Embed Size (px)
Citation preview

Life Sciences 77 (2005) 1993–2003
www.elsevier.com/locate/lifescie
pH effects on high conductance Ca2+-activated K+ channels (BKCa)
in human internal mammary artery smooth muscle cells
Jesica Raingoa, Alejandro Rebolledoa, Angela O. Grassi de Gendea, Nora Sanzb,
Juan Tommasib, Veronica Milesia,TaCatedra de Fisiologıa, Departamento de Ciencias Biologicas, Facultad de Ciencias Exactas,
Universidad Nacional de La Plata (UNLP). Calles 47 y 115, La Plata (1900), ArgentinabHospital San Juan de Dios, La Plata, Argentina
Received 4 January 2005; accepted 14 March 2005
Abstract
Introduction: In vascular smooth muscle cells, different types of K+ channels participate in the regulation of
membrane potential and consequently in the contractile behavior of the vessel. There is little information
about the properties and role of K+ channels in human internal mammary artery (HIMA), the vessel of
choice for coronary revascularization.
Methods: Patch-clamp technique on isolated HIMA smooth muscle cells was used.
Results: This work presents for the first time single-channel properties of the high conductance Ca2+-
activated K+ channel (BKCa) of HIMA. It presents a single-channel conductance of 228F4 pS (n =44,
8 cells), is sensitive to 100 nM iberiotoxin, and its open probability is Ca2+- and voltage-dependent. Inside-
out results show that BKCa channels in HIMA are directly activated by increasing the pH of intracellular
media (NPo=0.096F0.032 at pH 7.4 and NPo=0.459F0.111 at pH 7.6, n =12 cells, pb0.05) and inhibited
by lowering this pH (NPo=0.175F0.067 at pH 7.4 and NPo=0.051F0.019 at pH 6.8, n =13 cells,
p b0.05).
Conclusions: The evidences presented about single-channel properties and intracellular pH sensitivity of BKCa
from HIMA smooth muscle cells provide useful information to elucidate physiological or pathological
0024-3205/$ -
doi:10.1016/j.l
T Correspond
E-mail add
see front matter D 2005 Elsevier Inc. All rights reserved.
fs.2005.03.015
ing author. Tel.: +54 221 425 0497x30; fax: +54 221 489 0354.
ress: [email protected] (V. Milesi).

J. Raingo et al. / Life Sciences 77 (2005) 1993–20031994
mechanisms in this vessel, as well as for future studies where drugs could have BKCa channels as targets for
pharmacological therapies.
D 2005 Elsevier Inc. All rights reserved.
Keywords: BKCa channels; Human mammary artery; Patch-clamp; pH
Introduction
The human internal mammary artery (HIMA) is the vessel of choice for coronary artery bypass
grafting. In the last decade, the HIMA represented a superior long-term survival vessel compared to the
traditionally used saphenous vein bypass graft (Loop et al., 1986). HIMA spasms are frequent during
surgical manipulation, and were also reported to occur in the postoperative period (Green, 1979; Dincer
and Borman, 1983). Knowledge of regulatory mechanisms of the contractile state of this vessel is
important to understand the development of vasospasm. In HIMA, the production of endothelium-
derived factors and the reactivity of smooth muscle to them and other vasoactive substances has been
extensively studied in organ bath experiments (Verma et al., 2001; Cracowski et al., 1999, 2000; Pearson
et al., 1998). In order to avoid the vasospasms, several pharmacological strategies are used to induce
vasorelaxation and lower reactivity of the vessel (Rosenfeldt et al., 1999). Among these pharmacological
tools, the ATP-sensitive potassium channel openers (i.e., aprikalim or pinacidil) have been demonstrated
to be vasorelaxing drugs in HIMA (He and Yang, 1997; Gojkoviic Bukarica et al., 1997; Liu et al., 2001)
suggesting a possible role for this kind of channel in the regulation of contractile state of this vessel.
However, more than one type of potassium channels participate in the regulation of membrane
potential in vascular smooth muscle cells and consequently in the contractile behavior of blood vessels.
In particular, the high conductance calcium-activated K+ channel (BKCa) has been described as playing a
role in the regulation of membrane potential of human vascular smooth muscle cells (Zhang et al., 1996;
Milesi et al., 1999; Gokina et al., 1996), as well as being a target for endogenous and exogenous
vasoactive agonists (Bychkov et al., 1998; Gruhn et al., 2002). Their main electrophysiological
properties have been described in different tissues (Kaczorowski and Garcia, 1999), but these properties
and regulatory mechanisms may vary depending on the specific tissue considered. Description of these
channels at the single-channel level is sparse in human vascular smooth muscle, and absent in the case of
the HIMA.
Among the many different modulating factors of BKCa channels behavior (Dimitropoulou et al., 2001;
Thomson et al., 1998; Carvajal et al., 2000; Lang et al., 2000), changes in intracellular pH have been
shown to modify the channel activity. Inhibition of BKCa channels induced by intracellular acidification
has been reported in vascular smooth muscle cells (Schubert et al., 2001; Petrova-Kirova et al., 2000),
and a direct interaction between protons and constitutive or regulatory channel protein subunits could be
a mechanism responsible for these effects (Austin and Wray, 2000). Intracellular pH changes are
involved in the physiological responses to shear-stress (Stamatas et al., 1997), vasoactive agonists such
as angiotensin II (Hatori et al., 1987) and endothelin (Hubel and Highsmith, 1995), and pathological
states, such as cellular ischemia, which modify the contractile state of vascular smooth muscle (Smith,
G.L. et al., 1998).
In this study, we present for the first time single-channel properties of BKCa channels in freshly
dispersed human smooth muscle cells from internal mammary artery and their main electrophysiological

J. Raingo et al. / Life Sciences 77 (2005) 1993–2003 1995
properties. We show that these channels are directly regulated by internal pH and discuss its possible
implications on the mechanical response of HIMA to pH.
Materials and methods
Sample procurement and preparation
The vessels used in this study were obtained from 57 patients undergoing primary coronary artery
bypass surgery at the Service of Cardiothoracic Surgery of the Hospital San Juan de Dios, Ministerio de
Salud de la Provincia de Buenos Aires, La Plata, Argentina. The segments of HIMA used for this study
were classified as surgical discard specimens and thus exempted from patient consent requirements.
Additionally, the Scientific Research Commission of our institution reviewed and approved all the
experimental protocols involved in this study.
After the bypass surgery was finished, the remnant segments were transported to our laboratory in ice-
cold 0.9% NaCl solution and were studied within 3 h of arrival. The vessels were placed in a Petri dish
filled with 0.9% NaCl solution, cleaned of adherent connective tissue, and cut into small pieces for
enzymatic cell isolation.
Cell isolation procedure
Cell isolation procedure was based on the method described by Klockner (Klockner, 1993) and
currently used in our laboratory to obtain isolated smooth muscle cells from human saphenous vein
(Milesi et al., 1999) or, with slight modifications, to obtain cells from human umbilical artery (Milesi et
al., 2003). Briefly, small strips were placed in a dissociation medium (DM) containing: 140 mM NaCl, 5
mM KH2PO4, 5 mM MgCl2, 20 mM glucose, 5 mM HEPES, 2 mM dithiothreitol, 2 mg ml�1
collagenase, 2 mg ml�1 papain and 0.5 mg ml�1 protease type-I; pH 7.4 at 30 8C, and mechanically
stirred for 30 min. After the incubation period, the strips were washed with DM without enzymes and
single HIMA smooth muscle cells were obtained by gentle dispersion of the treated tissue using a
Pasteur pipette. The remaining tissue and the supernatant containing isolated cells were stored at room
temperature until used.
Patch-clamp recording
HIMA smooth muscle cells were allowed to settle onto the coverglass bottom of a 0.5 ml
experimental chamber. The cells were observed with a mechanically stabilized, inverted microscope
(Zeiss-Telaval3, Jena, Germany) equipped with a 40� objective lens. The chamber was perfused for
15 min with bath saline solution (BSS, see composition later) at 0.5 ml min�1 (by gravity) before
the patch-clamp experiment was started. Application of test solutions was performed through a
multibarreled pipette positioned close to the cell investigated. After each experiment on a single
cell, the experimental chamber was replaced by another one containing a new sample of cells. Only
well-relaxed, spindle-shaped smooth muscle cells were used for electrophysiological recording. Data
were collected within 4–6 h after cell isolation. All experiments were performed at room
temperature.

J. Raingo et al. / Life Sciences 77 (2005) 1993–20031996
The standard tight-seal cell-attached and inside-out configurations of the patch-clamp technique
(Hamill et al., 1981) were used. Glass pipettes were drawn from WPI PG52165-4 glass on a two-
stage vertical micropipette puller (PP-83, Narishige Scientific Instrument Laboratories, Tokyo, Japan)
and coated with Sylgard (Dow Corning Corporation, Midland, MI, USA) to reduce recording noise.
Pipette resistances ranged from 2 to 4 MV measured in BSS. Ionic currents were measured with an
Axopatch 200A amplifier (Axon Instruments, Foster City, CA, USA). Single channel currents were
filtered at 2 kHz and digitized at 16 kHz. Voltage-clamp 30–60 s recordings were obtained at
different membrane potential values. Open probability (Po) is expressed as NPo, where N is the
number of channels present in each patch. NPo values were calculated using the following expression
(Kajioka et al., 1991):
NPo ¼ A1 þ 2A2 þ 3A3 þ N þ nAnÞ= Ao þ A1 þ A2 þ N þ AnÞððwhere A0 is the area under the curve of the amplitude histogram which corresponds to the closed
state, and A1, A2. . . An are the areas corresponding to the n open-state levels of current. The
histogram parameters were obtained from least-squares Gaussian fits of experimental data.
The bath saline solution (BSS) used for single-channel experiments was (in mM): 140 KCl, 0.5
MgCl2, 1 EGTA, 10 HEPES, 10 glucose, pH was adjusted to 7.4 with NaOH. Free Ca2+
concentration in each solution was estimated by calculations made with an appropriate software
which considers total Ca2+ content, EGTA concentration, pH, presence of other salts and
temperature. For control BSS, a free Ca2+ concentration of 10�8 M was estimated considering
Ca2+ impurities from other salts (50 AM). Solutions with a free Ca2+ concentration of 10�7 and
10�6 M were prepared adding appropriate amounts of CaCl2 to the BSS. In the experiments
exploring intracellular pH effects on channel activity, a BSS with 10�7 M free Ca2+ and pH 7.4 was
used as control solution, and when pH was modified to 6.8 and 7.6, taking into account pH effects
on EGTA binding, free Ca2+ was maintained at the same concentration by changing the amount of
CaCl2 in this BSS.
The composition of the pipette solution (PS) was (in mM): 140 KCl, 0.5 MgCl2, 1 CaCl2, 10 HEPES
and 10 glucose, pH was adjusted to 7.4 with NaOH.
Reagents
Iberiotoxin (IBTX), ethylene glycol-bis(h-aminoethyl ether) N,N,NV,NV,-tetraacetic acid (EGTA) and
all the enzymes used for cell isolation were purchased from Sigma Chemical Co. All other reagents were
of analytical grade. When required by the experimental protocol, a fresh aliquot of a stock aqueous
solution of IBTX was added to the PS on the day of the experiment.
Statistics
The results are expressed as meansFSEM. Paired or unpaired Student’s t-tests were used to establish
statistically significant differences between two experimental groups. Chi-squared test was used to study
independence between groups of cells treated or untreated with IBTX. We used a Kruskal–Wallis one-
way ANOVA on ranks with Dunn’s multiple comparison procedure to test for differences between
groups of data that were not normally distributed (see Results). The level of statistically significant
differences was set to pb0.05.
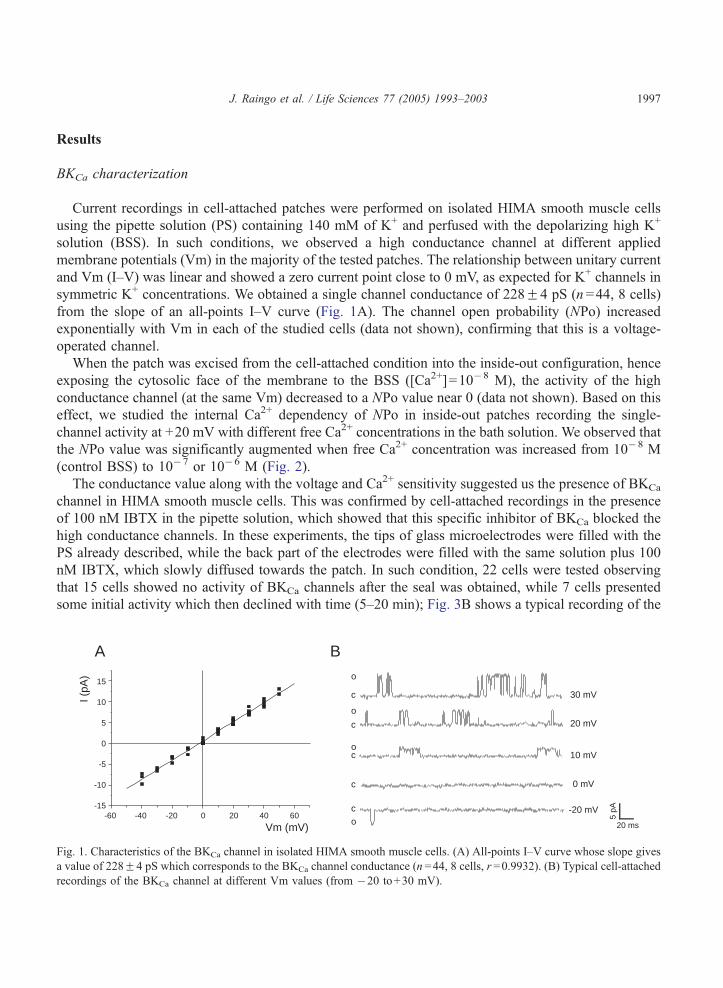
J. Raingo et al. / Life Sciences 77 (2005) 1993–2003 1997
Results
BKCa characterization
Current recordings in cell-attached patches were performed on isolated HIMA smooth muscle cells
using the pipette solution (PS) containing 140 mM of K+ and perfused with the depolarizing high K+
solution (BSS). In such conditions, we observed a high conductance channel at different applied
membrane potentials (Vm) in the majority of the tested patches. The relationship between unitary current
and Vm (I–V) was linear and showed a zero current point close to 0 mV, as expected for K+ channels in
symmetric K+ concentrations. We obtained a single channel conductance of 228F4 pS (n=44, 8 cells)
from the slope of an all-points I–V curve (Fig. 1A). The channel open probability (NPo) increased
exponentially with Vm in each of the studied cells (data not shown), confirming that this is a voltage-
operated channel.
When the patch was excised from the cell-attached condition into the inside-out configuration, hence
exposing the cytosolic face of the membrane to the BSS ([Ca2+]=10�8 M), the activity of the high
conductance channel (at the same Vm) decreased to a NPo value near 0 (data not shown). Based on this
effect, we studied the internal Ca2+ dependency of NPo in inside-out patches recording the single-
channel activity at +20 mV with different free Ca2+ concentrations in the bath solution. We observed that
the NPo value was significantly augmented when free Ca2+ concentration was increased from 10�8 M
(control BSS) to 10�7 or 10�6 M (Fig. 2).
The conductance value along with the voltage and Ca2+ sensitivity suggested us the presence of BKCa
channel in HIMA smooth muscle cells. This was confirmed by cell-attached recordings in the presence
of 100 nM IBTX in the pipette solution, which showed that this specific inhibitor of BKCa blocked the
high conductance channels. In these experiments, the tips of glass microelectrodes were filled with the
PS already described, while the back part of the electrodes were filled with the same solution plus 100
nM IBTX, which slowly diffused towards the patch. In such condition, 22 cells were tested observing
that 15 cells showed no activity of BKCa channels after the seal was obtained, while 7 cells presented
some initial activity which then declined with time (5–20 min); Fig. 3B shows a typical recording of the
A B
2mVm (mV)
I(pA
)
-60 -40 -20 0 20 40 60
15
10
5
0
-5
-10
-15
c
c
co
c
o
c
o
30 mV
20 mV
10 mV
0 mV
-20 mVo 5
pA
20 ms
Fig. 1. Characteristics of the BKCa channel in isolated HIMA smooth muscle cells. (A) All-points I–V curve whose slope gives
a value of 228F4 pS which corresponds to the BKCa channel conductance (n =44, 8 cells, r =0.9932). (B) Typical cell-attached
recordings of the BKCa channel at different Vm values (from �20 to+30 mV).
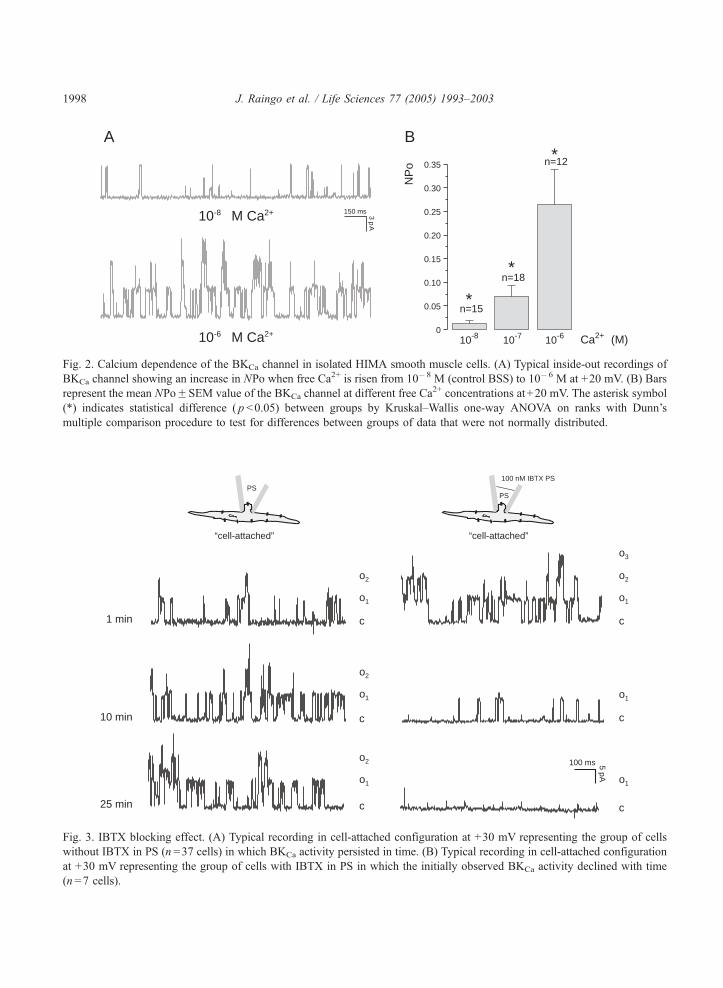
c1 min
10 min
25 min
“cell-attached”“cell-attached”
PS
5pA
100 ms
c
c
100 nM IBTX PS
PS
o1
o2
o1
o2
o1
o2
c
o1
o2
o3
c
o1
c
o1
Fig. 3. IBTX blocking effect. (A) Typical recording in cell-attached configuration at +30 mV representing the group of cells
without IBTX in PS (n =37 cells) in which BKCa activity persisted in time. (B) Typical recording in cell-attached configuration
at +30 mV representing the group of cells with IBTX in PS in which the initially observed BKCa activity declined with time
(n =7 cells).
A
10-8 10-7 10-6 Ca2+ (M)
NP
o
*
*
*B
n=15
n=18
n=120.35
0.30
0.25
0.20
0.15
0.10
0.05
0
150 ms 3p
A
10-8 M Ca2+
10-6 M Ca2+
Fig. 2. Calcium dependence of the BKCa channel in isolated HIMA smooth muscle cells. (A) Typical inside-out recordings of
BKCa channel showing an increase in NPo when free Ca2+ is risen from 10�8 M (control BSS) to 10�6 M at +20 mV. (B) Bars
represent the mean NPoFSEM value of the BKCa channel at different free Ca2+ concentrations at+20 mV. The asterisk symbol
(*) indicates statistical difference ( p b0.05) between groups by Kruskal–Wallis one-way ANOVA on ranks with Dunn’s
multiple comparison procedure to test for differences between groups of data that were not normally distributed.
J. Raingo et al. / Life Sciences 77 (2005) 1993–20031998
J. Raingo et al. / Life Sciences 77 (2005) 1993–2003 1999
attenuation of channel activity with time in one of these 7 cells. Conversely, when IBTX was absent
from the PS, BKCa activity was present in 37 out of 51 cells, without observing changes in activity with
time (Fig. 3A). In order to find statistically significant differences in the frequency of appearance of the
BKCa channel activity between the control group and the group with IBTX, we used a parametric Chi-
squared test. This analysis showed that the classificative variables (presence or absence of IBTX, and
activity or no activity of the BKCa channel) are not independent (Chi-squared test, pb0.001, n=73
cells), allowing us to conclude that the presence of IBTX statistically reduced the frequency of
appearance of the high conductance channel activity.
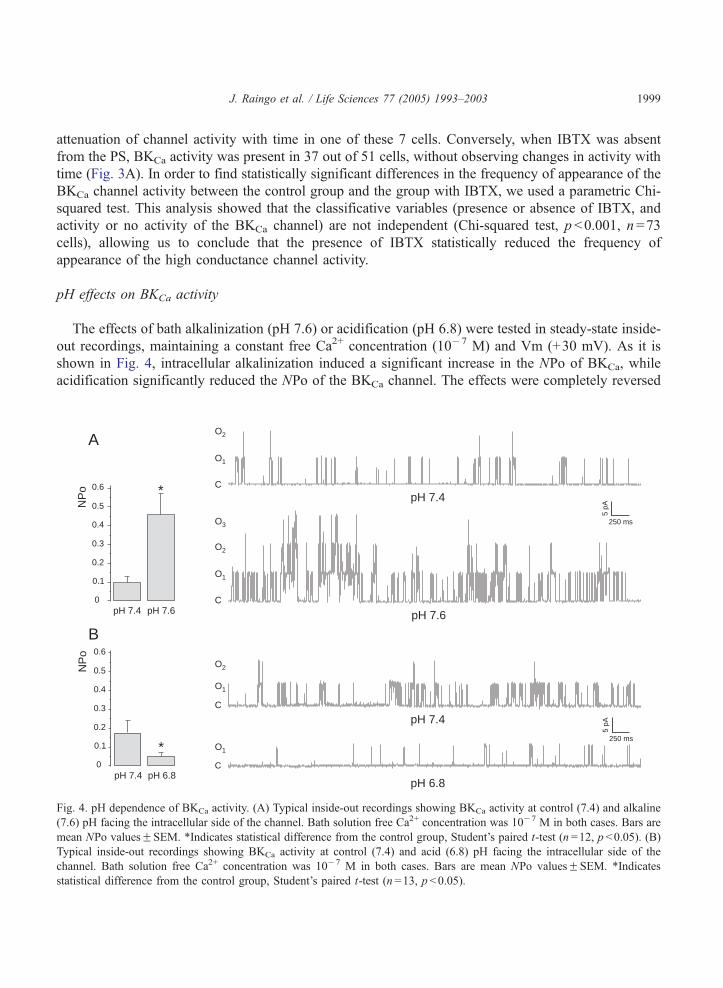
pH effects on BKCa activity
The effects of bath alkalinization (pH 7.6) or acidification (pH 6.8) were tested in steady-state inside-
out recordings, maintaining a constant free Ca2+ concentration (10�7 M) and Vm (+30 mV). As it is
shown in Fig. 4, intracellular alkalinization induced a significant increase in the NPo of BKCa, while
acidification significantly reduced the NPo of the BKCa channel. The effects were completely reversed
NP
o
pH 7.4 pH 7.6
*
NP
o
pH 7.4 pH 6.8
*
O2
O1
C
C
A
B
pH 7.4
pH 6.8
pH 7.6
pH 7.4
5pA
250 ms
5pA
250 ms
0.6
0.5
0.4
0.3
0.2
0.1
0
0.6
0.5
0.4
0.3
0.2
0.1
0
O2
O1
C
O3
O2
O1
C
O1
Fig. 4. pH dependence of BKCa activity. (A) Typical inside-out recordings showing BKCa activity at control (7.4) and alkaline
(7.6) pH facing the intracellular side of the channel. Bath solution free Ca2+ concentration was 10�7 M in both cases. Bars are
mean NPo valuesFSEM. *Indicates statistical difference from the control group, Student’s paired t-test (n =12, p b0.05). (B)
Typical inside-out recordings showing BKCa activity at control (7.4) and acid (6.8) pH facing the intracellular side of the
channel. Bath solution free Ca2+ concentration was 10�7 M in both cases. Bars are mean NPo valuesFSEM. *Indicates
statistical difference from the control group, Student’s paired t-test (n =13, p b0.05).

J. Raingo et al. / Life Sciences 77 (2005) 1993–20032000
after restoring control conditions (BSS at pH 7.4). In neither of the cases, we observed any changes in
the value of single-channel conductance.
Discussion
There is little information in the literature about the properties and role of K+ channels in HIMA,
the vessel of choice for myocardial revascularization. In this paper, we describe for the first time the
single-channel properties of BKCa channels present in cell-attached and inside-out patches obtained
from freshly dispersed HIMA smooth muscle cells. We observed a high frequency of occurrence of
BKCa activity (~73% of all patches studied) suggesting that these channels are highly expressed in this
vessel. In a symmetrical K+ concentration gradient, they show a single-channel conductance of 228
pS, are sensitive to 100 nM IBTX, and their open probability is Ca2+- and voltage-dependent. These
data are in accordance with the known properties of BKCa channels described in other vascular
smooth muscle cells.
An increase of BKCa activity can be an important mechanism to induce relaxation in response to
different physiological stimuli, such as shear-stress, vasoactive agonists or endothelial-derived factors
(Thomson et al., 1998; Carvajal et al., 2000; Lang et al., 2000). Archer et al. have recently demonstrated
that the 11,12-epoxyecosatrienoic acid is an endothelium-derived factor that hyperpolarizes and relaxes
HIMA by enhancing the activity of BKCa channels in this vessel (Archer et al., 2003). Moreover, as it
was reported by Rusch and Liu (1997), in pathological conditions such as hypertension BKCa channels
are up regulated providing a protective effect during acute or chronic episodes of high blood pressure.
Age also seems to influence the expression of BKCa channels: it has been recently reported that in human
coronary artery smooth muscle aging induces a reduction in the density of the alpha-subunit of these
channels (Toro et al., 2002).
The evidences presented in our work about single-channel BKCa activity in HIMA show that
changes in H+ concentration could directly modify the channel protein or its environment inducing an
increase or a decrease of BKCa open probability when intracellular pH augments or diminishes,
respectively. Kume et al. (1990) have also reported that in tracheal smooth muscle cells acidosis
inhibits Ca2+-activated K+ channels resulting in membrane depolarization, and Schubert et al. (2001)
have also given a detailed description showing that in rat vascular smooth muscle cells a decrease in
intracellular pH inhibits BKCa channels in inside-out patches and depresses whole-cell currents in
isolated cells. Then, changes in intracellular pH produced by physiological stimuli (i.e., vasoactive
agonists) or pathological conditions may trigger relaxing or contractile effects by modifications of K+
flux through BKCa channels.
The bibliographic references about pH and mechanical behavior of smooth muscle are extensive and
controversial in some aspects, results appearing to be dependent on the kind of smooth muscle tested and
the experimental conditions used in each case. Changes in H+ concentration may modify several cell
membrane Ca2+ transport pathways (Smith, G.L., et al., 1998; Smith, R.D. et al., 1998; Eto et al., 2003),
K+ channel activity (Smith et al., 1998), mobilization of Ca2+ from intracellular pools and Ca2+
sensitivity of contractile proteins (Smith, G.L. et al., 1998), as well as the activity of exchangers, i.e., Na/
H and Na/HCO3 (Neylon et al., 1990). Hence, the final effects of intracellular pH on the contractile state
of HIMAwill surely reflect the balance of different cellular mechanisms able to be modified directly or
indirectly by pH. Based on our electrophysiological results, we could speculate that BKCa channels

J. Raingo et al. / Life Sciences 77 (2005) 1993–2003 2001
would be inhibited or activated by low or high intracellular pH, respectively, contributing to determining
the final level of vessel contraction.
Conclusion
We characterized the single-channel properties of BKCa present in the HIMA and demonstrated its
direct activation by high and its inhibition by low intracellular pH. These data contribute to increase the
knowledge on the behavior of a vessel commonly used in myocardial revascularization.
Acknowledgements
The authors wish to thank Ms. Silvia Salemme for her excellent technical assistance. This work was
supported by grant PIP 4708 from the CONICET and PICT 0263 from the ANPCYT.
References
Archer, S.L., Gragasin, F.S., Wu, X., Wang, S., McMurtry, S., Kim, D.H., Platonov, M., Koshal, A., Hashimoto, K.,
Campbell, W.B., Falck, J.R., Michelakis, E.D., 2003. Endothelium-derived hyperpolarizing factor in human internal
mammary artery is 11,12-epoxieicosatrienoic acid and causes relaxation by activating smooth muscle BK(Ca) channels.
Circulation 107, 769–776.
Austin, C., Wray, S., 2000. Interaction between Ca2+ and H+ and functional consequences in vascular smooth muscle.
Circulation Research 86, 355–363.
Bychkov, R., Gollasch, M., Steinke, T., Ried, C., Luft, F.C., Haller, H., 1998. Calcium-activated potassium channels and nitrate-
induced vasodilation in human coronary arteries. Journal of Pharmacology and Experimental Therapeutics 285, 293–298.
Carvajal, J.A., Germain, A.M., Huidobro-Toro, J.P., Weiner, C.P., 2000. Molecular mechanism of cGMP-mediated smooth
muscle relaxation. Journal of Cell Physiology 184, 409–420.
Cracowski, J.L., Stanke-Labesque, F., Sessa, C., Hunt, M., Chavanon, O., Devillier, P., Bessard, G., 1999. Functional
comparison of the human isolated femoral artery, internal mammary artery, gastroepiploic artery, and saphenous vein.
Canadian Journal of Physiology and Pharmacology 77, 770–776.
Cracowski, J.L., Stanke-Labesque, F., Chavanon, O., Corompt, E., Veilt, S., Blin, D., Bessard, G., Devillier, P., 2000.
Thromboxane A2 modulates cyclic AMP relaxation and production in human internal mammary artery. European Journal of
Pharmacology 387, 295–302.
Dimitropoulou, C., White, R.E., Fuchs, L., Zhang, H., Catravas, J.D., Carrier, G.O., 2001. Angiotensin II relaxes microvessels
via the AT(2) receptor and Ca(2+)-activated K(+) (BK(Ca)) channels. Hypertension 37, 301–307.
Dincer, B., Borman, H.B., 1983. The occluded internal mammary artery graft. Restoration of patency after apparent occlusion
associated with progression of coronary disease. Journal of Thoracic and Cardiovascular Surgery 85, 318–320.
Eto, W., Hirano, K., Hirano, M., Nishimura, J., Kanaide, H., 2003. Intracellular alkalinization induces Ca2+ influx via non-
voltage-operated Ca2+ channels in rat aortic smooth muscle cells. Cell Calcium 34, 477–484.
Gojkoviic Bukarica, L., Kazic, T., Sajic, Z., Djukanovic, B., Panic, G., Peric, M., Bojic, M., 1997. The effects of
levocromakalim and pinacidil on the human internal mammary artery. Fundamental and Clinical Pharmacology 11,
550–560.
Gokina, N.I., Wellman, T.D., Bevan, R.D., Walters, C.L., Penar, P.L., Bevan, J.A., 1996. Role of Ca2+-activaded K+
channels in the regulation of membrane potential and tone of smooth muscle in human pial arteries. Circulation Research
79, 881–886.
Green, G.E., 1979. Techniques of internal coronary artery anastomosis. Journal of Thoracic and Cardiovascular Surgery 78,
455–459.

J. Raingo et al. / Life Sciences 77 (2005) 1993–20032002
Gruhn, N., Boesgaard, S., Eiberg, J., Bang, L., Thiis, J., Schroeder, T.V., Aldershvile, J., 2002. Effects of large conductance
Ca2+-activated K+ channels on nitroglycerin-mediated vasorelaxation in humans. European Journal of Pharmacology 446,
145–150.
Hamill, O.P., Marty, A., Neher, E., Sakmann, B., Sigworth, F.J., 1981. Improved patch clamp techniques for high-resolution
current readings from cells and cell-free membrane patches. Pflugers Archiv 391, 85–100.
Hatori, N., Fine, B.P., Nakamura, A., Cragoe Jr., E., Aviv, A., 1987. Angiotensin II-effect on cytosolic pH in cultured vascular
smooth muscle cells. Journal of Biological Chemistry 262, 5073–5078.
He, G.-W., Yang, C.-Q., 1997. Inhibition of vasoconstriction by potassium channel opener aprikalim in human conduit arteries.
British Journal of Clinical Pharmacology 44, 353–359.
Hubel, C.A., Highsmith, R.F., 1995. Endothelin-induced changes in intracellular pH and Ca2+ in coronary smooth muscle: role
of Na+–H+ exchange. Biochemical Journal 310, 1013–1020.
Kaczorowski, G.J., Garcia, M.L., 1999. Pharmacology of voltage-gated and calcium-activated potassium channels. Current
Opinion in Chemical Biology 3, 448–458.
Kajioka, S., Kitamura, K., Kuriyama, H., 1991. Guanosine diphosphate activates an adenosine 5V-triphosphate-sensitive K+
channel in the rabbit portal vein. Journal of Physiology 444, 397–418.
Klockner, U., 1993. Intracellular calcium ions activate a low-conductance chloride channel in smooth-muscle cells isolated from
human mesenteric artery. Pflugers Archiv 424, 231–237.
Kume, H., Takagi, K., Satake, T., Tokuno, H., Tomita, T., 1990. Effects of intracellular pH on calcium-activated potassium
channels in rabbit tracheal smooth muscle. Journal of Physiology 424, 445–457.
Lang, R.J., Harvey, G.J., McPhee, G.J., Klemm, M.F., 2000. Nitric oxide and thiol reagent modulation of Ca2+-activated K+
(BKCa) channels in myocites of guinea-pig taenia caeci. Journal of Physiology 525, 363–376.
Liu, M.-H., Storm Floten, H., Furnary, A.P., Yim, A.P.C., He, G.-W., 2001. Effects of potassium channel opener aprikalim on
the receptor-mediated vasoconstriction in the human internal mammary artery. Annals of Thoracic Surgery 71, 636–641.
Loop, F.D., Lytle, B.W., Cosgrove, D.M., Stewart, R.W., Goormastic, M., Williams, G.W., Golding, L.A., Gill, C.C., Taylor,
P.C., Sheldon, W.C., 1986. Influence of the internal mammary artery graft on 10-year survival and other cardiac events. New
England Journal of Medicine 314, 1–6.
Milesi, V., Aiello, E.A., Rebolledo, A., Gomez Alvis, A., Grassi de Gende, A.O., 1999. Role of a Ca2+-activated K+ current in
the maintenance of resting membrane potential of isolated, human, saphenous vein smooth muscle cells. Pflugers Archiv
437, 455–461.
Milesi, V., Raingo, J., Rebolledo, A., Grassi de Gende, A.O., 2003. Potassium channels in human umbilical artery cells. Journal
of the Society for Gynecologic Investigation 10, 339–346.
Neylon, C.B., Little, P.J., Cragoe Jr., E.J., Bobik, A., 1990. Intracellular pH in human arterial smooth muscle. Regulation by
Na+/H+ exchange and a novel 5-(N-ethyl-N-isopropyl)amiloride-sensitive Na+- and HCO3(�)-dependent mechanism.
Circulation Research 67, 814–825.
Pearson, P.J., Evora, P.R.B., Discigil, B., Schaff, H.V., 1998. Hypoxia increases vasodilator release from internal mammary
artery and saphenous vein grafts. Annals of Thoracic Surgery 65, 1220–1225.
Petrova-Kirova, P., Gagov, H., Krien, U., Duridanova, D., Noack, T., Schubert, R., 2000. 4-Aminopyridine affects rat smooth
muscle BKCa currents by changing intracellular pH. British Journal of Pharmacology 131, 1643–1650.
Rosenfeldt, F.L., He, G.-W., Buxton, B.F., Angus, J.A., 1999. Pharmacology of coronary artery bypass grafts. Annals of
Thoracic Surgery 67, 878–888.
Rusch, N., Liu, Y., 1997. Potassium channels in hypertension: homeostatic pathways to buffer arterial contraction. Journal of
Laboratory and Clinical Medicine 130, 245–251.
Schubert, R., Krien, U., Gagov, H., 2001. Protons inhibit the BK(Ca) channel of rat small artery smooth muscle cells. Journal of
Vascular Research 38, 30–38.
Smith, G.L., Austin, C., Crichton, C., Wray, S., 1998. A review of the action and control of intracellular pH in vascular smooth
muscle. Cardiovascular Research 38, 316–331.
Smith, R.D., Eisner, D.A., Wray, S., 1998. The effects of changing intracellular pH on calcium and potassium currents in
smooth muscle cells from the guinea-pig ureter. Pflugers Archiv 435, 518–522.
Stamatas, G.N., Patrick, C.W., McIntire, L.V., 1997. Intracellular pH changes of human aortic smooth muscle cells in response
to fluid shear stress. Tissue Engineering 3, 391–403.
Thomson, R.J., Nurce, C.A., 1998. Anoxia differentially modulates multiple K+ currents and depolarizes neonatal rat adrenal
chromaffin cells. Journal of Physiology 512, 421–434.

J. Raingo et al. / Life Sciences 77 (2005) 1993–2003 2003
Toro, L., Marijic, J., Nishimaru, K., Tanaka, Y., Song, M., Stefani, E., 2002. Aging, ion channel expression, and vascular
function. Vascular Pharmacology 38, 73–80.
Verma, S., Lovren, F., Dumont, A.S., Mather, K.J., Maitland, A., Kieser, T.M., Kidd, W., McNeill, J.H., Stewart, D.J., Triggle,
C.R., Anderson, T.J., 2001. Endothelin receptor blockade improves endothelial function in human internal mammary
arteries. Cardiovascular Research 49, 146–151.
Zhang, H., Li, P., Almassi, H., Nicolosi, A., Olinger, G.N., Rusch, N.J., 1996. Single-channel and functional characteristics
of a KCa channel in vascular muscle membranes of human saphenous veins. Journal of Cardiovascular Pharmacology 28,
611–617.





























