Embed Size (px)
Citation preview

Population dynamics of Aceodromus convolvuli (Acari:Mesostigmata: Blattisociidae) on spontaneous plantsassociated with Jatropha curcas in central Brazil
Wilton P. Cruz • Renato A. Sarmento • Marcal Pedro-Neto •
Adenir V. Teodoro • Diego M. Rodrigues • Gilberto J. de Moraes
Received: 5 December 2013 / Accepted: 5 June 2014 / Published online: 19 June 2014� Springer International Publishing Switzerland 2014
Abstract Spontaneously growing plants are commonly considered competitors of cul-
tivated plants. Owing to the lack of specificity of many arthropods, spontaneous plants may
be attacked by the same arthropods that attack cultivated plants and they may also harbor
natural enemies of organisms harmful to cultivated plants. Aceodromus convolvuli Muma
(Blattisociidae) has been reported recently in relatively large numbers in Tocantins state,
central Brazil, mostly on Helicteres guazumifolia Kunth (Malvaceae). Very little has been
reported about the population dynamics of blattisociid mites under field conditions. The
objective of this work was to study the population dynamics of A. convolvuli in Gurupi,
Tocantins state, to evaluate its possible interaction with associated mites. Monthly samples
were taken from leaves of the 11 most abundant and frequent spontaneous plants in a
Jatropha curcas L. (Euphorbiaceae) plantation. About 96.5 % of the specimens of A.
convolvuli were collected in the rainy season. The patterns of variation of the population of
A. convolvuli and of predators belonging to the family Phytoseiidae were similar, but A.
convolvuli was much more numerous than all phytoseiid specimens combined. Highly
W. P. Cruz � R. A. Sarmento (&) � M. Pedro-NetoUniversidade Federal de Tocantins (UFT), PO Box 66, Gurupi, State of Tocantins, Brazile-mail: [email protected]
W. P. Cruz � G. J. de MoraesPrograma de Pos-graduacao em Entomologia Agrıcola, Universidade Estadual Paulista Julio deMesquita Filho, UNESP, Jaboticabal, State of Sao Paulo, Brazil
A. V. TeodoroEmbrapa Tabuleiros Costeiros, Av. Beira Mar 3250, PO Box 44, Jardins, Aracaju, State of Sergipe,Brazil
D. M. RodriguesDepartamento de Ciencias Agrarias, Universidade Federal do Para (UFPA), Maraba, State of Para,Brazil
G. J. de MoraesDepartamento de Entomologia e Acarologia, Escola Superior de Agricultura ‘‘Luiz de Queiroz’’,Universidade de Sao Paulo (ESALQ/USP), Piracicaba, State of Sao Paulo, Brazil
123
Exp Appl Acarol (2014) 64:309–319DOI 10.1007/s10493-014-9828-5

significant correlations were observed between A. convolvuli densities and relative
humidity or diversity of spontaneous plants. When only mites on H. guazumifolia were
considered, highly significant correlation was also observed between densities of A. con-
volvuli and of mites of the family Tetranychidae. The results suggested that A. convolvuli
could be a predator of tenuipalpid and/or tetranychid mites. Studies about its biology are
needed to determine its preferred food sources and potential as biological control agent.
Keywords Ecology � Predation � Biological control � Blattisociid mite
Introduction
Spontaneously growing plants are often considered competitors of cultivated plants, able to
reduce the yield of the latter, being in those cases referred to as invasive plants or weeds.
Because of the lack of specificity of many arthropods, spontaneous plants may be attacked
by the same arthropods that attack cultivated plants and could thus serve as reservoirs of
pest organisms. However, they may also harbor natural enemies of organisms that are
harmful to cultivated plants (Altieri et al. 2003; Bellini et al. 2005).
The main mites of the order Mesostigmata found on plants are predators belonging to
the family Phytoseiidae. Some species of this family are considered to be important to the
control of several plant pests (McMurtry and Croft 1997; Gerson et al. 2003). Much less
common on plants are the members of the family Blattisociidae, which were previously
included in the family Ascidae (Lindquist and Evans 1965; Halliday et al. 1998; Lindquist
et al. 2009). Several blattisociids have been mentioned to prey on arthropods (Gerson et al.
2003; Lindquist et al. 2009).
Aceodromus Muma is a mite genus originally included in Phytoseiidae (Muma 1961)
and later transferred to Blattisociidae (Lindquist and Chant 1964). Aceodromus convolvuli
Muma was described from specimens collected in Florida, United States of America, on
leaves of Convolvulus sp. (Muma, 1961). It was later reported by Lindquist and Chant
(1964) in Jamaica on leaves of eggplant and Crotalaria sp., and by Fadamiro et al. (2009)
in Alabama, USA, on unidentified plants in citrus orchards. Aceodromus convolvuli was
first reported in Brazil by Moraes et al. (1993), on spontaneous plants growing in several
states of the northeastern part of the country. It was also reported on coffee in Sao Paulo
State, Brazil (Mineiro et al. 2006).
More recently, A. convolvuli was reported from Tocantins state, in central Brazil, in a
study conducted by Cruz et al. (2012) on the mite fauna of physic nut Jatropha curcas L.
(Euphorbiaceae) and associated spontaneous plants. Jatropha curcas has been considered
for extended cultivation in several tropical countries, mainly for biodiesel production
(Openshaw, 2000).
In the work conducted by Cruz et al. (2012), 1,141 specimens of A. convolvuli were
collected on samples of 11 species of spontaneous plants. About 69 % of those specimens
were found on Helicteres guazumifolia Kunth (Malvaceae). This plant has been reported
from southern Mexico to the central region of Brazil, in open, secondary and semidecid-
uous forests, gallery forests, pastures, zones of periodic fires and clearings, dry or moist
thickets and grassy or bushy slopes (Goldberg 2009). In Venezuela, this plant has been
reported mainly in open habitats with dry, sandy and rocky soils (Rondon and Cumana-
Campos 2007).
310 Exp Appl Acarol (2014) 64:309–319
123

The objective of this work was to conduct an analysis of the population dynamics of A.
convolvuli, based on the samplings conducted by Cruz et al. (2012), evaluating its possible
interaction with the associated mites on those 11 spontaneous plant species. This work is
one of the rare efforts to evaluate the population dynamics of species of the ascid sense
Lindquist and Evans (1965). Apparently, the only other study of this type refers to the
blattisociid Lasioseius parberlesei Bhattacharyya in Taiwan (Tseng 1984), where it was
considered an effective predator of Steneotarsonemus spinki Smiley (Tarsonemidae) on
rice. Thus, not much can be anticipated in terms of the response of A. convolvuli to the
varying climatic factors, or in terms of its possible natural prey. However, as other blat-
tisociid mites have a preference for humid environments, we anticipated that the same
occurs with A. convolvuli.
Materials and methods
The data used for the analysis presented here were collected in the study conducted by
Cruz et al. (2012). The work involved the evaluation of A. convolvuli and associated mite
species on spontaneous plants from a 4-year old cultivation of J. curcas of about 0.5 ha, in
Gurupi (11�4802900S, 48�5603900W; 280 m altitude), Tocantins state, in a subtropical region
with well defined rainy (November–April) and dry (May–October) seasons. The natural
vegetation of this locality is classified as ‘‘Cerrado’’, a very diverse savannah type of biome
covering slightly over 20 % of the Brazilian territory, where annual rainfall ranges between
800 and 2,000 mm, but with a pronounced dry season (Ratter et al. 1997). The experi-
mental field was mainly surrounded by pasture (mostly Brachiaria sp., Poaceae), except
for one side, edging an area of natural vegetation.
The height of the spontaneous plants varied from about 15 cm (dry season) to about
120 cm (wet season). They were not controlled during the conduction of the study or in the
previous year. No fertilization or pesticide spray was done during the study or in the period
of 2.5 years before it started (Cruz et al. 2012).
Mite sampling
Samples were taken from 15 sites (9 m2 each) randomly delimited within the experimental
field. Eleven plant species were selected for the evaluations, based on their abundance and
frequency in the area (Cruz et al. 2012). The selection was done by counting the plants of
each species within two quadrats (0.25 m2) randomly thrown in each sampling site (Mu-
eller-Dombois and Ellenberg 1974; Cruz et al. 2012). Selected species were those with
abundance of at least 2.5 plants per quadrat and which were present with a frequency of at
least 10 % of the quadrats. The same selected plant species were sampled throughout the
work, regardless of the variation in their abundance or frequency along the time. Identi-
fication of spontaneous plants was conducted with the use of a manual (Lorenzi 2006) and
the help of M. Ignacio, a botanist. The selected plants were: Fabaceae—Calopogonium
mucunoides Desv.; Lamiaceae—Hyptis suaveolens (L.) Poit.; Malvaceae—Peltaea riedelii
(Gurke) Standl., Sida urens L., Sida cordifolia L., Sida rhombifolia L., Urena lobata L.,
H. guazumifolia, Waltheria americana L.; Poaceae—Andropogon gayanus Kunth and
Urochloa mutica (Forssk.) T.Q. Nguyen (Angiosperm Phylogeny Group 2009). The
diversity of spontaneous plants was estimated at each sampling date by calculating the
Shannon–Wiener (H0) index with DIVES software version 2.0. Sampling of spontaneous
plants for diversity estimation was conducted using the methodology proposed for Mueller-
Exp Appl Acarol (2014) 64:309–319 311
123

Dombois and Ellenberg (1974), described above. Subsequently, the spontaneous plants’
richness and abundance were obtained.
The samples were collected once between the 20th and 25th of each month, from
February 2010 to January 2011. At each sampling date, 10 (Fabaceae, Lamiaceae and
Malvaceae) or 20 (Poaceae) leaves of each selected plant species present in each sampling
site were collected, constituting a sample. Thus, for each species, the number of samples
taken at each sampling date varied according to its respective frequency at that sampling
date, for a minimum of two and a maximum of 11 samples. The leaves of each species
from each sampling site were placed in a plastic bag that in turn was placed in a cool box
(about 20 �C) and transferred to the laboratory, where they were maintained in a refrig-
erator (about 10 �C) for up to 5 days, until examined under a stereomicroscope. For
quantification and identification, mites were mounted in Hoyer’s medium; however, when
a certain species was too abundant, the mites were counted and only part of the specimens
was mounted for confirmation of the identification. Identification of mite species was
carried out by the third and the last authors, using the original descriptions and relevant
complementary descriptions of the species of each group.
Leaf area was estimated at each sampling date. Climatic data (daily temperature, rel-
ative humidity and rainfall) were obtained from ‘‘Estacao Meteorologica de Gurupi (IN-
MET)’’, at Universidade Federal do Tocantins campus, about 20 km from the experimental
site. The analyses were conducted by taking into account monthly average temperature and
relative humidity and monthly total rainfall, in all cases for the period preceding each
sampling date (i.e., the most recent monthly reading was taken).
Statistical analysis
To estimate the main factors related to the fluctuation of A. convolvuli throughout the work,
data distribution was initially checked with Shapiro–Wilk normality test, using the Shap-
iro.test function procedure of R program. Since many of the variables did not have normal
distribution, Spearman’s test was used to determine simple correlations between variables
(population densities of A. convolvuli, phytoseiids, tetranychids and tenuipalpids, as well as
indexes of plant diversity, rainfall, relative humidity and temperature). This was done with
the use of the Spearman’s rank correlation (rho) function procedure or R program. A
multiple regression analysis relating the population densities of A. convolvuli with the
variables showing significant simple correlation with densities of A. convolvuli and not
related among themselves was performed, using ‘‘model’’ of R to obtain the coefficients of
the function and Summary (model) to determine the level of significance of the variables and
of the model as a whole, and R2 of the model (R Development Core Team 2006).
Results
A re-examination of the mites collected showed the need to correct or complement the
identification reported in Cruz et al. (2012), as indicated in Table 1.
Relation between Aceodromus convolvuli and environmental factors in rainy and dry
seasons
Considering all plants together, about 96.5 % of the specimens of A. convolvuli were
collected in the rainy season (Table 2). In the dry season, that species was only found on H.
312 Exp Appl Acarol (2014) 64:309–319
123

guazumifolia and C. mucunoides, at very low numbers. Total rainfall, average temperature
and average relative humidity were 964 mm, 26 ± 0.6 �C and 80 ± 1.1 % in the wet
season, and 80 mm, 26 ± 1.6 �C and 59 ± 3.9 % in the dry season, respectively.
Dynamics of Aceodromus convolvuli on all spontaneous plants
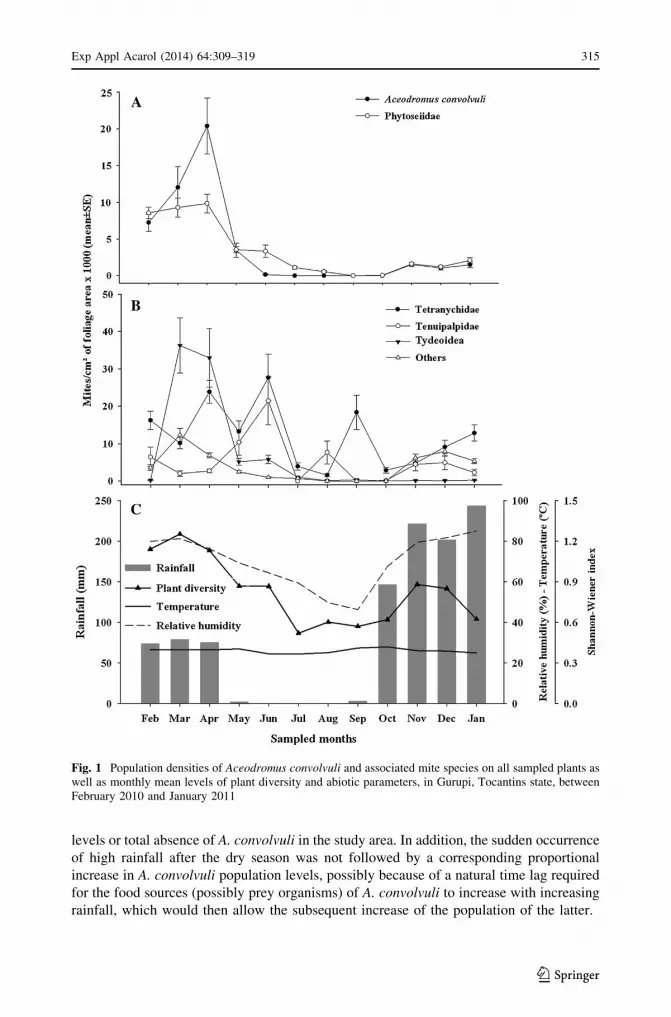
When all spontaneous plant species were considered together, the densities of A. convolvuli
increased between February and April, when the maximum level was reached (Fig. 1a).
Densities reduced drastically to nearly zero in June, remaining so until September; it again
increased progressively but slightly until November, remaining about the same level until
next January. The pattern of variation of the phytoseiid population was similar, except that
the decline of the population after reaching a peak density in April was more gradual.
Among other mite groups, the highest densities were attained by the Tydeoidea
(including Tydeidae and Iolinidae), which peaked in March, followed by the Tetranychidae
and the Tenuipalpidae, both reaching the maximum level in June (Fig. 1b). The highest
densities of A. convolvuli and phytoseiids coincided with the highest levels of plant
diversity and with the end of the rainy season (Fig. 1c).
Highly significant correlations were observed between the densities of A. convolvuli and
phytoseiids, as well as between the former and plant diversity indexes (Table 3); signifi-
cant correlations were observed between A. convolvuli densities and relative humidity. No
significant correlation between A. convolvuli densities and any of the remaining inde-
pendent variables was observed.
Owing to the significant correlations between phytoseiid densities and plant diversity as
well as between levels of relative humidity and plant diversity (Table 3), the latter
parameter was not included in the multiple regression analysis. In that analysis, the only
significant factor of the generated equation corresponded to phytoseiid densities, and the
coefficient of determination (R2) of the equation was 0.78.
Dynamics of Aceodromus convolvuli on Helicteres guazumifolia
The patterns of variation of the densities of A. convolvuli and phytoseiids were similar to
those when all spontaneous plant species were considered together (Fig. 2a). However, the
peak population density of A. convolvuli was much higher, while that of the phytoseiids
was only slightly higher. Differently from what was observed when all plants were con-
sidered together, densities of Tydeoidea were low during the whole observation period
Table 1 Correction and complementation of the identification of the mite species reported by Cruz et al.(2012), based on a re-examination of the specimens
Reported by Cruz et al. (2012) Corrected identification
Tetranychidae
Mononychellus sp.Panonychus sp.
Mononychellus planki (McGregor)Neotetranychus granifer Feres & Flechtmann
Phytoseiidae
Neoseiulus aff. mumaiNeoseiulus fallacis
Neoseiulus benjamini (Schicha)Neoseiulus pluridentatus Lofego & Moraes
Phytoseiulus sp.Typhlodromalus clavicus
Phytoseius guianensis De LeonAmblydromalus sp.
Exp Appl Acarol (2014) 64:309–319 313
123

(Fig. 2b). Among the remaining mite groups, the highest levels were attained by the
tetranychids and the tenuipalpids, also in June, being these about twice as high as deter-
mined for all plants together. Yet, variations in the populations of mites other than A.
convolvuli and phytoseiids were much more erratic than observed when all plants were
considered together.
Significant correlations (Table 3) were observed between A. convolvuli densities and
phytoseiid densities, tetranychid densities, relative humidity, and plant diversity. No sig-
nificant correlation was observed between the remaining independent variables with A.
convolvuli densities.
Because of the significant correlations between phytoseiid and tetranychid densities,
phytoseiid densities and plant diversity, relative humidity and plant diversity, and tetr-
anychid densities and plant diversity (Table 3), the multiple regression analysis included
only phytoseiid and tenuipalpid densities, temperature and rainfall. Similarly to what was
determined when all spontaneous plant species were considered together, the only sig-
nificant factor of the generated equation was phytoseiid densities, and the coefficient of
determination of the equation was 0.58.
Discussion
The analyses of the numbers of A. convolvuli in each of the two distinct seasons prevailing
in Gurupi as well as of the dynamics of each variable along the evaluation period suggested
a strong influence of rainfall and relative humidity on the densities of A. convolvuli. The
significant correlation between the population level of A. convolvuli and the levels of
relative humidity confirmed that assumption, but the non-significant correlation between
the first parameter and rainfall seemed not to support it. This apparent contradiction could
be due to the non-linear relation between A. convolvuli population densities and rainfall; in
this sense, there might be a minimum rainfall threshold level below which A. convolvuli is
about uniformly and negatively affected, independent of the amount of rain. Hence,
absence or incidence of very low rainfall between May and September resulted in very low
Table 2 Seasonal occurrence of Aceodromus convolvuli on spontaneous plants, in the dry and rainy sea-sons, in Gurupi, Tocantins state, Brazil, from February 2010 to January 2011
Spontaneous plants Dry season (May–October) Rainfall season (November–April) Total
Helicteres guazumifolia 38 746 784
Calopogonium mucunoides 2 135 137
Sida urens 0 48 48
Ureda lobata 0 41 41
Waltheria americana 0 41 41
Andropogon gayanus 0 23 23
Hyptis suaveolens 0 19 19
Sida cordifolia 0 18 18
Urochloa mutica 0 16 16
Peltaea riedelii 0 8 8
Sida rhombifolia 0 6 6
Total 40 1,101 1,141
314 Exp Appl Acarol (2014) 64:309–319
123
levels or total absence of A. convolvuli in the study area. In addition, the sudden occurrence
of high rainfall after the dry season was not followed by a corresponding proportional
increase in A. convolvuli population levels, possibly because of a natural time lag required
for the food sources (possibly prey organisms) of A. convolvuli to increase with increasing
rainfall, which would then allow the subsequent increase of the population of the latter.
Fig. 1 Population densities of Aceodromus convolvuli and associated mite species on all sampled plants aswell as monthly mean levels of plant diversity and abiotic parameters, in Gurupi, Tocantins state, betweenFebruary 2010 and January 2011
Exp Appl Acarol (2014) 64:309–319 315
123

With the onset of the dry season, the sharper decline of A. convolvuli population in
comparison with the phytoseiid population suggest the former to be more sensitive to lower
humidity than the latter. This pattern would be expected, as members of Ascidae sensu lato
Table 3 Significant correlations between the population densities of Aceodromus convolvuli and biotic andabiotic factors as well as between other combinations of factors for all spontaneous plants and on Helicteresguazumifolia alone, in Gurupi, Tocantins State, Brazil, from February 2010 to January 2011
Factors Spontaneous plants Helicteres guazumifolia
r P r P
Phytoseiidae 0.93 0.000009 0.80 0.002
Plant diversity 0.92 0.00001 0.87 0.0002
Relative humidity 0.66 0.01 0.71 0.01
Tetranychidae 0.66 0.01
Phytoseiidae 9 plant diversity 0.87 0.003 0.83 0.0007
Phytoseiidae 9 Tetranychidae 0.78 0.03
Relative humidity 9 plant diversity 0.60 0.04 0.61 0.04
Tetranychidae 9 plant diversity 0.61 0.03
Fig. 2 Population densities of Aceodromus convolvuli and associated mite species on Helicteresguazumifolia, in Gurupi, Tocantins state, between February 2010 and January 2011
316 Exp Appl Acarol (2014) 64:309–319
123

Lindquist and Evans (1965) are usually found on aerial plant parts only in regions of
prevailing high levels of relative humidity. In a study conducted in northeastern Brazil,
Rodrigues et al. (1996) determined the significant level of association A. convolvuli with
several other mites and plants. They determined the association to be negative with the
phytoseiid Neoseiulus (=Amblyseius) idaeus Denmark & Muma and positive with the
phytoseiid Amblydromalus (=Typhlodromalus) manihoti (Moraes), species that predomi-
nate respectively under conditions of low and high humidity levels (Moraes et al. 1990). As
commonly observed in areas of the Cerrado vegetation, the region where the study was
conducted has marked alternations of very dry and very humid seasons. Thus, the prev-
alence of A. convolvuli during part of the year in Gurupi leads to the conclusion that it has
an efficient mechanism to allow survival in the dry season, though at very low population
levels (Cruz et al. 2013).
Faunistic surveys on leaves of cultivated plants or plants of the natural vegetation have
usually shown outstandingly higher abundance of phytoseiids in comparison with other
mesostigmatid mites. Thus, the finding of a larger population density of A. convolvuli in
comparison with that of all phytoseiid species combined is uncommon, deserving detailed
analysis. A question to be answered is why A. convolvuli is so abundant in the region where
the study was conducted. Among other reasons, the relatively high abundance could be
related to the presence of H. guazumifolia, onto which A. convolvuli was by far the most
common Mesostigmata. However, in which way the plant affects the mite could not be
determined, because of the absence of information about its biology. The higher proportion
of A. convolvuli than of phytoseiids was not related to a conceivable more intense sur-
veying effort on this plant, as at each sampling date the examined leaf area of that plant
species corresponded to only about 20 % of all examined leaves.
Although not rare, H. guazumifolia was not one of the most common plants considered
in this study. It was only the eighth on the list of the most abundant, with a patchy
distribution in the study area. This species is a shrub whose leaves are covered by star-
shaped trichomes, under which A. convolvuli were found. Those structures seem to offer
some protection to the predators, as suggested by the fact that it was rather difficult to
collect them with a brush during the evaluations. Several factors have been related to the
presence of trichomes, including the maintenance of lower temperature on the leaf surface,
protection from the effect of UV light and the retention of a thicker boundary layer of
higher relative humidity over the leaf surface (Wagner et al. 2004; Coder and Warnell
2013). These characteristics might favor A. convolvuli.
As reported by Ordaz et al. (2011), several studies have been conducted on the mor-
phology, phenology, ecology and biochemical characteristics of H. guazumifolia, a plant of
medicinal importance in South America. Characteristics about patterns of nectar produc-
tion, morphology of floral nectaries, nectar composition and hummingbird association were
reported by Goldberg (2009). The chemical composition of essential oils of leaves of H.
guazumifolia was studied by Ordaz et al. (2011). The authors reported that the predominant
compounds were non-terpenoid volatile secondary metabolites. The role of alternative food
such as nectar and secondary compounds to the predominance of A. convolvuli on H.
guazumifolia in this study has not been determined. To our knowledge this is the first study
of the mite fauna of this plant.
Considering the possibility of using A. convolvuli for the control of pest organisms,
future studies should include detailed evaluations on its accepted food sources. The results
of the present study suggest that items to be investigated should include tetranychid and
tenuipalpid mites. The relatively low correlation between the densities of A. convolvuli and
tetranychids as well as the non significant correlation between the densities of A.
Exp Appl Acarol (2014) 64:309–319 317
123

convolvuli and tenuipalpids might not necessarily indicate that they do not interact. Instead,
that could be a function of a complex predator–prey interactions, with non-synchronous
population fluctuations, and/or due to the concurrent confusing effect of other environ-
mental factors. The inverse patterns of variation of A. convolvuli and tenuipalpids
graphically distinguishable between February and June further supports this possibility,
with an increase in the population level of the former being followed by a (slower)
reduction of the population of the latter during the end of the rainy season, and an inverse
trend at the onset of the dry season, which, as previously discussed, seems to disfavor A.
convolvuli. Detailed studies of predator–prey interactions should be stimulated, especially
in areas where mites have been barely investigated, as in central Brazil. These may lead to
the discovery of new prospective biological control agents of this relatively little known
group of mesostigmatid mites. Considering that another blattisociid mite, Lasioseius
lindquisti Nasr & Abou-Awad, has been reported to develop and reproduce when offered
pollen of Phoenix dactylifera as food source (Momen et al. 2011), pollen of H. guaz-
umifolia should also be investigated as a possible food source for this mite.
Acknowledgments The authors thank the National Council for Scientific and Technological Development(CNPq) for financial support (Projects 620028/2008-4 and 475408/2008-0) and Coordination of Improve-ment of Higher Education Personnel CAPES-Brazil for providing scholarships to the first and third authors(Projects: PROCAD-NF and CAPES-PNPD). We thank M. Ignacio (Universidade Federal do Tocantins,Brazil) for help with the identification of part of the plants collected in this study.
References
Altieri MA, Silva EM, Nicholls CI (2003) O papel da biodiversidade no manejo de pragas. Holos, RibeiraoPreto
Angiosperm Phylogeny Group [A.P.G.] (2009) An update of the angiosperm phylogeny group classificationfor the orders and families of flowering plants: APG III. Bot J Linnean Soc 161:105–121
Bellini MR, Moraes GJ, Feres RJF (2005) Plantas de ocorrencia espontanea como substratos alternativospara fitoseıdeos (Acari, Phytoseiidae) em cultivos de seringueira Hevea brasiliensis Muell. Arg.(Euphorbiaceae). Rev Bras Zool 22:35–42
Coder KD, Warnell DB (2013) Tree trichomes: big hairy tree? School of Forest Resources ExtensionPublication, The University of Georgia. http://warnell.forestry.uga.edu/SERVICE/LIBRARY/for99-006/for99-006.pdf. Accessed 30 Aug 2013
Cruz WP, Sarmento RA, Teodoro AV, Erasmo EAL, Pedro Neto M, Ignacio M, Ferreira Junior DF (2012)Acarofauna em cultivo de pinhao-manso e plantas espontaneas associadas. Pesq Agropec Bras47:319–327. doi:10.1590/S0100-204X2012000300002
Cruz WP, Sarmento RA, Teodoro AV, Pedro Neto M, Ignacio M (2013) Driving factors of the communitiesof phytophagous and predatory mites in a physic nut plantation and spontaneous plants associated. ExpAppl Acarol 60:509–519. doi:10.1007/s10493-013-9663-0
Fadamiro HY, Xiao Y, Nesbitt M, Childers CC (2009) Diversity and seasonal abundance of predaceousmites in Alabama Satsuma Citrus. Ann Entomol Soc Am 102:617–628
Gerson U, Smiley RL, Ochoa R (2003) Mites (Acari) for pest control. Blackwell Science, OxfordGoldberg L (2009) Patterns of nectar production and composition, and morphology of floral nectaries in
Helicteres guazumifolia and Helicteres baruensis (Sterculiaceae): two sympatric species from theCosta Rican tropical dry forest. Rev Bio Trop 57(Suppl 1):161–177
Halliday RB, Walter DE, Lindquist EE (1998) Revision of the Australian Ascidae (Acarina : Mesostigmata).Invert Taxon 12:1–54
Lindquist EE, Chant DA (1964) A redescription of Aceodromus Muma and its transfer to the Blattisociinae(Acarina: Blattisociidae). Can Entomol 96:500–507
Lindquist EE, Evans GO (1965) Taxonomic concepts in the Ascidae, with a modified setal nomenclature forthe idiosoma of the Gamasina (Acarina: Mesostigmata). Mem Entomol Soc Can 47:1–64
Lindquist EE, Krantz GW, Walter DE (2009) Classification. In: Krantz GW, Walter DE (eds) A manual ofacarology. Texas Tech Univer-sity Press, Lubbock, pp 97–103
318 Exp Appl Acarol (2014) 64:309–319
123

Lorenzi H (2006) Manual de identificacao e controle de plantas daninhas, 6th edn. Plantarum, Nova OdessaMcMurtry JA, Croft BA (1997) Life-styles of phytoseiid mites and their role in biological control. Annu Rev
Entomol 42:291–321Mineiro JLC, Sato ME, Raga A, Arthur V, Moraes GJ, Sarreta FO, Carrijo A (2006) Diversity of mites
(Arachnida: Acari) on Coffea arabica L. cv. ‘Mundo Novo’ in Jeriquara and Garca counties, state ofSao Paulo. Biota. doi:10.1590/S1676-06032006000200012
Momen FM, Abou-Elela AM, Metwally AM, Naser AK, Saleh KHM (2011) Biology and feeding habits ofthe predacious mite, Lasioseius lindquisti (Acari: Ascidae) from Egypt. Acta Phytopathol EntomolHung 46:151–163
Moraes GJ, Alencar JA, Wenzel Neto F, Mergulhao SMR (1990) Explorations for natural enemies of thecassava green mite in Brazil. In: Howeler RH (ed) Proceedings of VIII symposium of the internationalsociety of tropical root crops, Bangkok, Thailand, 1988. Departament of Agriculture of Thailand/CIAT/CIP, Bangkok, pp 351–353
Moraes GJ, Alencar JA, Lima JLS, Yaninek JS, Delalibera I Jr (1993) Alternative plant habitats for commonphytoseiid predators of the cassava green mite (Acari: Phytoseiidae, Tetranychidae) in northeast Brazil.Exp Appl Acarol 17:77–90. doi:10.1007/BF00156945
Mueller-Dombois D, Ellenberg H (1974) Aims and methods of vegetation ecology. Wiley, New York 547pMuma MH (1961) Subfamilies, genera, and species of Phytoseiidae (Acarina: Mesostigmata). Bull Fla State
Mus Biol Sci 5:267–302Openshaw K (2000) A review of Jatropha curcas: an oil plant of unfulfilled promise. Biomass Bioenergy
19:1–15. doi:10.1016/S0961-9534(00)00019-2Ordaz G, D’Armas H, Yanez D, Moreno S (2011) Composicion quımica de los aceites esenciales de las
hojas de Helicteres guazumifolia (Sterculiaceae), Piper tuberculatum (Piperaceae), Scoparia dulcis(Arecaceae) y Solanum subinerme (Solanaceae), recolectadas en Sucre, Venezuela. Rev Biol Trop59:585–595
Ratter JA, Ribeiro JF, Bridgwater S (1997) The Brazilian Cerrado vegetation and threats to its biodiversity.Ann Bot 80:223–230
Rodrigues GS, Paraıba LC, Moraes GJ (1996) Pairwise association as a criterion for the selection ofcollection sites of natural enemies of the cassava green mite, Mononychellus tanajoa (Bondar). SciAgric 53:2–3. doi:10.1590/S0103-90161996000200022
Rondon JB, Cumana-Campos LJ (2007) Revision taxonomica del genero Helicteres I. (Sterculiaceae) enVenezuela. Act Bot Venez 30:163–190
R Development Core Team (2006) R Foundation for Statistical Computing. Vienna, Austria. http://www.R-project.org
Tseng Y-H (1984) Mites associated with weeds, paddy rice and upland rice fields in Taiwan. In: GriffithsDA, Bowman CE (eds) Acarology VI. Ellis Howood, Chichester, pp 70–780
Wagner GJ, Wang E, Shepherd RW (2004) New approaches for studying and exploiting an old protuber-ance, the plant trichome. Ann Bot 93:3–11. doi:10.1093/aob/mch011
Exp Appl Acarol (2014) 64:309–319 319
123











![Coordinate changes in photosynthesis, sugar accumulation and antioxidative enzymes improve the performance of Jatropha curcas plants under drought stress [2012]](https://img.pdfslide.net/doc/110x75/632a95f9ebd37f1e740ae224/coordinate-changes-in-photosynthesis-sugar-accumulation-and-antioxidative-enzymes.jpg)

















