Embed Size (px)
Citation preview

www.elsevier.com/locate/yniox
Nitric Oxide 11 (2004) 201–207
NITRICOXIDE
Biology and Chemistry
Nitric oxide-scavenging activity of polyhydroxylated fullerenol,C60(OH)24
Snezana M. Mirkova, Aleksandar N. Djordjevicc, Nebojsa L. Andricb, Silvana A. Andricb,Tatjana S. Kosticb, Gordana M. Bogdanovicd, Mirjana B. Vojinovic-Miloradovc,
Radmila Z. Kovacevicb,*
a Department of Medicine, The University of Chicago, 5841 South Maryland Ave. MC 2115, Chicago, IL 60637, USAb Faculty of Sciences, Department of Biology and Ecology, University of Novi Sad, 21000 Novi Sad, Serbia and Montenegro, Yugoslavia
c Faculty of Sciences, Department of Chemistry, University of Novi Sad, 21000 Novi Sad, Serbia and Montenegro, Yugoslaviad Faculty of Medicine, Institute of Oncology, University of Novi Sad, 21000 Novi Sad, Serbia and Montenegro, Yugoslavia
Received 20 April 2004; revised 12 July 2004Available online 25 September 2004
Abstract
Investigation of the possible nitric oxide-scavenging activity of hydroxylated derivative of fullerene, fullerenol C60(OH)24, demon-strated that it expressed direct scavenging activity toward nitric oxide radical (NO) liberated within solution of sodium nitroprusside(SNP), a well known NO donor. In parallel, pre-treatment (30 0) with intratesticular injection of fullerenol (60 lg/each testis) pre-vented NO-induced decrease of catalase, glutathione transferase and glutathione peroxidase activities in the denucleated fractionof interstitial testicular cells of adult rats 2 h after intratesticular injection of SNP (20 lg/each testis). In addition, fullerenol decreasedformation of thiobarbituric acid-reactive substances (TBA-RS) with similar efficiency as butylated hydroxy toluen (BHT), a wellknown antioxidant. Also, fullerenol expressed certain scavenging activity toward superoxide anion ðO�
2 Þ in xanthine/xanthine oxidasesystem. In summary, results obtained in this study confirmed free radical-scavenging activity of fullerenol, and according to ourknowledge, it is the first evidence of direct NO-quenching activity of hydroxylated C60 derivative in different milieu.� 2004 Elsevier Inc. All rights reserved.
Keywords: Nitric oxide; Fullerenol; NO-scavenging activity; Antioxidant enzymes
Investigation of chemical and physical properties ofwater-soluble derivatives of fullerene C60 has yieldedan extensive amount of information, including its reac-tivity with reactive oxygen species (ROS) [1–5]. Combi-nation of the moderate electron affinity and the allylichydroxy functional group of water soluble fullerenolsmakes them appropriate candidate for application suchas free radical remover or water soluble antioxidant inbiological system [6]. It has been shown that polyhydr-oxylated fullerene (C60(OH)n n = 6–24) fullerenolprevented hydrogen peroxide and cumene hydroperox-
1089-8603/$ - see front matter � 2004 Elsevier Inc. All rights reserved.
doi:10.1016/j.niox.2004.08.003
* Corresponding author. Fax: 381 21 450 620.E-mail address: [email protected] (R.Z. Kovacevic).
ide-elicited damage in the hippocampus slices [3].Water-soluble fullerenol has shown excellent efficiencyin eliminating superoxide radical species [2,6–8]. Antiox-idative activity of fullerenol was demonstrated to be bet-ter than vitamin E in prevention of lipid peroxidationinduced by superoxide and hydroxyl radicals [9]. Alsoit has been demonstrated that C60 or water solublederivatives have inhibitory effects on lipid peroxides for-mation in differrent model system [10,11]. There is a lit-tle evidence concerning direct NO-scavenging activity offullerene derivatives. According to Satoh et al. [12] theinhibitory effect of dimalonic acid C60 on NO-inducedrelaxation of rabbit aorta could be explained by produc-tion of superoxide, and neutralization of NO by super-

202 S.M. Mirkov et al. / Nitric Oxide 11 (2004) 201–207
oxide anion, since such an effect was masked in the pres-ence of superoxid dismutase (SOD). The same effect wasdemonstrated for monomalonic acid C60 [13]. On theother hand, different derivatives of fullerene exert neuro-protective effects. According to Dugan et al. [4] carbox-ylic acid C60 derivatives with three malonic acid groupper molecule expressed a potent ROS-scavenging activ-ity and prevented the citotoxic death of cultured corticalneurons induced by exposure to NMDA-agonists. Thesame derivatives protected the nigro-striatal dopaminer-gic system from iron-induced oxidative injury [14]. Also,C3-fullero-tris-methanodicarboxylic acid preventedapoptosis in cerebellar granule cells caused by oxidativestress [15]. Free radical scavenging properties of fullere-nols are supposed to be responsible for their protectionfrom oxidative stress in cultured cortical neurons [16].Protective effects of C60(OH)18–20 toward neurotoxicityof glutamate was explained by glutamate receptorantagonistic activity of fullerol [17].
The aim of this study was to investigate a potentialscavenging activity of polyhydroxylated derivative offullerene, fullerenol C60(OH)24, toward NO, lipid perox-ides and superoxide radical. Activity of this compoundwas tested by co-incubation of C60(OH)24 with sodiumnitroprusside (SNP), a potent NO donor. The changesin the nitrite levels, a stable oxidative product of NO[18], were used as a measure of potential NO-scavengingactivity of C60(OH)24. We also investigated possible invivo NO-scavenging activity of this compound. We havepreviously shown that intratesticular injection of NO-donors, isosorbide dinitrate (ISDN) and SNP or exoge-nous substrate for nitric oxide synthase, LL-argininemethyl esther (LL-AME), resulted in decreased activitiesof catalase, glutathione peroxidase (GSH-Px) and gluta-thione transferase (GST) in the interstitial testicular tis-sue 2 h after treatment [19–21]. Therefore, antioxidantenzyme activities of the interstitial cells of adult rat testiswere used as a model system to follow NO-scavengingactivity of C60(OH)24. SNP and C60(OH)24, alone or incombination, were given by intratesticular injections toallow mainly local effect at the level of testicular tissue.The changes in the antioxidant enzyme activities wereused as a measure of potential NO-scavenging activityof C60(OH)24. For investigation of superoxide radicalquenching activity of C60(OH)24 we used well-describedxanthine/xanthine oxidase system for producing super-oxide radicals, while formation of lipid peroxides wasmonitored in liposomes/FeSO4/ascorbic acid system.
Experimental procedures
Chemicals
C60 was obtained from Alfa Aesar 99.5% (USA).Medium 199 was purchased from GIBCO Laboratories
(Gaithersburg, MD). Natriumazid, collagenase (Type I),hydrogen peroxide, xanthine, xanthine oxidase (buttermilk), cytochrome c, reduced glutathione (GSH), 1-chloro-2,4-dinitrobenzene (CDNB), tert-butyl hydro-peroxide, reduced nicotinamide adenine dinucleotidephosphate (bNADPH) were all purchased from SigmaChemical Co. (St. Louis, MI). All other reagents wereof analytical grade.
Synthesis of fullerenol C60(OH)24
Tested compound was synthesized and characterizedby Djordjevic et al. [22]. In brief, polybromine derivativeC60Br24 was synthetized with catalytical (FeBr3) reactionof C60 in Br2 [23]. Polyhydroxylated fullererenolC60(OH)24 was synthetized in alcaline media by completesubstitution of bromine atomes from C60Br24. Fifty mgof C60Br24was mixed in 5 cm3 of NaOH, pH 10, for 2 hat room temperature. After the reaction was completedthe solvent was evaporated at 40 �C, and the mixturewas five times rinsed with 10 cm3 portion of 80% ethanol.Water solution (20 ml) of fullerenol with residualamounts of NaOH and NaBr was applied to the top ofthe combined ion exchange resin (20 g DOWEX MB50QC121815 R1) and eluted with demineralized water untildiscoloration. Water solution of fullerenol (pH 7) wasevaporated under low pressure and a dark brown powdersubstance was left in the beaker. FTIR C60(OH)24: 3427,1627, 1419, 1080 cm�1; 13C NMR (D2O) reaction mix-ture: singlet peaks at d = 77.7 ppm and multiplet atd = 140 ppm, 13C NMR (D2O) C60(OH)24: singlet peaksd = 169.47 ppm and multiplet peak 160–110 ppm;MAL-DI (matrix a-cyano-4-hydroxycinnamic acid) (m/z): 720ðCþ
60Þ, 721 (C60H+), 722 ðC60H
þ2 Þ, 737 (C60(OH)+), 808
ðC60ðOHÞþ5 Þ, 839 ðC60ðOHÞþ7 Þ, 856 ðC60ðOHÞþ8 Þ, 1009ðC60ðOHÞþ17Þ, 1026 ðC60ðOHÞþ18Þ, 1094 ðC60ðOHÞþ22Þ andminor pik 1128 ðC60ðOHÞþ24Þ; DTG, DTA, TG: revealstwo thermal changes, in temperature of 120–395 �C, cor-responding to the loss of mass of 35.7% (23.7 OHgroups) and at the temperature of 430 �C loss of masswas 64.3%. (temperature of sublimation of C60). Elemen-tary analysis of fullerenol provided the following: C,63.0%; H, 2.0%; calc.: C, 63.83%; H, 2.13%. Obtainedresults are very similar to those previously published[24–26].
NO-scavenging activity of fullerenol C60(OH)24 in SNP
solution
To estimate possible NO-scavening activity of fullere-nol, different concentrations of SNP (1, 2 or 5 mM) wereincubated alone or in combination with different con-centrations of fullerenol (0.05 or 0.1 mM). All solutionswere made in 0.1 M phosphate buffer (pH 7.4) andadded directly into tubes. SNP is an inorganic complexwhere NO is found as NO+ and light irradiation is nec-

S.M. Mirkov et al. / Nitric Oxide 11 (2004) 201–207 203
essary for release of NO [27]. Therefore, incubation mix-tures were incubated on light, at room temperature, andnitrite levels were determined exactly after 30, 60, and120 min.
For nitrite measurements, sample aliquots of 1 mlwere mixed with an equal volume of Greiss reagent con-taining 1% sulphanilamide and 0.1% naphthylethylen-ediamine in 5% phosphoric acid. The mixture was thenincubated at room temperature for 10 min and theabsorbance was measured at 546 nm [28]. Nitrite con-centrations were determined relative to a standard curvederived from increasing concentrations of sodiumnitrite.
In vivo NO-scavening activity of fullerenol C60(OH)24
Experiments were performed in adult male Wistarrats bred in laboratory of Department of Biology andEcology and raised under controlled environmental con-ditions (temperature 22 ± 2 �C; 14 h light/10 h dark)with food and water ad libitum. Rats were handled dailyfor one week acclimation period prior to the experi-ments. Effects of NO-scavenging activity of fullerenolon the activities of testicular antioxidant enzymes wereinvestigated after intratesticular injection of SNP andfullerenol into each testis. Fullerenol (60 lg/testis) wasinjected at the beginning of experiment and SNP (20lg/testis) was injected 30 min later. The control animalswere injected with saline at the same time. Animals weresacrificed by decapitation 2 h after the injection of SNP.
All experiments were approved by the Local AnimalEthical Committee of the University of Novi Sad andwere conducted in accordance with principles and proce-dures of the NIH Guide for Care and Use of LaboratoryAnimals.
Right testes from control and experimental animalswere used for measuring antioxidant enzyme activities.Testes were decapsulated and dissociated with collage-nase (1.2 mg/ml) in M199-BSA as previously described[29]. Interstitial cells were resuspended in a buffer con-taining 17 mM Tris and 140 mM NH4Cl (pH 7.2) andincubated for 10 min at room temperature. This proce-dure eliminates red blood cells and interference ofhemoglobin. The cell pellets were washed twice withM199-BSA and once with 0.9% NaCl solution. Denucle-ated fraction of interstitial cells (separately for eachanimal) was obtained by homogenization of cell pelletsin 50 mM phosphate buffer containing 0.25 M sucrose(pH 7.4) by a glass-glass homogenizer and centrifuga-tion for 20 min at 1500g (4 �C). Protein content indenucleated fraction of interstitial cells was estimatedby Bradford method [30], using BSA as a standard.
The activity of superoxide dismutase was measuredby inhibition of xanthine/xanthine oxidase-dependentreduction of cytochrome c, according to the method de-scribed by McCord and Fridovich [31], with some mod-
ification. The catalase activity was determined by therate of H2O2 disappearance recorded at 240 nm, accord-ing to Claiborne [32]. The GST activity was assayedusing CDNB as a substrate and measuring the absor-bency change at 340 nm, according to Habig et al.[33]. Oxidation of NADPH at 340 nm, with tert-butylhydroperoxide as a substrate was employed in the mod-ified method of Paglia and Valentine [34] for measuringthe activity of glutathione peroxidase.
The results of SOD, catalase and GST assays were ex-pressed as units of enzyme activity per mg protein, (U/mg prot.) and for GSH-peroxidase as nM NADPH/min/mg of protein.
Prevention of lipid peroxidation by fullerenol C60(OH)24in liposomes model system
The concentration of thiobarbituric acid-reactivesubstances (TBA-RS) was used as a measure of lipidperoxidation according to the method of Buege andAust [35]. We used liposomes, a commercial preparationof biological membrane, as a model system. Briefly,tested substances and liposomes were added to FeSO4/ascorbic acid system and incubated 1 h at room temper-ature. After incubation thiobarbituric acid was addedand incubated again for 20 min at 100 �C. After coolingand centrifugation, levels of TBA-RS were monitoredspectrophotometrically at 532 nm. Antioxidant poten-tial of fullerenol toward lipid peroxides was comparedto BHT.
Superoxide radical-scavenging activity of fullerenol
C60(OH)24 in xanthine/xanthine oxidase system
To estimate possible superoxide radical-quenchingactivity of fullerenol, we used xanthine/xanthine oxidasesystem according to the method of McCord and Frido-vich [31] with some modification. Superoxide radicals,generated through the enzymatic activity of xanthineoxidase, reduce cytochrome c. Removing superoxideradicals from the system lead to decreased rate of cyto-chrome c reduction. Reaction mixture contained 0.01 MPO4 buffer, pH 7.8, 0.1 mM EDTA, 0.1 mM NaN3, 50lM xanthine, 20 lM cytochrome c and fullerenol in dif-ferent concentration (10�10–10�3 M). Reaction wasstarted by adding of xanthine oxidase and reduction rateof cytochrome c was monitored at 550 nm for 20 min.
Statistics
All results were expressed as means ± SE. Data wereanalysed using Mann–Whitney test, with p < 0.05 andby analysis of variance (ANOVA). Post hoc compari-sons between means were made by Duncan test, withp < 0.05. The dose of fullerenol which reduce the esti-
204 S.M. Mirkov et al. / Nitric Oxide 11 (2004) 201–207
mated response by 50%, i.e., ED50 was calculated by theleast-square method.
Results
NO-scavenging activity of fullerenol C60(OH)24 in SNP
solution
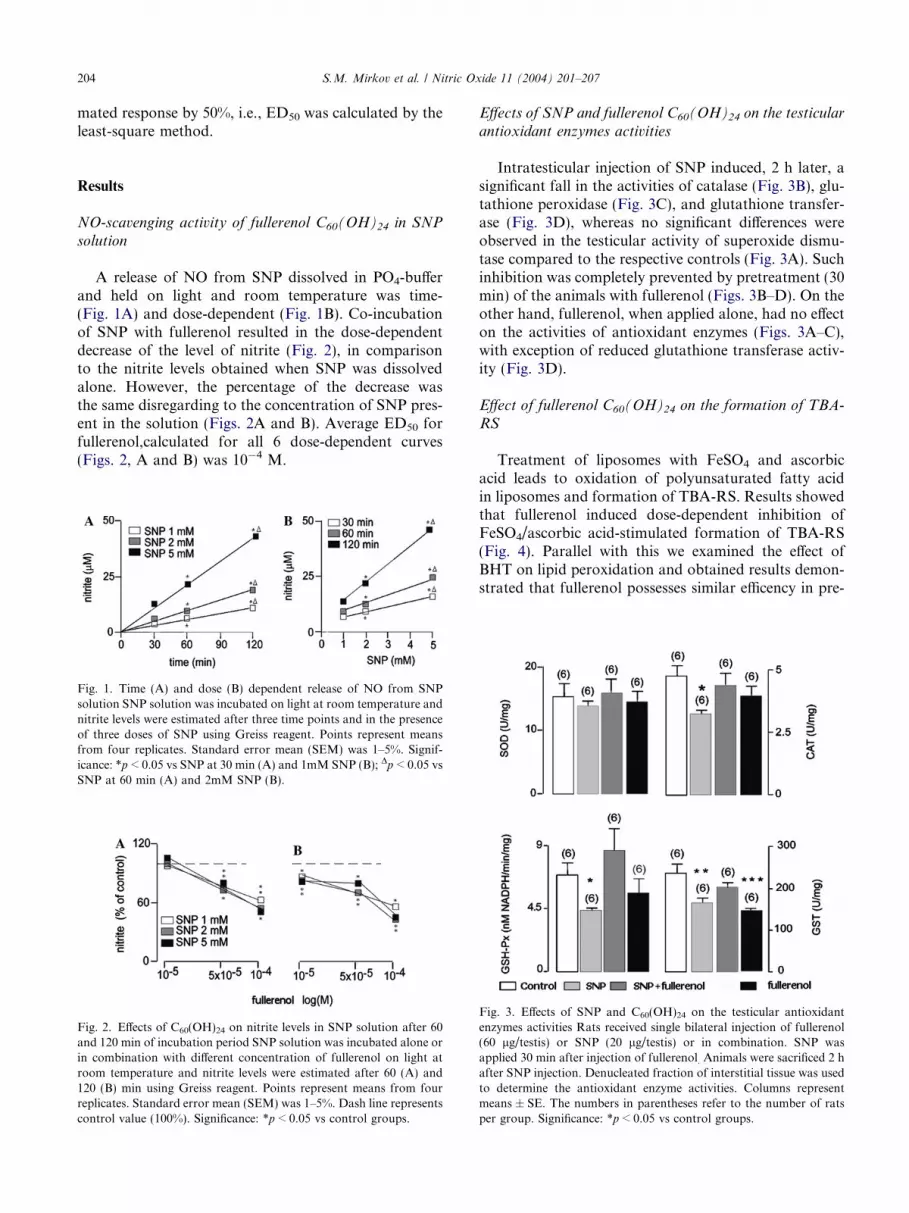
A release of NO from SNP dissolved in PO4-bufferand held on light and room temperature was time-(Fig. 1A) and dose-dependent (Fig. 1B). Co-incubationof SNP with fullerenol resulted in the dose-dependentdecrease of the level of nitrite (Fig. 2), in comparisonto the nitrite levels obtained when SNP was dissolvedalone. However, the percentage of the decrease wasthe same disregarding to the concentration of SNP pres-ent in the solution (Figs. 2A and B). Average ED50 forfullerenol,calculated for all 6 dose-dependent curves(Figs. 2, A and B) was 10�4 M.
Fig. 1. Time (A) and dose (B) dependent release of NO from SNPsolution SNP solution was incubated on light at room temperature andnitrite levels were estimated after three time points and in the presenceof three doses of SNP using Greiss reagent. Points represent meansfrom four replicates. Standard error mean (SEM) was 1–5%. Signif-icance: *p < 0.05 vs SNP at 30 min (A) and 1mM SNP (B); Dp < 0.05 vsSNP at 60 min (A) and 2mM SNP (B).
Fig. 2. Effects of C60(OH)24 on nitrite levels in SNP solution after 60and 120 min of incubation period SNP solution was incubated alone orin combination with different concentration of fullerenol on light atroom temperature and nitrite levels were estimated after 60 (A) and120 (B) min using Greiss reagent. Points represent means from fourreplicates. Standard error mean (SEM) was 1–5%. Dash line representscontrol value (100%). Significance: *p < 0.05 vs control groups.
Effects of SNP and fullerenol C60(OH)24 on the testicular
antioxidant enzymes activities
Intratesticular injection of SNP induced, 2 h later, asignificant fall in the activities of catalase (Fig. 3B), glu-tathione peroxidase (Fig. 3C), and glutathione transfer-ase (Fig. 3D), whereas no significant differences wereobserved in the testicular activity of superoxide dismu-tase compared to the respective controls (Fig. 3A). Suchinhibition was completely prevented by pretreatment (30min) of the animals with fullerenol (Figs. 3B–D). On theother hand, fullerenol, when applied alone, had no effecton the activities of antioxidant enzymes (Figs. 3A–C),with exception of reduced glutathione transferase activ-ity (Fig. 3D).
Effect of fullerenol C60(OH)24 on the formation of TBA-
RS
Treatment of liposomes with FeSO4 and ascorbicacid leads to oxidation of polyunsaturated fatty acidin liposomes and formation of TBA-RS. Results showedthat fullerenol induced dose-dependent inhibition ofFeSO4/ascorbic acid-stimulated formation of TBA-RS(Fig. 4). Parallel with this we examined the effect ofBHT on lipid peroxidation and obtained results demon-strated that fullerenol possesses similar efficency in pre-
Fig. 3. Effects of SNP and C60(OH)24 on the testicular antioxidantenzymes activities Rats received single bilateral injection of fullerenol(60 lg/testis) or SNP (20 lg/testis) or in combination. SNP wasapplied 30 min after injection of fullerenol. Animals were sacrificed 2 hafter SNP injection. Denucleated fraction of interstitial tissue was usedto determine the antioxidant enzyme activities. Columns representmeans ± SE. The numbers in parentheses refer to the number of ratsper group. Significance: *p < 0.05 vs control groups.

Fig. 4. Effect of C60(OH)24 on the formation of TBA-RS C60(OH)24and BHT were incubated in liposomes/FeSO4/ascorbic acid system indifferent concentrations and the level of lipid peroxidation wasdetermined spectrofotometrically at 532 nm as the amount of TBA-RS formed. Points represent means from three repeats. Significance:*p < 0.05 vs control groups.
S.M. Mirkov et al. / Nitric Oxide 11 (2004) 201–207 205
vention of lipid peroxidation as BHT (CalculatedED50 for fullerenol was 2.5 · 10�3 M, and for BHT1.2 · 10�3 M).
Superoxide radical-scavenging activity of fullerenol
C60(OH)24
Application of fullerenol into xanthine/xanthine oxi-dase system caused decrease in reduction rate of cyto-chrome c compared to control (Fig. 5). Obtainedresult demonstrated that fullerenol in the range ofnanomolar and micromolar concentrations decreasedreduction of cytochrome c beetwen 5 and 20%, whileconcentracion of 1 mM decreased reduction of cyto-chrome c for 40%.
Fig. 5. Effects of C60(OH)24 on cytochrome c reduction Superoxideradicals, generated through enzymatic activity of xanthine oxidase,reduced cytochrome c (Fe3+ in Fe2+) and rate of reduction wasmonitored as a change of absorbance at 550 nm. Different concentra-tion (10�10–10�3 M) of fullerenol were incubated in xanthine/xanthineoxidase system. Points represent means from two repeats that weremeasured every minute in a period of 20 min.
Discussion
In order to test scavenging activity of fullerenol inNO-donor solution, it is necessary to provide spontane-ous liberation of this radical into the solution. Thereforewe used SNP which is spontaneous liberator of NO inthe presence of light irradiation [27]. Besides SNP, thereare other spontaneous NO-donors with long lifetime,such as DETA/NO, or acetylpenicillamine (SNAP).However, data from the literature showed that severalauthors also used SNP as donor of NO, in order to testthe ability of different substances as NO-scavenger [36],or to prevent NO-mediated effects [37,38]. Also, in ourprevious study, SNP was efficient inhibitor of antioxi-dant enzyme activities in testicular tissue [20]. Obtainedresults showed that the presence of fullerenol in SNPsolution decreased levels of nitrite, a stable oxidationproduct of NO liberated from SNP, with practicallythe same EC50 regardless of SNP concentrations. Forall six curves (1, 2 and 5 mM of SNP in two time points)EC50 values was approximately 10�4 M (from 0.8 · 10�4
to 1.21 · 10�4M; average 0.98 · 10�4M), regardless ofSNP dose or time intervals.
To test possible in vivo NO-scavenging activity of ful-lerenol, antioxidant defence in adult rat testis was usedas a model system. It is known that adult rat testis ischaracterized by a relatively strong antioxidant defencesystem, including elevated levels of antioxidant vita-mins, as well as a presence of antioxidant enzyme bat-tery [39]. On the other hand, results from ourlaboratory showed that NO donors, ISDN or SNP, re-duced catalase, glutathione transferase and sglutathioneperoxidase activities [19,20] in the rat testicular intersti-tial cell compartment. As results in this study shown,pretreatment of the rats with it injection of fullerenolcompletely prevented SNP-induced reduction in theactivities of antioxidant enzymes. When applied alone,fullerenol induced no changes in the activity of antioxi-dant enzymes, with exception of decreased glutathionetransferase activity. This inhibition could be partially ex-plained by findings that certain derivatives of fullerenolinhibit glutathione reductase activity [2] which is neces-sary for normal activity of GST. These results suggestthat fullerenol possesses NO-scavenging activity in vivo.
In parallel with NO-scavenging activity, we investi-gated scavenging efficiency of our derivative toward lipidperoxides and superoxide radicals. Our results showedthat fullerenol blocked lipid peroxide formation in lipo-somes/FeSO4/ascorbic acid system. The calculatedED50 value was 2.5 · 10�3 M and it was very similar withED50 of BHT (1.23 · 10�3 M), a well known antioxidant.Concerning superoxide radical scavenging activity inxanthine/xanthine oxidase system, which was monitoredthrough reduction of cytochrome c, fullerenol expresseda relatively low activity in comparison to water-solublederivatives described in other reports [2,8,40].

206 S.M. Mirkov et al. / Nitric Oxide 11 (2004) 201–207
In summary, results obtained in this study confirmedfullerenol reactivity toward ROS, and according to ourknowledge, it is the first evidence of direct NO-quench-ing activity of hydroxylated C60 derivative in differentmilieu.
Acknowledgement
This work was supported by Ministry of Science,Technology and Development, Republic of Serbia,Grant No.1893
References
[1] P.J. Krusic, E. Wasserman, P.N. Keizer, J.R. Morton, K.F.Preston, Radical reactions of C60, Science 254 (1991) 1183–1185.
[2] K. Okuda, T. Mashino, M. Hirobe, Effects of a fullerenederivative C60-dimalonic acid, on active oxygen toxicity, FullereneSci. Technol. 3 (1996) 411–418.
[3] M.C. Tsai, Y.H. Chen, L.Y. Chiang, Polyhydroxylated C60,fullerenol, a novel free-radical trapper, prevented hydrogenperoxide-and cumene hydroperoxide-elicited changes in rat hip-pocampus in vitro, J. Pharm. Pharmacol. 49 (1997) 438–445.
[4] L.L. Dugan, D.M. Turetsky, C. Du, D. Lobner, M. Wheeler,C.R. Almli, C.K.F. Shen, T.Y. Luh, D.W. Choi, T.-S. Lin,Carboxyfullerenes as neuroprotective agents, Proc. Natl. Acad.Sci. USA (1997) 9434–9439.
[5] C.Y. Lu, S.D. Yao, W.Z. Lin, W.F. Wang, N.Y. Lin, Y.P. Tong,T.W. Rong, Studies on the fullerol of C60 in aqueous solution withlaser photolysis and pulse radiolysis, Radiat. Phys. Chem. 53(1998) 137–143.
[6] L.Y. Chiang, F.J. Lu, J.T. Lin, Free radical scavenging activity ofwater-soluble fullerenols, J. Chem. Soc. Chem. Commun. (1995)1283–1285.
[7] G. Bogdanovic, V. Kojic, A. -Dord-evic, J. Canadanovic-Brunet,M. Vojinovic-Miloradov, V.V. Baltic, Modulating activity offullerol C60(OH)22 on doxorubicin-induced cytotoxicity, Toxicol-ogy in Vitro, in press.
[8] K. Okuda, T. Hirota, M. Hirobe, T. Nagano, M. Mochizuki, T.Mashino, Synthesis of various water-soluble C60 derivatives andtheir superoxide-quenching activity, Fullerene Sci. Technol. 8(2000) 89–104.
[9] I.C. Wang, L.A. Tai, D.D. Lee, P.P. Kanakamma, C.K.F. Shen,T.-Y. Luh, C.H. Cheng, K.C. Hwang, C60 and water-solublefullerene derivatives as antioxidant against radical-initiated lipidperoxidation, J. Med. Chem. 42 (1999) 4614–4620.
[10] Y.T. Lee, L.Y. Chiang, W.J. Chen, H.C. Hsu, Water-solublehexasulfobutyl(60)fullerene inhibit low-density lipoprotein oxida-tion in aqueous and lipophilic phases, P.S.E.B.M. 224 (2000) 69–75.
[11] H.S. Lai, Y. Chen, W.J. Chen, K.J. Chang, L.Y. Chiang, Freeradical scavenging activity of fullerenol on grafts after small boweltransplatation in dogs, Transplant. Proc. 32 (2000) 1272–1274.
[12] M. Sato, K. Matsuo, H. Kiriya, T. Mashino, M. Hirobe, I.Takayanagi, Inhibitory effects of fullerene derivative, dimalonicacid C60, in nitric oxide-induced relaxation of rabbit aorta, Eur. J.Pharmacol. 327 (1997) 175–181.
[13] M. Sato, K. Matsuo, H. Kiriya, T. Mashino, M. Hirobe, I.Takayanagi, Inhibitory effects of fullerene derivative, monoma-lonic acid C60, in nitric oxide-dependent relaxation of aorticsmooth muscle, Eur. J. Pharmacol. 29 (1997) 345–351.
[14] A.M.Y. Lin, Recovery by NO of the iron-attenuated dopaminedynamics in nigrostriatal system of rat brain, Neurosci. Res. 34(1999) 133–139.
[15] M. Bisaglia, R. Natalini, R. Pellicciari, E. Straface, W. Malorni,D. Monto, C. Franceschi, G. Schettini, C3-fullero-tris-methanod-icarboxylic acid protects cerebellar granule cells from apoptosis, J.Neurochem. 74 (2000) 1197–1204.
[16] L.L. Dugan, J.K. Gabrielsen, S.P. Yu, T.-S. Lin, D.W. Choi,Buckminsterfullerenol free radical scavengers reduce excitotoxicand apoptotic death of cultured cortical neurons, Neurobiol. Dis.3 (1996) 129–135.
[17] H. Jin, W.Q. Chen, X.W. Tang, L.Y. Chiang, C.Y. Yang, J.V.Schloss, J.Y. Wu, Polyhydroxylated C(60), fullerenols, as gluta-mate receptor antagonists and neuroprotective agents, Neurosci.Res. 62 (2000) 600–607.
[18] L.J. Ignarro, J.M. Fukuto, J.M. Griscavage, N.E. Rogers,Oxidation of nitric oxide in aqueous solution to nitrite but notnitrate, comparison with enzymatically formed nitric oxide fromLL-arginine, Proc. Natl. Acad. Sci. USA 90 (1993) 8103–8107.
[19] T.S. Kostic, S.A. Andric, D. Maric, R.Z. Kovacevic, Inhibitoryeffects of stress-activated nitric oxide on antioxidant enzymes andtesticular steroidogenesis, J. Steroid Biochem. Mol. Biol. 75 (2000)299–306.
[20] N.L. Andric, S.A. Andric, T.S. Kostic, S.M. Dragisic, R.Z.Kovacevic, Inhibitory effects of LL-arginin methyl ester on antiox-idant enzymes and stress-impared steroidogenesis in rat testis,Rev. Res. Fac. Sci. 30 (2001) 43–57.
[21] N.L. Andric, The role of nitric oxide/nitric oxide synthase systemon stress-impaired steroidogenic function in rat testis. MSc.Thesis, University of Belgrade, Belgrade, 2001.
[22] A. Djordjevic, Synthesis of water soluble derivatives of fullereneC60. Ph. D. Thesis, University of Novi Sad, Novi Sad, 2000.
[23] A. Djordjevic, M. Vojinovic-Miloradov, N. Petranovic, A. Deve-cerski, D. Lazar, B. Ribar, Catalytical preparation and charac-terization of C60Br24, Fullerene Sci. Technol. 6 (1998) 689–694.
[24] Y.C. Chaing, Y.W. Swirtczewski, S. Chang, S.K. Hsu, S.Cameron, K. Creegan, Multi-hydroxy additions onto C60 fuller-ene molecules, J. Chem. Soc. Chem. Commun. (1992) 1791–1792.
[25] J. Li, A. Takeuchi, M. Ozawa, X. Li, K. Saigo, K. Kitazawa, C60
fullerenol formation catalysed by quaternary amonium hydrox-ides, J. Chem. Soc. Chem. Commun. (1993) 1784–1785.
[26] Y. Chen, R.F. Cai, S. Chen, Z.E. Huang, Synthesis andcharacterization of fullerol derived from C60 precursores, J. Phys.Chem. Sol. 62 (2001) 99–101.
[27] M. Feelisch, J.S. Stamler, Donors of nitrogen oxides, in: JohnWiley (Ed.), Methods in Nitric Oxide Research, John Wiley, 1996.
[28] L.C. Green, D.A. Wagner, J. Glogowski, P.L. Skipper, J.S.Wishnok, S.R. Tannenbaum, Analysis of nitrate, nitrite and [15N]nitrite in biological fluids, Anal. Biochem. 126 (1982) 131–138.
[29] R. Kovacevic, M. Sarac, Bromocriptine-induced inhibition ofhydroxylase/lyase activity of adult rat Leydig cells, J. SteroidBiochem. Mol. Biol. 46 (1993) 841–845.
[30] M.M. Bradford, A rapid and sensitive method quantitation ofmicrogram quantities of protein utilizing the principle of protein-dye binding, Anal. Biochem. 72 (1976) 248–254.
[31] J.M. McCord, I. Fridovich, The reduction of cytochrome c bymilk xanthine oxidase, J. Biol. Chem. 243 (1968) 5753–5760.
[32] A. Claiborne, Catalase activity, in: R.A. Greenwald (Ed.), CRCHandbook of Methods for Oxygen Radical Research, CRC PressInc, Boca Raton, FL, 1984.
[33] W.H. Habig, M.J. Pabst, W.B. Jakoby, Glutathione-S-transfer-ases, J. Biol. Chem. 249 (1974) 7130–7139.
[34] D.E. Paglia, N.W. Valentine, Studies on the quantitative andqualitative characterization of erythrocyte glutathione peroxi-dases, J. Lab. Clin. Med. 70 (1967) 74–77.
[35] A.L. Buege, D.S. Aust, Microsomal lipid peroxidation, Methodsin Enzimology, Academic Press, New York, 1978.

S.M. Mirkov et al. / Nitric Oxide 11 (2004) 201–207 207
[36] S. Kagota, Y. Yamaguchi, K. Nakamura, K. Shinozuka, M.Kunitomo, Chronic nitric oxide exposure alters the balancebetween endothelium-derived relaxing factors released from ratrenal arteries: prevention by treatment with NOX-100, a NOscavenger, Life Sci. 74 (2004) 2757–2767.
[37] S.L. Pan, J.H. Guh, Y.L. Chang, S.C. Kuo, F.Y. Lee, C.M.Teng, YC-1 prevents sodium nitroprusside-mediated apoptosis invascular smooth muscle cells, Cardiovasc. Res. 61 (2004) 152–158.
[38] Y. Huang, C.M. Wong, C.W. Lau, X. Yao, S.Y. Tsang, Y.L. Su,Z.Y. Chen, Inhibition of nitric oxide/cyclic GMP-mediatedrelaxation by purified flavonoids, baicalin and baicalein, in rataortic rings, Biochem. Pharmacol. 67 (2004) 787–794.
[39] V. Peltola, I. Huhtaniemi, M. Ahotupa, Antioxidant enzymeactivity in the maturing rat testis, J. Androl. 13 (1992) 450–455.
[40] C. Yu, J.B. Bhonsle, Y. Wang, L.J.G. Lin, B.J. Chen, L.Y.Chiang, Synthetic aspects and free-radical scavenging efficiency ofpolyhydroxylated C60, Fullerene Sci. Technol. 5 (1997) 1407–1421.









![Stereoselective synthesis of polyhydroxylated pyrrolidines: a route to novel 3,5-bis(hydroxymethyl)pyrrolidines from 2-azabicyclo[2.2.1]hept-5-enes](https://img.pdfslide.net/doc/110x75/635984f310f2bd48220e39e1/stereoselective-synthesis-of-polyhydroxylated-pyrrolidines-a-route-to-novel-35-bishydroxymethylpyrrolidines.jpg)


















![[Fullerenol – properties and applications in biomedical sciences]](https://img.pdfslide.net/doc/110x75/63124de2c32ab5e46f0be7ef/fullerenol-properties-and-applications-in-biomedical-sciences.jpg)
