Embed Size (px)
Citation preview

BIOLOGY OF REPRODUCTION 85, 1057–1065 (2011)Published online before print 3 August 2011.DOI 10.1095/biolreprod.111.092031
Neurons of the Lateral Preoptic Area/Rostral Anterior Hypothalamic Area AreRequired for Photoperiodic Inhibition of Estrous Cyclicity in Sheep1
Stanley M. Hileman,2,3 Christina J. McManus,3 Robert L. Goodman,3 and Heiko T. Jansen4
Department of Physiology and Pharmacology,3 West Virginia University, Morgantown, West VirginiaDepartment of Veterinary and Comparative Anatomy, Pharmacology, and Physiology,4 Washington State University,Pullman, Washington
ABSTRACT
Photoperiod determines the timing of reproductive activity inmany species, yet the neural pathways whereby day length istransduced to a signal influencing gonadotropin-releasinghormone (GnRH) release are not fully understood. Physicallesions of the lateral preoptic area (lPOA)/rostral anteriorhypothalamic area (rAHA) in female sheep extend the periodof estrous cyclicity during inhibitory photoperiods. In thepresent study we sought to determine whether destroying onlyneurons and not fibers of passage in this area would lead tosimilar resistance to photosuppression. Additionally, neuraltract-tracing was used to map connectivity between the lPOA/rAHA and other hypothalamic areas implicated in photoperiodicregulation of reproduction. Progesterone secretion was moni-tored in six sheep to determine estrous cycles for 90 days duringa short-day (permissive) photoperiod. Three sheep then receivedbilateral injections of the excitotoxic glutamate analog, n-methyl-aspartic acid, directed toward the lPOA/rAHA, whereasthree others served as controls. All were then exposed to a long-day (suppressive) photoperiod for 120 days. Control sheepceased cycling at 40 6 10 days (mean 6 SEM), whereas lesionedsheep continued cycling through the end of the study. Theresults of the tract-tracing study revealed both afferent andefferent projections to the medial POA, retrochiasmatic area,arcuate nucleus, and premammillary region. Furthermore, closeproximal associations with GnRH neurons from efferentprojections were observed. We conclude that neurons locatedwithin the lPOA/rAHA are important for timing cessation ofestrous cycles during photosuppression and that this areacommunicates directly with GnRH neurons and other hypotha-lamic areas involved in the photoperiodic regulation ofreproduction.
estrous cycle, neuroendocrinology, ovine/sheep, photoperiod,seasonal reproduction
INTRODUCTION
Changes in day length (photoperiod) regulate gonadotropinsecretion in vertebrates as diverse as fish and primates so thatreproductive activity is limited to a specific season [1]. Inspecies like sheep that have autumnal breeding seasons, a shiftfrom a short day length to longer day length (e.g., from10L:14D to 16L:8D) decreases gonadotropin secretion andcauses ovarian cycles to cease. In contrast, a shift from long toshort days results in increased gonadotropin secretion and onsetof ovarian cycles [2]. Furthermore, if animals are maintainedon constant photoperiods for a prolonged period (e.g., 150days), they undergo a spontaneous reversal in gonadotropinsecretion and reproductive activity, a phenomenon known asphotorefractoriness [3]. Finally, these photoperiod-inducedeffects interact with an endogenous circannual oscillator todetermine the timing and duration of the breeding season [4].
Day length is converted into a neural signal in the brain viathe pineal gland hormone, melatonin. Melatonin is releasednocturnally [2], thus duration of melatonin release is longer inshort days (long nights) than in long days (short nights). Thephotoperiod (melatonin) response is reflected in a change inresponsiveness to estradiol-negative feedback that in turn altersrelease of gonadotropin-releasing hormone (GnRH), and thusluteinizing hormone (LH). Only fragmentary informationcurrently exists about the anatomical sites of action formelatonin and which neurochemical systems it uses to alterGnRH secretion, although there does not appear to be a directeffect on the GnRH neuron [5].
A potential site through which melatonin may act to mediatethe effects of photoperiod (in particular, photosuppression) isthe lateral preoptic area (lPOA)/rostral anterior hypothalamicarea (rAHA). Melatonin binding has been demonstrated in thisregion [6], and lesions of the AHA block the inhibitory effectsof melatonin on blastocyst implantation in the skunk [7].Furthermore, lesions of the AHA render hamsters unresponsiveto short-day photoperiods, which are inhibitory in this species[8]. In sheep, we previously observed that radiofrequencylesions of the lPOA/rAHA inhibited the ability of a long-dayphotoperiod to suppress reproduction [9]. A limitation of thisprevious work was that the electrolytic lesions could haveablated not only neurons within the lPOA/rAHA, but neuralfibers of passage as well. To circumvent this issue, the currentstudy used n-methyl-aspartic acid (NMA) to induce excitotoxiclesions [10–12] and test the hypothesis that cells within thelPOA/rAHA are functionally important for control of seasonalbreeding. Once these lesions were confirmed to be effective,we then performed a second study to test the hypothesis thatthe lPOA/rAHA is anatomically linked with other brain regionsimplicated in the photoperiodic control of reproduction, inparticular the medial POA (mPOA), suprachiasmatic nucleus(SCN), retrochiasmatic area (RCH), arcuate nucleus (ARC),and premammillary region (PMR).
1Supported by US Department of Agriculture grants 01-10835 toS.M.H. and 05-15848 to H.T.J. Image analyses were performed in theWest Virginia University Imaging Facility, which is supported in part bythe Mary Babb Randolph Cancer Center and National Institutes ofHealth grant P20 RR016440.2Correspondence: Stanley M. Hileman, Department of Physiology andPharmacology, West Virginia University, P.O. Box 9229, 3047 HealthSciences North, Morgantown, WV 26506. FAX: 304 293 3850;e-mail: [email protected]
Received: 28 February 2011.First decision: 27 March 2011.Accepted: 13 July 2011.� 2011 by the Society for the Study of Reproduction, Inc.eISSN: 1529-7268 http://www.biolreprod.orgISSN: 0006-3363
1057
Dow
nloaded from w
ww
.biolreprod.org.
MATERIALS AND METHODS
All procedures described in the present study were performed with theapproval of the animal care and use committees at West Virginia University andWashington State University and were in accordance with the NationalInstitutes of Health’s ‘‘Guide for the Care and Use of Laboratory Animals.’’Before surgeries, animals were kept outdoors. Just prior to and after surgeries,animals were kept in either light-sealed photochambers (experiment 1) or inrooms with lighting adjusted to mimic natural day length (experiment 2).Temperature was maintained between 158C and 238C, and ewes weremaintained on alfalfa pellets supplemented with corn and had free access towater.
Experiment 1: Effect of Excitotoxic Lesions on EstrousCyclicity under an Inhibitory Long-Day Photoperiod
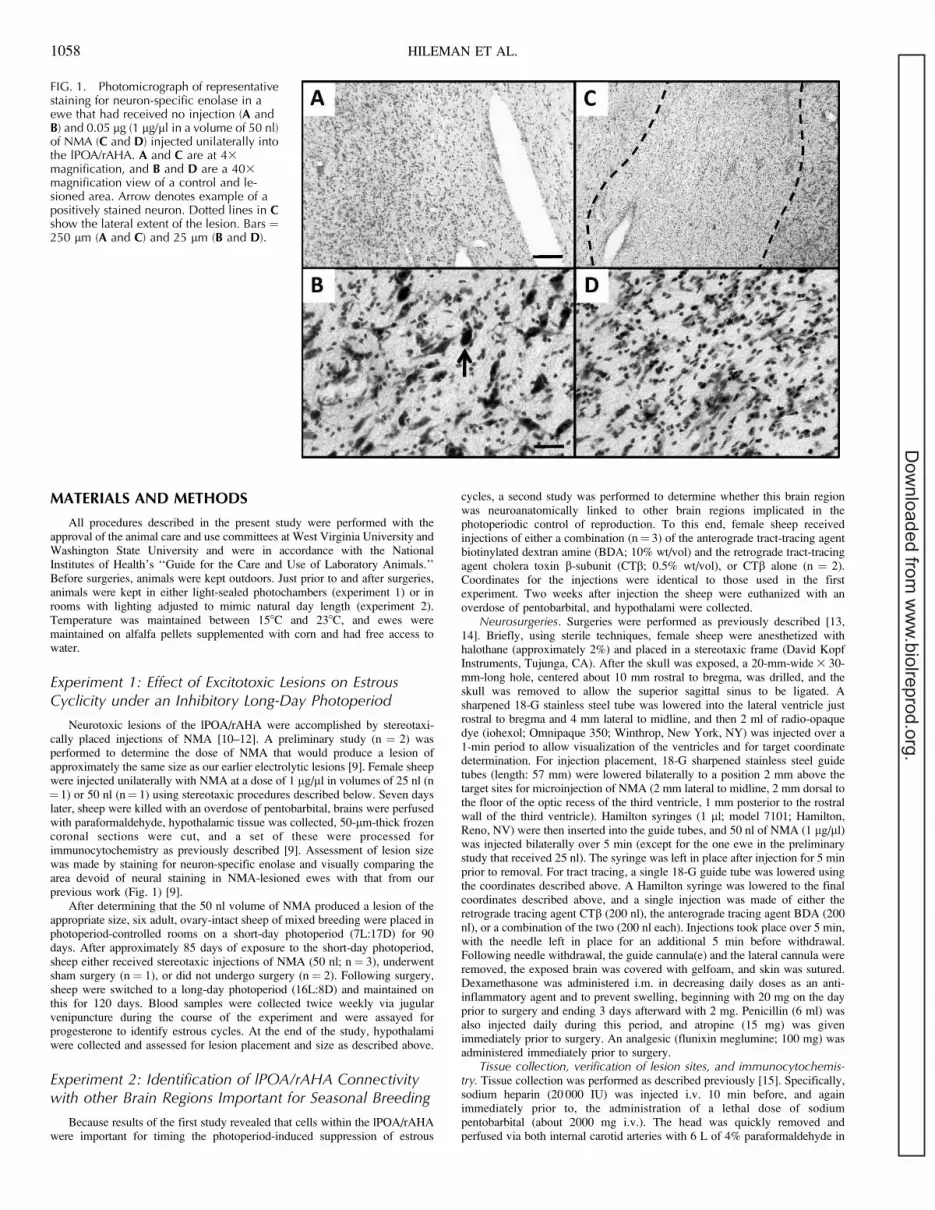
Neurotoxic lesions of the lPOA/rAHA were accomplished by stereotaxi-cally placed injections of NMA [10–12]. A preliminary study (n ¼ 2) wasperformed to determine the dose of NMA that would produce a lesion ofapproximately the same size as our earlier electrolytic lesions [9]. Female sheepwere injected unilaterally with NMA at a dose of 1 lg/ll in volumes of 25 nl (n¼ 1) or 50 nl (n¼ 1) using stereotaxic procedures described below. Seven dayslater, sheep were killed with an overdose of pentobarbital, brains were perfusedwith paraformaldehyde, hypothalamic tissue was collected, 50-lm-thick frozencoronal sections were cut, and a set of these were processed forimmunocytochemistry as previously described [9]. Assessment of lesion sizewas made by staining for neuron-specific enolase and visually comparing thearea devoid of neural staining in NMA-lesioned ewes with that from ourprevious work (Fig. 1) [9].
After determining that the 50 nl volume of NMA produced a lesion of theappropriate size, six adult, ovary-intact sheep of mixed breeding were placed inphotoperiod-controlled rooms on a short-day photoperiod (7L:17D) for 90days. After approximately 85 days of exposure to the short-day photoperiod,sheep either received stereotaxic injections of NMA (50 nl; n¼ 3), underwentsham surgery (n ¼ 1), or did not undergo surgery (n ¼ 2). Following surgery,sheep were switched to a long-day photoperiod (16L:8D) and maintained onthis for 120 days. Blood samples were collected twice weekly via jugularvenipuncture during the course of the experiment and were assayed forprogesterone to identify estrous cycles. At the end of the study, hypothalamiwere collected and assessed for lesion placement and size as described above.
Experiment 2: Identification of lPOA/rAHA Connectivitywith other Brain Regions Important for Seasonal Breeding
Because results of the first study revealed that cells within the lPOA/rAHAwere important for timing the photoperiod-induced suppression of estrous
cycles, a second study was performed to determine whether this brain regionwas neuroanatomically linked to other brain regions implicated in thephotoperiodic control of reproduction. To this end, female sheep receivedinjections of either a combination (n¼ 3) of the anterograde tract-tracing agentbiotinylated dextran amine (BDA; 10% wt/vol) and the retrograde tract-tracingagent cholera toxin b-subunit (CTb; 0.5% wt/vol), or CTb alone (n ¼ 2).Coordinates for the injections were identical to those used in the firstexperiment. Two weeks after injection the sheep were euthanized with anoverdose of pentobarbital, and hypothalami were collected.
Neurosurgeries. Surgeries were performed as previously described [13,14]. Briefly, using sterile techniques, female sheep were anesthetized withhalothane (approximately 2%) and placed in a stereotaxic frame (David KopfInstruments, Tujunga, CA). After the skull was exposed, a 20-mm-wide 3 30-mm-long hole, centered about 10 mm rostral to bregma, was drilled, and theskull was removed to allow the superior sagittal sinus to be ligated. Asharpened 18-G stainless steel tube was lowered into the lateral ventricle justrostral to bregma and 4 mm lateral to midline, and then 2 ml of radio-opaquedye (iohexol; Omnipaque 350; Winthrop, New York, NY) was injected over a1-min period to allow visualization of the ventricles and for target coordinatedetermination. For injection placement, 18-G sharpened stainless steel guidetubes (length: 57 mm) were lowered bilaterally to a position 2 mm above thetarget sites for microinjection of NMA (2 mm lateral to midline, 2 mm dorsal tothe floor of the optic recess of the third ventricle, 1 mm posterior to the rostralwall of the third ventricle). Hamilton syringes (1 ll; model 7101; Hamilton,Reno, NV) were then inserted into the guide tubes, and 50 nl of NMA (1 lg/ll)was injected bilaterally over 5 min (except for the one ewe in the preliminarystudy that received 25 nl). The syringe was left in place after injection for 5 minprior to removal. For tract tracing, a single 18-G guide tube was lowered usingthe coordinates described above. A Hamilton syringe was lowered to the finalcoordinates described above, and a single injection was made of either theretrograde tracing agent CTb (200 nl), the anterograde tracing agent BDA (200nl), or a combination of the two (200 nl each). Injections took place over 5 min,with the needle left in place for an additional 5 min before withdrawal.Following needle withdrawal, the guide cannula(e) and the lateral cannula wereremoved, the exposed brain was covered with gelfoam, and skin was sutured.Dexamethasone was administered i.m. in decreasing daily doses as an anti-inflammatory agent and to prevent swelling, beginning with 20 mg on the dayprior to surgery and ending 3 days afterward with 2 mg. Penicillin (6 ml) wasalso injected daily during this period, and atropine (15 mg) was givenimmediately prior to surgery. An analgesic (flunixin meglumine; 100 mg) wasadministered immediately prior to surgery.
Tissue collection, verification of lesion sites, and immunocytochemis-try. Tissue collection was performed as described previously [15]. Specifically,sodium heparin (20 000 IU) was injected i.v. 10 min before, and againimmediately prior to, the administration of a lethal dose of sodiumpentobarbital (about 2000 mg i.v.). The head was quickly removed andperfused via both internal carotid arteries with 6 L of 4% paraformaldehyde in
FIG. 1. Photomicrograph of representativestaining for neuron-specific enolase in aewe that had received no injection (A andB) and 0.05 lg (1 lg/ll in a volume of 50 nl)of NMA (C and D) injected unilaterally intothe lPOA/rAHA. A and C are at 43magnification, and B and D are a 403magnification view of a control and le-sioned area. Arrow denotes example of apositively stained neuron. Dotted lines in Cshow the lateral extent of the lesion. Bars ¼250 lm (A and C) and 25 lm (B and D).
1058 HILEMAN ET AL.
Dow
nloaded from w
ww
.biolreprod.org.

0.1 M PO4 buffer containing 1.0 IU/ml sodium heparin and 0.1% NaNO3 (avasodilator). The brains were removed, and tissue blocks containing thediencephalon were dissected out and stored at 48C in this fixative overnight,and then transferred to 0.1 M phosphate buffer containing 20% sucrose untilthey sank. Frozen coronal sections (50 lm thick) were cut on a freezingmicrotome into five series. Every fifth section was analyzed for neuron-specificenolase.
Immunocytochemical identification of neuron-specific enolase was per-formed on free-floating sections [16]. On Day 1, sections were washed 3 3 20min in 0.1 M PBS, followed by a 10-min incubation in 1% H2O2. Tissue wasthen washed 3 3 5 min in PBS, placed in 4% normal donkey serum (NDkS)with 0.1% Triton X for 1 h, and then placed overnight on a shaker table in anti-neuron-specific enolase antibody (MAB377; Millipore, Billerica, MA) diluted1:1000 in PBS with 4% NDkS and 0.1% Triton X. On Day 2, sections werewashed 3 3 5 min in PBS, followed by 1 h of incubation in 1:400 biotinylateddonkey anti-mouse immunoglobulin G (Vector Laboratories, Burlingame, CA)in 4% NDkS and 0.1% Triton X. Tissue was then washed 3 3 5 min in PBS,followed by a 1-h incubation in ABC diluted 1:400 (Vector Laboratories) inPBS. After 3 3 5 min washes in PBS, sections were incubated for 10 min innickel-enhanced diaminobenzidine (DAB) [17], washed again 3 3 5 min inPBS, mounted on slides, and dried overnight. On Day 3, sections were placedin acetate buffer for 30 sec, stained in 0.5% cresyl violet for 15 min, dehydratedthrough a series of alcohols, cleared in xylene, and then coverslipped usingPermount (Fisher Scientific, Pittsburgh, PA). This procedure produces a blue-black reaction product that is constrained to the nucleus. Lesion extent wasdetermined by identifying areas devoid of nuclear staining (Fig. 1) whenviewed at the light microscope level.
Biotinylated dextran amine and CTb were visualized as previouslydescribed [18]. For BDA, sections were washed 3 3 5 min in PBS, incubatedfor 10 min in 1% H2O2, washed again in PBS, and then incubated for 1 h inABC diluted 1:400 in PBS. After 3 3 5 min washes in PBS, sections wereincubated for 10 min in nickel-enhanced DAB. For CTb visualization on theBDA-labeled sections, tissues were then washed 3 3 5 min in PBS, followed byincubation for 1 h in PBS containing 0.4% Triton X and 4% normal goat serumfor 1 h. Tissues were then washed and incubated overnight at 48C on a shakertable in anti-rabbit CTb (C-3062; Sigma-Aldrich, St. Louis, MO) diluted1:15 000 in PBS containing 0.4% Triton X. The next day, sections were washedand then incubated for 1 h in goat anti-rabbit immunoglobulin G (1:200; VectorLaboratories). Tissue was then washed 3 3 5 min in PBS, followed by a 1-hincubation in ABC diluted 1:400 in PBS. After 3 3 5 min washes in PBS,sections were incubated for 10 min in DAB [17], washed again for 3 3 5 min inPBS, mounted on slides, and dried overnight. Tissues were then dehydratedthrough a series of alcohols, cleared in xylene, and coverslipped usingPermount. For mPOA tissue where GnRH was visualized in addition to BDA,tissues were incubated as described above except that mouse anti-GnRH (SMI-41; lot 3; Sternberger Monoclonals, Covance Inc., Princeton, NJ) was used asthe primary antibody at 1:3000.
Measurement of progesterone. Progesterone concentrations in plasmawere assessed by using a radioimmunoassay kit (Diagnostic Products Corp.,Los Angeles, CA). Sensitivity, as defined by 90% B/Bo, was 0.5 ng/ml. Allsamples were run in a single assay, with the intraassay coefficient of variationbeing 6.4%.
Statistics. Time to cessation of estrous cycle expression was defined to bethe time from the beginning of the long-day photoregimen to the first of fourconsecutive blood samples (i.e., a period of 2 wk) that contained less than 1 ng/ml progesterone. We originally had planned to compare time to cessation ofestrous cycles using a Student t-test. However, because none of the lesionedanimals became anestrous, there was no variability associated with thistreatment group. Thus, only means for each group are presented.
RESULTS
Experiment 1
Comparison of the control and NMA-injected side of thelPOA/rAHA in a female sheep injected with 50 nl of NMA canbe seen in Figure 1. Staining for neuron-specific enolaseshowed abundant neuronal nuclei in the uninjected side (Fig. 1,A and B). In contrast, NMA almost totally eliminated stainingin the area surrounding the injection site (Fig. 1, C and D). Thearea covered by the NMA-induced lesions within the lPOA/rAHA of each animal is shown in Figure 2. Both location andarea covered by these neurochemical lesions were very similarto those of our previous study [9]. Lesions were bilateral andconfined to the lPOA/rAHA.
Profiles of progesterone secretion for all animals are shownin Figure 3. All six sheep exhibited a normal pattern of estrouscycles during the short-day photoperiod. After placement in thelong-day photoperiod, nonsurgical controls and the singlesham-operated sheep showed only two complete estrouscycles, with a mean period to cycle cessation (anestrus) of 406 10 days. In contrast, NMA-injected ewes exhibited five toseven estrous cycles following the switch to the long-dayphotoperiod. Indeed, by our definition for cessation of estrouscycle expression, none of the three lesioned sheep had enteredanestrus by the end of the experiment.
Experiment 2
Schematics of the resultant CTb or BDA staining followingtract-tracer injections are shown in Figures 4 and 5,
FIG. 2. Schematic depicting the location of NMA-induced lesions.Lesioned areas, as determined by the absence of staining for neuron-specific enolase, are depicted by the black areas. AC, anteriorcommissure; FX, fornix; OC, optic chiasm; SON, supraoptic nucleus.
lPOA/rAHA AND SEASONAL BREEDING IN FEMALE SHEEP 1059
Dow
nloaded from w
ww
.biolreprod.org.

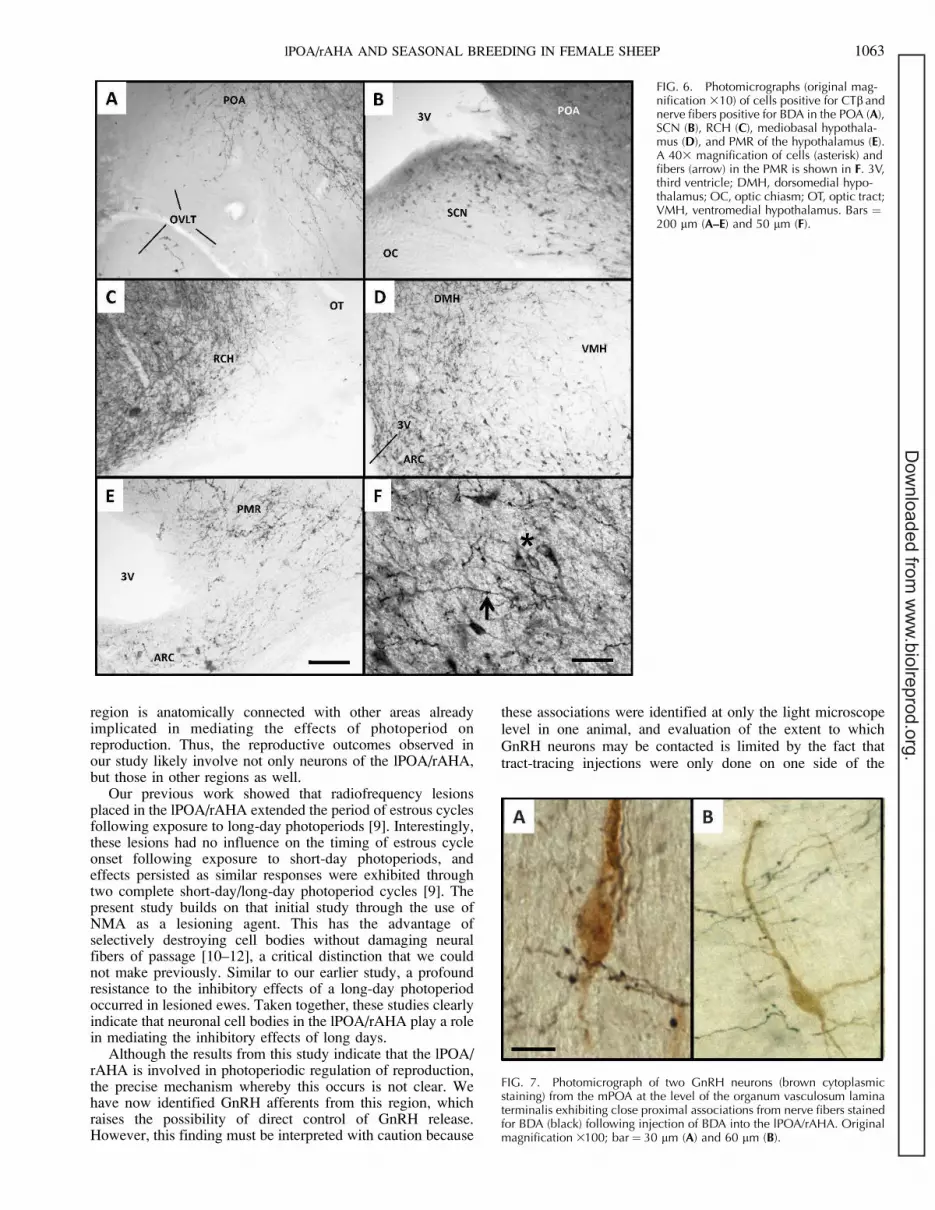
respectively. The injection site locations were very similar tothose of the excitotoxic lesions induced in experiment 1 and inour previous work. Importantly, injection of these amounts oftract-tracing agents resulted in an area of injection that wassimilar in scope to the area lesioned in this and our previousstudy. Cholera toxin b-subunit-labeled cells were foundthroughout the POA and the extent of the hypothalamus inan area generally bordered laterally by the fornix andmammillothalamic tracts. Injections were similarly placed inall three sheep, and a tracing from a representative sheep isshown in Figures 4 and 5. Retrogradely labeled cells (Fig. 4)were largely observed on the side that was ipsilateral to theinjection, with only scattered cells observed on the contralateralside. We observed CTb-labeled cells in several areas known tobe important to the photoperiodic regulation of reproduction.Numerous cells were observed in the mPOA, although theywere less abundant in the organum vasculosum lateralterminalis (OVLT; Fig. 6A). We also observed scattered cellsin the SCN (Fig. 6B) and the RCH just medial to the optictracts (Fig. 6C). There were also a large number of cells locatedwithin the mediobasal hypothalamus, namely the ARC,dorsomedial nucleus, and lateral hypothalamus, but relativelyfewer cells in the ventromedial nucleus (Fig. 6D). A highnumber of CTb-labeled cells were noted in the ventrolateralhypothalamus, and a relatively high density of cells was alsoobserved in the PMR (Fig. 6, E, higher magnification in F).There were also scattered cells extending laterally along thebase of the hypothalamus, and cells were also observed in thearea of the amygdala. Although not shown, the distribution ofcells in ewes receiving only CTb was virtually identical to thatof ewes receiving both CTb and BDA.
Examination of BDA staining (Fig. 5) revealed a pattern thatwas quite similar to that of CTb staining. A heavyconcentration of fibers was seen in the POA, with comparably
less in the region of the OVLT (Fig. 6A). A few fibers alsowere observed in the SCN (Fig. 6B). Fibers were evident in theRCH (Fig. 6C), and intermediate to heavy fiber densities wereobserved in the dorsomedial and lateral hypothalamus, withcomparatively fewer projections to the ARC and ventromedialnucleus (Fig. 6D). However, a heavy concentration of fiberswas evident in the ventrolateral hypothalamus. Intermediate toheavy projections were observed in the PMR (Fig. 6E). As withthe distribution of CTb-labeled cells, BDA fibers wereobserved extending along the base of the hypothalamus, withfibers clearly evident in the amygdala. Based on the highdensity of fibers in the POA, we also evaluated the possibilitythat BDA projections onto GnRH neurons were present. Twosections from one ewe containing the mPOA at the level of theOVLT were examined. As shown in Figure 7, close proximalassociations were observed between GnRH neurons and BDA-labeled fibers. Of 60 identified GnRH neurons, 27 (45%)appeared to have BDA-containing close proximal associations.
DISCUSSION
Control of seasonal reproduction by photoperiod involves acomplex interplay between various areas near or in thehypothalamus, such as the PMR, RCH, ARC, and POA, withhormones, such as estradiol, melatonin, and thyroid hormones[19–21]. The work presented in the present study supports thehypothesis that an additional area, the lPOA/rAHA, is animportant component of the circuitry of seasonal neuroendo-crine control. Specifically, we demonstrate that followingablation of cells within this region, female sheep exhibitresistance to the inhibitory effects of long-day photoperiods onovarian cycles. Furthermore, although our tract-tracing resultsraise the possibility of cells within the lPOA/rAHA directlyinfluencing GnRH neurons, they also clearly show that this
FIG. 3. Progesterone profiles for threeewes (Control) receiving either a shamsurgery (K54) or no surgery (K13, K46), andthree ewes receiving 0.05 lg (1 lg/ll in avolume of 50 nl) of NMA injected bilaterallyinto the lPOA/rAHA (NMA-lesioned). Eweswere exposed to 120 days of long-dayphotoperiod following surgery.
1060 HILEMAN ET AL.
Dow
nloaded from w
ww
.biolreprod.org.

FIG. 4. Tracings of cells positive for CTb insections from the POA through the hypo-thalamus of a ewe that had received aunilateral injection of the retrograde tracingagent CTb (200 nl) and the anterogradetracing agent BDA (200 nl). 3V, thirdventricle; AC, anterior commissure; AMG,amygdala; DMH, dorsomedial hypothala-mus; FX, fornix; INJ, injection site; MB,mammillary body; MT, mammillothalamictract; OC, optic chiasm; OT, optic tract; PD,pars distalis; PT, pars tuberalis; SON,supraoptic nucleus; ST, stria terminalis;VMH, ventromedial hypothalamus.
lPOA/rAHA AND SEASONAL BREEDING IN FEMALE SHEEP 1061
Dow
nloaded from w
ww
.biolreprod.org.

FIG. 5. Tracings of nerve fibers positive forBDA in sections from the POA through thehypothalamus of a ewe that had received aunilateral injection of the retrograde tracingagent CTb (200 nl) and the anterogradetracing agent BDA (200 nl). The sectionsshown here are from the same ewe as inFigure 4. Nerve fiber density: H, high; I,intermediate; L, light; VL, very light. AC,anterior commissure; AMG, amygdala;DMH, dorsomedial hypothalamus; FX, for-nix; INJ, injection site; MB, mammillarybody; MT, mammillothalamic tract; OC,optic chiasm; OT, optic tract; PD, parsdistalis; PT, pars tuberalis; SON, supraopticnucleus; ST, stria terminalis; 3V, thirdventricle; VMH, ventromedial hypothala-mus.
1062 HILEMAN ET AL.
Dow
nloaded from w
ww
.biolreprod.org.
region is anatomically connected with other areas alreadyimplicated in mediating the effects of photoperiod onreproduction. Thus, the reproductive outcomes observed inour study likely involve not only neurons of the lPOA/rAHA,but those in other regions as well.
Our previous work showed that radiofrequency lesionsplaced in the lPOA/rAHA extended the period of estrous cyclesfollowing exposure to long-day photoperiods [9]. Interestingly,these lesions had no influence on the timing of estrous cycleonset following exposure to short-day photoperiods, andeffects persisted as similar responses were exhibited throughtwo complete short-day/long-day photoperiod cycles [9]. Thepresent study builds on that initial study through the use ofNMA as a lesioning agent. This has the advantage ofselectively destroying cell bodies without damaging neuralfibers of passage [10–12], a critical distinction that we couldnot make previously. Similar to our earlier study, a profoundresistance to the inhibitory effects of a long-day photoperiodoccurred in lesioned ewes. Taken together, these studies clearlyindicate that neuronal cell bodies in the lPOA/rAHA play a rolein mediating the inhibitory effects of long days.
Although the results from this study indicate that the lPOA/rAHA is involved in photoperiodic regulation of reproduction,the precise mechanism whereby this occurs is not clear. Wehave now identified GnRH afferents from this region, whichraises the possibility of direct control of GnRH release.However, this finding must be interpreted with caution because
these associations were identified at only the light microscopelevel in one animal, and evaluation of the extent to whichGnRH neurons may be contacted is limited by the fact thattract-tracing injections were only done on one side of the
FIG. 6. Photomicrographs (original mag-nification 310) of cells positive for CTb andnerve fibers positive for BDA in the POA (A),SCN (B), RCH (C), mediobasal hypothala-mus (D), and PMR of the hypothalamus (E).A 403 magnification of cells (asterisk) andfibers (arrow) in the PMR is shown in F. 3V,third ventricle; DMH, dorsomedial hypo-thalamus; OC, optic chiasm; OT, optic tract;VMH, ventromedial hypothalamus. Bars ¼200 lm (A–E) and 50 lm (F).
FIG. 7. Photomicrograph of two GnRH neurons (brown cytoplasmicstaining) from the mPOA at the level of the organum vasculosum laminaterminalis exhibiting close proximal associations from nerve fibers stainedfor BDA (black) following injection of BDA into the lPOA/rAHA. Originalmagnification 3100; bar¼ 30 lm (A) and 60 lm (B).
lPOA/rAHA AND SEASONAL BREEDING IN FEMALE SHEEP 1063
Dow
nloaded from w
ww
.biolreprod.org.

lPOA/rAHA. Interestingly, cells in the lPOA/rAHA bindmelatonin [6] and express estrogen receptor-alpha (ESR1)[22, 23], raising the possibility that these hormones act locallyon cells within the lPOA/rAHA to influence GnRH release.Although we did not identify the phenotype of such cellswithin the lPOA/rAHA, neurons expressing tyrosine hydrox-ylase [24, 25] and dynorphin [26] in this area have beenreported in the sheep. The tyrosine hydroxylase-containingneurons are likely part of the A14 dopamine cell group, andlimited data in the sheep are consistent with a role for theseneurons in the photosuppression of LH secretion [25, 27]. Withregard to dynorphin, neurons expressing preprodynorphinmRNA are localized to the AHA, and GnRH neurons receivesynaptic input from dynorphin neurons [15]. PreprodynorphinmRNA expression in the AHA is decreased by ovariectomyand restored by progesterone treatment in ewes [28], and isincreased during a long-day photoperiod in rams by testoster-one [29]. An additional population of dynorphin neurons in theARC also plays a role in regulating GnRH/LH secretion [15,26], but it is not known whether they are involved inresponsiveness to long-day photoperiods.
As mentioned above, control of seasonal breeding undoubt-edly involves a complex interaction of a number of differentareas in or near the hypothalamus with the GnRH system [21,30]. Our tract-tracing results now reveal the connectivity of thelPOA/rAHA with several areas of known importance toseasonal breeding. Heavy innervations and high numbers oflabeled cells were found in the mPOA. This is not entirelysurprising because this region lies very close to the injectionsite; thus, some caution is required in interpreting this result. Insheep, this area contains the majority of GnRH cell bodies anda high number of ESR1-containing neurons [22–24, 31].Indeed, previous work showed that estradiol implants in theventral mPOA, an area just below the region of our lesions,suppressed LH in ovariectomized ewes during a long-dayphotoperiod [13]. Interestingly, melatonin implants placed inthis region, which should provide a short-day signal, did notreverse long-day photosuppression in ewes [32], suggestingthat this is not an area responsible for initiation of GnRH/LHsecretion in response to stimulatory photoperiods. However,this does not rule out a role for this area in melatonin-drivenchanges in GnRH/LH secretion during photosuppression.
Another area of note that received reciprocal input was theRCH. The RCH region contains the A15 group of dopamineneurons, and there is a large amount of evidence that theseneurons, located just medial to the optic tract, are involved inmediating steroid-induced suppression of GnRH secretionduring inhibitory photoperiods. Lesions of A15 dopamineneurons significantly compromise the ability of estradiol tosuppress LH secretion in ewes during the nonbreeding season,but not during the breeding season [27, 33]. Furthermore,ovariectomized ewes treated with estrogen exhibit an increasein the number of A15 dopamine cells expressing the immediateearly gene product FOS, a marker of neuronal stimulation, onlyduring the nonbreeding season [25]. Finally, estradiol implantsplaced directly into the RCH of the hypothalamus reduce LHpulse frequency during the nonbreeding season via ESR1, butnot ESR2 [14, 34]. Interestingly, A15 dopamine neurons do notexpress ESR1 [24, 35], so estradiol likely influences theseneurons indirectly. The identities of these indirect inputs arenot completely known but may include ESR1-containingneurons just dorsal to the A15 neurons or the ventral mPOA.
A large number of labeled cells and BDA-labeled fiberswere also observed within the mediobasal hypothalamus in theareas of the ARC and dorsomedial nucleus. The role of theARC in regulating GnRH/LH secretion recently has receivedattention because of a subset of neurons residing therein that
coexpress kisspeptin, neurokinin B, and dynorphin (termedKNDy neurons). KNDy neurons express ESR1 [36], andkisspeptin is a potent stimulator of GnRH/LH release in severalspecies [37–40]. Expression of kisspeptin is inhibited bygonadal steroids [36, 41] and is suppressed during an inhibitoryphotoperiod [42, 43]. These data support an important role forkisspeptin in the seasonal regulation of reproduction. Inaddition, we have recently shown that NKB stimulates LHrelease during the follicular phase of the estrous cycle [44] andin peripubertal female ewe lambs (Hileman and Goodman,unpublished results). With regard to the dorsomedial hypo-thalamus, cells in this area express the RFamide-relatedpeptide, gonadotropin-inhibiting hormone (GnIH) [42, 45].Although the role of GnIH in seasonal reproduction is notentirely clear for sheep [46], the number of GnIH-containingcells and the percentage of GnRH neurons exhibiting GnIH-containing close contacts are lower during the breeding seasonthan nonbreeding season [42], consistent with its role as aninhibitor of GnRH/LH secretion [46]. Thus, reciprocal contactsbetween these areas and the lPOA/rAHA could potentially beimportant for regulating the response to long-day photoperiods.
We observed dense reciprocal innervations of the PMR, anarea that appears to be very important in the actions of bothmelatonin and thyroxine (T4). T4 implants in the PMR wereeffective in inducing anestrus in ewes [47], and melatoninimplants in this site (presumably providing a short-day signal)during long days induced LH secretion [48]. Further, an area ofhigh melatonin binding in the caudal ARC/PMR [48]corresponds very well with an area receiving a high densityof BDA-labeled fibers in our study. Interestingly, the responsesobserved in our female sheep are somewhat similar to thoseobserved in the absence of thyroid hormones at the end of thebreeding season [20] (i.e., animals do not enter anestrus). Ourprevious study [9] showed that lesions of the lPOA/rAHA donot disrupt circulating T4 levels or melatonin secretorypatterns. Thus, input from the lPOA/rAHA may influence theresponse to T4 and/or melatonin, and therefore affect the timingof anestrus in response to long-day photoperiods.
The data presented in the present study clearly identify thelPOA/rAHA as playing a role in the photoperiodic control ofreproduction in the sheep. Neurotoxic lesions in this regionclearly interfered with the ability of a long-day photoperiod toinduce anestrus in ewes. Furthermore, our tract-tracing resultsunequivocally show that the lPOA/rAHA is intimatelyconnected with several regions of the hypothalamus that areinvolved in regulating seasonal reproduction. Additionalstudies will be required to identify the specific neural substratesinvolved and the functional relationship of this area with otherphotoperiod-sensitive regions of the hypothalamus.
REFERENCES
1. Gwinner E. Circannual Rhythms. New York: Springer-Verlag; 1986.2. Bittman EL, Karsch FJ, Hopkins JW. Role of the pineal gland in ovine
photoperiodism: regulation of seasonal breeding and negative feedbackeffects of estradiol upon luteinizing hormone secretion. Endocrinology1983; 113: 329–336.
3. Malpaux B, Moenter SM, Wayne NL, Woodfill CJ, Karsch FJ.Reproductive refractoriness of the ewe to inhibitory photoperiod is notcaused by alteration of the circadian secretion of melatonin. Neuroendo-crinology 1988; 48:264–270.
4. Robinson JE, Wayne NL, Karsch FJ. Refractoriness to inhibitory daylengths initiates the breeding season of the Suffolk ewe. Biol Reprod 1985;32:1024–1030.
5. Malpaux B, Viguie C, Skinner DC, Thiery JC, Chemineau P. Control ofthe circannual rhythm of reproduction by melatonin in the ewe. Brain ResBull 1997; 44:431–438.
6. Bittman EL, Weaver DR. The distribution of melatonin binding sites inneuroendocrine tissues of the ewe. Biol Reprod 1990; 43:986–993.
1064 HILEMAN ET AL.
Dow
nloaded from w
ww
.biolreprod.org.

7. Berria M, DeSantis M, Mead RA. Lesions to the anterior hypothalamusprevent the melatonin-induced lengthening of delayed implantation.Endocrinology 1989; 125:2897–2904.
8. Hastings MH, Roberts AC, Herbert J. Neurotoxic lesions of the anteriorhypothalamus disrupt the photoperiodic but not the circadian system of theSyrian hamster. Neuroendocrinology 1985; 40:316–324.
9. Hileman SM, Kuehl DE, Jackson GL. Effect of anterior hypothalamic arealesions on photoperiod-induced shifts in reproductive activity of the ewe.Endocrinology 1994; 135:1816–1823.
10. Hastings MH, Winn P, Dunnett SB. Neurotoxic amino acid lesions of thelateral hypothalamus: a parametric comparison of the effects of ibotenate,N-methyl-D,L-aspartate and quisqualate in the rat. Brain Res 1985;360:248–256.
11. Numan M, Corodimas KP, Numan MJ, Factor EM, Piers WD. Axon-sparing lesions of the preoptic region and substantia innominata disruptmaternal behavior in rats. Behav Neurosci 1988; 102:381–396.
12. Stewart GR, Price M, Olney JW, Hartman BK, Cozzari C. N-methylaspartate: an effective tool for lesioning basal forebrain cholinergicneurons of the rat. Brain Res 1986; 369:377–382.
13. Anderson GM, Connors JM, Hardy SL, Valent M, Goodman RL.Oestradiol microimplants in the ventromedial preoptic area inhibitsecretion of luteinizing hormone via dopamine neurones in anoestrousewes. J Neuroendocrinol 2001; 13:1051–1058.
14. Hardy SL, Anderson GM, Valent M, Connors JM, Goodman RL.Evidence that estrogen receptor alpha, but not beta, mediates seasonalchanges in the response of the ovine retrochiasmatic area to estradiol. BiolReprod 2003; 68:846–852.
15. Goodman RL, Coolen LM, Anderson GM, Hardy SL, Valent M, ConnorsJM, Fitzgerald ME, Lehman MN. Evidence that dynorphin plays a majorrole in mediating progesterone negative feedback on gonadotropin-releasing hormone neurons in sheep. Endocrinology 2004; 145:2959–2967.
16. Sliwowska JH, Billings HJ, Goodman RL, Coolen LM, Lehman MN. Thepremammillary hypothalamic area of the ewe: anatomical characterizationof a melatonin target area mediating seasonal reproduction. Biol Reprod2004; 70:1768–1775.
17. Adams JC. Heavy metal intensification of DAB-based HRP reactionproduct. J Histochem Cytochem 1981; 29:775.
18. Coolen LM, Jansen HT, Goodman RL, Wood RI, Lehman MN. A newmethod for simultaneous demonstration of anterograde and retrogradeconnections in the brain: co-injections of biotinylated dextran amine andthe beta subunit of cholera toxin. J Neurosci Methods 1999; 91:1–8.
19. Goodman RL, Jansen HT, Billings HJ, Coolen LM, Lehman MN. Neuralsystems mediating seasonal breeding in the ewe. J Neuroendocrinol 2010;22:674–681.
20. Karsch FJ, Dahl GE, Hachigian TM, Thrun LA. Involvement of thyroidhormones in seasonal reproduction. J Reprod Fertil Suppl 1995; 49:409–422.
21. Malpaux B, Tricoire H, Mailliet F, Daveau A, Migaud M, Skinner DC,Pelletier J, Chemineau P. Melatonin and seasonal reproduction: under-standing the neuroendocrine mechanisms using the sheep as a model.Reprod Suppl 2002; 59:167–179.
22. Blache D, Batailler M, Fabre-Nys C. Oestrogen receptors in the preoptico-hypothalamic continuum: immunohistochemical study of the distributionand cell density during induced oestrous cycle in ovariectomized ewe. JNeuroendocrinol 1994; 6:329–339.
23. Lehman MN, Ebling FJ, Moenter SM, Karsch FJ. Distribution of estrogenreceptor-immunoreactive cells in the sheep brain. Endocrinology 1993;133:876–886.
24. Lehman MN, Karsch FJ. Do gonadotropin-releasing hormone, tyrosinehydroxylase-, and beta-endorphin-immunoreactive neurons contain estro-gen receptors? A double-label immunocytochemical study in the Suffolkewe. Endocrinology 1993; 133:887–895.
25. Lehman MN, Durham DM, Jansen HT, Adrian B, Goodman RL.Dopaminergic A14/A15 neurons are activated during estradiol negativefeedback in anestrous, but not breeding season, ewes. Endocrinology1996; 137:4443–4450.
26. Foradori CD, Coolen LM, Fitzgerald ME, Skinner DC, Goodman RL,Lehman MN. Colocalization of progesterone receptors in parvicellulardynorphin neurons of the ovine preoptic area and hypothalamus.Endocrinology 2002; 143:4366–4374.
27. Havern RL, Whisnant CS, Goodman RL. Dopaminergic structures in theovine hypothalamus mediating estradiol negative feedback in anestrousewes. Endocrinology 1994; 134:1905–1914.
28. Foradori CD, Goodman RL, Adams VL, Valent M, Lehman MN.Progesterone increases dynorphin a concentrations in cerebrospinal fluid
and preprodynorphin messenger ribonucleic acid levels in a subset ofdynorphin neurons in the sheep. Endocrinology 2005; 146:1835–1842.
29. Scott CJ, Clarke IJ, Tilbrook AJ. The effect of testosterone and season onprodynorphin messenger RNA expression in the preoptic area-hypothal-amus of the ram. Domest Anim Endocrinol 2008; 34:440–450.
30. Lehman MN, Coolen LM, Goodman RL, Viguie C, Billings HJ, KarschFJ. Seasonal plasticity in the brain: the use of large animal models forneuroanatomical research. Reprod Suppl 2002; 59:149–165.
31. Lehman MN, Robinson JE, Karsch FJ, Silverman AJ. Immunocytochem-ical localization of luteinizing hormone-releasing hormone (LHRH)pathways in the sheep brain during anestrus and the mid-luteal phase ofthe estrous cycle. J Comp Neurol 1986; 244:19–35.
32. Malpaux B, Daveau A, Maurice F, Gayrard V, Thiery JC. Short-dayeffects of melatonin on luteinizing hormone secretion in the ewe: evidencefor central sites of action in the mediobasal hypothalamus. Biol Reprod1993; 48:752–760.
33. Thiery JC, Martin GB, Tillet Y, Caldani M, Quentin M, Jamain C, RavaultJP. Role of hypothalamic catecholamines in the regulation of luteinizinghormone and prolactin secretion in the ewe during seasonal anestrus.Neuroendocrinology 1989; 49:80–87.
34. Gallegos-Sanchez J, Delaleu B, Caraty A, Malpaux B, Thiery JC.Estradiol acts locally within the retrochiasmatic area to inhibit pulsatileluteinizing-hormone release in the female sheep during anestrus. BiolReprod 1997; 56:1544–1549.
35. Skinner DC, Herbison AE. Effects of photoperiod on estrogen receptor,tyrosine hydroxylase, neuropeptide Y, and beta-endorphin immunoreac-tivity in the ewe hypothalamus. Endocrinology 1997; 138:2585–2595.
36. Smith JT, Cunningham MJ, Rissman EF, Clifton DK, Steiner RA.Regulation of Kiss1 gene expression in the brain of the female mouse.Endocrinology 2005; 146:3686–3692.
37. Irwig MS, Fraley GS, Smith JT, Acohido BV, Popa SM, Cunningham MJ,Gottsch ML, Clifton DK, Steiner RA. Kisspeptin activation ofgonadotropin releasing hormone neurons and regulation of KiSS-1 mRNAin the male rat. Neuroendocrinology 2004; 80:264–272.
38. Jayasena CN, Dhillo WS, Bloom SR. Kisspeptins and the control ofgonadotropin secretion in humans. Peptides 2009; 30:76–82.
39. Messager S, Chatzidaki EE, Ma D, Hendrick AG, Zahn D, Dixon J,Thresher RR, Malinge I, Lomet D, Carlton MB, Colledge WH, Caraty A,et al. Kisspeptin directly stimulates gonadotropin-releasing hormonerelease via G protein-coupled receptor 54. Proc Natl Acad Sci U S A2005; 102:1761–1766.
40. Shahab M, Mastronardi C, Seminara SB, Crowley WF, Ojeda SR, PlantTM. Increased hypothalamic GPR54 signaling: a potential mechanism forinitiation of puberty in primates. Proc Natl Acad Sci U S A 2005;102:2129–2134.
41. Smith JT, Clay CM, Caraty A, Clarke IJ. KiSS-1 messenger ribonucleicacid expression in the hypothalamus of the ewe is regulated by sex steroidsand season. Endocrinology 2007; 148:1150–1157.
42. Smith JT, Coolen LM, Kriegsfeld LJ, Sari IP, Jaafarzadehshirazi MR,Maltby M, Bateman K, Goodman RL, Tilbrook AJ, Ubuka T, Bentley GE,Clarke IJ, et al. Variation in kisspeptin and RFamide-related peptide(RFRP) expression and terminal connections to gonadotropin-releasinghormone neurons in the brain: a novel medium for seasonal breeding in thesheep. Endocrinology 2008; 149:5770–5782.
43. Wagner GC, Johnston JD, Clarke IJ, Lincoln GA, Hazlerigg DG.Redefining the limits of day length responsiveness in a seasonal mammal.Endocrinology 2008; 149:32–39.
44. Billings HJ, Connors JM, Altman SN, Hileman SM, Holaskova I, LehmanMN, McManus CJ, Nestor CC, Jacobs BH, Goodman RL. Neurokinin Bacts via the neurokinin-3 receptor in the retrochiasmatic area to stimulateluteinizing hormone secretion in sheep. Endocrinology 2010; 151:3836–3846.
45. Dardente H, Birnie M, Lincoln GA, Hazlerigg DG. RFamide-relatedpeptide and its cognate receptor in the sheep: cDNA cloning, mRNAdistribution in the hypothalamus and the effect of photoperiod. JNeuroendocrinol 2008; 20:1252–1259.
46. Smith JT, Clarke IJ. Gonadotropin inhibitory hormone function inmammals. Trends Endocrinol Metab 2010; 21:255–260.
47. Anderson GM, Hardy SL, Valent M, Billings HJ, Connors JM, GoodmanRL. Evidence that thyroid hormones act in the ventromedial preoptic areaand the premammillary region of the brain to allow the termination of thebreeding season in the ewe. Endocrinology 2003; 144:2892–2901.
48. Malpaux B, Daveau A, Maurice-Mandon F, Duarte G, Chemineau P.Evidence that melatonin acts in the premammillary hypothalamic area tocontrol reproduction in the ewe: presence of binding sites and stimulationof luteinizing hormone secretion by in situ microimplant delivery.Endocrinology 1998; 139:1508–1516.
lPOA/rAHA AND SEASONAL BREEDING IN FEMALE SHEEP 1065
Dow
nloaded from w
ww
.biolreprod.org.




























