Embed Size (px)
Citation preview

Monitoring and isolation of blood dendritic cells from
apheresis products in healthy individuals: a platform
for cancer immunotherapy
J. Alejandro Lopez a, Georgina Crosbie a, Cathryn Kelly b, Ann Marie McGee a,Katrina Williams b, Slavica Vuckovic a, Robert Schuyler c, Robyn Rodwell b,
Sue J. Wright b, Kerry Taylor b, Derek N.J. Hart a,*
aMater Medical Research Institute, Aubigny Place, South Brisbane 4101, AustraliabMater Adult Hospital, Raymond Terrace, South Brisbane 4101, Australia
cGambro BCT, Lakewood, CO 80215, USA
Received 25 January 2002; received in revised form 19 April 2002; accepted 21 May 2002
Abstract
The fundamental role of dendritic cells (DC) in initiating and directing the primary immune response is well established.
Furthermore, it is now accepted that DC may be useful in new vaccination strategies for preventing certain malignant and
infectious diseases. As blood DC (BDC) physiology differs from that of the DC homologues generated in vitro from monocyte
precursors, it is becoming more relevant to consider BDC for therapeutic interventions. Until recently, protocols for the isolation
of BDC were laborious and inefficient; therefore, their use for investigative cancer immunotherapy is not widespread. In this
study, we carefully documented BDC counts, yields and subsets during apheresis (Cobe Spectra), the initial and essential
procedure in creating a BDC isolation platform for cancer immunotherapy. We established that an automated software package
(Version 6.0 AutoPBPC) provides an operator-independent reliable source of mononuclear cells (MNC) for BDC preparation.
Further, we observed that BDC might be recovered in high yields, often greater than 100% relative to the number of circulating
BDC predicted by blood volume. An average of 66 million (range, 17–179) BDC per 10-l procedure were obtained, largely
satisfying the needs for immunization. Higher yields were possible on total processed blood volumes of 15 l. BDC were not
activated by the isolation procedure and, more importantly, both BDC subsets (CD11c+CD123low and CD11c�CD123high) were
equally represented. Finally, we established that the apheresis product could be used for antibody-based BDC immunoselection
and demonstrated that fully functional BDC can be obtained by this procedure.
D 2002 Published by Elsevier Science B.V.
Keywords: Immunotherapy; Dendritic cells; Apheresis; Isolation
0022-1759/02/$ - see front matter D 2002 Published by Elsevier Science B.V.
PII: S0022 -1759 (02 )00185 -0
Abbreviations: APC, antigen-presenting cell; BDC, Blood DC; DC, dendritic cells; MNC, mononuclear cells; Mo-DC, monocyte-derived
DC; PBMC, peripheral blood mononuclear cells; SCF, stem cell factor; TBV, total blood volume.* Corresponding author. Tel.: +61-7-3840-2557; fax: +61-7-3840-2550.
E-mail address: [email protected] (D.N.J. Hart).
www.elsevier.com/locate/jim
Journal of Immunological Methods 267 (2002) 199–212

1. Introduction
Dendritic cells (DC) due to their unique properties
as initiators and modulators of the immune system
have become the focus of studies for the development
of vaccination strategies for malignant disease (Hart
and Hill, 1999; Fong and Engleman, 2000). Emerging
data from Phase I and Phase II studies show very
promising results (Hsu et al., 1996; Nestle et al., 1997;
Murphy et al., 1999; Geiger et al., 2000; Kugler et al.,
2000). However, there is a lack of consensus with
respect to the methods of DC isolation, the source cell
population for derivation of DC, the need for and
influence of exogenous cytokine stimulation and the
effects of the isolation strategy on the DC product
(Nestle et al., 2001). It is also now important to
employ Good Manufacturing Practice principles to
ensure consistency of the DC product and compliance
with an increasingly stringent regulatory environment.
DC plays a fundamental role both in innate and
cognate immune responses (Hart, 1997; Clark et al.,
2000). They circulate in blood as precursors with an
intermediate differentiation/activation phenotype and
represent a small (less than 1%) proportion of blood
mononuclear cells (MNC). DC moves from the cir-
culation into the tissues, where they acquire a fully
mature antigen-presenting cell (APC) phenotype after
interaction with pathogens or other stimuli. This is
characterized by the expression of key surface mole-
cules such as CD40, CD80 and CD86, which are
involved in costimulation as well as other markers of
DC differentiation/activation (Banchereau et al., 2000;
Hart et al., 2000). Various subsets of DC have been
described to date and they appear to have different
physiological functions (Rissoan et al., 1999); they are
defined by the expression of the CD123 (IL-3 recep-
tor) and the CD11c antigens (Olweus et al., 1997;
Robinson et al., 1999).
Various approaches have been used to achieve DC
isolation resulting in differences in the properties of the
generated DC preparations (Ferlazzo et al., 1999). DC-
like cells can be produced via transformation of blood
monocytes in vitro in the presence of various cytokines
and are referred to as monocyte-derived DC (Mo-DC)
(Romani et al., 1994; Sallusto and Lanzavecchia,
1994); additionally, DC can be expanded from
CD34+ progenitors obtained from bone marrow (Reid
et al., 1992; Egner et al., 1993; Szabolcs et al., 1995;
Young et al., 1995), peripheral blood (Mackensen et
al., 1995; Siena et al., 1995; Strunk et al., 1996; Monji
et al., in press) or neonatal cord blood (Caux et al.,
1992, 1996). Alternatively, DC and their precursors
present in the blood (blood dendritic cells, BDC) can
be enriched using density gradients (Mehta-Damani et
al., 1994; McLellan et al., 1995) and selected further
using monoclonal antibodies (Fearnley et al., 1997;
Schakel et al., 1998; Dzionek et al., 2000). This latter
approach encompasses potential theoretical advan-
tages over cytokine-mediated DC in as much as it
yields homogenous DC in a defined state of differ-
entiation, capable of responding to physiological stim-
uli and free from the influence of exogenous cytokines
(Hart and Hill, 1999). We have observed differences
between BDC and Mo-DC in their expression of
surface markers such as DC-SIGN, DEC-205 and the
mannose receptor (Hart et al., 2000; Kato et al., 2000).
More importantly, BDC appeared to be more efficient
in the induction of in vitro primary immune responses
(Osugi et al., in press. Blood, 2002), as well as in vitro
antigen uptake and processing (Ho et al., 2002).
Apheresis products have been used previously as a
source of BDC for cancer immunotherapy (Fong et
al., 2001), but no detailed studies on BDC phenotype
and yields have been published to date. Furthermore,
as the phenotype and function (e.g. adhesion mole-
cules and chemokine receptors) of BDC can be easily
modulated, it is important to investigate the effect of
apheresis on BDC phenotype and function. We have
demonstrated previously that BDC numbers can be
rapidly increased as a result of external modulators
such as surgical and physical stress (Ho et al., 2001)
and it is possible that BDC might ‘‘mobilize’’ during
the procedure.
Given the potential advantages of BDC, we have
developed a method for the generation of DC from the
mononuclear cell apheresis product via a single mag-
netic bead separation procedure using the monoclonal
antibodies CMRF-44 (Hock et al., 1994; Fearnley et
al., 1997) and 56 (Hock et al., 1999). Further descrip-
tion of the kinetics of CMRF-44/56 expression on
BDL and their purification will be provided elsewhere
(Lopez et al., submitted JIM, 2002). In order to
improve BDC yields and maximize their use in cancer
vaccination, it is essential to determine optimal aphe-
resis conditions. We describe a clinically applicable
technique using apheresis of mononuclear cells from
J. Alejandro Lopez et al. / Journal of Immunological Methods 267 (2002) 199–212200

unstimulated healthy donors for the purification of
BDC from the apheresis product via a single magnetic
bead separation procedure.
2. Materials and methods
2.1. Study population
The Mater Misericordiae Health Services Ethics
Committee approved the study. Healthy volunteers
(n=14; 10 males, 4 females), average age 41 (range
21–56), participated in this study. Medical evaluation
and complete blood cell counts were performed prior
to apheresis. No prior cytokine stimulation/mobiliza-
tion was attempted.
2.2. Apheresis isolation software
Two different programs on the Cobe Spectra
(Gambro BCT, Lakewood CO, USA) were compared
for BDC collection. The semi-automated (Version 4.7
software MNC Program) ‘‘manual’’ version, with
variable inlet flow rate from 20 to 150 ml/min allows
a continuous collection of the mononuclear cell
(MNC) layer forming at the interface between the
red cells and the plasma. The product is collected
when the operator judges the interface position to be
optimal based on collect line appearance (judged by
comparison with the Spectra WBC colorgram). The
hematocrit range of the collected products was 3–5%.
This protocol takes into account hematocrit, sex,
height and weight to determine the flow rates
required. The second protocol, the automated (Version
6.1 software, AutoPBPC ) version, accumulates MNC
at the interface position and periodically harvests them
according to the rate of movement of MNC into the
centrifuge channel. Input of subject MNC count,
hematocrit, height, weight and sex enables the Spectra
software to automatically control the collection over
three phases: accumulation, harvest and chase. We
configured the system to harvest 4-ml volumes of
MNC and a platelet-rich plasma chase of 7- or 9-ml
volumes.
Fig. 1. FACS profile of MNC labeled and analyzed for DC. Forward and side scatter profile selected viable cells (left panel) and live cells were
labeled with lineage markers and HLA-DR+. Lin�HLA-DR+ cells (region R2 in middle panel) were analyzed for BDC subsets in dot plots
(right panel).
J. Alejandro Lopez et al. / Journal of Immunological Methods 267 (2002) 199–212 201

2.3. Apheresis sample collection
Peripheral venous blood samples (20 ml) were taken
from a peripheral vein before apheresis (time 0), 30
min, 24 and 48 h after completion of the procedure.
Apheresis was performed in the Mater Hospital Clin-
ical Apheresis Unit. Additionally, at various time
points during the procedure, the products correspond-
ing to total processed blood volumes of 2.5, 5, 7.5, 10,
12.5 and 15 l were collected. Unless specifically stated,
samples were harvested at every planned volume,
except when no product had been collected by the
Fig. 2. Selection of harvesting software protocol. Data corresponding to seven experiments run with the automatic protocol Spectra Version 6.1,
AutoPBPC (shown in open bars, 1, 4, 10, 16, 19, 21 and 22) and four experiments run with the manual protocol Spectra Version 4.7, MNC (shown
in full bars, 2, 3, 9 and 20). (A) Bars show total MNC, volumes, platelets and DC recovered in 10-l apheresis products for each experiment. (B)
BDC efficiency of recovery in repeated experiments performed in three different donors, using the two software options. Placed horizontally,
experiments performed on the same individual with a minimum of 2 weeks interval. Calculated BDC (106) present in TBV before the procedure
(as described in Materials and methods) were compared with the total BDC recovered after apheresis and expressed as total DC recovered.
J. Alejandro Lopez et al. / Journal of Immunological Methods 267 (2002) 199–212202

machine. In some experiments, samples were taken
from the access vein immediately before each product
harvest. ACD-Awas used as anticoagulant at a ratio of
1:12 to the inlet flow. Normal healthy volunteers
participated in the study and they received standard
clinical apheresis care. The Mater Adults Hospital’s
Ethical Committee approved their participation.
2.4. Isolation of peripheral blood mononuclear cells
(PBMC) and labeling of BDC
MNC were isolated by Ficoll–Hypaque (Pharma-
cia, Sweden) density gradient and the interphase cells
washed three times. Cells were labeled in a 50-Almixture of hybridoma supernatants containing anti-
bodies specific for: CD14 (CMRF-31, IgG2a; pro-
duced in this laboratory), CD3 (OKT3, IgG2a,
American Type Tissue Collection, Rockville, MD),
CD16 (HuNK2, IgG2a) and CD19 (FMC63, IgG1),
gifts of Prof H. Zola, Adelaide, Australia. After one
washing step, fluorescein-conjugated sheep–mouse
immunoglobulin-specific (FITC-SAM, Amrad Bio-
tech, Victoria, Australia) was added, followed by
saturating concentrations of mouse immunoglobulin
(Sigma, St. Louis). Finally, PE.Cyanin5-conjugated
HLA-DR specific antibody (Immu375, IgG1, Immu-
notech, Marseille, France) was added together with
either PE-conjugated CD11c (S-HCL-3, IgG2a, BD
Bioscience, BDIS, San Jose, CA) or PE-conjugated
CD123 (a chain specific, 9F5, IgG2b, PharMingen
International, San Diego, CA). IgG1 (MOPC-21),
IgG2a (G155-178) and IgG2b (27-35) isotype con-
trols were purchased from PharMingen. For certain
experiments, the following antibodies were used: PE-
conjugated CD14 (leuM3, IgG2b), CD19 (leuM12,
IgG1) and CD80 (L307.4, IgG1) from BDIS and PE-
conjugated CD40 (5C3, IgG1), CD86 (24F, IgG1,
PharMingen). Labeled samples were analyzed by
FACSCalibur flow cytometer (BDIS).
2.5. DC counts
BDC counts (expressed as 106/l) were calculated
from the number of MNC/L blood (determined by the
automated cell counter Advia 120, Hematology Sys-
tem, Bayer, Tarrytown, NY) multiplied by the percent-
age of BDC (Ho et al., 2001). Alternatively, values
were also expressed as the total content of BDC
calculated on total blood volume (TBV) of every
volunteer.
2.6. DC isolation
The BDC purification procedure will be described
elsewhere (Lopez et al., submitted. JIM, 2002).
Briefly, magnetic bead separation of BDC was under-
taken using biotinylated CMRF-44 (Hock et al., 1994)
antibody and biotin-specific beads (Miltenyi Biotech,
Bergisch Gladbach, Germany). Positive and negative
fractions were collected and evaluated by flow cytom-
etry (level of BDC enrichment) and MLR (function).
For the experiment with frozen cells, apheresis mate-
rial was diluted in ice-cold DMEM medium to 50% of
the total final volume and supplemented with 10%
DMSO and 40% AB pooled serum, stored at �70 jCin an insulated styrofoam container and tested after 2
weeks of storage.
2.7. MLR with isolated BDC
A titration of irradiated MNC (cultured and fresh),
CMRF-44+ magnetically selected BDC or the nega-
tive fraction of the CMRF-44 isolation (5�103–
3�105 cells) was incubated with allogeneic T-lym-
Table 1
Efficiency of recovery of BDC: blood dendritic cells (BDC)
recovered after apheresis expressed as a percentage of total blood
volume (TBV)a
Experiment Software used
Automatic Manual
1 179
2 126
3 50
4 120
9 44
10 23
16 28
19 65
20 65
21 40
22 101
Mean 79.4 71.3
SE 21.6 18.8
a Percentage was established on the basis of the TBV (before
starting the procedure) and the calculated total DC recovered in the
apheresis product.
J. Alejandro Lopez et al. / Journal of Immunological Methods 267 (2002) 199–212 203

phocytes (5�104 cells) for 5 days in 96-well U-
bottom plates. Sixteen hours prior to harvesting the
cells, 0.5 ACi of 3H-thymidine was added to each
well. 3H-thymidine uptake was counted in a liquid h-scintillation counter (Wallac, MicroBeta Trilux Scin-
tillation Counter, Turku, Finland).
2.8. Statistical analysis
Paired statistical analysis was performed using the
Student’s two-tailed t-test.
3. Results
3.1. Definition of BDC for counting
Quantification of BDC was performed defining
class II expressing cells, lacking markers for specific
hemopoietic lineages. DC subsets were identified as
CD11c+ or CD123+ cells within the BDC gated
population (Fig. 1). For the purpose of calculations,
BDC were defined as the percentage of lineage
negative HLA-DR high expressing cells.
Fig. 3. Total BDC recovery. (A) Detailed evaluation of BDC present in 10-l (open bars) and 15-l apheresis (hatched) products; the numbers of
BDC (106) present at various harvesting points are shown as independent bars (right axis) and the percentages of BDC as a line (left axis). (B)
Percentage of BDC present in blood before, 0.5, 24 and 48 h after the procedure.
J. Alejandro Lopez et al. / Journal of Immunological Methods 267 (2002) 199–212204

3.2. Compliance with the apheresis procedure
As reported in the literature (Furuta et al., 1999;
Sato et al., 2001), apheresis was safe and well
tolerated. Of the 24 procedures undertaken, 1 was
aborted due to difficult venous access and 1 due to
mild hypocalcaemia symptoms (spontaneously revert-
ing); the rest were well tolerated with only local
discomfort that ceased at the end of the procedure.
Only a small (average 13%) reduction in the platelet
counts was detected at the end of the apheresis (mean
235�109/ml before and mean 201�109/ml after).
3.3. Optimal software for BDC isolation
Ten-liter collections were performed using the
semi-automated (Version 4.7 software MNC Program)
‘‘manual’’ or fully ‘‘automatic’’ (AutoPBSC) software
versions to control the COBE Spectra. A comparison
of the volume, total of MNC recovered, number of
platelets and total BDC recovered in the apheresis
product is shown in Fig. 2A. The automatic software
(n=7) resulted in consistently lower product volumes
(meanFS.E.=automatic: 45F5 ml versus manual:
116F8 ml; p<0.005) and the number of contaminating
platelets was also significantly lower than in the
manual (n=4) collections (automatic: 57F10�109
versus manual: 459F104�109; p<0.05). Although
the total number of MNC recovered from the manual
procedures was, as expected, higher than that obtained
with the automatic software (11.088F1.415�109 and
5.982F1.148�109, respectively; p<0.05), there was
no statistically significant difference in the number of
BDC recovered (automatic: 69F16�106 versus man-
ual: 102F35�106; p=0.34). Furthermore, the effi-
ciency to recover the available circulating BDC
(predicted blood volume�BDC count) was compara-
ble using the two software protocols (automatic:
80F22% versus manual: 71F19%; p=0.77). As
shown in Table 1, the efficiency of recovery differed
Fig. 3 (continued).
J. Alejandro Lopez et al. / Journal of Immunological Methods 267 (2002) 199–212 205

considerably between individual experiments possibly
as a reflection of the variations in BDC counts
documented previously (Ho et al., 2001). Three sets
of experiments were performed on the same individ-
uals utilizing automatic and manual software with a
minimum 2-week interval (experiments 1 and 2; 10,19
and 9; 21,22 and 20). In this group of experiments
(Fig. 2B), the automatic software (experiments 1, 10,
19, 21 and 22) produced an average 82% BDC
recovery as opposed to 78% with the manual proce-
dure (experiments 2, 9 and 20). The automatic soft-
ware was considered advantageous because of the
smaller product volume, lower platelet contamination
and reduced operator dependency, appropriate for
multicenter trials.
3.4. Optimal harvesting volume
We next evaluated the recovery of BDC obtained
using the automatic software at different total inlet
volumes. Firstly, we studied four 10-l and five 15-l
Fig. 4. Efficiency of BDC recovery. (A) The total BDC (106) present in the apheresis product from 10-l (open bars) and 15-l (shaded bars)
apheresis. (B) The percentage of the theoretical total BDC available obtained based on TBV is plotted.
Table 2
Activation status of blood dendritic (BDC) before, during and after
apheresis procedure: percentage of BDC
Sample % CD40+ % CD80+ %CD86+
Experiment
2 3 4 2 3 4 2 3 4
Before 7 47.4 25.4 0 4.5 1.1 0.97 38.8 57.5
Product 14 23.5 37.6 0.12 5.0 2.9 1.33 41.5 39.0
After 30 min 6 57.3 22.8 0 3.6 1.2 0.33 34.2 56.7
24 h 11.9 0.5 44.5
48 h 21.2 8.4 0.5 0.3 43.9 62.5
Fig. 5. BDC subset distribution in apheresis product. MeanFS.E. of
the percentage of CD11c+ and CD123+ in blood samples before and
after apheresis procedure (n=6, experiments 1, 2, 3, 4, 9 and 10) is
shown.
J. Alejandro Lopez et al. / Journal of Immunological Methods 267 (2002) 199–212206

procedures in detail. As shown in Fig. 3A, more than
50% of all the BDC recovery took place during the
later part of the procedure; this observation was made
both in the 10-l and the 15-l procedures. The higher
yields towards the end of the procedure resulted from
the combined effect of a slight increase in the
percentage of BDC (Fig. 3A, lines) and an increase
in the MNC in the product (not shown). There were
only minor changes in the percentage of BDC in
blood after the procedure. Although there was not a
defined pattern in all of the experiments, small
decreases in BDC% were detected 30 min after the
end of the procedure with a fast rebound at 24 h in
experiments 6, 8, 13 and 17 (Fig. 3B), i.e. four of
nine procedures.
3.5. Efficiency of BDC recovery
The efficiency of BDC recovery by the automatic
procedure was evaluated further by extending the
number of volunteers. As shown in Fig. 4A, the
efficiency of recovery was variable (n=17; mean
efficiency 73%; range 17–179%) and it reflected
volunteer variations. Recoveries that were greater than
Fig. 6. Isolation of CMRF-44 BDC from apheresis product. Dot plots show FACS analysis of cells before magnetic bead separation (upper
panels), in negative (middle panels) and positive (lower panels) fractions. Percentage of cells present in every quadrant of the dot plot is shown.
Left panels: experiments with freshly obtained apheresis product. Right panels: results obtained with the same cells frozen and thawed after 2
weeks.
J. Alejandro Lopez et al. / Journal of Immunological Methods 267 (2002) 199–212 207
100% of the predicted number of BDC in TBV
indicated possible BDC mobilization during the
procedures. Furthermore, the finding reinforced this
observation that higher recoveries were observed at
the latter harvesting points (data not shown). The
yield from the 5 15-l apheresis procedure was
112F21�106 (meanFS.E.), while that of the 12
10-l procedure was 78F17�106 ( p=0.2999). The
efficiency of the 15-l procedures was 97F10% and
that of the 10-l procedure, 66F14% ( p=0.202).
Evaluation of the BDC percentage and BDC counts
in peripheral blood obtained from the access line was
performed at times coinciding with the different
apheresis volume harvests. These remained constant
throughout the procedure (data not shown). This
finding was also observed in the 15-l experiments.
These data further reinforce the point that active
BDC mobilization or recruitment occurred during the
apheresis.
3.6. BDC costimulatory phenotype is not altered by
the apheresis procedure
As shown in Table 2, the apheresis procedure does
not generate significant changes in the expression of
CD40, CD80 and CD86 on lineage�HLA-DR+ DC
(n=3).
3.7. DC subset distribution remains unchanged after
apheresis
The BDC subset proportions remained unchanged
after the procedure. Fig. 5, summarizes the percent-
age CD11c+ DC and CD123+ subset distribution
observed in six experiments. In these experiments,
the CD11c+/CD123+ DC ratio before apheresis was
0.52F0.17 versus 0.45F0.18 in the product ( p=
0.502).
3.8. BDC can be successfully isolated from apheresis
product
We have optimized an isolation protocol for BDC,
based on the use of the CMRF-44 monoclonal anti-
body and magnetic bead technology to purify BDC
after overnight culture (Lopez et al., submitted for
publication). Following apheresis, CMRF-44 antigen
expression in vitro was not altered and purification
of the BDC was readily achieved (Fig. 6). Further-
more, the separation of BDC from both fresh and
frozen apheresis products yielded comparable BDC
purity (Fig. 6). Furthermore, repeated BDC isolations
(n=4) performed on the same frozen apheresis
material yielded very consistent results (data not
shown). The BDC purified in this way from aphe-
resis material were fully functional in an allogeneic
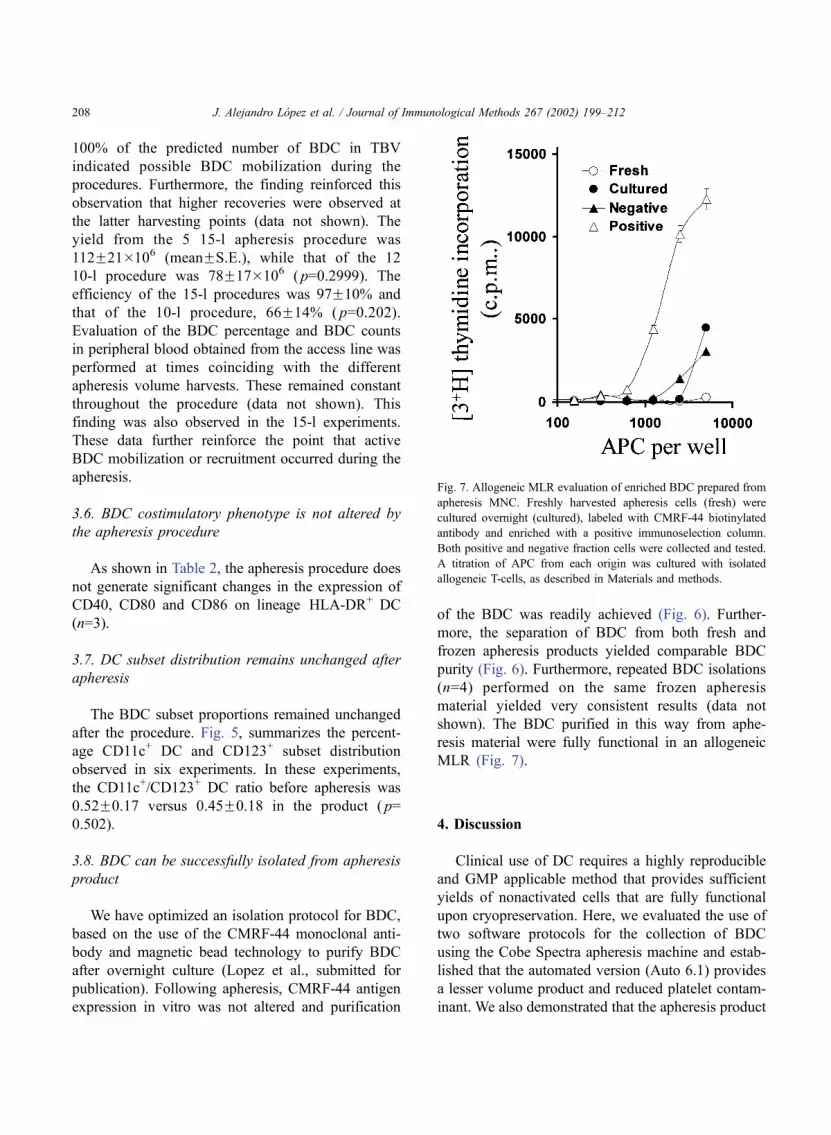
MLR (Fig. 7).
4. Discussion
Clinical use of DC requires a highly reproducible
and GMP applicable method that provides sufficient
yields of nonactivated cells that are fully functional
upon cryopreservation. Here, we evaluated the use of
two software protocols for the collection of BDC
using the Cobe Spectra apheresis machine and estab-
lished that the automated version (Auto 6.1) provides
a lesser volume product and reduced platelet contam-
inant. We also demonstrated that the apheresis product
Fig. 7. Allogeneic MLR evaluation of enriched BDC prepared from
apheresis MNC. Freshly harvested apheresis cells (fresh) were
cultured overnight (cultured), labeled with CMRF-44 biotinylated
antibody and enriched with a positive immunoselection column.
Both positive and negative fraction cells were collected and tested.
A titration of APC from each origin was cultured with isolated
allogeneic T-cells, as described in Materials and methods.
J. Alejandro Lopez et al. / Journal of Immunological Methods 267 (2002) 199–212208

contains sufficient numbers of BDC in a nonactivated
state suitable for immunotherapy. No skewing of
subset composition was observed allowing further
subset fractionation, if appropriate. The data suggests
that BDC are mobilized during the procedure and
although a 10-l apheresis procedure is sufficient, if
necessary, a 15-l apheresis procedure will generate an
improved BDC yield. Finally, we showed that BDC
can be isolated from the apheresis product and that
they are also fully functional after cryopreservation. A
detailed functional evaluation of BDC purified after a
magnetic bead separation following apheresis is pre-
sented elsewhere (Lopez et al., in preparation).
The selection of an automated protocol for the
harvest of BDC has various advantages. The repro-
ducibility and the operator independence permits the
standardization of the method across laboratories, a
particularly relevant issue when multi-center proto-
cols are envisaged, as will be important in evaluating
DC cancer immunotherapy protocols. Apheresis
using automated Cobe AutoPBSC software has also
been used to obtain MNC for the preparation of Mo-
DC and these were likewise fully functional; how-
ever, details on the apheresis procedure are not
available for comparison (Lewalle et al., 2000).
Apheresis MNC products have been used success-
fully for the production of Mo-DC from monocytes
(Thurner et al., 1999; Goxe et al., 2000; Lewalle et
al., 2000) and Langerhans cells from CD34+ progen-
itors but again, data on yields were not provided
(Gatti et al., 2000). Apheresed MNC preparations
have also been used for density gradient separation
of BDC but no information has been published
regarding the yields (Fong et al., 2001).
The reduced product volume facilitates further
BDC isolation procedures including the labeling with
monoclonal antibodies. The Cobe AutoPBPC pro-
gram provided lesser platelet contamination of the
sample. This feature is particularly relevant, since
platelets are known for their capacity to activate
BDC; for example, high platelet contamination has
been shown to hinder Mo-DC generation from aphe-
resis material both in melanoma patients and healthy
volunteers (Glaser et al., 1999).
Critical to the clinical use of the pheresed material
is the ability to handle the products in a closed clinical
grade environment. Clinical grade Mo-DC has been
prepared from pheresed MNC after processing with
the Cobe Spectra cell separator (Rouard et al., 2000).
This methodology can now be applied to BDC. The
product can then be enriched significantly for BDC,
whilst maintaining sterile Good Manufacturing Prac-
tice (GMP) procedures, using a BDC-specific mAb for
positive cell selection.
Cryopreservation of DC preparations will also be
highly advantageous for clinical scheduling. Protocols
examining the generation of Mo-DC have shown that
they may be stored safely frozen either as PBMC or as
matured Mo-DC (Thurner et al., 1999; Lewalle et al.,
2000). It has also been demonstrated that antigen-
loaded Mo-DC obtained from apheresis protocols can
be stored frozen, retaining their stimulatory function
(Feuerstein et al., 2000; Lewalle et al., 2000). Our
data shows that BDC can also be obtained from frozen
PBMC collected by apheresis. Thawing cells prior to
an overnight incubation before the antibody-mediated
separation of DC makes this a very convenient pro-
tocol.
Mo-DC has been generated from the apheresis
products obtained from patients with multiple mye-
loma (Tarte et al., 1997) and chronic myeloid leuke-
mia (Zheng et al., 2000). Indeed, we now have data
that establishes the optimal BDC harvesting time in
G-CSF and cyclophosphamide-treated patients with
non-Hodgkin lymphoma and multiple myeloma, who
are undergoing mobilization for blood stem cell col-
lection (Vuckovic et al., in preparation). The data also
predict that sufficient numbers of BDC will be
obtained from apheresis in those patients without the
need of additional mobilization. However, various
mobilizing conditions have been used to optimize
the generation of monocyte and CD34+-derived DC,
resulting in changes in the phenotype and/or function
of various cell types including T-lymphocytes NK and
DC (Gazitt, 2000; Roth et al., 2000). Stem cell factor
(SCF) mobilization in patients with breast cancer
resulted in minimal increases in BDC populations
(Menedez et al., 2001). Flt-3 ligand mobilizes BDL
in healthy volunteers (Maraskovsky et al., 2000) and
is currently being investigated by several groups for
its ability to mobilize BDC in patients.
DC are subject to various physiological influences
that may change their status of activation and/or
mobilization. We have observed that stress (surgery
and extreme exercise) prompts mobilization of BDC
without activation (Ho et al., 2001). Likewise, we
J. Alejandro Lopez et al. / Journal of Immunological Methods 267 (2002) 199–212 209

have documented changes in certain disease states
(Summers et al., 1999; Vuckovic et al., 1999; Brown
et al., 2001). Given their responses to environmental
influences, it was not inconceivable that apheresis
would change BDC differentiation/activation. Our
data on the evaluation of whole DC suggest that the
apheresis procedure does not induce the activation
state of collected BDC; a highly desirable feature
allowing for in vitro optimization of BDC function
for immunotherapy protocols. Differential activation
of a proportion of cells within certain DC subsets
(MacDonald et al., in press) may escape detection and
will be the subject of future analyses. In contrast,
maturation of Mo-DC from an apheresis product was
reported to occur earlier than for cells drawn from
fresh blood samples (Thurner et al., 1999). We have
shown here that BDC isolated from apheresis MNC
preparations exhibited similar functions to BDC
drawn by direct venipuncture.
In conclusion, this study describes critical infor-
mation required to generate sufficient, nonactivated
and fully functional BDC in a standard manner for
DC-based cancer immunotherapy studies.
Acknowledgements
We gratefully acknowledge the enthusiastic partic-
ipation of the volunteers recruited for this study; we
thank Drs. Cameron Turtle, Chris Ho, Geoff Hill and
Nick Murray for constructive discussions and Gilles
Bioley for help with the functional evaluation of
apheresis products. We acknowledge the support
Miltenyi Biotech (Bergisch Gladbach, Germany) for
immuno-isolation reagents. This work was funded by
the Mater Medical Research Institute and by GAM-
BRO BCT (Lakewood CO).
References
Banchereau, J., Briere, F., Caux, C., Davoust, J., Lebecque, S., Liu,
Y.J., Pulendran, B., Palucka, K., 2000. Immunobiology of den-
dritic cells. Annu. Rev. Immunol. 18, 767–811.
Brown, R.D., Pope, B., Murray, A., Esdale, W., Sze, D.M., Gibson,
J., Ho, P.J., Hart, D., Joshua, D., 2001. Dendritic cells from
patients with myeloma are numerically normal but functionally
defective as they fail to up-regulate CD80 (B7-1) expression
after huCD40LT stimulation because of inhibition by trans-
forming growth factor-beta(1) and interleukin-10. Blood 98,
2992–2998.
Caux, C., Dezutter-Dambuyant, C., Schmitt, D., Banchereau, J.,
1992. GM-CSF and TNF-cooperate in the generation of den-
dritic Langerhans cells. Nature 360, 258–261.
Caux, C., Vanbervliet, B., Massacrier, C., Dezutter-Dambuyant, C.,
de Saint-Vis, B., Jacquet, C., Yoneda, K., Imamura, S., Schmitt,
D., Banchereau, J., 1996. CD34+ hematopoietic progenitors
from human cord blood differentiate along two independent
dendritic cell pathways in response to GM-CSF+TNF alpha. J.
Exp. Med. 184, 695–706.
Clark, G.J., Angel, N., Kato, M., Lopez, J.A., Macdonald, K., Vuck-
ovic, S., Hart, D.N.J., 2000. The role of dendritic cells in the
innate immune system. Microbes Infect. 2, 257–272.
Dzionek, A., Fuchs, A., Schmidt, P., Cremer, S., Zysk, M., Miltenyi,
S., Buck, D.W., Schmitz, J., 2000. BDCA-2, BDCA-3, and
BDCA-4: three markers for distinct subsets of dendritic cells
in human peripheral blood. J. Immunol. 165, 6037–6046.
Egner, W., McKenzie, J.L., Smith, S.M., Beard, M.E.J., Hart,
D.N.J., 1993. Human bone marrow contains potent stimulatory
cells for the allogeneic MLR with the phenotype of dendritic
cells. Adv. Exp. Med. Biol. 329, 263–268.
Fearnley, D.B., McLellan, A.D., Mannering, S.I., Hock, B.D., Hart,
D.N.J., 1997. Isolation of human blood dendritic cells using the
CMRF-44 monoclonal antibody: implications for studies on
antigen presenting cell function and immunotherapy. Blood
89, 3708–3716.
Ferlazzo, G., Wesa, A., Wei, W.Z., Galy, A., 1999. Dendritic cells
generated either from CD34+ progenitor cells or from mono-
cytes differ in their ability to activate antigen-specific CD8+T
cells. J. Immunol. 163, 3597–3604.
Feuerstein, B., Berger, T.G., Maczek, C., Roder, C., Schreiner, D.,
Hirsch, U., Haendle, I., Leisgang, W., Glaser, A., Kuss, O.,
Diepgen, T.L., Schuler, G., Schuler-Thurner, B., 2000. A method
for the production of cryopreserved aliquots of antigen-pre-
loaded, mature dendritic cells ready for clinical use. J. Immunol.
Methods 245, 15–29.
Fong, L., Engleman, E.G., 2000. Dendritic cells in cancer immuno-
therapy. Annu. Rev. Immunol. 18, 245–273.
Fong, L., Brockstedt, D., Benike, C., Wu, L., Engleman, E.G.,
2001. Dendritic cells injected via different routes induce immun-
ity in cancer patients. J. Immunol. 166, 4254–4259.
Furuta, M., Shimizu, T., Mizuno, S., Kamiya, T., Ozawa, K., Na-
kase, T., Tadokoro, K., Takenaka, M., Ohkawa, T., Yokoyama,
S., Ogawa, Y., Kiyokawa, H., Shimizu, M., Sekine, N., Yoshi-
mura, I., 1999. Clinical evaluation of repeat apheresis donors in
Japan. Vox Sang. 77, 17–23.
Gatti, E., Velleca, M.A., Biedermann, B.C., Ma, W., Unternaehrer,
J., Ebersold, M.W., Medzhitov, R., Pober, J.S., Mellman, I.,
2000. Large-scale culture and selective maturation of human
Langerhans cells from granulocyte colony-stimulating factor-
mobilized CD34+ progenitors. J. Immunol. 164, 3600–3607.
Gazitt, Y., 2000. Immunologic profiles of effector cells and periph-
eral blood stem cells mobilized with different hematopoietic
growth factors. Stem Cells 18, 390–398.
Geiger, J., Hutchinson, R., Hohenkirk, L., McKenna, E., Chang, A.,
J. Alejandro Lopez et al. / Journal of Immunological Methods 267 (2002) 199–212210

Mule, J., 2000. Treatment of solid tumours in children with
tumour-lysate-pulsed dendritic cells. Lancet 356, 1163–1165.
Glaser, A., Zingsem, J., Zimmermann, R., Weisbach, V., Eckstein,
R., 1999. Collection of mononuclear cells in the Spectra for the
generation of dendritic cells. Transfusion 39, 661–662.
Goxe, B., Latour, N., Chokri, M., Abastado, J.P., Salcedo, M., 2000.
Simplified method to generate large quantities of dendritic cells
suitable for clinical applications. Immunol. Invest. 29, 319–
336.
Hart, D.N.J., 1997. Dendritic cells: unique leucocyte populations
which control the primary immune response. Blood 90,
3245–3287.
Hart, D., Hill, G., 1999. Dendritic cell immunotherapy for cancer:
application to low-grade lymphoma and multiple myeloma. Im-
munol. Cell Biol. 77, 451.
Hart, D.N.J., Clark, G.J., MacDonald, K., Kato, M., Vuckovic, S.,
Lopez, J.A., Wykes, M., 2000. In: Mason, D. (Ed.), 7th Leu-
cocyte Differentiation Antigen Workshop DC Section Sum-
mary. Leucocyte Typing VII. Oxford Univ. Press, Oxford,
pp. 283–294.
Ho, C.S.K., Lopez, J.A., Vuckovic, S., Pyke, C.M., Hockey, R.L.,
Hart, D.N.J., 2001. Surgical and physical stress increase circu-
lating blood dendritic cell counts independently of monocyte
counts. Blood 98, 140–145.
Ho, C.S.K., Munster, D., Pyke, C.M., Hart, D.N.J., Lopez, J.A.,
2002. Spontaneous generation and survival of blood dendritic
cells in mononuclear cell culture without exogenous cytokines.
Blood 99, 2897–2904.
Hock, B.D., Starling, G.C., Daniel, P.B., Hart, D.N., 1994. Charac-
terization of CMRF-44, a novel monoclonal antibody to an
activation antigen expressed by the allostimulatory cells within
peripheral blood, including dendritic cells. Immunology 83,
573–581.
Hock, B.D., Fearnley, D.B., Boyce, A., McLellan, A.D., Sorg, R.V.,
Summers, K.L., Hart, D.N.J., 1999. Human dendritic cells ex-
press a 95 kDa activation/differentiation antigen defined by
CMRF-56. Tissue Antigens 53, 320–334.
Hsu, F.J., Benike, C., Fagnoni, F., Liles, T.M., Czerwinski, D.,
Taidi, B., Engleman, E.G., Levy, R., 1996. Vaccination of pa-
tients with B-cell lymphoma using autologous antigen-pulsed
dendritic cells. Nat. Med. 2, 52–57.
Kato, M., Neil, T.K., Fearnley, D.B., McLellan, A.D., Vuckovic, S.,
Hart, D.N., 2000. Expression of multilectin receptors and com-
parative FITC-dextran uptake by human dendritic cells. Int.
Immunol. 12, 1511–1519.
Kugler, A., Stuhler, G., Walden, P., Zoller, G., Zobywalski, A.,
Brossart, P., Trefzer, U., Ullrich, S., Muller, C.A., Becker, V.,
Gross, A.J., Hemmerlein, B., Kanz, L., Muller, G.A., Ringert,
R.H., 2000. Regression of human metastatic renal cell carcino-
ma after vaccination with tumor cell –dendritic cell hybrids. Nat.
Med. 6, 332–336.
Lewalle, P., Rouas, R., Lehmann, F., Martiat, P., 2000. Freezing of
dendritic cells, generated from cryopreserved leukaphereses,
does not influence their ability to induce antigen-specific im-
mune responses or functionally react to maturation stimuli. J.
Immunol. Methods 240, 69–78.
MacDonald, K.P.A., Munster, D., Clark, G.C., Dzionek, A.,
Schmitz, J., Hart, D.N.J., 2002. Characterization of human
blood dendritic cell subsets. Blood (in press).
Mackensen, A., Herbst, B., Kohlter, G., Wolff-Vorbeck, G., Rosen-
thal, F., Veelken, H., Kulmburg, P., Schaefer, H.E., Mertels-
mann, R., Lindemann, A., 1995. Delineation of the dendritic
cell lineage by generating large numbers of birbeck granule-
positive Langerhans cells from human peripheral blood progen-
itor cells in vitro. Blood 86, 2699–2707.
Maraskovsky, E., Daro, E., Roux, E., Teepe, M., Maliszewski, C.R.,
Hoek, J., Caron, D., Lebsack, M.E., McKenna, H.J., 2000. In
vivo generation of human dendritic cell subsets by Flt3 ligand.
Blood 96, 878–884.
McLellan, A.D., Starling, G.C., Hart, D.N.J., 1995. Isolation of
human blood dendritic cells by discontinuous Nycodenz gra-
dient centrifugation. J. Immunol. Methods 184, 81–89.
Mehta-Damani, A., Markowicz, S., Engleman, E.G., 1994. Genera-
tion of antigen-specific CD8+ CTLs from naive precursors. J.
Immunol. 153, 996–1003.
Menedez, P., Prosper, F., Bueno, C., Arbona, C., San Miguel, J.F.,
Garcia-Conde, J., Sola, C., Hornedo, J., Cortes-Funes, H., Or-
fao, A., 2001. Sequential analysis of CD34+ and CD34-cell
subsets in peripheral blood and leukapheresis products from
breast cancer patients mobilized with SCF plus G-CSF and
cyclophosphamide. Leukemia 15, 430–439.
Monji, M., Tynes-Petersons, J., Saund, N.J., Vuckovic, S., Hart,
D.N.J., Auditore-Hargreaves, K., Risdon, G., 2002. Dendritic cell
progenitors are CMRF-44+. Immunol. Cell Biol. 80, 216–225.
Murphy, G.P., Tjoa, B.A., Simmons, S.J., Ragde, H., Rogers, M.,
Elgamal, A., Kenny, G.M., Troychak, M.J., Salgaller, M.L.,
Boynton, A.L., 1999. Phase II prostate cancer vaccine trial:
report of a study involving 37 patients with disease recurrence
following primary treatment. Prostate 39, 54–59.
Nestle, F.O., Gilliet, M., Alljagic, S., Wiesner, W., Grabbe, S.,
Dummer, R., Burg, G., Schadendorf, D., 1997. Vaccination of
melanoma patients with peptide-pulsed dendritic cells. Melano-
ma Res. 7, S14.
Nestle, F.O., Banchereau, J., Hart, D., 2001. Dendritic cells: on the
move from bench to bedside. Nat. Med. 7, 761–765.
Olweus, J., BitMansour, A., Warnke, R., Thompson, P.A., Carballi-
do, J., Picker, L.J., Lund-Johansen, F., 1997. Dendritic cell on-
togeny: a human dendritic cell lineage of myeloid origin. Proc.
Natl. Acad. Sci. U. S. A. 94, 12551–12556.
Reid, C.D., Stackpoole, A., Meager, A., Tikerpae, J., 1992. Interac-
tions of tumour necrosis factor with granulocyte–macrophage
colony-stimulating factor and other cytokines in the regula-
tion of dendritic cell growth in vitro from early bipotent
CD34+ progenitors in human bone marrow. J. Immunol. 149,
2681–2688.
Rissoan, M.C., Soumelis, V., Kadowaki, N., Grouard, G., Briere,
F., de Waal Malefyt, R., Liu, Y.J., 1999. Reciprocal control of
T helper cell and dendritic cell differentiation. Science 283,
1183–1186.
Robinson, S.P., Patterson, S., English, N., Davies, D., Knight, S.C.,
Reid, C.D., 1999. Human peripheral blood contains two distinct
lineages of dendritic cells. Eur. J. Immunol. 29, 2769–2778.
Romani, N., Gruner, S., Brang, D., Kampgen, E., Lenz, A., Trock-
enbacher, B., Konwalinka, G., Fritsch, P.O., Steinman, R.M.,
J. Alejandro Lopez et al. / Journal of Immunological Methods 267 (2002) 199–212 211

Hansson, M., 1994. Proliferating dendritic cell progenitors in
human blood. J. Exp. Med. 180, 83–93.
Roth, M.D., Gitlitz, B.J., Kiertscher, S.M., Park, A.N., Mendenhall,
M., Moldawer, N., Figlin, R.A., 2000. Granulocyte macrophage
colony-stimulating factor and interleukin 4 enhance the num-
ber and antigen-presenting activity of circulating CD14+and
CD83+cells in cancer patients. Cancer Res. 60, 1934–1941.
Rouard, H., Leon, A., Klonjkowski, B., Marquet, J., Tenneze, L.,
Plonquet, A., Agrawal, S.G., Abastado, J.P., Eloit, M., Farcet,
J.P., Delfau-Larue, M.H., 2000. Adenoviral transduction of hu-
man ‘clinical grade’ immature dendritic cells enhances costimu-
latory molecule expression and T-cell stimulatory capacity. J.
Immunol. Methods 241, 69–81.
Sallusto, F., Lanzavecchia, A., 1994. Efficient presentation of solu-
ble antigen by cultured human dendritic cells is maintained by
granulocyte/macrophage colony-stimulating factor plus interleu-
kin 4 and downregulated by tumour necrosis factor-a. J. Exp.
Med. 179, 1109.
Sato, H., Shiobara, S., Yasue, S., Chuhjo, T., Nakao, S., 2001.
Lymphocyte collection for donor leucocyte infusion from nor-
mal donors: estimation of the minimum processed blood volume
and safety of the procedure. Vox Sang. 81, 124–127.
Schakel, K., Mayer, E., Federle, C., Schmitz, M., Riethmuller, G.,
Rieber, E.P., 1998. A novel dendritic cell population in human
blood: one-step immunomagnetic isolation by a specific mAb
(M-DC8) and in vitro priming of cytotoxic T lymphocytes. Eur.
J. Immunol. 28, 4084–4093.
Siena, S., Di Nicola, M., Bregni, M., Mortarini, R., Anichini, A.,
Lombardi, L., Ravagnani, F., Parmiani, G., Gianni, A.M., 1995.
Massive ex vivo generation of functional dendritic cells from
mobilized CD34+ blood progenitors for anticancer therapy. Exp.
Hematol. 23, 1463–1471.
Strunk, D., Rappersberger, K., Egger, C., Strobl, H., Kromer, E.,
Elbe, A., Maurer, D., Stingl, G., 1996. Generation of human
dendritic cell/Langerhans cells from circulating CD34+ hema-
topoietic progenitor cells. Blood 87, 1292–1302.
Summers, K.L., O’Donnell, J.L., Heiser, A., Highton, J., Hart, D.N.,
1999. Synovial fluid transforming growth factor beta inhibits
dendritic cell – T lymphocyte interactions in patients with
chronic arthritis. Arthritis Rheum. 42, 507–518.
Szabolcs, P., Moore, M.A.S., Young, J.W., 1995. Expansion of
immunostimulatory dendritic cells among the myeloid progeny
of human CD34+ bone marrow precursors cultured with c-kit
ligand, granulocyte–macrophage colony-stimulating factor, and
TNF-a. J. Immunol. 154, 5851–5861.
Tarte, K., Lu, Z.Y., Fiol, G., Legouffe, E., Rossi, J.F., Klein, B.,
1997. Generation of virtually pure and potentially proliferating
dendritic cells from non-CD34 apheresis cells from patients with
multiple myeloma. Blood 90, 3482–3495.
Thurner, B., Roder, C., Dieckmann, D., Heuer, M., Kruse, M.,
Glaser, A., Keikavoussi, P., Kampgen, E., Bender, A., Schuler,
G., 1999. Generation of large numbers of fully mature and stable
dendritic cells from leukapheresis products for clinical applica-
tion. J. Immunol. Methods 223, 1–15.
Vuckovic, S., Fearnley, D.B., Gunningham, S., Spearing, R.L., Pat-
ton, W.N., Hart, D.N., 1999. Dendritic cells in chronic myelo-
monocytic leukaemia. Br. J. Haematol. 105, 974–985.
Young, J.W., Szabolcs, P., Moore, M.A.S., 1995. Identification of
dendritic cell colony-forming units among normal human
CD34+ bone marrow progenitors that are expanded by c-kit
ligand and yield pure dendritic cell colonies in the presence of
granulocyte/macrophage colony-stimulating factor and tumor
necrosis factor a. J. Exp. Med. 182, 1120.
Zheng, C., Pisa, P., Stromberg, O., Blennow, E., Hansson, M., 2000.
Generation of dendritic cells from peripheral blood of patients at
different stages of chronic myeloid leukemia. Med. Oncol. 17,
270–278.
J. Alejandro Lopez et al. / Journal of Immunological Methods 267 (2002) 199–212212





























