Embed Size (px)
Citation preview

Molecular Phylogenetics and Evolution 65 (2012) 149–162
Contents lists available at SciVerse ScienceDirect
Molecular Phylogenetics and Evolution
journal homepage: www.elsevier .com/ locate /ympev
Phylogeny reconstruction in the Caesalpinieae grade (Leguminosae) basedon duplicated copies of the sucrose synthase gene and plastid markers
Vincent Manzanilla, Anne Bruneau ⇑Institut de recherche en biologie végétale, Département de Sciences biologiques, Université de Montréal, 4101 Sherbrooke est, Montréal, Québec, Canada H1X 2B2
a r t i c l e i n f o
Article history:Received 29 July 2011Revised 30 May 2012Accepted 31 May 2012Available online 12 June 2012
Keywords:CaesalpinieaeCaesalpinioideaeLeguminosaeNodulationNuclear genesPhylogenySucrose synthase (SUSY)
1055-7903/$ - see front matter � 2012 Elsevier Inc. Ahttp://dx.doi.org/10.1016/j.ympev.2012.05.035
⇑ Corresponding author. Fax: +1 514 343 2288.E-mail addresses: [email protected] (V.
umontreal.ca (A. Bruneau).
a b s t r a c t
The Caesalpinieae grade (Leguminosae) forms a morphologically and ecologically diverse group of mostlytropical tree species with a complex evolutionary history. This grade comprises several distinct lineages,but the exact delimitation of the group relative to subfamily Mimosoideae and other members of subfam-ily Caesalpinioideae, as well as phylogenetic relationships among the lineages are uncertain. With theaim of better resolving phylogenetic relationships within the Caesalpinieae grade, we investigated theutility of several nuclear markers developed from genomic studies in the Papilionoideae. We clonedand sequenced the low copy nuclear gene sucrose synthase (SUSY) and combined the data with plastidtrnL and matK sequences. SUSY has two paralogs in the Caesalpinieae grade and in the Mimosoideae,but occurs as a single copy in all other legumes tested. Bayesian and maximum likelihood phylogeneticanalyses suggest the two nuclear markers are congruent with plastid DNA data. The Caesalpinieae gradeis divided into four well-supported clades (Cassia, Caesalpinia, Tachigali and Peltophorum clades), apoorly supported clade of Dimorphandra Group genera, and two paraphyletic groups, one with otherDimorphandra Group genera and the other comprising genera previously recognized as the Umtiza clade.A selection analysis of the paralogs, using selection models from PAML, suggests that SUSY genes are sub-jected to a purifying selection. One of the SUSY paralogs, under slightly stronger positive selection, maybe undergoing subfunctionalization. The low copy SUSY gene is useful for phylogeny reconstruction inthe Caesalpinieae despite the presence of duplicate copies. This study confirms that the Caesalpinieaegrade is an artificial group, and highlights the need for further analyses of lineages at the base of theMimosoideae.
� 2012 Elsevier Inc. All rights reserved.
1. Introduction
The evolutionary history of subfamily Caesalpinioideae in theLeguminosae remains poorly understood despite numerous recentanalyses of relationships within the subfamily and in the legumefamily as a whole (Doyle, 1994; Chappill, 1995; Käss and Wink,1996; Doyle et al., 1997, 2000; Kajita et al., 2001; Doyle and Luckow,2003; Herendeen et al., 2003a; Wojciechowski et al., 2004; Lavinet al., 2005; Bruneau et al., 2001, 2008). As described recently by Le-wis et al. (2005) subfamily Caesalpinioideae is paraphyletic; it in-cludes the monophyletic tribes Cercideae and Detarieae, and theparaphyletic tribes Cassieae and Caesalpinieae. Among these tribes,the Caesalpinieae is particularly problematic. Polhill and Vidal(1981) divided the Caesalpinieae into eight informal generic groups(Table 1), which were modified to nine groups by Polhill (1994).Tribe Caesalpinieae has since generally been resolved as paraphylet-
ll rights reserved.
Manzanilla), anne.bruneau@
ic and intergeneric relationships within the nine generic groupshave been adjusted with developing knowledge of the tribe (Lewisand Schrire, 1995; Du Puy et al., 1995; Doyle et al., 1997; Lewis,1998; Bruneau et al., 2001; Simpson and Lewis, 2003; Simpsonet al., 2004; Simpson and Ulibarri, 2006; Table 1). In their recentanalyses of subfamily Caesalpinioideae based on plastid sequences,Bruneau et al. (2008) found this tribe to comprise six informal cladesthat previously mostly were included in tribe Caesalpinieae (theUmtiza, Caesalpinia, Tachigali, Peltophorum, and two Dimorphan-dra Group lineages), as well as the Cassia clade (subtribe Cassiinae;Table 1), the latter historically placed in tribe Cassieae. Here we fo-cus on these seven lineages, which for simplicity will hereafter be re-ferred to as the Caesalpinieae grade. In addition, because thecaesalpinioid Dimorphandra Group (Polhill, 1994) shares numerousfloral, pollen and wood features with subfamily Mimosoideae andhas in the past been considered taxonomically as a ‘‘transitionallink’’ between the caesalpinioids and mimosoids (Polhill and Vidal,1981; Luckow et al., 2000, 2003), another interest of this study isto better determine the taxonomic delimitation of the Mimosoideaerelative to the Caesalpinioideae. In short, some caesalpinioid generamay best be placed within the Mimosoideae or may belong among
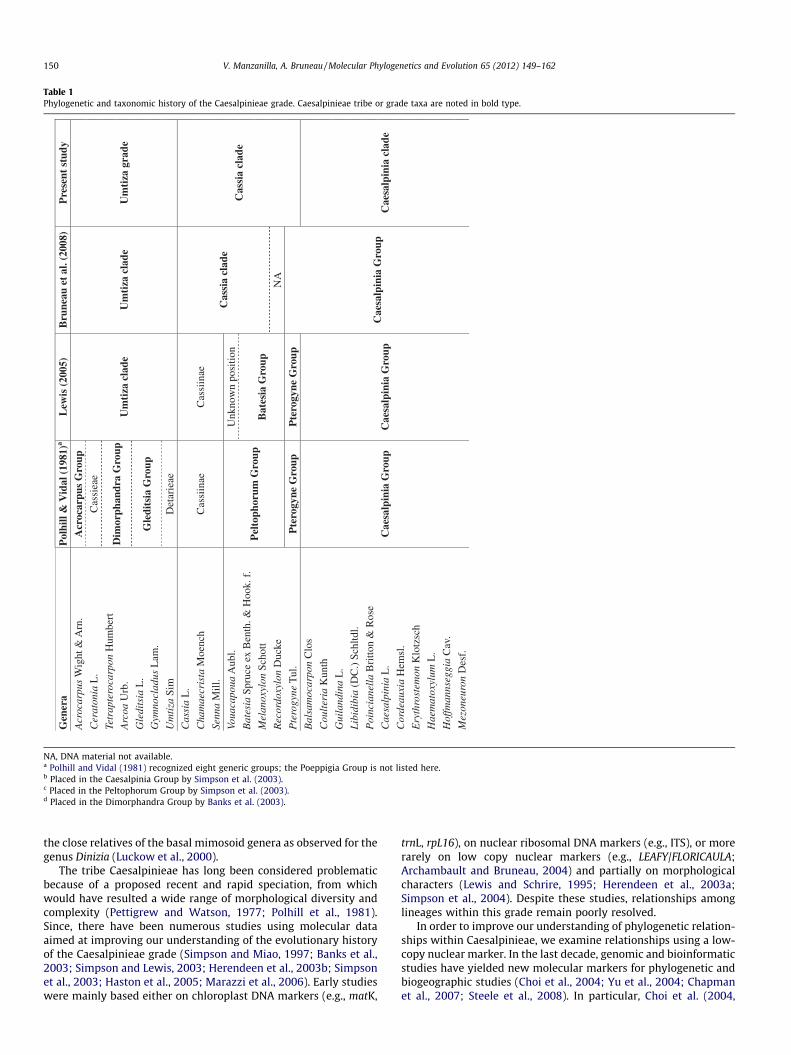
Table 1Phylogenetic and taxonomic history of the Caesalpinieae grade. Caesalpinieae tribe or grade taxa are noted in bold type.
NA, DNA material not available.a Polhill and Vidal (1981) recognized eight generic groups; the Poeppigia Group is not listed here.b Placed in the Caesalpinia Group by Simpson et al. (2003).c Placed in the Peltophorum Group by Simpson et al. (2003).d Placed in the Dimorphandra Group by Banks et al. (2003).
150 V. Manzanilla, A. Bruneau / Molecular Phylogenetics and Evolution 65 (2012) 149–162
the close relatives of the basal mimosoid genera as observed for thegenus Dinizia (Luckow et al., 2000).
The tribe Caesalpinieae has long been considered problematicbecause of a proposed recent and rapid speciation, from whichwould have resulted a wide range of morphological diversity andcomplexity (Pettigrew and Watson, 1977; Polhill et al., 1981).Since, there have been numerous studies using molecular dataaimed at improving our understanding of the evolutionary historyof the Caesalpinieae grade (Simpson and Miao, 1997; Banks et al.,2003; Simpson and Lewis, 2003; Herendeen et al., 2003b; Simpsonet al., 2003; Haston et al., 2005; Marazzi et al., 2006). Early studieswere mainly based either on chloroplast DNA markers (e.g., matK,
trnL, rpL16), on nuclear ribosomal DNA markers (e.g., ITS), or morerarely on low copy nuclear markers (e.g., LEAFY/FLORICAULA;Archambault and Bruneau, 2004) and partially on morphologicalcharacters (Lewis and Schrire, 1995; Herendeen et al., 2003a;Simpson et al., 2004). Despite these studies, relationships amonglineages within this grade remain poorly resolved.
In order to improve our understanding of phylogenetic relation-ships within Caesalpinieae, we examine relationships using a low-copy nuclear marker. In the last decade, genomic and bioinformaticstudies have yielded new molecular markers for phylogenetic andbiogeographic studies (Choi et al., 2004; Yu et al., 2004; Chapmanet al., 2007; Steele et al., 2008). In particular, Choi et al. (2004,

V. Manzanilla, A. Bruneau / Molecular Phylogenetics and Evolution 65 (2012) 149–162 151
2006) and Scherson et al. (2005) developed single copy nuclearphylogenetic markers for the Leguminosae based on genomic com-parisons. They provided a wide range of markers with putativelyconserved exonic regions and highly variable introns. Based onthese studies, we tested several single copy nuclear markers in or-der to resolve the complex evolutionary history of the Cae-salpinieae grade. We were particularly interested in the SUSYlocus, which is a part of the sucrose synthase gene (SUSY, SUS,SucS). This gene has a key role in crop plant metabolism, becauseit transforms sucrose into starch. Recent studies also have shownits implication in the functionality of the root nodules in legumes(Silvente et al., 2003; Baier et al., 2007; Horst et al., 2007). SUSYis necessary in the metabolism of legumes for the establishmentand maintenance of an efficient N-fixing symbiosis (Baier et al.,2007). Nitrogen fixation is commonly present in the Papilionoi-deae, but in the Caesalpinioideae is known to occur only in certainmembers of the Caesalpinieae grade (i.e., confirmed in CampsiandraBenth., Chamaecrista Moench, Dimorphandra Schott, ErythrophleumAfzel. ex R.Br., Melanoxylon Schott, Moldenhawera Schrad. and Tach-igali Aubl. (including Sclerolobium Vogel), and possibly in Recordox-ylon Ducke and Vouacapoua Aubl.; Sprent, 2001, 2009).
Based on previous studies, we postulate that the Caesalpinieaegrade consists of seven groups and test this hypothesis using bothnuclear and plastid markers. As part of this study, we evaluate theutility of seven single copy nuclear genes as phylogenetic markersfor further understanding the complex evolutionary history of lin-eages within this grade. Phylogenetic relationships are studiedusing the low-copy nuclear sucrose synthase locus, and the nucleardata are combined with two chloroplast loci, trnL and matK. Todetermine whether the SUSY gene evolved under the same evolu-tionary pressure within the Papilionoideae and the Caesalpinieaegrade, we examine the evolution of the SUSY gene based uponthe topology obtained.
2. Materials and methods
2.1. Taxon sampling
We sampled 50 of the 56 genera of the Caesalpinieae sensu Le-wis (2005; Table 2). The genera Stenodrepanum Harms, Lophocarpi-nia Bukart, Sympetalandra Stapf and Orphanodendron Barneby &J.W. Grimes are not included because we could not obtain leafmaterial; two others are not included because of poor quality ofthe material (Zuccagnia Cav. and Caesalpinia L.). Genera from sub-tribe Cassiinae (Cassieae) were also included in the study (Chamae-crista, Senna Mill., Cassia L.), as was Dinizia Ducke (Mimosoideae)because previous studies placed them as derived within the Cae-salpinieae grade (Bruneau et al., 2001, 2008). Adenanthera L., Calpo-calyx Harms, Cylicodiscus Harms, Entada Adans., PentaclethraBenth., Piptadenia Benth., and Parkia R.Br. were selected to repre-sent the basal Mimosoideae (Luckow et al., 2003). Two genera fromthe Dialiinae clade and nine from subfamily Papilionoideae werechosen as outgroup taxa. Most of the genera are represented by asingle species and each species by up to three individuals. Weuse the generic delimitation of Lewis et al. (2005). Most samplesare from fresh leaf material preserved in silica gel.
2.2. Loci sequenced
We selected nuclear markers developed by Choi et al. (2004,2006), Scherson et al. (2005) and J.J. Doyle (Cornell University,unpublished data). The objective of our analyses being phylogenyreconstruction at and below the subfamily level, we selected mark-ers with a relatively large coding region because we consideredthat the structure of the exon would be better preserved within
the large Caesalpinieae grade and that the intron sequences wouldbe too variable at this phylogenetic level. We evaluated the vari-ability of seven markers (Table 3) for a restricted sampling of 12Caesalpinieae grade taxa (Table 2). After sequencing, we manuallyedited chromatograms and performed a sequence similarity search(Blastn) (Altschul et al., 1997) on GenBank to ensure correspon-dence with the targeted locus. We aligned the sequences withthe Sequence-Tagged Sites (STS) from Choi et al. (2004) and DNAsequences from Scherson et al. (2005). We performed preliminaryBayesian analyses to test for paralogs and to establish marker var-iability within the grade. Following these initial surveys, we pur-sued our phylogenetic analyses with SUSY and PP1, butsubsequently only SUSY was retained for further analyses (see Re-sults). In addition, we sequenced the chloroplast trnL and matK locifor several taxa which had not been studied by Bruneau et al.(2008) (Table 2). The potential marker saturation for PP1 and SUSYwas assessed using a substitution saturation test (Xia et al., 2003),implemented in DAMBE version 5.0.52 (Xia and Xie, 2001).
2.3. DNA extraction, amplification and sequencing
DNA was isolated from leaf tissue using a modified CTAB proto-col (Joly et al., 2006) or the Qiagen Dneasy™ Kit (QIAGEN, Missis-sauga, Ontario, Canada). For the trnL region, the primers, PCRreaction mix, amplification, and sequencing procedures followedthose described by Bruneau et al. (2001). The matK region wasamplified using the conditions optimized by Bruneau et al. (2008).
Several of the primers for the nuclear markers, initially devel-oped for the Papilionoideae, were redesigned and optimized asspecific primers for the Caesalpinioideae, including those for SUSYand PP1 (Table 3). Using BioEdit 7.0.5.3 (Hall, 1999), we aligned allpublicly available SUSY sequence-tag sites (STS), express sequencetags (ESTs) and genomic sequences from the Leguminosae avail-able, as well as outgroup sequences from the Brassicaceae Arabid-opsis thaliana (L.) Heynh. Based on this alignment, conservedregions were identified and new primers were designed usingthe Amplify program version 3.1.4 (Engels, 2005). These new prim-ers increased the length of the targeted region for SUSY and PP1 by,respectively 211 bp and 209 bp, relative to the original fragmentsequenced by Choi et al. (2004).
The PCR amplification reaction mix for the nuclear markers con-tained 1� Roche Diagnostics Buffer (Laval, Quebec, Canada),100 lmol/L dNTP, 0.3 mmol/L primers, 1 unit Taq DNA polymerase,50–150 ng genomic DNA, and 2.5–5% Dimethyl sulfoxide (DMSO),0.05% Tween 20, or 2.5 lg Bovine serum albumin (BSA), toppedwith distilled water for a final reaction volume of 25 ll. Amplifica-tions were conducted with a ‘‘hot start’’ cycle (Taq DNA polymeraseadded at 95 �C), followed by 40 cycles consisting of 3 min denatur-ation at 95 �C, annealing at 48–64 �C (depending on the species) for30 s, and an extension step at 72 �C for 3 min. The sequencingreaction ended with an extension period at 10 �C. The PCR mixwas aliquoted into three separate PCR tubes prior to thermocyclingin order to minimize PCR recombination errors (Joly et al., 2006)and negative controls with no genomic DNA template were runon all amplifications.
The presence of polymorphisms in initial sequences obtainedfrom direct sequencing suggested that cloning was required.Therefore, all PCR products were cloned using a PGEM-T vectorand transformed into chemically competent E. coli DH5-a. Thetransformed bacteria were screened on selective solid LB mediacontaining 50 mg/ml kanamycin and 100 mg/ml ampicillin, andkept overnight at 37 �C. For each PCR product, six to 24 colonieswere selected. The positive colonies were grown overnight in LBbroth and then amplified. For each clone, PCR reactions first wereperformed to verify for the presence of the full-length desired se-quence, and then purified and sequenced. The amplification and
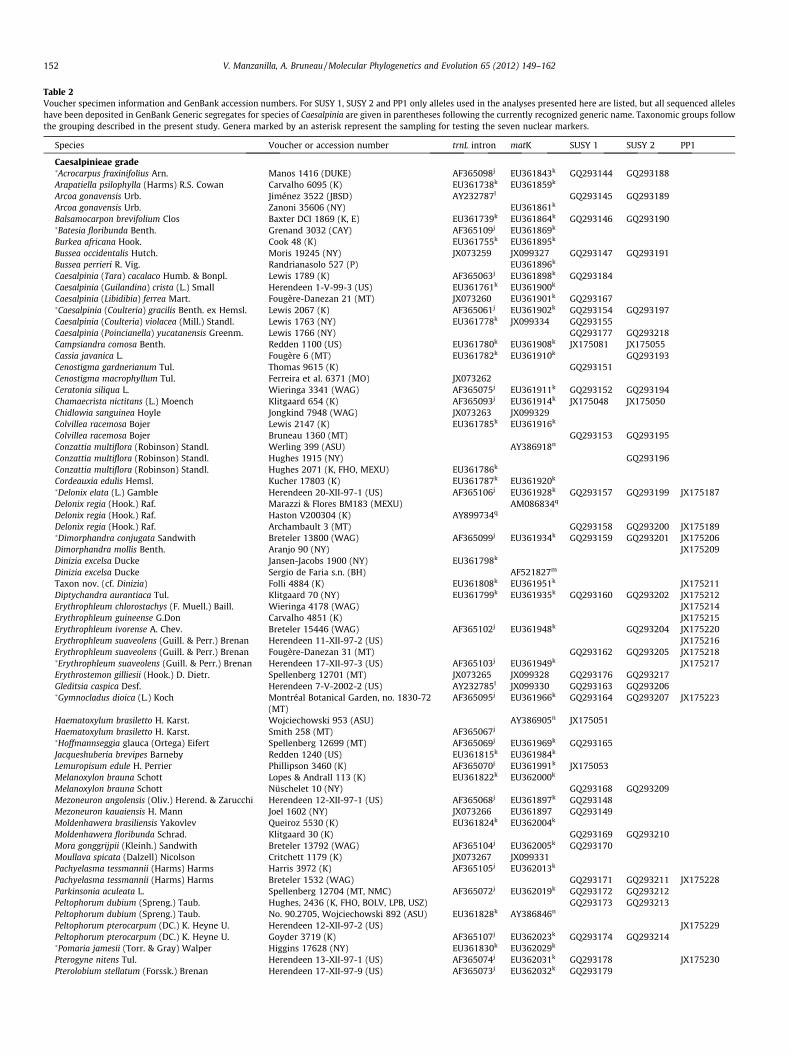
Table 2Voucher specimen information and GenBank accession numbers. For SUSY 1, SUSY 2 and PP1 only alleles used in the analyses presented here are listed, but all sequenced alleleshave been deposited in GenBank Generic segregates for species of Caesalpinia are given in parentheses following the currently recognized generic name. Taxonomic groups followthe grouping described in the present study. Genera marked by an asterisk represent the sampling for testing the seven nuclear markers.
Species Voucher or accession number trnL intron matK SUSY 1 SUSY 2 PP1
Caesalpinieae grade�Acrocarpus fraxinifolius Arn. Manos 1416 (DUKE) AF365098j EU361843k GQ293144 GQ293188Arapatiella psilophylla (Harms) R.S. Cowan Carvalho 6095 (K) EU361738k EU361859k
Arcoa gonavensis Urb. Jiménez 3522 (JBSD) AY232787l GQ293145 GQ293189Arcoa gonavensis Urb. Zanoni 35606 (NY) EU361861k
Balsamocarpon brevifolium Clos Baxter DCI 1869 (K, E) EU361739k EU361864k GQ293146 GQ293190�Batesia floribunda Benth. Grenand 3032 (CAY) AF365109j EU361869k
Burkea africana Hook. Cook 48 (K) EU361755k EU361895k
Bussea occidentalis Hutch. Moris 19245 (NY) JX073259 JX099327 GQ293147 GQ293191Bussea perrieri R. Vig. Randrianasolo 527 (P) EU361896k
Caesalpinia (Tara) cacalaco Humb. & Bonpl. Lewis 1789 (K) AF365063j EU361898k GQ293184Caesalpinia (Guilandina) crista (L.) Small Herendeen 1-V-99-3 (US) EU361761k EU361900k
Caesalpinia (Libidibia) ferrea Mart. Fougère-Danezan 21 (MT) JX073260 EU361901k GQ293167�Caesalpinia (Coulteria) gracilis Benth. ex Hemsl. Lewis 2067 (K) AF365061j EU361902k GQ293154 GQ293197Caesalpinia (Coulteria) violacea (Mill.) Standl. Lewis 1763 (NY) EU361778k JX099334 GQ293155Caesalpinia (Poincianella) yucatanensis Greenm. Lewis 1766 (NY) GQ293177 GQ293218Campsiandra comosa Benth. Redden 1100 (US) EU361780k EU361908k JX175081 JX175055Cassia javanica L. Fougère 6 (MT) EU361782k EU361910k GQ293193Cenostigma gardnerianum Tul. Thomas 9615 (K) GQ293151Cenostigma macrophyllum Tul. Ferreira et al. 6371 (MO) JX073262Ceratonia siliqua L. Wieringa 3341 (WAG) AF365075j EU361911k GQ293152 GQ293194Chamaecrista nictitans (L.) Moench Klitgaard 654 (K) AF365093j EU361914k JX175048 JX175050Chidlowia sanguinea Hoyle Jongkind 7948 (WAG) JX073263 JX099329Colvillea racemosa Bojer Lewis 2147 (K) EU361785k EU361916k
Colvillea racemosa Bojer Bruneau 1360 (MT) GQ293153 GQ293195Conzattia multiflora (Robinson) Standl. Werling 399 (ASU) AY386918n
Conzattia multiflora (Robinson) Standl. Hughes 1915 (NY) GQ293196Conzattia multiflora (Robinson) Standl. Hughes 2071 (K, FHO, MEXU) EU361786k
Cordeauxia edulis Hemsl. Kucher 17803 (K) EU361787k EU361920k
�Delonix elata (L.) Gamble Herendeen 20-XII-97-1 (US) AF365106j EU361928k GQ293157 GQ293199 JX175187Delonix regia (Hook.) Raf. Marazzi & Flores BM183 (MEXU) AM086834q
Delonix regia (Hook.) Raf. Haston V200304 (K) AY899734q
Delonix regia (Hook.) Raf. Archambault 3 (MT) GQ293158 GQ293200 JX175189�Dimorphandra conjugata Sandwith Breteler 13800 (WAG) AF365099j EU361934k GQ293159 GQ293201 JX175206Dimorphandra mollis Benth. Aranjo 90 (NY) JX175209Dinizia excelsa Ducke Jansen-Jacobs 1900 (NY) EU361798k
Dinizia excelsa Ducke Sergio de Faria s.n. (BH) AF521827m
Taxon nov. (cf. Dinizia) Folli 4884 (K) EU361808k EU361951k JX175211Diptychandra aurantiaca Tul. Klitgaard 70 (NY) EU361799k EU361935k GQ293160 GQ293202 JX175212Erythrophleum chlorostachys (F. Muell.) Baill. Wieringa 4178 (WAG) JX175214Erythrophleum guineense G.Don Carvalho 4851 (K) JX175215Erythrophleum ivorense A. Chev. Breteler 15446 (WAG) AF365102j EU361948k GQ293204 JX175220Erythrophleum suaveolens (Guill. & Perr.) Brenan Herendeen 11-XII-97-2 (US) JX175216Erythrophleum suaveolens (Guill. & Perr.) Brenan Fougère-Danezan 31 (MT) GQ293162 GQ293205 JX175218�Erythrophleum suaveolens (Guill. & Perr.) Brenan Herendeen 17-XII-97-3 (US) AF365103j EU361949k JX175217Erythrostemon gilliesii (Hook.) D. Dietr. Spellenberg 12701 (MT) JX073265 JX099328 GQ293176 GQ293217Gleditsia caspica Desf. Herendeen 7-V-2002-2 (US) AY232785l JX099330 GQ293163 GQ293206�Gymnocladus dioica (L.) Koch Montréal Botanical Garden, no. 1830-72
(MT)AF365095j EU361966k GQ293164 GQ293207 JX175223
Haematoxylum brasiletto H. Karst. Wojciechowski 953 (ASU) AY386905n JX175051Haematoxylum brasiletto H. Karst. Smith 258 (MT) AF365067j
�Hoffmannseggia glauca (Ortega) Eifert Spellenberg 12699 (MT) AF365069j EU361969k GQ293165Jacqueshuberia brevipes Barneby Redden 1240 (US) EU361815k EU361984k
Lemuropisum edule H. Perrier Phillipson 3460 (K) AF365070j EU361991k JX175053Melanoxylon brauna Schott Lopes & Andrall 113 (K) EU361822k EU362000k
Melanoxylon brauna Schott Nüschelet 10 (NY) GQ293168 GQ293209Mezoneuron angolensis (Oliv.) Herend. & Zarucchi Herendeen 12-XII-97-1 (US) AF365068j EU361897k GQ293148Mezoneuron kauaiensis H. Mann Joel 1602 (NY) JX073266 EU361897 GQ293149Moldenhawera brasiliensis Yakovlev Queiroz 5530 (K) EU361824k EU362004k
Moldenhawera floribunda Schrad. Klitgaard 30 (K) GQ293169 GQ293210Mora gonggrijpii (Kleinh.) Sandwith Breteler 13792 (WAG) AF365104j EU362005k GQ293170Moullava spicata (Dalzell) Nicolson Critchett 1179 (K) JX073267 JX099331Pachyelasma tessmannii (Harms) Harms Harris 3972 (K) AF365105j EU362013k
Pachyelasma tessmannii (Harms) Harms Breteler 1532 (WAG) GQ293171 GQ293211 JX175228Parkinsonia aculeata L. Spellenberg 12704 (MT, NMC) AF365072j EU362019k GQ293172 GQ293212Peltophorum dubium (Spreng.) Taub. Hughes, 2436 (K, FHO, BOLV, LPB, USZ) GQ293173 GQ293213Peltophorum dubium (Spreng.) Taub. No. 90.2705, Wojciechowski 892 (ASU) EU361828k AY386846n
Peltophorum pterocarpum (DC.) K. Heyne U. Herendeen 12-XII-97-2 (US) JX175229Peltophorum pterocarpum (DC.) K. Heyne U. Goyder 3719 (K) AF365107j EU362023k GQ293174 GQ293214�Pomaria jamesii (Torr. & Gray) Walper Higgins 17628 (NY) EU361830k EU362029k
Pterogyne nitens Tul. Herendeen 13-XII-97-1 (US) AF365074j EU362031k GQ293178 JX175230Pterolobium stellatum (Forssk.) Brenan Herendeen 17-XII-97-9 (US) AF365073j EU362032k GQ293179
152 V. Manzanilla, A. Bruneau / Molecular Phylogenetics and Evolution 65 (2012) 149–162
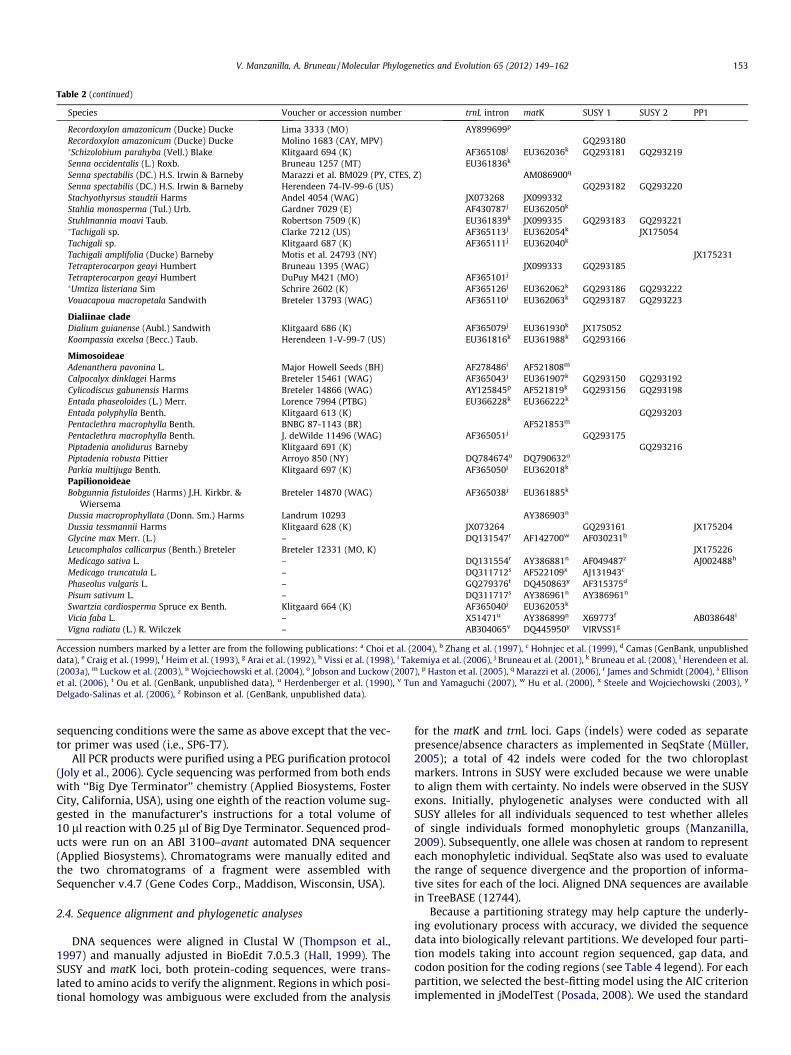
Table 2 (continued)
Species Voucher or accession number trnL intron matK SUSY 1 SUSY 2 PP1
Recordoxylon amazonicum (Ducke) Ducke Lima 3333 (MO) AY899699p
Recordoxylon amazonicum (Ducke) Ducke Molino 1683 (CAY, MPV) GQ293180�Schizolobium parahyba (Vell.) Blake Klitgaard 694 (K) AF365108j EU362036k GQ293181 GQ293219Senna occidentalis (L.) Roxb. Bruneau 1257 (MT) EU361836k
Senna spectabilis (DC.) H.S. Irwin & Barneby Marazzi et al. BM029 (PY, CTES, Z) AM086900q
Senna spectabilis (DC.) H.S. Irwin & Barneby Herendeen 74-IV-99-6 (US) GQ293182 GQ293220Stachyothyrsus staudtii Harms Andel 4054 (WAG) JX073268 JX099332Stahlia monosperma (Tul.) Urb. Gardner 7029 (E) AF430787j EU362050k
Stuhlmannia moavi Taub. Robertson 7509 (K) EU361839k JX099335 GQ293183 GQ293221�Tachigali sp. Clarke 7212 (US) AF365113j EU362054k JX175054Tachigali sp. Klitgaard 687 (K) AF365111j EU362040k
Tachigali amplifolia (Ducke) Barneby Motis et al. 24793 (NY) JX175231Tetrapterocarpon geayi Humbert Bruneau 1395 (WAG) JX099333 GQ293185Tetrapterocarpon geayi Humbert DuPuy M421 (MO) AF365101j
�Umtiza listeriana Sim Schrire 2602 (K) AF365126j EU362062k GQ293186 GQ293222Vouacapoua macropetala Sandwith Breteler 13793 (WAG) AF365110j EU362063k GQ293187 GQ293223
Dialiinae cladeDialium guianense (Aubl.) Sandwith Klitgaard 686 (K) AF365079j EU361930k JX175052Koompassia excelsa (Becc.) Taub. Herendeen 1-V-99-7 (US) EU361816k EU361988k GQ293166
MimosoideaeAdenanthera pavonina L. Major Howell Seeds (BH) AF278486i AF521808m
Calpocalyx dinklagei Harms Breteler 15461 (WAG) AF365043j EU361907k GQ293150 GQ293192Cylicodiscus gabunensis Harms Breteler 14866 (WAG) AY125845p AF521819k GQ293156 GQ293198Entada phaseoloides (L.) Merr. Lorence 7994 (PTBG) EU366228k EU366222k
Entada polyphylla Benth. Klitgaard 613 (K) GQ293203Pentaclethra macrophylla Benth. BNBG 87-1143 (BR) AF521853m
Pentaclethra macrophylla Benth. J. deWilde 11496 (WAG) AF365051j GQ293175Piptadenia anolidurus Barneby Klitgaard 691 (K) GQ293216Piptadenia robusta Pittier Arroyo 850 (NY) DQ784674o DQ790632o
Parkia multijuga Benth. Klitgaard 697 (K) AF365050j EU362018k
PapilionoideaeBobgunnia fistuloides (Harms) J.H. Kirkbr. &
WiersemaBreteler 14870 (WAG) AF365038j EU361885k
Dussia macroprophyllata (Donn. Sm.) Harms Landrum 10293 AY386903n
Dussia tessmannii Harms Klitgaard 628 (K) JX073264 GQ293161 JX175204Glycine max Merr. (L.) – DQ131547r AF142700w AF030231b
Leucomphalos callicarpus (Benth.) Breteler Breteler 12331 (MO, K) JX175226Medicago sativa L. – DQ131554r AY386881n AF049487z AJ002488h
Medicago truncatula L. – DQ311712s AF522109x AJ131943c
Phaseolus vulgaris L. – GQ279376t DQ450863y AF315375d
Pisum sativum L. – DQ311717s AY386961n AY386961n
Swartzia cardiosperma Spruce ex Benth. Klitgaard 664 (K) AF365040j EU362053k
Vicia faba L. – X51471u AY386899n X69773f AB038648i
Vigna radiata (L.) R. Wilczek – AB304065v DQ445950y VIRVSS1g
Accession numbers marked by a letter are from the following publications: a Choi et al. (2004), b Zhang et al. (1997), c Hohnjec et al. (1999), d Camas (GenBank, unpublisheddata), e Craig et al. (1999), f Heim et al. (1993), g Arai et al. (1992), h Vissi et al. (1998), i Takemiya et al. (2006), j Bruneau et al. (2001), k Bruneau et al. (2008), l Herendeen et al.(2003a), m Luckow et al. (2003), n Wojciechowski et al. (2004), o Jobson and Luckow (2007), p Haston et al. (2005), q Marazzi et al. (2006), r James and Schmidt (2004), s Ellisonet al. (2006), t Ou et al. (GenBank, unpublished data), u Herdenberger et al. (1990), v Tun and Yamaguchi (2007), w Hu et al. (2000), x Steele and Wojciechowski (2003), y
Delgado-Salinas et al. (2006), z Robinson et al. (GenBank, unpublished data).
V. Manzanilla, A. Bruneau / Molecular Phylogenetics and Evolution 65 (2012) 149–162 153
sequencing conditions were the same as above except that the vec-tor primer was used (i.e., SP6-T7).
All PCR products were purified using a PEG purification protocol(Joly et al., 2006). Cycle sequencing was performed from both endswith ‘‘Big Dye Terminator’’ chemistry (Applied Biosystems, FosterCity, California, USA), using one eighth of the reaction volume sug-gested in the manufacturer’s instructions for a total volume of10 ll reaction with 0.25 ll of Big Dye Terminator. Sequenced prod-ucts were run on an ABI 3100–avant automated DNA sequencer(Applied Biosystems). Chromatograms were manually edited andthe two chromatograms of a fragment were assembled withSequencher v.4.7 (Gene Codes Corp., Maddison, Wisconsin, USA).
2.4. Sequence alignment and phylogenetic analyses
DNA sequences were aligned in Clustal W (Thompson et al.,1997) and manually adjusted in BioEdit 7.0.5.3 (Hall, 1999). TheSUSY and matK loci, both protein-coding sequences, were trans-lated to amino acids to verify the alignment. Regions in which posi-tional homology was ambiguous were excluded from the analysis
for the matK and trnL loci. Gaps (indels) were coded as separatepresence/absence characters as implemented in SeqState (Müller,2005); a total of 42 indels were coded for the two chloroplastmarkers. Introns in SUSY were excluded because we were unableto align them with certainty. No indels were observed in the SUSYexons. Initially, phylogenetic analyses were conducted with allSUSY alleles for all individuals sequenced to test whether allelesof single individuals formed monophyletic groups (Manzanilla,2009). Subsequently, one allele was chosen at random to representeach monophyletic individual. SeqState also was used to evaluatethe range of sequence divergence and the proportion of informa-tive sites for each of the loci. Aligned DNA sequences are availablein TreeBASE (12744).
Because a partitioning strategy may help capture the underly-ing evolutionary process with accuracy, we divided the sequencedata into biologically relevant partitions. We developed four parti-tion models taking into account region sequenced, gap data, andcodon position for the coding regions (see Table 4 legend). For eachpartition, we selected the best-fitting model using the AIC criterionimplemented in jModelTest (Posada, 2008). We used the standard
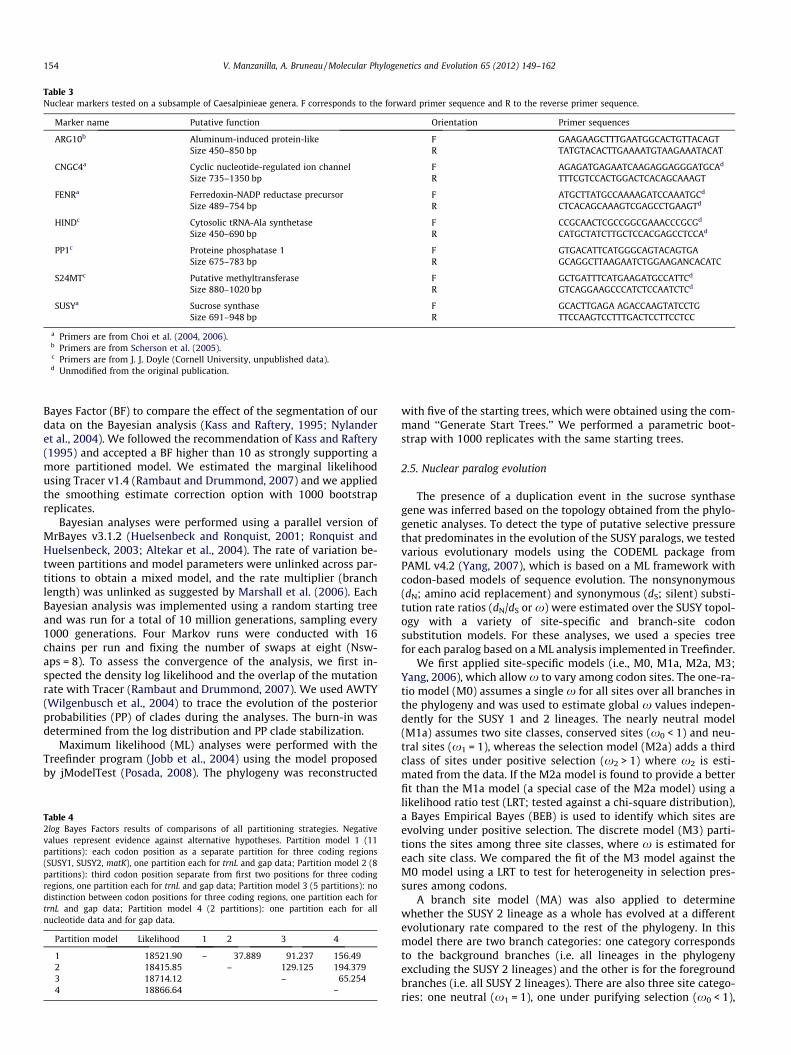
Table 3Nuclear markers tested on a subsample of Caesalpinieae genera. F corresponds to the forward primer sequence and R to the reverse primer sequence.
Marker name Putative function Orientation Primer sequences
ARG10b Aluminum-induced protein-like F GAAGAAGCTTTGAATGGCACTGTTACAGTSize 450–850 bp R TATGTACACTTGAAAATGTAAGAAATACAT
CNGC4a Cyclic nucleotide-regulated ion channel F AGAGATGAGAATCAAGAGGAGGGATGCAd
Size 735–1350 bp R TTTCGTCCACTGGACTCACAGCAAAGT
FENRa Ferredoxin-NADP reductase precursor F ATGCTTATGCCAAAAGATCCAAATGCd
Size 489–754 bp R CTCACAGCAAAGTCGAGCCTGAAGTd
HINDc Cytosolic tRNA-Ala synthetase F CCGCAACTCGCCGGCGAAACCCGCGd
Size 450–690 bp R CATGCTATCTTGCTCCACGAGCCTCCAd
PP1c Proteine phosphatase 1 F GTGACATTCATGGGCAGTACAGTGASize 675–783 bp R GCAGGCTTAAGAATCTGGAAGANCACATC
S24MTc Putative methyltransferase F GCTGATTTCATGAAGATGCCATTCd
Size 880–1020 bp R GTCAGGAAGCCCATCTCCAATCTCd
SUSYa Sucrose synthase F GCACTTGAGA AGACCAAGTATCCTGSize 691–948 bp R TTCCAAGTCCTTTGACTCCTTCCTCC
a Primers are from Choi et al. (2004, 2006).b Primers are from Scherson et al. (2005).c Primers are from J. J. Doyle (Cornell University, unpublished data).d Unmodified from the original publication.
154 V. Manzanilla, A. Bruneau / Molecular Phylogenetics and Evolution 65 (2012) 149–162
Bayes Factor (BF) to compare the effect of the segmentation of ourdata on the Bayesian analysis (Kass and Raftery, 1995; Nylanderet al., 2004). We followed the recommendation of Kass and Raftery(1995) and accepted a BF higher than 10 as strongly supporting amore partitioned model. We estimated the marginal likelihoodusing Tracer v1.4 (Rambaut and Drummond, 2007) and we appliedthe smoothing estimate correction option with 1000 bootstrapreplicates.
Bayesian analyses were performed using a parallel version ofMrBayes v3.1.2 (Huelsenbeck and Ronquist, 2001; Ronquist andHuelsenbeck, 2003; Altekar et al., 2004). The rate of variation be-tween partitions and model parameters were unlinked across par-titions to obtain a mixed model, and the rate multiplier (branchlength) was unlinked as suggested by Marshall et al. (2006). EachBayesian analysis was implemented using a random starting treeand was run for a total of 10 million generations, sampling every1000 generations. Four Markov runs were conducted with 16chains per run and fixing the number of swaps at eight (Nsw-aps = 8). To assess the convergence of the analysis, we first in-spected the density log likelihood and the overlap of the mutationrate with Tracer (Rambaut and Drummond, 2007). We used AWTY(Wilgenbusch et al., 2004) to trace the evolution of the posteriorprobabilities (PP) of clades during the analyses. The burn-in wasdetermined from the log distribution and PP clade stabilization.
Maximum likelihood (ML) analyses were performed with theTreefinder program (Jobb et al., 2004) using the model proposedby jModelTest (Posada, 2008). The phylogeny was reconstructed
Table 42log Bayes Factors results of comparisons of all partitioning strategies. Negativevalues represent evidence against alternative hypotheses. Partition model 1 (11partitions): each codon position as a separate partition for three coding regions(SUSY1, SUSY2, matK), one partition each for trnL and gap data; Partition model 2 (8partitions): third codon position separate from first two positions for three codingregions, one partition each for trnL and gap data; Partition model 3 (5 partitions): nodistinction between codon positions for three coding regions, one partition each fortrnL and gap data; Partition model 4 (2 partitions): one partition each for allnucleotide data and for gap data.
Partition model Likelihood 1 2 3 4
1 �18521.90 – �37.889 91.237 156.492 �18415.85 – 129.125 194.3793 �18714.12 – 65.2544 �18866.64 –
with five of the starting trees, which were obtained using the com-mand ‘‘Generate Start Trees.’’ We performed a parametric boot-strap with 1000 replicates with the same starting trees.
2.5. Nuclear paralog evolution
The presence of a duplication event in the sucrose synthasegene was inferred based on the topology obtained from the phylo-genetic analyses. To detect the type of putative selective pressurethat predominates in the evolution of the SUSY paralogs, we testedvarious evolutionary models using the CODEML package fromPAML v4.2 (Yang, 2007), which is based on a ML framework withcodon-based models of sequence evolution. The nonsynonymous(dN; amino acid replacement) and synonymous (dS; silent) substi-tution rate ratios (dN/dS or x) were estimated over the SUSY topol-ogy with a variety of site-specific and branch-site codonsubstitution models. For these analyses, we used a species treefor each paralog based on a ML analysis implemented in Treefinder.
We first applied site-specific models (i.e., M0, M1a, M2a, M3;Yang, 2006), which allow x to vary among codon sites. The one-ra-tio model (M0) assumes a single x for all sites over all branches inthe phylogeny and was used to estimate global x values indepen-dently for the SUSY 1 and 2 lineages. The nearly neutral model(M1a) assumes two site classes, conserved sites (x0 < 1) and neu-tral sites (x1 = 1), whereas the selection model (M2a) adds a thirdclass of sites under positive selection (x2 > 1) where x2 is esti-mated from the data. If the M2a model is found to provide a betterfit than the M1a model (a special case of the M2a model) using alikelihood ratio test (LRT; tested against a chi-square distribution),a Bayes Empirical Bayes (BEB) is used to identify which sites areevolving under positive selection. The discrete model (M3) parti-tions the sites among three site classes, where x is estimated foreach site class. We compared the fit of the M3 model against theM0 model using a LRT to test for heterogeneity in selection pres-sures among codons.
A branch site model (MA) was also applied to determinewhether the SUSY 2 lineage as a whole has evolved at a differentevolutionary rate compared to the rest of the phylogeny. In thismodel there are two branch categories: one category correspondsto the background branches (i.e. all lineages in the phylogenyexcluding the SUSY 2 lineages) and the other is for the foregroundbranches (i.e. all SUSY 2 lineages). There are also three site catego-ries: one neutral (x1 = 1), one under purifying selection (x0 < 1),

V. Manzanilla, A. Bruneau / Molecular Phylogenetics and Evolution 65 (2012) 149–162 155
and a third one for the foreground branches only that is under po-sitive selection (x2 > 1). To test for evidence of positive selection inSUSY 2, we compared this model with a more specific one with nopositive selection (x2 = 1) using a LRT. The same analysis was alsoperformed using the SUSY 1 lineage as foreground.
3. Results
3.1. Nuclear phylogenetic markers
Although seven low-copy nuclear loci were tested for their phy-logenetic utility in the Caesalpinioideae (Table 3), only one provedappropriate. The screening of ARGI and CNGC4 sequences showeda high degree of variability, with sequence length variations amongspecies of up to 400 bp and 300 bp, respectively. The FERN andS24MT loci amplifications yielded multiple bands and variationin length up to 500 bp. Amplification of the HIND marker wasunsuccessful. PP1 amplified easily and seemed to provide anappropriate level of variation for the group studied, but the phylo-genetic pattern obtained was chaotic (Manzanilla, 2009). By exam-ining the conceptual translation of the PP1 gene sequences, wefound a stop codon indicating the presence of a pseudogene andthe presence of paralogous copies of PP1. Despite the identificationof paralogs, we were unable to recover a coherent phylogenetic sig-nal. Moreover, this locus showed a significant level of saturationwith a proportion of invariable sites at 0.561, as calculated byjModelTest (Iss = 0.296, Iss.c = 0.707, p = 0.0918). The SUSY locushad a high variation rate and was sufficiently long, which corre-sponded to the criteria set for this study.
SUSY is composed of three exons (150 bp, 221 bp, 174 bp,respectively) and two introns (80–130 bp and 76–273 bp). Twocopies of SUSY (named SUSY 1 and SUSY 2) were detected for themajority of the taxa sampled here. We did not observe a secondcopy of the marker for genera within the Dialiinae clade and thePapilionoideae, despite extensive sampling of clones (24–48 clonesper sample). The translation into amino acid sequences of bothpartial copies did not reveal stop codons, suggesting that both cop-ies might be functional. We treated these paralogs as two phyloge-netic markers. The intron regions were removed from thealignment because they were hyper-variable; however these in-trons may prove useful in phylogenetic studies at the inter-specificlevel in caesalpinioid legumes.
The sequence comparisons using SeqState (Müller, 2005) sug-gested that overall among Caesalpinieae grade taxa, the SUSY cod-ing sequences were strongly divergent (range of up to 4% for SUSY1 sequences; up to 4.25% for SUSY 2 sequences), compared to 2.7%for the matK coding region. However, the PAML (Yang, 2007) anal-yses indicated that the 182 amino acid sequences are highly con-served with 0.35% and 0.5% divergence for SUSY 1 and SUSY 2across the Caesalpinieae grade, in comparison with 0.7% diver-gence for the matK gene. Approximately 13% of SUSY 1 and 11%of SUSY 2 sites are phylogenetically informative, compared to10% of sites for the matK coding region and 12% for the trnL intron.
3.2. Phylogenetic analyses
In the final matrix, at least one SUSY sequence was obtained for42 of 60 genera of the Caesalpinieae grade (47 species; 16 sampleswith only one of the two SUSY paralogs), six genera of subfamilyMimosoideae, two from the Dialiinae clade, and eight from thePapilionoideae (seven previously published). All sequences aredeposited in GenBank (Table 2). We were unable to obtain nuclearsequences for 12 genera for which we had plastid data, but thesegenera were included in the combined plastid plus nuclear matrixto achieve a better representation of the phylogeny of the group.
The combined matrix included 3437 characters, of which 10% wereinformative. The final alignment contained 8% missing data.
Using AWTY and Tracer, we estimated at around five milliongenerations per run the convergence of each run of the Bayesiananalysis to an apparent stationary position. The plot of the likeli-hood score for each run revealed that some runs were stuck on alocal maximum, leading us to re-start the analysis. The likelihoodscore of partition models 1 and 2 (that take into consideration co-don position) fitted the data better than partition models 3 and 4(see Table 4). The partition model 2, which distinguishes the thirdcodon position for all coding regions, had fewer parameters to esti-mate than partition model 1 and fit the data better.
3.3. Phylogenetic relationships
The nuclear gene phylogenetic analysis revealed the presence oftwo paralogous clades, SUSY 1 and SUSY 2 (Fig. 1). The tree isrooted with the Dialiinae and, in this analysis, the Papilionoideaeare weakly supported as sister to the SUSY 1 clade. Within boththe SUSY 1 and SUSY 2 clades, our analyses resolved four clades(Peltophorum, Cassia, Caesalpinia, Mimosoideae), with the samepattern of relationships in the two clades, albeit not always well-supported. The previously recognized Umtiza clade (Herendeenet al., 2003b) is not supported as monophyletic and relationshipsof genera in the Dimorphandra Group (Lewis, 2005) remain poorlyresolved relative to their placement with the Mimosoideae andPeltophorum clades.
No major dissimilarities between the ML and the Bayesian com-bined analyses were found except for the position of a few terminalnodes. In the combined nuclear and plastid analysis with bothBayesian and ML, we identified five strongly supported clades with-in the Caesalpinieae grade: Cassia, Caesalpinia, Peltophorum, Tach-igali, Mimosoideae; PP = 0.89–1.00, ML = 74–100 (Fig. 2). As in theSUSY only analysis, the Umtiza clade recognized by Herendeenet al. (2003b) is not resolved as monophyletic, but includes threeparaphyletic lineages that are each well supported. The Dimorphan-dra Group also is not supported as monophyletic.
These combined plastid and nuclear DNA analyses suggest thatCordeauxia and Stuhlmannia are sister to the other genera in a well-supported Caesalpinia clade. The remainder of the Caesalpiniaclade clearly separates into two strongly supported groups. Oneclade includes Pomaria, Erythrostemon and Poincianella as a mono-phyletic group sister to a group consisting of Libidibia, Hoffmann-seggia, Stahlia and Balsamocarpon. The other Caesalpinia claderesolves Tara and Coulteria as sister to a clade formed by Moullava,Pterolobium, Guilandina, Cenostigma, and Mezoneuron, but with noclear resolution among these two subclades and the genusHaematoxylum.
As resolved here, the Cassia clade groups together with moder-ate support values (PP = 0.89, ML = 74) seven genera that previ-ously had not been considered closely related. However,relationships among these genera are unclear. The topology pre-sents Cassia as sister to Senna, both sister to a clade that includesVouacapoua, Recordoxylon, Melanoxylon, Chamaecrista and Batesia.The genus Pterogyne is in an ambiguous position and, dependingon the marker or the analysis, it can be placed either in the Cae-salpinia clade (plastid data) or in the Cassia clade (nuclear andcombined data, Figs. 1 and 2). Moreover, Pterogyne distinguishes it-self from other taxa by a longer branch, both in the nuclear andchloroplast DNA topologies.
The Peltophorum clade has high support values and the rela-tionships within the clade are well supported. Nevertheless therelationships between Delonix, Lemuropisum and Colvillea remainunclear, and Delonix appears polyphyletic. The relationship of theTachigali clade (all three genera sampled; PP = 1.00, ML = 84) with
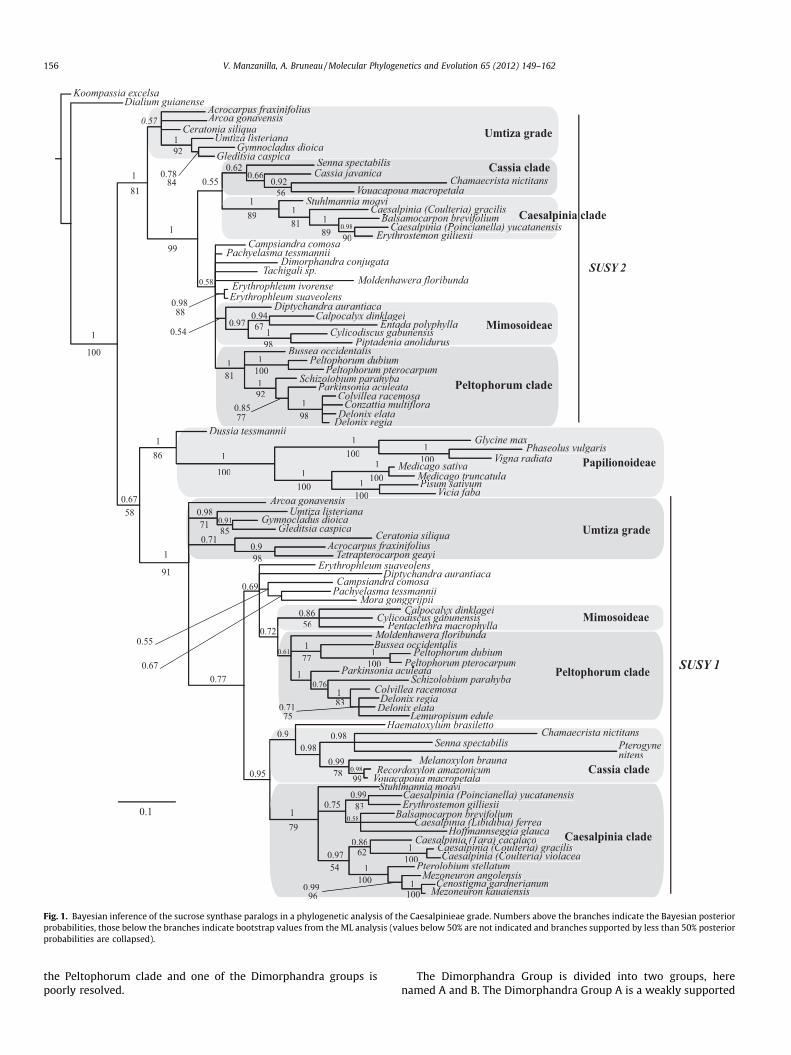
Fig. 1. Bayesian inference of the sucrose synthase paralogs in a phylogenetic analysis of the Caesalpinieae grade. Numbers above the branches indicate the Bayesian posteriorprobabilities, those below the branches indicate bootstrap values from the ML analysis (values below 50% are not indicated and branches supported by less than 50% posteriorprobabilities are collapsed).
156 V. Manzanilla, A. Bruneau / Molecular Phylogenetics and Evolution 65 (2012) 149–162
the Peltophorum clade and one of the Dimorphandra groups ispoorly resolved.
The Dimorphandra Group is divided into two groups, herenamed A and B. The Dimorphandra Group A is a weakly supported
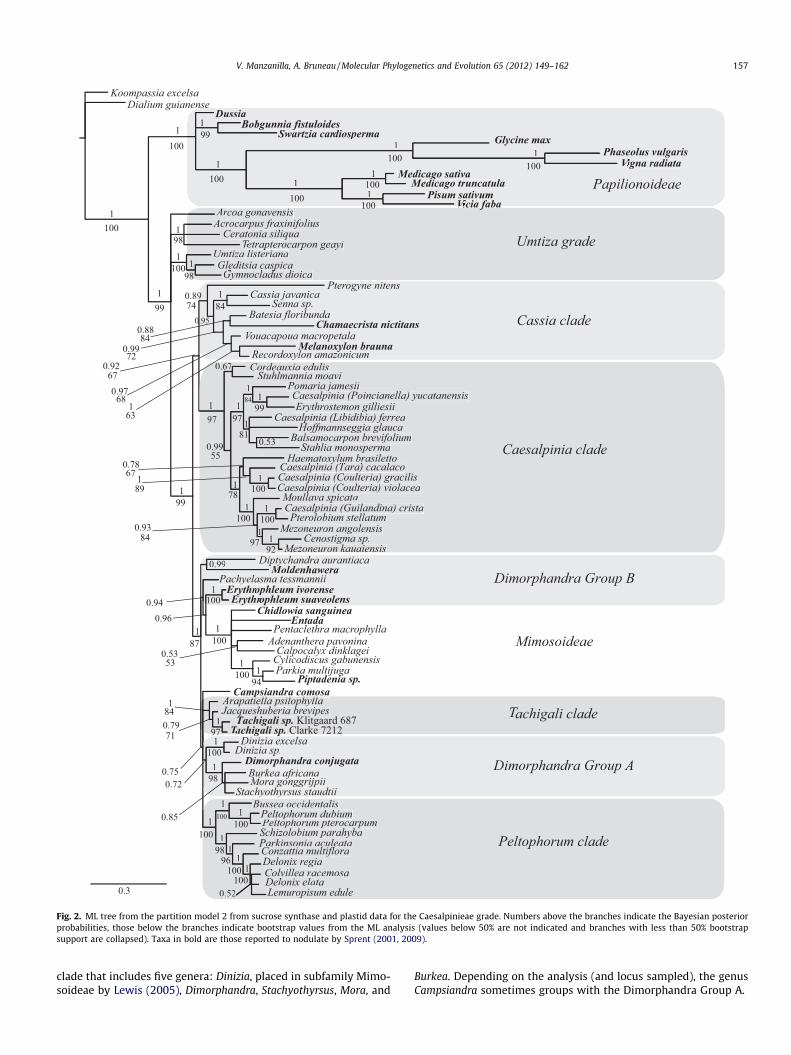
Fig. 2. ML tree from the partition model 2 from sucrose synthase and plastid data for the Caesalpinieae grade. Numbers above the branches indicate the Bayesian posteriorprobabilities, those below the branches indicate bootstrap values from the ML analysis (values below 50% are not indicated and branches with less than 50% bootstrapsupport are collapsed). Taxa in bold are those reported to nodulate by Sprent (2001, 2009).
V. Manzanilla, A. Bruneau / Molecular Phylogenetics and Evolution 65 (2012) 149–162 157
clade that includes five genera: Dinizia, placed in subfamily Mimo-soideae by Lewis (2005), Dimorphandra, Stachyothyrsus, Mora, and
Burkea. Depending on the analysis (and locus sampled), the genusCampsiandra sometimes groups with the Dimorphandra Group A.

Table 5Maximum likelihood parameter estimates for two copies of the SUSY gene in the Caesalpinieae grade. x, ratio of synonymous/non-synonymous substitutions; p, proportion ofsites estimated.
Model SUSY 1 SUSY 2
Loglikelihood
Parameter estimates Positively selectedsites
Loglikelihood
Parameter estimates Positively selectedsites
MO (one-ratio) �5595.83 x = 0.022 None �4591.39 x = 0.038 NoneM1a (neutral) �5476.04 p0 = 0.98 (p1 = 0.016) Not allowed �4456.68 p0 = 0.961 (p1 = 0.039) Not allowed
x0 = 0.01, x1 = 1 x0 = 0.017, x1 = 1M2a (selection) �5483.60 p0 = 0.984, p1 = 0.016 (p2 = 0) 103R (p > 0.5) �4443.08 p0 = 0.960, p1 = 0.035 (p2 = 0.005) 160R (p > 0.99)
x0 = 0.012, x1 = 1, x2 = 13.47 x0 = 0.01, x1 = 1 x2 = 3.99M3 (discrete) �5465.07 p0 = 0.971, p1 = 0.024 (p2 = 0.005) 103R (p > 0.99) �4427.19 p0 = 0.874, p1 = 0.109 (p2 = 0.017) 102K, 132N, 160R
(p > 0.99)Branch site
model A�5483.82 p0 = 0.918, p1 = 0.015, p2a = 0.066,
p2b = 0.001�4456.82 p0 = 0.878, p1 = 0.036, p2a = 0.083,
p2b = 0.003Background x0 = 0.012, x1 = 1.0 x0 = 0.017, x1 = 1.0Foreground x0 = 0.012, x1 = 1.0, x2a = 1.0,
x2b = 1.0x0 = 0.017, x1 = 1.0, x2a = 1.0,x2b = 1.0
158 V. Manzanilla, A. Bruneau / Molecular Phylogenetics and Evolution 65 (2012) 149–162
Our analysis of the closest relatives of subfamily Mimosoideaesuggests that a grade of several caesalpinioid genera may be atthe base of the Mimosoideae (Moldenhawera, Diptychandra, Pach-yelasma and Erythrophleum). This grade of Dimorphandra GroupB genera always occurs as close relatives of Mimosoideae, but rela-tionships are never well supported. The Mimosoideae clade isstrongly supported (PP = 1.00, ML = 100) and includes the caesal-pinioid genus Chidlowia.
3.4. Nuclear marker evolution
The codon selection model analyses with PAML (Yang, 2007)found low values for x, suggesting strong purifying selection forboth the SUSY1 and SUSY2 paralogs (xsusy1 = 0.022 and xsusy2
= 0.038, given the one-ratio model (M0); Table 5). The M2a modelfits significantly better the data than the M1a model(2Dlsusy1 = 15.12, 2Dlsusy2 = 27.2, p < 0.0001, df = 2) suggesting thatthere could be positive selection in both paralog lineages. The max-imum likelihood estimates under the selection model, M2a, suggestthat about 0% and 0.5% of the SUSY 1 and SUSY 2 sites, respectively,are under positive selection, with a much higher estimated x forSUSY 1 (x2 = 13.5) than for SUSY 2 (x2 = 4). The comparison ofthe one ratio (M0) model with the discrete model (M3) (2Dlsusy1 =261.5, 2Dlsusy2 = 328.38, p < 0.0001, df = 4) suggests heterogeneityacross sites in both SUSY lineages. The paralogous genes may haveexperienced different evolutionary pressures, which would be visi-ble with the estimation of different classes of sites. The M2a (andM3) model identifies four sites (codons) as potentially under posi-tive selection: 102K, 103R, 132N and 160R (Table 5). Because bothparalogs appear to evolve by positive selection for just one or a fewamino acid changes, the x ratios are not very different between theSUSY 1 and SUSY 2 clades.
The MA branch site model shows that 87% of the SUSY 2 se-quences are under strong purifying selection, against 97% for SUSY1, suggesting that SUSY 1 is under stronger purifying selection thanSUSY 2. The MA model did not provide a significantly better fit tothe data than the null hypothesis of neutrality with x2 = 1, provid-ing no evidence for directional selection either for the SUSY 2 line-age, or for the SUSY 1 lineage, relative to the background lineages.
4. Discussion
4.1. Nuclear loci evolution
The two SUSY paralog clades form well-supported gene subfam-ilies, in which the evolutionary patterns among lineages is similar.The Papilionoideae and Dialiinae clades do not appear to possess
the two copies of SUSY according to our sampling (see also Choiet al., 2004, 2006), but we cannot exclude the possibility that addi-tional paralogs could be present in earlier diverged lineages. TheMimosoideae sampled have both SUSY copies identified in thisstudy. In consequence, we consider that SUSY may have undergonea duplication event following the divergence of the Papilionoideaeclade (Fig. 3). However, it is also possible that the duplication oc-curred prior to the divergence of the Papilionoideae clade (Fig. 1)but this would imply SUSY 2 paralog loss in this subfamily. Singlegene duplication events occur stochastically and are known forcontributing to gene polymorphisms (Lynch and Conery, 2000;Moore and Purugganan, 2005). At higher phylogenetic levels, sim-ilar independent duplication events are known to have occurredregularly in the evolution of plant genes and have been identifiedin other groups, including other Leguminosae (e.g., Lavin et al.,1998; Citerne et al., 2003; Archambault and Bruneau, 2004).Although the observed conservation in the phylogenetic patternof the two paralogs is remarkable and particularly useful for phy-logeny reconstruction, congruent phylogenetic signals of paralogshave been noted elsewhere (e.g., Ree et al., 2004).
Immediately following a duplication event, paralogs are co-ex-pressed and preserve their function, but this functional redundancymay have several outcomes within an organism (Lynch and Conery,2000; Moore and Purugganan, 2005). Thus, we were interested inbetter understanding the evolutionary pressure experienced afterthe duplication event in both sucrose synthase copies. For example,following a duplication event, either paralog may accumulatedegenerative mutations to become a pseudogene. Pseudogeniza-tion implies relaxed evolution (x = 1), where the gene accumulatesneutral mutations, often at an observed accelerated rate. Alterna-tively the new gene may take on a new function driven by positiveselection due to new selective pressures (x > 1) with a predomi-nance of adaptive mutations (neofunctionalization). In addition,the duplicated genes may conserve the same function but with dif-ferent spatial or temporal actions due to differences in the coding orregulatory regions. This subfunctionalization may be the result ofthe accumulation of degenerate mutations (x > 1), where positiveselection may also have played a key role. The codon selection mod-el analyses suggest that the SUSY amino acid sequence is relativelyconserved with no major differences among lineages due to purify-ing selection. Assuming that an ancestrally duplicated copy was notsubsequently lost in the Papilionoideae, the phylogenetic analyses(Fig. 1) indicate that the SUSY 1 sequences, rather than the SUSY2 sequences, are more similar to the unduplicated sequences ofthe SUSY gene. Regardless, the absence of strong differences inselection pressure between the two paralogs, as noted by the codonmodel selection, suggests that subfunctionalization rather than

Papilionoideae
Caesalpinia clade
Cassia clade
Umtiza grade
Mimosoideae
Peltophorum clade
Tachigali clade
Dimorphandra group B
Dimorphandra group A
Documented nodulation
Dialiinae clade, Outgroup
Duplication of SUSY
0.3
1
100
1
99
981
1001
0.95
50
0.8974
971
0.9267
1001
991
184
0.72
1001
0.75
187
100
1
Fig. 3. Summary maximum likelihood (ML) tree using the partition model 2 (Table 4) for sucrose synthase and plastid data for the Caesalpinieae grade. Numbers above thebranches indicate the Bayesian posterior probabilities, while those below the branches indicate bootstrap values from the ML analysis. The major clades observed in Fig. 2 areindicated; branches with less than 50% bootstrap support are collapsed. Black stars indicate nodulating taxa for species and genera studied, following Sprent (2001, 2009).
V. Manzanilla, A. Bruneau / Molecular Phylogenetics and Evolution 65 (2012) 149–162 159
neofunctionalization may be a more plausible explanation for theevolution of the sucrose synthase gene is caesalpinioid legumes.
The SUSY gene is implicated in root nodule formation andmetabolism (Baier et al., 2007). The possible subfunctionalizationof this gene may be the consequence of particular morphological,structural or regulatory features of root nodules (Sprent, 2009).Our phylogenetic study supports multiple origins of nodules andtheir presence throughout all the Caesalpinieae clades and in theCassieae clade as well (Fig. 3). The evolutionary history of nodulesappears to be a complex problem within the Caesalpinieae grade,which could be further studied through phylogenies of genesimplicated in the N-fixating process (see Doyle, 2011).
4.2. Caesalpinieae grade phylogeny
The addition of nuclear sequence data provides a stronger phylo-genetic signal compared to previous plastid phylogenetic analyses ofsubfamily Caesalpinioideae (e.g., Bruneau et al., 2001, 2008). The nu-clear markers studied have a faster evolutionary rate than most plas-tid markers used to date. With the addition of these numerousvariable characters, we obtain a more complete view of phylogeneticrelationships in the Caesalpinieae grade and in the early diverginglineages of the Mimosoideae. The SUSY duplication event, which iscommon to the Caesalpinieae grade and the Mimosoideae, adds avaluable taxonomic character which supports the monophyly of

160 V. Manzanilla, A. Bruneau / Molecular Phylogenetics and Evolution 65 (2012) 149–162
these two groups together. In this phylogenetic analysis, we aremissing nuclear or plastid sequences for six genera generally placedin the tribe Caesalpinieae (Polhill, 1994; Lewis, 2005), four of whichrarely or never have been included in a molecular analysis because oflack of or poor quality of material (Stenodrepanum, Sympetalandra,Lophocarpinia, and Orphanodendron; Table 1). In the combined anal-yses, some groups are better supported and several have better re-solved intergeneric relationships. However, the positions ofcertain taxa differ between the nuclear and plastid analyses. The nu-clear topology suggests that Pterogyne belongs to the Cassia clade asalso suggested by the morphological analyses of Herendeen et al.(2003a), whereas previous plastid DNA analyses had shown Pterogy-ne as sister to the Caesalpinia clade, albeit with weak support values(Haston et al., 2003; Bruneau et al., 2008). This uncertainty may beattributed to the low variability of the chloroplast genome. The longbranch in Pterogyne is notable for all molecular markers (i.e., Bru-neau et al. (2008) for the chloroplast markers; Fig. 1 for SUSY), indi-cating a rapid evolutionary rate for this monospecific genus, which ispresently unexplained. The phylogeny obtained in previous molec-ular and morphological studies considered the Umtiza group to bea clade (Herendeen et al., 2003a, 2003b; Bruneau et al., 2008), butin our analyses this clade is not supported (Fig. 2). The combinedmorphological and molecular analyses of Herendeen et al. (2003a)also suggest that Tachigali is sister to the Caesalpinieae grade withthe exception of the Umtiza grade but all molecular analyses to datesuggest a nested position within the Caesalpinieae grade.
We infer the position of the genus Dinizia to be within the caesalpin-ioids, as previously suggested (Luckow et al., 2000; Wojciechowskiet al., 2004; Bruneau et al., 2008), and excluding this genus, our anal-yses support the monophyly of subfamily Mimosoideae (with verylimited sampling). Conversely, our analyses suggest for the first timethat the morphologically distinct monospecific West African genusChidlowia, generally considered a member of the Caesalpinioideae(Lewis, 2005), might best be placed in the Mimosoideae. The Dim-orphandra Group B grade contains genera that are morphologicallysimilar to some of the Mimosoideae, bringing into question the tax-onomic delimitation and the evolution of the Caesalpinioideae, andin particular of the Caesalpinieae grade. For example, should we in-clude these caesalpinioid lineages (Dimorphandra Group B grade)within the Mimosoideae as early diverging genera of mimosoids?Or should we include the whole Caesalpinieae grade within theMimosoideae? In summary, what is the distinction between Cae-salpinieae and Mimosoideae genera? The SUSY duplication eventis a good molecular synapomorphy that indicates a common evolu-tionary history for the Caesalpinieae grade and the Mimosoideae.This could be used to argue the Caesalpinieae grade should be partof one large subfamily Mimosoideae, in which the Caesalpinieaeclades would represent different tribes. Alternatively, the Mimosoi-deae could include the Dimorphandra Group B grade genera withperhaps some other closely related clades of the Caesalpinieae grade(i.e., Dimorphandra Group A, Peltophorum, Tachigali clades). Eachclade would be defined as a tribe of Mimosoideae and the remainingCaesalpinideae grade lineages could be defined as distinct subfami-lies of the Leguminosae. Of course, this alternative increases thenumber of subfamilies and the taxonomic complexity of the family.
Future phylogenetic developments may help to delimit thetransition between the Caesalpinieae grade and the Mimosoideae,with more extensive species-level sampling of the DimorphandraGroup and early diverging lineages of mimosoids in order to betterunderstand which caesalpinioid genera are sister to the Mimosoi-deae. The addition of numerous nuclear markers in the phylogenyof the legumes appears to be ideal to resolve these intergenericphylogenies. However, we need a better understanding of nucleargenome evolution of the caesalpinioids to obtain additional singlecopy nuclear markers (Cronk et al., 2006). The genomic project onChamaecrista fasciculata may help identify useful nuclear loci (Sing-
er et al., 2009; Cannon et al., 2010). To understand the evolution ofthe legumes, the addition of morphological data appears to be cru-cial to resolve the taxonomic dilemma surrounding the delimita-tion of the Caesalpinieae grade and of the subfamily Mimosoideae.
Acknowledgments
The authors thank Marielle Babineau, Marjorie Mercure andSébastien Renaud for laboratory assistance and members of the A.Bruneau and L. Brouillet labs, as well as Annie Archambault, SimonJoly and two reviewers for comments on the manuscript. We kindlyacknowledge the Réseau Québécois de Calcul Haute Performance(RQCHP) for providing computational facilities. This project wasfunded by grants from the Natural Sciences and Engineering Re-search Council (Canada) and National Science Foundation (USA)DEB-9527673.
References
Altekar, G., Dwarkadas, S., Huelsenbeck, J.P., Ronquist, F., 2004. Parallel Metropoliscoupled Markov chain Monte Carlo for Bayesian phylogenetic inference.Bioinformatics 20, 407–415.
Altschul, S.F., Madden, T.L., Schäffer, A.A., Zhang, J., Zhang, Z., Miller, W., Lipman, D.J.,1997. Gapped BLAST and PSI-BLAST: a new generation of protein databasesearch programs. Nucl. Acids Res. 25, 389–402.
Arai, M., Mori, H., Imaseki, H., 1992. Expression of the gene for sucrose synthaseduring growth of mung bean seedlings. Plant Cell Physiol. 33, 503–506.
Archambault, A., Bruneau, A., 2004. Phylogenetic utility of the LEAFY/FLORICAULAgene in the Caesalpinioideae (Leguminosae): gene duplication and a novelinsertion. Syst. Bot. 29, 609–626.
Baier, M.C., Barsch, A., Küster, H., Hohnjec, N., 2007. Antisense repression of theMedicago truncatula nodule-enhanced sucrose synthase leads to a handicappednitrogen fixation mirrored by specific alterations in the symbiotictranscriptome and metabolome. Plant Physiol. 145, 1600–1618.
Banks, H., Klitgaard, B.B., Lewis, G.P., Crane, P.R., Bruneau, A., 2003. Pollen and thesystematics of tribes Caesalpinieae and Cassieae (Caesalpinioideae:Leguminosae). In: Klitgaard, B., Bruneau, A. (Eds.), Advances in LegumeSystematics, Part 10. Royal Botanic Gardens, Kew, pp. 95–122.
Bruneau, A., Forest, F., Herendeen, P.S., Klitgaard, B.B., Lewis, G.P., 2001.Phylogenetic relationships in the Caesalpinioideae (Leguminosae) as inferredfrom chloroplast trnL intron sequences. Syst. Bot. 26, 487–514.
Bruneau, A., Mercure, M., Lewis, G.P., Herendeen, P.S., 2008. Phylogenetic patternsand diversification in the caesalpinioid legumes. Botany 86, 697–718.
Cannon, S.B., Ilut, D., Farmer, A.D., Maki, S.L., May, G.D., Singer, S.R., Doyle, J.J., 2010.Polyploidy did not predate the evolution of nodulation in all legumes. PLoS ONE5, e11630.
Chappill, J.A., 1995. Cladistic analysis of the Leguminosae: the development of anexplicit phylogenetic hypothesis. In: Polhill, R.M., Raven, P.H. (Eds.), Advances inLegume Systematics, Part 7. Royal Botanic Gardens, Kew, pp. 1–9.
Chapman, M.A., Chang, J., Weisman, D., Kesseli, R.V., Burke, J.M., 2007. Universalmarkers for comparative mapping and phylogenetic analysis in the Asteraceae(Compositae). Theor. Appl. Genet. 115, 747–755.
Choi, H.-K., Kim, D., Uhm, T., Limpens, E., Lim, H., Mun, J.-H., Kalo, P., Penmetsa, R.V.,Seres, A., Kulikova, O., Roe, B.A., Bisseling, T., Kiss, G.B., Cook, D.R., 2004. Asequence-based genetic map of Medicago truncatula and comparison of markercolinearity with M. sativa. Genetics 166, 1463–1502.
Choi, H.-K., Luckow, M.A., Doyle, J., Cook, D.R., 2006. Development of nuclear gene-derived molecular markers linked to legume genetic maps. Mol. Genet. Genom.276, 56–70.
Citerne, H.L., Luo, D., Pennington, R.T., Coen, E., Cronk, Q.C.B., 2003. A phylogenomicinvestigation of CYCLOIDEA-Like TCP genes in the Leguminosae. Plant Physiol.131, 1042–1053.
Craig, J., Barratt, P., Tatge, H., Dejardin, A., Handley, L., Gardner, C.D., Barber, L.,Wang, T., Hedley, C., Martin, C., Smith, A.M., 1999. Mutations at the rug4 locusalter the carbon and nitrogen metabolism of pea plants through an effect onsucrose synthase. Plant J. 17, 353-36.
Cronk, Q., Ojeda, I., Pennington, R.T., 2006. Legume comparative genomics: progressin phylogenetics and phylogenomics. Curr. Opin. Plant Biol. 9, 99–103.
Delgado-Salinas, A., Bibler, R., Lavin, M., 2006. Phylogeny of the genus Phaseolus(Leguminosae): a recent diversification in an ancient landscape. Syst. Bot. 31,779–791.
Doyle, J.J., 1994. Phylogeny of the legume family: an approach to understanding theorigins of nodulation. Ann. Rev. Ecol. Syst. 25, 325–349.
Doyle, J.J., 2011. Phylogenetic perspectives on the origins of nodulation. Mol. Plant–Microbe Interact. 24, 1289–1295.
Doyle, J.J., Doyle, J.L., Ballenger, J.A., Dickson, E.E., Kajita, T., Ohashi, H., 1997. Aphylogeny of the chloroplast gene rbcL in the Leguminosae: taxonomiccorrelations and insights into the evolution of nodulation. Am. J. Bot. 84, 541–554.

V. Manzanilla, A. Bruneau / Molecular Phylogenetics and Evolution 65 (2012) 149–162 161
Doyle, J.J., Chappill, J.A., Bailey, C.D., Kajita, T., 2000. Towards a comprehensivephylogeny of legumes: evidence from rbcL sequences and non-molecular data.In: Herendeen, P.S., Bruneau, A. (Eds.), Advances in Legume Systematics, Part 9.Royal Botanic Gardens, Kew, pp. 1–20.
Doyle, J.J., Luckow, M.A., 2003. The rest of the iceberg. Legume diversity andevolution in a phylogenetic context. Plant Physiol. 131, 900–910.
Du Puy, D.J., Phillipson, P.B., Rabevohitra, R., 1995. The genus Delonix (Leguminosae:Caesalpinioideae: Caesalpinieae) in Madagascar. Kew. Bull. 50, 445–475.
Ellison, N.W., Liston, A., Steiner, J.J., Williams, W.M., Taylor, N.L., 2006. Molecularphylogenetics of the clover genus (Trifolium–Leguminosae). Mol. Phylogenet.Evol. 39, 688–705.
Engels, W., 2005. Amplify 3. <http://engels.genetics.wisc.edu/amplify/index.html>.Hall, T.A., 1999. Bioedit: a user-friendly biological sequence alignment editor and
analysis program for Windows 95/98/NT. Nucl. Acids Symp. Ser. 41, 95–98.Haston, E.M., Lewis, G.P., Hawkins, J.A., 2003. A phylogenetic investigation of the
Peltophorum group (Caesalpinieae: Leguminosae). In: Klitgaard, B., Bruneau, A.(Eds.), Advances in Legume Systematics, Part 10. Royal Botanic Gardens, Kew,pp. 149–159.
Haston, E.M., Lewis, G.P., Hawkins, J.A., 2005. A phylogenetic reappraisal of thePeltophorum group (Caesalpinieae: Leguminosae) based on the chloroplasttrnL–F, rbcL and rps16 sequence data. Am. J. Bot. 92, 1359–1371.
Heim, U., Weber, H., Baumlein, H., Wobus, U., 1993. A sucrose-synthase gene of Viciafaba L.: expression pattern in developing seeds in relation to starch synthesisand metabolic regulation. Planta 191, 394–401.
Herdenberger, F., Pillay, D.T., Steinmetz, A., 1990. Sequence of the trnH gene and theinverted repeat structure deletion site of the broad bean chloroplast genome.Nucl. Acids Res. 18, 1297.
Herendeen, P.S., Lewis, G.P., Bruneau, A., 2003a. Phylogenetic relationships incaesalpinioid legumes: a preliminary analysis based on morphological andmolecular data. In: Klitgaard, B., Bruneau, A. (Eds.), Advances in LegumeSystematics, Part 10. Royal Botanic Gardens, Kew, pp. 37–62.
Herendeen, P.S., Lewis, G.P., Bruneau, A., 2003b. Floral morphology in caesalpinioidlegumes: testing the monophyly of the ‘‘Umtiza Clade’’. Int. J. Plant Sci. 164,393–407.
Hohnjec, N., Becker, J.D., Puhler, A., Perlick, A.M., Kuster, H., 1999. Genomicorganization and expression properties of the MtSucS1 gene, which encodes anodule-enhanced sucrose synthase in the model legume Medicago truncatula.Mol. Gen. Genet. 261, 514–522.
Horst, I., Welham, T., Kelly, S., Kaneko, T., Sato, S., Tabata, S., Parniske, M., Wang, T.L.,2007. TILLING mutants of Lotus japonicus reveal that nitrogen assimilation andfixation can occur in the absence of nodule-enhanced sucrose synthase. PlantPhysiol. 144, 806–820.
Hu, J.M., Lavin, M., Wojciechowski, M.F., Sanderson, M.J., 2000. Phylogeneticsystematics of the tribe Millettieae (Leguminosae) based on chloroplast trnK/matK sequences and its implications for evolutionary patterns inPapilionoideae. Am. J. Bot. 87, 418–430.
Huelsenbeck, J.P., Ronquist, F., 2001. MRBAYES: Bayesian inference of phylogenetictrees. Bioinformatics 17, 754–755.
James, D., Schmidt, A.-M., 2004. Use of an intron region of a chloroplast tRNA gene(trnL) as a target for PCR identification of specific food crops including sourcesof potential allergens. Food Res. Int. 37, 395–402.
Jobb, G., von Haeseler, A., Strimmer, K., 2004. TREEFINDER: a powerful graphicalanalysis environment for molecular phylogenetics. BMC Evol. Biol. 4, 18.
Jobson, R.W., Luckow, M., 2007. Phylogenetic study of the genus Piptadenia(Mimosoideae: Leguminosae) using plastid trnL-F and trnK/matK sequencedata. Syst. Bot. 32, 569–575.
Joly, S., Starr, J.R., Lewis, W.H., Bruneau, A., 2006. Polyploid and hybrid evolution inroses east of the Rocky Mountains. Am. J. Bot. 93, 412–425.
Kajita, T., Ohashi, H., Tateishi, Y., Bailey, D.C., Doyle, J.J., 2001. rbcL and legumephylogeny, with particular reference to Phaseoleae, Millettieae, and allies. Syst.Bot. 26, 515–536.
Käss, E., Wink, M., 1996. Molecular evolution of the leguminosae: phylogeny of thethree subfamilies based on rbcL-sequences. Biochem. Syst. Ecol. 24, 365–378.
Kass, R.E., Raftery, A.E., 1995. Bayes factors. J. Am. Stat. Assoc. 90, 773–795.Lavin, M., Eshbaugh, E., Hu, J.-M., Mathews, S., Sharrock, R.A., 1998. Monophyletic
subgroups of the tribe Millettieae (Leguminosae) as revealed by phytochromenucleotide sequence data. Am. J. Bot. 85, 412–433.
Lavin, M., Herendeen, P.S., Wojciechowski, M.F., 2005. Evolutionary rates analysis ofleguminosae implicates a rapid diversification of lineages during the tertiary.Syst. Biol. 54, 575–594.
Lewis, G.P., 1998. Caesalpinia: A Revision of the Poincianella–Erythrostemon Group.Royal Botanic Gardens, Kew.
Lewis, G.P., 2005. Caesalpinioideae. In: Lewis, G., Schrire, B., Mackinder, B., Lock, M.(Eds.), Legumes of the World. Royal Botanic Gardens, Kew, pp. 127–161.
Lewis, G.P., Schrire, B.D., 1995. A reappraisal of the Caesalpinia group(Caesalpinioideae: Caesalpinieae) using phylogenetic analysis. In: Crisp, M.D.,Doyle, J.J. (Eds.), Advances in Legume Systematics, Part 7. Royal BotanicGardens, Kew, pp. 41–52.
Lewis, G., Schrire, B., Mackinder, B., Lock, M. (Eds.), 2005. Legumes of the World.Royal Botanic Gardens, Kew.
Luckow, M., White, P.J., Bruneau, A., 2000. Relationships among the basal genera ofthe Mimosoid Legumes. In: Herendeen, P.S., Bruneau, A. (Eds.), Advances inLegume Systematics, Part 9. Royal Botanic Gardens, Kew, pp. 165–171.
Luckow, M., Miller, J.T., Murphy, D.J., Livshultz, T., 2003. A phylogenetic analysis ofthe Mimosoideae (Leguminosae) based on chloroplast DNA sequence data. In:
Klitgaard, B., Bruneau, A. (Eds.), Advances in Legume Systematics, Part 10. RoyalBotanic Gardens, Kew, pp. 197–211.
Lynch, M., Conery, J.S., 2000. The evolutionary fate and consequences of duplicategenes. Science 290, 1151–1155.
Manzanilla, V., 2009. Une phylogénie du grade des Caesalpinieae inférée à partir demarqueurs nucléaires et chloroplastiques. Master’s Thesis. Université deMontréal, Montréal, pp. 1–65.
Marazzi, B., Endress, P.K., Queiroz, L.P.d., Conti, E., 2006. Phylogenetic relationshipswithin Senna (Leguminosae, Cassiinae) based on three chloroplast DNA regions:patterns in the evolution of floral symmetry and extrafloral nectaries. Am. J. Bot.93, 288–303.
Marshall, D.C., Simon, C., Buckley, T.R., 2006. Accurate branch length estimation inpartitioned Bayesian analyses requires accommodation of among-partition ratevariation and attention to branch length priors. Syst. Biol. 55, 993–1003.
Moore, R.C., Purugganan, M.D., 2005. The evolutionary dynamics of plant duplicategenes. Curr. Opin. Plant Biol. 8, 122–128.
Müller, K., 2005. SeqState: primer design and sequence statistics for phylogeneticDNA datasets. Appl. Bioinform. 4, 65–69.
Nylander, J.A.A., Ronquist, F., Huelsenbeck, J.P., Nieves-Aldrey, J.L., 2004. Bayesianphylogenetic analysis of combined data. Syst. Biol. 53, 47–67.
Pettigrew, C.J., Watson, L., 1977. On the classification of Caesalpinioideae. Taxon 26,57–64.
Polhill, R.M., 1994. Classification of the Leguminosae. In: Bisby, F.A., Harborne, J.B.(Ed.), Phytochemical Dictionary of the Leguminosae. Chapman and Hall, NewYork, pp. 25–57.
Polhill, R.M., Raven, P.H., Stirton, C.H., 1981. Evolution and systematics of theLeguminosae. In: Polhill, R.M., Raven, P.H. (Eds.), Advances in LegumesSystematics, Part 1. Royal Botanic Gardens, Kew, pp. 1–26.
Polhill, R.M., Vidal, J.E., 1981. Caesalpinieae. In: Polhill, R.M., Raven, P.H. (Eds.),Advances in Legumes Systematics, Part 1. Royal Botanic Gardens, Kew, pp. 81–96.
Posada, D., 2008. JModelTest: phylogenetic model averaging. Mol. Biol. Evol. 25,1253–1256.
Rambaut, A., Drummond, A.J., 2007. Tracer v1.4. <http://beast.bio.ed.ac.uk/Tracer>.Ree, R.H., Citerne, H.L., Lavin, M., Cronk, Q.C.B., 2004. Heterogeneous selection on
legcyc paralogs in relation to flower morphology and the phylogeny of Lupinus(Leguminosae). Mol. Phylogenet. Evol. 21, 321–331.
Ronquist, F., Huelsenbeck, J.P., 2003. MrBayes 3: Bayesian phylogenetic inferenceunder mixed models. Bioinformatics 19, 1572–1574.
Scherson, R.A., Choi, H.K., Cook, D.R., Sanderson, M.J., 2005. Phylogenetics of NewWorld Astragalus: screening of novel nuclear loci for the reconstruction ofphylogenies at low taxonomic levels. Brittonia 57, 354–366.
Silvente, S., Camas, A., Lara, M., 2003. Heterogeneityof sucrose synthase genes inbean (Phaseolus vulgaris L.): evidence for a nodule-enhanced sucrose synthasegene. J. Exp. Bot. 54, 749–755.
Simpson, B.B., Miao, B.-M., 1997. The circumscription of Hoffmannseggia (Fabaceae,Caesalpinioideae, Caesalpinieae) and its allies using morphological and cpDNArestriction site data. Plant Syst. Evol. 205, 157–178.
Simpson, B.B., Larkin, L.L., Weeks, A., 2003. Progress towards resolving relationshipsof the Caesalpinia group (Caesalpinieae: Caesalpinioideae: Leguminosae). In:Klitgaard, B., Bruneau, A. (Eds.), Advances in Legume Systematics, Part 10. RoyalBotanic Gardens, Kew, pp. 123–148.
Simpson, B.B., Lewis, G.P., 2003. New combinations in Pomaria (Caesalpinioideae:Leguminosae). Kew. Bull. 58, 175–184.
Simpson, B.B., Tate, J.A., Weeks, A., 2004. Phylogeny and character evolution ofHoffmannseggia (Caesalpinieae: Caesalpinioideae: Leguminosae). Syst. Bot. 29,933–946.
Simpson, B.B., Ulibarri, E.A., 2006. A synopsis of the genus Hoffmannseggia(Leguminosae). Lundellia 9, 7–33.
Singer, S.R., Maki, S.L., Farmer, A.D., Ilut, D., May, G.D., Cannon, S.B., Doyle, J.J., 2009.Venturing beyond beans and peas: what can we learn from Chamaecrista? PlantPhysiol. 151, 1041–1047.
Sprent, J.I., 2001. Nodulation in Legumes. Royal Botanic Gardens, Kew, London.Sprent, J.I., 2009. Legume Nodulation: A Global Perspective. Wiley-Blackwell, West
Sussex, UK, pp. 3–7.Steele, K.P., Wojciechowski, M.F., 2003. Phylogenetic analyses of tribes Trifolieae
and Vicieae, based on sequences of the plastid gene matK (Papilionoideae:Leguminosae). In: Klitgaard, B., Bruneau, A. (Eds.), Advances in LegumeSystematics, Part 10. Royal Botanic Gardens, Kew, pp. 355–370.
Steele, P.R., Guisinger-Bellian, M., Linder, C.R., Jansen, R.K., 2008. Phylogeneticutility of 141 low-copy nuclear regions in taxa at different taxonomiclevels in two distantly related families of rosids. Mol. Phylogenet. Evol.48, 1013–1026.
Takemiya, A., Kinoshita, T., Asanuma, M., Shimazaki, K., 2006. Protein phosphatase 1positively regulates stomatal opening in response to blue light in Vicia faba.Proc. Natl. Acad. Sci. USA 103, 13549–13554.
Thompson, J.D., Gibson, T.J., Plewniak, F., Jeanmougin, F., Higgins, D.G., 1997. TheCLUSTAL_X windows interface: flexible strategies for multiple sequencealignment aided by quality analysis tools. Nucl. Acids Res. 25, 4876–4882.
Tun, Y.T., Yamaguchi, Y., 2007. Phylogenetic relationship of wild and cultivatedVigna (Subgenus Ceratotropis, Fabaceae) from Myanmar based on sequencevariations in non-coding regions of trnT-F. Breed Sci. 57, 271–280.
Vissi, E., Toth, E.C., Kovacs, I., Magyar, Z., Horvath, G.V., Bagossi, P., Gergely, P.,Dudits, D., Dombradi, V., 1998. Protein phosphatase 1 catalytic subunit isoformsfrom alfalfa: biochemical characterization and cDNA cloning. Arch. Biochem.Biophys. 360, 206–214.

162 V. Manzanilla, A. Bruneau / Molecular Phylogenetics and Evolution 65 (2012) 149–162
Wilgenbusch, J.C., Warren, D.L., Swofford, D.L., 2004. AWTY: A System for GraphicalExploration of MCMC Convergence in Bayesian Phylogenetic Inference. <http://ceb.csit.fsu.edu/awty>.
Wojciechowski, M.F., Lavin, M., Sanderson, M.J., 2004. A phylogeny of legumes(Leguminosae) based on analysis of the plastid matK gene resolves many well-supported subclades within the family. Am. J. Bot. 91, 1846–1862.
Xia, X., Xie, Z., 2001. DAMBE: software package for data analysis in molecularbiology and evolution. J. Hered. 92, 371–373.
Xia, X., Xie, Z., Salemi, M., Chen, L., Wang, Y., 2003. An index of substitutionsaturation and its application. Mol. Phylogenet. Evol. 26, 1–7.
Yang, Z., 2006. Computational Molecular Evolution. Oxford Series in Ecology andEvolution, Oxford, 357 pp.
Yang, Z., 2007. PAML 4: phylogenetic analysis by maximum likelihood. Mol. Biol.Evol. 24, 1586–1591.
Yu, J.K., La Rota, M., Kantety, R.V., Sorrells, M.E., 2004. EST derived SSR markers forcomparative mapping in wheat and rice. Mol. Genet. Genom. 271, 742–751.
Zhang, X.Q., Verma, D.P.S., Patil, S., Arredondo-Peter, R., Miao, G.H., Kuismanen, R.,Klucas, R.V., Chollet, R., 1997. The electronic plant gene register. Plant Physiol.115, 1729–1731.





![[2015] Copies of Filip Stanislavov’s Abagar (Rome, 1651)](https://img.pdfslide.net/doc/110x75/634ad92c9fa0ab3f6b082ff1/2015-copies-of-filip-stanislavovs-abagar-rome-1651.jpg)























