Embed Size (px)
Citation preview

441ZOOSYSTEMA • 2005 • 27 (3) © Publications Scientifiques du Muséum national d’Histoire naturelle, Paris. www.zoosystema.com
Neotropical Monogenoidea. 47. Phylogeny and coevolution of species of Rhinoxenus(Platyhelminthes, Monogenoidea,Dactylogyridae) and their Characiformes hosts(Teleostei, Ostariophysi) with description of four new species
Marcus V. DOMINGUESDepartamento de Zoologia, Universidade Federal do Paraná, Curitiba,
Caixa Postal 19073, PR 81.531-990 (Brazil)and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq)
Walter A. BOEGERDepartamento de Zoologia, Universidade Federal do Paraná, Curitiba,
Caixa Postal 19073, PR 81.531-990 (Brazil)and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq)
Domingues M. V. & Boeger W. A. 2005. — Neotropical Monogenoidea. 47. Phylogeny andcoevolution of species of Rhinoxenus (Platyhelminthes, Monogenoidea, Dactylogyridae)and their Characiformes hosts (Teleostei, Ostariophysi) with description of four newspecies. Zoosystema 27 (3) : 441-467.
ABSTRACTEight species of Rhinoxenus are reported from nasal cavities of characiform fish-es in the Neotropics: Rhinoxenus arietinus Kritsky, Boeger & Thatcher, 1988,from anostomid hosts; R. bulbovaginatus Boeger, Domingues & Pavanelli,1995, from characid hosts; R. nyttus Kritsky, Boeger & Thatcher, 1988, fromanostomid hosts; R. piranhus Kritsky, Boeger & Thatcher, 1988, from ser-rasalmin hosts; R. anaclaudiae n. sp., from characid hosts; R. euryxenus n. sp.,from serrasalmin and anostomid hosts; R. curimbatae n. sp., from prochilodon-tids hosts; and R. guianensis n. sp., from curimatid hosts. Furthermore, severalundeterminate species are reported from Hydrolicus scomberoides (Cyno-dontidae). Hosts were collected from rivers in Brazil and French Guiana. Thehypothesis on the sister-group relationships of species of Rhinoxenus (C.I. =66%; R.I. = 66%), based on 10 transformation series, is ((R. nyttus, R. curim-batae n. sp.) (R. bulbovaginatus (R. arietinus (R. guianensis n. sp., R. anaclaudiaen. sp. (R. euryxenus n. sp., R. piranhus))))). Coevolutionary analysis suggeststhat events of cospeciation, duplication, dispersion, and extinction are requiredto explain the observed host-parasite association. The origin of R. curimbataen. sp., R. nyttus, R. bulbovaginatus, R. anaclaudiae n. sp., and the ancestor ofR. piranhus + R. euryxenus n. sp. are putatively associated with events of cospe-ciation with the ancestors of their respective host families. Dispersion withsubsequent allopatric speciation to Anostomidae and Curimatidae appear asso-ciated with the origins of R. arietinus and R. guianensis n. sp., respectively.Extinction is necessary to explain the absence of Rhinoxenus spp. on severalother characiform families, although sampling density cannot be discarded.
KEY WORDSPlatyhelminthes, Monogenoidea, Dactylogyridae,
Ancyrocephalinae, Characiformes,
phylogeny, coevolution, new species.

INTRODUCTION
Characiformes (sensu Fink & Fink 1981) is divid-ed into 16 (Greenwood et al. 1966) or 14 families(Géry 1977) with species endemic to Africa (fourfamilies; about 200 species) and the Neotropics(12 families; more than 1200 species) (Ortí &Meyer 1997). The most important advances inthe systematics of the group occurred during thelatter part of the 19th and much of the 20th cen-tury. Some of the advances were based primarilyon anatomical features, which lacked a rigorousanalytical method, resulting in many divergentclassifications and largely untestable hypotheseson the evolutionary history of the group (Vari1998). However, the application of phylogenetic
methods and the use of other systems of charac-ters (e.g., molecular, cytogenetic) resulted inmore stable phylogenies, based on testablehypotheses. Vari (1998) provides an overview ofthe phylogenetic concepts utilized for proposal ofthe major lineages within what is now recognizedas the order Characiformes.Species of Rhinoxenus Kritsky, Thatcher &Boeger, 1988 are all parasites of the nasal cavityof characiform fishes. Boeger et al. (1995) suggestthat these species comprise a monophyletic groupwith the origin of their species associated with thedivergence of ancestor species of their hosts. Thissuggests the possible usefulness of these parasitesas evolutionary markers for the sister-group rela-tionships of the families of their characiform
Domingues M. V. & Boeger W. A.
442 ZOOSYSTEMA • 2005 • 27 (3)
RÉSUMÉMonogenoidea néotropicaux. 47. Phylogénie et coévolution des espèces deRhinoxenus (Plathelminthes, Monogenoidea, Dactylogyridae) et de leurs hôtesCharaciformes (Teleostei, Ostariophysi) et description de quatre nouvelles espèces.Huit espèces de Rhinoxenus sont rapportées des cavités nasales de poissonscharaciformes de la région néotropicale : Rhinoxenus arietinus Kritsky, Boeger& Thatcher, 1988, chez des Anostomidae ; R. bulbovaginatus Boeger,Domingues & Pavanelli, 1995, chez des Characidae ; R. nyttus Kritsky,Boeger & Thatcher, 1988, chez des Anostomidae ; R. piranhus Kritsky,Boeger & Thatcher, 1988, chez des Serrasalminae ; R. anaclaudiae n. sp., chezdes Characidae ; R. euryxenus n. sp., chez des Serrasalminae et desAnostomidae ; R. curimbatae n. sp., chez des Prochilodontidae ; et R. guianensisn. sp., chez des curimatidae. De plus, plusieurs espèces indéterminées deRhinoxenus sont rapportées chez Hydrolicus scomberoides (Cynodontidae). Leshôtes ont été récoltés dans des rivières du Brésil et de Guyane française.L’hypothèse sur les relations phylogénétiques des espèces de Rhinoxenus (C.I. =66 % ; R.I. = 66 %), basée sur 10 séries de transformations, est : ((R. nyttus,R. curimbatae n. sp.) (R. bulbovaginatus (R. arietinus (R. guianensis n. sp.,R. anaclaudiae n. sp. (R. euryxenus n. sp., R. piranhus))))). L’analyse coévolu-tive suggère que des événements de cospéciation, duplication, dispersion etextinction sont requis pour expliquer l’association hôte-parasite observée.Nous faisons l’hypothèse que l’origine de R. curimbatae n. sp., R. nyttus,R. bulbovaginatus, R. anaclaudiae n. sp. et l’ancêtre de R. piranhus + R. euryxenusn. sp. sont associés à des événements de cospéciation avec des ancêtres de leursfamilles d’hôtes respectives. La dispersion avec une spéciation allopatriquesubséquente vers les Anostomidae et des Curimatidae semble avoir été asso-ciée avec l’origine de R. arietinus et R. guianensis n. sp., respectivement.L’extinction est nécessaire pour expliquer l’absence de Rhinoxenus spp. chezplusieurs autres familles de Characiformes, bien qu’un biais dû à l’échan-tillonnage ne puisse être écarté.
MOTS CLÉSPlathelminthes, Monogenoidea, Dactylogyridae,
Ancyrocephalinae, Characiformes,
phylogénie, coévolution,
nouvelles espèces.

hosts. Thus, this study tests the hypothesis ofBoeger et al. (1995) by re-evaluating the hypoth-esis of cospeciation by the inclusion of newspecies described herein and a re-evaluation ofthe morphological matrix.
MATERIAL AND METHODS
Fish hosts were collected from Brazil and FrenchGuiana. A list of necropsied species for this studywith collection data and number of examinedspecimens is presented in the Appendix. Methodsof parasite collection, preparation, measurement,and illustration follow Kritsky et al. (1986, 1988).Measurements are in μm. Type specimens andvouchers are deposited in the parasite collectionsof the Coleção Helmintológica do InstitutoOswaldo Cruz, Rio de Janeiro, RJ, Brazil (CHIOC);the Instituto Nacional de Pesquisas da Amazônia,Manaus, AM, Brazil (INPA); Muséum national d’His-toire naturelle, Paris, France (MNHN); Museu deZoologia, Universidade de Sao Paulo (MZUSP);and the U.S. National Museum, HelminthologicalCollection, Beltsville, Maryland, USA (USNPC),as indicated in the respective descriptions.The phylogenetic relationship of Rhinoxenus spp.is determined using techniques of phylogeneticsystematics (sensu Hennig 1966). The transforma-tion series and character states were defined basedon the literature or through the study of voucherand type specimens. An initial hypothesis onevolutionary relationships of Rhinoxenus specieswas constructed manually using HennigianArgumentation (Wiley 1981) and tested with theprogram PAUP* 4.0b10 (Swofford 2002) usingHSEARCH, unlimited MAXTREE, and1000 random addition sequence (set maxtrees =10000, increase = auto, hsearch addseq = random,hold = 2, nreps = 1000, rstatus = yes) to confirmthat the tree obtained was a most-parsimonioustree. Bootstrap values were calculated based on1000 replicates and were considered supportivewhen ≥ 50%. A strict consensus cladogram wasdeveloped to evaluate branch support from allmost-parsimonious trees. All transformation serieswere considered initially unordered and were
polarized and optimized according to the tech-niques described by Watrous & Wheeler (1981)and Maddison et al. (1984). Protorhinoxenusprochilodi Domingues & Boeger, 2002 was used asoutgroup (Domingues & Boeger 2002).The coevolutionary analysis was performed usingBPA (Brooks Parsimony Analysis) as described byBrooks (1981, 1990), Brooks & McLennan(1991, 1993) and Brooks et al. (2001). The hostcladogram used is that proposed by Buckup(1993, 1998). Triportheus and Salminus areincluded in Characoidea (sensu Buckup 1998)based on their relationship with Brycon, as sug-gested by Malabarba (1998). Only taxa reportedas hosts of Rhinoxenus spp. or scrutinized in thisstudy were included in the cladogram. The para-site phylogeny was transformed into a matrix byaddictive binary coding; each parasite species wascoded as to indicate both its identity and ancestor(evolutionary history). A matrix for the hostgroups was then constructed based on this para-sites coding based on the parasites distribution(Table 10); hosts without parasites were coded bymissing or inapplicable data (?). Hosts with morethan one species of parasite were considered asindependent entities to allow the recognition ofthe mechanisms responsible for these associations(Brooks & McLennan 1991, 1993; Boeger &Kritsky 1997). A host phylogeny was then recon-structed based solely on parasitological informa-tion and compared with the phylogenetichypotheses based on host characters. The codedparasite cladogram was mapped onto the hostcladogram to determine congruence.
SYSTEMATICS
Subclass POLYONCHOINEA Bychowsky, 1937Order DACTYLOGYRIDEA Bychowsky, 1937
Family DACTYLOGYRIDAE Bychowsky, 1933
Genus RhinoxenusKritsky, Boeger & Thatcher, 1988
TYPE SPECIES. — Rhinoxenus piranhus Kritsky, Boeger& Thatcher, 1988, from Pygocentrus nattereri Kner,1858 (Characidae).
Phylogeny and coevolution of Rhinoxenus (Platyhelminthes, Monogenoidea) and their hosts
443ZOOSYSTEMA • 2005 • 27 (3)

OTHER SPECIES INCLUDED. — Rhinoxenus arietinusKritsky, Boeger & Thatcher, 1988 from Leporinusagassizii Steindachner, 1876, L. elongatus Valen-ciennes, 1849, L. friderici friderici (Bloch, 1794),L. lacustris Amaral Campos, 1945, L. obtusidens(Valenciennes, 1836), Schizodon altoparanae Garavello& Britski, 1990, S. borellii (Boulenger, 1990), S. fas-ciatus Spix & Agassiz, 1829, Schizodon sp. andRhytiodus argenteofuscus Kner, 1858 (Anostomidae);R. bulbovaginatus Boeger, Domingues & Pavanelli,1995 from Salminus brasiliensis (Cuvier, 1816)(Characidae); R. nyttus Kritsky, Boeger & Thatcher,1988 from Schizodon fasciatus and Schizodon sp.(Anostomidae); R. piranhus from Prystobricon sp.,Serrasalmus marginatus Valenciennes, 1836 and S. spi-lopleura Kner, 1858 (Characidae); R. anaclaudiaen. sp., from Triportheus cf. nematurus, Triportheus sp.and Brycon sp. (Characidae); R. curimbatae n. sp.,from Prochilodus cf. lineatus (Prochilodontidae);R. euryxenus n. sp. from Serrasalmus gouldingi Fink &Machado Allison, 1992, S. marginatus, S. rhombeus(Linnaeus, 1766), S. spliopleura, S. striolatusSteindachner, 1908 (Characidae), Leporinus agassizii(Anostomidae); R. guianensis n. sp., from Curimatacyprinoides (Linnaeus, 1766) (Curimatidae); and severalundeterminate Rhinoxenus species from Hydrolicusscomberoides (Cuvier, 1816) (Cynodontidae).
EMENDED DIAGNOSIS. — Dactylogyridae. Body divid-ed into cephalic region, trunk, haptor (peduncleabsent). Tegument smooth. Cephalic lobes, headorgans, cephalic glands present. Eyes four. Pharynxmuscular, glandular; intestinal ceca two, confluent inposterior trunk, lacking diverticula. Gonads overlap-ping; testis dorsal to germarium. Common genitalpore near bifurcation of intestinal caeca. Vas deferenslooping left intestinal cecum; seminal vesicle a dilationof vas deferens. Copulatory complex comprising a scle-rotized male copulatory organ (MCO), accessorypiece; MCO a coiled tube with counterclockwise rings(Kritsky et al. 1985, 1988), with conical base sur-rounded by two circular sclerotized tandem brims;proximal brim articulated to copulatory ligament ofaccessory piece. Copulatory ligament lying withinrings of MCO; distal portion of the accessory pieceexpanded. Vagina sclerotized, sinistral, proximal por-tion of vagina with one or more loops. Seminal recep-tacle present. Vitellaria follicular. Haptor with 14hooks, having ancyrocephaline distribution (Mizelle1936); hook comprising shank of two subunits; pair ofventral anchors, a pair of spike-like dorsal anchors;ventral bar; dorsal bar absent. Parasites of the nasalcavity of the Neotropical characiform fishes.
REMARKS
The general features of the internal morphologyof Rhinoxenus species are well documented byKritsky et al. (1988) and Boeger et al. (1995).
Diagnostic features of the genus include: 1) pres-ence of dorsal anchor modified into a spike-likesclerite; 2) absence of dorsal bar; 3) presence ofhook from pair 2 located in two bilateral lobes ofthe trunk; and 4) two circular sclerotized tandembrims on the base of the MCO. The originaldiagnosis of Rhinoxenus does not include the twocircular tandem sclerotized brims associated withthe base of the MCO. These structures have beenreported in species of Dawestrema (Kritsky et al.1985) and of Cacatuocotyle (Boeger et al. 1997)but probably represent independent evolutionaryevents. Thus, the presence of these brims is con-sidered a synapomorphy for Rhinoxenus. Kritskyet al. (1988) report that species of Rhinoxenuspresent all hooks with shank inflated proximally.Rhinoxenus curimbatae n. sp. is the only species ofthe genus that does not present this character.However, the absence of the shank inflationseems to be a second autapomorphic loss.
Rhinoxenus piranhusKritsky, Boeger & Thatcher, 1988
(Fig. 1A, B)
Rhinoxenus piranhus Kritsky, Boeger & Thatcher,1988: 88, 89, figs 1-8.
TYPE HOST AND LOCALITY. — Nasal cavities ofPygocentrus nattereri (Characidae): Ilha Marchantaria,Rio Solimões, near Manaus, Amazonas, Brazil,21.IX.1983, 15.VIII .1984, 14.IX.1985, 25-26.XI.1984.
MATERIAL EXAMINED. — Brazil. Rio Capucapu,Cachoeira das Garças, near Manaus, Amazonas, fromPristobrycon sp. (Characidae), 1.XI.1989, voucherspecimens (INPA 446). — Rio Paraná, near PortoRico, Paraná, from Serrasalmus marginatus(Characidae), 1992, voucher specimens (INPA 445);same locality, from Serrasalmus spilopleura(Characidae), 1992, voucher specimens (INPA 444).— Rio Uatumã, Lago Tapana, Santa Anna,Amazonas, from Serrasalmus spilopleura (Characidae),2-3.XI.1989, voucher specimens (INPA 441 a-f;MNHN 173 HG-T1 216-216 bis; USNPC 95241).French Guiana. Lagoa Manga, igarapé Tapardou,from Pygocentrus nattereri (Characidae), 15-16.X.1996, voucher specimens (INPA 442 a-b;MNHN 174 HG-T1 217; USNPC 95242); samelocality, from Serrasalmus spilopleura (Characidae),15-16.X.1996, voucher specimens (INPA 443;MNHN 175 HG-T1 217 bis).
Domingues M. V. & Boeger W. A.
444 ZOOSYSTEMA • 2005 • 27 (3)

Phylogeny and coevolution of Rhinoxenus (Platyhelminthes, Monogenoidea) and their hosts
445ZOOSYSTEMA • 2005 • 27 (3)
A
C
B
D
E
F
A-D, F
FIG. 1. — Sclerotized structures of selected Rhinoxenus species; A, R. piranhus Kritsky, Boeger & Thatcher, 1988, male copulatoryorgan (MCO); B, R. piranhus, vagina; C, R. nyttus Kritsky, Boeger & Thatcher, 1988, MCO; D, R. nyttus, detail of the base of the MCO(distal portion of MCO not shown); E, R. nyttus, ventral anchor; F, R. arietinus Kritsky, Boeger & Thatcher, 1988, MCO. Arrowindicates the circular tandem sclerotized brims. Scale bars: A-D, F, 20 μm; E, 50 μm.

Domingues M. V. & Boeger W. A.
446 ZOOSYSTEMA • 2005 • 27 (3)
A B
CD
E
G H
F
F-H
B-E
FIG. 2. — Rhinoxenus anaclaudiae n. sp.; A, holotype (ventral); B, male copulatory organ (MCO); C, hook from pair 2; D, hook frompairs 1, 3-7; E, vagina; F, ventral bar; G, dorsal anchor; H, ventral anchor. Scale bars: A, 100 μm; B-E, 10 μm; F-H, 30 μm.
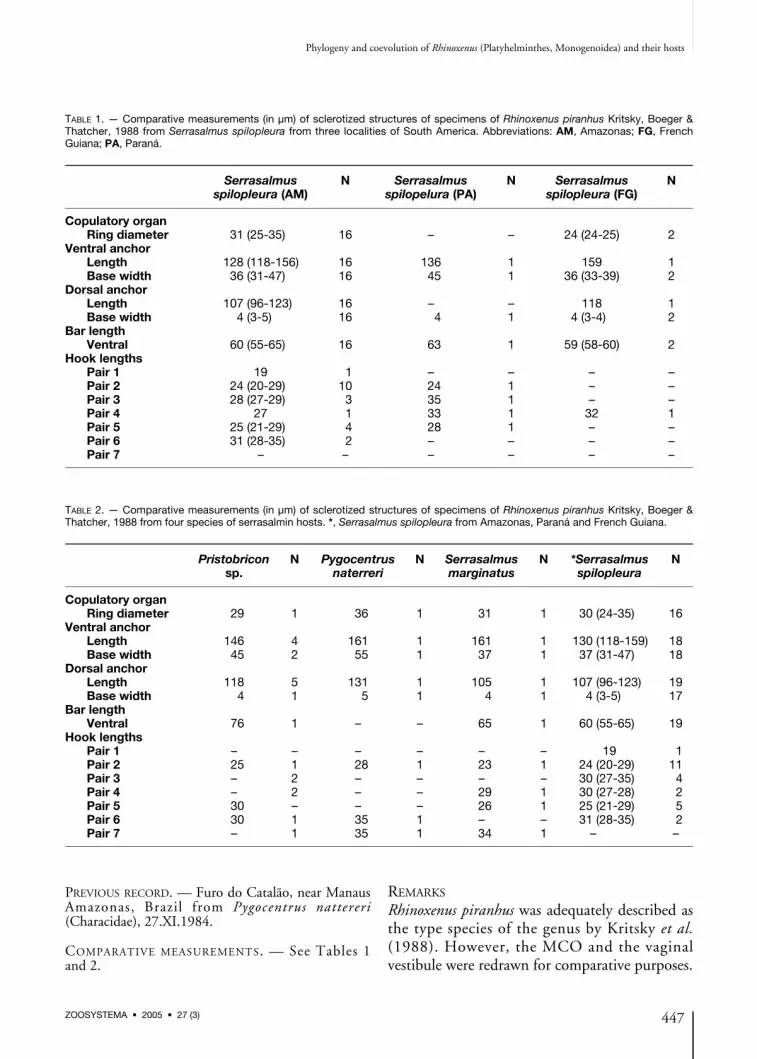
PREVIOUS RECORD. — Furo do Catalão, near ManausAmazonas, Brazil from Pygocentrus nattereri(Characidae), 27.XI.1984.
COMPARATIVE MEASUREMENTS. — See Tables 1and 2.
REMARKS
Rhinoxenus piranhus was adequately described asthe type species of the genus by Kritsky et al.(1988). However, the MCO and the vaginalvestibule were redrawn for comparative purposes.
Phylogeny and coevolution of Rhinoxenus (Platyhelminthes, Monogenoidea) and their hosts
447ZOOSYSTEMA • 2005 • 27 (3)
TABLE 1. — Comparative measurements (in μm) of sclerotized structures of specimens of Rhinoxenus piranhus Kritsky, Boeger &Thatcher, 1988 from Serrasalmus spilopleura from three localities of South America. Abbreviations: AM, Amazonas; FG, FrenchGuiana; PA, Paraná.
Serrasalmus N Serrasalmus N Serrasalmus Nspilopleura (AM) spilopelura (PA) spilopleura (FG)
Copulatory organRing diameter 31 (25-35) 16 – – 24 (24-25) 2
Ventral anchorLength 128 (118-156) 16 136 1 159 1Base width 36 (31-47) 16 45 1 36 (33-39) 2
Dorsal anchorLength 107 (96-123) 16 – – 118 1Base width 4 (3-5) 16 4 1 4 (3-4) 2
Bar lengthVentral 60 (55-65) 16 63 1 59 (58-60) 2
Hook lengthsPair 1 19 1 – – – –Pair 2 24 (20-29) 10 24 1 – –Pair 3 28 (27-29) 3 35 1 – –Pair 4 27 1 33 1 32 1Pair 5 25 (21-29) 4 28 1 – –Pair 6 31 (28-35) 2 – – – –Pair 7 – – – – – –
TABLE 2. — Comparative measurements (in μm) of sclerotized structures of specimens of Rhinoxenus piranhus Kritsky, Boeger &Thatcher, 1988 from four species of serrasalmin hosts. *, Serrasalmus spilopleura from Amazonas, Paraná and French Guiana.
Pristobricon N Pygocentrus N Serrasalmus N *Serrasalmus Nsp. naterreri marginatus spilopleura
Copulatory organRing diameter 29 1 36 1 31 1 30 (24-35) 16
Ventral anchorLength 146 4 161 1 161 1 130 (118-159) 18Base width 45 2 55 1 37 1 37 (31-47) 18
Dorsal anchorLength 118 5 131 1 105 1 107 (96-123) 19Base width 4 1 5 1 4 1 4 (3-5) 17
Bar lengthVentral 76 1 – – 65 1 60 (55-65) 19
Hook lengthsPair 1 – – – – – – 19 1Pair 2 25 1 28 1 23 1 24 (20-29) 11Pair 3 – 2 – – – – 30 (27-35) 4Pair 4 – 2 – – 29 1 30 (27-28) 2Pair 5 30 – – – 26 1 25 (21-29) 5Pair 6 30 1 35 1 – – 31 (28-35) 2Pair 7 – 1 35 1 34 1 – –

Rhinoxenus arietinusKritsky, Boeger & Thatcher, 1988
(Fig. 1F)
Rhinoxenus arietinus Kritsky, Boeger & Thatcher,1988: 88-92, figs 9-17.
TYPE HOST AND LOCALITY . — Nasal cavity ofSchizodon fasciatus (Anostomidae), Ilha Marchantaria,Rio Solimões, near Manaus, Amazonas, Brazil,25.XI.1983 (Kritsky et al. 1988).MATERIAL EXAMINED. — Brazil. Rio Capucapu,Cachoeira das Garças, Manaus, Amazonas, fromLeporinus agassizii (Anostomidae), 30.X.1989, voucher
Domingues M. V. & Boeger W. A.
448 ZOOSYSTEMA • 2005 • 27 (3)
A B
C
G H L
D E
F
M
IJ
K
C, D, F A, B
E, I, J
K-M
G, H
FIG. 3. — Sclerotized structures in Rhinoxenus; A-F, R. curimbatae n. sp.; A, male copulatory organ (MCO); B, vagina; C, ventralanchor; D, dorsal anchor; E, hook; F, ventral bar; G-M, R. guianensis n. sp.; G, male copulatory organ (MCO); H, vagina; I, hookfrom pair 2; J, hook from pairs 1, 3-7; K, ventral bar; L, ventral anchor; M, dorsal anchor. Scale bars: A, B, 20 μm; C, D, F, 30 μm;E, I, J, 15 μm; G, H, 10 μm; K-M, 50 μm.

specimens (CHIOC 36282 a-b). — Rio Paraná,Porto Rico, Paraná, from Leporinus elongatus(Anostomidae), 1992, voucher specimens (CHIOC36283 a-b; INPA 429; MNHN HG-T1 205-205 bis);same locality, from Leporinus obtusidens (Ano-stomidae), 1992, voucher specimens (INPA 428 a-c;MNHN HG-T1 204-240 bis; USNPC 95231). —Rio Paraná, Ressaco do pau véio, Porto Rico, Paraná,from Leporinus elongatus (Anostomidae), 1992,voucher specimens (INPA 430 a-c); from Leporinuselongatus (Anostomidae), 17.VI.1996, voucher speci-mens (USNPC 95232). — Rio Tibagi, nearJataizinho, Paraná, from Leporinus elongatus(Anostomidae), 28-29.V.1997, voucher specimens(CHIOC 35284 a-c; INPA 431 a-c; USNPC 95233);same locality, from Leporinus obtusidens , 28-29.V.1997, voucher specimens (CHIOC 36307 a-d;MNHN 164 HG-T1 206-206 bis). — Reservatório deItaipu, Foz do Iguassu, Paraná, from Leporinus fridericifriderici (Anostomidae), 15.XI.1996, voucher speci-mens (CHIOC 36286 a-b; INPA 432; USNPC95234); same locality, from Schizodon sp. (Ano-stomidae), 15.XI.1996, voucher specimens (CHIOC36291 a-c; INPA 426 a-b; MNHN 161 HG-T1 203-203 bis; USNPC 95229). — Rio Paraná, near PortoRico, Paraná, from Leporinus friderici friderici(Anostomidae), 1993, voucher specimens (CHIOC36285 a-b; MNHN 165 HG-T1 207); same locality,from Leporinus lacustris (Anostomidae), 1993, vouch-er specimens (CHIOC 36287 a-b; INPA 427;USNPC 95230); same locality, from Schizodonaltoparanae (Anostomidae), 1993, voucher specimens
(CHIOC 36288 a-c; INPA 424 a-b; MNHN 158HG-T1 201-201 bis; USNPC 95227); same locality,from Schizodon borelli (Anostomidae), 1992, voucherspecimens (CHIOC 36289 a-b; INPA 422; USNPC95224-95226); same locality, from Schizodon borelli(Anostomidae), 1993, voucher specimens (CHIOC36290 a-c; INPA 423 a-c; MNHN 156 HG-T1 198bis-199-199 bis, 157 HG-T1 200-200 bis); samelocality, from S. knerii (Steindachner, 1875) (Ano-stomidae), 1993, voucher specimens (USNPC95228).
PREVIOUS RECORD. — Aquarium INPA, Manaus,Amazonas, Brazil from Rhytiodus argenteofuscus(Anostomidae), 8.II.1984 (Kritsky et al. 1988).
COMPARATIVE MEASUREMENTS. — See Tables 3 and 4.
REMARKS
Rhinoxenus arietinus is known from nine speciesof Leporinus, Schizodon, and Rhytiodus. Leporinuselongatus and L. obtusidens are reported as hosts intwo different localities (both in the Rio Paranáand Rio Tibagi, PR, Brazil). Kritsky et al. (1988)considered the occurrence of Rhinoxenus arietinusin Rhytiodus argenteofuscus as accidental. However,this parasite occurs in other anostomids, andthus, it seems to represent a parasite specific tothis family.
Phylogeny and coevolution of Rhinoxenus (Platyhelminthes, Monogenoidea) and their hosts
449ZOOSYSTEMA • 2005 • 27 (3)
TABLE 3. — Comparative measurements (in μm) of sclerotized structures of specimens of Rhinoxenus arietinus Kritsky, Boeger &Thatcher, 1988 from Leporinus elongatus and L. obtusidens in two localities at Paraná State, Brazil. Abbreviations: PA, Rio Paraná;TI, Rio Tibagi.
Leporinus N Leporinus N Leporinus N Leporinus Nelongatus (PA) obtusidens (PA) elongatus (TI) obtusidens (TI)
Copulatory organRing diameter 36 (32-40) 5 32 (28-39) 12 30 (27-35) 9 30 1
Ventral anchorLength 138 (138-149) 4 140 (129-164) 12 127 (108-136) 9 102 1Base width 73 (68-78) 2 70 (63-75) 7 69 (65-73) 5 57 1
Dorsal anchorLength 154 (146-171) 5 157 (139-174) 12 147 (126-164) 8 23 1Base width 17 1 17 2 – – 14 1
Bar lengthVentral 66 1 54 (50-60) 7 55 (50-60) – 140 1
Hook lengthsPair 1 – – – – – – 33 1Pair 2 36 1 40 (35-47) 7 38 (33-42) 1 36 1Pair 3 – 2 48-49 2 – – 45 1Pair 4 42 2 42 (38-45) 2 – – 44 1Pair 5 – – – – – – 44 1Pair 6 46 1 47 1 – – 43 1Pair 7 48 1 48 1 – – 43 1

The specimens analyzed herein do not differ sig-nificantly from those described by Kritsky et al.(1988). However, the proximal projection ofthe accessory piece, that surrounds the first ringof the MCO (Kritsky et al. 1988: figs 11, 12),was not observed herein, and the two tandembrims associated to the base of the MCO(Fig. 1F) were not described by Kritsky et al.
(1988). The measurements of the sclerotizedstructures of the present material do not differfrom those presented in the original description.Rhinoxenus arietinus differs from the otherspecies of the genus by possessing two posteriormuscular lobes on the haptor and a ventralanchor with a conspicuous superficial and deeproot.
Domingues M. V. & Boeger W. A.
450 ZOOSYSTEMA • 2005 • 27 (3)
TABLE 4. — Comparative measurements (in μm) of sclerotized structures of specimens of Rhinoxenus arietinus Kritsky, Boeger &Thatcher, 1988 of eight anostomid hosts. *, Leporinus elongatus from Rio Paraná and Rio Tibagi; **, L. obtusidens from Rio Paranáand Rio Tibagi.
Leporinus N *Leporinus N Leporinus N Leporinus Nagassizii elongatus friderici lacustris
Copulatory organRing diameter 27 (21-33) 2 32 (27-40) 14 32 (30-35) 3 29 (27-30) 3
Ventral anchorLength 127 (99-156) 2 131 (108-149) 13 133 (118-14) 4 111 (106-113) 4Base width 63 (53-73) 2 70 (65-78) 7 58 (52-67) 3 49 (40-54) 4
Dorsal anchorLength 130 (106-154) 2 150 (126-171) 13 146 (116-159) 4 115 (120-131) 3Base width 12 (10-15) 1 17 1 15 (12-19) 4 14 (12-16) 4
Bar lengthVentral 63 1 60 (50-66) 7 58 5 50 1
Hook lengthsPair 1 – – – – 36 1 – –Pair 2 29 1 38 (33-42) 8 41 (36-45) 2 35 (34-38) 3Pair 3 – – – – 40 1 – –Pair 4 – – 42 1 48 (47-49) 2 39 (38-40) 2Pair 5 – – – – – – – –Pair 6 – – 46 1 – – – –Pair 7 – – 45 1 – – – –
**Leporinus N Schizodon N Schizodon N Schizodon Nobtusidens altoparanae borelli sp.
Copulatory organRing diameter 32 (28-39) 13 31 (30-33) 9 31 (27-35) 11 26-27 2
Ventral anchorLength 137 (102-164) 13 140 (134-154) 12 129 (121-144) 12 141 (134-147) 3Base width 68 (57-75) 8 66 (63-71) 7 72 (66-80) 12 65 (55-75) 2
Dorsal anchorLength 154 (123-174) 13 172 (164-186) 12 130 (113-141) 14 143 (134-159) 3Base width 16 (14-16) 3 20 (19-24) 6 19 (15-21) 4 – –
Bar lengthVentral 51 (40-60) 8 62 (58-65) 8 54 (50-58) 2 63 1
Hook lengthsPair 1 33 1 – – – – – –Pair 2 40 (35-47) 8 37 – 40 (37-44) 5 – –Pair 3 47 (45-49) 3 – – 50 1 – –Pair 4 42 (38-45) 3 – – 47 2 – –Pair 5 44 1 – – 43 1 – –Pair 6 47 (43-47) 2 – – – – – –Pair 7 48 (43-48) 2 – – 55 1 47 –

Rhinoxenus bulbovaginatusBoeger, Domingues & Pavanelli, 1995
Rhinoxenus bulbovaginatus Boeger, Domingues &Pavanelli, 1995: 696-698, figs 1-8.
HOST AND LOCALITY. — Nasal cavities of Salminusbrasiliensis (Characidae), Rio Paraná, near Porto Rico,Paraná, Brazil.
MATERIAL EXAMINED. — Brazil. Rio Miranda, Passodo Lontra, Mato Grosso do Sul, from Salminusbrasiliensis (Characidae), 5.VIII.1998, voucher speci-mens (INPA 421 a-c; MNHN 155 HG-T1 197-197bis-198; MZUSP 5934 a-b; USNPC 95223).
MEASUREMENTS. — Copulatory organ ring diameter27 (21-37; n = 5); ventral anchor 121 (117-122; n = 5)long, 32 (30-32; n = 5) wide; dorsal anchor 129 (125-132; n = 5) long; 5 (n = 5) wide; ventral bar 64 (60-67;n = 5) long; 12 (n = 5) wide; hook pair 1 37 (36-38;n = 2); hook pair 2 33 (30-35; n = 4); hook pair 3 48;hook pair 4 52; hook pair 5 45; hook pair 6 51; hookpair 7 49 (47-52; n = 3).
REMARKS
Rhinoxenus bulbovaginatus is characterized bypossessing the point of the ventral anchor in theshape of a “fish-hook”; posterolateral expansionsin the ventral bar for articulation to the ventralanchor; and vagina sclerotized with cup-shapevestibule.
Rhinoxenus nyttusKritsky, Boeger & Thatcher, 1988
(Fig. 1C-E)
Rhinoxenus nyttus Kritsky, Boeger & Thatcher, 1988:92, 93, figs 18-24.
TYPE HOST AND LOCALITY. — Nasal cavities ofSchizodon fasciatus (Anostomidae), Ilha Marchantaria,Rio Solimões, near Manaus, Amazonas, Brazil,25.XI.1983.
MATERIAL EXAMINED. — Brazil. Usina Hidrelétrica deITAIPU, Foz do Iguassu, Paraná, from Schizodon sp.(Anostomidae), 15.X.1996, voucher specimens(CHIOC 36292 a-c; INPA 433; MNHN 167 HG-T1208-208 bis; USNPC 95235). — Rio Capucapu,Cachoeira das Garças, near Manaus, Amazonas, fromLeporinus agassizii (Anostomidae), 30.X.1989, voucherspecimens (INPA 434; USNPC 95236).
COMPARATIVE MEASUREMENTS. — See Table 5.
REMARKS
Rhinoxenus nyttus is apparently restricted tomembers of Anostomidae. The specimens ana-lyzed here are similar to those described byKritsky et al. (1988), except by the possession of amore extensive sclerotized cap on the root ventralanchor (Fig. 1E). The original description does notmention the presence of the brims of the MCO,observed in all studied specimens (Fig. 1C, D).
Phylogeny and coevolution of Rhinoxenus (Platyhelminthes, Monogenoidea) and their hosts
451ZOOSYSTEMA • 2005 • 27 (3)
TABLE 5. — Comparative measurements (in μm) of sclerotized structures of specimens of Rhinoxenus nyttus Kritsky, Boeger &Thatcher, 1988 from Leporinus agassizii and Schizodon sp.
Leporinus agassizii N Schizodon sp. N
Copulatory organRing diameter 31 1 27 (23-30) 3
Ventral anchorLength 90 1 73 (69-85) 6Base width 45 1 36 (31-43) 6
Dorsal anchorLength 63 1 55 (51-59) 5Base width 3 1 3 (2-3) 6
Bar lengthVentral 62 1 45 (42-50) 7
Hook lengthsPair 1 15 1 18 (15-21) 4Pair 2 20 1 18 (17-22) 6Pair 3 25 1 23 (21-25) 5Pair 4 27 1 23 (21-25) 4Pair 5 19 1 18 (17-20) 18Pair 6 21 1 22 (19-24) 3Pair 7 26 1 21 (17-23) 3

Parasites of each host species present little varia-tion in size in almost all the analyzed structures.Rhinoxenus nyttus is characterized by possessing aventral bar with lateral projections that serve asarticulation to the ventral anchor; long chalice-shape vaginal vestibule; and ventral anchor withwavy point, representing 1/3 of the total lengthof the anchor.
Rhinoxenus anaclaudiae n. sp.(Fig. 2)
TYPE MATERIAL. — Holotype (CHIOC 36300);23 paratypes (CHIOC 36301 a-e; INPA 439 a-g;MNHN 171 HG-T1 212-212 bis-213; MZUSP 5933a-h; USNPC 95239).
ETYMOLOGY. — The species is named after Dr AnaClaudia dos Santos Brasil, a specialist of Polychaeta.
TYPE HOST AND LOCALITY. — Nasal cavities ofTriportheus cf. nematurus (Characidae), Rio Miranda,Passo do Lontra, Mato Grosso do Sul, Brazil,30.XI.1996.
OTHER MATERIAL EXAMINED. — Brazil. Rio Miranda,Passo do Lontra, Mato Grosso do Sul, from Brycon sp.(Characidae), 5.VIII.1998, voucher specimens(CHIOC 36302). — Baía da Medalha, Rio Miranda,Mato Grosso do Sul, from Triportheus sp. (Cha-racidae), 4.VIII.1998, voucher specimens (INPA 440
a-d; MNHN 172 HG-T1 214-214 bis-215; USNPC95240).
COMPARATIVE MEASUREMENTS. — See Table 6.
DESCRIPTION
Body pyriform, 254 (n = 1) long; greatest width98 (n = 1) at body midlength. Cephalic lobespoorly developed; three pairs of cephalic organs;cephalic glands posterolateral to pharynx.Members of posterior pair of eyes larger, closerthan those of anterior pair; eye granules elongate.Pharynx 19 (18-20; n = 5) in diameter. Haptorsubtriangular, 71 (n = 1) long, 57 (n = 1) wide.Ventral anchor with inconspicuous roots, sclero-tized cap of base with projection for articulationto ventral bar, shaft evenly curved, point short,strongly recurved. Dorsal anchor with bluntproximal end, pointed distal end, terminationswith conspicuous sclerotized caps. Ventral barflattened, with slightly thickened ends. Hookpair 2 with erect thumb, lightly curved shaft,short point, proximal 3/4 of shank inflated; fila-mentous hook (FH) loop extended to near begin-ning of shank dilation; remaining hooks witherect thumb, long shaft, lightly curved, shankinflated proximally; FH loop extended to nearbeginning of shank dilation. Male copulatory
Domingues M. V. & Boeger W. A.
452 ZOOSYSTEMA • 2005 • 27 (3)
TABLE 6. — Comparative measurements (in μm) of sclerotized structures of specimens of Rhinoxenus anaclaudiae n. sp. fromTriportheus cf. nematurus, Triportheus sp., and Brycon sp.
Tripotheus cf. nematurus N Tripotheus sp. N Brycon sp. N
Copulatory organRing diameter 22 (18-26) 23 18 (14-20) 9 13 1
Ventral anchorLength 74 (67-85) 22 73 (67-84) 9 87 1Base width 20 (17-23) 20 22 (18-26) 9 21 1
Dorsal anchorLength 65 (56-72) 17 67 (62-73) 8 68 1Base width 2 (2-3) 23 2 (2-3) 9 3 1
Bar lengthVentral 33 (30-36) 16 31 (30-33) 8 30 1
Hook lengthsPair 1 – – – – – –Pair 2 21 (19-24) 5 21 (20-22) 3 – –Pair 3 14 3 – – – –Pair 4 – – – – – –Pair 5 – – – – – –Pair 6 – – – – – –Pair 7 – – – – – –

organ a coiled tube with approximately two rings.Testis oval, 54 (43-64; n = 2) long, 24 (n = 1)wide; seminal vesicle fusiform. Germarium 49(42-54; n = 4) long, 29 (26-34; n = 4) wide.Ootype, uterus not observed. Vagina sclerotized,proximally wide, tapering distally, distal loop;vaginal vestibule sclerotized, cup shaped. Seminalreceptacle pyriform. Large dorsal prostate.Vitellaria coextensive with cecae; vitelline com-missure anterior to germarium. Egg not observed.
REMARKS
Rhinoxenus anaclaudiae n. sp. differs from itscongeners in possessing a ventral anchor withevenly curved and short shaft and stronglyrecurved point. Rhinoxenus anaclaudiae n. sp. isapparently restricted to members of Characoidea(sensu Buckup 1998).
Rhinoxenus curimbatae n. sp.(Fig. 3A-F)
TYPE MATERIAL. — Holotype (CHIOC 36303);paratypes (CHIOC 36304 a-b; INPA 419; MNHN153 HG-T1 195).
ETYMOLOGY. — The specific epithet refers to the localname of the host, “curimbatá”.
TYPE HOST AND LOCALITY. — Nasal cavities ofProchilodus cf. lineatus (Prochilodonidae), RepresaCapivari-Cachoeira, Municipality of Campina Grandedo Sul, metropolitan area of Curitiba, Paraná, Brazil,15.III.1995.
DESCRIPTION
Body 700 (n = 1) long; greatest width 130 (n = 1)at body midlength. Eyes usually present, equidis-tant; eye granules elongate. Pharynx spherical,30 (n = 1) in diameter. Haptor subtrapezoidal,82 (n = 1) long, 75 (n = 1) wide. Ventral anchor50 (42-56; n = 5) long, base 42 (30-53; n = 4)wide, superficial, roots inconspicuous, base withsclerotized cap articulated with ventral bar, knob-like projection on base, short recurved shaft,elongate straight point. Dorsal anchor 50 (31-59;n = 4) long, base 4 (3-5; n = 4) wide, robust, withblunt proximal end, straight shaft, distal enddiagonally truncated. Ventral bar 42 (30-53; n = 1)
long with expanded ends. Hooks 14 (11-17;n = 15) long, similar in shape, truncate thumb,evenly curved shaft, point, shank without infla-tion, FH loop extending to half of shank. Malecopulatory organ a coiled tube with approximate-ly 3.5 rings; greatest ring 27 (24-29; n = 4) indiameter; copulatory ligament twisted. Gonads,ootype, uterus, seminal receptacle not observed.Vagina sclerotized; distal portion tubular, sinu-ous; proximal end wide; vaginal vestibule heavilysclerotized, cup-shaped. Egg not observed.
REMARKS
Rhinoxenus curimbatae n. sp. resembles R. nyttusbased on the morphology of the MCO.However, it differs from this and other species ofthe genus by possessing dorsal anchors with distalend diagonally truncated, ventral anchors withshort recurved shafts and elongate straight points,heavily sclerotized cup-shaped vaginal vestibule,and hooks with shanks not inflated.
Rhinoxenus guianensis n. sp.(Fig. 3G-M)
TYPE MATERIAL. — Holotype (CHIOC 36305);paratypes (CHIOC 36306 a-d; INPA 420; MNHN154 HG-T1 196; USNPC 95222).
ETYMOLOGY. — The specific epithet refers to the typelocality from which the species was collected.
TYPE HOST AND LOCALITY. — Nasal cavities ofCurimata cyprinoides (Curimatidae), Iracoubo, DegradForian, French Guiana, 8-10.X.1996.
DESCRIPTION
Eyes usually present, equidistant, members ofposterior pair closer than those of anterior pair;eyes granules elongate. Ventral anchor delicate,115 (106-125; n = 5) long, base 19 (15-22; n = 3)wide, roots inconspicuous; base with sclerotizedcap articulated to ventral bar; straight shaft, pointshort, truncated, golf-club shaped. Dorsal anchordelicate, slender, 98 (95-100; n = 5) long; withblunt proximal, distal ends, slightly sinuous shaft;each end with conspicuous sclerotized cap.Ventral bar 35 (34-38; n = 5) long. Hook pair 221 (20-23; n = 4) long, erect thumb, shaft, point
Phylogeny and coevolution of Rhinoxenus (Platyhelminthes, Monogenoidea) and their hosts
453ZOOSYSTEMA • 2005 • 27 (3)

evenly curved, proximal 2/3 of shank inflation;FH loop extending to near beginning of shankdilation. Hook pairs 1, 3-7 22 (20-24; n = 3)long, elongate, erect thumb, lightly curved shaft,short point, proximal 2/3 of shank inflated; FHloop extending to near beginning of shank dila-tion. Male copulatory organ a coiled tube withapproximately 1.5 rings, 15 (14-17; n = 4) indiameter. Gonads, ootype, uterus, seminal recep-tacle not observed. Vagina sclerotized; with asubterminal loop, distally expanded; vestibulesclerotized, cup-shaped. Eggs not observed.
REMARKS
All specimens of Rhinoxenus guianensis n. sp. weremounted in Hoyer’s medium. Measurements anddescription of internal organs, therefore, are lim-ited. The new species differs from all other con-generic species by possessing ventral anchors withstraight shafts, short, truncated, club-shapedpoints and dorsal anchors with distal portionstruncated and curved.
Rhinoxenus euryxenus n. sp.(Fig. 4)
TYPE MATERIAL. — Holotype (CHIOC 36293);7 paratypes (CHIOC 36294 a-g; INPA 435 a-c;MNHN 168 HG-T1 209-209 bis).
ETYMOLOGY. — The specific ephitet is from Greek(eury = broad + xen/o = guest) and refers to the wideoccurrence of this parasite within Serrasalminae.
TYPE HOST AND LOCALITY. — Nasal cavities ofSerrasalmus marginatus (Characidae), Baía da Medalha,Rio Paraná, Matogrosso do Sul, Brazil, 18.VI.1996.
OTHER MATERIAL EXAMINED. — Brazil. Rio Uatumã,lago Tapana, Santa Anna, Amazonas, from Serrasalmusgouldingi (Characidae), 2-3.XI.1989, voucher speci-mens (CHIOC 36297). — Rio Miranda, Passo doLontra, Mato Grosso do Sul, from Serrasalmus mar-ginatus (Characidae), 31.XI.1996, voucher specimens(CHIOC 36299). — Baía da Medalha, Rio Paraná,Matogrosso do Sul, from Serrasalmus marginatus(Characidae), 18-19.VI.1996, voucher specimens(INPA 436 a-b; MNHN 169 HG-T1 210-210 bis;USNPC 95237). — Rio Iatapu, lago Maracanã,Manaus, Amazonas, from Serrasalmus rhombeus(Characidae), 2.XI.1989, voucher specimens (CHIOC36296; INPA 437 a-b; MNHN 170 HG-T1 211;USNPC 95238). — Rio Paraná, near Porto Rico,
Paraná, from Serrasalmus spilopleura (Characidae),1992, voucher specimens (CHIOC 36298). — RioCapucapu, near the confluence of the Rio Jatapu,Cachoeira das Garças, Amazonas, from Serrasalmusstriolatus (Characidae), 31.XI.1989, voucher speci-mens (CHIOC 36295); same locality, from Leporinusagassizii (Anostomidae), 31.XI.1989, voucher speci-mens (INPA 438 a-b).
COMPARATIVE MEASUREMENTS. — See Tables 7 and 8.
DESCRIPTION
Body fusiform, 607 (480-720; n = 10) long;greatest width 176 (150-210; n = 9) at bodymidlength. Cephalic lobes poorly developed;three pairs of cephalic organs; cephalic glandsposterolateral to pharynx. Eyes equidistant;members of anterior pair smaller than those ofposterior pair; eye granules elongate. Pharynxspherical, 42 (35-54; n = 11) in diameter. Haptorsubtrapezoidal, 96 (64-115; n = 11) long, 115(86-140; n = 11) wide. Ventral anchor withinconspicuous roots; base with sclerotized caparticulated with ventral bar; shaft recurved nearmidlength; point with saucer-like termination.Dorsal anchor with blunt proximal end coveredby conspicuous sclerotized cap, straight shaft,tapered distal end. Ventral bar with expandedends, irregularly sclerotized. Hook pair 2 robust,lying in small lobes on posterior trunk, withthumb erect, robust, shaft short, evenly shortcurved point; proximal 3/4 of shank inflated; FHloop extending to near beginning of shank dila-tion. Hooks pairs 1, 3-7 with erect thumb, deli-cate evenly curved shaft, short point; proximal4/5 of shank long inflated; FH loop extending tonear beginning of shank dilatation. Male copula-tory organ a coiled tube with approximately 1.5to 2 rings; copulatory ligament twisted. Testiselongate, 120 (n = 1) long, 25 (n = 1) wide; semi-nal vesicle fusiform; prostate in two dorsolateralfields. Germarium elongate, 63 (56-70; n = 2)long, 32 (29-37; n = 2) wide. Ootype, uterus notobserved. Vagina sclerotized, distal portion withloop; vestibule sclerotized, cup-shaped, ofteneverted (Fig. 4B). Seminal receptacle elongate.Vitellaria coextensive with cecae; vitellinecommissure anterior to germarium. Egg notobserved.
Domingues M. V. & Boeger W. A.
454 ZOOSYSTEMA • 2005 • 27 (3)

Phylogeny and coevolution of Rhinoxenus (Platyhelminthes, Monogenoidea) and their hosts
455ZOOSYSTEMA • 2005 • 27 (3)
A
F
B
C
D
E
G H
B-E
F-H
FIG. 4. — Rhinoxenus euryxenus n. sp.; A, holotype (ventral); B, vagina; C, male copulatory organ (MCO); D, hook from pairs 1, 3-7;E, hook from pair 2; F, ventral bar; G, ventral anchor; H, dorsal anchor. Scale bars: A, 100 μm; B-E, 10 μm; F-H, 50 μm.

REMARKS
Rhinoxenus euryxenus n. sp. closely resemblesR. piranhus based on the morphology of MCOand point of the ventral anchor. The new species,however, differs from R. piranhus by possessingventral anchors with shafts strongly recurved nearmidlength (relatively straight shaft in R. piranhus)
and lacking the small terminal protuberances onthe ventral bar as described by Kritsky et al.(1988: fig. 4) for R. piranhus. Rhinoxenus eury-xenus n. sp. appears ubiquitous to fishes belongingto Serrasalmus. The occurrence of R. euryxenusn. sp. on Leporinus agassizii (Anostomidae) isconsidered accidental.
Domingues M. V. & Boeger W. A.
456 ZOOSYSTEMA • 2005 • 27 (3)
TABLE 7. — Comparative measurements (in μm) of sclerotized structures of specimens of Rhinoxenus euryxenus n. sp. fromSerrasalmus marginatus from two localities of Brazil. *, Rio Paraná, Paraná; **, Rio Miranda, Matogrosso do Sul.
*Serrasalmus marginatus N **Serrasalmus marginatus N
Copulatory organRing diameter 33 (24-37) 23 29 1
Ventral anchorLength 152 (137-175) 24 144 1Base width 55 (44-63) 22 49 1
Dorsal anchorLength 129 (123-151) 16 123 1Base width 5 (4-6) 21 4 1
Bar lengthVentral 82 (71-82) 15 77 1
Hook lengthsPair 1 25 (16-34) 2 – –Pair 2 29 (27-30) 8 26 1Pair 3 40 (34-49) 6 – –Pair 4 37 (33-41) 6 – –Pair 5 33 (30-35) 3 – –Pair 6 34 (31-38) 3 – –Pair 7 36 (32-38) 3 – –
TABLE 8. — Comparative measurements (in μm) of sclerotized structures of specimens of Rhinoxenus euryxenus n. sp. from fiveserrasalmids and one anostomid hosts. *, Serrasalmus marginatus from Rio Paraná and Rio Miranda.
Serrasalmus N *Serrasalmus N Serrasalmus N Serrasalmus N Serrasalmus N Leporinus Ngouldini marginatus rhombeus striolatus spilopleura agassizii
Copulatory organRing diameter – – 33 (24-37) 24 37 (31-41) 3 37 1 34 1 32 (31-32) 2
Ventral anchorLength 142 1 153 (140-177) 25 150 (137-162) 3 124 1 150 1 147 (142-152) 2Base width 49 1 55 (49-63) 22 49 (48-50) 3 33 1 62 1 42 (39-46) 2
Dorsal anchorLength 123 1 128 (123-151) 17 129 (121-133) 3 119 1 133 1 133 (129-137) 2Base width 4 1 5 (4-6) 21 4 (4-5) 3 3 1 4 1 3 (3-4) 2
Bar lengthVentral 77 1 83 (76-90) 22 72 (61-81) 4 57 1 92 1 55 1
Hook lengthsPair 1 – – 25 (16-34) 2 21 1 – – – – 28 1Pair 2 29 1 29 (26-30) 9 27 2 27 1 39 1 26 1Pair 3 – – 40 (34-49) 6 – – – – – – 29 1Pair 4 – – 37 (33-41) 6 28 1 – – – – 28 1Pair 5 – – 33 (30-35) 3 32 (31-33) 2 – – 34 1 – –Pair 6 – – 34 (31-38) 4 – – – – – – – –Pair 7 36 1 36 (32-38) 3 30 2 – – – – 34 1

CHARACTER ANALYSIS
Numbers in parenthesis preceding the definitionof a character refers to coding in the matrix: (0)represents the plesiomorphic state and (?) denotesmissing or inapplicable data; numbers betweensquare brackets refer to respective evolutionarychanges depicted in cladogram (Fig. 5). Thematrix of characters used for this analysis is pre-sented in the Table 9. Transformation series 7and 9 were considered ordered in the analysis.A. Dorsal bar. (0) present; (1) absent [1].B. Shape of dorsal anchor. (0) shaft and pointclearly differentiated; (1) shaft and point not dif-ferentiated (Figs 2G; 3D, M; 4H) [2].C. Circular tandem sclerotized brims of Malecopulatory organ (MCO). (0) absent (Domin-gues & Boeger 2002: fig. 2); (1) present [3].
D. Shape of ventral anchor. (0) anchor elongate,shaft longer than point (Figs 2H; 3L; 4G) [4]; (1)anchor short, shaft shorter than point (Figs 1E;3C) [11, 18].E. Ventral bar. (0) bar with extremities lightlyexpanded (Figs 2F; 3F; 4F) [5]; (1) bar withproject ions for art iculat ion with ventra lanchor (Kritsky et al. 1988: figs 4, 23) [12,16, 21, 23].F. MCO. (0) MCO with more than three ringsfrom the base (Figs 1C; 3A) [6]; (1) MCO withless than three rings (Figs 1A, F; 2B; 3G; 4C)[14].G. Vaginal vestibule. (0) absent (Kritsky et al.1988: fig. 9) [19]; (1) vestibule chalice shaped,longer than wide (Kritsky et. al. 1988: fig. 18)[7]; (2) vestibule cup shaped, vestibule widerthan long (Figs 1B; 2E; 3B, H; 4B) [13, 20].
Phylogeny and coevolution of Rhinoxenus (Platyhelminthes, Monogenoidea) and their hosts
457ZOOSYSTEMA • 2005 • 27 (3)
TABLE 9. — Matrix used in reconstruction of evolutionary relationship of Rhinoxenus species.
Taxon Transformation seriesA B C D E F G H I J
Outgroup 0 0 0 0 0 0 0 0 0 0R. nyttus 1 1 1 1 1 1 0 0 0 0R. curimbatae n. sp. 1 1 1 1 0 0 2 0 0 ?R. bulbovaginatus 1 1 ? 0 1 1 1 0 0 1R. arietinus 1 1 1 1 0 1 0 1 1 1R. guianensis n. sp. 1 1 1 0 1 1 2 1 1 ?R. anaclaudiae n. sp. 1 1 1 0 0 1 2 1 1 ?R. piranhus 1 1 1 0 1 1 2 1 2 1R. euryxenus n. sp. 1 1 1 0 1 1 2 1 2 1
TABLE 10. — Data matrix listing binary codes indicating phylogenetic relationships among eight species of Rhinoxenus and their hosts.
Hosts Binary code
Acestrorhynchus ? ? ? ? ? ? ? ? ? ? ? ? ? ?Anostomidae 1 1 0 0 0 0 0 0 0 1 0 0 0 0 1Anostomidae 2 0 0 0 1 0 0 0 0 0 0 0 1 1 1Chilodontidae ? ? ? ? ? ? ? ? ? ? ? ? ? ?Crenuchidae ? ? ? ? ? ? ? ? ? ? ? ? ? ?Curimatidae 0 0 0 0 1 0 0 0 0 0 1 1 1 1Erythrinidae ? ? ? ? ? ? ? ? ? ? ? ? ? ?Gasteropelecidae ? ? ? ? ? ? ? ? ? ? ? ? ? ?Hemiodontidae ? ? ? ? ? ? ? ? ? ? ? ? ? ?Parodontidae ? ? ? ? ? ? ? ? ? ? ? ? ? ?Prochilodontidae 0 1 0 0 0 0 0 0 1 0 0 0 0 1Salminus 0 0 1 0 0 0 0 0 0 0 0 0 1 1Serrasalminae 1 0 0 0 0 0 0 1 0 0 1 1 1 1 1Serrasalminae 2 0 0 0 0 0 0 0 1 0 1 1 1 1 1Triportheus 0 0 0 0 0 1 0 0 0 0 1 1 1 1

H. Vagina. (0) sinuous with more than one loop(Fig. 3B) [8]; (1) sinuous with only one loop(Figs 1B; 2E; 3H; 4B) [15].I. Point of ventral anchor. (0) acute (Figs 1E;3C) [9]; (1) blunt (Figs 2H; 3L) [17]; (2) flat-tened (Fig. 4G) [22].J. Hook pair 2. (0) located within haptor; (1)located in two bilateral lobes in trunk (Kritskyet al. 1988: figs 1, 9, 18, 21; Boeger et al. 1995:fig. 1) [10].The phylogenetic hypothesis of Rhinoxenus isbased exclusively on sclerotized structures. Theinternal anatomy of Rhinoxenus species is redun-dant and, therefore, provides no evolutionaryinformation. Similarly, morphology of the hooksdoes not vary for species of the genus and repre-sents non-informative autapomorphies (e.g.,hooks with shank not inflated in Rhinoxenuscurimbatae n. sp.). The transformation series thatdescribes the relative position of the eyes (trans-formation series 1 of Boeger et al. 1995) was notincluded in this study because it was not possibleto define the state for some species.
PHYLOGENY
The hypothesis of the sister-group relationship ofthe studied species, one of four equally parsimo-nious trees produced through the analysis(consistency index = 66%; retention index =66%; length = 18), is presented in Figure 5.Monophyly of the genus is supported by threesynapomorphies: 1) absence of dorsal bar; 2) dor-sal anchor modified in probe form; and 3) pres-ence of circular tandem sclerotized brimsassociated with the base of the MCO.The previous hypothesis (Boeger et al. 1995) sug-gests that R. arietinus is sister group of (R. nyttus(R. bulbovaginatus, R. piranhus)). The presenthypotheses, with the inclusion of four new taxaand new transformation series, disagrees signifi-cantly from that proposed by Boeger et al.(1995). Rhinoxenus nyttus + R. curimbatae n. sp.appear as sister group of the common ancestor ofall other congeneric species. Boeger et al. (1995)suggest that R. bulbovaginatus is sister-group of
R. piranhus. However, in our hypotheses, R. bulbo-vaginatus is sister-group of (R. arietinus (R. ana-claudiae n. sp., R. guianensis n. sp. (R. piranhus,R. euryxenus n. sp.))). The relationship of thesefive taxa is supported by one synapomorphy,with 100% of consistence: 1) point of ventralanchor not acute (blunt or flattened).The sister relationships of R. nyttus, R. curimbataen. sp., R. bulbovaginatus n. sp., and R. arietinuswere identical in the four equally parsimonioustrees. Consensus support existed for these rela-tionships including the clade composed byR. guianensis n. sp. + R. anaclaudiae n. sp. +R. euryxenus n. sp. + R. piranhus. Bootstrap sup-port (≥ 50%) exists for the clade composed byR. arietinus + R. guianensis n. sp. + R. anaclaudiaen. sp. + R. euryxenus n. sp. + R. piranhus, and theclade composed by R. guianensis n. sp. + R. ana-claudiae n. sp. + R. euryxenus n. sp. + R. piranhus.The other three most parsimonious trees differfrom that in Figure 5 primarily by the phyloge-netic position of R. guianensis n. sp., R. piranhus,and R. euryxenus n. sp. The character “presence ofventral bar with articulation for ventral anchor”sometimes appears as a homoplasy for R. guianen-sis n. sp. and R. piranhus, sometimes as a synapo-morphy uniting these two taxa. When thetransformation series is removed from the analy-sis, the hypothesis presented in Figure 5 is thesingle most parsimonious cladogram produced.
COEVOLUTION OF RHINOXENUS SPECIES
The host cladogram based on host features(Fig. 6) has incongruences with that reconstruct-ed from parasitological data (Fig. 7). A summaryof the hypotheses of coevolution of Rhinoxenusspecies and their hosts is presented in the Figure 8.The analysis suggests that cospeciation cannottotally explain host-parasite relationships; eventsof extinction, dispersion and duplication are alsonecessary.Wiley (1988) proposes that absence of parasitesin certain host taxa can be associated with threehypotheses: 1) the ancestor of the group wasnever present in the host and no descendent ofthis ancestor dispersed to the host; 2) extinctionmay have occurred within certain host taxon; or
Domingues M. V. & Boeger W. A.
458 ZOOSYSTEMA • 2005 • 27 (3)

3) sampling errors exist (members of the parasitegroup are present in the host, but their presencewas not detected). In the present work, signifi-cant differences in the sampling among analyzedfish families exists. Thus, although our hypothe-ses depicts extinction (hypothesis 2) to explainthe absence of species of Rhinoxenus in species ofAcestrorhynchus Eigenmann & Kennedy, 1903,Chilodontidae, Crenuchidae, Erythrinidae,Gasteropelecidae, Hemiodontidae, Paro-
dontidae (see Fig. 8), sampling error (hypothesis3) is a likely explanation for some of theseabsences.The analysis suggests dispersion of R. guianensisn. sp. to members of Curimatidae and R. arietinusto those of Anostomidae. Rhinoxenus curimbataen. sp., R. nyttus, R. bulbovaginatus, and the ances-tor of R. euryxenus n. sp. + R. piranhus seeminglycospeciated with species of Prochilodontidae,Anostomidae, Salminus, and Serrasalminae,
Phylogeny and coevolution of Rhinoxenus (Platyhelminthes, Monogenoidea) and their hosts
459ZOOSYSTEMA • 2005 • 27 (3)
Rhinoxenus nyttus
Rhinoxenus bulbovaginatus
Rhinoxenus arietinus
Rhinoxenus guianensis n. sp.
Rhinoxenus anaclaudiae n. sp.
Rhinoxenus euryxenus n. sp.
Rhinoxenus piranhus
Rhinoxenus curimbatae n. sp.
12*
11*
16*
21*
23*
22
57
57
57
17
14-15
1-10
20*
18*-19*
13*
FIG. 5. — Phylogenetic hypotheses of the eight Rhinoxenus species. Slashes with numbers above the branches refer to postulatedevolutionary changes (respective homoplasies are identified by an asterisk). Numbers below branches indicate respective bootstrapsupport (nreps = 1000). Supported branches from strict consensus of all four equally parsimonious trees are indicated by doublelines: 1, absence of dorsal bar; 2, shaft and point of the dorsal anchor not differentiated; 3, circular tandem sclerotized brims associ-ated with the base of the male copulatory organ (MCO); 4, anchor elongate, shaft longer than point; 5, ventral bar with extremitieslightly expanded; 6, MCO with more than three rings beginning from the base; 7, vestibule of vagina in long “chalice” form; 8, vaginasinuous with more than one ring; 9, ventral anchor with point sharped; 10, hook from pair 2 located in two bilateral lobes in the trunk;11*, ventral anchor short, shaft shorter than the point; 12*, ventral bar with projections for articulation with the ventral anchor;13*, vestibule of vagina in “cup” form; 14, MCO with less than three rings; 15, vagina lightly sinuous with only one ring; 16*, ventralbar with projections for articulation with the ventral anchor; 17, point of the ventral anchor truncate; 18*, ventral anchor short, shaftshorter than the point; 19*, vestibule of vagina absent; 20*, vestibule of vagina in “cup” form; 21*, ventral bar with projections forarticulation with the ventral anchor; 22, point the ventral anchor flattened; 23*, ventral bar with projections for articulation with theventral anchor.

respectively. At this level of resolution of the hostcladogram, the origin of R. euryxenus n. sp. andR. piranhus is associated with duplication within
the Serrasalminae. Similarly, R. anaclaudiaecospeciated with the ancestor of Brycon +Salminus +Tripotheus with subsequent failure to
Domingues M. V. & Boeger W. A.
460 ZOOSYSTEMA • 2005 • 27 (3)
Chi
lodo
ntid
ae
Ano
stom
idae
Cur
imat
idae
Pro
chilo
dont
idae
Ery
thrin
idae
Ser
rasa
lmin
ae
Hem
iodo
ntid
ae
Cre
nuch
idae
Par
odon
tidae
Gas
tero
pele
cida
ein
cert
ae s
edis
Ace
stro
rhyn
chus
Trip
orth
eus
Bry
con
Sal
min
us
*
**
FIG. 6. — Cladogram showing phylogenetic relationship of Characiformes based on morphologic data, after Buckup (1991). Thecladogram represents only the families analyzed in the present work. *, Characoidea (sensu Buckup 1998); **, clade sensuMalabarba 1998.

speciate with the host group (shared withTriportheus + Brycon) with subsequent extinctionin Salminus.Coevolutionary analysis by BPA supports previ-ous hypotheses on the phylogenetic relationshipsof Anostomidae, Prochilodontidae and Cha-racidae (sensu Greenwood et al. 1966; Nelson1996). No parasitological evidence is available totest the sister-group relationships of Chilo-
dontidae, Curimatidae, Erythrinidae, Acestro-rhynchus, Gasteropelecidae, Hemiodontidae,Crenuchidae, and Parodontidae since no speciesof Rhinoxenus were collected from fish of thesetaxa.Within Characidae, our analysis suggests thatSerrasalminae, Salminus, Triportheus and Bryconcompose a monophyletic group, supporting thehypotheses of Buckup (1998) and of Malabarba
Phylogeny and coevolution of Rhinoxenus (Platyhelminthes, Monogenoidea) and their hosts
461ZOOSYSTEMA • 2005 • 27 (3)
Pro
chilo
dont
idae
+ R
hino
xenu
s cu
rimba
tae
n.sp
.
Ano
stom
idae
+ R
hino
xenu
s ny
ttus
Ano
stom
idae
+ R
hino
xenu
s ar
ietin
us
Cur
imat
idae
+ R
hino
xenu
s gu
iane
nsis
n.s
p.
Sal
min
us +
Rhi
noxe
nus
bulb
ovag
inat
us
Trip
orth
eus
+ R
hino
xenu
s an
acla
udia
e n.
sp.
Bry
con
+ R
hino
xenu
s an
acla
udia
e n.
sp.
Ser
rasa
lmin
ae +
Rhi
noxe
nus
pira
nhus
Ser
rasa
lmin
ae +
Rhi
noxe
nus
eury
xenu
s n.
sp.
FIG. 7. — Cladogram of hosts groups of Rhinoxenus species constructed from parasitological data and using Characiformes taxaanalyzed in the present work. Dashed lines postulate dispersion. Each host group is labeled with its taxonomic epithet followed bythe Rhinoxenus species parasitizing its members.

(1998). The phylogeny of fish hosts based solelyon parasitological evidence (Fig. 7) suggests thatBrycon and Triportheus are more closely relatedthan to Salminus, since both species harborRhinoxenus anaclaudiae n. sp. Parasites thus sup-port the hypothesis of Castro & Vari (1990),who suggested that Lignobrycon (= Moojenichthys)and Triportheus represent a monophyletic lineage
and this appears more related with a subunit ofBrycon. However, the current hypothesis of rela-tionships of Brycon, Salminus and Tripotheus isthat proposed by Malabarba (1998) who suggeststhat Salminus and Brycon are more closely relatedto each other than to Triportheus. Based in thishypothesis, the occurrence of R. anaclaudiaen. sp. in Brycon and Triportheus may be associat-
Domingues M. V. & Boeger W. A.
462 ZOOSYSTEMA • 2005 • 27 (3)
Gas
tero
pele
cida
e
Ser
rasa
lmin
ae*
Hem
iodo
ntid
ae
Cre
nuch
idae
Par
odon
tidae
Chi
lodo
ntid
ae
Ano
stom
idae
Cur
imat
idae
Pro
chilo
dont
idae
Ery
thrin
idae
Ace
stro
rhyn
chus
Trip
orth
eus
R. a
nacl
audi
aen.
sp.
R. e
uryx
enus
n. s
p.
R. p
iranh
us
R. c
urim
bata
en.
sp.
R. g
uian
ensi
sn.
sp.
R. a
rietin
us
R. n
yttu
s
Bry
con
R. a
nacl
audi
aen.
sp.
Sal
min
usR
. bul
bova
gina
tus
FIG. 8. — Summary of the proposed historical relationships of Rhinoxenus species and their hosts. The parasite cladogram (solidlines) is superimposed on that (broad grey lines) of their hosts taxa. Dotted lines indicate postulated extinction or sampling error ofparasites; dashed-dotted lines indicate postulated dispersion of parasite clades; circles indicate cospeciation events; triangle indi-cates duplication event. *, clade sensu Malabarba 1998.

ed with cospeciation followed by extinction inSalminus.The evolution of Rhinoxenus appears associated tohistorical events of the host clade. The origin ofmany species is linked to cospeciation or dispersionfollowed by speciation of the ancestor of theirrespective host family group. Curiously, mostRhinoxenus clades subsequently failed to cospeciatewithin the descendents of their original hostspecies, the ancestor of the respective fish clade.The only apparent exception observed in this studyis the clade of parasites of Serrasalminae, with twospecies, R. piranhus and R. euryxenus n. sp.As suggested above, some proposed extinctionevents are likely artifacts and denser sampling forparasites of members of all characiform familieswill likely recover species of Rhinoxenus. If theseputative new findings follow the proposed pat-tern of parasite distribution, obviously this willreinforce a much more adequate database foradditional testing of available host phylogeny.
AcknowledgementsThe authors thank the following individuals andagencies for supporting this study: GilbertoPavanelli, Ricardo Takemoto (NUPELIA,Universidade Estadual de Maringá, PR), MichelJegú (IRD, France), Celso Benites (UniversidadeFederal do Matogrosso do Sul, UFMS), DomingoFernandez (ITAIPU Binacional, Brazil), DulceAntoniutti (Universidade Federal do Paraná,UFPR), Almir P. Barreto, Euclides Grando Jrhelped during collection of hosts. Carlos A. S. deLucena (Museu de Ciência e Tecnologia,Pontifícia Universidade Católica, RS) identifiedthe fish hosts; Ralph Lichtenfels and Patricia Pillit(USNPC) allowed access to specimens under theircare; Delane C. Kritsky (Idaho State University,USA) provided a presubmission review of themanuscript, and helped sending material fromAmazonian and Trinidad-Tobago rivers. Thisstudy was partially supported by the ConselhoNacional de Desenvolvimento Científico eTecnológico (CNPq) and Fundação Coordenaçãode Aperfeiçoamento de Pessoal de Nível Superior(CAPES).
REFERENCES
BOEGER W. A. & KRITSKY D. C. 1997. — Co-evolution of Monogenoidea (Platyhelminthes)based on a revised hypotheses of parasite phylogeny.International Journal of Parasitology 27 (12):1495-1511.
BOEGER W. A., DOMINGUES M. V. & PAVANELLI G. C.1995. — Neotropical Monogenoidea. 24. Rhinoxenusbulbovaginatus n. sp. (Dactylogyridae, Ancyo-cephalinae) from the nasal cavity of Salminusmaxillosus (Osteichtyes, Characidae) from the RioParaná, Paraná, Brazil. Memórias do InstitutoOswaldo Cruz 90 (6): 695-698.
BOEGER W. A., DOMINGUES M. V. & KRITSKY D. C.1997. — Neotropical Monogenoidea 32. Cacatuo-cotyle paranaensis n. gen., n. sp. (Dactylogyridae,Ancyrocephalinae) from Characidium spp. from theState of Paraná, Brazil. Systematic Parasitology 36:75-78.
BROOKS D. R. 1981. — Hennig’s parasitologicalmethod: a proposed solution. Systematic Zoology 30:229-249.
BROOKS D. R. 1990. — Parsimony analysis inhistorical biogeography and coevolution: metho-dological and theorical update. Systematic Zoology42: 14-30.
BROOKS D. R. & MCLENNAN D. 1991. — Phylogeny,Ecology, and Behavior: a Research Program inComparative Biology. University of Chicago Press,Chicago, 434 p.
BROOKS D. R. & MCLENNAN D. 1993. — Parascript:Parasites and the Language of Evolution. Smith-sonian Institution Press, Washington; London,429 p.
BROOKS D. R., VAN VELLER M. G. P. & MCLENNAND. A. 2001. — How to do BPA, really. Journal ofBiogeography 28: 345-358.
BUCKUP P. A. 1991. — The Characidiinae: aPhylogenetic Study of South America Darters and theirRelationships with Other Characiform Fishes. Ph. D.Dissertation, University of Michigan, Ann Arbor,USA, 391 p.
BUCKUP P. A. 1993. — The monophyly of theCharacidiinae, a Neotropical group of characiformfishes (Teleostei: Ostariophysi). Zoological Journal ofthe Linnaean Society 108: 225-245.
BUCKUP P. A. 1998. — Relationships of theCharacidiinae and phylogeny of characiform fishes(Teleostei: Ostariophysi), in MALABARBA L. R., REISR. E., VARI R. P., LUCENA Z. M. & LUCENA C. A.(eds), Phylogeny and Classification of the NeotropicalFishes. Museu de Ciências e Tecnologia daPUCRS/Edipucrs, Porto Alegre: 123-144.
CASTRO R. M. C. & VARI R. P. 1990. — Moo-jenichthys Miranda Ribeiro (Pisces: Ostariophysi:Characidae), a phylogenetic reappraisal andredescription. Proceedings of the Biological Society ofWashington 103 (3): 525-542.
Phylogeny and coevolution of Rhinoxenus (Platyhelminthes, Monogenoidea) and their hosts
463ZOOSYSTEMA • 2005 • 27 (3)

DOMINGUES M. V. & BOEGER W. A. 2002. —Neotropical Monogenoidea. 40. Protorhinoxenusprochilodi n. gen., n. sp. (Monogenoidea: Ancyro-cephalinae), parasite of Prochilodus lineatus(Prochilodontidae: Characiformes) from SouthBrazil. Folia Parasitologica 49: 35-38.
FINK S. V. & FINK W. L. 1981. — Interrelationshipsof the ostariophysian fishes (Teleostei). ZoologicalJournal of the Linnaean Society 72: 297-353.
GÉRY J. 1977. — Characoids of the World. TFHPublications, Neptune City, New Jersey, 672 p.
GREENWOOD P. H., ROSEN D. E., WEITZMAN S. H.& MYERS G. S. 1966. — Phyletic studies ofteleostean fishes with provisional classification ofliving forms. Bulletin of the American Museumof Natural History 131 (4): 339-456.
HENNIG W. 1966. — Phylogenetic Systematics.University of Illinois Press, Urbana, 263 p.
KRITSKY D. C., BOEGER W. A. & THATCHER V. E.1985. — Neotropical Monogenea. 7. Parasites ofthe pirarucu, Arapaima gigas (Cuvier), with descrip-tions of two new species and redescription ofDawestrema cycloancistrium Price and Nowlin, 1967(Dactylogyridae, Ancyrocephalinae). Proceedings ofthe Biological Society of Washington 98 (2): 321-331.
KRITSKY D. C., THATCHER V. E. & BOEGER W. A.1986. — Neotropical Monogenea. 8. Revision ofUrocleidoides (Dactylogyridae, Ancyrocephalinae).Proceedings of the Helminthological Society ofWashington 53: 1-37.
KRITSKY D. C., BOEGER W. A. & THATCHER V. E.1988. — Neotropical Monogenea. 11. Rhinoxenus,new genus (Dactylogyridae: Ancyrocephalinae)with descriptions of three new species from thenasal cavities of Amazonian Characoidea.Proceedings of the Biological Society of Washington101 (1): 87-94.
MADDISON W. P., DONOGHUE M. J. & MADDISOND. R. 1984. — Outgroup analysis and parsimony.Systematic Zoology 33: 83-103.
MALABARBA L. R. 1998. — Monophyly of theCheirodontinae, characters and major clades(Ostarioshysi: Characidae), in MALABARBA L. R.,REIS R. E., VARI R. P., LUCENA Z. M. & LUCENAC. A. (eds), Phylogeny and Classification of theNeotropical Fishes. Museu de Ciências e Tecnologiada PUCRS/Edipucrs, Porto Alegre: 193-233.
MIZELLE J. D. 1936. — New species of trematodesform gills of Illinois fishes. American MidlandNaturalist 17: 785-806.
NELSON G. 1996. — Fishes of the World. 3rd ed.Wiley-Interscience, New York, 523 p.
ORTÍ G. & MEYER A. 1997. — The radiation ofcharaciform fishes and the limits of resolution ofmitochondrial ribosomal DNA sequences.Systematic Zoology 46 (1): 75-100.
SWOFFORD D. L. 2001. — Phylogenetic Analysis UsingParsimony (*and Other Methods). Version 4. SinauerAssociates, Sunderland, Massachusetts.
VARI R. P. 1998. — Higher lever phylogeneticconcepts within characiforms (Ostariophysi), a his-torical review, in MALABARBA L. R., REIS R. E.,VARI R. P., LUCENA Z. M. & LUCENA C. A. (eds),Phylogeny and Classification of the Neotropical Fishes. Museu de Ciências e Tecnologia daPUCRS/Edipucrs, Porto Alegre: 111-122.
WATROUS L. E. & WHEELER Q. D. 1981. — The out-group comparison method of character analysis.Systematic Zoology 30: 1-11.
WILEY E. O. 1981. — Phylogenetics: the Theory andPractice of Phylogenetic Systematics. Wiley, NewYork, 439 p.
WILEY E. O. 1988. — Parsimony analysis and vicari-ance biogeography. Systematic Zoology 37: 271-290.
Submitted on 19 January 2004;accepted on 1st December 2004.
Domingues M. V. & Boeger W. A.
464 ZOOSYSTEMA • 2005 • 27 (3)

Phylogeny and coevolution of Rhinoxenus (Platyhelminthes, Monogenoidea) and their hosts
465ZOOSYSTEMA • 2005 • 27 (3)
APPENDIX
List of hosts, number of specimens and locality examined during this study. Abbreviations: AM, Amazonas; BR, Brazil; FG, FrenchGuiana; GO, Goiás; MS, Mato Grosso do Sul; PR, Paraná; TT, Trinidad-Tobago; *, specimens collected by researchers of the Núcleode Pesquisas em Limnologia, Ictiologia e Aqüicultura (NUPELIA-Universidade Estadual de Maringá-Brazil) between 1992 and 1995.
Species Specimens Localities
ANOSTOMIDAE
Leporellus vittatus (Valenciennes, 1850) 4 * Rio Paraná, PR, BR; Rio Tibagi, desembocadura TrêsBocas, PR, BR, 24.IV.1997; Rio Tibagi, PR, BR, 29.V.1997
Leporinus agassizii Steindachner, 1876 1 Rio Capucapu, AM, BR, 30.X.1989Leporinus cf. amblirhynchus 2 Rio Tibagi, desembocadura Três Bocas, PR, BR, Garavello & Britski, 1987 24.IV.1997Leporinus elongatus Valenciennes, 1849 36 Rio Paraná, PR, BR, 17-19.VI.1996; Rio Tibagi, PR, BR,
24.IV.1997, 28.V.1997Leporinus friderici (Bloch, 1794) 66 Rio Paraná, PR, BR, 18.VI.1996; Rio Baía, MS, BR,
18.VI.1996; Foz do Iguaçu, PR, BR, 15.XI.1996; RioGuandu, RJ, BR, 22-25.IV.1991
Leporinus cf. friderici (Bloch, 1794) 1 Rio Tibagi, desembocadura Três Bocas, PR, BR,24.IV.1997
Leporinus grantii Eigenmann, 1912 3 Iracoubo, Degrad Forian, FG, 8-10.X.1996Leporinus lacustris Amaral Campos, 1945 64 * Rio Paraná, PR, BRLeporinus obtusidens (Valenciennes, 1836) 89 Foz do Iguaçu, PR, BR, 15.XI.1996; * Rio Paraná, PR, BRLeporinus octofasciatus Steidachner, 1915 3 Rio Tibagi, Barra do Ribeirão Tigrinho, PR, BR, 23.IV.1997;
Rio Tibagi, desembocadura Três Bocas, PR, BR,24.IV.1997; Rio Tibagi, PR, BR, 24-28.V.1997
Leporinus striatus Kner, 1858 3 Rio Tibagi, PR, BR, 29.V.1997Leporinus sp. 1 Rio Tibagi, PR, BR, 29.V.1997Schizodon altoparanae 38 Rio Paraná, PR, BR; Rio Paraná, PR, BR, 19.VI.1996Garavello & Britski, 1990Schizodon borelli (Boulenger, 1900) 144 * Rio Paraná, PR, BRSchizodon nasutus Kner, 1858 9 Rio Tibagi, PR, BR, 24.IV.1997, 28.V.1997Schizodon sp. 19 Foz do Iguaçu, PR, BR, 15.XI.1996
CHARACIDAE
Acestrorhynchus guianensis Menezes, 1969 2 Iracoubo, Degrad Forian, FG, 8-10.X.1996Acestrorhynchus lacustris (Lütken, 1875) 109 * Rio Paraná, PR, BR; Barra do Ribeirão Tigrinho, Rio
Tibagi, PR, BR, 23.IV.1997Aphiocharax sp. 1 Rio Guaporé, AM, BR, no dateAstyanax bimaculatus (Linnaeus, 1758) 55 Fish Farm, Sidrolândia, MS, BR, 3.XI.1996; Barra do
Ribeirão Tigrinho, Rio Tibagi, PR, BR, 23, 28.IV.1997Astyanax fasciatus (Cuvier, 1819) 10 Rio Tibagi, PR, BR, 28-29.V.1997Astyanax cf. janeiroensis Eigenmann, 1908 2 Rio Guandu, RJ, BR, 25.IV.1991Astyanax cf. scabripinnis (Jenyns, 1842) 12 Rio Cubatão, PR, BR, 10.V.1996Astyanax sp. 22 Lagoa Dourada, PR, BR, 27.X.1993; Rio Pequeno, PR, BR,
20.VI.1994; Lagoa da Petrosix, São Mateus do Sul, PR, BR,21.V.1997
Brycon orbignyanus (Valenciennes, 1850) 24 * Rio Paraná, PR, BR; Fish Farm, Estrada Alexandra-Matinhos, PR, BR, 1996
Bryconamericus stramineus Eigenmann, 1908 90 Foz do Iguaçu, PR, BR, 6.VII.1996, 15.X.1996Bryconamericus sp. 4 Desembocadura Três Bocas, Rio Tibagi, PR, BR,
24.IV.1997Characidium lanei Travassos, 1967 120 Rio Cubatão, PR, BR, 10.V.1996; Rio Vitória, PR, BR,
11.V.1996; Rio Peneira, PR, BR, 13.V.1996; Rio Perequê,PR, BR, 4.X.1994, 30.VI.1995, 3.VIII.1995; Rio Dois deFevereiro, PR, BR, 18.VII.1994, 15.III.1995; Rio Cacatu, PR,BR, 30.VI.1995
Characidium pterostictum Gomes, 1947 14 Rio Cacatu, PR, BR, 30.VI.1995Characidum sp. 3 Rio Cacatu, PR, BR, 18.VI.1996; Estrada do Lixão, PR, BR,
9.V.1994Deuterodon gaujo Lucena & Lucena, 1992 7 Rio Cubatão, PR, BR, 11.V.1996

Domingues M. V. & Boeger W. A.
466 ZOOSYSTEMA • 2005 • 27 (3)
Deuterodon sp. 6 Rio da Venda, PR, BR, 18.VII.1994; Rio Cubatão, PR, BR,11.V.1196
Galeocharax knerii (Steindachner, 1875) 24 * Rio Paraná, PR, BRHollandeichythys multifasciatus 2 Rio Cubatão, PR, BR, 10.V.1996(Eigenmann & Norris, 1900)Hyphessobrycon cf. bifasciatus Ellis, 1911 10 Rio da Venda, PR, BR, 18.VII.1994Hyphessobrycon griemi Hoedeman, 1957 12 Rio Perequê, PR, BR, 3.VIII.1995Hyphessobrycon cf. luetkeni (Boulenger, 1887) 12 Rio Perequê, PR, BR, 3.VIII.1995; Rio Cubatão, PR, BR,
10.V.1996Hyphessobrycon reticulatus Ellis, 1911 9 Rio Perequê, PR, BR, 2.III.1993, 20.VI.1994Hyphessobrycon cf. reticulatus Ellis, 1911 1 Unidentified localityHyphessobrycon sp. 2 Rio Caracu, PR, BR, 18.VI.1996Markiana nigripinnis (Perugia, 1891) 2 Rio Miranda, MS, BR, 30.XI.1996Mimagoniates cf. lateralis (Nichols, 1913) 14 Rio Dois de Fevereiro, PR, BR, 18.VII.1994; Rio da Venda,
PR, BR, 18.VII.1994Mimagoniates microlepis (Steidachner, 1876) 7 Rio Cubatão, PR, BR, 10.V.1996Mimagoniatis sp. 1 Rio Dois de Fevereiro, PR, BR, 18.VII.1997Moenkhausia dichoura (Kner, 1858) 18 Rio Cubatão, PR, BR, 10.V.1996; Rio Vitória, PR, BR,
11.V.1996; Rio Guaporé, AM, BROligosarcus hepsetus (Cuvier, 1829) 2 Rio Guandu, RJ, BR, 18.I.1991; Rio Cubatão, PR, BR,
12.V.1996Oligosarcus cf. hepsetus (Cuvier, 1829) 2 Rio Guandu, RJ, BR, 25.IV.1991Poptella paraguayensis (Eigenmann, 1907) 3 Rio Miranda, MS, BR, 30.XI.1996; Baía da Medalha, MS,
BR, 30.XI.1996Roeboides prognathus (Boulenger, 1895) 7 Foz do Iguaçu, PR, BR, 15.XI.1996; Baía da Medalha, MS,
BR, 1.XII.1996Salminus brasiliensis (Cuvier, 1816) 4 Rio Paraná, PR, BR; Baía da Medalha, MS, PR, 17.VI.1996Salminus hillari Valenciennes, 1850 1 * Rio Paraná, PR, BRSalminus sp. 1 Lagoa Dourada, PR, BR, 27.X.1993Triportheus cf. nematurus (Kner, 1858) 10 Rio Miranda, MS, BR, 30.XI.1996, 4-5.VIII.1998
SERRASALMINAE
Colossoma macropomum (Cuvier, 1818) 2 Fish Farm, Pariqueraçu, SP, BR, 6.V.1996Metynnis lippincottianus (Cope, 1870) 2 Lago Miri, Igarapé Taparabu, FG, 15-16.X.1996Myleus levis Eigenmann & McAtee, 1907 1 * Rio Paraná, PR, BRPiaractus mesopotamicus (Holmberg, 1887) 24 * Rio Paraná, PR, BR; Farm Fish, Toledo, PR, BR,
11.VII.1994; Fish Farm, Pescobrás, PR, BR, 17.VII.1995Pristobrycon sp. 1 Rio Capucapu, Cachoeira da Garça, AM, BR, 1.XI.1989Pristobrycon striolatus (Steindachner, 1908) 2 Rio Jatapu, Maracana, AM, BR, 1-2.XI.1989Pygocentrus naterreri Kner, 1858 7 Rio Vatumã, AM, BR, 2.XI.1989; Lago da Manga, Igarapé
Taparapu, FG, 15-16.X.1996Serrasalmus altuvei Ramírez, 1965 1 Anavilhanas, AM, BR, 6.VII.1996Serrasalmus eigenmanni Norman, 1929 1 Rio Jatapu, AM, BR, 1-2.XI.1989Serrasalmus elongatus Kner, 1858 1 Anavilhanas, AM, BR, 6.VII.1996Serrasalmus gouldingi Fink & Machado-Allison, 2 Rio Uatumã, Lago Tapana, Santa Anna, AM, BR, 1992 2-3.XI.1989Serrasalmus marginatus Valenciennes, 1836 148 * Rio Paraná, PR, BR; Rio Paraná, PR, BR, 18-19.VI.1996;
Rio Baía, MS, BR, 18.VI.1996; Rio Miranda, MS, BR,30.XI.1996
Serrasalmus rhombeus (Linnaeus, 1766) 15 Rio Iatapu, AM, BR, 2.XI.1989; Rio Pitanga, AM, BR,11.X.1989; Anavilhas, AM, BR, 11.X.1989; Iracoubo, DegradForian, FG, 8-10.X.1996; Serra da Mesa, GO, BR, 11.VIII.1996
Serrasalmus cf. serrulatus 2 Anavilhanas, AM, BR, 7.VII.1989Serrasalmus spilopleura Kner, 1858 50 * Rio Paraná, PR, BR; Rio Jatapu, Lago Maracana, AM, BR,
1-2.XI.1989; Rio Uatumã, Lago Tapana, Santa Anna, AM,BR, 2-3.XI.1989; Rio Baía, MS, BR, 18.VI.1996; Lago Miti,Igarapá Tapana, GF, 15-16.X.1996
Serrasalmus sp. 1 Represa Xavantes, SP, BR, 17.IV.1995
CHILODONTIDAE
Chilodus punctatus Müller & Troschel, 1844 9 Aquarium

CYNODONTIDAE
Rhaphiodon vulpinus Spix & Agassiz, 1829 76 Rio Paraná, PR, BR, 18.VI.1996; Rio Baía, MS, BR, 17-18.VII.1996
CURIMATIDAE
Curimata cyprinoides (Linnaeus, 1766) 3 Iracoubo, Degrad Forian, FG, 8-10.X.1996Cyphocharax gilbert (Quoy & Gaimard, 1824) 4 Rio Guandu, RJ, BR, 22.IV.1991Cyphocharax nagelli (Steindachner, 1881) 29 * Rio Paraná, PR, BRCyphocharax santacatarinae 6 Rio Cubatão, PR, BR, 12-13.V.1996(Fernández-Yépez, 1948)Cyphocharax sp. 11 Fish Farm, Sidrolândia, MS, BR, 3.XII.1996Steindachnerina argentea (Gill, 1858) 8 Rio Rouca, TT Steidachnerina brevipinna 2 Baía da Medalha, MS, BR, 30.XI.1996(Eigenmann & Eigenmann, 1889)Steidachnerina insculpita 89 * Rio Paraná, PR, BR; Barra do Ribeirão Tigrinho, Rio (Fernández & Yépez, 1948) Tibagi, PR, BR, 23-24.IV.1997; Desembocadura Três
Bocas, Rio Tibagi, PR, BR, 24.IV.1997; Rio Tibagi, PR, BR,28-29.V.1997
ERHYTHRINIDAE
Hoplias malabaricus (Bloch, 1794) 192 Lagoa Dourada, PR, BR, 26.X.1993; Rio Cubatão, PR,BR, 11.V.1996; Rio Baía, MS, BR, 17.VI.1996; Rio Paraná,PR, BR, 18.VI.1996; * Rio Paraná, PR, BR; Iracoubo,Degrad Forian, FG, 8-10.X.1996; Rio Guandu, RJ, BR,25.IV.1997
Hoplias cf. malabaricus (Bloch, 1794) 6 Represa Voçoroca, PR, BR, 16.IV.1998; Rio Piraquara, PR,BR, 28.IV.1998; Rio Pequeno, PR, BR, 8.VI.1998
Hoplias sp. 16 Rio Piraquara, PR, BR, 2.III.1993, 22.IV.1994; Rio Dois deFevereiro, PR, BR, 18.VII.1994; Rio da Venda, PR, BR,18.VII.1994; Rio Perequê, PR, BR, 4.VII.1994, 6.IX.1994;Represa Capivari-Cachoeira, PR, BR, 14.III.1995; * RioParaná, PR, BR
Hoplerythrinus unitaenatus (Agassiz, 1829) 10 * Rio Paraná, PR, BR
GASTEROPELECIDAE
Carnegiella striagata (Günther, 1864) 2 AquariumGasteropelecus levis (Eigenmann, 1909) 11 AquariumUnidentified specimen 3 Ilha de Marchantaria, AM, BR, no date
HEMIODONTIDAE
Hemiodus orthonops Eigenmann & Kennedy, 2 Baía da Medalha, MS, BR, 1.XII.19961903Hemiodopsis microlepis (Kner, 1858) 5 Furo do Catalão, AM, BR, 5.I.1989
PARODONTIDAE
Apareiodon affinis (Steindachner, 1879) 9 Desembocadura Três Bocas, Rio Tibagi, PR, BR,24.IV.1997; Rio Tibagi, PR, BR, 28-29.IV.1997
Apareiodon cf. affinis 3 Desembocadura Três Bocas, Rio Tibagi, PR, BR,24.IV.1997
Parodon nasus Kner, 1859 2 Desembocadura Três Bocas, Rio Tibagi, PR, BR,24.IV.1997
PROCHILODONTIDAE
Prochilodus lineatus (Valenciennes, 1836) 252 * Rio Paraná, PR, BR; Represa Capivari-Cachoeira, PR, BR,16.XII.1994, 13.III.1995, 23.V.1995, 6.VI.1995, 26.X.1995;Rio Paraná, PR, BR, 17-19.VI.1996; Rio Baía, PR, BR, 17-18.VI.1996, Foz do Iguaçu, PR, BR, 15.XI.1996; Barra doRibeirão Tigrinho, Rio Tibagi, PR, BR, 23.IV.1997;Desembocadura Três Bocas, Rio Tibagi, PR, BR,24.IV.1997; Rio Tibagi, PR, BR, 28-29.V.1997
Prochilodus sp. 6 Fish Farm, Bocaiúva do Sul, PR, BR, 7.VIII.1997; FishMarket, PR, BR, 12.VIII.1997
Phylogeny and coevolution of Rhinoxenus (Platyhelminthes, Monogenoidea) and their hosts
467ZOOSYSTEMA • 2005 • 27 (3)




























