Embed Size (px)
Citation preview

Immunity, Vol. 19, 571–582, October, 2003, Copyright 2003 by Cell Press
NAD-Induced T Cell Death:ADP-Ribosylation of Cell Surface Proteins by ART2Activates the Cytolytic P2X7 Purinoceptor
include withdrawal of survival factors as well as trig-gering of so-called “death receptors,” such as Fas andother members of the TNF receptor family (Krammer,2000). Here we delineate an alternative mechanism forinducing T cell death by extracellular NAD released from
Michel Seman,1 Sahil Adriouch,1,5
Felix Scheuplein,2,5 Christian Krebs,2,5
Dunja Freese,2 Gustavo Glowacki,2
Phillipe Deterre,3 Friedrich Haag,2,4
and Friedrich Koch-Nolte2,*injured cells.1Universite Denis Diderot
Beyond their central roles in energy metabolism, ATPF-75251 Parisand NAD also play multifaceted roles as second mes-Francesengers in the extracellular environment where they act2 Institute of Immunologyas substrates and ligands for diverse ectoenzymes andUniversity Hospitalreceptors. These nucleotides can be released from cellsMartinistr. 52by lytic and nonlytic mechanisms into the extracellularD-20246 Hamburgenvironment (Bruzzone et al., 2001; Contreras et al.,Germany2002), where they interact with specific receptors (e.g.,3 Hopital Pitie-SalpetriereP1 and P2 purinoceptors) and/or are degraded hydrolyti-75634 Paris Cedex 13cally by membrane-bound and secretory ectoenzymesFranceincluding NADases, ATPases, and phosphodiesterases4 Vaccine-Lab(e.g., CD38, CD39, and CD203) (Goding and Howard,18057 Rostock1998; Lund et al., 1995). ATP and NAD also serve asGermanysubstrates for posttranslational protein modifications,i.e., phosphorylation and ADP-ribosylation.
It has been shown that ecto-ATP can induce deathSummaryof T cells and macrophages (Canaday et al., 2002;Chvatchko et al., 1996; Di Virgilio et al., 2001; Labasi etT cells express a toxin-related ADP-ribosylating ec-al., 2002; Lammas et al., 1997; MacKenzie et al., 2001;toenzyme, ART2. Exposure of mature T cells to NAD,Surprenant et al., 1996; Zanovello et al., 1990). ATP-the substrate for ADP-ribosylation, induces cell death.induced activation of the cytolytic P2X7 purinoceptorART2-catalyzed ADP-ribosylation activates the cyto-induces calcium flux, formation of large membranelytic P2X7 purinoceptor, causing calcium flux, porepores, exposure of phosphatidylserine and uptake offormation, phosphatidylserine exposure, shedding ofpropidium iodide, ultimately resulting in cell death. How-CD62L, cell shrinkage, and propidium iodide uptake.ever, millimolar concentrations of ATP were required toInterestingly, much lower NAD than ATP concentra-elicit these responses. It has remained a mystery howtions are required to activate P2X7. NAD-induced cellsuch high ATP concentrations could be derived fromdeath (NICD) operates with endogenous sources ofnatural sources. Here we delineate a mechanism forNAD released upon cell lysis. These findings identifyactivating P2X7 by a ligand generated from endoge-P2X7 as a key effector of NICD and demonstrate thatnous sources.P2X7 can be activated by an endogenous ligand other
Cytotoxicity mediated by protein ADP-ribosylation isthan ATP. Our results delineate an alternative mecha-a common theme of many bacterial pathogens that posenism for inducing T cell death and set an interestingsignificant human health threats (C. diphtheriae, V. chol-precedent for immunoregulation via crosstalk betweenerae, E. coli, B. pertussis, S. entericae, C. botulinum, S.NAD-dependent ADP-ribosyltransferases and purino-aureus, P. aeruginosa) (Aktories and Just, 2000; Dome-
ceptors.nighini and Rappuoli, 1996). Symptoms of diphtheria,whooping cough, and severe diarrhea are caused by
Introduction ADP-ribosylating enzymes that translocate into mam-malian cells. These toxins interfere with protein synthe-
The maintenance of homeostasis in the immune system sis, signal transduction, or cytoskeletal functions byand the focusing of immune reactions on appropriate ADP-ribosylating key target proteins such as elongationtargets require the coordinated interplay of regulatory factor 2, G proteins, or actin. Mammalian ADP-ribosylat-mechanisms. Programmed cell death plays a major role ing ectoenzymes, designated ART1-ART5, have beenin many phases of the immune response (Van Parijs discovered that bear distant sequence similarity to theseand Abbas, 1998). Impairments in cell death signaling bacterial toxins (Glowacki et al., 2002; Haag and Koch-pathways can lead to autoimmune and lymphoprolifera- Nolte, 1997; Okazaki and Moss, 1998). Mammalian ecto-tive diseases. In the course of evolution, multiple mecha- ADP-ribosyltransferases (ARTs) have been shown tonisms of eliciting cell death have developed within the ADP-ribosylate T cell membrane proteins and secretoryimmune system that greatly enhance its flexibility to antibacterial factors (Liu et al., 1999; Paone et al., 2002).respond adequately to increasing challenges (Leist and The recently solved crystal structure of rat ART2 con-Jaattela, 2001). Death pathways in the immune system firms the close structural and evolutionary relationship
of mammalian ARTs and ADP-ribosylating bacterial tox-ins (Mueller-Dieckmann et al., 2002). The results pre-*Correspondence: [email protected]
5 These authors contibuted equally to this work. sented here demonstrate that ART2 and ADP-ribosylat-

Immunity572
ing bacterial toxins may share not only a common cubating cells with etheno-NAD followed by washingeffectively prevented subsequent NICD (Figures 2B andstructure and enzymatic activity but also a common
function: cytotoxicity. We show that micromolar con- 2D). This suggests that etheno-NAD acts by blockingprotein ADP-ribosylation via prior etheno-ADP-ribosyla-centrations of NAD derived from endogenous sources
are sufficient to kill T cells by activating P2X7 via ART2- tion rather than as a competitive inhibitor of NAD. Similarresults were obtained with the NAD analogs nicotin-catalyzed ADP-ribosylation.amide guanidine dinucleotide (NGD) and nicotinamidehypoxanthine dinucleotide (NHD) (Figures 2C and 2D).ResultsIn the experiment shown in Figure 2D, cells were prein-cubated with NGD and NHD and washed before treat-Exposure of T Cells to the ART Substrate, NAD,ment with NAD, suggesting that NGD and NHD, likeInduces Cell Death by Apoptosisetheno-NAD, serve as substrates for ecto-ARTs and ex-Exposure of phosphatidylserine (PS) is an early sign ofert their effects by GDP-ribosylation and HDP-ribosyla-apoptosis; propidium iodide (PI) staining is an indicatortion of cell surface proteins, respectively. Etheno-NAD,of cell death (Bossy-Wetzel and Green, 2000). In accordNGD, and NHD differ from NAD only in the adeninewith previous reports (Adriouch et al., 2001; Liu et al.,moiety (see Supplemental Data at http://www.immunity.2001), treatment of lymph node T cells with micromolarcom/cgi/content/full/19/4/571/DC1). These findings sug-concentrations of exogenous NAD induced PS exposuregested to us that an essential downstream effector ofand, ultimately, PI staining (Figure 1A). NAD-inducedcell surface protein ADP-ribosylation senses this differ-cell death (NICD) was accompanied by cell shrinkageence in the adenine group.(Figure 1D), indicating that cells were dying by apoptosis
rather than by necrosis, which is characterized by cellswelling (Dive et al., 1992; Okada et al., 2001). NAD induced NAD Induces Calcium Flux, Formation of Membraneother hallmarks of apoptosis, e.g., disruption of mitochon- Pores, and Shedding of CD62Ldrial membrane potential (Figure 1E) and DNA fragmen- Members of the purinoceptor family of cell surface pro-tation (Adriouch et al., 2001). teins are known to be sensitive to modifications of the
adenosine moiety of their ligands (North and Surprenant,2000). Among the purinoceptors, P2X7 is expressed onNICD Requires ART2-Catalyzed ADP-Ribosylationlymphocytes and is well known to mediate PS exposureof Cell Surface Proteinsupon triggering with high doses of ATP (Di Virgilio et al.,ADP-ribose, which can be generated from NAD by extra-2001; Zanovello et al., 1990). P2X7 thus represents acellular NAD-glycohydrolases (Lund et al., 1995), did notgood candidate for the downstream adenosine sensinginduce PS exposure or PI uptake, even at millimolareffector of NICD. Hallmarks of P2X7 triggering are induc-concentrations (Figure 1C), suggesting that NAD in-tion of calcium flux and formation of membrane poresduced cell death either directly or via catabolization bypermeable to molecules up to 800 Da. The latter can beNAD-dependent ADP-ribosyltransferases. Indeed, cellsmonitored by uptake of fluorescent DNA binding com-lacking ecto-ADP-ribosyltransferase ART2 (Ohlrogge etpounds such as ethidium bromide and YO-PRO-1 (Dial., 2002) were resistent to NICD (Figure 1B). Moreover,Virgilio et al., 2001; North and Surprenant, 2000). In orderpreincubating ART2-expressing cells with ART2-spe-to explore the possible role of P2X7 in NICD, we testedcific antibodies (Figure 1C) effectively prevented NICD.whether calcium flux and dye uptake can be elicitedSince these antibodies block ART2-catalyzed ADP-ribo-by NAD. Indeed, intracellular calcium levels increasedsylation of cell surface proteins (Adriouch et al., 2001),within minutes after addition of ecto-NAD (Figure 3A).the enzymatic activity of ART2 appears to be required forMoreover, as in the case of treatment with ATP, T cellsNICD. Consistent with this conclusion, other inhibitors ofbecame permeable to ethidium bromide and YO-PRO-1cell surface ADP-ribosylation (Banasik et al., 1992;within 30 min after treatment with NAD (Figures 3B andKoch-Nolte et al., 1996), e.g., agmatine, a low molecular3C). Another reported consequence of P2X7 receptorweight surrogate substrate of ecto-ARTs (Figure 1C),triggering by ecto-ATP is shedding of CD62L (Gu etand nicotinamide, a competitive inhibitor of NAD-depen-al., 1998; Jamieson et al., 1996). Similarily, treatment ofdent ADP-ribosylation (data not shown) also blockedT cells with NAD induced shedding of CD62L (Figure 3D).NICD. Taken together, these findings show that NICDConsistent with its effects on NICD (Figure 2), etheno-requires ART2-catalyzed ADP-ribosylation of cell sur-NAD did not induce CD62L shedding, and pretreatingface proteins.cells with etheno-NAD blocked subsequent NAD-inducedCD62L shedding (Figure 3D). Note that ART2�/� T cellsNAD Analogs with Modifications of the Adeninewere resistant to NAD-induced shedding of CD62L (Fig-Group Block NICDure 3D), calcium-flux, and dye uptake (data not shown).The NAD analog etheno-NAD is an efficient substrateNote also that both NAD-induced uptake of YO-PRO-1for ecto-ART2, and etheno-ADP-ribosylation of T celland NAD-induced CD62L shedding were restricted tosurface proteins can be monitored by flow cytometryT cells, whereas B cells—which do not express ART2and immunoblot analyses (Kahl et al., 2000; Krebs et(Koch-Nolte et al., 1999)—were completely resistant toal., 2003). Note that ART2-deficient cells do not etheno-these effects (Figures 3C and 3D).ADP-ribosylate cell surface proteins (Figure 2A). Re-
markably, in contrast to NAD, etheno-NAD did not in-duce PS exposure or PI staining of wild-type T cells P2X7 Antagonists Block NAD-Induced Calcium Flux
and PS Exposure(Figures 2B and 2C). Evidently, NICD can be inducedby ADP-ribosylated cell surface protein(s) but not by The involvement of P2X7 in NICD was further corrobo-
rated with known antagonists of P2X7 (Abbracchio andetheno-ADP-ribosylated protein(s). Furthermore, prein-

NAD-Induced Cell Death573
Figure 1. NAD Induces T Cell Death via ART2
(A and B) ART2�/� (A) or ART2�/� (B) T cells were incubated in the absence or presence of 10 �M NAD for 1 hr (top) or 20 hr (bottom). Cellswere then washed and stained with Annexin V and propidium iodide for FACS analysis.(C) ART2�/� T cells were incubated for 30 min in the absence (1) or presence of 2 mM ADP-ribose (2), 20 �M NAD (3–6), or 65 mM agmatine(7) and then stained with Annexin V/PI. Sixty minutes before and during NAD treatment, cells were also exposed to 0.1 �g/ml control antibodies(3), 0.1�g/ml ART2-specific antibodies (4), or 65 mM agmatine (6).(D) T cells were incubated for 16 hr in the absence or presence of 20 �M NAD and then monitored by FACS analyses for changes in forwardand side scatter (FSC, SSC). Sixty minutes before and during NAD treatment, cells in panel 3 were also exposed to 0.1 �g/ml ART2-specific antibodies.(E) T cells were incubated for 6 hr in the absence or presence of 50 �M NAD or 5 �M staurosporine and then stained with the cell permeantMitosensor dye whose fluorescence increases as the mitochondrial membrane potential decreases. Numbers indicate the percentage of cellsin the respective gated populations.
Burnstock, 1994; Gargett and Wiley, 1997; North and The Cytotoxic Effects of NAD Are Not MediatedSurprenant, 2000). Preincubation of cells with the P2X7 by ATP Released from Cellsantagonists KN-62 or oxidized ATP (oATP) effectively Since ATP was the only physiologic ligand for P2X7blocked both NAD-induced and ATP-induced PS expo- described so far, it was conceivable that the cytotoxicsure (Figure 3E) and calcium flux (Figure 3F). effects of NAD were mediated by ATP released from
cells. For example, ADP-ribosylation of membrane pro-teins might cause an increased release of ATP, and/orNAD Itself Is Not a Ligand for P2X7ADP-ribosylation of membrane proteins might dramati-To address the question whether NAD itself functionscally lower the threshold of P2X7 to ATP released fromas a ligand for P2X7, we analyzed the response of T cellscells in a constitutive fashion. In order to address thesefrom ART2�/� mice to NAD and ATP (Ohlrogge et al.,questions, we treated T cells with NAD or ATP in the2002). ART2-deficient T cells were resistant to NICD butpresence of the potent ATP-hydrolase apyrase (Figuresensitive to direct triggering of P2X7 by ATP (Figure 4A).4B). Apyrase treatment prevented ATP-induced PS ex-This rules out the possibility that NAD itself is a ligandposure even at very high concentrations of exogenousfor P2X7 and further supports the notion that P2X7 is
activated by ADP-ribosylation of membrane proteins. ATP, whereas it had no effect on NAD-induced PS expo-

Immunity574
Figure 2. NICD Is Prevented by Pretreating T Cells with Etheno-NAD or NAD Analogs Bearing Modifications in the Adenine Moiety
(A) Flow cytometric assay for etheno-ADP-ribosylation of membrane proteins. Panel 1: T cells were incubated for 30 min with or without 20�M etheno-NAD, washed, and then stained with fluorochrome-conjugated etheno-adenosine-specific mAb 1G4 (Krebs et al., 2003). Panel 2:ART2�/� and ART2�/� T cells were incubated for 10 min with 20 �M etheno-NAD, washed, and then stained with mAb 1G4.(B) ART2�/� T cells were incubated for 0.5 hr (top) or for 16 hr (bottom) with 20 �M NAD or 20 �M etheno-NAD and then stained with AnnexinV/PI. Cells in panel 4 were pretreated for 1 hr with etheno-NAD followed by washing prior to treatment with NAD.(C) T cells were incubated for 30 min with the indicated concentrations of NAD, etheno-NAD, NGD, or NHD and then stained as in (B).(D) T cells were preincubated for 30 min with the indicated concentrations of etheno-NAD, NGD, or NHD, washed, and then treated for 30min with 30 �M NAD before staining with Annexin V/PI as in (B).
sure. Conversely, treatment of cells with NAD in the by activating P2X7. Remarkably, the P2X7 antiserumcould also inhibit PS exposure in response to low levelspresence of NADase strongly reduced NICD but not
ATP-induced cell death (Figure 4B). NADase only re- of ATP (Figure 5B), but high concentrations of ATP couldbypass this inhibition (data not shown). Furthermore,duced PS exposure to background levels when NAD
rather than cells were preincubated with NADase (data following incubation of T cells with radiolabeled NAD,the P2X7 antiserum precipitated a radiolabeled 70 kdnot shown). In the latter case, evidently, ADP-ribosyla-
tion occurred before the NADase could hydrolyze the band corresponding to P2X7 (Figure 5C). These resultssuggest that P2X7 is activated directly by ADP-ribosy-substrate. We conclude that the effects of NAD are me-
diated by cell surface ADP-ribosylation and not by ATP lation.released from cells.
P2X7 Is ADP Ribosylated NAD Activates P2X7 at Lower ConcentrationsThan Does ATPThe obvious next question was whether P2X7 itself is a
target for ADP-ribosylation. To address this issue, we The results of dose-response analyses (Figure 6A) revealthat NAD can activate P2X7 at lower doses than ATPraised an antiserum to P2X7 by DNA vaccinating rabbit
“K1G” with an expression vector for P2X7. The specific- (EC50 for PS exposure 2 �M versus 100 �M). Moreover,the response to ATP shows a threshold effect, and dosesity of the K1G antiserum was verified by FACS (Figure
5A) and immunoblot analyses (Figure 5C). Importantly, below 50 �M ATP do not induce exposure of PS. Incontrast, NAD induces PS exposure of some cells atthe P2X7 antiserum, like the ART2 antiserum, blocked
NAD-induced PS exposure (Figure 5B). This further un- concentrations as low as 1 �M. With increasing concen-trations of NAD a steady increase in the number of cellsderscores the conclusion that NAD mediates its effects

NAD-Induced Cell Death575
Figure 3. NAD Induces Responses Characteristic of P2X7 which Are Blocked by P2X7 Antagonists
(A) Fura-2 loaded T cells were stirred in a fluorimeter cuvette. NAD was added (arrow) at the indicated concentrations, and changes in cytosolicCa2� were monitored by fluorometry with excitation at 340/380 nm and emission at 510 nm.(B) T cells were incubated for the indicated times in the presence of 10 �M NAD. Ethidium bromide (1 �g/ml) was added for the last 2 minprior to counterstaining with Annexin V.(C) Total lymph node cells were incubated for 30 min in the absence or presence of 100 �M NAD or 200 �M ATP. YO-PRO-1 (10 �g/ml) wasadded for the last 2 min prior to counterstaining with anti-mouse IgG.(D) Total lymph node cells were incubated for 30 min in the absence (panels 1 and 5) or presence of 100 �M NAD (panels 3, 4, and 7) or 1mM ATP (panels 2 and 6) and then stained with anti-CD3FITC and anti-CD62LPE. Cells in panel 4 were pretreated for 10 min with 100 �M etheno-NAD before treatment with NAD. Cells in panels 1–4 were from an ART2�/� mouse; cells in panels 5–7 were from an ART2�/� mouse.(E) T cells were preincubated for 120 min with P2X7 antagonists KN-62 or oxidized ATP, washed in the case of oxidized ATP, and incubatedfurther for 30 min in the presence of 10 �M NAD or 300 �M ATP. Cells were then washed and stained with Annexin V/PI.(F) Fura-2 loaded T cells were preincubated with or without KN-62, NHD, or ADP-ribose for 5 min in a stirred fluorimeter cuvette. NAD wasadded as indicated and changes in cytosolic Ca2� were monitored by fluorometry as in (A).
exposing PS is observed, but a fraction of cells remains (MacKenzie et al., 2001). To assess the reversibility ofNAD-induced PS exposure, we treated T cells brieflyresistant to NICD.with NAD or ATP, removed the nucleotides by washing,and incubated the cells further. The results show thatATP-Induced but Not NAD-Induced PS Exposure
Is Readily Reversed T cells, like macrophages, reverse PS exposure afterremoval of ATP (Figure 6B). In striking contrast, NAD-As shown recently, ATP-induced exposure of PS on
human macrophages is reversible upon removal of ATP induced PS exposure shows little if any recovery upon

Immunity576
Figure 4. NAD Itself Is Not a Ligand for P2X7, and NICD Is NotMediated by ATP Released from Cells
(A) ART2�/� T cells were incubated for 30 min in the absence orpresence of 25 �M NAD or 250 �M ATP and then stained withAnnexin V/PI.(B) ART2�/� T cells were incubated for 30 min with 25 �M NAD(top) or 250 �M ATP (bottom) in the absence or presence of potatoapyrase (ATPase) or Neurospora crassa NAD glycohydrolase(NADase), and cells were then stained with Annexin V/PI.
removal of NAD (Figure 6C). The reversibility of the re-sponse to ATP is compatible with that of a receptor toits soluble ligand. The prolonged reaction to NAD iscompatible with the covalent attachment of the ligand Figure 5. P2X7 Is a Target for ADP-Ribosylationto its receptor. (A) HEK cells stably transfected with P2X7 or ART2.2 were stained
with antisera (1:2000 dilution) derived from rabbits immunized withexpression vectors for ART2.2 (solid line) or P2X7 (dashed line).ADP-Ribosylation Agonizes and Etheno-ADP-Bound antibody was visualized by staining with PE-conjugated anti-Ribosylation Antagonizes P2X7 for Triggering by ATPrabbit Ig.The discovery that P2X7 is a target for ART2 raised(B) ART2�/� T cells were incubated for 30 min in the absence or
the important question whether NAD and ATP might presence of 25 �M NAD (top) or 125 �M ATP (bottom) and thenact synergistically, i.e., whether ADP-ribosylation could stained with Annexin V/PI. Sixty minutes before and during nucleo-
tide treatment, cells were also exposed to K1G preimmune serumsensitize P2X7 for activation by lower concentrations of(pIS) or immune serum (�P2X7) (each at 1:500 dilution).ATP, and conversely, whether etheno-ADP-ribosylation(C) MD27 lymphoma cells (lanes 1–4) and ART2�/� T cells (lanescould antagonize activation of P2X7 by ATP. To address5–7) were incubated for 30 min in the presence of 1 �M (5�Ci) 32P-this issue, we pretreated T cells briefly with NAD to allowNAD. Cells were washed and lysed in 1% Igepal, and proteins in
cell surface ADP-ribosylation, followed by treatment cell lysates were subjected to sequential immunoprecipitation withwith threshold levels of ATP. The results show that NAD, preimmune serum (lanes 2 and 5), P2X7 antiserum (lanes 3 and 6),
and anti-LFA-1 mAb (lanes 4 and 7). Lane 1 contains a control aliquotindeed, sensitizes cells slightly for ATP-induced calciumof the MD27 cell lysate before precipitation. Proteins were sizeinflux, albeit only at low ATP concentrations (Figure 6D).fractionated by SDS-PAGE and blotted onto a nitrocellulose mem-Conversely, pretreatment of cells with etheno-NAD in-brane. Radiolabeled proteins were visualized by autoradiography.hibited ATP-induced calcium influx (Figure 6E). The in-After quenching of radioactivity, proteins on the membrane were
hibitory effects of etheno-NAD could be overcome by subjected to immunoblot analyses with an anti-P2X7 peptide antise-high levels of ATP (data not shown). rum and peroxidase-conjugated goat anti-rabbit IgG. Bands corre-
sponding to the heavy chains of the precipitated rabbit antibodiesare marked by H; bands corresponding to P2X7 are marked byNAD Released by Cell Lysis Can Induce T Cell Deathan arrow.If NICD were to play a physiological role, the effects
described above should also be inducible with NADreleased from cells. While intracellular NAD levels are kept low by CD38 and other extracellular NAD glycohy-
drolases (Lund et al., 1995). Presumably, cell lysis duringin the range of 200–1000 �M, extracellular NAD levels,e.g., in serum, are in the submicromolar range and are tissue injury and inflammation can cause local ecto-NAD
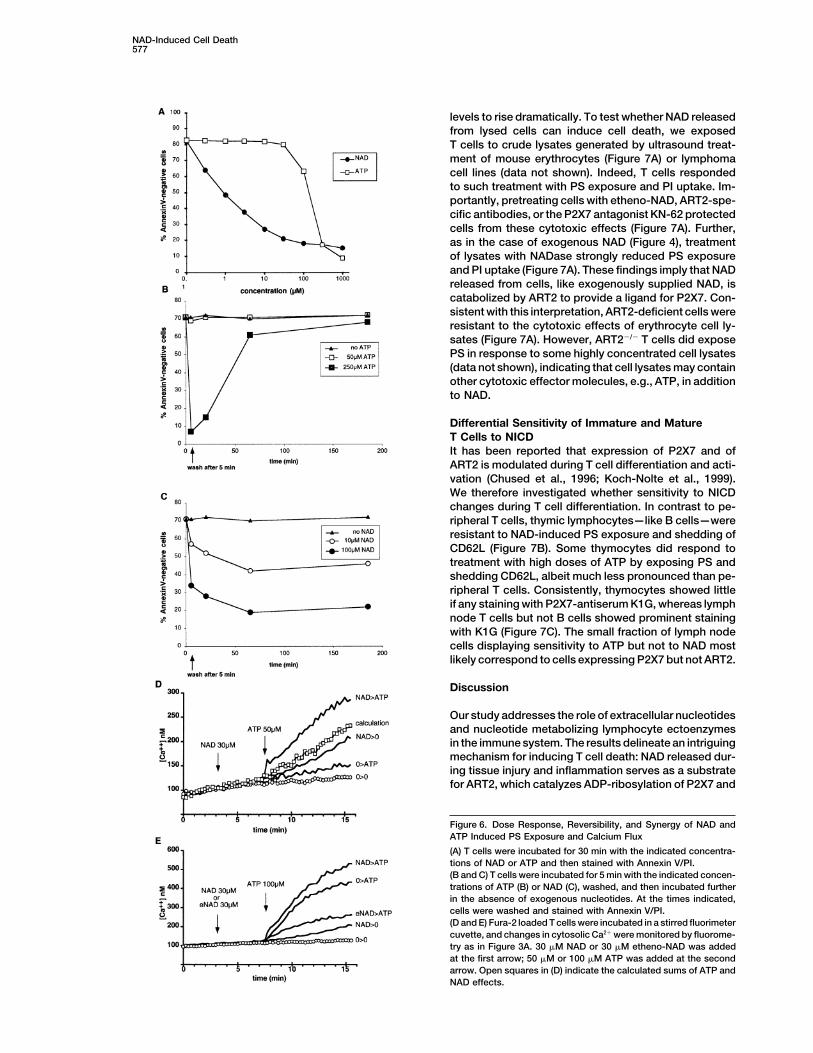
NAD-Induced Cell Death577
levels to rise dramatically. To test whether NAD releasedfrom lysed cells can induce cell death, we exposedT cells to crude lysates generated by ultrasound treat-ment of mouse erythrocytes (Figure 7A) or lymphomacell lines (data not shown). Indeed, T cells respondedto such treatment with PS exposure and PI uptake. Im-portantly, pretreating cells with etheno-NAD, ART2-spe-cific antibodies, or the P2X7 antagonist KN-62 protectedcells from these cytotoxic effects (Figure 7A). Further,as in the case of exogenous NAD (Figure 4), treatmentof lysates with NADase strongly reduced PS exposureand PI uptake (Figure 7A). These findings imply that NADreleased from cells, like exogenously supplied NAD, iscatabolized by ART2 to provide a ligand for P2X7. Con-sistent with this interpretation, ART2-deficient cells wereresistant to the cytotoxic effects of erythrocyte cell ly-sates (Figure 7A). However, ART2�/� T cells did exposePS in response to some highly concentrated cell lysates(data not shown), indicating that cell lysates may containother cytotoxic effector molecules, e.g., ATP, in additionto NAD.
Differential Sensitivity of Immature and MatureT Cells to NICDIt has been reported that expression of P2X7 and ofART2 is modulated during T cell differentiation and acti-vation (Chused et al., 1996; Koch-Nolte et al., 1999).We therefore investigated whether sensitivity to NICDchanges during T cell differentiation. In contrast to pe-ripheral T cells, thymic lymphocytes—like B cells—wereresistant to NAD-induced PS exposure and shedding ofCD62L (Figure 7B). Some thymocytes did respond totreatment with high doses of ATP by exposing PS andshedding CD62L, albeit much less pronounced than pe-ripheral T cells. Consistently, thymocytes showed littleif any staining with P2X7-antiserum K1G, whereas lymphnode T cells but not B cells showed prominent stainingwith K1G (Figure 7C). The small fraction of lymph nodecells displaying sensitivity to ATP but not to NAD mostlikely correspond to cells expressing P2X7 but not ART2.
Discussion
Our study addresses the role of extracellular nucleotidesand nucleotide metabolizing lymphocyte ectoenzymesin the immune system. The results delineate an intriguingmechanism for inducing T cell death: NAD released dur-ing tissue injury and inflammation serves as a substratefor ART2, which catalyzes ADP-ribosylation of P2X7 and
Figure 6. Dose Response, Reversibility, and Synergy of NAD andATP Induced PS Exposure and Calcium Flux
(A) T cells were incubated for 30 min with the indicated concentra-tions of NAD or ATP and then stained with Annexin V/PI.(B and C) T cells were incubated for 5 min with the indicated concen-trations of ATP (B) or NAD (C), washed, and then incubated furtherin the absence of exogenous nucleotides. At the times indicated,cells were washed and stained with Annexin V/PI.(D and E) Fura-2 loaded T cells were incubated in a stirred fluorimetercuvette, and changes in cytosolic Ca2� were monitored by fluorome-try as in Figure 3A. 30 �M NAD or 30 �M etheno-NAD was addedat the first arrow; 50 �M or 100 �M ATP was added at the secondarrow. Open squares in (D) indicate the calculated sums of ATP andNAD effects.

Immunity578
Figure 7. NICD with Endogenous NAD Released from Lysed Erythrocytes (A) and Differential Sensitivity of Immature and Mature T Cells toNICD and NAD-Induced CD62L Shedding (B)
(A) ART2�/� (top) or ART2�/� (bottom) T cells were incubated for 30 min in the absence or presence of erythrocyte lysates before stainingwith Annexin V/PI. Cells in panels 3–6 were preincubated with 0.1 �g/ml control antibodies (panel 3), 0.1 �g/ml ART2-specific antibodies(panel 4), 10 �M etheno-NAD (panel 5), or 2 mM KN-62 (panel 6) for 60 min prior to and during addition of erythrocyte lysates. Cells in panel7 were treated with erythrocyte lysate in the presence of NADase. The NAD concentration in the working solution was 11 �M.(B) Thymocytes (top) and lymph node T cells (bottom) were incubated for 30 min with 25 �M NAD or 250 �M ATP and then stained withAnnexin V/PI or with anti-CD3FITC and CD62LPE.(C) Thymocytes, total lymph node cells (LNC), and purified lymph node T cells were incubated with K1G preimmune (pIS) or immune (IS)serum (1:2000 dilution) for 30 min, washed, and stained with PE-conjugated goat anti-rabbit IgG and anti-CD3FITC prior to FACS analysis.
other cell surface proteins on T cells (Figure 5). This cell shrinkage, DNA fragmentation, and staining by pro-pidium iodide (Figures 1 and 3). Cells are protected fromactivates P2X7 and triggers a characteristic series of
responses ultimately resulting in T cell death by apopto- NICD both by ART2 antagonists and antibodies (Figure1) and by P2X7 antagonists and antibodies (Figures 3Esis: increased calcium flux, shedding of CD62L, expo-
sure of phosphatidylserine, opening of membrane and 5B). Further, NICD requires ADP-ribosylation (Figure1) but is blocked by etheno-ADP-ribosylation (Figurepores, breakdown of mitochondrial membrane potential,

NAD-Induced Cell Death579
2). Consistently, ADP-ribosylation agonizes and etheno- over, ADP-ribosylation would provide a longer lastingADP-ribosylation antagonizes P2X7 responses at low signal (Figures 6B and 6C).concentrations of ATP (Figures 6D and 6E). Sensitivity of T cells to NICD correlates with the re-
Proposed roles of P2X7 in immunity include the re- ported T cell differentiation-dependent expression oflease of IL-1�, killing of intracellular pathogens, and ART2 and P2X7 (Chused et al., 1996; Koch-Nolte et al.,cytotoxicity toward immune cells (Canaday et al., 2002; 1999). Immature thymocytes and B cells express littleChvatchko et al., 1996; Di Virgilio et al., 1989, 2001; if any ART2 or P2X7 and are resistant to NICD as wellLabasi et al., 2002; Lammas et al., 1997; MacKenzie et as to ATP-induced cell death (Figure 7B). Following mi-al., 2001; Zanovello et al., 1990). However, millimolar togenic stimulation, T cells show reduced sensitivity toconcentrations of exogenous ATP were required to elicit NICD consequent to shedding of ART2 (Kahl et al., 2000).these P2X7-mediated responses. It has remained a puz- It is not unlikely that NICD and other cell death pro-zle as to how such high ATP concentrations could be grams play distinct roles in immune responses. Sincederived from natural sources. Here we describe an alter- thymocytes are resistant to NICD (Figure 7B) and sincenative way of activating P2X7 on T cells: by NAD-depen- ART2KO mice show normal numbers and a normal distri-dent ADP-ribosylation. bution of thymic and peripheral T cells (Ohlrogge et al.,
The EC50 for NAD-mediated activation of P2X7 is in 2002), we do not expect NICD to play a decisive role inthe micromolar range (i.e., 2 �M for PS exposure) (Figure the generation and maintenance of the T cell repertoire.6A). We demonstrate that sufficient concentrations of Rather, we expect NICD to operate in settings whereecto-NAD to activate P2X7 can be reached upon rupture NAD is released from cells, for example, during mechani-of cells, e.g., erythrocytes. Our results show that NAD cal tissue injury or microbial inflammatory processesis a major cytotoxic principle in cell lysates, since cyto- with severe cytolysis. Under such circumstances, thetoxicity is blocked by NADase or by preincubation of massive release of intracellular antigens combined withcells with etheno-NAD or with ART2-antibodies and high local concentrations of inflammatory cytokinessince ART2-deficient T cells are resistant to lysate- would raise the danger of serendipitous activation ofinduced cytotoxicity (Figure 7A). Of note, freshly pre- autoreactive bystander T cells. Microbe-specific T cellspared ART2�/� T cells always contained a small fraction would become activated by professional antigen-pre-(5%–15%) of apoptotic, Annexin V�/PI� cells and some senting cells, and this strong stimulation would induce(5%–15%) dead, PI� cells (Figure 1A), whereas freshly shedding of ART2 (Kahl et al., 2000), thereby renderingprepared ART2�/� T cells usually contained few if any such cells resistant to NICD. NAD-dependent ADP-ribo-apoptotic cells but did contain some dead cells (Figure sylation, however, would occur on ART2-expressing by-1B). These dead cells most likely result from mechanical stander T cells—including cells bearing an irrelevantdamage incurred by tissue disruption during cell prepa- TCR or an autoantigen-specific TCR. We propose thatration. Since apoptotic cells appear in an ART2-depen- NICD thereby provides a safeguard mechanism againstdent manner, we surmise that apoptosis in these cells the undesirable activation of irrelevant and potentiallywas also triggered by NAD released from lysed cells. autoreactive bystander T cells during an inflammatoryThat NAD can be released into the extracellular environ- reaction. In this context it is of interest to note that ament in pathophysiological situations can also be sur- lack of ART2-expressing cells has been correlated withmised from the recently reported finding of ADP-ribosy- enhanced sensitivity to autoimmune disease in severallated extracellular proteins in inflammatory exudates, animal models (Ablamunits et al., 2001; Greiner et al.,i.e., broncheolar lavage of smokers (Paone et al., 2002). 1987).
How does ADP-ribosylation activate P2X7? Cova- This study uncovers the molecular mechanism of thelently immobilized ADP-ribose could function as a ligand activation of a purinoceptor by a toxin-related endoge-for the adenosine binding site. Alternatively, ADP-ribo- nous ecto-ADP-ribosyltransferase. The results providesylation could activate P2X7 by an allosteric conforma-
new insights into the role of ecto-ARTs in regulatingtional change independent of the ligand binding site.
T cell functions and identify P2X7 as a key downstreamThe former model more readily explains the observed
effector in NICD. These findings establish an importantdose responses of P2X7 to NAD and ATP (Figure 6A)precedent for further studies on links between ADP-as well as the agonizing effects of ADP-ribosylation (andribosylation and purinoceptors and open the excitingantagonizing effects of etheno-ADP ribosylation) onperspective of utilizing the pharmacology of NAD toP2X7 responses to ATP (Figures 6D and 6E). Cataboliza-modify the function of P2X7 and other purinoceptors.tion into a covalently immobilized ligand is compatible
with the effectivity of NAD at low concentrations. InExperimental Procedurescontrast, at saturating concentrations ATP induces
stronger P2X7 responses than NAD (e.g., compare mag- Reagents and Micenitudes of NAD versus ATP induced calcium flux and ADP-ribose, ATP, NAD, etheno-NAD, NGD, NHD, oxidized ATP,dye uptake in Figures 6E and 4B). This, in turn, is com- KN-62 (1-[N,O-bis(5-isoquinolinesulfonyl)-N-methyl-L-tyrosyl]-4-phenyl-patible with the notion that covalently attached ADP- piperazine), Neurospora crassa NAD glycohydrolase, and potato
apyrase were from Sigma Chemical Company; Fura-2, YO-PRO-1,ribose acts as a weaker agonist than ATP and is consis-ethidium bromide, and propidium iodide were from Moleculartent also with the finding that soluble ADP-ribose doesProbes; PE- and FITC-conjugated anti-B220, anti-CD62L, anti-CD3,not activate the P2X7 receptor whereas immobilizedanti-mouse Ig, and anti-rabbit Ig were from Becton Dickinson.
(covalently attached) ADP-ribose does (Figure 1C). Even Etheno-adenosine-specific mAb 1G4 (Krebs et al., 2003) was puri-if covalently attached ADP-ribose were a weaker agonist fied from hybridoma supernatant by affinity chromatography on Pro-than soluble ATP, it may be easier to produce the former tein G Sepharose (Pharmacia) and conjugated to Alexa488 (Molecu-
lar Probes) according to the manufacturer’s instructions. MD27in a physiological setting, e.g., upon lysis of cells. More-

Immunity580
lymphoma cells were sorted for high ART activity after etheno-ADP- analyses. NADase and Apyrase were added to the cells either 5 minbefore or simultaneously with NAD or ATP. All results shown areribosylation and staining with 1G4 as described previously (Krebs
et al., 2003). BALB/c mice were obtained from the Animal Resources representative of at least three similar and independent experi-ments.Units of the University Hospital Hamburg and of Diderot University,
Paris. ART2-deficient mice were generated by standard homologousrecombination procedures and backcrossed for five generations Immunoprecipitation and Western Blot Analysesonto the BALB/c background as described elsewhere (Ohlrogge et T cells (1 � 107/ml) and MD27 lymphoma cells (5 � 106/ml) wereal., 2002). Please note that ART2 is encoded by two closely related incubated for 30 min at 37�C in the presence of radiolabeled 32P-tandem genes in the mouse designated Art2.1 and Art2.2. The NAD (1 �M, 5 �Ci/ml) in RPMI medium containing 1 mM ADP-ribose.ART2.1 and ART2.2 gene products are coexpressed by T cells and Cells were then washed in cold PBS to remove free NAD. Cells wereshow similar enzymatic acitivities. All substrates, inhibitors, and lysed in 250 �l PBS, 1% Igepal, 1 mM AEBSF at 4�C for 30 min. Cellantibodies used in this study affect ART2.1 and ART2.2 to a similar lysates were precleared by centrifugation (14,000 � g at 4�C forextent, the ART2�/� mice are deficient for ART2.1 and ART2.2. For 30 min) followed by incubation with 20 �l protein G Sepharosesake of brevity “ART2” is used throughout this paper instead of (Pharmacia) for 60 min at 4�C. Immunoprecipitation was performed“ART2.1 and ART2.2”. sequentially with preimmune serum (1 �l), immune serum (1 �l), and
LFA-1-specific mAb M17/4 (1 �g), each immobilized on 20 �l proteinGeneration of ART2-Specific and P2X7-Specific Antisera G Sepharose. Proteins were size fractionated by SDS-PAGE onRabbits were immunized by repetitive gene gun immunization es- precast Nupage (10%) gels (Invitrogen) and blotted onto nitrocellu-sentially as described by Koch-Nolte et al. (1999) using expression lose (NC) membranes. Radioactivity was detected by autoradiogra-vectors for ART2.2 or P2X7 (Koch-Nolte et al., 1999, Adriouch et al., phy by exposing the membrane to Kodak X-omat Films at �80�C2001). Specificity of antisera was checked by FACS and immunopre- for 48 hr. The membrane was developed further with the enhancedcipitation analyses using HEK cells stably transfected with ART2.2 chemiluminescence system (Amersham) using a P2X7 peptide-spe-or P2X7 (Adriouch et al., 2001; Koch-Nolte et al., 1999). cific rabbit antiserum (Alomone) (1:1000) and peroxidase-conju-
gated secondary antibody (Amersham) (1:10.000) followed by a 60Assays for Phosphatidylserine Exposure, Pore Formation, s exposure to ECL hyperfilm (Amersham).Mitochondrial Membrane Potential, and Cell DeathSingle-cell suspensions from lymph nodes were prepared and pro- Acknowledgmentscessed for flow cytometry on a FACSCalibur (Becton Dickinson) asdescribed (Adriouch et al., 2001; Koch-Nolte et al., 1999). B cells This work was supported by grants from the Deutsche Forschungs-were depleted using magnetic cell separation with Dynabead-immo- gemeinschaft (SFB545/B9) and the Bundesministerium fur Wirt-bilized goat anti-mouse IgG (Dynal). Purity of T cells was always schaft und Technologie to F.H. and F.K.-N. and from the Ministeregreater than 95% as verified by FACS analyses using PE-conjugated de la Recherche et de la Technologie to M.S. S.A. was a recipientanti-B220 and FITC-conjugated anti-CD3. Following treatment with of a fellowship from the Association pour la Recherche sur le Cancer.NAD or ATP at 37�C, cells were washed in RPMI medium (Life Tech- C.K. is a recipient of a fellowship from the Werner Otto Foundation.nologies) supplemented with 2 mM CaCl2, and were then stained We thank Wiebke Ohlrogge, Claudia Dox, Fenja Braasch, Gudrunfor 20 min on ice with FITC-conjugated Annexin V (1 �g/ml) (Becton Dubberke, and Vivienne Welge (Hamburg) and S. Pechberty (Paris)Dickinson) and propidium iodide (10 �g/ml) or PE-conjugated anti- for technical assistance. Parts of the work described in this studymouse IgG. For assay of pore formation, YO-PRO-1 (10 �g/ml) or represent the partial fulfilment of the requirements for the graduateethidium bromide (1 �g/ml) was added for the last 2 min of incuba- theses of S.A., F.S., and C.K. F.K.-N., F.H., and M.S. designed andtion. For pulse-chase analyses, cells were treated with NAD or ATP supervised the study with essential contributions by P.D. to thefor 5 min at 37�C. Cells were then washed and incubated further for calcium flux experiments. F.S., C.K., D.F., F.H., and F.K.-N. per-the times indicated prior to staining with Annexin V/PI as above. For formed the experiments described in Figures 1, 2A, 2B, 3B–3D, 4,assay of mitochondrial membrane potential, cells were incubated for 5, 6A–6C, and 7; S.A. and M.S. those in Figures 2C, 2D, 3A, 3E, 3F,20 min at 37�C with the cationic dye MitoSensor (Becton Dickinson), 6D, and 6E; G.G. and F.H. made the P2X7-antiserum. F.K.-N. wrotewashed, and assayed by flow cytometry according to the manufac- the paper. The authors are indebted to the reviewers and the editorsturer’s protocol. For assay of calcium flux, Fura-2 loaded T cells of Immunity for helpful suggestions. We thank Drs. Edward H. Leiter,were stirred in a fluorimeter cuvette. KN-62, NAD, eNAD, or ATP Francesco Di Virgilio, Bernhard Fleischer, and Stefan Rothenburgwas added as indicated, and changes in cytosolic Ca2� were moni- for critical reading of the manuscript.tored by fluorometry with a SAFAS spectrofluorimeter with excita-tion at 340/380 nm and emission at 510 nm. Concentration of Ca2�
Received: June 27, 2003was calculated by using the equation of Grinkievich et al. (1985).Revised: August 13, 2003Accepted: August 20, 2003
Preparation of Cell LysatesPublished: October 14, 2003
Mouse erythrocytes were washed and resuspended in 5 volumesof PBS containing 1 mM ADP-ribose. Cells were ruptured either
Referencesby ultrasonic treatment or by two cycles of snap-freezing in liquidnitrogen and thawing in a 37�C waterbath. Cellular debris was re-
Abbracchio, M.P., and Burnstock, G. (1994). Purinoceptors: are theremoved by centrifugation (15.000 � g) for 15 min at 4�C, and aliquotsfamilies of P2X and P2Y purinoceptors? Pharmacol. Ther. 64,of clarified lysates (50 �l) were added to cell suspensions in RPMI445–475.medium (100 �l, 4 � 105 cells). NAD concentrations in cell lysates
were determined by a sensitive cycling assay (Jacobson and Jacob- Ablamunits, V., Bridgett, M., Duffy, T., Haag, F., Nissen, M., Koch-son, 1997). Nolte, F., and Leiter, H. (2001). Changing patterns of cell surface
mono (ADP-ribosyl) transferase antigen ART2.2 on resting versusProtection of Cells from Apoptosis with Antibodies, Etheno-NAD, cytopathically-activated T cells in NOD/Lt mice. Diabetologia 44,NGD, NHD, P2X7-Antagonists, NAD-Glycohydrolase, or Apyrase 848–858.Cells (4 � 105/100 �l) were preincubated at 37�C in microtiter plates Adriouch, S., Ohlrogge, W., Haag, F., Koch-Nolte, F., and Seman,in RPMI medium containing antibodies, etheno-NAD, NGD, NHD, M. (2001). Rapid induction of naive T cell apoptosis by ecto-nicotin-oxidized ATP, or KN-62 as indicated. In the case of antibodies and amide adenine dinucleotide: requirement for mono(ADP-ribosyl)-KN-62, NAD or ATP was then added as indicated, and incubation transferase 2 and a downstream effector. J. Immunol. 167, 196–203.continued in the presence of antibodies or KN-62 at 37�C prior to
Aktories, K., and Just, I. (2000). Bacterial Protein Toxins (Berlin:staining and FACS analyses. In the case of etheno-NAD, NGD, NHD,Springer Verlag).and oxidized ATP, cells were washed before addition of NAD and
ATP, and then incubated further at 37�C prior to staining and FACS Banasik, M., Komura, H., Shimoyama, M., and Ueda, K. (1992). Spe-

NAD-Induced Cell Death581
cific inhibitors of poly(ADP-ribose) synthetase and mono(ADP-ribo- enzymatically active mouse ecto-ADP-ribosyltransferase ART2.2syl)transferase. J. Biol. Chem. 267, 1569–1575. upon T cell activation. J. Immunol. 165, 4463–4469.
Bossy-Wetzel, E., and Green, D.R. (2000). Detection of apoptosis Koch-Nolte, F., Petersen, D., Balasubramanian, S., Haag, F., Kahlke,by annexin V labeling. Methods Enzymol. 322, 15–18. D., Willer, T., Kastelein, R., Bazan, F., and Thiele, H.G. (1996). Mouse
T cell membrane proteins Rt6-1 and Rt6-2 are arginine/proteinBruzzone, S., Guida, L., Zocchi, E., Franco, L., and De Flora, A.(2001). Connexin 43 hemi channels mediate Ca2�-regulated trans- mono(ADPribosyl)transferases and share secondary structure mo-membrane NAD� fluxes in intact cells. FASEB J. 15, 10–12. tifs with ADP-ribosylating bacterial toxins. J. Biol. Chem. 271, 7686–
7693.Canaday, D.H., Beigi, R., Silver, R.F., Harding, C.V., Boom, W.H.,and Dubyak, G.R. (2002). ATP and control of intracellular growth of Koch-Nolte, F., Duffy, T., Nissen, M., Kahl, S., Killeen, N., Ablamunits,mycobacteria by T cells. Infect. Immun. 70, 6456–6459. V., Haag, F., and Leiter, E.H. (1999). A new monoclonal antibody
detects a developmentally regulated mouse ecto-ADP-ribosyltrans-Chused, T.M., Apasov, S., and Sitkovsky, M. (1996). Murine T lym-phocytes modulate activity of an ATP-activated P2Z-type purino- ferase on T cells: subset distribution, inbred strain variation, andceptor during differentiation. J. Immunol. 157, 1371–1380. modulation upon T cell activation. J. Immunol. 163, 6014–6022.
Chvatchko, Y., Valera, S., Aubry, J.P., Renno, T., Buell, G., and Krammer, P.H. (2000). CD95’s deadly mission in the immune system.Bonnefoy, J.Y. (1996). The involvement of an ATP-gated ion channel, Nature 407, 789–795.P(2X1), in thymocyte apoptosis. Immunity 5, 275–283.
Krebs, C., Koestner, W., Nissen, M., Welge, V., Parusel, I., Malavasi,Contreras, J.E., Sanchez, H.A., Eugenin, E.A., Speidel, D., Theis, M., F., Leiter, E.H., Santella, R.M., Haag, F., and Koch-Nolte, F. (2003).Willecke, K., Bukauskas, F.F., Bennett, M.V., and Saez, J.C. (2002).
Flow cytometric and immunoblot assays for cell surface ADP-ribo-Metabolic inhibition induces opening of unapposed connexin 43
sylation using a monoclonal antibody specific for ethenoadenosine.gap junction hemichannels and reduces gap junctional communica-
Anal. Biochem. 314, 108–115.tion in cortical astrocytes in culture. Proc. Natl. Acad. Sci. USA99, 495–500. Labasi, J.M., Petrushova, N., Donovan, C., McCurdy, S., Lira, P.,
Payette, M.M., Brissette, W., Wicks, J.R., Audoly, L., and Gabel,Dive, C., Gregory, C.D., Phipps, D.J., Evans, D.L., Milner, A.E., andC.A. (2002). Absence of the P2X7 receptor alters leukocyte functionWyllie, A.H. (1992). Analysis and discrimination of necrosis and apo-and attenuates an inflammatory response. J. Immunol. 168, 6436–ptosis (programmed cell death) by multiparameter flow cytometry.6445.Biochim. Biophys. Acta 1133, 275–285.
Lammas, D.A., Stober, C., Harvey, C.J., Kendrick, N., Panchalingam,Di Virgilio, F., Bronte, V., Collavo, D., and Zanovello, P. (1989). Re-sponses of mouse lymphocytes to extracellular adenosine 5�-tri- S., and Kumararatne, D.S. (1997). ATP-induced killing of mycobac-phosphate (ATP). Lymphocytes with cytotoxic activity are resistant teria by human macrophages is mediated by purinergic P2Z(P2X7)to the permeabilizing effects of ATP. J. Immunol. 143, 1955–1960. receptors. Immunity 7, 433–444.
Di Virgilio, F., Chiozzi, P., Ferrari, D., Falzoni, S., Sanz, J.M., Morelli, Leist, M., and Jaattela, M. (2001). Four deaths and a funeral: fromA., Torboli, M., Bolognesi, G., and Baricordi, O.R. (2001). Nucleotide caspases to alternative mechanisms. Nat. Rev. Mol. Cell. Biol. 2,receptors: an emerging family of regulatory molecules in blood cells. 589–598.Blood 97, 587–600.
Liu, Z.X., Yu, Y., and Dennert, G. (1999). A cell surface ADP-ribosyl-Domenighini, M., and Rappuoli, R. (1996). Three conserved consen-
transferase modulates T cell receptor association and signaling. J.sus sequences identify the NAD-binding site of ADP-ribosylating
Biol. Chem. 274, 17399–17401.enzymes, expressed by eukaryotes, bacteria and T-even bacterio-phages. Mol. Microbiol. 21, 667–674. Liu, Z.X., Azhipa, O., Okamoto, S., Govindarajan, S., and Dennert,
G. (2001). Extracellular nicotinamide adenine dinucleotide inducesGargett, C.E., and Wiley, J.S. (1997). The isoquinoline derivativeT cell apoptosis in vivo and in vitro. J. Immunol. 167, 4942–4947.KN-62 a potent antagonist of the P2Z-receptor of human lympho-
cytes. Br. J. Pharmacol. 120, 1483–1490. Lund, F., Solvason, N., Grimaldi, J.C., Parkhouse, R.M., and Howard,Glowacki, G., Braren, R., Firner, K., Nissen, M., Kuhl, M., Reche, P., M. (1995). Murine CD38: an immunoregulatory ectoenzyme. Immu-Bazan, F., Cetkovic-Cvrlje, M., Leiter, E., Haag, F., and Koch-Nolte, nol. Today 16, 469–473.F. (2002). The family of toxin-related ecto-ADP-ribosyltransferases
MacKenzie, A., Wilson, H.L., Kiss-Toth, E., Dower, S.K., North, R.A.,in humans and the mouse. Protein Sci. 11, 1657–1670.and Surprenant, A. (2001). Rapid secretion of interleukin-1� by mi-
Goding, J.W., and Howard, M.C. (1998). Ecto-enzymes of lymphoid crovesicle shedding. Immunity 15, 825–835.cells. Immunol. Rev. 161, 5–10.
Mueller-Dieckmann, C., Ritter, H., Haag, F., Koch-Nolte, F., andGreiner, D.L., Mordes, J.P., Handler, E.S., Angelillo, M., Nakamura,
Schulz, G. (2002). Structure of the ecto-ADP-ribosyl transferaseN., and Rossini, A.A. (1987). Depletion of RT6.1� T lymphocytesART2.2 from rat. J. Mol. Biol. 322, 687–696.induces diabetes in resistant biobreeding/Worcester (BB/W) rats.
J. Exp. Med. 166, 461–475. North, R.A., and Surprenant, A. (2000). Pharmacology of cloned P2Xreceptors. Annu. Rev. Pharmacol. Toxicol. 40, 563–580.Gu, B., Bendall, L.J., and Wiley, J.S. (1998). Adenosine triphosphate-
induced shedding of CD23 and L-selectin (CD62L) from lymphocytes Ohlrogge, W., Haag, F., Lohler, J., Seman, M., Littman, D.R., Killeen,is mediated by the same receptor but different metalloproteases. N., and Koch-Nolte, F. (2002). Generation and characterization ofBlood 92, 946–951. ecto-ADP-ribosyltransferase ART2.1/ART2.2-deficient mice. Mol.Haag, F., and Koch-Nolte, F. (1997). ADP-Ribosylation in Animal Cell. Biol. 22, 7535–7542.Tissues: Structure, Function and Biology of Mono(ADP-Ribo-
Okada, Y., Maeno, E., Shimizu, T., Dezaki, K., Wang, J., and Mori-syl)transferases and Related Enzymes, Volume 419 (New York: Ple-shima, S. (2001). Receptor-mediated control of regulatory volumenum Press).decrease (RVD) and apoptotic volume decrease (AVD). J. Physiol.
Jacobson, E.L., and Jacobson, M.K. (1997). Tissue NAD as a bio- 532, 3–16.chemical measure of niacin status in humans. Methods Enzymol.
Okazaki, I.J., and Moss, J. (1998). Glycosylphosphatidylinositol-280, 221–230.anchored and secretory isoforms of mono-ADP-ribosyltransferases.Jamieson, G.P., Snook, M.B., Thurlow, P.J., and Wiley, J.S. (1996).J. Biol. Chem. 273, 23617–23620.Extracellular ATP causes of loss of L-selectin from human lympho-
cytes via occupancy of P2Z purinocepters. J. Cell. Physiol. 166, Paone, G., Wada, A., Stevens, L.A., Matin, A., Hirayama, T., Levine,637–642. R.L., and Moss, J. (2002). ADP ribosylation of human neutrophil
peptide-1 regulates its biological properties. Proc. Natl. Acad. Sci.Kahl, S., Nissen, M., Girisch, R., Duffy, T., Leiter, E.H., Haag, F.,and Koch-Nolte, F. (2000). Metalloprotease-mediated shedding of USA 99, 8231–8235.

Immunity582
Surprenant, A., Rassendren, F., Kawashima, E., North, R.A., andBuell, G. (1996). The cytolytic P2Z receptor for extracellular ATPidentified as a P2X receptor (P2X7). Science 272, 735–738.
Van Parijs, L., and Abbas, A.K. (1998). Homeostasis and self-toler-ance in the immune system: turning lymphocytes off. Science280, 243–248.
Zanovello, P., Bronte, V., Rosato, A., Pizzo, P., and Di Virgilio, F.(1990). Responses of mouse lymphocytes to extracellular ATP. II.Extracellular ATP causes cell type-dependent lysis and DNA frag-mentation. J. Immunol. 145, 1545–1550.





























