Leon, Spain) and Navy (from B & W Co-Op. Inc., Brekenridge, Michigan, USA).
The experiment was conducted in Santa Pau, with a volcanic soil; Castellar del Vallès, with soil originating from limestone decomposition; and El Prat de Llobregat, with alluvial soil (Table 1). All three locations are in Catalonia (NE Spain) and have a mild Mediterranean climate. A complete randomized block design (2 blocks per location) was used, where each plot consisted of two rows of 80 competitive plants. The distance between plants in a row was 50 cm and between rows 75 cm (greater than usual in order to avoid differences in competition among varieties). Irrigation was provided when necessary in Castellar and El Prat. The Santa Pau location was not irrigated, as this is the traditional way of cultivation. Table 1. Edaphic characteristics of the three locations. Santa Pau Castellar El Prat Soil pH 7.6 8.1 8.2 Oxidizable organic-matter (%) 4.4 2.8 2.2 Nitric nitrogen (mg N-NO3/kg) 43 15 53 Phosphorus (Olsen) (ppm) 55 97 50 Potassium (ppm) > 600 451 356 Magnesium (ppm) 466 332 306 Calcium carbonate equivalent (%) < 4 18 30 Calcium (ppm) 4,703 8,454 7,325 Cation exchange capacity (Meq/100 g) 26.3 13.3 7.2 Textural class (USDA) Sandy-loam Clay-sandy-
loam Loam
Raffinose and stachyose content were determined simultaneously as their trimethylsilyl
derivatives by gas chromatography. Verbascose was not assessed, but its content in beans, on average, is only 5 % of the total α-galactosides of sucrose (Muzquiz et al., 1999).
The traits were analysed using the linear equation:
Xijkl = µ + Gi + Lj + Bk(j) + GLij + ξl(ijk) where, Gi = population effect; Lj = location effect; Bk = block effect within location and ξl (ijk) = the residual effect (plot within population, block and location).
Data were recorded on a per plot basis. Calculations used the GLM procedure of the SAS statistical package (SAS Institute, 1985). Genetic and environmental correlations of raffinose and stachyose with other traits (Casañas et al., 2003) were calculated using the mean value of variety and trait, and the mean value of location and trait, respectively. Results and discussion The location effect was significant for stachyose (P < 0.02) (Table 2). The Santa Pau trial had the lowest value at 10 % lower (P < 0.05) than El Prat (the highest value). There was no significant difference in this trait for variety (Table 2) or for the variety × location interaction.

279
Raffinose, which accounted for less than 10 % of the total α-galactosides in our analysis (Table 2), had highly significant differences (P < 0.001) for location, variety, and the location × variety interaction. The ranking for raffinose content (significant, P < 0.05) was Santa Pau > Castellar ≈ El Prat, for location and Castellfollit del Boix ≈ Navy > Montcau > Genoll de Crist ≈ Canela, for variety.
Stachyose plus raffinose content (Table 2) was not significant for any effect (location, variety, or location x variety) because, in general, the higher the value was for stachyose, the lower it was for raffinose; thus, the two compensated for each other. Table 2. Variety and location means (g/kg) and least significant difference (LSD) (P <
0.05) of raffinose (Raf) and stachyose (Sta) and their total (Raf + Sta). Variety Raf Sta Raf +
Sta Location Raf Sta Raf + Sta
Genoll de Crist
0.84 23.41 24.25 Santa Pau
1.88 22.33 24.21
Montcau 1.81 22.70 24.51 Castellar 1.60 23.65 25.25 Castellfollit del Boix
2.39 23.30 25.69 El Prat 1.52 24.92 26.44
Canela 0.81 25.62 26.43 LSD 0.23 2.57 ns Navy 2.48 23.17 25.63 LSD 0.30 ns ns
The environmental correlation between stachyose and raffinose was -0.96 (P < 0.19). The corresponding genetic correlation was lower (-0.61) (Table 3). Similarly, stachyose and raffinose contents were more environmentally than genetically correlated with other chemical seed traits. Genetic correlations were only significant between stachyose and glucose, while environmental correlations of stachyose were significant with protein, glucose and citric acid (Table 3). Generally, the higher the correlation of a given trait with stachyose was, the lower was the corresponding one with raffinose. While this was true for the genetic correlations, it was especially evident for the environmental ones. The correlations of total α-galactosides (raffinose + stachyose) with other seed compounds were similar to the ones for stachyose, because of its predominant proportion of the total.
Cation exchange capacity was strongly correlated with α-galactosides, positively with raffinose and negatively with stachyose. As a high soil cation exchange capacity generally means less plant stress and α-galactosides are considered to have an anti-stress effect, this correlation makes sense. Other correlations with soil characteristics are more difficult to explain from a physiological point of view. A random association between cation exchange capacity and some chemical elements could account for these values.
Although the results show significant differences in some effects and traits, the magnitude of the differences does not seem enough to justify the use of these data (as the difference of flatulence induced) in the definition of Catalan PDOs.

280
Table 3. Genotypic and environmental correlations of raffinose and stachyose, with seed traits and soil characteristics of locations.
Raffinose Stachyose Raffi + Stachy Genetic Environ. Genetic Environ. Genetic Environ. Seed trait
Stachyose -0.61 -0.96 - - - - Malic acid -0.72 -0.80 -0.05 0.94 -0.70 0.96 Citric acid -0.33 -0.88 0.08 0.98* -0.19 0.99** Glucose 0.57 -0.92 -0.92*** 0.99** -0.65 1.00***
Lactose -0.34 0.62 0.65 -0.36 0.53 -0.32 Sucrose -0.78 -0.91 0.01 0.75 -0.68 0.72 Protein 0.41 0.98* -0.37 -0.99** -0.10 -0.99**
Soil characteristics Phosphorus -0.21 -0.09 -0.14 Potassium 0.98* -0.99** -0.99** Magnesium 1.00*** -0.94 -0.92 Calcium carbonate -0.96 1.00*** 1.00*** Calcium -0.87 0.69 0.65 Cation exchange capacity 0.99** -0.98* -0.97
Probabilities used. *P = 0.15; **P = 0.10; ***P = 0.05. References Alles, M.S., Pierson, M.D., Sathe, S.K. & Salunke, D.K. 1996. Fate of
fructooligosaccharides in the human intestine. British Journal of Nutrition 76, 211-221. Casañas, F., Bosch,L., Pujolà, M., Centelles, E., Gual, J., Florez, A., Beltràn, P. & Nuez, F.
2003. Hacia la descripción objetiva de denominaciones de origen para judía (Phaseolus vulgaris L.) en Cataluña. In A.M. de Ron (Ed.). Proceedings IIIer Seminario de Judía de la Península Ibérica, Lorenza, Spain, October 2003, 41-46.
Drumm, T.D., Gray, J.I. & Hosfield, G.L. 1990. Variability in the saccharide, protein, phenolic acid and saponin content of four market classes of edible dry beans. Journal of the Science of Food and Agriculture 51, 285-289.
Muzquiz, M., Burbano, C., Ayet, G., Pedrosa, M.M. & Cuadrado, C. 1999. The investigation of antinutritional factors in Phaseolus vulgaris. Environmental and varietal differences. Biotechnology, Agronomy Society and Environment 3, 210-216.
SAS Institute, 1985. SAS User's Guide: Statistics. SAS Institute, Cary, Nc.

281
Facile breeding markers to lower contents of vicine and convicine in faba bean seeds and trypsin inhibitors in pea seeds G. Duc1, P. Marget1, D. Page2 & C. Domoney3 1INRA, Unité de Génétique et Ecophysiologie des Légumineuses, URLEG, BP 86510, 21065 Dijon Cedex, France 2INRA, UMR A408, Qualité et Sécurité des Produits d'Origine Végétale, Domaine St-Paul, Site Agroparc, 84914 Avignon Cedex 9, France 3Department of Metabolic Biology, John Innes Centre, Colney, NR4 7UH, United Kingdom Summary Despite their low seed contents, vicine and convicine in faba bean (Vicia faba) and protease inhibitors in pea (Pisum sativum) are important feed constituents because of their negative effect on pig and poultry performance. The chemical determination of these compounds is time-consuming and expensive. We have developed simple markers to boost progress in breeding. In faba bean, the gene controlling hilum colour maps a small distance from the gene determining the seed vicine and convicine content and this phenotypic trait can therefore be used as a marker. In pea, gene polymorphism linked to variation in seed trypsin inhibitor activity (TIA) has allowed the definition of primers for polymerase chain reaction (PCR) assays, which can also be exploited in breeding programs. In both cases, these markers offer an efficient and cheap pre-screening procedure. Keywords: Vicia faba L., Pisum sativum L., marker-assisted selection, trypsin inhibitors,
vicine-convicine Introduction The development of simple markers is very useful to boost progress in breeding. Indeed, the easier the screening, the more diverse and numerous the crosses that can be manipulated by breeders. Tannins, protease inhibitors, vicine and convicine are legume seed components, which have antinutritional properties for pigs and poultry (Grosjean et al., 1999; Hedeman et al., 1999; Grosjean et al., 2000). Further, vicine and convicine cause favism (i.e. a haemolytic crisis) in some humans (Arese & de Flora, 1990). Tannin reduction in pea and faba bean seed, by breeding, is simplified by the fact that the genes involved are recessive genes which have a clear-cut pleiotropic effect on flower colour. Selecting for a white flower colour allows rapid screening for low tannin in breeding programs.
The situation is more complex for removal, by breeding, of other antinutritional factors (ANFs), such as vicine and convicine in faba bean and protease inhibitors in pea. Even though there are simplified chemical methods for both sets of compounds (Sixdenier et al., 1996; Page et al., 2000), these are still expensive for the screening large numbers of genotypes. Vicine and convicine are thermostable and can only be reduced by soaking seeds in water or a weak acid solution (Marquardt et al., 1983). This supports a breeding strategy to eliminate them from seed. Duc et al, (1989) reported a spontaneous mutation (vc-), which lowers the vicine + convicine (V+C) contents 10 to 20-fold and results in improved performance of laying hens (Grosjean et al., 2000). Similarly, expensive thermal treatments can reduce TIA in pea seed, but it has been shown that natural genetic variability in TIA can

282
be exploited for improved animal performance (Wiseman et al., 2003). These reasons motivated a search for facile markers to assist in plant breeding. Materials and methods Vicia faba genotype 1268 (Duc et al., 1989), an F4 line homozygous for both the vc- and the white hilum gene, was crossed with three lines, BPL938, BPL261 and L8-31811 (provided by M. Saxena, ICARDA, Syria) which had a black hilum and a high V + C content. The F1, F2 and F3 seeds were produced under cages to protect plants from insect pollination. Inheritance of low V + C is additive, with maternal determinism (Duc et al., 1989). Inheritance of the white hilum is recessive to the coloured hilum and the effect of this gene on seed phenotype is also maternal (Sirks, 1931). A semi-quantitative technique (Sixdenier et al., 1996) was used to characterize the F3 seeds from each F2 plant and defined three classes of V + C content: low (L) (0 - 0.1% of seed DM), intermediate (I) (0.2 - 0.6 % of seed DM) and high (H) (0.6 - 1.2 % of seed DM). Although L and I were not overlapping classes, in some crosses I and H overlapped in the 0.5 - 0.7 % range. In the I-H overlap situation, 25 % of the plants with higher V + C content were considered to be in the H group. Black (BH) or white (WH) seed hilum colour was recorded visually on F3 seed.
Page et al. (2002) described the development of a set of diagnostic PCR primers, based on sequence polymorphism in promoter and coding regions of pea TI genes. A total of 168 recombinant inbred lines (F7 RILs provided by I. Lejeune, INRA, Mons, France) derived from a cross between the cultivars Térèse (low TIA) and Champagne (high TIA) were screened, using this set of PCR primers as described by Page et al. (2002) and using a rapid biochemical method to determine TIA values (Page et al., 2000). Results and discussion Hilum colour as a marker for vicine and convicine content in faba bean seeds In this study, we identified and quantified a linkage between vc- and hilum colour gene, using several crosses involving the genotype 1268. Over all crosses, 481 and 1,469 plants were in the L and I + H classes for V + C content. This fits a Mendelian (1:3) segregation (χ2 = 0.11, P = 1.00) for the vc- gene, as reported by Duc et al. (1989). Over all crosses, 470 and 1,480 plants were in the white and coloured hilum classes, respectively.
This also fits a Mendelian (1:3) segregation (χ2 = 0.848, P = 0.32) for hilum colour, as reported by Sirks (1931). If the two genes segregated independently, a 1:3 ratio of white:black hilum would have been observed in L, I or H groups. This was not the case (Table 1); the L class showed a predominance of WH F2 plants, whereas the H class showed a low frequency of WH F2 plants. This indicates a close linkage between the two genes. A distance of 10.1 centimorgans was calculated for the two genes. This result suggests this facile marker is of value in pre-screening progenies for their V + C content where parental lines differ in their hilum colour. Such pre-screening could reduce by 75 % the number of chemical determinations required to identify alleles. However, due to recombinant genotypes, chemical analysis will still be necessary on lines, which are kept.

283
Table 1. Segregation of white hilum colour (WH) in F2 faba bean plants within low (L) and high (H) classes of vicine + convicine (V + C) content.
Cross V+C segregants
Number of F2 plants
Number of WH plants
χ2 value for (1 WH:3 BH)
Probability in χ2 test
L 96 78 159 P < 0.01 1268 x BPL261 H 106 4 26 P < 0.01
L 101 83 176 P < 0.01 BPL261 x 1268 H 98 5 21 P < 0.01
L 83 70 156 P < 0.01 1268 x BPL938 H 83 3 20 P < 0.01
L 97 78 159 P < 0.01 BPL938 x 1268 H 94 2 26 P < 0.01
L 104 80 150 P < 0.01 1268 x L8-31811 H 96 5 20 P < 0.01
Figure 1 clearly shows that, among the population of RILs derived from parents having high or low TIA, there is no case of a line with the 646 bp band having a high TIA. This provides a strong marker, directly located to TI genes and based on a simple PCR reaction. This promises to be of enormous value to breeders, and especially so if DNA extracted from populations can be screened simultaneously for additional markers.
Figure 1. Summary of PCR results for a population of 168 recombinant inbred lines from the pea cross Térèse (low TIA) x Champagne (high TIA). Two PCR bands were obtained (ordinate) of 646 and 821 bp. The TIA of each line is indicated on the abscissa; the higher the percentage, the higher the corresponding TIA.
Breeding perspectives The strategy of marker-assisted breeding is valuable because it can save a lot of the cost of phenotyping and, in some cases, can reduce errors due to environmental effects on gene expression. We have illustrated here, with two cases, how simplified tools can be provided to breeders that can be used to reduce the ANF contents of legume seeds. The present rapid development of finer genetic maps, with new types of markers and the development of robotics for genomics, should make molecular approaches to plant breeding even more efficient in the future. Selection may then be based on markers that are very close to the character (Gutierrez et al., 2004) Further, numerous seed compounds could be screened for simultaneously.
0% 10% 20% 30% 40% 50% 60% 70% 80% 90%
Trypsin inhibitor activity (% of control)
Size of PCR products (bp)
821
646

284
Acknowledgements UNIP (France) and DEFRA (UK) for financial support for the work on pea trypsin inhibitors and Isabelle Lejeune (INRA, Mons, France) for providing the population of pea recombinant inbred lines. References Arese, P. & Flora, A. de, 1990. Pathophysiology of hemolysis in glucose-6-phosphate
dehydrogenase deficiency. Seminars in Hematology 27, 1-40. Domoney, C., Welham, T., Ellis, N. & Hellens, R. 1994. Inheritance of qualitative and
quantitative trypsin inhibitor variants in Pisum. Theoretical and Applied Genetics 89, 387-391.
Domoney, C., Welham, T., Sidebottom, C. & Firmin, J. 1995. Multiple isoforms of Pisum trypsin inhibitors result from modification of two primary gene products. FEBS Letters 360, 15-20.
Duc, G., Sixdenier, G., Lila, M. & Furstoss, V. 1989. Search for genetic variability for vicine and convicine content in Vicia faba L. A first report of a gene which codes for nearly zero-vicine and zero-convicine contents. In Recent Advances of Research in Antinutritional Factors in Legumes Seeds. J. Huisman, T.F.B. van der Poel & I.E. Liener (Eds). Proceedings 1st International Workshop on Antinutritional Factors in Legume Seeds, Wageningen, 22-25 November 1988, 305-313.
Grosjean, F., Barrier-Guillot, B., Bastianelli, D., Rudeaux, F., Bourdillon, A. & Peyronnet, C. 1999. Feeding value of three categories of pea (Pisum sativum L.) for poultry. Animal Science 69, 591-599.
Grosjean, F., Bourdillon, A., Rudeaux, F., Bastianelli, C., Duc, G. & Lacassagne, L. 2000. Valeur alimentaire pour la volaille de féveroles isogéniques (Vicia faba L.) avec ou sans tannins et avec ou sans vicine. Sciences et Techniques Avicoles 32, 17-23.
Gutierrez, N., Duc, G., Marget, P., Avila, C., Cubero, J.I., Moreno, M.T. & Torres, A.M. 2004. Identification of molecular markers tightly linked to low tannins and vicine-convicine content in faba beans. (These Proceedings).
Hedeman, M.S., Welham, T., Boisen, S., Canibe, N., Bilham, L. & Domoney, C. 1999. Studies on the biological responses of rats to seed trypsin inhibitors using near-isogenic lines of Pisum sativum L. Journal of the Science of Food and Agriculture 79, 1647-1653.
Marquardt, R.R., Muduuli, D.S. & Frohlich, A., 1983. Purification and some properties of vicine and convicine isolated from faba bean (Vicia faba L.) protein concentrate. Journal of Agricultural Science and Food Chemistry 31, 839-844
Page, D., Aubert, G., Duc, G., Welham, T. & Domoney, C. 2002. Combinatorial variation in coding and promoter sequences of genes at the Tri locus in Pisum sativum accounts for variation in trypsin inhibitor activity in seeds. Molecular Genetics and Genomics 267, 359-369.
Page, D., Quillien, L. & Duc, G. 2000. Trypsin inhibitory activity measurement: simplifications of the standard procedure used for pea seeds. Crop Science 40, 1482-1485.
Sirks, M.J. 1931. Beitrage zu einer genotypischen analyse der ackerbohne Vicia faba L. Genetika 13, 210-631
Sixdenier, G., Cassecuelle, F., Guillaumin, L. & Duc, G. 1996. Rapid spectrophotometric method for reduction of vicine and convicine in faba bean seed. FABIS Newsletters 38/39, 42-43.

285
Wiseman, J., Al-Mazorqi, W., Welham, T. & Domoney, C. 2003. The apparent ileal digestibility, determined with young broilers, of amino acids in near-isogenic lines of peas (Pisum sativum L) differing in trypsin inhibitor activity. Journal of the Science of Food and Agriculture 83, 644-651.


287
Identification of molecular markers tightly linked to low tannin and vicine-convicine content in faba beans N. Gutierrez1, G. Duc3, P. Marget3, C.M. Avila2, M.J. Suso1, J.I. Cubero4, M.T. Moreno2 & A.M. Torres2 1Instituto de Agricultura Sostenible-CSIC, Apdo.4084, 14080 Córdoba, Spain 2CIFA-Alameda del Obispo, Departamento Mejora y Agronomía, Apdo. 3092, 14080 Córdoba, Spain 3INRA, Unité de Génétique et Ecophysiologie des Légumineuses URLEG, BP 86510, 21065 Dijon Cedex, France 4Departamento Genética, ETSIAM, Aptdo. 3048, 14080 Córdoba, Spain Summary The antinutritional factors (ANFs) present in the seed of Vicia spp. seeds are perceived to be major constraints to the wider utilisation of these promising crops as grain legumes. In case of Vicia faba L., breeding priorities are the absence polyphenols, broadly referred to as tannins, which diminish the biological value of the protein, and vicine-convicine responsible for favism in humans and for low egg production in laying hens. Tannin can be removed from seed by one of the two recessive genes (zt-1 and zt-2) associated with white flowers. The vicine-convicine content can be reduced 20 fold by the zvc gene linked to white hilum colour. The aim of this study was to identify markers linked to a low content of both tannin and vicine-convicine. Bulked Segregant Analysis (BSA) was used to identify RAPD markers linked to these genes. Qualitative quantification of vicine-convicine was by spectrophotometry of the parental lines and the F2 population. Two contrasting bulks were prepared per trait. Each contained equal amounts of DNA from homozygous F2 individuals (white vs coloured flower and high vs low vicine-convicine content). The RAPD markers tightly linked to the genes of interest will be sequenced to develop specific Sequence Characterised Amplified Regions (SCARs) that will aid selection in different breeding programs. Keywords: Vicia faba, molecular markers, tannins, vicine-convicine, nutrition Introduction Faba bean is one of the oldest crops grown by man and provides an excellent high-protein feed for human and animal nutrition. The protein level in faba bean seed is of about 200 g/kg. However, besides protein, legume seeds also contain ANFs, which negatively affect their digestibility. A recent increase in the demand for plant protein has emerged, partly because of Bovine Spongiform Encephalopathy (BSE) attributed to the use of animal-based cattle feed. The ban on the use of animal meals has increased the European deficit of protein rich material. At present legumes are only grown on 5 % of the agricultural land area in Europe compared with 20 - 30 % in North and South America, Australia and Asia. To increase interest in this legume in Western Europe the crop should be attractive both to the producers and to the users through the development of ANF free genotypes.
Priorities for breeding faba bean are an absence of polyphenols, broadly referred to as tannins and, vicine-convicine responsible for favism in humans and for low egg production in laying hens. The antinutritional effects of tannins are due to their affinity for certain

288
nutrients, especially proteins (Kuman & Singh, 1984) that inhibit digestive enzyme activity (Bartolomé et al., 1994). The zero-tannin character, which is monogenetically inherited in faba bean, is valuable in plant breeding to increase protein digestibility in monogastric animals. Two genes control the absence of tannins in faba beans, zt-1 and zt-2. They also determine the white flower character in the plant (Picard, 1976). The less common gene, zt-2, is associated with increased protein and energy levels and reduced fibre content. The seed has a superior feeding value (Duc et al., 1999; Grosjean et al., 2000). Tannin can be removed from seed by one of the two recessive genes. However, crosses between them produce coloured F1 plants with tannins that will continue to contaminate the crop. For this reason, it is important to identify the gene present in all tannin-free cultivars, and gene bank accessions, to enable plant breeders to chose suitable lines for their crosses (Crofton et al., 2000).
Two pyrimidine glucosides (vicine-convicine), which accumulate in the cotyledons have been linked with the aetiology of favism (Mager et al., 1965). This is an acute form of haemolytic anaemia associated with consumption of faba beans by humans carrying a genetic deficiency in of glucose-6-phosphate dehydrogenase in their red bloods cells. The glucosides also reduce egg production in laying hens (Marquardt, 1989). The discovery of a spontaneous mutant (zvc) that reduces 20 fold the seed vicine-convicine content raised the possibility of their removal from the seed (Duc et al., 1989). However, the cost, and difficulty, of chemical determination of these products seriously restricts advances in breeding programs.
Molecular markers tightly linked to desired genes have proved to be a valuable tool to increase the speed of detecting genotypes of interest thus saving time and resources. The aim of this work was the identification of markers, linked to low seed tannin and vicine-convicine content. These use of markers should speed up the production of new cultivars free of ANFs for human and animal consumption. Materials and methods Plant material Three F2 populations between an asynaptic mutant (Vf6) and lines with zero tannin genes (zt-1 y zt-2) and with the zero vicine-convicine mutant (zvc = line1268) were analysed (n = 88, 90 and 136, respectively). Since both traits are monogenic traits, BSA was used to identify RAPD markers linked to these genes (Michelmore et al., 1991). Homozygosity of F2 plants for white flower and hilum colour was determined in their corresponding F2 and F3 families. Qualitative quantification of vicine-convicine in the parental lines and the F2 population was by spectrophotometry. Based on these results, two contrasting bulks per trait were prepared, each containing equal amount of DNA from 7 to 10 homozygous F2 individuals. RAPD analysis Amplifications were as described by Williams et al. (1990) and Welsh & McClelland (1990) with slight modifications (Torres et al. 1993). A total of 700 RAPD primers from Operon Technologies (Alameda, California, USA) were surveyed. Primers generating marker polymorphisms between the bulks, are being tested individually on DNA from the homozygous F2 plants used in the bulk. When polymorphism is maintained, the primers will be screened on the F2 population for linkage analysis.

289
Reference method The reference method (Sixdenier et al., 1996) was HPLC adapted according to Quemener (1998). The HPLC was optimised for the best separation of vicine and convicine at 276 nm. Seeds were soaked for 3.5 h in 30 ml water in a 90 ºC water-bath. Normal HCl (100 µl) was mixed with 10 ml of the resulting solution centrifuged at 13,000 rpm for 15 min and then filtered on a Nalgene SFCA filter. The resulting solution was diluted with water (1:5 v/v) prior to HPLC injection. The chromatographic conditions were Licrospher 125-4, 100 RP-18 (5 m) column with precolumn, MILLI-Q water 1 ml/min as eluant, 276 nm wavelength of absorbance measurement. Standards of pure vicine and convicine were obtained from Dr Maquardt. Results and discussion Tannin content Segregation for white vs coloured flowers in the F2 plants from crosses Vf6 x zt-1 and Vf6 x zt-2 gave a good fit to a 3:1 ratio (χ2
0.05 = 0.24 and 0.91, respectively), further confirming the
monogenic control of the trait. The result was also verified in the corresponding F3 families based on a satisfactory fit to a 1:2:1 ratio. Thirty nine polymorphic markers were initially identified in the pools and are being further tested for consistent detection and subsequent analysis on the individuals that formed each bulk. Vicine-convicine content Fifteen seeds of both parental lines (Vf6 and 1268), as well as 15 from each 136 F3 families from the cross were analysed. The line Vf6 had a high vicine and convicine content (0.33 % and 0.21 %, respectively). In line 1268 the corresponding values were markedly lower (0.02 % and 0.01 %). On the scale used, the total vicine-convicine content in F3 individual plants ranged from 0.25 % to 0.02 %. Two contrasting DNA pools were used to identify polymorphic markers. Forty eight RAPD markers were present in only one of the bulks but, so far only 4 of them have displayed consistent detection in each of the individuals of the bulk.
After this all of the selected markers will be screened on the corresponding F2 individuals. The co-segregation data between RAPD markers and traits will be used to construct a linkage map. Finally, RAPD markers, which are tightly linked to the genes of interest, will be sequenced to develop specific Sequence Characterised Amplified Regions (SCARs) that will aid selection in different faba bean breeding programs (Paran & Michelmore, 1993). References Bartolomé, B., Quesada, C., Gómez-Cordobés, C., Hernández, T. & Estrella, I. 1994. New
contributions to the inhibition study of α-amylase and trypsin by phenolic compound. In H. Kozlowska, J. Fornal & J. Zdunezyc (Eds). Proceedings International Euro Food Tox IV Conference 'Bioactive Substances in Food of Plant Origin', Olsztyn, Poland, Vol. 1, 233-238.

290
Crofton, G.R.A., Bond D.A. & Duc, G. 2000. Potential seed multiplication problems arising from the existence of two genes for the absence of tannin in Vicia faba L. Plant Varieties and Seeds 13, 131-139.
Duc, G., Marget, P., Esnault, R., LeGuen, J. & Bastianelli, D. 1999. Genetic variability for feeding value of faba bean seeds (Vicia faba): Comparative chemical composition of isogenics involving zero-tannin and zero-vicine genes. Journal of Agricultural Science 133, 185-196.
Duc, G., Sixdenier, G., Lila, M. & Furtoss, V. 1989. Search of genetic variability for vicine and convicine content in Vicia faba L. A first report a gene which codes for nearly zero-vicine and zero-convicine contents. In Recent Advances of Research in Antinutritional Factors in Legume Seeds. J. Huisman, T.F.B. van der Poel & I.E. Leiner (Eds). Proceedings 1st International Workshop on Antinutritional Factors in Legume Seeds, Wageningen, 22-25 November 1988, 305-313.
Grosjean, F., Bourdillon, A.P., Rudeaux, F., Bastianelli, D., Peyronnet, C., Duc, G. & Lacassagne, L. 2000. Valeur alimentaire pour la volaille de féveroles isogéniques (Vicia faba L) avec ou sans tannins et avec ou sans vicine. Sciences et Techniques Avicoles 32, 17-23.
Kuman, R. & Singh, M. 1984. Tannins: their adverse role in human nutrition. Journal of Agricultural and Food Chemistry 32, 447.
Mager, J., Glaser, G., Razin, A., Izak, G., Bien, S. & Noam, M. 1965. Metabolic effects of pyrimidines derived from faba bean glycosides on human erythrocytes deficient in glucose-6-phosphate dehydrogenase. Biochemical and Biophysical Research Communications 29, 235-240.
Marquardt, R. 1989. Dietary effects of tannins, vicine and convicine. In Recent Advances of Research in Antinutritional Factors in Legume Seeds. J. Huisman, T.F.B. van der Poel & I.E. Leiner (Eds). Proceedings 1st International Workshop on Antinutritional Factors in Legume Seeds, Wageningen, 22-25 November 1988, 141-155.
Michelmore, R., Paran, I. & Keselli, V. 1991. Identification of markers linked to disease-resistance genes by bulk segregant analysis: A rapid method to detect markers in specific genomic regions by using segregating populations. Proceedings National Academy of Sciences USA 88, 9828-9832.
Paran, I. & Michelmore, R. 1993. Development of reliable PCR-based markers linked to downy mildew resistance genes in lettuce. Theoretical and Applied Genetics 85, 985-993.
Picard, J. 1976. Aperçu sur l’hérédité du caractère absence de tannins dans les graines de féverole (Vicia faba L.) Annales de l’Amélioration des Plantes 26, 101-106.
Torres, A.M., Weeden, N.F. & Martín, A. 1993. Linkage among isozyme, RFLP and RAPD markers in Vicia faba. Theoretical and Applied Genetics 85, 937-945.
Welsh, J. & McClelland, M. 1990. Fingerprinting genomes using PCR with arbitrary primers. Nucleic Acids Research 18, 7213-7218.
Williams, J.G.K., Kubelik, A.R., Livak, K.J.; Rafalski, J.A. & Tingey, S.V. 1990. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Research 18, 6531-6535.

Session V B


293
Technical and biotechnological modifications of antinutritional factors in legume and oilseeds Gy. Hajos1 and A.U. Osagie2 1Central Food Research Institute, H-1022 Budapest, Herman O. 15. Hungary 2Igbinedion University, Okada, Benin City, Edo State, Nigeria Summary Grain legumes such as peanut (Arachis hypogaea) pea (Pisum sativum), bean (Phaseolus spp), lentil (Lens culinaris), chickpea (Cicer arietinum), lupins (Lupinus spp) and soybean (Glycine max) and oilseeds: rapeseed (Brassica spp), sunflower (Helianthus annuus), cottonseed (Gossipium spp) and linseed (Linum usatissimum) are valuable sources of energy and protein for man and animals. However, their nutritional qualities are often limited by the presence of heat labile and heat stable antinutritional factors (ANFs). Further there is an inherent resistance to digestion of the major globulins. These ANFs need to be eliminated prior to human and animal consumption. Processing technologies have the potential for application as an effective means of improving the nutritional quality of legume and oil seed components. The site of digestion of starch and proteins in legume seed can also be manipulated by processing.
The emphasis in this paper is on the use of improved processing techniques for the removal and/or reduction of the ANFs in grain legumes and oil seeds. However, a number of related problems deserve future research attention. Keywords: antinutritional factors, processing technology, legume, oilseeds Introduction Legume seeds (pulses) and oil seeds have been consumed by humans since the beginning of agriculture and have been ascribed medicinal and cultural as well as nutritional roles. They are valuable sources of dietary protein to complement cereals, roots and tubers. However, their nutritional quality is limited by the presence of heat labile and heat stable ANFs as well as an inherent resistance to digestion of the major globulins. Consumption of legume protein has been shown to reduce plasma low-density lipoprotein. Legume starch is more slowly digested than cereal starch and on ingestion produces less abrupt changes in plasma glucose and insulin. Legume seeds and oil seeds are also valuable sources of dietary fibre and vitamins and minerals including folate, thiamine and riboflavin. They are important components of a prudent diet. However, their consumption is constrained by low yields, by the lack of convenient food applications and by undesirable physiological effects. These latter constraints can be addressed by the use of appropriate processing technology.
Legume seeds and oil seeds are valuable sources of energy and protein for humans and animals. However, the presence and effects of ANFs, which they may contain, often limits their use. The ANFs need to be eliminated prior to human or animal consumption. Fortunately, most of them are thermolabile and are destroyed by heating during usual processing and cooking. Suitable processing techniques (Chitra et al., 1996) should be used for the ANFs, which are heat stable. Simple processes like dehulling and decortication removes tannins as most tannin is in the seed testa.

294
The ANFs are a structurally varied class of compounds broadly divided into two categories; proteins (such lectins and protease inhibitors) and others such as glycosides, phenolics, phytate, oxalate and alkaloids. Polyphenols (tannins) decrease protein digestibility either by binding with digestive enzymes such as trypsin and chymotrypsin or by binding directly to the dietary protein. Phytic acid lowers the bioavailability of minerals and inhibits proteases and amylases. Oligosaccharides are involved in flatulence production, characterized by the production of CO2, H2 and CH4 and lead to abdominal rumbles. Saponins can affect animal performance in a variety of ways including erythrocyte haemolysis, reduction of blood and liver cholesterol, inhibition of smooth muscle activity, alteration of cell wall permeability and binding to the cells of the small intestine thereby affecting nutrient absorption.
The presence, distribution and negative effects of the ingestion of ANFs in grain legumes and oil seeds have been the subject of many papers (Hajós et al., 1995; Gelencsér et al., 1996, 2000; Pusztai et al., 1997; Jansman et al., 1998; Oboh et al., 1998; Sorensen et al., 1998; Burbano et al., 1999; El-Adawy et al., 2001; White et al., 2002; Olguin et al., 2003) and reviews (Liener, 1989; van der Poel, 1990; Udedibie & Carlini, 1998; Hanbury et al., 2000). More recent reviews also exist on this subject (D’Mello, 1995; Belmar et al., 1999; Wiryawan & Dingle, 1999; Phillips et al., 2003). Raw legume and oil seeds contain far higher levels of ANFs than their processed forms hence processing is necessary before the incorporation of legume and oil seeds into food or animal diets. Processing technologies Domestic processing Legumes and oilseeds have been processed in various ways by different investigators with varying results indicating a reduction in the quantity of the ANFs (Elsheikh et al., 2000, Egounlety & Aworh, 2003,). Soaking, washing, dehulling and boiling are generally effective in removing most of the ANFs. Generally, soaking in salt water (i.e. with bicarbonate added) appears to be more effective than soaking in distilled water for reducing the levels of ANFs especially the polyphenols. Soaking reduces the phytate content in most seed by about 10 %. The phytate loss is approximately doubled on cooking. This may be partly due to the formation of insoluble complexes between phytate and other components, such as phytate-protein and phytate-protein-mineral complexes (Egounlety & Aworh, 2003).
Cooking causes a greater reduction in the oligosaccharide level than soaking. Cooking was effective in inactivating tannin, phytic acid and trypsin inhibitor in faba been seeds and consequently improving protein digestibility (Saikia et al., 1999, Elsheikh et al., 2000). The efficacy of soaking, washing and boiling treatments seems to be related to the solubilisation of toxic components. Discarding the solubilised fraction probably means that more of the toxic factor is removed. Dehulling is an effective way of reducing the tannin content of grain legume seed. Hydrothermal treatments Antinutritional factors vary in their thermal stability and the extent to which they are destroyed by heat, in vivo; it is a function of several variables such as cultivar, particle size, and seed moisture content. Moist heat is more effective than dry heat as a method of inactivating lectins and antitryptic activity (Carlini & Udedibe, 1997).

295
Dry roasting, micronization, pressure toasting, extrusion, expander treatment, and pelleting are methods that have been found suitable for use in the heat processing of legume seed. Goelema (1999) and Goelema et al. (1999) reviewed heat processing methods.
Yu et al. (2002) reviewed recent information on the structural and compositional effects of heat processing of legume seeds, in relation to the legume seed characteristics, which affected digestion and nutrient utilisation.
Chymotrypsin inhibitor activity in soybean was more easily reduced by heat-treatment than trypsin inhibitory activity. Lectin activity was relatively heat-resistant. However, all of these activities could be eliminated by aqueous heat-treatment of fully imbibed seeds at 100 oC for 10 min (Armour et al., 1998). To use heat treatment effectively, the temperature and processing duration have to be carefully controlled. With prolonged or elevated heating, basic amino acids, such as lysine, undergo a Maillard reaction, making them less available for metabolism.
Autoclaving seems to be the best method for eliminating phytic acid, raffinose and stachyose (Umoren et al., 1997; Vijayakumari et al., 1997). It also improved the digestibility of the feed components and the feed energy value (Vijayakumari et al., 1998; Lucht, 2003). Extrusion Ismail & Zahran (2002) studied extrusion conditions of some cereals and legumes. The moisture content of the raw material, the extrusion temperature and the feed rate of the extruder were the most significant factors affecting the extrusion process. Suitable extrusion conditions for defatted soybean meal and chickpea flour were 20 % moisture content, 160 °C extrusion temperature and a feed rate of 25 rpm.
Extrusion reduced trypsin inhibitor activity and increased the protein digestibility of legume seed. Alonso et al. (1998, 2000, 2001) reported that extrusion was the best method of abolishing trypsin, chymotrypsin, α-amylase inhibitors and haemagglutinating activity without modifying the protein content. Protein isolates from commercial legumes had lower tannin and phytate content and haemagglunating activity than the original seed, but they had weak functional properties (Fernandez-Quintella et al., 1997).
Soaking and extrusion at a barrel temperature of 140 °C and 180 °C and a feed moisture content of 18 % and 22 %, on pea, chickpea, faba bean (Vicia faba) and kidney bean (Phaseolus vulgaris) seed significantly decreased ANFs such as phytic acid, tannins, phenols, α-amylase and trypsin inhibitors (El-Hady & Habiba, 2003). Germination Several recent reports have confirmed that seed germination is an effective method of reducing ANFs in legume seeds and oil seeds. Cserhalmi et al. (1998) investigated the effect of germination on trypsin inhibitor activity and oligosaccharide content of pea seed. Five days after germination commenced the protein, fat, ash and sucrose content had increased, and trypsin inhibitor activity, raffinose and stachyose content had decreased. Germination also decreased amylase inhibitor activity in legume samples (El-Sayed et al., 1997).
Cuadrado et al. (2000a, b) studied the effect of germination on the lectin level in Lens culinaris cv. Magda 20 seed. The lectin concentration was significantly higher after 6 d of germination: relative lectin levels were particularly high when germination was in the light and seeds were watered daily. The SDS-PAGE and immunoblotting results showed that lectin was not degraded during the first 6 d of germination. However, other storage proteins were broken down by proteolysis.

296
There was no reduction in legume seed germination in solar-heated seed. Farmers in tropical regions may consider using solar heating for disinfesting pea seed during storage before germination (Chauhan & Ghaffar, 2002). Microwave treatment The benefit of microwave treatment is shorter and improved drying. The combined effect of germination, microwave treatment and hot air drying to a final seed moisture content of 12 - 14 % and to a highest temperature of 80 oC can be recommended as a method of decreasing the α-galactooligosaccharides content and improving seed nutritional quality (Kadlec et al., 2001). Ionising radiation Siddhuraju et al. (2002a) reviewed the effect of ionising radiation on ANFs. The impact of gamma irradiation on the chemistry of various ANFs, including non-starch polysaccharides was considered. The potential effect of low irradiation dose levels of up of 10 KGY on the radiation of various ANFs was also reviewed.
A combination of soaking and ionising radiation was effective in reducing trypsin inhibitor activity in a nondose-dependent manner compared with raw seed samples (Siddhuraju et al., 2002b). Machaiah & Pednekar (2002) reported that radiation processing of six legumes: mung (Vigna radiata), Bengal gram (Cicer arietinum) (desi chickpea), horse bean (Vicia faba), horse gram (Dolichos biflorus), cowpea (Vigna unguiculata) and Rajma (Phaseolus vulgaris) at a disinfestation dose (0.25KGY) and germination for 0 - 2 d gave a rapid degradation of flatulence producing factors without affecting their sprout length.
Gamma irradiation was also reduced the level of trypsin inhibitor, phytic acid and oligosaccharides in broad bean (Vicia faba) (Al-Kaisey et al., 2003). Bioprocessing (fermentation) Cuadrado et al. (2002) investigated the effect of natural fermentation on the lectin in Lens culinaris seed. With increased fermentation time there was a rapid decline in haemagglutinating activity. After 72 h of natural fermentation under optimum conditions of flour concentration and temperature the lectin had almost disappeared.
Hoffman et al. (2003) showed that trypsin inhibitor was functionally inactivated as well as being physically degraded during rumen fermentation. Inactivation proceeds much faster than degradation, so it must be due to a preceding mechanism and is not directly related to proteolysis. These results support the view that heat treatment is not mandatory when feeding legume seed to ruminants.
Kiers et al. (2000) investigated the in vitro digestibility of Bacillus fermented soybean (using gastrointestinal enzymes). They concluded, that after 18 - 24 h of fermentation considerable substrate modification had occurred leading to a product with high nutrient availability in which the need for degradation of nutrients by gastro-intestinal enzymes was minimal.
The effect of soaking, dehulling, cooking and fermentation with Rhizopus oligosporus on the ANFs of different legumes were also investigated (Egounlety & Aworth, 2003). The study indicated, that these pre-treatment and fermentation eliminated the most flatulent oligosaccharide in soybean, cowpea and groundbean (Macrotyloma geocarpa).

297
Enzymatic modifications Enzymatic hydrolysis: Commercial enzymes (pepsin, papain, trypsin, ficin and hemicellulase) reduced ANFs in rapeseed (Brassica campestris) and improved its functional properties (Mahajan & Dua, 1998). The ANFs declined remarkably with the use of the different enzymes.
Native globulins of cowpea and algaroba (Ceratonia siliqua) seed were weakly digested by pepsin and were not digested by serine proteases. Heated cowpea proteins were digested more rapidly. Globulins from immature cowpea were more digestible than mature cowpea seed globulins (Araüjo et al., 2002). Enzymatic peptide modification: Enzymatic peptide modification of soy albumin fractions by transpeptidation particularly when combined with covalent amino acid (methionine) enrichment is improves the nutritional value of soy proteins. This is partly by compensating for their methionine deficiency and partly by modifying the structure of the soy ANFs. A reduction in the activity of soybean agglutinin and Kunitz trypsin inhibitor (Hajós et al., 1996) may possibly be the result of modification by proteolysis and the resynthesis of peptide sequences. Changes in the structure by proteolytic hydrolysis and synthesis of peptide chains can lead to alteration in protein conformation, resulting in modified biological activity and in increased nutritional value (Süle et al., 1997; Hajós et al., 1998, 2000; Calderón de la Barca et al., 2000). Combination of enzyme treatments: Walsh et al. (2003) investigated the effect of a limited hydrolysis with Alcalase, cross-linking with transglutaminase and a combination of these modifications on the nitrogen solubility (pH 3 - 8) of soy protein isolate. Their results demonstrate that a combination of these enzyme treatments and the order in which they are applied may have the potential for creating novel food ingredients with improved functional properties. The effect of these combined enzymic modification on soybean ANFs is still to be investigated. Action of phytase enzyme: Phytases belong to a special group of phosphatases, which are capable of hydrolysing phytate, the major storage form of phosphorus in plants, to a series of lower phosphate esters of myo-inositol and phosphate. Frias et al. (2003) studied the kinetics of inositol phosphate (IP) degradation during the action of naturally occurring endogenous phytases in pea and lentil flours. They obtained legume flour with a low IP6 and IP5 content and with a raised IP4 and IP3 content. The action of natural endogenous phytase for 30 min in lentil seed is recommended, as is the addition of commercial phytase enzyme for 60 min to pea seed.
Phytases are widely distributed in nature in plants and in microorganisms. There is a great interest in using enzymes for reducing the phytate content in feedstuffs and food. Various methods, and possible biotechnological applications of phytases, in the animal feed and food industry are discussed by Vohra & Satyanarayana (2003). Influence of malting: The influence of malting on selected seed components (α-galactosides including phytic acid, myo-inositols and lectins) of soybean, chickpea and black bean (Phaseolus vulgaris) samples was investigated by Muzquiz et al. (1999) and Trugo et al. (1999). Their results indicated, that short time malting might be useful for improving the nutritional characteristics of the samples.

298
High hydrostatic pressure High hydrostatic pressure processing is one of the main emerging preservation technologies in the food industry. High hydrostatic pressure can modify the activity of some enzymes and protein structure (Farkas et al., 2002, Hugas et al., 2002).
High pressure has been used to modify the functional properties of soy products. However, little is known about the effect of high pressure on the individual constituents of soy proteins (Galazka et al., 1999; Molina et al., 2001; Molina & Ledward, 2003). Molina et al. (2001) suggest, that a combination of heat and high-pressure treatments offers a range of possibilities for the development of new food products with novel textures. Harvest technique The trypsin inhibitor activity of the legumes (pea, chickling vetch (Lathyrus sativus), lentil, and soybean) was significantly altered by harvest technique. Pisulewska & Pisulewska (2000) concluded, that the improvement in nutritional value resulting from direct versus a two-phase harvest was limited to pea and chickling vetch. In the case of lentil and soybean, the reduction of trypsin inhibitor activity was too small to consider the proposed method feasible. Discussion Recent research has confirmed that a short-duration heat treatment is effective in reducing many ANFs in grain legume and oil seeds. Hydrothermal treatment is safe, efficient and economic for ANF reduction.
However, enzyme supplementation has potential and would be the method of choice in the future in the absence of heating facilities. Generally, a combination of methods is recommended for ANF removal e.g. germination and roasting, germination and pressure-cooking, soaking and autoclaving (Vaishali et al., 1998; Vijayakumari et al., 1998).
The choice of treatment will depend on the availability of facilities and on economic considerations. Methods designed to reduce ANFs frequently, alter protein, carbohydrate and other components of the seed leading to changes in their metabolisable energy and their protein and starch digestibility. It is therefore important to carefully evaluate products using animal experiments. This approach could open possible avenues for the utilisation of under-utilised and non-conventional crops as additional food and feed sources in near future (Vijakumari et al., 1997; Udedibie & Carlini, 1998; Siddhuraju et al., 2002a, b).
The emphasis in this paper has been on the use of improved processing techniques for the removal and/or reduction of ANFs in legume and oil seeds. However, there are a number of problems, which deserve further research. A legume or oil seed may contain two or more ANFs. Jack bean (Canavalia ensiformis) contains concanavaline – A (a lectin), conavanine and canaline (non-protein amino acids), urease and saponins. Similarly lima bean (Phaseolus lunatus) not only contains trypsin inhibitor and haemagglutinin but also phaseolunatin, which is a cyanogenic glycoside. These ANFs are generally believed to act in concert to produce the myriad of deleterious effects when raw or improperly processed beans are consumed. These ANFs act synergistically to exert their negative influences (Armour et al., 1998; Udedibe & Carlini, 1998). The contribution of each of the ANFs to the interactive effect needs to be properly assessed.

299
Common processing techniques, like dry and wet heating, extracting, etc., have been widely and successfully used to reduce the concentration of ANFs in plant foods and feeds. There is a need for caution when resorting to treatment methods because they can sometimes have unintended adverse effects on the nutritional quality of the food or feed materials, e.g. heat treatment can alter protein and carbohydrate structure.
Another important factor to consider is the interaction between the various ANFs in a particular substance. These interactions may lead to a decrease in the toxic effect of the interacting ANFs (Francis et al., 2001). A more detailed study of such interactions would be particularly useful. Studies are also needed to expose the effects of mixtures of ANFs in proportions similar to those found in plant derived nutritional sources.
High hydrostatic pressure can modify the activity of some enzymes and protein structure (Hugas et al., 2002). The primary structures of proteins are not very sensitive to high hydrostatic pressure. However, modification of weak bonds can lead to protein denaturation or on the contrary to enzyme activation. The effects vary with protein type and processing conditions. The effect of the high hydrostatic pressure on the ANFs of legumes still remains to be studied. New studies are envisaged to estimate pressure-induced alterations to the structure and activity of antinutritive proteins of legume and oil seeds.
Breeding legume and oil seeds for improved genotypes and improved processing technology will undoubtedly lead to changes in the content of both nutrients and ANFs, which must be carefully monitored. There is a general lack of sensitive methods for the quantitative determination of ANFs in foods and feeds. In many cases, identification and quantitative determination of the primary agent of toxicity in the seeds can be accomplished with considerable difficulty and not always with complete unanimity. For example, published phytic acid data often give over-estimates because the colourimetric methods used cause coprecipitation with other phosphorus compounds, which increases the value. High performance liquid chromatography has been recommended as the method of choice for phytic acid determination (Osagie et al., 1996).
Processing condition, chemical composition and cultivar can all influence starch digestibility in common beans (Phaseolus vulgaris) (Osorio-Diaz et al., 2002). Therefore, specific processing methods and formulations are needed for specific purposes. For principal foods a processing method leading to the highest starch digestibility, such as flour preparation, should be used. However, diabetic and hyperlipidemic subjects may be advised to consume whole cooked beans, with decreased starch bioavailability.
Most investigators have generally used bovine trypsin to measure the trypsin inhibitor activity of various grain legume and oil seeds. This is despite the fact that the nutritive value of the protein may have been evaluated in an entirely unrelated animal species. In vitro studies on the inhibition of proteases in the pancreatic juice of different animal species have shown marked differences in the degree to which these enzymes are inhibited by the protease inhibitors of the various legumes (Liener, 1989).
A novel concept, which meets new expectations of protein safety and food quality has been developed over the last few years. Proteins are needed with specific physiological functions and with particular protein quality for healthy nutrition. Enzymatically modified proteins (Hajós, 1996) will play a major role in producing foods in the future especially for controlling the antinutritive characters of legume- or oilseed-based food proteins.
Gravimetric, haemolytic and foam-based assays for saponins are non-specific and are liable to considerable error (Price et al., 1987). Thin layer chromatographic and spectrophotometric methods are also not suitable for estimating saponins in seed extracts. This is because the reactions are not specific and coloured products can be produced from other compounds such as flavonoids. It has been recommended that saponins should be

300
isolated with methanol, hydrolysed with acid, and the sapogenins derivatives quantitatively determined by gas chromatography and mass spectrometry (Osagie et al., 1996).
Methods of single enzymatic hydrolysis combined with microdialysis and micro-high–performance anion exchange chromatography with electrospray ionisation mass spectrometry (Okatch et al., 2003) have proved to be sufficient and reproducible for the characterisation of legume samples.
Thus it is likely that those who need information on the biotechnological modification of ANFs will need to generate reliable analytical methods. References Al-Kaisey, M.T., Alwan, A.K., Mohammad, M.H. & Saeed, A.H. 2003. Effect of gamma
irradiation on antinutritional factors in broad bean. Radiation Physics and Chemistry 67, 493-496.
Alonso, R., Aguirre, A. & Marzo, F. 2000. Effect of extrusion and traditional processing methods on antinutrients and in vitro digestibility of protein and starch in faba and kidney beans. Food Chemistry 68, 159-165.
Alonso, R., Orue, E & Marzo, F. 1998. Effect of extrusion and conventional processing methods on protein and antinutritional factor content in pea seeds. Food Chemistry 63, 505-512.
Alonso, R., Rubio, L.A., Muzquiz, M. & Marzo, F. 2001. The Effect of extrusion cooking on mineral bioavailability in pea and kidney bean seed meals. Animal Feed Science and Technology 94, 1-13.
Araüjo, A.H., Cardoso, P.C.B., Pereira, R.A., Lima, L.M., Oliveira, A.S., Miranda, M.R.A., Xavier-Filho, J. & Sales, M.P. 2002. In vitro digestibility of globulins from cowpea (Vigna unguiculata) and xerophytic algaroba (Prosopis juliflora) seeds by mammalian digestive proteinases: a comparative study. Food Chemistry 78, 143-147.
Armour, J.C., Chanaka Perera, R.L. Buchan, W.C. & Grant, G. 1998. Protease inhibitors and lectins in soya beans and effects of aqueous heat treatment. Journal of Science and Food Agriculture 78, 225-231.
Belmar, R., Nava-Mentero, R., Sandoval-Casto, C. & McNab, J.M. 1999. Jack bean (Canavalia ensifomis L. DC) in poultry diets: antinutritional factors and detoxicattion studies –a review. World’s Poultry Science Journal 55, 37-59.
Burbano, C., Muzquiz, M., Ayet, G., Cuadrado, C. & Pedrosa M.M. 1999. Evaluation of antinutritional factors of selected varieties of Phaseolus vulgaris. Journal of the Science of Food and Agriculture 79, 1468-1472,
Calderón de la Barca, A.M., Ruiz-Salazar, R.A. & Jara-Marini, M.E. 2000. Enzymatic hydrolysis and synthesis of soy protein to improve its amino acid composition and functional properties. Journal of Food Science 65, 246-253.
Carlini, C.R. & Udedibie, A.B. 1997. Comparative effects of processing methods on haemagglutinating and antitryptic activities of Canavalia ensiformis and Canavalia braziliensis seeds. Journal of Agriculture and Food Chemistry 45, 4372-4377.
Chauhan, Y.S. & Ghaffar, M.A. 2002. Solar heating of seeds-a low cost method to control bruchid (Callosobruchus spp.) attack during storage of pigeonpea. Journal of Stored Products Research 38, 87-91.
Chitra, U., Singh, U. & Rao, P.V. 1996. Phytic acid, in vitro protein digestibility, dietary fibre, and minerals of pulses as influenced by processing methods. Plant Foods For Human Nutrition 49, 307-316

301
Cuadrado, C., Burbano, C., Gelencser, E., Pedrosa, M.M., Ayet, G., Muzquiz, M. & Hajós, Gy. 2000a. Effect of germination on lectin in Lens culinaris seeds, Acta Alimentaria 29, 231-240.
Cuadrado, C., Burbano, C., Gelencser, E. Pedrosa, M.M., Ayet, G., Muzquiz, M., Pusztai, A., Szerdahelyi, E. & Hajós, Gy. 2000b. Influence of germination on lentil lectin. COST 98, Effects of Antinutrients on the Nutritional Value of Legume Diets. S. Pierzinowsky & E. Gelencser (Eds) Official Publications of the European Communities, Brussels. 9, 92-97.
Cuadrado, C., Hajós, Gy., Burbano, C., Pedrosa, M.M., Ayet, G., Muzquiz, M., Pusztai, Á. & Gelencsér, É. 2002. Effect of natural fermentation on the lectin of lentils measured by immunological methods. Food and Agricultural Immunology 14, 41-49.
Cserhalmi, Z., Schuster-Gajzago, I. & Czukor, B. 1998. Effect of germination on antrinutritive factors of pea. In AEP (Ed.). Proceeding of 3rd European Conference on Grain Legumes, Valladolid, Spain, 14-19 November 1998, 391.
D’Mello, J.P.F. 1995. Antinutritional substances in legume seeds. In Tropical legumes in Animal Nutrition. J.P.F. D’Mello & C. Devendra (Eds.). CAB International, Wallingford, 135-171.
Egounlety, M. & Aworh, O.C. 2003. Effect of soaking, dehulling, cooking and fermentation with Rhizopus oligosporus on the oligosaccharides, trypsin inhibitor, phytic acid and tannins of soybean (Glycine max Merr.), cowpea (Vigna unguiculata L. Walp) and groundbean (Macrotyloma geocarpa Harms). Journal of Food Engineering 56, 249-254.
El-Adawy, T.A., Rahma, E.H., El-Bedawey, A.A. & Gafar, A.F. 2001. Nutritional potential and functional properties of sweet and bitter lupin seed protein isolates. Food Chemistry 74, 455-462.
El-Hady, E.A.A. & Habiba, R.A. 2003. Effect of soaking and extrusion conditions on antinutrients and protein digestibility of legume seeds. Lebensmittel Wissenschaft and Technologie 36, 285-293.
El-Sayed, M.M., Ramadan, M.E., Ali, H.M. & El-Morsi, E.A. 1997. Biochemical studies on alpha amylase inhibitors in some legume seeds. Annals of Agriculture Science, Moshtohor 35, 357-374.
Elsheikh, E.A.E., Fadul, I.A. & El Tinay, A.H. 2000. Effect of cooking on anti-nutritional factors and in vitro protein digestibility (IVPD) of faba bean grown with different nutritional regimes. Food Chemistry 68, 211-212.
Farkas, J., Hajós, Gy., Szerdahelyi, E., Andrássy, É., Krommer J. & Mészáros, L. 2002. Protein changes in high pressure pasteurised raw sausage batter. In Congress Proceedings, 48th International Congress of Meat Science and Technology, Rome, 25-30 August 2002, P3/5.Vol.1, pp. 180-181.
Fernandez-Quintela, A., Macarulla, M.T., Barrio, A.S. del, Martinez J.A. & Del-Barrio, A.S. 1997. Composition and functional properties of protein isolates obtained from commercial legumes grown in northern Spain. Plant Foods for Human Nutrition 51, 331-342.
Francis, G., Makkar, H.P.S. & Becker, K. 2001. Antinutritional factors present in plant-derived alternate fish feed ingredients and their effects on fish. Aquaculture 199, 3-4, pp. 197-227.
Frias, J., Doblado, R., Antezana, J.R. & Vidal-Valverde, C. 2003. Inositol phosphate degradation by the action of phytase enzyme in legume seeds. Food Chemistry 81, 233-239.

302
Galazka, V.B., Dickinson, E. & Ledward, D.A. 1999. Emulsifying behaviour of 11S globulin Vicia faba in mixtures with sulphated polysaccharides: comparison of thermal and high-pressure treatments. Food Hydrocolloids 13, 425-435.
Gelencsér, É., Hajós, Gy., Pusztai, Á., Grant, G., Bardocz, S. & Ewen, S. 1996. Nutritional consequences of exposing rats to soya bean albumin diet. In Lectins. Biology, Biochemistry and Clinical Biochemistry. E. Van Driessche, J. Fischer, S. Beeckmans & T.C. Bog-Hansen (Eds). Textop. pp. 236-239.
Gelencsér É., Hajós Gy., Zduńczyk Z. & Jędrychowski L. 2000. Content of lectins in respect to the level of protein and other antinutritional factors in Polish pea varieties Polish Journal of Food Nutrition and Science 50, 91-94.
Goelema, J.O. 1999. Processing of legume seeds: effects of digestive behaviours in dairy cows. PhD Thesis. Wageningen Agricultural University, The Netherlands.
Goelema, J.O., Smits, A., Vaessen, L.M. & Wemmers, A. 1999. Effects of pressure toasting, expander treatment and pelleting on in vitro and in situ parameters of protein and starch in a mixture of broken peas, lupins and faba beans. Animal Feed Science and Technology 78, 109-126,
Hajós, Gy. 1996. Enzymatic modification as a tool for alteration of safety and quality of food proteins. In Surface activity of proteins: chemical and physicochemical modifications. S. Magdassi (Ed). Marcel Dekker, New York, 131-180.
Hajós, Gy., Farkas, Sz., Szerdahelyi, E. & Polgár, M. 1998. Effect of methionine enrichment on the biological activities of food proteins. Acta Biologica Hungarica 49, 201-207.
Hajós, Gy., Gelencsér, É., Ewen S.W.B., Pusztai, A., Grant G., Sakhri, M. & Bardocz, S. 1995. Biological effects and survival of trypsin inhibitors and the agglutinin from soya bean in the gut. Journal of Agriculture and Food Chemistry 43, 165-170.
Hajós, Gy., Gelencsér, É., Grant, G., Bardocz, S., Sakhri, M., Duguid, T.J., Newman A.M. & Pusztai A. 1996. Effect of proteolytic modification and methionine enrichment on the nutritional value of soya albumins for rats. Journal of Nutritional Biochemistry 7, 481-487.
Hajós, Gy., Szerdahelyi, E. & Gelencsér, É. 2000. Effect of enzymatic modification of the biological activities of soybean lectins. COST 98, Effects of Antinutrients on The Nutritional Value of Legume, Diets. S. Pierzinowsky & E. Gelencser (Eds). Official Publications of the European Communities, Brussels 9, 97-100.
Hanbury, C.D., White, C.L., Mullan, B.P. & Siddique, K.H.M. 2000. A review of the potential of Lathyrus sativus L. and L. cicera L. grain for use as animal feed. Animal Feed Science and Technology 87, 1-27.
Hoffmann, E.M., Muetzel, S. & Becker, K. 2003. The fermentation of soybean meal by rumen microbes in vitro reveals different kinetic features for the inactivation and the degradation of trypsin inhibitor protein. Animal Feed Science and Technology 106, 189-197.
Hugas, M., Garriga, M. & Monfort, J.M. 2002. New mild technologies in meat processing: high pressure as a model technology. Meat Science 62, 359-371.
Ismail, F.A. & Zahran, G.H. 2002. Studies on extrusion conditions of some cereals and legumes. Egyptian Journal of Food Science 30, 59-76.

303
Jansman, A.J.M., Huisman, J., Beelen, G.M. & Wiebenga, J. 1998. Effects of soya trypsin inhibitors on faecal nutrient digestibility and nitrogen retention in young piglets. In Recent Advances of Research in Antinutritional Factors in Legume Seeds and Rapeseed. A.J.M. Jansman, G.D. Hill & A.F.B. van der Poel (Eds.). Proceedings 3rd International Workshop on Antinutritional Factors in Legume Seeds and Rapeseed, Wageningen, 8-10 July 1998, 331-334.
Kadlec, P., Rubecova, A., Hinkova, A., Kaasova, J., Bubnik, Z. & Pour, V. 2001. Processing of yellow pea by germination, microwave treatment and drying. Innovative Food Science and Emerging Technologies 2, 133-137.
Kiers, J.L., van Iaeken, A.E.A., Rombouts, F.M. & Nout, M.J.R. 2000. In vitro digestibility of Bacillus fermented soya bean. International Journal of Food Microbiology 60, 163-169.
Liener, I.E. 1989. Antinutritional factors in legume seeds: state of the art. In Recent Advances of Research in Antinutritional Factors in Grain Legume Seeds. J. Huisman, T.F.B. van der Poel & I.E. Liener (Eds). Proceedings 1st International Workshop on Antinutritional Factors in Legume Seeds, Wageningen 13-25 November 1988, 6-13.
Lucht, H.W. 2003. Hydrothermal process for inactivation of antinutritional factors in oilseeds and legumes. Muhule-+-Mischfutter 140, 72-76.
Machaiah, J.P. & Pednekar, M.D. 2002. Carbohydrate composition of low dose radiation-processed legumes and reduction in flatulence factors. Food Chemistry 79, 293-301.
Mahajan, A. & Dua, S. 1998. Improvement of functional properties of rapeseed (Brassica campestris var toria) meal by reducing antinutritional factors employing enzymatic modification. Food Hydrocolloids 12, 349-355.
Molina, E. & Ledward, D.A. 2003. Effects of combined high-pressure and heat treatment on the textural properties of soya gels. Food Chemistry 80, 367-370.
Molina, E., Papadopoulou, A. & Ledward, D.A. 2001. Emulsifying properties of high-pressure treated soy protein isolate and 7S and 11S globulins. Food Hydrocolloids 15, 49-55.
Muzquiz, M., Pedrosa, M.M., Trugo, L., Ayet, G., Burbano, C. & Cuadrado, C. 1999. Influence of malting process on levels of non-nutritive compounds. In Effects of Antinutients on the Nutritional Value of Legume Diets (COST 98). European Communities, Brussels, 7, 153-159.
Oboh, H.A., Muzquiz, M., Burbano, C., Cuadrado, C., Pedrosa, M.M., Ayet, G. & Osagie, A.U. 1998. Anti-nutritional constituents of six underutilized legumes grown in Nigeria. Journal of Chromatography A 823, 307-312.
Okatch, H., Torto, N. & Armateifio, J. 2003. Characterisation of legumes by enzymatic hydrolysis, microdialysis sampling, and micro-high-performance anion-exchange chromatography with electrospray ionisation mass spectrometry. Journal of Chromatography A 992, 67-74.
Olguin, M.C., Hisano, N., D’Ottavio, A.E., Zingale, M.I., Revelant, G.C. & Calderari, S.A. 2003. Nutritional and antinutritional aspects of an Argentinean soy flour assessed on weanling rats. Journal of Food Composition and Analysis 16, 441-449.
Osagie A.U., Muzquiz, M., Burbano, C., Cuadrado C., Ayet, G. & Castano, A. 1996. Some antinutritional constituents in ten staple food items grown in Nigeria. Tropical Science (UK) 36, 109-115.
Osorio-Diaz, P., Bello-Pérez, L.A., Agama-Acevedo, E., Vargas-Torres, A., Tovar, J. & Paredes-López, O. 2002. In vitro digestibility and resistant starch content of some industrialized commercial beans (Phaseolus vulgaris L.). Food Chemistry 78, 333-337.

304
Philips, R.D., McWatters, K.H., Chinnan, M.S., Hung, Y.C., Beuchat, L.R., Sefa-Dedeh, S., Sakyi-Dawson, E., Ngoddy, P., Nnanyelugo, D., Enwere, J., Komey, N.S., Liu, K., Mensa-Wilmot, Y., Nnanna, H.A., Okeke, C., Prinyawiwatkul, W. & Saalia, F.K. 2003. Utilization of cowpeas for human food. Field Crops Research 82, 193-213.
Pisulewska, E. & Pisulewski, P.M. 2000. Trypsin inhibitor activity of legume seeds (peas, chickling vetch, lentils, and soya beans) as affected by the technique of harvest. Animal Feed Science and Technology 86, 261-265.
Price, K.R., Johnson, I.T. & Fenwick, G.R. 1987. The chemical and biological significance of saponins in foods and feeding stuffs. CRC Critical Reviews in Food Science and Nutrition 26, 27-135.
Pusztai, A., Grant, G., Bardocz, S., Gelencsér, É. & Hajós, Gy. 1997. Novel dietary strategy for overcoming the antinutritional effects of soya bean whey of high agglutinin content. British Journal of Nutrition 77, 933-945.
Saikia, P, Sarkar, C.R. & Borua, I. 1999. Chemical composition, antinutritional factors and effect of cooking on nutritional quality of rice bean Vigna umbellata (Thunb, Ohwi & Ohashi). Food Chemistry 67, 347-352.
Siddhuraju, P., Makkar, H.P.S. & Becker, K. 2002a. The effect of ionising radiation on antinutritional factors and the nutritional value of plant materials with reference to human and animal food. Food Chemistry 78, 187-205.
Siddhuraju, P., Osoniyi, O., Makkar, H.P.S. & Becker, K. 2002b. Effect of soaking and ionising radiation on various antinutritional factors of seeds from different species of an unconventional legume, Sesbania and a common legume, green gram (Vigna radiata). Food Chemistry 79, 273-281.
Sorensen, A.D., Hansen, A.B., Sorensen, S., Barkholt, V. & Frokiaer, H. 1998. Influence of industrial processing of peas on the content of antinutritional factors and the in vitro digestibility. In Opportunities for High Quality, Healthy and Added-Value Crops to Meet European Demands. Grain Legumes. AEP (Ed.). Proceedings 3rd European Conference on Grain Legumes, Valladolid, Spain 14-19 November 1998, 348-350.
Süle, E., Hajós, Gy. & Tömösközi. S. 1997. Functional properties of the enzymatically modified soya protein isolate. Acta Alimentaria 26, 279-285.
Trugo, L.C., Muzquiz, M., Pedrosa, M.M., Ayet, G., Burbano, C., Cuadrado, C. & Cavieres, E. 1999. Influence of malting on selected components of soya bean, black bean, chickpea and barley. Food Chemistry 65, 85-90.
Udedibie, A.B.I. & Carlini, C.R. 1998. Brazilian Mucuna pruriens seeds (velvet bean) lack hemagglutinating activity. Journal of Agriculture and Food Chemistry 46, 1450-1452.
Udedibie, A.B.I. & Carlini, C.R. 1998. Questions and answers to edibility problem of the Canavalia ensiformis seeds - A review. Animal Feed Science and Technology 74, 95-106.
Umoren, U.E., Tewe, O.O Bokanga, M. & Jackai, L.E.N. 1997. Protein quality of raw and autoclaved cowpeas: comparison between some insect resistant and susceptible varieties. Plant Food for Human Nutrition 50, 301-315.
Vaishali, A, Sadhana, J., Seema, K., Kishore, P. & Shashi, C. 1998. Effects of processing on phytate degradation & mineral solubility in pulses. Journal of Food Science and Technology – Mysore 35, 330-332.
Van der Poel, A.F.B. 1990. Effect of processing on antinutritional factors and protein nutritional value of dry beans (Phaseolus vulgaris). A review. Animal Feed Science and Technology 29, 179-208.

305
Vijayakumari, K., Siddhuraju, P. & Janardhanan, K. 1997. Effect of domestic processing on the levels of certain antinutrient in Prosopis chilensis (Molina) Stunz seeds. Food Chemistry 59, 367-371.
Vijayakumari, K., Siddhuraju, P., Pugalenthi, M. & Janardhanan K. 1998. Effect of soaking, cooking and autoclaving on the levels of antinutrients and digestible proteins in seeds of Vigna acontifolia and Vigna sinensis. Food Chemistry 63, 259-264.
Vohra, A. & Satyanarayana, T. 2003. Phytases: Microbial sources, production, purification and potential biotechnological applications. Critical Reviews in Biotechnology 23, 29-60.
Walsh, D.J., Cleary, D., McCarthy, E., Murphy, S. & Fitzgerald, R.J. 2003. Modification of the nitrogen solubility properties of soy protein isolate following proteolysis and transglutaminase cross-linking. Food Research International 36, 677-683.
White, C.L., Hanbury, C.D., Young, P., Phillips, N., Wiese, S.C., Milton, J.B., Davidson, R.H., Siddique, K.H.M. & Harris, D. 2002. The nutritional value of Lathyrus cicera and Lupinus angustifolius grain for sheep. Animal Feed Science and Technology 99, 45-64.
Wiryawan, K.G. & Dingle, J.G. 1999. Recent research on improving the quality of grain legumes for chicken growth. Animal Feed Science and Technology 76, 185-193.
Yu, P., Goelema, J.O., Leury, B.J., Tamminga, S. & Egan, A.R. 2002. An analysis of the nutritive value of heat processed legume seeds for animal production using the DVE/OEB model: A review. Animal Feed Science and Technology 99, 141-176.


307
Modifications of seed storage proteins during germination and seedling growth of faba bean cotyledons C. Cuadrado1, E. Guillamón1, C. Goyoaga1, M.M. Pedrosa1, P. Altares1, C. Burbano1, M. Muzquiz1 & C. Romero2 1Departamento Tecnología de Alimentos, SGIT-INIA, Carretera de la Coruña, Km 7,5, 28040 Madrid, Spain 2Departamento Genética, Facultad de Ciencias Biológicas, Universidad Complutense, Ciudad Universitaria s/n, 28040 Madrid, Spain Summary The SDS-PAGE patterns of two varieties of Vicia faba L. cotyledons were analysed to evaluate the qualitative changes of the major storage proteins, legumins (11S), vicilins (7S) and albumins (2S) during germination and seedling growth. The results confirm previous findings that legume storage proteins are mainly mobilised after germination and during seedling growth. Legumin electrophoretic pattern showed their most apparent and important changes 3 days after imbibition (DAI), because of the number of proteins affected. Vicilin breakdown started earlier, during the germination phase (1 - 2 DAI). Albumin mobilisation was related to the late seedling growth. Keywords: seed storage proteins, mobilisation, germination, seedling growth, Vicia faba Introduction Faba bean seed are used in human and in animal nutrition as sources of amino acids and energy. These legume seeds usually contain large amounts of stored materials in their cotyledons that are used as precursors for synthetic processes during germination and seedling growth. Starch, storage proteins and other proteins are enzymatically degraded to support early plant growth.
Legume storage proteins have been classified into three major groups on the basis of their sedimentation coefficients: legumins (11S), vicilins (7S) and albumins (2S) (Shewry et al., 1995). The 11S and 7S proteins are globulins and were first described by Osborne (1924).
According to Schlereth et al. (2000) germination in V. sativa ends 24 h after imbibition and then mobilisation of cotyledon globulins takes place, starting with vicilin breakdown. Such events are associated with seedling growth.
The aim of this work was to evaluate the qualitative changes in seed legumins, vicilins and albumins during the germination and seedling growth of two cultivars of Vicia faba. Materials and methods Plant material Seed of V. faba (cvs Alameda and Brocal) were obtained from C.I.F.A., Córdoba (Spain). The V. faba seed was screened for imperfections (40 seeds/tray) and were then placed in a constant environment chamber at 20 ºC with 8 h of light/dark, on sand-water (5:1 w/v) in covered photography trays. Germinated V. faba seedlings were rinsed daily and harvested at

308
intervals up to 9 d. The seedlings were sub-divided into their constituent parts and replicates were bulked into testa, cotyledon and axis fractions that were weighed, frozen and then freeze-dried. Only the cotyledons were analysed in this work. Protein extraction Fractionation of the seed storage proteins was by Danielsson’s (1949) method, with minor modifications. The method uses ammonium sulphate precipitation and isoelectric precipitation at pH 4.8 and fractionation based on differential protein solubility. The repeatability and reproducibility of the method was assessed by six extractions from raw V. faba meal from both cultivars. Comparable yield results were obtained. Protein analysis The freeze-dried samples were ground to pass through a 1 mm sieve (Tecator, Cyclotec 1093). Protein concentration was determined by the Bradford (1976) dye-binding assay using bovine serum albumin (BSA) as a standard. All assays were in duplicate. The SDS-PAGE analysis was as per Laemmli (1970), with minor modifications (Puzstai et al., 1981). The acrylamide:bisacrylamide ratios of running and stacking gels were 17.6:0.45 % and 3.95:1.42 %, respectively. Samples of 16 mg of cotyledon or seedling meal/ml of buffer were incubated at 100 oC for 30 min. in 0.01 M phosphate sample buffer containing 3 % (w/v) sodium dodecyl sulphate (SDS) and 2 % (v/v) β-mercaptoethanol before electrophoresis.
Gels were stained with 0.5 % (w/v) Coomassie Brilliant Blue R (Sigma, St Louis, Mo). The SDS-PAGE molecular weight markers were from BioRad. Gels were scanned and the molecular weight of the bands was assessed using the Quantity One Program supplied by Bio-Rad. Results and discussion The total protein in g/cotyledon pair of both cultivars of V. faba is shown in Figure 1. There was no difference in the amount of protein extracted per mg of dry weight (DW) during the germination of cv. Alameda (234.5 to 200.9 µg protein/mg DW for 0 and 9 DAI, respectively) or in cv. Brocal (210.5 to 195.5 µg/mg DW for 0 and 9 DAI, respectively). However, there was a decrease in total protein calculated on a per seed basis, implying that protein and dry matter (DM) hydrolysis proceeded in parallel during seed germination. There was a decrease from 0.18 to 0.10g protein/cotyledon pair in cv. Alameda when samples were compared at 0 and 9 DAI. There was a similar decrease in cv. Brocal (0.14 to 0.08 g protein/cotyledon pair at 0 and 9 DAI, respectively) (Figure 1). Figure 1. Total protein/seed (cotyledon pair) in Vicia faba, harvested up to 9 DAI (days after
imbibition). ▲ = cv. Alameda; • = cv. Brocal.
DAI0
0,05
0,1
0,15
0,2
0 0,5 1 2 3 4 5 6 7 8 9
Tota
l pro
tein
(g
)

309
Using this fractionation method the three main legume storage proteins (11S, 7S and 2S) were isolated from raw and germinated seeds of both cultivars of faba bean. The major fraction was the 11S (legumins) and the 7S (vicilins) proteins were the minor fraction in both cultivars when the protein content (g/100g total protein) of each fraction was measured. Further, the relative proportion of the three sub-populations was the same in both cultivars in ungerminated and germinated seed. Derbyshire et al. (1976) reported differences in these proportions in other cultivars of V. faba and intervarietal differences were also detected in the relative abundance of 11S and 7S Cicer arietinum proteins (data not shown).
The electrophoretic patterns of the total protein extract and for the 11S, 7S and 2S fractions during germination and seedling growth of cvs Alameda and Brocal are summarised in Figure 2. Both cultivars showed similar patterns and behaviour in their total extract (A, E) and in their protein fractions over the test period. The results showed that the 11S fractions (B, F) and the total protein extract had a similar band pattern. Eleven of the 12 bands observed at 0 DAI were the same. From 0.5 to 9 DAI there were changes in the legumins. There was a decrease in bands in the 58 - 34 kDa range. The biggest reduction of 50 % was in the 34 kDa band at 3 DAI. There was an increase in bands of about 27 kDa from 4 to 5 DAI. Between 22 - 16 kDa some bands decreased while others, with lower MW increased. This was probably due the presence of breakdown products. There was no change in the 22 kDa band during the experiment.
Figure 2. Analysis by SDS-PAGE of changes in protein patterns of total protein extract (A, E), 11S legumin (B, F), 7S vicilin (C, G) and 2S albumin (D, H) of Vicia faba cvs Alameda and Brocal during germination and seedling growth. The molecular weights (kDa) are given.
The vicilin 7S fraction (C, G) changed less during the experiment compared with the
11S fraction. A marked feature was the disappearance of the bands in the range 65 - 62 kDa and a decrease in the 27 kDa band at 1 - 2 DAI. The predominant 19 kDa band and the minor 16 kDa band were stable throughout the experiment.
Total protein extract legumin vicilin albumin 113 _
54 _
35 _
21 _
7 _
ALAMEDA
kDa
29 _
0 0.5 1 2 3 4 5 6 7 8 9 days after imbibition
0 0.5 1 2 3 4 5 6 7 8 9 days after imbibition
0 0.5 1 2 3 4 5 6 7 8 9 days after imbibition
BROCAL
0 0.5 1 2 3 4 5 6 7 8 9 days after imbibition
113 _54 _
35 _
21 _
7 _
kDa
29 _
A B C D
E F G H

310
The 2S albumins (D, H) had a wide range of molecular weights. There was an albumin protein of 75 kDa that decreased after 3 DAI, which was also present in the total protein extract (A, E). The bands at 35, 27 and 20 kDa were not degraded during the experiment. In the range 35 - 20 kDa the main changes that occurred were the appearance of a 32 kDa band and the breakdown of the 30 kDa band after 4 DAI. After 2 DAI there were reductions in the 26, 25 and 24 kDa bands. The same occurred in the 15 - 10 kDa bands. However, the breakdown of the 8 kDa band was later, at 6 DAI, when seedling growth was more advanced.
Our results agree with results for V. sativa, which indicated that vicilin mobilisation occurred earlier during germination than legumin breakdown (Schlereth et al., 2000; Müntz et al., 2001). However, in this work the legumin fraction (11S) in both faba bean cultivars was more abundant and changed more than the vicilin fraction (7S). From our results structural modifications of 2S proteins seems to be related to late seedling growth. In conclusion, our results confirm that legume storage proteins are mainly mobilised after germination and during seedling growth from 3 DAI. The legumin electrophoretic pattern of V. faba showed the most changes, due to the number of protein bands that were affected from 3 DAI. Vicilin breakdown started earlier, even during germination and albumin mobilisation was related more to late seedling growth. Acknowledgements This work was partly funded by INIA SC97-057. References Bradford, M.M. 1976. A rapid and sensitive method for the quantitation of microgram
quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry 72, 248-254
Danielsson, C.E. 1949. Seed globulins of the Gramineae and Leguminosae. Biochemistry Journal 44, 387-400.
Derbyshire, E., Wright, D.J. & Boulter, D. 1976. Legumin and vicilin, storage proteins of legume seeds. Phytochemistry 15, 3-24.
Laemmli, U.K. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227: 680-685
Müntz, K., Belozersky, M.A., Dunaevsky, Y.E., Schlereth, A. & Tiedeman, J. 2001. Stored proteinases and the initiation of storage protein mobilization in seeds during germination and seedling growth. Journal of Experimental Botany 52, 1741-1752.
Osborne, T.B. 1924. The Vegetable Proteins, 2nd Edition. Longmans-Green, London. Pusztai, A., Grant, G. & Steward, J.C. 1981. A new type of Phaseolus vulgaris (cv. Pinto
III) seed lectin: Isolation and characterization. Biochimica et Biophysica Acta 671, 146. Schlereth, A., Becker, C., Horstmann, C., Tiedemann, J. & Müntz, K. 2000. Comparison of
globulin mobilisation and cystine proteinases in embryonic axes and cotyledons during germination and seedling growth of vetch (Vicia sativa L.) Journal of Experimental Botany 51, 1423-1433.
Shewry, P.R., Napier, J.A. & Tatham, A.S. 1995. Seed storage proteins: structures and biosynthesis. The Plant Cell 7, 945-956.

311
Effects of exogenous enzymes on the content of bioactive compounds in lentils and peas M. Dueñas, T. Hernández & I. Estrella Instituto de Fermentaciones Industriales, CSIC, C/ Juan de la Cierva, 3. 28006 Madrid, Spain Summary The phenolic composition of lentil (Lens culinaris) and pea (Pisum sativum) was determined by HPLC-PAD-MS. The treatment of these legumes with the enzymes, phytase, α-galactosidase, tannase and viscozyme, modified their initial composition. The change observed in each legume depended on the enzyme and the chemical structure of the phenolics. The evaluation, by reaction with DPPH, of antioxidant activity established that the treatment of lentils with tannase, α-galactosidase and viscozyme increased this activity compared with untreated lentils, but decreased it in peas. Key words: Pisum sativum, Lens culinaris, phenolics, enzymes, antioxidant activity Introduction The use of commercial enzymes in the food industry has been recognised as a useful tool to improve the nutritional quality of legume seed. This process can decrease or eliminate antinutritional factors (ANFs) (Frías et al., 2003a,b), improve the nutritional value of the legume seed (Granito et al., 2002), and modify their content of bioactive compounds such as phenolic compounds.
Phenolic compounds are considered to be bioactive and have health giving properties, which may prevent the development of diseases such as atherosclerosis and cancer (Kahkonen et al., 1999). They may also act as protective factors against oxidative damage (Jovanovic et al., 1996; Castillo et al., 2000) to prevent disease in humans.
The aim of this work was to study the modification of the phenolic composition of lentil and pea flours as a result of adding exogenous enzymes such as α-galactosidase, phytase, viscozyme and tannase, and to evaluate the antioxidant activity, as a parameter for improving the nutritional quality of these legume flours. Materials and methods Legumes Lens culinaris cv. Pardina and Pisum sativum cv. Esla were ground in a ball mill and sieved to collect the 0.050 - 0.250 mm fraction. Enzymes The commercial enzymes, α-galactosidase, phytase, viscozyme were obtained from Novo Nordisk, and tannase from Juelich Enzyme Products.

312
Treatment with commercial enzymes Lentil and pea flours (300 g) were suspended in distilled water (3,000 ml) at pH 5.5, adjusted with 300 ml of a buffer solution (acetic acid/sodium hydroxide, 0.1 N). The suspensions were incubated in a stirred fermentor (MicroFerm Fermentor, MF-100, New Brunswick Scientific, USA), with the commercial enzymes at 37 ºC, under optimal concentration and incubation times, which had been established for each of them (Table 1). Under these conditions the assays were also carried out without enzymes (control). Table 1. Optimal conditions of enzymes to act on lentil and pea flour. Enzyme Phytase α-Galactosidase Viscozyme Tannase Incubation time (min) 60 90 601 120 Concentration (mg/g substrate) 1.60 1.16 1.50 0.50 1The incubation time of pea flour for the viscozyme treatment was 18 h; the other conditions were as for lentil flour. Analysis of phenolic compounds The samples were centrifuged and the residue was freeze-dried. Dueñas et al. (2002) extracted and analysed the phenolic compounds by HPLC-PAD and HPLC-MS (ESI). Antioxidant activity Antioxidant activity (IC50) was determined by the Brand-Williams et al. (1995) method with 2,2’diphenil-1-picrihidrazil (DPPH). Analysis of variance and comparison of treatment means (LSD, 5% level) were performed using Statgraphics Plus 5.0 v. (Graphics Software System, Rockville, Md). Results and discussion Effect of treatment of lentil flour with commercial enzymes There were qualitative and quantitative differences in the phenolic compound composition of the control, raw, and enzyme treated samples. Hydroxybenzoic and hydroxycinnamic compounds, glycosides of flavonols and flavones, various dimers and trimers of procyanidins and prodelphinidins, and the stilbene, resveratrol 3-O glucose were identified (Table 2) in the raw lentil flour. Some of these are reported in lentil for the first time.
There was a general decrease in the concentration of phenolic compounds in the raw lentil samples; this was probably due to the effect of endogenous enzymes in the raw seed. However, the commercial enzymes modified the phenolic compound composition compared to the corresponding control.
The hydroxybenzoic and hydroxycinamic compounds decreased, except protocatechuic acid that increased 47 % in the tannase treatment. The presence of the acid and the aldehyde gallic is remarkable, as they were not detected in raw lentil samples.
The glycosides of flavonols and flavones decrease in the control samples, except quercetin 3-O rutinoside and luteolin, which increased in all of them. The action of the α-galactosidase, phytase or viscozyme, decreased these compounds compared with the controls. However, in the presence of tannase they increased by 38 % and 77 % respectively.

313
Table 2. Concentration of phenolic compounds (µg/g) in raw lentil. Compound µg/g Compound µg/g Trimer PD (2GC-C) 31.05 ± 1.56 Trimer PC3 1.61 ± 0.23 Protocatechuic acid 1.45 ± 0.03 (-)-Epicatechin 4.17 ± 0.69 Dimer PD1 (GC-C) 8.03 ± 0.98 trans-p-Coumaric acid 5.74 ± 0.45 Dimer digallate PC 1.26 ± 0.15 Trimer PC4 1.55 ± 0.06 Dimer PD2 (GC-C) 6.02 ± 1.00 cis-p-Coumaric acid 0.73 ± 0.09 p-Hydroxybenzoic acid 3.25 ± 0.18 trans-Ferulic acid 0.74 ± 0.07 Dimer B3 10.61 ± 1.07 trans-Resveratrol 3-O-glucose* 0.93 ± 0.09 Trimer PC1 13.16 ± 1.12 Myricetin 3-O ramnose 5.79 ± 0.44 (+)-Catechin 3-O glucose 31.50 ± 1.98 Luteolin 7-O glucose 1.29 ± 0.07 (+)-Catechin 0.77 ± 0.05 Apigenin 7-O apioglucoside 6.19 ± 0.45 trans-p-Coumaroylmalic acid* 10.02 ± 1.07 Apigenin 7-O glucose 1.87 ± 0.07 trans-p-Coumaroylglicolic acid* 2.88 ± 0.15 Luteolin glycoside 1.35 ± 0.19 Dimer B2 13.80 ± 0.96 Quercetin 3-O rutinoside 5.24 ± 0.56 Trimer PC2 0.29 ± 0.05 Luteolin 0.33 ± 0.02 *Identified for the first time in lentil. Means ± SD (n = 3). PC: Procyanidin; PD: Prodelphinidin; GC: (epi)galocatechin; C: (epi)catechin.
It is important to note the formation of the stilbene, resveratrol trans in the control
samples, and an increase in the tannase and phytase treatments that were not detected in raw lentil flour. Generally the procyanidins and prodelphinidins decreased compared with the raw lentil flour, both in the control samples and in the enzyme treatments. However, (+)-catechin 3-O glucose increased in the enzyme treatments compared with the controls. It reached its highest level in the tannase treatment.
The effect of these exogenous enzymes on phenolic compounds was different. Generally, with tannase, some phenolic compounds increased compare with raw lentil and its control. Viscozyme produced a strong decrease compared with the raw and control lentil samples. Effect of treatment of pea flour with commercial enzymes In raw pea flour hydroxybenzoic compounds were identified, together with free and conjugated hydroxycinnamic compounds (Table 3) some of these are reported for the first time in pea. Table 3. Concentration of phenolic compounds (µg/g) in raw pea. Compound µg/g Compound µg/g Protocatechuic acid 2.13 ± 0.15 trans-p-Coumaric acid 0.64 ± 0.09 p-Hydroxibenzoic acid 1.20 ± 0.08 cis-p-Coumaric acid 0.54 ± 0.07 p-Hydroxibenzaldehyde 0.34 ± 0.02 trans-Feruloylmalic acid* 0.91 ± 0.10 Trans-p-Coumaroylmalic acid* 0.51 ± 0.09 *Identified for the first time in pea. Means ± SD (n = 3).
Hydroxybenzoic and hydroxycinnamic compounds decreased in all enzyme treatments.
The decrease was greatest in hydroxycinnamic compounds. Trans ferulic acid was present in the control samples and after enzyme treatment, but it was not detected in raw pea flour. The

314
action of each enzyme was different, and the viscozyme treatment yielded the highest concentration of free and conjugated hydroxycinnamic compounds. Antioxidant activity In the lentil flour antioxidant capacity was increased after enzyme treatment with viscozyme, α-galactosidase, and tannase. It was higher than in the pea flours in which it decreased with all of the enzyme treatments (Table 4). The higher activity in lentils is explained by their higher and different content of phenolic compounds than are present in peas. Table 4. Antioxidant activity (IC50) of raw lentil and pea flour and after treatment with
enzymes. Samples Raw Phytase Viscozyme α-Galactosidase Tannase Lentil 3.16 ± 0.10c 3.41 ± 0.07d 2.16 ± 0.04a 2.29 ± 0.05a 2.50 ± 0.08b
Pea 22.93 ± 1.00a 96.77 ± 1.12e 80.71 ± 1.07d 76.26 ± 1.23c 66.99 ± 0.98b
A smaller IC50 value corresponds with higher antioxidant activity. Means ± SD (n = 3); means followed by a different letter in a row are significantly different (LSD P < 0.05).
Free radical scavenging capacity (IC50) seems to be influenced not only by changes in the phenolic composition of lentil and pea, but also by other antioxidants present in the seed. Conclusions Under the conditions of these assays the contact of the legume flour with an aqueous solution, such as the buffer (control samples), modified legume phenolic composition by the action of their endogenous enzymes. After enzyme treatment, the phenolic composition of both lentil and pea flour was modified by the action of both, endogenous and exogenous enzymes, but in different ways depending on the legume matrix. Phytase, α-galactosidase, tannase and viscozyme act in a very different ways based on the structure of the phenolic compounds produced.
Taking the increase in antioxidant activity as a positive result, the addition of enzymes such as tannase, α-galactosidase and viscozyme could be an effective process for improving the functionality of the cv. Pardina lentil flour. However, it was a negative process in the pea cv. Esla. References Brand-Williams, W., Cuvelier, M.E. & Berset, C. 1995. Use of a free radical method to
evaluate antioxidant activity. Lebensmittel-Wissenschaft und-Technologie 28, 25-30. Castillo, J, Benavente-García, O., Lorente, J., Alcaraz, M., Redondo, A., Ortuño, A. & Del
Rio, J.A. 2000. Antioxidant activity and radioprotective effects against chromosomal damage induced in vivo by X-rays of flavan-3-ols (procyanidins) from grape seeds (Vitis vinifera): comparative study versus other phenolic and organic compounds. Journal of Agricultural and Food Chemistry 48, 1738-1745.
Dueñas ,M., Hernández, T. & Estrella, I. 2002. Phenolic composition of the cotyledon and the seed coat of lentils (Lens culinaris L). European Food Research and Technology 215, 478-483.

315
Frías, J., Doblado, R., Antezana, J.R. & Vidal-Valverde, C. 2003b. Inositol phosphate degradation by the action of phytase enzyme in legume seeds. Food Chemistry 81, 233-239.
Frías, J., Doblado, R. & Vidal-Valverde, C. 2003a. Kinetics of soluble carbohydrates by action of endo/exo α-galactosidase enzyme in lentils and peas. European Food Research and Technology 216, 199-203.
Granito, M., Frías, J., Doblado, R., Guerra, M., Champ, M. & Vidal-Valverde, C. 2002. Nutritional improvement of beans (Phaseolus vulgaris) by natural fermentation. European Food Research and Technology 214, 226-231.
Jovanovic, S.V., Steenken, S., Hara, Y. & Simic, M.G. 1996. Reduction potentials of flavonoid and model phenoxyl radicals. Which ring in flavonoids is responsible for antioxidant activity? Journal of the Chemical Society, Perkin Transactions 2, 2497-2504.
Kahkonen, M.P., Hopia, A.I., Vuorela, H.J., Rauha, J.P., Pihlaja, K., Kujala, T.S. & Heinonen, M. 1999. Antioxidant activity of plant extracts containing phenolic compounds. Journal of Agricultural and Food Chemistry 47, 3954-3962.


317
Content and distribution of vicine, convicine and L-DOPA through out germination and seedling growth of Vicia faba L. seeds. C. Goyoaga, C. Cuadrado, M.M. Pedrosa, E. Guillamón, P. Altares, M. Muzquiz & C. Burbano SGIT-INIA, Departamento Tecnología de Alimentos, Carretera de la Coruña Km 7.5, 28040, Madrid, Spain Summary This work examined the content and distribution in cotyledons pairs and the embryo axis of the pyrimidine glucosides and L-DOPA by high performance liquid chromatography (HPLC) in raw and germinated seed and seedling of Vicia faba cvs Alameda and Brocal. The results indicated that both are a similar but the of vicine and convicine content was higher in cv. Alameda. In the cotyledons pair, the vicine and convicine content decreased and L-DOPA was not found. In the embryo axis pyrimidine glucosides increased slightly and L-DOPA content increased rapidly and had increased 1,000 fold by the end of germination. A possible role for L-DOPA in treatment of Parkinson’s disease is discussed. Key words: Vicia faba, vicine, convicine, L-DOPA, pyrimidine glucosides Introduction Faba bean (V. faba) is cultivated widely in many countries of the old world. It has a major role in traditional diets of Mediterranean, Indian, Chinese, English, Middle Eastern, African and South American people. It is an excellent source of protein, complex carbohydrates, dietary fibre, choline, minerals and secondary metabolites such as phenolics and levo dihydroxy phenylalanine (L-DOPA). The latter is a precursor of the neurotransmitter dopamine, and occurs naturally in faba bean seedlings, green pods and seeds. The beans are used in the treatment of Parkinson’s disease, hypertension, renal failure and cirrhosis of the liver (Rabey et al., 1993; Randhir et al., 2002).
Nutritional studies showed that faba beans also contain antinutritional (ANFs) and/or toxic factors (Olaboro et al., 1981). Among the ANFs are the pyrimidine glucosides, vicine and convicine. These two compounds and their respective aglycons are implicated as the causative agents of favism. Favism is a metabolic disease that causes haemolytic anaemia in glucose-6-phosphate dehydrogenase deficient individuals who eat faba beans (Mager et al., 1980). However, vicine and convicine also appear to have beneficial properties including the prevention of cardiac arrhythmia and, under certain conditions, can inhibit the growth of the malaria parasite (Marquardt, 1989). The vicine or convicine concentration in faba bean is influenced by stage of maturity, environmental factors, and genetic variation.
The aim of this work was to determine the content and the distribution of vicine, convicine and L-DOPA in the cotyledon pair and the embryo axis during germination and the seedling growth of faba beans to obtain information that may help in the treatment of Parkinson’s disease.

318
Material and methods Plant Material Seeds of V. faba cvs Alameda y Brocal were obtained from CIFA, Córdoba (Spain). The seeds were screened for imperfection and 40 faba beans were placed per tray with sand/water (5:1 w/v) and were maintained in a constant environment chamber with a photoperiod of 8 h light/d at 20 ºC. There were two replicates. Seeds were removed after 12, 24, 48, 72, 96, 120, 144, 168, 192, and 216 h post-sowing. Ungerminated seeds (0 h) and seedlings were subdivided into testa, cotyledon and axis fractions. All samples were weighed individually and then frozen and freeze-dried. Samples were ground to pass through a 100-mesh sieve (Ciclotec 1093 Sample Mill; Tecator). Chemical analysis The concentration of vicine, convicine and L-DOPA in the plant parts was determined by HPLC (Marquardt & Fröhlich, 1981). Samples were extracted with 5 % perchloric acid (0.01 w/v) for 1 min. using an Ultraturrax homogenizer (T25 basic, IKA) at 4 ºC. The extract was centrifuged and filtered thought a Millipore filter (0.45 µm) to remove any suspended material prior to injection into the HPLC. A reverse phase Spherisorb ODS column was used (250 x 4.6 mm, 5 µm) with 0.05 M ammonium phosphate buffer (pH 2.0) as mobile phase. A Beckman System Gold HPLC machine with an absorbance detector (detection wavelength 280 nm) was used.
Dr R. Marquardt (University of Manitoba, Canada) provided purified reference samples of vicine and convicine and the L-DOPA standard was from Serva. Calibration curves were drawn for the three compounds. There was a linear response in the range 8.6 - 310 µg/ml for vicine, 3.7 - 134.0 µg/ml for convicine and 47.0 - 567.0 µg/ml for L-DOPA. Results and discussion Changes in dry matter, and the vicine, convicine and L-DOPA distribution during germination Cotyledons pair: During seedling development the cotyledons pair dry weight fell significantly. The greatest reduction was after 216 h of germination and was 30.5 % in cv. Alameda and 34.4 % in cv. Brocal (Fig. 1). The results obtained for the analysis of pyrimidine glucosides indicate that the initial amounts of vicine in cv. Alameda (3.55mg/cotyledon pair) and cv. Brocal (2.71 mg/cotyledon pair) were higher than those of convicine in both cultivars (2.11 mg/cotyledon pair and 1.32 mg/cotyledon pair respectively). The level of vicine was almost constant during the first 48 h in cv. Alameda and during the first 72 h in cv. Brocal. Afterwards it slowly declined as germination progressed (1.76 mg/cotyledon pair in cv. Alameda and 1.07 mg/cotyledon pair in cv. Brocal at the end of the process). The amount of convicine slowly and gradually decreased (0.72 mg/cotyledon pair in cv. Alameda and 0.42 mg/cotyledon pair in cv. Brocal at 216 h). The changes observed in these compounds run parallel to those in dry weight. We did not detected L-DOPA in germinated cotyledons.

319
Figure 1. Changes in dry weight and distribution of vicine and convicine per cotyledon pair of
V. faba cvs Alameda and Brocal. Embryo axis: The dry weight of the axis increased rapidly to 9 d in both cultivars of V. faba, reaching at the end of germination final levels 30 times the initial weight. In the axis, the amount of vicine was higher than convicine. The initial amounts of these pyrimidine glucosides were lower than in the cotyledons with a slight increment during germination. However, the L-DOPA level (approximately 0.015 mg/embryo axis in the ungerminated seed of both cultivars) had increased 1,000 times after 216 h (Fig. 2).
Figure 2. Changes in dry weight and distribution of vicine, convicine and L-DOPA in the
embryo axis of V. faba cvs Alameda and Brocal. Whole plant: The dry weight of the whole plant (cotyledon pair, embryo axis and testa) showed almost no variations during germination. There was a slight variation in the pyrimidine glucoside content (Fig. 3). This could be due to translocation of both compounds from the cotyledons to axis, as suggested by Griffiths & Ramsay (1996). The large increase in L-DOPA in the whole plant was parallel to the increase in embryo axis. It was probably due to de novo synthesis during seedling growth. Ramsay & Griffiths (1996) reported similar findings.
Clinical studies showed that consumption of unsprouted faba bean is effective in the treatment of Parkinson’s disease. However, large quantities are required to obtain the desired amount of L-DOPA and they caused flatulence in the patients (Apaydin, 2000). However, previous studies in our laboratory (Goyoaga et al., 2000) showed that a 48 h germination drastically reduced the flatulence factors (raffinose family oligosaccharides) in faba beans.
cv. Brocal
0
2
4
6
0 12 24 48 72 96 120 144 168 196 216 hours
mg
gluc
osid
e
0
200
400
600
800
1000
dry
wei
ght (
mg)
vicine convicine dry weight
cv. Alameda
0
2
4
6
0 12 24 48 72 96 120 144 168 196 216 hours
mg
gluc
osid
e
0
200
400
600
800
1000
dry
wei
ght (
mg)
vicine convicine dry weight
cv. Alameda
0
5
10
15
20
0 24 48 72 96 120 144 168 196 216
mg
com
poun
d
0
50
100
150
200
250
dry
wei
ght (
mg)
vicine convicineL-DOPA dry weight
cv. Brocal
0
5
10
15
20
0 24 48 72 96 120 144 168 196 216
mg
com
poun
d
0
50
100
150
200
250
dry
wei
ght (
mg)
vicine convicineL-DOPA dry weight
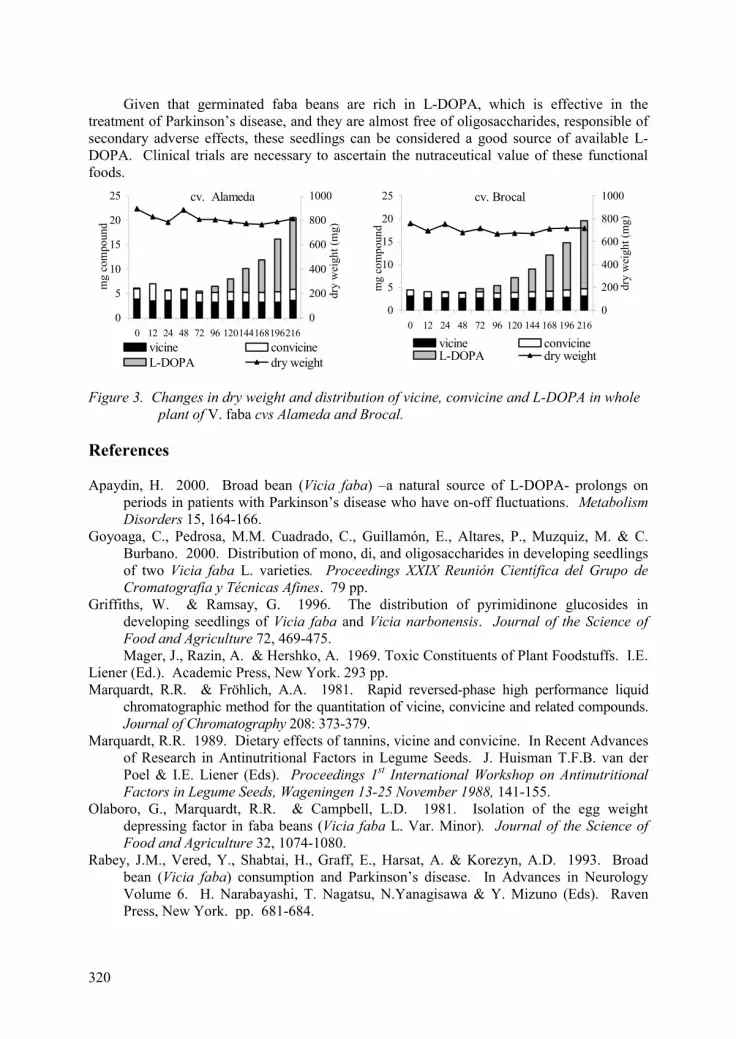
320
Given that germinated faba beans are rich in L-DOPA, which is effective in the treatment of Parkinson’s disease, and they are almost free of oligosaccharides, responsible of secondary adverse effects, these seedlings can be considered a good source of available L-DOPA. Clinical trials are necessary to ascertain the nutraceutical value of these functional foods.
Figure 3. Changes in dry weight and distribution of vicine, convicine and L-DOPA in whole
plant of V. faba cvs Alameda and Brocal. References Apaydin, H. 2000. Broad bean (Vicia faba) –a natural source of L-DOPA- prolongs on
periods in patients with Parkinson’s disease who have on-off fluctuations. Metabolism Disorders 15, 164-166.
Goyoaga, C., Pedrosa, M.M. Cuadrado, C., Guillamón, E., Altares, P., Muzquiz, M. & C. Burbano. 2000. Distribution of mono, di, and oligosaccharides in developing seedlings of two Vicia faba L. varieties. Proceedings XXIX Reunión Científica del Grupo de Cromatografía y Técnicas Afines. 79 pp.
Griffiths, W. & Ramsay, G. 1996. The distribution of pyrimidinone glucosides in developing seedlings of Vicia faba and Vicia narbonensis. Journal of the Science of Food and Agriculture 72, 469-475. Mager, J., Razin, A. & Hershko, A. 1969. Toxic Constituents of Plant Foodstuffs. I.E.
Liener (Ed.). Academic Press, New York. 293 pp. Marquardt, R.R. & Fröhlich, A.A. 1981. Rapid reversed-phase high performance liquid
chromatographic method for the quantitation of vicine, convicine and related compounds. Journal of Chromatography 208: 373-379.
Marquardt, R.R. 1989. Dietary effects of tannins, vicine and convicine. In Recent Advances of Research in Antinutritional Factors in Legume Seeds. J. Huisman T.F.B. van der Poel & I.E. Liener (Eds). Proceedings 1st International Workshop on Antinutritional Factors in Legume Seeds, Wageningen 13-25 November 1988, 141-155.
Olaboro, G., Marquardt, R.R. & Campbell, L.D. 1981. Isolation of the egg weight depressing factor in faba beans (Vicia faba L. Var. Minor). Journal of the Science of Food and Agriculture 32, 1074-1080.
Rabey, J.M., Vered, Y., Shabtai, H., Graff, E., Harsat, A. & Korezyn, A.D. 1993. Broad bean (Vicia faba) consumption and Parkinson’s disease. In Advances in Neurology Volume 6. H. Narabayashi, T. Nagatsu, N.Yanagisawa & Y. Mizuno (Eds). Raven Press, New York. pp. 681-684.
cv. Alameda
0
5
10
15
20
25
0 12 24 48 72 96 120144168196216
mg
com
poun
d
0
200
400
600
800
1000
dry
wei
ght (
mg)
vicine convicineL-DOPA dry weight
cv. Brocal
0
5
10
15
20
25
0 12 24 48 72 96 120 144 168 196 216
mg
com
poun
d
0
200
400
600
800
1000
dry
wei
ght (
mg)
vicine convicineL-DOPA dry weight

321
Ramsay, G. & Griffiths, D.W. 1996. Accumulation of vicine and convicine in Vicia faba and V. narbonensis. Phytochemistry l42, 63-67.
Randhir, R., Shetty, P. & Shetty, K. 2002. L-DOPA and total phenolic stimulation in dark germinated fava bean in response to peptide and phytochemical elicitors. Process Biochemistry 37, 1247-1256.


323
Detoxification of rapeseed meal by controlled instantaneous pressure drop J. Haddad & K. Allaf Laboratoire Maîtrise des Technologies Agro-Industrielles LMTAI, Université de La Rochelle, Avenue Michel Crepeau, 17042 La Rochelle cedex 01, France Summary The suitability of rapeseed (Brassica napus) as a protein supplement for human consumption and animal feed is limited due to toxicity problems caused by the presence of antinutritional factors (ANFs) such as glucosinolates. There are a number of procedures for removing or decreasing the quantity of glucosinolates and their hydrolysis products, but all these processes have major drawbacks such as protein loss, incomplete inactivation, cost and a lack of economic viability. In this study, controlled instantaneous pressure drop (Détente Instantanée Contrôlée DIC) treatment was evaluated on rapeseed meal. The results demonstrate the efficiency of DIC as a new process for the detoxification of rapeseed meal. Keywords: Brassica napus, rapeseed, glucosinolates, processing, controlled instantaneous
pressure drop Introduction Rapeseed is an important source of vegetable oil, with an oil content of 40 % to 46 % (Mohtadi-Nia et al., 1986). Numerous studies have aimed at obtaining rapeseed proteins for human and animal food. However, the use of rapeseed has been limited by the presence of ANFs and toxic substances such as glucosinolates and phytates. The genetic selection of varieties with a low glucosinolate content was the first approach to this problem, one example being the ‘00’ varieties. However, the concentration, in seed, of substances harmful to thyroid function in these varieties is still too high for human consumption (Bau et al., 1991).
The threshold fixed by the EU for glucosinolate sulphur compounds is 35 µmole/g of product at 9 % moisture (38.5g/100 g DM) (Bureau & Evrard, 1992). Mansour et al. (1993), Barrett et al. (1997) and Lucht (1998), and others have discussed the thermo-labile nature of the glucosinolates. This is why heat treatment, such as cooking, is frequently used to increase the nutritional quality of rapeseed meal. The aim of this work was to evaluate the effect of a the DIC process on the glucosinolate content of rapeseed. Materials and methods Raw material preparation A variety of rapeseed with a glucosinolate content of 23.11 µmole/g was used. The seed was moistened by grinding with water. It was then homogenised in polyethylene bags for at least 12 h at 4 °C before the DIC treatment.

324
Controlled instantaneous pressure drop Controlled instantaneous pressure drop (DIC) is distinguished by its capacity to provide a well-controlled hydro-thermo-mechanical treatment in terms of temperature (temperature up to 170 °C) and pressure (absolute steam pressure up to 8 bars).
The treatment duration is determined by a drop, generally instantaneous (< 0.1 s), to a low absolute pressure of about 50 millibars. The pressure drop generates, by self-vaporisation of water from the product, micro-mechanical constraints which lead to a modification of the product’s texture and results in a porous structure. The length of the operation (generally a few seconds) and the immediate decrease in temperature following the pressure drop stops all thermal deterioration and ensures conservation of quality in the end product (Haddad et al., 2001). DIC reactor The DIC machine has a capacity of 300 g. It consists of three main parts: 1) a processing chamber, in which the product is treated at high temperature and pressure (steam pressure); 2) a large vacuum reservoir (50 times the volume of the processing chamber) and 3) a large diameter valve (> 200 mm) that can be opened rapidly (< 0.1 s). Connection between the processing chamber and the vacuum tank is obtained by rapidly opening the valve. A measuring system with numerous pressure and temperature sensors, linked to a personal computer, ensures data collection and the manual and/or automatic control of the system (Fig. 1).
Figure 1. Diagrammatic layout of the DIC reactor (1: boiler; 2: DIC reactor; 3:
decompression valve; 4: vacuum tank; 5: vacuum pump; 6: condensation tank). Experimental design The operating parameters for the DIC treatments were: steam pressure (P) between 3 - 7 bars, treatment time (t) between 20 - 60 s, and initial water content (W) of between 30 - 50 g water/100 g DM. A central composite design of 22 points was used. A reference point representing the raw non-treated material was used. The results were statistically analysed using Windows Statgraphic/PC 4.1 software. The limit of significance between differences was set at P < 0.05 by Fisher’s test. Analysis The glucosinolate content was measured by high performance liquid chromatography (HPLC) using the NF EN ISO 9167-1 method.
1
23
5
6
4

325
Results and discussion Effect of DIC on glucosinolate content Depending on the operating parameters used, the reduction in glucosinolate content can be up to 40 % at a pressure of 7 bars, a processing time of 60 s and an initial water content of 30 % (sample number 13). This value compares favourably with literature values but has the advantage that DIC treatment only takes about 60 s (Mansour et al., 1993; Barrett et al., 1997; Lucht, 1998; Pal Vig & Walia, 2001). Table 1. Glucosinolate (GSL) (µmol/g DM) present in rapeseed after DIC processing. Sample numbe
r
Pressure (bar)
Time (s)
Moisture (%)
GSL (µmol/g)
Sample number
Pressure(bar)
Time (s)
Moisture (%)
GSL (µmol/g)
1 5 40 40 19.2 13 7 60 30 13.9 2 1.6 40 40 21.3 14 5 6 40 21.2 3 5 40 40 18.9 15 5 40 40 18.9 4 5 40 40 18.3 16 7 20 30 17.8 5 5 40 40 19.2 17 5 40 57 18.0 6 8 40 40 15.5 18 3 60 30 20.4 7 5 74 40 17.6 19 3 20 50 21.4 8 7 60 50 14.0 20 3 20 30 22.4 9 5 40 23 19.0 21 3 60 50 21.2 10 5 40 40 18.9 22 5 40 40 18.7 11 5 40 40 19.2 Control - - - 23.1 12 7 20 50 18.7
Figure 2 shows the effect of DIC treatment on glucosinolate content as a Pareto chart.
Within the domain of variation of the parameters chosen and for an α value of 5 %, the simple effects of pressure and processing time, as well as the effect of the interaction of pressure by processing time are significant.
Figure 2. Effect of DIC treatment on glucosinolate content in rapeseed.
For a better illustration of the DIC effect, a response surface representation was used (Fig. 3). The most marked decrease in glucosinolate content corresponds to higher values of steam pressure and treatment duration. Thus, to reduce the glucosinolate content of rapeseed, a short DIC treatment of a few seconds under high steam pressure should be sufficient.
0 3 6 9 12 15Standardized effect
C:Initial water contentBCACCCBBAAAB
B:Treatment durationA:Pressure

326
Figure 3. Response surface for glucosinolate content versus pressure and treatment
duration. Conclusions Although legumes and oilseeds are good sources of proteins, lipids and other essential nutrients their main drawback is their poor digestibility due to their high ANF content. To reduce the ANF content and obtain better utilisation of rapeseed the DIC treatment was studied. The reduction in glucosinolates after DIC treatment is similar to values of other workers (40 % maximum). However, DIC treatment is distinguished by a short processing time. These methods could easily be converted to an industrial scale. References Barret J.E., Klopfenstein C.F. & Leipold H.W. 1997. Detoxification of rapeseed meal by
extrusion with an added basic salt. Cereal Chemistry 74, 168-170. Bau, H.M., Villaume, C., Chandrasiri, V., Mejean L. & Nicolas, J-P. 1991. Process for
detoxifying substances of vegetable origin and foodstuffs obtained by means of this process. Brevet No FR2654585.
Bureau, J. & Evrard. J. 1992. Les graines entières de colza: une utilisation qui grimpe. Revue de L’alimentation Animale 460, 59-60.
Haddad, J., Louka, N., Gadouleau, M., Juhel, F. & Allaf, K. 2001. Application du nouveau procédé de séchage/texturation par Détente Instantanée Contrôlée DIC aux poissons: Impact sur les caractéristiques physico-chimiques du produit fini. Science des Aliments 21, 481-498.
Lucht, H.W. 1998. Reduction of glucosinolates and sinapine in rapeseed by technical treatment: Description of the technical procedure and effectiveness evaluation. In Recent Advances of Research in Antinutritional Factors in Legume Seeds and Rapeseed. A.J.M. Jansman, G.D. Hill & A.F.B. van der Poel (Eds.). Proceedings 3rd International Workshop on Antinutritional Factors in Legume Seeds and Rapeseed, Wageningen, 8-10 July 1998, 433-435.
Mansour, E.H., Dworschak, E., Lugasi, A., Gaal, O., Barna, E. & Gergely A. 1993. Effect of processing on the antinutritive factors and nutritive value of rapeseed products. Food Chemistry 47, 247-252.
Mohtadi-Nia, D.J., Bau, H.M., Giannangeli, F., Mejean, L. & Debry, G. 1986. Valorisation nutritionnelle des protéines de colza par un traitement hydro-thermique des graines. Canadian Institute of Food Science and Technology Journal 19, 95-103.
Estimated Response SurfaceInitial water content = 40%
Pressure (bar)Treatment duration (s)
Glu
cosi
nola
tes (
µmol
e/g
d.m
)
3 4 5 6 7 20 30 40 50 6014
16
18
20
22

327
Pal Vig A. & Walia A. 2001. Beneficial effects of Rhizopus oligosporus fermentation on reduction of glucosinolates, fibre and phytic acid in rapeseed (Brassica napus) meal. Bioresource Technology 78, 309-312.


329
The inactivation of soybean trypsin inhibitors by hydro-thermo-mechanical treatment J. Haddad & K. Allaf Laboratoire Maîtrise des Technologies Agro-Industrielles LMTAI, Université de La Rochelle, Avenue Michel Crepeau, 17042 La Rochelle cedex 01, France Summary Trypsin inhibitor activity (TIA) is one of the most important anti-nutritional factors in grain legumes. It decreases the utilisation of protein in legumes and therefore reduces the utility of grain legumes in human and animal nutrition. Heat processing of soybean (Glycine max) has proved to be an effective method to eliminate, or at least considerably reduce, the activity of antinutritional factors (ANFs). This paper deals with the effect of a new process called Détente Instantanée Contrôlée (DIC) on soybean trypsin inhibitors. The results demonstrate the efficiency of DIC for legume treatment. Keywords: soybeans, trypsin inhibitor activity, controlled instantaneous pressure drop Introduction The main antinutritional factors (ANFs) in soybean are trypsin inhibitors, which are an important obstacle to the use of untreated soybeans in human food. However, the advantage of soybeans compared with other grain legumes is that their ANFs are proteins and are thus heat sensitive (Baker & Mustakas, 1973; Pipa, 1988; Bau et al., 2001). Raw mature soybeans are not edible so various treatments, especially involving heating, are required (Ruales et al., 1988). Since severe heating can reduce protein digestibility and amino acid availability, over-processing should be avoided (Bau et al., 2001). Pipa (1988) reported that around 1 to 1.5 TIU/mg DM of trypsin inhibitor is optimal. However, other reports suggest a reduction in trypsin inhibitor activity of 79 to 87 % would be satisfactory (Leontowicz et al., 1998). The search for a new, simple, well-controlled technology for seed treatment has become a necessity for the food industry. Controlled instantaneous pressure drop (DIC) fulfils these criteria and the process is now tightly controlled. The aim of this work was to carry out a hydro-thermo-mechanical treatment on soybeans to reduce their ANF content. Materials and methods Raw material preparation A variety of soybean with an initial trypsin inhibitor content of 41.6 IU/mg was used. The seed was moistened and ground with water and then homogenised in polyethylene bags for at least 12 h at 4 °C before the DIC treatment. Controlled Instantaneous Pressure Drop (DIC) The DIC treatment consists of placing the moistened product in a processing chamber and exposing it to steam pressure (up to 8 bars) at high temperature (up to 170 °C), over a

330
relatively short time (a few seconds to one minute). The end of the thermal treatment involves a pressure drop, which is achieved by the instantaneous opening of a valve between the treatment chamber and a vacuum tank (50 millibars) with a volume, which is at least fifty times greater. There is a sharp fall in chamber pressure, which leads to a partial vaporisation of water in the product. This produces a porous structure, depending on the properties of the treated product and the operating conditions (e.g. temperature, amplitude of the pressure drop, processing duration). Thus, DIC re-texturises biological products, improving their functional properties and makes them easier to use in industrial processes (Haddad et al., 2001).
Figure 1. Pressure-time relationship in the DIC process. Experimental design The DIC operating parameters were: a steam pressure (P) of between 3 and 7 bars, a treatment duration (t) of between 20 and 60 s and an initial water content (W) of between 30 - 50 g water/100 g DM. A central composite design of 22 points was used. A point representing the non-treated raw material was taken as a control point. The results were statistically analysed using Windows Statgraphic/PC 4.1 software and the limit of significance between differences was set at P < 0.05 by Fisher’s test. Analysis Trypsin inhibitors were determined by spectrophotometry (410 nm) according to the Ba 12-75 AOCS method using benzoyl-DL-arginine-p-nitroanalide hydrochloride (BAPA) as a substrate for the trypsin. Results and discussion Effect of DIC treatment on trypsin inhibitors The trypsin inhibitor content of the raw material in this study is in line with published values (Elkowicz & Sosulski, 1982). The reduction in level after DIC treatment confirms the thermo-sensitive nature of these compounds (Baker & Mustakas, 1973; Pipa, 1988; Bau et al., 2001). The reduction was as much as 94 % for a DIC treatment at 7 bar, with a processing time of 60 s and an initial water content of 50 % (sample number 21). This level (2.65 TIU/mg) is lower than the most restrictive values for human consumption (3 TIU/mg). Baker & Mustakas (1973), Hafez & Singh (1983) and Rackis et al. (1986) reported similar values.
P atmospheric
Pressure
Pi (Ti)
P vacuum
Time
t

331
However, the advantage of the DIC treatment is the relatively short processing time required (60 s). Table 1. Trypsin inhibitor content in DIC processed soybeans. Sample number
Pressure (bar)
Time (s)
Moisture (%)
TI (IU/mg)
Sample number
Pressure(bar)
Time (s)
Moisture (%)
TI (IU/mg)
1 1.6 40 40 15.1 13 5 40 40 4.6 2 3 20 30 17.5 14 5 40 40 4.5 3 3 20 50 12.0 15 5 40 40 4.6 4 3 60 30 4.7 16 5 40 57 4.8 5 3 60 50 5.1 17 5 74 40 3.5 6 5 60 40 18.0 18 7 20 30 5.7 7 5 40 23 5.4 19 7 20 50 5.8 8 5 40 40 4.4 20 7 60 30 3.3 9 5 40 40 4.8 21 7 60 50 2.710 5 40 40 6.3 22 8 40 40 2.811 5 40 40 4.5 Control - - - 41.612 5 40 40 3.9
Based on the objective of reducing trypsin inhibitor activity by 80 % (8.2 TIU/mg),
which is the safety limit according to Leontowicz et al. (1998) all the DIC treatments tested were efficient with the exception of those carried out under mild conditions with low steam pressure and therefore relatively low temperature and short processing times.
Impact of the operating parameters Figure 2 displays a Pareto chart of the effect of the operating parameters used during DIC treatment on trypsin inhibitor factors. Almost all of the operating parameters affect trypsin inhibitor content, within the domain of variation of the parameters chosen. For a better illustration of the DIC effect, a response surface representation was used (Fig. 3). Higher steam pressure or treatment time will result in lower soybean trypsin inhibitor content.
Figure 2. Effect of DIC treatment on trypsin inhibitor content in soybean.
Conclusions
Like other heat treatments, DIC considerably reduces the ANF levels and ensures their inactivation. One of the major advantages of DIC is its short processing time. A DIC treatment of a few minutes is more than sufficient to meet the most restrictive norms with regard to the safe limit of trypsin inhibitor activity. The possibility of treating whole seeds is another advantage of DIC in for industrial applications.
0 2 4 6 8 10
CCAC
C:Initial water contentBCAAABBB
A:PressureB:Treatment duration

332
Figure 3. Response surface for trypsin inhibitors content (T.I.): variation with pressure and
treatment duration. References Baker, E.C. & Mustakas, G.C. 1973. Heat inactivation of trypsin inhibitor, lipoxygenase
and urease in soybeans: effect of acid and base additives. Journal of the American Oil Chemists Society 50, 137-141.
Bau, H.-M., Villaume, C., Giannangeli, F., Nicolas, J-P. & Mejean, L. 2001. Optimisation du chauffage et valeurs nutritionnelle et fonctionnelle des protéines de soja. Cahiers de Nutrition et de Diététique 36, 96-102.
Elkowicz, K. & Sosulski, F.W. 1982. Antinutritive factors in eleven legumes and their air-classified protein and starch fractions. Journal of Food Science 47, 1301-1304.
Haddad, J., Louka, N., Gadouleau, M., Juhel, F. & Allaf, K. 2001. Application du nouveau procédé de séchage/texturation par Détente Instantanée Contrôlée DIC aux poissons: Impact sur les caractéristiques physico-chimiques du produit fini. Science des Aliments 21, 481-498.
Hafez, Y.S. & Singh, G. 1983. Effects of microwave heating on nutritional quality of soybeans. Nutrition Reports International 28, 413-421.
Leontowicz, H., Kostyra H., Leontowicz, M. & Kulasek, G.W. 1998. The inactivation of legume seed haemagglutinin and trypsin inhibitors by boiling. In Recent Advances of Research in Antinutritional Factors in Legume Seeds and Rapeseed. A.J.M. Jansman, G.D. Hill & A.F.B. van der Poel (Eds). Proceedings 3rd International Workshop on Antinutritional Factors in Legume Seeds and Rapeseed, Wageningen, 8-10 July 1998, 429-432.
Pipa, F. 1988. Traitement hydro-thermique des matières premières. In Les Traitements Hydro-Thermiques des Matières Premières. Tecaliman, Nantes. pp. 59-69.
Rackis, J.J., Wolf, W.J. & Baker, E.C. 1986. Protease inhibitors in plant foods: content and inactivation. Advances in Experimental Medicine and Biology 199, 299-347.
Ruales, J., Polit, P. & Nair, B.M. 1988. Nutritional quality of blended foods of rice, soy and lupins, processed by extrusion. Food Chemistry 29, 309-321.
Estimated Response SurfaceInitial water content = 40%
Pressure (bar)Treatment duration (s)
T. I.
(IU
/g p
rodu
ct)
3 4 5 6 7 20 30 40 50 600369
121518
(X 1000)

333
Changes in phytate content of Lupinus albus and L. mutabilis seed during controlled instantaneous pressure drop treatment J. Haddad1, R. GreinerR2 & K. Allaf1
1Laboratoire Maîtrise des Technologies Agro-Industrielles LMTAI, Université de La Rochelle, Avenue Michel Crepeau, 17042 La Rochelle cedex 01, France 2Federal Research Centre for Nutrition, 76131 Karlsruhe, Germany Summary Although legumes provide health benefits, they also contain antinutritional factors (ANFs) like phytate. During food processing and digestion, phytate can be dephosphorylated to produce degradation products, such as myo-inositol pentakis-, tetrakis-, tris-, bis-, and monophosphates. Excessive amounts of phytate in the diet can lead to mineral deficiencies and phytate should be eliminated by processing. This paper reports the effect of controlled instantaneous pressure drop (Détente Instantanée Contrôlée (DIC)) treatment on the phytate content of Lupinus albus and L. mutabilis seed. The results show the potential of the DIC process for the treatment of lupin seed. Keywords: Lupinus albus, L. mutabilis phytate, anti-nutrients, controlled instantaneous
pressure drop, processing Introduction Lupin seed has one of the highest protein levels among the grain legumes. Lupins can grow in areas where the soybean cannot. However, a major drawback is the presence of heat-resistant ANFs like phytates. Phytic acid or inositolhexa-phosphoric acid (IHP) is very common in plants, in the form of salts (phytate or phytine) or in complexes with proteins. It reduces mineral availability and hinders zinc and iron absorption in humans (Greiner, 2001). According to some authors, phytates can be partially eliminated by heat treatment (Mohamed et al., 1986). De Boland et al. (1975) found 30 min steaming reduced the phytate content by 10 %.
Processes that are currently used for treating lupins aim at producing lupin protein concentrates or isolates. These are expensive, need heavy machinery and are mostly of interest where the aim of the process is to produce a high added value product, such as the functional protein isolates. However a new, simple, well-controlled technology for seed treatment is necessary for the food industry. Controlled instantaneous pressure drop (DIC) fulfils these criteria and the process is now tightly controlled. The aim of this work was to apply DIC treatments to L. albus and L. mutabilis lupin seed to improve their nutritional value by reducing their phytate content.

334
Materials and methods Raw material preparation The L. albus and L. mutabilis seed used had an initial phytate content of 16.95 and 22.68 mg/g DM respectively. Lupin seed was cracked, dehulled, moistened, and treated by DIC followed by hot air drying, at 40 °C to obtain a final seed moisture of 5 % (g/100 g DM). Controlled instantaneous pressure drop Controlled instantaneous pressure drop is a hydro-thermo-mechanical treatment for biological materials. Like extrusion cooking, DIC combines heat with simultaneous mechanical action. This increases the availability of some seed constituents such as starch and lipids in the intestine. Compared with extrusion cooking, the main advantage of the use of DIC for legume seed treatment is the possibility of treating and maintaining whole seeds as the end product. This permits the use of the seed in various culinary preparations.
In the DIC process, in which temperature and processing time are highly controlled, the end product has a porous texture and thus has better functional properties (Haddad et al., 2001). DIC parameters The operating parameters for the DIC treatment were: steam pressure (P) 7 bars, initial water content (W) at 50 % for L. albus and 30 % L. mutabilis while the treatment time (t) under high pressure and high temperature was varied from 1 to 7 minutes to study the kinetics of the thermal destruction of phytates. Analysis Phytate was determined by a combination of AOAC method No 986.11 and Sandberg & Ahderinne (1986). Results and discussion The effect of DIC on seed phytate content Lupinus albus: The results for processing times of 1 to 7 min are shown in Table 1. The myo-inositol hexakisphosphate (IP6) content decreased and the pentakis- (IP5), tetrakis- (IP4) and trisphosphate (IP3) compounds increased with increased processing time. Table 1. The effect of treatment time (min.) with controlled instantaneous pressure drop
(DIC) on the phytate content of L. albus seed (Pressure 7 bar, moisture content 50 %).
DIC Treatment time (min) 0 1 2 3 4 5 6 7 IP6 15.88 8.09 4.39 3.43 3.02 2.71 1.82 1.39 IP5+IP4+IP3 1.07 7.24 7.86 7.90 7.60 7.12 6.72 6.32 Total (mg/g DM) 16.95 15.33 12.25 11.33 10.63 9.84 8.53 7.72

335
The IP6 is dephosphorylated under thermal or enzymatic action and is transformed into IP5, IP4 and IP3. These forms are less harmful and bind minerals more weakly than IP6. The reduction in IP6 on one hand and the increase in IP5, IP4 and IP3 on the other hand depended on treatment duration.
The maximum total phytate (IP6 + IP5 + IP4 + IP3) reduction content was about 55 % (Fig. 1). This is greater than values reported by de Boland et al. (1975) and Chango et al. (1993).
Figure 1. The kinetics of the thermal destruction of phytates in L. albus and L. mutabilis seed
by controlled instantaneous pressure drop. Lupinus mutabilis: Table 2 shows that, as with L. albus, the IP6 content decreased while the IP5, IP4 and IP3 content increased with increased processing time. There was a 60 % decrease in total phytate (IP6 + IP5 + IP4 + IP3) content after 7 min of DIC treatment. This reduction is greater than reported by de Boland et al. (1975) or Chango et al. (1993). Table 2. The effect of treatment time (min) with controlled instantaneous pressure drop
(DIC) on the phytate content of L. mutabilis seed. (Pressure 7 bar, moisture content 30 %).
DIC Treatment time (min) 0 1 2 3 4 5 6 7 IP6 21.13 9.81 5.48 4.21 3.56 2.49 2.23 1.79 IP5+IP4+IP3 1.55 8.03 9.44 9.37 8.96 8.38 7.84 7.41 Total (mg/g DM) 22.68 17.84 14.92 13.57 12.51 10.87 10.07 9.19
The reduction in phytate content in L. mutabilis, with processing time is shown in
Figure 1. As discussed above under heat or enzymatic action, IP6 is transformed into less phosphorylated compounds such as IP5, IP4 or IP3 (Greiner, 2001; Vidal-Valverde, 2001). The IP6 is the first to bind minerals, and make them unavailable for intestinal absorption. The
Regression function (L . albus )y = 0,12x2 - 2,12x + 16,86
R2 = 0,98
Regression function (L . mutabilis )y = 0,26x2 - 3,54x + 21,86
R2 = 0,98
0
5
10
15
20
25
30
0 1 2 3 4 5 6 7 8Treatment duration (min)
Phyt
ates
(mg/
g d.
m)
Lupinus albusLupinus mutabilis

336
other inositol phosphates are less harmful and they may be sought-after for their medical and pharmaceutical properties (Cuadrado et al., 1996; Greiner, 2001). Conclusions This work shows that after DIC treatment, the level of the most harmful phytate compound, IP6, is decreased as processing time is increased. At the same time levels of IP5, IP4 and IP3 are increased. A decrease in total phytate (IP6 + IP5 + IP4 + IP3) content of 55 % in L. albus and 60 % in L. mutabilis was obtained. Thus, the search for new protein sources and new processes may make DIC, which is a highly controlled process, well suited for the treatment legume seed. References Chango, A., Bau, H.M., Villaume, C., Schwertz, A., Nicolas, J.P. & Mejean, L. 1993. Effets
des traitements (chauffage et fermentation par Rhizopus oligosporus sp-T3) de la graine de lupin blanc doux sur certains facteurs de son utilisation nutritionnelle. Reproduction Nutrition Development 33, 89-98.
Cuadrado, C., Ayet, G., Robredo, L.M., Muzquiz, M., Pedrosa, M.M. & Burbano, C. 1999. Occurrence of non-nutrient compounds in Andean Lupinus mutabilis. In Towards the 21st Century. G.D. Hill (Ed.). Proceedings 8th International Lupin Conference, Asilomar, Ca 11-16 May 1996, 120-124.
De Boland, A.R., Garner, G.B. & O’Dell, B.L. 1975. Identification and properties of “phytate” in cereal grains and oilseed products. Journal of Agricultural and Food Chemistry 23, 1186-1189.
Greiner R. 2001. Properties of phytate-degrading enzymes from germinated lupine seeds (Lupinus albus var. Amiga). In Towards the Sustainable Production of Healthy Food, Feed and Novel Products. AEP (Ed.). Proceedings 4th European Conference on Grain Legumes, Cracow 8-12 July 2001, 398-399.
Haddad, J., Louka, N., Gadouleau, M., Juhel, F. & Allaf, K. 2001. Application du nouveau procédé de séchage/texturation par Détente Instantanée Contrôlée DIC aux poissons: Impact sur les caractéristiques physico-chimiques du produit fini. Science des Aliments 21, 481-498.
Mohamed, A.I., Perera, P.A.J. & Hafez, Y. 1986. New chromophore for phytic acid determination. Cereal Chemistry 63, 475-478.
Sandberg, A.S. & Ahderinne, R. 1986. HPLC method for determination of inositol, tri-, tetra-, penta-, and hexaphosphates in foods and intestinal contents. Journal of Food Science 51, 547-550.
Vidal-Valverde, C., Frias, J., Lambein, F. & Kuo, Y-H. 2001. Increasing the functionality of legumes by germination. In Towards the Sustainable Production of Healthy Food, Feed and Novel Products. AEP (Ed.). Proceedings 4th European Conference on Grain Legumes, Cracow 8-12 July 2001, 422.

337
Effect of aqueous, acid and alkaline thermal treatments on the protein quality of Lupinus campestris seed C. Jiménez Martínez, R. Mora Escobedo & G. Dávila-Ortíz Departamento de Graduados e Investigación en Alimentos, Escuela Nacional de Ciencias Biológicas, Instituto Politécnico Nacional, México DF, 11340 México Summary Increasing importance is being given to the genus Lupinus (Lupinus spp) as a possible soybean (Glycine max) substitute in human food. Lupinus campestris seed has a similar chemical composition to soybean seed and contains 44 % protein and 13 % oil. Lupin protein is low in the sulphur amino acids. However, when supplemented with DL-methionine its biological value increases. Lupin seed utilisation has been limited because of the presence of toxic quinolizidine alkaloids (QAs). The objective of this work was to reduce, or eliminate, the alkaloids trough aqueous, acid or alkaline thermal treatments and to determine the improvement in the protein quality following treatment. The results showed that the most efficient treatment for alkaloid elimination was the alkaline one. The protein content increased with both treatments from 44 % to 50 - 53 %. Although the protein concentration was higher with all the treatments, it was not possible to determine its biological value because the experimental animals refused to eat any diet and finally died. Supplementation of debittered lupin meals, with 0.5 DL-methionine gave a PER value of 2.04. However, this was lower than the casein PER of 2.45. The PER results showed that the alkaline thermal treatment was the best. The apparent digestibility was more than 90 % for all treatments. Alkaline treated seed had the highest digestibility (93 %). Keywords: Lupinus campestris, detoxification, protein quality, digestibility Introduction Legume seed provides about 20 % of the world's total protein intake. Nutritional studies, with animals and humans, have shown that lupin can compare favourably with soybeans. Legume seed is widely incorporated into human diet. Lupin seeds are one the richest sources of plant protein. Although the protein content and amino acid profile vary among species, the intraspecific variability is low. Lupin is a member of the legume family and is native in the Mediterranean region and the Americans. For many centuries they have been grown as a source of food for both animals and humans (Yañez, 1979; Feldheim, 1994).
Lupinus campestris seed, like other lupins, has a high protein content (44 %). The limitation for the wider use of bitter lupins has been their high QA content. However, cooking, and other methods, can elimination toxic and antinutritional factors (ANFs) and improve the nutritive value of legume seed.
The nutritional value of a protein is determined by the quantity, availability and proportions of the essential amino acids it contains, and the presence, for optimum utilisation, of sufficient non-essential amino acids. Bioassays can measure the efficiency of biological utilisation of dietary proteins as sources of essential amino acids under standardised conditions (Friedman, 1996).

338
The aim of this study was to determine the protein efficiency ratio (PER), the corrected protein efficiency (cPER) and the digestibility of debittered L. campestris seed flour obtained by aqueous, alkaline and acid thermal treatments. Materials and methods Wild L. campestris seed was collected along 50 km of the Oaxtepec-Xochimilco highway in the Mexican state of Morelos.
Debittering of Lupinus campestris seeds. The process consisted of soaking and boiling of the seed. Firstly, immature and damage seed, plants remains, stones and other foreign material was eliminated. Boiling at 93 °C for 6 h, with water changes every 20 min hydrated the seed. The three debittering methods were aqueous, alkaline (0.5 % NaHCO3) and acid (0.1 % citric acid). Chemical analyses and protein quality Protein (N × 6.25, Method 955.04), lipids (Method 920.39), total dietary fibre (Method 985.29) and ash (Method 923.03), the PER, net protein ratio (RPN) and digestibility were determined according AOAC (1995) methods. The following brief definitions indicate the techniques used to measure protein nutritional quality.
(Weight gain of test group) Protein Efficiency Ratio (PER)
= (Total protein consumed)
(1)
(PER of test protein PER corrected = (PER of a casein control) (2)
[N consumed – faecal N + metabolic faecal N]Digestibility (TD) = N consumed × 100 (3)
Where:
Faecal N (protein-free animals) × food intake (test animals) Metabolic faecal N = Food intake (protein-free animals). (4)
Extraction and measurement of quinolizidine alkaloids The QAs were extracted as described by Muzquiz et al. (1994). Finely ground lupin seed (0.5 g) was homogenized three times in 5 ml of 5 % trichloroacetic acid with an Ultra Turrax and centrifuged at 10,000 rpm for 10 min. The supernatant was hydrolysed with 0.8 ml of 10M NaOH. The alkaloids were extracted three times with 15 ml of dichloromethane. The dichloromethane extracts were combined and evaporated to dryness at 30°C. Methanol (1 ml) was added to dissolve the alkaloids.
The QAs were measured by GC/MS using a Perkin Elmer Chromatography Autosystem equipped with a phosphorus-nitrogen detector (NDP). A Turbochrom program was used for instrument control and data analysis. An SPB-1 column (30 m × 0.25 mm ID, 0.25 mm film

339
thickness) was used with helium as the carrier gas (1.38 bar). The injector and detector temperatures were 240 °C and 300 °C, respectively. The temperature started at 150 °C and increased by 5° C/min to 235 °C with a 15-min holding time. The alkaloids were identified by their mass fragmentation pattern by mass spectrometry (MS). Calibration curves were prepared for pure lupanine. Responses were linear in the range 0 - 1.25 mg/ml. The determination coefficient of alkaloid content was > 0.99. Results Table 1 shows the proximate composition chemical of untreated L. campestris seed and the seed after aqueous, alkaline and acid thermal treatment. The protein level in L. campestris seed at 43 % was similar to the level in other species (Yañez et al., 1979; Feldheim, 1994). It was higher that the protein level in others grain legumes such as lentil (Lens culinaris) and bean (Phaseolus vulgaris), which have protein levels of 6 - 25 % (Bourges, 1987) and similar to soybean. The lipid content was similar to the level in L. albus seed (Yañez et al., 1979) and higher than in L. luteus seeds (8.49 %) according to Ballester et al., 1980). The total dietary fibre content at 14.7 % was similar to levels in L. angustifolius, L. luteus and L. notarius seed (13 - 19 %). Table 1. Chemical composition of L. campestris seed before and after being subjected to
aqueous, alkaline and acid thermal treatments (%). Treatment Component L. campestris seed Aqueous Alkaline Acid Protein (N × 6.25) 43.00 ± 0.23 54.00 ± 0.46 56.70 ± 0.29 56.30 ± 1.22 Lipids 10.84 ± 0.03 13.22 ± 0.10 15.83 ± 0.04 12.33 ± 0.15 Total dietary fibre 14.70 ± 0.10 12.20 ± 0.15 10.20 ± 0.11 10.80 ± 0.16 Extract free N* 20.22 ± 0.19 18.00 ± 0.50 14.00 ± 0.23 17.52 ± 0.28 Ash 3.24 ± 0.10 2.35 ± 0.07 2.46 ± 0.16 3.05 ± 0.06 *By difference
A consequence of the debittering treatments was that the different seed components were modified. Protein content increased from 43 % to 54.0, 56.7 and 56.3 % after aqueous, alkaline and acid treatment respectively. This increased could be due to the elimination of some carbohydrates, mineral and total dietary fibre treatment. The alkaline and acid treatments eliminated the most dietary fibre. The carbohydrate content was also reduced by the treatments. The greatest reduction being in the alkaline treatment. Quinolizidine alkaloid content of L. campestris seed The major hindrance to the wider use of bitter lupins as food and animal feed has been their bitter, toxic, QA content. The wild L. campestris seed contained an average of 2.73 mg/g hidroxiaphhylline and hidroxiaphyllidine, which were the principal QAs found in the L. campestris. (Jiménez et al., 2001). Based on alkaloid content L. campestris can be classified as bitter (Wink, 1998). Figure 1 shows the final QA concentration in the original and the debittered after aqueous, acid and alkaline treatment. Seed after alkaline treatment contained the least alkaloid at 0.14 mg/g.

340
Figure 1. Total quinolizidine alkaloid content of untreated L. campestris seed and seed
after aqueous, alkaline and acid thermal treatment. Protein quality Due at the low nutritional value of the flour without methionine it was necessary to add this amino acid to the test rations (Jiménez et al., 2001). Table 2 shows the PER results of the L. campestris after aqueous, alkaline and acid treatment. The addition of methionine and the treatments increased rat weight gain and the PER. Among the three treatments the highest PER was from rats fed the alkali treated seed (2.04), which was similar to the casein PER at (2.45). Table 2. Increase in weight, protein efficiency ratio (PER) and corrected protein efficiency
ratio (cPER) of L. campestris seed and flour obtained after aqueous, alkaline and acid thermal treatment.
Treatment DL-Met (%) Wt increase (g) PER cPER Aqueous 0.05 19.73 1.48 ± 0.37 1.51 ± 0.39 Alkaline 0.05 24.46 2.04 ± 0.40 2.08 ± 0.39 Acid 0.05 22.61 1.83 ± 0.35 1.87 ± 0.34 Casein 38.01 2.45 ± 0.43
A possible explanation to this result could be the presence of amino acid structures involving cystine, which are enzyme resistant, thus affecting its biological availability as reported in other legume (Kakade, 1974). The addition of methionine gave a good response in both rat growth and PER. Alkaline treatment had the greatest effect on seed ANF and this was reflected in increased weight gain and a higher PER.
Apparent and true digestibility (Table 3), also showed the benefit of the three treatments. The values, are close to the casein value. Of the three treatments, the lowest digestibility was in the water treatment, followed by acid treatment. The alkali treated flour had a digestibility, which was similar to casein (92 and 93 % respectively). Donovan et al. (1991) reported that the nutritional quality of two sweet L. albus cultivars with a low quinolizidine alkaloid content (0.05 %) and supplemented with 0.2 % L-methionine was similar to soybean meal.
The protein content of the L. campestris tested was similar to soybean. Lupin seed can contains up to twice as much protein than in grain legume seed normally consumed by humans. Further, lupin can yield 1,000 - 2,000 kg/ha compared with 580 - 620 kg/ha for
0.0
0.5
1.0
1.5
2.0
2.5
mg/
g (d
.m.) L. campestris seeds
AqueousAcidAlkaline

341
beans or 769 - 870 kg/ha for chickpeas (Cicer arietinum). These factors may explain why lupin has been used for centuries as a human food by certain cultures. However, as with many legumes, bitter lupins contain undesirable alkaloids that must be removed before their consumption. Table 3. Apparent and true digestibility of L. campestris seed after aqueous, alkaline and
acid thermal treatment. Treatment Apparent digestibility (%) True digestibility (%) Seed
Aqueous 91.96 ± 1.74 93.00 ± 1.71 Alkaline 92.90 ± 0.83 93.83 ± 0.90 Acid 90.96 ± 1.10 91.96 ± 1.25
Casein 92.37 ± 0.97 93.18 ± 0.98 Acknowledgements The Instituto Politécnico Nacional (IPN) and Consejo Nacional de Ciencia y Tecnología (CONACyT) through 33995 project, for financial support of this research. Cristian Jiménez Martínez acknowledges a study grant from CONACyT. References AOAC. 1995. Official Methods of Analysis, 15th Edition. Association of Analytical
Chemists, Washington DC. Ballester D. 1980. Chemical composition, nutritive value and toxicological evaluation of two
species of sweet lupine (L. albus and L. luteus). Journal of Agricultural and Food Chemistry 28, 402-405.
Donovan B. C.; McNiven, M.A.; McLeod, J.A.; Anderson, D.M. 1991. Protein quality of two cultivars of lupin seeds evaluated in weanling rats. Animal Feed Science and Technology 33, 87-95.
Feldheim, W. 1994. Fermentation of lupin fibre. In Advances in Lupin Research. J.M. Neves Martins & Ma Beirão de Costa (Eds). Proceedings VIIth International Lupin Conference, Évora, 18-23 April 1993, 445–450.
Friedman, M. 1996. Nutritional value of proteins from different food sources. A review. Journal of Agricultural and Food Chemistry44, 6-29.
Jiménez, M.C., Hernández, S.H., Alvarez, M.G., Robledo, Q.N., Martínez, H.J. & Dávila, O.G. 2001. Effect of aqueous and alkaline thermal treatment on chemical composition an oligosaccharide, alkaloid and tannin content of Lupinus campestris seeds. Journal of the Science of Food and Agriculture 81, 421-428.
Kakade, M.L. 1974. Biochemical basis for the differences in plant protein utilization. Journal of Agricultural and Food Chemistry 22, 550-555.
Muzquiz, M., Cuadrado, C., Ayet, G., Cuadra, C., de la, Burbano, C. & Osagie, A. 1994. Variation of alkaloid components of lupin seeds in 49 genotypes of Lupinus albus L. from different countries and locations. Journal of Agricultural and Food Chemistry 42, 1447-1450
Yañez, E. Gattas, V. & Ballester, D. 1979. Valor nutritivo del Lupinus y su potencial como alimento humano. Archivos Latinoamericanos de Nutrición 29, 510-520.
Wink M. 1998. Alkaloids: Biochemistry, Ecology and Medicinal Applications. Plenum Press, New York.


343
Elimination of quinolizidine alkaloids, αααα-galactosides and phenolic compounds from Lupinus campestris seed by aqueous, acid and alkaline thermal treatment C. Jiménez-Martínez1, M.M. Pedrosa2, M. Muzquiz2 & G. Dávila-Ortíz1 1Department of Graduates and Food Research, Escuela Nacional de Ciencias Biológicas-IPN, Prolongación de Carpio y Plan de Ayala, 11340 México DF, México 2Department of Food Technology, INIA, Apdo 811, 28080 Madrid, Spain Summary There has recently been a renewed interest in the seed of Lupinus spp as potential protein sources for human food and animal feed. Lupin seeds have a high protein content (35 - 48 %) and up to 15 % lipids, which compares favourably with soybean (Glycine max). However, their use has been limited, probably due to the presence of quinolizidine alkaloids (QA), α−galactosides (GAL) and phenolic compounds (PC). This study was done on the effect of aqueous, alkaline and acid thermal treatments in decreasing or eliminating toxic and antinutritional factors (ANFs) in Lupinus campestris seed. The three treatments eliminated more than 90 % of the original QA’s, 60 % of the GAL’s and 65 % of PC’s. The alkaline thermal treatment was the most efficient. Keywords: Lupinus campestris, debittered, alkaloids, α-galactosides, phenolic compounds Introduction Legumes are a significant component in a large proportion of human diets worldwide and provide about 20 % of worldwide protein intake (Kyle, 1995). Much of this protein is from soybean. Nutritional studies by Yañez et al. (1979), Gross et al. (1983) and Shoenenberger et al. (1982) with animals and humans have shown that lupin seed compares very favourably with soybean. In Mexico there are about 90 wild lupin species none of which are commercially exploited.
Like other lupins Lupinus campestris seed, has a high seed protein content (44 %). Lupins offer some nutritional advantages over soybean as they contain only small quantities of trypsin inhibitors, phenolic compounds, phytates and saponins (Kyle, 1995). The limiting factor for the wider use of lupin seed is the QA content of wild genotypes. The seed also contains relatively high levels of α-galactosides (Trugo et al., 1988). The latter cause flatulence, as they are indigestible by humans. Cooking and other methods, such as debittering, facilitate the elimination of ANFs and improve the nutritional value of legumes. Debittering is an ancient procedure used by peoples of the Andean Highlands to eliminate the lupin seed alkaloids (Gross et al., 1983).The objective of this study was to decrease, or eliminate, the alkaloids, oligosaccharides and phenolic compounds in L. campestris seed by aqueous, acid and alkaline thermal treatments. Materials and methods Wild L. campestris seed was collected along a 50 km section of the Oaxtepec-Xochimilco highway in the state of Morelos, Mexico. Immature and damaged seed, plants remains, stones and other foreign material were removed from the seed sample. The seed was then hydrated
344
by boiling (93 °C) in water for 6 h, with water changes every 20 min. Three debittering methods were applied: aqueous, alkaline (0.5 % NaHCO3) and acid (0.1 % citric acid).
Extraction and quantification of QA was as described by Muzquiz et al. (1994) using GC/SM. Extraction and quantification of GAL was as per Muzquiz et al. (1992) and was by HPLC. Finally PC extraction followed AOAC (1995). Results and discussion Quinolizidine alkaloids The initial total QA concentration L. campestris seed was 2.28 mg/g of seeds. This is comparable to concentrations found in other varieties of bitter lupin (Lupinus spp) (Muzquiz et al., 1994), and is slightly lower than in L. mutabilis seed (3.1 %) (Hatzold, et al., 1983) (Table 1). Five alkaloids were isolated by alkaloid separation. The main alkaloid present was hydroxyaphylline (1.87 mg/g seed), followed by hydroxyaphyllidine (0.27 mg/g seed).
After 6 h the QA concentration was 0.138 mg/g of seed in the alkaline treatment, 0.283 mg/g of seeds in the acid treatment and 0.259 mg/g of seeds in the water treatment. These percentages correspond to reductions in alkaloid level of 87.6 % in acid, 88.66 % in water and 93.96 % in the alkaline. These values are similar to those reported by Torres et al. (1980) using an aqueous thermal treatment (with an unspecified alkali), which reduced total QAs by 98.6 %. Ortiz & Murkerjei (1982) reported alkaline treatment reduced total QAs by 70 - 80 %. Table 1. Effect of acid, aqueous and alkaline thermal treatments on the quinolizidine alkaloid
content of L. campestris seed (mg/g seed). L. campestris Acid Aqueous Alkaline Dehydro-lupanine 0.042 ± 0.05 0 0 0 Hydroxyaphyiline 0.275 ± 0.10 0.102 ± 0.02 0.045±0.00 0.065 ± 0.00Hydroxyaphyillidine 1.867 ± 0.06 0.181 ± 0.00 0.214±0.01 0.073 ± 0.01Epi-hydroxyaphyilina 0.054 ± 0.01 0 0 0 Dehydroepihydroxiaphylina 0.047 ± 0.00 0 0 0 Total 2.285 ± 0.02 0.283 ±0.00 0.259±0.00 0.138 ± 0.00
Average of three determinations ± SD αααα-galactosides Stachyose was the most abundant of the three oligosaccharides in L. campestris seed. The GAL concentration of L. campestris at 11.65 mg/g seed was similar to that in L. luteus (11.8 mg/g seed) (Matherson & Saini, 1977). At the end of the 6 h the original GAL had been reduced by 92 % in the acid 96 % in the alkali and 97 % in the water (Table 2). This decrease was beneficial since stachyose, raffinose and verbascose were all included in the reduction. The elimination of these oligosaccharides is necessary as they cause flatulence in monogastric animals due to their lack of an enzyme, which can break them down (Saini & Lymbery, 1983).
The 92 % elimination achieved with acid treatment is efficient, especially compared with the results of Reddy and Salunke (1980) who reported a 25 % reduction in oligosaccharides in black gram (Phaseolus mungo) after heating for 40 min at 116 °C in water. In the alkaline treatment stachyose was reduced to 3 % of its initial value, and the other GAL’s were almost completely eliminated.

345
Table 2. Effect of acid, aqueous and alkaline thermal treatment on α-galactoside content of
L. campestris seed (mg/g seed). L. campestris Acid Aqueous Alkaline
Sucrose 21.45 ± 1.32 1.48 ± 0.17 0.71 ± 0.19 0.59 ± 0.10 Melibiose 5.83 ± 1.43 1.05 ± 0.00 Raffinose 11.65 ± 1.54 1.22 ± 0.14 1.13 ± 0.08 Ciceritol 4.82 ± 1.33 0 Stachyose 57.16 ± 1.65 5.55 ± 0.97 2.92 ± 0.60 3.20 ± 0.68 Verbascose 19.45 ± 1.78 0 Total 120.35± 1.58 9.19 ± 0.26 3.77 ± 1.56 4.78± 0.14
Average of three determinations ± SD Phenolic compounds The initial PC content of L. campestris was 5.1 mg/g seed (Fig. 1). Chavan et al. (1979) reported the soybean variety IS-2825 contained 3.4 % tannin, which is ten times more than the PC content in common varieties. There are also varieties with PC contents ranging from 0.40 - 2.46 % (Prince et al., 1980). The low PC content L. campestris (5.1 mg/g seed) indicates that the seed is possibly nutritionally better than soybean in terms of mineral bioavailability and protein digestibility (Adewusi & Falade,1996).
Figure 2. Effect of acid, aqueous and alkaline thermal treatment on the phenolic compound
content of L. campestris seed (mg/g seed). Acknowledgements The Instituto Politécnico Nacional (IPN) and the Consejo Nacional de Ciencia y Tecnología (CONACyT) for support via project 33995, and CYTED. Cristian Jiménez-Martínez was supported by a CONACyT study grant.
0
1
2
3
4
5
6
mg/
g db
Seed Acid
Treatments

346
References Adewusii, S.R.A. & Falade, O.S. 1996. The effects of cooking on the extractable tannins,
phytate, sugar and mineral solubility in some improved Nigerian legume seeds. Food Science and Technology International 2, 231-239A.
AOAC 1995. Official Methods of Analysis. 15th Edition Association of Analytical Chemists, Washington DC.
Chavan, J.K., Kadam, S.S., Ghonsikar, C.P. & Salunke, D.K. 1979. Removal of tannins and improvement of in vitro protein digestibility of sorghum seed by soaking in alkali. Journal of Food Science 44, 1319–1321.
Gross, U., Godomar, G.R. & Schoeneberger, H. 1983. The development and acceptability of lupine (Lupinus mutabilis) products. Qualitas Plantarum - Plant Foods in Human Nutrition 32, 155-164.
Hatzold, T., Ibraim, E., Gross, R., Wink, M, Harmannn, H. & Witte, L. 1983. Quinolizidine alkaloids in seed of Lupinus mutabilis. Journal of Agricultural and Food Chemistry 31, 934-938.
Kyle, A.S.W. 1994. The current and potential uses of Lupinus for human food. MSc Thesis, Department of Food Technology, Victoria University of Technology, Melbourne.
Matterson, N.K. & Saini, H.S. 1977. Polysaccharides and oligosaccharides changes in germination in lupin cotyledons. Phytochemistry 16, 59–66.
Muzquiz, M., Cuadrado, C., Ayet, G., Cuadra, C. de la, Burbano, C.C. & Osagie, A. 1994. Variation of alkaloid components of lupin seeds in 49 genotypes of Lupinus albus L. from different countries and locations. Journal of Agricultural and Food Chemistry 42, 1447-1450.
Muzquiz, M., Rey, C. & Cuadrado, C. 1992. Effect of germination on the oligosaccharide content of lupin species. Journal of Chromatography 607, 349-352.
Ologhobo, A.D. & Fetuga, B.L. 1982. Polyphenols, phytic acid and other phosphorus compounds of lima beans (Phaseolus lunatus). Nutrition Reports International 26, 605–611.
Ortiz, J.G.F. & Mukherjee, K.D. 1982. Extraction of alkaloids and oil from bitter lupine seed. Journal of the American Oil Chemists Society 59, 241–244.
Price, M.L., Hagerman, A.E. & Butler, L.G. 1980. Tannin content of cowpea, chickpeas, pigeonpeas and mung beans. Journal of Agricultural and Food Chemistry 28, 459–461.
Reddy, N.R. & Salunke, D.K. 1980. Changes in oligosaccharides during germination and cooking of black gram and fermentation of black gram/rice blend. Cereal Chemistry 57, 356–360.
Saini, H.S. & Lymbery, J. 1983. Soluble carbohydrates of developing lupine seed. Phytochemistry 22, 1367–1370.
Shoeneberger, H., Gross R., Cremer H.D., Elmadfa, I. 1982. Composition and protein quality of Lupinus mutabilis. Journal of Nutrition 112, 70-76.
Torres, T.F., Nagata, A. & Dreifua, S. 1980. Métodos de eliminación de alcaloides en la semilla de L. mutabilis. Archivos Latinoamericanos de Nutrición 30, 200-207.
Trugo, C.L., Almeida, D.C.F. & Gross, R. 1988. Oligosaccharides contents in the seed of cultivated Lupins. Journal of the Science of Food and Agriculture 45, 21-24.
Yañez, E., Gattas, V. & Ballester, D. 1979. Valor nutritivo del Lupinus y su potencial como alimento humano. Archivos Latinoamericanos de Nutrición 29, 510-520.
Ziena, H.M., Youssef, M.M. & El-Mahady. A.R. 1991. Amino acid composition and some antinutritional factors of cooked faba beans (Medammis): Effects of cooking temperature and time. Journal of Food Science 56, 1347-1352.

347
Limited phytate hydrolysis by endogenous phytase of sesame seed during germination K. Żyła1, T. Fortuna2, M. Mika1 & M. Czubak1 1Department of Food Biotechnology, University of Agriculture in Kraków, 29-Listopada Ave 46, 31-425 Kraków, Poland 2Department of Food Analysis and Food Quality Assessment, University of Agriculture in Kraków, ul. Balicka 122, 30-149 Kraków, Poland Summary It is well know that germination improves the nutritional quality of oilseeds. The objective of this work was to determine changes in phytase activity, in the chemical composition and the in vitro nutrient digestibility of germinated sesame seed (Sesamum indicum L.). The seed was germinated for 12, 24, 36, 48 and 60 h, at 25, 30, 35, 40, and 45 oC. Phytase activity was measured colourimetrically following cellulase and xylanase assisted extraction. Inositol phosphates were determined by HPLC and in vitro digestibility was measured by peptic and pancreatic digestion of samples.
The sesame phytase activity increased from 1.4 FTU/g in raw seeds to 5.0 FTU/g after 36 h of germination. The optimal germination temperature was 30 oC at pH 5,5. As a result of germination the sesame seeds fat and carbohydrates content decreased from 47 to 40 %, and from 6.05 to 2.03 %, respectively. However protein level was not altered (27 vs. 28 %). Total phosphorus, calcium and magnesium were unchanged, and the inorganic seed phosphorus increased from 0.6 to 1.4 mg/g. During germination IP6 concentration of IP6 decreased slightly (from 38.2 to 36.4 µM/g ddb1), the decrease in IP5 concentration was not significant, and only minor amounts of IP4 were produced. In the in vitro digestion test seed germination improved the digestibility of phosphorus, calcium, magnesium and carbohydrates, but did not affect protein digestibility. Keywords: Sesamum indicum seed, phytate, phytase myo-inositol phosphate, in vitro
digestibility, germination Introduction Sesame seed is primarily known for the high level of lignans (sesamin and sesamolin) in their oil (Sirato-Yasumoto et al., 2001). However, they are also an abundant source of phytic acid (myo-inositol hexakis-dihydrogen phosphate, IP6). During germination, cereal seeds and oilseeds synthesize the enzyme phytase (EC. 3.1.3.8), which dephosphorylates phytate to lower myo-inositol phosphates (IP1-5) releasing phosphate residues. The lower myo-inositol phosphates (mainly IP3 + 4) produced by the action of plant 6-phytases may have important physiological functions (Plaami, 1997; Vohra & Satyanarayana, 2003). The aim of this work was to try to convert sesame seed phytates into lower myo-inositol phosphates by seed germination and observe the changes caused by germination in seed composition and in the in vitro digestibility of protein, carbohydrate, phosphorus, calcium and magnesium.
1 ddb = defatted dry basis.

348
Materials and methods Materials Sesame seed was purchased from a local health food store. Cellulase, xylanase, pepsin and pancreatin were from Sigma Chemical, USA. The pancreatin had an activity of 8 x United States Pharmacopeia (USP). Dialysis tubing had a molecular weight cut-off 12 kDa. Sesame seed germination Sesame seeds were soaked in 0.2 % of potassium sorbate, placed on thin layers of wet cotton in Petri dishes and germinated at different temperatures in laboratory incubators. Germination times were 0, 12, 24, 36, 48, 72 and 96 h and germination temperatures were 25, 30, 35, 40, and 45 oC. Germinated seeds were washed free of sorbate solution, freeze-dried and ground to pass a 1 mm screen and stored at a room temperature. Phytase activity determinations Seed phytase activity was determined by a procedure that involved a 30 min extraction of a 0.25 g sample at 30 oC and pH 4.0 with 100 units of ultrafiltrated (membrane cut-off 30 kDa) Aspergillus niger cellulase and xylanase preparations. The extract was centrifuged and mixed with a 5 mM sodium phytate solution. The amount of phosphate liberated after 60 min of reaction at 40 oC and pH 4,0 was determined colourimetrically. Determination of myo-inositol phosphates Samples were defatted, mixed with 0.4 M HClO4 and extracted with a shaker for 2 h. The extract was centrifuged, neutralized with 1M K2CO3 and put on a HyperSep NH2 SPE column. A fraction eluted from the column by 0.6 ml 1M HCl was used for HPLC analysis of inositol phosphates. Inositol phosphates (IP6-4) were determined by a modified Lehrfeld (1994) procedure using ion-pair C18 reverse phase HPLC with refractive index detection. In vitro determination of nutrients digestibility An in vitro procedure, which was designed to simulate digestion in the intestinal tract, was used. Triplicate samples (0.4 ± 0.001 g) of ground sesame seed was weighed into plastic syringes without Luer-lock tips. Samples were hydrated with 1.0 ml of water and an a HCl solution so to obtain a pH of 2.0. Pepsin (6,200 units) was then added and the contents of each tube were mixed, sealed with parafilm, vortexed, and incubated in a water bath at 37 oC for 2 h. At the end of this period, 0.5 ml of NaHCO3 solution with 2 mg of pancreatin and 25 mg of bile extract (0.6 ml) were added to obtain a pH 7.0. The slurry was transferred quantitatively to segments of dialysis tubing by a syringe. The segments were placed in 250 ml Erlenmeyer flasks containing 40 ml of 0,1 M NaCl in a 0.05M imidazole buffer at pH 7.0 and were incubated in a shaking water bath. After 4 h samples of the dialysate were taken for the determination of inorganic phosphate, calcium, magnesium, protein and reducing sugars as detailed in Żyła et al. (2000).

349
Chemical analyses Basic chemical analyses were performed using a Tecator autoanalyser. Inorganic phosphorus was assayed using the procedure of Kessel (1972). Results and discussion Effects of temperature and time of germination on phytase activity in sesame seed During the first 36 h of sesame seed germination endogenous phytase activity increased 3.4 fold and then declined slightly (Table 1). Muzquiz et al. (1998) observed a constant increase in phytase activity in Lupinus albus and L. luteus seeds during 120 h of germination. The optimal temperature for phytase biosynthesis during germination was 30 oC at pH 5,5. The crude phytase extracted from the seed had an optimal pH of 4.0 and an optimal temperature of 40 oC (data not shown), which is common for phytases from oilseeds. Table 1. Phytase activity (FTU/g) in sesame seed at different germination times and
temperatures1. Germination time (Temperature 30 oC) Germination temperature (Time 30 h) (h) Mean Std Error (oC) Mean Std Error 0 1.45 0.05 25 3.66 0.08 12 1.92 0.04 30 4.89 0.05 24 4.21 0.11 35 4.67 0.07 36 4.99 0.09 40 3.01 0.09 48 4.25 0.08 45 0.87 0.10 60 4.30 0.12 1Values are means of five determinations. Effect of germination on chemical composition and nutrient in vitro digestibility of sesame seed As a result of germination (30 oC, 36 h) the fat and carbohydrate content decreased from 47 to 40 %, and from 6.05 to 2.03 %, respectively. However, the protein content was unchanged (27 vs. 28 % (NS); Table 2). Total phosphorus, calcium and magnesium content were unchanged. The seed inorganic phosphorus concentration of increased from 0.6 to 1.4 mg/g. During germination the IP6 concentration only decreased slightly (38.2 to 36.4 µM/g ddb.). The decrease in IP5 concentration was not significant and only minor amounts of IP4 were produced. In the in vitro digestion test seed germination improved phosphorus, calcium, magnesium and carbohydrate digestibility but had no effect on protein digestibility. Zduńczyk et al. (1982) obtained similar results germinating lentil (Lens culinaris), pea (Pisum sativum) and bean (Phaseolus vulgaris) seed.
It can be concluded that germination of sesame seed activates endogenous phytase, but this creates only minor changes in the seed myo-inositol phosphates profile. This is in agreement with results of Muzquiz et al. (1998) who observed similar phenomena in germinated Lupinus albus an L. luteus seed. We found that only at extreme temperatures (120 oC), in acidic pH, after prolonged reaction time can sesame seed IP6 be converted in substantial quantities to lower IPs. A practical application for such a procedure is, however, doubtful.

350
Table 2. Changes in the chemical composition, and nutrient in vitro digestibility of raw and germinated sesame seed1.
Sesame seed Sesame seed Raw Germinated Raw GerminatedAsh (%) 4.74 5.02 IP6 (µM/g ddb2) 38.20 36.37 Protein (%) 27.38 27.99 IP5 (µM/g ddb) 3.11 2.69 Fat (%) 47.20 39.85* IP4 (µM/g ddb) - 0.46 Carbohydrates (%) 6.05 2.03* P dial (mg/g) 0.23 0.53* P total (mg/g) 7.51 7.54 Ca dial (mg/g) 0.25 0.74* P inorganic (mg/g) 0.59 1.37* Mg dial (mg/g) 1.00 1.30* Ca (mg/g) 10.67 10.03 Protein dial (mg/g) 86.13 87.81 Mg (mg/g) 3.70 3.70 Sugars dial (mg/g) 6.01 51.31* 1Values are means of five determinations. 2ddb = defatted dry basis. *denotes significant P < 0.05 (t-test) Acknowledgement The authors acknowledge the financial support of the Polish Research Committee (Grant No. AR 73/31/PBZ/021/P06/99). References Kessel, H. 1972. Phosphatbestimmung in Phosphatstärken. Die Stärke 24 (2), 46-51. Lehrfeld, J. 1994. HPLC separation and quantification of phytic acid and some inositol
phosphates in foods: problems and solutions. Journal of Agricultural and Food Chemistry 42, 2726-2731.
Muzquiz, M., Pedrosa, M.M., Cuadrado, C., Ayet, G., Burbano, C., Brenes, A. 1998. Variation of alkaloids, alkaloids esters, phytic acid and phytase activity in germinated seeds of Lupinus albus and L. luteus. In Recent Advances of Research in Antinutritional Factors in Legume Seeds and Rapeseed. A.J.M. Jansman, G.D. Hill, J. Huisman & A.F.B. van der Poel (Eds). Proceedings 3rd International Workshop on Antinutritional Factors in Legume Seeds and Rapeseed, Wageningen 8-10 July 1998, 387-390.
Plaami, S. 1997. Myoinositol phosphates: Analysis, content in foods and effects in nutrition. Lebensmittel-Wissenschaft und Technologie 30, 633-647.
Sirato-Yasumoto, S., Katsuta, M., Okuyama, Y., Takahashi, Y. & Ide, T. 2001. Effect of sesame seeds rich in sesamin and sesamolin on fatty acid oxidation in rat liver. Journal of Agricultural and Food Chemistry 49, 2647-2651.
Vohra, A. & Satyanarayana, T. 2003. Phytases: Microbial sources, production, purification, and potential biotechnological applications. Critical Reviews in Biotechnology 23, 29-60.
Zduńczyk, Z., Godycka, I., Frejnagel, S., Juśkiewicz, J. & Milczak, M. 1982. Nutritional value of lentil seeds as compared with beans and peas. Polish Journal of Food and Nutrition Sciences 3, 74-78.
Żyła, K., Koreleski, J., Świątkiewicz, S., Wikiera, A., Kujawski, M., Piironen, J. & Ledoux, D.R. 2000. Effects of phosphorolytic and cell wall-degrading enzymes on the performance of growing broilers fed wheat-based diets containing different calcium levels. Poultry Science 79, 66-76.

Closing session


353
Grain legumes and oilseeds – the way ahead G.D. Hill Plant Sciences Group, Soil, Plant and Ecological Sciences Division, PO Box 84, Lincoln University, Canterbury, New Zealand Introduction This session concludes this 4th International Workshop on legume seeds and oilseeds. An advantage of having done this task at the end of the last meeting is that it gives me a good overview of what was discussed then and what has been discussed at this meeting. Since we last met plant proteins have become more important. The ban, in Europe, on the feeding of animal based protein meals back to animals has increased the demand for plant protein concentrates for the feeding of both monogastric and ruminant animals. While these proceedings were being prepared there were new reported outbreaks of Bovine Spongiform Encephalopathy (BSE) or ‘mad cow disease’ in both Canada and the United States. Such outbreaks, even if they are sporadic, will keep the demand for safe, plant based, proteins high.
Before considering the presented papers it is appropriate to review changes in global production of grain legumes and oilseeds since we met in Wageningen in 1998. Annual world production of soybean (Glycine max) increased from 160 million t to about 180 million t between 1998 and 2002. Annual production of peanuts (Arachis hypogaea) in shell, the other major leguminous oilseed, has been reasonably constant at about 34 million t. However, world production of the other grain legumes has almost been static. In 1998 total world production was 55.7 million t. Since then production has varied from 53.0 million t to 56.6 million t. In other words, among the grain legumes over the last five years, there has only been a significant increase in the production of soybean. As far as soybean is concerned production in Africa, Australia and Europe is insignificant. The biggest single producer is still the United States, which produced 74.3 million t in 2002. Other big producers in the Americas are Brazil, where production has increased from 31.3 to 41.9 million t over the five years and Argentina (18.7 to 30.0 million t). In Asia China is a major producer producing about 17 million t.
Why is it that the production of the other grain legumes has not increased over the same period? Probably one of the major reasons is the lack of concentrated research inputs on the legumes that are enjoyed by the cereal crops. Over the five years from 1998 to 2002 more than 2,000 million t of cereals were produced every year. Among the individual cereals, yearly, more than 500 million t of wheat (Triticum aestivum), rice (Oryza sativa) and maize (Zea mays) are produced. Both budgets and programs for grain legume research at the CGIAR institutes have been reduced and in many countries there has been a reduction in expenditure on agricultural research.
There have not been the same breakthroughs in grain legume seed production that occurred with the development of hybrid maize and sorghum (Sorghum vulgare) and the breeding of dwarf rice and wheat genotypes. As an example at the time of Indian independence in 1947 the average yield of wheat in India was about 800 kg/ha and the yield of chickpea (Cicer arietinum) about 500 kg/ha. From then until now the wheat yield has quadrupled to more than 3 t/ha. The chickpea yield is virtually unchanged. There are similar figures for pigeon pea (Cajanus cajan) yield.
At this meeting we further extended the crops we have considered from the rapeseed (Brassica spp) that was discussed in Wageningen in 1998 to consider other non-legume

354
oilseed species that frequently leave a high protein residue after oil is extracted from the seed. Some of these like linseed (Linum usitatissimum), sesame (Sesamum indicum) and sunflower (Helianthus annuus) seed are grown primarily for their seed oil. Others, like cottonseed, are a by-product of cotton (Gossypium spp) fibre production.
As with the grain legumes the total production of these other oilseed crops has not varied greatly over the last five years. Rapeseed production has varied between 41.5 and 33.2 (the current level) million t. Annual cottonseed production is about 53 million t and sunflower is about 24 million t and was as high as 29 million t in 1999. Sesame seed and linseed production are minor and are approximately 2.8 and 2 million t/annum.
Overall therefore the total amount of high protein plant seed material, which is available for both human and animal feeding has not increased greatly, particularly when compared with cereal production, and apart from the soybean production it is either static or decreasing.
With regard to this workshop the move to Spain from the Netherlands has seen a greater emphasis on the use of these seeds as a human food and the advantages and disadvantages of their consumption. Papers from Cuba and Mexico have extended the range of legume species, which are considered and add another new high oil species (Jatropha curcas) from the family Euphorbiaceae.
Biotechnology and the continued development of genetically modified plants (GMOs) have advanced considerably since 1998 and there are now commercially cultivars of genetically modified soybean, rapeseed and cottonseed. A number of other grain legumes have been transformed but are still in the experimental stage. However, this aspect of plant genetic manipulation is likely to be increasingly important in the future in relation to resistance to abiotic stress, plant protection and the modification of seed composition to improve nutritional quality and possibly to produce pharmaceutical products. Analysis of antinutritional factors in legume seeds and oilseeds As in the past the workshop opened with a session that considered analytical methods for the determination of antinutritional factors (ANFs) in seeds. A comprehensive review considered advances in the analysis of phenolic compounds. It appears that the analysis of simple phenolics is now reasonably straightforward. However, there are still major problems associated with tannin analysis. Low molecular weight polymers can be separated but complex hydrolysable tannins and condensed tannins are still difficult to separate. Other plant components can interfere with the colour development and often only condensed tannins are measured even though plant material contains both condensed and hydrolysable tannins. A further problem is that the analyses require expensive and complex equipment and are generally not suited to the requirements of plant breeders who need to be able to rapidly screen large segregating plant populations at minimal cost and preferably non-destructively. It is perhaps fortunate for the plant breeders, that in the legumes, the absence of seed tannins is generally associated with the presence of white flowers and pale seed coats.
The other papers presented in this session considered a range of techniques for the determination of specific ANFs in legume seeds and oilseeds. An exception was a paper from Poland, which investigated the perception of the bitter taste of lupin (Lupinus spp) alkaloids. The paper reported the construction of model of the molecular matrix of the taste receptor. It was suggested that unless the alkaloid molecule could fit into the taste receptor it would not be perceived as bitter. The hypothesis was supported by sensory tests.
Following the theme of the main paper one paper reported on the level of phenolic compounds and pyrimidine glycosides in twenty Spanish Vicia narbonensis genotypes from the germplasm collection in Cuencea. Although the mean polyphenol concentration was

355
2.78 % there was a wide variation (2.02 – 5.03 %). All of the genotypes from the germplasm collection contained lower levels of vicine (mean 0.025 %) and convicine (mean 0.005 %) than Vicia faba and the L-DOPA concentration in the seed was negligible.
A paper from Spain measured variation in protein solubility and the ANFs in eight pea (Pisum sativum) cultivars from France, the Netherlands, Spain and the United Kingdom. They were unable to detect α-amylase or chymotrypsin inhibitor in any of the pea cultivars. Lectin levels were low. However, there were marked differences in the nitrogen solubility in sodium dodecylsulphate among the cultivars. It recommended the use of the cultivars Progress-9, Frission and Deso based on their potential nutritional value.
One paper in this session considered the alkaloids, which were present and their concentration in the developing seed of the tropical leguminous tree Erythrina americana. Given the generally accepted role of alkaloids as a deterrent to herbivory it is not surprising that the alkaloids were present throughout seed development and that level was high in mature seed. The major alkaloid present in the seed was β-erythroidine.
Possible substitute plant species for rapeseed for oil production, and their nutritional quality, was the topic of a paper from Germany. Twelve different plant species were tested for glucosinolates, inositol phosphates, sinapine and condensed tannins. Of the twelve species tested only three Lepidium sativum, Sinapis alba because of high amounts of glucosinolates and sinapine and Lesquerella fendleri, which contained a high amount of condensed tannins were considered to be potentially unsuitable for animal nutrition.
In the lowland tropics many of the common grain legumes grown in temperate regions cannot be cultivated. A joint Cuban/Spanish paper reported on the ANFs in a range of tropical legume species compared with soybean. The results showed there was considerable variation in the ANF levels among the eight legumes evaluated. Generally cowpea (Vigna unguiculata) had the lowest level of ANFs.
Some legumes such as Lima bean (Phaseolus lunatus) contain significant amounts of cyanogenic glucosides. Velvet bean (Stizolobium deeringianum) is amongst these plants. A rat feeding trial in Mexico showed that provided the seed was processed it was safe to eat. However, consumption of raw seed for a prolonged period caused histological damage to the kidney, liver, pancreas and lymph nodes.
A major problem arising from the consumption of legumes is the flatulence caused by the presence of α-galactosides. Three papers in this session dealt with this problem. Two from Poland measured their development in Lupinus luteus and L. mutabilis seed. A paper from Mexico considered their levels in L. montanus and L. stipulatus seed and at the same time measured seed phytate level. The results from Mexico indicated that the levels of these compounds in the Mexican lupin species were similar to those reported for other lupin species.
Two papers in this group considered the problems of the cost of analysis, the equipment required for the determination of ANFs in Brassica seeds and in legume seed, and the importance of non-destructive test methods for plant breeders. The measurement of residual trypsin inhibitors after processing of legume seed for industrial feed formulation was considered in a paper from the Netherlands. After heat treatment samples were defatted and protein was extracted by stirring for one hour. A simple equation allowed the calculation of the amount of residual trypsin inhibitor. An advantage of the method besides its simplicity was the availability of the results within one day. Plant breeders in Spain used near infrared reflectance spectroscopy to non-destructively measure the erucic acid, glucosinolates and acid detergent fibre in Brassica (Brassica spp) seed. They concluded, based on a comparison with traditional analysis, that the method was sufficiently accurate for selection purposes with R2 values ranging from 0.82 to 0.91.

356
Mechanism of action of antinutritional factors in legume seeds and oilseeds This session opened with a major review of the mode of action of ANFs in the gastrointestinal tract and their effect on tract microflora. It was suggested that lectins had a potential role in the reversal of small bowel atrophy and also possibly by stimulating gut function in the treatment of obesity, which is reaching epidemic proportions in the developed world. Lectins also have major insecticidal properties. Lectins have differential effects on tumour cells. It appears that some can suppress tumour cells while other may stimulate them. Finally it appears that lectins have the capacity to positively modify the gut microflora.
Among the trypsin inhibitors there is also the potential for their use in the control of insect pests, which attack plants. However, of more interest is the potential of Bowman-Birk inhibitor to suppress the growth of cancer cells particularly in association with phtytoestrogens and phospholipids. In Wageningen the report of this work was mainly based on in vitro studies. However, since then human clinical trials have commenced. The review also considered the role of polyphenols. Phytate may have a role in the prevention of pulmonary carcinogenisis and is also possibly able to reduce cell damage by oxygen radicals. As mention above the α-galactosides are responsible for the production of flatus. However, they also have the capacity to increase the level of bifidobacteria in the colon, increasing colon health and reducing the possibility of colon cancer. The review concluded with considering the possible role of saponins in reducing blood cholesterol levels by forming complexes with cholesterol in the gut and thus reducing its absorption.
The other four papers in this session considered different aspects of the effects of ANFs on metabolism. A paper from Mexico considered the anitmutagenic effect of the ANFs in Lupinus campestris seed. The response was measured in Salmonella typhimurium strain YG-1024. The phenolic compounds in the seed gave an 86 % inhibition. Similarly, low concentrations of the quinolizidine alkaloids were inhibitory but higher alkaloid levels increased the number of revertants.
A joint paper from the Netherlands and the United Kingdom considered the effect of kidney bean (Phaseolus vulgaris) and wheat (Triticum aestivum) lectins on the expression of the heat shock response, as measured by heat shock proteins, on gut epithelial cells. Rats fed on the two lectins had lower levels of gut heat shock proteins than animals fed on a control casein diet. These result were supported by in vitro experiments using Cacao-2 cells. The effect of the lectins in soybean and evening primrose (Oenothera paradoxa) was measured in rats in a paper from Poland. When the lectins were fed to rats at 0.1 g/day per kg body weight for 14 days there was a reduction in pancreatic trypsin and α-amylase activity, but an increase in the α-amylase activity in the pancreatic juice. The lectins also improved the plasma lipid profile. The lectins from the two plants agglutinated human red blood cells in different ways and after different times.
The final paper in this session considered the negative effect of feeding of soybean as a protein source to Atlantic salmon (Salmo salar). Three experiments were reported. The first showed that even at a 10 % inclusion in fish rations the soybean decreased brush border enzyme activity and the decrease was greatest at high soy inclusion levels. The second experiment measured the effect of soybean meal on intestinal nutrient transport. The soybean meal decreased carrier-mediated transport and increased the permeability the distal intestinal epithelium. The final experiment considered the effect of soybean on the immune and histochemical phenotype of intestinal mucosal cells. The soy meal reduced enzyme activity in both brush border cells and in the intercellular structures. There was also an increase in the number of monocytes present. However, there was little effect of the soy in the diet in the mid intestine of the fish.

357
Beneficial effects of antinutritional factors in human nutrition Again this session opened with a major review. There was some overlap of the information presented in this review paper and in the review presented at the start of the previous session. However, it reported on the effect of changing gut polyamine levels by the feeding of lectins on tumour growth. When mice were fed on kidney bean lectin it stimulated gut development and reduced tumour growth compared with control animals. It was proposed that interorgan competition induced by the lectin suppressed tumour growth. The potential anticarcinogenic effects of Bowman-Birk inhibitor, phytic acid and saponins were also discussed. As was foreshadowed at our meeting 1998 it was considered that there is an important future role of legume ANFs in disease management.
A paper from the United Kingdom compared soybean and pea protease inhibitors on the proliferation, in vitro, of human colon cancer cells (HT29). Although the protease inhibitors from both legume species suppressed cancer cell growth the greatest suppression was from the pea proteases. A further paper in this session tested the hypothesis that orally administered kidney bean lectin could act as an mucosal adjuvant. The work showed that mice given the lectin had reduced levels of serum soluble marker antigen-IgG compared with control animals thus showing that the lectin could induce oral tolerance to an unrelated marker antigen.
A paper from Spain took a different approach to the benefits of legume. It investigated the effect of consumption of chickpea on hypercholesterolemia in rats. The rats were either fed on control diet (with 5 % olive oil) or on a ration with 25 % coconut oil and 1 % cholesterol. After 26 days the latter group were hypercholesterolemic. They were then split into two groups, which either continued on the high fat diet or were fed chickpea as their protein source with 5 % olive oil. In the rats fed chickpea there were significant reductions total cholesterol, LDL-cholesterol, the total cholesterol/HDL ratio and liver cholesterol compared with rats fed on the high fat diet. It was concluded that including chickpea in the diet can reduce saturated fats and cholesterol and therefore improve cardiac health.
The final paper in this group proposed the need to establishment national databanks to record the antinutrients present in food. Because in the past these were seen to have no positive nutritional value such information is scattered and difficult to find. There is a particular problem with the use of different units of measurement in the literature preventing ease of comparison. It was proposed that a first step would be to analyse the foods, which are most commonly consumed in a country. The need to use common units related to the weight of the food was also stressed. Negative effects of antinutritional factors in human nutrition At previous meetings the development of allergenicity to soybean in pre-ruminant calves has been discussed. However this session opened with a major review of the negative effects of the consumption of legume seed on humans. It appears that there is a negative relationship between legume protein quality and increased loss of nitrogen in the urine. Legume protein also changes plasma amino acid concentrations. Levels of arginine, glycine and histidine tended to be higher while leucine, lysine and threonine were lower. Compared with casein-based diets the rate of protein synthesis and amino acid absorption was higher in animals fed casein-based diets.
Consumption of legume protein also affects lipid metabolism. However, at this stage there is debate as to whether the observed effects are due to the legume proteins or to other factors in the seed such as fibre, phytates or saponins. When rats were fed a casein based high

358
cholesterol diet daily feeding with legume 7S or 11S legume globulins reduced the cholesterol level. This effect has also been observed in humans where lower cholesterol levels were observed even when soybean replaced only part of the animal protein in the diet. The exact reasons for the observed changes are not clear. However, it is considered possible the legume protein amino acids affect the plasma insulin/glucagons ratio or thyroid hormone levels.
Most experimental work on the development of allergenicity to legume proteins has been with animals. However, the compounds responsible have not yet been identified. In calves it has been shown that soy β-conglycinin is highly allergenic. However, it seems that α-conglycinin, glycinin, Bowman-Birk trypsin inhibitor and lectins are also involved. The legume proteins involved in immune reactions are those that remain undigested for longest in both the small intestine and in the digestive tract.
In humans, because of the high consumption of peanuts and soybean in North America, there is considerable information on allergic responses to these two legume species. However, grain legumes are a major dietary component in much of the developing world and around the Mediterranean basin. There is also observed allergenicity to beans, chickpea and lentil (Lens culinaris).
Peanut allergy is common in both the United Kingdom and the United States. Estimates of the level of allergy in the population are from 0.6 to 1.5 %. Peanuts are responsible for most of the fatalities and as little as 2 mg of peanut protein can induce shock symptoms. There is less information on the effects of other legumes but in Spain lentil allergy was more common than peanut allergy in children. It is important to note that the inhalation of legume protein can cause respiratory symptoms. Lupinus albus flour causes allergic sensitisation in exposed workers. It is considered that the legume storage proteins are responsible for the development of allergenic reactions.
In a study of legume allergenic patients in Spain the majority (45 %) were allergic to peanuts. These were followed by chickpea (11 %) lentil and green beans (both 9 %). Most of the patients only reacted to one legume species but many reacted to a wide range of plants from other plant families.
Two further papers in this session considered immune reactions in weaned piglets. In the first paper the piglets were fed on casein or on mixture of casein and raw pea, faba bean, narrow-leafed lupin (L. angustifolius), desi or Kabuli chickpea or soybean for 28 days. Western-blotting indicated that most of the immunogenic proteins were from the 7S protein family. The faba beans and the chickpeas were the most immunogenic legumes. The format of the second experiment was similar but in this case the ileal digesta was analysed using western-blotting. The results support the previously stated hypothesis in that the main immunogenic proteins, which had survived digestion, were from the 7S family of pea, faba bean and chickpea. However, in narrow-leafed lupin nearly intact 11S proteins were detected.
A further paper in this session considered the ANFs present in a new plant species Jatropha curcas form Mexico. This species is in the same family as castor bean (Ricinus communis), which is highly toxic, and normally so is Jatropha curcas. However, apparently non-toxic genotypes of Jatropha curcas exist and are consumed. After oil extraction the residual cake contains up to 58 % protein. The protein digestibility was 86 to 90 % but even the low toxin containing line contained alkaloids, trypsin inhibitors and agglutinated red blood cells.
The final paper in this group provides an object lesson in the potential danger of consuming products with pharmacological properties, which have not been fully tested. Apparently the alkaloid gramine, which is present in Lupus luteus and L. hartwegii, has been used in Europe as a dietary supplement in the treatment of depression, epilepsy and Alzheimer’s disease. It is also used as a sedative, to modulate blood pressure and to reduce

359
nicotine withdrawal symptoms. Oral administration to a range of animals did not cause observable pathological changes but did lower haemoglobin concentration and the number of white blood cells. In the experiment gramine was injected directly into the brain of rats. The administration of gramine induced histopathological changes in the brains of the treated animals. The degree of brain damage being dose related. As a note of caution however, this does not necessarily mean that ingested gramine would have the same effect as most lupin alkaloids are rapidly eliminated from the bloodstream in the urine.
Effects of antinutritional factors on monogastric animals This series of international workshops arose from the interest of the animal feed industry in minimising the effects of ANFs in the rations, which they formulated. This session considered the effects of ANFs on monogastric animal production. The opening review covered latest developments in the use of peas, faba beans, lupins, chickpeas, which are currently mainly consumed by humans, and rapeseed (Brassica spp). However, for the first time at one of these workshops the potential of a number of other plant species was considered. The new plant species reviewed were linseed (Linum usitatissimum), cottonseed (Gossypium spp), and sunflower (Helianthus annuus). Among the latter species each contained ANFs that limited their potential use in monogastric animal rations. Linseed contains high levels of mucilage, which can reduce nutrient availability. Inclusion of linseed meal in layer rations has a negative effect on egg quality. A further negative factor in linseed is the presence of cyanogenic glycosides, which may not be completely destroyed during oil extraction. In the case of cottonseed the major limitation to its use for monogastric feeding is the presence of toxic gossypol. However, there are also concerns about low protein lysine levels and a high fibre content. It is possible to treat cottonseed to minimise in the effect of gossypol and to reduce the effect by adding iron to the ration. Cottonseed meal in layer rations can produce eggs with pink albumen and brown yolks. A high fibre content, unless seed is decorticated, and a number of phenolic compounds limit the nutritional value of sunflower seed meal. There has however been some work on the feeding of full fat sunflower seed to animals to change the composition of their body fatty acids.
A further eight short papers were presented in this session and covered a range of topics. Most closely relating to the review was a paper that compared muscle growth in rats when fed on casein, raw or extruded faba bean as the sole protein source. Animals fed raw faba bean had reduced weight gain and a higher feed intake than control animals. At the end of the experiment there was no difference in the weight of the gastrocnemius muscle, protein, RNA and DNA levels in rats fed extruded faba beans and the control ration. However, all measured values were significantly lower in rats fed raw faba bean. The raw faba bean diet significantly reduced the rate of protein synthesis. Also considering the effect of extrusion on ANFs was a paper from Spain, which reported on the effect of extrusion of red kidney bean (Phaseolus vulgaris) seed on intestinal enzymatic activity and amino acid uptake in the brush border membrane vesicles in rats. The protein (11 %) was either provider entirely by casein or by 5 % from casein and 6 % from either raw or extruded kidney bean. Rats fed raw kidney bean made virtually no growth and consumed only about 30 % of the amount eaten by animals on the other two rations. However their intestinal weight was increased by nearly 300 %. Animals fed the extruded kidney beans also had significantly higher L-leucine absorption and intestinal enzymatic activity.
Two papers considered the effect of processing to reduce the effect of ANFs in rations. The first reported on the effect of thermal processing of low glucosinolate rapeseed meal on its digestibility for pig growth from weaning to market. The experiment compared the

360
feeding of 350 g/kg of rapeseed meal as meal, after pelleting at 80 oC or after expanding at 100 oC and then pelleting. Pelleting significantly improved growth rate and ileal digestibility of dry matter, protein, phosphorus and calcium. Expansion prior to pelleting did not give any further improvement in the nutritional quality of the seed. The treatments had no effect on soluble dietary fibre or the available lysine level. The second paper, also from Belgium, considered the effect of adding commercial enzyme preparations to a soybean based weaned piglet ration on their performance, digestion gut flora and gut function. Although the animals tended to perform better on the enzyme containing ration the response was not significant. As was the case with a number of the other measured parameters. However, the enzymes did have a significant effect on gut microflora and reduced the antigenic effect of the soy protein in the foregut of the piglets. The authors suggest that the use of enzymes may be a way of increasing the level of soybean meal in piglet rations without deleterious effects.
Also considering soybeans and their ANFs were two papers from Poland. The first compared them in vivo in pigs and rats and by in vitro analyses to ascertain if the latter was sufficiently accurate to determine potential trypsin inhibitor activity. It compared soybean meal with raw and extruded soybeans. Feeding raw soybean decreased standardised ileal protein digestibility. However, there was only a difference in rats between soybean meal and extruded soybeans. The in vitro tests did not detect differences in trypsin inhibitor activity and it was concluded that it would be best to use rats as a model because of their sensitivity to trypsin inhibitors. The second paper was also involved with trypsin inhibitor activity but in this case in various commercial products derived from soybeans measured in rats and in vitro. Trypsin inhibitor activity was highest in the extruded soybean. Generally the true digestibility and the biological value of the soy-based products was lower than for casein except for the digestibility of soybean isolate and the biological value of soybean oil meal. There was a relationship between the in vitro values and protein solubility in KOH. It was suggested that these could partially replace N balance tests but the final determination of nutritional value would still require long-term animal growth experiments.
Also from Poland was a paper that measured the in vivo, using pigs and rats, and the in vitro digestibility of pea cultivars with different seed tannin levels. Digestibility was highest in low tannin pea cultivars. However, unlike the situation with the trypsin inhibitors in soybean products the in vitro test gave decreased digestibility with increased tannin level in the peas. It was therefore concluded that it could be used to rank pea cultivars for their amino acid digestibility for pigs.
A paper, which was similar to the rat experiment (above) with cholesterol, measured the effect of the addition of Lupinus albus in chicken diets on their blood cholesterol level. The chickens were given 0, 20 or 40 % L. albus with or without 1 % cholesterol. As lupin was increased in the diet bird performance was decreased. However, so were live fat, cecal pH, cholesterol and total biliary salts. Thus as with the previous experiment addition of the legume to the diet reduced the serum cholesterol level.
The final paper in this session considered the effect of adding carob (Ceratonia siliqua) seed to the diet of broiler chickens at 60 and 90 g/kg of diet. Carob seed contains up to 46 % gum. Its addition to the diet depressed nutrient utilisation. At 90 g/kg of diet apparent protein digestibility was reduced by 29 % and fat by 24 %. At the same time there was a marked increase in the viscosity of the jejunal digesta to such an extent that viscosity could not be measured as no supernatant could be obtained by centrifugation. If carob gives similar effects in mammals it possibly could have a role in obesity control.

361
Application of biotechnology and processing to reduce ANFs and to increase nutritional quality. Potential use of GMOs Genetic modification and plant breeding Since we met in 1998 commercial cultivars of genetically modified plants have been released and have already brought about a reduction in sales of agrichemicals in North America. This session opened with a review of what has been achieved by the application of biotechnology to plants. Among the legume and oilseed species discussed at this meeting most have now been genetically modified in some way.
Many of the early cultivars of GMO plants have related to increasing their production. A common theme has been the production of lines, which are resistant to the herbicide glyphosate or that have had the Bt gene added to them to reduce attack by lepidopteran insects. However, work is in progress to induce disease resistance, resistance to nematodes and to produce hybrid pollination systems by inducing male sterility where it does normally exist. Despite the considerable negative publicity these plants have enjoyed it seems probable that these genetic modifications are likely to have little effect on their nutritional quality. However, work is now under way to improve the nutritional quality of plant protein by genetic modification in particular to increase the level of the sulphur amino acids. This has been achieved in rapeseed, in narbon bean (Vicia narbonensis) and in narrow-leafed lupin. A further aim has been the modification of plant oils and a number of the plant species considered at this meeting have been modified. Carbohydrate composition has also been changed. Modification of starch could be of major importance in modifying the glycemic index of a food. Gene technology should also allow for the modification of plant secondary metabolites. An example here, which has received considerable publicity, is the ‘golden rice’. which contains considerably increased levels of β-carotene. Of indirect importance to this meeting is the possibility of increasing the rate of photosynthesis, enhancing yield and increasing plant resistance to abiotic stresses such as drought, salinity, low or high temperatures, which will all potentially lead to increased food security particularly in the developing world.
The three short papers in this session were all concerned with traditional plant breeding and in two of them there was a return to the theme of the need for rapid and cheap testing of segregating populations. The papers were concerned with vicine convicine and tannin levels in faba bean and with trypsin inhibitor in pea seed. White flowered faba beans have a low tannin level and the vicine convicine level is considerably lower in plants containing the zyc gene, which is associated with a white seed hilum. Bulked sergeant analysis was used to identify RAPD markers that were closely linked in bulks of an F2. The results will be used to produce a linkage map and will assist in selection in faba bean breeding programs. The second paper sought facile markers and reported on the development of simple markers to speed breeding progress. As already indicated selection for white hilum in faba bean reduces vicine and convicine. In the peas gene polymorphism linked to trypsin inhibitor activity has allowed the definition of primers for polymerase chain reaction assays which can also be exploited in breeding programs.
The third paper in this group was different and investigated the possibility of using the α-galactoside levels in three Catalan landraces of Phaseolus vulgaris as a means of uniquely identifying them for registration with a Protected Designation of Origin. The three lines were grown at different locations and were compared. Unfortunately edaphic factors had a greater influence on seed α-galactoside level than the genetic differences. In particular the variation in the level of α-galactosides was highly correlated with the soil cation exchange capacity

362
(r = -0.97). It was therefore concluded that the results would not be of assistance in the registration of these landraces for Protected Designation of Origin. Processing While feed formulators are waiting for plant breeders to breed out the worst aspects of the ANFs from these plants they still have to be used in human diets and in animal feeding. To render them safe they therefore need to be processed in some way before their consumption. The final section opens with a review of processing techniques. As indicated in my summary at the end of the last meeting most seed ANFs are rendered safe for human consumption by the normal processing that occurs before they are eaten (Hill, 1998).
The effect of hydrothermal treatment depends on a number of factors, which include cultivar, particle size and seed moisture level. Generally moist heat is more effective than dry heat at reducing ANFs but the different ANFs tend to respond differently to heating. Extrusion can also be used to modify ANFs. Again the moisture content of the seed, extrusion temperature and the feed rate of the extruder were important. However, suitably selected extrusion conditions can decrease phytic acid, tannins, phenols, α-amylase and trypsin inhibitors.
It has long been known that germination modifies seed composition. Germination can reduce raffinose sugars and trypsin inhibitor activity. However, it has also been shown to increase lectin concentration in lentil as the lectins were not degraded during the first six days of germination while other proteins were being broken down. As a side benefit there is a marked increase in vitamin C levels with germination. Microwaving germinated seed followed by hot air drying reduced the α-galactoside level in seed. Soaking seed combined with ionising radiation can also improve seed nutritional quality. In Asia a number of fermentation processes are used to improve the nutritional quality of legume seed. A technique that is also being used to improve nutritional quality of seeds is the addition of exogenous enzymes and various enzyme combinations have been tested.
The short papers presented in session 5 B looked at various aspects of several of these methods of ANF reduction. Two papers from Mexico considered the effect of heat treatment in hot water, an acid and an alkali solution on the protein quality and the level of alkaloids, phenols and α-galactosides in Lupinus campestris seed. In the first experiment in the absence of methionine supplementation the rats died. However, once supplemented the seed gave protein efficiency ratios ranging from 1.48 to 2.04 compared with casein at 2.45. This would suggest that this species has a seed protein value similar to that of other lupin species. In the second experiment the treatments removed 90 % of the alkaloids, 60 % of the α-galactosides and 65 % of the phenols in the seed.
A series of three papers from France reported on the effect of an instantaneous drop in pressure (Détente Instantanée Contrôlée) on the ANFs in rapeseed meal, soybean and Lupinus albus and L. mutabilis. In the process moistened seed is placed in the chamber of the apparatus under pressure and is heated. The pressure is then instantaneously released. One advantage of the method is that the seeds can be left entire. Another is the short processing processing time that is required. The process reduced the glucosinolate level in rapeseed meal by 40 %, trypsin inhibitor level in soybean meal by up to 94 % and the phytate levels in the two lupin species by 55 to 60 %.
Three papers considered changes in seed composition during germination. In two papers the species used was Vicia faba. In the first paper the qualitative changes in the storage proteins of the cotyledons were monitored using SDS-PAGE electrophoresis. Major changes in the proteins occurred 3 days after imbibition and the vicilin breakdown of started 1 to 2

363
days after imbibition. The second experiment involved two cultivars of germinating V. faba seed. Changes in the vicine convicine and L-DOPA levels in the germinating seed were monitored. Apparently, because of their L-DOPA content, faba bean seed is taken for the treatment of Parkinson’s disease, hypertension, renal failure and cirrhosis. This is because L-DOPA is a precursor of the neurotransmitter dopamine. The results showed that during germination there was a considerable increase in L-DOPA level in the developing seedlings and a corresponding fall in the α-galactosides. Germinated faba bean seed is therefore potentially a good source of L-DOPA. The final germination paper reported on the hydrolysis of phytate by the endogenous enzymes present in sesame (Sesamum indicum) seed. The results indicated that the endogenous enzymes during germination had almost no effect on the seed phytate (IP6) concentration.
The final paper in this session reported on the effect of the addition of the enzymes α-galactosidase, tannase, and viscozyme on the phenolic compounds present in lentils and in peas. Treatment of lentil with the enzymes increased antioxidant activity but they decreased it in peas. General conclusions Like any good scientific meeting this one has probably created as many questions as it has answered. As foreshadowed at our last meeting the ANFs are now seen as being beneficial when consumed under certain circumstances. However, even if processed they can cause serious allergenicity in some individuals. As many of the ANFs are secondary metabolites such as the lupin alkaloids they often can be bred out without a major negative effect on plant growth. However, others such as the lectins have a major role in Rhizobium legume recognition and their removal would probably have a negative effect on biological nitrogen fixation. Similarly the breeding out of tannins from legume seed leaves stored seed much more prone to insect attack. The continued mapping of plant genomes should allow a decision as to which plant processes can be blocked without having a negative effect on plant growth and production. In the meantime these seeds will continue to be an important dietary item for both humans and the animals that we keep. The future It appears that there is still much work for everyone working in this field. For a start why is that a legume species which can yield 4 to 5 t/ha in a country such as France or New Zealand produces less than 1 t/ha in India or Bangladesh. Even more interesting is why do they produce up to 3 t/ha in these countries at a research station but only 500 to 700 kg/ha in a farmer’s field. For our medical friends what is the mechanism that makes a small proportion of the total population develop an allergy to a legume seed, which can be fatal while the vast majority do not?
Probably the biggest issue to be faced in the future will be the testing of GMOs to ascertain their safety for both human and animal feeding. This is an area of science, which has led to highly polarised opinions. The plant breeders assure us that with biotechnology they know exactly what has been transferred into the new genotype unlike the situation with conventional plant breeding where only the phenotype and not the genotype is known. However, the general public are no longer prepared to take the pronouncements of scientists on trust and increasingly want to be assured that their food is safe to eat whether it has been sprayed with a plant protection chemical or has been genetically modified. As biotechnology

364
is gaining increased acceptance in the world it is probably an area that will feature at our next meeting.
The other area that has been touched on at this meeting is the potential of currently under exploited high protein legume seeds particularly from the tropics such as Lathyrus sativus, where both protein and energy deficiency are probably at their greatest. Such new species will all require evaluation as it is highly probable that in their wild state they will contain high levels of ANFs.
It would also be interesting to know if some of the epidemiological observations made about the consumption of legume seed in human diets have a sound medical foundation. For instance is the lower incidence of bowel and breast cancer in Japan due to their consumption of soy protein compared with the rest of the developed world or to some other unrelated reason? Can the phytoestrogens in soybean really reduce the level of osteoporosis in post-menopausal women? Is the benefit attributed to the Mediterranean diet due to the amount of olive oil consumed or the eating of more legume seed than in most of the rest of the developed world?
In conclusion it appears there are still many questions, which we still need answers for. See you all in five years time. References FAO, 2004. FAOSTAT Database. Downloaded 7 January 2004. Hill, G.D, 1998. Overview and recommendations for the future. In Recent Advances of
Research in Antinutritional Factors in Legume Seeds and Rapeseed. A.J.M. Jansman, G.D. Hill & A.F.B. van der Poel (Eds.). Proceedings 3rd International Workshop on Antinutritional Factors in Legume Seeds and Rapeseed, Wageningen, 8-10 July 1998, 463-473.

365
Closing comments A. Brenes CSIC, Instituto del Frío, Departamento de Metabolismo y Nutrición, José Antonio Novais, 10, 28040 Madrid, Spain. Only five years after the 3rd ANF Workshop held in Wageningen, The Netherlands, we are pleased to say that interest in grain legume and oilseed research remains strong based on the number and quality of papers presented.
This 4th ANF Workshop is the first to be held outside of Wageningen and we appreciate the initiative of the all the Dutch scientists from the Animal Science Groups in Wageningen who invited us to organise this event.
Plant seeds proteins are a valuable commodity and widely used in both human and animal nutrition. Their latter role has assumed greater importance with recent restrictions on the use of animal-based protein sources. The presence of biologically active compounds in legume and oilseeds has long been recognized. This is particularly the case for the protein antinutrients and other compounds. However these are not always harmful, but are biologically active and can modify, and sometimes to improve, gut function and the body metabolism. They can also be used to alter the bacterial flora, health status and the general metabolism of humans and animals.
Oilseeds from other plant families are also emerging as key sources of functional phytochemicals (mucilage, oil), and in addition are the richest sources of α-linolenic acid that is an alternative means of increasing the long-chain n-3 polyunsaturated fatty acid in animal tissues.
A major area of interest to those of us who work with these compounds is the possibility of using biotechnology to modify their content. This has the potential to both improve the nutritional value of the seed for animal and human food or to increase their level to make the seed unattractive to insects, which consume much of our agricultural production between the field and the table. Already in some parts of the world the use of agricultural chemicals is declining as a result of the plant genetic modification of plants. This area of science has so far only been applied to a few grain legumes – particularly soybean in the United States. When we next meet, it would be good if we had more presentations from this area of science.
As with the objectives of the past three workshop, the aim of this book was to help food and feed producers understand the biological consequences of the presence of these compounds in food and feed, to stimulate interest in our scientific endeavours aimed at eliminating this major obstacle to the more efficient utilisation of nutrients in our diet and to improve both the nutritional value and safety of the diet.
Attaining the objectives of any organisation depends of the support and contribution of the participants. For that reason, we appreciate the high quality of your submitted work and the opportunity to discuss common aspects of our research.
As a member of the Organising Committee, I also wish to express my sincere appreciation to the sponsors of this ANF workshop. Without their financial donations to its organisation the workshop would not have been possible.
We hope that this 4th ANF workshop has been a success for all the participants and that everyone took advantage of both the formal presentations and informal exchanges for the development of future research collaboration.
Finally, I also hope that this workshop will have be followed by a 5th Workshop and I wish all the best for your research on ANFs.


367
Author index Allaf, K 323, 329, 333 Altares, P 307, 317 Alzueta, C 239 Aranguren, P 255 Arias, M 29 Arija, I 219 Ariza, N 35, 43 Avila, C.M 287 Bakke-McKellep, A.M 111 Bañuelos-Pineda, J 177 Bardocz, S 87, 107 Barrenetxe, J 235, 255 Barrientos, L 39 Beltran, P 277 Boer, H 77 Bosch, L 277 Brenes, A 195, 219, 365 Buraczewska, L 243, 247 Burbano, C 53, 307, 317 Casañas, F 277 Cazorla, A. 5 Centelles, E 277 Centeno, C 219, 239 Chel-Guerrero, L 185 Clemente, A 137 Crespo, J.F 159 Cuadrado, C 53, 307, 317 Cubero, J.I 287 Czerwiński, J.J 115 Czubak, M 347 Dale, T.M 123 Dávila, G 101 Dávila-Ortíz, G 337, 343 de Groot, J 77 De Haro-Bailón, A 49 de los Mozos, M 29 de Schrijver, R 225 de Vos, S 225 Decuypere, J 229 del Río, M 49 Díaz, M.F 43 Dierick, N 229 Domoney, C 137, 261, 281 Duc, G 281, 287 Dueñas, M 311 Esteban, R.M 35 Estrella, I 311
Ewen, S 107 Fernández, C 159 Florez, A 277 Font, R 49 Fortuna, T 347 Freire, J.P.B 181, 189 García-López, P.M 53, 177 García-Mateos, R 57 Garzón de la Mora, P 53, 177 Gelencser, E 143 Genovese, M.I 123 Górecki, R.J 73 Goyoaga, C 307, 317 Grant, G 143 Greiner, R 261, 333 Guillamón, E 307, 317 Gurrola Díaz, C.M 53 Gutierrez, N 287 Haddad, J 323, 329, 333 Hajos, Gy 293 Hendriks, H 107 Hernández, T 311 Hernández-Torres, A 177 Hill, G.D 353 Ibañez, F 235 Isaac Virgen, M.L 53 Jaime, L 43 Jansman, A.J.M 195 Jasiczak, J 81 Jiménez Martínez, C 337, 343 Jiménez, C 101 Johnson, I.T 137 Joyanes, M. 149 Kelly, D 143 Koninkx, J 107 Kostyra, H 115 Krogdahl, Å 111 Krzemiński, R 115 Lajolo, F.M 123 Lallès, J.P 181, 189 Leontowicz, H 115 Leontowicz, M 115 Loarca-Piña, G 101 López-Andréu, F.J 35, 43 López-Dellamary, F 39 Macarulla, M.T 153 MacKenzie, D.A 137

368
Makkar, H.P.S 11 Marget, P 281, 287 Marquardt, R.R 195 Martín-Cabrejas, M.A 35, 43, 87 Martínez, J.A 153 Martínez, M 57 Martínez-Ayala, A.L 185 Martínez-Herrera, J 185 Marzo, F 235, 255 Matthäus, B 63 Michalczyk, D.J 73 Mika, M 347 Mollá, E 35 Molly, K 229 Mora Escobedo, R 337 Moreno, M.T 287 Muzquiz, M 7, 53, 307, 317, 343 Navarro, F 39 Nuez, F 277 Ortiz, L.T 29, 239 Osagie, A.U 293 Ovelgönne, H 107 Page, D 281 Pastuszewska, B 251 Pedrosa, M.M 53, 307, 317, 343 Pineda, J 39 Piotrowicz-Cieślak, A.I 69, 73 Portillo, M.P 153 Pryme, I.F 123 Pujolà, M 277 Pusztai, A 87, 107 Rebolé, A 239 Rodríguez, A 39 Rodríguez, J 159 Rodríguez, M.L 239 Romero, C 307 Rubio, L.A 159 Ruiz Lopez, M.A 53, 177 Ruíz, M 39 Salgado, P 181, 189 Santidrian, S 235, 255 Soto-Hernández, M 57 Stobiecki, M 11 Suso, M.J 287 Święch, E 243, 247, 251 Taciak, M 243, 247, 251 Torres, A.M 287 Treviño, J 239 Urdaneta, E 235, 255 van der Poel, A.F.B 77
van Dijk, J 107 Vande Ginste, J 225 Vanderbeke, E 229 Vargas, J 39 Vidal–Valverde, C 43 Viveros, A 219 Wolters, I 77 Wysocka, W 81 Zulet, M.A 153 Żyła, K 347

369
Keyword index α-galactosides 53, 277, 343 ADF 49 alkaloids 57, 81, 343 allergy 159 amino acids 243, 247 Andean lupin 73 antibodies 229 antimutagenicity 101 anti-nutrients 333 antinutritional
factors 29, 35, 39, 43, 63, 293 antioxidant activity 311 anti-proliferation 137 Aspergillus niger 137 beneficial applications 87 Brassica 49 Brassica napus 323 carbohydrates 101 cell line 107 Ceratonia siliqua 239 chemoreception of taste 81 chickens 219 chicks 239 cholesterol 153, 219 chromatography 11 Cicer arietinum 153 conformation of bis-quinolizidine
alkaloids 81 controlled instantaneous pressure
drop 323, 329, 333 convicine 317 crop plants 261 cyanogenic glucosides 39 databank 149 D-chiro-inositol 69 debittered 343 detoxification 337 digestibility 239, 243, 247, 337 environmental correlations 277 enzymes 255, 311 erucic acid 49 Erythrina americana 57 expansion 225 extrusion 255 extrusion cooking 235 fat 153 galactosyl cyclitols 69, 73
gas chromatography-mass spectrometry 57
gastrocnemius muscle 235 genetic correlations 277 genetic modification 261 germination 307, 347 glucosinolates 49, 63, 323 Glycine max 115 gramine 177 haemagglutination 185 harmful effects 87 heat shock proteins 107 hypercholesterolemia 153 ileal digestion 189 immune functions 111 in vitro digestibility 347 in vitro 243, 247 indole alkaloids 177 inhibitory domains 137 inositol phosphates 63 input traits 261 intestinal absorption 255 intestinal viscosity 239 ionisation 11 Jatropha curcas 185 L. mutabilis phytate 333 L. stipulatus 53 landraces 277 L-DOPA 317 lectins 107, 115, 123 legumes 153, 159, 255, 293 legume agglutinin 143 legume proteins 181, 189 Lens culinaris 311 lipid metabolism 159 low molecular weight ANFs 87 lupin 177 Lupinus albus 219, 333 Lupinus campestris 337, 343 Lupinus campestris phenolic
compounds 101 Lupinus luteus 69 Lupinus montanus 53 Lupinus mutabilis 73 marker-assisted selection 281 mass spectrometry 11 maturing 73

370
mobilisation 307 molecular markers 287 morphological changes 177 mucosal adjuvant 143 mucosal antigen uptake 143 neurons 177 NIRS 49 nitrogen compounds 35 non-nutrients 149 nutrient transport 111 nutrition 287 nutritional quality 261 nutritional value 251 Oenothera paradoxa 115 oilseeds 63, 73, 293 oral tolerance 143 output traits 261 pancreatic enzyme activity 115 peas 35 pelleting 225 Phaseolus vulgaris 255, 277 phenolics 311 phenolic compounds 11, 39, 343 physiology 57 phytase myo-inositol phosphate 347 phytates 53, 347 phytic acid 123 piglet 181, 189 pigs 225, 243, 247 piñoncillo 185 Pisum sativum L. 137, 243, 281, 311 plasma IgG 181 polyphenolic compounds 29 processing 323, 333 processing technology 293 protease inhibitors 137 proteases 229 protein 87
metabolism 159 quality 337 solubility 251
pyrimidine glucosides 317 pyrimidine glycosides 29 quinolizidine alkaloids 101 raffinose family 73 rapeseed 323 rapeseed meal 225 rat 115, 235, 243, 247 rat small intestine 107 Salmo salar 111
salmonid digestion 111 saponins 123 seed storage proteins 307 seedling growth 307 Sesamum indicum seed 347 sinapine 63 soybean(s) 111, 329
meal 229 proteins 251
Stizolobium deeringianum 39 storage proteins 159 tannins 11, 243, 287 theory of taste 81 tropical legumes 43 trypsin inhibitor 123, 185, 281 trypsin inhibitor activity 247, 251, 329 tumour growth 123 varieties 35 Vicia faba L. 235, 281, 287, 307, 317 Vicia narbonensis 29 vicine 317 vicine-convicine 281, 287 weaning 181, 189 yellow lupin seed 69































