Embed Size (px)
Citation preview

NPT
GLAa
db
Pc
Pd
v
A(StaItdePcbmPtNd1GtgoirsinvtrA
*1EAaaghNn
Neuroscience 124 (2004) 147–160
0d
EUROKININ 1 RECEPTOR EXPRESSION AND SUBSTANCE PHYSIOLOGICAL ACTIONS ARE DEVELOPMENTALLY REGULATED IN
HE RABBIT RETINAKmm
SvnrpecmssidtiTr(p
atBtsnkctaemalloaoofi1lta
. CASINI,a* M. DAL MONTE,b F. FORNAI,c
. BOSCO,a D. WILLEMS,a Q. YANG,d Z. J. ZHOUd
ND P. BAGNOLIb
Dipartimento di Scienze Ambientali, Universita della Tuscia, Largoell’Universita/D, 01100 Viterbo, Italy
Dipartimento di Fisiologia e Biochimica “G. Moruzzi,” Universita diisa, 56127 Pisa, Italy
Dipartimento di Morfologia Umana e Biologia Applicata, Universita diisa, 56127 Pisa, Italy
Departments of Physiology and Biophysics and Ophthalmology, Uni-ersity of Arkansas for Medical Sciences, Little Rock, AR 72205, USA
bstract—We investigated the expression of the substance PSP) receptor (the neurokinin 1 receptor, NK1 receptor) andP functional effects in developing rabbit retinas. NK1 recep-
ors in adult retinas were in a population of cone bipolar cellsnd in dopaminergic amacrine cells, as previously described.n contrast, at birth and at postnatal day (PND) 6, NK1 recep-ors were exclusively expressed by cholinergic amacrine andisplaced amacrine cells. NK1 receptor expression in cholin-rgic cells was still observed at PND10 (eye opening), while atND21 it was confined to cholinergic cells of the inner nu-lear layer. Starting at PND10, NK1 receptors were also inipolar cells and in dopaminergic amacrine cells. A fullyature NK1 receptor expression pattern was observed atND35. Dopamine release was assessed in isolated retinas in
he presence of SP, the NK1 receptor agonist GR73632 or theK1 receptor antagonist GR82334. At PND35, extracellularopamine was significantly increased by 10 �M SP or 0.01–00 �M GR73632, and it was decreased by 0.01–10 �MR82334. No effects were detected in developing retinas up
o PND21. Ca2� imaging experiments were performed in sin-le cholinergic cells identified by their “starburst” morphol-gy in perinatal retinas. Intracellular Ca2� levels were signif-
cantly increased by 1 �M SP or GR73632. This effect waseversibly inhibited by 1 �M GR82334. These data demon-trate that both NK1 receptor expression and SP physiolog-cal actions are developmentally regulated in the retina. SPeurotransmission in the immature retina may subserve de-elopmental events, and SP is likely to represent an impor-ant developmental factor for the maturation of retinal neu-ons and circuitries. © 2004 IBRO. Published by Elsevier Ltd.ll rights reserved.
Corresponding author. Tel: �39-0761-357-040; fax: �39-0761-357-79.-mail address: [email protected] (G. Casini).bbreviations: CG-1, Calcium Green-1; ChAT, cholinecetyltransferase; DA, dopamine; DOPAC, dihydroxyphenylaceticcid; E30, embryonic day 30; F, average fluorescence intensity; GCL,anglion cell layer; HPLC, high-pressure liquid chromatography; HVA,omovanillic acid; INL, inner nuclear layer; IPL, inner plexiform layer;K1, neurokinin 1; OGB-1, Oregon green 488 BAPTA-1; PND, post-
Natal day; SP, substance P; TH, tyrosine hydroxylase.
306-4522/04$30.00�0.00 © 2004 IBRO. Published by Elsevier Ltd. All rights reseroi:10.1016/j.neuroscience.2003.10.049
147
ey words: neuropeptide receptor, tachykinin peptide, dopa-inergic amacrine cells, cholinergic amacrine cells, dopa-ine release, calcium imaging.
tudies of peptidergic systems in developing retinas pro-ide elements to postulate possible physiologic actions ofeuropeptides that may contribute to the differentiation ofetinal neurons and to the maturation of their functionalroperties (see Bagnoli et al., 2003, for review). The pres-nce of transient features in the developmental patterns ofertain peptidergic systems indicates that peptide actionsay vary in different periods of retinal development, and
uggests that retinal circuits are likely to function in aubstantially different manner before the retina becomes
nvolved in the processing of visual stimuli. However, toate there is only little evidence that transient changes inhe developmental expression of peptidergic systems are,n fact, related to functional changes in peptide actions.he present study shows that, in the developing rabbitetina, changes in the expression pattern of substance PSP) receptors are accompanied by modifications of SPhysiological effects.
SP is a member of the tachykinin peptide family, whichlso includes neurokinin A, two neurokinin A-related pep-ides (neuropeptide K and neuropeptide �) and neurokinin. These peptides are structurally related and derived from
wo tachykinin genes (see Maggio, 1988; Otsuka and Yo-hioka, 1993, for reviews). The cellular actions of SP,eurokinin A and neurokinin B are mediated by the neuro-inin receptors, belonging to the superfamily of G protein-oupled receptors and termed NK1, NK2 and NK3, respec-ively (Otsuka and Yoshioka, 1993). Immunohistochemicalnd in situ hybridization studies have reported the pres-nce of tachykinin peptides in the retinas of numerousammalian species. SP is localized to wide-field amacrinend displaced amacrine cells with processes arborizing in
aminae 1, 3 and 5 of the inner plexiform layer (IPL; IPLaminae are named after Cajal, 1893), and to ganglion cellsf rat, rabbit, hamster and primate retinas (see Casini etl., 1997a, 2002a, for references). Main functional actionsf SP in adult vertebrate retinas include excitatory effectsn ganglion cells that do not modify ganglion cell receptiveeld properties (Dick and Miller, 1981; Glickman et al.,982; Zalutsky and Miller, 1990). NK1 receptors have been
ocalized in adult rat and rabbit retinas using immunohis-ochemical and in situ hybridization techniques (Kondoh etl., 1996; Casini et al., 1997b, 2002a). In the rat retina,
K1 receptors are expressed by GABAergic interplexiformved.

adwtm
cacgiNieravbt2pfcfSotip
Nwpd8Ht
I
vPdcsdbwpg
eCi1iemrm
11w(
znBswcodtStpwa
sdmBtcPi
A
Rcrs(SmGtTeNatsw2dw((sOitmaswb(wa
G. Casini et al. / Neuroscience 124 (2004) 147–160148
nd amacrine cells, and by the adrenergic and most of theopaminergic amacrine cells (Casini et al., 1997b),hereas in the rabbit retina they are expressed by a dis-
inct population of cone bipolar cells and by all the dopa-inergic amacrine cells (Casini et al., 2002a).
In developing rat and rabbit retinas, SP-containingells have been detected at early postnatal ages, and theyre likely to be amacrine, displaced amacrine and ganglionells (Casini et al., 1997a, 2000). These observations sug-est that SP may be released in the developing retina and
t may interact, perhaps by paracrine mechanisms, withK1 receptors, whose developmental patterns have been
nvestigated in the rat retina (Oyamada et al., 1999; Casinit al., 2000). These receptors are detected in the newbornat retina, and their mature expression pattern is achievedround the time of eye opening. Interestingly, some obser-ations indicate a substantial rearrangement in the distri-ution of NK1-immunoreactive processes in the IPL be-ween postnatal day (PND) 7 and PND12 (Casini et al.,000), suggesting that SP and NK1 receptors may mediaterocesses in the developing rat retina that are differentrom those in the retina capable of visual information pro-essing. The purpose of the present study was to collecturther information on possible developmental actions ofP by investigating NK1 receptor expression in the devel-ping retina of a different animal model, the rabbit, and byesting some functional effects of SP in the immature andn the mature retina. Some preliminary results have beenresented in abstract form (Casini et al., 2002b).
EXPERIMENTAL PROCEDURES
ew Zealand albino rabbits obtained from commercial sourcesere used in these studies. Experiments were performed in com-liance with the Italian law on animal care N° 116/1992, in accor-ance with the European Community Council Directive (EEC/609/6) and with guidelines established by the National Institutes ofealth. All efforts were made to reduce both animal suffering and
he number of animals used.
mmunohistochemistry
Tissue preparation. Retinas were collected from animals atarious postnatal ages, ranging from the day of birth (PND0) toND35. The rabbits were killed with an overdose of chloral hy-rate (i.p.), the eyes were removed, the anterior segments wereut away, and the eyecups, containing the retinas, were immer-ion fixed in 4% paraformaldehyde for 2 h. Retinas were thenissected and transferred to 25% sucrose in 0.1 M phosphateuffer (pH 7.4) for storage until cryosectioning. Sections (10 �m)ere cut in a plane either perpendicular (coronal sections) orarallel (horizontal sections) to the vitreal surface, mounted ontoelatin-coated slides and stored at �20 °C.
Antibodies. NK1 receptors were labeled either with a guin-a-pig or with a rabbit polyclonal antibody (Chemicon, Temecula,A, USA), directed to a peptide sequence (393–407) correspond-
ng to the predicted C-terminus of rat NK1 receptor (Vigna et al.,994). Both antisera were used at 1:500 dilution. Double-label
mmunofluorescence experiments were performed using the guin-a-pig NK1 receptor antiserum in conjunction with: i) a mouseonoclonal antibody (clone 2/40/15; Chemicon) directed to ty-
osine hydroxylase (TH; Rohrer et al., 1986), which stains dopa-
inergic amacrine cells in the rabbit retina (Casini and Brecha, o992a,b), used at 1:200; ii) a mouse monoclonal antibody (cloneE6; Chemicon) directed to choline acetyltransferase (ChAT),hich stains cholinergic amacrine cells in rat and in chick retinas
Cellerino et al., 1998; Prada et al., 1999), used at 1:200.
Immunohistochemical procedures. Both coronal and hori-ontal sections were treated for single- and double-label immu-ofluorescence as previously described (Casini et al., 2002a).riefly, after overnight incubation with primary antibodies, theections were washed with phosphate buffer and incubatedith the appropriate secondary antibodies raised in goat andonjugated with Alexa Fluor fluorescent dyes (Alexa Fluor 546r Alexa Fluor 488; Molecular Probes, Eugene, OR, USA) at ailution of 1:100 for 2 h at room temperature. The slides were
hen coverslipped in a phosphate buffer– glycerin mixture.pecificity of immunostaining was evaluated by preadsorbing
he NK1 receptor antibodies with 10 �M of the NK1 receptoreptide fragment NK1393– 407 overnight at 4 °C. Immunostainingas not detected in sections incubated with the preadsorbedntibodies.
Figure preparation. Immunofluorescent material was ob-erved with an Olympus fluorescence microscope equipped with aigital photocamera and with a Leitz laser confocal scanningicroscope. Digital images were acquired and saved as TIF files.oth the images from the digital photocamera and those acquired
hrough the confocal microscope were adjusted to the final size,orrected for contrast and brightness and labeled using Adobehotoshop 5.0 (Adobe Systems, Mountain View, CA, USA). Final
mages were saved at a minimum of 300 dpi.
ssay of dopamine and metabolites
abbits at PND0, PND10, PND21 and PND35 were killed byervical dislocation. The eyes were rapidly enucleated and theetinas dissected in saline. Retinas were incubated in 1 ml ofaline alone or of saline containing various concentrations of SPSigma, St. Louis, MO, USA), a NK1 receptor agonist (GR73632;igma) or a NK1 receptor antagonist (GR82334; Sigma) for 15in at room temperature. Concentrations of SP, GR73632 andR82334 were 0.01, 0.1, 1, 10 or 100 �M. Three retinas from
hree different animals were used in each experimental condition.he wet weight of the retinas, determined at the end of eachxperiment, did not vary significantly within the single age groups.inety microliter aliquots of the incubation medium were collectednd added with 10 �l of 1 N perchloric acid. The samples werehen centrifuged at 30,000�g for 5 min at 4 °C. Finally, 40 �l of theupernatant were collected and stored at �80 °C until analysisith high-pressure liquid chromatography (HPLC). In particular,0 �l samples containing a constant amount (10 ng/ml) of 3,4-ihydroxybenzylamine (Sigma), used as an internal standard,ere injected into an HPLC system to measure levels of dopamine
DA), dihydroxyphenylacetic acid (DOPAC) and homovanillic acidHVA), as previously described (Fornai et al., 1997). The HPLCystem consisted of a reversed-phase column (250�4.5 mm;DS C18; Beckman, Palo Alto, CA, USA) and two electrochem-
cal detectors (analytical cell; ESA, 5011). The potentials of thewo electrodes were maintained at �0.35 and �0.35 V. Theobile phase consisted of a citrate-phosphate buffer (0.04 M citriccid, 0.06 M Na2HPO4·2H2O) containing 0.1 mM EDTA, 0.6 mModium 1-heptanesulphonate and 10% methanol. The flow rateas 1 ml/min. Values of DA and metabolites were calculatedased on regression curves obtained with known concentrationsexternal standard) for each compound. The reliability of the assayas based on previous pilot experiments from control retinas tossess the consistency and stability of retinal extracellular levels
f DA and metabolites.
C
E(rFlprb(Ncfib1
tCB((mtwoRfl3tu(smcUrilwCrcdew
(3GwacimS
I
Itnwasl
ictt
p(tpcrp(eNwwc(lidmoirwgwpml
FaFbbpp
G. Casini et al. / Neuroscience 124 (2004) 147–160 149
alcium imaging in the flat-mount rabbit retina
yes from rabbits aged from embryonic day 30 (E30) to PND1gestation period: 31 days) were used to study the effects of NK1eceptor agonists and antagonists on cholinergic (starburst;amiglietti, 1983) amacrine cells. Pre-term rabbit pups were de-
ivered by cesarian section. Rabbit retinal flat-mounts were pre-ared as described previously (Zhou and Z hao, 2000). In brief,etinas were isolated in a cold (4–10 °C), oxygenated, HEPES-uffered Ames medium, which was modified from Ames mediumAmes and Nesbett, 1981; Sigma Chemicals) by replacingaHCO3 with 20 mM HEPES (pH�7.4). Each isolated retina wasut into two to four pieces, mounted ganglion cell side up on blacklter paper (type: HABP; Millipore, Billerica, MA, USA) and incu-ated in HEPES-buffered Ames medium at room temperature for–6 h until recording.
Displaced starburst cells in the ganglion cell layer (GCL) ofhe flat-mount retina were loaded with the calcium indicator dyealcium Green-1 (CG-1; Molecular Probes) or Oregon Green 488APTA-1 (OGB-1; Molecular Probes) using a Helios gene gun
Bio Receptorad Laboratory, Hercules, CA, USA) as describedKettunen et al., 2002). Dye-coated particles were prepared byixing 25 mg tungsten particles (1.7 �m in diameter; Bio Recep-
orad Laboratory) with 10 mg CG-1 or OGB-1 in 25 �l water. Theyere dried on a glass slide, separated into fine grains, and loadedn the inner wall of a polyvinylpyrrolidone-coated PE tubing (Bioeceptorad Laboratory). The particles were bombarded into theat-mount retina through polycarbonate filter paper (pore size:�m; Millipore) at a gas pressure of 60–100 psi. The retina was
hen immediately placed in a recording chamber and examinednder the epifluorescence illumination of an upright microscopeBX51; Olympus USA, Melville, NY, USA). Dye-loaded displacedtarburst cells were identified based on their distinctive dendriticorphology (Zhou, 1998) and were imaged with a cooled CCD
amera (Cascade; Roper Scientific Instruments, Tuscan, AZ,SA) controlled by MetaFluo software (Universal Imaging Corpo-
ation, Downingtown, PA, USA). To monitor the excitability ofndividual cells, oval zones were drawn around the soma of dye-oaded cells. The average fluorescence intensity (F) in each zoneas plotted as a function of time. Changes in intracellular freea2� were represented by relative changes in the emission fluo-
escence intensity, �F/F. Spontaneous and drug-inducedhanges in fluorescence were observed in both the somata andendrites of starburst cells. Comparisons of �F/F between differ-nt cells were made at the somata only, by drawing a circular zoneithin the perimeter of the cell body.
During Ca2� imaging, the retina was superfused continuously3–4 ml/min) with Ames medium saturated with 95% O2/5%CO2 at5–37 °C. Pharmacological agents, including SP, GR73632,R82334 and a cocktail of receptor antagonists (see Results)ere applied to the retina by bath perfusion. The timing of drugpplication was marked in the figures as the time when a drug-ontaining perfusion line was switched on. Exchange of solutionsn the “dead space” of the perfusion system usually took about 2
in. All drugs used for Ca2� imaging experiments were fromigma Chemicals.
RESULTS
mmunohistochemistry
n retinas at PND35, the NK1 immunostaining pattern washe same as that previously described in adult rabbit reti-as (Casini et al., 2002a). In particular, NK1 receptorsere expressed by bipolar cells whose axonal terminalsrborized in the IPL adjacent to the GCL, and by large-ized amacrine cells located in the proximal inner nuclear
ayer (INL) whose processes were mainly confined to lam- dna 1 of the IPL (Fig. 1). These cells have been shown toonstitute a population of ON-type cone bipolar cells andhe population of TH-containing amacrine cells, respec-ively (Casini et al., 2002a).
There were marked changes in the NK1 receptor ex-ression pattern during postnatal development. At PND0Fig. 2A, B), NK1 immunolabeling was present throughouthe retina and it was localized to somata located in theroximal INL and in the GCL, while immunostained pro-esses were confined to two continuous bands best cor-esponding to laminae 2 and 4 of the IPL. The sameattern was also observed at PND6 (Fig. 2C). At PND10near eye opening), the characteristics of a mature NK1xpression pattern were detected, namely the presence ofK1-immunostained bipolar cells and of amacrine cellsith processes arborizing in lamina 1 of the IPL. Togetherith these mature features, NK1-immunolabeled pro-esses were still observed in laminae 2 and 4 of the IPLFig. 2D). At PND21 (Fig. 2E), the immunofluorescentabeling was similar to that at PND10; however, NK1-mmunoreactive profiles in lamina 2 of the IPL were lessensely distributed, and those in lamina 4 appeared to beainly associated with bipolar cell axonal terminals. Thesebservations indicate the presence of two distinct phases
n the developmental expression of NK1 receptors in theabbit retina. A first phase, which is before eye opening,hen NK1 receptors are localized to two mirror-symmetricroups of cells, one in the INL and the other in the GCL,ith processes in laminae 2 and 4 of the IPL; a secondhase, which is near and after eye opening, when theature features of NK1 receptor expression are estab-
ished and the pattern observed in the first phase gradually
ig. 1. Pattern of NK1 receptor immunostaining in a coronal section ofrabbit retina at PND35 (confocal image, 1 �m-thick optical section).luorescent immunolabeling is localized to the plasma membrane ofipolar cells (arrow) and large amacrine cells (double arrow). Theipolar cell axon arborizes near the GCL, while the amacrine cellrocesses are confined to the IPL adjacent to the INL. OPL, outerlexiform layer. Scale bar�10 �m.
isappears. The pattern of NK1 receptor distribution in the

ficZiectn
D
Aiflce
FasacrNe
G. Casini et al. / Neuroscience 124 (2004) 147–160150
rst phase strongly reminds that of cholinergic amacrineells in adult and developing mammalian retinas (seehou, 2001a, for references). Therefore, in double label
mmunofluorescence experiments, we tested the possiblexpression of NK1 receptors in ChAT-containing amacrineells. We also performed NK1/TH double labelings to de-ermine the timing of NK1 receptor expression in dopami-
ig. 2. A: Low power fluorescence photomicrograph showing the gerrows indicate immunolabeled somata in the INL, arrowheads indicateections) showing the pattern of NK1 receptor immunostaining in corot PND6 (C), NK1 receptor immunoreactivity is localized to cell bodieorresponding to laminae 2 and 4. At PND10 (D) and at PND21 (E), NKise to processes in lamina 1 of the IPL (arrows). At both these ages, NKK1 immunostaining in lamina 2 (arrowheads in D) is weaker than at PNndings. Numbers of IPL laminae are shown in B and in D. Abbreviat
ergic amacrine cells. l
ouble label immunofluorescence: First phase
s shown in Fig. 3, at PND0 and at PND6, NK1 receptormmunolabeling was clearly distinct from TH immuno-uorescence. As previously reported (Casini and Bre-ha, 1992a,b), TH-containing amacrine cells are pres-nt in the rabbit retina during this period. They are
earance of NK1 immunostaining in a coronal section at PND0. Thelabeled somata in the GCL. B–E: Confocal images (1 �m-thick opticalns of retinas at different developmental ages. At birth (B, PND0) andINL and in the GCL and in processes arborizing in IPL regions bestolabeling is also observed in bipolar cells and in amacrine cells givingopositive fibers are also seen in laminae 2 and 4 of the IPL. At PND21,ile in lamina 4 it seems to be mainly associated with bipolar cell axonaln Fig. 1. Scale bars�10 �m.
neral appimmuno
nal sectios in the1 immun1-immunD10, wh
ions as i
ocalized to the INL adjacent to the IPL and origi-

nlnfi
Cl
spcrcis
Fac
Faad
G. Casini et al. / Neuroscience 124 (2004) 147–160 151
ate short primary processes that remain confined toamina 1 of the IPL. TH-immunostained processes wereever observed to intermingle with NK1-immunolabeledbers.
In coronal sections treated with both NK1 receptor andhAT antibodies at PND0 and at PND6 (Fig. 4), double
abeled cell bodies were observed. Some ChAT-containing
ig. 3. Confocal images (1 �m-thick optical sections) of coronal sentibodies directed to NK1 receptors (green) and to TH (red). Overlayells are not labeled by NK1 antibodies. Abbreviations as in Fig. 1. S
ig. 4. Confocal images (1 �m-thick optical sections) of coronal sentibodies directed to NK1 receptors (green) and to ChAT (red). Overland fibers are also labeled by NK1 antibodies. However, some ChAT
ouble labeled cells; arrowheads indicate ChAT containing cells that are not labomata, particularly among those in the GCL, did not dis-lay NK1 receptor immunoreactivity, indicating that not allholinergic cells may express NK1 receptors or that theseeceptors are preferentially expressed on the cell pro-esses. The NK1-immunolabeled processes were local-
zed to the same IPL laminae where also ChAT-immuno-tained fibers were observed. Both immunostainings had a
PND0 (upper row) and at PND6 (bottom row) double labeled withNK1/TH) of both immunolabelings show that TH-containing amacrine10 �m.
PND0 (upper row) and at PND6 (bottom row) double labeled with(ChAT/TH) of both immunolabelings show that ChAT-containing cells
reactive somata are devoid of NK1 immunostaining. Arrows indicate
ctions atimages (cale bar�
ctions aty images-immuno
eled by NK1 antibodies. Abbreviations as in Fig. 1. Scale bar�10 �m.

pcttsm
aNgorpn
D
Aibisfipl
wnilotdcci
Fi(
FacNI
G. Casini et al. / Neuroscience 124 (2004) 147–160152
unctate appearance that prevented the detection of aomplete overlap of NK1 and ChAT immunostainings inhe IPL, although both NK1 receptors and ChAT were likelyo be expressed in the same processes. In horizontalections observed with conventional epifluorescence,ost cholinergic amacrine cell bodies were surrounded by
ig. 5. Overlay image of fluorescence photomicrographs from the INLn a horizontal section of a PND2 retina showing NK1 immunolabelinggreen) encircling ChAT-containing somata (red). Scale bar�20 �m.
ig. 6. Confocal images (1 �m-thick optical sections) of coronal secntibodies directed to NK1 receptors (green) and to TH (red). Overlayells at these ages display NK1 immunostaining both at the plasmaK1-immunoreactive fibers are still detected in laminae 2 and 4 of the
NL (arrow), indicating the presence of NK1-expressing bipolar cells (see also
rim of NK1 immunoreactivity, indicating the presence ofK1 receptors at their plasma membrane (Fig. 5). To-ether with the data of Ca2� imaging (see below), thesebservations indicate that during the first phase of NK1eceptor developmental expression, NK1 receptors are ex-ressed by cholinergic amacrine cells. In contrast, they areot present in dopaminergic amacrine cells.
ouble label immunofluorescence: Second phase
round the time of eye opening (PND10), NK1 receptormmunostaining was observed at or near the plasma mem-rane of TH-containing amacrine cells (Fig. 6) and NK1-
mmunoreactive puncta could be detected on TH-immuno-tained processes. In addition, the NK1-immunolabeledbers in laminae 2 and 4 of the IPL, observed in the firsthase, were still detected. A similar pattern of double
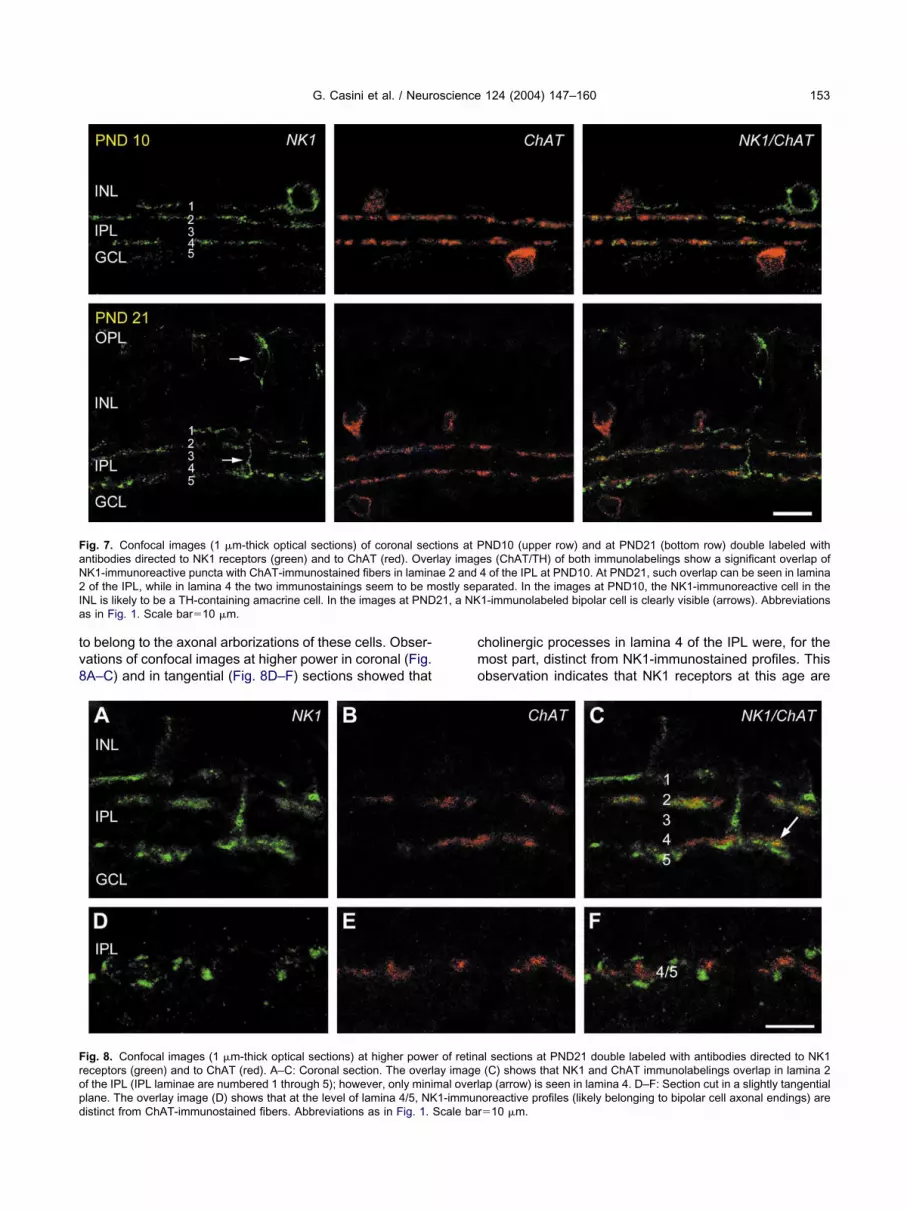
abeling was present at PND21.In NK1/ChAT double-labeled sections at PND10, there
as a significant overlap between NK1- and ChAT-immu-ostained fibers in laminae 2 and 4 of the IPL (Fig. 7),
ndicating that NK1 receptors are still expressed by cho-inergic amacrine cells at this age. At PND21, a similarverlap was only observed in lamina 2 of the IPL, while inhe IPL near the GCL cholinergic fibers appeared to beistributed slightly distal to NK1-immunolabeled pro-esses. Indeed, at this age, the NK1-expressing bipolarells are well developed (see also Fig. 2E) and the NK1-
mmunolabeled fibers in the IPL near the GCL were likely
ND10 (upper row) and at PND21 (bottom row) double labeled withNK1/TH) of both immunolabelings show that TH-containing amacrineane of their somata and in their processes in lamina 1 of the IPL.the images at PND21, NK1 immunolabeling can be seen in the distal
tions at Pimages (
membrIPL. In
Figs. 2 D and 7). Abbreviations as in Fig. 1. Scale bar�10 �m.
tv8
cmo
FaN2Ia
Fropd
G. Casini et al. / Neuroscience 124 (2004) 147–160 153
o belong to the axonal arborizations of these cells. Obser-ations of confocal images at higher power in coronal (Fig.A–C) and in tangential (Fig. 8D–F) sections showed that
ig. 7. Confocal images (1 �m-thick optical sections) of coronal secntibodies directed to NK1 receptors (green) and to ChAT (red). OveK1-immunoreactive puncta with ChAT-immunostained fibers in laminof the IPL, while in lamina 4 the two immunostainings seem to be m
NL is likely to be a TH-containing amacrine cell. In the images at PNDs in Fig. 1. Scale bar�10 �m.
ig. 8. Confocal images (1 �m-thick optical sections) at higher poweeceptors (green) and to ChAT (red). A–C: Coronal section. The overlf the IPL (IPL laminae are numbered 1 through 5); however, only minilane. The overlay image (D) shows that at the level of lamina 4/5, NK
istinct from ChAT-immunostained fibers. Abbreviations as in Fig. 1. Scale barholinergic processes in lamina 4 of the IPL were, for theost part, distinct from NK1-immunostained profiles. Thisbservation indicates that NK1 receptors at this age are
ND10 (upper row) and at PND21 (bottom row) double labeled withes (ChAT/TH) of both immunolabelings show a significant overlap of4 of the IPL at PND10. At PND21, such overlap can be seen in laminaarated. In the images at PND10, the NK1-immunoreactive cell in the1-immunolabeled bipolar cell is clearly visible (arrows). Abbreviations
l sections at PND21 double labeled with antibodies directed to NK1(C) shows that NK1 and ChAT immunolabelings overlap in lamina 2
ap (arrow) is seen in lamina 4. D–F: Section cut in a slightly tangentialoreactive profiles (likely belonging to bipolar cell axonal endings) are
tions at Prlay imagae 2 andostly sep21, a NK
r of retinaay imagemal overl1-immun
�10 �m.

mItt
roipaaes
D
Tmmodc5imli3r
irtdscmdiorlhrtebooGe
oiao2sntsdaPaPoob
Cc
Tcfor
Faiaa
T
PPPP
a
*
G. Casini et al. / Neuroscience 124 (2004) 147–160154
ostly absent in ChAT-containing fibers of lamina 4 of thePL and that NK1 receptor immunolabeling in this region ofhe IPL is mainly associated with bipolar cell axonalerminals.
At PND35 (Fig. 9), when the mature pattern of NK1eceptor expression is observed (see Fig. 1), there was noverlap between NK1 receptor and ChAT immunostain-
ngs. Together, these findings show that, in the secondhase of NK1 receptor developmental expression, thedult features of the NK1 receptor pattern are establishedround the time of eye opening. However, NK1 receptorxpression in cholinergic amacrine cells persists for a con-iderable period of postnatal life.
A assay
o determine whether NK1 receptors expressed by dopa-inergic amacrine cells mediate SP influences on dopa-inergic neurotransmission in the retina, the release levelsf DA and of its metabolites were measured in retinas atifferent developmental ages and in different experimentalonditions. In retinas at PND35, a 15 min exposure to0 mM KCl, led to significant increase of DA levels in the
ncubation medium, showing that DA is released by dopa-inergic amacrine cells upon stimulation. Basal (control)
evels of extracellular DA, DOPAC and HVA at the exam-ned developmental ages are reported in Table 1. At PND5, following bath addition of SP, a significant increase of
ig. 9. Confocal overlay image (1 �m-thick optical sections) of NK1nd ChAT immunostainings in a coronal section at PND35. The NK1
mmunostaining pattern (green) is fully mature (compare with Fig. 1)nd clearly distinct from ChAT-immunolabeled profiles (red). Abbrevi-tions as in Fig. 1. Scale bar�10 �m.
eleased DA was observed with 10 �M SP (Fig. 10A), P
ndicating that the peptide is effective in stimulating DAelease. The low level of statistical significance (P�0.05),ogether with the lack of significant effects with the otheroses of SP, indicated that the viability of the peptide ishort in these experimental conditions. Consistently,learer effects were obtained in experiments using theore stable NK1 receptor-specific agonist GR73632. At alloses tested, ranging from 0.01 to 100 �M, significant
ncreases of DA release around 150% of basal levels werebserved (Fig. 10B). In Fig. 10C, the effects of the NK1eceptor-specific antagonist GR82334 on basal DA re-ease are summarized. At all doses tested, except theighest dose of 100 �M, bath additions of this antagonisteduced DA release to undetectable levels, suggestinghat basal DA release is sustained in the retina by endog-nously released SP. The increase in DA release inducedy SP or GR73632 was accompanied by increased levelsf DOPAC (Fig. 10, inset), indicating increased DA metab-lism. Similarly, reduction of DA release induced byR82334 was accompanied by reduction of DOPAC lev-ls, which fell under detection limit (data not shown).
Basal levels of DA release varied during retinal devel-pment. In particular, extracellular DA and DOPAC in ret-
nas at PND0 were under detection limit, which in ourssay corresponds to 50 fmol (with a signal to noise ratiof 5). In contrast, HVA levels ranging from 0.57 to.04 ng/ml were measured, indicating that DA is synthe-ized and some DA metabolism is present in PND0 reti-as. At PND10 and PND21, basal levels of DA were higherhan at PND35, while metabolite control levels were ob-erved to increase from PND10 to PND35 (Table 1). Ineveloping retinas, SP, GR73632 and GR82334 did notppear to affect DA release. In particular, in retinas atND0, DA levels remained under detection limit after drugpplication, and, as shown in Fig. 11, at PND10 andND21 extracellular DA levels were not affected by 0.1 �Mr by 1.0 �M SP, GR73632 or GR82334. Similarly, levelsf DA metabolites did not significantly change followingath addition of pharmacological agents.
alcium imaging on developing cholinergic amacrineells
o determine whether the transiently expressed NK1 re-eptors in the developing cholinergic amacrine cells play aunctional role, Ca2� imaging experiments were performedn morphologically identified starburst cells in E30–PND1abbit retinal flat-mounts. Fig. 12 shows an example of a
able 1. Control levels of DA and its metabolites (ng/ml, meanS.D.)a
DA DOPAC HVA
ND0 — — 1.000.59***ND10 1.480.32*** 0.460.12*** 1.950.07**ND21 1.290.34*** 1.050.30* 2.010.53**ND35 0.610.10 1.720.30 4.510.53
Significantly different from PND35: * P�0.001, ** P�0.0005,** P�0.0001.
ND0 displaced starburst cell loaded with OGB-1. Most

dpi1nitr
cstf�AbGttbcsstitcrcdwarCss
1nistsbt
ntilrliTetc
Stmcn1a11tci
Fav**
G. Casini et al. / Neuroscience 124 (2004) 147–160 155
ye-filled starburst cells were functionally active and dis-layed spontaneous Ca2� transients (see Fig. 14), indicat-
ng the presence of spontaneous retinal waves (Zhou,998). Bath application of SP (1 �M) produced a pro-ounced increase in intracellular free Ca2� concentration
n starburst cells (Fig. 13A). This effect was found in all ofhe starburst cells (n�4) tested in three E30–PND0etinas.
In order to prove that the main responses of starburstells to SP were mediated by a direct SP action on thetarburst cells under imaging and not by indirect neuro-ransmitter actions from other cells, the effect of SP wasurther tested in the presence of a cocktail containing (inM) 100 hexamethonium, 1 atropine, 25 CNQX, 100 D-P7, 100 picrotoxin, 2 CGP55845, and 4 strychnine tolock nicotinic, muscarinic, non-NMDA, NMDA, GABAA/C,ABAB, and glycine receptors, respectively. The applica-
ion of this cocktail reduced the baseline Ca2� concentra-ion in starburst cells (Fig. 13B), as expected from thelockade of endogenous neurotransmitter inputs to theells, and completely blocked the spontaneous Ca2� tran-ients, which are known to rely on nicotinic neurotransmis-ion at this age (Zhou and Zhao, 2000). However, even inhe presence of this cocktail, SP (1 �M) remained effectiven evoking a robust Ca2� rise in starburst cells, suggestinghe presence of functional NK1 receptors directly on theseells (Fig. 13B). The effect of SP was mimicked by the NK1eceptor-specific agonist GR73632 (1 �M; Fig. 13C), indi-ating that the response of starburst cells to SP was me-iated by the activation of NK1 receptors. This conclusionas confirmed by the action of the NK1 receptor-specificntagonist GR82334. As shown in Fig. 14A and B, the NK1eceptor agonist GR73632 (1 �M) increased the baselinea2� level in a PND0 starburst cell (the same cell ashown in Fig. 12), together with an enhancement of the
2�
ig. 10. DA levels in the bath solution of retinas at PND35 treated wgonist GR73632 (B), or the NK1 receptor antagonist GR82334 (C). Talues); the dashed lines represent 100% of control values. Statisti** P�0.001. Inset: DOPAC levels in the bath solution of retinas tre* P�0.005.
pontaneous Ca transients (upward deflections in Fig. l
4A, B). Application of the antagonist GR82334 (0.5 �M)early completely and reversibly blocked the GR73632-
nduced increases of both the baseline Ca2� level and thepontaneous Ca2� transients (Fig. 14A, B). The applica-ion of GR82334, however, did not completely block thepontaneous transients, indicating that the SP action maye modulatory but not obligatory to the spontaneousransients.
Fig. 14C summarizes the effects of NK1 receptor ago-ists and antagonist on the intracellular Ca2� concentra-
ion of E30–PND1 starburst cells. SP (1 �M) significantlyncreased the fluorescence intensity at the soma of CG-1-oaded starburst cells, an effect mimicked by the NK1eceptor agonist GR73632 (1 �M) at the soma of OGB-1-oaded starburst cells. The effect of GR73632 was abol-shed by the NK1 receptor antagonist GR82334 (1 �M).hus, displaced starburst cells in the perinatal rabbit retinaxpressed functional NK1 receptors, and the activation ofhese receptors resulted in a significant increase in intra-ellular free Ca2�.
DISCUSSION
tudies of neuropeptides in developing retinas indicatehat the expression of these molecules is related to theaturation of retinal circuitry. Neuropeptides and their re-
eptors are generally expressed in the retina at late pre-atal or early postnatal ages (Ferriero and Sagar, 1987,989; Mitrofanis et al., 1989; Ferriero et al., 1990; Zhang etl., 1990; White and Chalupa, 1992; Casini et al., 1994,997a, 2000; Hutsler and Chalupa, 1995; Oyamada et al.,999; Skoglosa et al., 1999; Fontanesi et al., 2000), beforehe onset of visual information processing, when photore-eptor outer segments are still immature, synaptogenesis
s largely incomplete (McArdle et al., 1977) and before
nt doses of the NK1 receptor native ligand SP (A), the NK1 receptorrams represent DA values relative to controls (see Table 1 for controlcance was assessed with Student’s t-test. * P�0.05; ** P�0.005;different doses of the NK1 receptor agonist GR73632. * P�0.05;
ith differehe histogcal signifiated with
ight-evoked responses can be recorded in bipolar cells

(hrtrCaitoefr
rvsamif
S
Bp
FGd
G. Casini et al. / Neuroscience 124 (2004) 147–160156
Dacheux and Miller, 1981). In addition, transient changesave been observed in the expression patterns of neu-opeptides. For instance, a transient expression of soma-ostatin has been reported in ganglion cells of developingat and cat retinas (Mitrofanis et al., 1989; White andhalupa, 1992; Fontanesi et al., 1997; Xiang et al., 2001),nd both the levels of neuropeptide Y measured by radio-
mmunoassay and the number of cells expressing vasoac-ive intestinal peptide mRNA transiently increase near eyepening in the rat retina (Ferriero and Sagar, 1989; Casinit al., 1994). These observations suggest that there areunctional pathways in the developing retina where neu-
ig. 11. DA levels in the bath solution of retinas at PND10 (A) and atR73632 or the NK1 receptor antagonist GR82334. The histogramsashed lines represent 100% of control values. Statistically significan
opeptides mediate interactions that are not functionally c
elated to visual information processing but, likely, to de-elopmental events. In the present paper, we demon-trated that both the expression pattern of NK1 receptorsnd the functional actions of SP undergo substantial re-odeling during retinal development, indicating that in the
mmature retina SP plays a functional role that is differentrom that in the adult retina.
P and NK1 receptors in developing retinas
oth in rat and in rabbit retinas, SP-containing cells areresent at birth, and their morphological development is
) treated with 0.1 �M or with 1.0 �M of SP, the NK1 receptor agonistt DA values relative to controls (see Table 1 for control values); theces from control values were not observed in these experiments.
PND21 (Brepresent differen
ompleted by the time of eye opening (Sakiyama et al.,

1srN(Stdst2Ntrcrat
snsNctSpatcNabfssi
mrr
dtmtitarPpbc2os
S
Oaatrl2cbeTir
toatcGDtiauDo(emenct1tm
Faowl(
G. Casini et al. / Neuroscience 124 (2004) 147–160 157
984; Zhang and Yeh, 1992; Casini et al., 1997a, 2000),uggesting that SP may be released in the developingetina and it may interact with NK1 receptors. Consistently,K1 receptors are present at birth both in the rabbit
present paper) and in the rat (Casini et al., 2000) retina.ome features of the developmental pattern of NK1 recep-
ors in rat retinas, including a rearrangement of the laminaristribution of NK1-immunolabeled processes in the IPL,uggested that SP could mediate different interactions inhe developing and in the mature retina (Casini et al.,000). Our findings on the developmental expression ofK1 receptors in rabbit retinas clearly demonstrate that the
argets of SP actions are different in different phases ofetinal development. In particular, SP is likely to act onholinergic amacrine cells during the early postnatal pe-iod, while it may affect a population of cone bipolar cellsnd the dopaminergic amacrine cells at later developmen-al ages.
At PND0 and PND6, we observed that the somata ofeveral ChAT-containing cells were devoid of NK1 immu-ostaining. This finding may indicate that only a subset oftarburst cells express NK1 receptors or that, alternatively,K1 receptors are preferentially expressed on starburstell processes. Since in the Ca2� imaging experiments allhe randomly chosen starburst cells were responsive toP, we conclude that NK1 receptors are likely to be ex-ressed by all cholinergic amacrine cells, presumably withpredominance on their processes. We also observed that
he NK1 and ChAT immunostainings in the IPL did notompletely overlap. Although this result might indicate thatK1 receptors are also expressed by noncholinergic am-crine cells, such a possibility can be reasonably excludedecause all NK1-immunolabled somata were invariablyound to contain ChAT immunostaining. Instead, it is pos-ible that NK1 and ChAT immunostainings may be in theame processes, but the punctate appearance of both
ig. 12. Fluorescence image of a displaced starburst amacrine cell inPND0 flat-mount retina. The cell was loaded with OGB-1 by means
f ballistic delivery of indicator-coated particles. The image was takenith a cooled CCD camera under a 40� water immersion objective
ens. The white spots on the right are the out-of-focus somata of othernon starburst) dye-loaded cells. Scale bar�50 �m.
mmunostainings may prevent a complete overlap. In sum- i
ary, our findings are consistent with expression of NK1eceptors limited to all starburst cells in the early postnataletina.
The presence of NK1 receptors in bipolar cells and inopaminergic amacrine cells is first detected around theime of eye opening, indicating that the appearance ofature characteristics in the NK1 receptor pattern is func-
ionally related to the onset of visual information process-ng. On the other hand, the downregulation of NK1 recep-or expression in cholinergic cells gradually proceeds overn extended period after eye opening. In particular, NK1eceptor expression is first lost, between PND10 andND21, in the starburst cell population of the GCL, whoserocesses arborize in lamina 4 of the IPL. Subsequently,y PND35, it also disappears from the cholinergic ama-rine cells of the INL, whose processes arborize in laminaof the IPL. At present, our data do not allow hypotheses
n the functional status or significance of NK1 receptors intarburst cells at late developmental ages.
P, NK1 receptors and dopaminergic amacrine cells
ur observations demonstrate that, in the adult retina, SPcting at NK1 receptors not only induces DA release, butlso sustains basal DA levels. These findings are consis-ent with the hypothesis that SP may contribute to theegulation of DA levels in the retina, which, in turn, areikely to be involved in light adaptation (see Casini et al.,002a for discussion). The increase in released DA isoncomitant to an increase in DA metabolism, as indicatedy higher levels of DOPAC, which is commonly used tovaluate monoamine oxidase activity (Colzi et al., 1993).ogether, the data indicate SP as a factor of primary
mportance to modulate dopaminergic function in the adultetina.
In newborn retinas, SP or GR73632 was not expectedo have a direct effect on DA release, due to the absencef NK1 receptors in TH-containing amacrine cells at thisge. In fact, DA release in these retinas was below detec-ion limit and no changes were observed after drug appli-ation. At PND10 and at PND21, SP, GR73632 orR82334 was still unable to affect the levels of releasedA. This finding was unexpected and apparently in con-
radiction with the observed presence of NK1 receptormmunostaining in TH-containing amacrine cells at theseges. A plausible explanation may be the incomplete mat-ration of the dopaminergic system. Indeed, extracellularA levels decrease from PND10 to PND35, while metab-lite levels progressively increase during the same periodsee Table 1), suggesting an ongoing maturation of thenzymatic systems for DA metabolism. A similar develop-ental pattern has been reported in the rabbit retina for thextracellular levels of glutamate, which are high in theewborn retina (Haberecht and Redburn, 1996) and de-rease after the second postnatal week in coincidence withhe onset of Muller cell metabolic activity (Redburn et al.,992). In addition, previous studies showed that an ex-ended period of postnatal life is required for completeorphological maturation of TH-containing amacrine cells
n the rabbit retina (Casini and Brecha, 1992a). Together,

tpsTdfmrb
S
Aocceibwc
ii1seprit1alrcett1pp(tlpttots
FrsaCCtasal
cideaaca
G. Casini et al. / Neuroscience 124 (2004) 147–160158
hese observations indicate that some morpho-functionalroperties of dopaminergic amacrine cells, including re-ponsiveness to SP, are established late in development.he possibility exists that NK1 receptors expressed byopaminergic amacrine cells at PND10 and PND21 areunctional; however, the high levels of extracellular DAay sustain feedback mechanisms inhibiting further DA
elease and therefore preventing the observation of possi-le SP effects.
P, NK1 receptors and cholinergic amacrine cells
n obvious question following the immunohistochemicalbservation of the presence of NK1 receptors in starburstells in the early postnatal retina was whether these re-eptors are functional and what type of influence may bexerted by SP on these cells. The results of the Ca2�
maging experiments demonstrate that developing star-urst cells transiently express functional NK1 receptors,hich play an important role in regulating the Ca2� con-entration in these cells during development.
Cholinergic neurotransmission plays a fundamental rolen the spontaneous activity that is characteristically observedn the developing retina (Zhou, 1998, 2001a; Feller et al.,996). Many studies in different vertebrate retinas havehown that this activity is organized in spontaneous waves ofxcitation originating at random points and spreading in un-redictable and different directions (Wong, 1999). These cor-elated bursts of spontaneous electric activity are likely to playmportant roles in the refinement of specific neuronal connec-ivity in both the central visual pathways (Shatz and Stryker,988; Wong and Oakley, 1996; Penn et al., 1998; Stellwagennd Shatz, 2002) and the retina itself (Bodnarenko and Cha-
upa, 1993; Sernagor et al., 2001). In the early postnatalabbit retina, glutamatergic, GABAergic, cholinergic and gly-inergic neurotransmissions have been implicated in the gen-ration and modulation of the retinal waves, and the interac-
ions and transitions among these classical transmitter sys-ems underlying retinal waves have been described (Wong,999; Feller, 2002; Zhou, 2001b). At the ages tested in theresent study (E30–PND1), spontaneous activity mainly de-ends on cholinergic transmission at nicotinic receptorsZhou and Zhao, 2000). Our findings show that in the imma-ure retina SP is likely to play an important role in the regu-ation of cholinergic cell physiological activity. In particular, SPromotes both an increase in the intracellular Ca2� concen-ration of starburst cells, likely resulting in enhanced ace-ylcholine release, and an enhancement of the spontane-us bursts of activity. Together, these observations allowhe hypothesis that SP actions on starburst cells may affectpontaneous activity in the retina, although SP does not
hanges shown in this figure were obtained with nonratiometric imag-ng, and we cannot quantitatively compare the numbers obtained inifferent experiments because i) dye loading and intensity was differ-nt (random) in different cells; ii) we used CG-1 for SP experiments,nd OGB-1 for NK1 receptor agonist/antagonist experiments; iii) therea surrounding the cell selected to measure the average fluores-ence intensity (region of interest) was adjusted in each experimentnd it was different in different cells.
ig. 13. Imaging from displaced starburst amacrine cells in perinatalabbit retinal flat-mounts. A: Changes in fluorescence intensity mea-ured from an E30 starburst cell loaded with CG-1 during a 10 min 30 spplication of 1 �M SP, showing a SP-evoked rise in intracellulara2�. B: SP (1 �M, duration of application 4 min 45 s) increased thea2� concentration in another E30 starburst cell loaded with CG-1 in
he presence of a cocktail containing (in �M): 100 hexamethonium, 1tropine, 25 CNQX, 100 D-AP7, 100 picrotoxin, 2 CGP55845 and 4trychnine. C: A short (3 min) application of the NK1 receptor-specificgonist GR73632 (1 �M) mimicked the effect of SP on an OGB-1-
oaded starburst cell in another E30 retina. Note that the fluorescence

sTb
Tdcotrrpctipoi
AItN
A
B
B
CC
C
C
C
C
C
FioBs42Gc rent from
G. Casini et al. / Neuroscience 124 (2004) 147–160 159
eem to be essential for the generation of the activity itself.he possibility of SP regulation of retinal waves remains toe investigated in a separate study.
CONCLUSIONS
he present study shows that SP physiological roles un-ergo substantial changes during retinal development. Thehanges in functional actions of SP are mirrored by devel-pmentally regulated modifications of the expression pat-ern of NK1 receptors. In particular, SP is involved inegulation of dopaminergic neurotransmission in the adultetina, while it modulates cholinergic amacrine cells in theerinatal retina. The action exerted by SP on starburstells in the immature retina may contribute to the regula-ion of the retinal waves of spontaneous activity, thusnfluencing the development of the retina and retinofugalathways. If in the adult retina SP is involved in modulationf mechanisms for visual information processing, in the
mmature retina it is likely to act as a developmental factor.
cknowledgements—This work was supported by grants from thetalian Board of Education (MM05242912 to G.C. and L.B.), fromhe European Community (QLG3-1999-00908 to P.B.) and from
ig. 14. A, B: Effects of the NK1 receptor-specific agonist and antagon A, a brief application of the NK1-specific agonist GR73632 (1 �M) pf spontaneous Ca2� transients. These effects were completely and rshows blown-up views of the responses to GR73632 and GR82334
oma of starburst cells loaded with CG-1 or OGB-1. SP (1 �M) in8%17% (meanS.D., n�4). In cells loaded with OGB-1, the NK0%9% (meanS.D., n�3) under the control condition and by 2.4R82334 (0.5 �M). It should be noted that, because of the differenomparison between effects of SP and GR73632. * Significantly diffe
IH (EY10894) and RPB, Inc to Z.J.Z.
REFERENCES
mes A, Nesbett FB (1981) In vitro retina as an experimental model ofthe central nervous system. J Neurochem 37:867–877.
agnoli P, Dal Monte M, Casini G (2003) Expression of neuropeptidesand their receptors in the developing retina of mammals. HistolHistopathol 18:1219–1242.
odnarenko SR, Chalupa LM (1993) Stratification of ON and OFFganglion cell dendrites depends on glutamate mediated afferentactivity in the developing retina. Nature 364:144–146.
ajal SR (1893) La retine des vertebres. La Cellule 9:17–257.asini G, Brecha NC (1992a) Postnatal development of tyrosine hy-
droxylase immunoreactive amacrine cells in the rabbit retina: I.Morphological characterization. J Comp Neurol 326:283–301.
asini G, Brecha NC (1992b) Postnatal development of tyrosine hy-droxylase immunoreactive amacrine cells in the rabbit retina: II.Quantitative analysis. J Comp Neurol 326:302–313.
asini G, Molnar M, Brecha NC (1994) Vasoactive intestinal polypep-tide mRNA in the rat retina: adult distribution and developmentalexpression. Neuroscience 58:657–667.
asini G, Trasarti L, Andolfi L, Bagnoli P (1997a) Morphologic matu-ration of tachykinin peptide-expressing cells in the postnatal rabbitretina. Dev Brain Res 99:131–141.
asini G, Rickman DW, Sternini C, Brecha NC (1997b) Neurokinin 1receptor expression in the rat retina. J Comp Neurol 389:496–507.
asini G, Brecha NC, Bosco L, Rickman DW (2000) Developmentalexpression of neurokinin-1 and neurokinin-3 receptors in the rat
he same OGB-1-loaded starburst cell as shown in Fig. 13. As shownn increase in the baseline Ca2� level, together with an enhancementblocked by the NK1 receptor-specific antagonist GR82334 (0.5 �M).
mary of relative changes in fluorescence intensity measured from thethe fluorescence intensity (�F/F) of CG-1-loaded starburst cells byr-specific agonist GR73632 (1 �M) increased the fluorescence by(meanS.D., n�2) in the presence of the NK1 receptor antagonistdicator dyes used, this graph is not intended to give a quantitativecontrol, P�0.05.
nist on troduced aeversibly. C: Sumcreased1 recepto%1.8%t Ca2� in
retina. J Comp Neurol 421:275–287.

C
C
C
C
D
D
F
F
F
F
F
F
F
F
F
G
H
H
K
K
MM
M
O
O
P
P
R
R
S
S
S
S
S
V
W
W
W
X
Z
Z
Z
Z
Z
Z
Z
G. Casini et al. / Neuroscience 124 (2004) 147–160160
asini G, Sabatini A, Catalani E, Willems D, Bosco L, Brecha NC(2002a) Expression of the neurokinin 1 receptor in the rabbit retina.Neuroscience 115:1309–1321.
asini, G, Petrucci, C, Willems, D, Bosco, L, Bagnoli, P (2002b)Developmental expression of neurokinin 1 (NK1) receptors in rabbitretina. Program No. 236.9. 2002 Abstract Viewer/Itinerary Planner.Washington, DC: Society for Neuroscience, 2002. CD receptorOM.
ellerino A, Pinzon-Duarte G, Carroll P, Kohler K (1998) Brain-derivedneurotrophic factor modulates the development of the dopaminer-gic network in the rodent retina. J Neurosci 18:3351–3362.
olzi A, D’Agostini F, Cesura AM, Borroni E, Da Prada M (1993)Monoamine oxidase-A inhibitors and dopamine metabolism in ratcaudatus: evidence that an increased cytosolic level of dopaminedisplaces reversible monoamine oxidase-A inhibitors in vivo.J Pharmacol Exp Ther 265:103–111.
acheux RF, Miller RF (1981) An intracellular electrophysiologicalstudy of the ontogeny of functional synapses in the rabbit retina: I.Horizontal and bipolar cells. J Comp Neurol 198:307–326.
ick E, Miller RF (1981) Peptides influence retinal ganglion cells.Neurosci Lett 26:131–135.
amiglietti EV Jr (1983) “Starburst” amacrine cells and cholinergicneurons: mirror-symmetric on and off amacrine cells of rabbit ret-ina. Brain Res 261:138–144.
eller MB, Wellis DP, Stellwagen D, Werblin FS, Shatz CJ (1996)Requirement for cholinergic synaptic transmission in the propaga-tion of spontaneous retinal waves. Science 272:1182–1187.
eller MB (2002) The role of nAChR-mediated spontaneous retinalactivity in visual system development. J Neurobiol 53:556–567.
erriero DM, Sagar SM (1987) Development of somatostatin immuno-reactive neurons in rat retina. Brain Res 431:207–214.
erriero DM, Sagar SM (1989) Development of neuropeptide Y-immu-noreactive neurons in the rat retina. Dev Brain Res 48:19–26.
erriero DM, Head VA, Edwards RH, Sagar SM (1990) SomatostatinmRNA and molecular forms during development of the rat retina.Dev Brain Res 57:15–19.
ontanesi G, Casini G, Thanos S, Bagnoli P (1997) Transient soma-tostatin-immunoreactive ganglion cells in the developing rat retina.Dev Brain Res 103:119–125.
ontanesi G, Gargini C, Bagnoli P (2000) Postnatal development ofsomatostatin 2A (sst2A) receptors expression in the rabbit retina.Dev Brain Res 123:67–80.
ornai F, Alessandrı MG, Torracca MT, Bassi L, Corsini GU (1997)Effects of noradrenergic lesions on MPTP/MPP1 kinetics andMPTP-induced nigrostriatal dopamine depletions. J Pharmacol ExpTher 283:100–107.
lickman RD, Adolph AR, Dowling JE (1982) Inner plexiform circuits inthe carp retina: effects of cholinergic agonists, GABA, and sub-stance P on the ganglion cells. Brain Res 234:81–99.
aberecht MF, Redburn DA (1996) High levels of extracellular gluta-mate are present in retina during neonatal development. Neuro-chem Res 21:285–291.
utsler JJ, Chalupa LM (1995) Development of neuropeptide Y immu-noreactive amacrine and ganglion cells in the pre- and postnatal catretina. J Comp Neurol 361:152–164.
ettunen P, Demas J, Lohmann C, Kasthuri N, Gong Y, Wong RO,Gan WB (2002) Imaging calcium dynamics in the nervous systemby means of ballistic delivery of indicators. J Neurosci Methods119:37–43.
ondoh A, Houtani T, Ueyama T, Baba K, Ikeda M, Yamagishi K, MikiH, Uyama M, Nakanishi S, Sugimoto T (1996) In situ hybridizationanalysis of substance P receptor in the rat retina. Exp Brain Res112:181–186.
aggio JE (1988) Tachykinins. Annu Rev Neurosci 11:13–28.cArdle CB, Dowling JE, Masland RH (1977) Development of outersegments and synapses in the rabbit retina. J Comp Neurol 175:
253–274.(Accepted 24 Octob
itrofanis J, Robinson SR, Provis JM (1989) Somatostatinergic neu-rones of the developing human and cat retinae. Neurosci Lett104:209–216.
tsuka M, Yoshioka K (1993) Neurotransmitter functions of mamma-lian tachykinins. Phys Rev 73:229–308.
yamada H, Takatsuji K, Senba E, Manthy PW, Tohyama M (1999)Postnatal development of NK1, NK2 and NK3 neurokinin receptorsexpression in the rat retina. Dev Brain Res 117:59–70.
enn AA, Riquelme PA, Feller MB, Shatz CJ (1998) Competition inretinogeniculate patterning driven by spontaneous activity. Science279:2108–2121.
rada F, Medina JI, Lopez-Gallardo R, Quesada A, Spira A, Prada C(1999) Spatiotemporal gradients of differentiation of chick retinatypes i and ii cholinergic cells: identification of a common postmi-totic cell population. J Comp Neurol 410:457–466.
edburn DA, Agarwal SH, Messersmith EK, Mitchell CK (1992) De-velopment of the glutamate system in rabbit retina. Neurochem Res17:61–66.
ohrer H, Acheson AL, Thibault J, Thoenen H (1986) Developmentalpotential of quail dorsal root ganglion cells analyzed in vitro and invivo. J Neurosci 6:2616–2624.
akiyama T, Kuwayama Y, Ishimoto I, Sasaoka A, Shiosaka S, To-hyama M, Manabe R, Shiotani Y (1984) Ontogeny of substanceP-containing structures in the ocular tissue of the rat: an immuno-histochemical analysis. Brain Res 315:275–281.
ernagor E, Eglen SJ, Wong RO (2001) Development of retinal gan-glion cell structure and function. Prog Ret Eye Res 20:139–174.
hatz CJ, Stryker MP (1988) Prenatal tetrodotoxin infusion blockssegregation of retinogeniculate afferents. Science 242:87–89.
koglosa Y, Takei N, Lindholm D (1999) Distribution of pituitary ade-nylate cyclase activating polypeptide mRNA in the developing ratbrain. Mol Brain Res 65:1–13.
tellwagen D, Shatz CJ (2002) An instructive role for retinal waves in thedevelopment of retinogeniculate connectivity. Neuron 33:357–367.
igna SR, Bowden JJ, McDonald DM, Fisher J, Okamoto A, McVeyDC, Payan DJ, Bunnett NW (1994) Characterization of antibodiesto the rat substance P (NK-1) receptor and to a chimeric substanceP receptor expressed in mammalian cells. J Neurosci 14:834–845.
hite CA, Chalupa LM (1992) Ontogeny of somatostatin immunore-activity in the cat retina. J Comp Neurol 317:129–144.
ong ROL (1999) Retinal waves and visual system development.Annu Rev Neurosci 22:29–47.
ong ROL, Oakley DM (1996) Changing patterns of spontaneousbursting activity of On and Off retinal ganglion cells during devel-opment. Neuron 16:1087–1095.
iang Z, Jiang L, Kang Z (2001) Transient expression of somatostatinmRNA in developing ganglion cell layers of rat retina. Dev BrainRes 128:25–33.
alutsky RA, Miller RF (1990) The physiology of substance P in therabbit retina. J Neurosci 10:394–402.
hang D, Yeh HH (1992) Substance-P-like immunoreactive amacrine cellsin the adult and the developing rat retina. Dev Brain Res 68:55–65.
hang DR, Gallagher M, Sladek CD, Yeh HH (1990) Postnatal devel-opment of corticotropin releasing factor-like immunoreactive ama-crine cells in the rat retina. Dev Brain Res 51:185–194.
hou ZJ (1998) Direct participation of starburst amacrine cells inspontaneous rhythmic activities in the developing mammalian ret-ina. J Neurosci 18:4155–4165.
hou ZJ (2001a) The function of the cholinergic system in the devel-oping mammalian retina. Prog Brain Res 131:599–613.
hou ZJ (2001b) A critical role of the strychnine-sensitive glycinergicsystem in spontaneous retinal waves of the developing rabbit.J Neurosci 21:5158–5168.
hou ZJ, Zhao D (2000) Coordinated transitions in neurotransmittersystems for the initiation and propagation of spontaneous retinal
waves. J Neurosci 20:6570–6577.er 2003)





























