Embed Size (px)
Citation preview

Biological Journal of the Linnean Society (1999), 67: 97–118. With 1 figure
Article ID: bijl.1998.0291, available online at http://www.idealibrary.com on
Molecular evidence for the phylogeny ofAustralian gekkonoid lizards
STEPHEN C. DONNELLAN∗, MARK N. HUTCHINSON ANDKATHLEEN M. SAINT
South Australian Museum, North Terrace, Adelaide, Australia 5000
Received 29 January 1998; accepted for publication 22 September 1998
Partial sequences of the mitochondrial 12S rRNA and nuclear c-mos genes were determinedfor 12 species of gekkonoid lizards representing the four major taxa of the Australianregion, the Diplodactylini and Carphodactylini (forming the subfamily Diplodactylinae), thePygopodidae and the Gekkoninae. One further species represented a non-Australian gek-konoid lineage, the Eublepharinae. The combined sequence data were used to reconstructthe underlying molecular phylogeny. We used the molecular phylogeny to test the monophylyof the diplodactyline tribes and conflicting hypotheses of relationships of the pygopods andof the genus Oedura. Monophyly of the Diplodactylinae is supported, while pygopods forma monophyletic sister lineage to all Diplodactylinae. The molecular data support themonophyly of the Diplodactylini, with Oedura firmly placed as a diplodactylin. Monophylyof the Carphodactylini is not supported. The four carphodactylin genera form a paraphyleticcluster at the base of the Diplodactylini. Pygopods are nested within the traditional Gekkonidaeand pygopods plus diplodactylines form a well-supported monophyletic group with respectto the remaining gekkonoids, the gekkonines and eublepharines.
1999 The Linnean Society of London
ADDITIONAL KEY WORDS:—molecular systematics – reptile – mitochondrial DNA –nuclear gene – Gekkota – Gekkonidae – Pygopodidae – Sauria.
CONTENTS
Introduction . . . . . . . . . . . . . . . . . . . . . . . 98Material and methods . . . . . . . . . . . . . . . . . . . 100
DNA sources . . . . . . . . . . . . . . . . . . . . . 100PCR methods . . . . . . . . . . . . . . . . . . . . 100Analysis of sequence data . . . . . . . . . . . . . . . . . 101
Results . . . . . . . . . . . . . . . . . . . . . . . . 102Discussion . . . . . . . . . . . . . . . . . . . . . . . 105
Molecular evolution . . . . . . . . . . . . . . . . . . . 105Carphodactylini and the position of Oedura . . . . . . . . . . . 105The relationships of pygopods . . . . . . . . . . . . . . . 106Sequence availability . . . . . . . . . . . . . . . . . . 107
Acknowledgements . . . . . . . . . . . . . . . . . . . . 107References . . . . . . . . . . . . . . . . . . . . . . . 108Appendix . . . . . . . . . . . . . . . . . . . . . . . . 110
∗Corresponding author. E-mail: [email protected]
970024–4066/99/050097+22 $30.00/0 1999 The Linnean Society of London

S. C. DONNELLAN ET AL.98
INTRODUCTION
The lizards and snakes, order Squamata, include several major evolutionarylineages, each including hundreds of species and scores of genera. Most authorsnow recognize several major squamate lineages, Iguania, Gekkota, Scincomorpha,Anguinomorpha, Serpentes and Amphisbaenia. Strict monophyly of each of theselineages has yet to be established, and getting a clear evolutionary perspective oneven the major features of their phylogeny, both within and between lineages, hasproven to be a slow task. In this article we present molecular data which help toshed light on the relationships within one of these lineages, the Gekkota, originallyerected for the lizards commonly known as geckoes (geckos or gekkos) now usuallyplaced in a single family Gekkonidae.
McDowell & Bogert (1954) expanded the concept of the Gekkota to include theXantusiidae, the night lizards of the New World, and the Pygopodidae, a limblessgroup confined to Australia and New Guinea, as well as the Gekkonidae. Subsequentstudy of diverse morphological data sets (e.g. Estes et al., 1988; Presch, 1988) resultedin the transferral of xantusiids to the Scincomorpha (Estes, 1983; Estes et al., 1988),although recent reanalysis of the data of Estes et al. (1988) and Schwenk (1988) byMacey et al. (1997) again places xantusiids with the Gekkota rather than theScincomorpha. All these studies, however, plus additional work (e.g. Underwood,1957; Kluge, 1974, 1987), have confirmed the close affinity between pygopods andtraditional gekkonids.
Following pioneering work on the relationships of both geckoes (Underwood,1954; Stephenson, 1960; Holder, 1960) and pygopods (Underwood, 1957), the mostrecent research has its origins in the studies of Kluge (1967a, b). In these he revisedUnderwood’s subfamilial scheme, recognizing a single family, Gekkonidae, with foursubfamilies: Gekkoninae (cosmopolitan), Diplodactylinae (Australia–New Caledonia–New Zealand), Sphaerodactylinae (tropical America and Africa) and Eublepharinae(worldwide but excluding Australasia). The Diplodactylinae was subdivided intotwo tribes, Diplodactylini and Carphodactylini. Kluge later (1974, 1976) revisedPygopodidae, recognizing many new species, validating a generic scheme andproposing phylogenetic relationships within the family. The relationship betweenpygopods and gekkonids was explored in a subsequent review (Kluge, 1987), of majorgekkonoid lineage relationships. Kluge (1987) downgraded the Sphaerodactylinae toa lineage within the Gekkoninae and concluded that pygopods are the sister lineageof Diplodactylinae, rendering the Gekkonidae paraphyletic. He recommended ataxonomy which reassigned the Diplodactylinae as a subfamily of the Pygopodidaebut alternative taxonomies, such as submergence of Pygopodidae into Gekkonidae(Bauer, 1990) would still reflect the phylogeny.
Since Kluge’s revision, Russell (1979) analysed the probable course of evolutionof toe pad structures in diplodactylines and gekkonines, concluding that theDiplodactylinae was most likely primitively padless, and that pads had probablyarisen only once. This implies paraphyly for the Carphodactylini, with paddedcarphodactylins sharing a sister group relationship with the Diplodactylini, whilepadless taxa (the Australian genera Carphodactylus, Nephrurus, Phyllurus and Saltuarius)are the sister lineage of these two. Russell did not push his conclusions to the levelof a formal taxonomic change, and Bauer (1990) continued to accept Kluge’s (1967b)morphological diagnosis of the Carphodactylini.
During the 1980s, King’s karyological studies (King, 1985, 1987a, 1987b, 1990;

EVOLUTIONARY RELATIONSHIPS OF AUSTRALIAN GEKKONOID LIZARDS 99
King & Mengden, 1990) generated new data bearing on the problems of gekkonoidphylogeny. The karyotypic arguments revolve around the course of evolution of twoextreme conditions of a 2n=38 karyotype. Most diplodactylin gecko lineages includean all acrocentric karyotype. Most of Kluge’s carphodactylins have an all metacentric/submetacentric (=biarmed) condition. King (1987a) noted the existence of karyotypeswithin the diplodactylins where a few species which had mostly acrocentric karyotypeshave one or a few biarmed chromosomes. The fact that the identity of thesebiarmed chromosomes differs from species to species strongly suggested to King anevolutionary process of pericentric rearrangements modifying a primitively all-acrocentric karyotype. The all-biarmed karyotype of most carphodactylins (Nephrurusand Phyllurus have an all-acrocentric arrangement) was interpreted as the endpointof a series of pericentric rearrangements that had completely transformed anoriginally all-acrocentric karyotype. King concluded that the course of karyotypicevolution within the subfamily Diplodactylinae had started from an ancestral2n=38 acrocentric karyotype. This karyotype had been mostly retained by theDiplodactylini, with at most a few chromosomes becoming biarmed via pericentricrearrangements, but at an early stage, subsequent to the differentiation of theNephrurus-Phyllurus line, the Carphodactylini had completely transformed the karyo-type to an all-biarmed configuration. The fact that both Oedura, a diplodactylin inKluge’s (1967b) scheme, and the pygopods could also be characterised by thissupposed derived karyotype was evidence of their monophyly to the exclusion of theDiplodactylini. Bauer (1990) reviewed Kluge’s Carphodactylini without consideringOedura, except to comment on his doubts regarding the identification of karyotypesynapomorphies.
In their major review of the phylogeny of lizard families, Estes et al. (1988) didnot discover support for the treatment of pygopods as diplodactylines. They notedseveral features, including aspects of brain case anatomy, which they suggestedplaced pygopods as the sister lineage to all geckoes, not just diplodactylines.
At present, despite considerable attention from systematic herpetologists, thereremain several unresolved phylogenetic issues relating to the gekkonoid lizards ofthe Australian region. Pygopods are firmly regarded as gekkonoids and there isevidence from morphology and karyology for a close relationship to diplodactylines.However, their precise relationship to the diplodactyline genera remains unknown,and the possibility remains, according to Estes et al. (1988), that they are the sisterlineage of all traditional gekkonids. Within the Diplodactylinae, the monophyly andcontent of the two tribes proposed by Kluge (1967b) require more rigorous testing.King’s arguments over the likely course of karyotypic evolution would also be testedif an independent estimate of phylogeny could be reliably established.
We used nucleotide sequence data from a mitochondrial gene, 12S rRNA, and anuclear oncogene, c-mos to address several current areas of uncertainty regardingthe phylogenetic relationships among major lineages of the gekkonoid lizards (thetraditional families Gekkonidae and Pygopodidae) in the Australian region. Ourchoice of genes was influenced by the need to sample multiple genes to avoidpotential gene tree/species tree problems due either to lineage sorting or theinadvertent sampling of paralogous sequences. Sampling more than one mito-chondrial gene, while potentially improving the support for the mitochondrial genetree, will do nothing to improve our estimate of the species tree. Saint et al. (1998)have recently demonstrated the suitability of the c-mos gene for the reconstructionof reptile evolutionary relationships. Our choice of taxa for study was aimed at

S. C. DONNELLAN ET AL.100
T 1. Amplification and sequencing primers designed in thepresent study. L=light strand, H=heavy strand
Primer Sequence
L765 5′-CATGGAGCAGGCATCAGGCAC-3′H1219 5′-ATCGATTACAGAACAGGCTCCTCTA-3′L669 5′-ANAGGTTTGGTCCTRRCCTT-3′H1160 5′-TTAAGCTTTGCTCGTAGTTCYC-3′H1182 5′-GCACCGCCAAGTCCTTAGAGTTTT-3′
specifically addressing three questions: (1) What are the relationships of pygopodsto other gekkonoids? (2) Are the two diplodactyline tribes, Carphodactylini andDiplodactylini, reciprocally monophyletic? (3) Is Oedura a diplodactylin (sensu Kluge)or carphodactylin (sensu King) genus?
MATERIAL AND METHODS
DNA sources
To clarify communication, the adjectival form of Diplodactylinae used in thispaper is ‘diplodactyline’, while those of the Diplodactylini and Carphodactylini are‘diplodactylin’ and ‘carphodactylin’. Pygopodidae (traditional sense) or Pygopodinaeare referred to throughout as ‘pygopods’. Numbers after each taxon are museumvoucher numbers with the following acronyms: SAMA – South Australian Museum,Adelaide; CM – Craig Moritz collection, University of Queensland; AMS – AustralianMuseum, Sydney; and QM – Queensland Museum, Brisbane.
Two pygopods (Delma fraseri SAMA R32463 and Lialis burtonis SAMA R29312)were selected to represent relatively divergent members of this clade (Kluge,1976; Hutchinson, 1997). We included undoubted representatives of the AustralianDiplodactylini (Diplodactylus vittatus SAMA R37969, Rhynchoedura ornata SAMA R36873and Strophurus intermedius SAMA R28963) and the Australian Carphodactylini (Car-phodactylus laevis QM J8944, Nephrurus milii SAMA R38006 and Pseudothecadactyluslindneri AMS R90194), as well as a New Caledonian carphodactylin (Rhacodactylusleachianus AMS R118099), and a member of the genus Oedura (O. tryoni SAMAR33584) whose affinities with either the Diplodactylini or Carphodactylini havebeen disputed. As outgroups to these Australian taxa we used two unrelatedgekkonines (Gehyra variegata SAMA R37803 and Phyllodactylus marmoratus SAMAR42098) and a eublepharine (Eublepharis macularius CM200). The scincid Tiliquascincoides SAMA R30280 was used as an outgroup to root the phylogenetic trees.
PCR methods
Genomic DNA was extracted from frozen liver by the standard phenol-chloroformmethod (Sambrook et al., 1989). The gekkonoid 12S rRNA gene was amplified andsequenced in two overlapping fragments with the following primer pairs L1091/H1478 (Kocher et al., 1989) and either L669/H1160, L669/H1182 or L765/H1219(Table 1). Partial c-mos sequences were obtained with primers G73/G74 or G73/

EVOLUTIONARY RELATIONSHIPS OF AUSTRALIAN GEKKONOID LIZARDS 101
G78 (Saint et al., 1998). The polymerase chain reaction (PCR) conditions, templatepurification and sequencing are described in detail in Saint et al. (1998).
To test whether the mitochondrial DNA (mtDNA) sequences were mitochondrialin origin and not nuclear paralogues (Zhang & Hewitt, 1996), we carried out thefollowing procedure. MtDNA was enriched from frozen liver by a plasmid DNAisolation method modified from Welter et al., (1989), resulting in a solution ofmtDNA that still contains some nuclear DNA. Serial dilutions (e.g. neat to 10−6) ofthe enriched mtDNA were amplified with three sets of PCR primers: Ald1/Ald2specific for the nuclear locus aldolase (Lessa & Applebaum, 1993), L1091/H1478specific for 12S rRNA (Kocher et al., 1989), and the mitochondrial primers beingtested. For each primer pair, we determined the maximum dilution (the endpoint)that produced successful amplification. In practice, it was usual to observe a thousand-fold difference between the endpoints for the 12S rRNA primer pair and the aldolaseprimer pair.
Either of two outcomes were observed for the mitochondrial test primer pair. Ifthe endpoint for the test primers was the same endpoint as the aldolase primer pair,we concluded that a nuclear paralogue could have been amplified. In this case, newprimers were designed from gekkonoid sequences and tested for their ability toamplify mtDNA only. If the mitochondrial test primers and 12S rRNA primersamplified to a similar endpoint that was more dilute than for the nuclear primers,we concluded that the product was derived from mtDNA. The test primers may,however, be able to amplify a nuclear paralogue as well as the true mitochondrialproduct when used with total cellular DNA. To test this, we compared the sequenceof the product amplified from the maximum dilution of the enriched mtDNA withthat from total cellular DNA from the same individual. If these sequences were thesame, we concluded that the primers only amplified mtDNA. If the sequences weredifferent however, the primers may also have amplified nuclear DNA. If this occurredwe designed new primer pairs and tested their ability to amplify only mtDNA bythe procedure described here.
Analysis of sequence data
The nuclear c-mos sequences were aligned by eye. We used the secondary structuremodel of Springer and Douzery (1996) to align the mitochondrial 12S rRNAsequences, using their nomenclature for structural features (Appendix). We did thealignments by eye optimizing the match between our sequences and conservedstructural elements identified in the secondary structure model. For instance, possiblestems were invariably present in the reptile sequences, as Hickson et al. (1996) haveshown for the domain III section of this molecule. For the ribosomal RNA sequence,regions of uncertain alignment were deleted from the analyses (Appendix). The 12SrRNA and c-mos data sets were tested for their combinability using a test of characterincongruence, the ILD test of Farris et al. (1994). This test was implemented inversion 4.0d59 of PAUP∗, written by D. L. Swofford, in which it is called thePartition Homogeneity Test. Cunningham (1997) has shown that the ILD testperforms well in predicting the accuracy of combined data, in comparison with theTempleton (1983) test, another test of character incongruence. Phylogenetic tree-building algorithms based on both a parsimony analysis (MP) and a maximumlikelihood (ML) analysis were implemented in test version 4.0d59 of PAUP∗.

S. C. DONNELLAN ET AL.102
T 2. (A) Alternative phylogenies tested in (B). These trees were those found by MP that containedthe appropriate constraint. (B) Wilcoxon matched-pairs signed ranks tests of the null hypothesis thatthe alternative phylogenies fit the data as well as the MP phylogeny for the combined 12S rRNA andc-mos data. P∗ values for each tree found with the constraint compared with each of the two MP trees
found. Asterisked values in table indicate significant difference at P<0.05
(A)[1] (1,(2,(3,(4,((5,((6,7),8)),((9,10),((11,12),(13,14))))))))(1,(2,(3,(4,(((5,(6,7)),8),((9,10),((11,12),(13,14))))))))[2] (1,(2,((3,((4,((5,(7,8)),(((11,12),13),14))),6)),(9,10))))[3] (1,(2,(((3,((5,(7,8)),6)),4),(((9,10),14),((11,12),13)))))Key to taxon numbers: 1, Nephrurus milii; 2, Carphodactylus laevis; 3, Rhacodactylus leachianus; 4, Pseudothecadactylus lindneri;5, Diplodactylus vittatus; 6, Oedura tryoni; 7, Strophurus intermedius; 8, Rhynchoedura ornata; 9, Delma fraseri; 10, Lialis burtonis;11, Phyllodactylus marmoratus; 12, Gehyra variegata; 13, Eublepharis macularius; 14, Tiliqua scincoides.
(B)Hypothesis Z-value P∗(1) Reciprocal monophyly of the Diplodactylini and 0.7505, 0.7715 0.2265, 0.2202Carphodactylini 0.8190, 0.8454 0.2064, 0.1990(2) Oedura as a carphodactylin 2.0809, 1.9465 0.0187∗, 0.0258∗(3) Pygopods as the sister group of all Gekkonidae 2.5027, 2.2453 0.0062∗, 0.0124∗
Transition/transversion (ts/tv) ratios (1.5 for 12S rRNA and 2.5 for c-mos) wereestimated for each of the two data sets via ML (test version 4.0d59 of PAUP∗).These values were then used to down-weight the effects of transitions compared totransversions. Branch and bound searches with the furthest addition sequence optionwere used to find the most parsimonious tree(s). In the ML analysis, ts/tv ratio wasestimated for combined data sets (ts/tv ratio=1.696). The Hasegawa–Kishino–Yano(1985) two parameter model for unequal base frequencies was implemented withempirical base frequencies (A=0.31476, C=0.23839, G=0.21743, T=0.22943)and equal evolutionary rates assumed for all sites (Hasegawa et al., 1985). WeightedMP and ML analyses were tested for their robustness using bootstrapping (2000pseudoreplicates for MP and 200 pseudoreplicates for ML). Bremer decay indices(Bremer, 1988) for the MP analysis were estimated with the Autodecay program(Eriksson, 1997) and test version 4.0d59 of PAUP∗. To compare the weighted MPhypotheses with alternative phylogenetic hypotheses, weighted MP searches wereperformed on trees constrained to the alternative hypotheses. A statistical test ofwhether the weighted MP tree was significantly shorter than the MP trees foundwith the constraint was determined using the Templeton (1983) test. We used aone-tailed test rather than a two-tailed version as we tested whether the MP treewas significantly shorter than the constrained tree which a priori cannot be shorterthan the MP tree, as we used a branch and bound search to find MP trees. Theconstraint trees to the alternative hypotheses were 1, reciprocal monophyly ofthe Diplodactylini and Carphodactylini ((((5,6,10,9),(1,8,4,11)),(2,3)),7,(12,13),14); 2,Oedura as a carphodactylin ((1,8,11,5,2,3,4),(6,9,10),7,12,13,14); 3, pygopods as thesister group of all Gekkonidae ((5,6,10,9,1,8,4,11,7,12,13),(2,3),14); where numbersrefer to taxa as in Table 2.
RESULTS
Initial amplifications of the 12S rRNA gene with primers L669/H1182 in Oeduraand with primers L669/H1160 in Rhynchoedura gave an unexpectedly shorter length

EVOLUTIONARY RELATIONSHIPS OF AUSTRALIAN GEKKONOID LIZARDS 103
product. The sequence of the Oedura product had a substantial deletion relative tothe sequences of the other taxa whereas the sequence of Rhynchoedura could not bealigned with the other gekkonoid sequences. Titration of enriched mtDNA fromthese taxa with the above primer sets produced a product only at the same dilutionsas a nuclear gene primer pair, whereas the primer pair L1091/H1478 produced aproduct at 1000-fold greater dilution. The suspect products in Oedura and Rhynchoedurawere unlikely to be mitochondrial in origin. We therefore designed another primerpair, L765/H1219, based on our gekkonoid 12S rRNA sequences, which in titrationexperiments amplified products of expected size at dilutions well beyond that atwhich the nuclear primer set ceased to amplify.
The final alignment length of 736 base pairs (bp) was available for the 12S rRNAdata. Because of uncertain sequence alignment, 180 characters were excluded fromthe analysis (Appendix), leaving 179 parsimony-informative sites (287 invariant and90 uninformative). The final c-mos data set was 374 bp in length. The first nucleotideposition in our alignment is a first base position in codon 92 in the human sequence(Watson et al., 1982). Although the c-mos sequence alignment was unambiguous, thealignment required a 12 bp (four codon) putative deletion at position 189 in Oeduratryoni. The number of informative sites in the c-mos data set was 58 (239 invariantand 77 uninformative).
A partition homogeneity test for combinability of the final 12S rRNA and c-mosdata sets, using weighted parsimony parameters, gave P=0.31, indicating that thereis no evidence of incongruence among the two data sets than might be expected bychance alone.
Parsimony analysis yielded two trees (Fig. 1A) with both the unweighted data(tree length 922 steps) and with the data weighted using the combined ts/tv ratio.The two trees differed only in the placement of Oedura relative to Rhynchoedura andStrophurus, i.e. (Oedura(Rhynchoedura, Strophurus) versus (Rhynchoedura(Oedura, Strophurus).Only one of these trees is presented in Figure 1. Major features of the trees include:(1) the presence of Kluge’s Diplodactylini, i.e. Diplodactylus, Rhynchoedura, Strophurusand Oedura, as a monophyletic unit; (2) the paraphyly of the Carphodactylini withrespect to the Diplodactylini; (3) the pygopods as the sister-group of all theDiplodactylinae; and (4) the pygopods plus the Diplodactylinae forming a mon-ophyletic group. Figure 1B shows the results of the ML analysis, which differedfrom both the MP trees in reversing the order of the outgroups, Eublepharis branchingbefore the gekkonines. Relationships among the remaining taxa were the same asone of the MP trees. The same four major findings of the MP analysis are evidentin the ML tree.
Bootstrapping (Fig. 1) showed strong support for those key nodes noted above,but others, although present in both the MP and ML analyses, were less stronglysupported (appeared in fewer than 50% of the bootstrap pseudoreplicates). Ingeneral, Bremer decay indices were high ([6) when bootstrap proportions providedstrong support (Fig. 1A). Monophyly of the traditional subfamily Diplodactylinaewas supported, although at 56% bootstrap support this is the least robust of the‘supported’ nodes. Within the Diplodactylinae, relationships among the four generawere not well supported in the bootstrap pseudoreplicates, so that resolution ofrelationships within this clade requires further work. A monophyletic lineage leadingto Carphodactylus and Nephrurus was always recovered, and was always the basalbranch of the Diplodactylinae. However the branching order of the other twocarphodactylins, Rhacodactylus and Pseudothecadactylus, and their position relative to

S. C. DONNELLAN ET AL.104
Eublepharis macularius
Gehyra variegata
Phyllodactylus marmoratus
100
83/9913
Carphodactylini
Nephrurus milii
Carphodactylus laevis
Rhacodactylus leachianus
Pseudothecadactylus lindneri
Diplodactylus vittatus
Oedura tryoni
Strophurus intermedius
Rhynchoedura ornata
Delma fraseri
Lialis burtonis
Phyllodactylus marmoratus
Gehyra variegata
Eublepharis macularius
Tiliqua scincoides
Diplodactylini
pygopods
Gekkoninae
Eublepharinae
Carphodactylini
Nephrurus milii
Carphodactylus laevis
Rhacodactylus leachianus
Pseudothecadactylus lindneri
Diplodactylus vittatus
Oedura tryoni
Strophurus intermedius
Rhynchoedura ornata
Delma fraseri
Lialis burtonis
Tiliqua scincoides
Diplodactylini
pygopods
Gekkoninae
Eublepharinae
56
–/622 57
–/892 52
1
100
100/9212
100
95/9611
94
70/848
1
–/700
92
–/1006
–/642
100
96/98
50
–/86
96
78/8698
78/98
–/57
–/68
50/–100
100/89
98
–/100
A
B
Figure 1. MP (A) and ML (B) phylogenetic trees of relationships among Australasian gekkonoids basedon combined 12S rRNA and c-mos sequences. Bootstrap proportions from 2000 pseudoreplicates (MP)and 200 pseudoreplicates (ML) are indicated above the relevant branches. Bootstrap proportions from2000 pseudoreplicates (MP) and 200 pseudoreplicates (ML) for the individual 12S rRNA and c-mosdatasets analysed separately are indicated below the relevant branch, with the values for 12S rRNAfirst – indicates a bootstrap proportion of <50. Bremer decay indices are also indicated below therelevant branch.

EVOLUTIONARY RELATIONSHIPS OF AUSTRALIAN GEKKONOID LIZARDS 105
the Diplodactylini, are inconsistent. Our data strongly support (100% bootstrap)pygopods as sharing a common ancestor with all diplodactylines. Although diplo-dactyline monophyly is less strongly supported, our best trees support Kluge’s (1987)idea of pygopods being the sister group of the diplodactylines. Monophyly of thegekkonines was not seriously in question and Gehyra and Phyllodactylus do form awell-supported clade. The nodes determining the relative branching order of gek-konines and the eublepharine at the base of the tree have low bootstrap values andthis uncertainty is further reflected in the disagreement between the ML and MPtrees.
Alternative hypotheses of relationships, as suggested by different workers for sometaxa, were developed as constraints and MP trees were found that contained theindividual constrained relationships (Table 2A). These constrained trees inevitablyare longer than the two equally short MP trees recovered using the unconstrainedanalysis, and we used the Templeton (1983) test to determine whether this increasedlength represented a significantly worse fit of the data (Table 2B). Our best treesuggested that the Carphodactylini was paraphyletic, with at least three separatelineages (Carphodactylus + Nephrurus, Pseudothecadactylus and Rhacodactylus) but thealternative of a monophyletic Carphodactylini as the sister lineage of a monophyleticDiplodactylini could not be rejected. However, our data did not support transferringOedura to a closer relationship with any of the carphodactylin genera, or forcingpygopods to be the sister lineage of all gekkonids (the traditional view, supportedby Estes et al., 1988).
DISCUSSION
Molecular evolution
After adjustment for the sites excluded because of uncertain alignment, 12S rRNAstill provided twice as many parsimony-informative sites (32%) per equivalent lengthof sequence as the c-mos gene did (16%). However, on the criterion of relativebootstrap support, the c-mos gene appears to be more useful at all levels of divergencein the gekkonoid phylogeny, as bootstrap support from this gene alone was generallyequal to or better then that observed when the 12S rRNA data were analysed alone(Fig. 1A). In the present study, only the central third of the c-mos gene was sequenced.Judging from the level of sequence variability in alignments of the few c-mos genesequences presently available in GenBank, the remainder of the gene is likely toprovide a higher proportion of informative sites; however, its use is limited presentlyto taxa relatively closely related to those with representative sequences in GenBank.
Carphodactylini and the position of Oedura
Our sequence data unequivocally place Oedura among the Diplodactylini, sup-porting Kluge’s (1967b) allocation and contradicting King’s (1987a) interpretationof the karyotypic evidence. The arguments regarding Oedura as a carphodactylin ordiplodactylin have always tacitly assumed that these tribes were well-founded andreciprocally monophyletic. The case for a monophyletic Diplodactylini, with Oeduraas a member, is very strong, based on our sequence data which show very strongbootstrap support for the Diplodactylus + Oedura + Rhynchoedura + Strophurus clade,

S. C. DONNELLAN ET AL.106
and on the several morphological synapomorphies noted below. However, thejustification for continuing to recognize the Carphodactylini is more dubious.
The question of whether Carphodactylini is a clade or a grade has not beenseriously considered, even though this tribe had been erected by Kluge prior to theperiod where Kluge’s own approach to phylogenetic analysis had become explicitlycladistic. Kluge (1967b) diagnosed the Carphodactylini as being those diplodactylineswhich have numerous preanal pores grouped into a large irregular patch. Thispeculiar pore arrangement is arguably apomorphic, but is not present in allcarphodactylin genera. A single row of pores is present in Bavayia sauvageii, whilepores are absent in Phyllurus s.l. and Nephrurus. Diplodactylini were defined as havingthe plesiomorphic single preanal pore row, plus several synapomorphies: almostcomplete fusion of the premaxillae; maxilla broadly separated from frontal byprefrontal; absence or moderate development of the lateral and medial infraorbitalprocesses of the prefrontal; great reduction or absence of the jugal; reduction of theclavicle. Thus apart from the pore patch, carphodactylins were differentiated bypossessing the plesiomorphic states of the characters used to define the Diplodactylini.Bauer (1990) continued to use Kluge’s definition to justify recognition of the tribe,while King (1987a, 1990) added his conviction that the 2n=38 biarmed karyotypewas additional diagnostic feature.
Our data fail to strongly corroborate a monophyletic Carphodactylini. Supportfor crucial branches at the base of the pygopod + diplodactyline radiation are notstrongly established, as indicated both by the modest bootstrap support for some ofthese branches and the fact that forcing carphodactylin monophyly produced onlya slightly longer trees whose difference from the shortest tree was not significant. Atbest, carphodactylin monophyly rests on shaky ground, with no morphologicalsynapomorphy present in all taxa and no strong support from the sequence data.On the other hand, our shortest tree (carphodactylin paraphyly), is in good agreementwith Russell’s (1979) findings on toe pad evolution, with the primitively padlessCarphodactylus and Nephrurus lying basal to subsequent lineages with toe pads, onelineage being the Diplodactylini.
Our data suggest that Kluge’s caution regarding the karyological data was well-founded. The distribution of lineages dominated by either all-acrocentric or all-biarmed karyotypes make it difficult to confidently regard either as plesiomorphicfor geckoes overall. Either convergences or reversals, or both, would be necessaryto explain the evolution of the observed karyotypes if the karyotypic data are overlainon our preferred tree topology. However, for the diplodactylines and pygopods,interpreting the karyotypic data against our shortest tree topology implies that theall biarmed 2n=38 karyotype is plesiomorphic—i.e. the evolutionary polarity is thereverse of that favoured by King.
The relationships of pygopods
The gekkonoid nature of pygopods is no longer doubted (McDowell & Bogert,1954; Underwood, 1957; Kluge, 1974, 1987; Estes et al., 1988). However, the long-assumed position of pygopods as the sister lineage to all gekkonoids has only recentlybeen questioned (Kluge, 1987; King & Mengden, 1990; but see Estes et al., 1988).Our sequence data strongly support the general conclusion of both Kluge (1987)and King (1990) that diplodactylines and pygopods are each others’ closest relatives.

EVOLUTIONARY RELATIONSHIPS OF AUSTRALIAN GEKKONOID LIZARDS 107
Our best trees support Kluge’s conclusion, placing pygopods as the sister lineage ofall diplodactylines. King’s suggestion, that pygopods are probably nested within theCarphodactylini, is less favoured by our data, partly because our shortest topologyfailed to recover a monophyletic Carphodactylini. If carphodactylins are a para-phyletic assemblage, then the idea of pygopods being carphodactylins is meaningless.In any case, pygopods share their most recent ancestry with diplodactylines only,and if gekkonoids are monophyletic, they must be considered as nested within theGekkonidae.
Given this close relationship it is worth considering why it has taken so long torecognize it. Undoubtedly the reason lies with the anatomical gulf between typicalgeckoes and pygopods. In other lizard families, degenerate limbed and serpentinelineages have evolved several times without stimulating recognition of distinctfamilies, but the transitional forms are often present (e.g. Scincidae) or the bodyplan of the quadrupedal members of the family is very similar to that of the limblessmembers (e.g. Anguidae). In the Gekkota, the limbed and limbless taxa are highlydivergent from one another and from other lizards. The Gekkonidae have tra-ditionally stood out from other lizards, including pygopods, in major anatomicalfeatures. Chief among these is the nature of the vertebrae, persistently notochordal(amphicoelous) in nearly all geckoes, rather than procoelous as in all other livingsquamates. Kluge (1967a) supported Underwood’s (1957) conclusion that amphicoelywas a reversal within gekkonids, suggesting neoteny as the mechanism responsible, hisinterpretation reinforced by the presence of procoely in the primitive Eublepharinae.Debate regarding this feature’s interpretation has continued (reviewed by Bauer,1990), but neotenic amphicoely dovetails with other major aspects of gekkonidanatomy that suggest that heterochrony has been significant in gekkonoid evolution(Stephenson, 1960; Stephenson & Stephenson, 1956; Moffat, 1973). Neoteny hasbeen recognized in the poorly ossified skull (especially the parietal) and feebledevelopment of dermal scalation in most species. Pygopods, by contrast, have well-developed procoelous vertebrae, better-ossified skulls and large imbricate body scales,including enlarged ventrals and cephalic shields.
The phylogenetic position of pygopods suggested by Kluge (1987), King (1990)and by the present study, that is within traditional gekkonids, argues that pygopodsare legless geckoes, and should not be isolated in a separate family. The distinctivenessof pygopods stems from the lack of transitional forms and, we would suggest, a reversalof the typically gekkonoid paedomorphic developmental pattern. As suggestedby Rieppel (1984) and Bauer (1990), perhaps miniaturisation and developmentalacceleration have re-activated terminal steps in ossification and dermal developmentthat had been switched off early in gekkonoid evolution.
Sequence availability
All sequence data have been deposited in GenBank (accession numbers AF039462,AF039567–71, AF090174–87, AF090844–51).
ACKNOWLEDGEMENTS
We thank J. Birrell for the pilot work on 12S rRNA in lizards, M. Gardner forthe Tiliqua 12S rRNA sequence and P. Monis who designed some of the PCR primers.

S. C. DONNELLAN ET AL.108
M. King, K. McDonald, C. Moritz, and R. Sadlier supplied tissue samples. Earlierdrafts of the manuscript benefited from critical comments provided by S. Cooper,S. Keogh and L. Rawlings. This work was supported by an Australian ResearchCouncil Grant (A19602789) to MNH and SCD.
REFERENCES
Bauer AM. 1990. Phylogenetic systematics and biogeography of the Carphodactylini (Reptilia:Gekkonidae). Bonner Zoologische Monographien 30: 1–218.
Bremer K. 1988. The limits of amino acid sequence data in angiosperm phylogenetic reconstruction.Evolution 42: 795–803.
Cunningham C. 1997. Can three incongruence tests predict when data should be combined? MolecularBiology Evolution 14: 733–740.
Eriksson T. 1997. Autodecay version 2.9.10 (Hypercard stack distributed by the author). BotaniskaInstitutionen, Stockholm University. Stockholm.
Estes R. 1983. Sauria terrestria, Amphisbaenia. Handbuch der Palaoherpetologie 10A: 1–249.Estes R, De Queiroz K, Gauthier J. 1988. Phylogenetic relationships within Squamata. In: Estes
R, Pregill G, eds. Phylogenetic relationships of the lizard families: Essays commemorating Charles L Camp.Stanford: Stanford University Press, 119–281.
Farris JS, Kallersjo M, Kluge AG, Bult C. 1994. Testing significance of incongruence. Cladistics10: 315–319.
Hasegawa M, Kishino H, Yano T. 1985. Dating of the human-ape splitting by a molecular clockof mitochondrial DNA. Journal of Molecular Evolution 22: 160–174.
Hickson RE, Simon C, Cooper A, Spicer GS, Sullivan J, Penny D. 1996. Conserved sequencemotifs, alignment, and secondary structure for the third domain of animal 12S rRNA. MolecularBiology Evolution 13: 150–169.
Holder LA. 1960. The comparative morphology of the axial skeleton in the Australian Gekkonidae.Zoological Journal of the Linnean Society 44: 300–335.
Huelsenbeck JP, Bull JJ, Cunningham CW. 1996. Combining data in phylogenetic analysis. Trendsin Evolution and Ecology 11: 152–157.
Hutchinson MN. 1997. The first fossil pygopod (Squamata: Gekkota), and a review of mandibularvariation in living species. Memoirs of the Queensland Museum 41: 355–366.
King M. 1985. Chromosome markers and their use in phylogeny and systematics. In: Grigg G, ShineR, Ehmann H, eds. Biology of Australasian frogs and reptiles. Sydney: Surrey Beatty & Sons with theRoyal Zoological Society of New South Wales, 165–175.
King M. 1987a. Chromosomal evolution in the Diplodactylinae (Gekkonidae: Reptilia) I. Evolutionaryrelationships and patterns of change. Australian Journal of Zoology 35: 507–531.
King M. 1987b. Monophyleticism and polyphyleticism in the Gekkonidae: a chromosomal perspective.Australian Journal of Zoology 35: 641–654.
King M. 1990. Chromosomal and immunogenetic data: a new perspective on the origin of Australia’sreptiles. In: Olmo E, ed. Cytogenetics of amphibians and reptiles, Basle: Birkhauser Verlag, 153–180.
King M, Mengden G. 1990. Chromosomal evolution in the Diplodactylinae (Gekkonidae: Reptilia)II. Chromosomal variability between New Caledonian species. Australian Journal of Zoology 38:219–226.
Kluge AG. 1967a. Higher taxonomic categories of gekkonid lizards and their evolution. Bulletin of theAmerican Museum of Natural History 135: 1–60.
Kluge AG. 1967b. Systematics, phylogeny and zoogeography of the lizard genus Diplodactylus Gray(Gekkonidae). Australian Journal of Zoology 15: 1007–1108.
Kluge AG. 1974. A taxonomic revision of the lizard family Pygopodidae. Miscellaneous Publications ofthe Museum of Zoology, University of Michigan 147: 1–221.
Kluge AG. 1976. Phylogenetic relationships in the lizard family Pygopodidae: an evaluation of theory,methods and data. Miscellaneous Publications of the Museum of Zoology, University of Michigan 152: 1–72.
Kluge AG. 1987. Cladistic relationships in the Gekkonoidea (Squamata, Sauria). Miscellaneous Publicationsof the Museum of Zoology, University of Michigan 173: 1–54.
Kocher TD, Thomas WK, Meyer A, Edwards SV, Paabo SF, Villablanca FX, Wilson AC.1989. Dynamics of mt DNA evolution in animals: Amplification and sequencing with conservedprimers. Proceeding of the National Academy Science USA 86: 6196–6200.

EVOLUTIONARY RELATIONSHIPS OF AUSTRALIAN GEKKONOID LIZARDS 109
Lessa E, Applebaum G. 1993. Screening techniques for detecting allelic variation in DNA sequences.Molecular Ecology 2: 121–129.
Macey JR, Larson A, Ananjeva NB, Fang ZL, Papenfuss TJ. 1997. Two novel gene orders andthe role of light-strand replication in rearrangement of the vertebrate mitochondrial genome.Molecular Biology Evolution 14: 91–104.
McDowell SB, Bogert CM. 1954. The systematic position of Lanthanotus and the affinities of theanguimorphan lizards. Bulletin of the American Museum of Natural History 105: 1–142.
Moffat LA. 1973. The concept of primitiveness and its bearing on the phylogenetic classification ofthe Gekkota. Proceedings of the Linnean Society of New South Wales 97: 275–301.
Presch W. 1988. Cladistic relationships within the Scincomorpha. In: Estes R, Pregill G, eds. Phylogeneticrelationships of the lizard families: Essays commemorating Charles L Camp. Stanford: Stanford UniversityPress, 471–492.
Rieppel O. 1984. Miniaturisation of the lizard skull: its functional and evolutionary implications. In:Ferguson MWJ, ed. The structure, development and evolution of reptiles: Symposium No 52 of the ZoologicalSociety of London. London: Academic Press, 503–520.
Russell AP. 1979. Parallelism and integrated design in the foot structure of gekkonine and diplo-dactyline geckos. Copeia 1979: 1–21.
Saint KM, Austin CC, Donnellan SC, Hutchinson MN. 1998. C-mos, a nuclear marker forsquamate phylogenetic analysis. Molecular Phylogenetics and Evolution 10: 259–263.
Sambrook J, Fritsch EF, Maniatis T. 1989. Molecular Cloning. A Laboratory Manual. Cold SpringHarbour: Cold Spring Harbour Laboratory Press.
Schwenk K. 1988. Comparative morphology of the lepidosaur tongue and its relevance to squamatephylogeny. In: Estes R, Pregill G, eds. Phylogenetic relationships of the lizard families: Essays commemoratingCharles L Camp. Stanford: Stanford University Press, 571–598.
Springer M, Douzery E. 1996. Secondary structure and patterns of evolution among mammalianmitochondrial 12S rRNA molecules. Journal of Molecular Evolution 43: 357–373.
Stephenson NG. 1960. The comparative osteology of Australian geckoes and its bearing on theirmorphological status. Zoological Journal of the Linnean Society 44: 278–299.
Stephenson NG, Stephenson EM. 1956. The osteology of the New Zealand geckos and its bearingon their morphological status. Transactions of the Royal Society of New Zealand 84: 341–358.
Templeton AR. 1983. Phylogenetic inference from restriction endonuclease cleavage site maps withparticular reference to the humans and apes. Evolution 37: 221–244.
Underwood G. 1954. On the classification and evolution of geckos. Proceedings of the Zoological Societyof London 124: 469–492.
Underwood G. 1957. On the lizards of the family Pygopodidae, a contribution to the morphologyand phylogeny of the Squamata. Journal of Morphology 100: 207–268.
Watson R, Oskarson M, Vande Woude GF. 1982. Human DNA sequence homologous to thetransforming gene (mos) of Moloney murine sarcoma virus. Proceedings of the National Academy ofSciences, USA 79: 4078–4082.
Welter C, Dooley S, Blin N. 1989. A rapid protocol for the purification of mitochondrial DNAsuitable for studying restriction fragment length polymorphisms. Gene 83: 169–172.
Zhang D-X, Hewitt GM. 1996. Nuclear integrations: challenges for mitochondrial DNA markers.Trends in Evolution and Ecology 11: 247–251.

S. C. DONNELLAN ET AL.110
APPENDIX
Alignment of (a) 12S rRNA and (b) c-mos sequences of Australasian gekkonoids. –=gap inserted toimprovement alignment, ·=same as first sequence. Sites marked by asterisks were not included in thefinal analysis. Numbers above sequence alignment refer to secondary structural features (Springer &Douzery, 1996)
9′ 6′ 11 11′ 5′12S 1–46 **** **N. milii AAAAC ACC TAGCCTA--GC CAC ACC CC CACG GG --ACCCA GCAGTAC. laevis ..... ... ....TA.--.. ... ... .. .... .. --T.T.. ......R. leachianus ..... ... ....TGT--.. ... ..- .. TCAC .. G-..... ......P. lindneri ..G.. G.. ....G..--.. ... ... .. .... .. --.T... ......D. vittatus ..G.. G.. .....C.--.. ... ... .. .... .. --..... ......O. tryoni C.... ... ......T--.. ... ... .. .... .. --G.T.. ......S. intermedius ..... ... ......C--.. ... ... .. .... .. --.TT.. ......R. ornata ..G.. G.. .......--.. ... ..- .. ACAC .. G-..... ......D. fraseri ..G.. G.. .....A.AG.. ... ... .. .... .. --..... ......L. burtonis ..G.. G.. ......C--.. ... C.. .. .... .. --..... ......P. marmoratus ..... ... .C...A.T-.. ... ... .. ...- .. -A..... .....GG. variegata .C... G.. .....C.T-.. ... ... .. ...- .. -A..T.. .....GE. macularius .T... G.. .C..T-.T-.. ... .T. .. ...- .. -A..T.. .....GT. scincoides .C... ... .T..TAC--.. ... ..- .. ACAC .. GT..T.. .....G
12 12′12S 47–98 *******************N. milii GTAAACA TTAAGCAATGAGC GCCTT----------TTTTCAA GCTCGACTTAC. laevis ..T..T. .........A... ...CCA----AAACCCC..... ..........R. leachianus A..G... ............. ...CACTAATAACTT.A.C... ..........P. lindneri A.T.... ............A ...C.A-----TTAA...CA.. ..........D. vittatus A.T..T. .........A... .A.CAC-----------CC... ...T......O. tryoni A.T.... .........A... .A.CCC-----A------.... ..........S. intermedius A.T.... .........A... .ATACT-----ACTC--..T.. ...T......R. ornata A.T.... .........A... A..AAC-----ACCCC.A..-. ..........D. fraseri A...... .........A... .A.AAC------------CA.. ...T......L. burtonis A...... ............. .....C----------CCC.-. ..........P. marmoratus A.T.... ...G..C..A... CAAAGC------------.TG. C..A.T..C.G. variegata A.TG..C ..G...C....TT TA.A-----------------. C.......C.E. macularius A....TC .........A... ..TC.C----------C.C.-. ...T......T. scincoides AAT.... .....A...A... .------------------A.. ...T......
4′ 13 A 14 B A′12S 99–141 ******N. milii G TTACGATAA A---CC GGGTC GGT AAATT TCGTG CCA GC C ACCC. laevis . ...T..... .---TA ...C. ... ..... ..... ... .. . ...R. leachianus . ...T...TC .A--.A A..C. ... ..... ..... ... .. . ...P. lindneri . ...TAG..T G---.A ...C. ... ..... ..... ... .. . ...D. vittatus . ...T.G.TT .---.T ...C. ... ..... ..... ... .. . ...O. tryoni . ...T...TT C---.. A.... ... T.... ..... ... .. . ...S. intermedius . ...TAG..C C---.. C.... ... ..... ..... ... .. . ...R. ornata A ...T.G..T TA--A. A..C. ... ...C. ..... ... .. . ...D. fraseri . C..T..... .CAT.A ...CT ... ..... ..... ... .. . ...L. burtonis . ...T...GT .----A ...CT ... ..... ..... ... .. . ...P. marmoratus . .C.AAC... .C--AA ...C. ... ...C. C.... ... .. . ...G. variegata . C...AGCGC .ACAAT A..C. ... ...C. C.... ... .. . ...E. macularius . ...T.G.G. T---TA AA.C. ... ...C. ..... ... .. . ...T. scincoides . C..T...T. .G--TT T..C. ... ..... ..... ... .. . ...

EVOLUTIONARY RELATIONSHIPS OF AUSTRALIAN GEKKONOID LIZARDS 111
B′ 14′ 13′ 3′ 15 C12S 142–186N. milii GC GGTTA TACGA AA GACCC AT AATAATAGA ACCCC GGC GT AAAACC. laevis .. ..... ..... .. AG... .A G......AT .TA.- ... .. ...G.R. leachianus .. ..... ..... .. ..... .A ..C.....T .TA.- ... .. .....P. lindneri .. ..... ..... .. .G... .A .......AC C.T.- ... .. .....D. vittatus .. ..C.. ..... .. .G... .A G....CTA- .A..- ... AA ...G.O. tryoni .. ..... ..... .. ....T .A .......AT G...- ... AA .....S. intermedius .. ..... ..... .. ....A .A .......AT G...- ... AA .....R. ornata .. ..... ..... .G ....T TA .......A- ....- ... AC .....D. fraseri .. ..... C.... .. AG... .A .......CC CA..- ... .. ...G.L. burtonis .. ..... ..... .. AG... .A ......TT- TT..- ... .. ...G.P. marmoratus .. ..... C...C .G .G... .A G......A. CA.T. ... .. ....TG. variegata .. ..... ....G .G .G..A .A .G...C.CC CAA.- ... .. ...TTE. macularius .. ..... C.... CG .G.TT .A ......... .TT.- ... .. .....T. scincoides .. ..... ..... .. .G..A .A ..C...GAC C.T.- ... .. ...G.
16 17 18 1912S 187–233 **************** *****N. milii GTGA ?TAG AA CCACATCCTTTCCA-- AGAAACC CAAACAC-- CTGCCC. laevis .... C... .. .AC..AG...C.ACTT C...... ......... A....R. leachianus .... C... .G .T.A.AATA..AA... T..G... T....C... .G..TP. lindneri .... C... .G A....A....CAA... T....TT T....CT.. GAATTD. vittatus .... C... .G .ACACAA.CCA.A-.. T.CG..T A....CAT. GGA.TO. tryoni .... C... .G .ACACAAT...T..CG T..G..T A....CTTT TA.T.S. intermedius .... C... .G AT.T.AT..ACAA-.. T....TT T...TC... TAA..R. ornata .... C... .G .ACATATTAAC..... T..GGTA T....C... T.C..D. fraseri .... C..A .. AAGAG.TT.AAAA... GA..CTT T....C... .CT.TL. burtonis .... TC.. .. AATA.AAG.CCAT.AA T....TA .G...T... TAC.TP. marmoratus .... C... .G AATA..TTAAC.ATAA .....T. AT...TA.. .AATAG. variegata .... C... .G A.CACAAACAC.ATAA ...CG.T A.T..CTG. .C..AE. macularius ...G C... .G ATCTCA.A------.. T..G... GG..A.A.. AGA.AT. scincoides .... CC.. .. TA.TG..AAAAAA-.. .TTG.AG G...AC.A. AGC.A
20 20′ 19′ 2112S 234–279 **** ********************* **N. milii AA GT TATAAAAT AC ACCC GGCAC CCTTGAAGCCCACA------- --C. laevis .. .. ........ .. .T.. ..... .TC.....T..TACT...... ..R. leachianus .. .. .......C .. .AAA A.... G..G..CAA..CAC....... ..P. lindneri .. .. .......C .. T.A- A.T.. ATCA...AAT.TA........ ..D. vittatus C. .. .......C .. ..A- A.ACA .A..AG.A.TTC.-....... ..O. tryoni .. .. ........ .. .T.A .A... ATCAAG.ATA.C.TT...... ..S. intermedius .. .. .......C .. ...A ..ATA A.AG.G.AA.TC.TC...... ..R. ornata C. .. ........ .. .AA- ..TCT T.CG...A....--....... ..D. fraseri .. .. .......C .. .T.A A.AGA A.AG...A.T----....... ..L. burtonis .. .. .......C .. ..TA A.G.A G.AG...C.A..--....... ..P. marmoratus T. .. CG...... .. C.A- TAT.T GTAA..TCAA.CAT....... ..G. variegata C. .G CG.....A C. .AA- T..C. A.AG..CC...C..C...... ..E. macularius .. .A .G...... T. CAAA ACTTT TTAA....A...A-....... ..T. scincoides .G A. .G...... .T .T-- T.GCA .AAGA.CT..AC..ATAAAAT ..

S. C. DONNELLAN ET AL.112
21′ 18′ 17′ 16′12S 280–321 **** ** **************N. milii ---- -- CGTTTCA ACAAAACGAACCC- TT TTGAC TCAC GAAAC. laevis ---- CT G...... ....TCA.CC..-- .. .GAC. .... ....R. leachianus ---- CA AA.C... .TC..TATT..T-- C. .GAC. .... ....P. lindneri ---- CC AA..... ......GCC.T.T- C. .GA-. G... T...D. vittatus ---- CC A..CC.. TT....ATT.A.T- C. .GA-. .... ....O. tryoni ---- TA AA.C... .TG..CAATTAT.C C. .GAC. .... ....S. intermedius ---- AA AA..... TTT...ATT....- C. .GAT. .... ....R. ornata ---- TA AACC... TTT..CATT.TTT- C. .GA-. .... ....D. fraseri ---- CC AAA..TC ..CC..AA.CA.-- .. .GA-. .... ....L. burtonis ---- AC GA....C C..CT.A..C..-- .. .GA-. .... ....P. marmoratus ---- CA AA....T TTTGT.GA.TAA.- C. .GAC. .... ....G. variegata ---- CC A.CGC.T ...C..GCCC.A.- C. .GAC. C... A...E. macularius ---- TA G..C... ---C..AA------ C. .GA-. C... ....T. scincoides ---- GC .T.CAA. -T....GACTA--- .. CAAC. .... T...
22 23 23′ 22′12S 322–373N. milii ACCAAGAAACA AAC TGGG ATTAGATAC CCTA CTA TGCTTGGC CGTAACCCTAC. laevis ........... ... .... ......... .... ... .......T ..........R. leachianus ........... ... .... ......... .... ... ........ .....A..C.P. lindneri G......C... ... .... ......... .... ... ........ .A...A....D. vittatus ......G.... ... .... ......... .... ... .......T .A...A.T.TO. tryoni ........... ... .... ......... .... ... .......T .A...A...TS. intermedius ........... ... .... ......... .... T.. .......T .A...A...TR. ornata G......C... ... .... ......... .... ... .....A.. .A...A...TD. fraseri ..T........ ... .... ......... .... ... .....A.. .....A.A..L. burtonis ..T........ ... .... ......... .... ... .....A.T .A...A.A..P. marmoratus ..T....C... ... .... ......... .... ... .....AAT ...T.A..C.G. variegata G.T.G..C... ... .... ......... .... ... ...CAA.. ........A.E. macularius G.T........ ... .... ..C...... .... ... .....A.. .A...A....T. scincoides G.T.G...... ... .... ......... .... ... ...C.A.. .C...A.A..
24 C′ 24′ 15′ 2512S 374–422 ********** *****N. milii GACATTC CCCCCA---- AC TA--- -AATGTC C GCC AGAGAACTACA AGCC. laevis ...C--- T..AT.CT-- .. ACTAC ---GT.. . ... T.......... ..TR. leachianus ......- A.GTACC--- .. AC--- -...... . ... ....T...... ..TP. lindneri .G...C- AAA.A.T--- .. AC--- -C...C. . ... ....T...... ..TD. vittatus ....--- TA..------ .. ----- ---.... . ... ........... ..TO. tryoni ......- ...A.----- .. A---- --..... . ... ....C...... ..TS. intermedius .....-- ATTAA----- .. ----- --..... . ... ....T...... ..TR. ornata ......- .....----- .- ----- --..... T ... ....C...... ..TD. fraseri ..TT--- TTTTT.AGA- .. ACAAC ---.A.. . ... ......T...C ..TL. burtonis ..TC..- AATTAT---- .. A.--- -..AT.. . ... ....G.T...T ..TP. marmoratus .GT.G.- A..AT.---- .. C.--- -.C.AC. . ... ......T.... ..TG. variegata ..T.G-- A..T.CCCCC .. CCACA --C.A.. . ... ........... ...E. macularius .G..--- .AA.ATA--- .. ATGGA ---..C. . ... ........... ..TT. scincoides ..TGG.T AAA.AC---- .. A.--- A.CCA.T . ... ........... ...

EVOLUTIONARY RELATIONSHIPS OF AUSTRALIAN GEKKONOID LIZARDS 113
25′ 2′ 26 2712S 423–471N. milii GAAAA ACT CAAAACTCAAA AGA CT TGACGGTGCCCCA TATCAAC CTAGAC. laevis ..... ... .G......... G.. .. ............. C.C.... .....R. leachianus ..... ... AG......... G.. .. ........T.... C...... .....P. lindneri ..... ... T.......... G.. .. ........G.... C...... .....D. vittatus ..... ... T.......... ... .. ........T.... C...... .....O. tryoni ..... ... ........... G.. .. ........T.... C...... .....S. intermedius ..... ... T.......... G.. .. .....-..T.... C.C.... .....R. ornata ..... ... T.......... G.. .. ........T.... C.C..C. .....D. fraseri ..... ... T.......... G.. .. ........T.T.. ....... .....L. burtonis ..... ... T.......G.. G.. .. .....-.AT.T.. ....... .....P. marmoratus ..... G.. T.......... G.. .. ........TT... C.C.... .....G. variegata ACC.. G.C .G......... G.. .. ........T.... C.C.CC. .....E. macularius ...-- ... T--........ G.. .. .......T..... .....G. .....T. scincoides ..... G.. .G......... G.. .. ..G......T... C....T. .....
28 29 29′ 3012S 472–517N. milii GG AGCCTGTCCT ATA AACG ATGCCCCA CGAA AAACCTCA CCCCCCCC. laevis .. .......... ... .T.. ..A.T..C .... ........ .......R. leachianus .. .......... ... .T.. ..A..... .... ........ ....TT.P. lindneri .. .......... ... .T.. ..A..... .... .T...... ....TT.D. vittatus .. .......... ... .T.. ..A..... .... .T....TT ..TTTT.O. tryoni .. .......... ... .T.. ..A..... .... .C....T. ..T....S. intermedius .. .......... ... .T.. ..A..... .... .C....T. ..TTT..R. ornata .. .......... .C. .T.. ..A..... .... .C...... ..TTTT.D. fraseri .. .......... ..T .T.. ..A..... .... ........ .....ATL. burtonis .. .......... ... .T.. ..A.T... .... ........ ...T.TTP. marmoratus .. .......... ... .C.. ..A..... ...T .C...... ...TAA.G. variegata .. .......... .C. .T.. .CAA.... ...T .T...C.. .TT...TE. macularius .. .......... ... .T.. .C.AT... ...T ........ ...T...T. scincoides .. .......... ... .T.. ..A..... ..CT TT...C.. ..ATT.T
31 32 3312S 518–566 *****N. milii TAGTATAAC-TCA GCCTATATACCGCCGTCG-C CA GC TT ACCTT GCA-AC. laevis .C.CCACT.-C.. ..................-. .. .. C. ..... AT.-.R. leachianus .G.C.A.---A.. ..................-. AG .. C. ..... ATGG.P. lindneri ...C...---A.. .T................-. .. .. .. ..... ATG-.D. vittatus ...CC.CTAAC.. ..................-T .. .. C. ...C. A..-.O. tryoni ...C.CTT--... ..................-T .. .T C. ..... ATG-.S. intermedius .....C.T--... ..................-T .. .. C. ..... A.G-.R. ornata ...C.CT---... .T................-T .. .T C. ..... AT.-.D. fraseri ...C..G---... ..................-T .. .. C. ..... .TG-.L. burtonis ...C.A..T-... ..................GT .. .. C. ..... ATG-.P. marmoratus .T.C.A----C.. ..................-. .. .. .. ....A A..-.G. variegata ...A.A.C--... ..................-. .. .. CC ...C. .TG-.E. macularius .T.CT....-C.. ..................-. .. .. .C ....C ATG-.T. scincoides .T.C.AC---... ..................-T .. .. .. ...C. .TG..

S. C. DONNELLAN ET AL.114
33′ 33′ 32′ 34′ 34′12S 567–606 ******** *** ********N. milii AAG --CTTTAAA GT AG CC AAAATG- GG CCCTCCG- CC CAAC. laevis ... --.CCAC.. .. .. .. .C..A.- A. TT..T.C- .. ...R. leachianus ... --ACA.C.. A. .. .. ....C.- TA AATC.--- TA ...P. lindneri ... --....... .. .A .. .....AT .T ...C.--- A. A..D. vittatus .G. --.CACCG. A. .. A. ..GT.A- .C TTAC..-- G. A..O. tryoni ... --.AACT.. .. .. A. ...C.A- .C ...C.--- .. A..S. intermedius G.. --T.A.CT. .. .. A. .....CA .T TAAC.--- A. A..R. ornata ... -AT.AACT. .. .. A. .....A- .C ...CA.C- .. A..D. fraseri ... --ACCC... .. .. .. .C..CAA .T .AAA..-- A. A..L. burtonis ... --ACCCT.. .. .. A. .C..CTA .T TAAA..-- A. A..P. marmoratus T.. AAAAC..T. .. .A .. ......- .. ...CT.CC .. A..G. variegata .G. -ATACC.C. .. GA .. .G...A- .C ...CAACC A. T.GE. macularius G.. --A.G.... T. G. .. T...A.- .T ..TA..-- A. A..T. scincoides ... --AAAC... .. .. .. ....AA- .T TTACA--- A. T..
31′ 30′ 28′12S 607–657 ***N. milii AACGTCAGGTCAAGGTGTAGC ACAT GGGCGGG GCC AGAGATGGGCT ACA TTC. laevis ..................... .... ....... A.- T.......... ... ..R. leachianus ..................... .T.. .A.G... C-- ........... ... ..P. lindneri ..................... .... .AA.... CA- ........... ... ..D. vittatus ..................... .T.. .AAAA.. AG- ........... ... ..O. tryoni ..................... .T.. .AAGA.. AA- ........... ... ..S. intermedius ..................... .... ..AAA.. AA- ........... ... ..R. ornata ...................A. TA.A .AAAA.. AA- ........... ... ..D. fraseri ..................... .... AT.GC.. AA- ........... ... ..L. burtonis ..................... .T.. AA.A... AA- ........... ... ..P. marmoratus ..................... TA.. .ATA... C-- ........... ... ..G. variegata ..................... TA.C A..GAA. C-- ........... ... ..E. macularius .............C....... TT.. ...G... A-- ........... ... ..T. scincoides C.................... .G.. A.AAT.. A-- ........... ... ..
35 35′ 3612S 658–710 ****** ******** ********* *** ****N. milii TTCT ATAAA- AGAA AACAC--- CAACCC CTGC----- ---CCTGAAAA-----C. laevis .... .A..-- .... ....---- .C.A.. ...T----- ---........A----R. leachianus .... .A..-- .... ...----- ACTA.A ACA.----- ---.......CA----P. lindneri .... .A..-- .... ..T.---- .T.G.A GCA.----- ---.A......A----D. vittatus .... .GC.-- .... ..T.---- .C..AA .CA.----- ---....C...A----O. tryoni .... .A..-- .... C.A.---- .T.T.A TCAT----- ---........A----S. intermedius .... .AT.-- .... ..------ A.CAA. AATA----- --CAA......TAA--R. ornata .... .AT.-- .... ..T.---- .CCAA. ACA.----- ---........A----D. fraseri .C.. .A.--- ..G. C.A..A-- A..ATA T...----- --C.T......A----L. burtonis CC.. CA.--- ..GG ..A----- AC.G.A TATAC---- ---........A----P. marmoratus .... T....T .... C------- AC...G TAAGC---- -AATA.....CACAA-G. variegata .... .G.TT- ...C T.TCA--- .C.AAA G.------- -GC.A......AGC--E. macularius C... .A.TT- ...G -------- A.TTA. ACAAGG--- AAATA.....TTAAAAT. scincoides .... GCC.C- .... ..AT---- ACGAAA GGTGT---- ---AA.....CAC---

EVOLUTIONARY RELATIONSHIPS OF AUSTRALIAN GEKKONOID LIZARDS 115
36′ 27′12S 711–736 ******** *N. milii ----GTAG CCTCCGGTGGA TTTAG CAC. laevis ----AC.. ATCAA...... ..... ..R. leachianus ???????? ??????????? ????? ??P. lindneri ----..GC AAAA....... ..... ..D. vittatus ----..GC ..AAA.A.... ..... ..O. tryoni ----A.GA ...A....... ..... ..S. intermedius ----TATT -..A....... ..... ..R. ornata ----..GC .AAAA...... ..... ..D. fraseri ----.C.T AA.A....... ..... T.L. burtonis ---CTATA -AAAA...... ..... T.P. marmoratus ---GCC.A ---AA...... ..... ..G. variegata ------.C .AAAA..A... ..... ..E. macularius --CCAGTA ----A...... ....A GTT. scincoides ---ACACC -.AAA..A... ..... A.
c-mos 1–42N. milii TGT GGT AAG AAC AGT CTG GCA TCA CGG CAG AGC TTC TGG GCAC. laevis ??? ... ... ... ... T.. ... ... ... ... ... ... ... ...R. leachianus ..C ... ... ... ... T.A ... ..C ... ... ... ... ... ...P. lindneri A.C ... ... ... ... T.A ... ... ... ... ... ... ... ...D. vittatus ..C ... ... ... ... T.A ... ... ... ... ... ... ... ...O. tryoni ..C ... ... ... ... T.A ... ... ... ... ... ... ... ...S. intermedius ..C ... ... ... .A. T.. ... ... ... ... ... ... ... ...R. ornata ??? ?.. ... ... ... T.. ... ... ... ... ... ... ... ...D. fraseri ..C ... ... ... ... T.A ... ..T ... ... ... ... ... ...L. burtonis ??? ?.. ... ... ... T.A ... ... ... ... ... ... ... ...P. marmoratus ..C A.. ... ... ... T.A ... ... ... ... ... ... ... ...G. variegata ??? ??. ... ... ... T.A ... ... ... ... ... ... ... ...E. macularius ??? ??. ... ... ... T.A ... ... A.. ... ... ... ... ...T. scincoides ..C A.. ... ... C.. T.. ... ... ..A ..A ... ... ... ...
c-mos 43–84N. milii GAA TTA AAT GTG GCA CGC CTT GAT CAT AAA AAT GTG GTG CGTC. laevis ... ... ... ... ... .A. ... ... ... ... ..C ... ... ...R. leachianus ... ... ... ... ... ... ... ... ... ... ..C ... ... ...P. lindneri ... ... ... ... ... ... ... ... ... ... ..C ... ... ...D. vittatus ... ... ... ... ... ... ... ... ... ... ..C ... ... ...O. tryoni ... ... ... ... ... ... ... ... ... ... ... ... ... ...S. intermedius ... ..G ... ... ... ... ..G ... ... ... ..C ... ... ...R. ornata ... ... ... ... ... ... ... ... ... ... ..C ... ... ...D. fraseri ... ... ... ... ... .A. ... ... ... ... ..C ... ... .ACL. burtonis ... ... ... ... ... ..G ... ... ... ... ..C ... ... ...P. marmoratus ... ..C ... ..A ... ... ... ... ... C.. ... ... ... ...G. variegata ... ... ... ..A ... ... ... ... ... C.. ... ... ... ...E. macularius ... C.. ... ... ... TAT ... ... ... C.. ... ... ... ...T. scincoides ... C.G ..C ... ... ... ... AG. ... ..C ... ... ..A ...
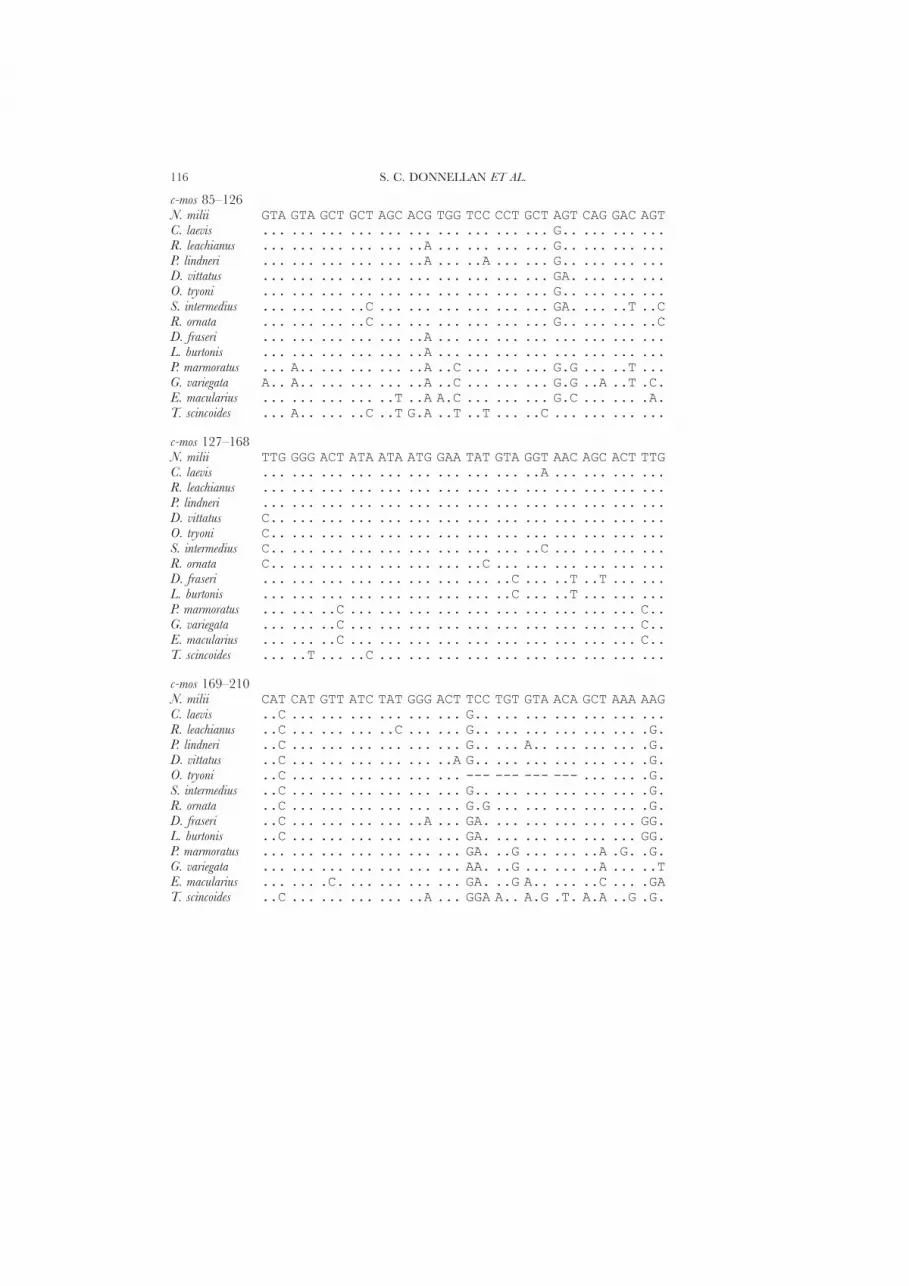
S. C. DONNELLAN ET AL.116
c-mos 85–126N. milii GTA GTA GCT GCT AGC ACG TGG TCC CCT GCT AGT CAG GAC AGTC. laevis ... ... ... ... ... ... ... ... ... ... G.. ... ... ...R. leachianus ... ... ... ... ... ..A ... ... ... ... G.. ... ... ...P. lindneri ... ... ... ... ... ..A ... ..A ... ... G.. ... ... ...D. vittatus ... ... ... ... ... ... ... ... ... ... GA. ... ... ...O. tryoni ... ... ... ... ... ... ... ... ... ... G.. ... ... ...S. intermedius ... ... ... ..C ... ... ... ... ... ... GA. ... ..T ..CR. ornata ... ... ... ..C ... ... ... ... ... ... G.. ... ... ..CD. fraseri ... ... ... ... ... ..A ... ... ... ... ... ... ... ...L. burtonis ... ... ... ... ... ..A ... ... ... ... ... ... ... ...P. marmoratus ... A.. ... ... ... ..A ..C ... ... ... G.G ... ..T ...G. variegata A.. A.. ... ... ... ..A ..C ... ... ... G.G ..A ..T .C.E. macularius ... ... ... ... ..T ..A A.C ... ... ... G.C ... ... .A.T. scincoides ... A.. ... ..C ..T G.A ..T ..T ... ..C ... ... ... ...
c-mos 127–168N. milii TTG GGG ACT ATA ATA ATG GAA TAT GTA GGT AAC AGC ACT TTGC. laevis ... ... ... ... ... ... ... ... ... ..A ... ... ... ...R. leachianus ... ... ... ... ... ... ... ... ... ... ... ... ... ...P. lindneri ... ... ... ... ... ... ... ... ... ... ... ... ... ...D. vittatus C.. ... ... ... ... ... ... ... ... ... ... ... ... ...O. tryoni C.. ... ... ... ... ... ... ... ... ... ... ... ... ...S. intermedius C.. ... ... ... ... ... ... ... ... ..C ... ... ... ...R. ornata C.. ... ... ... ... ... ... ..C ... ... ... ... ... ...D. fraseri ... ... ... ... ... ... ... ... ..C ... ..T ..T ... ...L. burtonis ... ... ... ... ... ... ... ... ..C ... ..T ... ... ...P. marmoratus ... ... ..C ... ... ... ... ... ... ... ... ... ... C..G. variegata ... ... ..C ... ... ... ... ... ... ... ... ... ... C..E. macularius ... ... ..C ... ... ... ... ... ... ... ... ... ... C..T. scincoides ... ..T ... ..C ... ... ... ... ... ... ... ... ... ...
c-mos 169–210N. milii CAT CAT GTT ATC TAT GGG ACT TCC TGT GTA ACA GCT AAA AAGC. laevis ..C ... ... ... ... ... ... G.. ... ... ... ... ... ...R. leachianus ..C ... ... ... ..C ... ... G.. ... ... ... ... ... .G.P. lindneri ..C ... ... ... ... ... ... G.. ... A.. ... ... ... .G.D. vittatus ..C ... ... ... ... ... ..A G.. ... ... ... ... ... .G.O. tryoni ..C ... ... ... ... ... ... --- --- --- --- ... ... .G.S. intermedius ..C ... ... ... ... ... ... G.. ... ... ... ... ... .G.R. ornata ..C ... ... ... ... ... ... G.G ... ... ... ... ... .G.D. fraseri ..C ... ... ... ... ..A ... GA. ... ... ... ... ... GG.L. burtonis ..C ... ... ... ... ... ... GA. ... ... ... ... ... GG.P. marmoratus ... ... ... ... ... ... ... GA. ..G ... ... ..A .G. .G.G. variegata ... ... ... ... ... ... ... AA. ..G ... ... ..A ... ..TE. macularius ... ... .C. ... ... ... ... GA. ..G A.. ... ..C ... .GAT. scincoides ..C ... ... ... ... ..A ... GGA A.. A.G .T. A.A ..G .G.

EVOLUTIONARY RELATIONSHIPS OF AUSTRALIAN GEKKONOID LIZARDS 117
c-mos 211–252N. milii GAG GAC AAT GGT CTT GGA GGT GGT CGA GAG TCC TTC AGC ATGC. laevis ... .GT G.. ... ... ... ... ..C ... ... ... ... ... ...R. leachianus A.. ..T G.. ..C ... ... ... ..C ... ... ... ..A ... C..P. lindneri A.. ..T G.. ..C ... ... ... ..C ... ... C.. ..A ... C..D. vittatus C.. ..T G.. ..C ... ... ... ..C ... ... ... ..A ... C..O. tryoni C.. ..T G.. ..C ... ... ... ..C ... ... ... ..A ... C..S. intermedius C.. ..T G.. ..C ... ... ... ..C ... ... ... C.A ... C..R. ornata C.. ..T G.. ..C ... ... ... ..C ... ... ... ..A ... C..D. fraseri A.. ..T G.. ... ... ... ... ..C .A. ... ... ..A ... ...L. burtonis A.. ..T G.. ... ... ... ... ..C ... ... ... ..A ... ...P. marmoratus A.. ..T G.. A.C ... ... T.. ..C ... ... ... ..A ..T T.AG. variegata A.. ..T G.A ..C ... ... T.. A.C .A. ... .TA ..A ... T.AE. macularius A.. ..T G.. ..G ... ... TA. ..C ... ... ..T ..A ... ...T. scincoides A.. ..T ... .AA ... ..T TA. ..C TAT ..A C.T ..G ..T ..A
c-mos 253–294N. milii TCT CAG TCT CTG AGC TAC TCC TGT GAC ATT GTG GCA GGC TTAC. laevis C.. ... ... ... ... ... ... ... ... ... ... ... ... ...R. leachianus C.. ... A.. ... ... ... ..T ..C ... ... ... ... ... ...P. lindneri C.. ... ... ... ... ... ... ... ... ... ... ... ... ...D. vittatus C.. ... ... ... ... ... ... ... ... ... ... ... ... C..O. tryoni C.. ... ... ... ... ... ... ... ... ... ... ... ... ...S. intermedius G.. ... G.. ... ... ... ... ... ... ... ... ... ... ...R. ornata C.. ... ... ... ... ... ... ... ... ... ... ... ... ...D. fraseri C.. ... ... ... ... ... ... ... ... ... ... ... ... ...L. burtonis C.. ... ... ... ... ..T ... ... ... ... ... ... ... ...P. marmoratus G.. ... ... ... CA. ... ... ... ... ... ... ... ..T C..G. variegata G.. ... ... ... CA. ... ... ... ... ... ... AA. ..T C..E. macularius GG. G.. .G. ... ... ... ... ... ... ... ... ... ... ...T. scincoides A.A ... ... ... ... ... ..A ... ... ... ... ... ..T ..G
c-mos 295–336N. milii GCC TTT CTC CAT TCC CAG TTA ATT GTG CAC CTG GAT TTA AAAC. laevis ... ... ... ... ... ... ... ... ... ... ... ... ... ...R. leachianus ... ... ... ... ... ... ... ... ... ... ... ... ..G ...P. lindneri ... ... ... ... ... ... ... ... ... ... ... ... ..G ...D. vittatus ... ... ... ... ... ... ..G ... ... ... ... ... ..G ...O. tryoni ... ... ... ... ... ... ... ... ... ... ... ... ..G ...S. intermedius ... ... ... ... ... ... ... ... ... ... ... ... ..G ...R. ornata ... ... ... ... ... ... ..G ..C ... ... T.A ... ..G ...D. fraseri ... ... ... ... ... ... ... ... ... ... ... ... ... ...L. burtonis ... ... ... ... ... ..A ... ... ... ... ... ... ... ...P. marmoratus .T. ... ... ... ..A ... ... ... ... ..T T.. ... ... ...G. variegata .T. ... ... ... ..A ... ... ... ... ... T.. ... ... ...E. macularius .TT ... ... ... ..A ... ..C ... ... ... ... ... ... ...T. scincoides CT. ... ... ... ..A ..A ... ... ... ..T ... ... ... ...

S. C. DONNELLAN ET AL.118
c-mos 337–374N. milii CCT GCC ?AC ATA TTC AT? ACC GAA CAA AAT ATT TGC AAC. laevis ... ... A.. ... ... ..C ... ... ... ... ... ... ??R. leachianus ... ... A.. ..T ... ..C ..T ... ... ... ... ... ..P. lindneri ... ... A.. ..T ... ..C ..T ... ... ... ... ... ..D. vittatus ... ... A.. ..T ... ..C ..T ... ... ... ... .?? ??O. tryoni ... ... A.. ..T ... ..C ..T ... ... ... ... ... ..S. intermedius ..G ... A.. ..T ... ..C ..T ... ... ... ... ... ..R. ornata ... ... A.. ..T ... ..C ..T ... ... ... ... .?? ??D. fraseri ... ... A.. ... ... ..C ..T ... ... ... ... ... ..L. burtonis ... ... A.. ... ... ..C ... ... ... ... ... ... ??P. marmoratus ... ..T A.. ... ... ..C ..T ... ... ... ... ... ..G. variegata ... ..T A.. ... ... ..C ..T ..C ... ... ... ??? ??E. macularius ... ... A.. ... ... ..C ..T ... ... ... ... G?? ??T. scincoides ... ... A.. ... ... ..C ..T ... ... ... G.. ... ..





























