Embed Size (px)
Citation preview

ENVIRONMENTAL ENGINEERING SCIENCEVolume 20 Number 3 2003copy Mary Ann Liebert Inc
Nitrification Inhibition by Ethylenediamine-Based Chelating Agents
Zhiqiang Hu1 Kartik Chandran1dagger Domenico Grasso2 and Barth F Smets1
1Environmental Engineering ProgramUniversity of Connecticut
Storrs CT 06269-20372Picker Engineering Program
Smith CollegeNorthampton MA 01063
ABSTRACT
Nitrification inhibition by the ethylenediamine-based chelating agents ethylenediaminetetraacetic acid(EDTA) diethylenetriaminepentaacetic acid (DTPA) and ethylenediamine (EDA) was evaluated in batchextant respirometric assays employing biomass from a continuously operated nitrifying enrichment reac-tor All three chelating agents inhibited ammonium oxidation but did not inhibit nitrite oxidation withinthe concentration range tested (0ndash3 mM) The order of inhibition (molar-based) was EDA EDTA DTPA The concentration causing 50 inhibition was estimated at 06 24 and 31 mM for EDA EDTAand DTPA respectively Inhibition by EDTA reached its ultimate value within 6 h whereas inhibition byEDA increased up to 6 h and then decreased Addition of multivalent cations (ie Fe31 Ca21 and Mg21)relieved inhibition by EDTA but not EDA Inhibition by both EDTA and DTPA but not EDA correlatedwith the depletion of cellular Ca21 Further inhibition by EDA correlated with substantial leakage of cel-lular K1 and disruption of cellular membrane integrity inferred from LIVEDEADreg Baclighttrade viabilityassays EDA appears to inhibit nitrifying activity by a different operative mechanism than EDTA andDTPA Finally the extant batch assays accurately predicted inhibition by EDTA in the continuous flowparent reactor suggesting the potential application of the batch assay to predict inhibition of nitrifying ac-tivity in full-scale biological treatment systems
Key words chelating agents nitrification inhibition EDTA DTPA ethylenediamine
219
Corresponding author Environmental Engineering Program University of Connecticut 261 Glenbrook Road CT 06269-2037 Phone 860-486-2270 Fax 860-486-2298 E-mail barthsmetsuconnedu
daggerCurrent address Metcalf amp Eddy 60 East 42 Street New York NY 10165
INTRODUCTION
AEROBIC NITRIFICATION IS MAINLY PERFORMED by twophysiologically distinct bacterial groups that carry
out NH41-N to NO2
2-N and NO22-N to NO3
2-N oxi-
dation respectively Nitrifying bacteria have low intrin-sic specific growth rates and are especially sensitive toenvironmental factors (eg pH temperature dissolvedoxygen) and toxic or inhibitory chemicals (Hockenburyand Grady 1977 Ford et al 1980 Shammas 1986) The
concentrations at which toxic substances inhibit nitrify-ing activity are often an order of magnitude lower thanthose required to inhibit heterotrophic activity in acti-vated sludge systems (Blum and Speece 1991) Hencenitrification is often the controlling step in biological ni-trogen removal (US EPA 1993 Daigger and Sadick1998)
Industrial waste streams containing chemical inhibi-tors commonly comprise varying portions of PubliclyOwned Treatment Works influent streams Chelatingagents which can complex physiologically essentialmetal cations via multidentate coordination are onechemical group that can inhibit nitrification For exam-ple allyl thiourea (ATU) is a chelating agent specificallyemployed as nitrification inhibitor with complete inhibi-tion observed at less than 5 mgL (Gernaey et al 1997Chandran and Smets 2000) Hydrocyanic acid (HCN)often found in the scrubber water of waste incineratorsis another strong nitrification inhibitor with 50 inhibi-tion in the range of 01 to 02 mgL (Daigger and Sadick1998) Both ATU and HCN are postulated to cause ni-trification inhibition because of Cu complexation as Cuis an essential part of the ammonia monooxygenase en-zyme in ammonium oxidizing bacteria (Painter 1970Vandevivere et al 1998)
Ethylenediamine-based chelating agents include ethyl-enediaminetetraacetic acid (EDTA) diethylenetriamine-pentaacetic acid (DTPA) and ethylenediamine (EDA) it-self (Fig 1) EDTA and DTPA are two of the most widelyused chelating agents in the pulp and paper photographicpharmaceutical and textile industries DTPA is a moreeffective chelator than EDTA at higher pH values(Bucheli-Witschel and Egli 2001 Sykora et al 2001)Their complexes are stable and have appeared to bepoorly biodegradable in wastewater treatment facilities(Alder et al 1990 Sykora et al 2001) ConsequentlyEDTA and DTPA have been found in the effluent of mu-nicipal wastewater treatment plants at concentrations ashigh as 05 mgL (Karl and Giger 1996 Bucheli-Witschel and Egli 2001) EDA itself is an industriallysignificant chemical used as an intermediate in the man-ufacture of EDTA herbicides (eg Komeenreg a copperndashethylenediamine complex) and various other chemicalproducts It is a polar highly water-soluble basic com-pound and appears biodegradable in wastewater treat-ment processes (Pietsch et al 2001)
Whereas the fate of chelating agents in the environ-ment especially in wastewater treatment facilities hasbeen investigated data on microbial inhibition by EDTAEDA and especially DTPA are sparse and certainly in-congruous with their industrial ubiquity (Painter 1970Hockenbury and Grady 1977) Moreover the mecha-nism of microbial inhibition is not well understood
Hence the objective of this study was to quantitativelyassess and elucidate the inhibitory effect of EDTADTPA and EDA on nitrifying activity
MATERIALS AND METHODS
Nitrifying bioreactor
Nitrifying biomass was cultivated in a continuouslystirred tank reactors (10 L) operated at solids retentiontime (SRT) of 20 days (wasted daily) and hydraulic re-tention time (HRT) of 1 day The reactor was fed an in-organic medium devoid of organic carbon with ammo-nium (300 mg NL (NH4)2SO4) as the sole energy sourcewith requisite macro- and micronutrients described else-where (Hu et al 2002a) Sodium carbonate (1 M) wasintermittently added to maintain a reactor pH of 74 601 and fulfilled both carbon and alkalinity requirementsFiltered laboratory air was provided to ensure adequatemixing and aeration Reactor performance was monitoredvia reactor and effluent COD effluent NH4
1-N NO22-N
and NO32-N measurements The reactor had been oper-
220 HU ET AL
Figure 1 Structural formulas of EDA EDTA and DTPA
ating for more than 2 years with its effluent concentra-tions of NH4
1-N NO22-N less than 01 mgL and NO3
2-N steady at 300 6 30 mgL Mixed liquor was periodi-cally withdrawn from the steady-state reactor and usedfor batch studies
Reagents
Stock solutions containing 50 mM EDTA (elec-trophoresis grade Fisher Scientific Fairlane NJ) DTPA(analytical grade Sigma-Aldrich St Louis MO) orEDA (analytical grade Sigma-Aldrich) were preparedwith Milli-Q purified water and the pH of EDTA orDTPA solution adjusted to 7 with 10 N NaOH prior touse From the stability constant information (Table 1) andequilibrium speciation calculation (MINEQL1 V 45)(Schecher and McAvoy 2001) the predominant forms(98) of these chelating agents in the test media at pH 75 were inferred as HEDTA32 HDTPA32 andH2EDA21
Nitrifier substrate oxidation activity
Maximum specific oxygen uptake rates (SOUR) weremeasured as surrogate of nitrifying activity in duplicateusing a batch extant respirometric assay described else-where (Chandran and Smets 2000 Hu et al 2002a)
The effects of chelating agents EDTA DTPA andEDA on nitrification kinetics were investigated sepa-rately in batch assays All batch inhibition assays wereconducted with a consistent chemical exposure time Thatis after the stock solutions containing chelating agentsin the absence or presence of coordination partner cationswere spiked into batch vessels a standard substrate oxi-dation assay ensued The actual time elapsed betweenchelator and complex spike and nitrogen substrate spikewas ca 15 min resulting in a total batch assay time of
approximately 60 min To mitigate the inhibition by theEDTA and EDA metals (as coordination partners) weremixed with EDTA or EDA stock solutions to yield finalconcentrations of Fe Cu Mg and Ca in the range of 0to 5 mM before they were spiked
To study the effect of chelator exposure time on nitri-fication inhibition reactor biomass samples were prein-cubated at room temperature (25 degC) and aerated by mag-netic stirring (100 rpm) with and without the chelatingagent addition At predetermined intervals (05 6 12 and24 h) aliquots were withdrawn and nitrification kineticswere determined Degree of inhibition was inferred from the difference in measured SOUR in the absence (SOURcontrol) vs the presence (SOURsample) of the testcompound (Hu et al 2002a)
Depletion of essential cellular cations
Biomass samples were removed from the continuousflow reactor supplemented with buffering capacity (20mM MOPS) pH adjusted to 75 6 005 and aliquots in-cubated for 30 min unamended (control) or spiked withEDTA DTPA or EDA (separately) to yield predeter-mined initial concentrations up to 3 mM Cellular cationdepletion was measured after earlier reports (Texier etal 2000 Hu et al 2003) Depletion was inferred fromthe difference in measured metal concentrations in theabsence vs the presence of the chelating agent
Bacterial viability
The fraction of nitrifying biomass with intact (viable)and damaged (dead) membranes were measured using aLIVEDEADreg Baclighttradeviability kit (Molecular ProbesEugene OR) and a FACS Calibur flow cytometer (Bec-ton Dickinson Mountain View CA) equipped with an Ar-gon-ion laser as described previously (Hu et al 2003)
NITRIFICATION INHIBITION BY ETHYLENEDIAMINE-BASED CHELATING AGENTS 221
ENVIRON ENG SCI VOL 20 NO 3 2003
Table 1 Stability constant (logK) for ethylenediamine-based chelating agents at ionic strength of 01 M and 25degC
Chelating agent
EDA EDTA DTPA
H1 HEDA1 99 HEDTA32 95 HDTPA32 99H2EDA21 170 H2EDTA22 156 H2DTPA22 183
H3EDTA2 183 H3DTPA2 226H4EDTA 203 H4DTPA 253H5EDTA1 218 H5DTPA1 273
Ca21 CaEDA21 01 CaEDTA22 108 CaDTPA22 108Mg21 MgEDA21 04 MgEDTA22 88 MgDTPA22 93Cu21 CuEDA21 105 CuEDTA22 188 CuDTPA22 212Fe21 FeEDA21 41 FeEDTA22 143 FeDTPA22 162Fe31 FeEDA31 NAa FeEDTA2 251b FeDTPA2 275b
All data are from Martel et al (1998) unless specified a not available bfrom Schowanek et al (1996)
Nitrifying cultures (20 mM MOPS pH 70 6 005)were incubated with individual chelating agents up to 2mM for 15 min Microbial cell pellets retained after cen-trifugation (1600 3 g 5 min) were resuspended in thesame volume of 01 mM NaCl followed by centrifuga-tion twice for 5 min at 1600 3 g Between each cen-trifugation the supernatant was decanted and pellets re-suspended in 01 mM NaCl Samples were prefilteredthrough 25-mm pore size filters (Calbiochem La JollaCA) to prevent clogging of the flow cytometer nozzleThe percentage of viable cells after the various chelatorspikes was then quantified using a constructed calibra-tion curve
EDTA shock load to continuous flow reactor
A shock load of EDTA was imposed by instanta-neously adding an aliquot of concentrated EDTA to thecontinuous flow reactor over a 5 min (50003HRT) in-terval Reactor or effluent aliquots were periodically col-lected to measure all nitrogen species as well as colloidaland soluble chemical oxygen demand (COD) The rou-tine batch respirometric assay was also performed withbiomass periodically removed during and after the shockload to determine the NH4
1 and NO22 oxidation kinet-
ics Floc size distribution was measured in duplicatewith an AccuSizer 770 Particle Optical Sizer (Santa Bar-bara CA) with resolution of 05ndash500 mm Each mea-surement lasted 30 s and employed a biomass volume of50 mL
Analytical procedures
Trace metal concentrations were measured accordingto standard methods (APHA et al 1998) Calcium ironmagnesium manganese and potassium concentrationswere measured by Inductively Coupled Plasma (ICP)spectroscopy (Optima 3300XL Perkin-Elmer Co Nor-walk CT) with method detection limit of 20 mgL foreach metal at the Environmental Research Institute (Uni-versity of Connecticut) Copper concentrations weremeasured via flame atomic absorption spectrometry(Model 5100 Perkin-Elmer Co with a method detectionlimit of 20 mgL Biomass concentrations were measuredas COD using commercially available reagents (HACHCOD vials 0ndash1500 mgL) Effluent concentrations ofcolloidal COD (0001ndash1 mm) were deduced from the dif-ference between the COD fraction less than 1 mm andthe truly soluble COD (measured after 05 mM LaCl3destabilization and subsequent 045-mm filtration) ac-cording to our earlier reported method (Hu et al 2002b)Effluent EDTA concentrations were approximated as ef-fluent truly soluble COD corrected for the preshock loadsoluble COD background (ca 20 mgL) NH4
1-N was
analyzed using an ammonia gas-sensing electrode (HNUSystems Newton MA) with an operating range from003 to 1400 mgL NH4
1-N NO22-N was measured col-
orimetrically with a method detection limit of 2 mgL andNO3
2-N via ion chromatography with MDL of 40 mgLall according to standard methods (APHA et al 1998)
Precautions were taken to avoid trace metal contami-nation Glassware and plastic ware when appropriatewere soaked in 1 M HNO3 overnight and rinsed with 5volumes of Milli-Q purified water before use
RESULTS AND DISCUSSION
Nitrification kinetics and dosendashresponse curves
In the short-term batch assays (about 1 h) the specificammonium oxidation rate (SOURNH4) decreased as theapplied EDTA or DTPA dose to the nitrifying biomassincreased whereas the specific nitrite oxidation rate(SOURNO2) was not adversely affected within the con-centration range tested (data not shown) A similar inhi-bition pattern was observed for EDA (data not shown)The higher sensitivity of ammonium oxidation comparedto nitrite oxidation is consistent with earlier observations(Hockenbury and Grady 1977 Blum and Speece 1991)From the SOUR data dosendashinhibitory response curvesfor ammonium oxidation were developed for the chelat-ing agents (Fig 2)
Ethylenediamine was inhibitory even at the lowest
222 HU ET AL
Figure 2 Dosendashresponse curves of chelating agent inhibitionto nitrifying cultures (1033 6 112 mgL COD pH 75) EDTA(d ) DTPA ( s ) EDA ( j ) Error bars indicate 1 standard de-viation (n 5 2)
concentrations tested The EC50 (defined as the concen-tration of chelating agent resulting in 50 inhibition) ofEDA was ca 06 mM This value is consistent with anearlier reported 61 inhibition of ammonia oxidation at05mM (Hockenbury and Grady 1977) EDTA has usu-ally been considered noninhibitory to ammonium oxida-tion at concentration less than approximately 1 mM(Tomlinson et al 1966 Painter 1970) However whenEDTA concentrations were higher than 15 mM signif-icant inhibition was observed in this work DTPA on theother hand has been reported to be more toxic thanEDTA in standard aquatic toxicity tests with fish inver-tebrates and algae (Schowanek et al 1996) In our ex-periments with nitrifying bacteria however DTPA wasfound to be less inhibitory than EDTA (Fig 2)
Overall under the same aquatic chemistry conditionsand employing nitrifying biomass with the same culture
NITRIFICATION INHIBITION BY ETHYLENEDIAMINE-BASED CHELATING AGENTS 223
ENVIRON ENG SCI VOL 20 NO 3 2003
Figure 3 Correlation of the inhibition to nitrifying cultures (1074 6 34 mgL COD pH 75) with the depletion of cellular es-sential elements (a) 5 S K 1 Ca 1 Mg 1 Mn 1 Fe 1 Cu (b) Ca (c) Mg (d) Cu Employed chelating agents were EDTA (d )DTPA ( s ) EDA (j ) The initial cellular concentrations were K 0112 6 0007 Ca 0032 6 0006 Mg 0022 6 0002 Mn0076 6 0002 Fe 0080 6 0003 Cu 0046 6 0001 mMg biomass COD (n 5 2) The error bars indicate 1 standard deviation
history the molar-based inhibitory character to ammo-nium oxidation followed EDA EDTA DTPAThe EC50 defined as the concentration of chelating agentcausing 50 inhibition was estimated at 06 24 and31 mM for EDA EDTA and DTPA respectively Theinhibitory character of tested chelating agents did not par-allel but in fact was opposite to their relative com-plexation potential for key multivalent cations (DTPA EDTA EDA) (Table 1) This surprising finding led usto investigate how cellular cation depletion by differentchelating agents compared
Depletion of essential cellular cation
Although C H O and N constitute the bulk of a bac-terial cellrsquos weight many other elements (K Ca Mg MnFe and Cu) are required in macro or microamounts The
K Ca Mg Mn Fe and Cu fraction for the nitrifying en-richment culture was measured at 049 6 003 014 6003 006 6 000 047 6 001 051 6 002 and 033 6001 dry weight respectively (n 5 2) with our prede-termined conversion factor 121 6 009 g biomass CODgbiomass DW (n 5 3) applied
After chelating agents were added to the nitrifying bio-mass substantial depletion (22) of essential cellularcations was observed (Fig 3a) The maximum percent ofdepletion of each cation was 21 67 25 15 21 and 64for K Ca Mg Mn Fe and Cu respectively Inhibitionby EDTA or DTPA correlated well with the depletion ofcellular Ca21 (presumably linear correlation R2 5 082)(Fig 3b) In contrast depletion of Ca21 by EDA was sig-nificantly less for the same extent of inhibition The in-hibition by all three chelating agents correlated well with
the depletion of cellular Mg21 (R2 5 093) (Fig 3c) al-though the maximum amount of Mg21 depletion wasmuch lower (ca 43) than that of Ca21 Finally nitrifi-cation inhibition did not correlate with the depletion ofcellular Cu21(R2 5 044) (Fig 3d) Similarly inhibitionwas not well correlated with the depletion of K Mn orFe (data not shown)
No effort was made to differentiate between the mem-brane-associated fraction and the intracellular fraction ofthe measured essential cellular cations However fromearlier experiments with Escherichia coli it had beenconcluded that EDTA does not bind to the cell surfaceor cross the cytoplasmic membrane (Ferris and Bev-eridge 1986) In most bacteria there would be no EDTAtransport across the membranes (Bucheli-Witschel andEgli 2001) and uncomplexed EDTA can destabilize the
224 HU ET AL
Figure 4 The depletion of cellular (a) Ca21 (b) Mg21 and (c) K1 (mMg XCOD) as a function of molar dose of chelationagents EDTA ( d ) DTPA ( s ) EDA ( j ) Conditions are the same as indicated in Fig 3
cell surface of Gram-negative bacteria by interactionswith essential cellular cations on the surface (Ferris1989) EDTA especially scavenges Ca21 which is pre-dominantly responsible for outer membrane stabilizationThis can cause a concomitant release of significantamount of lipopolysaccharide (Leive 1974 Ferris1989) The inhibition of ammonium oxidizing activity byEDTA observed in this work is probably attributed to asimilar loss of essential cations from the outer membranebecause ammonium oxidizing bacteria exhibit the typi-cal Gram-negative cell membrane structure and correla-tions between the extent of inhibition and cellular Ca21
and Mg21 depletion were observedDTPA has a similar chemical structure to and a higher
molecular weight than EDTA By analogy one can spec-ulate that its transport across cellular membranes is in-significant The mode of inhibition of ammonium oxi-dizing activity by DTPA observed in this work isprobably similar to EDTA as the strong correlations be-tween the extent of inhibition and cellular Ca21 andMg21 depletion existed for both EDTA and DTPA (Fig3b and c)
Based on equilibrium speciation calculations everymol of chelating agents will complex 1 mol of Ca21 orMg21 at pH 75 At the same molar concentration EDAresulted in less cellular Ca21 depletion (Fig 4a) but muchhigher inhibition than EDTA or DTPA (Fig 2) Secondat the same molar concentration EDA resulted in morecellular Mg21 depletion than EDTA or DTPA (Fig 4b)yet the stability constant of MgEDA (1004) is muchsmaller than that of Mg EDTA (1088) or MgDTPA (1093)(Table 1) The results with EDA suggest that cellularMg21 depletion by EDA did not derive primarily fromMgndashEDA complexation Instead it is speculated due toleakage of destabilized cell membrane as EDA resultedin substantially higher cellular K1 depletion than EDTAor DTPA (Fig 4c) K1 was the most abundant cellularcation (on a molar basis data not shown) and is gener-ally believed to play an important role in maintenance ofthe ionic environment in the cytoplasm Depletion of cel-lular K1 indicates a loss of cell membrane integrity andviability (Ohsumi et al 1988 Avery et al 1996 Howlettand Avery 1997a)
Bacterial viability
Because depletion of cellular K1 has been widely usedas a marker of plasma membrane disruption (Howlett andAvery 1997b) we investigated directly whether disrup-tion of the plasma membrane integrity was observed af-ter exposure to the different agents In the presence of01 mM EDA 745 6 31 cells remained viable after15-min exposure At 2 mM EDA less than 113 6 00cells were viable compared to 875 6 111 461 6
108 viable cells in the presence of similar concentra-tions of EDTA and DTPA The biological effect of EDAtherefore is different among the chelators studied andmay involve cytoplasmic membrane rupture The ob-served loss of the plasma membrane integrity by 2 mMDTPA addition was unexpectedly high in view of the un-affected nitrifying activity at the same concentration (Fig2) Further experiments are needed to better understandwhat factors are responsible for the inconsistency be-tween the changes in the measured cellular activity(SOUR) and viability (livedead staining) after DTPAexposure
NITRIFICATION INHIBITION BY ETHYLENEDIAMINE-BASED CHELATING AGENTS 225
ENVIRON ENG SCI VOL 20 NO 3 2003
Figure 5 Inhibition of NH41-N oxidation by (a) 25 mM
EDTA or (b) 025 mM EDA in presence of metal ions Cu21
(d ) Fe31 ( s ) Ca21 ( j ) Mg21 ( u ) Bold horizontal lines rep-resent mean percent of inhibition in the presence of (a) EDTAor (b) EDA without metal addition
Inhibition mitigation by complexation
Appropriate remedial strategies to relieve nitrificationinhibition by chelatina agents were subsequently ex-plored Simultaneous addition of metals as coordinationpartners reduced the biological effects of chelatingagents but actual trends were ligand and metal specific(Fig 5)
At a 1 mM Fe31 dose (as FeCl3) nitrification inhibi-tion by 25 mM EDTA was completely relieved (Fig 5a)Similarly addition of Ca21 (as CaCl2) or Mg21 (as
MgSO4) reduced nitrification inhibition However Ca21
and Mg21 were slightly less effective than Fe31 on a mo-lar basis This may derive from EDTArsquos ability to formstronger complexes with the Lewis acid Fe compared toCa or Mg (Table 1) On the other hand addition of Cu21
(as CuCl2) only partially reduced nitrification inhibitionat a 11 ratio of CuEDTA (predominant speciesCuEDTA22) Copper itself inhibited the nitrifying activ-ity at high concentrations
In contrast nitrification inhibition by 025 mM EDAwas not relieved at any Fe31 dose tested (Fig 5b) Sim-ilarly addition of Ca21 or Mg21 did not reduce nitrifi-cation inhibition at the highest dose applied (5 mM)which is in agreement with their weak complexation po-tential (CaEDA K 5 1001 MgEDA K 5 1004) Theweak complexation behavior of EDA by Mg21 and thehigh depletion of cellular Mg21 by EDA further sug-gests that EDA has a different mode of inhibition thanEDTA Again addition of Cu21 only partially reducedthe inhibition with Cu(EDA)2
21 as the predominantspecies according to equilibrium speciation calcula-tions
Effect of exposure time on nitrification inhibition
To quantify the effect of exposure time on nitrifica-tion inhibition by chelating agents a predetermined
226 HU ET AL
Figure 7 Distribution of EDTA ( d ) in a continuous flowbioreactor after a transient dose of 24 mM EDTA within min-utes The continuous line presents the prediction from the ex-ponential decay model where C C0 are the reactor EDTA con-centrations at time t (day) and zero respectively F is the feedflow rate to the continuous flow reactor (10 Lday) and V isthe reactor volume (10 L)
Figure 6 Effect of exposure time on nitrification inhibitionby (a) EDTA or (b) EDA in batch assays Initial chelating agentconcentrations are shown Inhibition is expressed with refer-ence of SOUR of a culture incubated without chelating agentsto same duration after removal from parent reactor
dose of EDTA or EDA was applied in the batch reac-tor and the extent of inhibition measured as a functionof time Nitrification inhibition by EDTA reached its ul-timate value within 6 h (paired t-test p 025) (Fig6a) whereas inhibition by EDA substantially increasedup to 6 h (paired t-test p 005) and then decreased(Fig 6b) Such discrepancy between EDTA and EDAfurther suggests different modes of action for EDTA andEDA It also suggests that changes of the intracellularcation pool by EDA may have been slow analogous toslow kinetics of metal internalization reported else-where (Hu et al 2003)
Nitrification inhibition by EDTA in thecontinuous flow parent reactor
A shock load of EDTA was imposed to the continu-ously operated parent nitrifying bioreactor at 24 mM aconcentration corresponding to 47 6 2 inhibition in thebatch assay An ideal continuous flow reactor model ad-equately described the effluent EDTA (complexed anduncomplexed) profile (R2 5 099) computed from the ef-fluent COD data corrected for prepulse concentrations(Fig 7) which indicates that EDTA essentially behavedas a conservative compound did not partition to the cellsurface and was not subject to significant decay (Ferrisand Beveridge 1986 Sykora et al 2001)
Substantial foaming in the reactor was observed andlasted for 1 day after the EDTA shock load Recovery ofthe biological activity in the reactor was rapid At 01day reactor EDTA concentration dropped to 19 mM and23 6 1 inhibition of ammonium oxidation was mea-sured consistent with the extent of inhibition predictedfrom the batch assay (Fig 2) Effluent concentrations (1day after the shock load) indicated a return to stable operation (NH4
1-N NO22-N less than 01 mgL and
NO32-N ca 300 mgL)
Concomitantly reactor colloidal COD increasedslightly (from 6 to 23 6 6 mgL COD) after EDTA shockloads and returned to background level after 2 days Bio-mass floc size remained unchanged at 40 (62) mm (num-ber weighted mean floc size) from 5-day measurementsafter EDTA load compared to 41 (61) mm before dose
Implications for full-scale biological nitrogenremoval operation
The batch extant respirometric assay employed in thiswork can be used to ensure that the microbial physiol-ogy and activity remain close to that in the sampled con-tinuous flow reactor (Ellis et al 1996 Grady et al1996) From the study of EDTA in the continuous flowreactor the measured SOUR and subsequent inhibitionwas indeed consistent with the predicted value from the
batch assay No further inhibition was noticed and therecovery of the nitrifying activity in the continuous flowreactor was rapid Hence at least for compounds likeEDTA (and presumably DTPA) batch extant respiro-metric assays may accurately portray the nitrifying ac-tivity and inhibition in the continuous flow reactors
Although we only observed nitrification inhibition inboth batch and continuous flow reactors by EDTA whenits concentration was higher than 15 mM caution shouldbe employed in directly extending these results to full-scalewastewater treatment plants Inhibition may be primarilydependent on the concentration of uncomplexed EDTAspecies (Bucheli-Witschel and Egli 2001) which amongother physicochemical factors is often mediated by aque-ous Ca21 and Mg21 concentrations For instance equilib-rium speciation calculation revealed that the concentrationof uncomplexed [EDTA42] in our reactor was 10255 mMat 15 mM total EDTA The same uncomplexed EDTAconcentration could be obtained at 07 mM total EDTA attypical wastewater conditions of 16 mgL Ca21 and 10mgL Mg21 (Tchobanoglous and Burton 1991)
ACKNOWLEDGMENTS
We are grateful to Prof Diane J Burgess in the Dept ofPharmacy and Dr Michele Barber in the Dept of Molec-ular and Cell Biology at the University of Connecticut forassistance with particle sizing and flow cytometry respec-tively This study was funded in part by the ConnecticutDepartment of Environmental Protection through a USEPA Long Island Sound Study grant under Section 119 ofthe Clean Water Act and by the Connecticut Institute ofWater Resources under the 104B program of the NationalInstitutes of Water Resources US Geological Survey asauthorized by the Water Research and Development Act of1978 (PL 95-467) contract number 01HQGR0078
REFERENCES
ALDER AC SIEGRIST H GUJER W and GIGGER W(1990) Behavior of NTA and EDTA in biological waste-water treatment Water Res 24 733ndash742
APHA AWWA and WEF (1998) Standard Methods for theExamination of Water and Wastewater 20th ed Washing-ton DC author
AVERY SV HOWLETT NG and RADICE S (1996)Copper toxicity towards Saccharomyces cerevisiae Depen-dence on plasma membrane fatty acid composition Appl En-viron Microbiol 62 3960ndash3966
BLUM DJW and SPEECE RE (1991) A database ofchemical toxicity to environmental bacteria and its use in in-
NITRIFICATION INHIBITION BY ETHYLENEDIAMINE-BASED CHELATING AGENTS 227
ENVIRON ENG SCI VOL 20 NO 3 2003
terspecies comparisons and correlations J Water PollutControl Fed 63 198ndash207
BUCHELI-WITSCHEL M and EGLI T (2001) Environ-mental fate and microbial degradation of aminopolycar-boxylic acids FEMS Microbiol Rev 25 69ndash106
CHANDRAN K and SMETS BF (2000) Single-step nitri-fication models erroneously describe batch ammonia oxida-tion profiles when nitrite oxidation becomes rate limitingBiotechnol Bioeng 68 396ndash406
DAIGGER GT and SADICK TE (1998) Evaluation ofmethods to detect and control nitrification inhibition withspecific application to incinerator flue-gas scrubber waterWater Environ Res 70 1248ndash1257
ELLIS TG BARBEAU DS SMETS BF and GRADYLCP (1996) Respirometric technique for determination ofextant kinetic parameters describing biodegradation WaterEnviron Res 68 917ndash926
FERRIS FG (1989) Metallic ion interactions with outer mem-brane of Gram-negative bacteria In TJ Beveridge and RJDoyle Eds Metal Ions and Bacteria New York John Wi-ley and Sons pp 295ndash323
FERRIS FG and BEVERIDGE TJ (1986) Physicochemi-cal roles of soluble metal cations in the outer membrane ofEscherichia coli K-12 Can J Microbiol 32 594ndash601
FORD DL CHURCHWELL RL and KACHTICK JW(1980) Comprehensive analysis of nitrification of chemicalprocessing wastewaters J Water Pollut Control Fed 522726ndash2736
GERNAEY K VERSCHUERE L LUYTEN L and VER-STRAETE W (1997) Fast and sensitive acute toxicity de-tection with an enrichment nitrifying culture Water EnvironRes 69 1163ndash1169
GRADY CPL JR SMETS BF and BARBEAU DS (1996)Variability in kinetic parameter estimates A review of possiblecauses and a proposed terminology Water Res 30 742ndash748
HOCKENBURY MR and GRADY CPL (1977) Inhibi-tion of nitrification-effects of selected organic compoundsJ Water Pollut Control Fed 49 768ndash777
HOWLETT NG and AVERY SV (1997a) Induction oflipid peroxidation during heavy metal stress in Saccha-romyces cerevisiae and influence of plasma membrane fattyacid unsaturation Appl Environ Microbiol 63 2971ndash2976
HOWLETT NG and AVERY SV (1997b) Relationship be-tween cadmium sensitivity and degree of plasma membranefatty acid unsaturation in Saccharomyces cerevisiae ApplMicrobiol Biotechnol 48 539ndash545
HU Z CHANDRAN K GRASSO D and SMETS BF(2002a) Effect of nickel and cadmium speciation on nitrifi-cation inhibition Environ Sci Technol 36 3074ndash3078
HU Z CHADRAN K GRASSO D and SMETS BF(2003) Impact of heavy metal sorption and internalization onnitrification inhibition Environ Sci Technol 37 728ndash734
HU Z CHANDRAN K SMETS BF and GRASSO D(2002b) Evaluation of a rapid physical-chemical method forthe determination of extant soluble COD Water Res 36617ndash624
KARI FG and GIGER W (1996) Speciation and fate of ethylenediaminetetraacetate (EDTA) in municipal waste-water treatment Water Res 30 122ndash134
LEIVE L (1974) The barrier function of Gram-negative en-velope Ann NY Acad Sci 235 109ndash127
MARTEL AE SMITH RM and MOTEKAITIS RJ(1998) NIST Critical Stability Constants of Metal ComplexesDatabase College Station TX Texas AampM Univ
OHSUMI Y KITAMOTO K and ANRAKU Y (1988)Changes induced in the permeability barrier of the yeastplasma membrane by cupric ion J Bacteriol 170 2676ndash2682
PAINTER HA (1970) A review of literature on inorganic nitrogen metabolism in microorganisms Water Res 4 393ndash450
PIETSCH J SACHER F SCHMIDT W and BRAUCHH-J (2001) Polar nitrogen compounds and their behaviorin the drinking water treatment process Water Res 35 3537ndash3544
SCHECHER WD and MCAVOY DC (2001) MINEQL1A Chemical Equilibrium Modeling System Version 45 forWindows Hallowell ME Environmental Research Software
SCHOWANEK D MCAVOY D VERSTEEG D andHANSTVEIT A (1996) Effects of nutrient trace metal spe-ciation on algal growth in the presence of the chelator [SS]-EDDS Aquatic Toxicol 36 253ndash275
SHAMMAS NK (1986) Interactions of temperature pH andbiomass on the nitrification process J Water Pollut Con-trol Fed 58 52ndash58
SYKORA V PITTER P BITTNEROVA I and LEDERERT (2001) Biodegradability of ethylenediamine-based com-plexing agents Water Res 35 2010ndash2016
TCHOBANOGLOUS G and BURTON FL (1991) Waste-water Engineering Treatment Disposal and Reuse 3rd edNew York Metcalf amp Eddy McGraw-Hill
TEXIER A ANDRES Y ILLEMASSENE M andCLOIREC PL (2000) Characterization of lanthanide ionsbinding sites in the cell wall of Pseudomonos aeruginosaEnviron Sci Technol 34 610ndash615
TOMLINSON TG BOON AG and TROTMAN CNA(1966) Inhibition of nitrification in the activated sludge pro-cess of sewage disposal J Appl Bacteriol 29 266ndash291
US EPA (1993) Nitrogen control In EPA625R-93-0101Washington DC Office of Water
VANDEVIVERE P FICARA E TERRAS C JULIES Eand VERSTRAETE W (1998) Copper-mediated selectiveremoval of nitrification inhibitors from industrial waste-waters Environ Sci Technol 32 1000ndash1006
228 HU ET AL

concentrations at which toxic substances inhibit nitrify-ing activity are often an order of magnitude lower thanthose required to inhibit heterotrophic activity in acti-vated sludge systems (Blum and Speece 1991) Hencenitrification is often the controlling step in biological ni-trogen removal (US EPA 1993 Daigger and Sadick1998)
Industrial waste streams containing chemical inhibi-tors commonly comprise varying portions of PubliclyOwned Treatment Works influent streams Chelatingagents which can complex physiologically essentialmetal cations via multidentate coordination are onechemical group that can inhibit nitrification For exam-ple allyl thiourea (ATU) is a chelating agent specificallyemployed as nitrification inhibitor with complete inhibi-tion observed at less than 5 mgL (Gernaey et al 1997Chandran and Smets 2000) Hydrocyanic acid (HCN)often found in the scrubber water of waste incineratorsis another strong nitrification inhibitor with 50 inhibi-tion in the range of 01 to 02 mgL (Daigger and Sadick1998) Both ATU and HCN are postulated to cause ni-trification inhibition because of Cu complexation as Cuis an essential part of the ammonia monooxygenase en-zyme in ammonium oxidizing bacteria (Painter 1970Vandevivere et al 1998)
Ethylenediamine-based chelating agents include ethyl-enediaminetetraacetic acid (EDTA) diethylenetriamine-pentaacetic acid (DTPA) and ethylenediamine (EDA) it-self (Fig 1) EDTA and DTPA are two of the most widelyused chelating agents in the pulp and paper photographicpharmaceutical and textile industries DTPA is a moreeffective chelator than EDTA at higher pH values(Bucheli-Witschel and Egli 2001 Sykora et al 2001)Their complexes are stable and have appeared to bepoorly biodegradable in wastewater treatment facilities(Alder et al 1990 Sykora et al 2001) ConsequentlyEDTA and DTPA have been found in the effluent of mu-nicipal wastewater treatment plants at concentrations ashigh as 05 mgL (Karl and Giger 1996 Bucheli-Witschel and Egli 2001) EDA itself is an industriallysignificant chemical used as an intermediate in the man-ufacture of EDTA herbicides (eg Komeenreg a copperndashethylenediamine complex) and various other chemicalproducts It is a polar highly water-soluble basic com-pound and appears biodegradable in wastewater treat-ment processes (Pietsch et al 2001)
Whereas the fate of chelating agents in the environ-ment especially in wastewater treatment facilities hasbeen investigated data on microbial inhibition by EDTAEDA and especially DTPA are sparse and certainly in-congruous with their industrial ubiquity (Painter 1970Hockenbury and Grady 1977) Moreover the mecha-nism of microbial inhibition is not well understood
Hence the objective of this study was to quantitativelyassess and elucidate the inhibitory effect of EDTADTPA and EDA on nitrifying activity
MATERIALS AND METHODS
Nitrifying bioreactor
Nitrifying biomass was cultivated in a continuouslystirred tank reactors (10 L) operated at solids retentiontime (SRT) of 20 days (wasted daily) and hydraulic re-tention time (HRT) of 1 day The reactor was fed an in-organic medium devoid of organic carbon with ammo-nium (300 mg NL (NH4)2SO4) as the sole energy sourcewith requisite macro- and micronutrients described else-where (Hu et al 2002a) Sodium carbonate (1 M) wasintermittently added to maintain a reactor pH of 74 601 and fulfilled both carbon and alkalinity requirementsFiltered laboratory air was provided to ensure adequatemixing and aeration Reactor performance was monitoredvia reactor and effluent COD effluent NH4
1-N NO22-N
and NO32-N measurements The reactor had been oper-
220 HU ET AL
Figure 1 Structural formulas of EDA EDTA and DTPA
ating for more than 2 years with its effluent concentra-tions of NH4
1-N NO22-N less than 01 mgL and NO3
2-N steady at 300 6 30 mgL Mixed liquor was periodi-cally withdrawn from the steady-state reactor and usedfor batch studies
Reagents
Stock solutions containing 50 mM EDTA (elec-trophoresis grade Fisher Scientific Fairlane NJ) DTPA(analytical grade Sigma-Aldrich St Louis MO) orEDA (analytical grade Sigma-Aldrich) were preparedwith Milli-Q purified water and the pH of EDTA orDTPA solution adjusted to 7 with 10 N NaOH prior touse From the stability constant information (Table 1) andequilibrium speciation calculation (MINEQL1 V 45)(Schecher and McAvoy 2001) the predominant forms(98) of these chelating agents in the test media at pH 75 were inferred as HEDTA32 HDTPA32 andH2EDA21
Nitrifier substrate oxidation activity
Maximum specific oxygen uptake rates (SOUR) weremeasured as surrogate of nitrifying activity in duplicateusing a batch extant respirometric assay described else-where (Chandran and Smets 2000 Hu et al 2002a)
The effects of chelating agents EDTA DTPA andEDA on nitrification kinetics were investigated sepa-rately in batch assays All batch inhibition assays wereconducted with a consistent chemical exposure time Thatis after the stock solutions containing chelating agentsin the absence or presence of coordination partner cationswere spiked into batch vessels a standard substrate oxi-dation assay ensued The actual time elapsed betweenchelator and complex spike and nitrogen substrate spikewas ca 15 min resulting in a total batch assay time of
approximately 60 min To mitigate the inhibition by theEDTA and EDA metals (as coordination partners) weremixed with EDTA or EDA stock solutions to yield finalconcentrations of Fe Cu Mg and Ca in the range of 0to 5 mM before they were spiked
To study the effect of chelator exposure time on nitri-fication inhibition reactor biomass samples were prein-cubated at room temperature (25 degC) and aerated by mag-netic stirring (100 rpm) with and without the chelatingagent addition At predetermined intervals (05 6 12 and24 h) aliquots were withdrawn and nitrification kineticswere determined Degree of inhibition was inferred from the difference in measured SOUR in the absence (SOURcontrol) vs the presence (SOURsample) of the testcompound (Hu et al 2002a)
Depletion of essential cellular cations
Biomass samples were removed from the continuousflow reactor supplemented with buffering capacity (20mM MOPS) pH adjusted to 75 6 005 and aliquots in-cubated for 30 min unamended (control) or spiked withEDTA DTPA or EDA (separately) to yield predeter-mined initial concentrations up to 3 mM Cellular cationdepletion was measured after earlier reports (Texier etal 2000 Hu et al 2003) Depletion was inferred fromthe difference in measured metal concentrations in theabsence vs the presence of the chelating agent
Bacterial viability
The fraction of nitrifying biomass with intact (viable)and damaged (dead) membranes were measured using aLIVEDEADreg Baclighttradeviability kit (Molecular ProbesEugene OR) and a FACS Calibur flow cytometer (Bec-ton Dickinson Mountain View CA) equipped with an Ar-gon-ion laser as described previously (Hu et al 2003)
NITRIFICATION INHIBITION BY ETHYLENEDIAMINE-BASED CHELATING AGENTS 221
ENVIRON ENG SCI VOL 20 NO 3 2003
Table 1 Stability constant (logK) for ethylenediamine-based chelating agents at ionic strength of 01 M and 25degC
Chelating agent
EDA EDTA DTPA
H1 HEDA1 99 HEDTA32 95 HDTPA32 99H2EDA21 170 H2EDTA22 156 H2DTPA22 183
H3EDTA2 183 H3DTPA2 226H4EDTA 203 H4DTPA 253H5EDTA1 218 H5DTPA1 273
Ca21 CaEDA21 01 CaEDTA22 108 CaDTPA22 108Mg21 MgEDA21 04 MgEDTA22 88 MgDTPA22 93Cu21 CuEDA21 105 CuEDTA22 188 CuDTPA22 212Fe21 FeEDA21 41 FeEDTA22 143 FeDTPA22 162Fe31 FeEDA31 NAa FeEDTA2 251b FeDTPA2 275b
All data are from Martel et al (1998) unless specified a not available bfrom Schowanek et al (1996)
Nitrifying cultures (20 mM MOPS pH 70 6 005)were incubated with individual chelating agents up to 2mM for 15 min Microbial cell pellets retained after cen-trifugation (1600 3 g 5 min) were resuspended in thesame volume of 01 mM NaCl followed by centrifuga-tion twice for 5 min at 1600 3 g Between each cen-trifugation the supernatant was decanted and pellets re-suspended in 01 mM NaCl Samples were prefilteredthrough 25-mm pore size filters (Calbiochem La JollaCA) to prevent clogging of the flow cytometer nozzleThe percentage of viable cells after the various chelatorspikes was then quantified using a constructed calibra-tion curve
EDTA shock load to continuous flow reactor
A shock load of EDTA was imposed by instanta-neously adding an aliquot of concentrated EDTA to thecontinuous flow reactor over a 5 min (50003HRT) in-terval Reactor or effluent aliquots were periodically col-lected to measure all nitrogen species as well as colloidaland soluble chemical oxygen demand (COD) The rou-tine batch respirometric assay was also performed withbiomass periodically removed during and after the shockload to determine the NH4
1 and NO22 oxidation kinet-
ics Floc size distribution was measured in duplicatewith an AccuSizer 770 Particle Optical Sizer (Santa Bar-bara CA) with resolution of 05ndash500 mm Each mea-surement lasted 30 s and employed a biomass volume of50 mL
Analytical procedures
Trace metal concentrations were measured accordingto standard methods (APHA et al 1998) Calcium ironmagnesium manganese and potassium concentrationswere measured by Inductively Coupled Plasma (ICP)spectroscopy (Optima 3300XL Perkin-Elmer Co Nor-walk CT) with method detection limit of 20 mgL foreach metal at the Environmental Research Institute (Uni-versity of Connecticut) Copper concentrations weremeasured via flame atomic absorption spectrometry(Model 5100 Perkin-Elmer Co with a method detectionlimit of 20 mgL Biomass concentrations were measuredas COD using commercially available reagents (HACHCOD vials 0ndash1500 mgL) Effluent concentrations ofcolloidal COD (0001ndash1 mm) were deduced from the dif-ference between the COD fraction less than 1 mm andthe truly soluble COD (measured after 05 mM LaCl3destabilization and subsequent 045-mm filtration) ac-cording to our earlier reported method (Hu et al 2002b)Effluent EDTA concentrations were approximated as ef-fluent truly soluble COD corrected for the preshock loadsoluble COD background (ca 20 mgL) NH4
1-N was
analyzed using an ammonia gas-sensing electrode (HNUSystems Newton MA) with an operating range from003 to 1400 mgL NH4
1-N NO22-N was measured col-
orimetrically with a method detection limit of 2 mgL andNO3
2-N via ion chromatography with MDL of 40 mgLall according to standard methods (APHA et al 1998)
Precautions were taken to avoid trace metal contami-nation Glassware and plastic ware when appropriatewere soaked in 1 M HNO3 overnight and rinsed with 5volumes of Milli-Q purified water before use
RESULTS AND DISCUSSION
Nitrification kinetics and dosendashresponse curves
In the short-term batch assays (about 1 h) the specificammonium oxidation rate (SOURNH4) decreased as theapplied EDTA or DTPA dose to the nitrifying biomassincreased whereas the specific nitrite oxidation rate(SOURNO2) was not adversely affected within the con-centration range tested (data not shown) A similar inhi-bition pattern was observed for EDA (data not shown)The higher sensitivity of ammonium oxidation comparedto nitrite oxidation is consistent with earlier observations(Hockenbury and Grady 1977 Blum and Speece 1991)From the SOUR data dosendashinhibitory response curvesfor ammonium oxidation were developed for the chelat-ing agents (Fig 2)
Ethylenediamine was inhibitory even at the lowest
222 HU ET AL
Figure 2 Dosendashresponse curves of chelating agent inhibitionto nitrifying cultures (1033 6 112 mgL COD pH 75) EDTA(d ) DTPA ( s ) EDA ( j ) Error bars indicate 1 standard de-viation (n 5 2)
concentrations tested The EC50 (defined as the concen-tration of chelating agent resulting in 50 inhibition) ofEDA was ca 06 mM This value is consistent with anearlier reported 61 inhibition of ammonia oxidation at05mM (Hockenbury and Grady 1977) EDTA has usu-ally been considered noninhibitory to ammonium oxida-tion at concentration less than approximately 1 mM(Tomlinson et al 1966 Painter 1970) However whenEDTA concentrations were higher than 15 mM signif-icant inhibition was observed in this work DTPA on theother hand has been reported to be more toxic thanEDTA in standard aquatic toxicity tests with fish inver-tebrates and algae (Schowanek et al 1996) In our ex-periments with nitrifying bacteria however DTPA wasfound to be less inhibitory than EDTA (Fig 2)
Overall under the same aquatic chemistry conditionsand employing nitrifying biomass with the same culture
NITRIFICATION INHIBITION BY ETHYLENEDIAMINE-BASED CHELATING AGENTS 223
ENVIRON ENG SCI VOL 20 NO 3 2003
Figure 3 Correlation of the inhibition to nitrifying cultures (1074 6 34 mgL COD pH 75) with the depletion of cellular es-sential elements (a) 5 S K 1 Ca 1 Mg 1 Mn 1 Fe 1 Cu (b) Ca (c) Mg (d) Cu Employed chelating agents were EDTA (d )DTPA ( s ) EDA (j ) The initial cellular concentrations were K 0112 6 0007 Ca 0032 6 0006 Mg 0022 6 0002 Mn0076 6 0002 Fe 0080 6 0003 Cu 0046 6 0001 mMg biomass COD (n 5 2) The error bars indicate 1 standard deviation
history the molar-based inhibitory character to ammo-nium oxidation followed EDA EDTA DTPAThe EC50 defined as the concentration of chelating agentcausing 50 inhibition was estimated at 06 24 and31 mM for EDA EDTA and DTPA respectively Theinhibitory character of tested chelating agents did not par-allel but in fact was opposite to their relative com-plexation potential for key multivalent cations (DTPA EDTA EDA) (Table 1) This surprising finding led usto investigate how cellular cation depletion by differentchelating agents compared
Depletion of essential cellular cation
Although C H O and N constitute the bulk of a bac-terial cellrsquos weight many other elements (K Ca Mg MnFe and Cu) are required in macro or microamounts The
K Ca Mg Mn Fe and Cu fraction for the nitrifying en-richment culture was measured at 049 6 003 014 6003 006 6 000 047 6 001 051 6 002 and 033 6001 dry weight respectively (n 5 2) with our prede-termined conversion factor 121 6 009 g biomass CODgbiomass DW (n 5 3) applied
After chelating agents were added to the nitrifying bio-mass substantial depletion (22) of essential cellularcations was observed (Fig 3a) The maximum percent ofdepletion of each cation was 21 67 25 15 21 and 64for K Ca Mg Mn Fe and Cu respectively Inhibitionby EDTA or DTPA correlated well with the depletion ofcellular Ca21 (presumably linear correlation R2 5 082)(Fig 3b) In contrast depletion of Ca21 by EDA was sig-nificantly less for the same extent of inhibition The in-hibition by all three chelating agents correlated well with
the depletion of cellular Mg21 (R2 5 093) (Fig 3c) al-though the maximum amount of Mg21 depletion wasmuch lower (ca 43) than that of Ca21 Finally nitrifi-cation inhibition did not correlate with the depletion ofcellular Cu21(R2 5 044) (Fig 3d) Similarly inhibitionwas not well correlated with the depletion of K Mn orFe (data not shown)
No effort was made to differentiate between the mem-brane-associated fraction and the intracellular fraction ofthe measured essential cellular cations However fromearlier experiments with Escherichia coli it had beenconcluded that EDTA does not bind to the cell surfaceor cross the cytoplasmic membrane (Ferris and Bev-eridge 1986) In most bacteria there would be no EDTAtransport across the membranes (Bucheli-Witschel andEgli 2001) and uncomplexed EDTA can destabilize the
224 HU ET AL
Figure 4 The depletion of cellular (a) Ca21 (b) Mg21 and (c) K1 (mMg XCOD) as a function of molar dose of chelationagents EDTA ( d ) DTPA ( s ) EDA ( j ) Conditions are the same as indicated in Fig 3
cell surface of Gram-negative bacteria by interactionswith essential cellular cations on the surface (Ferris1989) EDTA especially scavenges Ca21 which is pre-dominantly responsible for outer membrane stabilizationThis can cause a concomitant release of significantamount of lipopolysaccharide (Leive 1974 Ferris1989) The inhibition of ammonium oxidizing activity byEDTA observed in this work is probably attributed to asimilar loss of essential cations from the outer membranebecause ammonium oxidizing bacteria exhibit the typi-cal Gram-negative cell membrane structure and correla-tions between the extent of inhibition and cellular Ca21
and Mg21 depletion were observedDTPA has a similar chemical structure to and a higher
molecular weight than EDTA By analogy one can spec-ulate that its transport across cellular membranes is in-significant The mode of inhibition of ammonium oxi-dizing activity by DTPA observed in this work isprobably similar to EDTA as the strong correlations be-tween the extent of inhibition and cellular Ca21 andMg21 depletion existed for both EDTA and DTPA (Fig3b and c)
Based on equilibrium speciation calculations everymol of chelating agents will complex 1 mol of Ca21 orMg21 at pH 75 At the same molar concentration EDAresulted in less cellular Ca21 depletion (Fig 4a) but muchhigher inhibition than EDTA or DTPA (Fig 2) Secondat the same molar concentration EDA resulted in morecellular Mg21 depletion than EDTA or DTPA (Fig 4b)yet the stability constant of MgEDA (1004) is muchsmaller than that of Mg EDTA (1088) or MgDTPA (1093)(Table 1) The results with EDA suggest that cellularMg21 depletion by EDA did not derive primarily fromMgndashEDA complexation Instead it is speculated due toleakage of destabilized cell membrane as EDA resultedin substantially higher cellular K1 depletion than EDTAor DTPA (Fig 4c) K1 was the most abundant cellularcation (on a molar basis data not shown) and is gener-ally believed to play an important role in maintenance ofthe ionic environment in the cytoplasm Depletion of cel-lular K1 indicates a loss of cell membrane integrity andviability (Ohsumi et al 1988 Avery et al 1996 Howlettand Avery 1997a)
Bacterial viability
Because depletion of cellular K1 has been widely usedas a marker of plasma membrane disruption (Howlett andAvery 1997b) we investigated directly whether disrup-tion of the plasma membrane integrity was observed af-ter exposure to the different agents In the presence of01 mM EDA 745 6 31 cells remained viable after15-min exposure At 2 mM EDA less than 113 6 00cells were viable compared to 875 6 111 461 6
108 viable cells in the presence of similar concentra-tions of EDTA and DTPA The biological effect of EDAtherefore is different among the chelators studied andmay involve cytoplasmic membrane rupture The ob-served loss of the plasma membrane integrity by 2 mMDTPA addition was unexpectedly high in view of the un-affected nitrifying activity at the same concentration (Fig2) Further experiments are needed to better understandwhat factors are responsible for the inconsistency be-tween the changes in the measured cellular activity(SOUR) and viability (livedead staining) after DTPAexposure
NITRIFICATION INHIBITION BY ETHYLENEDIAMINE-BASED CHELATING AGENTS 225
ENVIRON ENG SCI VOL 20 NO 3 2003
Figure 5 Inhibition of NH41-N oxidation by (a) 25 mM
EDTA or (b) 025 mM EDA in presence of metal ions Cu21
(d ) Fe31 ( s ) Ca21 ( j ) Mg21 ( u ) Bold horizontal lines rep-resent mean percent of inhibition in the presence of (a) EDTAor (b) EDA without metal addition
Inhibition mitigation by complexation
Appropriate remedial strategies to relieve nitrificationinhibition by chelatina agents were subsequently ex-plored Simultaneous addition of metals as coordinationpartners reduced the biological effects of chelatingagents but actual trends were ligand and metal specific(Fig 5)
At a 1 mM Fe31 dose (as FeCl3) nitrification inhibi-tion by 25 mM EDTA was completely relieved (Fig 5a)Similarly addition of Ca21 (as CaCl2) or Mg21 (as
MgSO4) reduced nitrification inhibition However Ca21
and Mg21 were slightly less effective than Fe31 on a mo-lar basis This may derive from EDTArsquos ability to formstronger complexes with the Lewis acid Fe compared toCa or Mg (Table 1) On the other hand addition of Cu21
(as CuCl2) only partially reduced nitrification inhibitionat a 11 ratio of CuEDTA (predominant speciesCuEDTA22) Copper itself inhibited the nitrifying activ-ity at high concentrations
In contrast nitrification inhibition by 025 mM EDAwas not relieved at any Fe31 dose tested (Fig 5b) Sim-ilarly addition of Ca21 or Mg21 did not reduce nitrifi-cation inhibition at the highest dose applied (5 mM)which is in agreement with their weak complexation po-tential (CaEDA K 5 1001 MgEDA K 5 1004) Theweak complexation behavior of EDA by Mg21 and thehigh depletion of cellular Mg21 by EDA further sug-gests that EDA has a different mode of inhibition thanEDTA Again addition of Cu21 only partially reducedthe inhibition with Cu(EDA)2
21 as the predominantspecies according to equilibrium speciation calcula-tions
Effect of exposure time on nitrification inhibition
To quantify the effect of exposure time on nitrifica-tion inhibition by chelating agents a predetermined
226 HU ET AL
Figure 7 Distribution of EDTA ( d ) in a continuous flowbioreactor after a transient dose of 24 mM EDTA within min-utes The continuous line presents the prediction from the ex-ponential decay model where C C0 are the reactor EDTA con-centrations at time t (day) and zero respectively F is the feedflow rate to the continuous flow reactor (10 Lday) and V isthe reactor volume (10 L)
Figure 6 Effect of exposure time on nitrification inhibitionby (a) EDTA or (b) EDA in batch assays Initial chelating agentconcentrations are shown Inhibition is expressed with refer-ence of SOUR of a culture incubated without chelating agentsto same duration after removal from parent reactor
dose of EDTA or EDA was applied in the batch reac-tor and the extent of inhibition measured as a functionof time Nitrification inhibition by EDTA reached its ul-timate value within 6 h (paired t-test p 025) (Fig6a) whereas inhibition by EDA substantially increasedup to 6 h (paired t-test p 005) and then decreased(Fig 6b) Such discrepancy between EDTA and EDAfurther suggests different modes of action for EDTA andEDA It also suggests that changes of the intracellularcation pool by EDA may have been slow analogous toslow kinetics of metal internalization reported else-where (Hu et al 2003)
Nitrification inhibition by EDTA in thecontinuous flow parent reactor
A shock load of EDTA was imposed to the continu-ously operated parent nitrifying bioreactor at 24 mM aconcentration corresponding to 47 6 2 inhibition in thebatch assay An ideal continuous flow reactor model ad-equately described the effluent EDTA (complexed anduncomplexed) profile (R2 5 099) computed from the ef-fluent COD data corrected for prepulse concentrations(Fig 7) which indicates that EDTA essentially behavedas a conservative compound did not partition to the cellsurface and was not subject to significant decay (Ferrisand Beveridge 1986 Sykora et al 2001)
Substantial foaming in the reactor was observed andlasted for 1 day after the EDTA shock load Recovery ofthe biological activity in the reactor was rapid At 01day reactor EDTA concentration dropped to 19 mM and23 6 1 inhibition of ammonium oxidation was mea-sured consistent with the extent of inhibition predictedfrom the batch assay (Fig 2) Effluent concentrations (1day after the shock load) indicated a return to stable operation (NH4
1-N NO22-N less than 01 mgL and
NO32-N ca 300 mgL)
Concomitantly reactor colloidal COD increasedslightly (from 6 to 23 6 6 mgL COD) after EDTA shockloads and returned to background level after 2 days Bio-mass floc size remained unchanged at 40 (62) mm (num-ber weighted mean floc size) from 5-day measurementsafter EDTA load compared to 41 (61) mm before dose
Implications for full-scale biological nitrogenremoval operation
The batch extant respirometric assay employed in thiswork can be used to ensure that the microbial physiol-ogy and activity remain close to that in the sampled con-tinuous flow reactor (Ellis et al 1996 Grady et al1996) From the study of EDTA in the continuous flowreactor the measured SOUR and subsequent inhibitionwas indeed consistent with the predicted value from the
batch assay No further inhibition was noticed and therecovery of the nitrifying activity in the continuous flowreactor was rapid Hence at least for compounds likeEDTA (and presumably DTPA) batch extant respiro-metric assays may accurately portray the nitrifying ac-tivity and inhibition in the continuous flow reactors
Although we only observed nitrification inhibition inboth batch and continuous flow reactors by EDTA whenits concentration was higher than 15 mM caution shouldbe employed in directly extending these results to full-scalewastewater treatment plants Inhibition may be primarilydependent on the concentration of uncomplexed EDTAspecies (Bucheli-Witschel and Egli 2001) which amongother physicochemical factors is often mediated by aque-ous Ca21 and Mg21 concentrations For instance equilib-rium speciation calculation revealed that the concentrationof uncomplexed [EDTA42] in our reactor was 10255 mMat 15 mM total EDTA The same uncomplexed EDTAconcentration could be obtained at 07 mM total EDTA attypical wastewater conditions of 16 mgL Ca21 and 10mgL Mg21 (Tchobanoglous and Burton 1991)
ACKNOWLEDGMENTS
We are grateful to Prof Diane J Burgess in the Dept ofPharmacy and Dr Michele Barber in the Dept of Molec-ular and Cell Biology at the University of Connecticut forassistance with particle sizing and flow cytometry respec-tively This study was funded in part by the ConnecticutDepartment of Environmental Protection through a USEPA Long Island Sound Study grant under Section 119 ofthe Clean Water Act and by the Connecticut Institute ofWater Resources under the 104B program of the NationalInstitutes of Water Resources US Geological Survey asauthorized by the Water Research and Development Act of1978 (PL 95-467) contract number 01HQGR0078
REFERENCES
ALDER AC SIEGRIST H GUJER W and GIGGER W(1990) Behavior of NTA and EDTA in biological waste-water treatment Water Res 24 733ndash742
APHA AWWA and WEF (1998) Standard Methods for theExamination of Water and Wastewater 20th ed Washing-ton DC author
AVERY SV HOWLETT NG and RADICE S (1996)Copper toxicity towards Saccharomyces cerevisiae Depen-dence on plasma membrane fatty acid composition Appl En-viron Microbiol 62 3960ndash3966
BLUM DJW and SPEECE RE (1991) A database ofchemical toxicity to environmental bacteria and its use in in-
NITRIFICATION INHIBITION BY ETHYLENEDIAMINE-BASED CHELATING AGENTS 227
ENVIRON ENG SCI VOL 20 NO 3 2003
terspecies comparisons and correlations J Water PollutControl Fed 63 198ndash207
BUCHELI-WITSCHEL M and EGLI T (2001) Environ-mental fate and microbial degradation of aminopolycar-boxylic acids FEMS Microbiol Rev 25 69ndash106
CHANDRAN K and SMETS BF (2000) Single-step nitri-fication models erroneously describe batch ammonia oxida-tion profiles when nitrite oxidation becomes rate limitingBiotechnol Bioeng 68 396ndash406
DAIGGER GT and SADICK TE (1998) Evaluation ofmethods to detect and control nitrification inhibition withspecific application to incinerator flue-gas scrubber waterWater Environ Res 70 1248ndash1257
ELLIS TG BARBEAU DS SMETS BF and GRADYLCP (1996) Respirometric technique for determination ofextant kinetic parameters describing biodegradation WaterEnviron Res 68 917ndash926
FERRIS FG (1989) Metallic ion interactions with outer mem-brane of Gram-negative bacteria In TJ Beveridge and RJDoyle Eds Metal Ions and Bacteria New York John Wi-ley and Sons pp 295ndash323
FERRIS FG and BEVERIDGE TJ (1986) Physicochemi-cal roles of soluble metal cations in the outer membrane ofEscherichia coli K-12 Can J Microbiol 32 594ndash601
FORD DL CHURCHWELL RL and KACHTICK JW(1980) Comprehensive analysis of nitrification of chemicalprocessing wastewaters J Water Pollut Control Fed 522726ndash2736
GERNAEY K VERSCHUERE L LUYTEN L and VER-STRAETE W (1997) Fast and sensitive acute toxicity de-tection with an enrichment nitrifying culture Water EnvironRes 69 1163ndash1169
GRADY CPL JR SMETS BF and BARBEAU DS (1996)Variability in kinetic parameter estimates A review of possiblecauses and a proposed terminology Water Res 30 742ndash748
HOCKENBURY MR and GRADY CPL (1977) Inhibi-tion of nitrification-effects of selected organic compoundsJ Water Pollut Control Fed 49 768ndash777
HOWLETT NG and AVERY SV (1997a) Induction oflipid peroxidation during heavy metal stress in Saccha-romyces cerevisiae and influence of plasma membrane fattyacid unsaturation Appl Environ Microbiol 63 2971ndash2976
HOWLETT NG and AVERY SV (1997b) Relationship be-tween cadmium sensitivity and degree of plasma membranefatty acid unsaturation in Saccharomyces cerevisiae ApplMicrobiol Biotechnol 48 539ndash545
HU Z CHANDRAN K GRASSO D and SMETS BF(2002a) Effect of nickel and cadmium speciation on nitrifi-cation inhibition Environ Sci Technol 36 3074ndash3078
HU Z CHADRAN K GRASSO D and SMETS BF(2003) Impact of heavy metal sorption and internalization onnitrification inhibition Environ Sci Technol 37 728ndash734
HU Z CHANDRAN K SMETS BF and GRASSO D(2002b) Evaluation of a rapid physical-chemical method forthe determination of extant soluble COD Water Res 36617ndash624
KARI FG and GIGER W (1996) Speciation and fate of ethylenediaminetetraacetate (EDTA) in municipal waste-water treatment Water Res 30 122ndash134
LEIVE L (1974) The barrier function of Gram-negative en-velope Ann NY Acad Sci 235 109ndash127
MARTEL AE SMITH RM and MOTEKAITIS RJ(1998) NIST Critical Stability Constants of Metal ComplexesDatabase College Station TX Texas AampM Univ
OHSUMI Y KITAMOTO K and ANRAKU Y (1988)Changes induced in the permeability barrier of the yeastplasma membrane by cupric ion J Bacteriol 170 2676ndash2682
PAINTER HA (1970) A review of literature on inorganic nitrogen metabolism in microorganisms Water Res 4 393ndash450
PIETSCH J SACHER F SCHMIDT W and BRAUCHH-J (2001) Polar nitrogen compounds and their behaviorin the drinking water treatment process Water Res 35 3537ndash3544
SCHECHER WD and MCAVOY DC (2001) MINEQL1A Chemical Equilibrium Modeling System Version 45 forWindows Hallowell ME Environmental Research Software
SCHOWANEK D MCAVOY D VERSTEEG D andHANSTVEIT A (1996) Effects of nutrient trace metal spe-ciation on algal growth in the presence of the chelator [SS]-EDDS Aquatic Toxicol 36 253ndash275
SHAMMAS NK (1986) Interactions of temperature pH andbiomass on the nitrification process J Water Pollut Con-trol Fed 58 52ndash58
SYKORA V PITTER P BITTNEROVA I and LEDERERT (2001) Biodegradability of ethylenediamine-based com-plexing agents Water Res 35 2010ndash2016
TCHOBANOGLOUS G and BURTON FL (1991) Waste-water Engineering Treatment Disposal and Reuse 3rd edNew York Metcalf amp Eddy McGraw-Hill
TEXIER A ANDRES Y ILLEMASSENE M andCLOIREC PL (2000) Characterization of lanthanide ionsbinding sites in the cell wall of Pseudomonos aeruginosaEnviron Sci Technol 34 610ndash615
TOMLINSON TG BOON AG and TROTMAN CNA(1966) Inhibition of nitrification in the activated sludge pro-cess of sewage disposal J Appl Bacteriol 29 266ndash291
US EPA (1993) Nitrogen control In EPA625R-93-0101Washington DC Office of Water
VANDEVIVERE P FICARA E TERRAS C JULIES Eand VERSTRAETE W (1998) Copper-mediated selectiveremoval of nitrification inhibitors from industrial waste-waters Environ Sci Technol 32 1000ndash1006
228 HU ET AL

ating for more than 2 years with its effluent concentra-tions of NH4
1-N NO22-N less than 01 mgL and NO3
2-N steady at 300 6 30 mgL Mixed liquor was periodi-cally withdrawn from the steady-state reactor and usedfor batch studies
Reagents
Stock solutions containing 50 mM EDTA (elec-trophoresis grade Fisher Scientific Fairlane NJ) DTPA(analytical grade Sigma-Aldrich St Louis MO) orEDA (analytical grade Sigma-Aldrich) were preparedwith Milli-Q purified water and the pH of EDTA orDTPA solution adjusted to 7 with 10 N NaOH prior touse From the stability constant information (Table 1) andequilibrium speciation calculation (MINEQL1 V 45)(Schecher and McAvoy 2001) the predominant forms(98) of these chelating agents in the test media at pH 75 were inferred as HEDTA32 HDTPA32 andH2EDA21
Nitrifier substrate oxidation activity
Maximum specific oxygen uptake rates (SOUR) weremeasured as surrogate of nitrifying activity in duplicateusing a batch extant respirometric assay described else-where (Chandran and Smets 2000 Hu et al 2002a)
The effects of chelating agents EDTA DTPA andEDA on nitrification kinetics were investigated sepa-rately in batch assays All batch inhibition assays wereconducted with a consistent chemical exposure time Thatis after the stock solutions containing chelating agentsin the absence or presence of coordination partner cationswere spiked into batch vessels a standard substrate oxi-dation assay ensued The actual time elapsed betweenchelator and complex spike and nitrogen substrate spikewas ca 15 min resulting in a total batch assay time of
approximately 60 min To mitigate the inhibition by theEDTA and EDA metals (as coordination partners) weremixed with EDTA or EDA stock solutions to yield finalconcentrations of Fe Cu Mg and Ca in the range of 0to 5 mM before they were spiked
To study the effect of chelator exposure time on nitri-fication inhibition reactor biomass samples were prein-cubated at room temperature (25 degC) and aerated by mag-netic stirring (100 rpm) with and without the chelatingagent addition At predetermined intervals (05 6 12 and24 h) aliquots were withdrawn and nitrification kineticswere determined Degree of inhibition was inferred from the difference in measured SOUR in the absence (SOURcontrol) vs the presence (SOURsample) of the testcompound (Hu et al 2002a)
Depletion of essential cellular cations
Biomass samples were removed from the continuousflow reactor supplemented with buffering capacity (20mM MOPS) pH adjusted to 75 6 005 and aliquots in-cubated for 30 min unamended (control) or spiked withEDTA DTPA or EDA (separately) to yield predeter-mined initial concentrations up to 3 mM Cellular cationdepletion was measured after earlier reports (Texier etal 2000 Hu et al 2003) Depletion was inferred fromthe difference in measured metal concentrations in theabsence vs the presence of the chelating agent
Bacterial viability
The fraction of nitrifying biomass with intact (viable)and damaged (dead) membranes were measured using aLIVEDEADreg Baclighttradeviability kit (Molecular ProbesEugene OR) and a FACS Calibur flow cytometer (Bec-ton Dickinson Mountain View CA) equipped with an Ar-gon-ion laser as described previously (Hu et al 2003)
NITRIFICATION INHIBITION BY ETHYLENEDIAMINE-BASED CHELATING AGENTS 221
ENVIRON ENG SCI VOL 20 NO 3 2003
Table 1 Stability constant (logK) for ethylenediamine-based chelating agents at ionic strength of 01 M and 25degC
Chelating agent
EDA EDTA DTPA
H1 HEDA1 99 HEDTA32 95 HDTPA32 99H2EDA21 170 H2EDTA22 156 H2DTPA22 183
H3EDTA2 183 H3DTPA2 226H4EDTA 203 H4DTPA 253H5EDTA1 218 H5DTPA1 273
Ca21 CaEDA21 01 CaEDTA22 108 CaDTPA22 108Mg21 MgEDA21 04 MgEDTA22 88 MgDTPA22 93Cu21 CuEDA21 105 CuEDTA22 188 CuDTPA22 212Fe21 FeEDA21 41 FeEDTA22 143 FeDTPA22 162Fe31 FeEDA31 NAa FeEDTA2 251b FeDTPA2 275b
All data are from Martel et al (1998) unless specified a not available bfrom Schowanek et al (1996)
Nitrifying cultures (20 mM MOPS pH 70 6 005)were incubated with individual chelating agents up to 2mM for 15 min Microbial cell pellets retained after cen-trifugation (1600 3 g 5 min) were resuspended in thesame volume of 01 mM NaCl followed by centrifuga-tion twice for 5 min at 1600 3 g Between each cen-trifugation the supernatant was decanted and pellets re-suspended in 01 mM NaCl Samples were prefilteredthrough 25-mm pore size filters (Calbiochem La JollaCA) to prevent clogging of the flow cytometer nozzleThe percentage of viable cells after the various chelatorspikes was then quantified using a constructed calibra-tion curve
EDTA shock load to continuous flow reactor
A shock load of EDTA was imposed by instanta-neously adding an aliquot of concentrated EDTA to thecontinuous flow reactor over a 5 min (50003HRT) in-terval Reactor or effluent aliquots were periodically col-lected to measure all nitrogen species as well as colloidaland soluble chemical oxygen demand (COD) The rou-tine batch respirometric assay was also performed withbiomass periodically removed during and after the shockload to determine the NH4
1 and NO22 oxidation kinet-
ics Floc size distribution was measured in duplicatewith an AccuSizer 770 Particle Optical Sizer (Santa Bar-bara CA) with resolution of 05ndash500 mm Each mea-surement lasted 30 s and employed a biomass volume of50 mL
Analytical procedures
Trace metal concentrations were measured accordingto standard methods (APHA et al 1998) Calcium ironmagnesium manganese and potassium concentrationswere measured by Inductively Coupled Plasma (ICP)spectroscopy (Optima 3300XL Perkin-Elmer Co Nor-walk CT) with method detection limit of 20 mgL foreach metal at the Environmental Research Institute (Uni-versity of Connecticut) Copper concentrations weremeasured via flame atomic absorption spectrometry(Model 5100 Perkin-Elmer Co with a method detectionlimit of 20 mgL Biomass concentrations were measuredas COD using commercially available reagents (HACHCOD vials 0ndash1500 mgL) Effluent concentrations ofcolloidal COD (0001ndash1 mm) were deduced from the dif-ference between the COD fraction less than 1 mm andthe truly soluble COD (measured after 05 mM LaCl3destabilization and subsequent 045-mm filtration) ac-cording to our earlier reported method (Hu et al 2002b)Effluent EDTA concentrations were approximated as ef-fluent truly soluble COD corrected for the preshock loadsoluble COD background (ca 20 mgL) NH4
1-N was
analyzed using an ammonia gas-sensing electrode (HNUSystems Newton MA) with an operating range from003 to 1400 mgL NH4
1-N NO22-N was measured col-
orimetrically with a method detection limit of 2 mgL andNO3
2-N via ion chromatography with MDL of 40 mgLall according to standard methods (APHA et al 1998)
Precautions were taken to avoid trace metal contami-nation Glassware and plastic ware when appropriatewere soaked in 1 M HNO3 overnight and rinsed with 5volumes of Milli-Q purified water before use
RESULTS AND DISCUSSION
Nitrification kinetics and dosendashresponse curves
In the short-term batch assays (about 1 h) the specificammonium oxidation rate (SOURNH4) decreased as theapplied EDTA or DTPA dose to the nitrifying biomassincreased whereas the specific nitrite oxidation rate(SOURNO2) was not adversely affected within the con-centration range tested (data not shown) A similar inhi-bition pattern was observed for EDA (data not shown)The higher sensitivity of ammonium oxidation comparedto nitrite oxidation is consistent with earlier observations(Hockenbury and Grady 1977 Blum and Speece 1991)From the SOUR data dosendashinhibitory response curvesfor ammonium oxidation were developed for the chelat-ing agents (Fig 2)
Ethylenediamine was inhibitory even at the lowest
222 HU ET AL
Figure 2 Dosendashresponse curves of chelating agent inhibitionto nitrifying cultures (1033 6 112 mgL COD pH 75) EDTA(d ) DTPA ( s ) EDA ( j ) Error bars indicate 1 standard de-viation (n 5 2)
concentrations tested The EC50 (defined as the concen-tration of chelating agent resulting in 50 inhibition) ofEDA was ca 06 mM This value is consistent with anearlier reported 61 inhibition of ammonia oxidation at05mM (Hockenbury and Grady 1977) EDTA has usu-ally been considered noninhibitory to ammonium oxida-tion at concentration less than approximately 1 mM(Tomlinson et al 1966 Painter 1970) However whenEDTA concentrations were higher than 15 mM signif-icant inhibition was observed in this work DTPA on theother hand has been reported to be more toxic thanEDTA in standard aquatic toxicity tests with fish inver-tebrates and algae (Schowanek et al 1996) In our ex-periments with nitrifying bacteria however DTPA wasfound to be less inhibitory than EDTA (Fig 2)
Overall under the same aquatic chemistry conditionsand employing nitrifying biomass with the same culture
NITRIFICATION INHIBITION BY ETHYLENEDIAMINE-BASED CHELATING AGENTS 223
ENVIRON ENG SCI VOL 20 NO 3 2003
Figure 3 Correlation of the inhibition to nitrifying cultures (1074 6 34 mgL COD pH 75) with the depletion of cellular es-sential elements (a) 5 S K 1 Ca 1 Mg 1 Mn 1 Fe 1 Cu (b) Ca (c) Mg (d) Cu Employed chelating agents were EDTA (d )DTPA ( s ) EDA (j ) The initial cellular concentrations were K 0112 6 0007 Ca 0032 6 0006 Mg 0022 6 0002 Mn0076 6 0002 Fe 0080 6 0003 Cu 0046 6 0001 mMg biomass COD (n 5 2) The error bars indicate 1 standard deviation
history the molar-based inhibitory character to ammo-nium oxidation followed EDA EDTA DTPAThe EC50 defined as the concentration of chelating agentcausing 50 inhibition was estimated at 06 24 and31 mM for EDA EDTA and DTPA respectively Theinhibitory character of tested chelating agents did not par-allel but in fact was opposite to their relative com-plexation potential for key multivalent cations (DTPA EDTA EDA) (Table 1) This surprising finding led usto investigate how cellular cation depletion by differentchelating agents compared
Depletion of essential cellular cation
Although C H O and N constitute the bulk of a bac-terial cellrsquos weight many other elements (K Ca Mg MnFe and Cu) are required in macro or microamounts The
K Ca Mg Mn Fe and Cu fraction for the nitrifying en-richment culture was measured at 049 6 003 014 6003 006 6 000 047 6 001 051 6 002 and 033 6001 dry weight respectively (n 5 2) with our prede-termined conversion factor 121 6 009 g biomass CODgbiomass DW (n 5 3) applied
After chelating agents were added to the nitrifying bio-mass substantial depletion (22) of essential cellularcations was observed (Fig 3a) The maximum percent ofdepletion of each cation was 21 67 25 15 21 and 64for K Ca Mg Mn Fe and Cu respectively Inhibitionby EDTA or DTPA correlated well with the depletion ofcellular Ca21 (presumably linear correlation R2 5 082)(Fig 3b) In contrast depletion of Ca21 by EDA was sig-nificantly less for the same extent of inhibition The in-hibition by all three chelating agents correlated well with
the depletion of cellular Mg21 (R2 5 093) (Fig 3c) al-though the maximum amount of Mg21 depletion wasmuch lower (ca 43) than that of Ca21 Finally nitrifi-cation inhibition did not correlate with the depletion ofcellular Cu21(R2 5 044) (Fig 3d) Similarly inhibitionwas not well correlated with the depletion of K Mn orFe (data not shown)
No effort was made to differentiate between the mem-brane-associated fraction and the intracellular fraction ofthe measured essential cellular cations However fromearlier experiments with Escherichia coli it had beenconcluded that EDTA does not bind to the cell surfaceor cross the cytoplasmic membrane (Ferris and Bev-eridge 1986) In most bacteria there would be no EDTAtransport across the membranes (Bucheli-Witschel andEgli 2001) and uncomplexed EDTA can destabilize the
224 HU ET AL
Figure 4 The depletion of cellular (a) Ca21 (b) Mg21 and (c) K1 (mMg XCOD) as a function of molar dose of chelationagents EDTA ( d ) DTPA ( s ) EDA ( j ) Conditions are the same as indicated in Fig 3
cell surface of Gram-negative bacteria by interactionswith essential cellular cations on the surface (Ferris1989) EDTA especially scavenges Ca21 which is pre-dominantly responsible for outer membrane stabilizationThis can cause a concomitant release of significantamount of lipopolysaccharide (Leive 1974 Ferris1989) The inhibition of ammonium oxidizing activity byEDTA observed in this work is probably attributed to asimilar loss of essential cations from the outer membranebecause ammonium oxidizing bacteria exhibit the typi-cal Gram-negative cell membrane structure and correla-tions between the extent of inhibition and cellular Ca21
and Mg21 depletion were observedDTPA has a similar chemical structure to and a higher
molecular weight than EDTA By analogy one can spec-ulate that its transport across cellular membranes is in-significant The mode of inhibition of ammonium oxi-dizing activity by DTPA observed in this work isprobably similar to EDTA as the strong correlations be-tween the extent of inhibition and cellular Ca21 andMg21 depletion existed for both EDTA and DTPA (Fig3b and c)
Based on equilibrium speciation calculations everymol of chelating agents will complex 1 mol of Ca21 orMg21 at pH 75 At the same molar concentration EDAresulted in less cellular Ca21 depletion (Fig 4a) but muchhigher inhibition than EDTA or DTPA (Fig 2) Secondat the same molar concentration EDA resulted in morecellular Mg21 depletion than EDTA or DTPA (Fig 4b)yet the stability constant of MgEDA (1004) is muchsmaller than that of Mg EDTA (1088) or MgDTPA (1093)(Table 1) The results with EDA suggest that cellularMg21 depletion by EDA did not derive primarily fromMgndashEDA complexation Instead it is speculated due toleakage of destabilized cell membrane as EDA resultedin substantially higher cellular K1 depletion than EDTAor DTPA (Fig 4c) K1 was the most abundant cellularcation (on a molar basis data not shown) and is gener-ally believed to play an important role in maintenance ofthe ionic environment in the cytoplasm Depletion of cel-lular K1 indicates a loss of cell membrane integrity andviability (Ohsumi et al 1988 Avery et al 1996 Howlettand Avery 1997a)
Bacterial viability
Because depletion of cellular K1 has been widely usedas a marker of plasma membrane disruption (Howlett andAvery 1997b) we investigated directly whether disrup-tion of the plasma membrane integrity was observed af-ter exposure to the different agents In the presence of01 mM EDA 745 6 31 cells remained viable after15-min exposure At 2 mM EDA less than 113 6 00cells were viable compared to 875 6 111 461 6
108 viable cells in the presence of similar concentra-tions of EDTA and DTPA The biological effect of EDAtherefore is different among the chelators studied andmay involve cytoplasmic membrane rupture The ob-served loss of the plasma membrane integrity by 2 mMDTPA addition was unexpectedly high in view of the un-affected nitrifying activity at the same concentration (Fig2) Further experiments are needed to better understandwhat factors are responsible for the inconsistency be-tween the changes in the measured cellular activity(SOUR) and viability (livedead staining) after DTPAexposure
NITRIFICATION INHIBITION BY ETHYLENEDIAMINE-BASED CHELATING AGENTS 225
ENVIRON ENG SCI VOL 20 NO 3 2003
Figure 5 Inhibition of NH41-N oxidation by (a) 25 mM
EDTA or (b) 025 mM EDA in presence of metal ions Cu21
(d ) Fe31 ( s ) Ca21 ( j ) Mg21 ( u ) Bold horizontal lines rep-resent mean percent of inhibition in the presence of (a) EDTAor (b) EDA without metal addition
Inhibition mitigation by complexation
Appropriate remedial strategies to relieve nitrificationinhibition by chelatina agents were subsequently ex-plored Simultaneous addition of metals as coordinationpartners reduced the biological effects of chelatingagents but actual trends were ligand and metal specific(Fig 5)
At a 1 mM Fe31 dose (as FeCl3) nitrification inhibi-tion by 25 mM EDTA was completely relieved (Fig 5a)Similarly addition of Ca21 (as CaCl2) or Mg21 (as
MgSO4) reduced nitrification inhibition However Ca21
and Mg21 were slightly less effective than Fe31 on a mo-lar basis This may derive from EDTArsquos ability to formstronger complexes with the Lewis acid Fe compared toCa or Mg (Table 1) On the other hand addition of Cu21
(as CuCl2) only partially reduced nitrification inhibitionat a 11 ratio of CuEDTA (predominant speciesCuEDTA22) Copper itself inhibited the nitrifying activ-ity at high concentrations
In contrast nitrification inhibition by 025 mM EDAwas not relieved at any Fe31 dose tested (Fig 5b) Sim-ilarly addition of Ca21 or Mg21 did not reduce nitrifi-cation inhibition at the highest dose applied (5 mM)which is in agreement with their weak complexation po-tential (CaEDA K 5 1001 MgEDA K 5 1004) Theweak complexation behavior of EDA by Mg21 and thehigh depletion of cellular Mg21 by EDA further sug-gests that EDA has a different mode of inhibition thanEDTA Again addition of Cu21 only partially reducedthe inhibition with Cu(EDA)2
21 as the predominantspecies according to equilibrium speciation calcula-tions
Effect of exposure time on nitrification inhibition
To quantify the effect of exposure time on nitrifica-tion inhibition by chelating agents a predetermined
226 HU ET AL
Figure 7 Distribution of EDTA ( d ) in a continuous flowbioreactor after a transient dose of 24 mM EDTA within min-utes The continuous line presents the prediction from the ex-ponential decay model where C C0 are the reactor EDTA con-centrations at time t (day) and zero respectively F is the feedflow rate to the continuous flow reactor (10 Lday) and V isthe reactor volume (10 L)
Figure 6 Effect of exposure time on nitrification inhibitionby (a) EDTA or (b) EDA in batch assays Initial chelating agentconcentrations are shown Inhibition is expressed with refer-ence of SOUR of a culture incubated without chelating agentsto same duration after removal from parent reactor
dose of EDTA or EDA was applied in the batch reac-tor and the extent of inhibition measured as a functionof time Nitrification inhibition by EDTA reached its ul-timate value within 6 h (paired t-test p 025) (Fig6a) whereas inhibition by EDA substantially increasedup to 6 h (paired t-test p 005) and then decreased(Fig 6b) Such discrepancy between EDTA and EDAfurther suggests different modes of action for EDTA andEDA It also suggests that changes of the intracellularcation pool by EDA may have been slow analogous toslow kinetics of metal internalization reported else-where (Hu et al 2003)
Nitrification inhibition by EDTA in thecontinuous flow parent reactor
A shock load of EDTA was imposed to the continu-ously operated parent nitrifying bioreactor at 24 mM aconcentration corresponding to 47 6 2 inhibition in thebatch assay An ideal continuous flow reactor model ad-equately described the effluent EDTA (complexed anduncomplexed) profile (R2 5 099) computed from the ef-fluent COD data corrected for prepulse concentrations(Fig 7) which indicates that EDTA essentially behavedas a conservative compound did not partition to the cellsurface and was not subject to significant decay (Ferrisand Beveridge 1986 Sykora et al 2001)
Substantial foaming in the reactor was observed andlasted for 1 day after the EDTA shock load Recovery ofthe biological activity in the reactor was rapid At 01day reactor EDTA concentration dropped to 19 mM and23 6 1 inhibition of ammonium oxidation was mea-sured consistent with the extent of inhibition predictedfrom the batch assay (Fig 2) Effluent concentrations (1day after the shock load) indicated a return to stable operation (NH4
1-N NO22-N less than 01 mgL and
NO32-N ca 300 mgL)
Concomitantly reactor colloidal COD increasedslightly (from 6 to 23 6 6 mgL COD) after EDTA shockloads and returned to background level after 2 days Bio-mass floc size remained unchanged at 40 (62) mm (num-ber weighted mean floc size) from 5-day measurementsafter EDTA load compared to 41 (61) mm before dose
Implications for full-scale biological nitrogenremoval operation
The batch extant respirometric assay employed in thiswork can be used to ensure that the microbial physiol-ogy and activity remain close to that in the sampled con-tinuous flow reactor (Ellis et al 1996 Grady et al1996) From the study of EDTA in the continuous flowreactor the measured SOUR and subsequent inhibitionwas indeed consistent with the predicted value from the
batch assay No further inhibition was noticed and therecovery of the nitrifying activity in the continuous flowreactor was rapid Hence at least for compounds likeEDTA (and presumably DTPA) batch extant respiro-metric assays may accurately portray the nitrifying ac-tivity and inhibition in the continuous flow reactors
Although we only observed nitrification inhibition inboth batch and continuous flow reactors by EDTA whenits concentration was higher than 15 mM caution shouldbe employed in directly extending these results to full-scalewastewater treatment plants Inhibition may be primarilydependent on the concentration of uncomplexed EDTAspecies (Bucheli-Witschel and Egli 2001) which amongother physicochemical factors is often mediated by aque-ous Ca21 and Mg21 concentrations For instance equilib-rium speciation calculation revealed that the concentrationof uncomplexed [EDTA42] in our reactor was 10255 mMat 15 mM total EDTA The same uncomplexed EDTAconcentration could be obtained at 07 mM total EDTA attypical wastewater conditions of 16 mgL Ca21 and 10mgL Mg21 (Tchobanoglous and Burton 1991)
ACKNOWLEDGMENTS
We are grateful to Prof Diane J Burgess in the Dept ofPharmacy and Dr Michele Barber in the Dept of Molec-ular and Cell Biology at the University of Connecticut forassistance with particle sizing and flow cytometry respec-tively This study was funded in part by the ConnecticutDepartment of Environmental Protection through a USEPA Long Island Sound Study grant under Section 119 ofthe Clean Water Act and by the Connecticut Institute ofWater Resources under the 104B program of the NationalInstitutes of Water Resources US Geological Survey asauthorized by the Water Research and Development Act of1978 (PL 95-467) contract number 01HQGR0078
REFERENCES
ALDER AC SIEGRIST H GUJER W and GIGGER W(1990) Behavior of NTA and EDTA in biological waste-water treatment Water Res 24 733ndash742
APHA AWWA and WEF (1998) Standard Methods for theExamination of Water and Wastewater 20th ed Washing-ton DC author
AVERY SV HOWLETT NG and RADICE S (1996)Copper toxicity towards Saccharomyces cerevisiae Depen-dence on plasma membrane fatty acid composition Appl En-viron Microbiol 62 3960ndash3966
BLUM DJW and SPEECE RE (1991) A database ofchemical toxicity to environmental bacteria and its use in in-
NITRIFICATION INHIBITION BY ETHYLENEDIAMINE-BASED CHELATING AGENTS 227
ENVIRON ENG SCI VOL 20 NO 3 2003
terspecies comparisons and correlations J Water PollutControl Fed 63 198ndash207
BUCHELI-WITSCHEL M and EGLI T (2001) Environ-mental fate and microbial degradation of aminopolycar-boxylic acids FEMS Microbiol Rev 25 69ndash106
CHANDRAN K and SMETS BF (2000) Single-step nitri-fication models erroneously describe batch ammonia oxida-tion profiles when nitrite oxidation becomes rate limitingBiotechnol Bioeng 68 396ndash406
DAIGGER GT and SADICK TE (1998) Evaluation ofmethods to detect and control nitrification inhibition withspecific application to incinerator flue-gas scrubber waterWater Environ Res 70 1248ndash1257
ELLIS TG BARBEAU DS SMETS BF and GRADYLCP (1996) Respirometric technique for determination ofextant kinetic parameters describing biodegradation WaterEnviron Res 68 917ndash926
FERRIS FG (1989) Metallic ion interactions with outer mem-brane of Gram-negative bacteria In TJ Beveridge and RJDoyle Eds Metal Ions and Bacteria New York John Wi-ley and Sons pp 295ndash323
FERRIS FG and BEVERIDGE TJ (1986) Physicochemi-cal roles of soluble metal cations in the outer membrane ofEscherichia coli K-12 Can J Microbiol 32 594ndash601
FORD DL CHURCHWELL RL and KACHTICK JW(1980) Comprehensive analysis of nitrification of chemicalprocessing wastewaters J Water Pollut Control Fed 522726ndash2736
GERNAEY K VERSCHUERE L LUYTEN L and VER-STRAETE W (1997) Fast and sensitive acute toxicity de-tection with an enrichment nitrifying culture Water EnvironRes 69 1163ndash1169
GRADY CPL JR SMETS BF and BARBEAU DS (1996)Variability in kinetic parameter estimates A review of possiblecauses and a proposed terminology Water Res 30 742ndash748
HOCKENBURY MR and GRADY CPL (1977) Inhibi-tion of nitrification-effects of selected organic compoundsJ Water Pollut Control Fed 49 768ndash777
HOWLETT NG and AVERY SV (1997a) Induction oflipid peroxidation during heavy metal stress in Saccha-romyces cerevisiae and influence of plasma membrane fattyacid unsaturation Appl Environ Microbiol 63 2971ndash2976
HOWLETT NG and AVERY SV (1997b) Relationship be-tween cadmium sensitivity and degree of plasma membranefatty acid unsaturation in Saccharomyces cerevisiae ApplMicrobiol Biotechnol 48 539ndash545
HU Z CHANDRAN K GRASSO D and SMETS BF(2002a) Effect of nickel and cadmium speciation on nitrifi-cation inhibition Environ Sci Technol 36 3074ndash3078
HU Z CHADRAN K GRASSO D and SMETS BF(2003) Impact of heavy metal sorption and internalization onnitrification inhibition Environ Sci Technol 37 728ndash734
HU Z CHANDRAN K SMETS BF and GRASSO D(2002b) Evaluation of a rapid physical-chemical method forthe determination of extant soluble COD Water Res 36617ndash624
KARI FG and GIGER W (1996) Speciation and fate of ethylenediaminetetraacetate (EDTA) in municipal waste-water treatment Water Res 30 122ndash134
LEIVE L (1974) The barrier function of Gram-negative en-velope Ann NY Acad Sci 235 109ndash127
MARTEL AE SMITH RM and MOTEKAITIS RJ(1998) NIST Critical Stability Constants of Metal ComplexesDatabase College Station TX Texas AampM Univ
OHSUMI Y KITAMOTO K and ANRAKU Y (1988)Changes induced in the permeability barrier of the yeastplasma membrane by cupric ion J Bacteriol 170 2676ndash2682
PAINTER HA (1970) A review of literature on inorganic nitrogen metabolism in microorganisms Water Res 4 393ndash450
PIETSCH J SACHER F SCHMIDT W and BRAUCHH-J (2001) Polar nitrogen compounds and their behaviorin the drinking water treatment process Water Res 35 3537ndash3544
SCHECHER WD and MCAVOY DC (2001) MINEQL1A Chemical Equilibrium Modeling System Version 45 forWindows Hallowell ME Environmental Research Software
SCHOWANEK D MCAVOY D VERSTEEG D andHANSTVEIT A (1996) Effects of nutrient trace metal spe-ciation on algal growth in the presence of the chelator [SS]-EDDS Aquatic Toxicol 36 253ndash275
SHAMMAS NK (1986) Interactions of temperature pH andbiomass on the nitrification process J Water Pollut Con-trol Fed 58 52ndash58
SYKORA V PITTER P BITTNEROVA I and LEDERERT (2001) Biodegradability of ethylenediamine-based com-plexing agents Water Res 35 2010ndash2016
TCHOBANOGLOUS G and BURTON FL (1991) Waste-water Engineering Treatment Disposal and Reuse 3rd edNew York Metcalf amp Eddy McGraw-Hill
TEXIER A ANDRES Y ILLEMASSENE M andCLOIREC PL (2000) Characterization of lanthanide ionsbinding sites in the cell wall of Pseudomonos aeruginosaEnviron Sci Technol 34 610ndash615
TOMLINSON TG BOON AG and TROTMAN CNA(1966) Inhibition of nitrification in the activated sludge pro-cess of sewage disposal J Appl Bacteriol 29 266ndash291
US EPA (1993) Nitrogen control In EPA625R-93-0101Washington DC Office of Water
VANDEVIVERE P FICARA E TERRAS C JULIES Eand VERSTRAETE W (1998) Copper-mediated selectiveremoval of nitrification inhibitors from industrial waste-waters Environ Sci Technol 32 1000ndash1006
228 HU ET AL

Nitrifying cultures (20 mM MOPS pH 70 6 005)were incubated with individual chelating agents up to 2mM for 15 min Microbial cell pellets retained after cen-trifugation (1600 3 g 5 min) were resuspended in thesame volume of 01 mM NaCl followed by centrifuga-tion twice for 5 min at 1600 3 g Between each cen-trifugation the supernatant was decanted and pellets re-suspended in 01 mM NaCl Samples were prefilteredthrough 25-mm pore size filters (Calbiochem La JollaCA) to prevent clogging of the flow cytometer nozzleThe percentage of viable cells after the various chelatorspikes was then quantified using a constructed calibra-tion curve
EDTA shock load to continuous flow reactor
A shock load of EDTA was imposed by instanta-neously adding an aliquot of concentrated EDTA to thecontinuous flow reactor over a 5 min (50003HRT) in-terval Reactor or effluent aliquots were periodically col-lected to measure all nitrogen species as well as colloidaland soluble chemical oxygen demand (COD) The rou-tine batch respirometric assay was also performed withbiomass periodically removed during and after the shockload to determine the NH4
1 and NO22 oxidation kinet-
ics Floc size distribution was measured in duplicatewith an AccuSizer 770 Particle Optical Sizer (Santa Bar-bara CA) with resolution of 05ndash500 mm Each mea-surement lasted 30 s and employed a biomass volume of50 mL
Analytical procedures
Trace metal concentrations were measured accordingto standard methods (APHA et al 1998) Calcium ironmagnesium manganese and potassium concentrationswere measured by Inductively Coupled Plasma (ICP)spectroscopy (Optima 3300XL Perkin-Elmer Co Nor-walk CT) with method detection limit of 20 mgL foreach metal at the Environmental Research Institute (Uni-versity of Connecticut) Copper concentrations weremeasured via flame atomic absorption spectrometry(Model 5100 Perkin-Elmer Co with a method detectionlimit of 20 mgL Biomass concentrations were measuredas COD using commercially available reagents (HACHCOD vials 0ndash1500 mgL) Effluent concentrations ofcolloidal COD (0001ndash1 mm) were deduced from the dif-ference between the COD fraction less than 1 mm andthe truly soluble COD (measured after 05 mM LaCl3destabilization and subsequent 045-mm filtration) ac-cording to our earlier reported method (Hu et al 2002b)Effluent EDTA concentrations were approximated as ef-fluent truly soluble COD corrected for the preshock loadsoluble COD background (ca 20 mgL) NH4
1-N was
analyzed using an ammonia gas-sensing electrode (HNUSystems Newton MA) with an operating range from003 to 1400 mgL NH4
1-N NO22-N was measured col-
orimetrically with a method detection limit of 2 mgL andNO3
2-N via ion chromatography with MDL of 40 mgLall according to standard methods (APHA et al 1998)
Precautions were taken to avoid trace metal contami-nation Glassware and plastic ware when appropriatewere soaked in 1 M HNO3 overnight and rinsed with 5volumes of Milli-Q purified water before use
RESULTS AND DISCUSSION
Nitrification kinetics and dosendashresponse curves
In the short-term batch assays (about 1 h) the specificammonium oxidation rate (SOURNH4) decreased as theapplied EDTA or DTPA dose to the nitrifying biomassincreased whereas the specific nitrite oxidation rate(SOURNO2) was not adversely affected within the con-centration range tested (data not shown) A similar inhi-bition pattern was observed for EDA (data not shown)The higher sensitivity of ammonium oxidation comparedto nitrite oxidation is consistent with earlier observations(Hockenbury and Grady 1977 Blum and Speece 1991)From the SOUR data dosendashinhibitory response curvesfor ammonium oxidation were developed for the chelat-ing agents (Fig 2)
Ethylenediamine was inhibitory even at the lowest
222 HU ET AL
Figure 2 Dosendashresponse curves of chelating agent inhibitionto nitrifying cultures (1033 6 112 mgL COD pH 75) EDTA(d ) DTPA ( s ) EDA ( j ) Error bars indicate 1 standard de-viation (n 5 2)
concentrations tested The EC50 (defined as the concen-tration of chelating agent resulting in 50 inhibition) ofEDA was ca 06 mM This value is consistent with anearlier reported 61 inhibition of ammonia oxidation at05mM (Hockenbury and Grady 1977) EDTA has usu-ally been considered noninhibitory to ammonium oxida-tion at concentration less than approximately 1 mM(Tomlinson et al 1966 Painter 1970) However whenEDTA concentrations were higher than 15 mM signif-icant inhibition was observed in this work DTPA on theother hand has been reported to be more toxic thanEDTA in standard aquatic toxicity tests with fish inver-tebrates and algae (Schowanek et al 1996) In our ex-periments with nitrifying bacteria however DTPA wasfound to be less inhibitory than EDTA (Fig 2)
Overall under the same aquatic chemistry conditionsand employing nitrifying biomass with the same culture
NITRIFICATION INHIBITION BY ETHYLENEDIAMINE-BASED CHELATING AGENTS 223
ENVIRON ENG SCI VOL 20 NO 3 2003
Figure 3 Correlation of the inhibition to nitrifying cultures (1074 6 34 mgL COD pH 75) with the depletion of cellular es-sential elements (a) 5 S K 1 Ca 1 Mg 1 Mn 1 Fe 1 Cu (b) Ca (c) Mg (d) Cu Employed chelating agents were EDTA (d )DTPA ( s ) EDA (j ) The initial cellular concentrations were K 0112 6 0007 Ca 0032 6 0006 Mg 0022 6 0002 Mn0076 6 0002 Fe 0080 6 0003 Cu 0046 6 0001 mMg biomass COD (n 5 2) The error bars indicate 1 standard deviation
history the molar-based inhibitory character to ammo-nium oxidation followed EDA EDTA DTPAThe EC50 defined as the concentration of chelating agentcausing 50 inhibition was estimated at 06 24 and31 mM for EDA EDTA and DTPA respectively Theinhibitory character of tested chelating agents did not par-allel but in fact was opposite to their relative com-plexation potential for key multivalent cations (DTPA EDTA EDA) (Table 1) This surprising finding led usto investigate how cellular cation depletion by differentchelating agents compared
Depletion of essential cellular cation
Although C H O and N constitute the bulk of a bac-terial cellrsquos weight many other elements (K Ca Mg MnFe and Cu) are required in macro or microamounts The
K Ca Mg Mn Fe and Cu fraction for the nitrifying en-richment culture was measured at 049 6 003 014 6003 006 6 000 047 6 001 051 6 002 and 033 6001 dry weight respectively (n 5 2) with our prede-termined conversion factor 121 6 009 g biomass CODgbiomass DW (n 5 3) applied
After chelating agents were added to the nitrifying bio-mass substantial depletion (22) of essential cellularcations was observed (Fig 3a) The maximum percent ofdepletion of each cation was 21 67 25 15 21 and 64for K Ca Mg Mn Fe and Cu respectively Inhibitionby EDTA or DTPA correlated well with the depletion ofcellular Ca21 (presumably linear correlation R2 5 082)(Fig 3b) In contrast depletion of Ca21 by EDA was sig-nificantly less for the same extent of inhibition The in-hibition by all three chelating agents correlated well with
the depletion of cellular Mg21 (R2 5 093) (Fig 3c) al-though the maximum amount of Mg21 depletion wasmuch lower (ca 43) than that of Ca21 Finally nitrifi-cation inhibition did not correlate with the depletion ofcellular Cu21(R2 5 044) (Fig 3d) Similarly inhibitionwas not well correlated with the depletion of K Mn orFe (data not shown)
No effort was made to differentiate between the mem-brane-associated fraction and the intracellular fraction ofthe measured essential cellular cations However fromearlier experiments with Escherichia coli it had beenconcluded that EDTA does not bind to the cell surfaceor cross the cytoplasmic membrane (Ferris and Bev-eridge 1986) In most bacteria there would be no EDTAtransport across the membranes (Bucheli-Witschel andEgli 2001) and uncomplexed EDTA can destabilize the
224 HU ET AL
Figure 4 The depletion of cellular (a) Ca21 (b) Mg21 and (c) K1 (mMg XCOD) as a function of molar dose of chelationagents EDTA ( d ) DTPA ( s ) EDA ( j ) Conditions are the same as indicated in Fig 3
cell surface of Gram-negative bacteria by interactionswith essential cellular cations on the surface (Ferris1989) EDTA especially scavenges Ca21 which is pre-dominantly responsible for outer membrane stabilizationThis can cause a concomitant release of significantamount of lipopolysaccharide (Leive 1974 Ferris1989) The inhibition of ammonium oxidizing activity byEDTA observed in this work is probably attributed to asimilar loss of essential cations from the outer membranebecause ammonium oxidizing bacteria exhibit the typi-cal Gram-negative cell membrane structure and correla-tions between the extent of inhibition and cellular Ca21
and Mg21 depletion were observedDTPA has a similar chemical structure to and a higher
molecular weight than EDTA By analogy one can spec-ulate that its transport across cellular membranes is in-significant The mode of inhibition of ammonium oxi-dizing activity by DTPA observed in this work isprobably similar to EDTA as the strong correlations be-tween the extent of inhibition and cellular Ca21 andMg21 depletion existed for both EDTA and DTPA (Fig3b and c)
Based on equilibrium speciation calculations everymol of chelating agents will complex 1 mol of Ca21 orMg21 at pH 75 At the same molar concentration EDAresulted in less cellular Ca21 depletion (Fig 4a) but muchhigher inhibition than EDTA or DTPA (Fig 2) Secondat the same molar concentration EDA resulted in morecellular Mg21 depletion than EDTA or DTPA (Fig 4b)yet the stability constant of MgEDA (1004) is muchsmaller than that of Mg EDTA (1088) or MgDTPA (1093)(Table 1) The results with EDA suggest that cellularMg21 depletion by EDA did not derive primarily fromMgndashEDA complexation Instead it is speculated due toleakage of destabilized cell membrane as EDA resultedin substantially higher cellular K1 depletion than EDTAor DTPA (Fig 4c) K1 was the most abundant cellularcation (on a molar basis data not shown) and is gener-ally believed to play an important role in maintenance ofthe ionic environment in the cytoplasm Depletion of cel-lular K1 indicates a loss of cell membrane integrity andviability (Ohsumi et al 1988 Avery et al 1996 Howlettand Avery 1997a)
Bacterial viability
Because depletion of cellular K1 has been widely usedas a marker of plasma membrane disruption (Howlett andAvery 1997b) we investigated directly whether disrup-tion of the plasma membrane integrity was observed af-ter exposure to the different agents In the presence of01 mM EDA 745 6 31 cells remained viable after15-min exposure At 2 mM EDA less than 113 6 00cells were viable compared to 875 6 111 461 6
108 viable cells in the presence of similar concentra-tions of EDTA and DTPA The biological effect of EDAtherefore is different among the chelators studied andmay involve cytoplasmic membrane rupture The ob-served loss of the plasma membrane integrity by 2 mMDTPA addition was unexpectedly high in view of the un-affected nitrifying activity at the same concentration (Fig2) Further experiments are needed to better understandwhat factors are responsible for the inconsistency be-tween the changes in the measured cellular activity(SOUR) and viability (livedead staining) after DTPAexposure
NITRIFICATION INHIBITION BY ETHYLENEDIAMINE-BASED CHELATING AGENTS 225
ENVIRON ENG SCI VOL 20 NO 3 2003
Figure 5 Inhibition of NH41-N oxidation by (a) 25 mM
EDTA or (b) 025 mM EDA in presence of metal ions Cu21
(d ) Fe31 ( s ) Ca21 ( j ) Mg21 ( u ) Bold horizontal lines rep-resent mean percent of inhibition in the presence of (a) EDTAor (b) EDA without metal addition
Inhibition mitigation by complexation
Appropriate remedial strategies to relieve nitrificationinhibition by chelatina agents were subsequently ex-plored Simultaneous addition of metals as coordinationpartners reduced the biological effects of chelatingagents but actual trends were ligand and metal specific(Fig 5)
At a 1 mM Fe31 dose (as FeCl3) nitrification inhibi-tion by 25 mM EDTA was completely relieved (Fig 5a)Similarly addition of Ca21 (as CaCl2) or Mg21 (as
MgSO4) reduced nitrification inhibition However Ca21
and Mg21 were slightly less effective than Fe31 on a mo-lar basis This may derive from EDTArsquos ability to formstronger complexes with the Lewis acid Fe compared toCa or Mg (Table 1) On the other hand addition of Cu21
(as CuCl2) only partially reduced nitrification inhibitionat a 11 ratio of CuEDTA (predominant speciesCuEDTA22) Copper itself inhibited the nitrifying activ-ity at high concentrations
In contrast nitrification inhibition by 025 mM EDAwas not relieved at any Fe31 dose tested (Fig 5b) Sim-ilarly addition of Ca21 or Mg21 did not reduce nitrifi-cation inhibition at the highest dose applied (5 mM)which is in agreement with their weak complexation po-tential (CaEDA K 5 1001 MgEDA K 5 1004) Theweak complexation behavior of EDA by Mg21 and thehigh depletion of cellular Mg21 by EDA further sug-gests that EDA has a different mode of inhibition thanEDTA Again addition of Cu21 only partially reducedthe inhibition with Cu(EDA)2
21 as the predominantspecies according to equilibrium speciation calcula-tions
Effect of exposure time on nitrification inhibition
To quantify the effect of exposure time on nitrifica-tion inhibition by chelating agents a predetermined
226 HU ET AL
Figure 7 Distribution of EDTA ( d ) in a continuous flowbioreactor after a transient dose of 24 mM EDTA within min-utes The continuous line presents the prediction from the ex-ponential decay model where C C0 are the reactor EDTA con-centrations at time t (day) and zero respectively F is the feedflow rate to the continuous flow reactor (10 Lday) and V isthe reactor volume (10 L)
Figure 6 Effect of exposure time on nitrification inhibitionby (a) EDTA or (b) EDA in batch assays Initial chelating agentconcentrations are shown Inhibition is expressed with refer-ence of SOUR of a culture incubated without chelating agentsto same duration after removal from parent reactor
dose of EDTA or EDA was applied in the batch reac-tor and the extent of inhibition measured as a functionof time Nitrification inhibition by EDTA reached its ul-timate value within 6 h (paired t-test p 025) (Fig6a) whereas inhibition by EDA substantially increasedup to 6 h (paired t-test p 005) and then decreased(Fig 6b) Such discrepancy between EDTA and EDAfurther suggests different modes of action for EDTA andEDA It also suggests that changes of the intracellularcation pool by EDA may have been slow analogous toslow kinetics of metal internalization reported else-where (Hu et al 2003)
Nitrification inhibition by EDTA in thecontinuous flow parent reactor
A shock load of EDTA was imposed to the continu-ously operated parent nitrifying bioreactor at 24 mM aconcentration corresponding to 47 6 2 inhibition in thebatch assay An ideal continuous flow reactor model ad-equately described the effluent EDTA (complexed anduncomplexed) profile (R2 5 099) computed from the ef-fluent COD data corrected for prepulse concentrations(Fig 7) which indicates that EDTA essentially behavedas a conservative compound did not partition to the cellsurface and was not subject to significant decay (Ferrisand Beveridge 1986 Sykora et al 2001)
Substantial foaming in the reactor was observed andlasted for 1 day after the EDTA shock load Recovery ofthe biological activity in the reactor was rapid At 01day reactor EDTA concentration dropped to 19 mM and23 6 1 inhibition of ammonium oxidation was mea-sured consistent with the extent of inhibition predictedfrom the batch assay (Fig 2) Effluent concentrations (1day after the shock load) indicated a return to stable operation (NH4
1-N NO22-N less than 01 mgL and
NO32-N ca 300 mgL)
Concomitantly reactor colloidal COD increasedslightly (from 6 to 23 6 6 mgL COD) after EDTA shockloads and returned to background level after 2 days Bio-mass floc size remained unchanged at 40 (62) mm (num-ber weighted mean floc size) from 5-day measurementsafter EDTA load compared to 41 (61) mm before dose
Implications for full-scale biological nitrogenremoval operation
The batch extant respirometric assay employed in thiswork can be used to ensure that the microbial physiol-ogy and activity remain close to that in the sampled con-tinuous flow reactor (Ellis et al 1996 Grady et al1996) From the study of EDTA in the continuous flowreactor the measured SOUR and subsequent inhibitionwas indeed consistent with the predicted value from the
batch assay No further inhibition was noticed and therecovery of the nitrifying activity in the continuous flowreactor was rapid Hence at least for compounds likeEDTA (and presumably DTPA) batch extant respiro-metric assays may accurately portray the nitrifying ac-tivity and inhibition in the continuous flow reactors
Although we only observed nitrification inhibition inboth batch and continuous flow reactors by EDTA whenits concentration was higher than 15 mM caution shouldbe employed in directly extending these results to full-scalewastewater treatment plants Inhibition may be primarilydependent on the concentration of uncomplexed EDTAspecies (Bucheli-Witschel and Egli 2001) which amongother physicochemical factors is often mediated by aque-ous Ca21 and Mg21 concentrations For instance equilib-rium speciation calculation revealed that the concentrationof uncomplexed [EDTA42] in our reactor was 10255 mMat 15 mM total EDTA The same uncomplexed EDTAconcentration could be obtained at 07 mM total EDTA attypical wastewater conditions of 16 mgL Ca21 and 10mgL Mg21 (Tchobanoglous and Burton 1991)
ACKNOWLEDGMENTS
We are grateful to Prof Diane J Burgess in the Dept ofPharmacy and Dr Michele Barber in the Dept of Molec-ular and Cell Biology at the University of Connecticut forassistance with particle sizing and flow cytometry respec-tively This study was funded in part by the ConnecticutDepartment of Environmental Protection through a USEPA Long Island Sound Study grant under Section 119 ofthe Clean Water Act and by the Connecticut Institute ofWater Resources under the 104B program of the NationalInstitutes of Water Resources US Geological Survey asauthorized by the Water Research and Development Act of1978 (PL 95-467) contract number 01HQGR0078
REFERENCES
ALDER AC SIEGRIST H GUJER W and GIGGER W(1990) Behavior of NTA and EDTA in biological waste-water treatment Water Res 24 733ndash742
APHA AWWA and WEF (1998) Standard Methods for theExamination of Water and Wastewater 20th ed Washing-ton DC author
AVERY SV HOWLETT NG and RADICE S (1996)Copper toxicity towards Saccharomyces cerevisiae Depen-dence on plasma membrane fatty acid composition Appl En-viron Microbiol 62 3960ndash3966
BLUM DJW and SPEECE RE (1991) A database ofchemical toxicity to environmental bacteria and its use in in-
NITRIFICATION INHIBITION BY ETHYLENEDIAMINE-BASED CHELATING AGENTS 227
ENVIRON ENG SCI VOL 20 NO 3 2003
terspecies comparisons and correlations J Water PollutControl Fed 63 198ndash207
BUCHELI-WITSCHEL M and EGLI T (2001) Environ-mental fate and microbial degradation of aminopolycar-boxylic acids FEMS Microbiol Rev 25 69ndash106
CHANDRAN K and SMETS BF (2000) Single-step nitri-fication models erroneously describe batch ammonia oxida-tion profiles when nitrite oxidation becomes rate limitingBiotechnol Bioeng 68 396ndash406
DAIGGER GT and SADICK TE (1998) Evaluation ofmethods to detect and control nitrification inhibition withspecific application to incinerator flue-gas scrubber waterWater Environ Res 70 1248ndash1257
ELLIS TG BARBEAU DS SMETS BF and GRADYLCP (1996) Respirometric technique for determination ofextant kinetic parameters describing biodegradation WaterEnviron Res 68 917ndash926
FERRIS FG (1989) Metallic ion interactions with outer mem-brane of Gram-negative bacteria In TJ Beveridge and RJDoyle Eds Metal Ions and Bacteria New York John Wi-ley and Sons pp 295ndash323
FERRIS FG and BEVERIDGE TJ (1986) Physicochemi-cal roles of soluble metal cations in the outer membrane ofEscherichia coli K-12 Can J Microbiol 32 594ndash601
FORD DL CHURCHWELL RL and KACHTICK JW(1980) Comprehensive analysis of nitrification of chemicalprocessing wastewaters J Water Pollut Control Fed 522726ndash2736
GERNAEY K VERSCHUERE L LUYTEN L and VER-STRAETE W (1997) Fast and sensitive acute toxicity de-tection with an enrichment nitrifying culture Water EnvironRes 69 1163ndash1169
GRADY CPL JR SMETS BF and BARBEAU DS (1996)Variability in kinetic parameter estimates A review of possiblecauses and a proposed terminology Water Res 30 742ndash748
HOCKENBURY MR and GRADY CPL (1977) Inhibi-tion of nitrification-effects of selected organic compoundsJ Water Pollut Control Fed 49 768ndash777
HOWLETT NG and AVERY SV (1997a) Induction oflipid peroxidation during heavy metal stress in Saccha-romyces cerevisiae and influence of plasma membrane fattyacid unsaturation Appl Environ Microbiol 63 2971ndash2976
HOWLETT NG and AVERY SV (1997b) Relationship be-tween cadmium sensitivity and degree of plasma membranefatty acid unsaturation in Saccharomyces cerevisiae ApplMicrobiol Biotechnol 48 539ndash545
HU Z CHANDRAN K GRASSO D and SMETS BF(2002a) Effect of nickel and cadmium speciation on nitrifi-cation inhibition Environ Sci Technol 36 3074ndash3078
HU Z CHADRAN K GRASSO D and SMETS BF(2003) Impact of heavy metal sorption and internalization onnitrification inhibition Environ Sci Technol 37 728ndash734
HU Z CHANDRAN K SMETS BF and GRASSO D(2002b) Evaluation of a rapid physical-chemical method forthe determination of extant soluble COD Water Res 36617ndash624
KARI FG and GIGER W (1996) Speciation and fate of ethylenediaminetetraacetate (EDTA) in municipal waste-water treatment Water Res 30 122ndash134
LEIVE L (1974) The barrier function of Gram-negative en-velope Ann NY Acad Sci 235 109ndash127
MARTEL AE SMITH RM and MOTEKAITIS RJ(1998) NIST Critical Stability Constants of Metal ComplexesDatabase College Station TX Texas AampM Univ
OHSUMI Y KITAMOTO K and ANRAKU Y (1988)Changes induced in the permeability barrier of the yeastplasma membrane by cupric ion J Bacteriol 170 2676ndash2682
PAINTER HA (1970) A review of literature on inorganic nitrogen metabolism in microorganisms Water Res 4 393ndash450
PIETSCH J SACHER F SCHMIDT W and BRAUCHH-J (2001) Polar nitrogen compounds and their behaviorin the drinking water treatment process Water Res 35 3537ndash3544
SCHECHER WD and MCAVOY DC (2001) MINEQL1A Chemical Equilibrium Modeling System Version 45 forWindows Hallowell ME Environmental Research Software
SCHOWANEK D MCAVOY D VERSTEEG D andHANSTVEIT A (1996) Effects of nutrient trace metal spe-ciation on algal growth in the presence of the chelator [SS]-EDDS Aquatic Toxicol 36 253ndash275
SHAMMAS NK (1986) Interactions of temperature pH andbiomass on the nitrification process J Water Pollut Con-trol Fed 58 52ndash58
SYKORA V PITTER P BITTNEROVA I and LEDERERT (2001) Biodegradability of ethylenediamine-based com-plexing agents Water Res 35 2010ndash2016
TCHOBANOGLOUS G and BURTON FL (1991) Waste-water Engineering Treatment Disposal and Reuse 3rd edNew York Metcalf amp Eddy McGraw-Hill
TEXIER A ANDRES Y ILLEMASSENE M andCLOIREC PL (2000) Characterization of lanthanide ionsbinding sites in the cell wall of Pseudomonos aeruginosaEnviron Sci Technol 34 610ndash615
TOMLINSON TG BOON AG and TROTMAN CNA(1966) Inhibition of nitrification in the activated sludge pro-cess of sewage disposal J Appl Bacteriol 29 266ndash291
US EPA (1993) Nitrogen control In EPA625R-93-0101Washington DC Office of Water
VANDEVIVERE P FICARA E TERRAS C JULIES Eand VERSTRAETE W (1998) Copper-mediated selectiveremoval of nitrification inhibitors from industrial waste-waters Environ Sci Technol 32 1000ndash1006
228 HU ET AL

concentrations tested The EC50 (defined as the concen-tration of chelating agent resulting in 50 inhibition) ofEDA was ca 06 mM This value is consistent with anearlier reported 61 inhibition of ammonia oxidation at05mM (Hockenbury and Grady 1977) EDTA has usu-ally been considered noninhibitory to ammonium oxida-tion at concentration less than approximately 1 mM(Tomlinson et al 1966 Painter 1970) However whenEDTA concentrations were higher than 15 mM signif-icant inhibition was observed in this work DTPA on theother hand has been reported to be more toxic thanEDTA in standard aquatic toxicity tests with fish inver-tebrates and algae (Schowanek et al 1996) In our ex-periments with nitrifying bacteria however DTPA wasfound to be less inhibitory than EDTA (Fig 2)
Overall under the same aquatic chemistry conditionsand employing nitrifying biomass with the same culture
NITRIFICATION INHIBITION BY ETHYLENEDIAMINE-BASED CHELATING AGENTS 223
ENVIRON ENG SCI VOL 20 NO 3 2003
Figure 3 Correlation of the inhibition to nitrifying cultures (1074 6 34 mgL COD pH 75) with the depletion of cellular es-sential elements (a) 5 S K 1 Ca 1 Mg 1 Mn 1 Fe 1 Cu (b) Ca (c) Mg (d) Cu Employed chelating agents were EDTA (d )DTPA ( s ) EDA (j ) The initial cellular concentrations were K 0112 6 0007 Ca 0032 6 0006 Mg 0022 6 0002 Mn0076 6 0002 Fe 0080 6 0003 Cu 0046 6 0001 mMg biomass COD (n 5 2) The error bars indicate 1 standard deviation
history the molar-based inhibitory character to ammo-nium oxidation followed EDA EDTA DTPAThe EC50 defined as the concentration of chelating agentcausing 50 inhibition was estimated at 06 24 and31 mM for EDA EDTA and DTPA respectively Theinhibitory character of tested chelating agents did not par-allel but in fact was opposite to their relative com-plexation potential for key multivalent cations (DTPA EDTA EDA) (Table 1) This surprising finding led usto investigate how cellular cation depletion by differentchelating agents compared
Depletion of essential cellular cation
Although C H O and N constitute the bulk of a bac-terial cellrsquos weight many other elements (K Ca Mg MnFe and Cu) are required in macro or microamounts The
K Ca Mg Mn Fe and Cu fraction for the nitrifying en-richment culture was measured at 049 6 003 014 6003 006 6 000 047 6 001 051 6 002 and 033 6001 dry weight respectively (n 5 2) with our prede-termined conversion factor 121 6 009 g biomass CODgbiomass DW (n 5 3) applied
After chelating agents were added to the nitrifying bio-mass substantial depletion (22) of essential cellularcations was observed (Fig 3a) The maximum percent ofdepletion of each cation was 21 67 25 15 21 and 64for K Ca Mg Mn Fe and Cu respectively Inhibitionby EDTA or DTPA correlated well with the depletion ofcellular Ca21 (presumably linear correlation R2 5 082)(Fig 3b) In contrast depletion of Ca21 by EDA was sig-nificantly less for the same extent of inhibition The in-hibition by all three chelating agents correlated well with
the depletion of cellular Mg21 (R2 5 093) (Fig 3c) al-though the maximum amount of Mg21 depletion wasmuch lower (ca 43) than that of Ca21 Finally nitrifi-cation inhibition did not correlate with the depletion ofcellular Cu21(R2 5 044) (Fig 3d) Similarly inhibitionwas not well correlated with the depletion of K Mn orFe (data not shown)
No effort was made to differentiate between the mem-brane-associated fraction and the intracellular fraction ofthe measured essential cellular cations However fromearlier experiments with Escherichia coli it had beenconcluded that EDTA does not bind to the cell surfaceor cross the cytoplasmic membrane (Ferris and Bev-eridge 1986) In most bacteria there would be no EDTAtransport across the membranes (Bucheli-Witschel andEgli 2001) and uncomplexed EDTA can destabilize the
224 HU ET AL
Figure 4 The depletion of cellular (a) Ca21 (b) Mg21 and (c) K1 (mMg XCOD) as a function of molar dose of chelationagents EDTA ( d ) DTPA ( s ) EDA ( j ) Conditions are the same as indicated in Fig 3
cell surface of Gram-negative bacteria by interactionswith essential cellular cations on the surface (Ferris1989) EDTA especially scavenges Ca21 which is pre-dominantly responsible for outer membrane stabilizationThis can cause a concomitant release of significantamount of lipopolysaccharide (Leive 1974 Ferris1989) The inhibition of ammonium oxidizing activity byEDTA observed in this work is probably attributed to asimilar loss of essential cations from the outer membranebecause ammonium oxidizing bacteria exhibit the typi-cal Gram-negative cell membrane structure and correla-tions between the extent of inhibition and cellular Ca21
and Mg21 depletion were observedDTPA has a similar chemical structure to and a higher
molecular weight than EDTA By analogy one can spec-ulate that its transport across cellular membranes is in-significant The mode of inhibition of ammonium oxi-dizing activity by DTPA observed in this work isprobably similar to EDTA as the strong correlations be-tween the extent of inhibition and cellular Ca21 andMg21 depletion existed for both EDTA and DTPA (Fig3b and c)
Based on equilibrium speciation calculations everymol of chelating agents will complex 1 mol of Ca21 orMg21 at pH 75 At the same molar concentration EDAresulted in less cellular Ca21 depletion (Fig 4a) but muchhigher inhibition than EDTA or DTPA (Fig 2) Secondat the same molar concentration EDA resulted in morecellular Mg21 depletion than EDTA or DTPA (Fig 4b)yet the stability constant of MgEDA (1004) is muchsmaller than that of Mg EDTA (1088) or MgDTPA (1093)(Table 1) The results with EDA suggest that cellularMg21 depletion by EDA did not derive primarily fromMgndashEDA complexation Instead it is speculated due toleakage of destabilized cell membrane as EDA resultedin substantially higher cellular K1 depletion than EDTAor DTPA (Fig 4c) K1 was the most abundant cellularcation (on a molar basis data not shown) and is gener-ally believed to play an important role in maintenance ofthe ionic environment in the cytoplasm Depletion of cel-lular K1 indicates a loss of cell membrane integrity andviability (Ohsumi et al 1988 Avery et al 1996 Howlettand Avery 1997a)
Bacterial viability
Because depletion of cellular K1 has been widely usedas a marker of plasma membrane disruption (Howlett andAvery 1997b) we investigated directly whether disrup-tion of the plasma membrane integrity was observed af-ter exposure to the different agents In the presence of01 mM EDA 745 6 31 cells remained viable after15-min exposure At 2 mM EDA less than 113 6 00cells were viable compared to 875 6 111 461 6
108 viable cells in the presence of similar concentra-tions of EDTA and DTPA The biological effect of EDAtherefore is different among the chelators studied andmay involve cytoplasmic membrane rupture The ob-served loss of the plasma membrane integrity by 2 mMDTPA addition was unexpectedly high in view of the un-affected nitrifying activity at the same concentration (Fig2) Further experiments are needed to better understandwhat factors are responsible for the inconsistency be-tween the changes in the measured cellular activity(SOUR) and viability (livedead staining) after DTPAexposure
NITRIFICATION INHIBITION BY ETHYLENEDIAMINE-BASED CHELATING AGENTS 225
ENVIRON ENG SCI VOL 20 NO 3 2003
Figure 5 Inhibition of NH41-N oxidation by (a) 25 mM
EDTA or (b) 025 mM EDA in presence of metal ions Cu21
(d ) Fe31 ( s ) Ca21 ( j ) Mg21 ( u ) Bold horizontal lines rep-resent mean percent of inhibition in the presence of (a) EDTAor (b) EDA without metal addition
Inhibition mitigation by complexation
Appropriate remedial strategies to relieve nitrificationinhibition by chelatina agents were subsequently ex-plored Simultaneous addition of metals as coordinationpartners reduced the biological effects of chelatingagents but actual trends were ligand and metal specific(Fig 5)
At a 1 mM Fe31 dose (as FeCl3) nitrification inhibi-tion by 25 mM EDTA was completely relieved (Fig 5a)Similarly addition of Ca21 (as CaCl2) or Mg21 (as
MgSO4) reduced nitrification inhibition However Ca21
and Mg21 were slightly less effective than Fe31 on a mo-lar basis This may derive from EDTArsquos ability to formstronger complexes with the Lewis acid Fe compared toCa or Mg (Table 1) On the other hand addition of Cu21
(as CuCl2) only partially reduced nitrification inhibitionat a 11 ratio of CuEDTA (predominant speciesCuEDTA22) Copper itself inhibited the nitrifying activ-ity at high concentrations
In contrast nitrification inhibition by 025 mM EDAwas not relieved at any Fe31 dose tested (Fig 5b) Sim-ilarly addition of Ca21 or Mg21 did not reduce nitrifi-cation inhibition at the highest dose applied (5 mM)which is in agreement with their weak complexation po-tential (CaEDA K 5 1001 MgEDA K 5 1004) Theweak complexation behavior of EDA by Mg21 and thehigh depletion of cellular Mg21 by EDA further sug-gests that EDA has a different mode of inhibition thanEDTA Again addition of Cu21 only partially reducedthe inhibition with Cu(EDA)2
21 as the predominantspecies according to equilibrium speciation calcula-tions
Effect of exposure time on nitrification inhibition
To quantify the effect of exposure time on nitrifica-tion inhibition by chelating agents a predetermined
226 HU ET AL
Figure 7 Distribution of EDTA ( d ) in a continuous flowbioreactor after a transient dose of 24 mM EDTA within min-utes The continuous line presents the prediction from the ex-ponential decay model where C C0 are the reactor EDTA con-centrations at time t (day) and zero respectively F is the feedflow rate to the continuous flow reactor (10 Lday) and V isthe reactor volume (10 L)
Figure 6 Effect of exposure time on nitrification inhibitionby (a) EDTA or (b) EDA in batch assays Initial chelating agentconcentrations are shown Inhibition is expressed with refer-ence of SOUR of a culture incubated without chelating agentsto same duration after removal from parent reactor
dose of EDTA or EDA was applied in the batch reac-tor and the extent of inhibition measured as a functionof time Nitrification inhibition by EDTA reached its ul-timate value within 6 h (paired t-test p 025) (Fig6a) whereas inhibition by EDA substantially increasedup to 6 h (paired t-test p 005) and then decreased(Fig 6b) Such discrepancy between EDTA and EDAfurther suggests different modes of action for EDTA andEDA It also suggests that changes of the intracellularcation pool by EDA may have been slow analogous toslow kinetics of metal internalization reported else-where (Hu et al 2003)
Nitrification inhibition by EDTA in thecontinuous flow parent reactor
A shock load of EDTA was imposed to the continu-ously operated parent nitrifying bioreactor at 24 mM aconcentration corresponding to 47 6 2 inhibition in thebatch assay An ideal continuous flow reactor model ad-equately described the effluent EDTA (complexed anduncomplexed) profile (R2 5 099) computed from the ef-fluent COD data corrected for prepulse concentrations(Fig 7) which indicates that EDTA essentially behavedas a conservative compound did not partition to the cellsurface and was not subject to significant decay (Ferrisand Beveridge 1986 Sykora et al 2001)
Substantial foaming in the reactor was observed andlasted for 1 day after the EDTA shock load Recovery ofthe biological activity in the reactor was rapid At 01day reactor EDTA concentration dropped to 19 mM and23 6 1 inhibition of ammonium oxidation was mea-sured consistent with the extent of inhibition predictedfrom the batch assay (Fig 2) Effluent concentrations (1day after the shock load) indicated a return to stable operation (NH4
1-N NO22-N less than 01 mgL and
NO32-N ca 300 mgL)
Concomitantly reactor colloidal COD increasedslightly (from 6 to 23 6 6 mgL COD) after EDTA shockloads and returned to background level after 2 days Bio-mass floc size remained unchanged at 40 (62) mm (num-ber weighted mean floc size) from 5-day measurementsafter EDTA load compared to 41 (61) mm before dose
Implications for full-scale biological nitrogenremoval operation
The batch extant respirometric assay employed in thiswork can be used to ensure that the microbial physiol-ogy and activity remain close to that in the sampled con-tinuous flow reactor (Ellis et al 1996 Grady et al1996) From the study of EDTA in the continuous flowreactor the measured SOUR and subsequent inhibitionwas indeed consistent with the predicted value from the
batch assay No further inhibition was noticed and therecovery of the nitrifying activity in the continuous flowreactor was rapid Hence at least for compounds likeEDTA (and presumably DTPA) batch extant respiro-metric assays may accurately portray the nitrifying ac-tivity and inhibition in the continuous flow reactors
Although we only observed nitrification inhibition inboth batch and continuous flow reactors by EDTA whenits concentration was higher than 15 mM caution shouldbe employed in directly extending these results to full-scalewastewater treatment plants Inhibition may be primarilydependent on the concentration of uncomplexed EDTAspecies (Bucheli-Witschel and Egli 2001) which amongother physicochemical factors is often mediated by aque-ous Ca21 and Mg21 concentrations For instance equilib-rium speciation calculation revealed that the concentrationof uncomplexed [EDTA42] in our reactor was 10255 mMat 15 mM total EDTA The same uncomplexed EDTAconcentration could be obtained at 07 mM total EDTA attypical wastewater conditions of 16 mgL Ca21 and 10mgL Mg21 (Tchobanoglous and Burton 1991)
ACKNOWLEDGMENTS
We are grateful to Prof Diane J Burgess in the Dept ofPharmacy and Dr Michele Barber in the Dept of Molec-ular and Cell Biology at the University of Connecticut forassistance with particle sizing and flow cytometry respec-tively This study was funded in part by the ConnecticutDepartment of Environmental Protection through a USEPA Long Island Sound Study grant under Section 119 ofthe Clean Water Act and by the Connecticut Institute ofWater Resources under the 104B program of the NationalInstitutes of Water Resources US Geological Survey asauthorized by the Water Research and Development Act of1978 (PL 95-467) contract number 01HQGR0078
REFERENCES
ALDER AC SIEGRIST H GUJER W and GIGGER W(1990) Behavior of NTA and EDTA in biological waste-water treatment Water Res 24 733ndash742
APHA AWWA and WEF (1998) Standard Methods for theExamination of Water and Wastewater 20th ed Washing-ton DC author
AVERY SV HOWLETT NG and RADICE S (1996)Copper toxicity towards Saccharomyces cerevisiae Depen-dence on plasma membrane fatty acid composition Appl En-viron Microbiol 62 3960ndash3966
BLUM DJW and SPEECE RE (1991) A database ofchemical toxicity to environmental bacteria and its use in in-
NITRIFICATION INHIBITION BY ETHYLENEDIAMINE-BASED CHELATING AGENTS 227
ENVIRON ENG SCI VOL 20 NO 3 2003
terspecies comparisons and correlations J Water PollutControl Fed 63 198ndash207
BUCHELI-WITSCHEL M and EGLI T (2001) Environ-mental fate and microbial degradation of aminopolycar-boxylic acids FEMS Microbiol Rev 25 69ndash106
CHANDRAN K and SMETS BF (2000) Single-step nitri-fication models erroneously describe batch ammonia oxida-tion profiles when nitrite oxidation becomes rate limitingBiotechnol Bioeng 68 396ndash406
DAIGGER GT and SADICK TE (1998) Evaluation ofmethods to detect and control nitrification inhibition withspecific application to incinerator flue-gas scrubber waterWater Environ Res 70 1248ndash1257
ELLIS TG BARBEAU DS SMETS BF and GRADYLCP (1996) Respirometric technique for determination ofextant kinetic parameters describing biodegradation WaterEnviron Res 68 917ndash926
FERRIS FG (1989) Metallic ion interactions with outer mem-brane of Gram-negative bacteria In TJ Beveridge and RJDoyle Eds Metal Ions and Bacteria New York John Wi-ley and Sons pp 295ndash323
FERRIS FG and BEVERIDGE TJ (1986) Physicochemi-cal roles of soluble metal cations in the outer membrane ofEscherichia coli K-12 Can J Microbiol 32 594ndash601
FORD DL CHURCHWELL RL and KACHTICK JW(1980) Comprehensive analysis of nitrification of chemicalprocessing wastewaters J Water Pollut Control Fed 522726ndash2736
GERNAEY K VERSCHUERE L LUYTEN L and VER-STRAETE W (1997) Fast and sensitive acute toxicity de-tection with an enrichment nitrifying culture Water EnvironRes 69 1163ndash1169
GRADY CPL JR SMETS BF and BARBEAU DS (1996)Variability in kinetic parameter estimates A review of possiblecauses and a proposed terminology Water Res 30 742ndash748
HOCKENBURY MR and GRADY CPL (1977) Inhibi-tion of nitrification-effects of selected organic compoundsJ Water Pollut Control Fed 49 768ndash777
HOWLETT NG and AVERY SV (1997a) Induction oflipid peroxidation during heavy metal stress in Saccha-romyces cerevisiae and influence of plasma membrane fattyacid unsaturation Appl Environ Microbiol 63 2971ndash2976
HOWLETT NG and AVERY SV (1997b) Relationship be-tween cadmium sensitivity and degree of plasma membranefatty acid unsaturation in Saccharomyces cerevisiae ApplMicrobiol Biotechnol 48 539ndash545
HU Z CHANDRAN K GRASSO D and SMETS BF(2002a) Effect of nickel and cadmium speciation on nitrifi-cation inhibition Environ Sci Technol 36 3074ndash3078
HU Z CHADRAN K GRASSO D and SMETS BF(2003) Impact of heavy metal sorption and internalization onnitrification inhibition Environ Sci Technol 37 728ndash734
HU Z CHANDRAN K SMETS BF and GRASSO D(2002b) Evaluation of a rapid physical-chemical method forthe determination of extant soluble COD Water Res 36617ndash624
KARI FG and GIGER W (1996) Speciation and fate of ethylenediaminetetraacetate (EDTA) in municipal waste-water treatment Water Res 30 122ndash134
LEIVE L (1974) The barrier function of Gram-negative en-velope Ann NY Acad Sci 235 109ndash127
MARTEL AE SMITH RM and MOTEKAITIS RJ(1998) NIST Critical Stability Constants of Metal ComplexesDatabase College Station TX Texas AampM Univ
OHSUMI Y KITAMOTO K and ANRAKU Y (1988)Changes induced in the permeability barrier of the yeastplasma membrane by cupric ion J Bacteriol 170 2676ndash2682
PAINTER HA (1970) A review of literature on inorganic nitrogen metabolism in microorganisms Water Res 4 393ndash450
PIETSCH J SACHER F SCHMIDT W and BRAUCHH-J (2001) Polar nitrogen compounds and their behaviorin the drinking water treatment process Water Res 35 3537ndash3544
SCHECHER WD and MCAVOY DC (2001) MINEQL1A Chemical Equilibrium Modeling System Version 45 forWindows Hallowell ME Environmental Research Software
SCHOWANEK D MCAVOY D VERSTEEG D andHANSTVEIT A (1996) Effects of nutrient trace metal spe-ciation on algal growth in the presence of the chelator [SS]-EDDS Aquatic Toxicol 36 253ndash275
SHAMMAS NK (1986) Interactions of temperature pH andbiomass on the nitrification process J Water Pollut Con-trol Fed 58 52ndash58
SYKORA V PITTER P BITTNEROVA I and LEDERERT (2001) Biodegradability of ethylenediamine-based com-plexing agents Water Res 35 2010ndash2016
TCHOBANOGLOUS G and BURTON FL (1991) Waste-water Engineering Treatment Disposal and Reuse 3rd edNew York Metcalf amp Eddy McGraw-Hill
TEXIER A ANDRES Y ILLEMASSENE M andCLOIREC PL (2000) Characterization of lanthanide ionsbinding sites in the cell wall of Pseudomonos aeruginosaEnviron Sci Technol 34 610ndash615
TOMLINSON TG BOON AG and TROTMAN CNA(1966) Inhibition of nitrification in the activated sludge pro-cess of sewage disposal J Appl Bacteriol 29 266ndash291
US EPA (1993) Nitrogen control In EPA625R-93-0101Washington DC Office of Water
VANDEVIVERE P FICARA E TERRAS C JULIES Eand VERSTRAETE W (1998) Copper-mediated selectiveremoval of nitrification inhibitors from industrial waste-waters Environ Sci Technol 32 1000ndash1006
228 HU ET AL
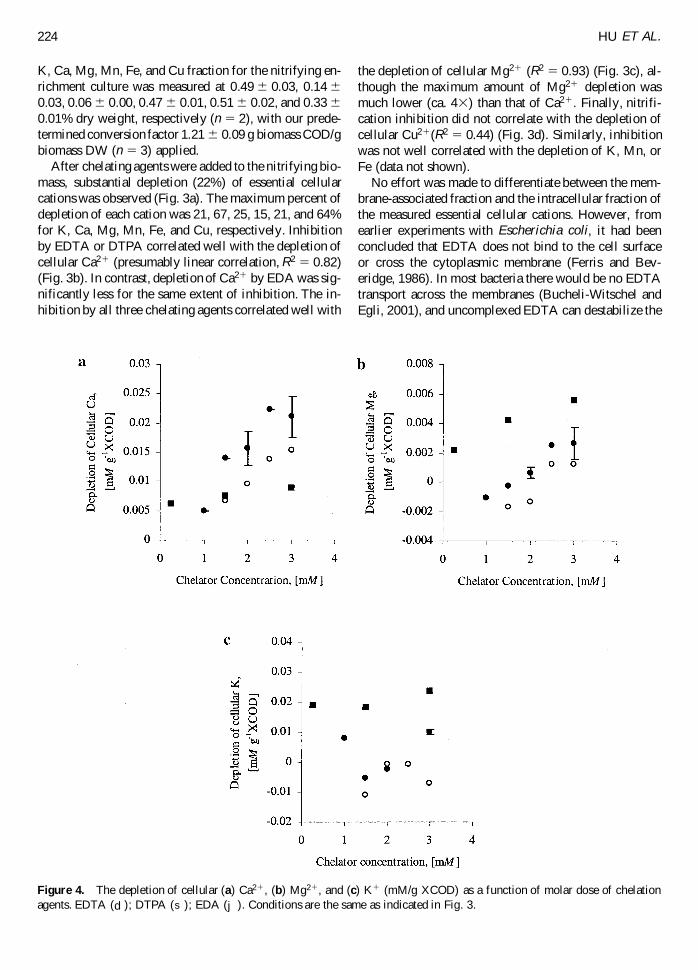
K Ca Mg Mn Fe and Cu fraction for the nitrifying en-richment culture was measured at 049 6 003 014 6003 006 6 000 047 6 001 051 6 002 and 033 6001 dry weight respectively (n 5 2) with our prede-termined conversion factor 121 6 009 g biomass CODgbiomass DW (n 5 3) applied
After chelating agents were added to the nitrifying bio-mass substantial depletion (22) of essential cellularcations was observed (Fig 3a) The maximum percent ofdepletion of each cation was 21 67 25 15 21 and 64for K Ca Mg Mn Fe and Cu respectively Inhibitionby EDTA or DTPA correlated well with the depletion ofcellular Ca21 (presumably linear correlation R2 5 082)(Fig 3b) In contrast depletion of Ca21 by EDA was sig-nificantly less for the same extent of inhibition The in-hibition by all three chelating agents correlated well with
the depletion of cellular Mg21 (R2 5 093) (Fig 3c) al-though the maximum amount of Mg21 depletion wasmuch lower (ca 43) than that of Ca21 Finally nitrifi-cation inhibition did not correlate with the depletion ofcellular Cu21(R2 5 044) (Fig 3d) Similarly inhibitionwas not well correlated with the depletion of K Mn orFe (data not shown)
No effort was made to differentiate between the mem-brane-associated fraction and the intracellular fraction ofthe measured essential cellular cations However fromearlier experiments with Escherichia coli it had beenconcluded that EDTA does not bind to the cell surfaceor cross the cytoplasmic membrane (Ferris and Bev-eridge 1986) In most bacteria there would be no EDTAtransport across the membranes (Bucheli-Witschel andEgli 2001) and uncomplexed EDTA can destabilize the
224 HU ET AL
Figure 4 The depletion of cellular (a) Ca21 (b) Mg21 and (c) K1 (mMg XCOD) as a function of molar dose of chelationagents EDTA ( d ) DTPA ( s ) EDA ( j ) Conditions are the same as indicated in Fig 3
cell surface of Gram-negative bacteria by interactionswith essential cellular cations on the surface (Ferris1989) EDTA especially scavenges Ca21 which is pre-dominantly responsible for outer membrane stabilizationThis can cause a concomitant release of significantamount of lipopolysaccharide (Leive 1974 Ferris1989) The inhibition of ammonium oxidizing activity byEDTA observed in this work is probably attributed to asimilar loss of essential cations from the outer membranebecause ammonium oxidizing bacteria exhibit the typi-cal Gram-negative cell membrane structure and correla-tions between the extent of inhibition and cellular Ca21
and Mg21 depletion were observedDTPA has a similar chemical structure to and a higher
molecular weight than EDTA By analogy one can spec-ulate that its transport across cellular membranes is in-significant The mode of inhibition of ammonium oxi-dizing activity by DTPA observed in this work isprobably similar to EDTA as the strong correlations be-tween the extent of inhibition and cellular Ca21 andMg21 depletion existed for both EDTA and DTPA (Fig3b and c)
Based on equilibrium speciation calculations everymol of chelating agents will complex 1 mol of Ca21 orMg21 at pH 75 At the same molar concentration EDAresulted in less cellular Ca21 depletion (Fig 4a) but muchhigher inhibition than EDTA or DTPA (Fig 2) Secondat the same molar concentration EDA resulted in morecellular Mg21 depletion than EDTA or DTPA (Fig 4b)yet the stability constant of MgEDA (1004) is muchsmaller than that of Mg EDTA (1088) or MgDTPA (1093)(Table 1) The results with EDA suggest that cellularMg21 depletion by EDA did not derive primarily fromMgndashEDA complexation Instead it is speculated due toleakage of destabilized cell membrane as EDA resultedin substantially higher cellular K1 depletion than EDTAor DTPA (Fig 4c) K1 was the most abundant cellularcation (on a molar basis data not shown) and is gener-ally believed to play an important role in maintenance ofthe ionic environment in the cytoplasm Depletion of cel-lular K1 indicates a loss of cell membrane integrity andviability (Ohsumi et al 1988 Avery et al 1996 Howlettand Avery 1997a)
Bacterial viability
Because depletion of cellular K1 has been widely usedas a marker of plasma membrane disruption (Howlett andAvery 1997b) we investigated directly whether disrup-tion of the plasma membrane integrity was observed af-ter exposure to the different agents In the presence of01 mM EDA 745 6 31 cells remained viable after15-min exposure At 2 mM EDA less than 113 6 00cells were viable compared to 875 6 111 461 6
108 viable cells in the presence of similar concentra-tions of EDTA and DTPA The biological effect of EDAtherefore is different among the chelators studied andmay involve cytoplasmic membrane rupture The ob-served loss of the plasma membrane integrity by 2 mMDTPA addition was unexpectedly high in view of the un-affected nitrifying activity at the same concentration (Fig2) Further experiments are needed to better understandwhat factors are responsible for the inconsistency be-tween the changes in the measured cellular activity(SOUR) and viability (livedead staining) after DTPAexposure
NITRIFICATION INHIBITION BY ETHYLENEDIAMINE-BASED CHELATING AGENTS 225
ENVIRON ENG SCI VOL 20 NO 3 2003
Figure 5 Inhibition of NH41-N oxidation by (a) 25 mM
EDTA or (b) 025 mM EDA in presence of metal ions Cu21
(d ) Fe31 ( s ) Ca21 ( j ) Mg21 ( u ) Bold horizontal lines rep-resent mean percent of inhibition in the presence of (a) EDTAor (b) EDA without metal addition
Inhibition mitigation by complexation
Appropriate remedial strategies to relieve nitrificationinhibition by chelatina agents were subsequently ex-plored Simultaneous addition of metals as coordinationpartners reduced the biological effects of chelatingagents but actual trends were ligand and metal specific(Fig 5)
At a 1 mM Fe31 dose (as FeCl3) nitrification inhibi-tion by 25 mM EDTA was completely relieved (Fig 5a)Similarly addition of Ca21 (as CaCl2) or Mg21 (as
MgSO4) reduced nitrification inhibition However Ca21
and Mg21 were slightly less effective than Fe31 on a mo-lar basis This may derive from EDTArsquos ability to formstronger complexes with the Lewis acid Fe compared toCa or Mg (Table 1) On the other hand addition of Cu21
(as CuCl2) only partially reduced nitrification inhibitionat a 11 ratio of CuEDTA (predominant speciesCuEDTA22) Copper itself inhibited the nitrifying activ-ity at high concentrations
In contrast nitrification inhibition by 025 mM EDAwas not relieved at any Fe31 dose tested (Fig 5b) Sim-ilarly addition of Ca21 or Mg21 did not reduce nitrifi-cation inhibition at the highest dose applied (5 mM)which is in agreement with their weak complexation po-tential (CaEDA K 5 1001 MgEDA K 5 1004) Theweak complexation behavior of EDA by Mg21 and thehigh depletion of cellular Mg21 by EDA further sug-gests that EDA has a different mode of inhibition thanEDTA Again addition of Cu21 only partially reducedthe inhibition with Cu(EDA)2
21 as the predominantspecies according to equilibrium speciation calcula-tions
Effect of exposure time on nitrification inhibition
To quantify the effect of exposure time on nitrifica-tion inhibition by chelating agents a predetermined
226 HU ET AL
Figure 7 Distribution of EDTA ( d ) in a continuous flowbioreactor after a transient dose of 24 mM EDTA within min-utes The continuous line presents the prediction from the ex-ponential decay model where C C0 are the reactor EDTA con-centrations at time t (day) and zero respectively F is the feedflow rate to the continuous flow reactor (10 Lday) and V isthe reactor volume (10 L)
Figure 6 Effect of exposure time on nitrification inhibitionby (a) EDTA or (b) EDA in batch assays Initial chelating agentconcentrations are shown Inhibition is expressed with refer-ence of SOUR of a culture incubated without chelating agentsto same duration after removal from parent reactor
dose of EDTA or EDA was applied in the batch reac-tor and the extent of inhibition measured as a functionof time Nitrification inhibition by EDTA reached its ul-timate value within 6 h (paired t-test p 025) (Fig6a) whereas inhibition by EDA substantially increasedup to 6 h (paired t-test p 005) and then decreased(Fig 6b) Such discrepancy between EDTA and EDAfurther suggests different modes of action for EDTA andEDA It also suggests that changes of the intracellularcation pool by EDA may have been slow analogous toslow kinetics of metal internalization reported else-where (Hu et al 2003)
Nitrification inhibition by EDTA in thecontinuous flow parent reactor
A shock load of EDTA was imposed to the continu-ously operated parent nitrifying bioreactor at 24 mM aconcentration corresponding to 47 6 2 inhibition in thebatch assay An ideal continuous flow reactor model ad-equately described the effluent EDTA (complexed anduncomplexed) profile (R2 5 099) computed from the ef-fluent COD data corrected for prepulse concentrations(Fig 7) which indicates that EDTA essentially behavedas a conservative compound did not partition to the cellsurface and was not subject to significant decay (Ferrisand Beveridge 1986 Sykora et al 2001)
Substantial foaming in the reactor was observed andlasted for 1 day after the EDTA shock load Recovery ofthe biological activity in the reactor was rapid At 01day reactor EDTA concentration dropped to 19 mM and23 6 1 inhibition of ammonium oxidation was mea-sured consistent with the extent of inhibition predictedfrom the batch assay (Fig 2) Effluent concentrations (1day after the shock load) indicated a return to stable operation (NH4
1-N NO22-N less than 01 mgL and
NO32-N ca 300 mgL)
Concomitantly reactor colloidal COD increasedslightly (from 6 to 23 6 6 mgL COD) after EDTA shockloads and returned to background level after 2 days Bio-mass floc size remained unchanged at 40 (62) mm (num-ber weighted mean floc size) from 5-day measurementsafter EDTA load compared to 41 (61) mm before dose
Implications for full-scale biological nitrogenremoval operation
The batch extant respirometric assay employed in thiswork can be used to ensure that the microbial physiol-ogy and activity remain close to that in the sampled con-tinuous flow reactor (Ellis et al 1996 Grady et al1996) From the study of EDTA in the continuous flowreactor the measured SOUR and subsequent inhibitionwas indeed consistent with the predicted value from the
batch assay No further inhibition was noticed and therecovery of the nitrifying activity in the continuous flowreactor was rapid Hence at least for compounds likeEDTA (and presumably DTPA) batch extant respiro-metric assays may accurately portray the nitrifying ac-tivity and inhibition in the continuous flow reactors
Although we only observed nitrification inhibition inboth batch and continuous flow reactors by EDTA whenits concentration was higher than 15 mM caution shouldbe employed in directly extending these results to full-scalewastewater treatment plants Inhibition may be primarilydependent on the concentration of uncomplexed EDTAspecies (Bucheli-Witschel and Egli 2001) which amongother physicochemical factors is often mediated by aque-ous Ca21 and Mg21 concentrations For instance equilib-rium speciation calculation revealed that the concentrationof uncomplexed [EDTA42] in our reactor was 10255 mMat 15 mM total EDTA The same uncomplexed EDTAconcentration could be obtained at 07 mM total EDTA attypical wastewater conditions of 16 mgL Ca21 and 10mgL Mg21 (Tchobanoglous and Burton 1991)
ACKNOWLEDGMENTS
We are grateful to Prof Diane J Burgess in the Dept ofPharmacy and Dr Michele Barber in the Dept of Molec-ular and Cell Biology at the University of Connecticut forassistance with particle sizing and flow cytometry respec-tively This study was funded in part by the ConnecticutDepartment of Environmental Protection through a USEPA Long Island Sound Study grant under Section 119 ofthe Clean Water Act and by the Connecticut Institute ofWater Resources under the 104B program of the NationalInstitutes of Water Resources US Geological Survey asauthorized by the Water Research and Development Act of1978 (PL 95-467) contract number 01HQGR0078
REFERENCES
ALDER AC SIEGRIST H GUJER W and GIGGER W(1990) Behavior of NTA and EDTA in biological waste-water treatment Water Res 24 733ndash742
APHA AWWA and WEF (1998) Standard Methods for theExamination of Water and Wastewater 20th ed Washing-ton DC author
AVERY SV HOWLETT NG and RADICE S (1996)Copper toxicity towards Saccharomyces cerevisiae Depen-dence on plasma membrane fatty acid composition Appl En-viron Microbiol 62 3960ndash3966
BLUM DJW and SPEECE RE (1991) A database ofchemical toxicity to environmental bacteria and its use in in-
NITRIFICATION INHIBITION BY ETHYLENEDIAMINE-BASED CHELATING AGENTS 227
ENVIRON ENG SCI VOL 20 NO 3 2003
terspecies comparisons and correlations J Water PollutControl Fed 63 198ndash207
BUCHELI-WITSCHEL M and EGLI T (2001) Environ-mental fate and microbial degradation of aminopolycar-boxylic acids FEMS Microbiol Rev 25 69ndash106
CHANDRAN K and SMETS BF (2000) Single-step nitri-fication models erroneously describe batch ammonia oxida-tion profiles when nitrite oxidation becomes rate limitingBiotechnol Bioeng 68 396ndash406
DAIGGER GT and SADICK TE (1998) Evaluation ofmethods to detect and control nitrification inhibition withspecific application to incinerator flue-gas scrubber waterWater Environ Res 70 1248ndash1257
ELLIS TG BARBEAU DS SMETS BF and GRADYLCP (1996) Respirometric technique for determination ofextant kinetic parameters describing biodegradation WaterEnviron Res 68 917ndash926
FERRIS FG (1989) Metallic ion interactions with outer mem-brane of Gram-negative bacteria In TJ Beveridge and RJDoyle Eds Metal Ions and Bacteria New York John Wi-ley and Sons pp 295ndash323
FERRIS FG and BEVERIDGE TJ (1986) Physicochemi-cal roles of soluble metal cations in the outer membrane ofEscherichia coli K-12 Can J Microbiol 32 594ndash601
FORD DL CHURCHWELL RL and KACHTICK JW(1980) Comprehensive analysis of nitrification of chemicalprocessing wastewaters J Water Pollut Control Fed 522726ndash2736
GERNAEY K VERSCHUERE L LUYTEN L and VER-STRAETE W (1997) Fast and sensitive acute toxicity de-tection with an enrichment nitrifying culture Water EnvironRes 69 1163ndash1169
GRADY CPL JR SMETS BF and BARBEAU DS (1996)Variability in kinetic parameter estimates A review of possiblecauses and a proposed terminology Water Res 30 742ndash748
HOCKENBURY MR and GRADY CPL (1977) Inhibi-tion of nitrification-effects of selected organic compoundsJ Water Pollut Control Fed 49 768ndash777
HOWLETT NG and AVERY SV (1997a) Induction oflipid peroxidation during heavy metal stress in Saccha-romyces cerevisiae and influence of plasma membrane fattyacid unsaturation Appl Environ Microbiol 63 2971ndash2976
HOWLETT NG and AVERY SV (1997b) Relationship be-tween cadmium sensitivity and degree of plasma membranefatty acid unsaturation in Saccharomyces cerevisiae ApplMicrobiol Biotechnol 48 539ndash545
HU Z CHANDRAN K GRASSO D and SMETS BF(2002a) Effect of nickel and cadmium speciation on nitrifi-cation inhibition Environ Sci Technol 36 3074ndash3078
HU Z CHADRAN K GRASSO D and SMETS BF(2003) Impact of heavy metal sorption and internalization onnitrification inhibition Environ Sci Technol 37 728ndash734
HU Z CHANDRAN K SMETS BF and GRASSO D(2002b) Evaluation of a rapid physical-chemical method forthe determination of extant soluble COD Water Res 36617ndash624
KARI FG and GIGER W (1996) Speciation and fate of ethylenediaminetetraacetate (EDTA) in municipal waste-water treatment Water Res 30 122ndash134
LEIVE L (1974) The barrier function of Gram-negative en-velope Ann NY Acad Sci 235 109ndash127
MARTEL AE SMITH RM and MOTEKAITIS RJ(1998) NIST Critical Stability Constants of Metal ComplexesDatabase College Station TX Texas AampM Univ
OHSUMI Y KITAMOTO K and ANRAKU Y (1988)Changes induced in the permeability barrier of the yeastplasma membrane by cupric ion J Bacteriol 170 2676ndash2682
PAINTER HA (1970) A review of literature on inorganic nitrogen metabolism in microorganisms Water Res 4 393ndash450
PIETSCH J SACHER F SCHMIDT W and BRAUCHH-J (2001) Polar nitrogen compounds and their behaviorin the drinking water treatment process Water Res 35 3537ndash3544
SCHECHER WD and MCAVOY DC (2001) MINEQL1A Chemical Equilibrium Modeling System Version 45 forWindows Hallowell ME Environmental Research Software
SCHOWANEK D MCAVOY D VERSTEEG D andHANSTVEIT A (1996) Effects of nutrient trace metal spe-ciation on algal growth in the presence of the chelator [SS]-EDDS Aquatic Toxicol 36 253ndash275
SHAMMAS NK (1986) Interactions of temperature pH andbiomass on the nitrification process J Water Pollut Con-trol Fed 58 52ndash58
SYKORA V PITTER P BITTNEROVA I and LEDERERT (2001) Biodegradability of ethylenediamine-based com-plexing agents Water Res 35 2010ndash2016
TCHOBANOGLOUS G and BURTON FL (1991) Waste-water Engineering Treatment Disposal and Reuse 3rd edNew York Metcalf amp Eddy McGraw-Hill
TEXIER A ANDRES Y ILLEMASSENE M andCLOIREC PL (2000) Characterization of lanthanide ionsbinding sites in the cell wall of Pseudomonos aeruginosaEnviron Sci Technol 34 610ndash615
TOMLINSON TG BOON AG and TROTMAN CNA(1966) Inhibition of nitrification in the activated sludge pro-cess of sewage disposal J Appl Bacteriol 29 266ndash291
US EPA (1993) Nitrogen control In EPA625R-93-0101Washington DC Office of Water
VANDEVIVERE P FICARA E TERRAS C JULIES Eand VERSTRAETE W (1998) Copper-mediated selectiveremoval of nitrification inhibitors from industrial waste-waters Environ Sci Technol 32 1000ndash1006
228 HU ET AL

cell surface of Gram-negative bacteria by interactionswith essential cellular cations on the surface (Ferris1989) EDTA especially scavenges Ca21 which is pre-dominantly responsible for outer membrane stabilizationThis can cause a concomitant release of significantamount of lipopolysaccharide (Leive 1974 Ferris1989) The inhibition of ammonium oxidizing activity byEDTA observed in this work is probably attributed to asimilar loss of essential cations from the outer membranebecause ammonium oxidizing bacteria exhibit the typi-cal Gram-negative cell membrane structure and correla-tions between the extent of inhibition and cellular Ca21
and Mg21 depletion were observedDTPA has a similar chemical structure to and a higher
molecular weight than EDTA By analogy one can spec-ulate that its transport across cellular membranes is in-significant The mode of inhibition of ammonium oxi-dizing activity by DTPA observed in this work isprobably similar to EDTA as the strong correlations be-tween the extent of inhibition and cellular Ca21 andMg21 depletion existed for both EDTA and DTPA (Fig3b and c)
Based on equilibrium speciation calculations everymol of chelating agents will complex 1 mol of Ca21 orMg21 at pH 75 At the same molar concentration EDAresulted in less cellular Ca21 depletion (Fig 4a) but muchhigher inhibition than EDTA or DTPA (Fig 2) Secondat the same molar concentration EDA resulted in morecellular Mg21 depletion than EDTA or DTPA (Fig 4b)yet the stability constant of MgEDA (1004) is muchsmaller than that of Mg EDTA (1088) or MgDTPA (1093)(Table 1) The results with EDA suggest that cellularMg21 depletion by EDA did not derive primarily fromMgndashEDA complexation Instead it is speculated due toleakage of destabilized cell membrane as EDA resultedin substantially higher cellular K1 depletion than EDTAor DTPA (Fig 4c) K1 was the most abundant cellularcation (on a molar basis data not shown) and is gener-ally believed to play an important role in maintenance ofthe ionic environment in the cytoplasm Depletion of cel-lular K1 indicates a loss of cell membrane integrity andviability (Ohsumi et al 1988 Avery et al 1996 Howlettand Avery 1997a)
Bacterial viability
Because depletion of cellular K1 has been widely usedas a marker of plasma membrane disruption (Howlett andAvery 1997b) we investigated directly whether disrup-tion of the plasma membrane integrity was observed af-ter exposure to the different agents In the presence of01 mM EDA 745 6 31 cells remained viable after15-min exposure At 2 mM EDA less than 113 6 00cells were viable compared to 875 6 111 461 6
108 viable cells in the presence of similar concentra-tions of EDTA and DTPA The biological effect of EDAtherefore is different among the chelators studied andmay involve cytoplasmic membrane rupture The ob-served loss of the plasma membrane integrity by 2 mMDTPA addition was unexpectedly high in view of the un-affected nitrifying activity at the same concentration (Fig2) Further experiments are needed to better understandwhat factors are responsible for the inconsistency be-tween the changes in the measured cellular activity(SOUR) and viability (livedead staining) after DTPAexposure
NITRIFICATION INHIBITION BY ETHYLENEDIAMINE-BASED CHELATING AGENTS 225
ENVIRON ENG SCI VOL 20 NO 3 2003
Figure 5 Inhibition of NH41-N oxidation by (a) 25 mM
EDTA or (b) 025 mM EDA in presence of metal ions Cu21
(d ) Fe31 ( s ) Ca21 ( j ) Mg21 ( u ) Bold horizontal lines rep-resent mean percent of inhibition in the presence of (a) EDTAor (b) EDA without metal addition
Inhibition mitigation by complexation
Appropriate remedial strategies to relieve nitrificationinhibition by chelatina agents were subsequently ex-plored Simultaneous addition of metals as coordinationpartners reduced the biological effects of chelatingagents but actual trends were ligand and metal specific(Fig 5)
At a 1 mM Fe31 dose (as FeCl3) nitrification inhibi-tion by 25 mM EDTA was completely relieved (Fig 5a)Similarly addition of Ca21 (as CaCl2) or Mg21 (as
MgSO4) reduced nitrification inhibition However Ca21
and Mg21 were slightly less effective than Fe31 on a mo-lar basis This may derive from EDTArsquos ability to formstronger complexes with the Lewis acid Fe compared toCa or Mg (Table 1) On the other hand addition of Cu21
(as CuCl2) only partially reduced nitrification inhibitionat a 11 ratio of CuEDTA (predominant speciesCuEDTA22) Copper itself inhibited the nitrifying activ-ity at high concentrations
In contrast nitrification inhibition by 025 mM EDAwas not relieved at any Fe31 dose tested (Fig 5b) Sim-ilarly addition of Ca21 or Mg21 did not reduce nitrifi-cation inhibition at the highest dose applied (5 mM)which is in agreement with their weak complexation po-tential (CaEDA K 5 1001 MgEDA K 5 1004) Theweak complexation behavior of EDA by Mg21 and thehigh depletion of cellular Mg21 by EDA further sug-gests that EDA has a different mode of inhibition thanEDTA Again addition of Cu21 only partially reducedthe inhibition with Cu(EDA)2
21 as the predominantspecies according to equilibrium speciation calcula-tions
Effect of exposure time on nitrification inhibition
To quantify the effect of exposure time on nitrifica-tion inhibition by chelating agents a predetermined
226 HU ET AL
Figure 7 Distribution of EDTA ( d ) in a continuous flowbioreactor after a transient dose of 24 mM EDTA within min-utes The continuous line presents the prediction from the ex-ponential decay model where C C0 are the reactor EDTA con-centrations at time t (day) and zero respectively F is the feedflow rate to the continuous flow reactor (10 Lday) and V isthe reactor volume (10 L)
Figure 6 Effect of exposure time on nitrification inhibitionby (a) EDTA or (b) EDA in batch assays Initial chelating agentconcentrations are shown Inhibition is expressed with refer-ence of SOUR of a culture incubated without chelating agentsto same duration after removal from parent reactor
dose of EDTA or EDA was applied in the batch reac-tor and the extent of inhibition measured as a functionof time Nitrification inhibition by EDTA reached its ul-timate value within 6 h (paired t-test p 025) (Fig6a) whereas inhibition by EDA substantially increasedup to 6 h (paired t-test p 005) and then decreased(Fig 6b) Such discrepancy between EDTA and EDAfurther suggests different modes of action for EDTA andEDA It also suggests that changes of the intracellularcation pool by EDA may have been slow analogous toslow kinetics of metal internalization reported else-where (Hu et al 2003)
Nitrification inhibition by EDTA in thecontinuous flow parent reactor
A shock load of EDTA was imposed to the continu-ously operated parent nitrifying bioreactor at 24 mM aconcentration corresponding to 47 6 2 inhibition in thebatch assay An ideal continuous flow reactor model ad-equately described the effluent EDTA (complexed anduncomplexed) profile (R2 5 099) computed from the ef-fluent COD data corrected for prepulse concentrations(Fig 7) which indicates that EDTA essentially behavedas a conservative compound did not partition to the cellsurface and was not subject to significant decay (Ferrisand Beveridge 1986 Sykora et al 2001)
Substantial foaming in the reactor was observed andlasted for 1 day after the EDTA shock load Recovery ofthe biological activity in the reactor was rapid At 01day reactor EDTA concentration dropped to 19 mM and23 6 1 inhibition of ammonium oxidation was mea-sured consistent with the extent of inhibition predictedfrom the batch assay (Fig 2) Effluent concentrations (1day after the shock load) indicated a return to stable operation (NH4
1-N NO22-N less than 01 mgL and
NO32-N ca 300 mgL)
Concomitantly reactor colloidal COD increasedslightly (from 6 to 23 6 6 mgL COD) after EDTA shockloads and returned to background level after 2 days Bio-mass floc size remained unchanged at 40 (62) mm (num-ber weighted mean floc size) from 5-day measurementsafter EDTA load compared to 41 (61) mm before dose
Implications for full-scale biological nitrogenremoval operation
The batch extant respirometric assay employed in thiswork can be used to ensure that the microbial physiol-ogy and activity remain close to that in the sampled con-tinuous flow reactor (Ellis et al 1996 Grady et al1996) From the study of EDTA in the continuous flowreactor the measured SOUR and subsequent inhibitionwas indeed consistent with the predicted value from the
batch assay No further inhibition was noticed and therecovery of the nitrifying activity in the continuous flowreactor was rapid Hence at least for compounds likeEDTA (and presumably DTPA) batch extant respiro-metric assays may accurately portray the nitrifying ac-tivity and inhibition in the continuous flow reactors
Although we only observed nitrification inhibition inboth batch and continuous flow reactors by EDTA whenits concentration was higher than 15 mM caution shouldbe employed in directly extending these results to full-scalewastewater treatment plants Inhibition may be primarilydependent on the concentration of uncomplexed EDTAspecies (Bucheli-Witschel and Egli 2001) which amongother physicochemical factors is often mediated by aque-ous Ca21 and Mg21 concentrations For instance equilib-rium speciation calculation revealed that the concentrationof uncomplexed [EDTA42] in our reactor was 10255 mMat 15 mM total EDTA The same uncomplexed EDTAconcentration could be obtained at 07 mM total EDTA attypical wastewater conditions of 16 mgL Ca21 and 10mgL Mg21 (Tchobanoglous and Burton 1991)
ACKNOWLEDGMENTS
We are grateful to Prof Diane J Burgess in the Dept ofPharmacy and Dr Michele Barber in the Dept of Molec-ular and Cell Biology at the University of Connecticut forassistance with particle sizing and flow cytometry respec-tively This study was funded in part by the ConnecticutDepartment of Environmental Protection through a USEPA Long Island Sound Study grant under Section 119 ofthe Clean Water Act and by the Connecticut Institute ofWater Resources under the 104B program of the NationalInstitutes of Water Resources US Geological Survey asauthorized by the Water Research and Development Act of1978 (PL 95-467) contract number 01HQGR0078
REFERENCES
ALDER AC SIEGRIST H GUJER W and GIGGER W(1990) Behavior of NTA and EDTA in biological waste-water treatment Water Res 24 733ndash742
APHA AWWA and WEF (1998) Standard Methods for theExamination of Water and Wastewater 20th ed Washing-ton DC author
AVERY SV HOWLETT NG and RADICE S (1996)Copper toxicity towards Saccharomyces cerevisiae Depen-dence on plasma membrane fatty acid composition Appl En-viron Microbiol 62 3960ndash3966
BLUM DJW and SPEECE RE (1991) A database ofchemical toxicity to environmental bacteria and its use in in-
NITRIFICATION INHIBITION BY ETHYLENEDIAMINE-BASED CHELATING AGENTS 227
ENVIRON ENG SCI VOL 20 NO 3 2003
terspecies comparisons and correlations J Water PollutControl Fed 63 198ndash207
BUCHELI-WITSCHEL M and EGLI T (2001) Environ-mental fate and microbial degradation of aminopolycar-boxylic acids FEMS Microbiol Rev 25 69ndash106
CHANDRAN K and SMETS BF (2000) Single-step nitri-fication models erroneously describe batch ammonia oxida-tion profiles when nitrite oxidation becomes rate limitingBiotechnol Bioeng 68 396ndash406
DAIGGER GT and SADICK TE (1998) Evaluation ofmethods to detect and control nitrification inhibition withspecific application to incinerator flue-gas scrubber waterWater Environ Res 70 1248ndash1257
ELLIS TG BARBEAU DS SMETS BF and GRADYLCP (1996) Respirometric technique for determination ofextant kinetic parameters describing biodegradation WaterEnviron Res 68 917ndash926
FERRIS FG (1989) Metallic ion interactions with outer mem-brane of Gram-negative bacteria In TJ Beveridge and RJDoyle Eds Metal Ions and Bacteria New York John Wi-ley and Sons pp 295ndash323
FERRIS FG and BEVERIDGE TJ (1986) Physicochemi-cal roles of soluble metal cations in the outer membrane ofEscherichia coli K-12 Can J Microbiol 32 594ndash601
FORD DL CHURCHWELL RL and KACHTICK JW(1980) Comprehensive analysis of nitrification of chemicalprocessing wastewaters J Water Pollut Control Fed 522726ndash2736
GERNAEY K VERSCHUERE L LUYTEN L and VER-STRAETE W (1997) Fast and sensitive acute toxicity de-tection with an enrichment nitrifying culture Water EnvironRes 69 1163ndash1169
GRADY CPL JR SMETS BF and BARBEAU DS (1996)Variability in kinetic parameter estimates A review of possiblecauses and a proposed terminology Water Res 30 742ndash748
HOCKENBURY MR and GRADY CPL (1977) Inhibi-tion of nitrification-effects of selected organic compoundsJ Water Pollut Control Fed 49 768ndash777
HOWLETT NG and AVERY SV (1997a) Induction oflipid peroxidation during heavy metal stress in Saccha-romyces cerevisiae and influence of plasma membrane fattyacid unsaturation Appl Environ Microbiol 63 2971ndash2976
HOWLETT NG and AVERY SV (1997b) Relationship be-tween cadmium sensitivity and degree of plasma membranefatty acid unsaturation in Saccharomyces cerevisiae ApplMicrobiol Biotechnol 48 539ndash545
HU Z CHANDRAN K GRASSO D and SMETS BF(2002a) Effect of nickel and cadmium speciation on nitrifi-cation inhibition Environ Sci Technol 36 3074ndash3078
HU Z CHADRAN K GRASSO D and SMETS BF(2003) Impact of heavy metal sorption and internalization onnitrification inhibition Environ Sci Technol 37 728ndash734
HU Z CHANDRAN K SMETS BF and GRASSO D(2002b) Evaluation of a rapid physical-chemical method forthe determination of extant soluble COD Water Res 36617ndash624
KARI FG and GIGER W (1996) Speciation and fate of ethylenediaminetetraacetate (EDTA) in municipal waste-water treatment Water Res 30 122ndash134
LEIVE L (1974) The barrier function of Gram-negative en-velope Ann NY Acad Sci 235 109ndash127
MARTEL AE SMITH RM and MOTEKAITIS RJ(1998) NIST Critical Stability Constants of Metal ComplexesDatabase College Station TX Texas AampM Univ
OHSUMI Y KITAMOTO K and ANRAKU Y (1988)Changes induced in the permeability barrier of the yeastplasma membrane by cupric ion J Bacteriol 170 2676ndash2682
PAINTER HA (1970) A review of literature on inorganic nitrogen metabolism in microorganisms Water Res 4 393ndash450
PIETSCH J SACHER F SCHMIDT W and BRAUCHH-J (2001) Polar nitrogen compounds and their behaviorin the drinking water treatment process Water Res 35 3537ndash3544
SCHECHER WD and MCAVOY DC (2001) MINEQL1A Chemical Equilibrium Modeling System Version 45 forWindows Hallowell ME Environmental Research Software
SCHOWANEK D MCAVOY D VERSTEEG D andHANSTVEIT A (1996) Effects of nutrient trace metal spe-ciation on algal growth in the presence of the chelator [SS]-EDDS Aquatic Toxicol 36 253ndash275
SHAMMAS NK (1986) Interactions of temperature pH andbiomass on the nitrification process J Water Pollut Con-trol Fed 58 52ndash58
SYKORA V PITTER P BITTNEROVA I and LEDERERT (2001) Biodegradability of ethylenediamine-based com-plexing agents Water Res 35 2010ndash2016
TCHOBANOGLOUS G and BURTON FL (1991) Waste-water Engineering Treatment Disposal and Reuse 3rd edNew York Metcalf amp Eddy McGraw-Hill
TEXIER A ANDRES Y ILLEMASSENE M andCLOIREC PL (2000) Characterization of lanthanide ionsbinding sites in the cell wall of Pseudomonos aeruginosaEnviron Sci Technol 34 610ndash615
TOMLINSON TG BOON AG and TROTMAN CNA(1966) Inhibition of nitrification in the activated sludge pro-cess of sewage disposal J Appl Bacteriol 29 266ndash291
US EPA (1993) Nitrogen control In EPA625R-93-0101Washington DC Office of Water
VANDEVIVERE P FICARA E TERRAS C JULIES Eand VERSTRAETE W (1998) Copper-mediated selectiveremoval of nitrification inhibitors from industrial waste-waters Environ Sci Technol 32 1000ndash1006
228 HU ET AL

Inhibition mitigation by complexation
Appropriate remedial strategies to relieve nitrificationinhibition by chelatina agents were subsequently ex-plored Simultaneous addition of metals as coordinationpartners reduced the biological effects of chelatingagents but actual trends were ligand and metal specific(Fig 5)
At a 1 mM Fe31 dose (as FeCl3) nitrification inhibi-tion by 25 mM EDTA was completely relieved (Fig 5a)Similarly addition of Ca21 (as CaCl2) or Mg21 (as
MgSO4) reduced nitrification inhibition However Ca21
and Mg21 were slightly less effective than Fe31 on a mo-lar basis This may derive from EDTArsquos ability to formstronger complexes with the Lewis acid Fe compared toCa or Mg (Table 1) On the other hand addition of Cu21
(as CuCl2) only partially reduced nitrification inhibitionat a 11 ratio of CuEDTA (predominant speciesCuEDTA22) Copper itself inhibited the nitrifying activ-ity at high concentrations
In contrast nitrification inhibition by 025 mM EDAwas not relieved at any Fe31 dose tested (Fig 5b) Sim-ilarly addition of Ca21 or Mg21 did not reduce nitrifi-cation inhibition at the highest dose applied (5 mM)which is in agreement with their weak complexation po-tential (CaEDA K 5 1001 MgEDA K 5 1004) Theweak complexation behavior of EDA by Mg21 and thehigh depletion of cellular Mg21 by EDA further sug-gests that EDA has a different mode of inhibition thanEDTA Again addition of Cu21 only partially reducedthe inhibition with Cu(EDA)2
21 as the predominantspecies according to equilibrium speciation calcula-tions
Effect of exposure time on nitrification inhibition
To quantify the effect of exposure time on nitrifica-tion inhibition by chelating agents a predetermined
226 HU ET AL
Figure 7 Distribution of EDTA ( d ) in a continuous flowbioreactor after a transient dose of 24 mM EDTA within min-utes The continuous line presents the prediction from the ex-ponential decay model where C C0 are the reactor EDTA con-centrations at time t (day) and zero respectively F is the feedflow rate to the continuous flow reactor (10 Lday) and V isthe reactor volume (10 L)
Figure 6 Effect of exposure time on nitrification inhibitionby (a) EDTA or (b) EDA in batch assays Initial chelating agentconcentrations are shown Inhibition is expressed with refer-ence of SOUR of a culture incubated without chelating agentsto same duration after removal from parent reactor
dose of EDTA or EDA was applied in the batch reac-tor and the extent of inhibition measured as a functionof time Nitrification inhibition by EDTA reached its ul-timate value within 6 h (paired t-test p 025) (Fig6a) whereas inhibition by EDA substantially increasedup to 6 h (paired t-test p 005) and then decreased(Fig 6b) Such discrepancy between EDTA and EDAfurther suggests different modes of action for EDTA andEDA It also suggests that changes of the intracellularcation pool by EDA may have been slow analogous toslow kinetics of metal internalization reported else-where (Hu et al 2003)
Nitrification inhibition by EDTA in thecontinuous flow parent reactor
A shock load of EDTA was imposed to the continu-ously operated parent nitrifying bioreactor at 24 mM aconcentration corresponding to 47 6 2 inhibition in thebatch assay An ideal continuous flow reactor model ad-equately described the effluent EDTA (complexed anduncomplexed) profile (R2 5 099) computed from the ef-fluent COD data corrected for prepulse concentrations(Fig 7) which indicates that EDTA essentially behavedas a conservative compound did not partition to the cellsurface and was not subject to significant decay (Ferrisand Beveridge 1986 Sykora et al 2001)
Substantial foaming in the reactor was observed andlasted for 1 day after the EDTA shock load Recovery ofthe biological activity in the reactor was rapid At 01day reactor EDTA concentration dropped to 19 mM and23 6 1 inhibition of ammonium oxidation was mea-sured consistent with the extent of inhibition predictedfrom the batch assay (Fig 2) Effluent concentrations (1day after the shock load) indicated a return to stable operation (NH4
1-N NO22-N less than 01 mgL and
NO32-N ca 300 mgL)
Concomitantly reactor colloidal COD increasedslightly (from 6 to 23 6 6 mgL COD) after EDTA shockloads and returned to background level after 2 days Bio-mass floc size remained unchanged at 40 (62) mm (num-ber weighted mean floc size) from 5-day measurementsafter EDTA load compared to 41 (61) mm before dose
Implications for full-scale biological nitrogenremoval operation
The batch extant respirometric assay employed in thiswork can be used to ensure that the microbial physiol-ogy and activity remain close to that in the sampled con-tinuous flow reactor (Ellis et al 1996 Grady et al1996) From the study of EDTA in the continuous flowreactor the measured SOUR and subsequent inhibitionwas indeed consistent with the predicted value from the
batch assay No further inhibition was noticed and therecovery of the nitrifying activity in the continuous flowreactor was rapid Hence at least for compounds likeEDTA (and presumably DTPA) batch extant respiro-metric assays may accurately portray the nitrifying ac-tivity and inhibition in the continuous flow reactors
Although we only observed nitrification inhibition inboth batch and continuous flow reactors by EDTA whenits concentration was higher than 15 mM caution shouldbe employed in directly extending these results to full-scalewastewater treatment plants Inhibition may be primarilydependent on the concentration of uncomplexed EDTAspecies (Bucheli-Witschel and Egli 2001) which amongother physicochemical factors is often mediated by aque-ous Ca21 and Mg21 concentrations For instance equilib-rium speciation calculation revealed that the concentrationof uncomplexed [EDTA42] in our reactor was 10255 mMat 15 mM total EDTA The same uncomplexed EDTAconcentration could be obtained at 07 mM total EDTA attypical wastewater conditions of 16 mgL Ca21 and 10mgL Mg21 (Tchobanoglous and Burton 1991)
ACKNOWLEDGMENTS
We are grateful to Prof Diane J Burgess in the Dept ofPharmacy and Dr Michele Barber in the Dept of Molec-ular and Cell Biology at the University of Connecticut forassistance with particle sizing and flow cytometry respec-tively This study was funded in part by the ConnecticutDepartment of Environmental Protection through a USEPA Long Island Sound Study grant under Section 119 ofthe Clean Water Act and by the Connecticut Institute ofWater Resources under the 104B program of the NationalInstitutes of Water Resources US Geological Survey asauthorized by the Water Research and Development Act of1978 (PL 95-467) contract number 01HQGR0078
REFERENCES
ALDER AC SIEGRIST H GUJER W and GIGGER W(1990) Behavior of NTA and EDTA in biological waste-water treatment Water Res 24 733ndash742
APHA AWWA and WEF (1998) Standard Methods for theExamination of Water and Wastewater 20th ed Washing-ton DC author
AVERY SV HOWLETT NG and RADICE S (1996)Copper toxicity towards Saccharomyces cerevisiae Depen-dence on plasma membrane fatty acid composition Appl En-viron Microbiol 62 3960ndash3966
BLUM DJW and SPEECE RE (1991) A database ofchemical toxicity to environmental bacteria and its use in in-
NITRIFICATION INHIBITION BY ETHYLENEDIAMINE-BASED CHELATING AGENTS 227
ENVIRON ENG SCI VOL 20 NO 3 2003
terspecies comparisons and correlations J Water PollutControl Fed 63 198ndash207
BUCHELI-WITSCHEL M and EGLI T (2001) Environ-mental fate and microbial degradation of aminopolycar-boxylic acids FEMS Microbiol Rev 25 69ndash106
CHANDRAN K and SMETS BF (2000) Single-step nitri-fication models erroneously describe batch ammonia oxida-tion profiles when nitrite oxidation becomes rate limitingBiotechnol Bioeng 68 396ndash406
DAIGGER GT and SADICK TE (1998) Evaluation ofmethods to detect and control nitrification inhibition withspecific application to incinerator flue-gas scrubber waterWater Environ Res 70 1248ndash1257
ELLIS TG BARBEAU DS SMETS BF and GRADYLCP (1996) Respirometric technique for determination ofextant kinetic parameters describing biodegradation WaterEnviron Res 68 917ndash926
FERRIS FG (1989) Metallic ion interactions with outer mem-brane of Gram-negative bacteria In TJ Beveridge and RJDoyle Eds Metal Ions and Bacteria New York John Wi-ley and Sons pp 295ndash323
FERRIS FG and BEVERIDGE TJ (1986) Physicochemi-cal roles of soluble metal cations in the outer membrane ofEscherichia coli K-12 Can J Microbiol 32 594ndash601
FORD DL CHURCHWELL RL and KACHTICK JW(1980) Comprehensive analysis of nitrification of chemicalprocessing wastewaters J Water Pollut Control Fed 522726ndash2736
GERNAEY K VERSCHUERE L LUYTEN L and VER-STRAETE W (1997) Fast and sensitive acute toxicity de-tection with an enrichment nitrifying culture Water EnvironRes 69 1163ndash1169
GRADY CPL JR SMETS BF and BARBEAU DS (1996)Variability in kinetic parameter estimates A review of possiblecauses and a proposed terminology Water Res 30 742ndash748
HOCKENBURY MR and GRADY CPL (1977) Inhibi-tion of nitrification-effects of selected organic compoundsJ Water Pollut Control Fed 49 768ndash777
HOWLETT NG and AVERY SV (1997a) Induction oflipid peroxidation during heavy metal stress in Saccha-romyces cerevisiae and influence of plasma membrane fattyacid unsaturation Appl Environ Microbiol 63 2971ndash2976
HOWLETT NG and AVERY SV (1997b) Relationship be-tween cadmium sensitivity and degree of plasma membranefatty acid unsaturation in Saccharomyces cerevisiae ApplMicrobiol Biotechnol 48 539ndash545
HU Z CHANDRAN K GRASSO D and SMETS BF(2002a) Effect of nickel and cadmium speciation on nitrifi-cation inhibition Environ Sci Technol 36 3074ndash3078
HU Z CHADRAN K GRASSO D and SMETS BF(2003) Impact of heavy metal sorption and internalization onnitrification inhibition Environ Sci Technol 37 728ndash734
HU Z CHANDRAN K SMETS BF and GRASSO D(2002b) Evaluation of a rapid physical-chemical method forthe determination of extant soluble COD Water Res 36617ndash624
KARI FG and GIGER W (1996) Speciation and fate of ethylenediaminetetraacetate (EDTA) in municipal waste-water treatment Water Res 30 122ndash134
LEIVE L (1974) The barrier function of Gram-negative en-velope Ann NY Acad Sci 235 109ndash127
MARTEL AE SMITH RM and MOTEKAITIS RJ(1998) NIST Critical Stability Constants of Metal ComplexesDatabase College Station TX Texas AampM Univ
OHSUMI Y KITAMOTO K and ANRAKU Y (1988)Changes induced in the permeability barrier of the yeastplasma membrane by cupric ion J Bacteriol 170 2676ndash2682
PAINTER HA (1970) A review of literature on inorganic nitrogen metabolism in microorganisms Water Res 4 393ndash450
PIETSCH J SACHER F SCHMIDT W and BRAUCHH-J (2001) Polar nitrogen compounds and their behaviorin the drinking water treatment process Water Res 35 3537ndash3544
SCHECHER WD and MCAVOY DC (2001) MINEQL1A Chemical Equilibrium Modeling System Version 45 forWindows Hallowell ME Environmental Research Software
SCHOWANEK D MCAVOY D VERSTEEG D andHANSTVEIT A (1996) Effects of nutrient trace metal spe-ciation on algal growth in the presence of the chelator [SS]-EDDS Aquatic Toxicol 36 253ndash275
SHAMMAS NK (1986) Interactions of temperature pH andbiomass on the nitrification process J Water Pollut Con-trol Fed 58 52ndash58
SYKORA V PITTER P BITTNEROVA I and LEDERERT (2001) Biodegradability of ethylenediamine-based com-plexing agents Water Res 35 2010ndash2016
TCHOBANOGLOUS G and BURTON FL (1991) Waste-water Engineering Treatment Disposal and Reuse 3rd edNew York Metcalf amp Eddy McGraw-Hill
TEXIER A ANDRES Y ILLEMASSENE M andCLOIREC PL (2000) Characterization of lanthanide ionsbinding sites in the cell wall of Pseudomonos aeruginosaEnviron Sci Technol 34 610ndash615
TOMLINSON TG BOON AG and TROTMAN CNA(1966) Inhibition of nitrification in the activated sludge pro-cess of sewage disposal J Appl Bacteriol 29 266ndash291
US EPA (1993) Nitrogen control In EPA625R-93-0101Washington DC Office of Water
VANDEVIVERE P FICARA E TERRAS C JULIES Eand VERSTRAETE W (1998) Copper-mediated selectiveremoval of nitrification inhibitors from industrial waste-waters Environ Sci Technol 32 1000ndash1006
228 HU ET AL

dose of EDTA or EDA was applied in the batch reac-tor and the extent of inhibition measured as a functionof time Nitrification inhibition by EDTA reached its ul-timate value within 6 h (paired t-test p 025) (Fig6a) whereas inhibition by EDA substantially increasedup to 6 h (paired t-test p 005) and then decreased(Fig 6b) Such discrepancy between EDTA and EDAfurther suggests different modes of action for EDTA andEDA It also suggests that changes of the intracellularcation pool by EDA may have been slow analogous toslow kinetics of metal internalization reported else-where (Hu et al 2003)
Nitrification inhibition by EDTA in thecontinuous flow parent reactor
A shock load of EDTA was imposed to the continu-ously operated parent nitrifying bioreactor at 24 mM aconcentration corresponding to 47 6 2 inhibition in thebatch assay An ideal continuous flow reactor model ad-equately described the effluent EDTA (complexed anduncomplexed) profile (R2 5 099) computed from the ef-fluent COD data corrected for prepulse concentrations(Fig 7) which indicates that EDTA essentially behavedas a conservative compound did not partition to the cellsurface and was not subject to significant decay (Ferrisand Beveridge 1986 Sykora et al 2001)
Substantial foaming in the reactor was observed andlasted for 1 day after the EDTA shock load Recovery ofthe biological activity in the reactor was rapid At 01day reactor EDTA concentration dropped to 19 mM and23 6 1 inhibition of ammonium oxidation was mea-sured consistent with the extent of inhibition predictedfrom the batch assay (Fig 2) Effluent concentrations (1day after the shock load) indicated a return to stable operation (NH4
1-N NO22-N less than 01 mgL and
NO32-N ca 300 mgL)
Concomitantly reactor colloidal COD increasedslightly (from 6 to 23 6 6 mgL COD) after EDTA shockloads and returned to background level after 2 days Bio-mass floc size remained unchanged at 40 (62) mm (num-ber weighted mean floc size) from 5-day measurementsafter EDTA load compared to 41 (61) mm before dose
Implications for full-scale biological nitrogenremoval operation
The batch extant respirometric assay employed in thiswork can be used to ensure that the microbial physiol-ogy and activity remain close to that in the sampled con-tinuous flow reactor (Ellis et al 1996 Grady et al1996) From the study of EDTA in the continuous flowreactor the measured SOUR and subsequent inhibitionwas indeed consistent with the predicted value from the
batch assay No further inhibition was noticed and therecovery of the nitrifying activity in the continuous flowreactor was rapid Hence at least for compounds likeEDTA (and presumably DTPA) batch extant respiro-metric assays may accurately portray the nitrifying ac-tivity and inhibition in the continuous flow reactors
Although we only observed nitrification inhibition inboth batch and continuous flow reactors by EDTA whenits concentration was higher than 15 mM caution shouldbe employed in directly extending these results to full-scalewastewater treatment plants Inhibition may be primarilydependent on the concentration of uncomplexed EDTAspecies (Bucheli-Witschel and Egli 2001) which amongother physicochemical factors is often mediated by aque-ous Ca21 and Mg21 concentrations For instance equilib-rium speciation calculation revealed that the concentrationof uncomplexed [EDTA42] in our reactor was 10255 mMat 15 mM total EDTA The same uncomplexed EDTAconcentration could be obtained at 07 mM total EDTA attypical wastewater conditions of 16 mgL Ca21 and 10mgL Mg21 (Tchobanoglous and Burton 1991)
ACKNOWLEDGMENTS
We are grateful to Prof Diane J Burgess in the Dept ofPharmacy and Dr Michele Barber in the Dept of Molec-ular and Cell Biology at the University of Connecticut forassistance with particle sizing and flow cytometry respec-tively This study was funded in part by the ConnecticutDepartment of Environmental Protection through a USEPA Long Island Sound Study grant under Section 119 ofthe Clean Water Act and by the Connecticut Institute ofWater Resources under the 104B program of the NationalInstitutes of Water Resources US Geological Survey asauthorized by the Water Research and Development Act of1978 (PL 95-467) contract number 01HQGR0078
REFERENCES
ALDER AC SIEGRIST H GUJER W and GIGGER W(1990) Behavior of NTA and EDTA in biological waste-water treatment Water Res 24 733ndash742
APHA AWWA and WEF (1998) Standard Methods for theExamination of Water and Wastewater 20th ed Washing-ton DC author
AVERY SV HOWLETT NG and RADICE S (1996)Copper toxicity towards Saccharomyces cerevisiae Depen-dence on plasma membrane fatty acid composition Appl En-viron Microbiol 62 3960ndash3966
BLUM DJW and SPEECE RE (1991) A database ofchemical toxicity to environmental bacteria and its use in in-
NITRIFICATION INHIBITION BY ETHYLENEDIAMINE-BASED CHELATING AGENTS 227
ENVIRON ENG SCI VOL 20 NO 3 2003
terspecies comparisons and correlations J Water PollutControl Fed 63 198ndash207
BUCHELI-WITSCHEL M and EGLI T (2001) Environ-mental fate and microbial degradation of aminopolycar-boxylic acids FEMS Microbiol Rev 25 69ndash106
CHANDRAN K and SMETS BF (2000) Single-step nitri-fication models erroneously describe batch ammonia oxida-tion profiles when nitrite oxidation becomes rate limitingBiotechnol Bioeng 68 396ndash406
DAIGGER GT and SADICK TE (1998) Evaluation ofmethods to detect and control nitrification inhibition withspecific application to incinerator flue-gas scrubber waterWater Environ Res 70 1248ndash1257
ELLIS TG BARBEAU DS SMETS BF and GRADYLCP (1996) Respirometric technique for determination ofextant kinetic parameters describing biodegradation WaterEnviron Res 68 917ndash926
FERRIS FG (1989) Metallic ion interactions with outer mem-brane of Gram-negative bacteria In TJ Beveridge and RJDoyle Eds Metal Ions and Bacteria New York John Wi-ley and Sons pp 295ndash323
FERRIS FG and BEVERIDGE TJ (1986) Physicochemi-cal roles of soluble metal cations in the outer membrane ofEscherichia coli K-12 Can J Microbiol 32 594ndash601
FORD DL CHURCHWELL RL and KACHTICK JW(1980) Comprehensive analysis of nitrification of chemicalprocessing wastewaters J Water Pollut Control Fed 522726ndash2736
GERNAEY K VERSCHUERE L LUYTEN L and VER-STRAETE W (1997) Fast and sensitive acute toxicity de-tection with an enrichment nitrifying culture Water EnvironRes 69 1163ndash1169
GRADY CPL JR SMETS BF and BARBEAU DS (1996)Variability in kinetic parameter estimates A review of possiblecauses and a proposed terminology Water Res 30 742ndash748
HOCKENBURY MR and GRADY CPL (1977) Inhibi-tion of nitrification-effects of selected organic compoundsJ Water Pollut Control Fed 49 768ndash777
HOWLETT NG and AVERY SV (1997a) Induction oflipid peroxidation during heavy metal stress in Saccha-romyces cerevisiae and influence of plasma membrane fattyacid unsaturation Appl Environ Microbiol 63 2971ndash2976
HOWLETT NG and AVERY SV (1997b) Relationship be-tween cadmium sensitivity and degree of plasma membranefatty acid unsaturation in Saccharomyces cerevisiae ApplMicrobiol Biotechnol 48 539ndash545
HU Z CHANDRAN K GRASSO D and SMETS BF(2002a) Effect of nickel and cadmium speciation on nitrifi-cation inhibition Environ Sci Technol 36 3074ndash3078
HU Z CHADRAN K GRASSO D and SMETS BF(2003) Impact of heavy metal sorption and internalization onnitrification inhibition Environ Sci Technol 37 728ndash734
HU Z CHANDRAN K SMETS BF and GRASSO D(2002b) Evaluation of a rapid physical-chemical method forthe determination of extant soluble COD Water Res 36617ndash624
KARI FG and GIGER W (1996) Speciation and fate of ethylenediaminetetraacetate (EDTA) in municipal waste-water treatment Water Res 30 122ndash134
LEIVE L (1974) The barrier function of Gram-negative en-velope Ann NY Acad Sci 235 109ndash127
MARTEL AE SMITH RM and MOTEKAITIS RJ(1998) NIST Critical Stability Constants of Metal ComplexesDatabase College Station TX Texas AampM Univ
OHSUMI Y KITAMOTO K and ANRAKU Y (1988)Changes induced in the permeability barrier of the yeastplasma membrane by cupric ion J Bacteriol 170 2676ndash2682
PAINTER HA (1970) A review of literature on inorganic nitrogen metabolism in microorganisms Water Res 4 393ndash450
PIETSCH J SACHER F SCHMIDT W and BRAUCHH-J (2001) Polar nitrogen compounds and their behaviorin the drinking water treatment process Water Res 35 3537ndash3544
SCHECHER WD and MCAVOY DC (2001) MINEQL1A Chemical Equilibrium Modeling System Version 45 forWindows Hallowell ME Environmental Research Software
SCHOWANEK D MCAVOY D VERSTEEG D andHANSTVEIT A (1996) Effects of nutrient trace metal spe-ciation on algal growth in the presence of the chelator [SS]-EDDS Aquatic Toxicol 36 253ndash275
SHAMMAS NK (1986) Interactions of temperature pH andbiomass on the nitrification process J Water Pollut Con-trol Fed 58 52ndash58
SYKORA V PITTER P BITTNEROVA I and LEDERERT (2001) Biodegradability of ethylenediamine-based com-plexing agents Water Res 35 2010ndash2016
TCHOBANOGLOUS G and BURTON FL (1991) Waste-water Engineering Treatment Disposal and Reuse 3rd edNew York Metcalf amp Eddy McGraw-Hill
TEXIER A ANDRES Y ILLEMASSENE M andCLOIREC PL (2000) Characterization of lanthanide ionsbinding sites in the cell wall of Pseudomonos aeruginosaEnviron Sci Technol 34 610ndash615
TOMLINSON TG BOON AG and TROTMAN CNA(1966) Inhibition of nitrification in the activated sludge pro-cess of sewage disposal J Appl Bacteriol 29 266ndash291
US EPA (1993) Nitrogen control In EPA625R-93-0101Washington DC Office of Water
VANDEVIVERE P FICARA E TERRAS C JULIES Eand VERSTRAETE W (1998) Copper-mediated selectiveremoval of nitrification inhibitors from industrial waste-waters Environ Sci Technol 32 1000ndash1006
228 HU ET AL

terspecies comparisons and correlations J Water PollutControl Fed 63 198ndash207
BUCHELI-WITSCHEL M and EGLI T (2001) Environ-mental fate and microbial degradation of aminopolycar-boxylic acids FEMS Microbiol Rev 25 69ndash106
CHANDRAN K and SMETS BF (2000) Single-step nitri-fication models erroneously describe batch ammonia oxida-tion profiles when nitrite oxidation becomes rate limitingBiotechnol Bioeng 68 396ndash406
DAIGGER GT and SADICK TE (1998) Evaluation ofmethods to detect and control nitrification inhibition withspecific application to incinerator flue-gas scrubber waterWater Environ Res 70 1248ndash1257
ELLIS TG BARBEAU DS SMETS BF and GRADYLCP (1996) Respirometric technique for determination ofextant kinetic parameters describing biodegradation WaterEnviron Res 68 917ndash926
FERRIS FG (1989) Metallic ion interactions with outer mem-brane of Gram-negative bacteria In TJ Beveridge and RJDoyle Eds Metal Ions and Bacteria New York John Wi-ley and Sons pp 295ndash323
FERRIS FG and BEVERIDGE TJ (1986) Physicochemi-cal roles of soluble metal cations in the outer membrane ofEscherichia coli K-12 Can J Microbiol 32 594ndash601
FORD DL CHURCHWELL RL and KACHTICK JW(1980) Comprehensive analysis of nitrification of chemicalprocessing wastewaters J Water Pollut Control Fed 522726ndash2736
GERNAEY K VERSCHUERE L LUYTEN L and VER-STRAETE W (1997) Fast and sensitive acute toxicity de-tection with an enrichment nitrifying culture Water EnvironRes 69 1163ndash1169
GRADY CPL JR SMETS BF and BARBEAU DS (1996)Variability in kinetic parameter estimates A review of possiblecauses and a proposed terminology Water Res 30 742ndash748
HOCKENBURY MR and GRADY CPL (1977) Inhibi-tion of nitrification-effects of selected organic compoundsJ Water Pollut Control Fed 49 768ndash777
HOWLETT NG and AVERY SV (1997a) Induction oflipid peroxidation during heavy metal stress in Saccha-romyces cerevisiae and influence of plasma membrane fattyacid unsaturation Appl Environ Microbiol 63 2971ndash2976
HOWLETT NG and AVERY SV (1997b) Relationship be-tween cadmium sensitivity and degree of plasma membranefatty acid unsaturation in Saccharomyces cerevisiae ApplMicrobiol Biotechnol 48 539ndash545
HU Z CHANDRAN K GRASSO D and SMETS BF(2002a) Effect of nickel and cadmium speciation on nitrifi-cation inhibition Environ Sci Technol 36 3074ndash3078
HU Z CHADRAN K GRASSO D and SMETS BF(2003) Impact of heavy metal sorption and internalization onnitrification inhibition Environ Sci Technol 37 728ndash734
HU Z CHANDRAN K SMETS BF and GRASSO D(2002b) Evaluation of a rapid physical-chemical method forthe determination of extant soluble COD Water Res 36617ndash624
KARI FG and GIGER W (1996) Speciation and fate of ethylenediaminetetraacetate (EDTA) in municipal waste-water treatment Water Res 30 122ndash134
LEIVE L (1974) The barrier function of Gram-negative en-velope Ann NY Acad Sci 235 109ndash127
MARTEL AE SMITH RM and MOTEKAITIS RJ(1998) NIST Critical Stability Constants of Metal ComplexesDatabase College Station TX Texas AampM Univ
OHSUMI Y KITAMOTO K and ANRAKU Y (1988)Changes induced in the permeability barrier of the yeastplasma membrane by cupric ion J Bacteriol 170 2676ndash2682
PAINTER HA (1970) A review of literature on inorganic nitrogen metabolism in microorganisms Water Res 4 393ndash450
PIETSCH J SACHER F SCHMIDT W and BRAUCHH-J (2001) Polar nitrogen compounds and their behaviorin the drinking water treatment process Water Res 35 3537ndash3544
SCHECHER WD and MCAVOY DC (2001) MINEQL1A Chemical Equilibrium Modeling System Version 45 forWindows Hallowell ME Environmental Research Software
SCHOWANEK D MCAVOY D VERSTEEG D andHANSTVEIT A (1996) Effects of nutrient trace metal spe-ciation on algal growth in the presence of the chelator [SS]-EDDS Aquatic Toxicol 36 253ndash275
SHAMMAS NK (1986) Interactions of temperature pH andbiomass on the nitrification process J Water Pollut Con-trol Fed 58 52ndash58
SYKORA V PITTER P BITTNEROVA I and LEDERERT (2001) Biodegradability of ethylenediamine-based com-plexing agents Water Res 35 2010ndash2016
TCHOBANOGLOUS G and BURTON FL (1991) Waste-water Engineering Treatment Disposal and Reuse 3rd edNew York Metcalf amp Eddy McGraw-Hill
TEXIER A ANDRES Y ILLEMASSENE M andCLOIREC PL (2000) Characterization of lanthanide ionsbinding sites in the cell wall of Pseudomonos aeruginosaEnviron Sci Technol 34 610ndash615
TOMLINSON TG BOON AG and TROTMAN CNA(1966) Inhibition of nitrification in the activated sludge pro-cess of sewage disposal J Appl Bacteriol 29 266ndash291
US EPA (1993) Nitrogen control In EPA625R-93-0101Washington DC Office of Water
VANDEVIVERE P FICARA E TERRAS C JULIES Eand VERSTRAETE W (1998) Copper-mediated selectiveremoval of nitrification inhibitors from industrial waste-waters Environ Sci Technol 32 1000ndash1006
228 HU ET AL
![Synthesis of chelating trimethylsiloxy diisonitrile ligands and their rhodium(1) complexes. Crystal and molecular structure of [Rh(t-BuDiNC)2]BPh4·1.5CH3CN (DiNC = chelating bidentate](https://img.pdfslide.net/doc/110x75/6352222a16edf97a790cce1e/synthesis-of-chelating-trimethylsiloxy-diisonitrile-ligands-and-their-rhodium1.jpg)


![Azido{ N , N ′-bis[3-(2-nitrophenyl)prop-2-enylidene]ethylenediamine}(triphenylphosphine)copper(I) chloroform solvate](https://img.pdfslide.net/doc/110x75/63200131962ed4ca8e03f617/azido-n-n-bis3-2-nitrophenylprop-2-enylideneethylenediaminetriphenylphosphinecopperi.jpg)












![μ-Peroxido-bis[acetonitrilebis(ethylenediamine)cobalt(III)] tetrakis(perchlorate)](https://img.pdfslide.net/doc/110x75/63259ac4545c645c7f09c93d/m-peroxido-bisacetonitrilebisethylenediaminecobaltiii-tetrakisperchlorate.jpg)









copper(I](https://img.pdfslide.net/doc/110x75/6320014ffdf36d7df6037ff7/bromido-n-n-bis2-nitrocinnamaldehydeethylenediaminetriphenylphosphinecopperi.jpg)


