Embed Size (px)
Citation preview

�-OPIOID RECEPTORS ARE LOCATED POSTSYNAPTICALLY ANDENDOMORPHIN-1 INHIBITS VOLTAGE-GATED CALCIUM CURRENTS INPREMOTOR CARDIAC PARASYMPATHETIC NEURONS IN THE RATNUCLEUS AMBIGUUS
M. IRNATEN,a S. A. AICHER,b J. WANG,a
P. VENKATESAN,a C. EVANS,a S. BAXIa ANDD. MENDELOWITZa*aDepartment of Pharmacology, George Washington University, 2300Eye Street NW, Washington, DC 20037, USAbNeurological Sciences Institute, Oregon Health & Science University,505 NW 185th Avenue, Beaverton, OR 97006, USA
Abstract—Activation of opioid receptors in the CNS evokes adramatic decrease in heart rate which is mediated by in-creases in inhibitory parasympathetic activity to the heart.Injection of opiates into the nucleus ambiguus, where premo-tor cardiac parasympathetic nucleus ambiguus neurons arelocated elicits an increase in parasympathetic cardiac activityand bradycardia. However, the mechanisms responsible foraltering the activity of premotor cardiac parasympathetic nu-cleus ambiguus neurons is unknown. This study examined atthe electron microscopic level whether premotor cardiacparasympathetic nucleus ambiguus neurons possesspostsynaptic opioid receptors and whether �-opioid receptoragonists alter voltage-gated calcium currents in these neu-rons.
Premotor cardiac parasympathetic nucleus ambiguusneurons were identified in the rat using retrograde fluores-cent tracers. One series of experiments utilized dual-labelingimmunocytochemical methods combined with electron mi-croscopic analysis to determine if premotor cardiac parasym-pathetic nucleus ambiguus neurons contain �-opioid recep-tors. In a second series of experiments whole cell patchclamp methodologies were used to determine whether acti-vation of postsynaptic opioid receptors altered voltage-gatedcalcium currents in premotor cardiac parasympathetic nu-cleus ambiguus neurons in brainstem slices.
The perikarya and 78% of the dendrites of premotor car-diac parasympathetic nucleus ambiguus neurons contain�-opioid receptors. Voltage-gated calcium currents in premo-tor cardiac parasympathetic nucleus ambiguus neurons werecomprised nearly entirely of �-agatoxin-sensitive P/Q-typevoltage-gated calcium currents. Activation of �-opioid recep-tors inhibited these voltage-gated calcium currents and thisinhibition was blocked by pretreatment with pertusis toxin.The �-opioid receptor agonist endomorphin-1, but not the�-opioid receptor agonist endomorphin-2, inhibited the cal-cium currents.
In summary, �-opioid receptors are located postsynapti-cally on premotor cardiac parasympathetic nucleus am-biguus neurons. The �-opioid receptor agonist endomor-phin1 inhibited the �-agatoxin-sensitive P/Q-type voltage-gated calcium currents in premotor cardiac vagal nucleusambiguus neurons. This inhibition is mediated via a G-pro-tein mediated pathway which was blocked by pretreatmentwith pertusis toxin. It is possible that the inhibition of calciumcurrents may act to indirectly facilitate the activity of premo-tor cardiac parasympathetic nucleus ambiguus neurons bydisinhibition, such as by a reduction in inhibitory calciumactivated potassium currents. © 2003 IBRO. Published byElsevier Science Ltd. All rights reserved.
Key words: baroreflex, heart rate, endomorphin, agatoxin,vagal, bradycardia.
Heart rate in healthy individuals is determined primarily bythe tonic and reflex control of parasympathetic activity thatinnervates the heart (Loewy and Spyer, 1990). However, inresponse to some cardiovascular challenges heart rateresponses are paradoxical and detrimental. Hypotensiontypically evokes an increase in sympathetic and a de-crease in parasympathetic activity which acts to restoreblood pressure to normal levels. In severe hemorrhagicand septic shock both the parasympathetic and sympa-thetic responses are initially compensatory but then a sec-ond paradoxical or “decompensatory” phase occurs inwhich the severe hypotension evokes a reflex bradycardiaand a decrease in vascular resistance causing blood pres-sure to fall precipitously (Evans et al., 1989; Ohnishi et al.,1997). The stimulus for this second phase has been attrib-uted to activation of vagal afferents from the heart while thereflex bradycardia is caused by an increase in efferentparasympathetic cardiac activity (Evans et al., 1989; Kwokand Dun, 1998). Application of low doses of opioid antag-onists into the CNS, but not intravenously, block the de-crease in heart rate during hemorrhagic shock indicatingthat activation of an endogenous opioid mechanism in theCNS is responsible for increasing cardiac parasympatheticactivity (Evans et al., 1989; Ohnishi et al., 1997; Ang et al.,1999).
Anatomical work has demonstrated that neurons in thenuclei where premotor cardiac parasympathetic neuronsare located, the nucleus ambiguus (NA), are densely in-nervated by opioid-containing pathways (Zamir et al.,1985; Morilak et al., 1989; Mansour et al., 1994; Batten,1995; Milner et al., 1995; Nomura et al., 1996). At the lightmicroscopic level enkephalin immunoreactive fibers and
*Corresponding author: Tel: �1-202-994-3466; fax: �1-202-994-2870.E-mail address: [email protected] (D. Mendelowitz).Abbreviations: AP5, D-2-amino-5-phosphonovalerate, CNQX,6-cyano-7-nitroquinoxaline; CTOP, D-Phe-Cys-Tyr-D-Trp-Orn-Thr-Pen-Thr-NH2, DIC, differential interference contrast; HEPES,4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid, MOR, �-opioidreceptor; NA, nucleus ambiguus, NMDA, N-methyl-D-asparate; NTS,nucleus tractus solitarius, PTX, pertusis toxin.
Neuroscience 116 (2003) 573–582
0306-4522/03$30.00�0.00 © 2003 IBRO. Published by Elsevier Science Ltd. All rights reserved.doi:10.1016/S0306-4522(02)00657-7
573

terminals are in close proximity to neurons in the NA,including premotor cardiac parasympathetic NA neurons(Milner et al., 1995; Nomura et al., 1996). These immuno-reactive terminals could be directly innervating postsynap-tic premotor cardiac parasympathetic NA neurons or thepresynaptic terminals of neurons that synapse upon pre-motor cardiac parasympathetic NA neurons. At least onesource of the opioid innervation of NA neurons originatesfrom the nucleus tractus solitarius (NTS) (Morilak et al.,1989).
Although there is a dense innervation of opioid-con-taining pathways in the NA, it is unknown whether premo-tor cardiac parasympathetic NA neurons possess postsyn-aptic opioid receptors. In addition the site and mechanismsof action of opioids on premotor cardiac parasympatheticNA neurons has also not been determined. This studyexamined whether premotor cardiac parasympathetic NAneurons possess postsynaptic opioid receptors. Since onecommon target of opioid receptor activation is voltage-gated calcium currents, the voltage-gated calcium currentsin premotor cardiac parasympathetic NA neurons wereidentified and the hypothesis that �-opioid receptor (MOR)agonists modulate these voltage-gated calcium currentswas tested.
EXPERIMENTAL PROCEDURES
Identification of premotor cardiac parasympatheticnucleus ambiguus neurons
All animal procedures were performed in compliance with theinstitutional guidelines at George Washington University, and arein accordance with the recommendations of the Panel on Eutha-nasia of the American Veterinary Medical Association and the NIHpublication “Guide for the Care and Use of Laboratory Animals.” Inan initial surgery, Sprague–Dawley rat pups (4–10 days old) wereanesthetized with the inhalation agent methoxyflurane and ex-posed to hypothermia during the surgery (10–15 min) to slow theheart and aid in recovery. A right thoracotomy was used to exposethe heart. For the anatomical experiments Fluoro-Gold (Fluoro-chrome Inc., 1% solution, 20 �l) was injected into the pericardialsac onto the surface of the cardiac tissue that contains the para-sympathetic ganglia. For electrophysiological experiments rhoda-mine (X-rhodamine-5-(and-6)-isothiocyanate (5(6), MolecularProbes, 1% solution, 20 �l) was used. Negative pressure wasapplied during the pericardial injection and until the thoracotomywas closed by a tight purse-string suture knot. Buprenorphine(0.1–0.5 mg/kg) was administered sub-Q for postoperative anal-gesia if there was any sign of stress such as labored breathing orlethargy. The animals were allowed 2–4 days to recover. Duringthe recovery period either Fluoro-Gold or rhodamine was trans-ported retrogradely and thereafter could be used to identify the cellbodies of these premotor cardiac parasympathetic NA neurons inthe nucleus ambiguus.
Perfusion and immunocytochemistry
Dual-labeling immunocytochemical methods were used to deter-mine if premotor cardiac parasympathetic NA neurons containMORs. Rats with prior cardiac injections of Fluoro-Gold (seeabove) were overdosed with pentobarbital sodium (150 mg/kg)and perfused through the ascending aorta with heparinized salinefollowed by a mixture of acrolein and paraformaldehyde (Aicher etal., 1999). The medulla oblongata was sectioned (40 �m) on avibrating microtome (Leica, Malvern, PA, USA) and collected into
0.1 M phosphate buffer. Tissue sections were processed for im-munocytochemical localization of Fluoro-Gold using an immuno-peroxidase method and MOR1 using an immunogold-silvermethod (Aicher et al., 1997, 1999). Tissue sections were incu-bated in a solution containing a rabbit anti-Fluoro-Gold IgG (1:10,000; Chemicon, Temecula, CA, USA) and a guinea-pig anti-MOR1 IgG (1:5000; Chemicon) for 40 h at 4 °C. The light micro-scopic distribution of MOR1 labeling with this antibody is identicalto the distribution previously reported using other MOR1 antibod-ies (Arvidsson et al., 1995; Ding et al., 1996). The distribution ofimmunocytochemical labeling obtained with the Fluoro-Gold anti-body was identical to the labeling detected with direct fluorescentvisualization of the tracer. The Fluoro-Gold antibody was visual-ized using a biotintylated goat anti-rabbit IgG (1:400; Vector Lab-oratories, Burlingame, CA, USA) and the avidin-biotin detectionmethod (Hsu and Raine, 1981). MOR1 was visualized using agoat anti-guinea-pig IgG conjugated to colliodal gold (1 nm; 1:50,BBI Goldmark, Phillipsburg, NJ, USA). Sections were rinsed incitrate buffer and colloidal gold particles were enhanced by silverintensification for 6–8 min using the IntenSEM kit (AmershamPharmacia Biotech, Piscataway, NJ, USA).
Electron microscopy
Following the immunocytochemical procedure, labeled tissue sec-tions were fixed for 1 h in 2.0% osmium tetroxide in 0.1 M PB,washed in 0.1 M PB, dehydrated through a graded series ofethanols, propylene oxide, and embedded between two sheets ofAclar plastic in EMBed (Aicher et al., 1999). Regions of the nu-cleus ambiguus containing cardiac premotor neurons were gluedto plastic blocks formed in Beem capsules and ultrathin sections(75 nm) through the outer surface of the tissue were collected ontocopper grids. The thin sections were counterstained with uranylacetate and Reynolds lead citrate and examined using a PhilipsCM10 electron microscope. Areas of tissue just below the tissue/plastic interface containing both labels were examined and pho-tographed. The cellular distribution of MOR1 relative to premotorcardiac parasympathetic NA neurons was assessed.
Electrophysiological experiments
On the day of the experiment, the animals were re-anesthetizedwith methoxyflurane and killed by rapid cervical dislocation. Thehindbrain was quickly removed and placed in cold (0–2 °C) bufferof the following composition (in mM): NaCl 140, KCl 5, CaCl2 2,glucose 5, HEPES 10, and continually gassed with 100% O2. Themedulla was then cut in transverse sections of 250-�m-thick slicesusing a vibratome. Slices were mounted in a perfusion chamberand submerged in a perfusate of the following composition (inmM): NaCl 125, KCl 3, BaCl2 5, NaHCO3 26, dextrose 5, HEPES5, constantly bubbled with gas (95% O2, 5% CO2), and maintainedat pH 7.4. Picrotoxin (100 �M), strychnine (1 �M), prazosin (10�M), and D-2-amino-5-phophono-valerate (50 �M) were added tothe bath perfusate to prevent GABAergic, glycine, �1 adrenergicand glutamatergic N-methyl-D-asparate (NMDA) postsynaptic cur-rents, respectively.
Individual vagal cardiac neurons were identified by the pres-ence of the fluorescent tracer. These identified vagal cardiacneurons were then imaged with differential interference contrast(DIC) optics, infrared illumination, and infrared-sensitive video-detection cameras to gain better spatial resolution and to visuallyguide and position the patch pipette onto the surface of theidentified neuron. Patch pipettes were mounted onto a pipetteholder and amplifier head stage (Axon Instruments, Axopatch200B) which were connected to micromanipulators (Narashige).The indifferent electrode was an Ag–AgCl plug submerged in thebath. The pipette was filled with a solution of the following com-position (in mM): CsCl 130, HEPES 10, EGTA 10, CaCl2 1, MgCl21, pH 7.4. QX-314 (5 mM). The pipette (2–5 megaohm resistance)
M. Irnaten et al. / Neuroscience 116 (2003) 573–582574

was advanced until it began to form a tight seal with the cellmembrane and whole cell access to the inside of the cell wasobtained by applying a brief period of suction to rupture the cellmembrane under the pipette tip.
Drug application
Drugs used in the experiments were: nimodipine (2 �M), �-conotoxinGVIA (1 �M), �-conotoxin MVIIC (5 �M) and �-agatoxin IVA (100nM), endomorphin-1 (1–100 �M), endomorphin-2 (100 �M), nalox-one (100 �M) and D-Phe-Cys-Tyr-D-Trp-Orn-Thr-Pen-Thr-NH2
(CTOP) (100 �M). All drugs were diluted with the perfusate solutionimmediately prior to use and were applied directly onto the neuronsfrom tubing positioned 2.0 mm above the neuron under study. Aconcentration of each drug was introduced using a pump at rate of0.3 ml/min and the solution was constantly suctioned out of thechamber at a fixed chamber height to keep the fluid within therecording chamber at a constant volume. Each concentration of eachdrug was maintained until a steady state response was obtained.Some slices were incubated in the presence of pertusis toxin (PTX,100 ng/ml) for 4–6 h at 37 °C and constantly bubbled with gas (95%O2, 5% CO2). All drugs were purchased from Sigma. To avoid anytolerance to opioids in the electrophysiological experiments only oneopioid agonist was applied at a single concentration to each cell.Each brainstem slice was used for only one experiment with a singleopioid agonist concentration.
Data acquisition and statistical analysis
Data acquisition and analysis were performed using pCLAMPsoftware (version 8.0, Axon Instruments) and Origin version 5. 0(Microcal Software, Northampton, MA, USA). Averaged data arepresented as mean�S.E.M., n�number of cells. Statistically sig-nificant differences were tested using paired t-tests for compari-sons of maximal responses and an analysis of variance (ANOVA)
with repeated measures was used to compare current-voltagerelationships. Differences were classified as significant at P�0.05.
RESULTS
Electron microscopy
The NA from five rats with successful retrograde and im-munocytochemical labeling and good to excellent ultra-structural morphology were included in the analysis. Asingle vibratome section through the NA was analyzed foreach rat and a total of 154 retrogradely labeled profileswere examined (12 perikarya, the rest were dendrites). Allof the retrogradely labeled perikarya and 78% of the den-drites of premotor cardiac parasympathetic NA neuronscontained immunoreactivity for MOR, see Fig. 1. MOR wasalso detected in other cells in the region, including presyn-aptic profiles of unknown origin. Although axons and axonterminals containing MOR occasionally contacted retro-gradely labeled neurons containing Fluoro-Gold, most ofthe MOR labeling was postsynaptic on the perikarya anddendrites of premotor cardiac parasympathetic NA neu-rons. A more detailed analysis of the distribution of MOR1in these and other premotor cardiac parasympathetic NAneurons was recently published (Aicher et al., 2001a).
Type of calcium channels present in premotorcardiac parasympathetic nucleus ambiguus neurons
Since the anatomical work demonstrated the majority ofMOR labeling was on perikarya and dendrites of premotor
Fig. 1. Dendrites of premotor cardiac parasympathetic NA neurons contain MORs. A large dendrite is shown which contains immunoperoxidaselabeling for Fluoro-Gold and immunogold labeling for MOR1 (FGold � MOR1-d). Immunogold particles for MOR1 were frequently associated withnon-synaptic portions of the plasma membrane (arrowheads). The dendrite shown received a synapse (curved arrow) from an unlabeled terminal (ut)and a direct apposition from another (ut). A dendrite containing only immunogold labeling for MOR1 (MOR-d1) and an unlabeled dendrite (ud) are inthe same field. Scale bar�0.5 �M.
M. Irnaten et al. / Neuroscience 116 (2003) 573–582 575

cardiac parasympathetic NA neurons the hypothesis thatMOR agonists alter voltage-gated calcium currents inthese neurons was tested. However, since there was noinformation on the types of voltage-gated calcium currentsin premotor cardiac parasympathetic NA neurons the cal-cium channel subtypes present first needed to be identi-fied. To pharmacologically identify the subtypes of voltage-gated calcium currents 4 specific calcium channel blockerswere utilized: �-conotoxin MVIIC (5 �M), nimodipine (2�M), �-agatoxin IVA (100 nM) and �-conotoxin GVIA (1�M), which are specific blockers of P/Q (Cav 2.1), L (Cav
1), P/Q (Cav 2.1), and N (Cav 2.2) type currents, respec-tively (Meir et al., 1999; Ertel et al., 2000). Calcium cur-rents were evoked by voltage steps from a holding poten-tial of �80 mV to voltages from �120 to �30 mV in 10-mVincrements. Application of 100 nM �-agatoxin IVA nearlyblocked the calcium currents in premotor cardiac parasym-pathetic NA neurons, Fig. 2A,B (n�9). Neither nimodipine,�-conotoxin MVIIC, or �-conotoxin GVIA had any signifi-cant effect on the inward calcium currents, Fig. 2B(P�0.05 compared with control). To insure the block with�-agatoxin IVA was not due to time-dependent rundown ofthe calcium currents, we examined calcium currents for 20min in the absence of calcium channel antagonists. Themaximum peak amplitude of the calcium currents was notsignificantly decreased during this period (Fig. 2C).
�-Opioid receptor modulation of �-agatoxinIVA-sensitive calcium channels
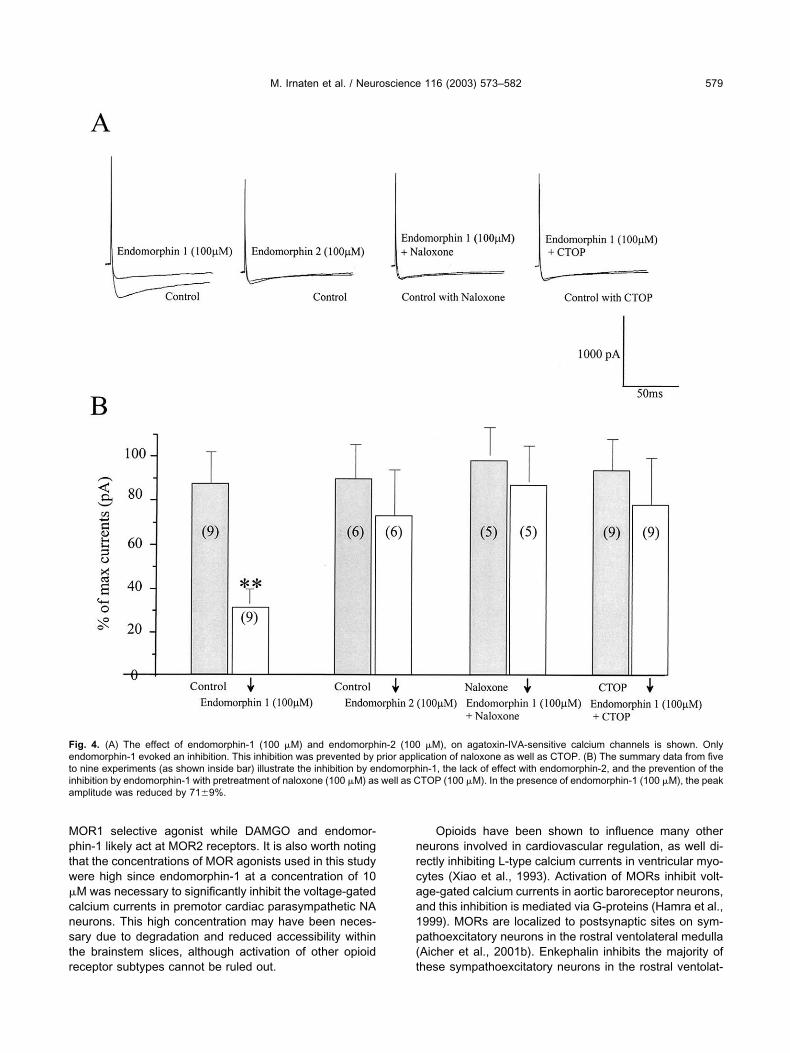
The modulation of �-agatoxin IVA-sensitive calciumchannels by MOR agonists was examined. Endomor-phin-1, at increasing concentrations from 1 �M to 100�M, evoked dose-dependent decreases in voltage-gated calcium currents (Fig. 3A,B). The attenuation wasdose-dependent and statistically significant at concen-trations of 10 �M and 100 �M (Fig. 3B). The peakcurrent amplitude of �-agatoxin IVA-sensitive calciumcurrents were decreased by 23�10%, 37�7% and71�9%, at endomorphin-1 concentrations of 1 �M, 10�M, and 100 �M, respectively.
To examine whether this inhibition could also beevoked by another MOR agonist the effect of endomor-phin-2 was tested. As shown from a typical experiment inFig. 4A, endomorphin-1 (100 �M) induced a decrease ofpeak amplitude calcium currents. However, application ofendomorphin-2 (100 �M) had no significant effect on thepeak amplitude of �-agatoxin IVA-sensitive calcium cur-rents in premotor cardiac parasympathetic NA neurons.Prior application of naloxone (100 �M), as well as CTOP(100 �M), prevented the inhibition by endomorphin-1. Thesummary data from these experiments is shown as histo-grams in Fig. 4B. Percentage of maximum peak amplitudeof �-agatoxin IVA-sensitive calcium currents was signifi-cantly reduced with endomorphin-1 by 71�9%, but notwith endomorphin-2 (16�17%). Pretreatment with nalox-one or CTOP prevented the inhibitory effect of endomor-phin-1.
Involvement of pertusis toxin-sensitive G-proteins
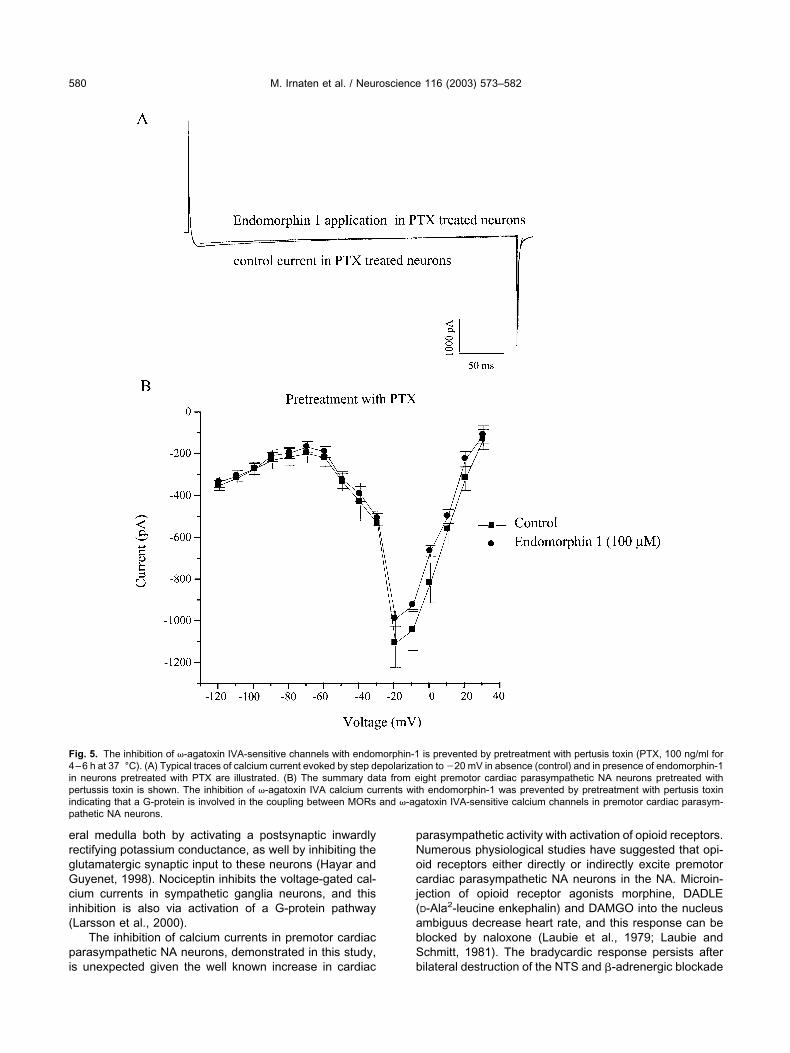
To determine whether or not endomorphin-1 modulates�-agatoxin IVA-sensitive calcium channels in premotorcardiac parasympathetic NA neurons via a G-protein cou-pled pathway, slices that contain premotor cardiac para-sympathetic NA neurons were preincubated with PTX (100ng/ml), an inhibitor of the �i/�o G-protein family (Aghaja-nian and Wang, 1986; Wimpey and Chavkin, 1991), for4–6 h at 37 °C. Figure 5A illustrates representative cal-cium currents in premotor cardiac parasympathetic NAneurons pretreated with PTX under control conditions andduring application of endomorphin-1. PTX abolished endo-morphin-1 elicited inhibition of �-agatoxin IVA-sensitivecalcium channels in premotor cardiac parasympathetic NAneurons, Fig. 5A,B. Figure 5B shows the average of I-Vrelationship of �-agatoxin-sensitive calcium currents inneurons pretreated with PTX. PTX pretreatment preventedthe inhibition by endomorphin-1, whereas in untreatedneurons endomorphin-1, at the same concentration (100�M), inhibited peak amplitude of calcium currents by71�9% (see Fig. 3B).
DISCUSSION
There are three major findings in this work. Using a com-bination of anatomical and electrophysiological tech-niques, this study demonstrates that MORs are locatedpostsynaptically on premotor cardiac parasympathetic NAneurons. Voltage-gated calcium currents in premotor car-diac parasympathetic NA neurons are comprised nearlyentirely of a P/Q-type voltage-gated calcium current that isblocked by �-agatoxin but is insensitive to �-conotoxinMVIIC. The MOR agonist endomorphin-1 inhibited thesevoltage-gated calcium currents in premotor cardiac para-sympathetic NA neurons via a G-protein mediated path-way.
It is somewhat surprising that the voltage-gated cal-cium currents in premotor cardiac parasympathetic NAneurons were blocked by the P/Q-type antagonist �-aga-toxin-IVA but were unaltered by �-conotoxin MVIIC. Inmany neurons, agatoxin IVA and �-Conotoxin MVIIC blockthe same P/Q type voltage-gated calcium current (Meir etal., 1999). However, in an earlier study of nicotinic excita-tion of premotor cardiac parasympathetic NA neurons thenicotinic responses were also blocked by �-agatoxin-IVAbut were unaltered by �-conotoxin MVIIC (Wang et al.,2001). In addition, there are other neurons in which apopulation of voltage-gated calcium currents are blockedby agatoxin IVA but are insensitive to �-conotoxin MVIIC.For example, in crayfish nerve terminals (Wright et al.,1996), peptidergic neurons (Garc et al., 1999) and teleostretinal cone horizontal neurons (Pfeiffer-Linn and Lasater,1996) agatoxin IVA nearly completely blocked voltage-gated calcium currents while these voltage-gated calciumcurrents were not altered by �-conotoxin MVIIC. In hu-mans there may be developmental changes in the sensi-tivity of voltage-gated calcium currents to blockers since ina 10-month-old child, but not in adults, hippocampal gran-ule cells are agatoxin IVA-sensitive but are insensitive to
M. Irnaten et al. / Neuroscience 116 (2003) 573–582576
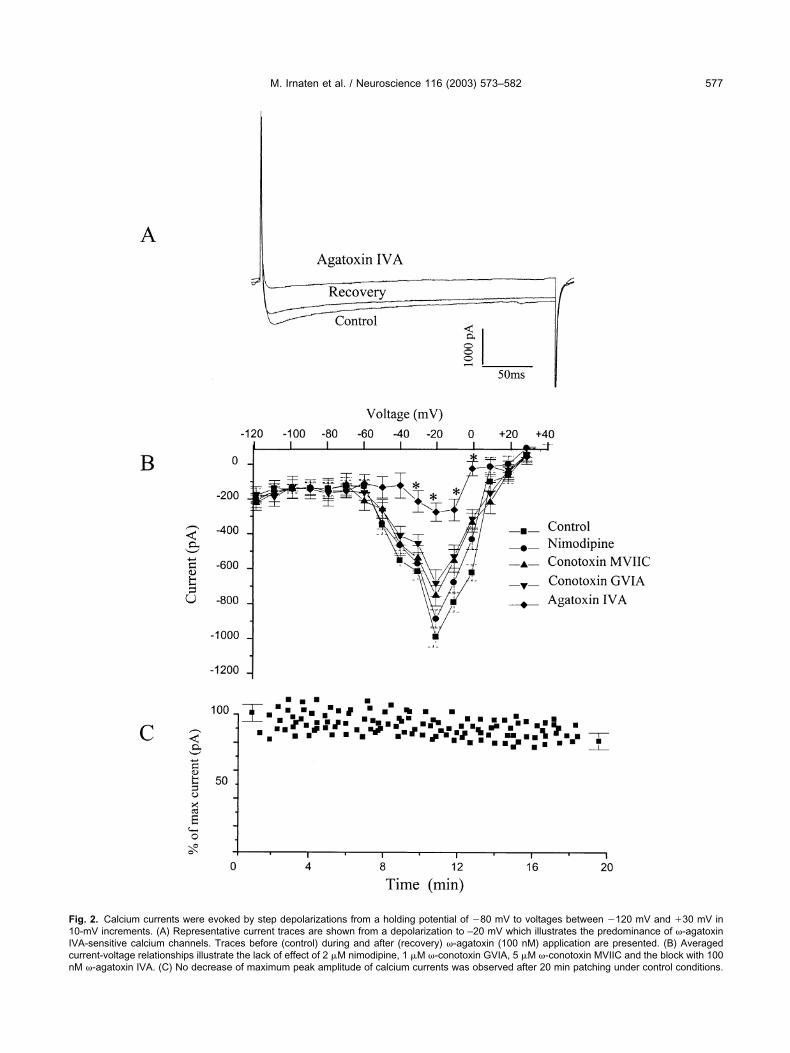
Fig. 2. Calcium currents were evoked by step depolarizations from a holding potential of �80 mV to voltages between �120 mV and �30 mV in10-mV increments. (A) Representative current traces are shown from a depolarization to –20 mV which illustrates the predominance of �-agatoxinIVA-sensitive calcium channels. Traces before (control) during and after (recovery) �-agatoxin (100 nM) application are presented. (B) Averagedcurrent-voltage relationships illustrate the lack of effect of 2 �M nimodipine, 1 �M �-conotoxin GVIA, 5 �M �-conotoxin MVIIC and the block with 100nM �-agatoxin IVA. (C) No decrease of maximum peak amplitude of calcium currents was observed after 20 min patching under control conditions.
M. Irnaten et al. / Neuroscience 116 (2003) 573–582 577

�-conotoxin MVIIC (Beck et al., 1997). It has recently beensuggested that there is a much greater diversity of voltagedependent calcium channel subtypes than previously rec-ognized and can be distinguished with the L, N, P, Q andR toxin based classification (Burley and Dolphin, 2000).
In this study endomorphin-1, but not endomorphin-2,inhibited P/Q-type calcium currents in premotor cardiacparasympathetic NA neurons. It is possible endomorphin-1and endomorphin-2 are agonists for different subtypes of
MORs. For example, endomorphin-2 has been proposedto be selective for MOR-1, but not MOR-2 receptors. Stud-ies of the antinociceptive activity of endomorphin-1 andendomorphin-2 (Kamei et al., 2000; Sakurada et al., 2001)as well as the actions of DAMGO (D-Ala2,MePhe4,Gly-ol5-enkephalin) on the hypothalamo-pituitary adrenal axis(Eisenberg, 1993) have found the actions of endomor-phin-2 were blocked by the selective MOR-1 antagonistnaloxonazine, indicating that endomorphin-2 can act as a
Fig. 3. (A) Typical current traces that illustrate the concentration-dependent inhibition of agatoxin-IVA-sensitive calcium currents during increasingendomorphin-1 concentrations is presented. Individual traces were evoked using 200 ms depolarizing pulses to �20 mV from a holding potential of�80 mV. (B) The graded inhibition by endomorphin-1 at increasing concentrations (1, 10, 100 �M) on the current-voltage relationship of voltage-gatedcalcium currents is illustrated.
M. Irnaten et al. / Neuroscience 116 (2003) 573–582578
MOR1 selective agonist while DAMGO and endomor-phin-1 likely act at MOR2 receptors. It is also worth notingthat the concentrations of MOR agonists used in this studywere high since endomorphin-1 at a concentration of 10�M was necessary to significantly inhibit the voltage-gatedcalcium currents in premotor cardiac parasympathetic NAneurons. This high concentration may have been neces-sary due to degradation and reduced accessibility withinthe brainstem slices, although activation of other opioidreceptor subtypes cannot be ruled out.
Opioids have been shown to influence many otherneurons involved in cardiovascular regulation, as well di-rectly inhibiting L-type calcium currents in ventricular myo-cytes (Xiao et al., 1993). Activation of MORs inhibit volt-age-gated calcium currents in aortic baroreceptor neurons,and this inhibition is mediated via G-proteins (Hamra et al.,1999). MORs are localized to postsynaptic sites on sym-pathoexcitatory neurons in the rostral ventolateral medulla(Aicher et al., 2001b). Enkephalin inhibits the majority ofthese sympathoexcitatory neurons in the rostral ventolat-
Fig. 4. (A) The effect of endomorphin-1 (100 �M) and endomorphin-2 (100 �M), on agatoxin-IVA-sensitive calcium channels is shown. Onlyendomorphin-1 evoked an inhibition. This inhibition was prevented by prior application of naloxone as well as CTOP. (B) The summary data from fiveto nine experiments (as shown inside bar) illustrate the inhibition by endomorphin-1, the lack of effect with endomorphin-2, and the prevention of theinhibition by endomorphin-1 with pretreatment of naloxone (100 �M) as well as CTOP (100 �M). In the presence of endomorphin-1 (100 �M), the peakamplitude was reduced by 71�9%.
M. Irnaten et al. / Neuroscience 116 (2003) 573–582 579
eral medulla both by activating a postsynaptic inwardlyrectifying potassium conductance, as well by inhibiting theglutamatergic synaptic input to these neurons (Hayar andGuyenet, 1998). Nociceptin inhibits the voltage-gated cal-cium currents in sympathetic ganglia neurons, and thisinhibition is also via activation of a G-protein pathway(Larsson et al., 2000).
The inhibition of calcium currents in premotor cardiacparasympathetic NA neurons, demonstrated in this study,is unexpected given the well known increase in cardiac
parasympathetic activity with activation of opioid receptors.Numerous physiological studies have suggested that opi-oid receptors either directly or indirectly excite premotorcardiac parasympathetic NA neurons in the NA. Microin-jection of opioid receptor agonists morphine, DADLE(D-Ala2-leucine enkephalin) and DAMGO into the nucleusambiguus decrease heart rate, and this response can beblocked by naloxone (Laubie et al., 1979; Laubie andSchmitt, 1981). The bradycardic response persists afterbilateral destruction of the NTS and -adrenergic blockade
Fig. 5. The inhibition of �-agatoxin IVA-sensitive channels with endomorphin-1 is prevented by pretreatment with pertusis toxin (PTX, 100 ng/ml for4–6 h at 37 °C). (A) Typical traces of calcium current evoked by step depolarization to �20 mV in absence (control) and in presence of endomorphin-1in neurons pretreated with PTX are illustrated. (B) The summary data from eight premotor cardiac parasympathetic NA neurons pretreated withpertussis toxin is shown. The inhibition f �-agatoxin IVA calcium currents with endomorphin-1 was prevented by pretreatment with pertusis toxinindicating that a G-protein is involved in the coupling between MORs and �-agatoxin IVA-sensitive calcium channels in premotor cardiac parasym-pathetic NA neurons.
M. Irnaten et al. / Neuroscience 116 (2003) 573–582580

(Laubie et al., 1979; Laubie and Schmitt, 1981). Endom-orphins administered intravenously also elicit decreases inheart rate and blood pressure (Kwok and Dun, 1998). Theheart rate responses can be blocked by atropine or bilat-eral vagotomy, suggesting cardiac parasympathetic neu-rons, as well as possibly vagal afferents, are involved(Kwok and Dun, 1998). It is possible that the inhibition ofcalcium currents in premotor cardiac parasympathetic NAneurons, rather than evoking an overall inhibition of pre-motor cardiac parasympathetic NA neurons, may act toindirectly facilitate their activity by disinhibition, such as bya reduction in inhibitory calcium activated potassium cur-rents. Inhibition of the apamin-sensitive calcium activatedpotassium currents has been shown to greatly increase theexcitability and firing in these neurons (Mendelowitz,1996). It is also possible that the inhibition of calciumcurrents is opposed by stronger opioid mediated excitatoryinfluences that act either on other voltage-gated currentsor synaptic pathways to these neurons. For example, re-cent work has shown both nociceptin and endomorphin-2inhibit GABAergic synaptic inputs to premotor cardiacparasympathetic NA neurons and might therefore disinhibitthese neurons (Venkatesan et al., 2002a, b). It is possiblethat opioid mediated changes in synaptic pathways topremotor cardiac parasympathetic NA neurons play amore important role than the opioid mediated effects onpostsynaptic currents in determining cardiac parasympa-thetic activity.
Acknowledgements—This work was supported from NIH grantsHL 49965 and 59895 to D.M. and HL 56301 to S.A.A.
REFERENCES
Aghajanian GK, Wang YY (1986) Pertussis toxin blocks the outwardcurrents evoked by opiate and alpha 2-agonists in locus coeruleusneurons. Brain Res 371:390–394.
Aicher SA, Sharma S, Pickel VM (1999) N-methyl-D-aspartate recep-tors are present in vagal afferents and their dendritic targets in thenucleus tractus solitarius. Neuroscience 91:119–132.
Aicher SA, Sharma S, Cheng PY, Pickel VM (1997) The N-methyl-D-aspartate (NMDA) receptor is postsynaptic to substance P-contain-ing axon terminals in the rat superficial dorsal horn. Brain Res772:71–81.
Aicher SA, Mitchell JL, Sharma S, Mendelowitz D (2001a) Distributionof mu-opioid receptors in rat visceral premotor neurons. Neuro-science 115:851–860.
Aicher SA, Schreihofer AM, Kraus JA, Sharma S, Milner TA, GuyenetPG (2001b) Mu-opioid receptors are present in functionally identi-fied sympathoexcitatory neurons in the rat rostral ventrolateral me-dulla. J Comp Neurol 433:34–47.
Ang KK, McRitchie RJ, Minson JB, Llewellyn-Smith IJ, Pilowsky PM,Chalmers JP, Arnolda LF (1999) Activation of spinal opioid recep-tors contributes to hypotension after hemorrhage in conscious rats.Am J Physiol 276:H1552–H1558.
Arvidsson U, Riedl M, Chakrabarti S, Lee JH, Nakano AH, Dado RJ,Loh HH, Law PY, Wessendorf MW, Elde R (1995) Distribution andtargeting of a mu-opioid receptor (MOR1) in brain and spinal cord.J Neurosci 15:3328–3341.
Batten TF (1995) Immunolocalization of putative neurotransmittersinnervating autonomic regulating neurons (correction of neurones)of cat ventral medulla. Brain Res Bull 37:487–506.
Beck H, Steffens R, Heinemann U, Elger CE (1997) Properties of
voltage-activated Ca2� currents in acutely isolated human hip-pocampal granule cells. J Neurophysiol 77:1526–1537.
Burley JR, Dolphin AC (2000) Overlapping selectivity of neurotoxinand dihydropyridine calcium channel blockers in cerebellar granuleneurones. Neuropharmacology 39:1740–1755.
Ding YQ, Kaneko T, Nomura S, Mizuno N (1996) Immunohistochem-ical localization of mu-opioid receptors in the central nervous sys-tem of the rat. J Comp Neurol 367:375–402.
Eisenberg RM (1993) DAMGO stimulates the hypothalamo-pituitary-adrenal axis through a mu-2 opioid receptor. J Pharmacol Exp Ther266:985–991.
Ertel EA, Campbell KP, Harpold MM, Hofmann F, Mori Y, Perez-ReyesE, Schwartz A, Snutch TP, Tanabe T, Birnbaumer L, Tsien RW,Catterall WA (2000) Nomenclature of voltage-gated calcium chan-nels. Neuron 25:533–535.
Evans RG, Ludbrook J, Potocnik SJ (1989) Intracisternal naloxoneand cardiac nerve blockade prevent vasodilatation during simulatedhaemorrhage in awake rabbits. J Physiol 409:1–14.
Garc IACJ, Valdiosera R, Garc IU (1999) P-type Ca2� current incrayfish peptidergic neurones. J Exp Biol 202:429–440.
Hamra M, McNeil RS, Runciman M, Kunze DL (1999) Opioid modu-lation of calcium current in cultured sensory neurons: Mu-modula-tion of baroreceptor input. Am J Physiol 277:H705–H713.
Hayar A, Guyenet PG (1998) Pre- and postsynaptic inhibitory actionsof methionine-enkephalin on identified bulbospinal neurons of therat RVL. J Neurophysiol 80:2003–2014.
Hsu SM, Raine L (1981) Protein A, avidin, and biotin in immunohisto-chemistry. J Histochem Cytochem 29:1349–1353.
Kamei J, Zushida K, Ohsawa M, Nagase H (2000) The antinociceptiveeffects of endomorphin-1 and endomorphin-2 in diabetic mice. EurJ Pharmacol 391:91–96.
Kwok EH, Dun NJ (1998) Endomorphins decrease heart rate andblood pressure possibly by activating vagal afferents in anesthe-tized rats. Brain Res 803:204–207.
Larsson KP, Olsen UB, Hansen AJ (2000) Nociceptin is a potentinhibitor of N-type Ca(2�) channels in rat sympathetic ganglionneurons. Neurosci Lett 296:121–124.
Laubie M, Schmitt H (1981) Indication for central vagal endorphinergiccontrol of heart rate in dogs. Eur J Pharmacol 71:401–409.
Laubie M, Schmitt H, Vincent M (1979) Vagal bradycardia produced bymicroinjections of morphine-like drugs into the nucleus ambiguus inanaesthetized dogs. Eur J Pharmacol 59:287–291.
Loewy A, Spyer K (1990) Central regulation of autonomic functions.Oxford University Press: London.
Mansour A, Fox CA, Thompson RC, Akil H, Watson SJ (1994) mu-Opioid receptor mRNA expression in the rat CNS: Comparison tomu-receptor binding. Brain Res 643:245–265.
Meir A, Ginsburg S, Butkevich A, Kachalsky SG, Kaiserman I, AhdutR, Demirgoren S, Rahamimoff R (1999) Ion channels in presynapticnerve terminals and control of transmitter release. Physiol Rev79:1019–1088.
Mendelowitz D (1996) Firing properties of identified parasympatheticcardiac neurons in nucleus ambiguus. Am J Physiol 271:H2609–H2614.
Milner TA, Okada J, Pickel VM (1995) Monosynaptic input from Leu5-enkephalin-immunoreactive terminals to vagal motor neurons in thenucleus ambiguus: Comparison with the dorsal motor nucleus ofthe vagus. J Comp Neurol 353:391–406.
Morilak DA, Somogyi P, McIlhinney RA, Chalmers J (1989) An en-kephalin-containing pathway from nucleus tractus solitarius to thepressor area of the rostral ventrolateral medulla of the rabbit. Neu-roscience 31:187–194.
Nomura S, Ding YQ, Kaneko T, Li JL, Mizuno N (1996) Localization ofmu-opioid receptor-like immunoreactivity in the central componentsof the vagus nerve: A light and electron microscope study in the rat.Neuroscience 73:277–286.
Ohnishi M, Kirkman E, Marshall HW, Little RA (1997) Morphine blocks
M. Irnaten et al. / Neuroscience 116 (2003) 573–582 581

the bradycardia associated with severe hemorrhage in the anes-thetized rat. Brain Res 763:39–46.
Pfeiffer-Linn CL, Lasater EM (1996) Whole cell and single-channelproperties of a unique voltage-activated sustained calcium currentidentified in teleost retinal horizontal cells. J Neurophysiol 75:609–619.
Sakurada S, Hayashi T, Yuhki M, Orito T, Zadina JE, Kastin AJ,Fujimura T, Murayama K, Sakurada C, Sakurada T, Narita M,Suzuki T, Tan-no K, Tseng LF (2001) Differential antinociceptiveeffects induced by intrathecally administered endomorphin-1and endomorphin-2 in the mouse. Eur J Pharmacol 427:203–210.
Venkatesan P, Wang J, Evans C, Irnaten M, Mendelowitz D (2002a)Endomorphin -2 inhibits GABAergic inputs to cardiac parasympatheticneurons in the nucleus ambiguus. Neuroscience 111:699–705.
Venkatesan P, Wang J, Evans C, Irnaten M, Mendelowitz D (2002b)Nociceptin inhibits gamma-aminobutyric acidergic inputs to cardiac
parasympathetic neurons in the nucleus ambiguus. J PharmacolExp Ther 300:78–82.
Wang J, Irnaten M, Mendelowitz D (2001) Agatoxin-IVA-sensitive cal-cium channels mediate the presynaptic and postsynaptic nicotinicactivation of cardiac vagal neurons. J Neurophysiol 85:164–168.
Wimpey TL, Chavkin C (1991) Opioids activate both an inward rectifierand a novel voltage-gated potassium conductance in the hip-pocampal formation. Neuron 6:281–289.
Wright SN, Brodwick MS, Bittner GD (1996) Presynaptic calcium cur-rents at voltage-clamped excitor and inhibitor nerve terminals ofcrayfish. J Physiol 496 Pt 2347–361.
Xiao RP, Spurgeon HA, Capogrossi MC, Lakatta EG (1993) Stimulationof opioid receptors on cardiac ventricular myocytes reduces L typeCa2� channel current. J Mol Cell Cardiol 25:661–666.
Zamir N, Palkovits M, Brownstein M (1985) Distribution of immunore-active Met-enkephalin-Arg6-Gly7-Leu8 and Leu-enkephalin in dis-crete regions of the rat brain. Brain Res 326:1–8.
(Accepted 30 August 2002)
M. Irnaten et al. / Neuroscience 116 (2003) 573–582582










![Endomorphin synthesis in rat brain from intracerebroventricularly injected [3H]-Tyr-Pro: A possible biosynthetic route for endomorphins](https://img.pdfslide.net/doc/110x75/6354e584922cbb7c550c51d8/endomorphin-synthesis-in-rat-brain-from-intracerebroventricularly-injected-3h-tyr-pro.jpg)


















