Embed Size (px)
Citation preview

Phloem transport of arsenic species from flag leaf tograin during grain filling
Anne-Marie Carey1, Gareth J. Norton1, Claire Deacon1, Kirk G. Scheckel2, Enzo Lombi3, Tracy Punshon4,
Mary Lou Guerinot4, Antonio Lanzirotti5, Matt Newville5, Yongseong Choi5, Adam H. Price1 and Andrew A.
Meharg1
1Institute of Biological and Environmental Sciences, University of Aberdeen, Cruickshank Building, St Machar Drive, Aberdeen AB24 3UU, UK; 2National
Risk Management Research Laboratory, US Environmental Protection Agency, 5995 Centre Hill Avenue, Cincinnati, OH 45224, USA; 3Centre for
Environmental Risk Assessment and Remediation, University of South Australia, Building X, Mawson Lakes Campus Mawson Lakes, S. Australia, SA-5095
Australia; 4Department of Biological Sciences, Dartmouth College, Hanover, NH 03755, USA; 5Centre for Advanced Radiation Sources, the University of
Chicago, Chicago, IL 60637, USA
Author for correspondence:Andrew A. Meharg
Tel: +44 1224 272264
Email: [email protected]
Received: 29 March 2011Accepted: 3 May 2011
New Phytologist (2011) 192: 87–98doi: 10.1111/j.1469-8137.2011.03789.x
Key words: arsenic, grain filling, phloem,rice, translocation.
Summary
• Strategies to reduce arsenic (As) in rice grain, below concentrations that
represent a serious human health concern, require that the mechanisms of As
accumulation within grain be established. Therefore, retranslocation of As species
from flag leaves into filling rice grain was investigated.
• Arsenic species were delivered through cut flag leaves during grain fill. Spatial
unloading within grains was investigated using synchrotron X-ray fluorescence
(SXRF) microtomography. Additionally, the effect of germanic acid (a silicic acid
analog) on grain As accumulation in arsenite-treated panicles was examined.
• Dimethylarsinic acid (DMA) and monomethylarsonic acid (MMA) were extre-
mely efficiently retranslocated from flag leaves to rice grain; arsenate was poorly
retranslocated, and was rapidly reduced to arsenite within flag leaves; arsenite dis-
played no retranslocation. Within grains, DMA rapidly dispersed while MMA and
inorganic As remained close to the entry point. Germanic acid addition did not
affect grain As in arsenite-treated panicles. Three-dimensional SXRF microtomog-
raphy gave further information on arsenite localization in the ovular vascular trace
(OVT) of rice grains.
• These results demonstrate that inorganic As is poorly remobilized, while organic
species are readily remobilized, from leaves to grain. Stem translocation of
inorganic As may not rely solely on silicic acid transporters.
Introduction
Rice (Oryza sativa), the staple food for over half theworld’s population, represents a significant dietary sourceof inorganic arsenic (As), a nonthreshold, class 1 humancarcinogen (Meharg et al., 2009). It is imperative thatstrategies to reduce grain As are developed and thosemechanisms which enable As to reach, and accumulatewithin, the rice grain be determined (Zhao et al., 2009,2010). The main species of As found in rice grain areinorganic As (arsenate and arsenite), and dimethylarsinicacid (DMA), with trace amounts of monomethylarsonicacid (MMA) and tetramethylarsonium sometimes present
(Williams et al., 2005; Hansen et al., 2011). Arsenate, thedominate soil species under aerobic conditions with a highaffinity for iron oxides, is an analog of phosphate, andenters rice roots through phosphate transporters, while themarkedly more mobile arsenite, the dominant redox formunder anaerobic conditions, enters the rice plant throughsilicic acid transporters (Abedin et al., 2002a; Ma &Yamaji, 2008; Ma et al., 2008). Consequently, as paddyrice cultivation relies on anaerobic soils, rice is particularlyeffective at accumulating As, compared with other cereals(Williams et al., 2007). Within the rice root, arsenate isreduced to arsenite, which then enters the stele via the Lsi2silicic acid effluxer (Ma & Yamaji, 2008; Xu et al., 2008;
NewPhytologist Research
� 2011 The Authors
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011) 192: 87–98 87www.newphytologist.com

Zhao et al., 2009). Su et al. (2010) reported that arsenitewas the main species found in the xylem sap of rice plantsfed with either arsenite or arsenate. Arsenite may becomplexed by phytochelatins, followed by sequestrationinto cell vacuoles (Bleeker et al., 2006; Raab et al., 2007;Zhao et al., 2009). Song et al. (2010) recently identifiedphytochelatin transporters responsible for transportingchelated arsenite into cell vacuoles, in Arabidopsis thaliana.The organic species DMA and MMA are assimilated at amuch slower rate by the root than inorganic As (Abedinet al., 2002b), with the protonated neutral forms also trans-ported through the silicic acid pathway (Li et al., 2009).Raab et al. (2007) and Li et al. (2009) found that methylat-ed As species are more readily translocated within the plantthan their inorganic counterparts. Once in the phloem ⁄xylem bundle of rice, DMA is translocated into grain withover an order of magnitude greater efficiency than inorganicAs species (Carey et al., 2010). Phloem transport accountedfor 90% of arsenite unloaded into grain, and for 55% ofDMA, in a study of shoot-to-grain translocation of arsenicspecies in excised panicles (Carey et al., 2010). A study ofAs transport in castor bean plants reported that arsenite wasnot complexed with thiols in the phloem, as thiol com-plexes are not stable in the alkaline conditions of thephloem (Ye et al., 2010).
Carey et al. (2010) used X-ray absorption near edgestructure (XANES) spectroscopy to examine the speciationof As in the developing rice grain for excised panicles thathad been exposed to either arsenate, arsenite or DMAthrough the cut stem (i.e. xylem plus phloem). DMA waspartially thiol-complexed within the grain and the redoxstatus of inorganic As seemed to be concentration-dependent(Carey et al., 2010). When arsenite was delivered to excisedrice panicles at 13.3 lM, arsenite was oxidized to arsenatewithin the rice grain, while under exposure to 133 lM,arsenite remained stable within the grain as free arsenite(Carey et al., 2010). Inorganic As is localized in the branlayer of rice grain, while the endosperm contains highconcentrations of DMA (Sun et al., 2008; Carey et al.,2010). The use of synchrotron X-ray fluorescence tech-niques to map the As distribution in rice grain with highconcentrations of inorganic As has shown As to be concen-trated in the region of the ovular vascular trace (OVT), thepoint of entry into the grain (Meharg et al., 2008; Lombiet al., 2009). Minerals are unloaded into the rice grain fromthe OVT, which contains both phloem and xylem cells,into the nucellar tissue and are then uploaded, via the apop-last, into the filial tissue (the aleurone and the endosperm)(Krishnan & Dayanandan, 2003). Lombi et al. (2009) sug-gested that this transport of material from the maternal tothe filial tissues may represent a physiological barrier whichAs species cross with differential efficiency. SynchrotronX-ray fluorescence (SXRF) microtomography of developingrice grains, that were fed arsenite or DMA through excised
stems, found that arsenite was localized in the OVT regionwhile DMA had rapidly spread throughout the outer layersand into the endosperm. These findings explain why inor-ganic As is concentrated in rice bran while the endospermcontains higher concentrations of DMA. Zheng et al.(2011) examined temporal variation in As concentrations inrice plants at different stages of growth and reported thatthe concentration of DMA decreased during grain fill,suggesting that DMA accumulation in grain occurred viathe retranslocation of DMA accumulated in the plant beforeflowering, while inorganic As concentrations remainedconstant, suggesting it was taken up during the period ofgrain fill. However, whether As is actually retranslocated tothe grain from flag leaves has yet to be established and theredox status of inorganic As within the flag leaves remainsunknown.
The aim of this study was to investigate the retransloca-tion of As species from flag leaves into the filling rice grain,by feeding As species directly into the flag leaves duringgrain fill, and to determine the spatial unloading of key Asspecies within the developing rice grain and establishwhether differences existed between leaf-retranslocated anddirect stem-transported As. Additionally, to investigate therole of silicic acid and arsenite pathways, germanic acid, asilicic acid analog (Ma et al., 2002; Nikolic et al., 2007),was used as a tracer in competitive inhibition in shoot andstem feeding studies. Furthermore, three-dimensional (3D)SXRF microtomography gave more insights into arsenitelocalization in the OVT after arsenite treatment.
Materials and Methods
Plant growth
For all experiments, a quick flowering Oryza sativa L. cvItalica Carolina was utilized, and grown to maturity, withhealthy panicles labelled at anthesis as described in Careyet al., 2010.
Leaf feeding of As species
To investigate the remobilization of As species from flagleaves during grain development, rice panicles were exposedto 333 lM of arsenite, arsenate, DMA or MMA at 10 dpost-anthesis (DPA). Treatment stock solutions of 13.3 mMwere prepared by dissolving the appropriate salt in Milli-Qdeionized water, and seven subsamples of each stock werefrozen for subsequent removal and use on the relevant day.Treatment vials were prepared by making 0.25 ml of thedefrosted treatment stock up to 1 ml with Milli-Q deionizedwater and then pipetting 0.1 ml of this diluted stock into aweighed Eppendorf vial, together with 2-(N-morpholino)ethanesulfonic acid (MES) buffer at a final concentration of5 mM, and rubidium (Rb) and strontium (Sr), as markers
88 Research
NewPhytologist
� 2011 The Authors
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011) 192: 87–98
www.newphytologist.com

for phloem and xylem transport, respectively, at final concen-trations of 1 mM (Kuppelwieser & Feller, 1991; Careyet al., 2010). Solutions were pH-adjusted to 6.4 withNaOH. This was then made up to 1 ml, by weight, by theaddition of Milli-Q, yielding a treatment concentration of333 lM arsenite, arsenate, DMA or MMA. For arsenite andDMA, additional treatments were included (0, 33 and133 lM) to generate a dose response and establish the bestconcentration to use in the primary experiments.
A sharp razor blade was used to remove the flag leaf tipand this cut end was inserted into the Eppendorf vial. Astrip of aluminum foil was applied to limit evaporation, anddust entry, and the vials were held in place by strong tape.Treatments were delivered through the cut panicle flag leafon intact plants for 7 d, with a fresh vial applied every 24 h.Each time a fresh vial was applied, the tip of the flag leafwas re-cut to ensure a fresh and open entry point.Treatment vials were frozen on removal from the flag leaves.For each treatment and control, three replicate panicleswere used. Control ± Rb and Sr were also included.
Inorganic As speciation in flag leaf
For flag leaves fed arsenite or arsenate, As speciation in thefresh flag leaf was analyzed to evaluate inorganic As status.Immediately following removal of the last treatment vial, asection of flag leaf lamina was cut from the ligule ⁄ sheathend of the treated leaf, at least 3–4 cm from solution con-tact, and ground rapidly in liquid nitrogen. This was thensuspended in 5 ml of chilled phosphate buffer (a solution of6.66 mM ammonium nitrate and 6.66 mM ammoniumhydrophosphate, adjusted to pH 6.2 using ammonia) and1 ml of the resulting slurry was decanted into a 1.5 mlmicrocentrifuge tube and centrifuged at 12 000 g for2 min. The supernatant was diluted (0.5 ml made up to5 ml) with chilled phosphate buffer solution and a 1 mlsubsample was taken for As speciation analysis, via anionexchange high-performance liquid chromatography coupledwith inductively coupled plasma mass spectrometry (HPLC-ICP-MS), as described in Sun et al. (2009). Each samplewas analyzed on the HPLC-ICP-MS within 20–30 min ofremoval from the rice plant. The remaining contents oftreatment vials were also speciated to confirm As species sta-bility. All treatment vials were frozen on removal from theplant and defrosted on the day of speciation analysis. Forthe arsenite and arsenate treatments, 0.1 ml was subsampledfrom each of the seven daily treatment vials for all threereplicates and diluted 1 : 1000 with chilled phosphatebuffer and analyzed for As speciation by HPLC-ICP-MS.
Total As analysis for leaf fed panicles
All treatments and controls were analyzed for total As viaICP-MS. Flag leaf sections were cut from the ligule ⁄ sheath
end of the blade, at least 3–4 cm from contact with tapeor solution where they had been leaf-fed. Grains wereselected from the top and middle regions of the panicleand husks were separated manually, with a portion of thegrain from each panicle being kept chilled for synchrotronanalysis and the remainder being oven-dried for ICP-MSanalysis. One fresh grain from one replicate of each treat-ment was then randomly selected for SXRF microtomo-graphy. Time limitations at the synchrotron prohibited theanalysis of more than one replicate per treatment. Oven-dried samples were microwave-digested and analyzed fortotal As, Rb and Sr concentrations, by ICP-MS (7500Agilent Technologies, Edinburgh, UK), as described inSun et al. (2008). Quality control procedures were asdescribed in Sun et al. (2008).
SXRF microtomography
For leaf-fed As-treated grain, SXRF microtomography wasconducted on fresh rice grain, pulsed with 333 lM of eitherarsenite, arsenate, DMA or MMA, at GeoSoilEnviroCARS(Sector 13) of the Advanced Photon Source, ArgonneNational Laboratory. Details were as described in Careyet al. (2010). The electron storage ring operated at 7 GeVwith a top-up fill mode. Fresh rice grain was suspendedfrom a rotation-translation stage in the path of a 3 lm X-ray beam and translated across in 3 lm steps with a dwelltime of 0.05 s per step. The grain was then rotated 0.5� andthe scan process repeated until a rotation of 0–180º wascomplete. Fluorescence data were collected using a multi-element fluorescence detector and the resulting two-dimen-sional (2D) sine wave plots were reconstructed as describedin McNear et al. (2005).
Additionally, as it was established that grain arsenite isnot retranslocated from flag leaves, further SXRF tomo-graphy was conducted for fresh developing rice grain pulsedwith 133 lM arsenite via the excised panicle stem, yieldingimages that offer an improved resolution compared withCarey et al. (2010). Fully rendered 3D microtomographicreconstructions of As in the grain were collected at beamlineX26A of the National Synchrotron Light Source(Brookhaven National Laboratory, Upton, NY, USA) withthe monochromator tuned to 12.5 keV to resolve the Asedge. Microtomography was carried out as described inKim et al. (2006). Installation of an additional silicon driftdetector at 180� to the existing multi-element germanium(Ge) array detector effectively removed self-absorptioneffects for the low Z elements, and allowed the use ofon-the-fly data processing. This reduced spectral collectiontimes to 50 ms per point. Arsenite-pulsed grain wasmounted on a Huber goniometer (x, y, z, h stage). A totalof 20 tomographic sections were collected from the awn tothe rachilla at intervals of 240 lm. The beam dimensionswere 10 lm horizontal · 6 lm vertical, with 20 lm pixel
NewPhytologist Research 89
� 2011 The Authors
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011) 192: 87–98
www.newphytologist.com

sizes. Data were rendered using tomographic display soft-ware written for the Interactive Data Language (IDLversion 8.0) program (ITT Visual Information Solutions,Boulder, CO, USA).
Phloem vs xylem transport of germanic acid in excisedpanicles
The comparative contributions of phloem and xylem in theshoot to grain translocation of germanic acid in ± stemgirdled plants was investigated, with panicles also exposedto Rb and Sr, as markers for phloem and xylem flow,respectively (Kuppelwieser & Feller, 1991; Carey et al.,2010). Panicle stems were girdled with a jet of steam1–2 cm below the panicle head, 24 h before excision fromthe plant, as described in Carey et al. (2010). This destroysthe phloem cells, preventing further phloem transport intothe rice grain while xylem vessels remain functional(Martin, 1982; Kuppelwieser & Feller, 1991; Chen et al.,2007; Carey et al., 2010).
At 10 DPA, plants were placed in darkness for 2 h toreduce transpiration, limiting the formation of air bubblesin the xylem during excision. Panicles were then excisedbelow the flag leaf node and placed in Pyrex tubes of auto-claved nutrient solution that had been spiked with theappropriate treatment, as described in Carey et al. (2010).Solutions contained Rb and Sr at a final concentration of1 mM. MES buffer, at a final concentration of 5 mM, wasadded to autoclaved nutrient solutions through a sterilefilter. Germanic acid treatment stocks were prepared by dis-solving GeO2 in Milli-Q deionized water containing 0.2%4 M NaOH to form germanic acid. Panicles were thentransferred to a growth chamber with a 12 h photoperiod, aday : night temperature of 28 : 23�C, relative humidity of80 : 60 and light intensity of 1200 lmol m)2 s)1. For eachtreatment and control three replicate panicles were used.Controls ± Rb and Sr were included. Following the 48 htreatment period flag leaves and grains were oven dried andgrains were selected from the top and middle regions of thepanicle and manually de-husked.
Additionally, to assess whether shoot-to-grain translocationof arsenite utilizes silicic acid transporters, a competitionexperiment was conducted by treating excised rice panicleswith a hydroponically administered 48 h pulse of 133 lMarsenite or arsenate (or Milli-Q for the control panicles),± 1.33 mM Ge during grain development.
Samples were microwave-digested and total Ge wasmeasured by ICP-MS as described earlier.
Retranslocation of Ge from flag leaf
To investigate the remobilization of Ge from flag leavesduring grain development, rice panicles were exposed to333 lM of Ge, via the cut flag leaf on intact plants, at
10 DPA. This was conducted alongside the leaf-fed Asexperiment and followed the same procedure.
Results
Leaf feeding of As species
Total solution uptake from feeding vials was less for theDMA and MMA treatments than for the control and inor-ganic treatments (Fig. 1). ANOVA demonstrated that Astreatment had a significant effect on solution uptake fromfeeding vials (P = 0.01) with inorganic As treatments andthe As control having significantly greater total solutionuptake than the MMA treatment, and the arsenate treat-ment having significantly greater uptake than DMA, withno difference between the two organic treatments.
Mean total As concentrations in flag leaf and grain forleaf-fed As species are reported in Fig 1. ANOVA revealed
Control
Arsen
ite
Arsen
ate
DMA
MMA
Fla
g le
af a
rsen
ic (
mg
kg–1
)
0
100
200
300
400
Sol
utio
n up
take
(m
l)
0
2
4
6
8
10
Leaf fed arsenic treatment
Control
Arsen
ite
Arsen
ate
DMA
MMA
Gra
in a
rsen
ic (
mg
kg–1
)
0
5
10
15
20
25
Sol
utio
n up
take
(m
l)
0
2
4
6
8
10
(a)
(b)
Fig. 1 Mean total arsenic (As) concentrations (bars) in flag leaf (a)and grain (b), for rice panicles exposed to 333 lM of one of fiveAs treatments together with 1 mM rubidium and strontium.Treatments were delivered through the cut flag leaf on intactplants for 7 d, with a fresh vial applied every 24 h. Total solutionuptake is also shown for each treatment (circles). Error barsrepresent ± SE of three replicates. DMA, dimethylarsinic acid;MMA, monomethylarsonic acid.
90 Research
NewPhytologist
� 2011 The Authors
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011) 192: 87–98
www.newphytologist.com

that As treatments had a highly significant effect on flag leafAs concentrations (P < 0.001), with mean flag leaf Assignificantly lower for As controls than all other As treat-ments, and no significant differences between the four Asspecies, although there was a large degree of variation inMMA-treated leaves. In the grain, ANOVA determinedthat for all As species, with the exception of arsenite, whichwas not significantly different to the control, mean grainAs was significantly increased by flag leaf exposure (P <0.001). Fisher’s pairwise comparisons showed that organicAs treatments, between which there was no significant dif-ference, led to significantly greater mean grain As thaninorganic treatments, with concentrations 100 times greaterthan those for arsenate-treated plants, and that leaf feedingof arsenate led to significantly higher grain As than feedingwith arsenite.
Mean total Rb and Sr concentrations in flag leaf and grainfor leaf-fed As species are detailed in the SupportingInformation (Methods S1 and Fig. S1). The results demon-strate that there is significant phloem transport to thedeveloping grain from the flag leaf, and that this is notaffected by the addition of As. There was no measurabletransport of Sr from the flag leaf to the grain, suggesting thatthere was little xylem transport from the flag leaf to the grain,although, in view of the large degree of variation betweenreplicates (Fig. S1), it cannot be definitively concluded thatthere is no xylem transport from the flag leaf to the grain.
As speciation in flag leaf
HPLC-ICP-MS analysis of As speciation in freshlyextracted flag leaves exposed to arsenite and arsenate for7 d, with feeding vials changed daily, showed that arseniteremained stable as arsenite in the leaf, with no conversionto other species, while arsenate was completely converted toarsenite (Fig. S4). Speciation of the remaining feeding vialcontents after 24 h attachment to the flag leaf found thatarsenite and arsenate treatments had remained stable withinthe vial for the duration of the feeding period, with As inarsenate feeding solutions predominantly arsenate (99%arsenate and 1% arsenite) and arsenite feeding vials contain-ing 100% arsenite (Fig. S5).
Imaging As unloading
Synchrotron X-ray fluorescence microtomography imagescollected for fresh developing rice grains fed with As speciesvia flag leaves are compared with those collected previouslyfor developing rice grains fed via the excised stem (Fig. S6).It appears that As species delivered to the rice grain via re-translocation from the flag leaf do not differ in theirunloading and distribution from those delivered via directstem transport. MMA and arsenate, not previously ana-lyzed, appear to be retained in the OVT region of the fresh
grain, whether fed via the cut flag leaf or the excised paniclestem (Fig. S6).
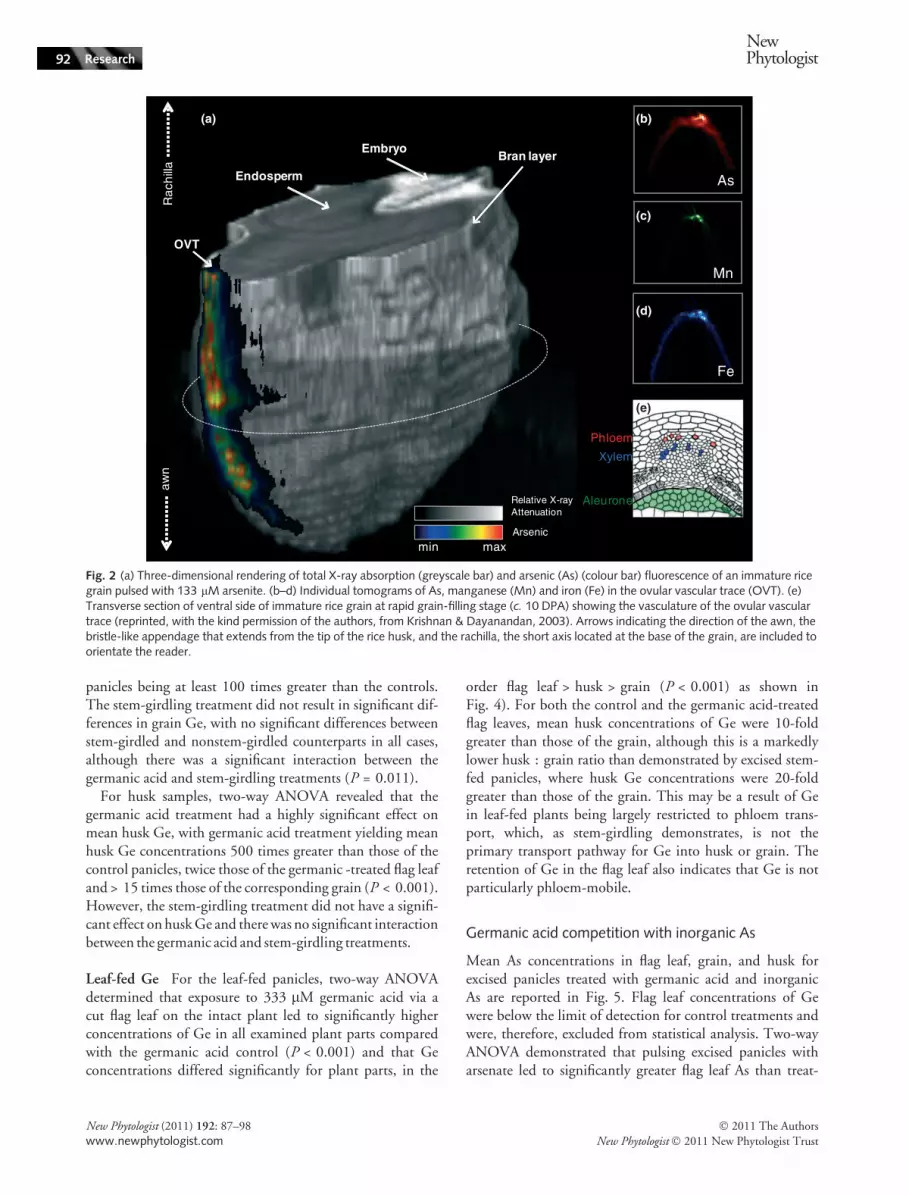
Synchrotron X-ray fluorescence microtomography imagesin Fig. 2 show As distribution, during unloading into therice grain, for an excised panicle pulsed with 133 lM arse-nite for 48 h, enhancing the information provided in Careyet al. (2010). In the 3D image in Fig. 2(a), As can clearlybe seen in the OVT, which runs down the ventral side ofthe grain (Krishnan & Dayanandan, 2003), apparentlycoincident with Fe and Mn, in agreement with previousstudies (Meharg et al., 2008; Lombi et al., 2009; Careyet al., 2010). However, closer inspection (Fig. 2b–d) revealsthat Fe and Mn appear to be located on either side of theAs. The 2D tomograms in Fig. 2(b–d) demonstrate that As,while remaining largely localized in the OVT, has begun tospread beyond it, indicating that arsenite is being unloadedinto the nucellar epidermis that encircles the endosperm,but much less efficiently than DMA. With regard to Fe andMn, Fe appears to be more efficiently unloaded into thenucellar epidermis than arsenite, while Mn is more stronglyretained within the OVT.
Ge phloem and xylem transport
Excised stem fed Ge Mean concentrations of the phloemmarker, Rb, and the xylem marker, Sr, in the grain, huskand flag leaves for the stem-girdling experiment are detailedin Methods S1 and Fig. S7. These results demonstrate thatthe stem-girdling treatment was effective in interruptingphloem transport without damaging xylem transport andthat addition of germanic acid did not significantly affectphloem or xylem transport to the rice grain and husk.
Silicic acid itself was originally utilized to examine silicicacid ⁄ arsenite competition in shoot-to-grain transport ofinorganic As; however, addition of silicic acid at 10 timesthe molar concentration of As proved so phytotoxic to theexcised panicles that the data could not be meaningfullyinterpreted. Therefore, germanic acid was used. Meanconcentrations of Ge in the flag leaf, grain, and husk forthe ± stem-girdled panicles stem-fed germanic acid arereported in Fig. 3. Since stem-girdled panicles were gener-ally smaller and took up less solution than nonstem-girdledpanicles, solution uptake has been included in Fig. 3. Thisdemonstrates that any observed differences between stem-girdled and nonstem-girdled germanic acid treatments ingrain, husk and flag leaf Ge concentration corresponds withdifferences in solution uptake. Two-way ANOVA showedthat germanic acid treatment had a highly significant effecton mean flag leaf Ge concentrations (P < 0.001), resultingin 100 times more Ge than control panicles, and that therewas a significant effect of stem-girdling (P = 0.037).
Two-way ANOVA revealed that the germanic acid treat-ment had a highly significant effect on mean grain Geconcentrations (P < 0.001), with germanic acid-treated
NewPhytologist Research 91
� 2011 The Authors
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011) 192: 87–98
www.newphytologist.com
panicles being at least 100 times greater than the controls.The stem-girdling treatment did not result in significant dif-ferences in grain Ge, with no significant differences betweenstem-girdled and nonstem-girdled counterparts in all cases,although there was a significant interaction between thegermanic acid and stem-girdling treatments (P = 0.011).
For husk samples, two-way ANOVA revealed that thegermanic acid treatment had a highly significant effect onmean husk Ge, with germanic acid treatment yielding meanhusk Ge concentrations 500 times greater than those of thecontrol panicles, twice those of the germanic -treated flag leafand > 15 times those of the corresponding grain (P < 0.001).However, the stem-girdling treatment did not have a signifi-cant effect on husk Ge and there was no significant interactionbetween the germanic acid and stem-girdling treatments.
Leaf-fed Ge For the leaf-fed panicles, two-way ANOVAdetermined that exposure to 333 lM germanic acid via acut flag leaf on the intact plant led to significantly higherconcentrations of Ge in all examined plant parts comparedwith the germanic acid control (P < 0.001) and that Geconcentrations differed significantly for plant parts, in the
order flag leaf > husk > grain (P < 0.001) as shown inFig. 4). For both the control and the germanic acid-treatedflag leaves, mean husk concentrations of Ge were 10-foldgreater than those of the grain, although this is a markedlylower husk : grain ratio than demonstrated by excised stem-fed panicles, where husk Ge concentrations were 20-foldgreater than those of the grain. This may be a result of Gein leaf-fed plants being largely restricted to phloem trans-port, which, as stem-girdling demonstrates, is not theprimary transport pathway for Ge into husk or grain. Theretention of Ge in the flag leaf also indicates that Ge is notparticularly phloem-mobile.
Germanic acid competition with inorganic As
Mean As concentrations in flag leaf, grain, and husk forexcised panicles treated with germanic acid and inorganicAs are reported in Fig. 5. Flag leaf concentrations of Gewere below the limit of detection for control treatments andwere, therefore, excluded from statistical analysis. Two-wayANOVA demonstrated that pulsing excised panicles witharsenate led to significantly greater flag leaf As than treat-
Relative X-rayAttenuation
Arsenic min max
awn
Rac
hill
aEndosperm
EmbryoBran layer
(a)
As
Mn
Fe
(b)
(c)
(d)
OVT
(e)
Phloem
Xylem
Aleurone
Fig. 2 (a) Three-dimensional rendering of total X-ray absorption (greyscale bar) and arsenic (As) (colour bar) fluorescence of an immature ricegrain pulsed with 133 lM arsenite. (b–d) Individual tomograms of As, manganese (Mn) and iron (Fe) in the ovular vascular trace (OVT). (e)Transverse section of ventral side of immature rice grain at rapid grain-filling stage (c. 10 DPA) showing the vasculature of the ovular vasculartrace (reprinted, with the kind permission of the authors, from Krishnan & Dayanandan, 2003). Arrows indicating the direction of the awn, thebristle-like appendage that extends from the tip of the rice husk, and the rachilla, the short axis located at the base of the grain, are included toorientate the reader.
92 Research
NewPhytologist
� 2011 The Authors
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011) 192: 87–98
www.newphytologist.com

ment with arsenite (P = 0.019), although at 1.5-fold, thisdifference was not as marked as that observed in grain andhusk samples, while germanic acid addition had no signifi-cant effect. There was no interaction between As andgermanic acid treatments.
Two-way ANOVA showed that As treatments had a sig-nificant effect on grain As concentrations (P < 0.001), witharsenite and arsenate addition both yielding significantly
higher grain As than the controls and arsenate, leading tosignificantly higher (three- to fourfold more) grain As thanthe arsenite treatment. Germanic acid, fed at 10 times themolar concentration of the As species, had no significanteffect on grain As concentrations and there was no signifi-cant interaction between As and germanic acid treatments.
For the husk, two-way ANOVA showed that treatmentwith As had a highly significant effect on husk As concen-trations (P < 0.001) while germanic acid treatment did notlead to significant differences in husk As and there wasno significant interaction between the As and germanicacid treatments. As with the grain, pairwise comparisonsrevealed that treatment with arsenate led to significantlygreater (three- to fourfold more) husk As than the arsenitetreatment (P < 0.001).
There were no significant differences in solution uptake(Fig. 5); two-way ANOVA confirmed that the As and ger-manic acid treatments did not lead to any significantdifferences in solution uptake and there was no significantinteraction between treatments.
Discussion
Retranslocation of As from shoot to grain
This study investigated the retranslocation of As speciesfrom flag leaves to rice grain during grain filling, by feedingAs species directly into the panicle flag leaf of intact plants.
Retranslocation of the organic As species, DMA andMMA, was extremely efficient, despite lower solution uptakecompared with the inorganic treatments and controls. Flagleaves treated with DMA took up 25–30% less feeding solu-tion than the inorganic As and control treatments, andMMA-treated leaves took up 35–40% less feeding solution.This is consistent with their use as herbicides and defoliants(desiccants) (Marcus Wyner & Rains, 1982; Mastradone &
Gra
in G
e (m
g kg
–1)
0
2
4
6
8
10
12
14H
usk
Ge
(mg
kg–1
)
0
50
100
150
200
250
300
Contro
l-Rb-
Sr –SG
Contro
l-Rb-
Sr +SG
Ge Con
trol –
SG
Ge Con
trol +
SG
Ge tre
atm
ent –
SG
Getre
atm
ent +
SG
Fla
g le
af G
e (m
g kg
–1)
0
20
40
60
80
100
120
140
Sol
utio
n up
take
(g)
0
10
20
30
Sol
utio
n up
take
(g)
0
10
20
30
Sol
utio
n up
take
(g)
10
20
30
(a)
(b)
(c)
Fig. 3 Mean total concentrations of germanium (Ge) in grain (a),husk (b), and flag leaf (c) for excised rice panicles subjected to a (±)stem-girdling (± SG) treatment and hydroponically fed, over a 48 hperiod, nutrient solution amended with 133 lM germanic acid and1 mM rubidium (Rb) and strontium (Sr); 1 mM Rb and Sr only(germanic acid controls); or, for zero exposure controls, noamendment. Total solution uptake is also shown for each treatment(circles). Error bars represent the ± SE of three replicates.
Grain Husk Flag leaf
Ger
man
ium
(m
g kg
–1)
0
100
200
300
400
Sol
utio
n up
take
(m
l)
0
2
4
6
8
10
Fig. 4 Mean total germanium (Ge) concentrations in flag leaf, grain,and husk for rice panicles exposed to 333 lM germanic acid, and1 mM rubidium (Rb) and strontium (Sr), through the cut flag leaf onintact plants for 7 d, with a fresh vial applied every 24 h. Totalsolution uptake is also shown for each treatment (circles). Error barsrepresent ± SE of three replicates.
NewPhytologist Research 93
� 2011 The Authors
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011) 192: 87–98
www.newphytologist.com

Woolson, 1983; Marin et al., 1993; Valette-Silver et al.,1999; Rosen & Liu, 2009). The dose response conducted forDMA and arsenite (Fig. S2) confirmed that DMA was effi-ciently transported to the grain, regardless of leaf dose, incontrast to arsenite, which displayed no measureable retrans-location from flag leaf to grain. Mean total solution uptake,for each treatment in the dose response experiment, isreported in the Supporting Information (Fig. S3). The rela-tively high mobility of DMA and MMA compared with
inorganic As agrees with previous studies (Raab et al., 2007;Li et al., 2009; Carey et al., 2010). Raab et al. (2007) sug-gested this might be because of poorer thiol complexationlimiting retardation during translocation. A recent studyshowed that DMA and MMA both remained stable and un-complexed in the phloem and xylem (Ye et al., 2010).
Zheng et al. (2011) reported that the DMA concentrationin rice grain decreased during grain filling. The results pre-sented here appear to contradict that study, as DMA wasactively translocated into filling grain to reach relatively highconcentrations. As DMA is so rapidly retranslocated fromflag leaves to rice grain, DMA that has accumulated withinthe plant before panicle formation will swiftly partition intothe panicle and ovary, during development, leading to a highproportion of DMA in the initial stages of panicle develop-ment and grain fill. As DMA is slowly taken up by rice roots(Abedin et al., 2002b), in contrast to inorganic As, theamount of DMA taken up by the plant throughout the grainfill period will be comparatively low, and further diluted bythe carbohydrate filling the grain. Oparka & Gates (1981)suggested that water entering the endosperm of developinggrains, with the mass flow of assimilates, leaves the grain viathe xylem of the pericarp bundle. It is possible that someDMA, being xylem-mobile, does so too, although this wouldnecessitate a corresponding increase in plant tissues else-where and Zheng et al. (2011) did not detect this.
While MMA is similar to DMA in its rapid retransloca-tion from flag leaf to rice grain, SXRF microtomographydemonstrates that MMA and DMA have very differentdegrees of mobility within the developing grain. DMA rap-idly spreads throughout the outer layers and into theendosperm, whether arriving via direct stem transport or viaremobilization from flag leaves. MMA, conversely, isretained within the OVT region in a similar pattern toinorganic As species (Fig. S6).
With regard to inorganic As, arsenate was relativelypoorly retranslocated to the rice grain, but proved moremobile than arsenite, which displayed no measureableremobilization from flag leaf to grain (Fig. 1). Arsenate wasalso three to four times more efficiently translocated intothe rice grain than arsenite when fed through excised pani-cles at concentrations of 133 lM (Fig. 5). Despiteexhibiting a similar lack of mobility within the rice grain(Fig. S6), arsenate is clearly more mobile within the shootthan arsenite, probably because of arsenate’s low affinity forthiol groups (Raab et al., 2007). However, arsenate israpidly reduced to arsenite within the flag leaf and, in ahydroponic study, arsenite was the predominant speciesfound in the xylem of rice plants fed arsenate (Su et al.,2010). Carey et al. (2010) did not find any significantdifferences in shoot-to-grain transport of arsenite and arse-nate when fed through excised panicles at concentrations of13.3 lM. In that study, XANES speciation of developingrice grains demonstrated that, at 13.3 lM exposure,
Gra
in a
rsen
ic (
mg
kg–1
)
0
2
4
6
8H
usk
arse
nic
(mg
kg–1
)
0
5
10
15
20
25
As and Ge treatments
Contro
l
Contro
l + G
eAsII
I
AsIII +
Ge
AsV
AsV +
Ge
Fla
g le
af a
rsen
ic (
mg
kg–1
)
0
50
100
150
200
250
300
Sol
utio
n up
take
(g)
0
5
10
15
20
25
30
Sol
utio
n up
take
(g)
0
5
10
15
20
25
30
Sol
utio
n up
take
(g)
0
5
10
15
20
25
30
(a)
(b)
(c)
Fig. 5 Mean total arsenic (As) concentrations in rice grain (a), husk(b), and flag leaf (c) for excised rice panicles hydroponically fed, overa 48 h period, nutrient solution amended with either 133 lMarsenite (AsIII) or arsenate (AsV) together with 1.33 mM germanicacid (Ge). Total solution uptake is shown for each treatment(circles). Error bars represent ± SE of three replicates.
94 Research
NewPhytologist
� 2011 The Authors
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011) 192: 87–98
www.newphytologist.com

arsenite in the grain was oxidized to arsenate, while at133 lM exposure, arsenite remained stable (Carey et al.,2010). The interpretation that arsenate is retranslocatedfrom the flag leaf to the grain, albeit in small quantities,while arsenite is not, suggests that some of the initial arse-nate is exported to grain, potentially through the phosphatepathways, before reduction takes place. Within the flag leaf,arsenite is likely detoxified through complexation withthiol-rich peptides, including phytochelatins and glutathi-one, and sequestered into leaf cell vacuoles (Bleeker et al.,2006; Raab et al., 2007; Zhao et al., 2009). Song et al.(2010) recently identified phytochelatin transportersresponsible for facilitating the sequestration of chelatedarsenite into cell vacuoles, in A. thaliana. In this study, Asspeciation would not have distinguished between free arse-nite and thiol-complexed arsenite. A number of studieshave shown that translocation of As from shoot to rice grainbecomes less efficient at higher shoot As concentrations,indicating some threshold (Williams et al., 2007; Adomakoet al., 2009; Lu et al., 2009; Norton et al., 2009). Nortonet al. (2009) reported that, for a range of cultivars, the per-centage of DMA in rice grain increased with increasingtotal As, while the percentage of arsenite decreased (Nortonet al., 2009). This may be, in part, the result of a greaterproportion of arsenite being retained within leaf cell vacu-oles. Zheng et al. (2011) reported that the nodes of riceplants distribute 40% of any As they receive to the closestleaf. The results presented here suggest that any arsenite dis-tributed to the flag leaf will remain there, while a significantamount of any DMA transported to the flag leaf will be re-translocated to the grain. The rapid reduction of arsenate toarsenite within the flag leaf means that any arsenate accu-mulated within the rice plant during growth will have beenreduced to arsenite and effectively immobilized. Therefore,arsenite and arsenate accumulating in the developing grainmust arrive via direct transport through the stem duringgrain fill, while DMA (and MMA) will be delivered to therice grain both from flag leaf stores and via the direct trans-port through the stem of any taken up during grain fill.
Spatial unloading of As
The 3D and 2D images of grains from the leaf-fed andexcised stem agree with previous images of As-fed rice grains(Carey et al., 2010). In addition, in Fig. 2, arsenite canclearly be seen in the OVT, closely localized, with Fe andMn apparently on each side of arsenite. Since phloem cellsare situated on the outside of the xylem cells in the OVT(Krishnan & Dayanandan, 2003), it is likely that arsenite islocalized in the phloem while Fe and Mn are present inboth phloem and xylem, although the resolution is not highenough to definitively conclude this. Also, Fig. 2 demon-strates that a portion of the As is being unloaded into thenucellar epidermis, albeit much less efficiently than DMA.
The concentration of inorganic As, particularly arsenite,within the bran layer of mature rice grain would suggest thatarsenite does not move beyond the nucellar epidermis intothe endosperm, as demonstrated for DMA (Meharg et al.,2008; Sun et al., 2008). Moore et al. (2010) showed that Aswas concentrated both in hotspots in the OVT region and inthe subaleurone layer that surrounds the endosperm for a ricegrain containing c. 30% inorganic As and 70% organic As.
Ge phloem ⁄ xylem mobility
The highly efficient translocation of germanic acid to theSi-accumulating tissues, particularly the husk, observed forexcised panicles, agrees with the role of germanic acid as asilicic acid analog. Stem-girdling demonstrated that phloemtransport is not important in germanic acid, and analo-gously, silicic acid translocation to the rice panicle. It maybe that, as germanic acid is extremely mobile in xylem,interruption of the percentage that is distributed to tissuesvia phloem is masked by the substantial xylem flow. In con-trast to ‘leaf-locked’ arsenite and phloem-immobile Sr,there was some translocation of germanic acid from treatedflag leaves to rice grain. This was limited and the markedlylower husk : grain ratio than that demonstrated by excisedstem-fed panicles indicated that this is not the primarypathway for germanic acid, and thus silicic acid, to thegrain. As background Ge in tissues is low, even trace leveltranslocation of germanic acid to grain can be detected, butit should be stressed that grain concentrations are 277-foldlower than those in the leaf. It may be that silicic and ger-manic acids are phloem-mobile, but at the flag leaf node ahigh proportion, if not all, is transferred to the xylem, andtherefore stem-girdling at the base of the panicle (above thenode) has no effect. Nodal vascular anastomoses in rice pan-icle nodes connect the enlarged vascular bundles to thesurrounding diffuse vascular bundles and this intervascularconnection is necessary for translocation to panicle tissues(Yamaji & Ma, 2009). Yamaji & Ma (2009) noted thatintervascular transport via nodal vascular anastomoses doesnot sufficiently account for the rapid accumulation of Si inrice husks and demonstrated that Lsi6 transporters, inxylem transfer cells, mediate Si transport from enlarged vas-cular bundles in the panicle node to the diffuse vascularbundles that transport material to the panicle.
With regard to differential mobility of As species, organicAs species are perhaps rapidly transferred via xylem transfercells from enlarged vascular bundles to diffuse vascular bun-dles, while arsenite, which is far less mobile in the xylem, isnot.
Arsenite-germanic acid competition
Germanic acid has been shown to be an effective analog ofsilicic acid in rice (Ma et al., 2002; Nikolic et al., 2007)
NewPhytologist Research 95
� 2011 The Authors
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011) 192: 87–98
www.newphytologist.com

and silicic acid transporters have been implicated in thetransport of arsenite (and methylated As species) into riceroots (Ma & Yamaji, 2008; Ma et al., 2008; Li et al.,2009). Germanic acid does not significantly reduce shoot tograin transport of arsenite, indicating that arsenite may notrely solely on silicic acid transporters for shoot-to-graintransport, or that competition is poor for these transporters,as may be the case for channels. In a study of field-grownrice cultivars, Norton et al. (2010) found a positive correla-tion between shoot As and shoot silicon but no correlationbetween shoot silicon and grain As concentrations. Maet al. (2008) showed that Lsi6 transporters are not involvedin arsenite uptake into the roots of rice plants, although thismay simply be the result of low expression of Lsi6 in riceroots. Arsenite–silicic acid competition may be significantin rice roots, where there is strong control on uptake intothe plant, but is perhaps less important in the rice shootwhere transport is largely driven by mass flow. When Liet al. (2009) found that silicic acid did not reduce rootuptake of MMA and DMA, they suggested that aquaporinssuch as Lsi1 may allow rapid fluxes of solutes so competi-tion will be less obvious (Maure et al., 2008; Li et al.,2009); this may also be the case for Lsi6 but further investi-gation is required.
In conclusion, this study demonstrates that DMA andMMA are extremely efficiently retranslocated, via thephloem, from the panicle flag leaf to the filling rice grain. Incontrast to inorganic As and MMA, DMA is extremelymobile within the grain, whether arriving via direct trans-port though the stem or retranslocation from flag leaves.Arsenate was relatively poorly retranslocated to the ricegrain and was rapidly reduced to arsenite within the flagleaf. Leaf-fed arsenite was not translocated to the rice grain,and was potentially sequestered into leaf cell vacuoles.Consequently, arsenite and arsenate accumulation in thedeveloping grain must occur solely via direct transportthrough the stem during grain fill, while DMA will also bedelivered to the rice grain from flag leaf stores. The resultsof this study also show that arsenite does not compete withgermanic acid for direct transport to the rice grain throughthe stem, suggesting that arsenite does not rely solely onsilicic acid transporters for shoot-to-grain translocation and,therefore, that shoot silicon status may not be important inrice grain As accumulation. Alternatively, arsenite–silicicacid competition is perhaps less obvious in the rice shootwhere transport is largely driven by mass flow and ⁄ or therate of transport through channels is too rapid to allowcompetition to take place.
Acknowledgements
This work was supported by a Biotechnology andBiological Sciences Research Council Doctoral Training
Grant and by grants from the National Institute ofEnvironmental Health Sciences, Superfund ResearchProgram (grant no. P42 ES007373-14) to T.P. andM.L.G. Its contents are solely the responsibility of theauthors and do not necessarily represent the official viewsof NIH. Portions of this work were performed atGeoSoilEnviroCARS (Sector 13), Advanced Photon Source(APS), Argonne National Laboratory. GeoSoilEnviroCARSis supported by the National Science Foundation – EarthSciences (EAR-0622171) and Department of Energy –Geosciences (DE-FG02-94ER14466). Use of the AdvancedPhoton Source was supported by the US Department ofEnergy, Office of Science, Office of Basic Energy Sciences,under contract no. DE-AC02-06CH11357. A portion ofthis work was performed at Beamline X26A, NationalSynchrotron Light Source (NSLS), Brookhaven NationalLaboratory. X26A is supported by the Department ofEnergy (DOE) – Geosciences (DE-FG02-92ER14244 toThe University of Chicago – CARS) and DOE – Office ofBiological and Environmental Research, EnvironmentalRemediation Sciences Div. (DE-FC09-96-SR18546 to theUniversity of Kentucky). Use of the NSLS was supported byDOE under contract no. DE-AC02-98CH10886. The USEnvironmental Protection Agency, through its Office ofResearch and Development, funded and managed a portionof the research; it has not been subject to Agency reviewand, therefore, does not necessarily reflect the views of theAgency, no official product endorsement should be inferred.We would like to express our sincere gratitude to NormanLittle and Dave Hadwen of the University of Aberdeen andto all the staff of Sector 13 at the APS for their support.
References
Abedin J, Cresser MS, Meharg AA, Feldmann J, Cotter-Howells J.
2002a. Arsenic accumulation and metabolism in rice (Oryza sativa L.).
Environmental Science and Technology 36: 962–968.
Abedin MJ, Feldmann J, Meharg AA. 2002b. Uptake kinetics of arsenic
species in rice plants. Plant Physiology 128: 1120–1128.
Adomako EE, Solaiman ARM, Williams PN, Deacon C, Rahman
GKMM, Meharg AA. 2009. Enhanced transfer of arsenic to grain for
Bangladesh grown rice compared to US and EU. EnvironmentInternational 35: 476–479.
Bleeker PM, Hakvoort HWJ, Bliek M, Souer E, Schat H. 2006.
Enhanced arsenate reduction by a CDC25-like tyrosine phosphatase
explains increased phytochelatin accumulation in arsenate-tolerant
Holcus lanatus. Plant Journal 45: 917–929.
Carey AM, Scheckel KG, Lombi E, Newville M, Choi Y, Norton GJ,
Charnock JM, Feldmann J, Price AH, Meharg AA. 2010. Grain
unloading of arsenic species in rice (Oryza sativa L.). Plant Physiology152: 309–319.
Chen F, Wu F, Dong J, Vincze E, Zhang G, Wang F, Huang Y, Wei K.
2007. Cadmium translocation and accumulation in developing barley
grains. Planta 227: 223–232.
Hansen HR, Raab A, Price AH, Duan G, Zhu Y, Norton GJ, Feldmann
J, Meharg AA. 2011. Identification of tetramethylarsonium in rice
96 Research
NewPhytologist
� 2011 The Authors
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011) 192: 87–98
www.newphytologist.com

grains with elevated arsenic content. Journal of EnvironmentalMonitoring 13: 32–34.
Kim SA, Punshon T, Lanzirotti A, Li A, Alonso JM, Ecker JR, Kaplan J,
Guerinot ML. 2006. Localization of iron in Arabidopsis seed requires
the vacuolar membrane transporter VIT1. Science 314: 1295–1298.
Krishnan S, Dayanandan P. 2003. Structural and histochemical studies on
grain-filling in the caryopsis of rice (Oryza sativa L.). Journal ofBiosciences 28: 455–469.
Kuppelwieser H, Feller U. 1991. Transport of Rb and Sr to the ear in
mature, excised shoots of wheat: effects of temperature and stem length
on Rb removal from the xylem. Plant and Soil 132: 281–288.
Li RY, Ago Y, Liu WJ, Mitani N, Feldmann J, McGrath SP, Ma JF, Zhao
FJ. 2009. The rice aquaporin lsi1 mediates uptake of methylated arsenic
species. Plant Physiology 150: 2071–2080.
Lombi E, Scheckel KG, Pallon J, Carey AM, Zhu YG, Meharg AA. 2009.
Speciation and distribution of arsenic and localization of nutrients in
rice grains. New Phytologist 184: 193–201.
Lu Y, Adomako EE, Solaiman ARM, Islam MR, Deacon C, Williams
PN, Rahman GKMM, Meharg AA. 2009. Baseline soil variation is a
major factor in arsenic accumulation in Bengal delta paddy rice.
Environmental Science and Technology 43: 1724–1729.
Ma JF, Tamai K, Ichii M, Wu GF. 2002. A rice mutant defective in Si
uptake. Plant Physiology 130: 2111–2117.
Ma JF, Yamaji N. 2008. Functions and transport of silicon in plants.
Cellular and Molecular Life Sciences 65: 3049–3057.
Ma JF, Yamaji N, Mitani N, Xu XY, Su YH, McGrath SP, Zhao FJ.
2008. Transporters of arsenite in rice and their role in arsenic
accumulation in rice grain. Proceedings of the National Academy ofScience, USA 105: 9931–9935.
Marcus Wyner L, Rains DW. 1982. Uptake, accumulation, and
translocation of arsenical compounds by cotton. Journal ofEnvironmental Quality 11: 715–719.
Marin AR, Masscheleyn PH, Patrick WH Jr. 1993. Soil redox-pH
stability of arsenic species and its influence on arsenic uptake by rice.
Plant and Soil 152: 245–253.
Martin P. 1982. Stem xylem as a possible pathway for mineral
retranslocation from senescing leaves to the ear in wheat. AustralianJournal of Plant Physiology 9: 197–207.
Mastradone PJ, Woolson EA. 1983. Levels of arsenical species in cotton
after field application of a cacodylic acid defoliant. Bulletin ofEnvironmental Contamination and Toxicology 31: 216–221.
Maure C, Verdoucq L, Luu DT, Santoni V. 2008. Plant aquaporins:
membrane channels with multiple integrated functions. Annual Reviewof Plant Biology 59: 595–624.
McNear DH Jr, Peltier E, Everhart J, Chaney RL, Sutton S, Newville M,
Rivers M, Sparks DL. 2005. Application of quantitative fluorescence
and absorption-edge computed microtomography to image metal
compartmentalization in Alyssum murale. Environmental Science andTechnology 39: 2210–2218.
Meharg AA, Lombi E, Williams PN, Scheckel KG, Feldmann J, Raab A,
Zhu YG, Islam R. 2008. Speciation and localization of arsenic in white
and brown rice grains. Environmental Science and Technology 42:
1051–1057.
Meharg AA, Williams PN, Adomako E, Lawgali YY, Deacon C, Villada
A, Cambell RCJ, Sun GX, Zhu YG, Feldmann J et al. 2009.
Geographical variation in total and inorganic arsenic content of polished
(white) rice. Environmental Science and Technology 43: 1612–1617.
Moore KL, Schroder M, Lombi E, Zhao FJ, McGrath SP, Hawkesford
MJ, Shewry PR & Grovenor CRM. 2010. NanoSIMS analysis of
arsenic and selenium in cereal grain. New Phytologist 185: 434–445.
Nikolic M, Nikolic N, Liang Y, Kirkby EA, Romheld V. 2007.
Germanium-68 as an adequate tracer for silicon transport in plants.
Characterization of silicon uptake in different crop species. PlantPhysiology 143: 495–503.
Norton GJ, Islam MR, Deacon C, Zhao FJ, Stroud JL, Mcgrath SP,
Islam S, Jahiruddin M, Feldmann J, Price AH et al. 2009.
Identification of low inorganic and total grain arsenic rice cultivars from
Bangladesh. Environmental Science and Technology 43: 6070–6075.
Norton GJ, Islam MR, Duan G, Lei M, Zhu YG, Deacon CM, Moran
AC, Islam S, Zhao FJ, Stroud JL et al. 2010. Arsenic shoot-grain
relationships in field grown rice cultivars. Environmental Science andTechnology 44: 1471–1477.
Oparka KJ, Gates P. 1981. Transport of assimilates in the developing
caryopsis of rice (Oryza sativa L.) – The pathways of water and
assimilated carbon. Planta 152: 388–396.
Raab A, Williams PN, Meharg AA, Feldmann J. 2007. Uptake and
translocation of inorganic and methylated arsenic species by plants.
Environmental Chemistry 4: 197–203.
Rosen BP, Liu Z. 2009. Transport pathways for arsenic and selenium: a
minireview. Environment International 35: 512–515.
Song WY, Park J, Mendoza-Cozatl DG, Suter-Grotemeyer M, Shima D,
Hortensteiner S, Geisler M, Weder B, Rea PA, Rentsch D et al. 2010.
Arsenic tolerance in Arabidopsis is mediated by two ABCC-type
phytochelatin transporters. Proceedings of the National Academy ofSciences, USA 107: 21187–21192.
Su YH, McGrath SP, Zhao FJ. 2010. Rice is more efficient in arsenite
uptake and translocation than wheat and barley. Plant and Soil 328:
27–34.
Sun GX, Williams PN, Carey AM, Zhu YG, Deacon C, Raab A,
Feldmann J, Islam RM, Meharg AA. 2008. Inorganic arsenic in rice
bran and its products are an order of magnitude higher than in bulk
grain. Environmental Science and Technology 42: 7542–7546.
Sun GX, Williams PN, Zhu YG, Deacon C, Carey AM, Raab A,
Feldmann J, Meharg AA. 2009. Survey of arsenic and its speciation in
rice products such as breakfast cereals, rice crackers and Japanese rice
condiments. Environment International 35: 473–475.
Valette-Silver NJ, Riedel GF, Crecelius EA, Windom H, Smith RG,
Dolvin SS. 1999. Elevated arsenic concentrations in bivalves from the
southeast coasts of the USA. Marine Environmental Research 48:
311–333.
Williams PN, Price AH, Raab A, Hossain SA, Feldmann J, Meharg AA.
2005. Variation in arsenic speciation and concentration in paddy rice
related to dietary exposure. Environmental Science and Technology 39:
5531–5540.
Williams PN, Villada A, Deacon C, Raab A, Figuerola J, Green AJ,
Feldmann J, Meharg AA. 2007. Greatly enhanced arsenic shoot
assimilation in rice leads to elevated grain levels compared to wheat and
barley. Environmental Science and Technology 41: 6854–6859.
Xu XY, McGrath SP, Meharg AA, Zhao FJ. 2008. Growing rice
aerobically markedly decreases arsenic accumulation. EnvironmentalScience and Technology 42: 5574–5579.
Yamaji N, Ma JF. 2009. A transporter at the node responsible for
intervascular transfer of silicon in rice. Plant Cell 21: 2878–2883.
Ye WL, Wood BA, Stroud JL, Andralojc PJ, Raab A, McGrath SP,
Feldmann J, Zhao FJ. 2010. Arsenic speciation in phloem and xylem
exudates of castor bean. Plant Physiology 154: 1505–1513.
Zhao FJ, Ma JF, Meharg AA, McGrath SP. 2009. Arsenic uptake and
metabolism in plants. New Phytologist 181: 777–794.
Zhao FJ, McGrath SP, Meharg AA. 2010. Arsenic as a food chain
contaminant: mechanisms of plant uptake and metabolism and
mitigation strategies. Annual Review of Plant Biology 61: 535–559.
Zheng MZ, Cai C, Hu Y, Sun GX, Williams PN, Cui HJ, Li G, Zhao FJ,
Zhu YG. 2011. Spatial distribution of arsenic and temporal variation of
its concentration in rice. New Phytologist 189: 200–209.
NewPhytologist Research 97
� 2011 The Authors
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011) 192: 87–98
www.newphytologist.com

Supporting Information
Additional supporting information may be found in theonline version of this article.
Fig. S1 Mean total concentrations of rubidium (Rb) andstrontium (Sr) in flag leaf and grain, for rice paniclesexposed to 333 lM of one of five arsenic treatments,together with 1 mM Rb and Sr, or, in the case of the zeroexposure controls, ultrapure, MilliQ water only.
Fig. S2 Dose–response curves showing mean arsenicconcentrations in grain and flag leaf for panicles exposed toincreasing levels of arsenite or DMA (0, 33, 133 and333 lM) via the cut flag leaf on intact plants.
Fig. S3 Mean total solution uptake for panicles treated with0, 33, 133 and 333 lM arsenite or DMA via the cut flagleaf on intact plants.
Fig. S4 Chromatograms showing arsenic speciation in freshleaf extracts of arsenite- and arsenate-fed flag leaves togetherwith a standard mix.
Fig. S5 Chromatograms showing arsenic speciation in leaffeeding vials of arsenite and arsenate.
Fig. S6 Fluorescence microtomography images showingdistributions of arsenic for virtual cross-sections of rice grainexposed to arsenite, arsenate, MMA or DMA via the excisedpanicle stem or the cut flag leaf on intact plants.
Fig. S7 Mean total concentrations of Rb and Sr in ricegrain, husk and flag leaf for excised rice panicles subjectedto a stem-girdling treatment and hydroponically fed, over a48 h period, nutrient solution.
Methods S1 Phloem transport of arsenic species from flagleaf to grain during grain filling.
Please note: Wiley-Blackwell are not responsible for thecontent or functionality of any supporting informationsupplied by the authors. Any queries (other than missingmaterial) should be directed to the New Phytologist CentralOffice.
98 Research
NewPhytologist
� 2011 The Authors
New Phytologist � 2011 New Phytologist Trust
New Phytologist (2011) 192: 87–98
www.newphytologist.com










![pkS/kjh j.kchj flag fo'ofo|ky;]thUn - CRSU](https://img.pdfslide.net/doc/110x75/633d9c9a5fdfc7998b01fac0/pkskjh-jkchj-flag-foofokythun-crsu.jpg)


















