Embed Size (px)
Citation preview

Behavioral Ecology Vol. 9 No. 4: 360-366
Presampling sensory information and preydensity assessment by wolf spiders(Araneae, Lycosidae)
Matthew H. Persons and George W. UetzDepartment of Biological Sciences, University of Cincinnati, Cincinnati, OH 45221-0006, USA
Decisions regarding foraging patch residence time and the assessment of patch quality may be mediated by various sources ofinformation. This study examined the use of sensory cues by hunting spiders to assess prey density in the absence of preycapture. Adult female wolf spiders [Schixocosa ocmeata (Hentz); Lycosidae] had food withheld for 4 days and then were exposedto artificial foraging patches containing four densities of crickets (0, 3,10, 20) with different sensory stimuli (visual and vibratoryinformation, visual only, and vibratory only). The spiders were not allowed to feed during trials, and patch residence time wasrecorded. The spiders varied patch residence time based on sensory cues alone and spent more time in patches with higherprey density. With visual information only, spiders could apparently distinguish among prey densities almost as well as with visualand vibratory cues combined, but residence time did not differ among prey densities when only vibratory information waspresented. Measurements of vibration levels produced by cricket activity under experimental sensory treatments conform to testresults, suggesting that visual detection of crickets is important in patch assessment used in determining patch residence time.Key words: patch quality, prey density, residence time, Schixocosa ocreata, sensory information, wolf spiders. [Behav Ecol 9:360-366 (1998)]
How animals assess foraging patch quality and determinehow long to remain in a patch before leaving has been
of considerable interest in foraging theory (Charnov, 1976;Cook and Hubbard, 1977; Iwasa et al., 1981; Oaten, 1977;Pyke, 1984). Many species of sit-and-wait predators remainlonger in patches where encounters with prey are more fre-quent (Janetos, 1982; Turnbull, 1964; Zhang and Sanderson,1993), but the proximate mechanisms mediating the decisionto leave or stay needs further investigation because the typeand accuracy of information used to make foraging decisionsmay affect predictions of patch residence time and patchchoice (Morse, 1989).
For predators, each stage of the predation process consti-tutes a b«tter estimator of prey and patch quality. Endler(1991) outlined six stages of predation: encounter, detection,identification, approach, subjugation, and consumption. Var-ious forms of information are gathered at different stages ofthis process, including search time, prey density, subjugationand handling costs, prey palatabiHty, and energy value. Themajority of empirical and theoretical foraging studies havelargely emphasized the foraging decisions of animals as a re-sult of cumulative information gathered through the laststages of predation (subjugation and consumption). However,animals may make foraging decisions with less information,particularly if the time costs associated with increased accuracyoutweigh the value of the information.
Sources of information used in assessing foraging patcheshave been largely divided into two types: presampling andsampling information. Although most sources of sampling in-formation include some measure of prey capture rate per unittime (Stephens and Krebs, 1986), individuals may also esti-mate patch quality before exploitation. Presampling infor-
Address correspondence to M. H. Persons, who is now at the De-partment of Zoology, Miami Univenitjr, Oxford, OH 45056, USA. E-maiL personmhftmuohio.edu.
Received 14 February 1997; first revision 19 August 1997; secondrevision 9 December 1997; accepted 22 December 1997.O 1998 International Society for Behavioral Ecology
mation includes sensory cues, memory of patch location andquality from previous patch sampling,, or information on therelative distribution of resources within patch subtypes (Baye-sian foraging) (Valone, 1991). Foragers that use sensory cuesor memory in predicting patch quality in spatially or tempo-rally variable environments are known as prescient foragers(Valone arid Brown, 1989).
Several mechanisms for the assessment of patch quality viasampling have been suggested, and most fall under one ofseveral categories suggested by Formanowicz and Bradley(1987): (1) prey capture rates or intercapture interval (Char-nov, 1976); (2) gut fullness or other postingestion feedbackmechanisms (DeBenedictis et al., 1978; Johnson et aL, 1975;Nakamura, 1968; Provenzaand Cincotta, 1993; Schuler, 1990);and (3) encounter rate with prey (Hughes, 1979; Pulliam,1974). The type of sampling information used to make for-aging decisions may significantly affect predictions of patchresidence time or other foraging decisions.
Other empirical and theoretical foraging studies have ex-amined the use of presampling sources of information includ-ing public information (Iivoreil and Giraldeau, 1997; Tem-pleton and Giraldeau, 1995), prior patch experience (Valone,1992), or the interaction of sampling and presampling infor-mation (Bayesian foraging) (Iwasa et aL, 1981; Mangel andClark, 1983; McNamara and Houston, 1980, 1982, 1987; Oat-en, 1977), but few studies have examined the use of sensoryinformation specifically (Cordon and Bell, 1991; Marden,1984; Morse, 1989). The majority of empirical treatments ofinformation use, residence time, and patch assessment haveused avian models (Krebs et aL, 1974, 1978; Nishimura,1991a,b; Nishimura and Abe, 1988; Pyke, 1978b; Templetonand Giraldeau, 1995; Valone, 1991; Wolf and Hainsworth,198*t ZasfeandEsuls, 1976) or Hymenoptera (Heinrich, 1979,1983; Hodges, 1981; Pyke. 1978a, 1980), both of wnieh havebeen found to use prior patch experience under some cir-cumstances.
Wolf spiders (Schixocosa ocrtata) have not been demonstrat-ed to base residence times in food patches on prior feedingexperience (Persons and Uetz, 1997b) or prior sensory infor-
by guest on June 6, 2012http://beheco.oxfordjournals.org/
Dow
nloaded from

Persons and Ueti • Prey density assessment by wolf spiders 361
mation (Persons and Uetz, 1996a), and have been found tomove randomly between patches that differ in the presenceof prey (Persons and Uetz, 1996a). Studies show that wolf spi-der patch residence decisions may be based on the intensityof current sensory stimuli without using prey capture, feedinginformation, or prior patch experience. Although sensorycues alone may be of central importance in determining res-idence time, almost no study has separated this mechanismexplicitly from energy gain. If an animal uses sensory assess-ment rather than energy gain to make residence time deci-sions, it should vary its patch time with different numbers offood items in a patch, even if no feeding occurs. In previousstudies, we demonstrated that wolf spiders vary their patchresidence time based on different types of sensory informa-tion in the absence of feeding (Persons and Uetz, 1996a,b).Sensory cues from prey result in longer foraging patch resi-dence time for 5. ocrtata than when prey are captured in apatch (Persons and Uetz, 1997b). Studies have also found pre-feeding cues to be important determinants of choosing for-aging sites among orb weaving spiders (Pasquet et al., 1994)funnel-web spiders (Riechert, 1985), and crab spiders (Morse,1993), indicating that sensory cues may serve as a better pre-dictor of foraging behavior than feeding among other spidersas well. Further, patch residence time in spiders may be de-termined more by prey perception than the more commonlyassumed feeding rates or prior patch experience that havebeen studied in birds. We examined the influence of two sen-sory channels, visual and vibratory, in the absence of feeding,on patch residence time of S. ocreata under different preydensities.
METHODS
Study species
Schizocosa ocreata (Araneae, Lycosidae), like most wolf spiders,is a mobile adt-and-wait forager with frequent changes in for-aging site (Cady, 1984; Ford, 1978) and does not build websto snare prey. These ground-dwelling arthropods are foundin complex leaf litter of deciduous forests in the eastern Unit-ed States. Their foraging behavior has been studied largelywithin the context of distribution and population dynamics(Cady, 1984; Wise, 1993; Wise and Wagner, 1992). Some wolfspiders locate and attack prey using both substratum-coupledvibrations and visual cues (lizotte and Rovner, 1988; Personsand Uetz, 1996a), but the threshold level of different types ofsensory stimuli necessary to elicit an increase in residencetime is unknown. Under natural conditions, these spiders fre-quently are presented with only visual or vibratory cues fromprey. Prey vibrations may be transmitted only the distance ofa single leaf (Scheffer et. a]., 1996), but spiders are able toperceive prey visually at greater distances (T. Valerius and J.Renneker, personal communication). Spiders may also be lo-cated on the opposite side of the same leaf that a potentialprey item occupies, where it can detect vibrations from itsmovement but cannot see the prey.
Spider collection and twi
We caught 28 immature female S. ocrtata in April 1993 at ther.inHnnari Nature Center, Qermont County, Ohio, USA. Eachspider was housed in its own opaque container, provided wa-ter ad libitum, and fed three 1-week-old cricket nymphs every4 days to standardize hunger level for testing. We kept allspiders under identical controlled conditions at room tem-perature (23-25°C) in an environment with stable humidityand a 12:12 h lightdark photoperiod. The spiders were al-lowed to mature before being subjected to experiments.
Experimental protocol
We conducted three experiments. The first experiment testedif spiders modified their patch residence time in the presenceof increasing density of prey without prey consumption orcapture but with both visual and vibratory information. Thesecond and third experiments tested if spiders altered theirpatch residence time when visual or vibratory informationalone was present to determine if sensory channels were useddifferentially or if the spider's accuracy to perceive differencesin density varied. Thus, each spider was presented with fourdensity treatments (0, 3, 10, 20 crickets) and diree sensorytreatments (visual and vibratory information, visual informa-tion alone, and vibratory alone) in a fully crossed (non-nest-ed) design. We analyzed each experiment separately.
The experimental apparatus controlled for visual and vibra-tory cues used in prey detection (Figure 1). The test apparatusfor the first experiment (visual and vibratory information)consisted of four containers made of white foam-core board.Each container housed two round chambers 20 cm in diam-eter with transparent 0.08-mm acetate walls. One chamberserved as a neutral chamber into which the spider was intro-duced, and the other chamber contained cricket stimuli (sen-sory chamber). In the sensory chamber, behind each acetatewall, crickets were introduced at four prey densities: 0, 3, 10,and 20. We randomly assigned each spider to a container andtested it under all four density levels. For the second experi-ment, only visual information was presented to the spider.This treatment had the cricket enclosure mounted on a sep-arate foam-core block from that of the spider so that vibra-tions could not be transmitted to the substratum that the spi-der occupied. The third experiment presented only vibratoryinformation, as the acetate wall separating the spider andcrickets was covered with an opaque paper barrier.
An experimental trial consisted of a single spider intro-duced into the neutral (no cricket stimuli) chamber under aclear plastic vial. Before testing, spiders were maintained onthree crickets every 4 days. All spiders tested were used afterhaving prey witheld for 4 days and were therefore under sim-ilar hunger levels. After a 5-min acclimation period, the vialwas removed and the spider was allowed to enter and exit thesingle sensory chamber freely for 30 min. We videotaped eachtrial from above and determined duration and number ofchamber visits by analyzing the videotape. Clean white paperwas placed on die floor of each chamber of the apparatusbefore each trial. The chambers were also swabbed with dean,dry cotton between trials to remove any silk draglines fromprevious spiders that might affect subsequent foraging behav-ior. We tested each spider under each density treatment andall three types of sensory stimuli.
Statistical methods and analysis
Many spiders fail to move into a sensory chamber at all duringan entire trial. Others do not visit all density treatment sen-sory chambers at least twice, which is required for an ANOVAanalysis of individual variation and interactions with prey den-sity effects (because there is no replication within the individ-ual factors). Therefore, for statistical reasons, we decided apriori to analyze only residence times of spiders that visitedeach density treatment and sensory treatment at least fourtimes. This produced a more balanced data set for ease ofinterpreting statistical results (Shaw and Mitchell-Olds, 1993).The last visit into a sensory chamber was omitted from analysisif the spider was in the chamber when the trial time expiredbecause including it would underestimate the true patch res-idence time. Only IS of the 28 spiders tested matched allanalysis criteria, and although it may have biased the data set
by guest on June 6, 2012http://beheco.oxfordjournals.org/
Dow
nloaded from

362 Behavioral Ecology Vol. 9 No. 4
Figure 1The artificial foraging environ-ment used for spider testing.Spiders are placed in the "neu-tral" chamber before each ex-periment (spider* shown instimulus chambers) and al-lowed to move freely betweenthe two chambers after a 5-minacclimation period. Usingcricket nymphs as stimuli, eachpair of chambers differs in thedensity treatments presentedto the spider. From left, thetreatments are no crickets, 3,10, and 20 crickets. Figure rep-resents visual and vibratorystimuli presented. Other sen-sory treatments include visualstimuli only and vibratory stim-uli only.
toward more mobile spiders, this procedure did allow morereliable interpretation of the analysis of variance.
For each experiment, we used a fully crossed, mixed-modeltwo-way ANOVA to analyze the variation in duration of patchvisits. Each of the three experiments was analyzed separately.Patch residence time was natural log transformed to conformto ANOVA assumptions of normality. The primary parameter,patch residence time, was tested using individual (random ef-fects) and density (fixed effects) as categorical variables. TheF ratio for prey density effect was constructed with the inter-action term mean squares in the denominator (Zar, 1984) forthe appropriate F ratio for a mixed model. We used a mixed-model ANOVA in this analysis because it is more conservativethan a repeated-measures analysis and accurately accounts forindividual spiders as a random effect (see Bennington andThayne, 1994). All computations were performed with a main-frame version of SAS software (version 6.07). Differencesamong individual spiders are likely to occur and may affectinferences that may be drawn about patch assessment abilitiesof individuals when treating them as groups. This design over-comes this problem (Martin and Kraemer, 1987). We usedrepeated visits of individual spiders as replicates for the indi-vidual factor. Repeated visits by an individual spider have beentested statistically for independence in previous studies ofpatch residence using an identical or similar test apparatus(Persons and Uetz, 1996a, 1997a,b). Spiders have shown noevidence that prior experience biases their residence time(Persons and Uetz, 1996a, 1997a,b), and lengths of sequentialvisits to the same patch are not correlated (Persons and Uetz,1996a, 1997a, b). Thus, sequential visits by spiders to a partic-ular aeatmont wexe used as replicates for the individual spi-der factor as a measure of individual variation.
Despite the apparent independence of sequential visits intoa chamber by an individual spider, use of sequential visits maybe construed as pseudoreplication (Hurlbcrt, 1984). To avoidthis problem, we conducted two other analyses of the data.Individual variation was collapsed to a mean value for each ofthe 13 spiders and a one-way ANOVA was done for each of
the sensory experiments (vibratory only, visual only, visual andvibratory together). A Tukey post-hoc comparison of meansanalysis determined differences in density treatments withineach experiment. To examine more closely an individual spi-der's ability to perceive differences in patch quality, we con-ducted one-way ANOVAs for each individual spider to deter-mine significant differences in residence time for each densitytreatment in the presence of visual and vibratory cues fromprey.
Vibration testing
Vibration levels were expected to increase with cricket density.Vibration levels for each sensory and density treatment weremeasured in decibels using a Bruel and Kjaer accelerometer(type 4366) high-sensitivity vibration pickup, loaded to a Brueland Kjaer sound level meter (type 2203). Dedbel level wasmeasured every 15 s for 30 measurements under each exper-imental treatment. We placed the accelerometer along theedge of each chamber nearest the crickets because most spi-ders had a strong tendency to move along die edges of thechambers. A one-way ANOVA was performed and a Tukeypost-hoc comparison of means test was used to distinguishbetween all sensory and density treatments for the stimulusand neutral chambers. We compared 24 chambers: 3 sensorytreatments with 4 density treatments each, and 2 chambers(neutral and stimulus) for each density.
RESULTS
Patch residence times of spiders varied significantly with preydensity aad type of sensory information (Figure 2). Whenboth visual and vibratory information were presented to thespider, significant density effects were found (.Fj,» = 18.34, p=• .0001) as well as significant variation among individual spi-ders (Fa u 7 - 5.57, p - .0001; Table 1). Using individualspiders as die level of replication and a mean value of resi-dence time for each density treatment, spiders spent signifi*
by guest on June 6, 2012http://beheco.oxfordjournals.org/
Dow
nloaded from
Persons and Uetz • Prey density assessment by wolf spiders
BOO
500
400
300
200
100
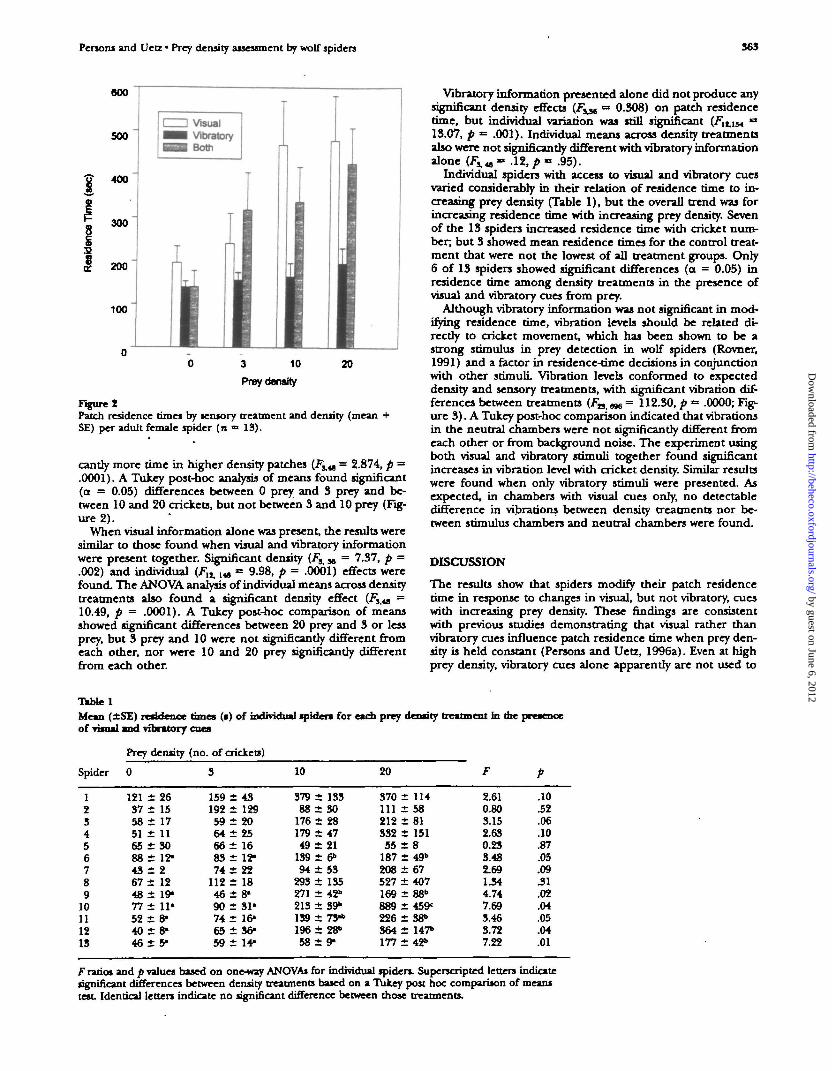
Figure ZPatch residence times by sensory treatment and density (meanSE) per adult female spider (n => 13).
candy more time in higher density patches (F^^ = 2.874, p =.0001). A Tukey post-hoc analysis of means found significant(a = 0.05) differences between 0 prey and 3 prey and be-tween 10 and 20 crickets, but not between 3 and 10 prey (Fig-ure 2).
When visual information alone was present, the results weresimilar to those found when visual and vibratory informationwere present together. Significant density (F^ x = 7.37, p =.002) and individual (Flit lia = 9.98, p = .0001) effects werefound. The ANOVA analysis of individual means across densitytreatments also found a significant density effect (FSM =10.49, p = .0001). A Tukey post-hoc comparison of meansshowed significant differences between 20 prey and 3 or lessprey, but 3 prey and 10 were not significantly different fromeach other, nor were 10 and 20 prey significantly differentfrom each other.
Vibratory information presented alone did not produce anysignificant density effects ( f j j , = 0.308) on patch residencetime, but individual variation was still significant (Flt,iM »13.07, p = .001). Individual means across density treatmentsalso were not significantly different with vibratory informationalone (Fxv ™ .12, p = .95).
Individual spiders with access to visual and vibratory cuesvaried considerably in their relation of residence time to in-creasing prey density (Table 1), but the overall trend was forincreasing residence time with increasing prey density. Sevenof the 13 spiders increased residence time with cricket num-ber; but 3 showed mean residence times for the control treat-ment that were not the lowest of all treatment groups. Only6 of 13 spiders showed significant differences (a = 0.05) inresidence time among density treatments in the presence ofvisual and vibratory cues from prey.
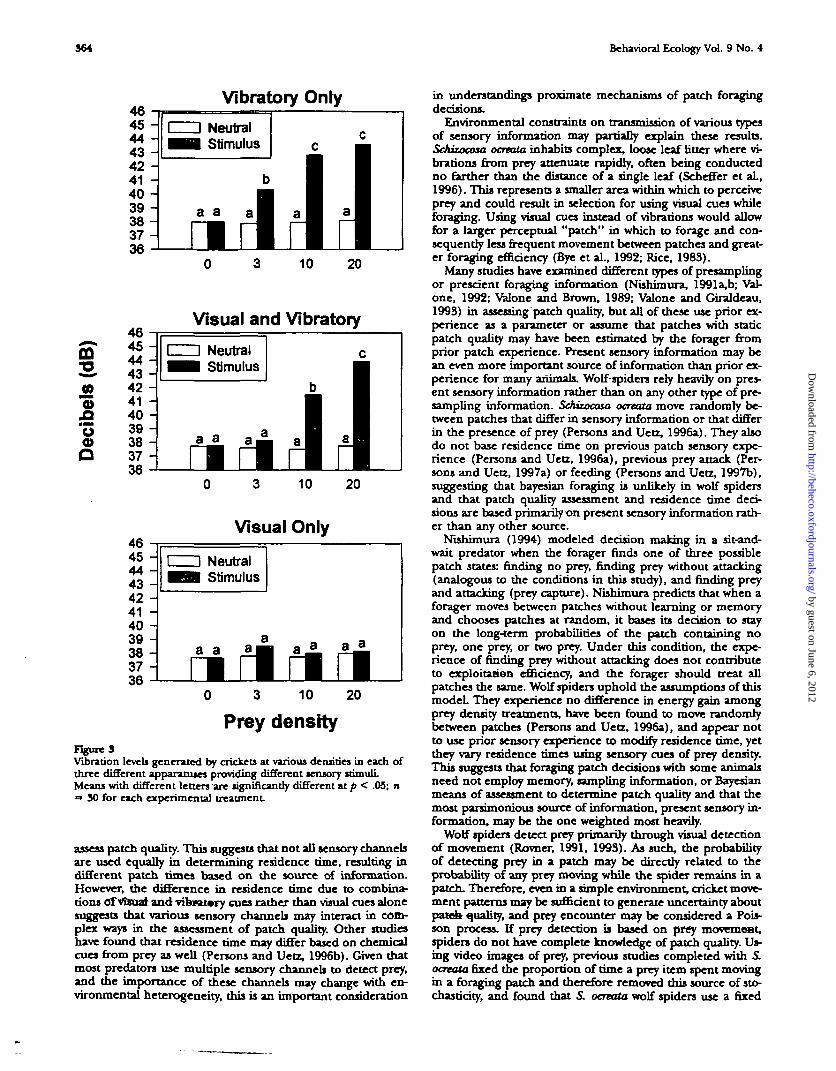
Although vibratory information was not significant in mod-ifying residence time, vibration levels should be related di-rectly to cricket movement, which has been shown to be astrong stimulus in prey detection in wolf spiders (Rovner,1991) and a factor in residence-time decisions in conjunctionwith other stimuli Vibration levels conformed to expecteddensity and sensory treatments, with significant vibration dif-ferences between treatments (FBet6 = 112.30, p = .0000; Fig-ure 3). A Tukey post-hoc comparison indicated that vibrationsin the neutral chambers were not significantly different fromeach other or from background noise. The experiment usingboth visual and vibratory stimuli together found significantincreases in vibration level with cricket density. Similar resultswere found when only vibratory stimuli were presented. Asexpected, in chambers with visual cues only, no detectabledifference in vibrations between density treatments nor be-tween stimulus chambers and neutral chambers were found.
DISCUSSION
The results show that spiders modify their patch residencetime in response to changes in visual, but not vibratory, cueswith increasing prey density. These findings are consistentwith previous studies demonstrating that visual rather thanvibratory cues influence patch residence time when prey den-sity is held constant (Persons and Uetz, 1996a). Even at highprey density, vibratory cues alone apparently are not used to
Table 1Mean (±SE) reridence times (•) of individual spiden for each prey density treatment in die presenceof visual and vibratory cues
Prey density (no. of crickets)
Spider
I23456789
10111213
0
121 237 258 251 265 288 243 d67 24 8 277 252 240 246 2
t 26: 15i 17: 11:30l: 12*: 2: 12: 19*1 11*:8*:8*: 5*
3
159 :192 25 9 :6 4 :6628 3 :7 4 :
112 :4 6 :90 :7 4 :65 :59 :
t 43t 129t 20t 251 16t 12*!: 22t 181 8*1 31*1 16*I 36*i 14*
10
379 :8 8 :
176 :179 :492
139 :9 4 :
293 2271 :213:139:1%:582
t 133t 30!: 28i 47:21iffi 53i 135i 42"139*t 73*tiSf: 9 *
20
370 :ill :212 :3 3 2 :55 :
187:208 :527 :169:889 :226 :3642177 2
1 114158t 81£ 151'.8t 49"i 67I: 407• 8 8 "H459*1 38"1 147": 42"
F
2.610.803.152.630.233.482.691.344.747.693.463.727.22
P
.10
.52
.06
.10£7.05.09.31.02.04.05.04.01
F ratios and p values based on one-way ANOVAs for individual spiders. Superscripted letters indicatefignifjr?m differences between density treatments based on a Tukey post hoc comparison of meanstest Identical letters indicate no significant difference between those treatments.
by guest on June 6, 2012http://beheco.oxfordjournals.org/
Dow
nloaded from
564 Behavioral Ecology Vol. 9 No. 4
Vibratory Only
m
"3"ooa
4645444342414039383736
4645444342414039383736
3 10 20
Visual and Vibratory
20
Visual Only
a a a
10 20
Prey densityFigure 3Vibration levels generated by crickets at various densities in each ofthree different apparatuses providing different sensory wimuliMeans with different letters are significantly different at p < .05; n=> 30 for each experimental treatment.
assess patch quality. This suggests that not all sensory channelsare used equally in determining residence time, resulting indifferent patch times based on the source of information.However, the difference in residence time due to combina-tions of Visual and vibratory cues rather than visual cues alonesuggests that various sensory channels may interact in com-plex ways in the assessment of patch quality. Other studieshave found that residence time may differ based on chemicalcues from prey as well (Persons and Uetz, 1996b). Given thatmost predators use multiple sensory channels to detect prey,and the importance of these channels may change with en-vironmental heterogeneity, this is an important consideration
in understandings proximate mechanisms of patch foragingdecisions.
Environmental constraints on transmission of various typesof sensory information may partially explain these results.Schixocosa ocreata inhabits complex, loose leaf litter where vi-brations from prey attenuate rapidly, often being conductedno farther than the distance of a single leaf (Scheffer et aL,1996). This represents a smaller area within which to perceiveprey and could result in selection for using visual cues whileforaging. Using visual cues instead of vibrations would allowfor a larger perceptual "patch" in which to forage and con-sequently less frequent movement between patches and great-er foraging efficiency (Bye et al., 1992; Rice, 1983).
Many studies have examined different types of presamplingor prescient foraging information (Nishimura, 1991a,b; Val-one, 1992; Valone and Brown, 1989; Valone and Giraldeau,1993) in assessing patch quality, but all of these use prior ex-perience as a parameter or assume that patches with staticpatch quality may have been estimated by the forager fromprior patch experience. Present sensory information may bean even more important source of information than prior ex-perience for many animals. Wolf spiders rely heavily on pres-ent sensory information rather than on any other type of pre-sampling information. Schizocosa ocreata move randomly be-tween patches that differ in sensory information or that differin the presence of prey (Persons and Uetz, 1996a). They alsodo not base residence time on previous patch sensory expe-rience (Persons and Uetz, 1996a), previous prey attack (Per-sons and Uetz, 1997a) or feeding (Persons and Uetz, 1997b),suggesting that bayesian foraging is unlikely in wolf spidersand that patch quality assessment and residence time deci-sions are based primarily on present sensory information rath-er than any other source.
Nishimura (1994) modeled decision makfng in a sit-and-wait predator when the forager finds one of three possiblepatch states: finding no prey, finding prey without attacking(analogous to the conditions in this study), and rinding preyand attacking (prey capture). Nishimura predicts that when aforager moves between patches without learning or memoryand chooses patches at random, it bases its decision to stayon the long-term probabilities of the patch containing noprey, one prey, or two prey. Under diis condition, the expe-rience of finding prey without attacking does not contributeto exploitation efficiency, and the forager should treat allpatches the same. Wolf spiders uphold the assumptions of thismodel. They experience no difference in energy gain amongprey density treatments, have been found to move randomlybetween patches (Persons and Uetz, 1996a), and appear notto use prior sensory experience to modify residence time, yetthey vary residence times using sensory cues of prey density.This suggests that foraging patch decisions with some animalsneed not employ memory, sampling information, or Bayesianmeans of assessment to determine patch quality and that themost parsimonious source of information, present sensory in-formation, may be the one weighted most heavily.
Wolf spiders detect prey primarily through visual detectionof movement (Rovner, 1991, 1993). As such, the probabilityof detecting prey in a patch may be directly related to theprobability of any prey moving while the spider remains in apatch. Therefore, even in a simple environment, cricket move-ment patterns may be sufficient to generate uncertainty aboutpateb quality, and prey encounter may be considered a Pois-son process. If prey detection is based on prey movemest,spiders do not have complete knowledge of patch quality. Us-ing video images of prey, previous studies completed with S.ocreata fixed die proportion of time a prey item spent movingin a foraging patch and therefore removed this source of sto-chasticity, and found that S. ocreata wolf spiders use a fixed
by guest on June 6, 2012http://beheco.oxfordjournals.org/
Dow
nloaded from

Persons and Uetz • Prey density assessment by wolf spiders 365
probability of leaving a patch that is influenced by both visualdetection of cricket movement and stochasticity inherent inthe decision rule (i.e., not all variation in residence time isaccounted for by stochasticity in prey movement) (Personsand Uetz, 1997a). It is important to note that in this study,even 25 sequential visits into a patch with a standardizedamount of cricket movement (using a video-generated crick-et) failed to modify the behavior of the spider (Persons andUetz, 1997b). This also emphasizes the lack of importance ofprior sensory experience in a patch. However, lunging at preywithout feeding does significantly modify wolf spider resi-dence time (Persons and Uetz, 1997a). This may simply reflectthat crickets may not be detected on every patch visit Studieson residence time in crab spiders found that they also exhibita fixed probability of leaving a patch even though this doesnot result in optimal prey encounter (Kareiva et aL, 1989).Kareiva et aL suggest that this is due to extreme stochasticityin prey encounter. Morse (1993) later found that crab spiderdecisions regarding patch choices may be modified by dis-tance cues radier than by prey capture directly.
Chamov's marginal value theorem (Charnov, 1976) as-sumes a continuous and deterministic gain function that ac-celerates negatively over time in a patch, and the decision toleave is partially contingent on the gain function for alterna-tive patches. The energetic gain for patches in this study iszero, and residence time is largely determined by the proba-bility of detection of prey movement, which is likely to be alinear function of residence time. Differences in vibration lev-els across density treatments reflect linear increases in cricketmovement and therefore increases in the probability of de-tecting prey for any given patch visit The spiders had a choicewith regard to the patch: remain in the presence of sensorystimuli or leave for another patch void of sensory cues. Thetwo patches are mutually exclusive with respect to sensorystimuli, but identical with respect to energy gain. To deter-mine if sensory cues function in an analogous way to energygain with respect to the marginal value theorem would re-quire alternative patches being offered with greater or lesserdegrees of sensory stimuli radier than an empty neutral patch.
The threshold of response for spider detection of prey vi-brations may be higher than that for any foraging patch usedfor this study. However, spiders in patches with vibratory in-formation alone were occasionally observed lunging at theacetate screen when a cricket was walking nearby. This strong-ly suggests diat the vibration levels were sufficient for preydetection but were not used to determine residence time.
This laboratory study demonstrates diat spiders are at leastcapable of responding to differences in prey density even ifprey capture success is minimal for a particular site. It alsodemonstrates diat a postingestion feedback mechanism is notnecessary to explain foraging time at a particular site and thatsensory information should be considered more strongly inproximate and theoretical studies of residence time decisions.
This research was supported in part by funds from the National Sci-ence Foundation through grant IBN-94142S9 (support for G.U.), theDepartment of Biological Sciences, the Arachnological Research Fundof the University of Cincinnati, and a University of Cincinnati Re-search Council Fellowship. Portions of this research were submittedin partial fulfillment of the requirements for M5. and Ph.D. degreesin Biological Sciences at the University of Cincinnati. We are gratefulto B. C Jayne for «t3p«rirai advice and technical criticism. We thankD. Wise, A. Cady, J. Shann, and two anonymous reviewers for theirhelpful comments on this manuscript. We acknowledge the people ofthe Cincinnati Nature Center for their willingness to provide a col-lecting site for Schizocosa ocrtala. Additional thanks go to W. Mc-diniock, A. DeLay, K. Cook, D. Kroeger, and A. McCrate for theiradvice and assistance with spider husbandry.
REFERENCES
Bennington CC, Thayne WV, 1994. Use and misuse of mixed modelanalysis of variance in ecological studies. Ecology 75:717-722.
Bye FN, Jacobsen BV, Sonerud GA, 1992. Auditory prey location in apause-travel predator search height, search time, and attack rangeof Tengmalm's owls (AtgoHusfurunus). Behav Ecol 3:266-276.
Cady AB, 1984. Microhabiat selection and locomotor activity of Scfa-zoeosa ocrtata (Walckenaer) (Araneae: Lycosidae). J Arachnol 11:297-307.
Charnov EL, 1976. Optimal foraging, the marginal value theorem.Theor Popul Biol 9:129-136.
Cordon D, Bell WJ, 1991. The use of visual information by house-flies,Musca domtstita (Diptera, Muscidae), foraging in resource patches.J Comp Physiol A 168:365-371.
Cook RM, Hubbard SF, 1977. Adaptive searching strategies in insectparasites. J Anim Ecol 46:115-126.
DeBenedictis PA. Gill FB, Hainsworth RR, Pyke GH, Wolf LL, 1978.Optimal meal size in hummingbirds. Am Nat 112:301-316.
EndlerJA, 1991. Interactions between predators and prey. In: Behav-ioural ecology: an evolutionary approach, 3rd ed (Krebs JR, DaviesNB eds). Oxford: Blackwell Scientific Publications; 169-196.
Ford MJ, 1978. Locomotory activity and the predation strategy of thewolf spider Pardosa amentata (Qerck) (Lycosidae). Anim Behav 26:31-35.
Formanowicz DR, Bradley PJ, 1987. Fluctuations in prey density: ef-fects on the foraging tactics of scolopendrid centipedes. Anim Be-hav 35:453-461.
Heinrich B, 1979. Resource heterogeneity and patterns of movementin foraging bumblebees. Oecologia 40:235-245.
Heinrich B, 1983. Do bumblebees forage optimally, and does it mat-ter? Am Zool 23:273-281.
Hodges CM, 1981. Optimal foraging in bumblebees—hunting by ex-pectation. Anim Behav 29:1166-1171.
Hughes RN, 1979. Optimal diets under the energy maximizationpremise: die effects of recognition time and learning. Am Nat 113:209-221.
Hurlbert SH, 1984. Pseudoreplication and the design of ecologicalfield experiments. Ecol Monogr 54:187-211.
Iwasa Y, Higashi M, Yanamura N, 1981. Prey distribution as a factordetermining die choice of optimal foraging strategy. Am Nat 117:710-723.
Janetos AC, 1982. Foraging tactics of two guilds of web-spinning spi-ders. Behav Ecol Sociobiol 10:19-27.
Johnson DM, Akre BG, Crowley PH, 1975. Modeling arthropod pre-dation: wasteful killing by damselfly naiads. Ecology 56:1081-1093.
Kareiva P, Morse.DH, Ecdeston J, 1989. Stochastic prey arrivals andcrab spider residence times: simulations of spider performance us-ing two simple 'rules of thumb.' Oecologia 78:547-549.
Krebs JR, Ryan JC Charnov EL, 1974. Hunting by expectation oroptimal foraging? A study of patch use by chickadees. Anim Behav22553-964.
Krebs JR. Kacelnik A, Taylor P, 1978. Test of optimal sampling byforaging great tits. Nature 275:27-31.
Livoreil B, Giraldeau L-A, 1997. Patch departure decisions by spicefinches foraging singly or in groups. Anim Behav 54.-967-9T7.
Lizoue RS, Rovner JS, 1988. Nocturnal capture of fireflies by tycosidspiders: visual versus vibratory stimuli. Anim Behav 36:1809-1815.
Mangel M, Clark CW, 1983. Uncertainty, search and information infisheries. J Intern Council Explor Seas 41:93-103.
Marden JH, 1984. Remote perception of floral nectar by bumblebees.Oecologia 64:232-240.
Martin P, Kraemer HC, 1987. Individual differences in behaviour andtheir statistical consequences. 35:1366-1375.
McNamaraJM, Houston AI, 1980. The application of statistical deci-sion theory to animal behavior. J Theor Biol 85: 673-690.
McNamaraJM, Houston AI, 1982. Optimal patch use in a stochasticenvironment. Theor Popul Biol 21:269-288.
McNamara JM, Houston AI, 1987. Memory and the efficient use ofinformation. J Theor Biol 125:385-395.
Morse DH, 1989. Cues associated with patch-choice decisions by for-aging crab spiders Misumena vatia. Behaviour 107:297-312.
Morse DH, 1993. Choosing hunting sites with little information:patch-choice responses of crab spiders to distant cues. Behav Ecol4:61-65.
by guest on June 6, 2012http://beheco.oxfordjournals.org/
Dow
nloaded from

S66 Behavioral Ecology Vol. 9 No. 4
Nakamura K, 1968. The ingestion in wolf spiders. I. capacity of gutof Lycosa pstudoatmulata. Ret Popul Ecol 10:45-53.
Niihimura K, 1991a. Optimal patch residence time of a sit-and-waitforager. Bchav Ecol 2:283-294.
NUhimura K, 1991b. Utilization of different prey type patches in theUral owl (Strix urairruis): a sit-and-wait predator. Behav Ecol 2:99-105.
Nishimura K, 1994. Decision making of a sit-and-wait forager in anuncertain environment: learning and memory load. Am Nat 143:656-676.
Nishimura K, Abe TM, 1988. Prey susceptibilities, prey utilization andvariable attack efficiencies of Ural owls. Oecologia 77:414-422.
Oaten A, 1977. Optimal foraging in patches: a case for uochastitiry.Theor Popul Biol 12:263-285.
Pasquet A, Ridwan A, LeBorgne R, 1994. Presence of potential preyaffects web-building in an orb-weaving spider Zyguila x-notata.Anim Behav 47:477-480.
Persons MH, Uetz GW, 1996a. The influence of sensory informationon die patch residence time in wolf spiders (Araneae: Lycosidae).Anim Behav 51:1285-1293.
Persons MH, Uetz GW, 1996b. Wolf spiders vary patch residence timein the presence of chemical cues from prey (Araneae, Lycosidae).J Arachnol 24:76-79,
Persons MH, Uetz GW, 1997a. The effect of prey movement and at-tack behavior on the patch residence decision rules of wolf spiders(Araneae: Lycosidae). J Insect Behav 10:737-751.
Persons MH, Uetz GW, 1997b. Residence time decisions in wolf spi-ders: is perceiving prey as important as eating prey? Ecoscience 4:1^5.
Provenza FD, Qncotta RP, 1993. Foraging as a self-organizationallearning process: accepting adaptability at the expense of predict-ability. In: Diet selection: an interdisciplinary approach to foragingbehaviour (Hughes RN, ed). Oxford: Blackwell Scientific Publica-tions; 78-101.
Pulliam HR, 1974. On the dieory of optimal diets. Am Nat 108:59-7.4.
Pyke GH, 1978a. Optimal foraging in bumblebees and coevolutionwith their plants. Oecologia 36:281-293.
Pyke GH, 1978b. Optimal foraging in hummingbirds: testing the mar-ginal value theorem. Am Zool 18:739-52.
Pyke GH, 1980. Optimal foraging in bumblebees: calculation of netrate of energy intake and optimal patch choice. Theor Popul Biol17:232-246.
Pyke GH, 1984. Optimal foraging theory: a critical review. Annu RevEcol Syst 15:523-575.
Rice WR, 1983. Sensory modality: an example of its effect on optimalforaging behavior. Ecology 64:403-406.
Riechert SE, 1985. Decisions in multiple goal contexts: habitat selec-
tion of the spider, AgtUnopsis aperta (Gertsch). Z Tierpsychol 70:53-69.
Rovner JS, 1991. Evidence for idiothetically controlled turns and ex-traocular photoreception in tycosid spiders. J Arachnol 19:169-173.
Rovner JS, 1993. Visually mediated responses in the lycosid spiderRabidosa rabidcc the roles of different pairs of eyes. Mem Queens-land Mus 55*35-638.
Scheffer SJ, Uetz GW, Stranon GE, 1996. Sexual selection, male mor-phology, and the efficacy of courtship signalling in two wolf spiders(Araneae: Lycosidae). Behav Ecol Sociobiol 38:17-23.
Schuler W, 1990. Are rules of thumb sufficient for the starling'schoice of food according to gain in energy? In: Behavioural mech-anisms of food selection (Hughes RN, ed). New York: Springer;127-142.
Shaw R, Mitcheu-Olds T, 1993. ANOVA for unbalanced data; an over-view. Ecology 74:1638-1645.
Stephens DW, Krebs KR, 1986. Foraging theory. Princeton, New Jer-sey: Princeton University Press.
Templeton JJ, Giraldeau L-A, 1995. Patch assessment in foragingflocks of European starlings: evidence for die use of public infor-mation. Behav Ecol 6:65-72.
Turn bull AL, 1964. The search for prey by a web-building spider,Achatararua upidariorum (CL. Koch) (Araneae: Theridiidae). CanEntomol 96-368-579.
Valone TJ, 1991. Bayesian and prescient assessment: foraging with pre-harvest information. Anim Behav 41:569-577.
Valone TJ, 1992. Information for patch assessment: a field investiga-tion with black-chinned hummingbirds. Behav Ecol 3:211-221.
Valone TJ, Brown JS, 1989. Measuring patch assessment abilities ofdesert granivores. Ecology 70:1800-1810.
Valone TJ, Giraldeau L-A, 1993. Patch estimation by group foragers:what information is used? Anim Behav 45:721-728.
Wise DH, 1993. Spiders in ecological webs. New York; Cambridge Uni-versity Press.
Wise DH, Wagner JD, 1992. Exploitative competition for prey amongyoung stages of the wolf spider Schaocosa ocrtata. Oecologia 91:7-13.
Wolf LL, Hainswordi FR, 1983. Economics of foraging strategies insunbirds and hummingbirds. In: Behavioral Eenergetics: the costof survival in vertebrates (Aspey WP, Lustick SI, eds). Columbus:Ohio State University Press.
Zach R, Falls JB, 1976. Do ovenbirds (Aves: Parulidae) hunt by ex-pectation? Can J Zool 54:1894-1903.
Zar JH, 1984. Biostatistical analysis, 2nd ed. Englewood Cliffs, NewJersey: Prentice-Hall; 470-476.
Zhang Z-Q, Sanderson JP, 1993. Behavioral responses to prey densityby three acarine predator species with different degrees of polyph-agy. Oecologia 96:147-156.
by guest on June 6, 2012http://beheco.oxfordjournals.org/
Dow
nloaded from





























