Embed Size (px)
Citation preview

ORIGINAL PAPER
Modulation of transcription factor and metabolic pathwaygenes in response to water-deficit stress in rice
Swatismita Ray & Prasant K. Dansana & Jitender Giri & Priyanka Deveshwar &
Rita Arora & Pinky Agarwal & Jitendra P. Khurana & Sanjay Kapoor &
Akhilesh K. Tyagi
Received: 3 April 2010 /Revised: 10 August 2010 /Accepted: 16 August 2010 /Published online: 7 September 2010# Springer-Verlag 2010
Abstract Water-deficit stress is detrimental for rice growth,development, and yield. Transcriptome analysis of 1-week-old rice (Oryza sativa L. var. IR64) seedling under water-deficit stress condition using Affymetrix 57 K GeneChip®has revealed 1,563 and 1,746 genes to be up- anddownregulated, respectively. In an effort to amalgamatedata across laboratories, we identified 5,611 differentiallyexpressing genes under varying extrinsic water-deficitstress conditions in six vegetative and one reproductivestage of development in rice. Transcription factors (TFs)involved in ABA-dependent and ABA-independent path-ways have been found to be upregulated during water-deficit stress. Members of zinc-finger TFs namely, C2H2,C2C2, C3H, LIM, PHD, WRKY, ZF-HD, and ZIM, alongwith TF families like GeBP, jumonji, MBF1 and ULTexpress differentially under water-deficit conditions. NAC(NAM, ATAF and CUC) TF family emerges to be apotential key regulator of multiple abiotic stresses. Amongthe 12 TF genes that are co-upregulated under water-deficit,salt and cold stress conditions, five belong to the NAC TFfamily. We identified water-deficit stress-responsive genes
encoding key enzymes involved in biosynthesis of osmo-protectants like polyols and sugars; amino acid andquaternary ammonium compounds; cell wall looseningand structural components; cholesterol and very long chainfatty acid; cytokinin and secondary metabolites. Compari-son of genes responsive to water-deficit stress conditionswith genes preferentially expressed during panicle and seeddevelopment revealed a significant overlap of transcriptomealteration and pathways.
Keywords Oryza sativa L. var. IR64 .Microarray .
Transcriptome analysis .Water-deficit stress .
Metabolic pathways . Transcription factors
Introduction
Rice is a moisture-loving plant and, thus, its production isadversely affected by drought, which alone causes loss ofapproximately 18 million metric tons of rice productionworld-wide (Widawsky and O’Toole 1990; Evenson 1996).Moreover, global climate change is increasing the threat ofwater stress in near future (Tuba and Lichtenthaler 2007).Hence, identification of key genes, involved in regulation ofthe complex trait of drought tolerance and their manipulationby molecular breeding and/or genetic engineering fordevelopment of drought tolerant varieties, have becomestrategically important. Several quantitative trait loci associ-ated with drought tolerance have been identified throughphenotyping and marker-assisted selection (Tuberosa andSalvi 2006). Generation of ESTs from rice tissues subjectedto drought has also been an effective approach foridentifying genes associated with drought stress (Babu etal. 2002; Reddy et al. 2002; Markandeya et al. 2005;Gorantla et al. 2007; Varshney et al. 2009). The completion
Electronic supplementary material The online version of this article(doi:10.1007/s10142-010-0187-y) contains supplementary material,which is available to authorized users.
S. Ray : P. K. Dansana : J. Giri : P. Deveshwar : R. Arora :P. Agarwal : J. P. Khurana : S. Kapoor :A. K. Tyagi (*)Interdisciplinary Centre for Plant Genomics and Department ofPlant Molecular Biology, University of Delhi South Campus,New Delhi 110021, Indiae-mail: [email protected]
Present Address:A. K. TyagiNational Institute of Plant Genome Research,Aruna Asaf Ali Marg,New Delhi 110067, India
Funct Integr Genomics (2011) 11:157–178DOI 10.1007/s10142-010-0187-y

of genome sequences from several plant species, likeArabidopsis, rice, poplar, grape, papaya, Medicago, lotus,tomato, sorghum, and maize, and advances in the high-throughput techniques have revolutionized the gene discov-ery process followed by global gene expression (Vij andTyagi 2007; Paterson et al. 2009). Expression Microarrayplatform has been widely used in identifying genes involvedin abiotic stress tolerance (Rensink and Buell 2005).
In 2001, Seki et al. identified 44 cDNAs expressingdifferentially in Arabidopsis under drought and cold stressusing a microarray of ~1,300 full-length cDNAs. Subse-quently, Seki et al. (2002b) identified 277 drought-induciblegenes from Arabidopsis. Other, more extensive microarraystudies in Arabidopsis have revealed that initial perception ofstress might not be very specific to individual stress but laterthey attain specificity by involving transcription factors,signaling components and metabolic pathways (Kreps et al.2002; Oono et al. 2003; Kilian et al. 2007).
Abscisic acid (ABA) is known to have important role indrought stress response. Seki et al. (2002a) identified 299ABA responsive genes, 155 of which were upregulatedunder drought stress condition as well. Effect of ABAtreatment on guard cells revealed 64 differentially expressinggenes (Leonhardt et al. 2004). A comparative study of water-deficit stress-induced genes from microarray experiments ofSeki et al. (2002a, b), Kreps et al. (2002), and Kawaguchi etal. (2004) helped identify 27 genes that were induced underall three conditions, falling in six functional categories, e.g.,metabolism, transport, signaling, transcription, hydrophilicproteins and unknown proteins (Bray 2004).
Transcriptome analysis of rice under water-deficit stresshas been carried out earlier (Cooper et al. 2003; Rabbani etal. 2003; Hazen et al. 2005; Lan et al. 2005; Wang et al.2007; Zhou et al. 2007). Lan et al. (2004) identified that ofthe 253 cDNAs involved in pollination/fertilization in rice,136 genes (Lan et al. 2005) coding for regulatory proteinsinvolved in signal transduction and gene expression werealso involved in dehydration. Under drought stress condi-tion, the regulation of transcriptome of two divergentcultivars with high- and low-osmotic adjustment capacitywas found to be remarkably distinct. Genes encoding forsucrose synthase, a pore protein, a heat shock protein, and aLEA protein, were found exclusively upregulated in high-osmotic adjustment variety which could be contributing tothe tolerance trait (Hazen et al. 2005). In another study,much higher number of genes was found to be upregulatedin sensitive variety of rice than tolerant variety underdrought stress condition. Interestingly, ribosomal andplastid protein synthesis genes were downregulated insensitive variety but not in tolerant variety; whereas, genesfor protein degradation, amino acid degradation, lipiddegradation were triggered in sensitive variety but not intolerant variety. Moreover, genes encoding for cytochrome
P450 were significantly upregulated in the tolerant varietybut not in sensitive variety. All these factors cumulativelycould be contributing to drought tolerance (Degenkolbe etal. 2009). Transcriptome analysis under water-deficit stresshas also been studied in various other species like chickpea,sunflower, barley, tobacco, Physcomitrella patens, Pinustaeda, Populus trichocarpa, Populus euphratica, Sorghumbicolor, Zea mays, Thellungiella salsuginea, and Vitis vinifera(Oztur et al. 2002; Rizhsky et al. 2002; Watkinson et al.2003; Yu and Setter 2003; Zheng et al. 2004; Brosche et al.2005; Buchanan et al. 2005; Andjelkovic and Thompson2006; Street et al. 2006; Wong et al. 2006; Cramer et al.2007; Cuming et al. 2007; Mantri et al. 2007; Roche et al.2007; Talame et al. 2007; Tattersall et al. 2007; Varshney etal. 2009). However, a comprehensive analysis of stress-responsive genes during various stages of plant life cycle isnot yet available.
In this study, an attempt has been made to identify andenlist the water-deficit stress-responsive genes from micro-array gene expression profiling of 1-week-old rice seedlingunder water-deficit stress condition. To achieve a consensuson water-deficit stress-responsive genes, a cumulativeanalysis has been performed considering various otherhigh-throughput studies across the globe. The water-deficit stress-responsive genes have thus been analyzed inseven developmental (vegetative and reproductive) stages,although water-deficit conditions varied widely. Here, wealso report a repertoire of genes commonly triggered bywater-deficit, cold, and salt stress conditions. Moreover, anoverlap between genes differentially expressed duringvarious stages of panicle and seed development and thoseresponsive to water-deficit stress condition has beenestablished.
Materials and methods
Plant material, growth condition, and stress treatment
The seeds of indica rice (Oryza sativa L. var. IR64,IET9671), after disinfection with 0.1% HgCl2 and thoroughwashing with reverse-osmosis (RO) water, were soakedovernight in RO water. Next day, seeds were spread on ameshed float and grown hydroponically at 28±1°C inculture room with a daily photoperiodic cycle of 14 h lightand 10 h dark. After 1 week of growth, the seedlings weregiven different stress treatments (Mukhopadhyay et al.2004). One-week-old seedling stage was chosen foranalysis to emphasize on transcriptome regulation atseedling survival in early stage of development underwater-deficit stress condition. Specifically, for cold stress,the seedlings were kept at 4±1°C in RO water, for saltstress the seedlings were transferred into a beaker contain-
158 Funct Integr Genomics (2011) 11:157–178

ing 200 mM NaCl solution and for water-deficit stress theywere air-dried on a Whatman 3 mm sheet at 28±1°C, andall stresses were given for 3 h. For control, 1-week-oldseedlings were maintained in water in 100 ml beaker for3 h. Different developmental stages of rice panicle wereobtained from field-grown rice (O. sativa ssp. indica var.IR64). The young panicles were taken out from the sheathand measured to be categorized in six groups (P1, 0–3 cm;P2, 3–5 cm; P3, 5–10 cm; P4, 10–15 cm; P5, 15–22 cm,and P6, 22–30 cm) based on length of the panicle and thelandmark developmental events (Itoh et al. 2005), andfrozen in liquid nitrogen. The rice seed were tagged fromthe day of pollination (DAP), and developing seeds werecollected on each DAP from 0 to 30 DAP. These werepooled into S1, S2, S3, S4, and S5, representing, 0–2, 3–4,5–10, 11–20, and 21–29 DAP, respectively. Mature leaveswere harvested from the same plants.
Affymetrix GeneChip hybridization and data collection
Total RNAwas isolated from vegetative tissue of 1-week-oldrice seedlings (root and shoot tissue), the stress samples,mature leaf and panicle of rice and quality of the RNA wasascertained as described previously (Jain et al. 2006). Forisolating total RNA from rice seed, RNA isolation methodfrom carbohydrate-rich seeds was followed (Sharma et al.2003). The microarray analysis using Affymetrix GeneChip®Rice Genome Array was carried out according to Affymetrixmanual for one-cycle target labeling and control reagents(Affymetrix, Santa Clara, CA) using 5 μg of RNA as startingmaterial. Target preparation, hybridization to arrays, washing,staining, and scanning were carried out as described earlier(Jain et al. 2007; Ray et al. 2007). The cell intensity data files(*.cel) generated by the Gene Chip Operating Software(GCOS 1.2) (Schadt et al. 2001) were imported in ArrayAssist® software (Strandgenomics, Bangalore, India) forsubsequent data processing. The data from 12 chips werenormalised by using GeneChip robust multi-array average(GCRMA) algorithm (Wu et al. 2004). The correlationbetween the biological replicates were assessed usingPearson’s correlation coefficient (R) on the signal intensitiesand the R values between the three replicates were≥0.95 forfour stress experimental stages. For further data analysis, thethree replicates under each stress condition (water-deficit,cold, or salt) and control tissue were normalized as individualexperimental pairs by using GCRMA algorithm. The finaldataset after normalization contained 57,381 probesets fromwhich hybridization controls, TE-related and redundantprobesets, were removed after an extensive manual curation.The final number of unique probeset was determined to be37,927 (mentioned as genes in this study). The unique probesets include gene loci identified by TIGR (The Institute forGenomic Research; http://www.tigr.org/) and the KOME
(Knowledge-based Oryza Molecular biological Encyclopedia;http://red.dna.affrc.go.jp/cDNA/) cDNAs which were notpresent in the sequenced genome. The normalized data werelog2 transformated, and differential expression analysis wereperformed, using paired t test method. A gene was designatedas up- or downregulated if the signal ratios were ≥2 at pvalue <0.005 with respect to 1-week-old unstressed seedlingfor stress samples and mature leaf was considered as controlfor the reproductive developmental stages. To avoid lowexpressing genes under water-deficit, cold, and salt stresscondition, from the differentially up- and downregulatedgenes list, genes having average normalized intensity value≥50 in stress sample and control sample, respectively, wereconsidered for further analysis. The respective log trans-formed intensity values were used for hierarchical clusteringby using Euclidean distance matrix and K-Means. Forcomparative study of genes regulated by extrinsic andintrinsic (developmental) water-deficit stress, cell intensitydata files for mature leaf, six panicle stages (P1, P2, P3, P4,P5, and P6), and five seed stages (S1, S2, S3, S4, and S5)were used to make a project along with the cell intensity datafiles of stressed and unstressed 1-week-old seedlings.GCRMA normalized data was analyzed for differentialexpression and genes showing twofold changes at a p value≥0.05 were called as differentially expressed genes. Benja-mini–Hoschberg correction was applied for all t testsperformed. Co-regulation of water-deficit stress-responsivegenes with seed preferential expression was identified asthose genes that were at least two-fold upregulated in any ofthe seed stage with respect to any of the panicle stage suchthat the maximum signal intensity amongst the five seeddevelopment stages is higher than the maximum signalintensity amongst the six panicle development stages.Furthermore, they were filtered for those which had at leasttwo-fold upregulation in any of the seed stages with respectto mature leaf, which served as the vegetative control.Similarly, co-regulation with panicle preferential genes wasidentified where expression of panicle was up in comparisonto seed and mature leaf in water-deficit-induced genes.Further analyses were carried out in Microsoft Excel. Micro-array data from this article have been deposited in the GeneExpression Omnibus database at the National Center forBiotechnology Information under the series accession numb-ers GSE6893 and GSE6901.
Literature database analysis
Literature search was made to compile a list of alreadyknown water-deficit stress-related genes from differentcultivars, tissue, time points, level of stress, and platformsstudied. Only those studies were taken into considerationfor which data was available in retrievable form (Cooper etal. 2003; Rabbani et al. 2003; Wang et al. 2007; Zhou et al.
Funct Integr Genomics (2011) 11:157–178 159

2007). Gene IDs provided in these papers were mapped togene loci of TIGR version 5. Corresponding Affymetrixprobeset IDs for genes retrieved from published sourceswere searched from Rice Multi-platform Microarray Search(http://www.ricearray.org/matrix.search.shtml). Probe IDscorresponding to the 37,927 unique probe set were usedfor further analysis. The up- and downregulation of thesegenes were determined after comparing the data providedfor treated tissue and control tissue. Few genes we report tobe up- and downregulated in same tissue as we arereporting a concise data of all time course experiments.
Functional classification
Gene Ontology-based functional analysis
Genes responsive to water-deficit stress condition wereclassified according to the function of the protein theyencode according to Gene Ontology (GO) database (http://www.geneontology.org/). The underlying fact of the GOclassification is that every gene could be part of all threemain classification types i.e., molecular function, cellularprocess, and biological process. However, these numberswould change depending on the current state of ourunderstanding about that particular gene or protein.
Metabolic pathway analysis
From RiceCyc in GRAMENE (Jaiswal et al. 2006), wedownloaded metabolic pathway-associated genes whose IDwas clustering on metabolic pathways. Pathways werereconstructed using Adobe Illustrator® software.
Results
Identification and analysis of genes responsive to extrinsicwater-deficit stress during rice development
IR64 variety of indica rice was chosen for transcriptomeanalysis under water-deficit stress condition using RiceGenome Array. IR64, a semi-dwarf lowland variety withgood yield potential, is widely grown in irrigated area intropical Asia (Khush 1995; Narciso and Hossain 2002).Sampling for microarray gene expression study was doneafter 3 h of water-deficit stress to 1-week-old rice seedlingwhen relative water content (Barr and Weatherley 1962)was approximately 31–37%. Most of the previouslyidentified water-deficit stress-responsive genes, includingLEA protein (LOC_Os01g12580), dehydrin-Rab16B(LOC_ Os11g26780), Rab21 (LOC_Os11g26790),COR410 (LOC_Os02g44870), aquaporin-TIP3.1(LOC_Os10g35050), and DREB1 (LOC_Os04g55520)
showed increased transcript accumulation in our microarrayexperiment. The microarray data for abiotic stress-responsive genes of selected gene families have beenalready validated in our previous studies by quantitativePCR analysis (Agarwal et al. 2007; Arora et al. 2007; Jainet al. 2007; Ray et al. 2007; Nijhawan et al. 2008; Vij et al.2008). Under water-deficit stress condition, 1,563 and1,746 genes were differentially up- and downregulated,respectively. Initiative for identification of water-deficitstress-responsive genes in different cultivars of rice (japon-ica and indica), at varied developmental stages (2-, 4-, and6-week-old, 1-week-before-heading, and 4-tiller stage), andexperimental conditions has been taken-up across laborato-ries (Cooper et al. 2003; Rabbani et al. 2003; Wang et al.2007; Zhou et al. 2007) as listed in Table 1.
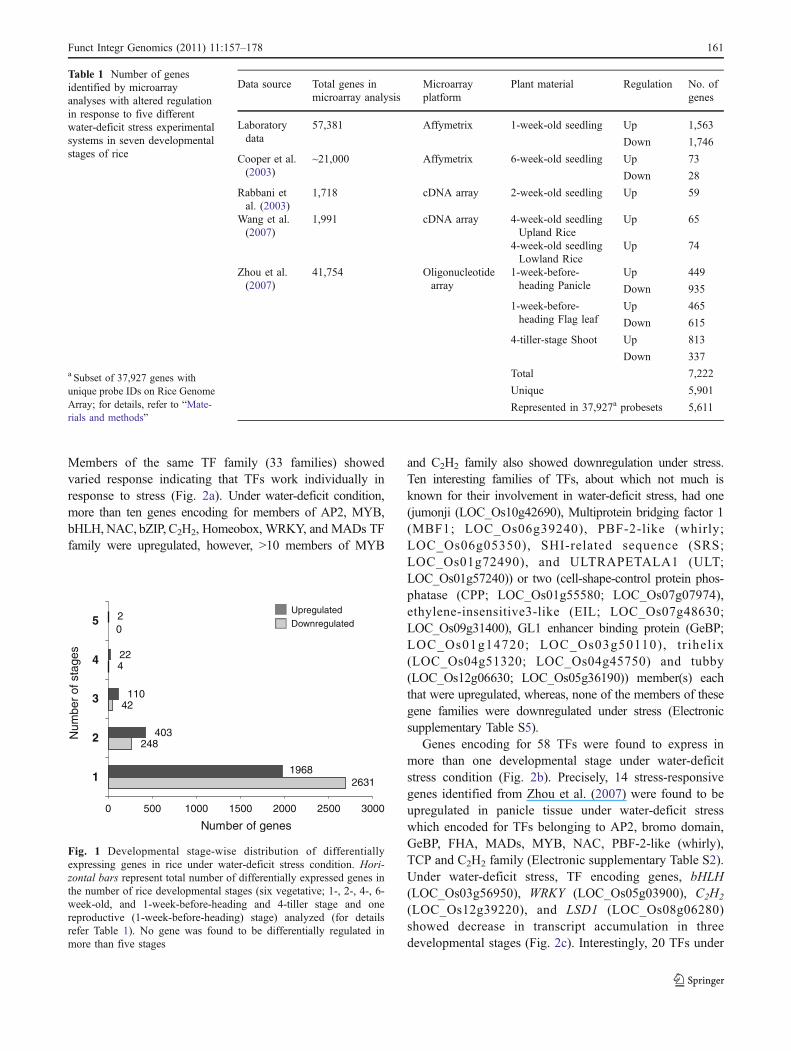
A total of 5,901 unique genes were thus identified afterremoving the redundant ones from the cumulative list of 7,222differentially regulated genes (Table 1). Among the 5,901unique genes, 5,611 were found to be represented in the37,927 subset of unique genes on Rice Genome Array whichhas been used for further analysis (Electronic supplementaryTable S1). Expression of genes under water-deficit stressresponse in seven developmental stages, including sixvegetative (1-, 2-, 4-, 6-week, 1-week-before-heading, and4-tiller stage) and one reproductive (panicle: 1-week-before-heading) stage, was found to be spatially and temporallyregulated. Under water-deficit stress condition, 2,505 and2,925 genes were found to be up- and downregulated,respectively, wherein, 181 genes were found to be up- aswell as downregulated in different studies implying that theirdifferential regulation in response to water-deficit stress maybe developmental stage-specific (Electronic supplementaryTables S2, S3, and S4). Maximum number of genes (1,968and 2,631 up- and downregulated, respectively) was found tobe expressing differentially under individual experimentalcondition, followed by those that showed differentialexpression under two or more experimental conditions(Fig. 1). A set of 24 genes (22 and 2 genes in four andfive developmental stages, respectively) were found to beinduced at many developmental stages under water-deficitstress. They were also involved in regulation (transcriptionfactors), signaling (kinase), and metabolism (CTP synthase,epimearse), although few genes remain to be assignedspecific function (Fig. 1).
Transcription factors expressing differentiallyunder water-deficit condition
The rice genome has been found to code for 2,314 TFsbelonging to 68 TF families (unpublished data). Out of 68TF families, at least one member of each of the 58 TFfamilies showed differential expression under water-deficitstress condition (Electronic supplementary Table S5).
160 Funct Integr Genomics (2011) 11:157–178
Members of the same TF family (33 families) showedvaried response indicating that TFs work individually inresponse to stress (Fig. 2a). Under water-deficit condition,more than ten genes encoding for members of AP2, MYB,bHLH, NAC, bZIP, C2H2, Homeobox, WRKY, and MADs TFfamily were upregulated, however, >10 members of MYB
and C2H2 family also showed downregulation under stress.Ten interesting families of TFs, about which not much isknown for their involvement in water-deficit stress, had one(jumonji (LOC_Os10g42690), Multiprotein bridging factor 1(MBF1; LOC_Os06g39240), PBF-2-like (whirly;LOC_Os06g05350), SHI-related sequence (SRS;LOC_Os01g72490), and ULTRAPETALA1 (ULT;LOC_Os01g57240)) or two (cell-shape-control protein phos-phatase (CPP; LOC_Os01g55580; LOC_Os07g07974),ethylene-insensitive3-like (EIL; LOC_Os07g48630;LOC_Os09g31400), GL1 enhancer binding protein (GeBP;LOC_Os01g14720; LOC_Os03g50110), tr ihel ix(LOC_Os04g51320; LOC_Os04g45750) and tubby(LOC_Os12g06630; LOC_Os05g36190)) member(s) eachthat were upregulated, whereas, none of the members of thesegene families were downregulated under stress (Electronicsupplementary Table S5).
Genes encoding for 58 TFs were found to express inmore than one developmental stage under water-deficitstress condition (Fig. 2b). Precisely, 14 stress-responsivegenes identified from Zhou et al. (2007) were found to beupregulated in panicle tissue under water-deficit stresswhich encoded for TFs belonging to AP2, bromo domain,GeBP, FHA, MADs, MYB, NAC, PBF-2-like (whirly),TCP and C2H2 family (Electronic supplementary Table S2).Under water-deficit stress, TF encoding genes, bHLH(LOC_Os03g56950), WRKY (LOC_Os05g03900), C2H2
(LOC_Os12g39220), and LSD1 (LOC_Os08g06280)showed decrease in transcript accumulation in threedevelopmental stages (Fig. 2c). Interestingly, 20 TFs under
248
42
4
403
110
22
20
0 500 1000 1500 2000 2500 3000
1
2
3
4
5
Number of genes
Upregulated
Downregulated
Num
ber
of s
tage
s
19682631
Fig. 1 Developmental stage-wise distribution of differentiallyexpressing genes in rice under water-deficit stress condition. Hori-zontal bars represent total number of differentially expressed genes inthe number of rice developmental stages (six vegetative; 1-, 2-, 4-, 6-week-old, and 1-week-before-heading and 4-tiller stage and onereproductive (1-week-before-heading) stage) analyzed (for detailsrefer Table 1). No gene was found to be differentially regulated inmore than five stages
Data source Total genes inmicroarray analysis
Microarrayplatform
Plant material Regulation No. ofgenes
Laboratorydata
57,381 Affymetrix 1-week-old seedling Up 1,563
Down 1,746
Cooper et al.(2003)
~21,000 Affymetrix 6-week-old seedling Up 73
Down 28
Rabbani etal. (2003)
1,718 cDNA array 2-week-old seedling Up 59
Wang et al.(2007)
1,991 cDNA array 4-week-old seedlingUpland Rice
Up 65
4-week-old seedlingLowland Rice
Up 74
Zhou et al.(2007)
41,754 Oligonucleotidearray
1-week-before-heading Panicle
Up 449
Down 935
1-week-before-heading Flag leaf
Up 465
Down 615
4-tiller-stage Shoot Up 813
Down 337
Total 7,222
Unique 5,901
Represented in 37,927a probesets 5,611
Table 1 Number of genesidentified by microarrayanalyses with altered regulationin response to five differentwater-deficit stress experimentalsystems in seven developmentalstages of rice
a Subset of 37,927 genes withunique probe IDs on Rice GenomeArray; for details, refer to “Mate-rials and methods”
Funct Integr Genomics (2011) 11:157–178 161
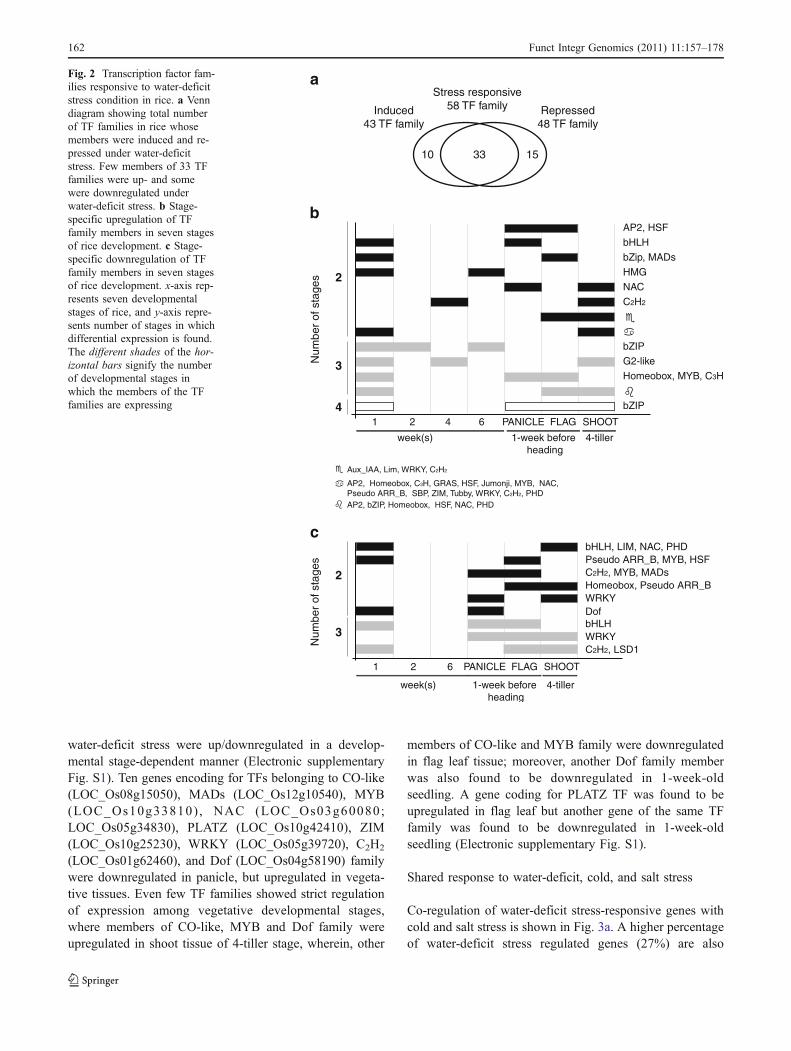
water-deficit stress were up/downregulated in a develop-mental stage-dependent manner (Electronic supplementaryFig. S1). Ten genes encoding for TFs belonging to CO-like(LOC_Os08g15050), MADs (LOC_Os12g10540), MYB(LOC_Os10g33810) , NAC (LOC_Os03g60080;LOC_Os05g34830), PLATZ (LOC_Os10g42410), ZIM(LOC_Os10g25230), WRKY (LOC_Os05g39720), C2H2
(LOC_Os01g62460), and Dof (LOC_Os04g58190) familywere downregulated in panicle, but upregulated in vegeta-tive tissues. Even few TF families showed strict regulationof expression among vegetative developmental stages,where members of CO-like, MYB and Dof family wereupregulated in shoot tissue of 4-tiller stage, wherein, other
members of CO-like and MYB family were downregulatedin flag leaf tissue; moreover, another Dof family memberwas also found to be downregulated in 1-week-oldseedling. A gene coding for PLATZ TF was found to beupregulated in flag leaf but another gene of the same TFfamily was found to be downregulated in 1-week-oldseedling (Electronic supplementary Fig. S1).
Shared response to water-deficit, cold, and salt stress
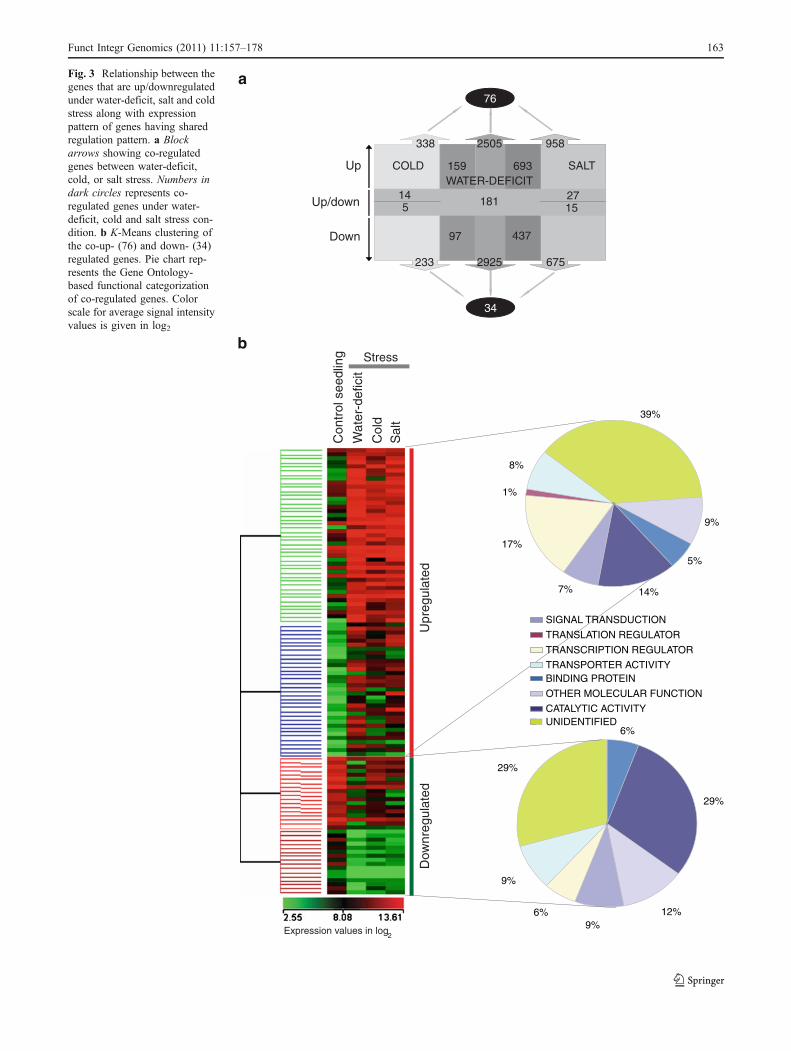
Co-regulation of water-deficit stress-responsive genes withcold and salt stress is shown in Fig. 3a. A higher percentageof water-deficit stress regulated genes (27%) are also
3310 15
Repressed48 TF family
Induced43 TF family
Stress responsive58 TF family
2
3
Num
ber
of s
tage
s
Dof
WRKY
WRKY
C2H2, LSD1
C2H2, MYB, MADs
bHLH, LIM, NAC, PHD
bHLH
Homeobox, Pseudo ARR_B
Pseudo ARR_B, MYB, HSF
week(s) 1-week before heading
4-tiller
1 2 6 PANICLE FLAG SHOOT
3
2
4
Num
ber
of s
tage
s
1 2 4 6
week(s)
PANICLE FLAG
1-week before heading
4-tiller
SHOOT
bZIP
bZIP
C2H2
NAC
HMG
bZip, MADs
bHLH
AP2, HSF
Homeobox, MYB, C3H
G2-like
Aux_IAA, Lim, WRKY, C2H2
AP2, bZIP, Homeobox, HSF, NAC, PHD
AP2, Homeobox, C3H, GRAS, HSF, Jumonji, MYB, NAC, Pseudo ARR_B, SBP, ZIM, Tubby, WRKY, C2H2, PHD
a
b
c
Fig. 2 Transcription factor fam-ilies responsive to water-deficitstress condition in rice. a Venndiagram showing total numberof TF families in rice whosemembers were induced and re-pressed under water-deficitstress. Few members of 33 TFfamilies were up- and somewere downregulated underwater-deficit stress. b Stage-specific upregulation of TFfamily members in seven stagesof rice development. c Stage-specific downregulation of TFfamily members in seven stagesof rice development. x-axis rep-resents seven developmentalstages of rice, and y-axis repre-sents number of stages in whichdifferential expression is found.The different shades of the hor-izontal bars signify the numberof developmental stages inwhich the members of the TFfamilies are expressing
162 Funct Integr Genomics (2011) 11:157–178
COLD SALTWATER-DEFICIT
338
159
2505
693
958
233
97
2925
437
675
145
2715
76
34
181
Up
Up/down
Down
Con
trol
see
dlin
g
Wat
er-d
efic
it
Col
dS
alt
Stress
Upr
egul
ated
Dow
nreg
ulat
ed
5%
14%7%
17%
1%
8%
39%
9%
6%
29%
12%9%
6%
9%
29%
SIGNAL TRANSDUCTION
TRANSLATION REGULATOR
TRANSCRIPTION REGULATOR
TRANSPORTER ACTIVITYBINDING PROTEIN
OTHER MOLECULAR FUNCTION
CATALYTIC ACTIVITYUNIDENTIFIED
Expression values in log2
a
b
Fig. 3 Relationship between thegenes that are up/downregulatedunder water-deficit, salt and coldstress along with expressionpattern of genes having sharedregulation pattern. a Blockarrows showing co-regulatedgenes between water-deficit,cold, or salt stress. Numbers indark circles represents co-regulated genes under water-deficit, cold and salt stress con-dition. b K-Means clustering ofthe co-up- (76) and down- (34)regulated genes. Pie chart rep-resents the Gene Ontology-based functional categorizationof co-regulated genes. Colorscale for average signal intensityvalues is given in log2
Funct Integr Genomics (2011) 11:157–178 163

affected by salt stress, however, only 6.3% genes werefound to be co-regulated between water-deficit and coldstress dataset. It was also observed that a homeobox-leucinezipper protein (LOC_Os02g43330), water stress-inducibleprotein Rab21 (LOC_Os11g26790) and seed maturationprotein (LOC_Os08g23870) encoding genes are among thefive genes showing highest differential regulation in 1-week-old rice seedling under water-deficit and salt stress(Electronic supplementary Table S6). Furthermore, amongthe genes co-regulated by both salt and cold stress, aMKKK2 (LOC_Os01g50420) and expressed protein(LOC_Os06g46140) genes exhibit greater degree of differ-ential expression under cold stress condition. Of the 181genes, which showed both up/downregulation of transcriptunder water-deficit stress in a tissue and developmentalstage-dependent manner, 14 and 5 genes, respectively, wereup- and downregulated under cold stress condition, where-as, more genes were co-regulated under salt stress from thiscategory of genes.
Of the 693 co-regulated genes under water-deficit and saltstress conditions, 76 were also upregulated under cold stress(Fig. 3a). These 76 genes code for proteins involved in signaltransduction, transcription and translation regulation, trans-porters and catalytic activity; function to a few proteins is yetto be assigned (Fig. 3b). The signal transduction componentgenes include proteins belonging to calcium-regulated cascade(namely calmodulin, EF-hand family protein and ATPases),kinases, phosphatases, heat shock proteins, transporters, andhormone action. More genes encoding for TFs were co-upregulated (17%) than co-downregulated (6%). One or twomembers of TF families namely, bHLH (LOC_Os08g42470),CPP (LOC_Os07g07974), C2H2 (LOC_Os03g60570;LOC_Os03g60560), MYB (LOC_Os04g43680), WRKY(LOC_Os06g44010), and AP2 (LOC_Os01g58420), wereamong the co-regulated genes; additionally, five genesencoded for NAC TF family (LOC_Os01g60020;LOC_Os11g03370; LOC_Os01g15640; LOC_Os01g66120;LOC_Os11g03300). Moreover, among these five NAC TFs,one gene (LOC_Os01g66120) was upregulated in uplanddrought tolerant variety of rice under water-deficit stress(Wang et al. 2007). These co-regulated TFs might be keyplayers in downstream responses induced by different kind ofabiotic stresses. Water-deficiency limits photosynthesis, salin-ity leads to ion toxicity and low-temperature directly affectscellular functioning, thus, collectively, they affect normalmetabolic processes of plant which is reflected in the declineof transcript accumulation of genes having catalytic activity.Among these downregulated genes, cytochrome p450 86A2(LOC_Os03g04530) , phosphoe thano lamine N-methyltransferase (LOC_Os05g47540), two expressed pro-teins (LOC_Os04g11120 and LOC_Os04g11060) andCRK10 (LOC_Os07g43570) encoding genes showed maxi-mum decline in transcript accumulation under water-deficit
stress in 1-week-old seedling (Electronic supplementary TableS6). More genes encoding for proteins having catalyticactivity were downregulated (29%) than upregulated (14%).
Functional categorization and pathway determinationof the water-deficit responsive genes
Gene Ontology-based analysis showed that 2,408 differen-tially regulated genes were involved in biological processesand molecular function could be assigned to 1,382 and1,611 up- and downregulated genes, respectively; however,a large number of differentially regulated genes (1,857)remains to be annotated (Electronic supplementary Fig. S2).
Ricecyc (http://www.gramene.org/pathway) databasewas further used for metabolic profiling of the up anddownregulated genes under water-deficit condition. Carbo-hydrate, energy, lipid, amino acid, nucleotide, cofactor,vitamin, and secondary metabolite metabolism were alteredunder water-deficit stress condition. Moreover, processeslike transcription, translation, replication and repair, fold-ing/sorting/degradation, transport and signaling were alsofound to be affected. While many functional categorieswere similarly represented in the up- and downregulatedgroups, not every regulated gene has a role in water-deficitstress tolerance and the change in expression in some ofthem may simply be the result of damages caused by stress(Bray 1997; Chaves et al. 2003).
The detailed list of the pathways affected under water-deficit stress condition is provided as Electronic supplemen-tary Tables S7, S8, and S9. A large range of osmolytes havebeen implicated in preventing damage to proteins caused bywater-deficit stress. Among the seven upregulated genesencoding for enzymes involved in trehalose biosynthesis,three of them represented isomers of trehalose-6-phosphatesynthase (TPS; LOC_Os02g54820; LOC_Os08g34580;LOC_Os09g23350) and two for isomers of trehalosesynthase (LOC_Os01g53000; LOC_Os01g54560). Thegenes encoding for trehalose-phosphate phosphatase (TPP;LOC_Os10g40550) and trehalase (LOC_Os10g37660) werealso upregulated (Fig. 4; Electronic supplementary Fig. S3A).In an alternative trehalose biosynthesis pathway, where glyco-gen is converted to trehalose, gene encoding for isoamylase-type starch debranching enzyme (LOC_Os05g32710) wasupregulated by fourfold in 1-week-old seedling under water-deficit stress, however, hydrolase was downregulated in thepanicle (Zhou et al. 2007), suggesting that under water-deficitstress condition this alternative pathway might be cell-typespecific.
Sucrose synthesis utilizes glucose-1-phosphate, glucose-6-phosphate, fructose-6-phosphate, and sugar nucleotide UDP-D-glucose as hexose phosphate pool (Hoekstra et al. 2001).Under water-deficit stress, gene encoding for sucrose synthase1 (LOC_Os03g22120) was upregulated which mediated
164 Funct Integr Genomics (2011) 11:157–178

OS
MO
PR
OT
EC
TAN
TC
ELL
WA
LLC
OM
PO
NE
NT
S
HO
RM
ON
ES
LIP
ID M
ETA
BIL
OS
M
CA
RB
O-
HY
DR
ATE
SE
CO
ND
AR
YM
ETA
BO
LIT
E
PO
LYO
LS
AN
D S
UG
AR
S
Trehalose biosynthesis I
Fructan biosynthesis
Arginine biosynthesis II Arginine biosynthesis IV Proline biosynthesis I Proline biosynthesis V
Stachyose biosynthesis
myo-inositol biosynthesis
Sucrose biosynthesis
Trehalose biosynthesis V
AM
INO
AC
IDS
OLI
GO
SA
C-
CH
AR
IDE
QA
Cs
Sucrose degradation to ethanol and lactate
β-alanine betaine biosynthesisβ-alanine biosynthesis II
GDP-mannose metabolism
Salicylate biosynthesis
Flavonoid biosynthesis
Coumarin biosynthesis
Very long chain fatty acid biosynthesis
Cholesterol biosynthesisIIIIII
Flavonol biosynthesis
Phenylpropanoid biosynthesis
(initial reaction)
Phenylpropanoid biosynthesis
GDP-L-fucose biosynthesis
GDP-D-rhamnose biosynthesis
UDP-D-glucuronate biosynthesis
UDP-D-xylose biosynthesis
CMP-KDO biosynthesis II
Cytokinins 7-N-glucoside biosynthesis
Cytokinins 9-N-glucoside biosynthesis
Cytokinins-O-glucoside biosynthesis
Cytokinins degradation
Upregulated in any of the seven developmental stages of rice under water-deficit stress
Downregulated in any of the seven developmental stages
Strictly regulated with specificity to developmental stage i.e. upregulated as well as downregulated in any of the developmental stages
Fig. 4 Regulation of transcript and metabolite profiling during water-deficit stress condition. Selected pathways which were found to besignificantly influenced by water-deficit stress condition have been shown (Electronic supplementary Tables S7–S9)
Funct Integr Genomics (2011) 11:157–178 165

conversion of UDP-D-glucose to sucrose. However, regulationof other isoform of sucrose synthase 1 and 2 gene(LOC_Os03g28330; LOC_Os06g09450) was found to bestage specific. Moreover, genes encoding for sucrose-phosphate synthase (LOC_Os01g69030, LOC_Os01g27880),which mediates synthesis of sucrose utilizing fructose-6-phosphate, were downregulated. This would cause increasein the hexose phosphate pool (Fig. 4; Electronic supplementaryFig. S3B). Transcript level of gene encoding for ß-fructofur-anosidase (LOC_Os02g01590), involved in biosynthesis offructan, a polysaccharide functioning as storage carbohydrate,was found to have strict regulation depending on develop-mental stage and organ, i.e. sixfold upregulated in 1-week-oldseedling, whereas it was downregulated in panicle and flagleaf (Fig. 4; Electronic supplementary Fig. S3B). Transcriptlevel of stachyose synthase enzyme encoding gene(LOC_Os01g07530) involved in biosynthesis of anotheroligosaccharide, stachyose, which is used by plants as storagematerial and known to act as protective agent during seedmaturation and cold stress (Bentsink et al. 2000; Gilmour etal. 2000), was found to be upregulated by 11-fold in 1-week-old seedling under water-deficit stress condition (Fig. 4;Electronic supplementary Fig. S3C). In mannitol biosynthesispathway, isomerization of fructose-6-phosphate to mannose-6-phosphate is mediated by mannose-6-phosphate isomerase(LOC_Os01g03710), whose transcript level increased byeightfold in 1-week-old seedling under water-deficit stresscondition (Fig. 4; Electronic supplementary Fig. S4A)establishing a positive correlation between accumulation ofmannitol and water-deficit stress.
Genes encoding for enzymes involved in biosynthesis ofprimary cell wall components were upregulated under water-deficit stress condition. Synthesis of GDP-D-rhamnose andGDP-L-fucose were favored under stress condition. Moreover,transcript level of hexokinase gene (LOC_Os06g45980) thatmediates conversion of GDP-D-mannose to mannose-6-phosphate in GDP-mannose metabolism was sixfold down-regulated in 1-week-old seedling, which in turn could facilitatechanneling of more GDP-D-mannose into synthesis of GDP-D-rhamnose and GDP-L-fucose (Fig. 4; Electronic supplemen-tary Fig. S4A). UDP-galacturonate and UDP-xylose are twoother cell wall components derived from UDP-glucuronate,which is synthesized via inositol oxidation pathway (Fig. 4;Electronic supplementary Fig. S4B). Increased accumulationof transcript of myo-inositol oxygenase encoding gene(LOC_Os06g3656) occurred under stress condition whichwould convert myo-inositol to glucuronate.Moreover, inositol-3-phosphate synthase gene (LOC_Os03g09250) involved inmyo-inositol biosynthesis pathway showed six fold increase inits transcript levels (Fig. 4; Electronic supplementary Fig. S4B).Interestingly, it was also observed that the synthesis of myo-inositol from α-D-glucose-6-phosphate was favored oversynthesis of UDP-D-glucose under stress condition. 3-deoxy-
D-manno-octulosonate (KDO) is a component of rhamnoga-lacturonanII pectin fraction of the primary cell wall. The genecoding for 3-deoxy-manno-octulosonate-cytidylyltransferase(CKS; LOC_Os05g48750), which activates KDO by couplingit to CMP (cytidine monophosphote), was found to beupregulated under stress condition (Fig. 4; Electronic supple-mentary Fig. S4C).
Genes encoding for enzymes involved in amino acidmetabolism pathways were found to be differentially regulat-ed under water-deficit stress condition (Electronic supple-mentary Tables S7, S8, and S9). Genes involved inbiosynthesis of ß-alanine, histidine, and serine were prefer-entially upregulated under water-deficit stress. Inproline biosynthesis, genes encoding for enzymes δ-1-pyrroline-5-carboxylate synthase (LOC_Os05g38150;LOC_Os01g62900), oxidoreductase (LOC_Os01g12710),aldehyde dehydrogenase (LOC_Os09g26880), and NADP-dependent glyceraldehydes-3-phosphate dehydrogenase(LOC_Os08g34210) were upregulated under stress condition(Fig. 4; Electronic supplementary Fig. S5). Transcriptaccumulation of six enzymes (LOC_Os11g39220;LOC_Os06g23870; LOC_Os06g24704; LOC_Os05g03480;LOC_Os05g07090; LOC_Os05g46480) involved in synthe-sis of β-alanine from propionate was also found to beincreased (Fig. 4; Electronic supplementary Fig. S6). Thepathways related to biosynthesis of cholesterol, very longchain fatty acid, flavonol and isoflavonol derivatives werealso stimulated under water-deficit stress condition (Fig. 4).
Analysis of commonly regulated genes by extrinsic(environmental) and intrinsic (developmental) water-deficitconditions
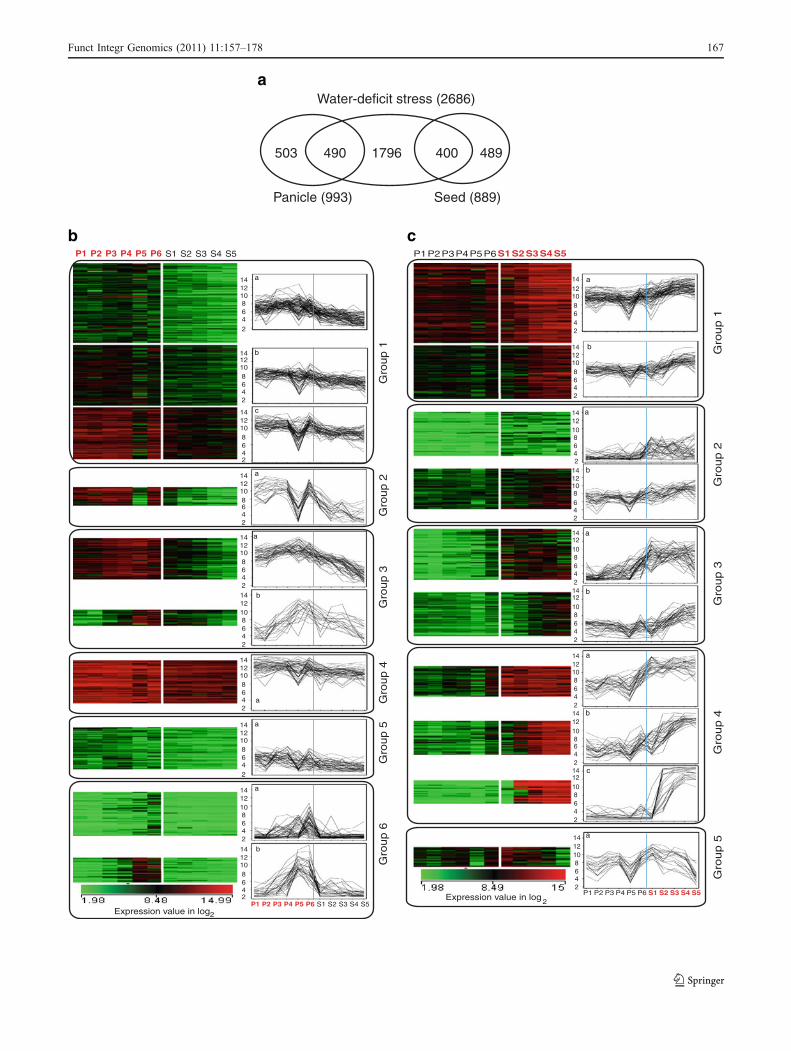
The reproductive developmental stages of rice were categorizedinto six stages of panicle development followed by five stagesof seed development, which have been described in detail inmaterials and methods section. Among the 2,686 water-deficitstress-induced genes, 490 genes were found to be inducedduring panicle development (Electronic supplementary TableS10) and 400 genes (Electronic supplementary Table S11)showed upregulation during seed development (Fig. 5a).
Fig. 5 Correlation between extrinsic and intrinsic water-deficit stressresponse during six stages of panicle (P1–P6) and five stages of seeddevelopment (S1–S5). a Venn diagram showing relationship amongwater-deficit stress-responsive genes having panicle and seed devel-opmental stage-specific regulation. b K-Means clustering of 490panicle preferential genes under water-deficit stress condition. Clusterswith similar trend of expression during panicle and seed developmenthave been grouped together into six groups. c K-Means clustering of400 seed preferential genes under water-deficit stress condition.Clusters with similar trend of expression during panicle and seeddevelopment have been grouped together into five groups. Color scalefor average log ratio values is given in log2
b
166 Funct Integr Genomics (2011) 11:157–178
1412108642
a
1412108642
a
1412108642
b
1412108642
a
1412108642
a
P1 P2 P3 P4 P5 P6 S1 S2 S3 S4 S5
Gro
up
1G
rou
p 2
Gro
up
3G
rou
p 4
Gro
up
5G
rou
p 6
Expression value in log2
P1P2P3P4P5P6S1S2S3S4S5
Gro
up
1G
rou
p 2
Gro
up
3G
rou
p 4
Gro
up
5
Expression value in log 2
Seed (889)Panicle (993)
Water-deficit stress (2686)
1796503 490 489400
141210864
2
a 1412108642
a
1412108642
b1412108642
b
1412108642
c
1412108642
a
1412108642
b
P1 P2 P3 P4 P5 P6 S1 S2 S3 S4 S5
S4 S5P5 P6 S1 S2 S3P1 P2 P3 P4
1412108642
a
b14121086421412108642
c
1412108642
a
1412108642
a
1412108642
a
1412108642
b
1412108642
b
a
b c
Funct Integr Genomics (2011) 11:157–178 167

Overlap of gene expression between water-deficit stresscondition and panicle development
Zhou et al. (2007) identified 408 genes to be inducible inpanicle under water-deficit stress condition. Ninety of thesegenes were also found to be differentially regulated duringnormal course of panicle development, indicating towards anunderlying connection in panicle development and water-deficit stress response. In this study, 490 water-deficit stress-responsive genes with preferential expression in panicle werecategorized into six major groups using K-means clustering(Group 1–6; Fig. 5b). Genes in group 3a (48) showed hightranscript accumulation uniformly from P1 to P6 stages ofpanicle development with decrease in expression duringseed maturation. Among these, only seven genes(LOC_Os01g54080, LOC_Os04g47520, LOC_Os07g01600,LOC_Os07g37850, LOC_Os09g31031, LOC_Os08g35710,and LOC_Os06g06780) showed upregulation in panicle tissueunder water-deficit stress (Zhou et al. 2007), whereas, othercluster members showed transcript accumulation at variousvegetative developmental stages under water-deficit stress.The P5 and P6 stages of panicle development includematuration of stamen, carpel, pollen, and anther dehiscence,which involves natural dehydration process. Two clusters (6aand 6b) of group 6 include genes expressing in the later stages(P5 and P6) of panicle development. Interestingly, a largefraction of (41.38% and 31.37%) genes belonging to cluster6a and b, respectively, showed upregulation in panicle underwater-deficit stress (Zhou et al. 2007) and the remaininggenes were responsive to external water-deficit stress atvarious vegetative stages. Taken together, among the genesshowing specific upregulation during various stages ofpanicle development, 490 genes showed correlation withthe stress upregulated genes and these may participate innatural desiccation process related to panicle development.
Overlap of gene expression between water-deficit stressand seed development
The ten K-means derived clusters, representing 400 seedpreferential genes upregulated under water-deficit stress, couldbe grouped in five distinct groups (Fig. 5c). Group 4 genesshowed higher level of transcript accumulation in seedcompared to panicle development. Group 4c genesmostly encoding for LEA proteins (LOC_Os06g23350;LOC_Os01g12580), dehydrin (LOC_Os11g26750), em-bryonic protein (LOC_Os05g28210; LOC_Os04g52110;LOC_Os11g26570), aquaporin (LOC_Os10g35050),phosphatase (LOC_Os06g04790), l ipid- t ransferprotein (LOC_Os03g02050), calcium-sensory protein(LOC_Os10g09850), and NAC transcription factor(LOC_Os03g21060) were sharply upregulated after S1 stageof seed development. By 5th day after anthesis (S3), the
embryo sac is filled with endosperm cells and its gradualmaturation continues till S5 stage of seed development. Genesbelonging to cluster (4b) were found to be encoding for proteinsmostly involved in synthesis of storage proteins such asglobulin (LOC_Os05g41970), glutelin (LOC_Os02g16830;LOC_Os02g15169; LOC_Os02g25640), prolamine(LOC_Os11g33000; LOC_Os06g31070) along with oleosin(LOC_Os03g49190), and patatin (LOC_Os01g67310), as wellas aquaporin (LOC_Os04g44570), NAC transcription factor(LOC_Os02g12310) and LEA protein (LOC_Os12g43140).Endoplasmic reticulum (ER) stress-related genes were foundclustered in group 4a, namely heat shock 70 kDa protein 1(LOC_ Os03g11910), heat shock 22 kDa protein(LOC_Os02g52150), 17.4 kDa class I heat shock protein 2(LOC_Os03g16020), heat shock cognate 70 kDa protein(LOC_Os05g38530), and heat shock protein 101(LOC_Os05g44340), which started accumulating from P6stage of panicle development till last stage of seed maturation.In group 5a cluster, transcript accumulation is maximum in S1–S2 stages and then decreases with seed maturity, which is adiagonally opposite expression profile to the group 4 genes.MYB (LOC_Os05g37060) and C2H2 (LOC_Os03g60560)transcription factors, ABC-transporter (LOC_Os01g03144),ATPase-3 (LOC_Os03g58790), GA inactivating enzyme(gibberellin 2-beta-dioxygenase; LOC_Os01g55240)and defense related proteins, acidic endochitinase(LOC_Os01g47070) and immediate-early fungal elicitorprotein CMPG1 (LOC_Os02g50460) encoding genes arerepresented in group 5 cluster. Therefore, it is apparent fromthis analysis that the genes known traditionally to be involvedin reproductive development could also be potential players inabiotic stress response.
Overlap in metabolic pathways between water-deficit stressand reproductive development
Metabolic pathways regulated under water-deficit stress andvarious stages of panicle/seed development are given inElectronic supplementary Tables S12 and S13. Genesencoding for proteins involved in secondary metabolitebiosynthesis pathways namely, flavonol, maackiain, medi-carpin, phenylpropanoid, salicylate, divinyl ether, coumarinbiosynthesis were upregulated during panicle developmentand under water-deficit stress. However, stachyose synthaseenzyme encoding gene (LOC_Os01g07530), genes encodingfour enzymes (metabolite transport protein-csbC, ADP-glucose pyrophosphorylase large subunit 3, starch synthaseand 1,4-α-glucan branching enzyme) involved in starchbiosynthesis pathway and transketolase (LOC_Os04g19740)enzyme involved in pentose phosphate pathway, glucosefermentation to lactate II, Calvin cycle and xylulose-monophosphate cycle, were preferentially upregulated dur-ing seed development as well as under water-deficit stress.
168 Funct Integr Genomics (2011) 11:157–178

Discussion
Our study primarily focused on the identification andanalysis of water-deficit stress-responsive genes from ricecultivars at different developmental stages including sixvegetative (1-, 2-, 4-, 6-week, 1-week-before-heading, and4-tiller stage) and one reproductive (1-week-before-head-ing) stages with the aim to identify important componentsof water-deficit stress response. Data across laboratorieswas used for this analyses as it were thought to enrich thelist of water-deficit stress-responsive genes under variedexperimental conditions (Bray 2004). While preparing themanuscript, another substantial study on the effect of long-term drought stress on rice cultivars was published byDegenkolbe et al. (2009); most of the differentiallyexpressed genes identified in this study were found to berepresented in the comprehensive non-redundant gene listprepared in this study.
Transcription factors regulating water-deficit stressresponse
Previous studies have shown that transcription factors areimportant regulators involved in plant response to environmen-tal stress (Chinnusamy et al. 2004; Mahajan and Tuteja 2005;Yamaguchi-Shinozaki and Shinozaki 2006; Shinozaki andYamaguchi-Shinozaki 2007; Nakashima et al. 2009). Both,ABA-independent and ABA-dependent regulatory pathwaysexist for water-deficit stress-responsive genes in plants. It wasobserved that under water-deficit stress condition, members ofTF families involved in both ABA-independent (AP2/ERF,bHLH and NAC) and ABA-dependent (MYB, bZIP, bHLH(MYC), NAC and homeodomain) pathways are upregulatedin rice. However, members of a gene family are differentiallyup or downregulated indicating that TFs work individually inresponse to stress. TFs belonging to these families have beenknown to interact with specific cis-elements and/or proteinsand their overexpression conferred stress tolerance in heterol-ogous systems (Abe et al. 1997, 2003; Jaglo-Ottosen et al.1998; Liu et al. 1998; Kasuga et al. 1999; Kang et al. 2002;Fujita et al. 2004, 2005; Tran et al. 2004; Furihata et al. 2006;Hu et al. 2006; Ito et al. 2006; Sakuma et al. 2006; Chen et al.2007; Dai et al. 2007; Jung et al. 2008; Zhou et al. 2009).Expression of TFs in an organ-specific manner was noted byZhou et al. (2007). In our study also, the regulation of TFswas found to be very precise in terms of spatial and temporaldistribution, as detailed in Fig. 2 and Electronic supplemen-tary Fig. S1.
Certain TF gene families (NAC, zinc-finger) have beenshown to play important role during stress (Ciftci-Yilmazand Mittler 2008; Nakashima et al. 2009). Involvement ofNAC TF in biotic and abiotic stress response is well known(Olsen et al. 2005; Hu et al. 2006; Nakashima et al. 2009;
Seo and Park 2010). Transgenic rice overexpressingOsNAC10, OsNAC6, and ONAC045 showed improveddrought and salt tolerance (Zheng et al. 2004; Nakashimaet al. 2009; Jeong et al. 2010). SNAC1 gene was found tobe involved in stomatal closure leading to drought tolerance(Hu et al. 2006). Fang et al. (2008) reported 20 rice NACTF genes to be drought-inducible, however, our studyrevealed 22 NAC TF genes to be upregulated under water-deficit stress (Electronic supplementary Table S2). Five ofthe NAC TF genes shared upregulation under water-deficit,cold and salt stress condition. Moreover, two of these NACTF genes (LOC_Os11g03370; LOC_Os11g03300) showedpanicle preferential regulation; however, they were notinducible by water-deficit stress condition in panicle tissue(Zhou et al. 2007). Six other water-deficit stress-inducibleNAC TF genes (LOC_Os02g36880; LOC_Os07g37920;LOC_Os06g46270; LOC_Os03g21060; LOC_Os02g56600;LOC_Os02g12310) showed seed preferential expression.Involvement of NAC TF in development as well as in stresstolerance has been reported in earlier studies (Sablowski andMeyerowitz 1998; Guo and Gan 2006; Peng et al. 2009).Arabidopsis NAC TF gene ANAC092 demonstrated anintricate overlap of ANAC092-mediated gene regulatorynetworks during salt-promoted senescence and seed matura-tion (Balazadeh et al. 2010). Hence, these studies highlightinterplay of pathways regulated by NAC TFs during stressand developmental stages.
Out of zinc-finger motif containing TFs, C2H2, C2C2-Dof, C3H, PHD, WRKY and ZIM are particularly repre-sented in water-deficit stress-responsive transcriptome.Genes belonging to these categories have been shown toconfer stress tolerance on overexpression in transgenicsystems (Chen et al. 1996; Bowman 2000; Eliasson et al.2000; Rao et al. 2000; Rizhsky et al. 2002; Kim et al. 2004;Narusaka et al. 2004; Reyes et al. 2004; Rizhsky et al.2004; Sakamoto et al. 2004; Yanagisawa 2004; Davletovaet al. 2005; Zhang and Wang 2005; Jiang and Deyholos2006; Major and Constabel 2006; Agarwal et al. 2007;Huang et al. 2007; Park et al. 2007; Sun et al. 2007; VanHolme et al. 2007; Ciftci-Yilmaz and Mittler 2008).
Members of ten TF families were only induced (none of themembers were repressed) under water-deficit stress implyingthat they might play special role in stress tolerance. Amongthese families, members of SRS, CPP, EIL, Tubby and trihelixfamily have been earlier reported to be induced in rice roottissue under osmotic stress condition (Ma et al. 2009). Amongthe other TF families, PBF-2-like (whirly) proteins are mostlyknown to play role in defense response and could alsofunction in the chloroplast as well as the nucleus (Desveaux etal. 2005). Members of the remaining four TF families(jumonji, MBF1, ULT, and GeBP), which are conventionallynot known to be stress-responsive and are primarily involvedin developmental processes and phytohormone responses
Funct Integr Genomics (2011) 11:157–178 169

(Curaba et al. 2003; Noh et al. 2004; Tsuda et al. 2004; Carleset al. 2005; Chevalier et al. 2008; Yu et al. 2008) wereupregulated under water-deficit stress condition. The preciserole of these gene products in water-deficit stress responserequires more detailed investigations.
Modulation of metabolic pathways under water-deficitstress condition
Osmoprotectants
One of the most important class of molecules known toprotect plant cells from dehydration represents osmopro-tectants (Ramanjulu and Bartels 2002). Osmoprotectantsfall in several groups―polyols and sugars (mannitol,trehalose, sucrose, and fructan), amino acids (e.g., proline)and quaternary ammonium compounds (glycine betaine).Trehalose functions in the stabilization of biologicalstructures under abiotic stress in bacteria, fungi andinvertebrates (Ramanjulu and Bartels 2002). We observedthat biosynthesis of trehalose is favored under water-deficitstress condition by more accumulation of transcripts ofseven genes coding for trehalase, TPS, trehalose synthaseand TPP enzymes (trehalase: LOC_Os10g37660;TPS: LOC_Os02g54820, LOC_Os08g34580 andLOC_Os09g23350; trehalose synthase: LOC_Os01g53000and LOC_Os01g54560; TPP: LOC_Os10g40550). Abioticstress tolerance was successfully achieved in rice byoverexpression of Escherichia coli trehalose biosyntheticgenes, otsA and otsB, as a fusion gene (encoding for TPSand TPP, respectively; Garg et al. 2002). TPS1 is alsorequired for normal vegetative development and floraltransition in Arabidopsis (Ramon and Rolland 2007). Thus,trehalose, along with its protective role in stabilizingproteins, might also be helping plants in sustaining normalvegetative and reproductive growth by maintaining normalcell division, cellular differentiation and associated tran-scriptional changes under water-deficit stress condition.The alternative pathway of trehalose showed tissue-specificregulation. This kind of cell type-specific regulation hasbeen reported earlier, however, a possible physiological rolefor such tissue-specific accumulation is unclear (Leyman etal. 2001). However, the alternative trehalose pathway is notyet characterized in rice, but it is well characterized inRhizobium sp. M-11 (Iturriaga et al. 2009). The genesrelated to biosynthesis of sucrose, fructan and mannitol arealso stimulated under water-deficit stress; for example,sucrose synthase (SUS) gets upregulated. SUS expressionwas reported earlier to be induced by cold, dehydration, andosmotic stress (Hesse and Willmitzer 1996; Dejardin et al.1999; Kleines et al. 1999). The transcript level of AtSUS3from Arabidopsis was found to increase under droughtstress and mannitol treatment, as well as during seed
maturation (Baud et al. 2004). Enhanced sucrose biosyn-thesis under temperature shock and cold acclimation inArabidopsis has been reported (Kaplan et al. 2004, 2007).Resurrection plants have been shown to accumulate sucroseand trehalose when dehydrating (Whittaker et al. 2001;Moore et al. 2007). Recently, it has also been found thatglucose and sucrose accumulate in specific locations inresurrection plant tissue during dehydration from desicca-tion (Martinelli 2008). Fructans are known to prevent lipidcondensation during the phase transition and are believed toprotect biological membranes under stress (Hincha et al.2002; Vereyken et al. 2003). Sugars, which play versatilerole in plant development, could also trigger an oxidativeburst in tissues under abiotic stress conditions (reviewed byVan den Ende and Valluru 2009). Under osmotic stress,accumulation of proline helps in stabilizing proteins,membranes and subcellular structures; it also protectscellular metabolism by scavenging reactive oxygen species(Ramanjulu and Bartels 2002). One of the genes upregu-lated during water-deficit stress was δ-1-pyrroline-5-car-boxylate synthetase. It has been reported earlier thatPetunia plants expressing δ-1-pyrroline-5-carboxylate syn-thetase genes (AtP5CS from Arabidopsis or OsP5CS fromrice) accumulated proline and the transgenic plants couldtolerate 14 days of drought stress (Yamada et al. 2005).
Synthesis of non-protein amino acid β-alanine may beenhanced, as seven genes encoding for enzymes involved inbiosynthesis of β-alanine from propionate were upregulatedunder water-def ic i t s t ress (LOC_Os02g17390,LOC_Os05g46480, LOC_ Os05g07090, LOC_Os05g03480,LOC_Os06g24704, LOC_Os06g23870, LOC_Os11g 39220;Electronic supplementary Fig. S6). β-alanine in turn isconverted to β-alanine betaine, which acts as an osmopro-tectant in most members of the highly stress tolerant plantfamily Plumbaginaceae (Rathinasabapathi et al. 2001), medi-ated by N-methyltransferase. Two isomeric genes encodingfor N-methyltransferase were up (LOC_Os06g06560;LOC_Os07g42280) and downregulated (LOC_Os07g49300;LOC_Os09g29710), respectively, under water-deficit stresscondition.
Cell wall components
A number of transcripts encoding for enzymes involved insynthesis of primary cell wall component and enzymesresponsible for cell wall loosening like xyloglucan endo-transglycosylase (XET: LOC_Os08g13920) and six expansins(LOC_Os02g16730, LOC_Os10g39110, LOC_Os02g44108,LOC_Os10g39640, LOC_Os10g40710, LOC_Os06g50400)were upregulated under water-deficit stress condition. It hasbeen shown in earlier studies that cell wall plays crucial role incell enlargement, which is indispensable part of plant growthand development (Cosgrove 2001). Moreover, it has been
170 Funct Integr Genomics (2011) 11:157–178

found that the tensile property of cell wall helps inameliorating the shearing force generated during droughtstress in grasses (Balsamo et al. 2006). Hence, loosening ofcell wall and synthesis of structural constituents together couldhelp in coping with the water-deficit stress.
Lipid metabolism
The gene coding for squalene synthase (LOC_Os03g59040),involved in cholesterol biosynthesis, was found to be upregu-lated in the present study. In parallel to this observation,squalene synthase ESTwere found to be overrepresented undera variety of abiotic stresses (Houde et al. 2006). Recent studieshave shown that sterols are required for secretory vesicletargeting which might be facilitating stress adaptation process(Carter et al. 2004). Plant sterols have been shown to cyclebetween plasma membrane and endosomes in an actin-dependent manner (Grebe et al. 2003). Abiotic stress causessignificant intracellular restructuring in plants which leads totransportation, removal and compartmentalization of certainmolecule via vesicle trafficking. Very long chain fatty acidbiosynthesis-related gene (long-chain-3-hydroxyacyl-CoA de-hydrogenase; LOC_Os02g17390) was also upregulated underwater-deficit stress. VLCAFs are known to mainly function asprecursors for components of the cuticle, such as cutin andepicuticular waxes (Post-Beittenmiller 1996), which areknown to prevent water loss at leaf surface.
Secondary metabolite
Accumulation of anthocyanin pigments in vegetative tissueis hallmark of plant stress (Winkel-Shirley 2002). Increasein transcript accumulation of phenylalanine ammonia-lyase(PAL; LOC_Os02g41670) gene was observed, whichmediates conversion of L-phenylalanine to trans-cinnamate,favoring salicylate, flavonoid, coumarin and phenylpropa-noid biosynthesis (Fig. 4; Electronic supplementary Fig.S7). Induction of PAL genes under salt, drought, cold stress(Guo and Wang 2009), wounding, and fungal elicitortreatment (Zhu et al. 1995) have been observed in variousplant species. Flavonol synthase (FLS; LOC_Os02g52840),a key enzyme in flavonoid biosynthetic pathway, wasupregulated in the present study. Flavonoids are known tohave role in providing UV protection (Bharti and Khurana1997; Ryan et al. 2001, 2002), conferring resistance toaluminium toxicity in maize (Kidd et al. 2001) and haveantimicrobial as well as antifungal properties (Dixon andSteele 1999). Isoforms of FLS gene in Arabidopsis showtissue-specific expression as well as differential expressionin response to different environmental conditions (Owens etal. 2008). Recently, a R2R3-MYB transcription factor,MYB12, from Arabidopsis, has been found to be thetranscriptional regulator of flavonol synthase (Mehrtens et
al. 2005). AtMYB60, another member of R2R3-MYBfamily has been assigned the role of transcriptionalmodulator of physiological responses in guard cells whichcould help plants survive desiccation (Cominelli et al.2005). Interestingly, the overexpression of a rice R2R3-MYB transcription factor, OsMYB3R-2, in Arabidopsisconferred tolerance to cold, drought and salt stress (Dai etal. 2007). It has also been reported that overexpression ofCpMYB10 from resurrection plant, Craterostigma planta-gineum, in Arabidopsis led to desiccation and salt toleranceof transgenic lines by altering ABA and Glc signaling(Villalobos et al. 2004). Thus, among the 25 MYB TFsupregulated under water-deficit stress condition in ourstudy, some could be regulating secondary metabolitebiosynthesis helpful in stress response.
Hormone metabolism
Genes coding for enzymes involved in cytokinin biosyn-thesis showed increased transcript accumulation underwater-deficit stress condition (Fig. 4). However, cytokinindegradation-related genes were also found to be upregu-lated under water-deficit stress. Cytokinin is essential fornormal functioning of plants as well as mediating stressresponse by stimulating accumulation of anthocyanin,proline and ethylene as reviewed by Hare et al. (1997).Studies showed that overexpression of isopentenyltransfer-ase (IPT) gene, involved in cytokinin synthesis, on theonset of senescence resulted in suppression of drought-induced leaf senescence leading to drought tolerance oftransgenic tobacco plants (Rivero et al. 2007), however, ithas also been found that overexpression of IPT gene intobacco under light-inducible promoter resulted in elevatedaccumulation of cytokinin level which induced wiltingsymptom as observed during salinity stress (Thomas et al.1995). In Arabidopsis, cytokinin receptor histidine kinases,AHK2, AHK3, and CRE1, have been found to act asnegative regulators in stress responses in a cytokinin-dependent manner (Tran et al. 2007). Thus, it is apparentthat cytokinin mediated regulation might be dependent onthe nature of target cells and precise developmental stages.
Resistant variety (upland rice), on exposure to stress, hasalso been found to trigger biosynthesis of osmoprotectants,cell wall strengthening components, oxidation protectionmolecules, secondary metabolite, and ion transport compo-nents to ensure normal cellular functioning under stresscondition (Chao et al. 2005; Walia et al. 2005; Wang et al.2007). We noted in metabolic pathway analysis that genesinvolved in proline biosynthesis (LOC_Os01g12710;LOC_Os09g26880), sucrose (LOC_Os03g22120), and cellwall component (LOC_Os06g36560; LOC_Os07g04690)were upregulated in upland rice variety (Wang et al. 2007).This observation strengthens the fact that along with such
Funct Integr Genomics (2011) 11:157–178 171

genes, other upregulated genes involved in these pathways might be responsible for conferring stress tolerance.Our study also revealed upregulation of metallothionein(LOC_Os12g38051), methionine sulfoxide reductase(LOC_Os03g24600) , respi ra tory burs t oxidase(LOC_Os11g33120; LOC_Os05g45210), calcium transportingATPase (LOC_Os05g02940; LOC_Os04g51610) and ninecalmodulin binding proteins (Electronic supplementary TableS1). Genes belonging to same groups were upregulated in rootof upland rice variety (Prata Ligeiro) and not in lowlandvariety (IRAT20) after drought stress at anthesis stage(Rabello et al. 2008).
Physiological parameters like root development andstomatal movement have utmost importance in water-deficit stress tolerance. Development of root in bothlowland and upland variety of rice is intricately related todrought tolerance (Fukai and Cooper 1995). Two genes(LOC_Os12g01550 (upregulated in 1-week-old seedling)and LOC_Os03g45750 (downregulated in panicle)) codingfor LOB domain proteins, which are essential for adventi-tious root formation in rice (Liu et al. 2005), weredifferentially regulated under water-deficit stress condition.Involvement of the vacuolar Ca2+-activated channel TPC1and protein phosphatase 2C, are already known in stomatalmovement (Peiter et al. 2005; Pandey et al. 2007). Ourstudy identified more genes coding for homologues of calciumchannel protein TPC1 (LOC_Os01g48680), protein phospha-tase 2C ABI1 (LOC_Os05g49730; LOC_Os01g46760), pro-tein phosphatase 2C ABI2 (LOC_Os05g46040;LOC_Os05g51510; LOC_Os01g40094), and 11 more phos-phatase 2C to be upregulated under water-deficit stress(Electronic supplementary Table S1). These genes,which could be essential for root development andstomatal movement, are still not assigned any pathway,hence, further study is needed for their functionalcharacterization.
Regulation of genes in relation to oxidativeand water-deficit stress
On comparing our 5,611 differentially regulated genes with1,062 oxidative stress-responsive genes from rice, we foundthat 4% of water-deficit stress-responsive genes are alsoresponsive to oxidative stress (Liu et al. 2010). However, weadd more genes coding for enzymes with antioxidant proper-ties, namely, ascorbate peroxidase (LOC_Os04g14680), andsuperoxide dismutase (LOC_Os07g46990). Developmentalstage-specific expression of antioxidant genes under water-deficit stress has also been observed (Electronic supplementaryTables S2 and S3). Jain et al. (2010) reported sevenglutathione-S-transferases (GST) genes to be upregulatedunder water-deficit stress in 1-week old seedling. Weidentified another isoform of GST gene (LOC_Os01g70770)
to be upregulated only in shoot tissue under water-deficit stress,emphasizing on tissue-specific regulation of oxidative stress-related gene expression. Hence, we suggest that specificpathways are operational under water-deficit stress to counter-act oxidative stress in a developmental stage-specific manner.
Downregulation of genes
In higher plants, foliar photosynthetic rate is known todecrease with low relative water content (Lawlor 2002).Stomatal limitation, decrease in ATP content and CO2
concentration, limited metabolic processes, and loss ofrubisco are considered to be the determinant of reducedphotosynthesis under drought condition (Cornic et al. 2000;Lawlor 2002; Parry et al. 2002; Vu and Allen 2009).Present study also showed downregulation of transcriptsrelated to photosynthesis (i.e., photosynthesis-antennaprotein, porphyrin, and chlorophyll metabolism; Electronicsupplementary Table S14) under water-deficit stress. More-over, genes encoding for proteins related to normal geneticfunctioning like, components of RNA polymerase,aminoacyl-tRNA biosynthesis, DNA replication and repairwere also found widely downregulated under stress. Similarobservation was made by Seki et al. (2002b), where genesrelated to photosynthesis were downregulated underdrought stress. They also reported that transcripts involvedin DNA damage repair showed decrease in accumulationunder drought. The downregulation of these genes could infact mean switching between the productivity and thesustenance mode.
Members of TF families, zf_DHHC (four members),SNF2 (three members), SET (two members), and sigma70(two members) were downregulated under water-deficitstress but none were upregulated (Electronic supplementaryTable S15). SET, SNF2 and zf_DHHC TFs are also knownto be involved in development of reproductive organs(Farrona et al. 2004; Thorstensen et al. 2008; Verdier et al.2008). Moreover, six of the 17 members of CO-like TFfamily were found to be downregulated under water-deficitstress (Electronic supplementary Table S3), however, threeother members (LOC_Os06g19444; LOC_Os02g49230;LOC_Os08g15050) were found to have tissue-specific up/downregulation, indicating their involvement in tissuedependent stress response in rice (Electronic supplementaryFig. S1). CONSTANS (CO) gene is known to be involved inthe photoperiodic regulation of flowering (Imaizumi andKay 2006; Kobayashi and Weigel 2007; Kim et al. 2008).However, in earlier studies interplay between stress- andcircadian-regulated gene expression have also been shown(Kreps et al. 2002; Hannah et al. 2005; Achard et al. 2007).Hence, we suggest that the signaling cascade or pathways,which are downregulated under stress, could be targeted byregulated overexpression of these downregulated genes,
172 Funct Integr Genomics (2011) 11:157–178

which might help in sustaining normal functioning understress.
Correlation between water-deficit, cold, and salt stress
Among the genes which are responsive to water-deficit,cold, and salt stress, more overlap was found between saltand water-deficit stress-responsive genes, whereas, water-deficit and cold stress showed least number of commonlyregulated genes. Similar trend of regulation was also notedby Rabbani et al. (2003). Drought and salt stress ultimatelyresult in dehydration of the cell causing osmotic imbalance,which leads to de-regulation of almost every aspect ofcellular physiology and metabolism. Therefore, prominentcross-talk of components regulated by these two stressconditions is not surprising. Even cold stress induces severemembrane damage due to dehydration associated withfreezing, which might be responsible for regulation ofcommon genes under these two stress conditions. The geneexpression data indicate that the major abiotic stresses,water-deficit, low temperature, and salinity, are complexstimuli, because they possess common, yet differentattributes, resulting in signaling cascades that are uniqueto each stress condition and/or have complex networking(Xiong and Zhu 2002; Chinnusamy et al. 2004;Yamaguchi-Shinozaki and Shinozaki 2006; Nakashima etal. 2009).
Genetic overlap in development and water-deficit stress
Complex genetic network functions during development oforgan in plants and it has been found that substantialoverlaps exist between the developmental pathways and thestress-response pathways (Cooper et al. 2003). Some of thegenes are expressed under stress as well as in embryoniccells or during seed desiccation (Sivamani et al. 2000;Medina et al. 2001). Lan et al. (2005) showed that a largefraction of genes regulated by dehydration is also upregu-lated by pollination/fertilization. Similar conclusions weredrawn from analyses of promoter-GUS fusions of cold-inducible RD29A, COR15A, KIN1, and COR6.6 genes inArabidopsis which were regulated during plant develop-ment under both stressed (cold) and unstressed conditions(Yamaguchi-Shinozaki and Shinozaki 1993; Baker et al.1994; Wang and Cutler 1995).
In cereals, it has been seen that water-deficit stressduring flower induction and inflorescence developmentleads to a delay or complete inhibition in flowering (Winkelet al. 1997). Interestingly, TFs belonging to AP2, MYB andPBF2-like (whirly) family were found to be involved inpanicle development as well as water-deficit stress re-sponse, implying that they may represent the cross-talkcomponents between development and stress.
Acknowledgement This work is supported by the Department ofBiotechnology, Government of India, the Council of Scientific andIndustrial Research (research fellowship to P.D.), and the UniversityGrants Commission (research fellowship to P.K.D.).
References
Abe H, Yamaguchi-Shinozaki K, Urao T, Iwasaki T, Hosokawa D,Shinozaki K (1997) Role of Arabidopsis MYC and MYBhomologs in drought- and abscisic acid-regulated gene expres-sion. Plant Cell 9:1859–1868
Abe H, Urao T, Ito T, Seki M, Shinozaki K, Yamaguchi-Shinozaki K(2003) Arabidopsis AtMYC2 (bHLH) and AtMYB2 (MYB)function as transcriptional activators in abscisic acid signaling.Plant Cell 15:63–78
Achard P, Baghour M, Chapple A, Hedden P, Van Der Straeten D,Genschik P, Moritz T, Harberd NP (2007) The plant stresshormone ethylene controls floral transition via DELLA-dependent regulation of floral meristem-identity genes. Proc NatlAcad Sci USA 104:6484–6489
Agarwal P, Arora R, Ray S, Singh AK, Singh VP, Takatsuji H, KapoorS, Tyagi AK (2007) Genome-wide identification of C2H2 zinc-finger gene family in rice and their phylogeny and expressionanalysis. Plant Mol Biol 65:467–485
Andjelkovic V, Thompson R (2006) Changes in gene expression inmaize kernel in response to water and salt stress. Plant Cell Rep25:71–79
Arora R, Agarwal P, Ray S, Singh AK, Singh VP, Tyagi AK, KapoorS (2007) MADS-box gene family in rice: genome-wide identi-fication, organization and expression profiling during reproduc-tive development and stress. BMC Genomics 8:242
Babu PR, Sekhar AC, Ithal N, Markandeya G, Reddy AR (2002)Annotation and BAC/PAC localization of non-redundant ESTs fromdrought-stressed seedlings of an indica rice. J Genet 81:25–44
Baker SS, Wilhelm KS, Thomashow MF (1994) The 5′-region ofArabidopsis thaliana cor15a has cis-acting elements that confercold-, drought- and ABA-regulated gene expression. Plant MolBiol 24:701–713
Balazadeh S, Siddiqui H, Allu AD, Matallana-Ramirez LP, Caldana C,Mehrnia M, Zanor MI, Kohler B, Mueller-Roeber B (2010) Agene regulatory network controlled by the NAC transcriptionfactor ANAC092/AtNAC2/ORE1 during salt-promoted senes-cence. Plant J 62:250–264
Balsamo RA, Willigen CV, Bauer AM, Farrant J (2006) Droughttolerance of selected Eragrostis species correlates with leaftensile properties. Ann Bot (Lond) 97:985–991
Barr HD, Weatherley PE (1962) A re-examination of the relativeturgidity technique for estimating water deficit in leaves. Aust JBiol Sci 15:413–428
Baud S, Vaultier MN, Rochat C (2004) Structure and expressionprofile of the sucrose synthase multigene family in Arabidopsis. JExp Bot 55:397–409
Bentsink L, Alonso-Blanco C, Vreugdenhil D, Tesnier K, Groot SP,Koornneef M (2000) Genetic analysis of seed-soluble oligosac-charides in relation to seed storability of Arabidopsis. PlantPhysiol 124:1595–1604
Bharti AK, Khurana JP (1997) Mutants of Arabidopsis as tools tounderstand the regulation of phenylpropanoid pathway and UVBprotection mechanisms. Photochem Photobiol 65:765–776
Bowman JL (2000) The YABBY gene family and abaxial cell fate.Curr Opin Plant Biol 3:17–22
Bray E (1997) Plant responses to water deficit. Trends Plant Sci 2:48–54Bray EA (2004) Genes commonly regulated by water-deficit stress in
Arabidopsis thaliana. J Exp Bot 55:2331–2341
Funct Integr Genomics (2011) 11:157–178 173

Brosche M, Vinocur B, Alatalo ER, Lamminmaki A, Teichmann T,Ottow EA, Djilianov D, Afif D, Bogeat-Triboulot MB, AltmanA, Polle A, Dreyer E, Rudd S, Paulin L, Auvinen P, KangasjarviJ (2005) Gene expression and metabolite profiling of Populuseuphratica growing in the Negev desert. Genome Biol 6:R101
Buchanan CD, Lim S, Salzman RA, Kagiampakis I, Morishige DT,Weers BD, Klein RR, Pratt LH, Cordonnier-Pratt MM, Klein PE,Mullet JE (2005) Sorghum bicolor’s transcriptome response todehydration, high salinity and ABA. Plant Mol Biol 58:699–720
Carles CC, Choffnes-Inada D, Reville K, Lertpiriyapong K, FletcherJC (2005) ULTRAPETALA1 encodes a SAND domain putativetranscriptional regulator that controls shoot and floral meristemactivity in Arabidopsis. Development 132:897–911
Carter CJ, Bednarek SY, Raikhel NV (2004) Membrane trafficking inplants: new discoveries and approaches. Curr Opin Plant Biol7:701–707
Chao DY, Luo YH, Shi M, Luo D, Lin HX (2005) Salt-responsivegenes in rice revealed by cDNA microarray analysis. Cell Res15:796–810
Chaves MM, Maroco JP, Pereira JS (2003) Understanding plantresponses to drought-from genes to the whole plant. Funct PlantBiol 30:239–264
Chen W, Chao G, Singh KB (1996) The promoter of a H2O2-inducible, Arabidopsis glutathione S-transferase gene containsclosely linked OBF- and OBP1-binding sites. Plant J 10:955–966
Chen M, Wang QY, Cheng XG, Xu ZS, Li LC, Ye XG, Xia LQ, MaYZ (2007) GmDREB2, a soybean DRE-binding transcriptionfactor, conferred drought and high-salt tolerance in transgenicplants. Biochem Biophys Res Commun 353:299–305
Chevalier F, Perazza D, Laporte F, Le Henanff G, Hornitschek P,Bonneville JM, Herzog M, Vachon G (2008) GeBP and GeBP-like proteins are noncanonical leucine-zipper transcription factorsthat regulate cytokinin response in Arabidopsis. Plant Physiol146:1142–1154
Chinnusamy V, Schumaker K, Zhu JK (2004) Molecular geneticperspectives on cross-talk and specificity in abiotic stresssignalling in plants. J Exp Bot 55:225–236
Ciftci-Yilmaz S, Mittler R (2008) The zinc finger network of plants.Cell Mol Life Sci 65:1150–1160
Cominelli E, Galbiati M, Vavasseur A, Conti L, Sala T, Vuylsteke M,Leonhardt N, Dellaporta SL, Tonelli C (2005) A guard-cell-specific MYB transcription factor regulates stomatal movementsand plant drought tolerance. Curr Biol 15:1196–1200
Cooper B, Clarke JD, Budworth P, Kreps J, Hutchison D, Park S,Guimil S, Dunn M, Luginbuhl P, Ellero C, Goff SA, GlazebrookJ (2003) A network of rice genes associated with stress responseand seed development. Proc Natl Acad Sci USA 100:4945–4950
Cornic G, Bukhov NG, Wiese C, Bligny R, Heber U (2000) Flexiblecoupling between light-dependent electron and vectorial protontransport in illuminated leaves of C3 plants. Role of photosystemI-dependent proton pumping. Planta 210:468–477
Cosgrove DJ (2001) Wall structure and wall loosening. A lookbackwards and forwards. Plant Physiol 125:131–134
Cramer GR, Ergul A, Grimplet J, Tillett RL, Tattersall EA, BohlmanMC, Vincent D, Sonderegger J, Evans J, Osborne C, Quilici D,Schlauch KA, Schooley DA, Cushman JC (2007) Water andsalinity stress in grapevines: early and late changes in transcriptand metabolite profiles. Funct Integr Genomics 7:111–134
Cuming AC, Cho SH, Kamisugi Y, Graham H, Quatrano RS (2007)Microarray analysis of transcriptional responses to abscisic acidand osmotic, salt, and drought stress in the moss, Physcomitrellapatens. New Phytol 176:275–287
Curaba J, Herzog M, Vachon G (2003) GeBP, the first member of anew gene family in Arabidopsis, encodes a nuclear protein withDNA-binding activity and is regulated by KNAT1. Plant J33:305–317
Dai X, Xu Y, Ma Q, Xu W, Wang T, Xue Y, Chong K (2007)Overexpression of an R1R2R3 MYB gene, OsMYB3R-2,increases tolerance to freezing, drought, and salt stress intransgenic Arabidopsis. Plant Physiol 143:1739–1751
Davletova S, Schlauch K, Coutu J, Mittler R (2005) The zinc-fingerprotein Zat12 plays a central role in reactive oxygen and abioticstress signaling in Arabidopsis. Plant Physiol 139:847–856
Degenkolbe T, Do PT, Zuther E, Repsilber D, Walther D, Hincha DK,Kohl KI (2009) Expression profiling of rice cultivars differing intheir tolerance to long-term drought stress. Plant Mol Biol69:133–153
Dejardin A, Sokolov LN, Kleczkowski LA (1999) Sugar/osmoticumlevels modulate differential abscisic acid-independent expressionof two stress-responsive sucrose synthase genes in Arabidopsis.Biochem J 344(Pt 2):503–509
Desveaux D, Marechal A, Brisson N (2005) Whirly transcriptionfactors: defense gene regulation and beyond. Trends Plant Sci10:95–102
Dixon RA, Steele CL (1999) Flavonoids and isoflavonoids—a goldmine for metabolic engineering. Trends Plant Sci 4:394–400
Eliasson A, Gass N, Mundel C, Baltz R, Krauter R, Evrard JL,Steinmetz A (2000) Molecular and expression analysis of a LIMprotein gene family from flowering plants. Mol Gen Genet264:257–267
Evenson RE (1996) Priority setting methods. In: Evenson RE, HerdtRW, Hossain M (eds) Rice research in Asia: progress andpriorities. CAB International, Wallingford, pp 91–105
Fang Y, You J, Xie K, Xie W, Xiong L (2008) Systematic sequenceanalysis and identification of tissue-specific or stress-responsivegenes of NAC transcription factor family in rice. Mol GenetGenomics 280:547–563
Farrona S, Hurtado L, Bowman JL, Reyes JC (2004) The Arabidopsisthaliana SNF2 homolog AtBRM controls shoot development andflowering. Development 131:4965–4975
Fujita M, Fujita Y, Maruyama K, Seki M, Hiratsu K, Ohme-TakagiM, Tran LS, Yamaguchi-Shinozaki K, Shinozaki K (2004) Adehydration-induced NAC protein, RD26, is involved in anovel ABA-dependent stress-signaling pathway. Plant J39:863–876
Fujita Y, Fujita M, Satoh R, Maruyama K, Parvez MM, Seki M,Hiratsu K, Ohme-Takagi M, Shinozaki K, Yamaguchi-ShinozakiK (2005) AREB1 is a transcription activator of novel ABRE-dependent ABA signaling that enhances drought stress tolerancein Arabidopsis. Plant Cell 17:3470–3488
Fukai S, Cooper M (1995) Development of drought resistant cultivarsusing physio-morphological traits in rice. Field Crops Res 40:67–86
Furihata T, Maruyama K, Fujita Y, Umezawa T, Yoshida R, ShinozakiK, Yamaguchi-Shinozaki K (2006) Abscisic acid-dependentmultisite phosphorylation regulates the activity of a transcriptionactivator AREB1. Proc Natl Acad Sci USA 103:1988–1993
Garg AK, Kim JK, Owens TG, Ranwala AP, Choi YD, Kochian LV,Wu RJ (2002) Trehalose accumulation in rice plants confers hightolerance levels to different abiotic stresses. Proc Natl Acad SciUSA 99:15898–15903
Gilmour SJ, Sebolt AM, Salazar MP, Everard JD, Thomashow MF(2000) Overexpression of the Arabidopsis CBF3 transcriptionalactivator mimics multiple biochemical changes associated withcold acclimation. Plant Physiol 124:1854–1865
Gorantla M, Babu PR, Lachagari VB, Reddy AM, Wusirika R,Bennetzen JL, Reddy AR (2007) Identification of stress-responsive genes in an indica rice (Oryza sativa L.) using ESTsgenerated from drought-stressed seedlings. J Exp Bot 58:253–265
Grebe M, Xu J, Mobius W, Ueda T, Nakano A, Geuze HJ, Rook MB,Scheres B (2003) Arabidopsis sterol endocytosis involves actin-
174 Funct Integr Genomics (2011) 11:157–178

mediated trafficking via ARA6-positive early endosomes. CurrBiol 13:1378–1387
Guo Y, Gan S (2006) AtNAP, a NAC family transcription factor, hasan important role in leaf senescence. Plant J 46:601–612
Guo J, Wang MH (2009) Characterization of the phenylalanineammonia-lyase gene (SlPAL5) from tomato (Solanum lycopersi-cum L.). Mol Biol Rep 36:1579–1585
Hannah MA, Heyer AG, Hincha DK (2005) A global survey of generegulation during cold acclimation in Arabidopsis thaliana. PLoSGenet 1:e26
Hare PD, Cress WA, Van Staden J (1997) The involvement ofcytokinins in plant responses to environmental stress. PlantGrowth Regul 23:79–103
Hazen SP, Pathan MS, Sanchez A, Baxter I, Dunn M, Estes B, ChangHS, Zhu T, Kreps JA, Nguyen HT (2005) Expression profiling ofrice segregating for drought tolerance QTLs using a rice genomearray. Funct Integr Genomics 5:104–116
Hesse H, Willmitzer L (1996) Expression analysis of a sucrosesynthase gene from sugar beet (Beta vulgaris L.). Plant Mol Biol30:863–872
Hincha DK, Zuther E, Hellwege EM, Heyer AG (2002) Specificeffects of fructo- and gluco-oligosaccharides in the preservationof liposomes during drying. Glycobiology 12:103–110
Hoekstra FA, Golovina EA, Buitink J (2001) Mechanisms of plantdesiccation tolerance. Trends Plant Sci 6:431–438
Houde M, Belcaid M, Ouellet F, Danyluk J, Monroy AF, Dryanova A,Gulick P, Bergeron A, Laroche A, Links MG, MacCarthy L,Crosby WL, Sarhan F (2006) Wheat EST resources for functionalgenomics of abiotic stress. BMC Genomics 7:149
Hu H, Dai M, Yao J, Xiao B, Li X, Zhang Q, Xiong L (2006)Overexpressing a NAM, ATAF, and CUC (NAC) transcriptionfactor enhances drought resistance and salt tolerance in rice. ProcNatl Acad Sci USA 103:12987–12992
Huang J, Yang X, Wang MM, Tang HJ, Ding LY, Shen Y, Zhang HS(2007) A novel rice C2H2-type zinc finger protein lacking DLN-box/EAR-motif plays a role in salt tolerance. Biochim BiophysActa 1769:220–227
Imaizumi T, Kay SA (2006) Photoperiodic control of flowering: notonly by coincidence. Trends Plant Sci 11:550–558
Ito Y, Katsura K, Maruyama K, Taji T, Kobayashi M, Seki M,Shinozaki K, Yamaguchi-Shinozaki K (2006) Functional analysisof rice DREB1/CBF-type transcription factors involved in cold-responsive gene expression in transgenic rice. Plant Cell Physiol47:141–153
Itoh J, Nonomura K, Ikeda K, Yamaki S, Inukai Y, Yamagishi H,Kitano H, Nagato Y (2005) Rice plant development: from zygoteto spikelet. Plant Cell Physiol 46:23–47
Iturriaga G, Suarez R, Nova-Franco B (2009) Trehalose metabolism:from osmoprotection to signaling. Int J Mol Sci 10:3793–3810
Jaglo-Ottosen KR, Gilmour SJ, Zarka DG, Schabenberger O,Thomashow MF (1998) Arabidopsis CBF1 overexpressioninduces COR genes and enhances freezing tolerance. Science280:104–106
Jain M, Nijhawan A, Tyagi AK, Khurana JP (2006) Validation ofhousekeeping genes as internal control for studying geneexpression in rice by quantitative real-time PCR. BiochemBiophys Res Commun 345:646–651
Jain M, Nijhawan A, Arora R, Agarwal P, Ray S, Sharma P, Kapoor S,Tyagi AK, Khurana JP (2007) F-box proteins in rice. Genome-wide analysis, classification, temporal and spatial gene expres-sion during panicle and seed development, and regulation bylight and abiotic stress. Plant Physiol 143:1467–1483
Jain M, Ghanashyam C, Bhattacharjee A (2010) Comprehensiveexpression analysis suggests overlapping and specific roles ofrice glutathione S-transferase genes during development andstress responses. BMC Genomics 11:73
Jaiswal P, Ni J, Yap I, Ware D, Spooner W, Youens-Clark K, Ren L,Liang C, Zhao W, Ratnapu K, Faga B, Canaran P, Fogleman M,Hebbard C, Avraham S, Schmidt S, Casstevens TM, Buckler ES,Stein L, McCouch S (2006) Gramene: a bird’s eye view of cerealgenomes. Nucleic Acids Res 34:D717–D723
Jeong JS, Kim YS, Baek KH, Jung H, Ha SH, Do Choi Y, Kim M,Reuzeau C, Kim JK (2010) Root-specific expression ofOsNAC10 improves drought tolerance and grain yield in riceunder field drought conditions. Plant Physiol 153:185-197
Jiang Y, Deyholos MK (2006) Comprehensive transcriptional profil-ing of NaCl-stressed Arabidopsis roots reveals novel classes ofresponsive genes. BMC Plant Biol 6:25
Jung C, Seo JS, Han SW, Koo YJ, Kim CH, Song SI, Nahm BH, ChoiYD, Cheong JJ (2008) Overexpression of AtMYB44 enhancesstomatal closure to confer abiotic stress tolerance in transgenicArabidopsis. Plant Physiol 146:623–635
Kang JY, Choi HI, Im MY, Kim SY (2002) Arabidopsis basic leucinezipper proteins that mediate stress-responsive abscisic acidsignaling. Plant Cell 14:343–357
Kaplan F, Kopka J, Haskell DW, Zhao W, Schiller KC, Gatzke N,Sung DY, Guy CL (2004) Exploring the temperature-stressmetabolome of Arabidopsis. Plant Physiol 136:4159–4168
Kaplan F, Kopka J, Sung DY, Zhao W, Popp M, Porat R, Guy CL(2007) Transcript and metabolite profiling during cold acclima-tion of Arabidopsis reveals an intricate relationship of cold-regulated gene expression with modifications in metabolitecontent. Plant J 50:967–981
Kasuga M, Liu Q, Miura S, Yamaguchi-Shinozaki K, Shinozaki K(1999) Improving plant drought, salt, and freezing tolerance bygene transfer of a single stress-inducible transcription factor. NatBiotechnol 17:287–291
Kawaguchi R, Girke T, Bray EA, Bailey-Serres J (2004) DifferentialmRNA translation contributes to gene regulation under non-stress and dehydration stress conditions in Arabidopsis thaliana.Plant J 38:823–839
Khush GS (1995) Modern varieties: their real contribution to foodsupply and equity. GeoJournal 35:275–284
Kidd PS, Llugany M, Poschenrieder C, Gunse B, Barcelo J (2001)The role of root exudates in aluminium resistance and silicon-induced amelioration of aluminium toxicity in three varieties ofmaize (Zea mays L.). J Exp Bot 52:1339–1352
Kilian J, Whitehead D, Horak J, Wanke D, Weinl S, Batistic O,D’Angelo C, Bornberg-Bauer E, Kudla J, Harter K (2007) TheAtGenExpress global stress expression data set: protocols,evaluation and model data analysis of UV-B light, drought andcold stress responses. Plant J 50:347–363
Kim SH, Hong JK, Lee SC, Sohn KH, Jung HW, Hwang BK (2004)CAZFP1, Cys2/His2-type zinc-finger transcription factor genefunctions as a pathogen-induced early-defense gene in Capsicumannuum. Plant Mol Biol 55:883–904
Kim SY, Yu X, Michaels SD (2008) Regulation of CONSTANS andFLOWERING LOCUS T expression in response to changinglight quality. Plant Physiol 148:269–279
Kleines M, Elster RC, Rodrigo MJ, Blervacq AS, Salamini F, BartelsD (1999) Isolation and expression analysis of two stress-responsive sucrose-synthase genes from the resurrection plantCraterostigma plantagineum (Hochst.). Planta 209:13–24
Kobayashi Y, Weigel D (2007) Move on up, it’s time for change–mobile signals controlling photoperiod-dependent flowering.Genes Dev 21:2371–2384
Kreps JA, Wu Y, Chang HS, Zhu T, Wang X, Harper JF (2002)Transcriptome changes for Arabidopsis in response to salt,osmotic, and cold stress. Plant Physiol 130:2129–2141
Lan L, Chen W, Lai Y, Suo J, Kong Z, Li C, Lu Y, Zhang Y,Zhao X, Zhang X, Zhang Y, Han B, Cheng J, Xue Y (2004)Monitoring of gene expression profiles and isolation of
Funct Integr Genomics (2011) 11:157–178 175

candidate genes involved in pollination and fertilization inrice (Oryza sativa L.) with a 10 K cDNA microarray. PlantMol Biol 54:471–487
Lan L, Li M, Lai Y, Xu W, Kong Z, Ying K, Han B, Xue Y(2005) Microarray analysis reveals similarities and variationsin genetic programs controlling pollination/fertilization andstress responses in rice (Oryza sativa L.). Plant Mol Biol59:151–164
Lawlor DW (2002) Limitation to photosynthesis in water-stressedleaves: stomata vs. metabolism and the role of ATP. Ann Bot(Lond) 89 Spec No:871–885
Leonhardt N, Kwak JM, Robert N, Waner D, Leonhardt G, SchroederJI (2004) Microarray expression analyses of Arabidopsis guardcells and isolation of a recessive abscisic acid hypersensitiveprotein phosphatase 2C mutant. Plant Cell 16:596–615
Leyman B, Van Dijck P, Thevelein JM (2001) An unexpected plethoraof trehalose biosynthesis genes in Arabidopsis thaliana. TrendsPlant Sci 6:510–513
Liu Q, Kasuga M, Sakuma Y, Abe H, Miura S, Yamaguchi-ShinozakiK, Shinozaki K (1998) Two transcription factors, DREB1 andDREB2, with an EREBP/AP2 DNA binding domain separatetwo cellular signal transduction pathways in drought- and low-temperature-responsive gene expression, respectively, in Arabi-dopsis. Plant Cell 10:1391–1406
Liu H, Wang S, Yu X, Yu J, He X, Zhang S, Shou H, Wu P (2005)ARL1, a LOB-domain protein required for adventitious rootformation in rice. Plant J 43:47–56
Liu F, Xu W, Wei Q, Zhang Z, Xing Z, Tan L, Di C, Yao D, Wang C,Tan Y, Yan H, Ling Y, Sun C, Xue Y, Su Z (2010) Geneexpression profiles deciphering rice phenotypic variation be-tween Nipponbare (Japonica) and 93-11 (Indica) during oxida-tive stress. PLoS ONE 5:e8632
Ma T, Chen R, Yu R, Zeng H, Zhang D (2009) Differential globalgenomic changes in rice root in response to low-, middle-, andhigh-osmotic stresses. Acta Physiol Plant 31:773–785
Mahajan S, Tuteja N (2005) Cold, salinity and drought stresses: anoverview. Arch Biochem Biophys 444:139–158
Major IT, Constabel CP (2006) Molecular analysis of poplar defenseagainst herbivory: comparison of wound- and insect elicitor-induced gene expression. New Phytol 172:617–635
Mantri NL, Ford R, Coram TE, Pang EC (2007) Transcriptionalprofiling of chickpea genes differentially regulated in response tohigh-salinity, cold and drought. BMC Genomics 8:303
Markandeya G, Babu PR, Lachagari VBR, Feltus FA, PatersonAH, Reddy AR (2005) Functional genomics of drought-stressresponse in rice: transcript mapping of annotated unigenes ofan indica rice (Oryza sativa L. cv. Nagina 22). Curr Sci89:496–514
Martinelli T (2008) In situ localization of glucose and sucrose indehydrating leaves of Sporobolus stapfianus. J Plant Physiol165:580–587
Medina J, Catala R, Salinas J (2001) Developmental and stressregulation of RCI2A and RCI2B, two cold-inducible genes ofArabidopsis encoding highly conserved hydrophobic proteins.Plant Physiol 125:1655–1666
Mehrtens F, Kranz H, Bednarek P, Weisshaar B (2005) TheArabidopsis transcription factor MYB12 is a flavonol-specificregulator of phenylpropanoid biosynthesis. Plant Physiol138:1083–1096
Moore JP, Lindsey GG, Farrant JM, Brandt WF (2007) An overviewof the biology of the desiccation-tolerant resurrection plantMyrothamnus flabellifolia. Ann Bot 99:211–217
Mukhopadhyay A, Vij S, Tyagi AK (2004) Overexpression of a zinc-finger protein gene from rice confers tolerance to cold,dehydration, and salt stress in transgenic tobacco. Proc NatlAcad Sci USA 101:6309–6314
Nakashima K, Ito Y, Yamaguchi-Shinozaki K (2009) Transcriptionalregulatory networks in response to abiotic stresses in Arabidopsisand grasses. Plant Physiol 149:88–95
Narciso J, Hossain M (2002) World Rice Statistics [Online]. Availableby IRRI http://www.irri.org/science/ ricestat (posted 2002)
Narusaka Y, Narusaka M, Seki M, Umezawa T, Ishida J, Nakajima M,Enju A, Shinozaki K (2004) Crosstalk in the responses to abioticand biotic stresses in Arabidopsis: analysis of gene expression incytochrome P450 gene superfamily by cDNA microarray. PlantMol Biol 55:327–342
Nijhawan A, Jain M, Tyagi AK, Khurana JP (2008) Genomic surveyand gene expression analysis of the basic leucine zippertranscription factor family in rice. Plant Physiol 146:333–350
Noh B, Lee SH, Kim HJ, Yi G, Shin EA, Lee M, Jung KJ, Doyle MR,Amasino RM, Noh YS (2004) Divergent roles of a pair ofhomologous jumonji/zinc-finger-class transcription factor pro-teins in the regulation of Arabidopsis flowering time. Plant Cell16:2601–2613
Olsen AN, Ernst HA, Leggio LL, Skriver K (2005) NAC transcriptionfactors: structurally distinct, functionally diverse. Trends PlantSci 10:79–87
Oono Y, Seki M, Nanjo T, Narusaka M, Fujita M, Satoh R, Satou M,Sakurai T, Ishida J, Akiyama K, Iida K, Maruyama K, Satoh S,Yamaguchi-Shinozaki K, Shinozaki K (2003) Monitoring ex-pression profiles of Arabidopsis gene expression during rehydra-tion process after dehydration using ca 7000 full-length cDNAmicroarray. Plant J 34:868–887
Owens DK, Alerding AB, Crosby KC, Bandara AB, Westwood JH,Winkel BS (2008) Functional analysis of a predicted flavonolsynthase gene family in Arabidopsis. Plant Physiol 147:1046–1061
Oztur ZN, Talame V, Deyholos M, Michalowski CB, Galbraith DW,Gozukirmizi N, Tuberosa R, Bohnert HJ (2002) Monitoringlarge-scale changes in transcript abundance in drought- and salt-stressed barley. Plant Mol Biol 48:551–573
Park HC, Kim ML, Lee SM, Bahk JD, Yun DJ, Lim CO, Hong JC,Lee SY, Cho MJ, Chung WS (2007) Pathogen-induced bindingof the soybean zinc finger homeodomain proteins GmZF-HD1and GmZF-HD2 to two repeats of ATTA homeodomain bindingsite in the calmodulin isoform 4 (GmCaM4) promoter. NucleicAcids Res 35:3612–3623
Parry MA, Andralojc PJ, Khan S, Lea PJ, Keys AJ (2002) Rubiscoactivity: effects of drought stress. Ann Bot (Lond) 89 SpecNo:833–839
Paterson AH, Bowers JE, Bruggmann R, Dubchak I, Grimwood J,Gundlach H, Haberer G, Hellsten U, Mitros T, Poliakov A,Schmutz J, Spannagl M, Tang H, Wang X, Wicker T, Bharti AK,Chapman J, Feltus FA, Gowik U, Grigoriev IV, Lyons E, MaherCA, Martis M, Narechania A, Otillar RP, Penning BW, SalamovAA, Wang Y, Zhang L, Carpita NC, Freeling M, Gingle AR,Hash CT, Keller B, Klein P, Kresovich S, McCann MC, Ming R,Peterson DG, Mehboob ur R, Ware D, Westhoff P, Mayer KF,Messing J, Rokhsar DS (2009) The Sorghum bicolor genome andthe diversification of grasses. Nature 457:551–556
Pandey S, Zhang W, Assmann SM (2007) Roles of ion channels andtransporters in guard cell signal transduction. FEBS Lett581:2325–2336
Peiter E, Maathuis FJ, Mills LN, Knight H, Pelloux J, HetheringtonAM, Sanders D (2005) The vacuolar Ca2+-activated channelTPC1 regulates germination and stomatal movement. Nature434:404–408
Peng H, Cheng HY, Chen C, Yu XW, Yang JN, Gao WR, Shi QH,Zhang H, Li JG, Ma H (2009) A NAC transcription factor geneof chickpea (Cicer arietinum), CarNAC3, is involved in droughtstress response and various developmental processes. J PlantPhysiol 166:1934–1945
176 Funct Integr Genomics (2011) 11:157–178

Post-Beittenmiller D (1996) Biochemistry and molecular biology ofwax production in plants. Annu Rev Plant Physiol Plant Mol Biol47:405–430
Rabbani MA, Maruyama K, Abe H, Khan MA, Katsura K, Ito Y,Yoshiwara K, Seki M, Shinozaki K, Yamaguchi-Shinozaki K(2003) Monitoring expression profiles of rice genes under cold,drought, and high-salinity stresses and abscisic acid applicationusing cDNA microarray and RNA gel-blot analyses. PlantPhysiol 133:1755–1767
Rabello AR, Guimaraes CM, Rangel PH, da Silva FR, Seixas D, deSouza E, Brasileiro AC, Spehar CR, Ferreira ME, Mehta A(2008) Identification of drought-responsive genes in roots ofupland rice (Oryza sativa L). BMC Genomics 9:485
Ramanjulu S, Bartels D (2002) Drought- and desiccation-inducedmodulation of gene expression in plants. Plant Cell Environ25:141–151
Ramon M, Rolland F (2007) Plant development: introducing trehalosemetabolism. Trends Plant Sci 12:185–188
Rao MV, Koch JR, Davis KR (2000) Ozone: a tool for probingprogrammed cell death in plants. Plant Mol Biol 44:345–358
Rathinasabapathi B, Fouad WM, Sigua CA (2001) β-Alanine betainesynthesis in the Plumbaginaceae. Purification and characteriza-tion of a trifunctional, S-adenosyl-L-methionine-dependent N-methyltransferase from Limonium latifolium leaves. Plant Physiol126:1241–1249
Ray S, Agarwal P, Arora R, Kapoor S, Tyagi AK (2007) Expressionanalysis of calcium-dependent protein kinase gene family duringreproductive development and abiotic stress conditions in rice(Oryza sativa L. ssp. indica). Mol Genet Genomics 278:493–505
Reddy AR, Ramakrishna W, Sekhar AC, Ithal N, Babu PR, BonaldoMF, Soares MB, Bennetzen JL (2002) Novel genes are enrichedin normalized cDNA libraries from drought-stressed seedlings ofrice (Oryza sativa L. subsp. indica cv. Nagina 22). Genome45:204–211
Rensink WA, Buell CR (2005) Microarray expression profilingresources for plant genomics. Trends Plant Sci 10:603–609
Reyes JC, Muro-Pastor MI, Florencio FJ (2004) The GATA family oftranscription factors in Arabidopsis and rice. Plant Physiol134:1718–1732
Rivero RM, Kojima M, Gepstein A, Sakakibara H, Mittler R,Gepstein S, Blumwald E (2007) Delayed leaf senescence inducesextreme drought tolerance in a flowering plant. Proc Natl AcadSci USA 104:19631–19636
Rizhsky L, Liang H, Mittler R (2002) The combined effect of droughtstress and heat shock on gene expression in tobacco. PlantPhysiol 130:1143–1151
Rizhsky L, Davletova S, Liang H, Mittler R (2004) The zinc fingerprotein Zat12 is required for cytosolic ascorbate peroxidase 1expression during oxidative stress in Arabidopsis. J Biol Chem279:11736–11743
Roche J, Hewezi T, Bouniols A, Gentzbittel L (2007) Transcriptionalprofiles of primary metabolism and signal transduction-relatedgenes in response to water stress in field-grown sunflowergenotypes using a thematic cDNA microarray. Planta 226:601–617
Ryan KG, Swinny EE, Winefield C, Markham KR (2001) Flavonoidsand UV photoprotection in Arabidopsis mutants. Z Naturforsch C56:745–754
Ryan KG, Swinny EE, Markham KR, Winefield C (2002) Flavonoidgene expression and UV photoprotection in transgenic andmutant Petunia leaves. Phytochemistry 59:23–32
Sablowski RW, Meyerowitz EM (1998) A homolog of NO APICALMERISTEM is an immediate target of the floral homeotic genesAPETALA3/PISTILLATA. Cell 92:93–103
Sakamoto H, Maruyama K, Sakuma Y, Meshi T, Iwabuchi M,Shinozaki K, Yamaguchi-Shinozaki K (2004) Arabidopsis
Cys2/His2-type zinc-finger proteins function as transcriptionrepressors under drought, cold, and high-salinity stress con-ditions. Plant Physiol 136:2734–2746
Sakuma Y, Maruyama K, Osakabe Y, Qin F, Seki M, Shinozaki K,Yamaguchi-Shinozaki K (2006) Functional analysis of anArabidopsis transcription factor, DREB2A, involved in drought-responsive gene expression. Plant Cell 18:1292–1309
Schadt EE, Li C, Ellis B, Wong WH (2001) Feature extraction andnormalization algorithms for high-density oligonucleotide geneexpression array data. J Cell Biochem Suppl 37:120–125
Seki M, Narusaka M, Abe H, Kasuga M, Yamaguchi-Shinozaki K,Carninci P, Hayashizaki Y, Shinozaki K (2001) Monitoring theexpression pattern of 1300 Arabidopsis genes under drought andcold stresses by using a full-length cDNA microarray. Plant Cell13:61–72
Seki M, Ishida J, Narusaka M, Fujita M, Nanjo T, Umezawa T,Kamiya A, Nakajima M, Enju A, Sakurai T, Satou M, AkiyamaK, Yamaguchi-Shinozaki K, Carninci P, Kawai J, Hayashizaki Y,Shinozaki K (2002a) Monitoring the expression pattern of around7, 000 Arabidopsis genes under ABA treatments using a full-length cDNA microarray. Funct Integr Genomics 2:282–291
Seki M, Narusaka M, Ishida J, Nanjo T, Fujita M, Oono Y, Kamiya A,Nakajima M, Enju A, Sakurai T, Satou M, Akiyama K, Taji T,Yamaguchi-Shinozaki K, Carninci P, Kawai J, Hayashizaki Y,Shinozaki K (2002b) Monitoring the expression profiles of 7000Arabidopsis genes under drought, cold and high-salinity stressesusing a full-length cDNA microarray. Plant J 31:279–292
Seo PJ, Park CM (2010) A membrane-bound NAC transcription factoras an integrator of biotic and abiotic stress signals. Plant SignalBehav 5:481–483
Sharma AD, Gill PK, Singh P (2003) RNA isolation from plant tissuesrich in polysaccharides. Anal Biochem 314:319–321
Shinozaki K, Yamaguchi-Shinozaki K (2007) Gene networks involvedin drought stress response and tolerance. J Exp Bot 58:221–227
Sivamani E, Bahieldin A, Wraith JM, Al-Niemi T, Dyer WE, Ho TD, QuR (2000) Improved biomass productivity and water use efficiencyunder water deficit conditions in transgenic wheat constitutivelyexpressing the barley HVA1 gene. Plant Sci 155:1–9
Street NR, Skogstrom O, Sjodin A, Tucker J, Rodriguez-Acosta M,Nilsson P, Jansson S, Taylor G (2006) The genetics and genomicsof the drought response in Populus. Plant J 48:321–341
Sun J, Jiang H, Xu Y, Li H, Wu X, Xie Q, Li C (2007) The CCCH-type zinc finger proteins AtSZF1 and AtSZF2 regulate salt stressresponses in Arabidopsis. Plant Cell Physiol 48:1148–1158
Talame V, Ozturk NZ, Bohnert HJ, Tuberosa R (2007) Barleytranscript profiles under dehydration shock and drought stresstreatments: a comparative analysis. J Exp Bot 58:229–240
Tattersall EA, Grimplet J, DeLuc L, Wheatley MD, Vincent D,Osborne C, Ergul A, Lomen E, Blank RR, Schlauch KA,Cushman JC, Cramer GR (2007) Transcript abundance profilesreveal larger and more complex responses of grapevine tochilling compared to osmotic and salinity stress. Funct IntegrGenomics 7:317–333
Thomas JC, Smigocki AC, Bohnert HJ (1995) Light-inducedexpression of ipt from Agrobacterium tumefaciens results incytokinin accumulation and osmotic stress symptoms in trans-genic tobacco. Plant Mol Biol 27:225–235
Thorstensen T, Grini PE, Mercy IS, Alm V, Erdal S, Aasland R, AalenRB (2008) The Arabidopsis SET-domain protein ASHR3 isinvolved in stamen development and interacts with the bHLHtranscription factor ABORTED MICROSPORES (AMS). PlantMol Biol 66:47–59
Tran LS, Nakashima K, Sakuma Y, Simpson SD, Fujita Y, MaruyamaK, Fujita M, Seki M, Shinozaki K, Yamaguchi-Shinozaki K(2004) Isolation and functional analysis of Arabidopsis stress-inducible NAC transcription factors that bind to a drought-
Funct Integr Genomics (2011) 11:157–178 177

responsive cis-element in the early responsive to dehydrationstress 1 promoter. Plant Cell 16:2481–2498
Tran LS, Urao T, Qin F, Maruyama K, Kakimoto T, Shinozaki K,Yamaguchi-Shinozaki K (2007) Functional analysis of AHK1/ATHK1 and cytokinin receptor histidine kinases in response toabscisic acid, drought, and salt stress in Arabidopsis. Proc NatlAcad Sci USA 104:20623–20628
Tsuda K, Tsuji T, Hirose S, Yamazaki K (2004) Three ArabidopsisMBF1 homologs with distinct expression profiles play roles astranscriptional co-activators. Plant Cell Physiol 45:225–231
Tuba Z, Lichtenthaler HK (2007) Long-term acclimation of plants toelevated CO2 and its interaction with stresses. Ann NYAcad Sci1113:135–146
Tuberosa R, Salvi S (2006) Genomics-based approaches to improvedrought tolerance of crops. Trends Plant Sci 11:405–412
Van den Ende W, Valluru R (2009) Sucrose, sucrosyl oligosaccharides,and oxidative stress: scavenging and salvaging? J Exp Bot 60:9–18
Van Holme B, Grunewald W, Bateman A, Kohchi T, Gheysen G(2007) The tify family previously known as ZIM. Trends PlantSci 12:239–244
Varshney RK, Hiremath PJ, Lekha P, Kashiwagi J, Balaji J, DeokarAA, Vadez V, Xiao Y, Srinivasan R, Gaur PM, Siddique KH,Town CD, Hoisington DA (2009) A comprehensive resource ofdrought- and salinity- responsive ESTs for gene discovery andmarker development in chickpea (Cicer arietinum L.). BMCGenomics 10:523
Verdier J, Kakar K, Gallardo K, Le Signor C, Aubert G, Schlereth A,Town CD, Udvardi MK, Thompson RD (2008) Gene expressionprofiling of M. truncatula transcription factors identifies putativeregulators of grain legume seed filling. Plant Mol Biol 67:567–580
Vereyken IJ, Chupin V, Islamov A, Kuklin A, Hincha DK, de KruijffB (2003) The effect of fructan on the phospholipid organizationin the dry state. Biophys J 85:3058–3065
Vij S, Tyagi AK (2007) Emerging trends in the functional genomics ofthe abiotic stress response in crop plants. Plant Biotechnol J5:361–380
Vij S, Giri J, Dansana PK, Kapoor S, Tyagi AK (2008) The receptor-like cytoplasmic kinase (OsRLCK) gene family in rice: organi-zation, phylogenetic relationship, and expression during devel-opment and stress. Mol Plant 1:732–750
Villalobos MA, Bartels D, Iturriaga G (2004) Stress tolerance andglucose insensitive phenotypes in Arabidopsis overexpressing theCpMYB10 transcription factor gene. Plant Physiol 135:309–324
Vu JCV, Allen LH Jr (2009) Growth at elevated CO(2) delays theadverse effects of drought stress on leaf photosynthesis of the C(4) sugarcane. J Plant Physiol 166:107–116
Walia H, Wilson C, Condamine P, Liu X, Ismail AM, Zeng L,Wanamaker SI, Mandal J, Xu J, Cui X, Close TJ (2005)Comparative transcriptional profiling of two contrasting ricegenotypes under salinity stress during the vegetative growthstage. Plant Physiol 139:822–835
Wang H, Cutler AJ (1995) Promoters from KIN1 and COR66, twoArabidopsis thaliana low-temperature- and ABA-induciblegenes, direct strong beta-glucuronidase expression in guard cells,pollen and young developing seeds. Plant Mol Biol 28:619–634
Wang H, Zhang H, Gao F, Li J, Li Z (2007) Comparison of geneexpression between upland and lowland rice cultivars underwater stress using cDNA microarray. Theor Appl Genet115:1109–1126
Watkinson JI, Sioson AA, Vasquez-Robinet C, Shukla M, Kumar D,Ellis M, Heath LS, Ramakrishnan N, Chevone B, Watson LT, vanZyl L, Egertsdotter U, Sederoff RR, Grene R (2003) Photosyn-thetic acclimation is reflected in specific patterns of gene
expression in drought-stressed loblolly pine. Plant Physiol133:1702–1716
Whittaker A, Bochicchio A, Vazzana C, Lindsey G, Farrant J (2001)Changes in leaf hexokinase activity and metabolite levels inresponse to drying in the desiccation-tolerant species Sporobolusstapfianus and Xerophyta viscosa. J Exp Bot 52:961–969
Widawsky DA, O’Toole JC (1990) Prioritizing the rice biotechnologyresearch agenda for eastern India. The Rockefeller Foundation,New York, p 86
Winkel-Shirley B (2002) Biosynthesis of flavonoids and effects ofstress. Curr Opin Plant Biol 5:218–223
Winkel T, Renno JF, Payne WA (1997) Effect of the timing of waterdeficit on growth, phenology and yield of pearl millet (Pennise-tum glaucum (L.) R. Br.) grown in Sahelian conditions. J ExpBot 48:1001–1009
Wong CE, Li Y, Labbe A, Guevara D, Nuin P, Whitty B, Diaz C,Golding GB, Gray GR, Weretilnyk EA, Griffith M, Moffatt BA(2006) Transcriptional profiling implicates novel interactionsbetween abiotic stress and hormonal responses in Thellungiella, aclose relative of Arabidopsis. Plant Physiol 140:1437–1450
Wu Z, Irazarry RA, Gentleman R, Murillo FM, Spencer F (2004) Amodel based background adjustment for oligonucleotide expres-sion arrays. Johns Hopkins University, Dept. of BiostatisticsWorking Papers paper 1
Xiong L, Zhu JK (2002) Molecular and genetic aspects of plantresponses to osmotic stress. Plant Cell Environ 25:131–139
Yamada M, Morishita H, Urano K, Shiozaki N, Yamaguchi-ShinozakiK, Shinozaki K, Yoshiba Y (2005) Effects of free prolineaccumulation in petunias under drought stress. J Exp Bot56:1975–1981
Yamaguchi-Shinozaki K, Shinozaki K (1993) Arabidopsis DNAencoding two desiccation-responsive rd29 genes. Plant Physiol101:1119–1120
Yamaguchi-Shinozaki K, Shinozaki K (2006) Transcriptional regula-tory networks in cellular responses and tolerance to dehydrationand cold stresses. Annu Rev Plant Biol 57:781–803
Yanagisawa S (2004) Dof domain proteins: plant-specific transcriptionfactors associated with diverse phenomena unique to plants. PlantCell Physiol 45:386–391
Yu LX, Setter TL (2003) Comparative transcriptional profiling ofplacenta and endosperm in developing maize kernels in responseto water deficit. Plant Physiol 131:568–582
Yu X, Li L, Guo M, Chory J, Yin Y (2008) Modulation ofbrassinosteroid-regulated gene expression by Jumonji domain-containing proteins ELF6 and REF6 in Arabidopsis. Proc NatlAcad Sci USA 105:7618–7623
Zhang Y, Wang L (2005) The WRKY transcription factor superfamily:its origin in eukaryotes and expansion in plants. BMC Evol Biol5:1
Zheng J, Zhao J, Tao Y, Wang J, Liu Y, Fu J, Jin Y, Gao P, Zhang J,Bai Y, Wang G (2004) Isolation and analysis of water stressinduced genes in maize seedlings by subtractive PCR and cDNAmacroarray. Plant Mol Biol 55:807–823
Zhou J, Wang X, Jiao Y, Qin Y, Liu X, He K, Chen C, Ma L, Wang J,Xiong L, Zhang Q, Fan L, Deng XW (2007) Global genomeexpression analysis of rice in response to drought and high-salinitystresses in shoot, flag leaf, and panicle. Plant Mol Biol 63:591–608
Zhou J, Li F, Wang JL, Ma Y, Chong K, Xu YY (2009) Basic helix-loop-helix transcription factor from wild rice (OrbHLH2)improves tolerance to salt- and osmotic stress in Arabidopsis. JPlant Physiol 166:1296–1306
Zhu Q, Dabi T, Beeche A, Yamamoto R, Lawton MA, Lamb C (1995)Cloning and properties of a rice gene encoding phenylalanineammonia-lyase. Plant Mol Biol 29:535–550
178 Funct Integr Genomics (2011) 11:157–178





























