Embed Size (px)
Citation preview

NORTH AFRICAN COASTAL LAGOONS
Environmental influences on the qualitative and quantitativecomposition of phytoplankton and zooplankton in NorthAfrican coastal lagoons
M. Ramdani Æ N. Elkhiati Æ R. J. Flower ÆJ. R. Thompson Æ L. Chouba Æ M. M. Kraiem ÆF. Ayache Æ M. H. Ahmed
Published online: 22 January 2009
� Springer Science+Business Media B.V. 2009
Abstract Within the framework of the international
research project MELMARINA, seasonal dynamics of
plankton communities in three North African coastal
lagoons (Merja Zerga, Ghar El Melh, and Lake
Manzala) were investigated. The sampling period
extended from July 2003 to September 2004 with the
aim of evaluating hydrological and other influences on
the structure, composition and space-time develop-
ment of these communities in each lagoon.
Phytoplankton in Merja Zerga showed a quasi-perma-
nent predominance of marine diatoms in the open sea
station and in the marine inlet channel. Dinoflagellates
were abundant in summer and early autumn in the
marine inlet and extended into the central lagoon
station. In Ghar El Melh, marine species (especially
diatoms and dinoflagellates) dominated despite occa-
sional winter inflows of freshwater. In Lake Manzala,
freshwater species generally predominated and the
planktonic communities were comparatively very
diverse. Chlorophyceae contributed 39% of the total
species recorded and diatoms and cyanophyceans were
also common; the Dinophyceae, Euglenophyceae,
Chrysophyceae and Cryptophyceae less so. Zooplank-
ton communities in both Ghar El Melh and Merja
Zerga were dominated by marine copepods. Rotifera,
Copepoda, Ostracoda, and Cladocera were recorded in
both lagoons as were meroplanktonic larvae of Poly-
chaeta, Cirripedia, Mysidacea and Gastropoda and free
living nematodes. Ghar El Melh was the more
productive of these two lagoons with spring and early
Guest editors: J. R. Thompson & R. J. Flower
Hydro-ecological Monitoring and Modelling of North African
Coastal Lagoons
Electronic supplementary material The online version ofthis article (doi:10.1007/s10750-008-9678-4) containssupplementary material, which is available to authorized users.
M. Ramdani (&)
Department of Zoology & Animal Ecology, Institut
Scientifique, University Mohamed V, Charia Ibn Batouta,
Rabat Agdal BP703, Morocco
e-mail: [email protected]
N. Elkhiati
Faculte des Sciences, Biologie, Universite Hassan II Ain
Chock, Km 8, Route El Jadida, Casablanca, Morocco
R. J. Flower � J. R. Thompson
UCL Department of Geography, Environmental Change
Research Centre/Wetland Research Unit, University
College London, Gower Street, London WC1E 6BT, UK
L. Chouba � M. M. Kraiem
INSTM, Rue de 2 mars 1934, No. 28, Salammbo, Tunis
2025, Tunisia
F. Ayache
Department de Geography, Faculte des Lettres & Sciences
Humaines, Universite de Sousse, Sousse 4029, Tunisia
M. H. Ahmed
Department of Marine Resources, National Authority for
remote sensing and Space Sciences, 23 Josef Burrows
Tito Street, El Nozha, Al Gadida, PO Box 1654, Cairo,
Egypt
123
Hydrobiologia (2009) 622:113–131
DOI 10.1007/s10750-008-9678-4

summer being the productive seasons. Zooplankton
communities in Lake Manzala were generally domi-
nated by rotifers and highest zooplankton abundances
occurred in April (2003). Sampling stations near the
marine inlets showed the highest diversity and the
zooplankton communities showed considerable spatial
variation within this large lagoon. The three lagoons
represent very different water bodies contrasted
strongly in terms of tidal effects and freshwater
availability. Yet, there are some similarities in eco-
system structure. Space-time development of the
plankton communities was similar especially in Merja
Zerga and Ghar El Melh. Species abundances and
specific diversities indicated that seasonal changes in
salinity and nutrient concentrations were the main
influential factors. Lake Manzala was the most
productive lagoon and all the three sites supported
toxic algal species. Relatively low plankton biomass in
Merja Zerga and Ghar El Melh probably resulted from
a combination of factors including highly episodic
nutrient inputs, light suppression (by turbidity) and
nutrient competition with benthic algae. Water quality
variables were largely driven by the hydrological
regime specific to each lagoon. Nutrient enrichment
and, particularly for Lake Manzala, sea level rise
threaten the sustainability of the planktonic ecosys-
tems in all three lagoons.
Keywords Coastal lagoons � Water chemistry �Plankton � Hydrology � Monitoring
Introduction
Plankton communities in coastal lagoons are of major
importance in food web structures and for ecosystem
health. Coastal lagoons are often heavily impacted by
human activities and eutrophication is one of the
several processes that can degrade water quality and so
alter plankton species abundances (e.g. Delgado,
1990). Precise species changes are difficult to predict
(e.g. Gamito et al., 2005), but hydrological modifica-
tions of freshwater inflows and of marine inlet channels
can bring about major changes in lagoon ecosystems
and particularly in the plankton (Borja, 2005). The
structure of plankton communities in any coastal water
body is especially important, not the least for com-
mercial fisheries in relation to environmental change.
Phytoplankton is the main primary producer in most
lagoon ecosystems (Khalil, 1990; Romdhane et al.,
1998; Scheffer, 1998; Daly Yahia et al., 2001; Sakka
Hlaili et al., 2003; Mageed, 2006) and, as well as
biomass, the species composition of phytoplankton
communities influences the ecological status of a site,
with particular species being good indicators of either
high or low ecological status (Hutchinson, 1967, 1975;
Talling, 1976; Talling & Lemoalle, 1998; Fathi et al.,
2001; Borja, 2005; EC Water Framework Directive -
2000/60/E).
As a consequence of a dry climate and intensive
land use, coastal lagoons in the Southern Mediterra-
nean Region (SMR) are particularly vulnerable to
disturbance and environmental change. Notably, agri-
cultural exploitation of land around many SMR lakes
and lagoons has expanded strongly during the twen-
tieth Century (e.g. Flower, 2001). Freshwater
resources in the region have been generally impacted
by increasing water demand from a growing local
population and associated land use changes (Ayache
et al., 2009; Thompson et al., 2009). In the future these
impacting factors will be exacerbated by predicted
global climate change, including sea level rise.
Relatively little research has been undertaken on
aquatic ecosystems within SMR lagoons, although
some environmental factors influencing aquatic com-
munities have been investigated. Some studies
demonstrated human pressures and declining envi-
ronment quality in some lagoons (e.g. Meininger &
Mullie, 1981; Saad et al., 1985). In the late 1990s, the
CASSARINA Project (Flower, 2001) used sediment
records to reveal past changes in aquatic communities
in several North African lagoons (e.g. Birks et al.,
2001; Flower et al., 2001) and linked studies
evaluated the status of plankton (Fathi et al., 2001;
Ramdani et al., 2001a, b, c) and fish (Kraıem et al.,
2001). In 2002, follow-up work was initiated as part
of the MELMARINA (Monitoring and Modelling
Coastal Lagoons: Making Management Tools for
Aquatic Resources in North Africa) Project (Flower
& Thompson, 2009). The project (funded through the
EU INCO-Med programme) set base lines and
undertook integrated hydro-ecological monitoring at
selected sites to ascertain the impacts of environment
and management changes and to enable setting of
future scenarios through hydroecological modelling.
This article undertakes assessment of water quality
and describes the results of qualitative and quantita-
tive sampling of both phytoplankton and zooplankton
114 Hydrobiologia (2009) 622:113–131
123

communities in three lagoons, Merja Zerga (Mor-
occo), Ghar El Melh (Tunisia) and Lake Manzala
(Egypt).
Sites, materials and methods
The MELMARINA primary lagoons
Detailed accounts of the three MELMARINA
primary lagoons are provided by Ayache et al.
(2009). All the three lagoons have considerable value
for biodiversity, which include both fish and birds.
The Moroccan site, Merja Zerga (Fig. 1a), is linked
to the Atlantic Ocean by a single channel (the
‘‘gullet’’), that is strongly tidal and, at high tide, is
connected with a smaller lagoon, the Merja Khala.
Open water area at high tide is approximately
13.2 km2 (2003) and at low tide, large areas of mud
flats are exposed. Macroalgae (Enteromorpha and
Ulva) are very common on these mud flats and in
intertidal depressions. The mud flats are exploited for
edible molluscs (Ruditapes decussatus). The margins
of the lagoon are managed for agriculture, including
cattle and sheep grazing and cash crops. Merja Zerga
is well known as a bird reserve and as an area of
outstanding natural beauty. Its hydrology and vege-
tation have been studied since the 1970s and the
plankton since 1986 (Ramdani, 1988; Ramdani et al.,
2001a, b; Fraikech et al., 2005).
Ghar El Melh, on the northeast coast of Tunisia
(Fig. 1b), has also received considerable attention
regarding water quality and plankton and the large
migrant bird populations (Morgan & Boy, 1980; Daly
Yahia & Romdhane, 1994; Souissi et al., 2000; Daly
Yahia et al., 2001; Benrejeb-Jenhani & Romdhane,
2002). The lagoon, which has a total area of approx-
imately 35.6 km2, is separated from the Mediterranean
by sand bars through which there is one main and
several smaller connections to the sea. The lagoon is
subject to the relatively small Mediterranean tidal
regime. It is mainly surrounded by agricultural land
and the littoral margins support abundant growth of
macroalgae (Enteromorpha and Ulva), especially on
the north and western shores. In summer months, a
mat of filamentous green algae develops over much
of the submerged surface sediments. Occasional large
winter freshwater inflows from the diverted Mejerda
River occur (Ayache et al., 2009) and can temporarily
lower lagoon salinity. In general, however, salinity
remains near that of seawater, fluctuating between 27
and 48 % annually. The freshwater inflows deliver
sediment and agro-chemicals from farmland. Birds
frequent the shallow SE part of the lagoon and the
small fishery comprises eel, mullet, sea-bass and sole
(Kraıem et al., 2009).
Egyptian Lake Manzala is the largest of the four
Nile Delta lakes with an area of c. 700 km2 (Fig. 1c). It
has been much reduced by twentieth Century recla-
mation, and Meininger & Mullie (1981) noted that the
lake formerly occupied 1710 km2. It is shallow
(average depth c. 1 m with 25% of the area \ 60 cm
deep, Shaheen & Yosef, 1987). The lake is linked to
the Mediterranean by several channels the largest of
which is at El Gamil. In the southern sector, there are a
number of drains, including the grossly polluted Bahr
Fig. 1 Sample locations within the MELMARINA lagoons (a) Merja Zerga (Morocco), (b) Ghar El Melh (Tunisia), (c) Lake
Manzala (Egypt)
Hydrobiologia (2009) 622:113–131 115
123

El Baqar, which discharge freshwater to the lake.
Much of the northwestern and eastern sectors are now
wholly or partly reclaimed or converted to fish
farming. Similarly, extensive areas of reclaimed lake
bed, which are not intensively farmed, border the lake
to the south. The lagoon still retains extensive
emergent and submerged aquatic plant communities
and provides approximately 50% of the total inland
fisheries within Egypt. It is highly threatened by
pollution (Khalil & Salib, 1986; Shaheen and Yosef,
1987), and sea level rise and geological subsidence
(Stanley & Warne, 1994; Saad et al., 1985). A number
of studies of Lake Manzala’s plankton have been
undertaken (El-Hawary 1960; Samaan & Aleem,
1972; Samaan, 1977; El-Sherif & Aboul-Ezz, 1988;
Radwan & Popiolek, 1989; Khalil, 1990; Guerguess,
1993; Aboul-Ezz, 1995; El-Naggar et al., 1997;
Gharib & Soliman, 1998; Aboul-Ezz & Soliman,
2000; Zaghloul & Hussein, 2000; El-Sherif & Gharib,
2001; Fathi et al., 2001, Khalifa & Mageed, 2002;
Abdel-Aziz & Aboul-Ezz, 2004, Mageed, 2006;
Zakaria et al., 2007).
Water quality monitoring
Within each lagoon representative water sampling
stations were selected: four in Merja Zerga, five in
Ghar El Melh, and ten in Manzala. All stations were
sampled at approximately monthly intervals from July
2003 to September 2004. For tidal Merja Zerga,
sampling was undertaken at approximately the same
time in the tidal cycle. Water clarity was assessed by
Secchi disc. Dissolved oxygen was determined by the
azide modification of the Winkler method (according
to APHA, 1995). Water temperature, pH, conductiv-
ity, total dissolved solids (g l-1) and dissolved oxygen
were measured using field probes (EXTECH, YSI or
Mettler Toledo). Water quality and phytoplankton
samples were collected from the subsurface water
column by hand dipping a one-litre sample bottle.
Samples for nitrate, total nitrogen (TN), inorganic
phosphorus and chlorophyll-a were treated according
to Grasshoff (1976). Before assay, samples were
filtered through 47 mm GF/C filters (nominal porosity
1.2 lm). In the laboratory, analyses were made using
a LKB 4050 spectrophotometer according to standard
procedures (Parsons et al., 1984) and results were
expressed in mg l-1. Total phosphorus (TP) was
assayed by spectrophotometry after reaction with
ammonium molybdate and reduction by ascorbic acid
(Murphy & Riley, 1962). Nitrate was assayed by ionic
chromatography using a Shimadzu ion chromatograph
HIC-6A (limit of detection for NO3 was less than
1.0 mg l-1). This apparatus was equipped with an ion-
exchange resin that is stable to a wide range of salt
concentrations. Appropriate blanks and standards
were used to calibrate each analytical method (APHA,
1995; Souissi et al., 2000; Fathi et al., 2001).
Plankton sampling
Phytoplankton samples were concentrated by sedi-
mentation (Sournia 1986; Hallegraeff et al., 1995).
Identification and enumeration were done under an
inverted microscope (Axiovert 25C—Egypt and
Olympus C-K2—Morocco and Tunisia) at 6009
magnification. Identification of the species and their
taxonomic categories were according to Tregouboff &
Rose (1957), Sournia (1986), Ricard (1987), Delgado
& Fortuna (1991), Hallegraeff et al. (1995) and Tomas
(1997). All cell counts were corrected to abundances in
1 litre original sampling volume. Zooplankton sam-
ples were collected at the same time as phytoplankton.
For Ghar El Melh and Merja Zerga, plankton samples
for Copepods, Cladocera and Rotifera were collected
with a standard 20 cm diameter plankton net (55-lm
mesh size) drawn through c. 20 m of open water at
each water quality sampling station (Fig. 1). For Lake
Manzala, zooplankton samples were collected by
filtering 50 l of lake water through a standard plankton
net (55-lm mesh size) and fixing in 4% formalin. A
Dollfus Vat was used to fractionate the samples and to
facilitate counting of the zooplankton (Ramdani, 1988;
Paterson, 1993, Ramdani et al., 2001b). The number of
species of Copepoda, Cladocera and Rotifera as well
as other zooplanktoners was estimated using tech-
niques from Dussart (1969), Rougier & Lam Hoai
(1997) and Ramdani et al. (2001c). For Manzala,
samples were concentrated to 100 ml prior to micro-
scopical examination. For specific enumeration
purposes, at least two 2-ml aliquots were used and
the total number of zooplankton determined so that
abundances and total numbers per m-3 could be
calculated. Species identifications and their taxonomic
categories were according to Tregouboff & Rose
(1957), Hutchinson (1967, 1975) and Dussart (1969).
Toxic phytoplankton species were identified using the
Hallegraeff et al. (1995) classification.
116 Hydrobiologia (2009) 622:113–131
123

Statistical analyses
A matrix of the normalized physical, chemical and
biological data was produced and analysed by means
of principal components analysis (PCA) using SPSS
software version 9.01n. This type of statistical
analysis is used when uniform linear relationships
are likely to exist between variables (Doledec &
Chessel, 1989). Differences between the three study
lagoons were tested by one-way analysis of variance
(ANOVA).
Results
Hydrochemistry
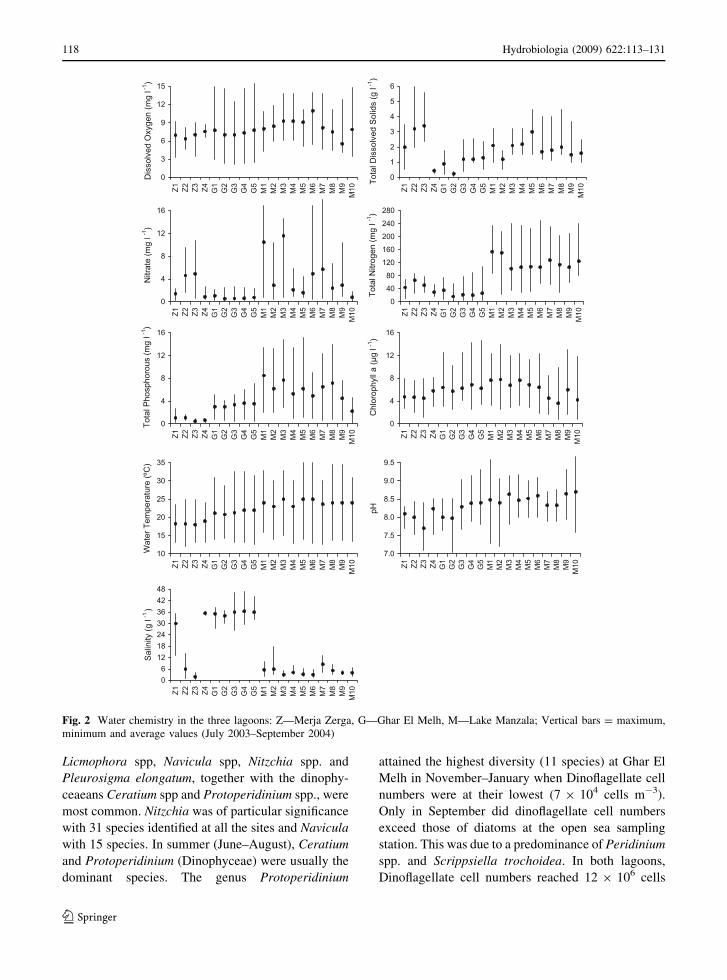
Water quality data are summarized for each sampling
station at all three sites in Fig. 2 and Supplementary
Material Table 1. This figure provides the maximum,
minimum and average values. The principal charac-
teristics of these data are described as follows:
Merja Zerga: Water temperature (12.1–23.5�C)
and pH values (7.7–8.4) varied seasonally. Open water
salinity was higher in summer (35.7 %) and the lowest
in autumn (13.2 %) in station Z1. Open water showed
only slight variations in salinity during the year (c. 35
%). Sample sites Z2 and Z3 near drainage inflows
were fresher and the salinity fluctuated between 2 and
12 % (Fig. 2). The maximum oxygen concentration
was recorded in autumn (12.6 mg l-1). Chlorophyll-a
peaked in spring–summer at 7.6 lg l-1. Total soluble
salts were higher in summer but dropped to minimum
levels through the autumn. Total phosphorus concen-
trations showed no regular trend but were the lowest in
the autumn (at c. 0.5 mg l-1). The maximum nitrate
concentration (10.7 mg l-1) was recorded in spring
adjacent to the Nador Canal (Fig. 2). Water transpar-
ency was higher in summer and lower in autumn and
winter at all stations and, during the flood period
(November and December); Secchi disc depth at Z2
and Z3 was nearly 1 cm but reached 45 cm in the
summer period.
Ghar El Melh: The maximum water temperature
(32�C) was recorded in summer (July). Water pH
values were from 7.3 (winter) to 9.3 (summer) and
salinities ranged from 43.5 % (summer) to below 29
% (winter). Oxygen concentration was around
15 mg l-1 in late autumn but increased during summer
when phytoplankton activity was high. Chlorophyll-a
values showed only small fluctuations during the year
(3.0–12.3 lg l-1). Total phosphorus concentrations
were higher (0.36–7.04 mg l-1) than in Merja Zerga
(0.5–1.6 mg l-1). Maximum nitrate concentrations (c.
50 mg l-1) were recorded in spring. Water transpar-
ency was greater during spring and summer when
Secchi disc depths were[50 cm.
Lake Manzala: Water temperature ranged from
15�C in winter to a maximum of 31.3�C in summer.
pH varied from 6.3 to 9.3. Salinity was low and
varied from 1.4 % in spring to 20.3 % in summer (at
station M2). Open water was always well oxygenated
during day time and varied from 6.7 mg l-1 in
summer to 15.3 mg l-1 in winter. The total soluble
salts were higher in summer and dropped to minimum
levels through the autumn. Total phosphorus concen-
trations showed no regular trends but were high at all
stations at around 8.5 mg l-1. The highest nitrates
concentration (10.7 mg l-1) was recorded in spring.
Water transparency was higher in the northwest part
of the lagoon where macrophytes were present
(Secchi disc depth usually[60 cm or to the sediment
surface) and lower in the turbid eastern part of the
lake (typically \25 cm).
Phytoplankton composition and abundance
A total of 314 phytoplankton taxa were identified at
the three lagoons. These included 68 Dinophyceae,
131 Bacillariophyceae, 29 Cyanophyceae, 67 Chlo-
rophyceae, 8 Euglenophyceae, 3 Chrysophyceae, 6
Cryptophyceae and 2 Silicoflagellate taxa (see
Supplementary Material Tables 2–5).
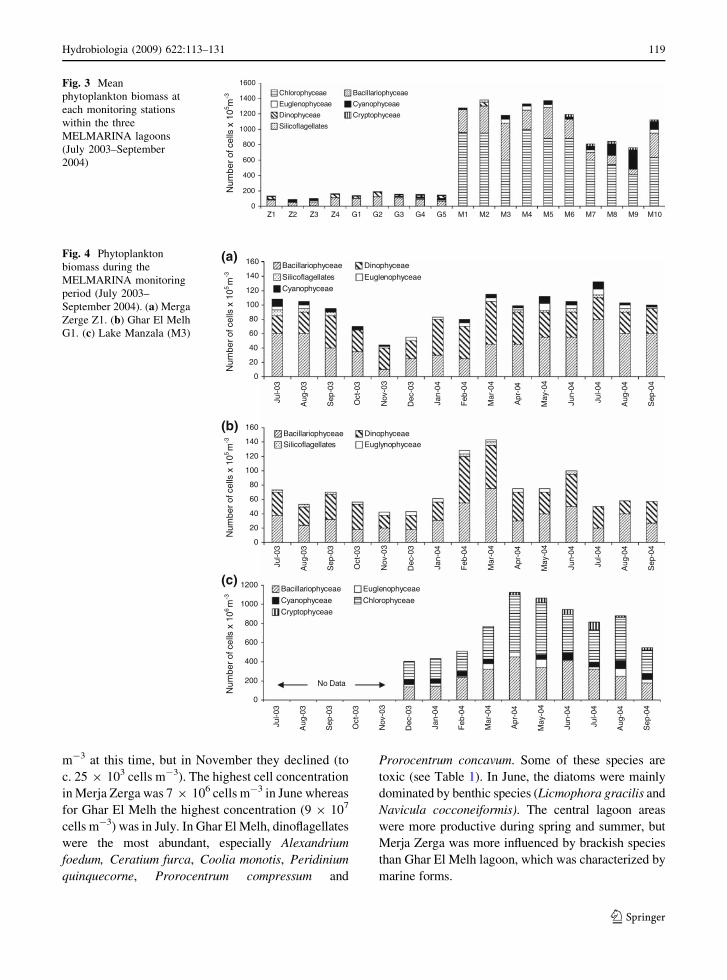
Merja Zerga and Ghar El Melh: Diatoms were
dominant in terms of the number of species and
abundance in these two lagoons (Fig. 3). The highest
number of species occurred in spring and summer in
both lagoons, but in summer and autumn the number
of Dinophyceae species usually exceeded that of the
diatoms. The lowest number of phytoplankton species
in Merja Zerga occurred in November and December
(Fig. 4). In terms of phytoplankton species succession,
the diatoms were most abundant during April–May
(Pseudonitzschia pseudodelicatissima, Nitzschia spp.,
Rhizosolenia sp.) as was the dinophyceaean Ceratium.
In early April, species of Chaetoceros and Navicula
were dominant in terms of both abundance and
the number of the species. In summer the diatoms
Hydrobiologia (2009) 622:113–131 117
123
Licmophora spp, Navicula spp, Nitzchia spp. and
Pleurosigma elongatum, together with the dinophy-
ceaeans Ceratium spp and Protoperidinium spp., were
most common. Nitzchia was of particular significance
with 31 species identified at all the sites and Navicula
with 15 species. In summer (June–August), Ceratium
and Protoperidinium (Dinophyceae) were usually the
dominant species. The genus Protoperidinium
attained the highest diversity (11 species) at Ghar El
Melh in November–January when Dinoflagellate cell
numbers were at their lowest (7 9 104 cells m-3).
Only in September did dinoflagellate cell numbers
exceed those of diatoms at the open sea sampling
station. This was due to a predominance of Peridinium
spp. and Scrippsiella trochoidea. In both lagoons,
Dinoflagellate cell numbers reached 12 9 106 cells
Fig. 2 Water chemistry in the three lagoons: Z—Merja Zerga, G—Ghar El Melh, M—Lake Manzala; Vertical bars = maximum,
minimum and average values (July 2003–September 2004)
118 Hydrobiologia (2009) 622:113–131
123
m-3 at this time, but in November they declined (to
c. 25 9 103 cells m-3). The highest cell concentration
in Merja Zerga was 7 9 106 cells m-3 in June whereas
for Ghar El Melh the highest concentration (9 9 107
cells m-3) was in July. In Ghar El Melh, dinoflagellates
were the most abundant, especially Alexandrium
foedum, Ceratium furca, Coolia monotis, Peridinium
quinquecorne, Prorocentrum compressum and
Prorocentrum concavum. Some of these species are
toxic (see Table 1). In June, the diatoms were mainly
dominated by benthic species (Licmophora gracilis and
Navicula cocconeiformis). The central lagoon areas
were more productive during spring and summer, but
Merja Zerga was more influenced by brackish species
than Ghar El Melh lagoon, which was characterized by
marine forms.
0
200
400
600
800
1000
1200
1400
1600
Z1 Z2 Z3 Z4 G1 G2 G3 G4 G5 M1 M2 M3 M4 M5 M6 M7 M8 M9 M10
Num
ber
of c
ells
x 1
05 m-3 Chlorophyceae Bacillariophyceae
Euglenophyceae Cyanophyceae
Dinophyceae Cryptophyceae
Silicoflagellates
Fig. 3 Mean
phytoplankton biomass at
each monitoring stations
within the three
MELMARINA lagoons
(July 2003–September
2004)
0
20
40
60
80
100
120
140
160
Jul-0
3
Aug
-03
Sep
-03
Oct
-03
Nov
-03
Dec
-03
Jan-
04
Feb
-04
Mar
-04
Apr
-04
May
-04
Jun-
04
Jul-0
4
Aug
-04
Sep
-04
Num
ber
of c
ells
x 1
06m
-3N
umbe
r of
cel
ls x
105
m-3
Num
ber
of c
ells
x 1
05m
-3
Bacillariophyceae Dinophyceae
Silicoflagellates Euglenophyceae
Cyanophyceae
0
20
40
60
80
100
120
140
160
Jul-0
3
Aug
-03
Sep
-03
Oct
-03
Nov
-03
Dec
-03
Jan-
04
Feb
-04
Mar
-04
Apr
-04
May
-04
Jun-
04
Jul-0
4
Aug
-04
Sep
-04
Bacillariophyceae DinophyceaeSilicoflagellates Euglynophyceae
0
200
400
600
800
1000
1200
Jul-0
3
Aug
-03
Sep
-03
Oct
-03
Nov
-03
Dec
-03
Jan-
04
Feb
-04
Mar
-04
Apr
-04
May
-04
Jun-
04
Jul-0
4
Aug
-04
Sep
-04
Bacillariophyceae Euglenophyceae
Cyanophyceae Chlorophyceae
Cryptophyceae
No Data
(a)
(b)
(c)
Fig. 4 Phytoplankton
biomass during the
MELMARINA monitoring
period (July 2003–
September 2004). (a) Merga
Zerge Z1. (b) Ghar El Melh
G1. (c) Lake Manzala (M3)
Hydrobiologia (2009) 622:113–131 119
123

The phytoplankton communities at Lake Manzala
were diverse and were of essentially freshwater
species. They were represented by 163 species
belonging to seven classes (Supplementary Material
Tables 2–5). The green algae were dominant, but
diatoms and cyanophytes were common at stations
M6–M10 (Fig. 3). Other groups (7 Dinophyceae, 5
Euglenophyceae, 2 Chrysophyceae, 2 Cryptophyceae
and 1 Silicoflagellate species) were minor constitu-
ents of the phytoplankton. The green algae
constituted c. 48% of the total species recorded (67
species) and c. 77% of the total phytoplankton
abundance during spring with 91 9 106 cells m-3
at station M4. The western part of Lake Manzala
(stations M7, M8 and M9) supported lower phyto-
planktonic abundances compared to the other
stations, while stations 1, 2 and 5 attained maximum
population densities during the period of investiga-
tion. A gradual increase in phytoplankton population
densities was observed from March till June; the
highest density (780 9 107 cells m-3) was recorded
in April at M3. Diatoms ranked second in terms of
both species occurrence (c. 27%; 48 species) and the
total abundance (c. 18%; 98 9 107 cells m-3)
(Fig. 4). Cyanophytes formed c. 15% of the total
species recorded and c. 4 9 107 cells m-3 of the total
phytoplankton abundance. The green algae Ankis-
trodesmus spp, Dictyosphaerium pulchellum,
Kirchneriella spp, Pyramimonas spp, Scenedesmus
spp and Selenastrum spp were the most important
algae in the Lake Manzala communities. Diatoms
were typically freshwater or brackish water species
and mainly represented by the three taxa Cyclotella
meneghiniana, Nitzschia closterium and N. frustulum
var. perpusilla (at stations M3, M4 and M5). The
most important cyanophyceans were Microcystis aeru-
ginosa, Rhabdoderma lineare var. unicella and
Spirulina major and these characterized the western
part of the lake (stations M7 to M10). Cryptophyceae
were represented by Chroomonas acuta, C. erosa,
C. vata, Cryptomonas sp. and Rhodomonas sp. Euglen-
ophyceae (Euglena acus, E. clara, E. hemichromata,
E. proxima and E. viridi) were also represented, mostly
in the western and central parts of the lake.
Toxic species of phytoplankton were recorded in
all the three lagoons and, following the UNESCO
(Hallegraeff et al., 1995) classification, seven groups
of harmful algae were identified (Table 1). These
occurred mainly during summer and autumn. Dino-
flagellates and, to a lesser extent cyanophyceans,
comprised the most harmful groups and particularly
noteworthy are the dinoflagelate Gonyaulax spinifera
Table 1 Phytoplankton toxic species identified in the three lagoons following the classification of UNESCO (Hallegraeff et al.,
1995)
Useful,
but mostly
harmless
Oxygen
depletion
Paralytic
shellfish
poisoning
Diarrheic
shellfish
poisoning
Neurotoxic
shellfish
poisoning
Amnesic
shellfish
poisoning
Ciguatera
fish
poisoning
Gonyaulax spinifera GEM
Prorocentrum micans GEM, Zerga
Dinophysis caudata GEM, Zerga
Alexandrium tamarense GEM
Gymnodinium catenatum Zerga
Dinophysis acuminata GEM, Zerga
Dinophysis caudata GEM, Zerga
Gymnodinium breve Zerga
Pseudonitzschia spp GEM, Zerga
Coolia monotis GEH
Osteopsis sp. Zerga
Prorocentrum lima GEM, Zerga
Amphidinium sp. Zerga
Microcystis flos-aque Manzala
Microcystis aeruginosa Manzala
120 Hydrobiologia (2009) 622:113–131
123

in Ghar El Melh, Dinophysis species in both Ghar El
Melh and Merja Zerga, and Microcystis aeroginosa
and M. flos-aquae in Lake Manzala.
Zooplankton composition and abundance
Analysis of the zooplankton communities in the three
lagoons revealed five major groups: Copepoda,
Rotifera, Cladocera, Ostracoda and Protozoa
(Supplementary Material Tables 6–8). Other marine
representatives (meroplanktonic larval Polychaeta,
Cirripedia, Mysidacea and Gastropoda and the free-
living nematods) were also occasionally encountered
in some samples. Merja Zerga and Ghar El Melh
lagoons were both dominated by copepods (48 and 55
species, respectively). Copepods were mainly marine
species (Supplementary Material Table 6) in both
these lagoons, but more freshwater Cladocera and
rotifers were found in samples from the stations (Z2
and Z3) near the Oued Drader and Nador Canal in
Merja Zerga (Supplementary Material Tables 7–8).
These taxa were either very rare or totally absent in
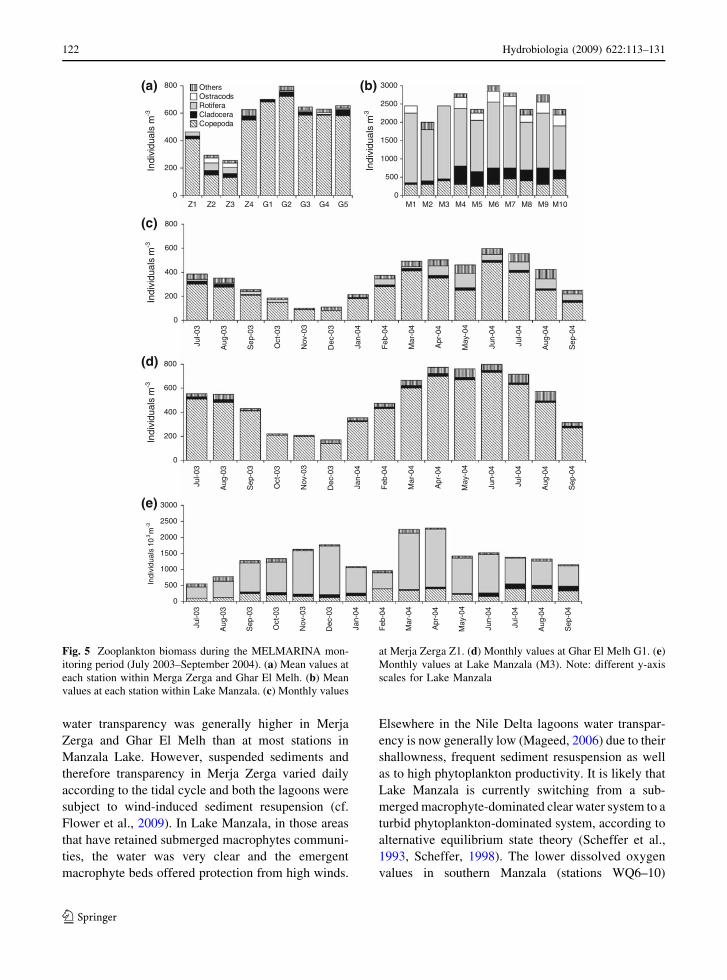
Ghar El Melh. The minimum zooplankton standing
crop was 75 ind m-3 in Merja Zerga (Z1) during
November and December (Fig. 5). Peaks occurred
during spring and early summer, with a minimum
standing crop of 130 ind m-3 and maximum abun-
dance of c. 700 ind m-3 in April, May and June.
Open sea stations in both Merja Zerga and Ghar El
Melh were often more diverse and more productive
than in the central areas of each lagoon.
At Lake Manzala, Rotifera were remarkably abun-
dant and constituted the main group of zooplanktoners
comprising c. 80% of the total number in all the
sampled stations (Fig. 5). Five species dominated the
Rotifera: Brachionus angularis, B. urceolaris,
B. calyciflorus, B. plicatilis and Colurella adriatica
and were most common in the southern part of the
lagoon. The genus Brachionus was represented by
eight species in the western part of the lake (stations
M6–M10). The Copepods constituted the second most
abundant group forming c. 14% of the total number of
zooplankton individuals. The average zooplankton
standing crop at Manzala was relatively high with a
minimum of 50 9 104 ind m-3 in February and
August and a maximum of c. 300 9 104 ind m-3 in
April at M6. The marine copepod species Euterpina
acutifrons and Oithona nana were only recorded in
the northern part of the lake near the El Gamil
connection with the Mediterranean.
Discussion
All the three of the MELMARINA primary lagoons
share some similarities in terms of location and near
catchment landscape. They all occur at the termini of
inland drainage systems where land is modified
principally for agriculture by land drainage. Hydrol-
ogy exerts a major influence on the environmental
conditions at each lagoon. Merja Zerga is subjected to
daily flushing by Atlantic tides whereas tidal effects
are much less at Ghar El Melh and Lake Manzala
because of the smaller Mediterranean tidal regime.
Lake Manzala is differentiated by its freshwater
character as a result of large discharges from land
drains along the lake’s southern margin (Ayache
et al., 2009; Thompson et al., 2009). These differ-
ences have major implications for both the water
quality and the plankton of the three lagoons, and
these are now considered and relationships are
evaluated using statistical methods.
Water quality
The concentrations of nutrients in the three lagoons
and their variations are largely controlled by the
quality of drainage water entering and leaving the
lagoon and the uptake of nutrients by plant assimila-
tion. The concentrations and distribution of nutrients
can be strongly affected by the mixing of the water
through the lagoon–sea connection (Thompson et al.,
2009). Absorption on suspended sediment and sub-
sequent deposition/resuspension may also play an
important role in determining nutrient concentrations
in lagoon water. Significant sources of nitrate and total
phosphorus for Merja Zerga are the drainage waters
from the Nador Canal and Oued Drader inflows. For
Ghar El Melh, nutrients enter from drainage of the
agricultural Utica floodplain as well from several
significant point sources that include the town of Ghar
El Melh and, exceptionally, from overflow of the
Mejerda River. Within Lake Manzala, the Bahr El
Baqar drain is a well-known source of urban waste
water with high concentrations of total phosphorus
and nitrate (Fathi et al., 2001; Mageed, 2006). Except
during freshwater floods and otherwise poor weather,
Hydrobiologia (2009) 622:113–131 121
123
water transparency was generally higher in Merja
Zerga and Ghar El Melh than at most stations in
Manzala Lake. However, suspended sediments and
therefore transparency in Merja Zerga varied daily
according to the tidal cycle and both the lagoons were
subject to wind-induced sediment resupension (cf.
Flower et al., 2009). In Lake Manzala, in those areas
that have retained submerged macrophytes communi-
ties, the water was very clear and the emergent
macrophyte beds offered protection from high winds.
Elsewhere in the Nile Delta lagoons water transpar-
ency is now generally low (Mageed, 2006) due to their
shallowness, frequent sediment resuspension as well
as to high phytoplankton productivity. It is likely that
Lake Manzala is currently switching from a sub-
merged macrophyte-dominated clear water system to a
turbid phytoplankton-dominated system, according to
alternative equilibrium state theory (Scheffer et al.,
1993, Scheffer, 1998). The lower dissolved oxygen
values in southern Manzala (stations WQ6–10)
0
200
400
600
800
Z1 Z2 Z3 Z4 G1 G2 G3 G4 G5
Indi
vidu
als
m-3
Indi
vidu
als
m-3
Indi
vidu
als
m-3
OthersOstracodsRotiferaCladoceraCopepoda
0
500
1000
1500
2000
2500
3000
M1 M2 M3 M4 M5 M6 M7 M8 M9 M10
Indi
vidu
als
m-3
0
200
400
600
800
Jul-0
3
Aug
-03
Sep
-03
Oct
-03
Nov
-03
Dec
-03
Jan-
04
Feb
-04
Mar
-04
Apr
-04
May
-04
Jun-
04
Jul-0
4
Aug
-04
Sep
-04
0
200
400
600
800
Jul-0
3
Aug
-03
Sep
-03
Oct
-03
Nov
-03
Dec
-03
Jan-
04
Feb
-04
Mar
-04
Apr
-04
May
-04
Jun-
04
Jul-0
4
Aug
-04
Sep
-04
0
500
1000
1500
2000
2500
3000
Jul-0
3
Aug
-03
Sep
-03
Oct
-03
Nov
-03
Dec
-03
Jan-
04
Feb
-04
Mar
-04
Apr
-04
May
-04
Jun-
04
Jul-0
4
Aug
-04
Sep
-04
Indi
vidu
als
103m
-3
(a) (b)
(c)
(d)
(e)
Fig. 5 Zooplankton biomass during the MELMARINA mon-
itoring period (July 2003–September 2004). (a) Mean values at
each station within Merga Zerga and Ghar El Melh. (b) Mean
values at each station within Lake Manzala. (c) Monthly values
at Merja Zerga Z1. (d) Monthly values at Ghar El Melh G1. (e)
Monthly values at Lake Manzala (M3). Note: different y-axis
scales for Lake Manzala
122 Hydrobiologia (2009) 622:113–131
123

provide further evidence of eutrophication and decom-
position of excess organic matter. Because of its size
and array of nutrient sources, Lake Manzala showed
the highest values and the greatest spatial variation in
nutrients compared to the Merja Zerga and Ghar El
Melh lagoons.
The concentrations of total phosphorus indicated
the pattern of drainage water distribution in Lake
Manzala. Higher concentrations of total phosphorus
generally occurred in the southern and eastern zones of
the lake (near to the drainage canals and to the waste
water treatment works). Nitrate concentrations were
also relatively high in stations near the drains. They
reached their highest values during spring–summer
while total phosphorus concentrations peaked after-
wards. Nutrient concentrations in general declined in
late summer probably due to increased uptake by
phytoplankton and benthic algae and by the sub-
merged and emergent macrophytes. In Lake Manzala,
nutrient concentrations generally decreased towards
the sea connections and probably indicate several
processes: adsorption and sedimentation, plant assim-
ilation and flushing through the El Gamil channels.
Guerguess (1993) and Ahmed et al. (2009) identify the
drains as the main source of phosphate to the lake, and
the data presented here indicate that the nutrient-rich
drainage water is diluted and distributed across the
lake in a northerly direction. This pattern is modified
by emergent vegetation stands and water circulation
(Rasmussen et al., 2009a, b). Contaminated freshwater
is undoubtedly the major source of nutrients in all the
lagoons although in some periods nutrients may be
transported in from the coastal zone (i.e. as suspected
for Ghar El Melh, Rasmussen et al., 2009b). Nutrient
concentrations recorded in Ghar El Melh were lower
than the previous findings for Bizerte lagoon (Sakka
Hlaili et al., 2003). For Merja Zerga, the highest
nutrient concentrations were recorded adjacent to the
Canal Nador and Oued Drader. As indicated by
modelling results (Rasmussen et al., 2009a, b),
reduction of nutrient inputs, especially phosphorus,
is needed in these coastal lagoons.
In order to reveal correlations between the various
physico-chemical parameters measured in the three
lagoons, ADE4 software (Data Analysis functions to
analyse Ecological and Environmental data in the
framework of Euclidean Exploratory methods) was
applied. Twelve environmental variables (descriptors)
were used: temperature (�C), pH, salinity (%),
dissolved oxygen (mg l-1), suspended sediment
(mg l-1), nitrate (mg l-1), total nitrogen (mg l-1),
total phosphorus (mg l-1), chlorophyll-a (lg l-1),
alkalinity (mg l-1), mean depth (cm) and transparency
(cm). A correlation matrix between the components
was constructed which gave each variable equal
importance and simplified analysis of a large data set
(Ayadi et al., 2002). The relationship of these variables
with the first two axes of variation indicates the most
influential variables. Principal Component Analysis
(PCA) was applied and the environmental data com-
ponents biplot of the first two axes are shown in Fig. 6.
These highlight the close correlation between salinity
and transparency, which is in contrast to alkalinity,
chlorophyll-a, total phosphorus, nitrate and total nitro-
gen along the first axis. Mean depth, salinity and
temperature are more correlated with the second axis.
Variables associated with axis 1 indicate a eutrophica-
tion gradient and the analysis illustrates clearly the
tendency of Lake Manzala towards eutrophication.
Axis 2 is related to hydrologically linked factors
(Fig. 6, left). PCA ordination of the data according to
the 19 reference stations (Fig. 6, right) highlights the
water quality differences between both the sites and the
sampling stations. Sample points are all well discrim-
inated according to each lagoon. In Lake Manzala,
stations are clearly discriminated by being fresher and
richer in nutrients (phosphate and nitrate). The Merja
Zerga sampling stations are differentiated by salinity
and (moderate) nutrient concentrations; similarly Ghar
El Melh stations are characterised by (lower) nutrients
and salinity differences.
Phytoplankton
Phytoplankton abundance in all the three lagoons
displayed significant fluctuations according to season
and to location. Cell numbers decreased in November
and January and then increased in March. The highest
cell concentrations were found during the summer
period when relatively large individuals dominated.
The summer increases were attributed to higher cell
numbers of Pseudonitzschia spp, Licmophora spp,
Navicula spp and Pleurosigma elongatum. These
diatoms reached 24,400 cells l-1 in Ghar El Melh and
35,000 cells l-1 in Merja Zerga. The latter three
species are predominantly benthic in habit, and their
abundance in the plankton samples probably reflects
benthic productivity and turbulent suspension of the
Hydrobiologia (2009) 622:113–131 123
123

shallow water surface sediments. Pseudonitschia spp
have long needle-like cells, and Sournia (1986) noted
that larger phytoplankton species respond more
quickly to favourable conditions and tend to domi-
nate the community at the time of bloom. Diatoms
and dinoflagellates comprised 82.5% and 15.5% of
cell numbers, respectively, and their diversity was the
lowest in winter. The 211 identified phytoplankton
taxa (Supplementary Material Table 3) include spe-
cies previously identified elsewhere in Mediterranean
lagoons and bays by Delgado (1990), Fathi et al.
(2001), Benrejeb-Jenhani & Romdhane (2002), Sakka
Hlaili et al. (2003) and Fraikech et al. (2005).
Overall, phytoplankton cell numbers recorded in
this study for Ghar El Melh and Merja Zerga were
lower than those previously reported by Sakka Hlaili
et al. (2003) for the Gulf of Tunis. In general, large
phytoplankton species were dominant but they did
not reach high concentrations in Merja Zerga. In Ghar
El Melh, the concentrations reached 9 9 105 cells l-1
during July–August when benthic diatoms (Licmo-
phora gracilis and Navicula sp. were common).
However, it is likely that the small size groups, such
as picophytoplankton (\2 lm), have great impor-
tance in these warm water sites (cf. Agawin et al.,
2000). Dinoflagellate abundance diversity was quite
low during the study period but, on a seasonal basis,
overall phytoplankton species diversity was the
highest in spring and summer. Delgado & Fortuna
(1991) also noted that one or a few species were
dominant in phytoplankton blooms in the early winter
period. In contrast to other findings (Delgado, 1990;
Ayadi et al., 2002) from the Mediterranean, a
significant positive relationship was found in the
SMR lagoons between temperature and phytoplankton
abundance (r = 0.72, P \ 0.01).
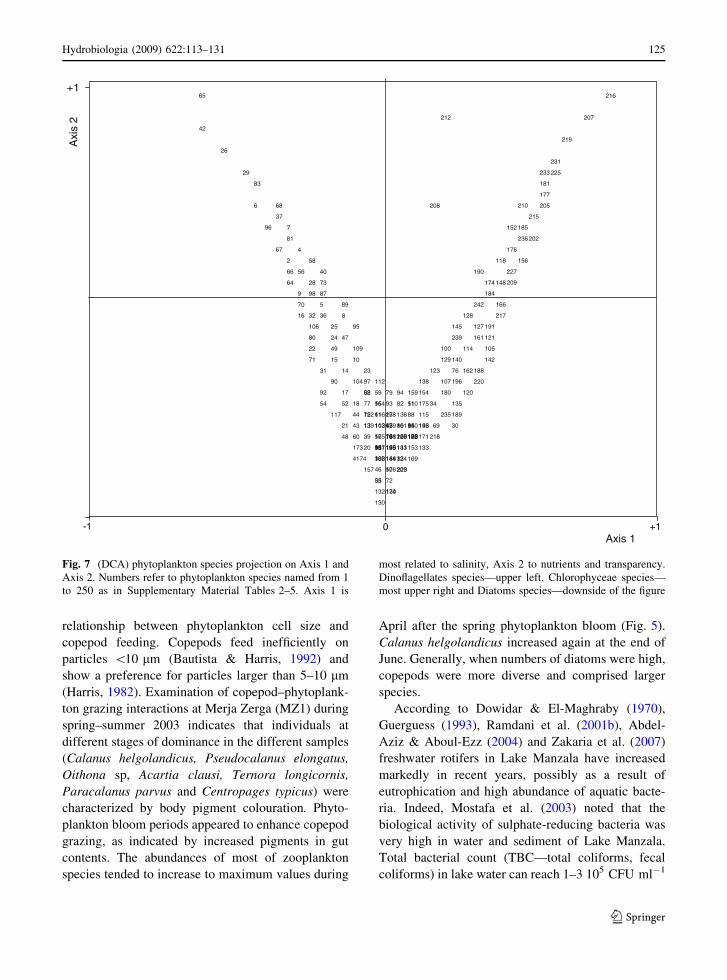
Combined ordinations (DCA) of the phytoplank-
ton data (samples and species) for the three lagoons
(Fig. 7) clearly separated Merja Zerga and Ghar El
Melh where the marine species (dinoflagellates and
diatoms) were often dominated from Lake Manzala
where the freshwater species (Chlorophyceae, fresh-
water diatoms and Cyanophyceae) were abundant.
Zooplankton
Zooplankton can influence the phytoplankton com-
munity through preferential grazing. In turn, nuisance
cyanobacteria (e.g. Microcystis aeroginosa) may
affect copepod community structure by allowing
certain species to out-compete others (Bautista &
Harris, 1992). Many authors have reported a direct
M8
Z4
G4
G3G1
G5
Component 1 C
ompo
nent
2 Z3
Z2
Z1
G2
M9
M7
M6 M5
M4M3
M1
M2
M10
Transparency
pH
Depth
Com
pone
nt 2
Total Nitrogen
Salinity Nitrates
Dissolved Oxygene
Total Phosphorus
Chlorophyll
Temperature
Alkalinity
Suspended matter
Component 1
Fig. 6 Left: PCA components projection on the Axis 1 and
Axis 2. Chlorophyll, Temperature and Total Phosphorus are
most closely related to PCA Axis 1 and salinity and nitrates to
Axis 2. Right: PCA stations projection in the Axis 1 and Axis 2
(Z—Merja Zerga, G—Ghar El Melh, M—Lake Manzala)
124 Hydrobiologia (2009) 622:113–131
123
relationship between phytoplankton cell size and
copepod feeding. Copepods feed inefficiently on
particles \10 lm (Bautista & Harris, 1992) and
show a preference for particles larger than 5–10 lm
(Harris, 1982). Examination of copepod–phytoplank-
ton grazing interactions at Merja Zerga (MZ1) during
spring–summer 2003 indicates that individuals at
different stages of dominance in the different samples
(Calanus helgolandicus, Pseudocalanus elongatus,
Oithona sp, Acartia clausi, Ternora longicornis,
Paracalanus parvus and Centropages typicus) were
characterized by body pigment colouration. Phyto-
plankton bloom periods appeared to enhance copepod
grazing, as indicated by increased pigments in gut
contents. The abundances of most of zooplankton
species tended to increase to maximum values during
April after the spring phytoplankton bloom (Fig. 5).
Calanus helgolandicus increased again at the end of
June. Generally, when numbers of diatoms were high,
copepods were more diverse and comprised larger
species.
According to Dowidar & El-Maghraby (1970),
Guerguess (1993), Ramdani et al. (2001b), Abdel-
Aziz & Aboul-Ezz (2004) and Zakaria et al. (2007)
freshwater rotifers in Lake Manzala have increased
markedly in recent years, possibly as a result of
eutrophication and high abundance of aquatic bacte-
ria. Indeed, Mostafa et al. (2003) noted that the
biological activity of sulphate-reducing bacteria was
very high in water and sediment of Lake Manzala.
Total bacterial count (TBC—total coliforms, fecal
coliforms) in lake water can reach 1–3 105 CFU ml-1
-1
Axi
s 2
+1
Axis 1+1 0
242
239
236
235
233
231
230
229
227
225
220
219
218
217
216
215
212
210
209
208
207
205
203
202
196
195
193
191
190
189
188
187
186
185
184
182
181
180
179
178
177
176
175
174
173
172 171
170
169
168 167
166
165
164
162
161
160
159
158
157
156
155
154
153
152
150
149
148
146
145
144
143
142
141
140
139
138
136
135
134
133
132
131
130
129
128
127
126125
124
123
122
121
120
118
117 116 115
114
113
112
111
110
109
108
107
106
105
104
103
102 101
100
99
98
97
96
95
94
93
92
91
90
89
88
87
86
85
84
83
82
81
80
79
78
77
76
75
74
73
72
71
70
69
68
67
66
65
64
63
62
61
60
59
58
57
56
5554
53
52 51
50
49
48
47
46
45
44
43
42
41
40
39
38
37
36
35
34
33
32
31
30
29
28
27
26
25
24
23
22
21
20
19
18
17
16
15
14
13
10
9
8
7
6
5
4
3
2
1
Fig. 7 (DCA) phytoplankton species projection on Axis 1 and
Axis 2. Numbers refer to phytoplankton species named from 1
to 250 as in Supplementary Material Tables 2–5. Axis 1 is
most related to salinity, Axis 2 to nutrients and transparency.
Dinoflagellates species—upper left, Chlorophyceae species—
most upper right and Diatoms species—downside of the figure
Hydrobiologia (2009) 622:113–131 125
123
in summer (Mostafa et al., 2003 and Sabae, 2006). In
the other two lagoons, Merja Zerga and Ghar El
Melh, the zooplankton are mainly marine species,
and compared to other Meditterean lagoons there was
a slight tendency towards species poor-communities.
Salinity and nutrients probably have the greatest
effect on intra-annual changes in each lagoon.
Combined ordinations (DCA) of the zooplankton
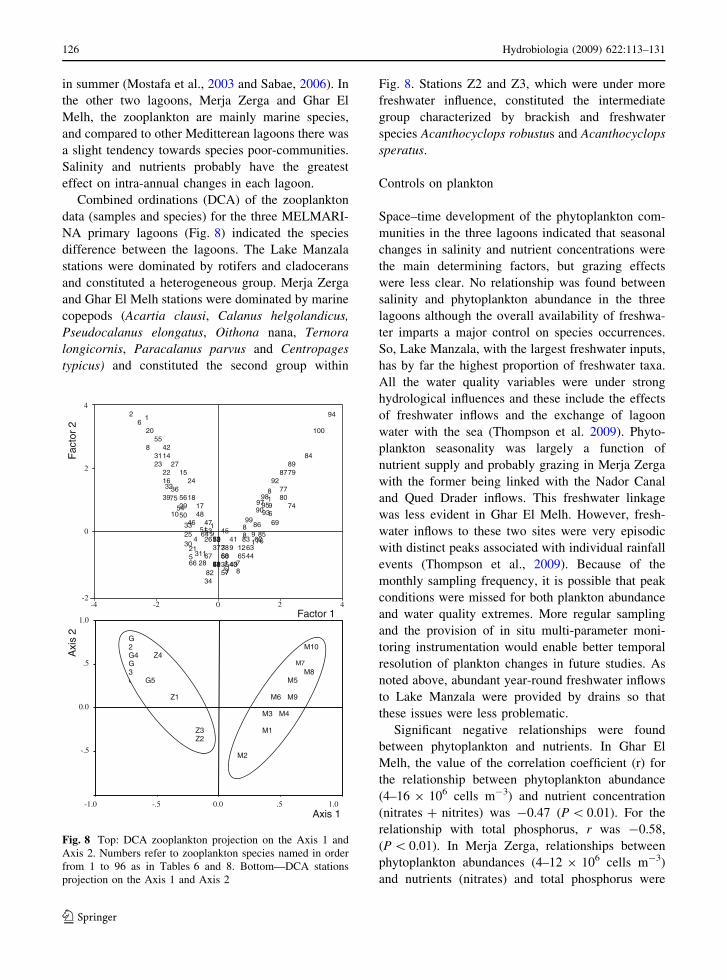
data (samples and species) for the three MELMARI-
NA primary lagoons (Fig. 8) indicated the species
difference between the lagoons. The Lake Manzala
stations were dominated by rotifers and cladocerans
and constituted a heterogeneous group. Merja Zerga
and Ghar El Melh stations were dominated by marine
copepods (Acartia clausi, Calanus helgolandicus,
Pseudocalanus elongatus, Oithona nana, Ternora
longicornis, Paracalanus parvus and Centropages
typicus) and constituted the second group within
Fig. 8. Stations Z2 and Z3, which were under more
freshwater influence, constituted the intermediate
group characterized by brackish and freshwater
species Acanthocyclops robustus and Acanthocyclops
speratus.
Controls on plankton
Space–time development of the phytoplankton com-
munities in the three lagoons indicated that seasonal
changes in salinity and nutrient concentrations were
the main determining factors, but grazing effects
were less clear. No relationship was found between
salinity and phytoplankton abundance in the three
lagoons although the overall availability of freshwa-
ter imparts a major control on species occurrences.
So, Lake Manzala, with the largest freshwater inputs,
has by far the highest proportion of freshwater taxa.
All the water quality variables were under strong
hydrological influences and these include the effects
of freshwater inflows and the exchange of lagoon
water with the sea (Thompson et al. 2009). Phyto-
plankton seasonality was largely a function of
nutrient supply and probably grazing in Merja Zerga
with the former being linked with the Nador Canal
and Qued Drader inflows. This freshwater linkage
was less evident in Ghar El Melh. However, fresh-
water inflows to these two sites were very episodic
with distinct peaks associated with individual rainfall
events (Thompson et al., 2009). Because of the
monthly sampling frequency, it is possible that peak
conditions were missed for both plankton abundance
and water quality extremes. More regular sampling
and the provision of in situ multi-parameter moni-
toring instrumentation would enable better temporal
resolution of plankton changes in future studies. As
noted above, abundant year-round freshwater inflows
to Lake Manzala were provided by drains so that
these issues were less problematic.
Significant negative relationships were found
between phytoplankton and nutrients. In Ghar El
Melh, the value of the correlation coefficient (r) for
the relationship between phytoplankton abundance
(4–16 9 106 cells m-3) and nutrient concentration
(nitrates ? nitrites) was -0.47 (P \ 0.01). For the
relationship with total phosphorus, r was -0.58,
(P \ 0.01). In Merja Zerga, relationships between
phytoplankton abundances (4–12 9 106 cells m-3)
and nutrients (nitrates) and total phosphorus were
Factor 1 420-2-4
4
2
0
-2
100
99
9897 9
695
94
93
92
91
90
89
88
87
8685
84
83
82
81 80
79
78
77
76
7574
73
72 71
69
6867
6665
64
6362
6160
59
58
57
56
55
54
5352
51
50
49
484746
45
4443
42
41
40
39
3837
36
35
34
33
32
31
30
29
28
27
2625
24
2322
21
20
19
1817
1615
14
13
1211
10
9
8
7
6
5
4
3
2 1
Fac
tor
2
G1
1.0Axis 1
.50.0-.5
Axi
s 2
1.0
.5
0.0
-.5
-1.0
M10
M8M7
M6
M5
M4M3
M1
M2
Z3
Z1
Z4
G5
G4G3
G2
Z2
M9
Fig. 8 Top: DCA zooplankton projection on the Axis 1 and
Axis 2. Numbers refer to zooplankton species named in order
from 1 to 96 as in Tables 6 and 8. Bottom—DCA stations
projection on the Axis 1 and Axis 2
126 Hydrobiologia (2009) 622:113–131
123

-0.53 (P \ 0.01) and -0.42, (P \ 0.01), respec-
tively. The corresponding values for Lake Manzala,
where phytoplankton abundances were between 40
and 110 9 107 cells m-3, were -0.67, (P \ 0.01)
and -0.58 (P \ 0.01). These relationships suggest
that phytoplankton development accounts for the
decline in dissolved nutrient concentrations in each
lagoon. Nutrient fluxes in the drainage water were not
measured, but it seems very likely that little nutrient
transport into Merja Zerga and Ghar El Melh
occurred during the dry season. This was in sharp
contrast to the situation in Lake Manzala where
perennial irrigation means that the nutrient-rich
freshwater was available throughout the year and
the phytoplankton was clearly the most eutrophic in
nature. At the other two sites, phytoplankton abun-
dance was generally low with relatively small
seasonal changes. The daily flushing of Merja Zerga
diminishes any significant crop development. Ghar El
Melh normally receives very little freshwater inflow,
so nutrients were not regularly distributed within the
lagoon and, even though the tidal range was small,
additional nutrient input from the Bay of Tunis
(Rasmussen et al., 2009b) (as well as from sediments
and domestic drainage water) is suspected.
Recent changes in plankton communities
Evidence from previous studies indicates that the
plankton, especially the phytoplankton, has changed
significantly in many North African lagoons during
the twentieth Century. During this time, both hydro-
logical modifications and excess nutrients arising
from agricultural development increased markedly in
the region (e.g. Flower, 2001, Ayache et al., 2009).
Sediment records (Flower, 2001; Ramdani et al.,
2001a) indicate a trend towards more freshwater
plankton species within Lake Manzala from the 1960s
onwards. Documentary records show that the density
of zooplankton in Lake Manzala also increased over
the last few decades. In 1987, the zooplankton density
was 100–200 9 103 ind m-3 (El-Sherif & Gharib,
2001). Results from this study indicate densities in the
range 50–300 9 104 ind m-3. This particular ecosys-
tem is highly productive when compared with other
Mediterranean lagoons (Aboul-Ezz, 1995; Daly
Yahya et al., 2001; Mageed, 2006). Khalil (1990)
showed that the phytoplankton communities in Lake
Manzala were dominated by diatoms (68%), followed
by Chlorophyceae (22%) and Cyanophyceae (3%).
Dinoflagellates and Euglinophyceae were rarely and
sporadically present. Previous work indicated that the
diatoms were mainly represented by the genera
Synedra, Nitzschia, Melosira [Aulacoseira] and Cos-
cinodiscus, the Chlorophyceae by Tetraspora,
Scenedesmus and Pediastrum and the Cyanophyceae
by the filamentous Spirulina and Anabaena. El-Sherif
& Gharib (2001) suggested that there has been a
continuous increase in the standing crop of phyto-
plankton in this lake. They identified 129 species that
were either fresh or brackish, whereas 12 species were
purely marine, supporting the suggestion of a move
towards lower salinity conditions. Elsewhere in
Egypt, Zaghloul & Hussein (2000) studied the impact
of the large inputs of pollutants on the phytoplankton
communities in Lake Edku. They suggested that
nutrient pollution created a rich resource spectrum for
algal growth making the lake biologically productive.
In Merja Zerga, a trend to more freshwater conditions
in the south of the lagoons has also been noted
(Ayache et al., 2009). Sediment cores from this
location (Ramdani et al., 2001a; Flower et al., 2001)
showed a decline in marine species and a change from
attached diatoms to those indicative of soft sediments.
Implications
Pollution and salinity changes have already brought
about major shifts in the occurrence of both planktonic
and benthic species in coastal lagoons of the SMR (e.g.
Birks et al., 2001; Reynolds et al., 2002) and without
water quality regulation these changes will become
increasingly detrimental to aquatic resources, espe-
cially to fisheries and biodiversity. The drainage
systems to all the three lagoons have been strongly
modified through construction of barrages, abstraction
and drainage. It is likely that availability of silica, an
essential nutrient for diatom phytoplankton, has
declined at all the three sites (cf. Humborg et al.,
1997). One consequence of this silica depletion
hypothesis is that non-diatom phytoplankton are
favoured, and these algae (especially the dinoflagel-
lates) are often less favourable to lagoon ecosystems.
Toxic phytoplankton species identified in this study
included Gymnodinium catenatum, Alexandrium spp.,
Dinophysis spp. Lingulodinium polyedra, Pseudo-
nitzschia ssp. Gymnodinium aureolum, G. sanguineum,
Prorocentrum micans, P. depressum, Ceratium furca,
Hydrobiologia (2009) 622:113–131 127
123

Gyrodinium spirale, G. impudicum and Scrippsiella
spp. It is thought possible that the presence of these
species is encouraged in the SMR lagoons by changing
nutrient ratios as well as by increasing enrichment and
salinity.
Excess nutrients and discharge of waste or agri-
cultural return water together with land reclamation
are probably the most serious current and common
problems confronting sustainability of the SMR
lagoons. Such changes have caused eutrophication
and loss of ecological capacity in all the three
monitored lagoons. Climate change further threatens
to degrade these lagoon ecosystems as the twenty-first
Century progresses. Lagoon ecosystem surveillance
through regular monitoring is needed not only to better
understand the relationships between phytoplankton
and environmental change, but also for detecting
invasive species. Predictive models to aid water body
management will benefit from the enhanced field data,
especially regarding the plankton. Also, deployment of
automated water quality loggers is a key requirement in
future studies. Similarly, enhanced spatial studies using
remote sensing applications will facilitate monitoring
of phytoplankton distributions (cf. Simis et al., 2005,
Ahmed et al., 2009). Nevertheless, species level
information can only be provided by microscopical
examination, and this remains an essential tool for
monitoring planktonic communities.
Conclusions
Although Merja Zerga, Ghar El Melh and Lake
Manzala displayed major differences in their hydro-
logical regimes which influence salinity and suspended
sediment dynamics, they all were affected to some
extent by excess nutrients. The relatively high inflows
of both freshwater and nutrients into Lake Manzala
produced a rich phytoplankton community dominated
by green algae. At the more saline lagoons, Merja
Zerga and Ghar El Melh, diatoms often predominated
in the phytoplankton during the summer period.
Harmful species in the two latter lagoons were
identified, and these species raise phytosanitation
issues. Future management options would need to
curb the development of harmful blooms by, in the first
instance, reducing nutrient enrichment locally and in
the coastal zone more generally. Zooplankton showed
a high marine influence at Merja Zerga and Ghar El
Melh, but the densities of zooplanktoners tended to be
low in comparison with Lake Manzala and other
Mediterranean lagoons. A major influence by fresh-
water occurs at Lake Manzala with a high diversity of
rotifers, cladocerana and green algae. The historical
records for Lake Manzala indicate that zooplankton
density has increased strongly since the 1970s,
confirming the trend towards eutrophication. The
status of planktonic communities is of crucial impor-
tance for maintaining ecosystem quality. Not only are
the plankton communities a keystone component of
aquatic food webs that support fisheries but they also
function for the dispersal of many delicate larval forms
that are essential for the survival of communities
elsewhere. Careful surveillance of these communities
in relation to water quality and freshwater availability
is required to form environmental management deci-
sions that must be taken in the near future, if these
coastal resources are to be sustained.
Acknowledgements The MELMARINA Project was financed
by the EU Framework V INCO-Med Programme under their
Grant No. ICA3-CT2002-10009. The authors acknowledge the
assistance of all the partner institutions involved in this project.
The generous help provided by the enthusiastic field teams from
each of the countries in North Africa, where the studies were
conducted is also duly acknowledged. Several individuals, in
particular, Dr A. Berraho and Dr. M. Serghini (INRH), Dr S.
Duvail and Ms C. Chambers (UCL), Dr S. Zaghloul (NARSS),
Mr M. Mansour (Merja Zerga), R. M’Rabet (INSTM) also
deserve the authors’ gratitude for their special contributions to
the success of this study.
References
Abdel-Aziz, N. E. & S. M. Aboul-Ezz, 2004. The structure of
zooplankton community in Lake Maryout, Alexandria,
Egypt. Egyptian Journal of Aquatic Research 30(A):
160–170.
Aboul-Ezz, S. M., 1995. Zooplankton of Lake Buroullus.
Bulletin of the National Institute of Oceanography &
Fisheries, Alexandria 21: 233–261.
Aboul-Ezz, S. M. & A. M. Soliman, 2000. Zooplankton com-
munity in Lake Edku. Bulletin of the National Institute of
Oceanography and Fisheries, Alexandria 26: 71–99.
Ahmed, M. H., B. M. El Leithy, J. R. Thompson, R. J. Flower,
M. Ramdani, F. Ayache & S. M. Hassan, 2009. Appli-
cation of remote sensing to site characterisation and
environmental change analysis of North African coastal
lagoons. Hydrobiologia. doi:10.1007/s10750-008-9682-8.
American Public Health Association (APHA), 1995. Standard
methods for the examination of water and wastewater,
128 Hydrobiologia (2009) 622:113–131
123

16th ed. American Public Health Association, Washing-
ton, DC.
Ayache, F., J. R. Thompson, R. J. Flower, A. Boujarra, F.
Rouatbi & H. Makina, 2009. Environmental characteris-
tics, landscape history and pressures on three coastal
lagoons in the Southern Mediterranean Region: Merja
Zerga (Morocco), Ghar El Melh (Tunisia) and Lake
Manzala (Egypt). Hydrobiologia. doi:10.1007/s10750-
008-9676-6.
Ayadi, H., O. Abid, A. Sellami Kamoun, R. Amdouni, A.
Bouain, T. Sime-Ngando & C. Amblard, 2002. Applica-
tion of the Principal Components Analysis to the study of
the phytoplankton and the physicochemical descriptors of
a saltwater marsh. Littoral 2002, the Changing Coast.
EUROCOAST/EUCC, Porto – Portugal.
Bautista, B. & R. P. Harris, 1992. Copepod gut contents,
ingestion rates and grazing impact on phytoplankton in
relation to size structure of zooplankton and phytoplank-
ton during a spring bloom. Marine Ecology Progress
Series 82: 41–50.
Benrejeb-Jenhani, A. M. & S. Romdhane, 2002. Impact des
perturbations anthropiques sur l’evolution du phytop-
lancton de la lagune de Boughrara, (Tunisie). Bulletin
Institut National Sciences et Technplogie de la Mer,
Salammbo 29: 65–75.
Birks, H. H., S. M. Peglar, I. Boomer, R. J. Flower, M. Ram-
dani, P. G. Appleby, A. E. Bjune, S. T. Patrick, M. M.
Kraiem, A. A. Fathi & H. M. A. Abdelzaher, 2001. Pal-
aeolimnological responses of nine North African Lakes to
recent environmental changes and human impacts detec-
ted by macrofossil and pollen analyses. Aquatic Ecology
35: 405–430.
Borja, A., 2005. The European water framework directive: a
challenge for nearshore, coastal and continental shelf
research. Continental Shelf Research 25: 1768–1783.
Daly Yahia, M. N. & M. S. Romdhane, 1994. Contribution a la
connaissance des cycles saisonniers des Copepodes
pelagiques (Mer de Bou Grara). Bulletin de la Societe des
Sciences Naturelles Tunis 10: 1–10.
Delgado, M., 1990. Phytoplankton distribution along the
Spanish coast of the Alboran Sea. Sciecias del Mar Bar-
celona 54: 169–178.
Delgado, M. & J. M. Fortuna, 1991. Atlas de fitoplancton del
Mar Mediterraneo. Sciecias del Mar Barcelona 55.
Doledec, S. & D. Chessel, 1989. Rythmes saisonniers et
composantes stationnelles en milieu aquatique II- Prise en
compte et elimination d’effets dans un tableau faunistique.
Acta Œcologica, Œcologia Generalis 10: 207–232.
Dowidar, N. & A. El-Maghraby, 1970. The neritic zooplankton of
the south eastern Mediterranean at Alexandria. I. Distribution
and ecology of zooplankton organisms with special reference
to Copepoda. Bulletin of the National Institute of Oceanog-
raphy & Fisheries, Alexandria 1: 225–273.
Dussart, B., 1969. Les Copepodes des eaux continentales.
Tome II, Cyclopoıdes et Biologie quantitative. CNRS
Publisher. Editions N. Boubee & Cie, Paris.
El-Hawary, M. A., 1960. The zooplankton of the Egyptian
lakes. A preliminary study on the zooplankton of Lake
Maryut and Lake Edku. Institute of Hydrobiology &
Fisheries, Alexandria. Notes & Memoires 52: 1–12.
El-Naggar, M. E., S. A. Shaaban-Dessouki, M. I. Abdel-Hamid
& E. M. Aly, 1997. Effect of treated sewage on the water
quality and phytoplankton populations of Lake Manzala
(Egypt) with emphasis on biological assessment of water
quality. New Microbiologica 20: 253–276.
El-Sherif, Z. M. & S. Gharib, 2001. Spatial and temporal
patterns of phytoplankton communities in Manzala
Lagoon. Bulletin Institute Oceanography & Fisheries,
Alexandria 27: 217–239.
El-Sherif, Z. M. & S. M. Aboul Ezz, 1988. Preliminary study
on phytoplankton-zooplankton relationship in Lake Bu-
rollus, Egypt. Bulletin Institute Oceanography &
Fisheries, Alexandria 14: 23–30.
Fathi, A. A., H. M. A. Abdelzaher, R. J. Flower, M. Ramdani &
M. M. Kraiem, 2001. Phytoplankton communities in
North African wetland lakes: the CASSARINA Project.
Aquatic Ecology 35: 303–318.
Flower, R. J., 2001. Change, Stress, Sustainability and Aquatic
ecosystem Resilience ln tlorth African wetland lakes
during the 20th century: An introduction to integrated
biodiversity studies within the CASSARINA Project.
Aquatic Ecology 35: 261–280.
Flower, R. J. & J. R. Thompson, 2009. An overview of inte-
grated hydro-ecological studies in the MELMARINA
Project: monitoring and modelling coastal lagoons—
making management tools for aquatic resources in North
Africa. Hydrobiologia. doi:10.1007/s10750-008-9674-8.
Flower, R. J., S. Dobinson, M. Ramdani, M. M. Kraiem, C.
Ben Hamza, A. A. Fathi, H. M. A. Abdelzaher, P. G.
Appleby, H. H. Birks, J. A. Lees, E. Shilland & S. T.
Patrick, 2001. Recent environmental change in North
African wetland lakes: diatom and other stratigraphic
evidence from nine sites in the CASSARINA Project.
Aquatic Ecology 35: 369–388.
Flower, R. J., P. G. Appleby, J. R. Thompson, M. H. Ahmed,
M. Ramdani, L. Chouba, N. Rose, R. Rochester, F. Ay-
ache, M. M. Kraiem, N. Elkhiati, S. El Kafrawy, H. Yang
& E. K. Rasmussen, 2009. Sediment distribution and
accumulation in lagoons of the Southern Mediterranean
region (the MELMARINA Project) with special reference
to environmental change and aquatic ecosystems. Hyd-
robiologia. doi:10.1007/s10750-008-9677-5.
Fraikech, M., A. Berraho, M. Ramdani, A. Chafik, M. Serghini
& A. Moukrim, 2005. Evolution spatio-temporelle des
dinoflagelles dans la baie d’Agadir, Maroc. Marine Life
15: 19–27.
Gamito, S., J. Gilaberyt, C. M. Diego & A. Perez-Ruzafa,
2005. Effects of changing environmental conditions on
lagoon ecology. In Gonenc, I. E. & J. P. Wolfin (eds),
Coastal Lagoons. CRC Press, Boca Raton: 193–230.
Gharib, S. M. & S. M. Soliman, 1998. Some water character-
istics and phyto-zooplankton relationship in Lake Edku
(Egypt) and adjacent sea. Bulletin Faculty of Sciences,
Alexandria 38: 25–44.
Grasshoff, K., 1976. Filtration and Storage. In Grasshoff, K.
(ed.), Methods of Sea Water Analysis. Verlag Chemie,
Weinheim and New York: 21–30.
Guerguess, Sh. K., 1993. Distribution of some rotifers in the
Egyptian Inland waters. Bulletin Institute Oceanography
& Fisheries, Alexandria 19: 249–275.
Hydrobiologia (2009) 622:113–131 129
123

Hallegraeff, G. M., D. M. Anderson & M. B. Cembella (eds),
1995. Manual on harmful marine microalgae. UNESCO,
Paris.
Harris, R. P., 1982. Comparison of the feeding behavior of
Calanus and Pseudocalanus in two experimentally
manipulated enclosed systems. Journal of the Marine
Biology Association, UK 62: 71–91.
Humborg, C., V. Ittekot, A. Cociasu & B. von Bodungen,
1997. Effect of Danube river dam on Black Sea biogeo-
chemistry and ecosystem structure. Nature 386: 385–388.
Hutchinson, G. E., 1967. A Treatise on Limnology and Limno-
plankton. Vol. II-Limnological Botany. Wiley, New York.
Hutchinson, G. E., 1975. A Treatise on Limnology. Vol. III-
Limnological Botany. Wiley, New York.
Khalifa, N. & A. Mageed, 2002. Some ecological aspects on
the zooplankton in Lake Manzala, Egypt. Journal of
Zoology 38: 293–307.
Khalil, M., 1990. Plankton and primary productivity of Lake
Manzala, Egypt. Hydrobiologia 196: 201–207.
Khalil, M. & E. A. Salib, 1986. Effects of some water quality
parameters on fish composition and productivity in Lake
Manzala, Egypt. Proceedings of the Zoology Society of
London 12: 101–109.
Kraıem, M. M., C. Ben Hamza, M. Ramdani, A. A. Fathy,
H. M. A. Abdelzaher & R. J. Flower, 2001. Some
observations on the age and growth of the thin-lipped
Grey Mullet, Liza ramada (Pisces, Mugilidae) in three
North African wetland lakes: Merja Zerga (Morocco),
Garaat Ichkeul (Tunisia) and Edku lake (Egypt). Aquatic
Ecology 35: 335–345.
Kraıem, M. M., L. Chouba, M. Ramdani, M. H. Ahmed,
J. R. Thompson & R. J. Flower, 2009. The fish fauna of three
North African lagoons: specific inventories, ecological
status and production. Hydrobiologia. doi:10.1007/s10750-
008-9679-3.
Mageed, A. A., 2006. Spatio-temporal variations of zoo-
plankton community in the hypersaline lagoon of
Bardawil, North Sinai–Egypt. Egyptian Journal of Aqua-
tic Research 32: 168–183.
Meininger, P. L. & W. C. Mullie, 1981. Egyptian wetlands as
threatened wintering areas for water birds. Sandgrouse 3:
62–77.
Morgan, N. C. & V. Boy, 1980. An ecological survey of
standing water in North West Africa: I: rapid classifica-
tion. Biology Conservation 24: 81–101.
Mostafa, S. A., M. A. Hewedy, H. E. Touliabah, S. M. Ashour
& S. A. Abdallah, 2003. Comparative study among
microflora in El-Manzala Lake water and Rashid (Rosetta)
estuary of Nile River, Egypt. Pakistan Journal of Bio-
logical Sciences 6: 671–679.
Murphy, J. & J. P. Riley, 1962. A modified single solution
method for the determination of phosphate in natural
waters. Analytica Chimca Acta 27: 31–36.
Parsons, T. R., Y. Maita & C. M. Lalli, 1984. A Manual of
Chemical and Biological Methods for Seawater and
Analysis. Pergamon Press, New York.
Paterson, M., 1993. The distribution of micro-crustacea in the lit-
toral zone of a freshwater lake. Hydrobiologia 263: 173–183.
Radwan, S. & B. Popiolek, 1989. Percentage of rotifers in
spring zooplankton in lakes of different trophy. Hydro-
biologia 186(187): 235–238.
Ramdani, M., 1988. Les eaux stagnantes au Maroc: etudes
biotypologique et biogeographique du zooplancton.
Travaux Institut Scientifique Rabat, Serie Zooloie 43:
1–40.
Ramdani, M., N. Elkhiati, R. J. Flower, M. M. Kraiem, A. A.
Fathi, H. H. Birks & S. T. Patrick, 2001a. Open water
zooplankton communities in North African wetland lakes:
the CASSARINA Project. Aquatic Ecology 35: 319–333.
Ramdani, M., R. J. Flower, N. Elkhiati, H. H. Birks, M. M.
Kraiem & A. A. Fathi, 2001b. Zooplankton (Cladocera-
Ostracoda) Chironomidae and other benthic faunal
remains in sediment cores from nine wetland lakes: the
Cassarina Project. Aquatic Ecology 35: 389–403.
Ramdani, M., R. J. Flower, N. Elkhiati, M. M. Kraiem, A. A.
Fathi, H. H. Birks & S. T. Patrick, 2001c. North African
wetland lakes: characterization of nine sites included in
the CASSARINA Project. Aquatic Ecology 35: 281–302.
Rasmussen, E. K., O. S. Petersen, J. R. Thompson, R. J. Flower &
M. H. Ahmed, 2009a. Hydrodynamic-ecological model
analyses of the water quality of Lake Manzala (Nile Delta,
Northern Egypt). Hydrobiologia. doi:10.1007/s10750-008-
9683-7.
Rasmussen, E. K., O. S. Petersen, J. R. Thompson, R. J.
Flower, F. Ayache, M. Kraiem & L. Chouba, 2009b.
Model analyses of the future water quality of the eu-
trophicated Ghar El Melh lagoon (Northern Tunisia).
Hydrobiologia. doi:10.1007/s10750-008-9681-9.
Reynolds, C. S., V. Huszar, C. Kruk, L. Naselli-Flores & S.
Melo, 2002. Towards a functional classification of the
freshwater phytoplankton. Journal of Plankton Research
24: 417–428.
Ricard, M., 1987. Atlas du Phytoplancton Marin, Diatom-
ophycees, Vol. II. CNRS, Paris.
Romdhane, M. S., H. C. Eilertsen, O. Kefi Daly Yahia & M. N.
Daly Yahia, 1998. Toxic Dinoflagellate blooms in Tunisia
lagoons: Causes, and consequences for aquaculture. In
Reguera, B., J. Blance, M. L. Fernandez & T. Wyatt (eds),
Harmful Algae. Xunta de Galicia and Intergovernmental
Oceanographic Commission of UNESCO, Paris: 80–83.
Rougier, C. & T. Lam Hoai, 1997. Biodiversity through two
groups of micro zooplankton in a coastal lagoon (Etang de
Thau, France). Vie & Milieu 47: 387–394.
Saad, M. A. H., S. R. McComas & S. J. Eisenreich, 1985.
Metals and chlorinated hydrocarbons in surficial sedi-
ments of three Nile Delta lakes, Egypt. Water Air Soil
Pollution 24: 27–39.
Sabae, S. Z., 2006. Spatial and temporal variations of sapro-
phytic bacteria, faecal indicators and the nutrient. Cycle
bacteria in Lake Bardawil, Egypt. International Journal of
Agriculture & Biology 8: 178–185.
Sakka Hlaili, A., M. A. Chikhaoui, B. El-Grami & H. Hadj
Mabrouk, 2003. Variation hiverno-estivale de la com-
munaute phytoplanctonique de la lagune de Bizerte en
milieux naturel et fertilise en nutriments. Revue Faculty
Science Bizerte 2: 37–49.
Samaan, A. A., 1977. Distribution of zooplankton in Lake
Edku. Bulletin Institute Oceanography & Fisheries, Cairo
6: 157–196.
Samaan, A. A. & R. R. Aleem, 1972. The Ecology of zoo-
plankton in Lake Maryut. Bulletin Institute Oceanography
& Fisheries, Cairo 4: 234–273.
130 Hydrobiologia (2009) 622:113–131
123

Scheffer, M., 1998. Ecology of Shallow Lakes. Chapman &
Hall publishers, London.
Scheffer, M., S. H. Hosper, M. L. Meijer, B. Moss & E. Je-
ppesen, 1993. Alternative equilibria in shallow lakes.
Trends in Ecology & Evolution 8: 275–279.
Shaheen, A. H. & S. F. Yosef, 1987. The effect of the cessation
of the Nile flood on the hydrographic features of Lake
Manzala, Egypt. Archive fur Hydrobiologie 84: 339–367.
Simis, S. G. H., S. W. M. Peters & H. J. Gons, 2005. Remote
sensing of the cyanobacterial pigment phycocyanin in
turbid inland water. Limnology and Oceanography 50:
237–245.
Souissi, S., O. Daly. Yahia-Kefi & M. N. Daly. Yahia, 2000.
Spatial characterisation of nutrient dynamics in the Bay of
Tunis (south-western Mediterranean) using multivariate
analyses: consequences for phyto- and zooplankton dis-
tribution. Journal Plankton Research 22: 2039–2059.
Sournia, A., 1986. Atlas du Phytoplancton Marin. I: Cya-
nophycees, Dictyochophycees, Dinophycees,
Raphidophycees. CNRS Editions, Paris.
Stanley, D. J. & A. G. Warne, 1994. Nile Delta: recent geo-
logical evolution and human impact. Science 260:
628–634.
Talling, J. F., 1976. Phytoplankton: composition, development
and productivity. In Rzoska, J. (ed.), The Nile, Biology of
an Ancient River. Monographiae Biologia 29. Junk, The
Hague: 385–402.
Talling, J. F. & J. Lemoalle, 1998. Ecological dynamics of
tropical inland waters. Cambridge University Press,
Cambridge.
Thompson, J. R., R. J. Flower, M. Ramdani, F. Ayache, M. H.
Ahmed, E. K. Rasmussen & O. S. Petersen, 2009.
Hydrological characteristics of three North African
coastal lagoons: insights from the MELMARINA project.
Hydrobiologia. doi:10.1007/s10750-008-9680-x.
Tomas, C. R., 1997. Identifying Marine Phytoplankton. Aca-
demic Press, New York.
Tregouboff, G. & M. Rose, 1957. Manuel de planctonologie
Mediterraneenne. CNRS, Paris.
Zaghloul, F. & N. Hussein, 2000. Impact of pollution on
phytoplankton communities’ structure in Lake Edku,
Egypt. Bulletin Institute Oceanography & Fisheries,
Alexandria 26: 297–318.
Zakaria, H. Y., M. H. Ahmed & R. J. Flower, 2007. Envi-
ronmental assessment of spatial distribution of
zooplankton community in Lake Manzalah, Egypt. Acta
Adriatica 48: 161–172.
Hydrobiologia (2009) 622:113–131 131
123





























