Embed Size (px)
Citation preview

Positionally-Dependent Chondrogenesis Induced by BMP4Is Co-Regulated by Sox9 and Msx2ICHIRO SEMBA, KAZUAKI NONAKA, ICHIRO TAKAHASHI, KATSU TAKAHASHI, RALPH DASHNER,LILLIAN SHUM, GLEN H. NUCKOLLS, AND HAROLD C. SLAVKIN*Craniofacial Development Section, National Institute of Arthritis and Musculoskeletal and Skin Diseases,National Institutes of Health, Bethesda, Maryland
ABSTRACT Cranial neural crest cells emi-grate from the posterior midbrain and anterior hind-brain to populate the first branchial arch and even-tually differentiate into multiple cell lineages in themaxilla and mandible during craniofacial morpho-genesis. In the developing mouse mandibular pro-cess, the expression profiles of BMP4, Msx2, Sox9,and type II collagen demonstrate temporally andspatially restrictive localization patterns suggestiveof their functions in the patterning and differentia-tion of cartilage. Under serumless culture condi-tions, beads soaked in BMP4 and implanted into em-bryonic day 10 (E10) mouse mandibular explantsinduced ectopic cartilage formation in the proximalposition of the explant. However, BMP4-soakedbeads implanted at the rostral position did not havean inductive effect. Ectopic chondrogenesis was as-sociated with the up-regulation of Sox9 and Msx2expression in the immediate vicinity of the BMP4beads 24 hours after implantation. Control beadshad no effect on cartilage induction or Msx2 andSox9 expression. Sox9 was induced at all sites ofBMP4 bead implantation. In contrast, Msx2 expres-sion was induced more intensely at the rostral posi-tion when compared with the proximal position, andsuggested that Msx2 expression was inhibitory tochondrogenesis. To test the hypothesis that over-ex-pression of Msx2 inhibits chondrogenesis, we ectopi-cally expressed Msx2 in the mandibular process or-gan culture system using adenovirus gene deliverystrategy. Microinjection of the Msx2-adenovirus tothe proximal position inhibited BMP4-induced chon-drogenesis. Over-expression of Msx2 also resulted inthe abrogation of endogenous cartilage and thedown-regulation of type II collagen expression.Taken together, these results suggest that BMP4 in-duces chondrogenesis, the pattern of which is posi-tively regulated by Sox9 and negatively by Msx2.Chondrogenesis only occurs at sites where Sox9 ex-pression is high relative to that of Msx2. The combi-natorial action of these transcription factors appearto establish a threshold for Sox9 function andthereby restricts the position of chondrogenesis. DevDyn 2000;217:401–414. Published 2000 Wiley-Liss, Inc.†
Key words: BMP4; Sox9; Msx2; type II collagen;Meckel’s cartilage; chondrogenesis;
mouse embryo; mandibular process;bead implantation; organ culture;whole-mount in situ hybridization;Alcian Blue staining; competitiveRT-PCR; adenovirus gene delivery
INTRODUCTION
Bone morphogenetic factor was used to describe theinduction of cartilage and bone formation when semi-purified bone matrix was introduced into the subecto-dermal and myogenic tissues of the adult rat and rabbit(Urist, 1965). Thereafter, these factors were character-ized (Wozney et al., 1988) and revealed more than 15related proteins belonging to the family of bone mor-phogenetic proteins (BMPs); which in turn belongs toan even larger group of growth and differentiation fac-tors, the transforming growth factor beta (TGFb) su-perfamily (see recent reviews Reddi, 1997; Vortkamp,1997; Urist, 1997; Wozney and Rosen, 1998). BMPssignal through a system of type I and II transmem-brane receptor serine/threonine kinases, which arethen modulated by cytoplasmic Smad proteins (see re-views Derynck et al., 1998; Kawabata et al., 1998). Inthe adult, elevated expression of BMP4 has been asso-ciated with the pathological condition afflicting theskeleton, fibrodysplasia ossificans progressiva (Shaf-ritz et al., 1996; Kaplan and Shore, 1998; Lanchoney etal., 1998). In the embryo, BMP2 and BMP4 have beenimplicated to function during skeletogenesis (see re-view Hall and Miyake, 1995), as well as in tissues inwhich epithelial-mesenchymal interactions mediatemorphogenesis and differentiation (Nuckolls et al.,1998).
Despite these significant contributions, the molecu-lar mechanisms of bone or cartilage induction by anymember of the family remain elusive, partly due tolimitations associated with using either in vivo systems
The first two authors contributed equally to this project and areconsidered co-first authors.
Grant sponsor: National Institutes of Health; Grant number: Z01-AR41114.
Ichiro Semba’s present address is Department of Oral Pathology,Kagoshima University Dental School, Kagoshima, 890-8544, Japan.
*Correspondence to: Dr. Harold C. Slavkin, Craniofacial Develop-ment Section, NIAMS, NIH, 6 Center Drive, Building 6, Room 324,MSC-2745, Bethesda, MD, 20892-2745.
Received 16 August 1999; Accepted 7 January 2000
DEVELOPMENTAL DYNAMICS 217:401–414 (2000)
Published 2000 WILEY-LISS, Inc. †This article is a US governmentwork and, as such, is in the public domain in the United States of America.

or serum-containing in vitro systems in which the in-teractive effects of many other growth and differentia-tive factors cannot be isolated from the direct actions ofBMP itself. Further, the functions of BMP2 or BMP4 inthe embryo cannot as yet be fully studied in geneticallyengineered mouse models because targeted disruptionof either molecule or their type I receptor resulted inearly embryonic lethality rendering the role of BMPsduring organogenesis, including skeletogenesis unin-formative (Mishina et al., 1995; Winnier et al., 1995;Zhang and Bradley, 1996). Finally, BMPs are a diversegroup of proteins with pleiotrophic functions in multi-ple differentiation programs and developmental stages(see recent reviews Hogan, 1996; Graff, 1997). Theprecise microenvironmental milieu regulating a spe-cific biological action of a specific BMP molecule is oftendifficult to identify.
Therefore, in order to address what are the positionaland temporal controls required for BMP-induced skel-etogenesis, and what specific molecular events are as-sociated with the developmental process, we employeda unique in vitro culture system in which embryonicmouse mandibular processes were explanted under se-rum-free conditions and subjected to a variety of mi-cromanipulations. We report that exogenous BMP4 de-livered by a protein-soaked bead implantation methodinduced ectopic cartilage formation, yet the extent ofchondrogenesis was dependent on the dose of BMP4administered, the developmental stage of the embry-onic tissue, and the position within the developingmandible. These temporal and positional tissue re-sponses were attributable to the induction and combi-natorial expression of two transcription factors, Sox9and Msx2. The combinatorial and antagonistic actionsof these molecules appeared to restrict ectopic cartilageformation during mandibular morphogenesis.
RESULTSExogenous BMP4 Induced Ectopic CartilageFormation in Embryonic Mandibular ProcessCultured Under Serumless Conditions
Mouse embryos at embryonic day 10 (E10) (Fig. 1a),corresponding to Theiler stage 18 with 40–44 somitepairs, are at a stage of development during which theanlagen for major organs are shaped and overt cyto-differentiation is initiated. The mandibular process ofthe developing first branchial arch was isolated bymicrodissection and explanted into an organ culturesystem supported by serumless, chemically-definedmedium. The mandibular process (Fig. 1b) consisted ofbilateral prominences of epithelium enclosing ectomes-enchymal cells, derivatives of cranial neural crest cellsthat emigrated from the posterior midbrain and ante-rior segments of the hindbrain (rhombomeres 1 and 2)(Lumsden et al., 1991; Sechrist et al., 1993; Osumi-Yamashita et al., 1994), and head mesoderm. The se-rumless and chemically-defined culture system facili-tates the investigation of time- and positional-dependent autocrine and paracrine growth anddifferentiation factors, without the numerous con-
Fig. 1. Embryonic mandibular process explant culture. Embryonicday 10 (E10) mouse embryo (a) was microdissected to isolate the man-dibular process (arrow) and placed in serumless, chemically-definedexplant culture. At the beginning of the culture period (b), the mandibularprocess consisted of two halves fused at the midline. Gross morphogen-esis and cytodifferentiation occurred by day 6 of culture (c). Meckel’scartilage was identified by Alcian blue staining (d). Protein-soaked beads(arrowheads) were placed within the developing tissue to deliver exoge-nous proteins (b, c, d). Exogenous BMP4 delivered by this methodinduced ectopic cartilage formation as revealed by Alcian blue staining(d). Scale bar for (a) 5 1mm, and for (b–d) 5 150mm.
402 SEMBA & NONAKA ET AL.

Fig. 2. BMP4-soaked beads induced ectopic cartilage formation onlywhen placed at the proximal position of the developing mandibular pro-cess. 100ng/ml BMP4 was delivered by bead implantation method intothe rostral (a, d) or proximal (b, e) positions and allowed to develop invitro for 6 days. Chondrogenesis was assayed by whole-mount Alcianblue staining (a, b, c) and toluidine blue staining on histological sections
d, e, f). Only beads implanted into the proximal position induced ectopiccartilage formation (b, e). PBS-soaked bead had no effect on chondro-genesis (c). The morphology of BMP4-induced cartilage (e) was similar toendogenous Meckel’s cartilage (f); arrowheads indicating perichondrium.All beads are circled. Rostral orientation is top of panel in (a–c). Scale barfor (a, b, c) 5 150mm, and for (d, e, f) 5 50mm.
Fig. 3. BMP4, Msx2, and Sox9 exhibited restrictive expression patterns. Whole-mount in situ hybridizationfor BMP4 (a–d), Msx2 (e–h) and Sox9 (i–l ) on 0 (a, e, i), 2 (b, f, j), 4 (c, g, k), and 6 days (d, h, i) of culture.Whole-mount Alcian blue staining demonstrated Meckel’s cartilage formation at 4 (m) and 6 days (n) ofculture. Rostral orientation is top for all panels. Scale bars for (a, e, i) and (b–d, f–h, j–n) 5 100mm.
403Sox9 and Msx2 CO-REGULATE BMP4-INDUCED CHONDROGENESIS

founding variables found in serum supplement and thematernal circulation. Growth and differentiation of themandibular process in vitro are delayed but not re-tarded when compared with in vivo development(Slavkin et al., 1989). Further, the explant system sup-ports micromanipulations such as bead implantationand the microinjections used in this study.
Beads soaked in BMP4 were positioned efficientlyand reproducibly in the mandibular process at the be-ginning of culture (Fig. 1b) and the biological effectsobserved over a period of six days. By six days inculture (Slavkin et al., 1999; Fig. 1c), differentiationand the formation of cartilage, osteoid, incisor and mo-lar tooth buds and tongue were observed. Whole-mountAlcian blue staining demonstrated the formation ofMeckel’s cartilage which was composed of a rostraltriangular-shaped piece, bilateral rod-shaped piecesand proximal pieces representing the future middle earossicles (Fig. 1d). Beads soaked in 100ng/ml BMP4 con-sistently induced ectopic cartilage formation (Fig. 1d).Ectopic cartilage was observed to fuse with endogenouscartilage formation patterns resulting in expansion orbifurcation morphology, or as observed as isolated ec-topic cartilage pieces.
Ectopic Chondrogenesis Induced by BMP4 WasPositional-, Temporal-, and Dose-Dependent
Further experiments demonstrated that beadssoaked in 100ng/ml BMP4 only induced ectopic carti-lage formation when the bead was placed in the prox-imal position of the developing mandibular processwhen compared with the rostral position (Fig. 2a, b;Table 1). Control beads soaked in PBS (Fig. 2c), orbeads soaked in 100ng/ml FGF8 (data not shown), didnot induce ectopic chondrogenesis or disrupt endoge-nous cartilage formation. Because Alcian blue stainingdemonstrated positive reaction to sulfated proteogly-can suggestive of cartilage, we additionally performedroutine histological evaluation on frozen sections of thedeveloping mandiblular explants at the site of beadimplantation. Consistent with the whole-mount Alcianblue staining results, cartilage was observed envelop-ing beads implanted at the proximal position (Fig. 2e),whereas cartilage was not detectable surroundingbeads implanted at the rostral position (Fig. 2d). Car-tilage induced by BMP4 has morphological and histo-logical characteristics similar to that of normal carti-lage (Fig. 2f), including the presence of an intactperichondrium and mature chondrocytes residingwithin lacunae of the cartilagenous matrix. No signif-
icant apoptosis was detected associated with bead im-plantation.
The incidence of ectopic chondrogenesis was directlydependent on the concentration of BMP4 used in theseinvestigations (Table 1). At 10ng/ml BMP4, no ectopiccartilage was induced, whereas at 100ng/ml the inci-dence was 83.3% when beads were implanted at theproximal position.
BMP4 Induced Different Levels of Sox9 andMsx2 Expression Dependent on PositionalInformation
To explain positional dependency of BMP4 inducedchondrogenesis, whole-mount in situ hybridizations,using several gene probes at various time points, wasperformed to identify candidate molecules associatedwith the phenomenon. Mandibular processes at thebeginning of culture, and after 2, 4, or 6 days of devel-opment were fixed and processed. Endogenous BMP4transcripts were detected in the E10 mandibular pro-cess, with a higher level of expression on the medialaspect when compared with the lateral (Fig. 3a); con-sistent with previous reports (Bennett et al., 1995;Neubuser et al., 1997). Thereafter, BMP4 exhibitedcomplex localization patterns (Figure 3b–d), presum-ably due to its functions in multiple cell lineages withinthe developing mandibular process—chondrogenic andodontogenic. The localization pattern of Msx2 in themandibular process in culture in part overlapped withthat of BMP4 expression especially in the rostral area,yet was more restrictive (Fig. 3e–h). At day 2 of cul-ture, Msx2 was expressed in rostral-medial and bilat-erally symmetrical proximal-lateral sites, which ap-peared to delineate the rostral and caudal extent offuture Meckel’s cartilage formation (Fig. 3f). Further,at day 4 of culture Msx2 expression was observed alonga boundary interface with sites destined to becomecartilage (Fig. 3g).
Sox9 has been shown to be a direct transcriptionalactivator of the cartilage-specific type II collagen gene(Lefebvre et al., 1997). The localization of endogenousSox9 in the developing mandibular process was associ-ated with Meckel’s cartilage development (Figure 3i–l).At day 2 of culture, prior to cartilage differentiation,Sox9 was localized in a rostral triangular-shaped site,bilateral sigmoidal-shaped sites and at the proximallateral margins of the explants (Fig. 3j). This patternresembled the Alcian blue staining pattern of Meckel’scartilage at day 6 of culture (Fig. 3n), and presumablyserved as a morphogenetic template for this cartilage.Sox9 expression was somewhat decreased but still detect-able at day 4 and 6 of culture (Fig. 3k, l) at which timescartilage differentiation was observed (Fig. 3m, n).
Because the endogenous expression pattern of Msx2overlapped with that of BMP4, and that it also ap-peared to define boundary interfaces suggestive of apatterning role during chondrogenesis, the expressionof Msx2 in mandibular processes implanted withBMP4-soaked beads was examined after 24 hours ofculture (Fig. 4a, b). Consistent with the responses of
TABLE 1. Frequency of Ectopic Cartilage Induced byBMP4-Soaked Beads in Mandibular Processes in
Explant Cultures
Concentration ofBMP4 (ng/ml) 10 50 100Rostral 0/15 (0%) 0/15 (0%) 0/18 (0%)Proximal 0/15 (0%) 6/15 (40%) 15/18 (83.3%)
404 SEMBA & NONAKA ET AL.

many other developmental stages and tissues, BMP4induced Msx2 expression in the immediate vicinity ofthe source. However, the level of induced Msx2 expres-sion appeared to be higher when the BMP4-soakedbead was implanted in the rostral as compared withthat in the proximal position. This expression of Msx2in response to BMP4 was transient and down-regu-lated in the following 24 hours (Fig. 4c, d).
Because Sox9 is a direct transcription activator ofthe type II collagen gene, we examined its expression inresponse to exogenous BMP4. One day after bead im-plantation, Sox9 expression was induced surroundingthe BMP4-soaked bead (Fig. 4e, f ). Contrary to Msx2induced expression, the amount of Sox9 was higheraround the BMP4-soaked bead implanted at the prox-imal as compared with the rostral positions. Similar tothe Msx2 response, induced Sox9 expression was de-creased by day 2 of explant culture (Fig. 4g, h).
The expression of type II collagen, presumably me-diated by the activation of Sox9, was detected sur-rounding the BMP4-soaked bead at 48 hours of culture;when Sox9 expression was already down-regulated(Fig. 4j). This observation is consistent with previousreports of Sox9 expression preceding that of type IIcollagen (Ng et al., 1997; Takahashi et al., 1998a). Theexpression of type II collagen at 24 and 48 hoursshowed a pattern that was predictable of the differen-tiated phenotype as detected by Alcian blue staining onday 4. Type II collagen expression was detected aroundthe developing Meckel’s cartilage. In addition, expres-sion was also detected around the BMP4-soaked beadimplanted at the proximal position, resulting in anexpansion of the domain (Fig. 4j, l). Type II collagenexpression, though detected surrounding the BMP4-soaked bead implanted at the rostral position at 24hours of culture (Fig. 4i), was observed to be down-regulated to non-existence by 48 hours (Fig. 4k). Thischange in expression level was repeatedly consistentwith the phenotypic outcome at day 6, when Alcianblue staining did not detect any ectopic chondrogenesiswhen BMP4-soaked beads were introduced at this po-sition.
The molecular responsiveness of the tissue to BMP4was restricted to the first 24 hours of culture. BMP4-soaked beads implanted into the mandibular processafter the explants had been in culture for one dayshowed no induction of Msx2 nor Sox9, and no ectopiccartilage (data not shown). Taken together, these datademonstrate that tissue response to exogenous BMP4is positionally and temporally dependent.
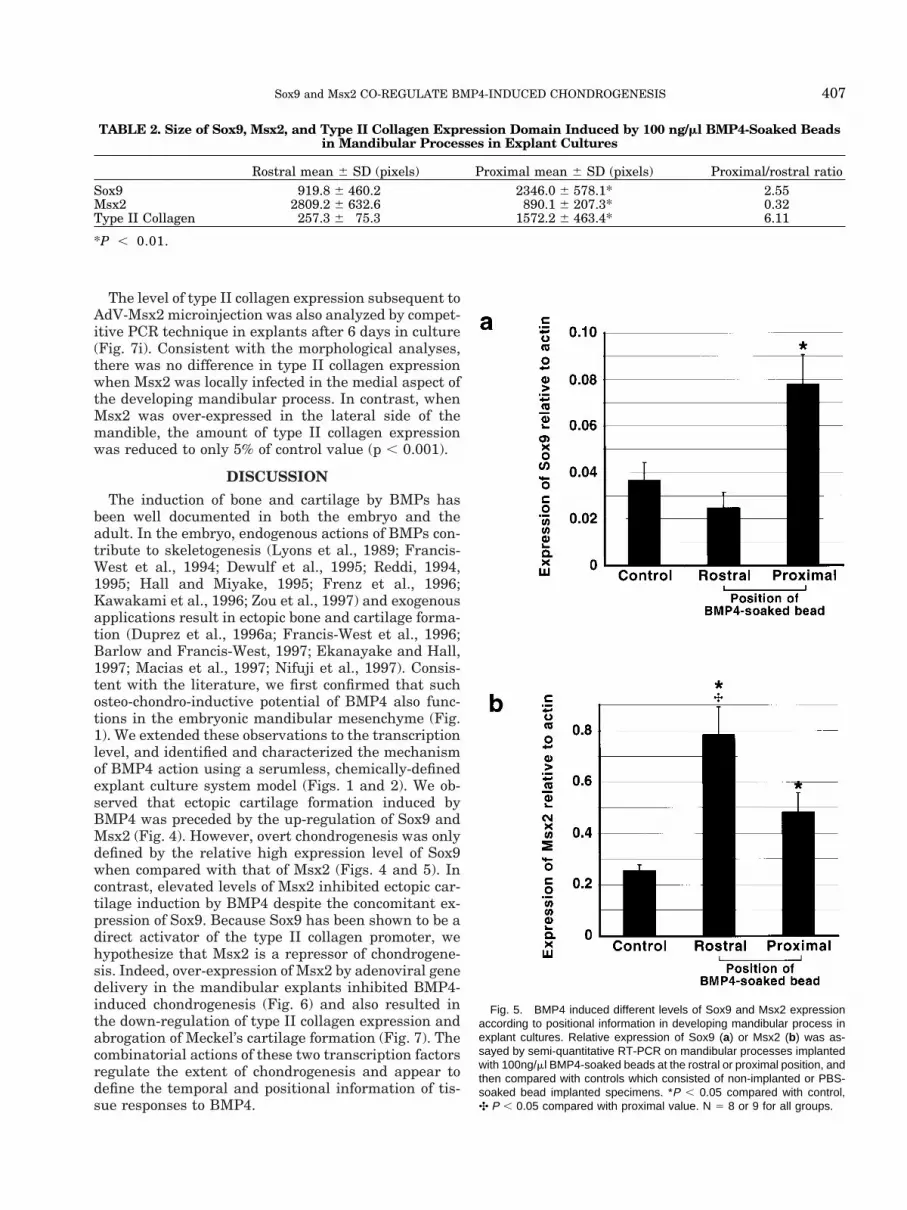
The observed difference in the expression of BMP4inducible genes was further analyzed using quanti-tated morphometric (Table 2) and semi-quantitativeRT-PCR approaches (Fig. 5). First, to minimize varia-tions in developmental stage and thereby responsive-ness among different E10 explants, two beads wereimplanted into the same piece of tissue; one in each ofthe rostral and proximal positions, respectively. Con-sistent with the single bead implantation experiments,the rostral position demonstrated a more intense re-
sponse to Msx2 (Fig. 4m), and a weaker signal for Sox9(Fig. 4n), when compared with the proximal position.Type II collagen expression was detected at both sitesof implantation (Fig. 4o). Control beads soaked in PBShad no effect on gene expression level or pattern ofeither Msx2 (Fig. 4p), Sox9 (Fig. 4q) or type II collagen(Fig. 4r). Subsequently, explants processed for whole-mount in situ hybridization were subjected to morpho-metric analyses in which the area of induced geneexpression was converted to the number of pixels pos-itive for the hybridization signal (Table 2). With overnine explants analyzed for each gene pattern, thequantitative analysis was consistent with the qualita-tive analysis. Expression of Sox9 surrounding theBMP4-bead implanted at the proximal position was 2.6times higher than that at the rostral position. In con-trast, Msx2 expression was three-fold weaker, compar-ing the corresponding positions. Expression of type IIcollagen was 6.1-fold higher surrounding the proxi-mally placed BMP4-beads than those placed rostrally.The differences in size of induced Sox9, Msx2, and typeII collagen expression between the rostral and proxi-mal positions were statistically significant (P , 0.01).
Semi-quantitative RT-PCR results also supportedpositionally-dependent differential levels of inductionof Sox9 and Msx2 by BMP4. Consistent with data fromboth whole-mount in situ hybridization and morpho-metric analyses, a three-fold elevation of Sox9 inducedby BMP4 was detectable when beads were implantedproximally as compared with beads implanted ros-trally, or controls using PBS-soaked beads or no beads(Fig. 5a). We could not detect a significant increase inSox9 expression induced by BMP4-soaked beads placedat the rostral position, possibly due to the high endog-enous level of Sox9 expression marking the morphoge-netic template of Meckel’s cartilage. Significant induc-tion of Msx2 expression by BMP4 was also detected atboth rostrally or proximally implanted beads; morethan three-fold and almost two-fold, respectively, whencompared with controls (Fig. 5b). The difference be-tween rostral and proximal expression levels was sta-tistically significant (P , 0.05).
Direct Over-Expression of Msx2 InhibitedChondrogenesis
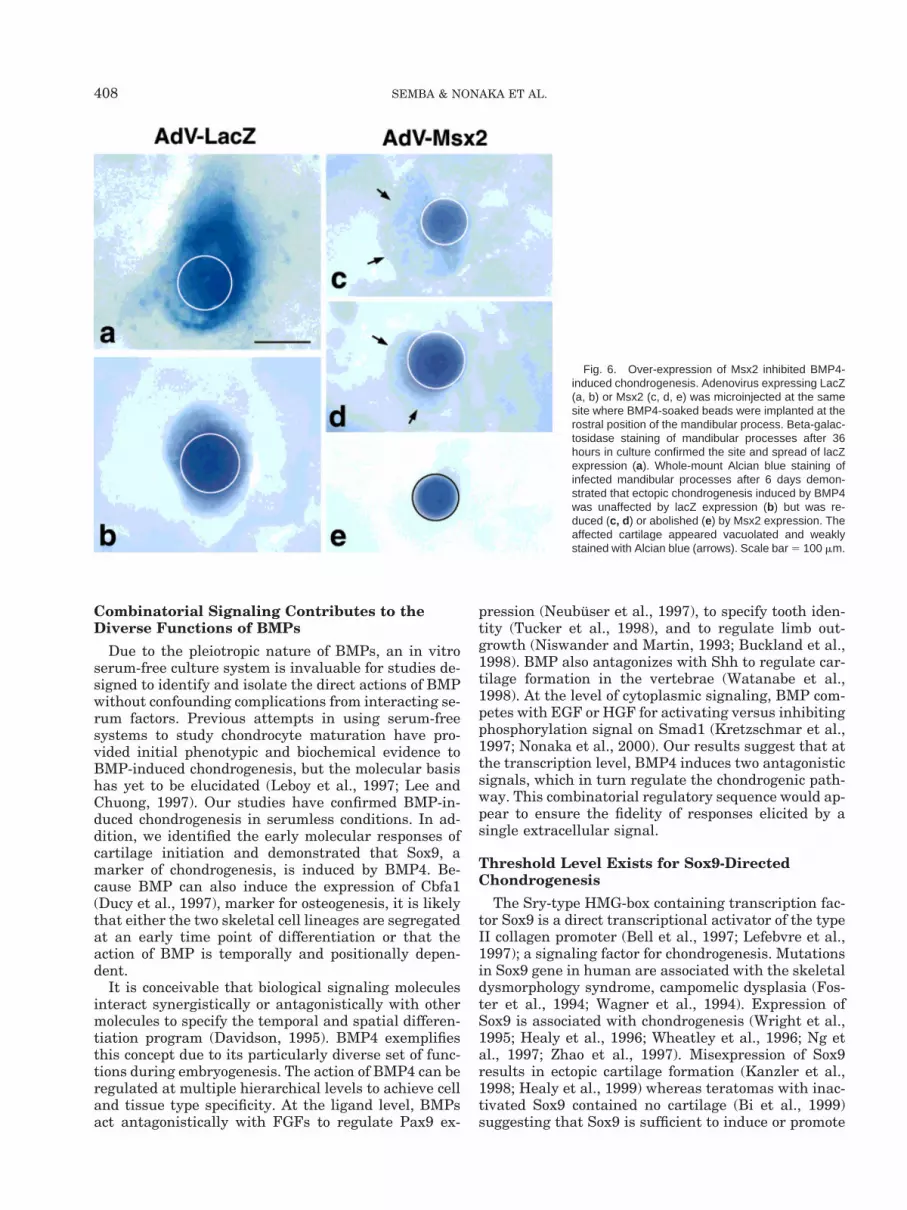
Sox9 expression preceded that of type II collagen, yettype II collagen expression was not maintained aroundthe BMP4-soaked bead implanted at the rostral posi-tion, resulting in no ectopic cartilage. This could be dueto the expression of Msx2, which was much moreheavily detected at the rostral rather than the proxi-mal position. To test the hypothesis that Msx2 inhibitsBMP4-induced chondrogenesis, a recombinant adeno-virus was engineered to over-express the Msx2 gene(AdV-Msx2). The adenovirus was microinjected intothe developing mandibular process and its actions eval-uated against explants microinjected with control ade-novirus bearing LacZ expression (AdV-LacZ). Consis-tent with previous reports (Takahashi et al., 1998b),microinjection of AdV-LacZ resulted in high levels of
405Sox9 and Msx2 CO-REGULATE BMP4-INDUCED CHONDROGENESIS

expression detectable 36 hours subsequent to injection(Fig. 6a). AdV-LacZ did not affect BMP4-induced ec-topic cartilage formation in the proximal position of themandibular process (Fig. 6b). When AdV-Msx2 wasmicroinjected at the same site where BMP4-soakedbead was implanted, ectopic cartilage formation wasreduced (Fig. 6c, d) or abolished (Fig. 6e). The cartilagematrix was weakly stained with Alcian blue and ap-peared vacuolated.
In addition, we also tested whether over-expressionof Msx2 inhibits endogenous cartilage formation. Therecombinant adenovirus was injected into the medialor lateral side of the developing mandibular process.Similarly, high expression of lacZ was detectable at 36
hours subsequent to injection (Fig. 7a, b). AdV-LacZinjected explants had no deleterious effects and Meck-el’s cartilage developed normally (Fig. 7c, d). In con-trast, AdV-Msx2 injected into the lateral side of themandibular process resulted in an abrogation of Meck-el’s cartilage formation, whereas neither AdV-Msx2 in-jected into the medial part of the explant, nor theuninjected side produced Meckel’s cartilage dysmor-phogenesis. Histological analyses confirmed that nor-mal cartilage formed in the control non-injected side(Fig. 7g), yet no evidence of cartilage was observed atthe AdV-Msx2 injected side of the explant (Fig. 7h). Nosignificant apoptosis was detected in adenovirus in-jected specimens.
Fig. 4. BMP4 induced Sox9 and Msx2 expression in developingmandibular process in explant cultures. Beads soaked in 100ng/ml BMP4were implanted into the rostral (a, c, e, g, i, k) or proximal (b, d, f, h, j, l)positions of mandibular processes and explanted into cultures. Whole-mount in situ hybridization was performed for Msx2 (a–d), Sox9 (e–h),and type II collagen (i–l), after 24 (a, b, e, f, i, j), or 48 (c, d, g, h, k, l) hoursin culture. The induced expression of Msx2 and Sox9 was detected after24 hours, whereas the inductive effect was down-regulated after 48 hoursin culture. Sox9 expression appeared higher surrounding the BMP4-soaked bead placed at the proximal (f) than that at the rostral (e) position,whereas this response appeared to be reverse for that of Msx2. Type II
collagen was only observed surrounding BMP4-soaked beads implantedat the proximal postition (j). In order to demonstrate that positional-dependent differential gene induction was not due to differences amongindividual mandibles, multiple BMP4-soaked beads were implanted atvarious positions within the same mandible and whole-mount in situhybridization was performed for Msx2 (m), Sox9 (n) and type II collagen(o) after 1 day in culture. The results were similar to that of the singlebead implantation experiments. Control PBS-soaked beads implanted atany position did not induce Msx2 (p), Sox9 (q) or type II collagen (r) after24 hours in culture. All beads are circled. Rostral orientation is top for allpanels. Scale bars 5 100mm.
406 SEMBA & NONAKA ET AL.
The level of type II collagen expression subsequent toAdV-Msx2 microinjection was also analyzed by compet-itive PCR technique in explants after 6 days in culture(Fig. 7i). Consistent with the morphological analyses,there was no difference in type II collagen expressionwhen Msx2 was locally infected in the medial aspect ofthe developing mandibular process. In contrast, whenMsx2 was over-expressed in the lateral side of themandible, the amount of type II collagen expressionwas reduced to only 5% of control value (p , 0.001).
DISCUSSION
The induction of bone and cartilage by BMPs hasbeen well documented in both the embryo and theadult. In the embryo, endogenous actions of BMPs con-tribute to skeletogenesis (Lyons et al., 1989; Francis-West et al., 1994; Dewulf et al., 1995; Reddi, 1994,1995; Hall and Miyake, 1995; Frenz et al., 1996;Kawakami et al., 1996; Zou et al., 1997) and exogenousapplications result in ectopic bone and cartilage forma-tion (Duprez et al., 1996a; Francis-West et al., 1996;Barlow and Francis-West, 1997; Ekanayake and Hall,1997; Macias et al., 1997; Nifuji et al., 1997). Consis-tent with the literature, we first confirmed that suchosteo-chondro-inductive potential of BMP4 also func-tions in the embryonic mandibular mesenchyme (Fig.1). We extended these observations to the transcriptionlevel, and identified and characterized the mechanismof BMP4 action using a serumless, chemically-definedexplant culture system model (Figs. 1 and 2). We ob-served that ectopic cartilage formation induced byBMP4 was preceded by the up-regulation of Sox9 andMsx2 (Fig. 4). However, overt chondrogenesis was onlydefined by the relative high expression level of Sox9when compared with that of Msx2 (Figs. 4 and 5). Incontrast, elevated levels of Msx2 inhibited ectopic car-tilage induction by BMP4 despite the concomitant ex-pression of Sox9. Because Sox9 has been shown to be adirect activator of the type II collagen promoter, wehypothesize that Msx2 is a repressor of chondrogene-sis. Indeed, over-expression of Msx2 by adenoviral genedelivery in the mandibular explants inhibited BMP4-induced chondrogenesis (Fig. 6) and also resulted inthe down-regulation of type II collagen expression andabrogation of Meckel’s cartilage formation (Fig. 7). Thecombinatorial actions of these two transcription factorsregulate the extent of chondrogenesis and appear todefine the temporal and positional information of tis-sue responses to BMP4.
Fig. 5. BMP4 induced different levels of Sox9 and Msx2 expressionaccording to positional information in developing mandibular process inexplant cultures. Relative expression of Sox9 (a) or Msx2 (b) was as-sayed by semi-quantitative RT-PCR on mandibular processes implantedwith 100ng/ml BMP4-soaked beads at the rostral or proximal position, andthen compared with controls which consisted of non-implanted or PBS-soaked bead implanted specimens. *P , 0.05 compared with control,✣ P , 0.05 compared with proximal value. N 5 8 or 9 for all groups.
TABLE 2. Size of Sox9, Msx2, and Type II Collagen Expression Domain Induced by 100 ng/ml BMP4-Soaked Beadsin Mandibular Processes in Explant Cultures
Rostral mean 6 SD (pixels) Proximal mean 6 SD (pixels) Proximal/rostral ratioSox9 919.8 6 460.2 2346.0 6 578.1* 2.55Msx2 2809.2 6 632.6 890.1 6 207.3* 0.32Type II Collagen 257.3 6 75.3 1572.2 6 463.4* 6.11
*P , 0.01.
407Sox9 and Msx2 CO-REGULATE BMP4-INDUCED CHONDROGENESIS
Combinatorial Signaling Contributes to theDiverse Functions of BMPs
Due to the pleiotropic nature of BMPs, an in vitroserum-free culture system is invaluable for studies de-signed to identify and isolate the direct actions of BMPwithout confounding complications from interacting se-rum factors. Previous attempts in using serum-freesystems to study chondrocyte maturation have pro-vided initial phenotypic and biochemical evidence toBMP-induced chondrogenesis, but the molecular basishas yet to be elucidated (Leboy et al., 1997; Lee andChuong, 1997). Our studies have confirmed BMP-in-duced chondrogenesis in serumless conditions. In ad-dition, we identified the early molecular responses ofcartilage initiation and demonstrated that Sox9, amarker of chondrogenesis, is induced by BMP4. Be-cause BMP can also induce the expression of Cbfa1(Ducy et al., 1997), marker for osteogenesis, it is likelythat either the two skeletal cell lineages are segregatedat an early time point of differentiation or that theaction of BMP is temporally and positionally depen-dent.
It is conceivable that biological signaling moleculesinteract synergistically or antagonistically with othermolecules to specify the temporal and spatial differen-tiation program (Davidson, 1995). BMP4 exemplifiesthis concept due to its particularly diverse set of func-tions during embryogenesis. The action of BMP4 can beregulated at multiple hierarchical levels to achieve celland tissue type specificity. At the ligand level, BMPsact antagonistically with FGFs to regulate Pax9 ex-
pression (Neubuser et al., 1997), to specify tooth iden-tity (Tucker et al., 1998), and to regulate limb out-growth (Niswander and Martin, 1993; Buckland et al.,1998). BMP also antagonizes with Shh to regulate car-tilage formation in the vertebrae (Watanabe et al.,1998). At the level of cytoplasmic signaling, BMP com-petes with EGF or HGF for activating versus inhibitingphosphorylation signal on Smad1 (Kretzschmar et al.,1997; Nonaka et al., 2000). Our results suggest that atthe transcription level, BMP4 induces two antagonisticsignals, which in turn regulate the chondrogenic path-way. This combinatorial regulatory sequence would ap-pear to ensure the fidelity of responses elicited by asingle extracellular signal.
Threshold Level Exists for Sox9-DirectedChondrogenesis
The Sry-type HMG-box containing transcription fac-tor Sox9 is a direct transcriptional activator of the typeII collagen promoter (Bell et al., 1997; Lefebvre et al.,1997); a signaling factor for chondrogenesis. Mutationsin Sox9 gene in human are associated with the skeletaldysmorphology syndrome, campomelic dysplasia (Fos-ter et al., 1994; Wagner et al., 1994). Expression ofSox9 is associated with chondrogenesis (Wright et al.,1995; Healy et al., 1996; Wheatley et al., 1996; Ng etal., 1997; Zhao et al., 1997). Misexpression of Sox9results in ectopic cartilage formation (Kanzler et al.,1998; Healy et al., 1999) whereas teratomas with inac-tivated Sox9 contained no cartilage (Bi et al., 1999)suggesting that Sox9 is sufficient to induce or promote
Fig. 6. Over-expression of Msx2 inhibited BMP4-induced chondrogenesis. Adenovirus expressing LacZ(a, b) or Msx2 (c, d, e) was microinjected at the samesite where BMP4-soaked beads were implanted at therostral position of the mandibular process. Beta-galac-tosidase staining of mandibular processes after 36hours in culture confirmed the site and spread of lacZexpression (a). Whole-mount Alcian blue staining ofinfected mandibular processes after 6 days demon-strated that ectopic chondrogenesis induced by BMP4was unaffected by lacZ expression (b) but was re-duced (c, d ) or abolished (e) by Msx2 expression. Theaffected cartilage appeared vacuolated and weaklystained with Alcian blue (arrows). Scale bar 5 100 mm.
408 SEMBA & NONAKA ET AL.

chondrogenesis. It has also been reported that Sox9may bind and activate the promoters of two other car-tilage markers, aggrecan and type X collagen (Haradaet al., 1996; Sekiya et al., 1997). Despite evidence sup-porting the critical role of Sox9 during early chondro-genesis, further upstream regulatory elements of thispathway have been elusive.
It has been implicated that the level of Sox9 ex-pression is significant in determining whether chon-drogenesis is to occur or not. High levels of Sox9 areobserved to correlate with all sites of cartilage for-mation. Low levels of Sox9 are detected in a broadarray of non-chondrogenic embryonic tissues such asnotochord, neural tube, heart and lung (Ng et al.,1997; Zhao et al., 1997). It is yet unclear whether an
absolute threshold level of Sox9 expression is re-quired to induce chondrogenesis, nor how this level isdetermined and controlled. Our data suggests that arelative threshold level may exist. The expression ofSox9 precedes that of type II collagen expression, anindication that the responding mesenchymal cellscan and have initiated the chondrogenic programregardless of the absolute expression level of Sox9.However, overt chondrogenesis only occurs at siteswhere Sox9 expression is high relative to that of therepressor Msx2. Therefore, the antagonism betweenSox9 and Msx2 appears to establish a relativethreshold for Sox9 function in cartilage induction,and that this relationship is site specific. Thisthreshold may also be modulated positively or nega-
Fig. 7. Over-expression of Msx2 in developing mandibular processesinhibited Meckel’s cartilage formation. Adenovirus-expressing LacZ (a–d)or Msx2 (e, f) was microinjected to the medial (a, c, e) or lateral portions(b, d, f) of the right half of the E10 mandibular process as shown in thefigure. Beta-galactosidase staining of mandibular processes after 36hours in culture confirmed restrictive gene expression delivered by ade-novirus microinjection and infection method (a, b). Whole-mount Alcianblue staining of infected mandibular processes after 6 days demonstratedno effect on Meckel’s cartilage formation by LacZ expression (c, d ) orMsx2 expression to the medial portion (e). Expression of Msx2 at thelateral portion of the mandibular process inhibited Meckel’s cartilage
formation (f). Toluidine blue staining of histological sections after 6 daysrevealed Meckel’s cartilage (outlined in white) on the uninjected side (g)but absence of chondrogenesis or excessive apoptosis on the Msx2expressing side (h). Scale bars for (a–f) 5 100mm, and for (g, h) 5 50mm.Expression level of type II collagen was assayed by quantitative RT-PCRin mandibular processes infected with adenovirus expressing LacZ(hatched bars) or Msx2 (solid bars) at the medial or lateral portion (i).Consistent with morphological analyses, Msx2 expression at the lateralportion of the mandibular process inhibited type II collagen expression.*P , 0.001. N 5 6.
409Sox9 and Msx2 CO-REGULATE BMP4-INDUCED CHONDROGENESIS

tively by the presence of other transcription factorssuch as other Sox proteins (Lefebvre et al., 1998).
Msx2 Is a Repressor of Chondrogenesis
The Msx family of transcription factors is multifunc-tional, although very little is known of Msx3. Msx1 andMsx2 are largely co-expressed in the developing em-bryo, and are thought to be redundant in functions(Davidson, 1995; Maas et al., 1996). Disruption of Msxfunctions results in multiple malformations includingthat of the craniofacial skeleton (Satokata and Maas,1994; Liu et al., 1995; Foerst-Potts and Sadler, 1997;Winograd et al., 1997). In many cases, Msx genes me-diate the function of BMPs including cranial suturemorphogenesis (Kim et al., 1998), apoptosis (Marazzi etal., 1997; Takahashi et al., 1998b), tooth formation(Tureckova et al., 1995; Jernvall et al., 1998), dorsal-ization of midline structures along the neural tube(Takahashi et al., 1996), vertebral development (Mon-soro-Burq et al., 1996; Watanabe and Le Douarin,1996), and mammary gland development (Phippard etal., 1996). Further, Msx1 and Msx2 are considered tobe general repressors of transcription (Catron et al.,1995; Semenza et al., 1995), directly modulating thefunction of the promoter for osteocalcin (Newberry etal., 1997), type I collagen (Dodig et al., 1996), andMyoD enhancer (Woloshin et al., 1995).
Consistent with a repressor function of Msx, ourinvestigations have provided direct evidence that ex-pression of Msx2 is inhibitory in the chondrogenicpathway; including both endogenous as well as ectopiccartilage formation. This is supported by previous re-ports in which application of antisense oligonucleotidesdirected against Msx2 resulted in an increase in carti-lage formation in the avian mandible (Mina et al.,1996). The repressor action of Msx2 is likely to becomplex. For example, it is unclear how Msx2 over-expression eliminated endogenous cartilage from thelateral side of the mandibular process and not themedial side. We speculate that because the medialportion is a site of high endogenous expression of Msx1and Msx2, our adenovirus-driven ectopic expressionmight not have achieved an additionally increasedlevel. The endogenous ratio of Sox9 to Msx2 would thenalready determine the outcome of chondrogenesis. It isalso possible that chondroprogenitor cells at the medialposition are less committed in phenotype and less re-sponsive to Msx2 than those at the lateral. The lateralpart of Meckel’s cartilage was formed by day 4 of cul-ture (Fig. 4m), whereas the medial portion was Alcianblue positive only by day 6 (Fig. 4n). Because adenovi-rus expression in our system is maintained for 48–72hours, there is the potential for compensatory growthwhen expression level declines. In addition, other re-sults indicate that Msx1 or Msx2 can interact withother transcription factors and compete for DNA bind-ing (Catron et al., 1995; Zhang et al., 1996; Wu et al.,1997; Zhang et al., 1997; Bendall et al., 1998). Theseadditional regulatory networks may be different lo-cally. Taken together, these results suggest that the
regulation of a single extracellular signaling moleculeby combinations of transcription factors would appearto be required to achieve a specific response in a specificdevelopmental context.
The Developing Mandibular Process IsCompartmentalized
Compartmentalization of the developing mandibularmesenchyme is associated with early complex molecu-lar patterning. The cranial to caudal and proximal torostral axial plans are established and maintained bythe expression of a sequence of homeobox-containinggenes. The oral to aboral axis along the maxillary andmandibular processes of the first branchial arch is as-sociated with the expression of Lim-domain homeopro-teins, Lhx6 and Lhx7 (Grigoriou et al., 1998). The oralmesenchyme is thus competent to respond to epithelialodontogenic signals such as FGFs. These axes are fur-ther refined by homeobox containing transcription fac-tors of the Msx and Dlx families, and may specify theformation of incisor versus molar teeth within the firstbranchial arch (Qiu et al., 1997; Thomas et al., 1997).Other domains have been suggested to limit the man-ifestation of other phenotypes, such as bone, cartilageand muscle within the embryonic craniofacial mesen-chyme. We propose that further compartmentalizationalong the rostral-proximal and medial-lateral axes ex-ists that are defined by the chondrogenic potential inthe forming mandible. This assertion is supported byexperiments in which meticulous microdissections fol-lowed by grafting, tissue recombination or micromasscultures of different segments of the developing man-dible, identified varying potentials to form cartilage(Hall, 1982; Langille, 1994). Similar positional infor-mation also exists in the developing limb bud and hasbeen defined by the proximodistal, dorsoventral, andanteroposterior axes (Cottrill et al., 1987; Ros et al.,1997). Although not assayed in this study, it is likelythat BMP2, BMP7, and Msx1 may also be involved indeveloping mandibular cartilage based on their se-quence homologies and overlapping expression patternwith BMP4 and Msx2, respectively. The specificity ofthese families of growth and transcription factors to-wards chondrogenesis versus odontogenesis remains tobe characterized.
During early development, progressively restrictivemolecular patterning emerges from a sequence of tran-scriptional activations and inactivations. Our resultssuggest that within the presumptive myogenic regiondestined to be the tongue, chondrogenic potential existsand that mesenchymal cells can be induced to becomechondrocytes in response to BMP4. The induction isdose-dependent on exogenous BMP4, and suggests thatthe endogenous developmental program consists of si-lencers to the chondrogenic pathway in the tuberculumimpar. Indeed, the induction of cartilage by BMP maybe the result of simultaneous regulation of two path-ways; stimulation of chondrogenesis and inhibition ofmyogenesis. In chick limb bud micromass cultures,BMP2 specifically promotes chondrogenesis presum-
410 SEMBA & NONAKA ET AL.

ably at the expense of myogenesis (Duprez et al.,1996b). In the developing somite, BMP signaling inhib-its the activation of MyoD and Myf5 (Reshef et al.,1998). Further, in vitro studies support that endoge-nous Msx1 can bind to and repress the MyoD enhancer(Woloshin et al, 1995). These findings and our resultsindicate that BMP is instructive to both chondrogene-sis as well as myogenesis in undifferentiated mesen-chymal cells.
CONCLUSION
In summary, the evidence supports the concept thatthe emergence of the chondrogenic potential of BMP4from the pleiotropic nature of this growth factor ismediated by the induced expression of Sox9 and Msx2.Sox9 and Msx2 function as positive and negative reg-ulators of chondrogenesis, respectively. The combina-torial action of these transcription factors appears todefine the extent of embryonic mouse mandibularchondrogenesis.
EXPERIMENTAL PROCEDURESMouse Embryo and Mandibular ProcessExplant Culture
Timed pregnant Swiss Webster mice were obtained(Harlan Bioproducts for Science, Indianapolis, IN) andembryos were collected at embryonic day 10 (E10) orE12. Isolated embryos were further staged by externalmorphology and number of somite pairs according toTheiler staging system (Theiler, 1989). Organ cultureof mandibular processes from E10 embryos was per-formed according to previously reported protocols(Shum et al., 1993; Slavkin et al., 1999). Briefly, themandibular processes were isolated by microdissectionand explanted into a Trowell-type organ culture systemusing serumless, chemically-defined BGJb medium(Life Technologies Inc., Gaithersburg, MA) supple-mented with 100 mg/ml ascorbic acid and 100 U each ofpenicillin and streptomycin, and allowed to develop at37°C and 5% CO2.
Bead Implantation
Affi-Gel blue agarose beads (Bio-Rad Labs., Her-cules, CA) at 100–200 mesh, which correspond to adiameter of 50–75 mm were selected, washed twice inphosphate buffered saline (PBS), and soaked in 10, 50,or 100 ng/ml human recombinant BMP4 (Genetics In-stitute Inc., Cambridge, MA), or PBS as control for 1 hrat room temperature. BMP4-soaked or PBS controlbeads were positioned by fine forceps and implantedinto the mandibular process explants using a mouth-controlled micropipette under the stereomicroscope(Slavkin., et al. 1999).
Whole-Mount Alcian Blue Staining
The presence of sulfated proteoglycans indicative ofcartilage formation was detected by Alcian Blue stain-ing as previously described (Shum et al., 1993), using0.04% Alcian Blue 8GX (Sigma, St. Louis, MO) in acid
ethanol followed by tissue clearing with a graded seriesof potassium hydroxide and glycerol.
Histology
Specimens were fixed in 4% paraformaldehyde in 0.1M PBS (pH 7.4) for 12 hr at 4°C. Subsequently, speci-mens were rinsed thoroughly in 0.01 M PBS, infiltratedthrough a graded series of sucrose up to 30% and fi-nally in 10% acrylamide and 30% sucrose in 0.01 MPBS, embedded first in 10% acrylamide and then inTissue Freezing Medium (Triangle Biomedical Sci-ences, Durham, NC). Seven micron-thick frozen sec-tions were collected and stained with toluidine blue.
Whole-Mount In Situ Hybridization
Probes for BMP4 (GenBank X56848: 776-1248nt),Sox9 (Wright et al., 1995, 926-1683nt), and type IIcollagen (Metsaranta et al., 1991, GenBank M65161:29648-31343nt) were obtained by RT-PCR method andconfirmed by direct sequencing. Probe for Msx2 was anAluI to EcoRI fragment of the full length Msx2 cDNA(Bell et al., 1993, GenBank L11739: 251-708nt). Digoxi-genin (DIG)-labeled sense and antisense riboprobes formouse BMP4, Sox9, Msx2, and type II collagen wereprepared by in vitro transcription of linearized pBlue-Script (Stratagene, La Jolla, CA) or pCRII (InvitrogenCorp., Carlsbad, CA) phagemids containing cDNA in-sert using RNA Transcription Kit (Stratagene, LaJolla, CA) according to specifications from the manu-facturer.
Embryos and mandibular process explants werefixed overnight in 4% paraformaldehyde in PBS at 4°C,and processed according to the protocol described byRosen and Beddington (1993) with minor modifica-tions. Briefly, after fixation, the specimens werebleached for 1 hr with 6% hydrogen peroxide in PBScontaining 0.1% Tween-20 at room temperature. Man-dibular explants and E10 embryos were permeabilizedwith RIPA buffer and E12 embryos were treated with10 mg/ml proteinase K for 15 min at room temperatureto enhance permeabilization. Following post-fixationand prehybridization, the specimens were hybridizedovernight with 1 mg/ml DIG-labeled riboprobes at70°C. The specimens were washed, blocked, and incu-bated with anti-DIG alkaline phosphatase conjugatedantibody (Boehringer Mannheim Corp., Indianapolis,IN) at a dilution of 1:2000 for 90 min at 4°C. Thespecimens were washed extensively after which endog-enous alkaline phosphatase activities were blocked byovernight incubation in 0.48 mg/ml levamisol (Sigma,St. Louis, MO). The color reaction was developed usingnitro blue tetrazolium (NBT) and 5-bromo-4-chloro-3-indolyl phosphate (BCIP) as substrates (Sigma, St.Louis, MO). The area of gene expression induced byBMP4-soaked beads and visualized by whole-mount insitu hybridization was determined by first photograph-ing the processed specimens and then subjecting theimages to morphometric analyses using NIH ImageVersion 1.6.1 (NIH, Bethesda, MA). Signal to back-ground was determined by density slice method within
411Sox9 and Msx2 CO-REGULATE BMP4-INDUCED CHONDROGENESIS

the software. Student’s t-test was performed on numer-ical data.
Semi-Quantitative RT-PCR
In order to maximize the sensitivity of detection,multiple PBS- or 100 ng/ml BMP4-soaked beads wereimplanted at either the rostral or proximal positions onone side of the E10 mandibular process and allowed togrow in explant culture system. Twenty-four hr later,each mandible was bisected along the midline in orderto separate the implanted half from the non-implantedhalf. Four halves of the same treatment were pooled asone sample and subjected to RT-PCR for Sox9, Msx2,and beta-actin (Stratagene, La Jolla, CA). Amplimersfor Sox9 were 59-AAGATAAGTTCCCCGTGTGC-39 and59-GTAGTGAGGAAGGTTGAAGG-39, and those forMsx2 were 59-TGTTTTCGTCGGATGAGGAG-39 and59-GTCGCTTAGGGTGACAATGC-39. Thirty, 28, and18 cycles were empirically determined for Sox9, Msx2,and beta-actin PCR, respectively to optimize for ampli-fication linearity and signal. The expression levels ofSox9 and Msx2 were normalized against beta-actin.The results were subjected to Student’s t-test.
Adenovirus and Microinjection
Recombinant adenovirus carrying haemagglutin(HA) epitope-tagged mouse Msx2 gene (AdV-Msx2)(Takahashi et al., 1998b) or LacZ gene (AdV-LacZ) ascontrol were prepared. Ten nl of the recombinant ade-novirus solution (titer of 1 3 1010 pfu/ml) accompaniedby 1% tetramethylrhodamine dextran (MolecularProbes Inc., Eugene, OR), which was used as a visiblemarker, were microinjected into the mandibular ex-plants using a glass micropipette and a Transjector5246 (Eppendorf, Hamburg, Germany) under a ste-reomicroscope. After 36 hr of culture, explants wereprocessed for HA immunostaining and b-galactosidasestaining as previously described (Takahashi et al.,1998b) to confirm the site and spread of infection.
Quantitative RT-PCR for Type II CollagenmRNA
Type II collagen mRNA expression levels were ana-lyzed by quantitative RT-PCR method as previouslydescribed (Takahashi et al., 1998a). Six explants werepooled for each analysis. The amount of type II collagenmRNA in the adenovirus microinjected half of the man-dibular explants was compared with the control unin-jected half for each AdV-LacZ and AdV-Msx2 infectedspecimens after 6 days in culture and reported as theirrelative ratio. The data were subjected to statisticalanalyses using Student’s t-test.
ACKNOWLEDGMENTS
We are grateful to Dr. Yi-Hsin Liu (University ofSouthern California) for mouse Msx2 cDNA, Dr. SilvioGutkind (National Institute of Dental and CraniofacialResearch) for AdV-LacZ and Ms. Audra Wright forassistance with Figure 3. We also thank Dr. Hari Reddiand Dr. Lee Niswander for valuable suggestions to this
project. Human recombinant BMP4 is a gift from Ge-netics Institute.
REFERENCES
Barlow AJ, Francis-West PH. 1997. Ectopic application of recombi-nant BMP-2 and BMP-4 can change patterning of developing chickfacial primordia. Development 124:391–398.
Bi W, Deng JM, Zhang A, Behringer RR, de Crombrugghe B. 1999.Sox9 is required for cartilage formation. Nat Genet 22:85–89.
Buckland RA, Collinsosn JM, Graham E, Davidson DR, Hill RE. 1998.Antagonistic effects of FGF4 on BMP induction of apoptosis andchondrogenesis in the chick limb bud. Mech Dev 71:143–150.
Bell DM, Leung KKH, Wheatley SC, Ng LJ, Zhou S, Ling KW, ShamMH, Koopman P, Tam P PL, Cheah KSE. 1997. SOX9 directlyregulates the type-II collagen gene. Nat Genet 16:174–178.
Bell JR, Noveen A, Liu YH, Ma L, Dobias S, Kundu R, Luo W, Xia Y,Maxson R. 1993. Genomic structure, chromosomal location, andevolution of the mouse Hox 8 gene. Genomics 16:123–131.
Bendall AJ, Rincon-Limas DE, Botas J, Abate-Shen C. 1998. Proteincomplex formation between Msx1 and Lhx2 homeoproteins is in-compatible with DNA binding activity. Differentiation 63:151–157.
Bennett JH, Hunt P, Thorogood P. 1995. Bone morphogenetic pro-tein-2 and -4 expression during murine orofacial development. ArchOral Biol 40:847–854.
Catron KM, Zhang H, Marshall SC, Inostroza JA, Wilson JM, AbateC. 1995. Transcriptional repression by Msx-1 does not require ho-meodomain DNA-binding sites. Mol Cell Biol 15:861–871.
Cottrill CP, Archer CW, Hornbruch A, Wolpert L. 1987. The differen-tiation of the normal and muscle-free distal chick limb bud mesen-chyme in micromass culture. Dev Biol 119:143–151.
Davidson D. 1995. The function and evolution of Msx genes: pointersand paradoxes. Trends Genet 11:405–411.
Derynck R, Zhang Y, Feng XH. 1998. Smads: transcriptional activa-tors of TGF-beta responses. Cell 95:737–740.
Dewulf N, Verschueren K, Lonnyoy O, Moren A, Grimsby S, VandeSpiegle K, Miyazono K, Huylebroeck D, Ten Dijke P. 1995. Distinctspatial and temporal expression patterns of two type I receptors forbone morphogenetic proteins during mouse embryogenesis. Endo-crinology 136: 2652–2663.
Dodig M, Kronenberg MS, Bedalov A, Kream BE, Gronowicz G, ClarkSH, Mack K, Liu YH, Maxon R, Pan ZZ, Upholt WB, Rowe DW,Lichtler AC. 1996. Identification of a TAAT-containing motif re-quired for high level expression of the COL1A1 promoter in differ-entiated osteoblasts of transgenic mice. Biol Chem 271:16422–16429.
Ducy P, Zhang R, Geoffroy V, Ridall AL, Karsenty G. 1997. Osf2/Cbfa1: a transcriptional activator of osteoblast differentiation. Cell89:747–754.
Duprez D, Bell EJ, Richardson MK, Archer CW, Wolpert L, BrickellPM, Francis-West PH. 1996a. Overexpression of BMP-2 and BMP-4alters the size and shape of developing skeletal elements in thechick limb. Mech Dev 57:145–157.
Duprez DM, Colety M, Amthor H, Brickell PM, Tickle C. 1996b. Bonemorphogenetic protein-2 (BMP-2) inhibits muscle development andpromotes cartilage formation in chick limb bud cultures. Dev Biol174:448–452.
Ekanayake S, Hall BK. 1997. The in vivo and in vitro effects of bonemorphogenetic protein-2 on the development of the chick mandible.Int J Dev Biol 41:67–81.
Foerst-Potts L, Sadler TW. 1997. Disruption of Msx-1 and Msx-2reveals roles for these genes in craniofacial, eye, and axial develop-ment. Dev Dyn 209:70–84.
Foster JW, Dominguez-Steglich MA, Guioli S, Kwok C, Weller PA,Weissenbach J, Mansour S, Young ID, Goodfellow PN, Brook JD,Schafer AJ. 1994. Campomelic dysplasia and autosomal sex rever-sal caused by mutations in an SRY-related gene. Nature 372:525–530.
Francis-West PH, Tatla T, Brickell PM. 1994. Expression pattern ofthe bone morphometric protein genes Bmp-4 and Bmp-2 in thedeveloping chick face suggest a role in outgrowth of the primordia.Dev Dyn 201:168–178.
Francis-West PH, Richardson MK, Bell E, Chen P, Luyten F, Adel-fattah A, Barlow AJ, Brickell PM, Wolpert L, Archer CW. 1996. The
412 SEMBA & NONAKA ET AL.

effect of overexpression of BMPs and GDF-5 on the development ofchick limb skeletal elements. Ann NY Acad Sci 785:254–255.
Frenz DA, Liu W, Capparelli M. 1996. Role of BMP-2a in otic capsulechondrogenesis. Ann NY Acad Sci 785:256–258.
Graff JM. 1997. Embryonic patterning: to BMP or not to BMP, this isthe question. Cell 89:171–174.
Grigoriou M, Tucker AS, Sharpe PT, Pachnis V. 1998. Expression andregulation of Lhx6 and Lhx7, a novel subfamily of LIM homeodo-main encoding genes, suggests a role in mammalian head develop-ment. Development 125:2063–2074.
Hall BK. 1982. Distribution of osteo- and chondrogenetic neural crest-derived cells and of osteogenically inductive epithelia in mandibu-lar arches of embryonic chicks. J Embryol Exp Morphol 68:127–136.
Hall BK, Miyake T. 1995. Divide, accumulate, differentiate: cell con-densation in skeletal development revisited. Int J Dev Biol 39:881–893.
Harada S, Rodan SB, Goodfellow PA, Rodan GA. 1996. SOX9, adevelopmental transcription factor, stimulates type X collagen pro-moter via unique recognition motifs. J Bone Mineral Res 11(supp.1):145–145.
Healy C, Uwanogho D, Sharpe PT. 1996. Expression of the chickenSox9 gene marks the onset of cartilage differentiation. Ann NYAcad Sci 785:261–262.
Healy C, Uwanogho D, Sharpe PT. 1999. Regulation and role of Sox9in cartilage formation. Dev Dyn 215:69–78.
Hogan BL. 1996. Bone morphogenetic proteins: multifunctional reg-ulators of vertebrate development. Genes Dev 10:1580–1594.
Jernvall J, Aberg T, Kettunen P, Keranen S, Thesleff I. 1998. The lifehistory of an embryonic signaling center: BMP-4 induces p21 and isassociated with apoptosis in the mouse tooth enamel knot. Devel-opment 125:373–382.
Kaplan FS, Shore EM. 1998. Encrypted morphogens of skeletogen-esis: biological errors and pharmacologic potentials. Biochem Phar-macol 55:373–382.
Kanzler B, Kuschert SJ, Liu YH, Mallo M. 1998. Hoxa-2 restricts thechondrogenic domain and inhibits bone formation during develop-ment of the branchial area. Development 125:2587–2597.
Kawabata M, Imamura T, Miyazono K. 1998. Signal transduction bybone morphogenetic proteins. Cytokine Growth Factor Rev 9:49–61.
Kawakami Y, Ishikawa T, Shimabara M, Tanda N, Enomoto-IwamotoM, Iwamoto M, Kuwana T, Ueki A, Noji S, Nohno T. 1996. BMPsignaling during bone pattern determination in the developinglimb. Development 122:3557–3566.
Kim HJ, Rice D, Kettunen P, Thesleff I. 1998. FGF-, BMP- andShh-mediated signalling pathways in the regulation of cranial su-ture morphogenesis and calvarial bone development. Development125:1241–1251.
Kretzschmar M, Doody J, Massague J. 1997. Opposing BMP and EGFsignalling pathways converge on the TGF-beta family mediatorSmad1. Nature 389:618–622.
Lanchoney TF, Olmsted EA, Shore EM, Gannon FA, Rosen V, ZasloffMA, Kaplan FS. 1998. Characterization of bone morphogeneticprotein 4 receptor in fibrodysplasia ossificans progressiva. ClinOrthop 346:38–45.
Langille RM. 1994. In vitro analysis of the spatial organization ofchondrogenic regions of avian mandibular mesenchyme. Dev Dyn201:55–62.
Leboy PS, Sullivan TA, Nooreyazdan M, Venezian RA. 1997. Rapidchondrocyte maturation by serum-free culture with BMP-2 andascorbic acid. J Cell Biochem 66:394–403.
Lee YS, Chuong CM. 1997. Activation of protein kinase A is a pivotalstep involved in both BMP-2- and cyclic AMP-induced chondrogen-esis. J Cell Physiol 170:153–165.
Lefebvre V, Huang W, Harley VR, Goodfellow PN, de Crombrugghe B.1997. Sox9 is a potent activator of the chondrocyte-specific enhancerof the pro alpha1(II) collagen gene. Mol Cell Biol 17:2336–2346.
Lefebvre V, Li P, de Crombrugghe B. 1998. A new long form of Sox5(L-Sox5), Sox6 and Sox9 are coexpressed in chondrogenesis andcooperatively activate the type II collagen gene. EMBO J 17:5718–5733.
Liu YH, Kundu R, Wu L, Luo W, Ignelzi MR Jr, Snead ML, MaxsonRE, Jr. 1995. Premature suture closure and ectopic cranial bone in
mice expressing Msx2 transgenes in the developing skull. Proc NatlAcad Sci USA 92:6137–6141.
Lumsden A, Sprawson N, Graham A. 1991. Segmental origin andmigration of neural crest cells in the hindbrain region of the chickembryo. Development 113:1281–1291.
Lyons KM, Pelton RW, Hogan BL. 1989. Patterns of expression ofmurine Vgr-1 and BMP-2a RNA suggest that transforming growthfactor-beta-like genes coordinately regulate aspects of embryonicdevelopment. Genes Dev 3:1657–1668.
Maas R, ChenYP, Bei M, Woo I, Satokata I. 1996. The role of Msxgenes in mammalian development. Ann NY Acad Sci 785:171–181.
Macias D, Ganan Y, Sampath TK, Piedra ME, Ros MA, Hurle JM.1997. Role of BMP-2 and OP-1 (BMP-7) in programmed cell deathand skeletogenesis during chick limb development. Development124:1109–1117.
Marazzi G, Wang Y, Sasoon D. 1997. Msx2 is a transcriptional regu-lator in the BMP4-mediated programmed cell death pathway. DevBiol 186:127–138.
Metsaranta M, Toman D, de Crombrugghe B, Vuorio E. 1991. Mousetype II collagen gene. Complete nucleotide sequence, exon struc-ture, and alternative splicing. J Biol Chem 266:16862–16869.
Mina M, Gluhak J, Roger B. 1996. Downregulation of Msx-2 expres-sion results in chondrogenesis in the medial region of the avianmandible. Connect Tissue Res 35:79–84.
Mishina Y, Suzuki A, Ueno N, Behringer RR.1995. Bmpr encodes atype I bone morphogenetic protein receptor that is essential forgastrulation during mouse embryogenesis. Genes Dev 9:3027–3037.
Monsoro-Burq AH, Duprez D, Watanabe Y, Bontoux M, Vincent C,Brickell P, Le Douarin N. 1996. The role of bone morphogeneticproteins in vertebral development. Development 122:3607–3616.
Neubuser A, Peters H, Balling R, Martin GR. 1997. Antagonisticinteractions between FGF and BMP signaling pathways: a mecha-nism for positioning the sites of tooth formation. Cell 90:247–255.
Newberry EP, Boudreaux JM, Towler DA. 1997. Stimulus-selectiveinhibition of rat osteocalcin promoter induction and protein-DNAinteractions by the homeodomain repressor Msx2. Biol Chem 272:29607–29613.
Ng LJ, Wheatley S, Muscat GE, Conway-Campbell J, Bowles J,Wright E, Bell DM, Tam PP, Cheah KS, Koopman P. 1997. Sox9binds DNA, activates transcription and coexpresses with type IIcollagen during chondrogenesis in the mouse. Dev Biol 183:108–121.
Nifuji A, Kellermann O, Kuboki Y, Wozney JM, Noda M. 1997. Per-turbation of BMP signaling in somitogenesis resulted in vertebraland rib malformations in the axial skeletal formation. J Bone MinerRes 12:332–342.
Niswander L, Martin GR. 1993. FGF-4 and BMP-2 have oppositeeffects on limb growth. Nature 361:68–71.
Nonaka K, Shum L, Takahashi I, Takahashi K, Ikura T, Dashner R,Nuckolls GH, Slavkin HC. 1999. Convergence of the BMP and EGFsignaling pathways on Smad1 in the regulation of chondrogenesis.Int J Dev Biol 43:795–807.
Nuckolls GH, Slavkin HC, Shum L. 1998. Bone morphogenetic proteinsignaling in limb and craniofacial development. In: Davidowitch Z,editor. Proceedings of the biological mechanisms of tooth eruption,resorption and replacement by implants. Birmingham, Alabama:Harvard Society for the Advancement of Orthodontics, EBSCOMedia. p 39–47,
Osumi-Yamashita N, Ninomiya Y, Doi H and Eto K. 1994. The con-tribution of both forebrain and midbrain crest cells to the mesen-chyme in the frontonasal mass of mouse embryos. Dev Biol 164:409–419.
Phippard, DJ, Weber-Hall SJ, Sharpe PT, Naylor MS, Jayatalake H,Maas R, Woo I, Roberts-Clark D, Francis-West PH, Liu YH, MaxsonR, Hill RE, Dale TC. 1996. Regulation of Msx-1, Msx-2, Bmp-2 andBmp-4 during foetal and postnatal mammary gland development.Development 122:2729–2737.
Qiu M, Bulfone A, Ghattas I, Meneses JJ, Christensen L, Sharpe PT,Presley R, Pedersen RA, Rubenstein JL. 1997. Role of the Dlxhomeobox genes in proximodistal patterning of the branchial arch-es: mutations of Dlx-1, Dlx-2, and Dlx-1 and -2 alter morphogenesisof proximal skeletal and soft tissue structures derived from the firstand second arches. Dev Biol 185:165–184.
413Sox9 and Msx2 CO-REGULATE BMP4-INDUCED CHONDROGENESIS

Reddi AH. 1994. Bone and cartilage differentiation. Curr Opin GenetDev 4:737–744.
Reddi AH. 1995. Cartilage morphogenesis: role of bone and cartilagemorphogenetic proteins, homeobox genes and extracellular matrix.Matrix Biol 14:599–606.
Reddi AH. 1997. Bone morphogenetic protein: an unconventionalapproach to isolation of first mammalian morphogens. CytokineGrowth Factor Rev 8:11–20.
Reshef R, Maroto M, Lassar AB. 1998. Regulation of dorsal somiticcell fates: BMPs and Noggin control the timing and pattern ofmyogenic regulator expression. Genes Dev 12:290–303.
Ros MA, Piedra ME, Fallon JF, Hurle JM. 1997. Morphogeneticpotential of the chick leg interdigital mesoderm when diverted fromthe cell death program. Dev Dyn 208:406–419.
Rosen B, Beddington RSP. 1993. Whole-mount in situ hybridization inthe mouse embryo: gene expression in three dimensions. TrendsGenet 9:162–167.
Satokata I, Maas R. 1994. Msx1 deficient mice exhibit cleft palate andabnormalities of craniofacial and tooth development. Nat Genet6:348–356.
Sechrist J, Serbedzija GN, Scherson T, Fraser SE, Bronner-Fraser M.1993. Segmental migration of the hindbrain neural crest does notarise from its segmental generation. Development 118:691–703.
Sekiya I, Koopman P, Watanabe H, Ezura Y, Yamada Y, Noda M.1997. SOX9 enhances aggrecan gene expression via the promoterregion containing a single HMG-BOX-sequence in a chondrogeniccell line, TC6. J Bone Miner Res 12 (supp.1):P222–P222.
Semenza GL, Wang GL, Kundu R. 1995. DNA binding and transcrip-tional properties of wild-type and mutant forms of the homeodo-main protein Msx2. Biochem Biophys Res Commun 209:257–262.
Shafritz AB, Shore EM, Gannon FH, Zasloff MA, Taub R, Muenke M,Kaplan FS. 1996. Overexpression of an osteogenic morphogen infibrodysplasia ossificans progressiva. N Engl J Med 335:555–561.
Shum L, Sakakura Y, Bringas P, Luo W, Snead ML, Mayo M, CrohinC, Millar S, Werb Z, Buckely S, Hall FL, Warburton D, Slavkin HC.1993. EGF abrogation induced fusilli-form dysmorphogenesis ofMeckel’s cartilage during embryonic mouse mandibular morpho-genesis in vitro. Development 118:903–917.
Slavkin HC, Bringas P, Sasano Y, Mayo M. 1989. Early embryonicmouse mandibular morphogenesis and cytodifferentiation in se-rumless, chemically-defined medium: A model for studies of auto-crine and/or paracrine regulatory factors. J Craniofac Genet DevBiol 9:185–205.
Slavkin HC, Nuckolls GH, Shum L. 1999. Craniofacial developmentand patterning. In: Tuan RS, Lo CW, editors. Methods in molecularbiology, vol. 136: Developmental biology protocols, vol. II. HumanaPress Inc., Totowa, NJ. p 45–54.
Takahashi I, Nuckolls GH, Takahashi K, Tanaka O, Semba I, Dash-ner R, Shum L, Slavkin HC. 1998a. Compressive force inducesSox9, Type II collagen and Aggrecan and inhibits IL-1b expressionresulting in chondrogenesis in mouse embryonic limb bud mesen-chymal cells. J Cell Sci 111:2067–2076.
Takahashi K, Nuckolls GH, Tanaka O, Semba I, Takahashi I, Dash-ner R, Shum L, Slavkin HC. 1998b. Adenovirus mediated ectopicexpression of Msx2 in even-numbered rhombomeres induces apo-ptotic elimination of cranial neural crest cells en ovo. Development125:1627–1635.
Takahashi Y, Tonegawa A, Matsumoto K, Ueno N, Kuroiwa A, NodaM, Nifuji A. 1996. BMP-4 mediates interacting signals between theneural tube and skin along the dorsal midline. Genes Cells 1:775–783.
Theiler K. 1989. The house mouse: atlas of embryonic development .New York: Springer-Verlag. p 73–77.
Thomas BL, Tucker AS, Qui M, Ferguson CA, Hardcastle Z, Ruben-stein JL, Sharpe PT. 1997. Role of Dlx-1 and Dlx-2 genes in patter-ing of the murine dentition. Development 124:4811–4818.
Tucker AS, Matthews KL, Sharpe PT. 1998. Transformation of toothtype induced by inhibition of BMP signaling. Science 282:1136–1138.
Tureckova J, Sahlberg C, Aberg T, Ruch JV, Thesleff I, Peterkova R.1995. Comparison of expression of the msx-1, msx-2, BMP-2 andBMP-4 genes in the mouse upper diastemal and molar tooth pri-mordia. Int J Dev Biol 39:459–468.
Urist MR. 1965. Bone: formation by autoinduction. Science 150:893–899.
Urist MR. 1997. Bone morphogenetic protein: the molecularization ofskeletal system development. J Bone Miner Res 12:343–346.
Vortkamp A. 1997. Defining the skeletal elements. Curr Biol 7:R104–R107.
Wagner T, Wirth J, Meyer J, Zabel B, Held M, Zimmer J, Pasantes J,Bricarelli FD, Keutel J, Hustert E, Wolf U, Tommeruo N, SchemppW, Scherer G. 1994. Autosomal sex reversal and campomelic dys-plasia are caused by mutations in and around the SRY-related geneSOX9. Cell 79:1111–1120.
Watanabe Y, Le Douarin NM. 1996. A role for BMP-4 in the develop-ment of subcutaneous cartilage. Mech Dev 57:69–78.
Watanabe Y, Duprez D, Monsoro-Burq AH, Vincent C, Douarin NM.1998. Two domains in vertebral development: antagonistic regula-tion by SHH and BMP4 proteins. Development 125:2631–2639.
Wheatley S, Wright E, Jeske Y, McCormack A, Bowles J, Koopman P.1996. Aetiology of the skeletal dysmorphology syndrome cam-pomelic dysplasia: expression of the Sox9 gene during chondrogen-esis in mouse embryos. Ann NY Acad Sci 785:350–352.
Winnier G, Blessing M, Labosky PA, Hogan BLM. 1995. Bone mor-phogenetic protein-4 is required for mesoderm formation and pat-terning in the mouse. Genes Dev 9:2105–2116.
Winograd J, Reilly MP, Roe R, Lutz J, Laughner E, Xu X, Asakura T,vander Kolk C, Strandberg JD, Semenza GL. 1997. Perinatal le-thality and multiple craniofacial malformations in MSX2 trans-genic mice. Hum Mol Genet 6:369–379.
Woloshin P, Song K, Degnin C, Killary AM, Goldhamer DJ, SassoonD, Thayer MJ. 1995. MSX1 inhibits myoD expression in fibroblast x10T1/2 cell hybrids. Cell 82:611–620.
Wozney JM, Rosen V. 1998. Bone morphogenetic protein and bonemorphogenetic protein gene family in bone formation and repair.Clin Orthop 346:26–37.
Wozney JM, Rosen V, Celeste AJ, Mitsock LM, Whitters MJ, Kriz RW,Hewick RM, Wang EA. 1988. Novel regulators of bone formation:molecular clones and activities. Science 242:1528–1534.
Wright E, Hargrave MR, Christiansen J, Cooper L, Kun J, Evans T,Gangadharan U, Greenfield A, Koopman P. 1995. The Sry-relatedgene Sox-9 is expressed during chondrogenesis in mouse embryos.Nat Genet 9:15–20.
Wu L, Wu H, Ma L, Sangiorgi F, Wu N, Bell JR, Lyons GE, Maxson R.1997. Miz1, a novel zinc finger transcription factor that interactswith Msx2 and enhances its affinity for DNA. Mech Dev 65:3–17.
Zhang H, Bradley A. 1996. Mice deficient for BMP2 are nonviable andhave defects in amnion/chorion and cardiac development. Develop-ment 122:2977–2986.
Zhang H, Catron KM, Abate-Shen C. 1996. A role for the Msx-1homeodomain in transcriptional regulation: residues in the N-ter-minal arm mediate TATA binding protein interaction and tran-scriptional repression. Proc Natl Acad Sci USA 93:1764–1769.
Zhang H, Hu G, Wang H, Sciavolino P, Iler N, Shen MM, Abate-ShenC. 1997. Heterodimerization of Msx and Dlx homeoproteins resultsin functional antagonism. Mol Cell Biol 17:2920–2932.
Zhao Q, Eberspaecher H, Lefebvre V, De Crombrugghe B. 1997.Parallel expression of Sox9 and Col2a1 in cells undergoing chon-drogenesis. Dev Dyn 209:377–386.
Zou H, Wieser R, Massague J, Niswander L. 1997. Distinct roles oftype I bone morphogenetic protein receptors in the formation anddifferentiation of cartilage. Genes Dev 11:2191–2203.
414 SEMBA & NONAKA ET AL.

















