Embed Size (px)
Citation preview

The Prostate 69:706 ^718 (2009)
Over-Expressionof IkB-Kinase-e (IKKe/IKKi) InducesSecretionof InflammatoryCytokines in Prostate
CancerCell Lines
Benjamin Peant,1 Jean-Simon Diallo,1 Florent Dufour,1 Cecile Le Page,1
Nathalie Delvoye,1 Fred Saad,1,2,3 and Anne-Marie Mes-Masson1,4*1Centre derecherche duCentrehospitalierdel’Universite¤ deMontre¤ al (CR-CHUM)/InstitutducancerdeMontre¤ al,
Que¤ bec,Canada2De¤ partementdeChirurgie del’Ho“ pitalNotre-Dame (CHUM),Que¤ bec,Canada
3De¤ partementdeChirurgie del’Universite¤ deMontre¤ al,Que¤ bec,Canada4De¤ partementdeMe¤ decine del’Universite¤ deMontre¤ al,Que¤ bec,Canada
BACKGROUND. Elevated inflammatory cytokine levels in serum have been associated withadvanced stage metastasis-related morbidity in prostate cancer. Several studies have shownthat IL-6 and IL-8 can accelerate the growth of human prostate cancer cell lines. Previous studies,in murine embryonic fibroblasts, have shown that Ik-B kinase-epsilon (IKKe/IKKi)-deficiencyresults in the reduction of lipopolysaccharide-mediated expression of IL-6.RESULTS. In this study, we report that over-expression of IKKe in hormone-sensitive 22Rv1and LNCaP prostate cancer cells induces the secretion of several inflammatory cytokinesincluding IL-6 and IL-8. Both of these cytokines are secreted by hormone-refractory PC-3prostate cancer cells and IKKe knock-down in these cells correlates with a strong decrease inIL-6 secretion. Furthermore, we demonstrate that IKKe over-expression does not induce theactivation of the IKKe classical targets NF-kB and IRF-3, two transcription factors involved in theregulation of several cytokines. Finally, we observe that high IKKe expression results in itsnuclear translocation, a phenomena that is TBK1-independent.CONCLUSIONS. This study identifies IKKe as a potential prostate cancer gene that may favorchronic inflammation and create a tumor-supporting microenvironment that promotes prostatecancer progression, particularly by the induction of IL-6 secretion that may act as a positivegrowth factor in prostate cancer. Prostate 69: 706–718, 2009. # 2009 Wiley-Liss, Inc.
KEY WORDS: prostate cancer; IKKe/IKKi; cytokine secretion; IL-6; IL-8
INTRODUCTION
Prostate cancer (PCa) typically progresses from ahormone sensitive (HS) to a hormone refractory (HR)/chemoresistant state [1]. Given the high incidence ofPCa (1/7 men) and the limited therapeutic optionsavailable to treat HR disease, it is necessary to improveour understanding of the biological mechanismsinvolved in HR tumor progression. Recurrent orchronic inflammation has been linked to various stepsof carcinogenesis including cellular transformation,survival, proliferation, invasion, angiogenesis, andmetastasis. Inflammation is thought to be involved in
the development of several types of human cancerincluding liver, stomach, intestine, prostate, andbladder cancers [2,3]. Among other cancers, the release
*Correspondence to: Anne-Marie Mes-Masson, CR-CHUM—Institutdu Cancer de Montreal, Hopital Notre-Dame (CHUM), 1560 rueSherbrooke Est (Y-4609), Montreal, QC, Canada H2L 4M1.E-mail: [email protected] 1 December 2008; Accepted 5 December 2008DOI 10.1002/pros.20912Published online 23 January 2009 in Wiley InterScience(www.interscience.wiley.com).
0 2009 Wiley-Liss, Inc.

of cytokines plays a major role in mediating inflamma-tion and the immune response [4,5].
In the tumor microenvironment, cytokines aresecreted both by the tumor-infiltrating immune cellsas well as the tumor cells themselves. Several pro-inflammatory cytokines, including interleukin (IL)-1a,IL-1b, IL-6, IL-8, and regulated on activation of normalT cells expressed and secreted (RANTES) have beenshown to be involved in carcinogenesis (reviewed inRef. [6]). In human breast cancer cells, IL-1a expressioncorrelates with both poor cell differentiation anddecreased epithelial estrogen receptor-a expression[7,8]. Elevated IL-6 [9,10], RANTES [11], IL-8 [12,13],and IL-1b [14] serum levels are detected in breast cancerpatients with a high recurrence rate and unfavorableprognosis. Comparing serum cytokine levels amongmen with HS versus HR disease revealed a significantelevation in inflammatory cytokine levels, such as IL-6and IL-8, in men with HR PCa [15–20]. There is alsogrowing evidence to suggest that IL-6, a pleiotropiccytokine originally identified as a regulator of immuneand inflammatory responses, can act as an autocrinegrowth factor in PCa progression [21–23]. IL-6 has alsobeen shown to increase nuclear androgen receptor (AR)translocation in PCa cells and activate AR-mediatedtranscriptional activity in HS LNCaP cells in theabsence of androgen [24–27]. In PCa, the levels ofIL-6 and its receptor gp130 are increased in organ-confined tumors and IL-6 has a growth-promoting roleand acts as a positive growth factor for most PCa cells[28]. Increased IL-6 levels have been found in both PCapatient serum [21,29] and PCa tissues [15,30]. Inaddition, there is a direct correlation between elevatedprostate specific antigen (PSA) and IL-6 in HR disease[31]. Therefore, the role of inflammatory cytokines inPCa may be dual: cytokines could have a trans-formation-promoting role during prostate inflamma-tion and may also act as a positive growth factor inresponse to androgen deprivation therapy and thuscontribute to HR PCa.
The transcription factor nuclear factor-kB (NF-kB) isa major regulator of pro-inflammatory genes includingcytokines and chemokines (IL-6, RANTES, IL-8, andGro-a) [32]. This transcription factor is constitutivelyactive in PCa cell lines lacking AR expression (i.e., PC-3and DU145), whereas in HS cell lines (i.e., LNCaP and22Rv1) only very low levels of NF-kB activity aremeasured [33]. Therefore constitutive NF-kB activationmay play a role in PCa progression and contributeto PCa cell survival following androgen withdrawal[34–40]. In line with this hypothesis, we have foundthat NF-kB nuclear translocation in primary PCatissues correlates with poor patient outcome andlymph node metastasis [41–43]. Recently, we showedthat IKKe (IkB kinase-e/IKKi) expression, but not its
partner Tank Binding Kinase-1 (TBK-1), is under thecontrol of NF-kB in PCa cells and we noted high IKKeexpression in HR PC-3 cells which exhibit constitutiveNF-kB activity. We also observed that AR stimulationinduced a de novo IkBa synthesis and, indirectly, adownregulation of IKKe expression in a LNCaP cellline [44].
The cytosolic heterodimer IKKe-TBK1 is involved intoll like receptor 3- (TLR3) and TLR4-mediatedinterferon INF-b production through activation ofIRF-3 (IFN regulatory factor) and IRF-7 [45,46]. Like-wise, the IRF-3-responsive promoters of IFN-a4 andRANTES are also stimulated in response to IKKeexpression [47], an effect that is blocked by a dominantnegative form of IRF-3 [48]. IKKe is constitutivelyexpressed in human-like synoviocytes from rheuma-toid arthritis patients and could be associated with therheumatoid arthritis inflammation phenotype [49].Finally, IKKe-deficiency results in the reduction ofLPS-mediated IL-6 gene expression in murine embry-onic fibroblasts (MEFs) [50]. Combined, this datasuggests that IKKe can impact the expression andregulation of inflammatory cytokines.
Despite a constitutive IKKe expression in HR PCacell lines and its association with the processes ofinflammation, little is known about the potential role ofIKKe in the context of prostate inflammation and in PCaprogression to a HR phenotype. In the present study,we observe that TBK-1-independent nuclear trans-location of IKKe correlates with inflammatorycytokine secretions in HS PCa cell lines by an NF-kB-and IRF-3-independent mechanism.
MATERIALSANDMETHODS
Cell Lines andCell Culture
HS LNCaP and 22Rv1 cells and HR PC-3 cells werepurchased from the American Type Culture Collection(ATCC CRL-1740, ATCC CRL-2505 and ATCC CRL-1435 respectively; Manassas, VA). Cells were grown inRPMI 1640 (Wisent, Inc., St-Bruno, Quebec, Canada)supplemented with 100 mg/ml gentamicin, 0.25 mg/mlamphotericin B (Invitrogen, Paisley, United Kingdom),and 10% fetal calf serum (FCS). Tetracycline, Zeocinand Blasticidin S were purchased from Invitrogen andhuman recombinant TNF-a was obtained from RocheApplied Science (Indianapolis, IN).
Plasmids Construction
pUNO-hIKKe was obtained from Invitrogen andpUNO control vector was derived from pUNO-hIKKeby deletion of the hIKKe gene. For the construction ofthe pENTRTM/H1/TO-shIKKe vector, small interfer-ing oligonucleotides specific for IKKewere synthesized
The Prostate
IKKeExpression Induces Cytokine Secretion 707

(Forward: 50-caccgcaccacatctatatccatgcttcaagagagcatg-gatatagatgtggtgc-30, Reverse: 50 aaaagcaccacatctatatc-catgctctcttgaagcatggatatagatgtggtgc-30) and annealedto generate a double-stranded oligo (ds-oligo) accord-ing to the manufacturer’s instructions (Invitrogen). Theds-oligo was cloned into the pENTRTM/H1/TO vector(Invitrogen) and transformed into competent TOP10bacteria for selection and amplification. Plasmids wereverified by sequencing.
Transfection andCreation of Stable Cell Lines
Transient transfection of LNCaP and 22Rv1 cellswith the pUNO and pUNO-hIKKe plasmids was doneusing LipofectAMINE reagent (Invitrogen) accordingto the manufacturer’s instructions. Double strandRNA mimetic polyinosinic/polycytidylic acid(pI/pC) (Calbiochem, San Diego, CA) was heated to908C prior to transient transfection using Lipofect-AMINE reagent (Invitrogen).
For the creation of stable cell lines, PC-3 cells weretransfected with pcDNA6/TR (Invitrogen). Selection ofPC-3-pcDNA6/TR clones was done in RPMI supple-mented with 20% FCS and 12 mg/ml Blasticidin S.Several clones were probed for Tetracyclin-repressor(TetR) expression in comparison to wild type PC-3 cellsby western blot using anti-TetR antibody. A clone over-expressing TetR was selected and used to create stablePC-3/6TR-shIKKe cell lines as recommended by themanufacturer. PC-3/6TR cells were transfected withthe pENTRTM/H1/TO-shIKKe vector and selected inRPMI-20% FCS supplemented with 1 mg/ml Zeocin.Clones were isolated by serial dilution. Three PC-3/6TR-shIKKe clones with different efficiencies of tetra-cycline-inducible IKKe repression were selected forstudy.
LuciferaseAssays
NF-kB activity was measured using the 3kB-conA-Firefly plasmid or conA-Firefly vector as control (200ng/well) in the presence of the pUNO-hIKKe or pUNOvector (200 ng/well) according to previously publishedconditions [44]. Luciferase activity was normalizedusing the pCMV-Renilla plasmid (20 ng/well)which constitutively expresses the Renilla luciferasegene (Promega, Madison, WI). TNF-a stimulation(10 ng/ml) was used as a control for NF-kB activitymeasurement in 3kB-conA-Firefly and conA-Fireflytransfected PCa cells. All transfection experimentswere carried out in duplicate and repeated three times.
ELISA
IL-1a, IL-6, IL-8, CXCL-10/IP-10, RANTES, andMIF concentrations in culture media were
measured according to manufacturer’s conditionsusing Quantikine and DuoSet ELISA kits from R&DSystems (Minneapolis, MN). IFN-b secretion wasevaluated using ELISA kit from PBL Interferon Source(Piscataway, NJ). The detection limit was 3.12 pg/mlfor IL-6, 3.9 pg/ml for IL-1a, 6.25 pg/ml for IP-10,25 pg/ml for IFN-b and 31.25 pg/ml for IL-8,RANTES and MIF. Cell supernatants were filtratedusing 0.2 mm syringe filter and stored at �808C.Remaining cells were scraped and proteins wereextracted. Cytokine secretion was standardized bysample protein concentration. All assays were carriedout in triplicate and each experiment was repeatedthree times.
Protein Extraction
Cell membranes were lysed with ice cold buffer I (10mM HEPES, 50 mM NaCl, 10 mM EDTA, 5 mM MgCl2)with freshly added protease and phosphatase inhib-itors (protease inhibitor cocktail from Roche, 5 mMNaF, 200 mM Na3VO4) and 1% Igepal CA-630 (Sigma–Aldrich, St. Louis, MI), cytosolic fractions werecollected. Ice cold buffer II (10 mM HEPES, 400 mMNaCl, 0.1 mM EDTA, 0.5 mM DTT) with freshly addedprotease and phosphatase inhibitors was then added tothe nuclear fraction. Each fraction was immediatelystored at �808C. Whole cell extracts (WCE) wereobtained after 30 min in lysis buffer (1% IgepalCA-630, 10% Glycerol, 50 mM Tris, 2 mM EDTA,5 mM NaF, 150 mM NaCl) with freshly added proteaseand phosphatase inhibitors. Protein concentration wasmeasured using a Bradford assay (Bio-Rad Laborato-ries, Inc., Hercules, CA) according to the manufac-turer’s instructions.
Western BlotAnalysis
For Western blot analysis, 15–30 mg of proteins fromWCE or cell fractions were resolved on 7.5–10%polyacrylamide gels and then transferred onto PVDFmembranes.
Anti-IKKe/TBK-1 antibodies were purchased fromIMGENEX (San Diego, CA) while the anti-AR antibodywas obtained from NeoMarkers (Fremont, CA). Anti-bodies recognizing IRF-3, p65, c-Rel, and IkBa wereobtained from Santa Cruz Biotechnology, Inc. (SantaCruz, CA), whereas antibodies specific for phospho-Ser536-p65, p100/p52 and p105/p50 were purchasedfrom Upstate Cell Signaling (Charlottesville, VA). Toensure equal protein loading, membranes were probedwith an anti-b-actin antibody (Abcam, Cambridge,UK). Cytosolic contamination of nuclear fractions wasevaluated using an anti-a-tubulin antibody (Santa CruzBiotechnology, Inc.).
The Prostate
708 Pe¤ ant et al.
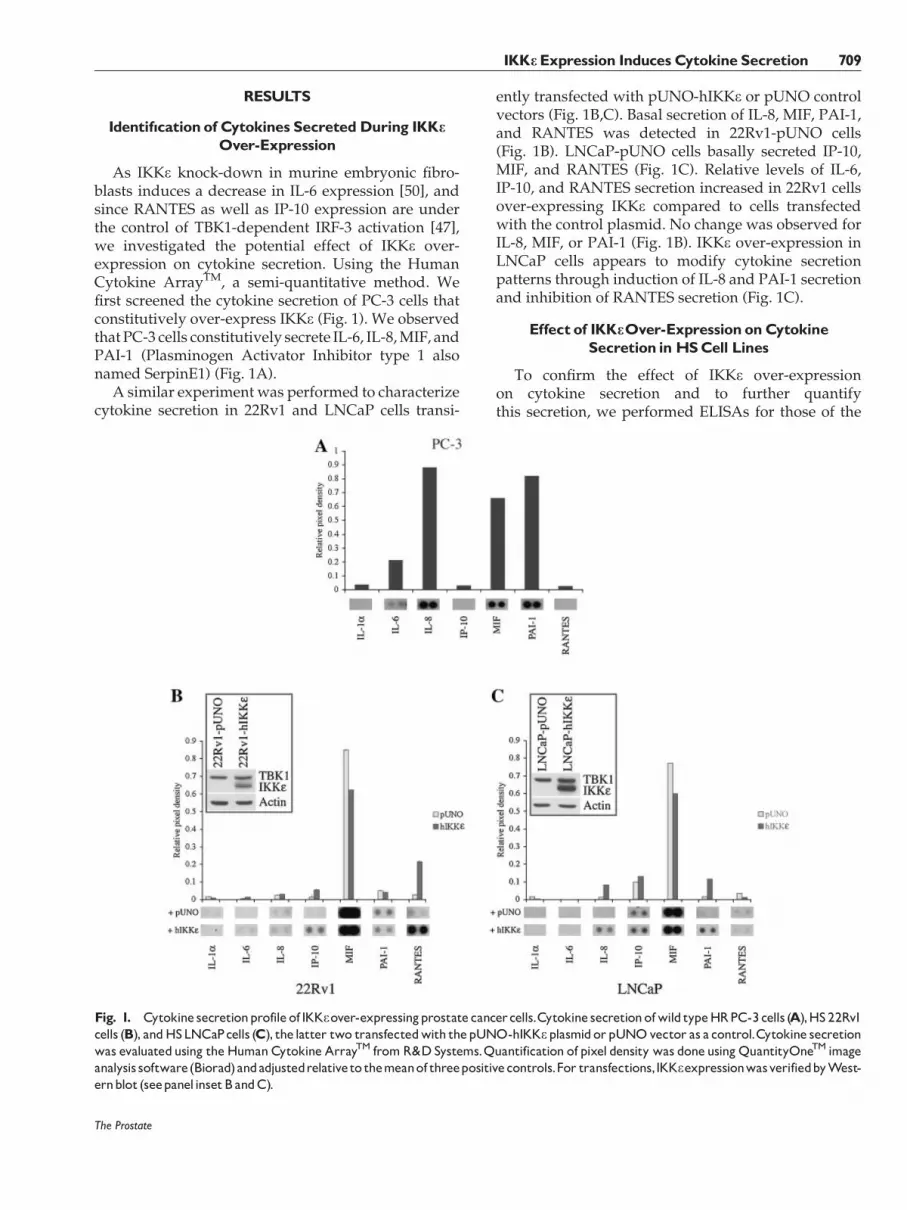
RESULTS
Identif|cation of Cytokines SecretedDuring IKKeOver-Expression
As IKKe knock-down in murine embryonic fibro-blasts induces a decrease in IL-6 expression [50], andsince RANTES as well as IP-10 expression are underthe control of TBK1-dependent IRF-3 activation [47],we investigated the potential effect of IKKe over-expression on cytokine secretion. Using the HumanCytokine ArrayTM, a semi-quantitative method. Wefirst screened the cytokine secretion of PC-3 cells thatconstitutively over-express IKKe (Fig. 1). We observedthat PC-3 cells constitutively secrete IL-6, IL-8, MIF, andPAI-1 (Plasminogen Activator Inhibitor type 1 alsonamed SerpinE1) (Fig. 1A).
A similar experiment was performed to characterizecytokine secretion in 22Rv1 and LNCaP cells transi-
ently transfected with pUNO-hIKKe or pUNO controlvectors (Fig. 1B,C). Basal secretion of IL-8, MIF, PAI-1,and RANTES was detected in 22Rv1-pUNO cells(Fig. 1B). LNCaP-pUNO cells basally secreted IP-10,MIF, and RANTES (Fig. 1C). Relative levels of IL-6,IP-10, and RANTES secretion increased in 22Rv1 cellsover-expressing IKKe compared to cells transfectedwith the control plasmid. No change was observed forIL-8, MIF, or PAI-1 (Fig. 1B). IKKe over-expression inLNCaP cells appears to modify cytokine secretionpatterns through induction of IL-8 and PAI-1 secretionand inhibition of RANTES secretion (Fig. 1C).
Effect of IKKeOver-Expression onCytokineSecretion inHSCell Lines
To confirm the effect of IKKe over-expressionon cytokine secretion and to further quantifythis secretion, we performed ELISAs for those of the
The Prostate
Fig. 1. Cytokine secretionprofile of IKKeover-expressingprostate cancercells.Cytokine secretionofwild typeHRPC-3cells (A),HS22Rv1cells (B), andHSLNCaPcells (C), the latter two transfectedwith thepUNO-hIKKeplasmidorpUNOvector as a control.Cytokine secretionwas evaluatedusing theHumanCytokine ArrayTM fromR&DSystems.Quantification of pixel density was doneusingQuantityOneTM imageanalysis software(Biorad)andadjustedrelativeto themeanof threepositivecontrols.For transfections,IKKeexpressionwasverifiedbyWest-ernblot (seepanelinsetBandC).
IKKeExpression InducesCytokine Secretion 709

six cytokines identified for which reagents were readilyavailable (Fig. 2). We failed to detect any IL-1a secretionby 22Rv1 or LNCaP cells, in the presence or absence ofpUNO-hIKKe vector. As observed using the humancytokine array, IKKe over-expression in 22Rv1 cellsinduced high secretion of RANTES (510 pg/ml) andIP-10 (590 pg/ml), two cytokines that were not secretedby 22Rv1-pUNO cells (Fig. 2A) and wild-type 22Rv1cells. IKKe over-expression also induced an increasein MIF secretion (2,150 pg/ml) in 22Rv1 cells whencompared to the basal expression of this cytokine(680 pg/ml). Wild type (data not shown) and pUNOtransfected LNCaP cells constitutively secreted IP-10(320 pg/ml), MIF (950 pg/ml), and RANTES (25 pg/ml). IKKe over-expression in LNCaP cells induced amoderate but significant increase in RANTES secretion(37 pg/ml), but no change in IP-10 and MIF secretion(Fig. 2B). While IL-6 and IL-8 were not secreted in wild-type (data not shown) or control transfected cells, IKKeover-expression resulted in IL-6 [130 pg/ml (22Rv1-pUNO-hIKKe), 22 pg/ml (LNCaP-pUNO-hIKKe)] andIL-8 [170 pg/ml (22Rv1-pUNO-hIKKe), 175 pg/ml(LNCaP-pUNO-hIKKe)] secretion in both cell lines.
Effectof IKKeKnock-DownonCytokineSecretion inHRPC-3 Cells
These observations suggest a correlation betweencytokine secretion and IKKe over-expression in HS celllines. Because HR PC-3 cells constitutively expressIKKe and secrete high levels of IL-6 and IL-8 [44,51,52],we studied the effect of IKKe knock-down oncytokine secretion patterns in the HR PC-3 cell line. Inorder to be able to transiently control IKKe expressionin PC-3 cells, we engineered a PC-3-6TR cell linetransformed with a pcDNA-6TR vector which stablyexpresses the Tet repressor (TetR). Then, we designedand cloned a shRNA targeting IKKe and obtained threePC-3-6TR-shIKKe clones with different efficiencies oftetracycline-inducible IKKe knock-down (Fig. 3A).
Using our PC-3-6TR-shIKKe clones, we followed thealteration of cytokine secretion in the absence or presenceof IKKe as monitored by ELISAs (Fig. 3B). RANTES or IP-10 secretion by PC-3 cells was not detected as expectedbased on results obtained using the human cytokine array(data not shown). We found no effect of IKKe knock-down on IL-8, IL-1a or MIF secretion, three cytokinesconstitutively secreted by PC-3 cells. In contrast, weobserved a 50% decrease (P¼ 0.0005, t-test) in IL-6 secretion after transient and complete knock-down ofIKKe expression in PC-3-6TR-shIKKe clone A (Fig. 3B)while a similar but not statistically significant resultswas seen for clone B (P¼ 0.07, t-test). Partial IKKe knock-down in clone C resulted in a 20% decrease ofIL-6 secretion (P¼ 0.01, t-test).
IKKeOver-ExpressionDoesNot InducePhosphorylation andNuclearTranslocation
of IRF-3 inHSPCaCells
The IRF-3-responsive promoters of IFN-b IP-10 andRANTES are stimulated in response to IKKe-TBK1heterodimer formation in MEFs [47] and IKKe expres-sion is sufficient to induce phosphorylation and nucleartranslocation of IRF-3 in B cell lines [45]. We thereforeanalyzed IRF-3 nuclear status in HS LNCaP and 22Rv1cell lines transiently transfected with the pUNO-hIKKevector (Fig. 4). IKKe over-expression in LNCaP and22Rv1 cells did not induce IRF-3 phosphorylation andnuclear accumulation compared to LNCaP cells treatedwith dsRNA mimetic polyinosinic/polycytidylic (pI/pC) acid (Fig. 4A). Moreover, IKKe over-expression didnot significantly increase IFN-b secretion in LNCaP and22Rv1 cells compared to the same cells treated with pI/pC (Fig. 4B,C).
Cytokine Secretion Inducedby IKKeOver-Expression IsNF-kB-Independent
To determine if IKKe over-expression and nucleartranslocation could control cytokine expression/secretion via activation of the NF-kB pathway, wemonitored the nuclear translocation of four NF-kBsubunits after transient transfection of pUNO orpUNO-hIKKe in LNCaP and 22Rv1 cells (Fig. 5A).TNF-a treatment induced IKKe expression in LNCaPcells. This expression could only be detected byWestern blot after extended film exposure [44]. TNF-atreatment resulted in a decrease in p65/Rel-A, p50 andp52 cytosolic levels after 30 min. This treatment alsoinduced the complete degradation of the NF-kBinhibitor IkB-a and a decrease in p105 cytosolic levels,the inactive form of p50 (Fig. 5A). We also observed anincrease in p100/p52 cytosolic levels and p100 nuclearlevels after 8 hr of TNF-a stimulation. In comparison,IkB-a p100, and p105 cytosolic levels were not affectedby IKKe over-expression. Moreover, IKKe over-expres-sion did not induce either the nuclear translocation ofp65/Rel-A or p50, nor p65/RelA phosphorylationcompared to TNF-a stimulation of LNCaP cells(Fig. 5B). No change in p100/p52 or c-Rel nuclearlevels were observed when IKKe was over-expressed.Finally, using a 3kB-conA-luciferase plasmid co-trans-fected with pUNO or pUNO-hIKKe vectors in LNCaPand 22Rv1 cells, we measured the effect of IKKe over-expression on the activation of a NF-kB-dependentpromoter (Fig. 5C). As expected, TNF-a induced NF-kBactivity as measured by luciferase assay in both LNCaPand 22Rv1 cells, at 11- and 18-fold respectively. Bycontrast, IKKe over-expression failed to increase theNF-kB activity.
The Prostate
710 Pe¤ ant et al.

The Prostate
Fig. 2. Characterizationofcytokine secretioninducedby IKKeover-expressionin22Rv1andLNCaPcells.Cytokine secretionwasevaluatedusingELISAswhichwereperformedoncellculturesupernatants transiently transfectedfor40hr.IL-6,IL-8,IP-10,MIF,andRANTESsecretionsweremeasuredbyELISA from supernatants of 22Rv1-pUNO(graybox) and22Rv1-hIKKe (blackbox) cell cultures (A) orLNCaP-pUNO(graybox) and LNCaP-hIKKe (black box) cell cultures (B). Sample concentrationwas calculatedusing standard curves. Eachmeasurewas done intriplicate and each experiment was repeated three times. Error bars represent the standard error of the mean. *P� 0.05 (Paired t-test).***P� 0.001 (Pairedt-test).
IKKeExpression InducesCytokine Secretion 711

IKKeOver-Expression Is CorrelatedWith itsNuclearTranslocation
Recently, three members of the IkB-kinase family(IKKa, IKKb, and IKKg) were found to partially localizeto the nucleus in human neutrophils [53]. Nucleartranslocation of IKKe had not previously beenreported. In looking at nuclear and cytoplasmic extractsof HR PC-3 cells, we observed substantial IKKe
expression in the nuclear fractions (Fig. 6A). Moreover,nuclear localization of IKKe appears to be independentfrom TBK1 (Fig. 6A,B). TNF-a stimulation of HS LNCaPand 22Rv1 cells failed to induce nuclear translocationof IKKe (data not shown). On the other hand, whenwe forced IKKe over-expression in these cells bytransient transfection with the pUNO-hIKKe vector,we observed a TBK1-independent nuclear accumula-tion of IKKe (Fig. 6B).
The Prostate
Fig. 3. Effect of IKKe knockdown on cytokine secretion in PC-3 cells.Characterization of IKKeexpression in three different PC-3-6TR-shIKKe clones (A).Cellsweregrownin thepresence or absence of1mg/ml tetracycline for 40 hr.Whole cell extracts fromparental PC-3-6TRcell line or stable clones derived from the pENTR/H1/TO-shIKKe transfectionwere separatedby SDS^PAGE, transferred onto PVDFmem-branes andprobedwith anti-IKKe antibody.Equal loadingwas testedwith an anti-b-actin antibody.Theinfluence of tetracycline treatmentonIL-6,IL-8,IL-1a,andMIFsecretionsintheparentalPC-3-6TRcelllineandinthestableinducibleshIKKecloneswas tested(B).CytokinesecretionwasmeasuredusingELISAperformedon supernatants following40hrofculturewith (blackbox) andwithout (graybox) tetracycline. Sampleconcentrationwascalculatedusingstandardcurves.Cytokinesecretionwasadjustedfor1mlofcellculturesurpernatantandfor1mgof totalcellproteins.Eachmeasurewasdonein triplicateandeachexperimentwasrepeated three times.Errorbars show the standarderrorof themean.*P� 0.05(Pairedt-test). **P� 0.01 (Pairedt-test).
712 Pe¤ ant et al.

DISCUSSION
In the present study, we used a Human CytokineArrayTM to determine that IKKe over-expression in HSPCa cell lines correlates with increased secretion ofseveral cytokines. Secretion of IP-10, MIF, and RANTESwas easily detected (Fig. 2) and confirmed by ELISA. IP-10 and RANTES secretion was induced during IKKeover-expression in 22Rv1 cells (590 and 510 pg/mlrespectively). In LNCaP cells, transient transfection ofpUNO-hIKKe increased RANTES secretion (1.5-fold)but did not change IP-10 secretion, which is constitutive(320 pg/ml). However, this observation contradicts arecent study which failed to detect any IP-10 secretionin unstimulated LNCaP cells [54]. Very weak signalsfrom some cytokines (such as IL-1a, IL-2, IL-18, andSDF-1) detected by the Human Cytokine ArrayTM werenot confirmed by ELISA (data not shown). NotablyPAI-1 expression could not be validated due to theabsence of a sensitive commercial ELISA kit. Thesecytokines are well characterized and some of them areassociated with inflammatory diseases. Elevated MIF isobserved in rheumatoid arthritis [55], PAI-1 is detected
and involved in the pathogenesis of sepsis whichresults from a systemic inflammatory response [56,57]while IP-10 is implicated in artherosclerosis develop-ment [58]. High secretion of IL-1a, PAI-1 and MIF wasalso detected in HR PC-3 cells but did not appear to bemodulated by IKKe levels in this cell line (Fig. 4B).
We also observed an effect of IKKe over-expressionon IL-6 (130 and 22 pg/ml respectively) and IL-8 (170and 175 pg/ml respectively) secretion in 22Rv1 andLNCaP cells. These cytokines are not produced by HSPCa cell lines but were secreted by HR PCa PC-3(Fig. 2A) and DU145 cells [17,59]. Both are associatedwith the progression of prostate cancer and poorprognosis [23,60,61]. IKKe expression has already beenassociated with IL-6 expression in mice, but the resultsare contradictory. An initial study showed that IKKe-deficiency results in a reduction of LPS-mediated IL-6gene expression in MEFs [50]. To our knowledge, ourstudy is the first to show an increase in IL-6 andIL-8 secretion in response to IKKe over-expression inHS PCa cells. Moreover, in PC-3 cells which constitu-tively over-express IKKe and produce IL-6, a decreasein IKKe expression correlated with a decrease in
The Prostate
Fig. 4. IKKeover-expressionandIRF-3activity.Cellsweregrownto80%confluencyandtransfectedwithpUNO-hIKKeorpUNOvectorasacontrol (A).Fortyhourslater,cellswerecollectedandnuclearprotein fractionswereextracted.ProteinsweremigratedbySDS^PAGE,trans-ferredontoPVDFmembranesandprobedwithanti-IRF-3antibodies.Nuclearextracts fromLNCaPcells following4or24hr transfectionwithpI/pC (10mg/ml)wereusedaspositive controls for IRF-3phosphorylation andnuclear translocation.Equal loadingwasverifiedusing ananti-b-actin antibody.To confirm IRF-3 activation, IFN-b secretionwasmeasuredbyELISA from supernatants of LNCaP (B) and 22Rv1 (C) cells tran-siently transfectedwithpUNO(40hrstimulation,lightgraybox),pUNO-hIKKe (40hrstimulation,graybox)andpI/pC(24hr stimulation,blackbox) cell cultures. Sample concentrationwas calculatedusing standard curves.Eachmeasurewas done in duplicate andeach experimentwasrepeatedfour times.Errorbars show thestandarderrorof themean.**P� 0.01 (t-test).
IKKeExpression Induces Cytokine Secretion 713

IL-6 secretion but did not modulate IL-8 secretion(Fig. 4). Near complete knock-down of IKKe did notresult in complete loss of IL-6 secretion in PC-3 cells aspreviously observed in IKKe �/� MEFs [62]. Theseresults suggest that IKKe expression is not necessary forbasal IL-6 secretion but that IKKe over-expression canparticipate in increased IL-6 secretion.
Altogether, these observations suggest that IKKeover-expression promotes the secretion of severalinflammatory cytokines in a cell line specific manner.Given this data, we then focused on the molecularpathways activated by IKKe over-expression. RANTESand IP-10 genes are under the control of IRF-3 and IRF-7[45,46,62–64] and it is known that IKKe expression is
The Prostate
Fig. 5. Effectof IKKeover-expressiononNF-kBactivity.Cellsweregrown to80%confluencyandtransfectedwithpUNO-hIKKeorpUNOvector as a control (A). Forty hours later, cells were collected and cytosolic/nuclear protein fractions were extracted.Nuclear (10 mg) andcytosolic(20mg)proteinswereseparatedby7.5%SDS^PAGE,transferredontoPVDFmembranesandsuccessivelyprobed(afterstripping)withanti-p105/p50, anti-p100/p52, anti-TBK-1, anti-IKKe, anti-c-Rel, anti-p65, anti-phospho-p65, and anti-Ik B-a antobodies. Cytosolic/nuclearextracts fromLNCaPcells following30minor8hr stimulationwithTNF-a (10ng/ml)wereusedaspositive control forNF-kB activation.Equalloadingwasverifiedusingananti-b-actinantibodyandcytosoliccontaminationwasmeasuredwithananti-a-tubulinantibody.NF-kBtranscrip-tional activity wasmeasuredby luciferase assay (B).Cells were grown to 80% confluency and co-transfectedwith pCMV-Renilla, 3kB-conA-FireflyandpUNOorpUNO-hIKKe.TransfectionsusingconA-FireflyandpUNOweredoneas controls. Sixteenhours later, cellswereassayedfor luciferase activity.Transfection efficiency was normalized to thatofRenilla luciferase andcompared to conA-Firefly and 3kB-conA-Fireflytransfectedcells treatedduring8hrwithTNF-a (10ng/ml).Errorbars show thestandarderrorof themean.*P� 0.05 (t-test). **P� 0.01 (t-test).
714 Pe¤ ant et al.

sufficient to induce IRF-3-dependent gene transcrip-tion [45,47,48]. However, PC-3 cells which constitu-tively over-express IKKe, did not secrete IP-10 orRANTES (data not shown). Moreover, IKKe over-expression and IKKe knock-down did not affect IL-1a secretion (Figs. 2 and 4) which is under the control ofTLR3- and TLR4-dependent pathways [65]. Finally,during IKKe over-expression in LNCaP (Fig. 4) and22Rv1 cells (data not shown), we did not detect anydifferences in IRF-3 nuclear translocation and IFN-b secretion used as reporter for IRF-3 activity. Based onthese observations, we concluded that IKKe over-expression does not induce IRF-3 phosphorylationand nuclear translocation in PCa cells.
As IL-6 and IL-8 gene expression is thought to beunder the control of NF-kB [66,67], we tested whetherthe high secretion of these cytokines was due to NF-kBactivation by IKKe. Several studies presented IKKe asan important regulator of NF-kB signaling pathway byinducing phosphorylation and nuclear translocation ofp65 [68] and c-Rel [69,70]. It has also been proposed thatIKKe could have an oncogenic function through theactivation of c-Rel. In particular, IKKe deregulationcorrelates with up regulation of NF-kB-responsivegenes and IkB degradation in breast cancer cell lines[70]. However, we previously found that the p65 NF-kBsubunit did not translocate to the nucleus following anincrease in IKKe expression after TNF-a treatment ofLNCaP cells [44]. Consistent with our previous study,we found that IKKe over-expression in LNCaP and22Rv1 PCa cell lines did not correlate with p65/p50 orc-Rel nuclear translocation or with IkB-a degradation(Fig. 5A). Similarly, IKKe knock-down in PC-3 cells didnot change nuclear p65 levels or IkB-a stability (datanot shown). Moreover, there was no increase intranscription from the kB site within 3kB-conA pro-moter, as well as no nuclear translocation and
phosphorylation of p65 during IKKe over-expressionin LNCaP and 22Rv1 cells (Fig. 5). As we did not detectany change in NF-kB activity, we propose that IKKe-dependent IL-6 and IL-8 deregulation is not due to theactivation of the NF-kB signaling pathway in prostatecancer.
As IRF-3 and NF-kB were not induced during IKKeover-expression in PCa cell lines, we speculated analternative IKKe activity that could be dependent notonly on its expression levels but also on its cellularlocalization. We found that IKKe can accumulate in thenucleus while TBK1 expression is strictly cytosolic inthe HR PC-3 cell line that has a high level of constitutiveIKKe expression. To our knowledge, our study is thefirst report of nuclear IKKe. This phenotype was alsoobserved for the DU145 cell line (data not shown),another HR PCa cell line that constitutively expresseshigh levels of IKKe. Importantly, we observed thatIKKe transient over-expression in HS 22Rv1 andLNCaP cell lines led to nuclear accumulation of IKKe,independent of TBK-1 localization. While TNF-astimulation induces moderate IKKe expression inLNCaP and 22Rv1 cells [44], it does not induce TBK-1or IKKe nuclear translocation (data not shown). Similarobservations were reported in human neutrophilswhere TNF-a stimulation failed to provoke TBK-1 orIKKe nuclear translocation [53]. Based on theseobservations we speculate that IKKe nuclear activityleads to the activation of inflammatory cytokine genes,possibly through the activation of currently unidenti-fied transcription factors. Although, IL-6 gene expres-sion is known to be regulated by several transcriptionfactors (AP-1, NF-kB, NF-IL6, CREB), two of which,NF-kB and NF-IL6 (also known as C/EBP-b), arerequired for strong induction of IL-6 expression [71].NF-kB alone is unable to fully activate IL-6 genetranscription, and requires additional transcription
The Prostate
Fig. 6. IKKe localizationinprostatecancercelllines.Nuclear(10mg)andcytosolic (20mg)proteinswereresolvedbySDS^PAGE,transferredonto PVDF and probed with anti-IKKe and anti-TBK-1 antibodies. Equal loading was verified with an anti-b-actin antibody and cytosoliccontaminationofnuclearextractwas controlledwith ananti-a-tubulin antibody.Westernblot analysis of IKKeexpression innuclear/cytosolicfractions of PCa cell lines (A). IKKe subcellular localization after transient transfectionofpUNOorpUNO-hIKKevector inLNCaP and22Rv1cells (B).
IKKeExpression InducesCytokine Secretion 715

factors to induce complete activation [72,73]. In PC-3cells, a previous study showed that over expression ofNF-IL6 or NF-kB induced IL-6 promoter activity but thestrongest induction occurred with the simultaneousover-expression of both transcription factors. Thisstrong induction was also impaired by the inhibitionof NF-IL6 or NF-kB activity [71]. IL-6 expression in PC-3 cells, with high NF-kB activity and constitutive IKKeexpression, was at least ten times higher than in 22Rv1and LNCaP cells which over-express IKKe but show noactivation of the NF-kB pathway (Figs. 2, 3B, and 5). Aswe observed that IKKe expression is not necessary forbasal IL-6 secretion in PC-3 cells and IKKe does notactivate the NF-kB pathway, it is tempting to speculatethat IKKe nuclear translocation leads to the activationof the nuclear transcription factor NF-IL6 to increase IL-6 promoter activity. Interestingly, the IL-8 promoter isalso regulated by the same pool of transcription factorsin breast cancer cells [74]. More detailed studies areneeded to address this hypothesis.
CONCLUSION
We propose that IKKe over-expression leads to animbalance between pro- and anti-inflammatory cyto-kines which can contribute to the initiation andprogression of prostate cancer in two ways: (1) IKKeover-expression stimulates IL-8 and particularly IL-6 secretion contributing to disease as positive growthfactors in PCa [28], specifically in the context ofandrogen independent progression and (2) it mayfavor chronic inflammation and create a tumor-supporting microenvironment that promotes the neo-plastic process [55,75]. Xenograft mouse models inwhich IKKe expression has been modulated may beuseful to resolve the contribution of this kinase toprostate cancer development. Its contribution to pros-tate cancer initiation would ultimately best be studiedin an appropriate transgenic mouse model.
Finally, we propose that IKKe induced cytokineexpression and secretion may be linked to its nucleartranslocation. Our results suggest that the proposednuclear activity of IKKe does not target IRF-3 or NF-kB.The study of the IKKe-dependent control of pro-inflammatory cytokine secretion is certainly warrantedand will likely further our understanding of PCa and itsprogression towards a hormone-refractory state.
ACKNOWLEDGMENTS
The authors would like to thank the CHUM urologydepartment for their support, the laboratory membersfor helpful discussions and Genevieve St-Onge, JasonMadore and Kate Morris for proof-reading the manu-script. This work was supported by a grant from theProstate Cancer Research Foundation of Canada
awarded to Anne-Marie Mes-Masson. Jean-SimonDiallo is a recipient of Canderel and Marc BourgieFoundation studentships. Fred Saad is the recipient ofthe Universite de Montreal Chair in Prostate CancerResearch.
REFERENCES
1. Landis SH, Murray T, Bolden S, Wingo PA. Cancer statistics,1999. CA Cancer J Clin 1999;49(1):8–31, 31.
2. Coussens LM, Werb Z. Inflammation and cancer. Nature 2002;420(6917):860–867.
3. Mantovani A. Cancer: Inflammation by remote control. Nature2005;435(7043):752–753.
4. Liu RS, Ta D, Payne J, Coccia M, Kuus-Reichel K. Generation ofmurine monoclonal antibodies in serum-free medium. Hybrid-oma 1998;17(1):69–72.
5. Schindler C, Strehlow I. Cytokines and STAT signaling. AdvPharmacol 2000;47:113–174.
6. Aggarwal BB, Shishodia S, Sandur SK, Pandey MK, Sethi G.Inflammation and cancer: How hot is the link? BiochemPharmacol 2006;72(11):1605–1621.
7. Miller LJ, Kurtzman SH, Anderson K, Wang Y, Stankus M,Renna M, Lindquist R, Barrows G, Kreutzer DL. Interleukin-1family expression in human breast cancer: Interleukin-1 receptorantagonist. Cancer Invest 2000;18(4):293–302.
8. Singer CF, Kronsteiner N, Hudelist G, Marton E, Walter I,Kubista M, Czerwenka K, Schreiber M, Seifert M, Kubista E.Interleukin 1 system and sex steroid receptor expression inhuman breast cancer: Interleukin 1alpha protein secretion iscorrelated with malignant phenotype. Clin Cancer Res 2003;9(13):4877–4883.
9. Kurebayashi J. Regulation of interleukin-6 secretion from breastcancer cells and its clinical implications. Breast Cancer 2000;7(2):124–129.
10. Knupfer H, Schmidt R, Stanitz D, Brauckhoff M, Schonfelder M,Preiss R. CYP2C and IL-6 expression in breast cancer. Breast2004;13(1):28–34.
11. Niwa Y, Akamatsu H, Niwa H, Sumi H, Ozaki Y, Abe A.Correlation of tissue and plasma RANTES levels with diseasecourse in patients with breast or cervical cancer. Clin Cancer Res2001;7(2):285–289.
12. Kozlowski L, Zakrzewska I, Tokajuk P, Wojtukiewicz MZ.Concentration of interleukin-6 (IL-6), interleukin-8 (IL-8) andinterleukin-10 (IL-10) in blood serum of breast cancer patients.Rocz Akad Med Bialymst 2003;48:82–84.
13. Benoy IH, Salgado R, Van Dam P, Geboers K, Van Marck E,Scharpe S, Vermeulen PB, Dirix LY. Increased serum interleukin-8 in patients with early and metastatic breast cancer correlateswith early dissemination and survival. Clin Cancer Res 2004;10(21):7157–7162.
14. Mettler L, Salmassi A, Heyer M, Schmutzier A, Schollmeyer T,Jonat W. Perioperative levels of interleukin-1beta and interleu-kin-6 in women with breast cancer. Clin Exp Obstet Gynecol2004;31(1):20–22.
15. Hobisch A, Rogatsch H, Hittmair A, Fuchs D, Bartsch G Jr,Klocker H, Bartsch G, Culig Z. Immunohistochemical local-ization of interleukin-6 and its receptor in benign, premalignantand malignant prostate tissue. J Pathol 2000;191(3):239–244.
16. Hobisch A, Ramoner R, Fuchs D, Godoy-Tundidor S, Bartsch G,Klocker H, Culig Z. Prostate cancer cells (LNCaP) generated
The Prostate
716 Pe¤ ant et al.

after long-term interleukin 6 (IL-6) treatment express IL-6 andacquire an IL-6 partially resistant phenotype. Clin Cancer Res2001;7(9):2941–2948.
17. Chung TD, Yu JJ, Spiotto MT, Bartkowski M, Simons JW.Characterization of the role of IL-6 in the progression of prostatecancer. Prostate 1999;38(3):199–207.
18. Chung TD, Yu JJ, Kong TA, Spiotto MT, Lin JM. Interleukin-6activates phosphatidylinositol-3 kinase, which inhibitsapoptosis in human prostate cancer cell lines. Prostate 2000;42(1):1–7.
19. Uehara H, Troncoso P, Johnston D, Bucana CD, Dinney C, DongZ, Fidler IJ, Pettaway CA. Expression of interleukin-8 gene inradical prostatectomy specimens is associated with advancedpathologic stage. Prostate 2005;64(1):40–49.
20. Murphy C, McGurk M, Pettigrew J, Santinelli A, Mazzucchelli R,Johnston PG, Montironi R, Waugh DJ. Nonapical and cytoplas-mic expression of interleukin-8, CXCR1, and CXCR2 correlateswith cell proliferation and microvessel density in prostatecancer. Clin Cancer Res 2005;11(11):4117–4127.
21. Shariat SF, Andrews B, Kattan MW, Kim J, Wheeler TM, SlawinKM. Plasma levels of interleukin-6 and its soluble receptor areassociated with prostate cancer progression and metastasis.Urology 2001;58(6):1008–1015.
22. Smith PC, Hobisch A, Lin DL, Culig Z, Keller ET. Interleukin-6and prostate cancer progression. Cytokine Growth Factor Rev2001;12(1):33–40.
23. Michalaki V, Syrigos K, Charles P, Waxman J. Serum levels of IL-6 and TNF-alpha correlate with clinicopathological features andpatient survival in patients with prostate cancer. Br J Cancer2004;91(6):1227.
24. Chen T, Wang LH, Farrar WL. Interleukin 6 activates androgenreceptor-mediated gene expression through a signal transducerand activator of transcription 3-dependent pathway in LNCaPprostate cancer cells. Cancer Res 2000;60(8):2132–2135.
25. Lin DL, Whitney MC, Yao Z, Keller ET. Interleukin-6 inducesandrogen responsiveness in prostate cancer cells through up-regulation of androgen receptor expression. Clin Cancer Res2001;7(6):1773–1781.
26. Ueda T, Bruchovsky N, Sadar MD. Activation of the androgenreceptor N-terminal domain by interleukin-6 via MAPK andSTAT3 signal transduction pathways. J Biol Chem 2002;277(9):7076–7085.
27. Ueda T, Mawji NR, Bruchovsky N, Sadar MD. Ligand-independent activation of the androgen receptor by interleu-kin-6 and the role of steroid receptor coactivator-1 in prostatecancer cells. J Biol Chem 2002;277(41):38087–38094.
28. Culig Z, Steiner H, Bartsch G, Hobisch A. Interleukin-6regulation of prostate cancer cell growth. J Cell Biochem 2005;95(3):497–505.
29. Drachenberg DE, Elgamal AA, Rowbotham R, Peterson M,Murphy GP. Circulating levels of interleukin-6 in patients withhormone refractory prostate cancer. Prostate 1999;41(2):127–133.
30. Giri D, Ozen M, Ittmann M. Interleukin-6 is an autocrine growthfactor in human prostate cancer. Am J Pathol 2001;159(6):2159–2165.
31. Wise GJ, Marella VK, Talluri G, Shirazian D. Cytokine variationsin patients with hormone treated prostate cancer. J Urol2000;164(3 Pt 1):722–725.
32. Barnes PJ, Karin M. Nuclear factor-kappaB: A pivotal tran-scription factor in chronic inflammatory diseases. N Engl J Med1997;336(15):1066–1071.
33. Le Page C, Koumakpayi IH, Lessard L, Mes-Masson AM, Saad F.EGFR and Her-2 regulate the constitutive activation of NF-kappaB in PC-3 prostate cancer cells. Prostate 2005;65(2):130–140.
34. Chen CD, Sawyers CL. NF-kappa B activates prostate-specificantigen expression and is upregulated in androgen-independentprostate cancer. Mol Cell Biol 2002;22(8):2862–2870.
35. Suh J, Rabson AB. NF-kappaB activation in human prostatecancer: Important mediator or epiphenomenon? J Cell Biochem2004;91(1):100–117.
36. Gasparian AV, Yao YJ, Kowalczyk D, Lyakh LA, Karseladze A,Slaga TJ, Budunova IV. The role of IKK in constitutive activationof NF-kappaB transcription factor in prostate carcinoma cells.J Cell Sci 2002;115(Pt 1):141–151.
37. Ayala GE, Dai H, Ittmann M, Li R, Powell M, Frolov A, WheelerTM, Thompson TC, Rowley D. Growth and survival mecha-nisms associated with perineural invasion in prostate cancer.Cancer Res 2004;64(17):6082–6090.
38. Ross JS, Kallakury BV, Sheehan CE, Fisher HA, Kaufman RP Jr,Kaur P, Gray K, Stringer B. Expression of nuclear factor-kappa Band I kappa B alpha proteins in prostatic adenocarcinomas:Correlation of nuclear factor-kappa B immunoreactivity withdisease recurrence. Clin Cancer Res 2004;10(7):2466–2472.
39. Sweeney C, Li L, Shanmugam R, Bhat-Nakshatri P, JayaprakasanV, Baldridge LA, Gardner T, Smith M, Nakshatri H, Cheng L.Nuclear factor-kappaB is constitutively activated in prostatecancer in vitro and is overexpressed in prostatic intraepithelialneoplasia and adenocarcinoma of the prostate. Clin Cancer Res2004;10(16):5501–5507.
40. Shukla S, MacLennan GT, Fu P, Patel J, Marengo SR, Resnick MI,Gupta S. Nuclear factor-kappaB/p65 (Rel A) is constitutivelyactivated in human prostate adenocarcinoma and correlateswith disease progression. Neoplasia 2004;6(4):390–400.
41. Lessard L, Mes-Masson AM, Lamarre L, Wall L, Lattouf JB, SaadF. NF-kappa B nuclear localization and its prognostic signifi-cance in prostate cancer. BJU Int 2003;91(4):417–420.
42. Lessard L, Karakiewicz PI, Bellon-Gagnon P, Alam-Fahmy M,Ismail HA, Mes-Masson AM, Saad F. Nuclear localization ofnuclear factor-kappaB p65 in primary prostate tumors is highlypredictive of pelvic lymph node metastases. Clin Cancer Res2006;12(19):5741–5745.
43. Fradet V, Lessard L, Begin LR, Karakiewicz P, Masson AM, SaadF. Nuclear factor-kappaB nuclear localization is predictive ofbiochemical recurrence in patients with positive margin prostatecancer. Clin Cancer Res 2004;10(24):8460–8464.
44. Peant B, Diallo JS, Lessard L, Delvoye N, Le Page C, Saad F, Mes-Masson AM. Regulation of IkappaB kinase epsilon expression bythe androgen receptor and the nuclear factor-kappaB tran-scription factor in prostate cancer. Mol Cancer Res 2007;5(1):87–94.
45. Sharma S, tenOever BR, Grandvaux N, Zhou GP, Lin R, Hiscott J.Triggering the interferon antiviral response through an IKK-related pathway. Science 2003;300(5622):1148–1151.
46. Fitzgerald KA, McWhirter SM, Faia KL, Rowe DC, Latz E,Golenbock DT, Coyle AJ, Liao SM, Maniatis T. IKKepsilon andTBK1 are essential components of the IRF3 signaling pathway.Nat Immunol 2003;4(5):491–496.
47. McWhirter SM, Fitzgerald KA, Rosains J, Rowe DC, GolenbockDT, Maniatis T. IFN-regulatory factor 3-dependent geneexpression is defective in Tbk1-deficient mouse embryonicfibroblasts. Proc Natl Acad Sci USA 2004;101(1):233–238.
48. Hiscott J, Grandvaux N, Sharma S, Tenoever BR, Servant MJ, LinR. Convergence of the NF-kappaB and interferon signaling
The Prostate
IKKeExpression InducesCytokine Secretion 717

pathways in the regulation of antiviral defense and apoptosis.Ann NY Acad Sci 2003;1010:237–248.
49. Aupperle KR, Yamanishi Y, Bennett BL, Mercurio F, Boyle DL,Firestein GS. Expression and regulation of inducible IkappaBkinase (IKK-i) in human fibroblast-like synoviocytes. CellImmunol 2001;214(1):54–59.
50. Kravchenko VV, Mathison JC, Schwamborn K, Mercurio F,Ulevitch RJ. IKKi/IKKepsilon plays a key role in integratingsignals induced by pro-inflammatory stimuli. J Biol Chem 2003;278(29):26612–26619.
51. Lou W, Ni Z, Dyer K, Tweardy DJ, Gao AC. Interleukin-6 inducesprostate cancer cell growth accompanied by activation of stat3signaling pathway. Prostate 2000;42(3):239–242.
52. Konig JE, Senge T, Allhoff EP, Konig W. Analysis of theinflammatory network in benign prostate hyperplasia andprostate cancer. Prostate 2004;58(2):121–129.
53. Ear T, Cloutier A, McDonald PP. Constitutive nuclear expressionof the I kappa B kinase complex and its activation in humanneutrophils. J Immunol 2005;175(3):1834–1842.
54. Nagpal ML, Davis J, Lin T. Overexpression of CXCL10 in humanprostate LNCaP cells activates its receptor (CXCR3) expressionand inhibits cell proliferation. Biochim Biophys Acta 2006;1762(9):811–818.
55. McInnes IB, Schett G. Cytokines in the pathogenesis ofrheumatoid arthritis. Nat Rev Immunol 2007;7(6):429–442.
56. Mesters RM, Florke N, Ostermann H, Kienast J. Increase ofplasminogen activator inhibitor levels predicts outcome ofleukocytopenic patients with sepsis. Thromb Haemost 1996;75(6):902–907.
57. Menges T, Hermans PW, Little SG, Langefeld T, Boning O, EngelJ, Sluijter M, de Groot R, Hempelmann G. Plasminogen-activator-inhibitor-1 4G/5G promoter polymorphism and prog-nosis of severely injured patients. Lancet 2001;357(9262):1096–1097.
58. Braunersreuther V, Mach F, Steffens S. The specific role ofchemokines in atherosclerosis. Thromb Haemost 2007;97(5):714–721.
59. Kooijman R, Himpe E, Potikanond S, Coppens A. Regulation ofinterleukin-8 expression in human prostate cancer cells byinsulin-like growth factor-I and inflammatory cytokines.Growth Horm IGF Res 2007;17(5):383–391.
60. Ferrer FA, Miller LJ, Andrawis RI, Kurtzman SH, Albertsen PC,Laudone VP, Kreutzer DL. Angiogenesis and prostate cancer: Invivo and in vitro expression of angiogenesis factors by prostatecancer cells. Urology 1998;51(1):161–167.
61. Inoue K, Slaton JW, Eve BY, Kim SJ, Perrotte P, Balbay MD, YanoS, Bar-Eli M, Radinsky R, Pettaway CA, Dinney CP. Interleukin 8expression regulates tumorigenicity and metastases in andro-gen-independent prostate cancer. Clin Cancer Res 2000;6(5):2104–2119.
62. Hemmi H, Takeuchi O, Sato S, Yamamoto M, Kaisho T, Sanjo H,Kawai T, Hoshino K, Takeda K, Akira S. The roles of two IkappaBkinase-related kinases in lipopolysaccharide and doublestranded RNA signaling and viral infection. J Exp Med 2004;199(12):1641–1650.
63. Sato M, Tanaka N, Hata N, Oda E, Taniguchi T. Involvement ofthe IRF family transcription factor IRF-3 in virus-inducedactivation of the IFN-beta gene. FEBS Lett 1998;425(1):112–116.
64. Lin R, Heylbroeck C, Genin P, Pitha PM, Hiscott J. Essential roleof interferon regulatory factor 3 in direct activation of RANTESchemokine transcription. Mol Cell Biol 1999;19(2):959–966.
65. Krasowska-Zoladek A, Banaszewska M, Kraszpulski M, KonatGW. Kinetics of inflammatory response of astrocytes induced byTLR 3 and TLR4 ligation. J Neurosci Res 2007;85(1):205–212.
66. Sanceau J, Kaisho T, Hirano T, Wietzerbin J. Triggering of thehuman interleukin-6 gene by interferon-gamma and tumornecrosis factor-alpha in monocytic cells involves cooperationbetween interferon regulatory factor-1, NF kappa B, and Sp1transcription factors. J Biol Chem 1995;270(46):27920–27931.
67. Wang W, Ye L, Li B, Gao B, Zeng Y, Kong L, Fang X, Zheng H, WuZ, She Y. Up-regulation of IL-6 and TNF-alpha induced by SARS-coronavirus spike protein in murine macrophages via NF-kappaB pathway. Virus Res 2007;128(1–2):1–8.
68. Adli M, Baldwin AS. IKK-i/IKKepsilon controls constitutive,cancer cell-associated NF-kappaB activity via regulation of Ser-536 p65/RelA phosphorylation. J Biol Chem 2006;281(37):26976–26984.
69. Harris J, Oliere S, Sharma S, Sun Q, Lin R, Hiscott J, GrandvauxN. Nuclear accumulation of cRel following C-terminal phos-phorylation by TBK1/IKK epsilon. J Immunol 2006;177(4):2527–2535.
70. Boehm JS, Zhao JJ, Yao J, Kim SY, Firestein R, Dunn IF, SjostromSK, Garraway LA, Weremowicz S, Richardson AL, Greulich H,Stewart CJ, Mulvey LA, Shen RR, Ambrogio L, Hirozane-Kishikawa T, Hill DE, Vidal M, Meyerson M, Grenier JK, HinkleG, Root DE, Roberts TM, Lander ES, Polyak K, Hahn WC.Integrative genomic approaches identify IKBKE as a breastcancer oncogene. Cell 2007;129(6):1065–1079.
71. Xiao W, Hodge DR, Wang L, Yang X, Zhang X, Farrar WL. Co-operative functions between nuclear factors NFkappaB andCCAT/enhancer-binding protein-beta (C/EBP-beta) regulatethe IL-6 promoter in autocrine human prostate cancer cells.Prostate 2004;61(4):354–370.
72. Asschert JG, De Vries EG, De Jong S, Withoff S, Vellenga E.Differential regulation of IL-6 promoter activity in a humanovarian-tumor cell line transfected with various p53 mutants:Involvement of AP-1. Int J Cancer 1999;81(2):236–242.
73. Steiner GE, Newman ME, Paikl D, Stix U, Memaran-Dagda N,Lee C, Marberger MJ. Expression and function of pro-inflam-matory interleukin IL-17 and IL-17 receptor in normal, benignhyperplastic, and malignant prostate. Prostate 2003;56(3):171–182.
74. Freund A, Jolivel V, Durand S, Kersual N, Chalbos D, Chavey C,Vignon F, Lazennec G. Mechanisms underlying differentialexpression of interleukin-8 in breast cancer cells. Oncogene2004;23(36):6105–6114.
75. Lin WW, Karin M. A cytokine-mediated link between innateimmunity, inflammation, and cancer. J Clin Invest 2007;117(5):1175–1183.
The Prostate
718 Pe¤ ant et al.













![evidence for AGN feedback in galaxies with CIII]-λ1908 Ε](https://img.pdfslide.net/doc/110x75/63249bfcbe5419ea700f1fc4/evidence-for-agn-feedback-in-galaxies-with-ciii-l1908-e-.jpg)















