Embed Size (px)
Citation preview

Journal of Ecology
2009,
97
, 472–483 doi: 10.1111/j.1365-2745.2009.01479.x
© 2009 Landcare Research. Journal compilation © 2009 British Ecological Society
Blackwell Publishing Ltd
Multi-stemmed trees in montane rain forests:
their frequency and demography in relation to elevation,
soil nutrients and disturbance
Peter J. Bellingham
1
* and Ashley D. Sparrow
2
1
Landcare Research, PO Box 40, Lincoln 7640, New Zealand; and
2
Department of Natural Resources
and Environmental Science, University of Nevada, Mail Stop 186, 1000 Valley Road, Reno, Nevada, 89152, USA
Summary
1.
Multi-stemmed trees are a common component of tropical and temperate montane rain forests,where they co-occur with single-stemmed trees. We hypothesized that multi-stemmed architectureshould predominate in less productive sites (such as at high elevation or with low soil nutrientavailability) and that, because it frequently results from sprouting, it should prevail in sites that arefrequently disturbed. We also hypothesized that where multi-stemmed architecture predominates,there should be lower rates of mortality and recruitment of individuals.
2.
We tested these hypotheses using data from permanent plots in tropical montane rain forests inJamaica (14 years) and in temperate montane rain forests in New Zealand (19 years). The frequencyof multi-stemmed trees varied across plots in both forests (4–34% in Jamaica; 0–21% in New Zealand)along gradients of elevation and soil nutrients.
3.
Analyses examined the direct and indirect effects of the various environmental variables that arelikely to drive site productivity and disturbance. We showed contrasting predictors of the frequencyof multi-stemmed trees in the tropical and temperate forests. Multi-stemmed trees in Jamaican forestswere most frequent on sites of low soil nutrient status (especially available soil phosphorus, P)whereas in New Zealand they were most frequent at high elevation sites, especially towards tree line.In both forests there was no relationship between multi-stemmed tree frequency and slope.
4.
Turnover (the mean of mortality and recruitment rates) of multi-stemmed trees in Jamaicanforests was lowest on sites of low soil nutrient status (especially available P) but was unrelated toenvironmental predictors in New Zealand. In both forests, turnover rates of multi-stemmed treesoverall were 60% lower than those for co-occurring single-stemmed trees, offering support for thehypothesis that multi-stemmed architecture favours persistence.
5.
Synthesis
. Our study demonstrates that multi-stemmed trees can predominate in low productivitysites in montane rain forests. Their low turnover compared with co-occurring single-stemmed treesconfirms the importance of evaluating the persistence niche as a mechanism promoting woodyspecies coexistence in forests.
Key-words:
Jamaica, mortality, New Zealand, recruitment, soil nitrogen, soil phosphorus,sprouting, structural equation model, temperature, tree line
Introduction
Multi-stemmed trees are common in many forests (Johnston &Lacey 1983; Del Tredici 2001). Multiple stems originate at ornear the ground (Givnish 1984; Bellingham & Sparrow 2000)and often arise by sprouting (Vesk & Westoby 2004; Fujiki &Kikuzawa 2006). Disturbance can promote multi-stemmed
architecture by stimulating growth of existing or new sproutsafter damage to existing stems (Peterson & Jones 1997); forexample, after wind storms (Bellingham
et al
. 1994; Miura &Yamamoto 2003) and after fire (Bond & van Wilgen 1996).Disturbance is not a requirement for multi-stemmedarchitecture to develop (Johnston & Lacey 1983; Woolley
et al
. 2008) and, in the absence of disturbance, replacement ofstems by new sprouts can benefit tree fitness through enhancedcarbon gain or sexual reproductive output (Chamberlin &
*Correspondence author. E-mail: [email protected]

Multi-stemmed trees in montane rain forests
473
© 2009 Landcare Research. Journal compilation © 2009 British Ecological Society,
Journal of Ecology
,
97
, 472–483
Aarsen 1996; Fujiki & Kikuzawa 2006). Whether promotedby disturbance or not, multi-stemmed architecture often enablespersistence at a site (Pigott 1989; Bond & Midgley 2001; DelTredici
et al
. 1992).Multi-stemmed trees are particularly prevalent in tropical
and temperate montane rain forests (van Steenis 1935; Midgley& Cowling 1993). Low productivity constrains maximumcanopy height in many montane rain forests below the 12-mthreshold above which multi-stemmed architecture is lessstructurally stable (McMahon 1973; Givnish 1984, 1995).Because height growth is relatively unconstrained in productivelowland forests, multi-stemmed architecture becomes lesscompetitive because multi-stemmed trees are shaded (Midgley1996). Multi-stemmed architecture in lowland forests isusually competitive in early succession where canopy heightsare low (Lutz & Halpern 2006), but can be maintained intaller forests that are frequently disturbed such as floodplainforests (e.g.
Cercidiphyllum japonicum
maintains a sprout bankaround a tall emergent stem in Japanese floodplain forests;Kubo
et al
. 2005). Multi-stemmed architecture can persist inlowland forest subcanopies when the canopies above are thin(Grier & Logan 1977) and also when they are intact, in whichcase multi-stemmed trees can trade off the smaller maximumheight they attain against more rapid growth rates and earliermaturation (Kohyama 1992).
Dominance of multi-stemmed trees in low-productivitysites was predicted in a theoretical model (Bellingham &Sparrow 2000) and is consistent with empirical evidence forforests (e.g. Kruger
et al
. 1997). However, the model requiresfurther testing because, in some moisture- or nutrient-limitedshrublands and grasslands, the capacity to sprout (often aprerequisite for multi-stemmed architecture) predominates inrelatively resource-rich sites (Clarke
et al
. 2005; Dalgleish &Hartnett 2006; but see Pausas & Bradstock 2007). Thereis little evidence to evaluate demographic patterns alongenvironmental gradients in forests where multi-stemmedarchitecture is prevalent. Most demographic studies in theseforests report the dynamics of stems only, even though manyarise from the same root stock and are not independent(e.g. Tanner & Bellingham 2006). There are few demographicevaluations of multi-stemmed woody plants (e.g. Tanner 2001;Fujiki & Kikuzawa 2006; Alfonso-Corrado
et al
. 2007) andthe significance of stems of sprout origin to plant demographyis often speculative (Harper 1977). A recent review suggestedthat assessments of the demography of woody plant stems ofsprout origin require evaluation over at least 5-year intervals(Cornelissen
et al
. 2003).In this study, we used long-term data from tropical (Jamaica,
14 years) and temperate (New Zealand, 19 years) montanerain forests to examine the environmental drivers of dominanceby multi-stemmed trees. Tropical montane rain forests andcool-temperate Southern Hemisphere rain forests arephysiognomically very similar (Richards 1952; Dawson &Sneddon 1969) but our sites have contrasting disturbanceregimes (see Methods). We present the first evaluation ofmortality and recruitment of multi-stemmed and single-stemmed individuals in rain forests. Our aim is to determine
whether there are consistent relationships between thedominance and dynamics of multi-stemmed trees, and siteproductivity and disturbance.
A conceptual model for multi-stemmed trees in
montane rain forests
Bellingham & Sparrow (2000) predicted greater frequencyof sprouting, and of multi-stemmed trees, in forests asproductivity declines and frequency of disturbance increases.In a montane rain forest, productivity is linked to gradientsof elevation (Kitayama & Aiba 2002; Moser
et al
. 2007) andfrequency of disturbance to topography, especially slope(Nagamatsu & Miura 1997). A conceptual model (Fig. 1) outlinesexpected influences on the frequency of multi-stemmed treesin rain forests.
Elevation exerts a direct influence on forest productivitybecause temperature decreases with increasing elevation.On temperate mountains, declining temperature limits treegrowth (Körner 1999; Körner & Paulsen 2004) such that attree line forest stature is often dominated by stunted multi-stemmed architecture (krummholz). New Zealand tree linesare very low (
c
. 1200–1300 m) compared with continental sitesat the same latitudes (
c
. 1800–3300 m; Körner & Paulsen2004; Wardle 2008), thus forest stature reduces over a narrowelevational range. Declining forest stature with increasingelevation on wet tropical mountains, as in Jamaica, is drivenby the combination of declining temperature and light, thelatter linked to increasing cloudiness with elevation (Bruijnzeel &Veneklaas 1998). Increasing cloudiness (and decreasing light)is further likely to limit forest stature on some wet temperatemountains (Wardle 1986). Elevation also exerts indirectinfluences on forest stature. In particular, nutrient availability,especially N and P, declines with increasing elevation on wettropical mountains (Grubb 1977) mediated by increasingprecipitation. Slower rates of decomposition with increasingaltitude also lead to increased soil C : N and declining pH(Grubb 1977). These indirect effects of elevation are likely tobe most pronounced on tropical mountains. On temperatemountains nutrients are not necessarily limiting at tree line;disturbance history (notably past glaciation) is a major deter-minant of nutrient supply rates (Prescott
et al
. 1992; Tonkin& Basher 2001; Bowman
et al
. 2003).As slope increases there is a higher incidence of ground–
surface disturbances. Landslides occur on steep slopes inboth the Jamaican and New Zealand sites (Dalling 1994; Reif& Allen 1988), but are more frequent in the New Zealandsites, driven by high rainfall events (Tonkin & Basher 2001)and periodic movements of the Alpine fault (Wells
et al
. 1999).Multi-stemmed architecture can be more prevalent as slopeinstability increases (Tang & Ohsawa 2002). Damage to treeson steep slopes caused by erosion of bedrock can promotesprouting; resources are remobilized from damaged above-ground parts to form new stems (Del Tredici
et al
. 1992; Sakai
et al
. 1997). Slope is also likely to exert indirect effects on theproportion of multi-stemmed trees by influencing nutrientavailability. On ridge crests with little slope, soil organic

474
P. J. Bellingham & A. D. Sparrow
© 2009 Landcare Research. Journal compilation © 2009 British Ecological Society,
Journal of Ecology
,
97
, 472–483
matter accumulates and C : N increases; P supply can beparticularly deficient (Grubb 1977; Richardson
et al
. 2008).On steep slopes, where primary successions occur, as on land-slides, P availability is relatively high and N availability verylow (Walker & del Moral 2003). Differential erosion of bedrockunder forest canopies according to steepness of slopes andlocal deposition also results in different levels of soil N and Pavailability (Vitousek
et al
. 2003).Other disturbances promote multi-stemmed architecture in
montane forests, in particular sprouting after wind disturbances.Hurricanes affect Jamaican montane forests on average every25 years and promote growth of sprouts to form new stems(Bellingham
et al
. 1994). In Jamaica, hurricane winds canaffect forests on ridge crests more than those on slopes(Bellingham 1991) and this could result in a greater frequencyof multi-stemmed trees on ridge crests. Less intense winddisturbances, including cyclones, affect New Zealand montaneforests and many dominant trees sprout after wind damage(Martin & Ogden 2006), but unlike Jamaica, the return intervalto most sites is > 100 years. Fire is part of the disturbanceregime in some temperate montane forests; some species sproutin response and form multi-stemmed trees (e.g. riparianangiosperms in largely coniferous North American forests;Pettit & Naiman 2007). However, globally very few montanerain forests are affected by fire. Fire disturbance regimes inthose forests are predominantly anthropogenic and havealtered the forests’ composition (e.g. Corlett 1984). Fire is notpart of the disturbance regime in New Zealand temperatemontane rain forests (Ogden
et al
. 1998). Jamaican montanerain forests also lack a history of fire disturbance, andanthropogenic fires at their margins result in their destruction(McDonald
et al
. 2002).As a step towards a comprehensive test of the model (Fig. 1),
we tested some of the key components, that is the effects
of elevation, soil nutrients, and slope (the latter a proxyfor one component of the disturbance regime) on multi-stemmed architecture and the dynamics of multi-stemmedtrees (Fig. 2).
1.
Multi-stemmed architecture in rain forests will becomemore prevalent as productivity declines (Bellingham &Sparrow 2000). Multi-stemmed architecture will also beprevalent in sites subject to frequent disturbance (Nzunda
et al
. 2007a), such as steep, unstable slopes where multiplestems are likely to be a response to uprooting and physicaldamage (Sakai
et al
. 1997; Nzunda
et al
. 2007b).
2.
Population turnover (the mean of mortality and recruitmentrates) of both single- and multi-stemmed individuals will belowest on the lowest productivity sites (Russo
et al
. 2005;Stephenson & van Mantgem 2005). In low-productivity sites,where we expect multi-stemmed architecture to predominate,there will be lower population turnover of multi-stemmedindividuals than of single-stemmed individuals. If the turnoverof multi-stemmed trees is lower than that of single-stemmedindividuals in the same site (Bond & Midgley 2003; Zobel2008), then this would give further support to multi-stemmedarchitecture being an expression of a ‘persistence niche’ (Bond &Midgley 2001).
Our approach to examining these hypotheses was to fitstructural equation models (SEMs; Grace 2006) because thisanalysis is appropriate for examining the direct and indirecteffects of the various environmental variables that are likelyto drive site productivity and disturbance. In species-richrain forests, a possible confounding influence is that thepropensity of species to sprout or form multi-stemmedindividuals is not uniform across species (Bellingham
et al
.1994) and species are not uniformly distributed alongenvironmental and disturbance gradients (Tanner 1977;Reif & Allen 1988). Therefore we also tested whether there are
Fig. 1. Complete conceptual model to test hypotheses of drivers of dominance and dynamics of multi-stemmed trees in rain forests.

Multi-stemmed trees in montane rain forests
475
© 2009 Landcare Research. Journal compilation © 2009 British Ecological Society,
Journal of Ecology
,
97
, 472–483
phylogenetic gradients that confounded our ability to test ourhypotheses.
Methods
STUDY
S ITES
Tropical montane forests in this study were in the western BlueMountains, Jamaica (18
°
05
′
N, 76
°
38–40
′
W). The geology of themountains is complex, with at least eight rock types in highly faultedstrata including schists, shales, amphibolites and, locally, limestone.Rainfall is
c.
2800 mm year
−
1
, with a relatively dry period during July–August (Shreve 1914). Composition of the lower and upper montanerain forests reflects elevation, variation in soils, and moisture gradientsaccording to the prevailing trade winds, (Shreve 1914; Grubb & Tanner1976). Primary successions occur on landslides (Dalling 1994) andsecondary successions occur after hurricanes (Tanner & Bellingham2006).
Temperate montane forests in this study were in the WhitcombeRiver valley on the western side of the Southern Alps, South Island,
New Zealand (43
°
3–7
′
S, 170
°
58
′
–171
°
3
′
E). Geology in the site is schistand rainfall is
c
. 7000–8000 mm year
−
1
, distributed evenly throughoutthe year with many high-intensity events. Forest composition changeswith elevation, variation in soils with topography, and with disturbance.Primary successions on landslides and floodplains are commonplace(Reif & Allen 1988) and secondary successions occur after death ofold-growth canopies (Bellingham & Lee 2006).
PERMANENT
PLOT
MEASUREMENTS
We used data from permanently-tagged stems, all identified tospecies,
≥
3 cm d.b.h. (diameter at 1.3 m) in permanent plots in bothsites. In each plot, measurements were recorded for all tagged stems,including single-stemmed individuals and multi-stemmed individuals,that is, those that arose from a common rootstock and which wereseparate below the point of measurement (1.3 m). Thus we recordeddiameter data, mortality and recruitment for stems and for individuals(both single- and multi-stemmed).
Jamaican tropical montane forests were sampled using 16 permanent200-m
2
plots measured in 1990 and 2004 (mean census interval14.0 ± 0.02 year). The plots were placed at regular intervals, samplingthe northern windward slopes, the crest, and the southern leewardslopes of the mountains within
c
. 2.5 km
2
(1300–1900 m a.s.l.; locationsmostly shown in Bellingham 1991). There were no major disturbancesto the forests during the census period, but Hurricane Gilbert in1988 had disturbed forests in the plots (Bellingham 1991) and this islikely to have affected demographic processes during the current study(Tanner & Bellingham 2006). The New Zealand temperate montaneforests were sampled using 400-m
2
plots established along randomcompass lines from valley floor to tree line: 23 plots along 8 lines(250–850 m), measured in 1980 and 1999 (mean census interval18.9 ± 0.02 year). Tropical montane forests in Jamaica were located1100 m higher in elevation than the New Zealand forests (Table 1)but the spanned elevation range was similar. The Jamaican tropicalforests had higher tree diversity than the New Zealand forests (higherFisher’s
α
, Shannon H
′
and higher
β
diversity; Table 1 and Table S1in Supporting Information).
In all plots, we recorded elevation and slope, and sampled soils.Soils, including organic horizons, were sampled to 10 cm depth at asingle random point in each Jamaican plot, and to 15 cm depth ineach New Zealand plot (mean values from four random samples perplot). Soils were air-dried, sieved then analysed for pH, N, C andavailable P (Bray 1-extractable P from Jamaica, Olsen P from NewZealand).
DATA
ANALYSIS
We computed the percentage of individuals that were multi-stemmedin each plot in both forests at the earliest census, and computed themean number of stems per multi-stemmed individual in each plot.We carried out detrended correspondence analysis (DCA) of the basalarea contributed by plant orders in each plot at the earliest census(using CANOCO for Windows version 4.54, ter Braak &
S
milauer1997) to determine whether there were phylogenetic gradientsalong major environmental gradients (orders follow Heywood
et al
.2007).
Mortality and recruitment rates of stems and individuals betweencensuses were determined for each plot in each forest. We alsodetermined these rates for both multi-stemmed and single-stemmedindividuals in each plot; 3 Jamaican plots and 9 New Zealandplots, where multi-stemmed individuals
n
≤
10 at the earlier census,were excluded from these analyses. Mortality as percentage is 100
Fig. 2. Models to test hypotheses for relationships between quantifiedenvironmental variables and (a) frequency of multi-stemmed individualsand (b) turnover (average of mortality and recruitment) of multi- andsingle-stemmed individuals.

476
P. J. Bellingham & A. D. Sparrow
© 2009 Landcare Research. Journal compilation © 2009 British Ecological Society,
Journal of Ecology
,
97
, 472–483
(
m
= 1
−
[1
−
(
N
0
−
N
1
)/
N
0
]
1/
t
), where
N
0
= the number at the earliestcensus,
N
1
= the number of survivors at the most recent census, and
t
= time in years. Recruitment as percentage is 100 (
r
= 1
−
(1
−
N
r
/
N
t
)
1/
t
), where
N
r
= the number recruited between censuses,
N
t
= thenumber at the most recent census. We also report annual turnoverrates as the average of annual mortality and recruitment rates ineach plot (e.g. Phillips & Gentry 1994; Stephenson & van Mantgem2005).
Hypotheses were tested by fitting data to the conceptual model asSEMs (Grace 2006) using AMOS student version 5.0.1 software(AMOS Development Corp., 2003). Before analysis, variables werelog-transformed as required for meeting assumptions of normality.Before fitting the SEMs, environmental data were tested for correlation(Table S2) and analysed by principal components analysis, separatelyfor both forests, using S-plus version 8.0.4 software (Insightful Corp.2007). Separate model predictions were made for abundance of multi-stemmed individuals (hypothesis 1; Fig. 2a) and turnover ofmulti-stemmed and single-stemmed individuals and the ratio ofthese turnovers (hypothesis 2; Fig. 2b). Analyses were conducted forall species combined. We also tested within-species patterns in thefrequency of multi-stemmed individuals. Only two species, both inNew Zealand, were sufficiently widespread and abundant (with
n
≥
10individuals in
n
≥
10 plots at the earliest census) to conduct thesetests. These were the canopy tree
Weinmannia racemosa
(Cunoniaceae)and the understorey tree
Pseudowintera colorata
(Winteraceae). Nospecies was sufficiently frequent to examine within-species relationshipsin Jamaica. SEM pathways were forced simplifications of the conceptualmodel that excluded all latent (i.e. unmeasured, but inferred) variablesbecause the relatively small number of plots in each forest constrainedthe total degrees of freedom available for model fitting. Models werefitted using maximum likelihood estimation and additional linkageswere added to models where overall model diagnostics indicatedsignificant covariance not accounted for by the preliminary model(Grace 2006; Table S2). For ease of comparison between forests, fittedSEMs were not further simplified by removal of non-significantrelationships. SEMs are presented as diagrams in the same manneras in Vile
et al
. (2006), with the unstandardized regression coefficient and
its significance level shown for each relationship, and the proportionof total variance explained (
R
2
) given for the final response variable(i.e. frequency of multi-stemmed trees or turnover of trees).
Results
MULTI
-
STEMMED
TREES
AND
RELATIONSHIPS
WITH
ELEVATION
,
SOIL
NUTRIENTS
AND
DISTURBANCE
Multi-stemmed trees were nearly twice as frequent in tropicalmontane forests in Jamaica as they were in temperatemontane forests in New Zealand (Table 1). The frequency ofmulti-stemmed trees (%) was strongly positively related toelevation in temperate montane forests in New Zealand (allspecies combined, Fig. 3b) but was unrelated to elevation inJamaican tropical montane rain forests (Fig. 3a). In temperatemontane rain forests in New Zealand, multi-stemmed P. coloratatrees increased in frequency with elevation (as predicted), butthe opposite was the case for W. racemosa (Fig. S1).
The frequency of multi-stemmed trees was negatively relatedto Bray-extractable soil P in tropical montane forests inJamaica, as predicted by the conceptual model (Fig. 3a), butin temperate montane forests in New Zealand the frequencyof multi-stemmed trees (across all species) was positivelyrelated to soil Olsen P (Fig. 3b), contrary to prediction.Soil N and other soil variables were unrelated to the frequencyof multi-stemmed trees in both forests. In New Zealand,relationships between the frequency of multi-stemmedW. racemosa trees and soil pH, C : N and Olsen P wereconsistent with our model predictions (although againstprediction in relationship to soil N; Fig. S1). In contrast,multi-stemmed P. colorata tree frequency was positively relatedto soil pH and negatively to soil C : N ratio (both contraryto prediction). Soil C : N was lower in Jamaican tropical
Table 1. Soil and site characteristics, tree diversity, characteristics of multi-stemmed individuals and demography of individuals in tropical(Jamaica) and temperate (New Zealand) montane rain forests: mean ± SE (range)
Jamaica New Zealand
Soil and site characteristicsElevation 1680 ± 44 (1300–1920) 560 ± 40 (250–850)Slope 29 ± 3.5 (6–45) 26 ± 3.0 (2–44)Soil pH 4.2 ± 0.21 (3.1–5.7) 4.1 ± 0.09 (3.3–5.2)Soil C : N 13.8 ± 0.51 (10.6–17.4) 20.3 ± 0.92 (13.8–31.6)Soil N (%) 1.26 ± 0.09 (0.54–1.86) 0.71 ± 0.11 (0.17–1.79)Soil P (mg kg−1)* 24.4 ± 3.7 (10.0–54.0) 18.4 ± 2.3 (8.8–50.6)
Tree diversityAlpha 10.95 ± 1.23 (4.91–20.09) 3.81 ± 0.22 (1.78–6.37)Shannon H′ 2.65 ± 0.09 (1.89–3.13) 1.92 ± 0.22 (1.48–2.35)Beta (DCA length; half-change units) 3.7153 3.3496
Multi-stemmed individualsPercentage of multi-stemmed individuals 15.7 ± 1.9 (3.8–34.0) 8.6 ± 1.1 (0–20.6)Stems/multi-stemmed individuals 2.6 ± 0.07 (2.3–3.5) 2.3 ± 0.07 (2.0–3.2)
Demography of individualsMortality (% per year)† 1.90 ± 0.21 (0.92–3.20) 1.56 ± 0.21 (0.77–3.82)Recruitment (% per year)† 2.09 ± 0.26 (0.69–3.59) 1.95 ± 0.42 (0.16–5.69)
*Soil P = Bray-extractable P for Jamaica and Olsen-extractable P for New Zealand.†Calculated excluding three plots in Jamaica and nine plots in New Zealand where multi-stemmed individuals n ≤ 10 at t0.
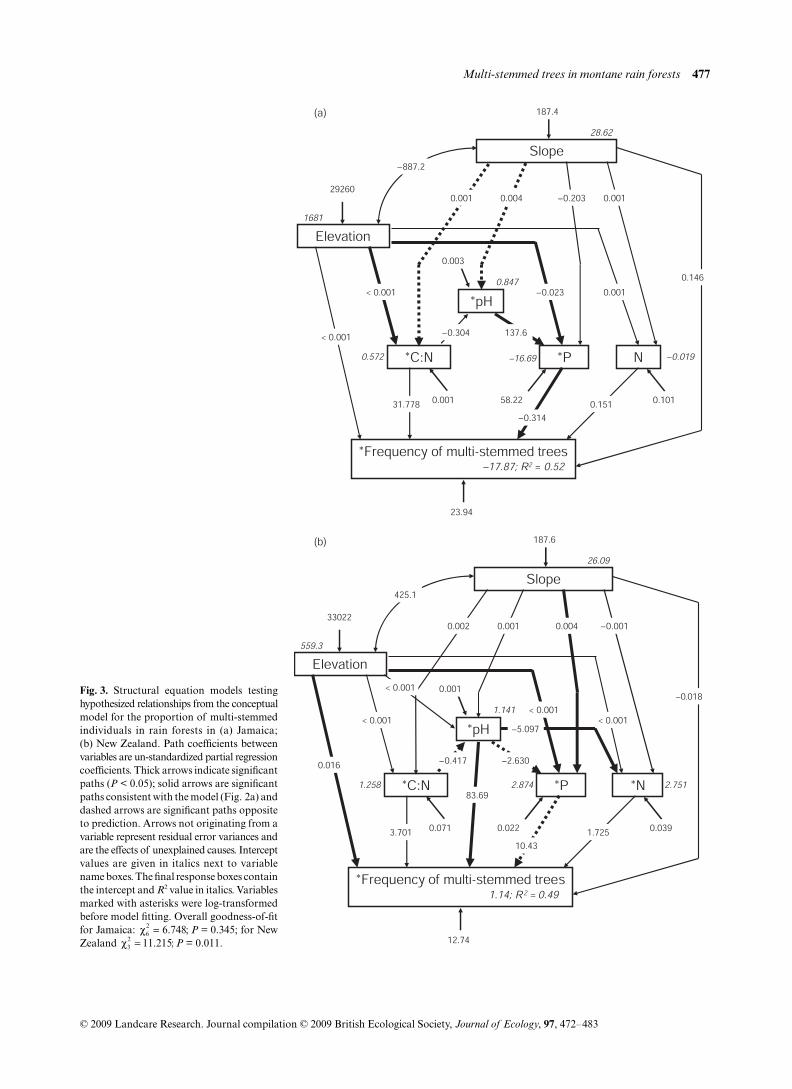
Multi-stemmed trees in montane rain forests 477
© 2009 Landcare Research. Journal compilation © 2009 British Ecological Society, Journal of Ecology, 97, 472–483
Fig. 3. Structural equation models testinghypothesized relationships from the conceptualmodel for the proportion of multi-stemmedindividuals in rain forests in (a) Jamaica;(b) New Zealand. Path coefficients betweenvariables are un-standardized partial regressioncoefficients. Thick arrows indicate significantpaths (P < 0.05); solid arrows are significantpaths consistent with the model (Fig. 2a) anddashed arrows are significant paths oppositeto prediction. Arrows not originating from avariable represent residual error variances andare the effects of unexplained causes. Interceptvalues are given in italics next to variablename boxes. The final response boxes containthe intercept and R2 value in italics. Variablesmarked with asterisks were log-transformedbefore model fitting. Overall goodness-of-fitfor Jamaica: P = 0.345; for NewZealand P = 0.011.
χ62 6 748 . ;=
χ32 11 215 . ;=

478 P. J. Bellingham & A. D. Sparrow
© 2009 Landcare Research. Journal compilation © 2009 British Ecological Society, Journal of Ecology, 97, 472–483
montane forests than in New Zealand temperate montaneforests, and soil N% was greater in Jamaica (Table 1). SoilpH was similar in both forests. Available soil P was notcomparable between the Jamaican and New Zealand sitesbecause different measures of extractable P were used(Bray-extractable in Jamaica, Olsen in New Zealand).
Slope, a proxy for frequency of ground–surface disturbance,was similar in both forests and there was no relationshipbetween multi-stemmed tree frequencies across all speciescombined and slope in either forest (Fig. 3). Of the twocommon New Zealand trees, the frequency of multi-stemmedW. racemosa trees increased with slope (consistent with themodel) but there was no relationship in the case of P. colorata(Fig. S1).
PHYLOGENIES OF MULTI-STEMMED TREES AND RELATIONSHIPS WITH ENVIRONMENT
Plant orders differed in their percentages of multi-stemmedindividuals in both forests (range in Jamaica 0.2–23.8%; NewZealand 0–22.2%; Table S1, Fig. S2). However, the percentageof multi-stemmed individuals in plant orders was not correlatedwith the DCA Axis 1 score of the basal area of orders in any ofthe forests (Jamaica r = 0.36, P = 0.22; New Zealand r = 0.32,P = 0.44; Fig. S2). We interpret this to mean that althoughdifferent plant orders dominated along major environmentalgradients in each of the forests, there was no phylogenetic biasin the frequency with which multi-stemmed trees occurredalong the same gradient.
EFFECTS OF ELEVATION, SOIL NUTRIENTS, AND DISTURBANCE ON TREE TURNOVER
The turnover of both multi- and single-stemmed trees intropical montane forests in Jamaica was positively related toelevation (contrary to prediction; Fig. 4), but as elevationincreased, the turnover of multi-stemmed trees relative tosingle-stemmed trees in the same site declined (Fig. 4),consistent with our predictions. In temperate montane rainforests in New Zealand there were no relationships betweentree turnover and elevation (single- or multi-stemmed trees).
Turnover of multi- and single-stemmed trees was positivelyrelated to soil Bray-extractable P (as predicted) in tropicalmontane forests in Jamaica (Fig. 4). Turnover of multi- andsingle-stemmed trees in Jamaica was negatively related tosoil C : N (as predicted), although the relationship for multi-stemmed tree turnover relative to that of single-stemmed treesincreased, contrary to prediction (Fig. 4). In New Zealandtemperate montane forests, the SEMs fitted poorly to treeturnover data: soil Olsen P and C : N had only weak positiveeffects on turnover of single-stemmed trees, both contrary tomodel predictions. Multi- and single-stemmed tree turnoverwas unrelated to slope in tropical and temperate montaneforests.
Mortality and recruitment rates of individuals (combiningsingle- and multi-stemmed individuals) were similar in thetropical and temperate rain forests (Fig. S3). Multi-stemmedindividuals had much lower turnover (average of mortalityand recruitment) rates than single-stemmed individuals in
Fig. 4. Structural equation model testinghypothesized relationships from the conceptualmodel for the turnover of multi-stemmedindividuals in Jamaican montane rain forests(no significant relationships were found inNew Zealand rain forests). Path coefficientsbetween variables are un-standardized partialregression coefficients. Thick arrows indicatesignificant paths (P < 0.05); solid arrows aresignificant paths consistent with the model(Fig. 2b) and dashed arrows are significantpaths opposite to prediction. Other SEMdiagram annotations as in Fig. 3. Overallgoodness-of-fit P = 0.159.χ6
2 9 266 . ;=

Multi-stemmed trees in montane rain forests 479
© 2009 Landcare Research. Journal compilation © 2009 British Ecological Society, Journal of Ecology, 97, 472–483
both forests (Fig. 5). Typically, turnover rates are reported forstems rather than individuals. In both forests, turnover rateswere similar for stems and for all individuals (combiningsingle- and multi-stemmed trees) because more abundantsingle-stemmed individuals diluted the contribution of slowturnover of rarer multi-stemmed individuals, althoughindividual turnover rates were lower than stem turnoverrates in Jamaica (Fig. S3).
Discussion
MULTI-STEMMED TREES AND RELATIONSHIPS WITH ELEVATION, SOIL NUTRIENTS AND DISTURBANCE
The frequency of multi-stemmed trees in both the tropicaland the temperate montane rain forests was driven by just onekey environmental variable and one SEM pathway. However,the single environmental determinant differed between thetropical and the temperate forest. As elevation increased intemperate montane forests in New Zealand, multi-stemmedindividuals were more frequent. This is consistent with ourexpectation that temperature (declining with elevation) ratherthan soil nutrients would be the principal determinant intemperate forests. It is also consistent with expectations ofgreater frequency of krummholz architecture as temperaturelimits forest growth near tree line (Körner 1999). This patternwas apparent for one of two common trees in New Zealand(P. colorata) and, similarly, multi-stemmed Nothofaguscunninghamii trees increased in frequency with elevation in cooltemperate Australian rain forests (Johnston & Lacey 1983).
In contrast, elevation had no effect on the frequency ofmulti-stemmed trees in tropical montane forests in Jamaica.
Instead, multi-stemmed architecture was more frequent asavailable P (mediated by pH and slope) declined. This isconsistent with the theory that P, as well as N, limits pro-ductivity and forest stature on tropical mountains (Grubb1977; Tanner et al. 1992, 1998; Moser et al. 2007). That thepathway is mediated by slope (Fig. 3a) is again consistentwith general models of P availability in montane forests.On flat or gently sloping terrain, high precipitation and poordrainage lead to soil podsolisation and low P availability(Grubb 1977), whereas on steeper terrain, erosion exposesbedrock and thereby brings new supplies of P to the surface(Vitousek et al. 2003; Porder et al. 2007). Our results alsosuggest that in some temperate forests P supply can dictatethe frequency of multi-stemmed trees at lower elevations.Multi-stemmed W. racemosa trees in New Zealand weremost frequent on low P soils and at low elevation, the latterin contrast to the trend across all species combined. Moreinvestigations are needed in low-stature but less temperature-limited temperate forests to determine whether soil nutrientlimitation, especially of P, determines architecture, forexample across nutrient gradients or soil chronosequences(Wardle et al. 2004).
Multi-stemmed architecture can be a response to chronicdisturbance regimes (Smale 1994). For example, on forestson unstable dunes in subtropical South Africa, 39% ofindividuals were multi-stemmed (Nzunda et al. 2007a), amuch greater frequency than in either of the forests in ourstudy. Multi-stemmed trees are often common on steep slopes,where many trees have a high sprouting capacity to counteractground–surface disturbances and can obtain resources forsprouting from their above-ground parts that often survivedisturbances (Del Tredici et al. 1992; Sakai et al. 1995, 1997;Nzunda et al. 2007b). However, slope was not a direct predictorof abundance of multi-stemmed trees in either the tropical orthe temperate montane forest in our study; instead its effectswere indirect (by mediating nutrient availability, see above).Our interpretation of slope as a driver of disturbance ishampered by our inability to discriminate between long-termfrequency of disturbance and time since last disturbance, twovariables that are not orthogonal and that are often confoundedin non-experimental studies of disturbance ecology (e.g. Connell1978). More detailed investigations of soil profiles or analysisof isotopes in our study sites could reveal time since large-scaleprimary disturbance in plots (e.g. Hewitt 1996), a superiormeasure of disturbance frequency and, in turn, perhaps apredictor of multi-stemmed architecture.
EFFECTS OF ELEVATION, SOIL NUTRIENTS, AND DISTURBANCE ON TREE TURNOVER
Our data from forests, where up to 34% of individuals aremulti-stemmed, demonstrate that there is a small difference inreporting stand dynamics at an individual level vs. at a stemlevel (Fig. S3). In contrast to overall stand dynamics patterns,it is clear that turnover of multi-stemmed individuals wassubstantially less than that of single-stemmed individuals(Fig. 5). This result offers strong support for the hypothesis
Fig. 5. Annual mortality and recruitment rates of single-stemmedindividuals (closed symbols) and multi-stemmed individuals (opensymbols) in Jamaican (circle; n = 13 plots) and New Zealand (triangle;n = 14) montane rain forests. Values are means across plots ± SEM.In both forests, turnover rates (average of mortality and recruitmentrates) of multi-stemmed individuals were less than those of single-stemmed individuals (paired t-tests; P ≤ 0.005).

480 P. J. Bellingham & A. D. Sparrow
© 2009 Landcare Research. Journal compilation © 2009 British Ecological Society, Journal of Ecology, 97, 472–483
that multi-stemmed architecture in trees, especially whenmaintained by sprouting, enables long-term persistence ofindividuals (Bond & Midgley 2001; Vesk & Westoby 2004).Persistence after major disturbance has been shown inmontane forests: a hurricane caused low turnover of individualsin the very nutrient-limited Jamaican Mor Ridge forestbecause most trees were multi-stemmed and death of thesestems was offset by recruitment of newly sprouted stems(Tanner & Bellingham 2006). Multi-stemmed architectureresults in lower population dynamism and greater persistence,but an implication of our study is that population dynamicsare likely to be variable within species. Multi-stemmedarchitecture in two widespread trees in New Zealandmontane forests ranged in frequency among plots (0–23% forP. colorata and 0–35% for W. racemosa), and this is also thecase for other trees (Cao & Peters 1998; Nanami et al. 2004).Moreover, within species there can be genetic differencesbetween populations that are predominantly multi-stemmedcompared with those that are single-stemmed, thus multi-stemmed architecture in local populations may be reinforced bylimited gene flow (Steinke et al. 2008). Because of variabilitywithin species, assignment of species traits with respect topersistence may not be as readily achieved in montane rainforests as in other systems where sprouting is a common responseafter disturbance, especially fire (e.g. Cornelissen et al. 2003;Pausas & Lavorel 2003; Klimesova & Klimes 2007).
In generally P-limited tropical montane forests (see above),sites with low available soil P are likely to be among the leastproductive sites. Thus our finding that the lowest turnover oftrees, both single- and multi-stemmed, in Jamaican montaneforests is in sites with the lowest soil Bray-extractable P, isconsistent with other studies which have shown lowestturnover rates in forests on sites of low productivity (Russoet al. 2005; Stephenson & van Mantgem 2005). Highermortality of multi-stemmed trees in more productive sites,as in Jamaica (i.e. with high soil Bray-extractable P), is alsoconsistent with studies which have shown that there can behigh mortality, even of tree species that sprout, in high-productivity sites (Lutz & Halpern 2006). Overall, however,there was little support for our conceptual model linkingmulti-stemmed tree turnover to environmental variables andin the New Zealand temperate forests the variance explainedwas low. More data on turnover of multi-stemmed treesfrom other forests will be needed to determine links betweenpersistence (Bond & Midgley 2001) and site productivity.
L IGHT AND INFREQUENT DISTURBANCES AS INFLUENCES ON MULTI-STEMMED TREES
Our original model emphasized that site productivity is a keydeterminant of sprouting in woody plants (Bellingham &Sparrow 2000). In the test of the model for multi-stemmedtrees in rain forests, we have only examined one component ofsite productivity – soil nutrients – as a driver of multi-stemmedarchitecture and turnover (Fig. 1). Light availability is anotherkey component of productivity which probably explains someof the variance not accounted for in our tests of the model
(Figs 3 and 4). Elevation can provide a coarse surrogate forlight availability in temperate and tropical mountains becauseas elevation increases there is increasing cloudiness thatresults in lower light levels and dominance of diffuse radiation(Wardle 1986; Bruijnzeel & Veneklaas 1998). This oversimpli-fies matters because light competition is mediated by soilresource availability (sensu Grime 2002). Forest stature, leafsize and leaf area indices in tropical montane rain forests alldecline with increasing elevation as a result of nutritionalconstraints (Grubb & Tanner 1976; Tanner et al. 1998;Kitayama & Aiba 2002; Moser et al. 2007), thus light levelscan be higher below canopies on mountaintops with lowestsoil nutrient availability, as is the case in Jamaica (Bellinghamet al. 1996). Similarly, leaf area and stature decline withelevation and decreasing temperature towards tree lines ontemperate mountains. While our model predicts that low lightlevels and reduced productivity should favour multi-stemmedarchitecture, Dunphy et al. (2000) proposed that high lightlevels promote multi-stemmed architecture in tropical dryforests. Therefore further tests of our model will require directmeasurements of light availability in plots.
Disturbance is also a central driver of sprouting and multi-stemmed architecture in our original model (Bellingham &Sparrow 2000; Fig. 1). Chronic, low-level disturbance as aresult of soil movement on steep slopes did not predict theincidence of multi-stemmed architecture in our two sites,but large periodic disturbances are probably influential.Hurricanes promote sprouting and favour persistence ofmulti-stemmed trees in some Jamaican sites (Tanner &Bellingham 2006). They also change resource availability,for example increased light levels for up to 2 years afterwards(Bellingham et al. 1996), which may alter competitive balancesbetween growth of single-stemmed vs. multi-stemmed trees.Less frequent but larger-magnitude tectonic disturbances inboth sites are likely to favour multi-stemmed architectureduring early succession (Reif & Allen 1988) and also alter siteproductivity substantially (Walker & del Moral 2003). Periodicfire, too, is likely to drive multi-stemmed architecture in manyother forests. Further tests of our model would benefit throughincorporation of documented or modelled frequency andintensity of periodic disturbances at each plot.
Conclusions
We have shown that multi-stemmed architecture is drivenprincipally by soil nutrient availability on tropical mountainsand by elevation as a likely index of temperature on temperatemountains. It may be that lower light availability, because ofhigher elevation, has a stronger effect on multi-stemmedarchitecture than we have realised. Future tests of our model(Fig. 1) will require determination of the circumstances inwhich the frequency of multi-stemmed trees is related to lightcompetition and/or low-temperature stress. This could bedetermined along latitudinal gradients and should considerforests where cold-temperature extremes are greater than thoseencountered in New Zealand forests (e.g. rain forests at highlatitudes in South America, Barrera et al. 2000).

Multi-stemmed trees in montane rain forests 481
© 2009 Landcare Research. Journal compilation © 2009 British Ecological Society, Journal of Ecology, 97, 472–483
Multi-stemmed trees have lower turnover than single-stemmed trees and hence longer generation times. When longergeneration times result in long-term persistence of specieswith poor litter quality and slow litter decomposition rates,then this is likely to affect the rate of carbon and nutrientcycling. For example, the montane Mor forests in Jamaica aredominated by multi-stemmed trees of species that producelitter that decomposes more slowly than that of nearby forestswith fewer multi-stemmed trees (Tanner 1981; Tanner &Bellingham 2006) and in the Mor forests c. 50-cm deep, acidichumus accumulates with a soil C : N twice as that of soils innearby forests (Tanner 1977; Tanner & Bellingham 2006).Longer generation times could also result in lower speciationrates in genera or families with high frequency of multi-stemmedarchitecture compared with shorter generation times forco-occurring genera with high frequency of single-stemmedarchitecture (Rohde 1992; see also Wells 1969). There is noevidence that this is the case in fire-prone Mediterraneanshrublands worldwide; speciation rates of persistent sprouters,with long-generation times, are not different from those ofseeders (Bond & Midgley 2003; Lamont & Wiens 2003).However, persistence through multi-stemmed architecturemay be a reason for low levels of speciation in some genera inmontane forests where the disturbance regimes are less severeand frequent than in Mediterranean shrublands. Monotypicgenera in montane forests with high frequencies of multi-stemmed architecture include Cyrilla (Jamaica), andDavidia, Cercidiphyllum and Tetracentron (China; Tang &Ohsawa 2002). This is unlikely to be a general explanationbecause they co-occur with other species-rich genera thatalso have high frequencies of multi-stemmed architecture(e.g. Vaccinium in Jamaica and Prunus in China; Tang &Ohsawa 2002). Future studies could examine the geneticdifferences among co-occurring multi-stemmed and single-stemmed species in forests to ascertain the evolutionaryconsequences of these different life-history strategies (Bond& Midgley 2003).
Acknowledgements
New Zealand rain forest data derive from the National Vegetation Surveydatabank (http://nvs.landcareresearch.co.nz/). The Jamaican plots werere-measured in 2004 with funds from a Manaaki Whenua fellowship and fromGonville and Caius College, Cambridge (to Edmund Tanner). This studyreceived financial support from the Marsden Fund, Royal Society of NewZealand and the Foundation for Research, Science and Technology (Ecosystemresilience OBI). We thank Dave Kelly for his support of the study. SarahRichardson, Matt McGlone and Edmund Tanner made valuable criticisms ofthe manuscript.
References
Alfonso-Corrado, C., Clark-Tapia, R. & Mendoza, A. (2007) Demography andmanagement of two clonal oaks: Quercus eduardii and Q. potosina (Fagaceae)in central México. Forest Ecology and Management, 251, 129–141.
Barrera, M.D., Frangi, J.L., Richter, L.L., Perdomo, M.H. & Pinedo, L.B.(2000) Structural and functional changes in Nothofagus pumilio forestsalong an altitudinal gradient in Tierra del Fuego, Argentina. Journal ofVegetation Science, 11, 179–188.
Bellingham, P.J. (1991) Landforms influence patterns of hurricane damage:evidence from Jamaican montane forests. Biotropica, 23, 427–433.
Bellingham, P.J. & Lee, W.G. (2006) Distinguishing natural processes from
impacts of invasive mammalian herbivores. Biological Invasions in NewZealand (eds R.B. Allen & W.G. Lee), pp. 323–336. Springer, Berlin.
Bellingham, P.J. & Sparrow, A.D. (2000) Resprouting as a life history strategyin woody plant communities. Oikos, 89, 409–416.
Bellingham, P.J., Tanner, E.V.J. & Healey, J.R. (1994) Sprouting of trees inJamaican montane forests after a hurricane. Journal of Ecology, 82, 747–758.
Bellingham, P.J., Tanner, E.V.J., Rich, P.M. & Goodland, T.C.R. (1996)Changes in light below the canopy of a Jamaican montane rainforest after ahurricane. Journal of Tropical Ecology, 12, 699–722.
Bond, W.J. & Midgley, J.J. (2001) The persistence niche: ecology of sprouting inwoody plants. Trends in Ecology and Evolution, 16, 45–51.
Bond, W.J. & Midgley, J.J. (2003) The evolutionary ecology of sprouting inwoody plants. International Journal of Plant Sciences, 164, S103–S114.
Bond, W.J. & van Wilgen, B.W. (1996) Fire and Plants. Chapman & Hall,London.
Bowman, W.D., Bahnj, L. & Damm, M. (2003) Alpine landscape variation infoliar nitrogen and phosphorus concentrations and the relationship to soilnitrogen and phosphorus availability. Arctic, Antarctic and Alpine Research,35, 144–149.
Bruijnzeel, L.A. & Veneklaas, E. (1998) Climatic conditions and tropicalmontane forest productivity: the fog has not lifted yet. Ecology, 79, 3–9.
Cao, K. & Peters, R. (1998) Structure and stem growth of multi-stemmed treesof Fagus engleriana in China. Plant Ecology, 139, 211–220.
Chamberlin, E.A. & Aarsen, L.W. (1996) The cost of apical dominance in whitepine (Pinus strobus L.): growth in multi-stemmed versus single-stemmedtrees. Bulletin of the Torrey Botanical Club, 123, 268–272.
Clarke, P.J., Knox, K.J.E., Wills, K.E. & Campbell, M. (2005) Landscape patternsof woody plant response to crown fire: disturbance and productivity influencesprouting ability. Journal of Ecology, 93, 544–555.
Connell, J.H. (1978) Diversity in tropical rain forests and coral reefs. Science,199, 1302–1310.
Corlett, R.T. (1984) Human impact on the subalpine vegetation of Mt Wilhelm,Papua New Guinea. Journal of Ecology, 72, 841–854.
Cornelissen, J.H.C., Lavorel, S., Garnier, E., Díaz, S., Buchmann, N., Gurvich, D.E.,et al. (2003) A handbook of protocols for standardised and easy measurementof plant functional traits worldwide. Australian Journal of Botany, 51, 335–380.
Dalgleish, H.J. & Hartnett, D.C. (2006) Below-ground bud banks increasealong a precipitation gradient of the North American Great Plains: a test ofthe meristem limitation hypothesis. New Phytologist, 171, 81–89.
Dalling, J.W. (1994) Vegetation colonization of landslides in the Blue Mountains,Jamaica. Biotropica, 26, 392–399.
Dawson, J.W. & Sneddon, B.V. (1969) The New Zealand rain forest. Acomparison with tropical rain forest. Pacific Science, 23, 131–147.
Del Tredici, P. (2001) Sprouting in temperate trees: a morphological andecological review. Botanical Review, 67, 121–140.
Del Tredici, P., Ling, H. & Yang, G. (1992) The Ginkgos of Tian Mu Shan.Conservation Biology, 6, 202–209.
Dunphy, B.K., Murphy, P.G. & Lugo, A.E. (2000) The tendency for trees to bemultiple-stemmed in tropical and subtropical dry forests: studies of Guanicaforest, Puerto Rico. Tropical Ecology, 41, 161–167.
Fujiki, D. & Kikuzawa, K. (2006) Stem turnover strategy of multiple-stemmedwoody plants. Ecological Research, 21, 380–386.
Givnish, T.J. (1984) Leaf and canopy adaptations in tropical forests.Physiological Ecology of Plants of the Wet Tropics (eds E. Medina, H.A.Mooney & C. Vázquez-Yánes), pp. 51–84. Dr W. Junk, The Hague.
Givnish, T.J. (1995) Plant stems: biomechanical adaptation for energy captureand influence on species distribution. Plant Stems: Physiology and FunctionalMorphology (ed. B.L. Gartner), pp. 3–49. Academic Press, London.
Grace, J.B. (2006) Structural Equation Modeling and Natural Systems. CambridgeUniversity Press, Cambridge.
Grier, C.G. & Logan, R.S. (1977) Old-growth Pseudotsuga menziesii communitiesof a western Oregon watershed: biomass distribution and production budgets.Ecological Monographs, 47, 373–400.
Grime, J.P. (2002) Plant Strategies, Vegetation Processes, and Ecosystem Properties.Wiley, Chichester.
Grubb, P.J. (1977) Control of forest growth and distribution on wet tropicalmountains with special reference to mineral nutrition. Annual Review ofEcology and Systematics, 8, 83–107.
Grubb, P.J. & Tanner, E.V.J. (1976) The montane forests and soils of Jamaica:a reassessment. Journal of the Arnold Arboretum, 57, 313–368.
Harper, J.L. (1977) Population Biology of Plants. Academic Press, London.Hewitt, A.E. (1996) Estimating surface erosion using 137Cs at a semi-arid site
in Central Otago, New Zealand. Journal of the Royal Society of New Zealand,26, 107–118.

482 P. J. Bellingham & A. D. Sparrow
© 2009 Landcare Research. Journal compilation © 2009 British Ecological Society, Journal of Ecology, 97, 472–483
Heywood, V.H., Brummitt, R.K., Culham, A. & Seberg, O. (2007) FloweringPlant Families of the World. Firefly Books, Richmond, Ontario.
Insightful Corporation (2007) S-Plus version 8.0.4 for Windows. Seattle, Washington.Johnston, R.D. & Lacey, C.J. (1983) Multi-stemmed trees in rainforest.
Australian Journal of Botany, 31, 189–195.Kitayama, K. & Aiba, S. (2002) Ecosystem structure and productivity of trop-
ical rain forests along altitudinal gradients with contrasting soil phosphoruspools on Mount Kinabalu, Borneo. Journal of Ecology, 90, 37–51.
Klimesová, J. & Klimes, L. (2007) Bud banks and their role in vegetativeregeneration – a literature review and proposal for simple classificationand assessment. Perspectives in Plant Ecology, Evolution and Systematics, 8,115–129.
Kohyama, T. (1992) Size-structured multi-species model of rain forest trees.Functional Ecology, 6, 206–212.
Körner, C. (1999) Alpine plant life. Springer, Berlin.Körner, C. & Paulsen, J. (2004) A worldwide study of high altitude treeline
temperatures. Journal of Biogeography, 31, 713–732.Kruger, L.M., Midgley, J.J. & Cowling, R.M. (1997) Resprouters vs. reseeders
in South African forest trees; a model based on forest canopy height.Functional Ecology, 11, 101–105.
Kubo, M., Sakio, H., Shimano, K. & Ohno, K. (2005) Age structure anddynamics of Cercidiphyllum japonicum sprouts based on growth ring analysis.Forest Ecology and Management, 213, 253–260.
Lamont, B.B. & Wiens, D. (2003) Are seed set and speciation rates always lowamong species that resprout after fire, and why? Evolutionary Ecology, 17,277–292.
Lutz, J.A. & Halpern, C.B. (2006) Tree mortality during early forest development:a long-term study of rates, causes, and consequences. Ecological Monographs,76, 257–275.
Martin, T.J. & Ogden, J. (2006) Wind damage and response in New Zealandforests: a review. New Zealand Journal of Ecology, 30, 295–310.
McDonald, M.A., Healey, J.R. & Stevens, P.A. (2002) The effects of secondaryforest clearance and subsequent land-use on erosion losses and soil properties inthe Blue Mountains of Jamaica. Agriculture, Ecosystems and Environment,92, 1–19.
McMahon, T.A. (1973) Size and shape in biology. Science, 179, 1201–1204.Midgley, J.J. (1996) Why the world’s vegetation is not totally dominated
by resprouting plants: because resprouters are shorter than reseeders.Ecography, 19, 92–95.
Midgley, J.J. & Cowling, R.M. (1993) Regeneration patterns in Cape subtropicaltransition thicket – where are all the seedlings? South African Journal of Botany,59, 496–499.
Miura, M. & Yamamoto, S.-I. (2003) Structure and dynamics of a Castanopsiscuspidata var. sieboldii population in an old-growth, evergreen, broad-leaved forest:the importance of sprout regeneration. Ecological Research, 18, 115–129.
Moser, G., Hertel, D. & Leuschner, C. (2007) Altitudinal change in LAI andstand leaf biomass in tropical montane forests: a transect study in Ecuadorand a pan-tropical meta-analysis. Ecosystems, 10, 924–935.
Nagamatsu, D. & Miura, O. (1997) Soil disturbance regime in relation to micro-scale landforms and its effects on vegetation structure in a hilly area in Japan.Plant Ecology, 133, 191–200.
Nanami, S., Kawaguchi, H., Tateno, R., Li, C. & Katagiri, S. (2004) Sproutingtraits and population structure of co-occurring Castanopsis species in anevergreen broad-leaved forest in southern China. Ecological Research, 19,341–348.
Nzunda, E.F., Griffiths, M.E. & Lawes, M.J. (2007a) Multi-stemmed trees insubtropical coastal dune forest: survival strategy in response to chronicdisturbance. Journal of Vegetation Science, 18, 693–700.
Nzunda, E.F., Griffiths, M.E. & Lawes, M.J. (2007b) Resprouting versus turningup of leaning trees in a subtropical dune forest in South Africa. Journal ofTropical Ecology, 23, 289–296.
Ogden, J., Basher, L. & McGlone, M. (1998) Fire, forest regeneration and linkswith early human habitation: evidence from New Zealand. Annals of Botany,81, 687–696.
Pausas, J.G. & Bradstock, R.A. (2007) Fire persistence traits of plants along aproductivity and disturbance gradient in mediterranean shrublands ofsouth-east Australia. Global Ecology and Biogeography, 16, 330–340.
Pausas, J.G. & Lavorel, S. (2003) A hierarchical deductive approach for functionaltypes in disturbed ecosystems. Journal of Vegetation Science, 14, 409–416.
Peterson, C.J. & Jones, R.H. (1997) Clonality in woody plants: a review andcomparison with clonal herbs. The ecology and evolution of clonal plants (edsH. de Kroon & J. van Groenendael), pp. 263–289. Backhuys, Leiden.
Pettit, N.E. & Naiman, R.J. (2007) Fire in the riparian zone: characteristics andecological consequences. Ecosystems, 10, 673–687.
Phillips, O.L. & Gentry, A.H. (1994) Increasing turnover through time intropical forests. Science, 263, 954–958.
Pigott, C.D. (1989) Factors controlling the distribution of Tilia cordata Mill atthe northern limits of its geographical range. IV. Estimated ages of the trees.New Phytologist, 112, 117–121.
Porder, S., Vitousek, P., Chadwick, O.A., Chamberlain, C.P. & Hilley, G.E.(2007) Uplift, erosion and phosphorus limitation in terrestrial ecosystems.Ecosystems, 10, 158–170.
Prescott, C.E., Corbin, J.P. & Parkinson, D. (1992) Availability of nitrogen andphosphorus in the forest floors of Rocky-Mountain coniferous forests.Canadian Journal of Forest Research, 22, 593–600.
Reif, A. & Allen, R.B. (1988) Plant communities of the steepland conifer–broadleaved hardwood forests of central Westland, South Island, NewZealand. Phytocoenologia, 16, 145–224.
Richards, P.W. (1952) The tropical rain forest. Cambridge University Press,Cambridge.
Richardson, S.J., Allen, R.B. & Doherty, J.E. (2008) Shifts in leaf N : P ratioduring resorption reflect soil P in temperate rainforest. Functional Ecology,22, 738–745.
Rohde, K. (1992) Latitudinal gradients in species diversity: the search for theprimary cause. Oikos, 65, 514–527.
Russo, S.E., Davies, S.J., King, D.A. & Tan, S. (2005) Soil-related performancevariation and distributions of tree species in a Bornean rain forest. Journal ofEcology, 93, 879–889.
Sakai, A., Ohsawa, T. & Ohsawa, M. (1995) Adaptive significance of sproutingof Euptelea polyandra, a deciduous tree growing on steep slopes with shallowsoil. Journal of Plant Research, 108, 377–386.
Sakai, A., Sakai, S. & Akiyama, F. (1997) Do sprouting trees on erosion-pronesites carry large reserves of resources? Annals of Botany, 79, 625–630.
Shreve, F. (1914) A montane rain-forest. A contribution to the physiologicalplant geography of Jamaica. Carnegie Institute of Washington Publication,199, 1–110.
Smale, M.C. (1994) Structure and dynamics of kanuka (Kunzea ericoides var.ericoides) heaths on sand dunes in Bay of Plenty, New Zealand. New ZealandJournal of Botany, 32, 441–452.
Steinke, L.R., Premoli, A.C., Souto, C.P. & Hedrén, M. (2008) Adaptive andneutral variation of the resprouter Nothofagus antarctica growing in distincthabitats in north-western Patagonia. Silva Fennica, 42, 177–188.
Stephenson, N.L. & van Mantgem, P.J. (2005) Forest turnover rates follow globaland regional patterns of productivity. Ecology Letters, 8, 524–531.
Tang, C.Q. & Ohsawa, M. (2002) Tertiary relic deciduous forests on a humidsubtropical mountain, Mt. Emei, Sichuan, China. Folia Geobotanica, 37,93–106.
Tanner, E.V.J. (1977) Four montane rain forests of Jamaica: a quantitativecharacterization of the floristics, the soils and foliar mineral levels, and adiscussion of the interrelations. Journal of Ecology, 65, 883–918.
Tanner, E.V.J. (1981) The decomposition of leaf litter in Jamaican montanerain forests. Journal of Ecology, 69, 263–275.
Tanner, E.V.J. & Bellingham, P.J. (2006) Less diverse forest is more resistant tohurricane damage: evidence from montane rain forests in Jamaica. Journalof Ecology, 94, 1003–1010.
Tanner, E.V.J., Kapos, V. & Franco, W. (1992) Nitrogen and phosphorusfertilization effects on Venezuelan montane forest trunk growth andlitterfall. Ecology, 73, 78–86.
Tanner, E.V.J., Vitousek, P.M. & Cuevas, E. (1998) Experimental investigationsof nutrient limitation of forest growth on wet tropical mountains. Ecology,79, 10–22.
Tanner, J.E. (2001) The influence of clonality on demography: patterns inexpected longevity and survivorship. Ecology, 82, 1971–1981.
ter Braak, C.J.F. & Smilauer, P. (1998) Canoco 4.0 for Windows. Centre forBiometry, Wageningen, The Netherlands.
Tonkin, P.J. & Basher, L.R. (2001) Soil chronosequences in subalpine super-humid Cropp Basin, western Southern Alps, New Zealand. New ZealandJournal of Geology and Geophysics, 44, 37–45.
van Steenis, C.G.G.J. (1935) On the origin of the Malaysian mountain flora.Part 2. Altitudinal zones, general considerations and renewed statement ofthe problem. Bulletin du Jardin Botanique de Buitenzorg, 13, 289–419.
Vesk, P.A. & Westoby, M. (2004) Sprouting by plants: the effects of modularorganization. Functional Ecology, 18, 939–945.
Vile, D., Shipley, B. & Garnier, E. (2006) A structural equation model to integratechanges in functional strategies during old-field succession. Ecology, 87,504–517.
Vitousek, P., Chadwick, O., Matson, P., Allison, S., Derry, L., Kettley, L.,Luers, A., Mecking, E., Monastra, V. & Porder, S. (2003) Erosion and therejuvenation of weathering-derived nutrient supply in an old tropicallandscape. Ecosystems, 6, 762–772.
Walker, L.R. & del Moral, R. (2003) Primary Succession and EcosystemRehabilitation. Cambridge University Press, Cambridge.

Multi-stemmed trees in montane rain forests 483
© 2009 Landcare Research. Journal compilation © 2009 British Ecological Society, Journal of Ecology, 97, 472–483
Wardle, P. (1986) Frequency of cloud cover on New Zealand mountains inrelation to subalpine vegetation. New Zealand Journal of Botany, 24, 553–565.
Wardle, P. (2008) New Zealand forest to alpine transitions in global context.Arctic, Antarctic and Alpine Research, 40, 240–249.
Wardle, D.A., Walker, L.R. & Bardgett, R.D. (2004) Ecosystem propertiesand forest decline in contrasting long-term chronosequences. Science, 305,509–513.
Wells, P.V. (1969) The relation between mode of reproduction and extent ofspeciation in woody genera of the California chaparral. Evolution, 23,264–267.
Wells, A., Yetton, M.D., Duncan, R.P. & Stewart, G.H. (1999) Prehistoricdates of the most recent Alpine fault earthquakes, New Zealand. Geology,27, 995–998.
Woolley, L.P., Henkel, T.W. & Sillett, S.C. (2008) Reiteration in the mono-dominant tropical tree Dicymbe corymbosa (Caesalpinaceae) and itspotential adaptive significance. Biotropica, 40, 32–43.
Zobel, K. (2008) On the forces that govern clonality versus sexuality in plantcommunities. Evolutionary Ecology, 22, 487–492.
Received 4 December 2008; accepted 12 January 2009Handling Editor: Hans Cornelissen
Supporting Information
Additional Supporting Information may be found in theonline version of this article:
Figure S1. Structural equation models testing for the pro-portion of multi-stemmed individuals of Pseudowinteracolorata and Weinmannia racemosa in New Zealand montanerain forests.
Figure S2. Detrended correspondence analysis of compositionof montane rain forests according to basal area in orders in 2montane rain forests.
Figure S3. Annual mortality and recruitment rates of stemsand of individuals in 2 montane rain forests.
Table S1. Each species frequency, frequency of multi-stemmedindividuals, mean and range of number of multi-stemmedindividuals in Jamaican and New Zealand forests
Table S2. Correlations between physiographic and soilvariables for two montane rain forests
Please note: Wiley-Blackwell are not responsible for the con-tent or functionality of any supporting materials supplied bythe authors. Any queries (other than missing material) shouldbe directed to the corresponding author for the article.





























