Embed Size (px)
Citation preview

Planktonic Communities in Florida Softwater Lakes of Varying pH
Patrick L. ~rezonik, ' Thomas L. Crisman, and Randy L. Schulze Department of Environmental Erlgineering Sciences, University of Florida, Cainesville, FL. 326 1 1, USA
Brenonik, P. L., -6. L. Crisman, and R. L. Schulze. 1984. Planktonic communities in Florida softwater lakes of varying pH. Can. 8. Fish. Aquat. Sci. 41 : 46-56.
In 20 softwater Florida lakes (pH range4.7-6.8), phytoplankton diversity was significantly higher in the less acidic lakes; mean numbers of species in two groups (1, pH < 5.3; 2, p H > 5.6), each consisting of 10 lakes, was 10.8 and 16.5, respectiveiy. Blue-green algae were dominant in less acidic lakes, while green algae were the most common group in acidic lakes. Phytoplankton abundance also was lower in acidic lakes, but covariation of phosphorus concentrations with pH confounded interpretation. Trends in zooplankton communities along the p H gradient were less pronounced. More acidic lakes had more species than the less acidic lakes, but three measures of species diversity showed no significant differences between the two groups. Zooplankton abundance was lower in the acidic lakes, but the relationship between abundance and pH showed much scatter. Lake productivity probably affects~ooplankton abundance more than p H does in the sample group. No clear trends were observed in relative abundance of major zooplankton groups across the pH gradient. Six acid-tolerant species (including Diaptomus d/orbdanus, Eubosrnina terbicen, and Daphnia ambigera) were dominant in all 20 lakes.
Dans 20 lacs d'eau douce (ou le pH varie de 4,7 a 63 ) de la Floride, la diversite du phytoplancton etait sensiblement plus elevee dans les lacs les moins acides. Le nombre moyen d'especes reparties en deux groupes (4, pH 5,3; 2, p H 5,6), chacun comprenant '80 lacs, s'elevait respectivement a 10,8 et a 46,5. Les algues bleues-vertes dominaient dans les lacs moins acides tandis que les algues vertes etaient les plus communes dans les lacs acides. L'abondance du phytoplancton etait en outre rnoins elevee dans ces derniers mais la covariation des concentrations de phosphore avec le pH a brouille I'interpretation. Les tendances des communautes de zooplancton par rapport au gradient du pH etaient moins prononcees. Les lacs plus acides contenaient davantage d'especes que les moins acides, rnais trois quantifications de la diversite des especes n'ont pas rev6le de differences significatives entre ies deux groupes. La quantite de zooplancton etait moins elevee dans les lacs acides mais la relation entre I'abondance et le p H a revele une grande dispersion. La productivite Bacustre influe probablement plus sur I'abondance du zooplancton que le p H dans les lacs echantilionnes. Aucune tendance precise n'a ete observee pour ce qkei est de l'abon- dance relative des principaux groupes de nooplancton par rapport au gradient du pH. Six esp&ces tolerantes par rapport aux conditions acides (y compris Diaptsrners floridanus, Eerbosrnina tubicen et Daphnia ambigua) etaient en nombre dominant dans les 26 lacs.
Received Februay 4, 1983 Accepted September 12, 1983
A cid precipitation is a well-documented phenomenon in northeastern North America, and numerous studies have been undertaken to evaluate its effects on aquatic ecosystems of that region (see Haines 1981; Cowling
1982). The effects of increased acidity on the composition of planktonic and fish communities are fairly well understood for temperate lakes, but many questions remain concerning the effects of acidification on biological processes such as primary production and mineral cycling.
Acid rain is not limited to northeastern North America. In the United States, the entire area east of the Mississippi River receives acid loadings from the atmosphere (e.g. see Likens et al. 1979; Cowling 19821, and other areas also are affected. Extensive studies in Florida (Brezonik et al. 1980, 1983) have shown that the northern two-thirds of the state receives bulk precipitation with an annual average pH < 4.7; data from
l~hhuthor to whom correspondence should be addressed. Present address: Department of Civil and Mincral Engineering, University of Minnesota, Minneapolis. M N 55455, U S A .
Recu le 4 fevrier 1983 Accepte be 12 septembre 1983
wet/dry collectors indicate that rainfall pH throughout the state is 0.2-0.6 units lower than bulk precipitation pH. Excess (nonmarine) sulfate loadings in northern Florida amount to 7- 11 kg. ha-' yr- ', or roughly 50-88% of the loadings in the northeastern United States.
Acid precipitation is of particular concern in northern and central Florida because highly sandy soils are common through- out the region's interior uplands. The soils have low cation exchange capacities and low base saturation ratios. Similarly, many small lakes in upland sandhill areas have very soft waters and correspondingly low alkalinities. These lakes are sensitive to acidification from atmospheric loadings, as recent maps have illustrated (8memik and Powers 1982). The effects of pH on community structure and ecosystem processes in warm temper- ate and subtropical lakes such as those in Florida are not well understood. No previous studies have evaluated trends in planktonic abundance and composition of Florida lakes over the pH range associated with acid precipitation.
As part of a broad effort to evaluate the significance of acid precipitation in Florida (Brezonik et al. 19831, we undertook a
46 Can. J . Fish. Aqlmt. Sci., Vo1. 41, 1984
Can
. J. F
ish.
Aqu
at. S
ci. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y 19
9.20
1.12
1.12
on
06/0
5/13
For
pers
onal
use
onl
y.

cross-sectional analysis of chemical and biological conditions dong a pH gradient in softwater sandhill lakes. Description of variations in chemical conditions and nutrient status relative to pH and analysis of the sensitivity of the lakes to acidification is presented elsewhere (Hendry 1983; Hendry and Brezonik 1984). This paper describes the abundance and species composition of phytoplankton and zooplankton communities in the lakes in relation to pH.
Two groups of softwater lakes were selected for the study: a group of 13 lakes in the Trail Ridge physiographic region of northcentral Florida (Alachua, Clay, and Putnam counties, 29"401N, 82"001W) and a group of 7 lakes in the Highlands Ridge physiographic region in southcentral Florida (Highlands County, 27'21 'N, 8l028'W) (see Fig. 6-1 in Brezonik et al. 1983). The lakes range in pH from about 4.7 to 6.8, but they are otherwise similar in water chemistry. None of the lakes is eutrophic. Selection was based primarily on two criteria: soft water, making them sensitive to atmospheric loadings of acidity, and availability of data to determine whether pH andlor alkalinity had changed over the past 18-28 yr. Detailed descrip- tions of the two regions and each lake are given by Brezonik et al. (1983).
The 20 lakes were sampled quarterly beginning in April and May, 1978, and ending in January and February, 1979. A midlake sampling station was selected in each lake at either the deepest part of the lake or at the center of the largest basin, if the lake consisted of several basins. The same station was used on all four sampling dates.
Sampling and analytical methods for chemical parameters are described by Brezonik et al. (1 983). Phytoplankton samples were collected with a 2.2-L Kemmerer bottle at 0.5 m in lakes less than 5 m deep and at the top. middle, and bottom of the water column of lakes greater than 5 rn in depth. Equal volume aliquots were combined in $8-mL bottles and preserved with 2 mL of acidified Lugo19 s solution. In the laboratory, aliquots were settled in an Utemohl chamber and examined with a Unitron inverted microscope. Taxonomic identifications were based on keys in Whitford and Schumacher (1973), Cocke (1966), Weber (197 I), Prescott (1954), and Smith (1950). All counts represent at least 200 individuals.
Zooplankton samples were obtained by vertical tow with a No. 20 Wisconsin plankton net (80-km mesh) through the entire water column and were preserved in 10% buffered fomalin. At least 100 individuals were counted for each sample under a dissecting microscope at 2 5 0 ~ . Zooplankton biomass (milli- grams C per cubic metre) was obtained by multiplying species counts by biomass conversion factors (Dumont et al. 1975). Taxonomic keys in Edmondson (1959) and Pennak (1953) were used to confirm identifications,
Descriptive statistics, correlations, and analyses of variance were calculated by the Statistical Analysis System (SAS) computer package (Barr et al. 1976). Three diversity indices (Simpson's, Shannon- Weaver, and evenness ( E = PillH~,,)) were calculated and cluster analyses were performed with a computer package of Bloom et al. (1977). Cluster analyses were done on log-transformed data (X' = log ( X + 1 ), where X is the abundance of a species (number per litre) in the sample). Czekanowski's index (Connack 197 1) was used as the measure of similarity between zooplankton communities in the 20 lakes.
One-way analysis of variance (ANOVA) and Duncan's multiple range test of pH data on the 20 lakes showed that they could be grouped into two nonoverlapping categories (statisti- cally different at P < 0.05): an acidic group (pH < 5.3) and a nonacidic group (pH > 5.6). The two descriptors are intended to be relative terms md are used here for ease of identification. Statistically significant differences were found between mean values of many chemical parameters for the two groups (Hendry 1983). The groupings based on pH correspond fairly well to geographical groups (see Table l ) , except that the three largest lakes in the Trail Ridge region (Mingsley, Geneva, and Altho) were grouped with the less acidic Highlands Ridge group. The higher pH values of the large Trail Ridge lakes probably reflect differences in shoreline cultural development and hydrologic conditions compared with the other Trail Ridge lakes.
The ensuing analysis compares planktonic population and species composition data on the two groups of lakes. To examine trends along a pH gradient in more detail, the lakes also were grouped into five categories based on divisions of 0.5 pH units as follows: pH 4.50-5.00 (four), pH 5.01-5.50 (six), pH 5.51-6.00 (two), pH 6.01-6.50 (four), and pH 6.51-7.00 (four); the numbers of lakes in each group are given in parentheses. Mean values for various biological parameters were averaged for the lakes in each category to smooth out variability among the individual lakes and clarify trends with pH*
Results and Discussion
Phytoplankton
Species richness and abundance Mean numbers of phytoplankton species in the acidic and
nonacidic lakes were 10.8 (range 8-13) and 16.5 (range 11-22), respectively. The means are significantly different (t-test, P < 0.01). The mean number of phytoplankton species for each lake was significantly correlated with pH (P < 0.05) (Fig. lA), but considerable scatter exists in the relationship, especially for lakes with pH > 6.0. Three nonacidic lakes (Annie, Geneva, and Mingsley) had numbers of species similar to those of the acidic lakes; this trend may be explained by lower levels of total phosphorus (TP) in these lakes compared with the other nonacidic lakes.
The decrease in species numbers with decreasing pH is seen more clearly when lakes are grouped into pH intervals (Fig. 1C). One-way ANOVA and Duncan's test showed that the means of the two lowest intervals (4.5-5.5) were significantly lower than the means for the two highest intervals (6.0-7.0). Although the number of algal taxa often is greater in temperate lakes than in comparable Florida lakes, our results agree generally with those of synoptic surveys in Scandinavia (Almer et al. 1974; Leivestad et al. 1976) and Canada (Kwiatkowski and Woff 1976; Yan and Stokes 1978). These studies all show that species richness and diversity decrease with decreasing pH, and the largest changes occur in lakes with pH < 5.6.
Mean numbers of phytoplankton individuals for the 20 lakes displayed patterns similar to those for species richness. Mean abundance in the 18 acidic lakes was 5700cells/mL (range 2900-10 900/mL), while the mean for the 10 nonacidic lakes was 14 880 cells/mL (range 4700-25 60WmL). The means are different at P < 0.05 (t-test). Phytoplankton abundance was positively correlated with pH (P < 0.05), but again the trend shows much scatter (Fig. 1B). A plot of mean abundance in the
Can. .I. Fish. Aquae. Sci., Vol. 41, 1983
Can
. J. F
ish.
Aqu
at. S
ci. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y 19
9.20
1.12
1.12
on
06/0
5/13
For
pers
onal
use
onl
y.

TABLE 1. Summary of phytoplankton and zooplankton statistics over four quarters for 20 Florida lakes (Nos. 1 - 10, acidic; Nos. 11-20, nonacidic), Lake regions: T, Trail Ridge; H, Highlands Ridge.
- - - - - - - - - - - -- -- - - - -
Phytoplankton Zooplankton
Mean Mean Lake Mean No. abundance Mean No. abundance Biomass
No, region Name pH of Species (No. /mL) of Species (NO. /m3) (mg/m")
McCloud Cowpen Anderson-Cue Galilee Brooklyn Rosa Magnolia Sheeler Lowery Johnson
Annie Altho Josephine Geneva Letta Placid June Clay Kingsley Francis
4.5-5.8 5.01-5.5 5.51-6.0 6.01-6.5 6.51-9.0
pH Intervals
4 6 2 4 4
Number of Lakes
I . 1 (A) Mean annual species richness and (B) abundance of phytoplankton vs pH. (C) Mean values of the same parameters over 5 pH intervals. Bars denote ranges in each interval.
Can. J . Fish. A q u a . Sci., Vol. 41 , 1984
Can
. J. F
ish.
Aqu
at. S
ci. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y 19
9.20
1.12
1.12
on
06/0
5/13
For
pers
onal
use
onl
y.

five pH intervals illustrates the trend m r e clearly (Fig. 16). One-way ANOVA and Duncan's test showed that the means of the two lowest pH intervals were significantly lower than the means of the two highest intervals.
Scatter in the correlation between phytoplankton abundance and pH is at least partly explained by the wide range of TB concentrations found in the lakes (Hendry 1983). TP and pH both affect phytoplankton abundance (Fig. 29. A trend of increasing abundance with increasing TP levels is apparent at all pH levels but is most noticeable in the less acidic lakes. The paucity of acidic lakes with high TP levels precludes definition of the complete response surface. Phytoplankton abundance d s o increased with increasing pH for constant levels of TP in the data set.
Although several synoptic studies have reported trends of lower planktonic abundance with decreasing pH (e.g . Alnler et al. 1974; Kwiatkowski and roff 1976), more recent reports indicate that acidification of temperate lakes is not necessarily accompanied by a significant reduction in phytoplankton biomass (Almer et al. 1978; National Research Council of Canada 1981). Several investigators (Hendrey et al. 1976; Yan and Stokes 1978) have suggested that phytoplankton biomass is controlled more by phosphorus concentrations than by pH. This hypothesis is supported by results of enclosure and whole-lake experiments in which acidic water has been enriched with phosphorus (Dillon et al. 1979; Fee 1980; Wilcox and BeCosta 1982; DeCosta et al. 1983).
Consistently low TP Bevels in all acidic lakes except Anderson-Cue (TP = 40 pg/L) suggest that a causal relation- ship may exist between TP and pH. Almer et al. (1978) suggested that nutrient cycling is diminished in acidic lakes, but other explanations are possible for the trend between TP and pH (see Hendry 1983). Acidic lakes may have lower TP input rates from both cultural and natural sources. For example, acidic lakes may have generally lower watershed to lake surface area ratios than those of nonacid lakes. Evaluation of the hypothesis that acidic lakes have lower nutrient loading rates would require accurate nutrient budget data, but such information is not available for most of the lakes. Because s f the lakes' seepage character, such budgets would be difficult to determine accu- rately. Consequently, causes of the trends in TP and phytoplank- ton abundance with pH in Florida Eakes remain undetermined.
specie^ corngositisst The most obvious trend in the composition of phytoplankton
communities in the present investigation was a replacement of blue-green algae as the dominant group at high pH by green algae as the dominant group below pH 5.5-6.0 (Fig. 3). With the exception of an increase in pyrrhophytes (Peridinium) in the most acidic lakes (pH 4.5-5.0), the subdominant algal groups displayed little trend with pH. Diatoms were rare in all Bakes, and no apparent trend in silica levels was evident along the pH gradient (Hendry 1983).
A decrease in the number of rare species was associated with decreasing pH (Table 2). Highly acidic lakes were dominated by Staurastrum sp., Scenedesmus sp . , Avtkistrotiesmus fascatus, Peridinium inconspicuum, and several species of small coccoid green algae. Euglena sp., Oscystis spp., and Cla/arnydomonas sp. also were fairly common (but not dominant) in acidic lakes. Blue-green algae were sparse in acidic lakes and were represen- ted mainly by Bscillatoria kirnnetica and Anacys~is incerta The less acidic lakes contained more rare species and a greater variety of blue-green species, such as AmigdaHum quatlraden-
FIG. 2. Relationships among total phosphorus, pH, and phyto- plankton abundance for 219 Florida lakes.
Chlorophyta a Cyonophyta 8 All others
FIG. 3. Relative abundance s f thee major groups of phytoplankton for the 20 lakes grouped in 5 pH intervals.
tala, Anabaena spp., Spirulina kaxissima, and Microcjstis aerug inssa .
Oligotrophic nonacidic lakes in Canada typically are domi- nated by either chrysophytes (Duthie and Socha 8976; Findlay and Mling 1979) or diatoms (Yan et al. 1973, but dominance in acidic l&es shifts to pyrrhophytes (Yaw and Stokes 1978; Yan 1979). Similar trends have been reported in Sweden (Almer et al. 1974); according to Haines ( 198 1) the same species often are dominant in acidic lakes of both countries.
Although similar taxa characterize acidic lakes in both Florida and the temperate zone, and the increased importance of Peridinkurn inconspicuum under acidic conditions is a common feature of both regions, significant differences in phytoplankton community structure are apparent. In Florida we observed a dominance of chlorophytes in acidic lakes, while pyrrhophytes, ckrysophytes, and diatoms generally were of low importance across the pH range of oligotrophic lakes. Reasons for the differences in subtropical Florida Bakes are not certain but may
Can. J . Fish. Aquat. Sci., Vol. 41, 1984
Can
. J. F
ish.
Aqu
at. S
ci. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y 19
9.20
1.12
1.12
on
06/0
5/13
For
pers
onal
use
onl
y.

TABLE 2. Phytoplankton distribution versus pH ( x indicates species found in lake at least once during the four quarters).
Taxona Phytoplankton 4.5-5.0 5.01-5.5 5.51-6.0 6.01-6.5 6.51-6.7
Anacystis incerta x Ankistrodesmus falcatus x Chlamydornonas sp. x Chroomonas sp. x Closterium sp. x Bin~bryon sp. x Euglena sp. x Gloeotheca linearis x Bocystis pusilla x Bscillatoria limnetica x Pelanate diatom x Peridinium incorzspicuum X
Scenedesmsts x Selenastrum minuturn x Small gr. coccoids x Staurastrum spp. x Amigdalum quadridentata Anacystis montana Centric diatom Chlorella sp. Closteriopsis sp . Cosmarium spp. Crkscigenia tetrapodia Bac~lococcopis rhaphidioides Gomphosphaeria lacustris Kkrchneriella contorta Microcystis aeruginssa Phaczls sp. Pseudotetraspora sp. Rhabdoderma sp. Synedra sp. Tetradon minimum Anabaena sp. Aphanothece nidulans Asterionella formosa Chrossoccus limneticus Gottyostomum semen Melosira sp. Rhizosolenia sp. Anaqstis sp. Chlorogonium elongoturn Echinosphaerilla sp. Elakatothrix gelatinosa Gloeocapsa rupestris Mallomonas sp. Bscillotoria angustissima Schiaothrix sulcicola Sraurodesmus dijectus
"B, Bacillaiophyta; C, Chloromonadophyta; Ch, Chlorophyta; Cr, Cryptophyta; Cs, Chrysophyta; Cy , Cyanophyta; E, Euglenophyta; P, Pyrrhophyta.
be related to the fact that Florida zooplarnkton communities are not as seriously changed by pH as those in temperate lakes. Unfortunately, our synoptic data are not sufficient to determine whether the role of zooplankton as grazers and nutrient cyclers is significantly altered by acidification.
Zooplankton
Species richness and abundance As was the case for phytoplankton populations, the diversity
and abundance of zooplankton were lower in more acidic Bakes, but trends were smaller and less consistent. The mean number of zooplankton species found in the 10 nonacidic lakes was 19, while the 18 acidic lakes had an average of only 14 species; the means are significantly different (t-test, B < 0.08 1). The mean number of species in each Bake also was correlated positively with pH ( B < 8.801). Species richness over the five pH intervals (Fig. 4) increased from the second-lowest interval (5.8-5 .§) to the highest interval (6.5-7.8), but ANOVA showed no signifi-
50 Can. J . Fish. Ayuat. St-i., Vol. 41, 1984
Can
. J. F
ish.
Aqu
at. S
ci. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y 19
9.20
1.12
1.12
on
06/0
5/13
For
pers
onal
use
onl
y.

a Number of Species Abundance
4 . 5 - 5 . 0 5.01-5.5 5.51-6.0 6.01-6.5 6.51-7.0
pH Intervals
FIG. 4. Mean species richness and abundance of zooplankton in 5 pH intervals. Bars denote ranges in each interval.
15 i Similarity Cutoff
2 I 8
1 1 I I I I
90 €30 ao I
IQQ
Level sf Sirn i l~s i ty 4.5- 5.8 5.01-5.5 5.51-6.Q 6.81-6.5 6.51-7.0 FIG. 6 . Classification of 20 Florida lakes by Czekanowski's simi-
pH Interval 1arity index and mean annual zooplankton data (see Table 1 for key to
I . 5 . Relative abundance of major zooplankton groups in 5 pH lakes). Vertical line shows similarity level where lakes form two
intervals. chsters. An asterisk indicates a nonacidic lake.
cant differences among the five groups. The range of species richness was relatively small over the entire pH spectrum, suggesting that pH does not have an overwhelming effect on zooplankton diversity in Florida lakes.
Several investigators have reported a progressive decline in numbers of species with decreasing pH in temperate lakes (e.g. Sprules 1995; Waddum et al. 1 980; Confer et al. 1983), but the trend is less obvious in Florida lakes. In addition, the number of species found in the present study is much higher than numbers reported in temperate lakes of comparable pH. In a study of acidic lakes near Sudbury , Ontario, Yan and Strus ( 1980) found about half as many crustacean species in lakes with pH values similar to the acidic Florida lakes. Leivestad et al. (1976) reported numbers of zooplankton species in acidic Norwegian lakes that are a third to a half of those observed in this study.
Reasons for the higher diversity in Florida lakes are uncer- tain. Concentrations of heavy metals may be lower in Florida lakes, which are in highly weathered sandy soils compared with Scandinavian and temperate North American lakes, which are in granitic terrain. Lower concentrations of toxic metals may pennit higher species diversity in acidic Florida lakes, but
further studies are necessary to evaluate this hypothesis. In addition, lakes in Florida are not subjected to large seasonal fluctuations that occur in temperate lakes. The temperature range in the lakes during the year of sample collection was about 12-30°C. All the lakes are oligotrophic or mesotrophic, and none exhibited anoxic conditions in the water column. The adaptive potential of organisms to increased acidity may be enhanced by the otherwise moderate environment. Finally, lakes in the study areas probably have been moderately acidic (for natural reasons) for many years. Historical data (Hendry and Brezsnik 1984) show that some Trail Ridge lakes had pH values of 5.0-5.5 more than 20 yt ago. Because the region's soil is naturally acidic and low in buffering ability, natural pH levels in many of the lakes probably were in or near this range. The high diversity of the populations thus may reflect natural selec- tion for acid-tolerant species.
The mean abundance of zooplankton in the 10 acidic Florida lakes was 7.5 x lo4 individuals/m3 (range 3.0 X lo4 to 13.8 x lo4), while the corresponding value for the nonacidic lakes was 14.5 X 104(range3.8 x 104t032.6 X 104).Themeanvaluefor the acidic lakes is significantly lower than the mean of the nonacid group (t-test, P < 0.05). Zooplankton abundance over
Can. J . Fish. Aquat. Sci., kTol. 41, 1984 5 1
Can
. J. F
ish.
Aqu
at. S
ci. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y 19
9.20
1.12
1.12
on
06/0
5/13
For
pers
onal
use
onl
y.

TABLE 3. Composite zoksplankt~n species list from 20 Florida lakes (see Table 1 for lake names and pH). X indicates species found in lake at least once during the four quarters.
- - - -
Acidic lakes (pH < 5.3) Nonacidic lakes (pH > 5.6)
1 2 3 4 5 6 7 8 9 10 1 1 12 13 14 1 % 16 17 18 19 20
C~pepsda Cyciops varicuns x x x x x x x x x x X X X X X X X X ~ X
Diaptomus floridanus x x x x x x x x x x x x x x x x x x x x Ergasilus sp. x x x x x x x x x x x x MesocycCops edax x x x x x x x x x x x x x x x x x x x x Trqocyclops pmsinaks x x x x x x x x x x x x x x x x x Copepdite x x x x x x x x x x x x x x x x x x x x Nauplii x x x x x x x x x x x x x x x x x x x x
Cladocera Eubosmina tubicen Dsrphpaia ambigua Leptsdora kkndfii Chydorus sphaericus Immature cladxeran Simocephalus sp. Holspedium arnaaonkcuan Bosmiraopsis dieters; Ceriodaphnia reticulafa Diaphanossmn br~chyttrurn
X X X X X X X X X X X X X X X X X X X X
X X X X X X X X X X X X X X X X X X X X
X X X X X X X X X X X X X X X X X X X X X X X X
X X X X X X X X X X X X X X
X X
X X
X
X
Rotifera Keratelta cochiearis x x x x x x x x x x x x x x x x x x x x Trichocerca multicrinus X x x x x x x x x x x x x x x x Pslyarrhra vulgaris x x x x x x x x x x x x x x x x x x Trichocerca iotzgicefu x x x X X, x x x x x x x x x C~nochilus unicornis X X X X x x x X x x x x x X
Kerateltw taurocephuln x x x x x x x x x x x x x Mon~s@la sp. x x x x Kelbocottia sp. x x x x x x x x x x x x x x Brachionus quadridentata x x x x x Monoma fus sp. x Kerwtella quadrata x x Filinia sg. x x X X X X X X x x Brachionmts havanaensis x x x x X X
Brachionus calyc$orus x x Pdeosepmks sp. x x A.~planchna sp. x x x x x x x x x Gastropus sp. x Brachionus angularis x x x x Platyins patutus x Conschiloides sp. x Eecarae sp. x
the five pH intervals displayed a somewhat inconsistent pattern (Fig. 4). The lowest population occurred in the range pH 5.0-5.5, and the highest occurred in the range pH 6.0-6.5. ANOVA and Duncan's test showed significant differences in means between pH intervals; the mean for pH 6.0-6.5 is significantly higher than the means for the two intervals from pH 4.5 to 5.5 (P < 0.05). The annual average (dry weight) biomass in the 10 acidic lakes was 93 mg/rn3 (range 24-175), and the average biomass in the 10 nonacidic lakes was 123rng/m3 (range 49-268). Biomass trends were similar to those observed for abundance, but because of large within-group variances, differences in means were not statistically significant.
Beaver and Crisman (1981) reported biomass values for ciliated protozoa in 10 of the survey lakes; their samples were collected concurrently with ours. The biomass of this often ignored zooplankton group displayed much scatter between lakes, but values generally increased with increasing pH.
Although comparison with total zooplankton is limited to the acidic lake group, results indicate that ciliates are less tolerant of Bow pH than are other zooplankton groups. Ciliates contributed 2-7% of the total zooplankton biomass in the most acidic lakes (pH 4.7-4.9) and 20-30% in lakes near pH 5 -2.
Zooplankton abundance and biomass generally decrease with decreasing pH in temperate lakes (Yan 1979; Roff and Kwiat- kowski 1977; Confer et al. 1983). The latter authors found a significant correlation with pH, but Yan and Strus (1980) did not. Although pH affects the abundance of zooplankton in Florida lakes to some extent, the great variability in the data (Fig. 4) suggests that additional factors are important. For example, mean zooplankton biomass in the 20 lakes displayed a stronger correlation with chlorophyll (r = 0 5 3 ; P < 8.0 1) than with pH ( r = 0.25; P < 0.01). Gsnelations using zooplankton abundance strengthened both relationships somewhat ( r = 0.62 and 0.44, respectively; P < 0-0 1 ) but the trend remains. Since
52 Can. /. Fish. Aquur. Sci., k'ol. $ 1 , 1984
Can
. J. F
ish.
Aqu
at. S
ci. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y 19
9.20
1.12
1.12
on
06/0
5/13
For
pers
onal
use
onl
y.
Similarity Cutoffs
Level of Similarity
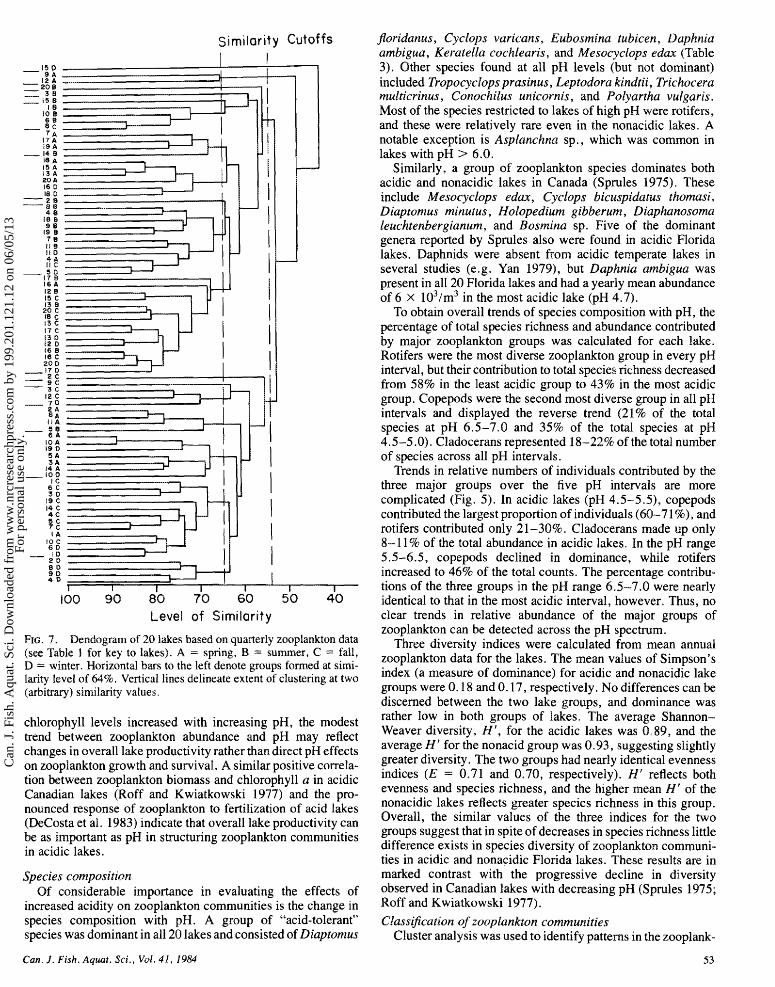
FIG. 7 . Dendogram of 20 lakes based on quarterly zooplankton data (st32 Table I for key to lakes). A = spring, B = summer, C -- fall, D = winter. Horizontal bars to the left denote groups formed at simi- larity level of 64%. Vertical lines delineate extent of clustering at two (arbitrary) similarity values.
chlorophyll levels increased with increasing pH, the modest trend between zooplankton abundance and pH may reflect changes in overall lake productivity rather than direct effects on zooplankton growth and survival. A similar positive correla- tion between zocsplankton biomass and chlorophyll a in acidic Canadian lakes (Roff and Kwiatkowski 1977) and the pro- nounced response of zooplankton to fertilization of acid lakes (DeCosta et al. 1983) indicate that overall lake productivity can be as important as pH in structuring zooplankton comnnunities in acidic lakes.
Species composition - Of considerable importance in evaluating the effects of
increased acidity on zooplankton communities is the change in species composition with pH. A group of "acid-tolerant" species was dominant in all 20 lakes and consisted of Diaptomus
Joridanus, Cyclops varicans, Eubosmina tubicen, Duphnia arnbigua, Keratelka cochlearis , and Mesocyclops edax ('Fable 3). Other species found at all pH levels (but not dominant) included Tropocyc Hops prase'nus, Leptodora kindtii, Trichocera multlerinus , Conochikus unicornis, and Pokyartha vulgaris. Most of the species restricted to lakes sf high pH were rotifers, and these were relatively rare even in the nonacidic lakes. A notable exception is Asplanchna sp., which was common in lakes with pH > 6.0.
Similarly, a group of zooplankton species dominates both acidic and nonacidic lakes in Canada (Sprules 1975). These include Mesocyclops @dm, Cyclops bicuspidatus thomasi, Diaptomus minutus, Holopedium gibberurn, Diaphanosoma leuchtenbergianum, and Bosmirza sp. Five of the dominant genera reported by Sprules also were found in acidic Florida lakes. Daphnids were absent from acidic temperate lakes in several studies (e.g. Yan 1979), but Daphnza ambzgua was present in all 20 Florida lalees and had a yearly mean abundance of 6 x l ~ ~ l m ' in the most acidic lake (pH 4.7).
To obtain overall trends of species composition with pH, the percentage of total species richness and abundance contributed by major zooplankton groups was calculated for each lake. Rotifers were the most diverse zooplankton group in every pH interval, but their contribution to total species richness decreased from 58% in the least acidic group to 43% in the most acidic group. Copepods were the second most diverse group in a11 pH intervals and displayed the reverse trend (21% of the total species at pH 6.5-7.0 and 35% of the total species at pH 4.5-5.0). Gladocerans represented 113-2276 of the total number of species across all pH intervals.
Trends in relative numbers of individuals contributed by the three major groups over the five pH intervals are more complicated (Fig. 5). In acidic lakes (pH 4.5-5.5), copepods contributed the largest proportion of individuals (68-71%), and rotifers contributed only 2 1-30%. Cladocerans made cap only 8- 1 1% of the total abundance in acidic lakes. In the pH range 5.5-6.5, copepods declined in dominance, while rotifers increased to 46% of the total counts. The percentage contribu- tions of the three groups in the pH range 6.5-7.0 were nearly identical to that in the most acidic interval, however. Thus, no clear trends in relative abundance of the major groups of zooplankton can be detected across the pH spectrum.
Three diversity indices were calculated from mean annual zooplankton data for the lakes. The mean values of Simpson's index (a measure of dominance) for acidic and nonacidic lake groups were 0.18 and 0.17, respectively. No differences can be discerned between the two lake groups, and dominance was rather low in both groups of lakes. The average Shannon- Weaver diversity, H ' , for the acidic lakes was 0.89, and the average H ' for the nonacid group was 0.93, suggesting slightly greater diversity. The two groups had nearly identical evenness indices (E = 0.71 and 0.70, respectively). H' reflects both evenness and species richness, and the higher mean M' of the nonacidic lakes reflects greater species richness in this group. Overill, the similar values of the three indices for the two groups suggest that in spite of decreases in species richness little difference exists in species diversity of zooplankton communi- ties in acidic and nonacidic Florida lakes. These results are in marked contrast with the progressive decline in diversity observed in Ganadian lakes with decreasing pH (Sprules 1975; Roff and Kwiatkowski 1977). CKass$cation of zocdplhnkton communities
Cluster analysis was used to identify patterns in the zooplank-
Can. .I. Fish. Aquat. Sci., Vo1. 41, 1984
Can
. J. F
ish.
Aqu
at. S
ci. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y 19
9.20
1.12
1.12
on
06/0
5/13
For
pers
onal
use
onl
y.

MONOM SP . . . d
............ D Bff ACH
PLEOS SP ........
L PATEL ..........
P PATUL .......... CON0 SP ..........
SlMO SP ........
B QUAD ...........
€3 CALY ............
C SPHAEW ........
MONOS SP ....... B HAVAN ............
ERGAS SP .......... JClV CLAD ......... K TAUR ............
KELLl SP .........
X C UNI .. . . . . . . . . . . .
c R T M U L T ............ 8 ............ X
T LONG 0 % T P R A S ............. I- KIND .........
AMBBG ...........
M EDAX ...........
............. C VARl
D FLOR ............
....... COPEPODl ........... NAbBPLl
COCHk .........
ASPL SP ..........
FlLlN SP .........
.......... B ANGUL B DIETER .
GASTff SP .......
K QUAD ............ 1 I I I
Level of Similar it y
FIG. 8 . Classification of zooplankton species based on mean abundance in the 20 lakes (see Table 3 for complete names sf organisms). Dominant "acid-tolerant" species found in all lakes form cluster A. Species indicated with arm asterisk are acid tolerant but not dominant.
ton communities in relation to pH. Two types of classification were generated: standard (Q-mode) analysis, which clustered lakes according to the similarity of species composition, and reverse (R-mode) analysis, which clustered species according to similarity of occurrence. Both analyses were performed using quantitative (actual count) data.
To assess overall trends in zooplankton communities, cluster- ing was performed on yearly mean data. Two major groups were formed at the 68% similarity level (Fig. 6). One contains seven nonacidic lakes (pH values in the range 6.03-6. $0). The second contains the 18 acidic lakes plus 3 nonacidic lakes (Geneva, Kingsley, and Annie) that are more oligotrophic than the other nonacidic lakes. This trend, along with the general trend of lower phytoplankton levels in acidic lakes, suggests that phytoplankton abundance has an important influence on zoo- plankton community structure.
A dendrogram also was generated from Q-mode classifica-
tion of quarterly data on the 20 lakes (Fig. 7), and results show some effects of seasonality on zooplankton communities. Eleven multiobject groups (and seven single-object groups) were formed at a similarity level of 64%; these were further clustered into two main groups at a similarity level of 55%. One of the latter groups was composed primarily of acidic lakes clustered into subgroups according to a single season (fall, winter, or spring). Two low-productivity nonacidic lakes and one moderately productive, colored, and nonacidic lake also fell into this group. The second major group included clusters made up primarily of nonacidic lakes from all four quarters plus summer quarter data for the acidic lakes. Overall, the results suggest that differences exist between the acidic and nonacidic lakes during fall, winter, and spring, but their zooplankton communities are similar during summer.
R-mode cluster analysis of abundance data on species in the lakes grouped the six dominant, acid-tolerant species together
54 Can. J . Fish. Aquut. Sci., Vol. 41, 1984
Can
. J. F
ish.
Aqu
at. S
ci. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y 19
9.20
1.12
1.12
on
06/0
5/13
For
pers
onal
use
onl
y.

with copepodites and nauplii at a high degree of similarity (Fig. 8). Other species or groups of species were added to this group in order of decreasing occurrence and abundance.
Concluding Remarks
Although trends were observed in planktonic communities over the pH range 4.7-6.8, it is difficult to separate direct effects of pH from covarying factors related to competition, predation, and nutrient status. For example, less acidic lakes tended to have higher nutrient concentrations and thus greater abundance of phytoplankton. Trends in zooplankton communi- ties were relatively slight and less consistent than those for phytoplankton. Species richness and abundance for both groups were lower in acidic lakes. Mean values of three diversity indices, on the other hand, were nearly identical for zooplank- ton in the acidic and nonacidic lake groups. A group of acid-tolerant zooplankton was dominant at all pH levels, and zooplankton diversity generally was much higher than in temperate lakes of comparable pH.
Eriksson et al. ($988) attributed the increased importance of pyrrhophytes in acidic Scandinavian lakes to an increase in large herbivorous zooplankton brought about by elimination of fish and the subsequent shift to invertebrate predators. As a group, pyrrhophytes are assumed to be poorly grazed by zooplankton (Porter 1973). Major regional differences in trophic-level interactions with pH may be a reflection of the available species pool. In contrast with the situation described for Scandinavian lakes, Confer et al. ($983) noted that small species dominate the macrozooplankton assemblages of North American acidic lakes, and predaceous cyclopoid copepods are not abundant even in fishless lakes. DeCosta and Sanicki (1978) attributed the importance of Bosrninu longirostris in an acidic West Virginia lake to the near absence of a major invertebrate predator, Messcyclops edax.
Regardless of cause, Florida macrozooplankton tend to be smaller bodied than those in comparable temperate assemblages (Crisman 19811. Large daphnids are absent, and Dayhnku ambigua, one of the smallest daphnid species in North America is dominant. Predation intensity appears to be less altered in acidic Florida lakes than that in comparable temperate lakes. Total fish biomass decreases with decreasing pH in Florida lakes, but no major species replacements have been noted down to pH 4.5 (T. L. Crisman, PI. Schramm, and A. Keller, unpub- lished data). In addition, no species of cyclopoid copepods were eliminated with decreasing pH. We suggest that the small response of zooplankton to pH, influenced by the regional absence of large-bodied species and lack of alteration in predation, may explain the dominance of chlorophytes rather than relatively inedible pyrrhophytes in the phytoplankton assemblages of acidic Florida lakes.
Acknowledgments
Assistance of S. Bloom (Department of Zoology, University of Florida) in statistical analyses and of C. D. Wendry in sampling is appreciated. We thank W. G. Spnmles and an anonymous reviewer for their helpful comments. This work was supported by grant no. $85560 from the USEPA, Conlallis, OR, and T. Maloney and C. Powers, project officers. Conclu- sions and opiniom do not necessarily reflect the views or policies of the USEBA. Manuscript preparation was supported
by a grant to the senior author from the Graduate School of the University of Minnesota.
References
ALMER, B. , W. DICKSON, C. EKSTROM, AND E. HBRNSTROM. 1978. Sulfur pollution and the aquatic ecosystems, p. 273-31 1. Hta J. 0 . Nriagu bed.] Sulfur in the environment. Part 11. Ecological impacts. J. Wiley & Sons, New York, NY.
ALMER, B . , W. DICKSON, C. EKSTROM, E. HORNSTROM, AND U. MILLER. 1974. Effects of acidification on Swedish lakes. Ambio 3: 30-36.
BARR, A. J., J. PI. GOODNIGHT, J. P. SALL, AND J. T. HELWIG. 1976. A user's guide to statistical analysis system. SAS Institute Inc., Raleigh, NC. 329 p.
BEAVER, J. R., AND T. L. CRISMAN. 1981. Acid precipitation and the response sf ciliated protozoans in Florida lakes. Verh. Int. Ver. Limsl. 21: 353-358.
BLOOM, S. A,, J. L. SIMON, AND V. D. HUNTER. 1977. A package of computer programs for benthic community analysis. Bull. Mar. Sci. 27: 577-580.
BREZONIK, B. L., E. S. EDGERTON, AND C. D. HENDRY. 1980. Acid precipitation and sulfate deposition in Florida. Science (Washing- ton, DC) 288: 1027-1029.
BREZONIK, P. L., C. D. WENDRY, E. S. EDGERTON, R. L. SCHULZE, AND T. L. CRISMAN. 1983. Acidity, nutrients, and minerals in atmospheric precipitation over Florida: deposition patterns, mech- anisms, and ecological effects. Rep. EBA 6001 3-83-004, Id .S. EPA, Corvallis, OR.
COCKE, E. C. 1966. The myxophyceae of North Carolina. Edwards Brothers, Inc., Ann Arbor, MI. 340 p.
CONFER, J. L., T. KAARET, AND G. E. LIKENS. 1983. Zooplankton diversity and biomass in recently acidified lakes. Can. J. Fish. Aquat. Sci. 40: 36-42.
CORMACK, W. H. 197 1. A review of classification. J. R. Stat. Soc. 134: 321 -367.
COWLING, E. B. 1982. Acid precipitation in historical perspective. Environ. Sci. Technol. 16: 1 10A- 123A.
CRISMAN, T. L. 198 1 . Algal control though trophic-level interactions: a subtropical perspective. p. 13 1 - 145. Proceedings of Workshop on Algal Management and Control. U.S. Amy Waterways Experiment Station, Vicksburg, MS. 313 p.
DECOSTA, J., AND A. JANICKI. 1978. The population dynamics and age structure of Bosnzina longirostrfs in an acid water impound- ment. Verh. Int. Ver. Limnol. 28: 2479-2483.
DECOSTA, J., A. JANICKI, G. SHELLITO, AND G. WILCBX. 1983. The effect of phosphorus additions in enclosures on the phytoplankton and zooplankton of an acid lake. Oikos 40: 283-294.
DILLON, P. J., N. D. YAN, W. A. SCHEIDER, AND N. CONROY. 1979. Acidic lakes in Ontario, Canada: characterization, extent and responses to base and nutrient additions. Arch. Hydrobiol. Beih. Ergegn. Limnol. 1 3: 317-336.
DUMONT, H. J., I. VAN DE VELDE, AND S. DUMONT. 1975. The dry weight estimate of biomass in a selection of Cladwera, Copepsda, and Rotifera from the plankton, periphyton, and benthos of continental waters. Oecologia 19: 75-97.
DUTHIE, H. C., AND R. SOCHA. 1976. A checklist of the freshwater algae of Ontario, exclusive of the Great Lakes. Nat. Can. 103: 83- 109.
EDMONBS~N, W. T. [ED.] 1959. Fresh-water biology. J. Wiley & Sons, New York, NY. 1249 p.
ERIKSSON, M. 0 . G., L. HENRIKSEN, B. I. NILSSBN, G. NYMAN, H. G. OSCARSBN , AND A. E. STENSON. 1980. Predator-prey relations important for the biotic changes in acidified lakes. Ambio 9: 248-249.
FEE, E. J. 1988. The important factors controlling annual phytoplank- ton production in the Experimental Lakes Area. Can, J. Fish. Aquat. Sci. 37: 513-522.
RNDLAY, D. L., AND PI. J. KLING. 1979. A species Iist and pictorial
Can. J . Fish. Aquat. Sci., Vol. 41, 1984
Can
. J. F
ish.
Aqu
at. S
ci. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y 19
9.20
1.12
1.12
on
06/0
5/13
For
pers
onal
use
onl
y.

reference to the phytoplankton of central and northern Canada. Dep. Fish. Environ., Fish. Mar. Sew. Manage. Rep. No. 1503: 619 p.
HAINES, 9. A. 1981. Acidic precipitation and its consequences for aquatic ecosystems: a review. Trans. Am. Fish. Soc. 110: 669-707.
HENDREY, G . R., K. BAALSRUD, T. TRAAEW, M. LAAKE, AND G. WADDUM. 1976. Acid precipitation: some hydrobiological changes. Ambio 5: 224-227.
HENDRY, C, B. 1983. Spatial and temporal variations in bulk, wet, and dry atn~ospheric deposition of acidity and minerals across Florida. Ph.D. thesis, University of Florida, Gainesvil fe, FL. 2 10 p.
HENDRY, C. D., AND P. L. BREZONBK. 1984. Chemical composition of softwater Florida lakes and their sensitivity to acid precipita- tion. Water Resour. Bull. 20. (In press)
KWIATKOWSKI, R. E., AND I. C, WOFF. 1976. Effects of acidity on the phytoplankton and primary productivity of selected northern Ontario lakes. Can. J. Bot. 54: 2546-2561.
LEIVESTAD, M . , G . R. HEWDREY, I. P. MUNIZ, AND E. SNEKVHK. 1976. Effects of acid precipitation on freshwater organisms, p. 87- 1 1 8 . lfi F, PI. Braekke [ed. ] Impact of acid precipitation on forest and freshwater ecosystems in Norway. SNSF Project FR 6176, Aas, Norway.
LIKENS, G . E., R. F. WRIGHT, J. N. GALLOWAY, AND T. J . BUTLER. 1979. Acid rain. Sci. Am. 241 (4): 43-5 1 .
NATIONAL RESEARCH COUNCIL OF CANADA. 1981. Acidification in the Canadian aquatic environment: scientific criteria for assessing the effects of acidic deposition on aquatic ecosystems. Publ. NO. 18475: 369 p.
OMERNHK, J. M., AND C. F. POWERS. 1982. Total alkalinity of surface waters. EPA-600lD-82-333. I%. S. EPA, Corvallis, OR.
~ N N A R , R. W. 1953. Fresh-water invertebrates of the United States. Ronald Press, New York, NY. 769 p.
PORTER, K. G. 1973. Selective grazing and differential digestion of agae by zooplankton. Nature (London) 244: 179-180.
P~ESCOTT, G. W. 1954. How to know the fresh-water algae. W. C. Brown Publishers, Dubuque, IA. 272 p.
RABDUM, G. G., A. HOBAEK, E. R. LOMSLAND, AND T. JOHNSON. 1980. Phytoplankton and zooplankton in acidified lakes in south Norway, p. 332-333. In D. Drablos and A. Tollan Bed.] Proc. Hnt. Conf. Ec01. Imp. Acid Precip., Sandefjord, Norway. SNSF Project, Aas, Norway.
ROOF, J. R., AND R. E. KWIATKOWSKI. 1977. Zooplankton and zoobenthos communities sf selected northern Ontario lakes of different acidities. Can. J. Zool. 55: 899-91 1 .
SMITH, 6. M. 1950. The fresh-water algae of the United States. McGraw-Hill Book Company, New York, NY. 719 p.
SPRULES, W. G. 1975. Midsummer crustacean zooplankton communi- ties in acid-stressed lakes. J . Fish. Res. Board Can. 32: 389-395.
WEBER, C. 1. 197 1. A guide to the common diatoms at water pollution surveillance system stations. U.S. EPA, Cincinnati, OH. 100 p.
WHITFORD, L. A., AND G. J. SCHUMAGHER. 1973. A manual of the fresh-water algae in North Carolina. Sparks Press, Raleigh, NC. 324 p.
WILCOX, G., AND J. DECOSTA. 1982. The effect of phosphorus and nitrogen addition on the algal biomass and species composition of an acidic lake. Arch. Hydrobiol. 94: 393-424.
YAN, N. D. 1979. Phytoplankton community of an acidified, heavy metal contaminated lake near Sudbury, Ontario: 1973-77. Water Air Soil Pollut. B I : 43-55.
YAN, N. D., W. A. SCHEIDER, AND P. J. DILLON. 1977. Chemical and biological changes in Nelson Lake, Ontario, following experi- mental elevation of lake pH. Prsc. Can. Symp. Water Pollut. Res. 12: 213-231.
YAN, N. D., AND P. STOKES. 1978. Phytoplankton of an acidic lake, and its response to experimental alterations of pH. Environ. Conserv. 5: 93- 100.
YAN, N. D., AND R. STRUS. 1980. Crustacean zooplankton cammuni- ties of acidic, metal contaminated lakes near Sudbury, Ontario. Can. J. Fish. Aquat. Sci. 37: 2282-2293.
Can. J . Fish. ,4quar. Sci., Vol. 41, 1984
Can
. J. F
ish.
Aqu
at. S
ci. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y 19
9.20
1.12
1.12
on
06/0
5/13
For
pers
onal
use
onl
y.





























