Embed Size (px)
Citation preview

Research Article
Olive oil protects against 2,4-dichlorophenoxyaceticacid-induced oxidative renal dysfunction in adult rats
Amel Nakbi1, Wafa Tayeb1, Samia Dabbou1, Issam Chargui2, Manel Issaoui1, Ameur Ferih1,
Zohra Ben Ali2, Mohammed A. Alsaif3 and Mohamed Hammami1,3
1 Biochemistry Laboratory, UR03ES08 ‘Human Nutrition & Metabolic Disorders’, Faculty of Medicine,
University of Monastir, Monastir, Tunisia2 Laboratory of Histology and Cytogenetic, Faculty of Medicine of Monastir, Monastir, Tunisia3 College of Applied Medical Sciences, VPP Unit, King Saud University, Riyadh, Saudi Arabia
This study was aimed to evaluate the protective effect of extra virgin olive oil (EVOO) and its fractions
(hydrophilic (HF) and lipophilic (LF)) treatment on 2,4-dichlorophenoxiacetic acid (2,4-D)-induced
nephrotoxicity in rats. The Wistar rats (n ¼ 80, 210–230 g) were divided into eight groups having ten
animals, i.e., control group (C), (2,4-D) group that received 2,4-D (5 mg/kg b.w.), (2,4-D/EVOO)
group was treated with 2,4-D plus EVOO, (2,4-D/olive oil hydrophilic fraction, OOHF) group that
received 2,4-D plus HF, (2,4-D/olive oil lipophilic fraction, OOLF) group was treated with 2,4-D plus
LF, (EVOO) group that received only EVOO, (OOHF) group was given HF and (OOLF) group was
treated with the LF. These components were administered daily by gavage for 4 wk. 2,4-D administration
affected plasma urea and creatinine levels, which increased while uric acid significantly decreased. A
marked increase of malondialdehyde level and with a significant decrease in enzymatic antioxidant’s
activities, were also observed in 2,4-D treated rats. The co-administration of EVOO and its fractions
along with 2,4-D resulted in a reversal of 2,4-D induced biochemical changes in kidney accompanied by a
significant decrease in lipid peroxidation and an increase in the level of the antioxidant defence system.
Light microscopy investigations revealed that 2,4-D exposure induced numerous histopathological
alterations in the kidney. EVOO administration with 2,4-D severely reduced the toxicity of 2,4-D
and preserved the normal histological architecture of the renal tissue. However, a milder
histopathological improvement was observed in animals co-treated with HF or LF plus 2,4-D. In
conclusion, the present study suggest that the nephroprotective potential of EVOO against 2,4-D
toxicity might be due to the synergic effect of the two fractions, which could be useful for achieving
optimum effects in 2,4-D-induced renal damage.
Keywords: 2,4-D / Kidney / Olive oil / Oxidative stress / Rat
Received: June 20, 2011 / Revised: October 13, 2011 / Accepted: November 8, 2011
DOI: 10.1002/ejlt.201100213
1 Introduction
2,4-Dichlorophenoxyacetic acid (2,4-D) is a selective herbi-
cide of the phenoxyacetic acid group, with weak aromatic acid
properties [1]. Its herbicidal activity is mediated by an auxin
like capacity to alter normal protein synthesis and cell division
in plant meristems and leaves [2, 3]. Although it has recently
been suggested that its herbicidal activity may also be due to
an increase in the production of reactive oxygen species
(ROS) leading to the generation of oxidative stress in the
weed [4, 5]. Exposure of humans and animals occurs through
Additional corresponding author: Amel Nakbi, [email protected]
Correspondence: Professor Mohamed Hammami, Laboratory of
Biochemistry, Faculty of Medicine, UR03/ES-08, ’Human Nutrition and
Metabolic Disorders’, Monastir 5019, Tunisia
E-mail: [email protected]
Fax: þ216 73 460733
Abbreviations: CAT, catalase; DC, conjugated dienes; 2,4-D, 2,4-
dichlorophenoxiacetic acid; EVOO, extra virgin olive oil; GPx,
glutathione peroxidase; GR, glutathione reductase; LDL, low density
lipoprotein; MDA, malondialdehyde; OOHF, olive oil hydrophilic fraction;
OOLF, olive oil lipophilic fraction; ROS, reactive oxygen species; SFA,
saturated fatty acid; SOD, superoxide dismutase; UFA, unsaturated fatty
acid
Eur. J. Lipid Sci. Technol. 2012, 114, 469–478 469
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

contaminated air, drinking water, soil, and foodstuff or
during production of the herbicide. It has been reported that
2,4-D may cause a health-risk [1, 6]. Dose-dependent toxic
effects include damage to the eye, thyroid, kidney, adrenals,
and ovaries or testes. In addition, researchers have observed
neurotoxicity, reproductive toxicity and developmental
toxicity [7, 8]. The kidney is the critical target organ of
chronic 2,4-D toxicity [6, 9] and is well documented by a
number of studies in experimental animals. In fact, subacute
2,4-D administration induces dose-dependent histopatho-
logical degenerative effects in the rat kidney cortex [10].
Several lines of evidence indicate that oxidative stress and
ROS formed in the presence of 2,4-D could be responsible for
its toxic effects in many settings in vitro [11, 12] and in vivo
[13, 14]. Consequently, increased tissue oxidative stress can
lead to cell damage.
Most works have focused on the beneficial effect of plant
foods on oxidative stress. Constituents of some dietary
plant products, such as unsaturated fatty acids (UFAs)
and phenolics, endowed with antioxidant and anti-
inflammatory properties, which may play a protective role
in several human pathologies [15–17]. Most of the studies
comparing the effects of a MUFA-rich diet with PUFA-rich
diet on low density lipoprotein (LDL) oxidation parameters has
found a higher resistance of LDL particles to oxidation after the
consumption of MUFA-rich diet [15, 16]. Furthermore, in
vitro studies suggest that PUFAs are more pro-inflammatory
than MUFAs and saturated fatty acids (SFAs) [18].
Phenolics are also powerful scavengers of superoxide
anions and hydrogen peroxide and are capable of preventing
the generation of ROS by intact leukocytes as it has been
demonstrated by other authors [19, 20]. Numerous in vitro
studies have shown these phenolics to possess strong radical
scavenging activity at least equal in potency with other
important dietary antioxidants, such as ascorbic acid and
a-tocopherol [21, 22]. Phenolic compounds are also potent
in vitro inhibitors of LDL oxidation and are capable of break-
ing the peroxidative chain reaction [23, 24].
One from the Mediterranean diet, olive oil appears to be a
functional food with various components such as MUFAs that
may have health benefits and is also a good source of phyto-
chemicals, including polyphenolic compounds. Thus, it is likely
that extra virgin olive oil (EVOO) may be useful to alleviate or
protect against oxidant-induced various disease conditions,
including nephrotoxicity caused by 2,4-D. The present study
was established to examine the possible beneficial effect of
EVOO and its fractions on 2,4-D-induced renal functional
and structural abnormalities in experimental animals.
2 Materials and methods
2.1 Chemicals
2,4-D commercial formulation (Desormone Lourd) consist-
ing of 600 g/L 2,4-D ester butylglycol, register number
H.96064 was used in the experiments. 2-Thiobarbituric
acid (TBA) was obtained from Sigma Chemicals Co
(Taufkirchen, Germany). 1,1,3,3-Tetramethoxypropane
were purchased from Sigma Chemical Co. (St. Louis,
MO). Folin-Ciocalteu phenol reagent was purchased from
Fluka Biochemika (Buchs, Switzerland). All other chemicals
used were of analytical grade and were obtained from Sigma
Chemicals Co or Merck (Darmstadt, Germany).
2.2 Oil sample analysis
The EVOO used was harvested from the North of Tunisia.
The olive oil hydrophilic fraction (OOHF) was extracted
from EVOO by the method of Montedoro et al. [25]. The
olive oil lipophilic fraction (OOLF) was obtained from
EVOO as follows: the EVOO was homogenized for 1 min
with water (1:1, v/v), and the oil was separated by centrifu-
gation; this procedure was repeated six times. Then, the oil
fraction (OOLF) was filtered through a cellulose acetate
membrane [26, 27].
Extra virgin olive oil and its fractions analysis were per-
formed as reported in our previous paper [27]. In fact, the
determination of fatty acids was accomplished through the
quantification of their methyl esters (FAMEs) by gas chroma-
tography using a model 5890 series II instruments (Hewelett-
Packard Ca Palo Alto, California, USA) as described in
ref. 28. Carotenoids and chlorophylls (mg/kg oil) were deter-
mined spectrophotometrically according to Minguez-
Mosquera’s method [29]. Phenolic compounds were
extracted, estimated colorimetrically and expressed as
hydroxytyrosol equivalents as reported by Montedoro et al.
[25]. a-Tocopherol was evaluated according to Gimeno et al.
[30] on a Hewlett-Packard system (Waldbronn, Germany) as
described in ref. [31].
2.3 Animal treatment
This study was carried out in healthy, male adult Wistar rats,
weighing 200–230 g. The animals were housed under stand-
ard laboratory conditions of light (12-h light-dark periods),
temperature (22 � 38C), and relative humidity of 40%. The
animals were given standard diet (SICO, Sfax Tunisia) and
tap water ad libitum. According to our previous papers [26, 27],
the animals were randomly divided into eight experimental
groups: C (control), D (5 mg/kg b.w. of 2,4-D), D/EVOO
(5 mg/kg b.w. of 2,4-D þ 300 mL of EVOO), D/OOHF
(5 mg/kg b.w. of 2,4-D þ 1 mL of OOHF), D/OOLF
(5 mg/kg b.w. of 2,4-D þ 300 mL of OOLF), EVOO
(300 mL of EVOO), OOHF (1 mL of OOHF, the same
amount of phenols that we found in 300 mL of
oil ¼ 0.16 mg), and OOLF (300 mL of OOLF), each group
consisted of ten rats.
At the end of the experimental period of 4 wk, the animals
were sacrificed under diethyl ether anaesthesia; plasma
samples were collected and stored at �808C for analysis.
470 A. Nakbi et al. Eur. J. Lipid Sci. Technol. 2012, 114, 469–478
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

Kidney tissues were excised from sacrificed animals,
cleaned and individually weighted. Some samples were
minced and homogenized (10%, w/v) in an appropriate buf-
fer (pH 7.4) and centrifuged. The resulting supernatants
were collected and stored at �808C until used for enzyme
assays. The others were immediately fixed in Bouin liquid for
histological studies.
2.4 Kidney biomarkers analysis
The levels of urea, uric acid, creatinine, and albumin in
plasma were estimated spectrophotometrically using the
available diagnostic kits supplied by Randox Laboratories
(Ardmore, Northern Ireland, UK). The protein content
was determined according to Bradford [32].
2.5 Kidney lipoperoxidation
The level of lipid peroxidation products was measured as
thiobarbituric acid reactive metabolites (TBA-rm) according
to Yagi [33]. One hundred and twenty-five microliters of
plasma or supernatants were homogenized by sonication with
50 mL of TBS, 125 mL of TCA-BHT in order to precipitate
proteins and then centrifuged (1000g, 10 min, 48C). Two
hundred microliters of the supernatant were mixed with
40 mL of HCl (0.6 M) and 160 mL of TBA dissolved in
Tris, and the mixture was heated at 808C for 10 min. The
absorbance of the resultant supernatant was read at 530 nm.
The TBA-rm amount was calculated using a 156 mM/cm
extinction coefficient.
2.6 Kidney activities of antioxidant enzymes
Antioxidant enzyme activities were analyzed using a BioRad
UV–Visible spectrophotometer with a ‘‘kinetics’’ program
(BioRad, Mares la Coquette, France). The measurement
of superoxide dismutase (SOD), glutathione peroxidase
(GSH-Px), and glutathione reductase (GR) activities in
supernatants were performed by the commercially available
diagnostic kits supplied by Randox Laboratories. Catalase
(CAT) activity was measured at 258C according to Aebi’s
method [34] by measuring H2O2 concentration decrease at
240 nm.
2.7 Histopathological studies
For qualitative analysis of kidney histology, the tissue samples
were fixed for 48 h in Bouin liquid. The specimens were
washed and dehydrated through a graded series of ethanol
and were sequentially embedded in paraffin wax blocks.
Blocks were made and sectioned at 5 mm thickness using a
rotary microtome. Sections were rehydrated in distilled water
and stained with hematoxylin–eosin (H–E) then examined
under light microscopy. The images were obtained by a
digital camera system attached to the microscope. A mini-
mum of three fields for each kidney slide were examined and
scored semiquantitatively for severity of changes by a path-
ologist unaware of the type of treatment. The scoring was
done as none (�), mild (þ), moderate (þþ), and severe
(þþþ).
2.8 Statistical analysis
The data obtained were expressed as means (�SD) and
analyzed using the Statistical Package for the Social
Sciences (SPSS) program, release 11.0 for Windows
(SPSS, Chicago, IL, USA). Tukey’s test was used to deter-
mine any significant differences between analytical
parameters means of supplemented diet (one way analysis
of variance, ANOVA; 95% confidence interval). Student’s
t-test was used when comparison between two groups was
required. Statistical significance was considered at p<0.05.
3 Results
3.1 Analytical parameters of extra virgin olive oiland its fractions
Figure 1 shows that EVOO and OOLF presented approxi-
mately the same fatty acid composition with 17% of SFA,
65% of MUFA, and 15% of PUFA, whereas the standard diet
was more abundant in PUFA (55%) [26, 27]. The minor
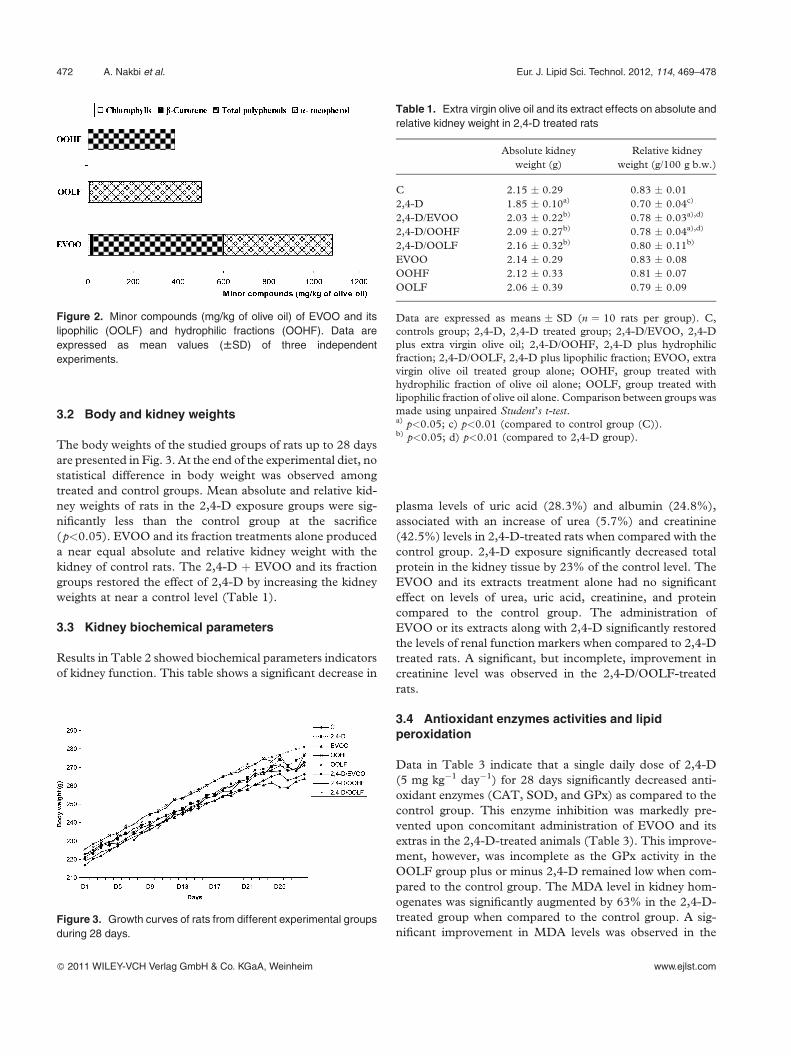
compounds of EVOO and its fractions are present in Fig. 2.
Data showed that EVOO and OOHF were rich in phenols
(580 and 384 mg/kg of olive oil, respectively) while the
OOLF was deprived from phenols but preserved the higher
content of a-tocopherol (490 mg/kg of olive oil) and other
minor compounds as pigments (Fig. 2).
Figure 1. Fatty acids composition (%) of extra virgin olive oil
(EVOO), lipophilic fractions (OOLF), and standard diet fed to rats.
Data are expressed as mean values (WSD) of three independent
experiments. SFA, saturated fatty acids; MUFA; PUFA. Values fol-
lowed by same letters are not significantly different (Tukey’s test,
p<0.05).
Eur. J. Lipid Sci. Technol. 2012, 114, 469–478 Olive oil protects against oxidative renal dysfunction 471
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com
3.2 Body and kidney weights
The body weights of the studied groups of rats up to 28 days
are presented in Fig. 3. At the end of the experimental diet, no
statistical difference in body weight was observed among
treated and control groups. Mean absolute and relative kid-
ney weights of rats in the 2,4-D exposure groups were sig-
nificantly less than the control group at the sacrifice
(p<0.05). EVOO and its fraction treatments alone produced
a near equal absolute and relative kidney weight with the
kidney of control rats. The 2,4-D þ EVOO and its fraction
groups restored the effect of 2,4-D by increasing the kidney
weights at near a control level (Table 1).
3.3 Kidney biochemical parameters
Results in Table 2 showed biochemical parameters indicators
of kidney function. This table shows a significant decrease in
plasma levels of uric acid (28.3%) and albumin (24.8%),
associated with an increase of urea (5.7%) and creatinine
(42.5%) levels in 2,4-D-treated rats when compared with the
control group. 2,4-D exposure significantly decreased total
protein in the kidney tissue by 23% of the control level. The
EVOO and its extracts treatment alone had no significant
effect on levels of urea, uric acid, creatinine, and protein
compared to the control group. The administration of
EVOO or its extracts along with 2,4-D significantly restored
the levels of renal function markers when compared to 2,4-D
treated rats. A significant, but incomplete, improvement in
creatinine level was observed in the 2,4-D/OOLF-treated
rats.
3.4 Antioxidant enzymes activities and lipidperoxidation
Data in Table 3 indicate that a single daily dose of 2,4-D
(5 mg kg�1 day�1) for 28 days significantly decreased anti-
oxidant enzymes (CAT, SOD, and GPx) as compared to the
control group. This enzyme inhibition was markedly pre-
vented upon concomitant administration of EVOO and its
extras in the 2,4-D-treated animals (Table 3). This improve-
ment, however, was incomplete as the GPx activity in the
OOLF group plus or minus 2,4-D remained low when com-
pared to the control group. The MDA level in kidney hom-
ogenates was significantly augmented by 63% in the 2,4-D-
treated group when compared to the control group. A sig-
nificant improvement in MDA levels was observed in the
Figure 2. Minor compounds (mg/kg of olive oil) of EVOO and its
lipophilic (OOLF) and hydrophilic fractions (OOHF). Data are
expressed as mean values (WSD) of three independent
experiments.
Figure 3. Growth curves of rats from different experimental groups
during 28 days.
Table 1. Extra virgin olive oil and its extract effects on absolute and
relative kidney weight in 2,4-D treated rats
Absolute kidney
weight (g)
Relative kidney
weight (g/100 g b.w.)
C 2.15 � 0.29 0.83 � 0.01
2,4-D 1.85 � 0.10a) 0.70 � 0.04c)
2,4-D/EVOO 2.03 � 0.22b) 0.78 � 0.03a),d)
2,4-D/OOHF 2.09 � 0.27b) 0.78 � 0.04a),d)
2,4-D/OOLF 2.16 � 0.32b) 0.80 � 0.11b)
EVOO 2.14 � 0.29 0.83 � 0.08
OOHF 2.12 � 0.33 0.81 � 0.07
OOLF 2.06 � 0.39 0.79 � 0.09
Data are expressed as means � SD (n ¼ 10 rats per group). C,
controls group; 2,4-D, 2,4-D treated group; 2,4-D/EVOO, 2,4-D
plus extra virgin olive oil; 2,4-D/OOHF, 2,4-D plus hydrophilic
fraction; 2,4-D/OOLF, 2,4-D plus lipophilic fraction; EVOO, extra
virgin olive oil treated group alone; OOHF, group treated with
hydrophilic fraction of olive oil alone; OOLF, group treated with
lipophilic fraction of olive oil alone. Comparison between groups was
made using unpaired Student’s t-test.a) p<0.05; c) p<0.01 (compared to control group (C)).b) p<0.05; d) p<0.01 (compared to 2,4-D group).
472 A. Nakbi et al. Eur. J. Lipid Sci. Technol. 2012, 114, 469–478
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

EVOO or its extracts administered along with 2,4-D-treated
rats (Table 3).
3.5 Kidney histological studies
Histopathological studies showed that control rats pre-
sented normal rat kidney with normal tubular brush-bor-
ders and intact glomeruli (Fig. 4A). Contrarily, histology of
the 2,4-D treated groups revealed renal corpuscular
degeneration, which was marked by glomeruli atrophy,
epithelial cell degeneration of distal convoluted tubules
(DCTs), and dilatation with dislocation of brush-borders
of proximal convoluted tubules. A congestion, necrosis,
and vascular/tissular inflammation were observed in this
group (Fig. 4B and Table 4). However, the pathological
lesions induced by 2,4-D were remarkably reduced by the
administration of EVOO and its extracts (Fig. 4C–E).
A higher improvement was observed in the group supple-
mented with EVOO along with 2,4-D with a moderate
amelioration for the 2,4-D/OOLF and 2,4-D/OOHF
groups.
The histopathological changes were in agreement with
biochemical parameters. There were no histological altera-
tions in the kidney of EVOO, OOHF, and OOLF groups
showing the same histological structure of kidney (Fig. 4A
and Table 4) when compared to control.
Table 2. The effect of extra virgin olive oil and its fractions on 2,4-D induced changes in renal functional markers
Urea
(mmol/L)
Uric acid
(mmol/L)
Creatinine
(mmol/L)
Albumin
(g/dL)
Total protein
(g/dL)
C 6.97 � 0.62 202.22 � 49.06 105.85 � 16.26 4.44 � 0.44 1.56 � 0.11
2,4-D 7.37 � 0.69 145.00 � 29.89b) 150.87 � 5.51c) 3.34 � 0.36c) 1.19 � 0.13c)
2,4-D/EVOO 6.95 � 0.67 193.00 � 47.08a) 117.33 � 24.54d) 4.32 � 0.48d) 1.71 � 0.19d)
2,4-D/OOHF 6.71 � 1.07 199.66 � 48.90a) 116.75 � 18.39d) 4.20 � 0.47d) 1.59 � 0.29a)
2,4-D/OOLF 6.45 � 0.83a) 204.66 � 43.69a) 136.00 � 3.67c),d) 4.67 � 0.42d) 1.57 � 0.24a)
EVOO 6.43 � 0.83 210.85 � 53.61 106.00 � 19.07 4.11 � 0.42 1.48 � 0.10
OOHF 6.58 � 1.15 184.33 � 53.81 103.00 � 17.01 4.03 � 0.61 1.48 � 0.17
OOLF 6.18 � 0.76 205.20 � 55.71 109.28 � 16.69 4.22 � 1.21 1.73 � 0.20
Data are expressed as means � SD (n ¼ 10 rats per group). C, controls group; 2,4-D, 2,4-D treated group; 2,4-D/EVOO, 2,4-D plus extra
virgin olive oil; 2,4-D/OOHF, 2,4-D plus hydrophilic fraction; 2,4-D/OOLF, 2,4-D plus lipophilic fraction; EVOO, extra virgin olive oil
treated group alone; OOHF, group treated with hydrophilic fraction of olive oil alone; OOLF, group treated with lipophilic fraction of olive oil
alone. Comparison between groups was made using unpaired Student’s t-test.a) p<0.05; c) p<0.01 (compared to control group (C)).b) p<0.05; d) p<0.01 (compared to 2,4-D group).
Table 3. The effect of extra virgin olive oil and its fractions on kidney antioxidant enzyme activities and MDA levels of 2,4-D treated rats
SOD
(U/mg protein)
GPx
(U/mg protein)
CAT
(mmol/min mg protein)
MDA
(mM/g protein)
C 5.28 � 0.79 0.86 � 0.18 19.92 � 1.82 1.00 � 0.14
2,4-D 3.80 � 0.53a) 0.69 � 0.17 12.56 � 0.91c) 1.63 � 0.11c)
2,4-D/EVOO 6.50 � 2.42b) 0.89 � 0.20 17.00 � 2.54b) 0.99 � 0.10d)
2,4-D/OOHF 6.49 � 2.21b) 0.75 � 0.17 17.84 � 0.35b) 1.02 � 0.19d)
2,4-D/OOLF 4.29 � 1.93 0.51 � 0.14 17.39 � 1.42 0.93 � 0.27d)
EVOO 5.33 � 1.19 0.87 � 0.18 20.66 � 2.54 0.93 � 0.27
OOHF 5.41 � 1.88 0.72 � 0.22 18.22 � 3.78 1.07 � 0.15
OOLF 6.11 � 2.26 0.52 � 0.07 18.63 � 2.01 0.95 � 0.15
Data are expressed as means � SD (n ¼ 10 rats per group). C, controls group; 2,4-D, 2,4-D treated group; 2,4-D/EVOO, 2,4-D plus extra
virgin olive oil; 2,4-D/OOHF, 2,4-D plus hydrophilic fraction; 2,4-D/OOLF, 2,4-D plus lipophilic fraction; EVOO, extra virgin olive oil
treated group alone; OOHF, group treated with hydrophilic fraction of olive oil alone; OOLF, group treated with lipophilic fraction of olive oil
alone. Comparison between groups was made using unpaired Student’s t-test.a) p<0.05; c) p<0.01 (compared to control group (C)).b) p<0.05; d) p<0.01 (compared to 2,4-D group).
Eur. J. Lipid Sci. Technol. 2012, 114, 469–478 Olive oil protects against oxidative renal dysfunction 473
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

Figure 4. Histopathology of rat kidney cortex
from control and experimental groups. Kidney
sections were stained using the hematoxylin–
eosin method (H&E 32T). (A) Kidney section,
from control, EVOO, OOHF, and OOLF groups,
showing a normal appearance of glomeruli (G)
and an intact Bowman’s capsule (BC) with adja-
cent proximal (PCT) and distal convoluted
tubules (DCT). (B) 2,4-D (5 mg/kg) treated rat
revealed renal corpuscular degeneration show-
ing a glomeruli atrophy (G At), tubular dilatation
(TD) with an expansion (exp) of the space
between glomeruli and the Bowman’s capsule.
(C) 2,4-D/EVOO group showing a higher
improvement of the histological alteration of
the kidneys induced by 2,4-D. (D and E) 2,4-
D/OOHF and 2,4-D/OOLF groups showing a
moderate improvement with manifestation of
the congestion (Cg).
Table 4. Histopathological findings in the kidney of the control group and experimental groups
Glomerular
atrophy
Tubular
dilatation
Tissular and
vascular congestion
Tubular
degeneration Necrosis
C S S S S S2,4-D þþþþ þþþþ þþþþ þþþþ þþ2,4-D/EVOO ### ### ## ### ###2,4-D/OOLF # ## # ## #2,4-D/OOHF # ## # ## #EVOO S S S S SOOHF � � � � �OOLF � � � � �
�, none; þþ, moderate; þþþ, severe; #, mild improvement; ##, moderate improvement; ###, severe improvement.
Data were carried from six rats per group.
474 A. Nakbi et al. Eur. J. Lipid Sci. Technol. 2012, 114, 469–478
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

4 Discussion
There are various chemical products and drugs, which cause
damage to renal tissues by ROS production [35]. The phe-
noxyacetic acid herbicides have previously been shown to
accumulate at high levels in liver and kidney via the organic
acid transport system, and this accumulation decreases the
oxygen uptake by the renal cortical slices and results in
uncoupling of renal mitochondria raising the possibility of
organ-specific toxicity secondary to transport [10]. Seeing the
deleterious effects of chlorophenoxy compounds on kidney,
the current study was investigated on the reno-protective
effect of EVOO and its fractions against 2,4-D-induced oxi-
dative stress in rats.
Among the chlorophenoxy herbicides used for pest con-
trol with their higher pesticidals activity and considerably
lower mammalian toxicity compared to other pesticides
[10], 2,4-D is the most used in Tunisia. The precise mech-
anism of its acute toxicity is not elucidated but can comprise
the disruption of plasma and intracellular membranes or
uncoupling of oxidative phosphorylation [36]. This last
mechanism may be involved in the generation of oxidative
stress and subsequent induction of cytotoxic and genotoxic
effects, which may lead, among others to reproductive and
developmental toxicity [5]. In the current study, 2,4-D
induced a kidney dysfunction which was demonstrated by
a decrease of kidney weight, alterations of biochemical
parameters, and histological damages. This kidney injury
was recovered by treatment of EVOO and its fractions.
The presence of high levels of plasma urea and creatinine
and lower level of uric acid and albumin are possible
indicators of kidney injuries induced through 2,4-D treat-
ment. Moreover, 2,4-D exposure caused a significant
reduction in kidney total protein level, which indicated the
disorders in protein synthesis and metabolism [3].
Furthermore, the decreases in the plasma total protein and
albumin values may be due to hepatic injury, including
degenerative and necrotic changes.
The results of the present study have demonstrated that sub-
acute treatment with 2,4-D could affect the antioxidant defense
systems of animals. In fact, it has been observed that 2,4-D
induced a decrease in SOD, CAT, and GPx activities and
enhanced MDA level in kidney. These data confirmed the
findings of Celik et al. [14] indicating that 2,4-D at 50 and
100 ppm produced substantial systemic organ toxicity the eryth-
rocyte, liver, brain, heart, and kidney, during the period of a
25-day subacute exposure. In fact, the results indicated that
2,4-D might affect antioxidant potential enzymes and lipid per-
oxidation in the kidney. Moreover, it has been suggested that a
decrease in the activities of primary antioxidants, CAT and SOD,
may be due to accumulation of ROS induced by 2,4-D. An
observation that strengthens this hypothesis is that SOD activity
can be inhibited by hydrogen peroxide treatment [35, 37].
The current study revealed that supplementation of
EVOO and its fractions significantly restored the level of
biochemical markers of kidney function as well as the renal
antioxidant defence system. In fact, the biochemical
parameters such as total protein, albumin, and uric acid were
increased after administration of EVOO and its fractions,
where the better improvement was observed in 2,4-D/
EVOO group for total protein. However, urea and creatinine
levels in plasma were decreased in 2,4-D treated rats supple-
mented with EVOO or its fractions compared to 2,4-D
group. These data were in agreement with previous reports,
which proved that the beneficial effects of oils were evidenced
by reduced plasma urea and creatinine concentrations in the
group receiving oils compared to the non-oil treatment
animals receiving gentamicin only [38]. While the present
results showed that the creatinine level remained significantly
higher in the 2,4-D/OOLF compared to controls which can
be due to the absence of phenolic compounds in the LF
fraction.
Supplementation of EVOO, in the present study, signifi-
cantly improved the tissue level of enzymatic antioxidants
compared to 2,4-D-treated rats. These effects have been
attributed to its high MUFA content, mainly the oleic acid.
Due to its high MUFA content, olive oil seems to have
protective properties with regard to LDL oxidation.
Indeed, several investigators have compared the influence
of dietary MUFA and PUFA on LDL oxidation. It could
first be shown in rabbits that oleate-rich LDL particles are
remarkably resistant to oxidative modification [39, 40].
Moreover, dietary fatty acids can influence the susceptibility
of cells to oxidative stress, probably by changing cell mem-
brane fatty acid composition. In fact, cells enriched with
MUFA have been shown to be less susceptible to oxidative
damage, whereas n-6 PUFA increased the susceptibility to
oxidative damage. Additionally, dietary MUFA from EVOO
has been shown to protect mitochondrial membranes from
rat heart cells against ageing-related per oxidative damage
[41]. In addition, in rats, low levels of 5% lipid-enriched diet
(canola oil, fish oil, and butter) in a chronic nutritional
regimen can modify the age-related changes in renal function
and that the impact of different types of lipid-supplemented
diets on renal function depends on the kind of lipid present in
the diet [42, 43]. However, OOLF did not show any improve-
ment in the SOD and GPx antioxidant enzyme activities.
This might indicate the usefulness of EVOO, as an excellent
source of antioxidants, in modulating 2,4-D-induced neph-
rotoxicity since EVOO has other components with antioxi-
dant properties, mainly polyphenol [44]. The antioxidant
efficiency of olive phenols has been assessed in various tests
such as the inhibition of LDL oxidation [45]. Phenolic com-
pounds have been shown to possess free radical-scavenging
and metal-chelating activities in addition to the reported
anticarcinogenic properties [46, 47]. As a very rich source
of polyphenol compounds, the strong antioxidant and oxygen
radicals scavenging effects of tea have been documented
[48–50]. Hydroxytyrosol, the main phenolic compound in
olives and their by-products, is a phenolic compound which
Eur. J. Lipid Sci. Technol. 2012, 114, 469–478 Olive oil protects against oxidative renal dysfunction 475
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

has been shown to possess diverse healing properties for its
antioxidant and anti-inflammatory activities [51, 52].
Moreover, low rate in free radicals and important activity
in antioxidant enzymes such as SOD, CAT, and GPX in liver
and kidney tissues were observed in diabetic rats [53].
Additionally, the hypoglycemic and antioxidant activities of
hydroxytyrosol prevent oxidative stress; in fact administration
of hydroxytyrosol to diabetic rats prevented kidney toxicity
observed by low rates in creatinine and urea in plasma, which
increased in diabetic rats [53].
2,4-D administration mediated lipid peroxidation of lipid
structures of renal tissues, resulting in subcellular damages as
observed in the histopathological examination. In this study,
The optic microscopic findings have shown characteristic
morphological alterations such as glomeruli atrophy, epithelial
cell degeneration of DCTs, and dilatation with dislocation of
brush-borders of proximal convoluted tubules in kidneys of
2,4-D treated rats. A congestion, necrosis, and vascular/tissu-
lar inflammation were also observed. Similar histopathological
changes were observed by Uyanikgil et al. [10] in renal of rats
treated with 2,4-D. The severe changes were not observed in
the groups treated with EVOO and its fractions suggesting the
protective effects of EVOO in attenuating 2,4-D-induced
morphological changes. In fact, previous researches have
shown that usual glomerulosclerosis, glomerular enlargement,
and glomeruli loss in spontaneously hypertensive rats have
been prevented (fish, canola, and palm oils) or attenuated
(olive and soybean oils) by this oil long-term administration
[43, 54]. However, cyclosporine-dependent changes in renal
histology were only partially reversed by the administration of
the hydroxytyrosol in rats [55]. In the current study, a higher
improvement was observed in the 2,4-D/EVOO group with a
moderate amelioration for the 2,4-D/OOLF and 2,4-D/
OOHF groups. The histopathological changes were in agree-
ment with biochemical parameters. There were no histological
alterations in the kidney of EVOO, OOHF, and OOLF groups
showing the same histological structure of kidney when
compared to control.
5 Conclusions
This study substantiated the scientific evidence in favor of the
pharmacological use of olive oil in renal disorders in folk
medicine. According to the data obtained in the present
study, the administration of 2,4-D to rats at a dose of
5 mg/kg b.w./day for a period of 28 days caused significant
changes in oxidative stress markers and lipid peroxidation in
kidney rats. These changes were observed in the form of the
improvement of values in the groups, which were adminis-
tered EVOO and its fractions. However, the LF and HF
fractions supplemented separately showed a moderate
improvement in the histology kidney toxicity induced by
2,4-D. While EVOO supplemented to rats showed better
improvement. These data should be explained in terms of
cooperation between the two fractions.
This research was supported by a grant from the ‘‘Ministere de
l’Enseignement Superieur et de la Recherche Scientifique’’ Tunisia
(UR03/ES-08 ‘‘Nutrition Humaine et Desordres Metaboliques’’
et USCR-Spectrometrie de Masse, University of Monastir).
The authors have declared no conflict of interest.
References
[1] Aydin, H., Ozdemir, N., Uzunoren, N., Investigation of theaccumulation of 2,4-dichlorophenoxyacetic acid (2,4-D) inrat kidneys. Foren. Sci. Int. 2005, 153, 53–56.
[2] Stevens, J. T., Breckenridge, C. B., in: Hayes, W. A. (Ed.),Principles and Methods of Toxicology, Taylor & Francis,Philadelphia 2001, pp. 565–648.
[3] Tayeb, W., Nakbi, A., Trabelsi, M., Attia, N. et al.,Hepatotoxicity induced by sub-acute exposure of rats to2,4-dichlorophenoxyacetic acid based herbicide‘‘Desormone lourd’’. J. Hazard. Mater. 2010, 180, 225–233.
[4] Romero-Puertas, M. C., Carthy, M. C., Gomez, M.,Sandalio, L. M. et al., Reactive oxygen species-mediatedenzymatic systems involved in the oxidative action of 2,4-dichlorophenoxyacetic acid. Plant Cell Environ. 2004, 27,1135–1148.
[5] Dinamarca, V. M., Hidalgo, M. E., Cavieres, M. F., Lack ofeffects of 2,4-dichlorophenoxyacetic acid administration onmarkers of oxidative stress during early pregnancy in mice.Toxicology 2007, 237, 104–110.
[6] Balague, C. E., Ruız, C. S., Rey, R., De Duffard, A. M. E.,Nader-Macıas, M. E., Effect of herbicide 2,4-dichlorophe-noxyacetic acid on uropathogenic Escherichia coli virulencefactors. Toxicology 2002, 177, 143–155.
[7] Charles, J. M., Bond, D. M., Jeffries, T. K., Yano, B. L. et al.,Chronic dietary toxicity/oncogenicity studies on 2,4-dichlor-phenoxyacetic acid in rodents. Fund. Appl. Toxicol. 1996, 33,166–172.
[8] Registration Eligibility Decision (RED) 2,4-D; EPA 738-R-05-002; U.S, Environmental Protection Agency, Office ofPrevention, Pesticides and Toxic Substances, Office ofPesticide Programs, U.S. Government Printing Office:Washington, DC. 2005.
[9] Amer, S. M., Aly, F. A. E., Genotoxic effect of 2,4-dichlor-ophenoxyacetic acid and its metabolite 2,4-Dichlorophenolin mouse. Mutat. Res. 2001, 494, 1–12.
[10] Uyanikgil, Y., Ates, U., Baka, M., Bicer, S. et al.,Immunohistochemical and histopathological evaluation of2,4-dichlorophenoxyacetic acid-induced changes in ratkidney cortex. Bull Environ. Contam. Toxicol. 2009, 82,749–755.
[11] Bukowska, B., Chajdys, A., Duda, W., Duchnowic, P., Effectof phenoxyherbicides and their metabolites on the activity ofcatalase in the human erythrocyte. Cell Biol. Int. 2000, 24,705–711.
[12] Duchnowicz, P., Koter, M., Damage to the erythrocytemembrane caused by chlorophenoxyacetic herbicides. Cell.Mol. Biol. Lett. 2003, 8, 25–30.
[13] Ozcan, E., Sevgiler, Y., Uner, N., Tissue-specific oxidativestress responses in fish exposed to 2,4-D and azinphosmethyl.Comp. Biochem. Physiol., Part C 2004, 137, 43–51.
476 A. Nakbi et al. Eur. J. Lipid Sci. Technol. 2012, 114, 469–478
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

[14] Celik, I., Tuluce, Y., Isik, I., Influence of subacute treatmentof some plant growth regulators on serum marker enzymesand erythrocyte and tissue antioxidant defense and lipidperoxidation in rats. J. Biochem. Mol. Toxicol. 2006, 20,174–182.
[15] Aguilera, C. M., Mesa, M. D., Ramirez-Tortosa, M. C.,Nestares, M. T. et al., Sunflower oil does not protect againstLDL oxidation as virgin olive oil does in patients withperipheral vascular disease. Clin. Nutr. 2004, 23, 673–681.
[16] Kratz, M., Cullen, P., Kannenberg, F., Kassner, A. et al.,Effects of dietary fatty acids on the composition and oxidiz-ability of low-density lipoprotein. Eur. J. Clin. Nutr. 2002, 56,72–81.
[17] Visioli, F., Galli, C., Plasmati, E., Viappiani, S. et al., Olivephenol hydroxytyrosol prevents passive smoking-inducedoxidative stress. Circulation 2000, 102, 2169–2171.
[18] Williams, C. M., Maitin, V., Jackson, K. G., Triacylglycerol-rich lipoprotein–gene interactions in endothelial cells.Biochem. Soc. Trans. 2004, 32, 994–998.
[19] Visioli, F., Bellomo, G., Galli, C., Free radical-scavengingproperties of olive oil polyphenols. Biochem. Biophys. Res.Commun. 1998, 247, 60–64.
[20] De la Puerta, R., Ruiz-gutierrez, V., Hoult, J. R. S.,Inhibition of leukocyte 5-lipoxygenase by phenolicsfron virgin olive oil. Biochem. Pharmacol. 1999, 57, 445–449.
[21] Manna, C., Galletti, P., Cucciolla, V., Moltedo, O. et al.,The protective effect of the olive oil polyphenol (3,4-dihy-droxyphenyl)-ethanol counteracts reactive oxygen metab-olite-induced cytotoxicity in Caco-2 cells. J. Nutr. 1997,127, 286–292.
[22] Vissers, M. N., Zock, P. L., Katan, M. B., Bioavailability andantioxidant effects of olive oil phenols in humans. Eur. J.Clin. Nutr. 2004, 58, 955–965.
[23] Fito, M., Covas, M. I., Lamuela-Raventos, R. M., Vila, J.et al., Protective effect of olive oil and its phenolic com-pounds against low density lipoprotein oxidation. Lipids2000, 35, 633–638.
[24] Visioli, F., Bellomo, G., Montedoro, G., Galli, C., Lowdensity lipoprotein oxidation is inhibited in vitro by oliveoil constituents. Atherosclerosis 1995, 117, 25–32.
[25] Montedoro, G. F., Servili, M., Baldioli, M., Miniati, E.,Simple and hydrolyzable phenolic compounds in virgin oliveoil. Their extraction, separation and quantitative and semi-quantitative evaluation by HPLC. J. Agric. Food Chem. 1992,40, 1571–1576.
[26] Nakbi, A., Tayeb, W., Dabbou, S., Issaoui, M. et al., Dietaryolive oil effect on antioxidant status and fatty acid profile inthe erythrocyte of 2,4-D-exposed rats. Lipid Heal. Dis. 2010,9, 89–99.
[27] Nakbi, A., Tayeb, W., Kasdallah-Grissa, A., Issaoui, M.et al., Effects of olive oil and its fractions on oxidative stressand the liver’s fatty acid composition in 2,4-dichlorophe-noxyacetic acid-treated rats. Nutr. Metab. 2010, 7, 80–91.
[28] Issaoui, M., Flamini, G., Ben Hassine, K., Chehab, H., etal.Improvement of Chemlali olive oil oxidative stability byblending with Chetoui and Rekhami cultivars. Int. J. FoodSci. Technol. 2009, 44, 1323–1332.
[29] Minguez-Mosquera, M. I., Rejano, L., Gandul, B., Sanchez,A. H., Garrido, J., Color-pigment correlation in virgin oliveoil. J. Am. Oil Chem. Soc. 1991, 68, 332–336.
[30] Gimeno, E., Castellote, A. I., Lamuela-Raventos, R. M., DeLa Torre, M. C., Lopez-Sabater, M. C., Rapid determi-nation of vitamin E in vegetable oils by reversed-phasehigh-performance liquid chromatography. J. Chromatogr. A2000, 881, 251–254.
[31] Nakbi, A., Issaoui, M., Dabbou, S., Koubaa, N. et al.,Evaluation of antioxidant activities of phenolic compoundsfrom two extra virgin olive oils. J. Food Compos. Anal. 2010,23, 711–715.
[32] Bradford, M. M., A rapid and sensitive method for thequantitation of microgram quantities of protein utilizingthe principle of protein-dye binding. Anal. Biochem. 1976,72, 248–254.
[33] Yagi, K., A simple fluorometric assay for lipoperoxide inblood plasma. Biochem. Med. 1976, 15, 212–216.
[34] Aebi, H., Catalase in vitro. Methods Enzymol. 1984, 105,121–126.
[35] Khan, R. A., Khan, M. R., Sahreen, S., Bokhari, J.,Prevention of CCl-4-induced nephrotoxicity with Sonchusasper in rat. Food Chem. Toxicol. 2010, 48, 2469–2476.
[36] Bradberry, S. M., Watt, B. E., Proudfoot, A. T., Vale, J. A.,Mechanisms of toxicity, clinical features, and management ofacute chlorophenoxy herbicide poisoning. J. Toxicol. Clin.Toxicol. 2000, 38, 111–122.
[37] Miguel, F., Augusto, A. C., Gurgueira, S. A., Effect of acutevs. chronic H2O2-induced oxidative stress on antioxidantenzyme activities. Free Radical Res. 2009, 43, 340–347.
[38] Rashid, F., Sheema, K. M., Bano, B., Comparative effect ofolive oil and fish oil supplementation in combating gentami-cin induced nephrotoxicity in rats. Ind. J. Clin. Biochem.2005, (20/1), 109–114.
[39] Aviram, M., Eias, K., Dietary olive oil reduces low-densitylipoprotein uptake by macrophages and decreases thesusceptibility of the lipoprotein to undergo lipid peroxi-dation. Ann. Nutr. Metab. 1993, 37, 75–84.
[40] Berry, E. M., Eisenberg, S., Friedlander, Y., Harats, D. et al.,Effects of diets rich in monounsaturated fatty acids on plasmalipoproteins—the Jerusalem Nutrition Study. II.Monounsaturated fatty acids vs. carbohydrates. Am. J.Clin. Nutr. 1992, 56, 394–403.
[41] Huertas, J. R., Martinez, V. E., Ibanez, S., Lopez, F. M.et al., Virgin olive oil and coenzyme Q10 protect heart mito-chondria from peroxidative damage during aging. Biofactors1999, 9, 337–343.
[42] Valente Gamba, C., Zeraib Caraviello, A., Matsushita, A.,Alves, G. M., et al., Effects of dietary lipids on renal functionof aged rats. Braz. J. Med. Biol. Res. 2001, 34, 265–269.
[43] Medeiros, F. J., Aguila, M. B., Mandarim-de-Lacerda, C. A.,Renal cortex remodeling in streptozotocin-induced diabeticspontaneously hypertensive rats treated with olive oil, palmoil and fish oil from Menhaden. Prostaglandins, LeukotrienesEssent. Fatty Acids 2006, 75, 357–365.
[44] Perona, J. S., Cabello-Moruno, R., Ruız-Gutierrez, V., Therole of virgin olive oil components in the modulation ofendothelial function. J. Nutr. Biochem. 2006, 17, 429–445.
[45] Leenen, R., Roodenburg, A. J. C. R., Vissers, M. N.,Schurbiers, J. A. E. et al., Supplementation of plasma witholive oil phenols and extracts: Influence on LDL oxidation.J. Agric. Food Chem. 2002, 50, 1290–1297.
[46] Middleton, E. Jr., Effect of plant flavonoids on immune andinflammatory cell function. Adv. Exp. Med. Biol. 1998, 439,175–182.
Eur. J. Lipid Sci. Technol. 2012, 114, 469–478 Olive oil protects against oxidative renal dysfunction 477
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com

[47] Nakbi, A., Dabbou, S., Champion, S., Fouchier, F. et al.,Modulation of the superoxide anion production and MMP-9expression in PMA stimulated THP-1 cells by olive oil minorcomponents: Tyrosol and hydroxytyrosol. Food Res. Int.2011, 44, 575–581.
[48] Yang, C. S., Maliakal, P., Meng, X., Inhibition of carcinogen-esis by tea. Annu. Rev. Pharmacol. Toxicol. 2002, 42, 25–54.
[49] Arts, M. J., Haenen, G. R., Wilms, L. C., Beetstra, S. A.et al., Interactions between flavonoids and proteins: Effect onthe total antioxidant capacity. J. Agric. Food Chem. 2002, 50,1184–1187.
[50] Mohamadin, A. M., El-Beshbishya, H. A., El-Mahdy, M. A.,Green tea extract attenuates cyclosporine A-induced oxi-dative stress in rats. Pharmacol. Res. 2005, 51, 51–57.
[51] Allouche, N., Fki, I., Sayadi, S., The use of olive mill waste-waters as a cheap source of natural antioxidants: Biologicalactivities of a continuous solvent extract and purified hydrox-ytyrosol. Polyphenols Actual. 2003, 23, 16–19.
[52] Zhang, X., Cao, J., Zhong, L., Hydroxytyrosol inhibits pro-inflammatory cytokines, iNOS, and COX-2 expression inhuman monocytic cells. Naunyn Schmiedebergs Arch.Pharmacol. 2009, 379, 581–586.
[53] Hamden, K., Allouche, N., Damak, M., Elfeki, A.,Hypoglycemic and antioxidant effects of phenolic extractsand purified hydroxytyrosol from olive mill waste in vitro andin rats. Chem. Biol. Int. 2009, 180, 421–432.
[54] Aguila, M. B., Pinheiro, A. R., Aquino, J. C., Gomes, A. P.,Mandarim-de-Lacerda, C. A., Different edible oil beneficialeffects (canola oil, fish oil, palm oil, olive oil, and soybean oil)on spontaneously hypertensive rat glomerular enlargementand glomeruli number. Prostaglandins Other Lipid Mediators2005, 76, 74–85.
[55] Capasso, G., Di Gennaro, C. I., Ragione, F. D., Manna, C.et al., In vivo effect of the natural antioxidant hydroxytyrosolon cyclosporine nephrotoxicity in rats. Nephrol. Dial. Transpl.2008, 23, 1186–1195.
478 A. Nakbi et al. Eur. J. Lipid Sci. Technol. 2012, 114, 469–478
� 2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.ejlst.com











![3-[( E )-(2,4-Dichloropbenzylidene)amino]benzoic acid](https://img.pdfslide.net/doc/110x75/6323a62a48d448ffa006c621/3-e-24-dichloropbenzylideneaminobenzoic-acid.jpg)

















