Embed Size (px)
Citation preview

1
2
3
45
6
7
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
34
35
36
37
38
39
40
41
42
43
44
45
46
European Journal of Pharmacology xxx (2008) xxx-xxx
EJP-65340; No of Pages 9
Contents lists available at ScienceDirect
European Journal of Pharmacology
j ourna l homepage: www.e lsev ie r.com/ locate /e jphar
ARTICLE IN PRESS
Potent antioxidant role of Pirfenidone in experimental cirrhosis
Adriana Salazar-Montes a, Luis Ruiz-Corro a, Alberto López-Reyes a,Eugenio Castrejón-Gómez a, Juan Armendáriz-Borunda a,b,⁎a Institute for Molecular Biology in Medicine and Gene Therapy, CUCS, University of Guadalajara, Guadalajara, Jalisco, Mexico
^b OPD Hospital Civil, Guadalajara, Jalisco, Mexico^
FUAbbreviations: CCl4, carbon tetrachloride; TGF-β,GAPDH, glyceraldehyde-3P-dehydrogenase; SOD, superoiNOS, inducible nitric oxide synthase; HSC, hepatic stell⁎ Corresponding author. Department of Molecular
University of Guadalajara, Apartado Postal 2-123, GuadTel.: +52 33 1058 5317; fax: +52 33 1058 5318.
E-mail address: [email protected] (J. Armendá
0014-2999/$ – see front matter © 2008 Elsevier B.V. Aldoi:10.1016/j.ejphar.2008.06.110
Please cite this article as: Salazar-Montes, Adoi:10.1016/j.ejphar.2008.06.110
OA B S T R A C T
A R T I C L E I N F OArticle history:
Three important features Received 15 April 2008Received in revised form 19 June 2008Accepted 27 June 2008Available online xxxxKeywords:PirfenidoneExperimental liverGastrointestinal and urogenital pharmacology
CTED
PROmust be considered when proposing therapeutic strategies in liver cirrhosis:
inflammation, oxidative stress and fibrogenesis. Pirfenidone is a synthetic molecule which oxidative actionhas not been tested in cirrhosis. Cirrhosis was induced in rats by ligation of the common bile duct or carbontetrachloride (CCl4) chronic intoxication and treated with Pirfenidone or Diphenyleneiodonium (a potentknown anti
^oxidant) for the last two weeks for bile duct ligation model or for the last three weeks for CCl4
chronic intoxication. A 60% reduction in fibrosis index for bile duct ligation model and 42% for CCl4 alongwith reduced inflammation was observed. Considerable reduction on hepatic enzymes and total and directbilirubins were detected with Pirfenidone in both models. Pirfenidone antioxidant capacity rendered a 28%and 30% reduction in nitrites and Malonyldealdehide concentration in bile duct ligation and 52% and 38% inCCl4. With respect to gene expression, fibrotic genes like transforming growth factor-β (TGF-β) and CollagenIα (Col-1α) were down-regulated by Pirfenidone and increased expression of regenerative geneslike hepatocyte growth factor (HGF) and c-met . superoxide dismutase (SOD), catalase (CAT) and induciblenitric oxide synthase (iNOS) gene expressionwere importantly down-regulated where nuclear factor
^kappa B
(NF-κB^) binding activity also decreased with Pirfenidone treatment. Also, SOD and CAT functional activity
decreased after Pirfenidone action. On the other hand, Diphenyleneiodonium induced a drop in oxidativestress similar in extent to Pirfenidone, but it was not as effective as Pirfenidone in reducing fibrosis.In this work, we showed antioxidant properties of Pirfenidone beyond its well-known antifibrotic effect.These features make Pirfenidone an attractive drug for trying fibrotic diseases accompanied by oxidativestress processes.
© 2008 Elsevier B.V. All rights reserved.
E1. Introduction
4748
49
50
51
52
53
54
55
56
57
58
59
60
NCOR
RHepatic cirrhosis ends up in distortion of normal tissue architecturethat develops when liver is chronically damaged (Friedman, 2000;Sokol, 2002). Primary damage mechanisms involve mainly oxidativestress and free radicals generation (Tapia et al., 1997). It has beenestablished that inflammation in the liver is initiated by an oxidativeburst in Kuppfer cells (Vrba and Mondriasky, 2002; Flora, 2007). Afterthese events, a fibrogenic process sets inmotion driven by the paracrineaction of specific pro-inflammatory and pro-fibrogenic cytokines, aswell as free radicals generation by Kuppfer cells and hepatocytes which,in turn, stimulate Hepatic Stellate Cells (HSC) to produce exacerbatedamount of collagens, which constitutes the main component of liverfibrosis (Tsukamoto et al., 1995; Britton and Bacon, 1999; Poli, 2000).
61
62
63
64
65
66
67
transforming growth factor;xide dismutase; CAT, catalase;ate cells.Biology and Genomics, CUCS,alajara, Jalisco, 44281
^, Mexico.
riz-Borunda).
l rights reserved.
., et al., Potent antioxidant ro
Regarding these facts, Pirfenidone (5 methyl-1-phenyl-2-(1H)-pyridone), is a synthetic molecule with lowmolecular weight which isrecognized by its anti-inflammatory and anti-fibrogenic properties(Tada et al., 2001) demonstrated in both animal models and humans(Iyer et al., 1999). Additional piece of data has shown that prevents thedevelopment of progressive renal disease by down-regulating ofmessenger ribonucleic acid (mRNA) levels of Collagen Iα and IV (Col-Iα and IV) and transforming growth factor-β (TGF-β) (Schwart et al.,1997; Shimizu et al., 1998).
Our group has demonstrated that Pirfenidone is an anti-inflam-matory and anti-fibrogenic agent in experimental models of cirrhosis(García et al., 2002) and in humans affected with hepatic fibrosiscaused by different etiologies (Armendariz-Borunda et al., 2003).Moreover, Pirfenidone showed to improve necro-inflammation,steatosis and liver regeneration in a sub-set of patients withestablished advanced liverfibrosis causedbyHepatitis C virus infection(Armendariz-Borunda et al., 2006). We also have demonstrated thatPirfenidone has an anti-fibrogenic action inhibiting capsular contrac-ture in mammary implants (Gancedo et al., 2008). Thus, it was ourintention to explore whether Pirfenidone possesses additional anti-oxidant role in different experimental models of cirrhosis. As reference
le of Pirfenidone in experimental cirrhosis, Eur. J. Pharmacol. (2008),

C
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
2 A. Salazar-Montes et al. / European Journal of Pharmacology xxx (2008) xxx-xxx
ARTICLE IN PRESS
UNCO
RRE
we used the well known antioxidant Diphenyleneiodonium. Wefound that Pirfenidone has a potent antioxidant action and is morepronounced than Diphenyleneiodonium. Here, we also described keymolecular mechanisms involved in fibrosis resolution by Pirfenidonein two different animal models: bile duct ligation and chronicintoxication of carbon tetrachloride (CCl4).
2. Materials and methods
2.1. Materials
Pirfenidone was provided by Cell Therapy and Technology,Diphenyleneiodonium was purchased from Sigma Aldrich Co., StLouis Missouri, carbon tetrachloride was purchased from MerckCompany, Darmstadt, Germany, Nuclear factor kappa B (NF-κB
^) anti-
body was obtained from Santa Cruz Biotechnologies, Santa Cruz CA.
2.2. Animals
Wistar rats used in this study were obtained from Charles Rivers(Boston, MA, USA) and housed according to the Animal Care protocolestablished byUniversity of Guadalajara. Fibrosiswas induced by ligationof the common bile duct in animals or by CCl4 chronic intoxication inanimals weighing 80
^g as previously reported by our group (Bueno et al.,
2000; Salgado et al., 2000). Three groups of five rats each were used forcontrol, Pirfenidone
^and Diphenyleneiodonium treatment group in each
animal model. Rats in all groups were sacrificed four weeks after bileduct-ligationoreightweeks after CCl4 intoxication.Dosingof PirfenidoneandDiphenyleneiodoniumusedwere chosenon thebasis of ourpreviousexperience and early reports in the literature (Wheeler et al., 2000;García et al., 2002). In Pirfenidone
^group, bile duct-ligation and CCl4 rats
were treated with 200 mg/kg of Pirfenidone, for the last two weeks forbile duct-ligation and for the last threeweeks for CCl4 animals. A parallelgroup was treated with 1 mg/kg Diphenyleneiodonium daily for thesame period. The control groups were treated with vehicle only.
2.3. Biochemical assays
Blood was obtained from control and experimental cirrhoticanimals at the moment of sacrifice and serum transaminases weredetermined in an automated Sincron
^-7 analyzer.
2.4. Fibrosis index determination
Liver sections of approximately 0.5^cm2 were taken from each rat
liver for histology analysis with Masson's trichrome. Percentage offibrosis in the liver tissue was determined using a computer assistedautomated image analyzer Pro Plus (Cambridge Instruments, Cam-bridge) byanalyzing twenty randomfields per slide and calculating theratio of connective tissue to the whole liver area (Salgado et al., 2000).
2.5. Griess reaction
Total nitrite was analyzed colorimetrically by using Griess reactionand its absorbancewasdeterminedat550nmusinga spectrophotometer.
2.6. Malonyldealdehide (MDA) determination
Lipid peroxidation was determined according to the protocolpreviously reported (Callaway et al., 1998) and the absorbance wasread at 532 nm.
2.7. Real time polymerase chain reaction (PCR)
2 μg of total RNA extracted from all samples according to modifiedChomzynsky and Sacchi technique (Chomznsky and Sacchi, 1987)
Please cite this article as: Salazar-Montes, A., et al., Potent antioxidant rodoi:10.1016/j.ejphar.2008.06.110
TEDPR
OOF
1were added with 240 ng of random primers, 5 mM dithiothreitol1(DTT), 1 mM deoxyribonucleotide triphosphate (dNTP's) mix,
^40 units
1of RNase inhibitor, 200 units^of moloney murine Leukemia Virus
1Reverse Transcriptase (MMLV-RT) and incubated for 10 min at 25 °C,160 min at 37 °C and 10 min at 95 °C. After incubation samples were1stored at −
^70 °C.
1Quantitative real^time PCR was performed using a Rotor Gene RG
13000 Sequence Detector (Corbett Research, Sydney Australia) under the1following conditions:
^1 cycle of 50 °C for 2min,1 cycle
^of 95 °C for 10min
1and 40 cycle^s of 95 °C for 30 s and 60 °C for 40 s. The total reaction was
1made in 10 μl containing 2 μl of cDNA, 1X of Universal PCR Master Mix1(Applied Biosystems) and 1X of final concentration of primers and1TaqMan probe from experimental and control genes synthesized by1Applied Biosystems (assay on demand). Multiplex PCR Real Time for1Collagen Iα (Col-1α), transforming growth factor (TGF-β), hepatocyte1growth factor (HGF), hepatocyte growth factor receptor (c-met),1inducible nitric oxide synthase (iNOS), catalase (CAT) and superoxide1dismutase (SOD) were performed in duplicate for each animal using1glyceraldehyde-3P-dehydrogenase (GAPDH) as house keeping gene.1Data analysis was carried out according to theUser Bulletin Number 2 of1Applied biosystems (Bulletin 2 Applied Byosistem). Using sequence1detection software we calculated the threshold cycle (Ct) for each1reaction which was used to quantify the amount of starting template1reaction. A difference in Ct values (ΔCt) was calculated for each gene by1duplicate. The amount of the target gene normalized to the endogenous1gene GAPDH and relative calibrator is given by: 2
^−ΔΔCt where ΔΔCt is
1ΔCt^−ΔCtGAPDH and ΔCt is CTgene target^
−^CtGAPDH.
12.8. Superoxide dismutase activity
1SOD activity was quantitatively determined in tissue homogenate1loaded in polyacrylamide gel. The gel was stained and incubated for1SOD activity as described by Misra Fridovich (Burke and Oliver, 1992).1Incubation took place in the dark and then exposed to light under a1400 W high pressure sodium lamp 60 cm above the gel. SOD activity1was observed like achromatic zone in the gel.
12.9. Catalase activity
1Catalase activity was determined according to the zymographic1method reported by Gennady P. Manchenko (1994). The method is1based on the starch–iodine reaction. Thiosulfate in the staining1solution is inactivated by hydrogen peroxide except at the sites of CAT1activity, where hydrogen peroxide is destroyed enzymatically. The1iodide is oxidized by hydrogen peroxide to iodine which forms a1chromatophore with the starch and sites of CAT localization remain1achromatic.
12.10. Obtention of nuclear extracts
1Isolation of nuclear proteins was carried out with little modifica-1tions according to the method described by Andrews (Andrews and1Faller, 1991). Supernatant fraction containing DNA binding proteins1was recollected and quantified by the Bradford method (Bradford11976). Supernatant was stored at −70 °C in aliquots until use.
12.11. Gel mobility shift assays
1Electrophoretic mobility shift assays (EMSA) were performed as1described elsewhere (Armendariz-Borunda et al., 1994). Binding1reactions were made using 2 μg of nuclear extracts from experimental1and control animals. 100,000 cpm of radioactive NF-κB probe and11.2 μg of poly (dI–dC) as a non-specific DNA competitor were included1in the binding reactions. Mixtures were incubated for 30 min at room1temperature in binding buffer containing 10 mM 4-(2-hydroxyethyl)-11-piperazineethanesulfonic acid (HEPES) (pH 7.5), 50 mM NaCl, 1 mM
le of Pirfenidone in experimental cirrhosis, Eur. J. Pharmacol. (2008),
OF
177
178
179
180Q1181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
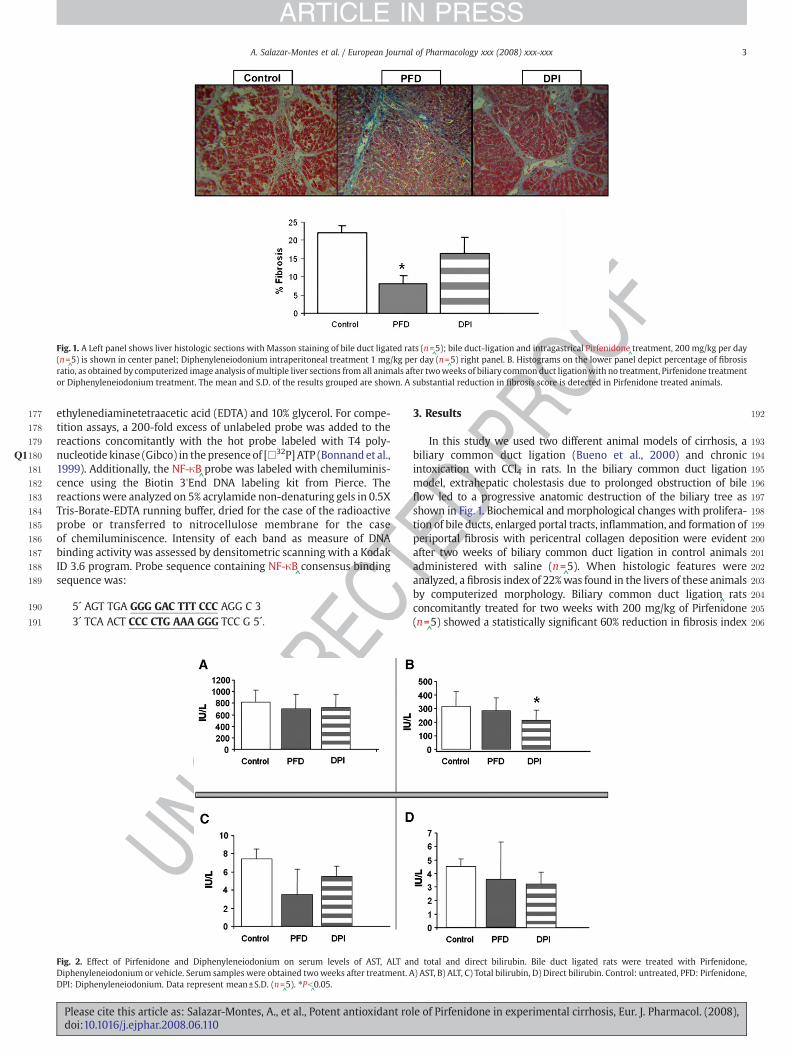
Fig. 1. A Left panel shows liver histologic sections with Masson staining of bile duct ligated rats (n=^5); bile duct-ligation and intragastrical Pirfenidone
^treatment, 200 mg/kg per day
(n=^5) is shown in center panel; Diphenyleneiodonium intraperitoneal treatment 1 mg/kg per day (n=
^5) right panel. B. Histograms on the lower panel depict percentage of fibrosis
ratio, as obtained by computerized image analysis ofmultiple liver sections from all animals after twoweeks of biliary common duct ligationwith no treatment, Pirfenidone treatmentor Diphenyleneiodonium treatment. The mean and S.D. of the results grouped are shown. A substantial reduction in fibrosis score is detected in Pirfenidone treated animals.
3A. Salazar-Montes et al. / European Journal of Pharmacology xxx (2008) xxx-xxx
ARTICLE IN PRESS
Cethylenediaminetetraacetic acid (EDTA) and 10% glycerol. For compe-tition assays, a 200-fold excess of unlabeled probe was added to thereactions concomitantly with the hot probe labeled with T4 poly-nucleotide kinase (Gibco) in the presence of [□32P] ATP (Bonnand et al.,1999). Additionally, the NF-κB
^probe was labeled with chemiluminis-
cence using the Biotin 3'End DNA labeling kit from Pierce. Thereactionswere analyzed on 5% acrylamide non-denaturing gels in 0.5XTris-Borate-EDTA running buffer, dried for the case of the radioactiveprobe or transferred to nitrocellulose membrane for the caseof chemiluminiscence. Intensity of each band as measure of DNAbinding activity was assessed by densitometric scanning with a KodakID 3.6 program. Probe sequence containing NF-κB
^consensus binding
sequence was:
5´ AGT TGA GGG GAC TTT CCC AGG C 33´ TCA ACT CCC CTG AAA GGG TCC G 5´.
UNCO
RRE
Fig. 2. Effect of Pirfenidone and Diphenyleneiodonium on serum levels of AST, ALT anDiphenyleneiodonium or vehicle. Serum samples were obtained twoweeks after treatment. ADPI: Diphenyleneiodonium. Data represent mean±S.D. (n=
^5). ⁎Pb
^0.05.
Please cite this article as: Salazar-Montes, A., et al., Potent antioxidant rodoi:10.1016/j.ejphar.2008.06.110
TEDPR
O
3. Results
In this study we used two different animal models of cirrhosis, abiliary common duct ligation (Bueno et al., 2000) and chronicintoxication with CCl4 in rats. In the biliary common duct ligationmodel, extrahepatic cholestasis due to prolonged obstruction of bileflow led to a progressive anatomic destruction of the biliary tree asshown in Fig. 1. Biochemical and morphological changes with prolifera-tion of bile ducts, enlarged portal tracts, inflammation, and formation ofperiportal fibrosis with pericentral collagen deposition were evidentafter two weeks of biliary common duct ligation in control animalsadministered with saline (n=
^5). When histologic features were
analyzed, a fibrosis index of 22%was found in the livers of these animalsby computerized morphology. Biliary common duct ligation
^rats
concomitantly treated for two weeks with 200 mg/kg of Pirfenidone(n=
^5) showed a statistically significant 60% reduction in fibrosis index
d total and direct bilirubin. Bile duct ligated rats were treated with Pirfenidone,) AST, B) ALT, C) Total bilirubin, D) Direct bilirubin. Control: untreated, PFD: Pirfenidone,
le of Pirfenidone in experimental cirrhosis, Eur. J. Pharmacol. (2008),
F
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
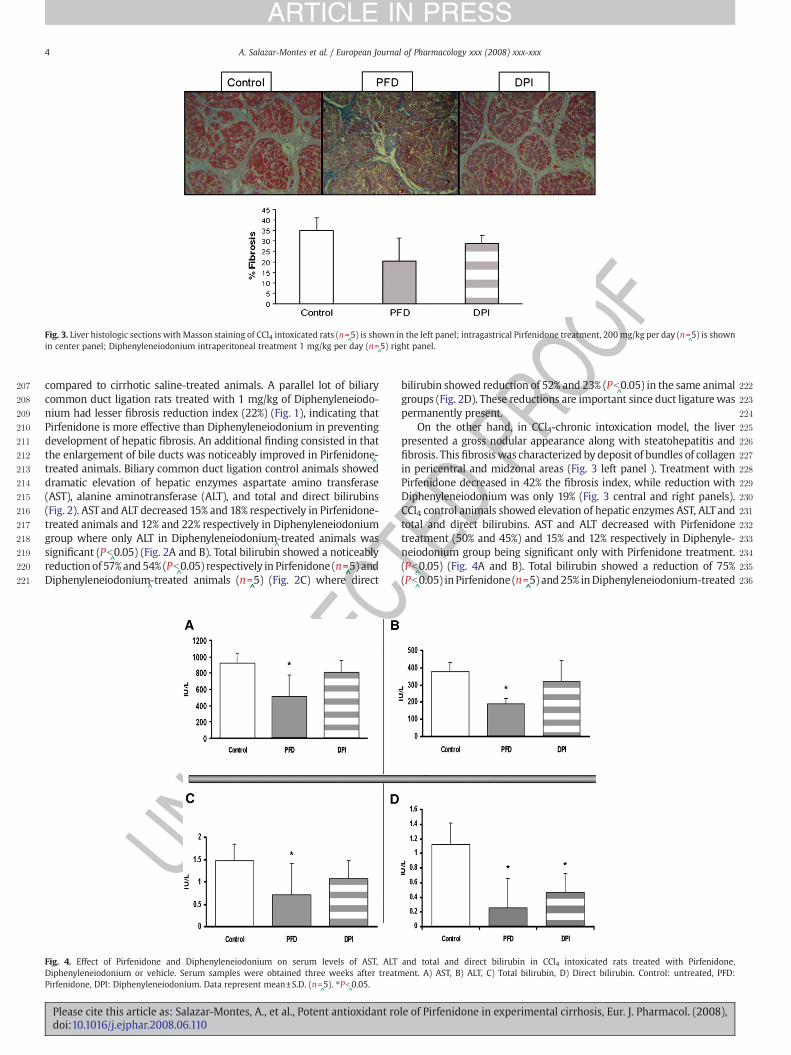
Fig. 3. Liver histologic sections with Masson staining of CCl4 intoxicated rats (n=^5) is shown in the left panel; intragastrical Pirfenidone treatment, 200 mg/kg per day (n=
^5) is shown
in center panel; Diphenyleneiodonium intraperitoneal treatment 1 mg/kg per day (n=^5) right panel.
4 A. Salazar-Montes et al. / European Journal of Pharmacology xxx (2008) xxx-xxx
ARTICLE IN PRESS
compared to cirrhotic saline-treated animals. A parallel lot of biliarycommon duct ligation rats treated with 1 mg/kg of Diphenyleneiodo-nium had lesser fibrosis reduction index (22%) (Fig. 1), indicating thatPirfenidone is more effective than Diphenyleneiodonium in preventingdevelopment of hepatic fibrosis. An additional finding consisted in thatthe enlargement of bile ducts was noticeably improved in Pirfenidone
^-
treated animals. Biliary common duct ligation control animals showeddramatic elevation of hepatic enzymes aspartate amino transferase(AST), alanine aminotransferase (ALT), and total and direct bilirubins(Fig. 2). ASTand ALT decreased 15% and 18% respectively in Pirfenidone-treated animals and 12% and 22% respectively in Diphenyleneiodoniumgroup where only ALT in Diphenyleneiodonium
^-treated animals was
significant (Pb^0.05) (Fig. 2A and B). Total bilirubin showed a noticeably
reductionof 57%and54% (Pb^0.05) respectively in Pirfenidone (n=
^̂5) and
Diphenyleneiodonium^-treated animals (n=
^̂5) (Fig. 2C) where direct
UNCO
RREC
Fig. 4. Effect of Pirfenidone and Diphenyleneiodonium on serum levels of AST, ALTDiphenyleneiodonium or vehicle. Serum samples were obtained three weeks after treatmPirfenidone, DPI: Diphenyleneiodonium. Data represent mean±S.D. (n=
^5). ⁎Pb
^0.05.
Please cite this article as: Salazar-Montes, A., et al., Potent antioxidant rodoi:10.1016/j.ejphar.2008.06.110
EDPR
OO 2bilirubin showed reduction of 52% and 23% (Pb^0.05) in the same animal
2groups (Fig. 2D). These reductions are important since duct ligaturewas2permanently present.2On the other hand, in CCl4-chronic intoxication model, the liver2presented a gross nodular appearance along with steatohepatitis and2fibrosis. This fibrosiswas characterized by deposit of bundles of collagen2in pericentral and midzonal areas (Fig. 3 left panel ). Treatment with2Pirfenidone decreased in 42% the fibrosis index, while reduction with2Diphenyleneiodonium was only 19% (Fig. 3 central and right panels).2CCl4 control animals showed elevation of hepatic enzymes AST, ALT and2total and direct bilirubins. AST and ALT decreased with Pirfenidone2treatment (50% and 45%) and 15% and 12% respectively in Diphenyle-2neiodonium group being significant only with Pirfenidone treatment.2(Pb
^0.05) (Fig. 4A and B). Total bilirubin showed a reduction of 75%
2(Pb^0.05) in Pirfenidone (n=
^̂5) and25% inDiphenyleneiodonium-treated
Tand total and direct bilirubin in CCl4 intoxicated rats treated with Pirfenidone,ent. A) AST, B) ALT, C) Total bilirubin, D) Direct bilirubin. Control: untreated, PFD:
le of Pirfenidone in experimental cirrhosis, Eur. J. Pharmacol. (2008),
OOF
237
238
239
240
241
242
243
244
245
246
247
248
249
250
251
252
253
254
255
256
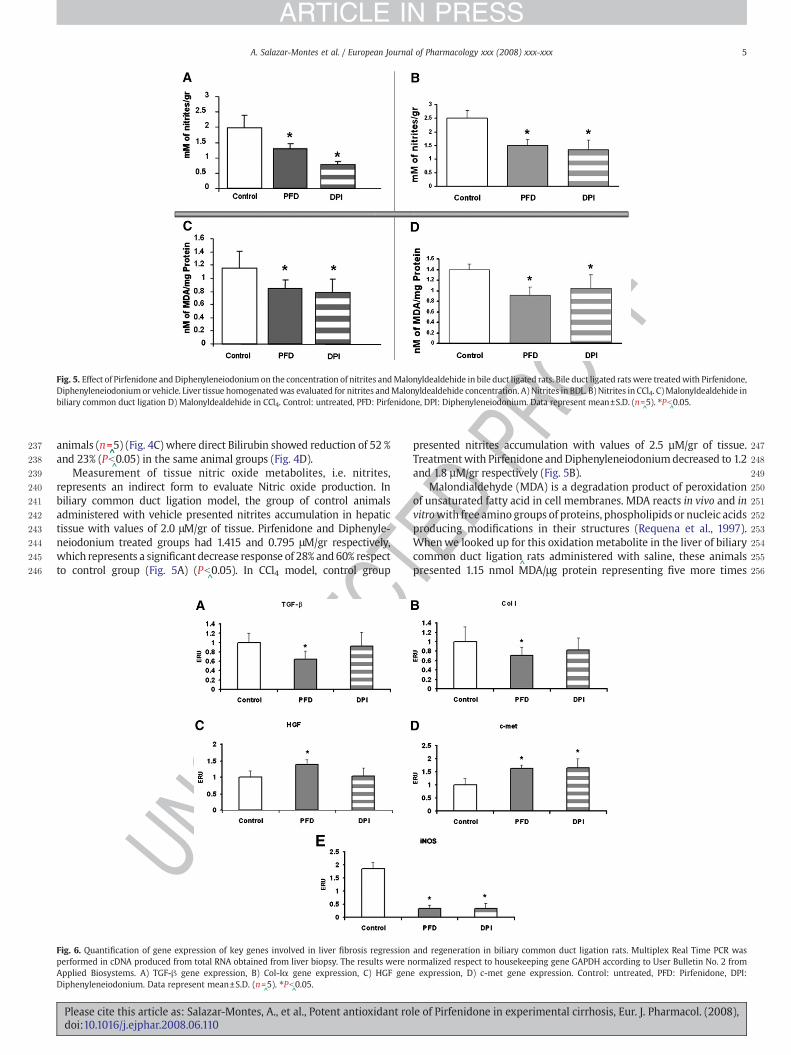
Fig. 5. Effect of Pirfenidone and Diphenyleneiodoniumon the concentration of nitrites andMalonyldealdehide in bile duct ligated rats. Bile duct ligated rats were treatedwith Pirfenidone,Diphenyleneiodoniumor vehicle. Liver tissue homogenatedwas evaluated for nitrites andMalonyldealdehide concentration. A) Nitrites in BDL. B) Nitrites in CCl4. C)Malonyldealdehide inbiliary common duct ligation D) Malonyldealdehide in CCl4. Control: untreated, PFD: Pirfenidone, DPI: Diphenyleneiodonium. Data represent mean±S.D. (n=
^5). ⁎Pb
^0.05.
5A. Salazar-Montes et al. / European Journal of Pharmacology xxx (2008) xxx-xxx
ARTICLE IN PRESS
animals (n=^̂5) (Fig. 4C) where direct Bilirubin showed reduction of 52 %
and 23% (Pb^0.05) in the same animal groups (Fig. 4D).
Measurement of tissue nitric oxide metabolites, i.e. nitrites,represents an indirect form to evaluate Nitric oxide production. Inbiliary common duct ligation model, the group of control animalsadministered with vehicle presented nitrites accumulation in hepatictissue with values of 2.0 μM/gr of tissue. Pirfenidone and Diphenyle-neiodonium treated groups had 1.415 and 0.795 μM/gr respectively,which represents a significant decrease response of 28% and 60% respectto control group (Fig. 5A) (Pb
^0.05). In CCl4 model, control group
UNCO
RREC
Fig. 6. Quantification of gene expression of key genes involved in liver fibrosis regressionperformed in cDNA produced from total RNA obtained from liver biopsy. The results were nApplied Biosystems. A) TGF-β gene expression, B) Col-Iα gene expression, C) HGF genDiphenyleneiodonium. Data represent mean±S.D. (n=
^5). ⁎Pb
^0.05.
Please cite this article as: Salazar-Montes, A., et al., Potent antioxidant rodoi:10.1016/j.ejphar.2008.06.110
EDPRpresented nitrites accumulation with values of 2.5 µM/gr of tissue.
Treatmentwith Pirfenidone and Diphenyleneiodoniumdecreased to 1.2and 1.8 µM/gr respectively (Fig. 5B).
Malondialdehyde (MDA) is a degradation product of peroxidationof unsaturated fatty acid in cell membranes. MDA reacts in vivo and invitrowith free amino groups of proteins, phospholipids or nucleic acidsproducing modifications in their structures (Requena et al., 1997).Whenwe looked up for this oxidation metabolite in the liver of biliarycommon duct ligation
^rats administered with saline, these animals
presented 1.15 nmol MDA/μg protein representing five more times
Tand regeneration in biliary common duct ligation rats. Multiplex Real Time PCR wasormalized respect to housekeeping gene GAPDH according to User Bulletin No. 2 frome expression, D) c-met gene expression. Control: untreated, PFD: Pirfenidone, DPI:
le of Pirfenidone in experimental cirrhosis, Eur. J. Pharmacol. (2008),
C
ROOF
257
258
259
260
261
262
263
264
265
266
267
268
269
70
71
72
73
74
75
76
77
78
79
80
81
82
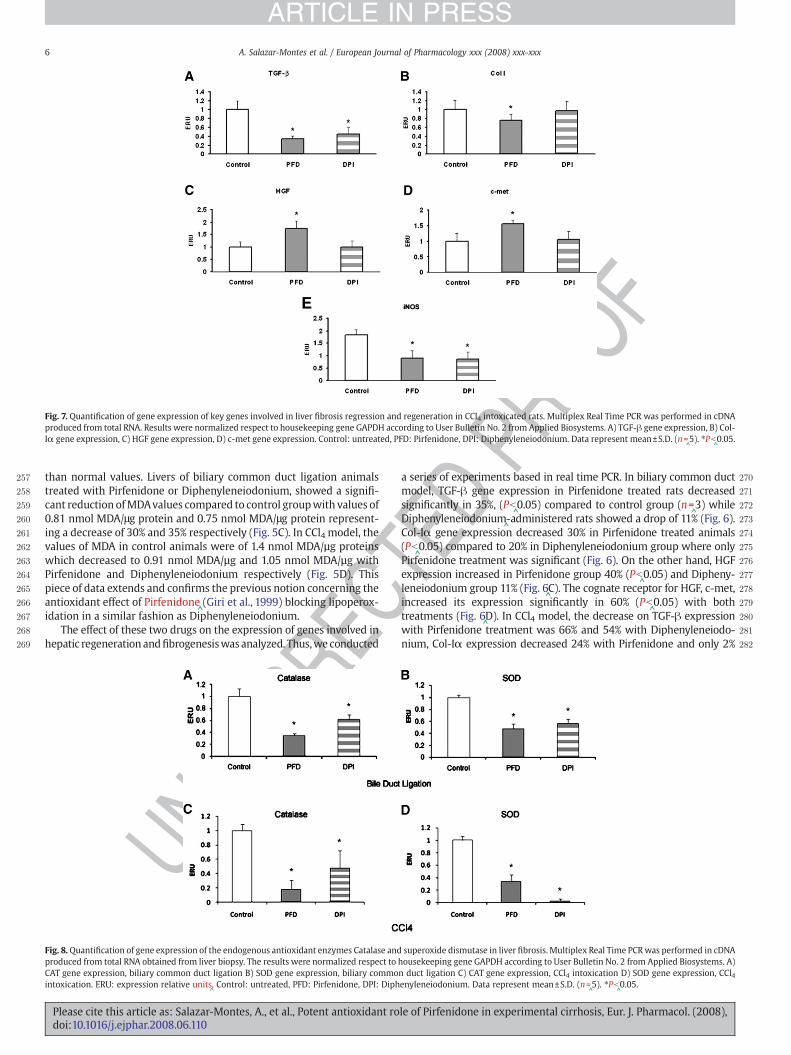
Fig. 7. Quantification of gene expression of key genes involved in liver fibrosis regression and regeneration in CCl4 intoxicated rats. Multiplex Real Time PCR was performed in cDNAproduced from total RNA. Results were normalized respect to housekeeping gene GAPDH according to User Bulletin No. 2 from Applied Biosystems. A) TGF-β gene expression, B) Col-Iα gene expression, C) HGF gene expression, D) c-met gene expression. Control: untreated, PFD: Pirfenidone, DPI: Diphenyleneiodonium. Data represent mean±S.D. (n=
^5). ⁎Pb
^0.05.
6 A. Salazar-Montes et al. / European Journal of Pharmacology xxx (2008) xxx-xxx
ARTICLE IN PRESS
than normal values. Livers of biliary common duct ligation animalstreated with Pirfenidone or Diphenyleneiodonium, showed a signifi-cant reduction ofMDAvalues compared to control groupwith values of0.81 nmol MDA/μg protein and 0.75 nmol MDA/μg protein represent-ing a decrease of 30% and 35% respectively (Fig. 5C). In CCl4 model, thevalues of MDA in control animals were of 1.4 nmol MDA/µg proteinswhich decreased to 0.91 nmol MDA/µg and 1.05 nmol MDA/µg withPirfenidone and Diphenyleneiodonium respectively (Fig. 5D). Thispiece of data extends and confirms the previous notion concerning theantioxidant effect of Pirfenidone
^(Giri et al., 1999) blocking lipoperox-
idation in a similar fashion as Diphenyleneiodonium.The effect of these two drugs on the expression of genes involved in
hepatic regeneration andfibrogenesiswasanalyzed. Thus,we conducted
UNCO
RRE
Fig. 8.Quantification of gene expression of the endogenous antioxidant enzymes Catalase andproduced from total RNA obtained from liver biopsy. The results were normalized respect toCAT gene expression, biliary common duct ligation B) SOD gene expression, biliary commointoxication. ERU: expression relative units
^. Control: untreated, PFD: Pirfenidone, DPI: Diph
Please cite this article as: Salazar-Montes, A., et al., Potent antioxidant rodoi:10.1016/j.ejphar.2008.06.110
TEDP 2a series of experiments based in real time PCR. In biliary common duct
2model, TGF-β gene expression in Pirfenidone treated rats decreased2significantly in 35%, (Pb
^0.05) compared to control group (n=
^3) while
2Diphenyleneiodonium^-administered rats showed a drop of 11% (Fig. 6).
2Col-Iα gene expression decreased 30% in Pirfenidone treated animals2(Pb
^0.05) compared to 20% in Diphenyleneiodonium group where only
2Pirfenidone treatment was significant (Fig. 6). On the other hand, HGF2expression increased in Pirfenidone group 40% (Pb
^0.05) and Dipheny-
2leneiodonium group 11% (Fig. 6^C). The cognate receptor for HGF, c-met,
2increased its expression significantly in 60% (Pb^0.05) with both
2treatments (Fig. 6^D). In CCl4 model, the decrease on TGF-β expression
2with Pirfenidone treatment was 66% and 54% with Diphenyleneiodo-2nium, Col-Iα expression decreased 24% with Pirfenidone and only 2%
superoxide dismutase in liver fibrosis. Multiplex Real Time PCRwas performed in cDNAhousekeeping gene GAPDH according to User Bulletin No. 2 from Applied Biosystems. A)n duct ligation C) CAT gene expression, CCl4 intoxication D) SOD gene expression, CCl4enyleneiodonium. Data represent mean±S.D. (n=
^5). ⁎Pb
^0.05.
le of Pirfenidone in experimental cirrhosis, Eur. J. Pharmacol. (2008),
OF
283
284
285
286
287
288
289
290
291
292
293
294
295
296
297
298
299
300
301
302
303
304
305
306
307
308
309
310
311
312
313
314
315
316
317
318
319
320
321
322
323
324
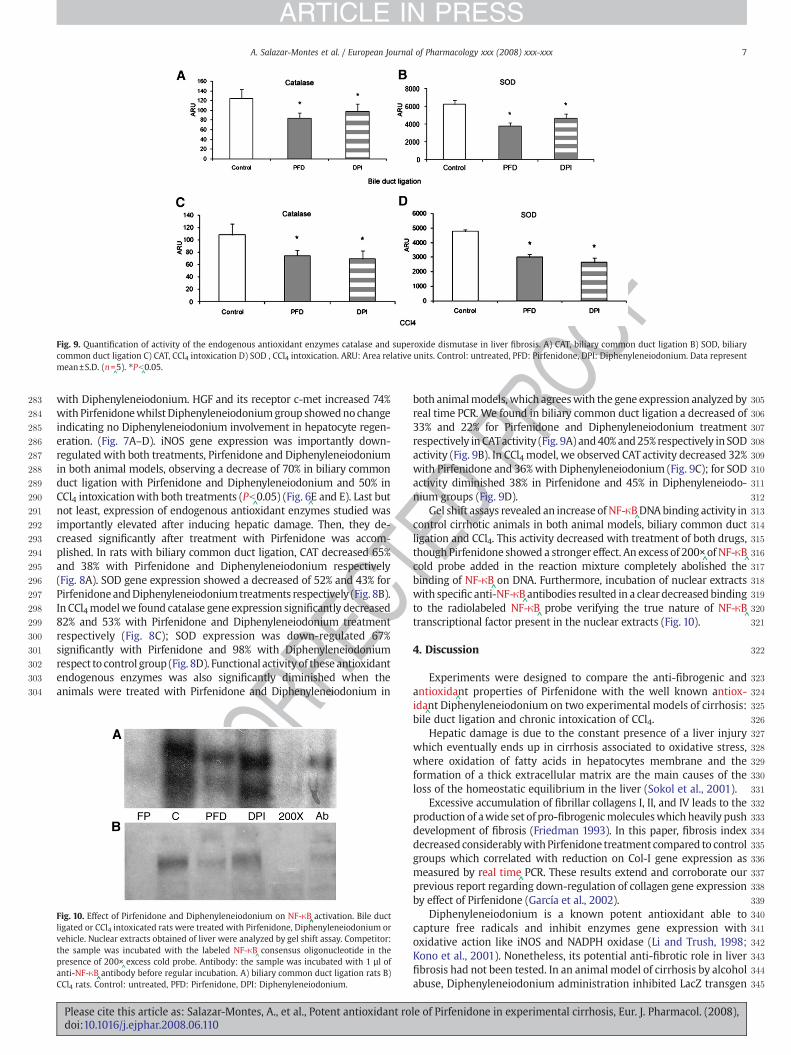
Fig. 9. Quantification of activity of the endogenous antioxidant enzymes catalase and superoxide dismutase in liver fibrosis. A) CAT, biliary common duct ligation B) SOD, biliarycommon duct ligation C) CAT, CCl4 intoxication D) SOD , CCl4 intoxication. ARU: Area relative units. Control: untreated, PFD: Pirfenidone, DPI: Diphenyleneiodonium. Data representmean±S.D. (n=
^5). ⁎Pb
^0.05.
7A. Salazar-Montes et al. / European Journal of Pharmacology xxx (2008) xxx-xxx
ARTICLE IN PRESS
REC
with Diphenyleneiodonium. HGF and its receptor c-met increased 74%with Pirfenidonewhilst Diphenyleneiodoniumgroup showednochangeindicating no Diphenyleneiodonium involvement in hepatocyte regen-eration. (Fig. 7A–D). iNOS gene expression was importantly down-regulated with both treatments, Pirfenidone and Diphenyleneiodoniumin both animal models, observing a decrease of 70% in biliary commonduct ligation with Pirfenidone and Diphenyleneiodonium and 50% inCCl4 intoxicationwith both treatments (Pb
^0.05) (Fig. 6
^E and E). Last but
not least, expression of endogenous antioxidant enzymes studied wasimportantly elevated after inducing hepatic damage. Then, they de-creased significantly after treatment with Pirfenidone was accom-plished. In rats with biliary common duct ligation, CAT decreased 65%and 38% with Pirfenidone and Diphenyleneiodonium respectively(Fig. 8A). SOD gene expression showed a decreased of 52% and 43% forPirfenidoneandDiphenyleneiodoniumtreatments respectively (Fig. 8B).In CCl4modelwe found catalase gene expression significantly decreased82% and 53% with Pirfenidone and Diphenyleneiodonium treatmentrespectively (Fig. 8C); SOD expression was down-regulated 67%significantly with Pirfenidone and 98% with Diphenyleneiodoniumrespect to control group (Fig. 8D). Functional activityof these antioxidantendogenous enzymes was also significantly diminished when theanimals were treated with Pirfenidone and Diphenyleneiodonium in
UNCO
R 325
326
327
328
329
330
331
332
333
334
335
336
337
338
339
340
341
342
343
344
345
Fig. 10. Effect of Pirfenidone and Diphenyleneiodonium on NF-κB^̂activation. Bile duct
ligated or CCl4 intoxicated rats were treated with Pirfenidone, Diphenyleneiodonium orvehicle. Nuclear extracts obtained of liver were analyzed by gel shift assay. Competitor:the sample was incubated with the labeled NF-κB
^consensus oligonucleotide in the
presence of 200×^excess cold probe. Antibody: the sample was incubated with 1 µl of
anti-NF-κB^̂antibody before regular incubation. A) biliary common duct ligation rats B)
CCl4 rats. Control: untreated, PFD: Pirfenidone, DPI: Diphenyleneiodonium.
Please cite this article as: Salazar-Montes, A., et al., Potent antioxidant rodoi:10.1016/j.ejphar.2008.06.110
TEDPR
Oboth animalmodels, which agreeswith the gene expression analyzed byreal time PCR. We found in biliary common duct ligation a decreased of33% and 22% for Pirfenidone and Diphenyleneiodonium treatmentrespectively in CATactivity (Fig. 9A) and40% and25% respectively in SODactivity (Fig. 9B). In CCl4 model, we observed CATactivity decreased 32%with Pirfenidone and 36%with Diphenyleneiodonium (Fig. 9C); for SODactivity diminished 38% in Pirfenidone and 45% in Diphenyleneiodo-nium groups (Fig. 9D).
Gel shift assays revealed an increase of NF-κB^DNAbinding activity in
control cirrhotic animals in both animal models, biliary common ductligation and CCl4. This activity decreased with treatment of both drugs,though Pirfenidone showed a stronger effect. An excess of 200×
^of NF-κB
^cold probe added in the reaction mixture completely abolished thebinding of NF-κB
^on DNA. Furthermore, incubation of nuclear extracts
with specific anti-NF-κB^antibodies resulted in a clear decreased binding
to the radiolabeled NF-κB^probe verifying the true nature of NF-κB
^transcriptional factor present in the nuclear extracts (Fig. 10).
4. Discussion
Experiments were designed to compare the anti-fibrogenic andantioxida
^nt properties of Pirfenidone with the well known antiox-
ida^nt Diphenyleneiodonium on two experimental models of cirrhosis:
bile duct ligation and chronic intoxication of CCl4.Hepatic damage is due to the constant presence of a liver injury
which eventually ends up in cirrhosis associated to oxidative stress,where oxidation of fatty acids in hepatocytes membrane and theformation of a thick extracellular matrix are the main causes of theloss of the homeostatic equilibrium in the liver (Sokol et al., 2001).
Excessive accumulation of fibrillar collagens I, II, and IV leads to theproduction of awide set of pro-fibrogenicmoleculeswhich heavily pushdevelopment of fibrosis (Friedman 1993). In this paper, fibrosis indexdecreased considerablywithPirfenidone treatment compared to controlgroups which correlated with reduction on Col-I gene expression asmeasured by real time
^PCR. These results extend and corroborate our
previous report regarding down-regulation of collagen gene expressionby effect of Pirfenidone (García et al., 2002).
Diphenyleneiodonium is a known potent antioxidant able tocapture free radicals and inhibit enzymes gene expression withoxidative action like iNOS and NADPH oxidase (Li and Trush, 1998;Kono et al., 2001). Nonetheless, its potential anti-fibrotic role in liverfibrosis had not been tested. In an animal model of cirrhosis by alcoholabuse, Diphenyleneiodonium administration inhibited LacZ transgen
le of Pirfenidone in experimental cirrhosis, Eur. J. Pharmacol. (2008),

C
346
347
348
349
350
351
352
353
354
355
356
357
358
359
360
361
362
363
364
365
366
367
368
369
370
371
372
373
374
375
376
377
378
379
380
381
382
383
384
385
386
387
388
389
390
391
392
393
394
395
396
397
398
399
400
401
402
403
404
405
406
407
408
409
410
411
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27 Q2
28
29
30
31
32
33
34
35
36
37
38
39404142434445464748495051525354555657585960616263646566676869707172737475767778798081
8 A. Salazar-Montes et al. / European Journal of Pharmacology xxx (2008) xxx-xxx
ARTICLE IN PRESS
UNCO
RRE
expression, probably due to the fact that, oxidative stress is necessaryfor transgen expression when the used vectors are adenoassociates(Wheeler et al., 2000). In our study, the principal effect of Dipheny-leneiodoniumwas observed on nitrites, MDA concentration, and iNOSgene expression, parameters significantly diminished compared tocontrol groups. We also observed a slight effect of Diphenyleneiodo-nium on Col-Iα gene expression which could be explained by itsprotector effect against oxidative stress damage preventing scarring. Inthis line of reasoning, TGF-β, the most potent inductor of fibrosis,decreased insignificantly with Diphenyleneiodonium treatment, sug-gesting that Diphenyleneiodonium has slight or no action on pro-fibrogenic molecules and its effect is mostly antioxidant. In contrastPirfenidone diminished significantly TGF-β gene expression confirm-ing its antifibrotic role reported previously for us (García et al., 2002).Regarding hepatic regeneration process, it is already known thathepatocyte proliferation inhibition, is associated with down-regula-tion of HGF and its cognate receptor, c-met (Schoedel et al., 2003).During Pirfenidone treatment, we observed an increase on HGF geneexpression suggesting the induction of hepatocyte proliferation andtherefore, liver regeneration (Shiota et al., 2000; Armendariz-Borundaet al., 2006).
A liver-protector effect of Pirfenidone was observed along with animprovement onmarkers of hepatic damage ALT, ASTand bilirubins inthese experimental groups.
In the other hand, iNOS expression is up-regulated during liverdamage when oxidative state is present (Gaudiot et al., 2000). In ourexperiments, iNOS expression decreased in the presence of bothantioxidant drugs showing Pirfenidone a protective effect similar toDiphenyleneiodonium. In the same context, the decreased activity andgene expression of the endogenous antioxidant enzymes SOD and CATwith both treatments indicate lower levels of oxidative stress intreated animals with respect to controls as is reported by Want T andothers (Wang et al., 2008, Battisti et al., 2008) where the use of severalantioxidants decreased endogenous antioxidant enzymes activity.These results confirm the previous reports for us and others on theantioxidant role of Pirfenidone and support its use in preventing liverdamage and liver fibrosis.
NF-κB^activity determined by gel-shift assay decreased in the
presence of both drugs, being more effective Pirfenidone in thisreduction. These results are consistent with previous reports whereNakanishi et al. (2004)
^reported an inhibition of NF-κB
^activity in
hepatocytes in the presence of Pirfenidone (Nakanishi et al., 2004..This inhibition correlates with iNOS expression decrease. Transfectionexperiments revealed that Pirfenidone decreased transcriptionalactivation of iNOS gene promoter. In addition, iNOS promoter hasconsensus sequence for NF-κB
^(Oh et al., 2007). This information
correlates with our results of iNOS and NF-κB^, and suggests that
Pirfenidonemight inhibit induction of iNOS gene expression via NF-κB^binding activity inhibition (Pinzani 2004).
The antioxidant effect of Pirfenidone is observed also as areduction of nitrite and MDA concentrations. MDA is a product ofpoly-unsaturated fatty acids of cell membranes oxidation which reactwith the free radical peroxinitrite originated by the reaction betweenanion superoxide and nitric oxide (Requena, 1997). Results gained inthis study for MDA concentrations are in harmony with previousreports in pulmonary fibrosis with Pirfenidone treatment where asignificant decrease on lipid peroxidation with Pirfenidone treatmentwas reported (Iyer et al., 1999).
In our study, MDA and nitrite^s levels match as lipid peroxidation
markers. The antioxidant effect observed of Pirfenidone and Dipheny-leneiodonium is probably due to iNOS expression inhibition, and inconsequence, oxide nitric production failed to decrease oxidative stress.
Furthermore, in this work we showed antioxidant properties ofPirfenidone above and beyond its well-known antifibrotic effect. Thebest antifibrogenic treatmentwould be capable of eliminating the causeof parenchymal damage, metabolic overload or excessive oxidative
Please cite this article as: Salazar-Montes, A., et al., Potent antioxidant rodoi:10.1016/j.ejphar.2008.06.110
TEDPR
OOF
4stress (Pinzani 2004). Along with this, cirrhosis is characterized by4oxidative stress, inflammation and fibrosis. The use of drugs for cirrhosis4treatmentmight be focused to attack the causes of fibrosis like oxidative4stress and inflammation preventing fibrosis development. Thus, use of4drugs like Pirfenidone and Diphenyleneiodonium might be adequate.4Nevertheless, if it is an establishedfibrosis it will be necessary to remove4the extracellular matrix besides to prevent the liver damage caused by4the oxidative stress and inflammation. In this context, Pirfenidone have4these two properties: prevents fibrosis due to its antioxidant and anti-4inflammatory actions, and attacks fibrosis because turn on and turn off4genes are involved in fibrosis synthesis. Because Pirfenidone presents4both features, this is, preventive and curative drug, represents an4attractive drug for trying fibrotic diseases accompanied by an oxidative4process like many degenerative chronic diseases, i.e. renal fibrosis,4cardiac diseases and hepatic cirrhosis (Pinzani 2004).
45. Uncited reference
4http://dna-9.int-med.uiowa.edu/RealtimePCRdocs/Bulletin 24Applied Byosistem
4Acknowledgments
4The present work was supported by COECyTJal grant # 08-2004 to4Juan Armendariz-Borunda. The authors are indebted to Dr. Pedro Diaz4for providing care to the experimental animals, to Nuclear Medicine4Department from Hospital Civil de Guadalajara “Fray Antonio Alcalde”4for the facilities to use radioactivity and to Rosa Lina Torres-Rodríguez4fromHospital Civil of Guadalajara “Juan IMenchaca” for their invaluable4technical help in hepatic function test determination.
4References
4Andrews, N.C., Faller, D.V., 1991. A rapid micropreparation technique for extraction of4DNA binding proteins from limiting numbers of mammalian cells. Nucleic Acids4Res. 19, 2499.4Armendariz-Borunda, J., Simkevich, C.P., Roy, N., Raghow, R., Kang, A.H., Seyer, J.M.,1994.4Activation of Ito cells involves regulation of AP-1 binding proteins and induction of4type I collagen gene expression. Biochem. J. 304, 817–824.4Armendáriz-Borunda, J., Islas-Carvajal, M., Meza, E., Rincón, A.R., Alvarez, A., Goodman,4Z.D., Sandoval, A.S., Covarrubias, A., Aréchiga, G., Garcia, L., 2003. A pilot study of a4novel antiinflamatory and anti-fibrotic agent, Pirfenidone, in patients with liver4cirrhosis. Hepatology 38, 308A.4Armendáriz-Borunda, J., Islas-Carvajal, M.C., Meza-García, E., Rincón, A.R., Sandoval, A.S.,4Salazar, A., Berumen, J., Alvarez, A., Alvarez, A., Covarrubias, A., Aréchiga, G., García,4L., 2006. A pilot study in cirrhotic patients using a new anti-inflammatory and anti-4fibrotic agent, Pirfenidone. Gut. 551, 663–665.4Battisti, V., Maders, L.D., Bagatini, M.D., Santos, K.F., Spanevello, R.M., Maldonado, P.A.,4Brulé, A.O., Araújo, M.D., Schetinger, M.R., Morsch, V.M., 2008. Measurement of4oxidative stress and antioxidant status in acute lymphoblastic leukemia patients.4Clin. Biochem. 41, 511–518.4Bonnand, A.M., Heathcote, E.J., Lindor, K.D., Poupon, R.E., 1999. Clinical significance of4serum bilirubin levels under ursodeoxycholic acid therapy in patients with primary4biliary cirrhosis. Hepatology 29, 39–43.4Bradford, M.M., 1976. A rapid and sensitive method for the quantification of microgram4quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem.472, 248–254.4Britton, R.S., Bacon, B.R., 1999. Intracellular signaling pathways in stellate cell activation.4Alcohol. Clin. Exp. Res. 23, 922–925.4Bueno, R., Daneri, A., Armendariz-Borunda, J., 2000. Cholestiasis-induced fibrosis is4reduced by interferon α-2a and is associated with elevated liver metalloprotease4activity. J. Hepatol. 33, 915–925.4Burke, J.J., Oliver, M.J., 1992. Differential temperature sensitivity of pea superoxide4dismutases. Plant. Physiol. 100, 1595–1598.4Callaway, J.K., Beart, P.M., Jarott, B., 1998. A reliable procedure for comparison of4antioxidants in rat brain homogenates. J. Pharmacol. Toxicol. Methods 39, 155–162.4Chomzynsky, P., Sacchi, N., 1987. Single-stepmethod of RNA isolation by acid guanidium4thiocyanate-phenol-cloroform extraction. Annal. Biochem. 162, 156–159.4Flora, S.J., 2007. Role of free radicals and antioxidants in health and disease. Cell. Mol.4Biol. 53, 1–2.4Friedman, S.L.,1993. The cellular basis of hepatic fibrosis. N. Engl. J. Med. 328,1828–1835.4Friedman, S.L., 2000. Molecular regulation of hepatic fibrosis an integrated cellular4response to tissue injury. J. Biol. Chem. 275, 2247–2250.4Gancedo, M., Ruiz-Corro, L., Salazar-Montes, A., Rincón, A.R., Armendáriz-Borunda, J.,42008. Pirfenidone prevents capsular contracture post mammary implantation.4Aesthetic. Plast. Surg. 32, 32–40.
le of Pirfenidone in experimental cirrhosis, Eur. J. Pharmacol. (2008),

482483484485486487488489490491492493494495496497498499500501502503504505506507508509510511512513514515516517518
519520521522523524525526527528529530531532533534535536537538539540541542543544545546547548549550551552553
554
555
9A. Salazar-Montes et al. / European Journal of Pharmacology xxx (2008) xxx-xxx
ARTICLE IN PRESS
Garcia, L., Hernandez, I., Sandoval, A., Salazar, A., Garcia, J., Vera, J., Grijalva, G., Muriel, P.,Margolin, S., Armendariz-Borunda, J., 2002. Pirfenidone effectively reversesexperimental liver fibrosis. J. Hepatol 37, 797–805.
Gaudiot, N., Ribiere, C., Jaubert, A.M., Giudicelli, Y., 2000. Endogenous nitric oxide isimplicated in the regulation of lipolysis through antioxidant related effect. Am.J. Physiol. 279, 1603–1610.
Giri, S.N., Leonard, S., Shi, X.,Margolin, S.B., Vallyathan, V.,1999. Effect of Pirfenidone on thegeneration of reactive oxygen species in vitro. J. Environ. Pathol. Toxicol. Oncol. 18,169–177.
http://dna-9.int-med.uiowa.edu/RealtimePCRdocs/Bulletin 2 Applied Byosistem.Iyer, S.N., Gurujeyalakshmi, G., Giri, S.N., 1999. Effects of pìrfenidone on transforming
growth factor-beta gene expression at the transcriptional level in bleomycinhamster model of lung fibrosis. J. Pharmacol. Exp. Ther 291, 367–373.
Kono, H., Rusyn, I., Uesugi, T., Yamashina, S., Connor, H., Dikalova, A., Mason, R.,Thurman, R., 2001. Diphenyleneiodonium sulfate, an NADPH oxidase inhibitor,prevents early alcohol-induced liver injury in the rat. Am. J. Physiol. Gastrointest.Liver. Physiol. 280, G1005–G1012.
Li, Y., Trush, M.A., 1998. Diphenyleneiodonium, an NAD(P)H oxidase inhibitor, alsopotently inhibits mitochondrial reactive oxygen species production. Biochem.Biophys. Res. Commun. 253, 295–299.
Manchenko, G.P., 1994. Handbook of Detection of Enzymes on Electrophoretic Gels. CRCPress, p. 92.
Nakanishi, H., Kaibori, M., Teshima, S., Yoshida, H., Kwon, A.H., Kamiyama, Y., Nishizawa,M., Ito, S., Okumura, T., 2004. Pirfenidone inhibits the induction of iNOS stimulatedby interleukin-1 beta at a step of NF-kappa B DNA binding in hepatocytes.J. Hepatol. 41, 730–736.
Oh, I., Ozaki, K., Sato, K., Meguro, R., Tatara, R., Hatanaka, K., Nagai, T., Muroi, K., Ozawa,K., 2007. Interferon g and NF-κB
^mediate nitric oxide production by mesenchymal
stromal cells. Biochem. Biophys. Res. Commun. 355, 956–962.Pinzani, M., 2004. Therapies for hepatic fibrosis: real hope or just academic exercise?
Dig. Liver. Dis 36, 714–716.Poli, G., 2000. Pathogenesis of liver fibrosis: role of oxidative stress. Mol. Aspects. Med.
21, 49–98.Requena, J.R., Fu, M.X., Ahmed, M.U., Jenkins, A.J., Lyons, T.J., Baynes, J.W., Thorpe, S.R.,
1997. Quantification of malondialdehyde and 4-hydroxynonenal adducts to lysineresidues in native and oxidized human low-density lipoprotein. Biochem. J. 322,317–325.
UNCO
RREC
Please cite this article as: Salazar-Montes, A., et al., Potent antioxidant rodoi:10.1016/j.ejphar.2008.06.110
ROOF
Salgado, S., García, J., Vera, J., Siller, F., Bueno, M., Miranda, A., Armendáriz-Borunda, J.,2000. Liver cirrhosis is reverted by urokinase-
^type plasminogen activator gene
therap^y. Mol. Ther. 2, 545–551.
Schoedel, K.E., Tyner, V.Z., Kim, T.H., Michalopoulos, G.K., Mars, W.M., 2003. HGF, metand matrix-related proteases in hepatocellular carcinoma, fibrolamellar variant,cirrhotic and normal liver. Mod. Pathol. 16, 14–21.
Schwart, D., Mendonca, M., Schwartz, I., Xia, Y., Satriano, J., Wilson, C., Blantz, R., 1997.Inhibition of constitutive nitric oxide synthase (NOS) by nitric oxide generated byinducible NOS after lipopolysaccharide administration provokes renal dysfunctionin rats. J. Clin. Invest. 100, 439–448.
Shimizu, T., Kuroda, T., Hata, S., Fukagawa, M., Margolin, S.B., Kurokawa, K., 1998.Pirfenidone improves renal function andfibrosis in thepost-obstructedkidney. KidneyInt. 54, 99–109.
Shiota, G., Kunisada, T., Oyama, K., Udagawa, A., Nomi, T., Tanaka, K., Tsutsumi, A., Isono,M., Nakamura, T., Hamada, H., Sakatani, T., Sell, S., Sato, K., Ito, H., Kawasaki, H., 2000.In vivo transfer of hepatocyte growth factor gene accelerate proliferation oval cells ina 2-acetylaminofluorene/partial hepatectomy model rats. FEBS Lett. 470, 325–330.
Sokol, R.J., 2002. Liver cell injury and fibrosis. J. Pediatr. Gastroenterol. Nutr. 35, 7–10.Sokol, R., Straka, M., Dahl, R., Devereaux, M.W., Verushalmi, B., Gumpricht, E., Elkins, N.,
Everson, G., 2001. Role of oxidant stress in the permeability transition induced in rathepatic mitochondria by hydrophobic bile acids. Pediatr. Res. 49, 519–531.
Tada, S., Nakamuta, M., Enjoji, M., 2001. Pirfenidone inhibits dimethylnitrosamine-induced hepatic fibrosis in rats. Clin. Exp. Pharmacol. Physiol. 28, 522–527.
Tapia, G., Pepper, I., Smok, G., Videla, L.A., 1997. Kupffer cell function in thyroid hormoneinduced liver oxidative stress in the rat. Free Rad. Res 26, 267–279.
Tsukamoto, H., Rippe, R., Niemela, O., Lin, M., 1995. Roles of oxidative stress in activationof Kupffer and Ito cells in liver fibrogenesis. J. Gastroenterol. Hepatol. 10, S50–S53.
Vrba, J., Mondriasky, M., 2002. Oxidative burst of Kupffer cells: target for liver injurytreatment. Biomed. Pap.Med. Fac. Univ. Palacky. Olomouc. Czech. Repub.146,15–20.
Wang, T., Sun, N.L., Zhang, W.D., Li, H.L., Lu, G.C., Yuan, B.J., Jiang, H., She, J.H., Zhang, C.,2008. Protective effects of dehydrocavidine on carbon tetrachloride-induced acutehepatotoxicity in rats. J. Ethnopharmacol. 117, 300–308.
Wheeler, M.D., Kono, H., Rusyn, I., Arteel, G.E., McCarty, D., Samuliski, R.J., Thurman, R.,2000. Chronic ethanol increases adeno-associated viral transgene expression in ratliver via oxidant and NFκB-dependent mechanisms. Hepatology 32, 1050–1059.
P TEDle of Pirfenidone in experimental cirrhosis, Eur. J. Pharmacol. (2008),





























