Embed Size (px)
Citation preview

REVIEW
Presence and detection of anaerobic ammonium-oxidizing(anammox) bacteria and appraisal of anammox processfor high-strength nitrogenous wastewater treatment: a review
Akihiko Terada • Sheng Zhou • Masaaki Hosomi
Received: 30 September 2010 / Accepted: 15 February 2011
� Springer-Verlag 2011
Abstract Until now, anaerobic ammonium oxidation
(anammox) has been widely applied as an alternative
method to the conventional nitrification–denitrification
pathway for biological nitrogen removal from wastewater.
Since their discovery in a denitrifying fluidized bed reactor
in the Netherlands in the early 1990s, anammox bacteria
have also been detected in natural environments. Anam-
mox is one of the newly found drivers known to contribute
to the biogeochemical nitrogen cycle. In the marine envi-
ronment, more than 50% of nitrogenous compounds are
reportedly converted into nitrogen gas via the anammox
pathway. These observations were made using state-of-the-
art techniques for detecting anammox bacteria based on
their lipids, small-subunit ribosomal RNA genes, func-
tional genes, and unique reaction pathways. The research
objectives for anammox bacteria are quite diverse, ranging
from the application of anammox processes to various
wastewater types, to anammox biochemistry and phylog-
eny, to elucidating how anammox bacteria have evolved.
Since the genome of the anammox bacterium Kuenenia
stuttgaritiensis was deciphered, anammox bacteria have
proved to be quite versatile. The next challenge is to enrich
knowledge of anammox bacterial physiology and phylog-
eny to improve their use in engineered and natural envi-
ronmental systems and minimize nitrogen loads to
downstream water bodies. Furthermore, rapid startup of the
anammox process for engineered systems is required to
broadly harness the benefits of anammox bacteria. This
review article summarizes the physiology and phylogeny
of anammox bacteria, detection methods of anammox
bacteria and reactions, the behavior of anammox bacteria
in natural environments, and recent developments in their
use for engineered systems.
Keywords Anaerobic ammonium oxidation � Anaerobic
ammonium-oxidizing bacteria � Biofilm � Microbial
ecology � Planctomycetes
Introduction
Nitrogenous compounds, discharged in municipal and
industrial wastewaters, are drivers of eutrophication, which
enhances excessive growth of green algae and cyanobac-
teria, in many developing and developed countries. The
excessive growth of the algae and cyanobacteria reduces
the oxygen concentration in the receiving waters and,
additionally, some cyanobacteria excrete toxins (Sangolkar
et al. 2006). This leads to decreases in the dissolved oxygen
concentration, water stagnation and the eventual death of
fish and shellfish in the water body. Nitrite and nitrate
contamination of water and groundwater is an emerging
problem worldwide, and originates from anthropogenic
nitrogen pollution.
To reduce the nitrogen loading to water bodies, bio-
logical nitrogen removal which comprises nitrification and
denitrification has been an essential biochemical step.
Nitrification is a key process for nitrogen removal in most
municipal and industrial wastewater treatment plants
(WWTPs) and constructed wetlands as well as natural
environments. The term ‘‘nitrification’’ refers to the aero-
bic, sequential oxidation of ammonium to nitrite, and
nitrite to nitrate. These two steps are catalyzed by spe-
cialized chemolithoautotrophic prokaryotes known as
A. Terada (&) � S. Zhou � M. Hosomi
Department of Chemical Engineering,
Tokyo University of Agriculture and Technology,
2-24-16 Naka-cho, Koganei, Tokyo 184-8588, Japan
e-mail: [email protected]
123
Clean Techn Environ Policy
DOI 10.1007/s10098-011-0355-3

ammonia-oxidizing bacteria (AOB) (Wiesmann 1994),
ammonia-oxidizing archaea (AOA) (Konneke et al. 2005;
Francis et al. 2007), and nitrite-oxidizing bacteria (NOB)
(Wiesmann 1994). Currently, no microorganisms have been
reported to be capable of mediating both autotrophic nitrifi-
cation steps. Denitrification is defined as the anoxic process in
which nitrite and nitrate are reduced to gaseous nitrogen
oxides (nitric oxide (NO), nitrous oxide (N2O), and free
nitrogen (N2)) by facultative anaerobic heterotrophic deni-
trifying bacteria. These conventional nitrification–denitrifi-
cation pathways have reportedly been central to biological
nitrogen removal in engineered systems and natural envi-
ronments (Ahn and Choi 2006; Van Hulle et al. 2010) and are
considered to be of major importance in the global nitrogen
cycle (Jetten 2008; Jetten et al. 2009; Zhu et al. 2010).
In 1995, anaerobic ammonium oxidation (anammox)
was accidently discovered in a fluidized bed denitrifying
reactor where influent ammonium was suddenly lost con-
comitant with nitrate reduction following long-term oper-
ation of the reactor (Mulder et al. 1995). The discovery of
the anammox reaction was corroborated from the thermo-
dynamic calculations of Broda (1977), which revealed that
ammonia oxidation in the absence of oxygen but in the
presence of nitrite/nitrate is thermodynamically feasible. It
has been 15 years since two papers on the discovery of the
anammox reaction were published in 1995 (Mulder et al.
1995; van de Graaf et al. 1995).
Initially, the electron acceptor for the anammox reaction
was considered to be nitrate; but this was corrected to
nitrite and the following anammox stoichiometry was
proposed in 1998 (Strous et al. 1998):
1NHþ4 þ 1:32NO�2 þ 0:066HCO�3 þ 0:13Hþ ! 1:02N2
þ 0:26NO�3 þ 0:066CH2O0:5N0:15 þ 2:03H2O
ð1Þ
A few papers have also reported nitrogen loss in the
absence of organic carbon to support the anammox reaction
(Kuai and Verstraete 1998; Siegrist et al. 1998); however, the
conclusions as to whether anammox is a biologically
mediated reaction or not has been disputed for some time.
This is because the anammox bacteria have an extremely
slow growth rate, making it difficult to confirm the anammox
reactions reported by other researchers. However, anammox
bacteria were successfully enriched to 70% in a sequencing
batch reactor (SBR) (Strous et al. 1998) and concentrated up
to 99.6% by density-gradient centrifugation (Strous et al.
1999a), leading to the conclusion that anammox is a
biological reaction. Strous et al. (1999a) successfully
enriched a new autotrophic member of the order
Planctomycetales (in the phylum Planctomycetes), and
identified it as anammox bacteria. This paper revolutionized
research on better uses of anammox bacteria for nitrogen
removal in engineered systems. Since then, many engineered
systems for biological nitrogen removal have been developed
and have been reviewed elsewhere (Kuenen and Jetten 2001;
Ahn 2006). Currently, anammox-related technology has the
potential to achieve an energy-neutral or even energy-positive
process (Kartal et al. 2010).
The number of scientific papers related to anammox,
published in journals registered in the ISI Web of Science
(Thomson Reuters Inc.) has been increasing year by year as
shown in Fig. 1, broadening the research field from engi-
neered systems to the exploration of anammox to natural
environments. Studies on natural environments have required
us to reconsider the global nitrogen cycle, as anammox is one
of the major biogeochemical processes in marine and estua-
rine environments (Francis et al. 2007). Many surprising and
exciting discoveries regarding anammox ubiquity in aquatic
environments have been reported (Jetten et al. 2003), mainly
due to rapidly developing molecular techniques which mainly
focus on small-subunit ribosomal RNA (16S rRNA) and
functional genes in a culture-independent manner (Wagner
et al. 2006; Amann and Fuchs 2008). This review article
summarizes the physiology and phylogeny of anammox
bacteria, detection methods for anammox bacteria and reac-
tions, the presence and behavior of anammox bacteria in
natural systems, and recent developments on the use of these
bacteria for wastewater treatment.
Involvement of anaerobic ammonium oxidation
in the biological nitrogen cycle
Microbiologically, nitrogen compounds are involved in
five biological processes: nitrification which involves two
0
20
40
60
80
100
120
140
160
1994 1996 1998 2000 2002 2004 2006 2008 2010
The
num
ber
of I
SI p
ublic
atio
n
Year
Discovery of anammox in marine sediment (Thamdrup and Dalsgaard, 2002)
Completion of the genome sequence of K. stuttgartiensis(Strous et al., 2006)
Discovery of anammox in the ocean (Dalsgaard et al., 2003; Kuypers et al., 2003)
Discovery of anammox (Mulder et al., 1995)
Proof of anammox as biologically mediated process (Strous et al., 1999)
The number of papers on anammox research
The number of papers on anammox research for bioreactors
Fig. 1 Increase in the number of publications related to anaerobic
ammonium oxidation since 1995. Comments represent ground-
breaking papers on anammox research
A. Terada et al.
123

metabolic processes, i.e., ammonium and nitrite oxidation,
denitrification, dissimilatory nitrate reduction to ammo-
nium (DNRA), and anammox. In engineered systems, the
nitrogenous constituents, mainly organic nitrogen and
ammonium, are converted to nitrogen gas through con-
ventional nitrification–denitrification and partial nitrifica-
tion to nitrite (nitritation) via anammox pathways (Ahn
2006). Typical nitrogen removal processes are summarized
in Fig. 2. The nitrification–denitrification pathway has been
commonly utilized in engineered systems such as con-
structed wetlands (Knight et al. 1999; Chavan et al. 2008;
Lee et al. 2009a) and WWTPs (van Loosdrecht 2008).
Recently, intensive efforts have focused on nitrogen
removal via nitrite rather than nitrate due to the lower
energy input requirement. Nitrogen removal via nitrite, a
short-cut nitrogen removal process, allows for a lower
oxygen requirement with a stoichiometric value of 3.43
g-O2/g-N (via nitrate: 4.57 g-O2/g-N); a lower organic
carbon requirement as a denitrification electron donor with
a stoichiometric value of 1.71 g-COD/g-N (via nitrate:
2.86 g-COD/g-N); and less sludge production with 0.73
g-volatile suspended solid (VSS)/g-N (via nitrate: 1.19
g-VSS/g-N) compared with nitrogen removal via nitrate
(Rittmann and McCarty 2001). Applications using the
anammox process are more beneficial than nitrogen
removal via nitrite. The partial ammonium oxidation to
nitrite, the stoichiometric value suitable for the subsequent
anammox reaction of an ammonium/nitrite molar ratio of
1/1.32 (Strous et al. 1998), requires much less oxygen at
1.75 g-O2/g-N (Terada et al. 2007), no organic carbon as
an electron donor, and produces less sludge at 0.14 g-VSS/
g-N (Strous et al. 1998). Therefore, this approach reduces
power requirements by 64% (due to reduced aeration),
COD demands by 100% and sludge production by 80–90%
compared with nitrogen removal via nitrate, creating sig-
nificant benefits for the treatment of high-strength nitrog-
enous wastewaters (van Loosdrecht 2008). Thus, nitritation
coupled to anammox is an innovative process for biological
nitrogen removal from wastewater (Sliekers et al. 2002;
Pynaert et al. 2003; van der Star et al. 2007; Kuenen 2008).
One engineering challenge for nitrogen removal via a
nitritation–anammox pathway is to stabilize nitritation
performance. Suppression of nitrite oxidation is a key for
success of the nitritation–anammox pathway. Indeed,
nitrification system perturbation/malfunction, e.g., exces-
sive oxygen supply, potentially leads to conversion of
nitrite into nitrate which cannot be used for anammox
bacteria. This eventually impairs the anammox activity,
resulting in system failure of a nitritation–anammox pro-
cess (Strous et al. 1999b; Third et al. 2005; Terada et al.
2007). Given that the metabolic workload during nitrifi-
cation is divided between two physiologically and phylo-
genetically distinct groups of microorganisms, AOB
(or AOA) and NOB (Francis et al. 2007), AOB need to
over-grow NOB so that nitrite can successfully accumulate
in the bioreactor. Several technical approaches have been
used to preferentially enrich AOB over NOB by employing
their different eco-physiological properties, e.g., tempera-
ture (Balmelle et al. 1992; Jetten et al. 1997), solid reten-
tion time (SRT) (Hellinga et al. 1998), free ammonia (FA)
(Kim et al. 2006; Ganigue et al. 2007), and dissolved
oxygen (Bernet et al. 2005; Ahn et al. 2008; Terada et al.
2010). Not only suspended cultures, but also biofilms can
take advantage of these strategies for successful nitritation.
Especially, biofilm systems operating at high FA concen-
trations and/or limited oxygen loading (Pynaert et al. 2004)
plus the inoculum composition (Terada et al. 2010) can
allow for nitritation, but the challenge of fine adjustment
and control of the oxygen loading remains. In addition, the
long-term nitritation stability is still questionable, which
warrants future challenge (Fux et al. 2004). The details of
various criteria on successful nitritation are summarized in
several review articles (Schmidt et al. 2003; Khin and
Annachhatre 2004; Ahn 2006; Sun et al. 2010; Van Hulle
et al. 2010).
Phylogeny of anammox bacteria
The 16S rRNA sequence of an anammox microorganism,
Candidatus Brocadia anammoxidans, was identified for the
first time by Strous et al. (1999a), showing that Ca. Bro-
cadia anammoxidans is affiliated to the phylum Plancto-
mycetes. Numerous research has been carried out since
then. As shown in Fig. 3, which shows the 16S rRNA gene-
based phylogenetic tree as reconstructed by ARB software
NH4+
NO2- NO3
-
NON2O
NitrificationAerobic
DenitrificationAnoxic
NOB
Denitrifying bacteria
Nitritation-anammox
N2
AnammoxAnammox bacteria
AOB and AOA
DNRAOrg-N
NH2OH
PONDON
Nit
roge
n fi
xati
on
Fig. 2 Microbial nitrogen transformation in natural and engineered
systems. Methane-dependent denitrification is excluded here. AOAammonia-oxidizing archaea, AOB ammonia-oxidizing bacteria,
DNRA dissimilatory nitrate reduction to ammonium, DON dissolved
organic nitrogen, PON particulate organic nitrogen, and NOB nitrite-
oxidizing bacteria
Review of anammox detection, presence, and application
123

(Ludwig et al. 2004), anammox bacteria have been divided
into five genera, Ca. Brodacia, Ca. Kuenenia, Ca. Scalin-
dua, Ca. Anammoxoglobus, and Ca. Jettenia (Jetten et al.
2005; Kartal et al. 2007b; Quan et al. 2008). Generally, Ca.
Scalindua have been detected as the predominant anammox
bacteria in marine environments such as sediment and the
oxygen minimum zone (OMZ) (Kuypers et al. 2003; Sch-
mid et al. 2003; Dong and Reddy 2010; Li et al. 2010b),
and high-temperature oil resorviors (Li et al. 2010a) while
the other anammox genera have generally been found in
bioreactor systems for wastewater treatment (Egli et al.
2001; Pynaert et al. 2003; Kindaichi et al. 2007; van der
Star et al. 2008), hot spring (Jaeschke et al. 2009a), and
terrestrial environments (Amano et al. 2007; Dale et al.
2009; Humbert et al. 2010), exhibiting a ubiquitous pres-
ence (Kuenen 2008; Schmid et al. 2007). These anammox
bacteria are deeply branched in the Planctomycetes phy-
lum, with low 16S rRNA similarities to other Planctomy-
cetales genera, e.g., Gemmata, Isosphaera, Planctomyces,
and Pirellula (below 80%) as shown in Fig. 3 (Schmid
et al. 2005). Even in the branch of anammox bacteria, each
distinct genus has as low as a 90% similarity in 16S rRNA
sequences. The detailed reason for this lack of similarity
remains unresolved, but the Planctomycetes phylum seems
to have evolved very slowly and may be less sensitive to
evolutional changes. This has the potential to permit the
16S rRNA gene of each individual species and genera to
become separated. It is also possible that Planctomycetes
could be the first bacterial group to emerge (Brochier and
Philippe 2002). After the deciphering the genome of Ca.
Kuenenia stuttgartiensis (Strous et al. 2006), the detailed
anammox reaction mechanisms and the metabolic versa-
tility of anammox bacteria have been investigated in depth.
The detailed results can be found in a review article (Jetten
et al. 2009).
Anammox versatility has been recently reported, with
Ca. Brocadia fulgida and Ca. Anammoxoglobus propio-
nicus capable of taking up acetate (Kartal et al. 2008) and
propionate (Kartal et al. 2007b) along with ammonium,
respectively. These anammox bacteria are likely to pre-
vail over other anammox bacteria when such volatile fatty
acids are present with ammonium. Furthermore, Ca.
Kuenenia is able to mediate DNRA and, potentially, so
are the other anammox bacterial genera (Kartal et al.
2007a). These different physiological characteristics
could be a key for determining the predominant anammox
bacteria in both natural environments and engineered
systems.
Detection of anammox bacteria and reactions
The discovery of anammox reactions in natural environ-
ments and advances in anammox-related technologies are
attributed to the development of diversified methods
focusing on anammox bacterial cells/genome and anam-
mox reactions. The overview of methods which have been
used for detecting anammox reactions and bacteria are
shown in Fig. 4. In this chapter, each method used for
anammox detection is introduced.
Planctomyces
Candidatus Scalindua wagneri, AY254882Candidatus Scalindua brodae, AY257181
Candidatus Kuenenia stuttgartiensis, AF375995
Candidatus Anammoxoglobus propionicus, DQ317601
Candidatus Jettenia asiatica, DQ301513
Candidatus Brocadia fulgida, DQ459989
Candidatus Brocadia anammoxidans, AF375994
Isosphaera
Gemmata
Pirellula
0.10
To outgroups
Fig. 3 Phylogenetic tree based
on 16S rRNA gene for the
phylum Planctomycetesconsisting of five genera of
previously reported anaerobic
ammonium-oxidizing bacteria
and the genera Planctomyces,
Pirellula, Gemmata, and
Isophaera. The tree
reconstruction was carried out
by neighbor-joining analysis
with a 50% conservation filter
for bacteria. The bar represents
10% sequence divergence. This
tree was reconstructed using the
ARB software package (Ludwig
et al. 2004)
A. Terada et al.
123

Polymerase chain reaction for 16S rRNA and functional
genes
As of January 2011, the isolation of an anammox bacterium
has yet to be successful. Therefore, culture-independent
methods, targeting either 16S rRNA or functional genes,
are the most widely used approaches. To detect anammox
bacteria in environmental samples, PCR amplification with
general 16S rRNA gene-targeted primers and subsequent
quantitative or phylogenetic analyses are a central method,
in common use. Bacterial diversity studies have also been
used, and these are summarized elsewhere (Muyzer 1999;
Fromin et al. 2002; Schutte et al. 2008).
The 16S rRNA or functional gene-based approach
without cultivation is a very powerful technique when
anammox bacteria have low activity or samples contain a
lot of inert particulates, both of which hamper the appli-
cation of the fluorescence in situ hybridization (FISH)
method which will be explained in ‘‘Fluorescence in situ
hybridization’’ section (Schmid et al. 2005). Generally,
there are two drawbacks to the PCR method: low DNA
extraction yield and the production of artifacts after PCR.
The low DNA recovery after extraction is generally
dependent on the method and type of samples. Different
protocols for DNA extraction are reported to provide dif-
ferent degrees of DNA yield, potentially underestimating
the microbial community in respect of AOA (Urakawa
et al. 2010). There are currently no reports on the rela-
tionship between the degree of DNA yield and microbial
community diversity and density for anammox bacteria.
However, the precaution of selection of an appropriate
DNA extraction method should be undertaken. Numerous
primer sets are available for targeted anammox bacteria in
different environmental systems (Schmid et al. 2000, 2003,
2005; Penton et al. 2006).
The use of 16S rRNA as a biomarker of anammox
bacteria lacks a direct relationship to their physiologies,
which might be a disadvantage when investigating the
microbial community responsible for anammox. The
information obtained from 16S rRNA only identifies the
existence of DNA for targeted bacteria but not their
activities. The use of the 16S–23S rRNA intergenic spacer
region (ISR) could be a good candidate to identify anam-
mox activity. Anammox bacteria, particularly Ca. ‘Broca-
dia anammoxidans’ and Ca. ‘Kuenenia stuttgartiensis’, are
reported to have an ISR of approximately 450 bp, which
divides the 16S and 23S rRNA genes (Schmid et al. 2001).
In contrast, most other members of the Planctomycetales,
i.e., the genera Pirellula, Planctomyces, Isophaera, and
Gemmata do not have that ISR but possess unlinked 16S
and 23S rRNA genes. The difference can be detected by
molecular based quantification methods such as FISH
(Schmid et al. 2001) or quantitative PCR (Park et al. 2010).
The benefit of using the ISR region to detect anammox
Environmental Samples
Target:anammox reaction
In situ activity in a sediment or biofilm
Microsensor technique
Elucidation of anammox reaction
pathway
Tracer technique by stable isotope
Target:anammox bacteria
DNA/RNA in a bacterial cell
16S rRNA gene
Intergenic spacer region gene
Identification (cloning, finger printing),
quantification (quantitative PCR)
Functional gene
Identification (cloning, finger printing),
quantification (quantitative PCR)
Bacterial cell
16S rRNA gene
Intergeneicspacer region gene
FISH, CARD-FISH etc
Protein (cytochrome c)
Confocal Raman microscopy
Membrane lipid
GC/MS or NMR
Advantage
in situ measurement of chemical species without sample destruction
Able to distinguish anammox reaction from denitrification by 15N labeled NH4
+ or NO2
-
Able to identify and quantify anammox bacteria based on bacterial phylogeny without cultivation
Able to identify and quantify anammox bacteria based on bacterial function without cultivation
Able to detect activity based on transcriptome analysis (mRNA)
in situ detection of anammox bacteria without cultivation and sample destruction
Able to quantify anammox bacteria with spatial information kept
in situ detection of anammox bacteria based on cytochrome c
No need for pretreatment like FISH
Able to explicitly distinguish anammox bacteria (‘ladderane lipid’ is only a biomaker for anammox bacteria)
Diisadvantage
Difficult to specify a single reaction (necessary to link microsensor technique with a molecular method)
Incapable of maintaining spatial information (if a sample is an aggregate, sediment or biofilm)
Need to consider biases due to PCR amplification
Need to consider bias due to PCR amplification
Difficult to detect bacteria with lowactivity (FISH)
Potential artifacts due to amplification of negative signal (CARD-FISH)
Extremelyexpensive to procure a confocal Raman microscopy
Extremelyexpensive to procure an apparatus
Complicated to set up the apparatus
Fig. 4 Overview of methodologies for anammox reactions and bacterial detection
Review of anammox detection, presence, and application
123

bacteria occurs because the ISR allows monitoring of the
anammox bacteria activity in their natural and engineered
environments. For instance, Park et al. (2010) reported that
the ISR tracks and precedes the autotrophic nitrogen
removal rate, suggesting that ISR expression is a molecular
biomarker for predicting anammox reactor performances.
Functional gene markers, genes which encode key
enzymes involved in a specific metabolic pathway, are
another alternative to reflect bacterial physiology (Junier
et al. 2010). For anammox bacteria, the hydrazine oxido-
reductase (hzo) gene, which encodes for hydrazine oxido-
reductase (HZO), is a representative functional gene
biomarker. HZO, a member of the octaheme cytochrome
c hydroxylamine oxidoreductase protein family (Klotz
et al. 2008), is reported to be a key player in the anammox
biochemical process (Klotz and Stein 2008). The role of
HZO is to dehydrogenate hydrazine, an intermediate in the
anammox reaction, and to convert it into N2. HZO has been
purified from an anammox-enriched bioreactor and its role
has been investigated (Shimamura et al. 2007). The iden-
tification of the hzo gene has been used as a biomarker for
anammox bacteria (Quan et al. 2008; Schmid et al. 2008;
Park et al. 2010). A new primer set for the hzo gene has
been investigated with several environmental samples, and
is of promise for identifying the gene in ubiquitous natural
environments (Li et al. 2010b). These studies do not target
genuine anammox bacterial activity. Hence, transcriptome
analysis is essential to identify the gene activity by moni-
toring messenger RNA (mRNA), which may illuminate
anammox activity more accurately. It has been reported
that hzo mRNA, in addition to ISR, is a potential molecular
biomarker for predicting anammox reactor performances
(Park et al. 2010). Additionally, the cytochrome cd1-con-
taining nitrite reductase gene (nirS), which is unique to Ca.
Scalindua but distinct from the heterotrophic denitrifier,
nirS, has also been used with transcriptome analysis
(mRNA monitoring) to quantify the activity of Ca. Scal-
indua (Lam et al. 2009).
Application of real time quantitative PCR
Although conventional PCR amplification cannot quantify
the amount of a target gene, qPCR, and competitive PCR
(cPCR) allow targeted bacterial density to be quantified
based on 16S rRNA, ISR, or functional genes. The prin-
ciples behind these techniques are described elsewhere
(Sharkey et al. 2004; Smith and Osborn 2009). Competitive
PCR is less likely to be applied due to its lower accuracy
and the availability of cheaper qPCR apparatus. Several
researchers have applied the qPCR technique to quantify
anammox 16S rRNA (Tsushima et al. 2007a, b; Quan et al.
2008; Li et al. 2009) and functional gene (Lam et al. 2009;
Park et al. 2010) copy numbers. Quantitative PCR provides
higher throughput, more reliability and more sensitive
quantification compared with quantitative FISH. Therefore,
this technique has been used to quantify the maximum
growth rate of anammox bacteria by using a growth curve
for the bacteria (Tsushima et al. 2007a; van der Star et al.
2007). The doubling time of anammox bacteria was first
reported to be 11 days (Strous et al. 1998) but estimations
by qPCR have indicated a broader range for the maximum
growth rate of anammox bacteria, ranging from 3.6 to
17 days (Tsushima et al. 2007a; van der Star et al. 2007).
However, the disadvantage of qPCR is the fact that the
copy number of a targeted gene is quantified but not the
actual bacterial cell number. Bacteria have different copy
numbers of 16S rRNA genes in each bacterial cell
depending on the bacterial species (Klappenbach et al.
2001; Lee et al. 2009b). The 16S rRNA gene copies cannot
be directly converted into cell numbers, indicating that the
copy numbers acquired by qPCR do not necessarily indi-
cate the density of the bacteria in the microbial community.
This also occurs for anammox bacteria: they may have
different 16S rRNA or hzo operon copy numbers depend-
ing on species, which warrants future study. Therefore, the
results obtained by qPCR should be double-checked using
known amounts of the clone-derived positive control to
confirm the putative effect of competition/inhibition of a
positive control with an actual sample (Tsushima et al.
2007a).
FISH
FISH relies on DNA/RNA hybridizations occurring within
whole microbial cells in situ. In situ hybridizations with
DNA oligonucleotides designed for the detection of spe-
cific bacteria are performed with fluorescent-labeled com-
pounds. Currently, FISH is well known as a powerful
diagnostic tool with widespread environmental and medi-
cal applications, and has been reviewed by numerous
researchers (Wagner et al. 2006; Amann and Fuchs 2008).
Numerous FISH probes have been described in literature
for the detection of phylogenetically different bacteria
based on the 16S rRNA gene in mixed bacterial commu-
nities, most of which are listed in probeBase (Loy et al.
2003) (http://www.microbial-ecology.de/probebase/). With
FISH, abundant rRNA molecules in bacterial ribosomes are
probed with fluorescently labeled DNA oligonucleotides
which target specific sites on the rRNA of targeted Bac-
teria, Archaea, and Eukarya. The DNA probes hybridize
with their target sites by fluorescence after either excess or
non-hybridized probes are removed. The target fluorescent
bacterial cells can be detected by epi-fluorescent micros-
copy, confocal laser scanning microscopy or flow cytom-
etry. There are fluorochrome variations with different
excitation/emission wavelengths. Multiple staining of
A. Terada et al.
123

different probes with different fluorochromes permits
simultaneous detection of different bacteria. Precaution is
required to check if the signal from one probe interferes the
signal from another, but the general protocol has been
validated (Amann 1995). Anammox bacterial probes are
available from phylum-specific (e.g., Neef et al. 1998) to
member-specific (e.g. Schmid et al. 2003) and genus-spe-
cific probes (e.g., Schmid et al. 2000, 2001; Kuypers et al.
2003; Kartal et al. 2007a, b), which are chosen depending
on the research objective. Probes that are widely used for
the detection of anammox bacteria are reviewed elsewhere
(Schmid et al. 2005; Daims et al. 2009).
The advantages of the FISH technique are to allow a
reliable result to be obtained; a reduced likelihood of
misinterpret artifacts than PCR-related techniques because
FISH does not amplify bacterial cells; and to provide the
spatial location of bacteria and their density in a limited
region such as biofilm or bacterial aggregate without the
destruction of the sample. A requirement for the success of
FISH is that a sample does not contain a large amount of
inert material, which can interfere with a positive signal
due to their auto-fluorescent nature. The basic morphology
of anammox bacteria are coccoid and the bacteria tend to
form a cluster-like structure (Egli et al. 2001) as also seen
with nitrifying bacteria (Schramm et al. 1996; Okabe et al.
1999).
When the activity in a targeted bacterial cell is low (i.e.,
low numbers of rRNA), the FISH technique may not pro-
vide bacterial cell detection using microscopy. In this case,
fluorescent signal amplification shows promise for detect-
ing bacterial cells. Catalyzed reporter deposition (CARD)-
FISH is one technique for dramatically increasing the
sensitivity of the FISH signal. This technique was first
introduced to microbial ecology in 2002 (Pernthaler et al.
2002). With CARD-FISH, the probes are not directly
labeled with fluorochromes but are covalently bound to
horseradish peroxidase (HRP). In an amplification step, this
enzyme catalyzes the formation of fluorochrome-labeled
tyramide radicals. These highly reactive molecules bind to
tyrosine rich regions of the ribosome and other proteins in
the vicinity. With this additional catalytic step, a cell
containing only a single ribosome can theoretically be
detected since the fluorescence signal is amplified many
times by the massive deposition of labeled tyramide. The
sensitivity can reportedly be amplified by a factor of 26–41
(Hoshino et al. 2008).
To summarize, CARD-FISH is ideal when conventional
FISH techniques have a low signal and/or high background
from hybridized cells. In particular, CARD-FISH has been
used to detect anammox bacteria in oligotrophic environ-
ments such as lakes (Hamersley et al. 2009) and marine
environments (Woebken et al. 2007; Galan et al. 2009).
The CARD-FISH methods have been optimized for the in
situ detection of anammox bacteria in biological waste-
water treatment (Pavlekovic et al. 2009).
Other modified FISH techniques have also been devel-
oped to minimize probe penetration problems and increase
hybridization efficiency. For example, peptide nucleic acid
FISH (PNA FISH) (Perry-O’Keefe et al. 2001) and locked
nucleic acid FISH (LNA FISH) (Kubota et al. 2006) could
be used for the detection of anammox bacteria under oli-
gotrophic conditions. The detailed principles underlying
these methods can be found in a review article (Amann and
Fuchs 2008).
Confocal Raman microscopy
In microbiology, it is feasible to differentiate bacteria down
to the strain level by Raman microscopy (Kirschner et al.
2001). The use of confocal Raman microscopy allows for
identification of the three-dimensional distribution of
anammox bacteria in aggregates, e.g., biofilms. Anammox
bacteria (enriched Ca. Brocadia anammoxidans) have been
identified using their Raman vibrational signature in a non-
invasive manner (Patzold et al. 2006). The modified tech-
nique, i.e., confocal resonance Raman microscopy
(CRRM), permits the use of the Raman resonance effects
on cytochrome c, which is abundant in an anammox cell
comprising more than 10% of the protein content of
anammox bacteria (Cirpus et al. 2005), so that anammox
bacteria can be non-invasively identified (Patzold et al.
2008). Unlike FISH, CRRM does not require pretreatment,
i.e., bacterial cell fixation by paraformaldehyde. Further-
more, it allows for the acquisition of reliable species-level
identification, which is an alternative for detecting anam-
mox bacteria in biofilms or granules if economically
feasible.
Lipid analysis as a biomarker for anammox bacteria
The lipid membrane composition of anammox bacteria has
been investigated by gas chromatography mass spectrom-
etry (GC–MS) and high field nuclear magnetic resonance
(NMR) spectrometry (Damste et al. 2002, 2004). Anam-
mox bacteria contain a membrane-bound intracytoplasmic
compartment, i.e., anammoxosome, where ammonium and
nitrite are converted to N2 gas via hydrazine, a toxic and
energy-rich intermediate (Damste et al. 2002). Unprece-
dented lipids, known as ladderane lipids, were discovered
in both Ca. Brocadia anammoxidans and Ca. Kuenenia
stuttgartiensis cells (Damste et al. 2002). Details can be
found in the review article (Jetten et al. 2009). The
ladderane lipids in anammox bacteria can be used as a
biomarker to show their presence. They have been detected
in natural environments, such as oceanic OMZs (Kuypers
et al. 2003, 2005) and marine sediments (Jaeschke et al.
Review of anammox detection, presence, and application
123

2009b, 2010). Recently, it has been revealed that the
composition of ladderane lipids in anammox bacteria
depends on applied temperatures, which may indicate the
possibility of determining the origin of ladderane lipids in
sediments (Rattray et al. 2010).
Microsensors
Microsensors, in the form of microelectrodes and mic-
rooptodes (fiberoptic microsensors), are very powerful
tools to determine a hot spot where anammox reactions
take place in sediments or biofilms at the micrometer level.
The tip diameter for microsensors can be fabricated down
to a few micrometers. Because the minimum resolution of
microsensors is twice as large as the tip diameter, the
interval of microsensor measurement, the minimum sample
depth is at the micrometer level. The microsensor tech-
nique allows the concentrations of various dissolved
compounds to be measured without sample destruction.
To measure the depth profiles of nitrogen species, i.e.,
ammonium, nitrite, and nitrate ions, ion-selective micro-
electrodes based on Ag/AgCl have been used with engi-
neered biofilms (Schramm et al. 1996; Okabe et al. 1999).
To measure oxygen, a Clark-type dissolved oxygen
microelectrode is commercially available (http://www.
unisense.com/). Decreases in the ammonium and nitrite
concentrations concomitant with nitrate production at the
same location indicate evidence of anammox reactions in a
biofilm (Kindaichi et al. 2007; Tsushima et al. 2007b; Cho
et al. 2010). The combination of nitrite (or nitrate) and
dissolved oxygen microelectrodes has been used to identify
hot spots of anammox reactions in a biofilm during com-
plete autotrophic nitrogen removal (coupling of partial
nitritation and anammox) (Nielsen et al. 2005), in man-
groves (Meyer et al. 2005) and in estuary sediments
(Risgaard-Petersen et al. 2004). The combination of FISH
with microsensor techniques provides information on the
identity and activity of anammox bacteria (Nielsen et al.
2005; Kindaichi et al. 2007; Cho et al. 2010).
15N isotope labeling technique
The use of the stable isotope 15N is a method for quanti-
fying anammox activity in an environmental sample. The
initial observation of nitrogen loss in a bioreactor system
has been confirmed by tracer experiments by applying 15N-
labeled ammonium (Mulder et al. 1995; van de Graaf et al.
1995). Since then, isotope experiments with 15N-labeled
ammonium (or 15N-labeled nitrate) and non-labeled nitrite
(14N) have been performed to detect anammox activity
(Thamdrup and Dalsgaard 2002; Hamersley et al. 2007;
Schmid et al. 2007; Dale et al. 2009). Given the anammox
reaction pathway, combining 15N-labeled ammonium with
14N-labeled nitrite or nitrate results in the expected mixed
labeled 14?15N2, which is evidence of the anammox reac-
tion and can be differentiated from the denitrification
pathway, which produces 14?14N2. In addition to the
detection of anammox activity, the use of 15N-labeled
nitrate has revealed anammox versatility participating in
DNRA (Kartal et al. 2007a). Furthermore, an 15N tracer
study has been used to test anammox inhibition in marine
sediments (Jensen et al. 2007).
Anaerobic ammonium oxidation in natural
environments
Although anammox was speculated as a possible pathway
for producing N2 in the ocean several decades ago, the first
observation of anammox bacteria was confirmed in a
wastewater treatment reactor (Mulder et al. 1995). Since
then, a number of studies have reported the presence of
anammox in natural environments such as marine sedi-
ments (Thamdrup and Dalsgaard 2002; Dalsgaard et al.
2003; Kuypers et al. 2003), marine sponges (Mohamed
et al. 2010), estuarine/tidal river sediments (Trimmer et al.
2003; Rysgaard et al. 2004; Meyer et al. 2005; Dale et al.
2009), deep-sea hydrothermal vents (Byrne et al. 2009), hot
spring (Jaeschke et al. 2009a), and some freshwater eco-
systems (Schubert et al. 2006; Erler et al. 2008; Hamersley
et al. 2009). The discovery and potential contribution of
anammox bacteria to N2 production is important because it
may necessitate a re-evaluation of nitrogen transformation
processes in the global nitrogen budget. For example, some
evidence from marine ecosystems suggests that anammox
may in fact be the dominant nitrogen loss process in anoxic
marine water columns (Dalsgaard et al. 2003; Kuypers
et al. 2003).
Inland freshwater ecosystems such as lakes, rivers, and
wetlands have an important role in the ecological systems
on earth and may provide suitable habitats for diverse
nitrogen cycle bacteria including anammox bacteria. In
particular, ecosystems with low dissolved oxygen concen-
trations in surface water and the availability of both
ammonium and nitrite/nitrate are considered as prime
environments for the anammox process. The presence
of the anammox process has been confirmed in lakes
(Schubert et al. 2006) and anammox bacteria have also been
detected in inland rivers (Zhang et al. 2007). Furthermore,
nitrogen transformations in soil planted with forage rice
plants (Zhou et al. 2009) and bulrushes (Dong and Reddy
2010) have also indicated the occurrence of the anammox
process. As anthropogenic eutrophication increases nutrient
loading to freshwater bodies, there is an increasing need to
understand the anammox process in freshwater ecosystems
in more detail to prevent eutrophication.
A. Terada et al.
123

Diversity of anammox bacteria in various natural
ecosystems
The first marine anammox bacteria (Ca. Scalindua sor-
okinii) were reported in the Black Sea (Kuypers et al.
2003). Since then, Ca. Scalindua-like bacteria have been
detected in various marine ecosystems (Risgaard-Petersen
et al. 2004; Amano et al. 2007; Galan et al. 2009). A new
cluster, Ca. Scalindua arabiaca, was also observed in the
Arabian Sea (Woebken et al. 2008). Compared with the Ca.
Brocadia and Ca. Kuenenia genera that usually dominate in
wastewater treatment systems, Ca. Scalindua are mainly
present in natural environments, particularly in anoxic
marine environments. Studies on 11 anoxic marine sedi-
ments and water column samples from different seas found
that all detected anammox species were affiliated with the
genus Ca. Scalindua, and were grouped into Ca. Scalindua
brodae and Ca. Scalindua sorokinii (Schmid et al. 2007).
Similarly, most of the anammox bacteria identified by
cloning the 16S rRNA gene from the Seto Inland Sea are
also proximate to these two genera (Amano et al. 2007).
Tidal river estuaries and freshwater ecosystems appear
to harbor a greater diversity of anammox bacteria com-
pared with anoxic marine environments (Dale et al. 2009;
Humbert et al. 2010). These environments have various
zones where different nitrogen loading, oxygen concen-
tration, organic content, and salinity are present. In the
Cape Fear River estuary in the United States, an increase in
anammox bacterial diversity was observed along a salinity
gradient. Ca. Scalindua are the most halotolerant to high
salinity of the anammox genera, while Ca. Brocadia and
Ca. Kuenenia are less tolerant to salinity (Dale et al. 2009).
A similarly high anammox bacterial diversity was found in
the Yodo River estuary, where sequences affiliated with
Ca. Scalindua, Ca. Brocadia, and Ca. Kuenenia have been
recovered (Amano et al. 2007), and Ca. Scalindua wagneri
was predominant in the samples recovered from the same
river (Amano et al. 2007). In addition to a Ca. Scalindua-
affiliated clone identified by the 16S rRNA gene, Ca.
Brocadia-affiliated counterparts have also been recovered
from sediments of the inland Xinyi River in China,
showing a high anammox diversity in the samples (Zhang
et al. 2007). The anammox 16S rRNA gene sequences
recovered from Lake Rassnitzer in Germany are proximal
to Ca. Scalindua sorokinii/brodae, Ca. Scalindua wagneri,
and Ca. Brocadia fulgida (Hamersley et al. 2009), while
Ca. Scalindua brodae was identified in Lake Tanganyika in
Africa (Schubert et al. 2006). These results suggest a high
anammox bacterial diversity in freshwater ecosystems,
which is comparable to estuarine sediments.
Despite the detection of anammox bacteria at psychro-
philic and mesophilic temperatures (4–37�C) in natural
environments, anammox bacteria are recently reported to
be able to thrive at even higher temperature than 43�C
(Byrne et al. 2009; Jaeschke et al. 2009a; Li et al. 2010a),
which is reported to be the highest temperature to allow
enriched anammox culture Ca. Brocadia anammoxidans in
a laboratory to maintain the activity (Strous et al. 1999b).
These reports have revealed that anammox bacteria were
present and active at 60–85�C in deep-hydrothermal vents
(Byrne et al. 2009), at 52�C in California and Nevada hot
springs (Jaeschke et al. 2009a) and at 55–75�C in petro-
leum reservoirs (Li et al. 2010a). These anammox bacteria
are affiliated to not only the known anammox bacterial
genera, i.e., Ca. Brocadia, Ca. Kuenenia, and Ca. Scalin-
dua but also, intriguingly, new deep-branching phylotypes
(Byrne et al. 2009; Jaeschke et al. 2009a; Li et al. 2010a).
The phylogeny and physiology of the new anammox clades
should be elucidated in a stably controlled environment,
leading to better understanding of the nitrogen cycle in
natural environments and, in future, use of these bacteria
for engineered systems.
Contribution of anammox to total N2 flux in various
natural ecosystems
Nitrogen production via the anammox reaction has been
detected using 15N isotope labeling techniques in various
natural environments, which are typically performed with
homogenized anoxic sediment or anoxic water. The con-
tribution of anammox to total N2 flux ranges from a small
percentage to approximately 70% depending on different
environmental conditions (Table 1). At the sites in the
Celtic Sea (water depth: 500–2000 m), the highest contri-
bution of anammox to total N2 production was 65%, while
the mean anammox contribution to total N2 production at
an Irish Sea shelf (water depth: 50–100 m) was 33% (Ja-
eschke et al. 2009b). In Skagerrak, at two typical conti-
nental shelf sites (water depths: 380 and 695 m) in the
Danish belt seaway, N2 production via anammox accoun-
ted for 24 and 67% of the total N2 production, indicating
that anammox is occasionally more dominant than deni-
trification for the removal of fixed nitrogen from deep
sediments (Thamdrup and Dalsgaard 2002). Similarly, the
anammox contribution to N2 production accounted for
19–35% of the total N2 production in the water column at
Golfo Dulce (water depth: 200 m), Costa Rica (Dalsgaard
et al. 2003), and accounted for 1–35% in coastal sediments
in East and West Greenland (water depth: 36–100 m)
(Rysgaard et al. 2004).
Sediments in estuaries also allow anammox bacteria to
grow and contribute to nitrogen loss; however, the degree
of contribution of anammox reaction to nitrogen loss is
quite diverse. The contribution of anammox to total N2
production ranged from 5 to 24% in the sediment of
Review of anammox detection, presence, and application
123

Randers Fjord (water depth: 1 m) (Risgaard-Petersen et al.
2004). On the other hand, marginal anammox activities
were observed relative to denitrification in an eutrophic
coastal bay in Aarhus (water depth: 16 m) (Thamdrup and
Dalsgaard 2002) and a shallow-water estuary in Norsminde
Fjord (water depth: 0.5 m) (Risgaard-Petersen et al. 2004).
Estuaries also provide spatial gradients of nitrogen com-
pounds and dissolved oxygen, creating different niches for
anammox bacteria. In the Thames River estuary, anammox
was responsible for 8% of the total N2 production at the
inland side but this decreased along the estuarine gradient
to \1% at the mouth of the estuary, congruent with the
decrease in anammox activity (Trimmer et al. 2003). A
similar trend was discovered in a subtropical tidal river
system (Logan/Albert River), where the anammox rate
increased with increasing distance from the mouth of the
river (Meyer et al. 2005). In the Cape Fear River estuary,
the anammox activity was somewhat lower than in the
Thames River, but the contribution of anammox to the total
N2 production was higher (3.8–16.5%) (Dale et al. 2009).
These findings indicate that environmental conditions such
as salinity variation probably influence the diversity, dis-
tribution, and eventually, activity of anammox bacteria in
estuaries. Different anammox bacterial communities,
comprised of Ca. Brocadia, Ca. Kuenenia, Ca. Jettenia or
Ca. Scalindua, were detected at sites along an estuarine
gradient with the highest anammox rate found at the site
where Ca. Scalindua dominated with the highest anammox
bacterial abundance (Dale et al. 2009).
Recently, a few reports have been published relating to
anammox in inland freshwater ecosystems. In Lake
Tanganyika, the rate of N2 production through anammox
in an anoxic water column at both 100 and 110 m depths
was up to 0.01 nmol N ml-1 h-1 (Schubert et al. 2006),
similar to those measured in Golfo Dulce (Dalsgaard et al.
2003). The contribution of anammox to total N2 produc-
tion at the different depths was 9 and 13%, respectively.
It appears that anammox activity depends on the season.
In another anoxic water column at Lake Rassnitzer, the
anammox activity in January and October (max.
0.0225 nmol N ml-1 h-1) was significantly lower than in
May (Hamersley et al. 2009). The anammox activity
varied from 0.13 to 3.91 nmol ml-1 h-1 in a constructed
wetland and the presence of oxygen determined the
anammox contribution to N2 production from 0.1 to 30%,
indicating the significance of oxygen for anammox
activity (Erler et al. 2008). However, information on the
contribution of anammox activity to total N2 production
in inland ecosystems has yet to be well understood and
compared with marine and estuarine environments.
Table 1 Anammox activity in various natural ecosystems
Location Anammox rate
(nmol N ml-1 h-1)
A/T (%) NH4?
(lM)
NOx-
(lM)
Salinity
(psu)
Org. C
(%)
Reference
Marine ecosystems
Celtic Sea 1.3–2.8a 65 \130 \140 ND 0.04–0.49 Jaeschke et al. (2009a, b)
Irish Sea 2.1–25.7a 33 \130 \100 ND 0.82–1.36 Jaeschke et al. (2009a, b)
Arctic Ocean 0.17–15 1–35 ND 0.3–15.3 31.7–33.2 0.3–3.2 Rysgaard et al. (2004)
Golfo Dulce 0.042–0.061b 19–35 0–0.3 0.5–15 35 ND Dalsgaard et al. (2003)
Skagerrak 1.25–4.1 24–67 1.4–3.6 35 34.7 ND Thamdrup and Dalsgaard (2002)
Estuaries and tidal rivers
Aarhus Bay 3.46 2 ND \19 ND ND Thamdrup and Dalsgaard (2002)
Randers Fjord 3.8–5.9 5–20 ND 15–300 3–15 4.2 Risgaard-Petersen et al. (2004)
Norsminde Fjord 0 0 ND \430 3–23.5 4.4 Risgaard-Petersen et al. (2004)
Cape Fear 0.033–0.33c 3.8–16.5 4–5.2 5.5–6.9 0.4–9.9 ND Dale et al. (2009)
Thames River 0.4–9.9 \1–8 ND ND 2–30% 0.26–3.6 Trimmer et al. (2003)
Logan/Albert River 0–8 0–9 0.3–53 0.8–93 ND ND Meyer et al. (2005)
Yodo River 0.14–0.33 1–2 ND ND ND ND Amano et al. (2007)
Inland water bodies
Lake Tanganyika 0–0.01 9–13 0–27 0–10 ND ND Schubert et al. (2006)
Lake Rassnitzer 0–0.0225 20 or 50 0–300 0–200 3–23 ND Hamersley et al. (2009)
Constructed wetland 0.04–3.91 0.1–30 ND 10 ND ND Erler et al. (2008)
A/T contribution of anammox to total N2 production, ND no dataa nmol 29N2 ml-1 (overnight)b lmol N m-2 day-1
c nmol N g-1 h-1
A. Terada et al.
123

Variations in the contribution of anammox to N2 pro-
duction relative to denitrification is attributed to changes in
sediment reactivity, i.e., the availability of organic matter
in sediment for denitrifying bacteria. Following investiga-
tions in Aarhus Bay and Norsminde Fjord estuary, anam-
mox activity seems to become insignificant when sediment
reactivity increases in enriched estuarine sediments.
Although anammox rates in Randers Fjord were compa-
rable with those measured previously in the Skagerrak
sediment (Thamdrup and Dalsgaard 2002), the contribution
of anammox to total N2 production in Randers Fjord was
significantly lower than in Skagerrak sediment. This indi-
cates that the denitrification rates in Randers Fjord are
10–15 times higher than in the Skagerrak sediment, prob-
ably due to the greater availability of organic matter in the
Randers Fjord sediment.
The supply of nitrite is another key driver for both
anammox and denitrification. In natural environments,
nitrite is provided by ammonium oxidation by AOB or
AOA, or by nitrate reduction by heterotrophic bacteria
(Meyer et al. 2005). When nitrite is available, the anam-
mox bacteria consume ammonium from sediments
(Thamdrup and Dalsgaard 2002). The presence and
absence of anammox bacteria in Randers Fjord and
Norsminde Fjord, respectively, stems from differences in
the availability of nitrite in the suboxic zone of the sedi-
ment (Risgaard-Petersen et al. 2004). In OMZs, the nitro-
gen cycle including anammox has been substantially
revised. Lam et al. (2009) have identified that anammox
bacteria obtained[67% of nitrite from nitrate reduction in
the Eastern Tropical South Pacific OMZ, which is much
larger contribution than nitrite from ammonium oxidation.
Furthermore, DNRA is important in OMZs to provide
anammox bacteria with ammonium, which concludes the
complicated picture of nitrogen cycling in the Eastern
Tropical South Pacific OMZ (Lam et al. 2009).
Anaerobic ammonium oxidation in engineered systems
The paper by Strous et al. (1999a), which concluded that
the anammox reaction is biologically mediated by anam-
mox bacteria, paved the way for the application of anam-
mox-related technologies and studies on anammox
biochemistry and phylogeny. As shown in Fig. 1, the
number of publications especially related to the application
of anammox-related bioreactors for wastewater treatment
has increased explosively since 2000. Comparisons of
bioreactor system configurations, objectives and perfor-
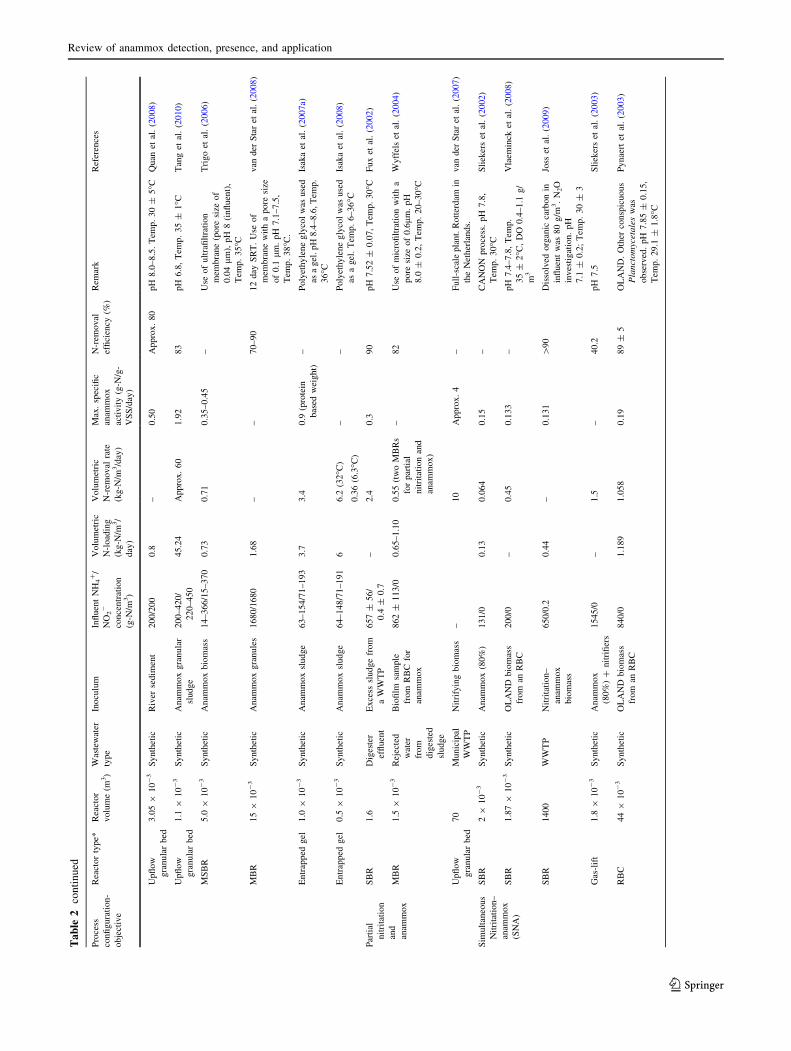
mance are tabulated in Table 2. Numerous biochemical
processes have been developed and various wastewater
types have been used to test the feasibility of anammox
processes.
Bioreactor systems involving enrichment of anammox
bacteria
Currently, there are five known genera of anammox bac-
teria in the Planctomycetes phylum (Kuenen 2008; Jetten
et al. 2009). Although an anammox bacterium has not been
successfully isolated, the physiological properties of these
anammox bacteria have been revealed. The enrichment of
Ca. Kuenenia stuttgartiensis was successful up to 73%,
allowing the genome of the uncultured bacterium Kunuenia
stuttgartiensis to be assembled (Strous et al. 2006). Deci-
phering an environmental genome illustrates the evolu-
tional history of the bacterium and allows speculation into
the genetic blueprint of the bacterium properties. Taking
advantage of the special properties of anammox bacteria
allows the targeted anammox genus to be enriched pref-
erentially in a bioreactor system. For example, the adap-
tation of anammox biomass to high salinity conditions
allows Ca. Scalindua to be enriched in an SBR (Kartal
et al. 2006). This observation has been followed by reports
where Ca. Scalindua has been preferentially grown by
inoculating marine sediments (van de Vossenberg et al.
2008; Kawagoshi et al. 2009). The addition of acetate to
ammonium, nitrite, and nitrate provides an environment in
a bioreactor where Ca. Brocadia fulgida can outcompete
other bacteria (Kartal et al. 2008), while the presence of
both propionate and ammonium enriches Ca. Anam-
moxoglobus propionicus (Kartal et al. 2007b). Two species
affiliated with Ca. Scalindua and enriched in SBRs at 15
and 20�C, are also able to consume acetate, formate, and
propionate along with nitrate (van de Vossenberg et al.
2008).
Optimum temperature at which anammox bacteria show
the highest anammox activity depends on enriched species.
Ca. Brocadia-enriched biomass exhibited the highest
anammox activity at 40�C (Strous et al. 1999b) whereas the
highest activity was observed at 37�C in Ca. Kuenenia-
enriched biomass in a rotating biological contactor (RBC)
(Egli et al. 2001) and in multi-groups of anammox bacteria
entrapped in a polyethylene glycol gel carrier (Isaka et al.
2008). The optimum temperature range disagrees with the
counterpart at which the highest anammox activity is
attained in natural environments. Anammox bacteria dis-
covered in marine sediments exhibited the highest activity
at 25�C and lost the activity at 37�C (Dalsgaard and
Thamdrup 2002), indicating presence of anammox bacteria
with physiologically and phylogenetically different prop-
erties. Although no anammox activity was observed above
45�C in engineered systems, anammox bacteria are repor-
ted to be present even above 50�C in deep-hydrothermal
vents (Byrne et al. 2009), hot springs (Jaeschke et al.
2009a), and petroleum reservoirs (Li et al. 2010a). The
enrichment of these anammox bacteria and elucidation of
Review of anammox detection, presence, and application
123

Ta
ble
2P
roce
ssco
nfi
gu
rati
on
san
dp
erfo
rman
ceo
fan
amm
ox
-rel
ated
bio
reac
tors
Pro
cess
confi
gura
tion-
obje
ctiv
e
Rea
ctor
type*
Rea
ctor
volu
me
(m3)
Was
tew
ater
type
Inocu
lum
Infl
uen
tN
H4?
/
NO
2-
conce
ntr
atio
n
(g-N
/m3)
Volu
met
ric
N-l
oad
ing
(kg-N
/m3/
day
)
Volu
met
ric
N-r
emoval
rate
(kg-N
/
m3/d
ay)
Max
.sp
ecifi
c
anam
mox
acti
vit
y(g
-N/g
-
VS
S/d
ay)
N-r
emoval
effi
cien
cy(%
)
Rem
ark
Ref
eren
ces
Anam
mox
FB
R23
910
-3
Bak
ers
yea
st
pro
duct
ion
pla
nt
(aft
er
met
han
ogen
ic
pro
cess
)
Den
itri
fyin
g90–130/n
.d.
(for
NO
3-
)
0.7
–1.5
0.4
n.d
.n.d
.N
O3-
was
use
das
elec
tron
acce
pto
r.pH
7,
Tem
p.
36�C
Muld
eret
al.
(1995
)
FB
R2.5
910
-3
Synth
etic
Den
itri
fyin
g(f
rom
Muld
eret
al.
1995
)
70–420/7
04.8
3.1
0.0
21
65
NO
3-
was
also
added
.S
and
par
ticl
esw
ere
use
das
a
carr
ier.
pH
7,
Tem
p.
36�C
van
de
Gra
afet
al.(1
995)
FB
R2.5
910
-3
Synth
etic
/slu
dge
dig
esti
on
effl
uen
t
Den
itri
fyin
g(f
rom
Muld
eret
al.
1995
)
70–840/7
0–/8
40
1.5
0.7
–1.5
0.1
584
(NH
4?
)/99
(NO
2-
)
San
dpar
ticl
esw
ere
use
das
a
carr
ier.
pH
8,
Tem
p.
36�C
Str
ous
etal
.(1
997)
Fix
edbed
2.5
910
-3
Synth
etic
Den
itri
fyin
g(f
rom
Muld
eret
al.
1995
)
70–840/7
0–/8
40
1.0
0.7
0.1
582
(NH
4?
)/99
(NO
2-
)
Sin
tere
dgla
ssw
asm
ounte
d.
pH
8,
Tem
p.
36�C
Str
ous
etal
.(1
997)
SB
R15
910
-3
Synth
etic
Den
itri
fyin
g(f
rom
Muld
eret
al.
1995
)
63–420/6
3–420
3.2
3n.d
.0.0
54
n.d
.T
he
anam
mox
stoic
hio
met
ry
was
eval
uat
ed.
pH
7–8,
Tem
p.
32–33
�C
Str
ous
etal
.(1
998)
SB
R1
910
-3
Synth
etic
Munic
ipal
WW
TP
28–400/1
5–520
1.4
8–
0.1
882
pH
7.8
–8,
Tem
p.
35�C
Dap
ena-
Mora
etal
.
(2004a)
SB
R1
910
-3
Synth
etic
Nit
rify
ing
gra
nule
s50–375/5
0–375
0.7
5–
0.4
478
pH
7.8
–8,
Tem
p.
35�C
Dap
ena-
Mora
etal
.
(2004b
)
SB
R2
910
-3
Synth
etic
Anam
mox
(80%
)257/2
03
0.4
60.3
15
0.2
5–
pH
7.8
,T
emp.
30�C
Sli
eker
set
al.
(2002)
SB
R20
910
-3
Synth
etic
Mix
edbio
mas
s
from
leac
hat
ean
d
urb
antr
eatm
ent
pla
nts
250–1268/
263–1661
1.6
0–
–79.9
–99.9
Anam
mox
abundan
ce85.8
%
atth
een
d.
pH
7.2
–8.7
,
Tem
p.
36.0
±0.3
�C
Lopez
etal
.(2
008
)
Gas
-lif
t7
910
-3
Synth
etic
Nit
rify
ing
gra
nule
s28–900/1
4–1100
2.0
–0.9
988
pH
7.8
–8,
Tem
p.
30�C
Dap
ena-
Mora
etal
.
(2004b
)
Gas
-lif
t1.8
910
-3
Synth
etic
Anam
mox
(80%
)?
nit
rifi
ers
1360/1
370
–8.9
–93
pH
7.5
Sli
eker
set
al.
(2003)
UA
SB
6.4
910
-3
Synth
etic
Met
han
ogen
ic
gra
nule
s
65–510/6
8–700
–2.9
––
pH
7.5
,T
emp.
30�C
Imaj
oet
al.
(2004)
UA
SB
0.2
Synth
etic
Met
han
ogen
ic
gra
nule
s
65–510/6
8–700
–6.4
0.1
2–0.1
386.5
–92.3
pH
7.5
,T
emp.
30�C
Imaj
oet
al.
(2004)
Upfl
ow
colu
mn
bed
14
910
-3
Synth
etic
Den
itri
fyin
g
bio
mas
s
250/2
50
1.6
1.0
3–
69
±8
Use
of
non-w
oven
shee
t.pH
7.8
9±
0.5
6,
Tem
p.3
0�C
Furu
kaw
aet
al.
(2003)
Upfl
ow
colu
mn
bed
0.8
910
-3
Synth
etic
Munic
ipal
WW
TP
20–550/2
0–460
58.5
26.0
1.6
Appro
x.
60
Choic
eof
11
dif
fere
nt
WW
TP
sas
inocu
lum
.pH
7.0
–7.5
,T
emp.
37�C
.
Tsu
shim
aet
al.
(2007a)
Upfl
ow
colu
mn
bed
0.5
910
-3
Synth
etic
(mim
ickin
gse
a
wat
er)
Mar
ine
sedim
ent
21–100/6
.4–100
0.4
0.3
n.d
.A
ppro
x.
80
Ca.
Sca
lindua
was
enri
ched
.
Tem
p.
30
�CK
awag
osh
iet
al.
(2009)
Upfl
ow
gra
nula
rbed
1.2
59
10
-3
Synth
etic
Anam
mox
bio
mas
s100–300/
100–300
Appro
x.
32.0
17.4
–60
±7
pH
8,
Tem
p.
37
±1
�CC
ho
etal
.(2
010
)
A. Terada et al.
123
Ta
ble
2co
nti
nu
ed
Pro
cess
confi
gura
tion-
obje
ctiv
e
Rea
ctor
type*
Rea
ctor
volu
me
(m3)
Was
tew
ater
type
Inocu
lum
Infl
uen
tN
H4
?/
NO
2-
conce
ntr
atio
n
(g-N
/m3)
Volu
met
ric
N-l
oad
ing
(kg-N
/m3/
day
)
Volu
met
ric
N-r
emoval
rate
(kg-N
/m3/d
ay)
Max
.sp
ecifi
c
anam
mox
acti
vit
y(g
-N/g
-
VS
S/d
ay)
N-r
emoval
effi
cien
cy(%
)
Rem
ark
Ref
eren
ces
Upfl
ow
gra
nula
rbed
3.0
59
10
-3
Synth
etic
Riv
erse
dim
ent
200/2
00
0.8
–0.5
0A
ppro
x.
80
pH
8.0
–8.5
.T
emp.
30
±5�C
Quan
etal
.(2
008)
Upfl
ow
gra
nula
rbed
1.1
910
-3
Synth
etic
Anam
mox
gra
nula
r
sludge
200–420/
220–450
45.2
4A
ppro
x.
60
1.9
283
pH
6.8
,T
emp.
35
±1�C
Tan
get
al.
(2010
)
MS
BR
5.0
910
-3
Synth
etic
Anam
mox
bio
mas
s14–366/1
5–370
0.7
30.7
10.3
5–0.4
5–
Use
of
ult
rafi
ltra
tion
mem
bra
ne
(pore
size
of
0.0
4l
m),
pH
8(i
nfl
uen
t),
Tem
p.
35
�C
Tri
go
etal
.(2
006)
MB
R15
910
-3
Synth
etic
Anam
mox
gra
nule
s1680/1
680
1.6
8–
–70–90
12
day
SR
T.
Use
of
mem
bra
ne
wit
ha
pore
size
of
0.1
lm
.pH
7.1
–7.5
,
Tem
p.
38
�C.
van
der
Sta
ret
al.
(2008)
Entr
apped
gel
1.0
910
-3
Synth
etic
Anam
mox
sludge
63–154/7
1–193
3.7
3.4
0.9
(pro
tein
bas
edw
eight)
–P
oly
ethyle
ne
gly
col
was
use
d
asa
gel
.pH
8.4
–8.6
,T
emp.
36�C
Isak
aet
al.
(2007a)
Entr
apped
gel
0.5
910
-3
Synth
etic
Anam
mox
sludge
64–148/7
1–191
66.2
(32�C
)
0.3
6(6
.3�C
)
––
Poly
ethyle
ne
gly
col
was
use
d
asa
gel
.T
emp.
6–36�C
Isak
aet
al.
(2008)
Par
tial
nit
rita
tion
and
anam
mox
SB
R1.6
Dig
este
r
effl
uen
t
Exce
sssl
udge
from
aW
WT
P
657
±56/
0.4
±0.7
–2.4
0.3
90
pH
7.5
2±
0.0
7,
Tem
p.
30�C
Fux
etal
.(2
002
)
MB
R1.5
910
-3
Rej
ecte
d
wat
er
from
dig
este
d
sludge
Bio
film
sam
ple
from
RB
Cfo
r
anam
mox
862
±113/0
0.6
5–1.1
00.5
5(t
wo
MB
Rs
for
par
tial
nit
rita
tion
and
anam
mox)
–82
Use
of
mic
rofi
ltra
tion
wit
ha
pore
size
of
0.6
lm
.pH
8.0
±0.2
,T
emp.
20–30
�C
Wyff
els
etal
.(2
004)
Upfl
ow
gra
nula
rbed
70
Munic
ipal
WW
TP
Nit
rify
ing
bio
mas
s–
10
Appro
x.
4–
Full
-sca
lepla
nt.
Rott
erdam
in
the
Net
her
lands.
van
der
Sta
ret
al.
(2007)
Sim
ult
aneo
us
Nit
rita
tion–
anam
mox
(SN
A)
SB
R2
910
-3
Synth
etic
Anam
mox
(80%
)131/0
0.1
30.0
64
0.1
5–
CA
NO
Npro
cess
.pH
7.8
,
Tem
p.
30
�CS
liek
ers
etal
.(2
002)
SB
R1.8
79
10
-3
Synth
etic
OL
AN
Dbio
mas
s
from
anR
BC
200/0
–0.4
50.1
33
–pH
7.4
–7.8
,T
emp.
35
±2�C
,D
O0.4
–1.1
g/
m3
Vla
emin
cket
al.
(2008)
SB
R1400
WW
TP
Nit
rita
tion–
anam
mox
bio
mas
s
650/0
.20.4
4–
0.1
31
[90
Dis
solv
edorg
anic
carb
on
in
infl
uen
tw
as80
g/m
3.
N2O
inves
tigat
ion.
pH
7.1
±0.2
;T
emp.
30
±3
Joss
etal
.(2
009
)
Gas
-lif
t1.8
910
-3
Synth
etic
Anam
mox
(80%
)?
nit
rifi
ers
1545/0
–1.5
–40.2
pH
7.5
Sli
eker
set
al.
(2003)
RB
C44
910
-3
Synth
etic
OL
AN
Dbio
mas
s
from
anR
BC
840/0
1.1
89
1.0
58
0.1
989
±5
OL
AN
D.
Oth
erco
nsp
icuous
Pla
nct
om
ycet
ale
sw
as
obse
rved
.pH
7.8
5±
0.1
5,
Tem
p.
29.1
±1.8
�C
Pynae
rtet
al.
(2003
)
Review of anammox detection, presence, and application
123

Ta
ble
2co
nti
nu
ed
Pro
cess
confi
gura
tion-
obje
ctiv
e
Rea
ctor
type*
Rea
ctor
volu
me
(m3)
Was
tew
ater
type
Inocu
lum
Infl
uen
tN
H4?
/
NO
2-
conce
ntr
atio
n
(g-N
/m3)
Volu
met
ric
N-l
oad
ing
(kg-N
/m3/
day
)
Volu
met
ric
N-r
emoval
rate
(kg-N
/
m3/d
ay)
Max
.sp
ecifi
c
anam
mox
acti
vit
y(g
-N/
g-V
SS
/day
)
N-r
emoval
effi
cien
cy(%
)
Rem
ark
Ref
eren
ces
RB
C50
910
-3
Synth
etic
?dig
este
r
effl
uen
t
Nit
rify
ing
bio
mas
s,
foll
ow
edby
anae
robic
gra
nula
rsl
udge
880/0
.15
–1.7
95
n.d
.–
pH
7.7
8±
0.2
6,
22.4
±1.9
�CP
ynae
rtet
al.
(2004
)
RB
C2.8
910
-3
Synth
etic
?B
lack
wat
er
OL
AN
Dbio
mas
s
from
anR
BC
1000/0
(CO
D
600
g/m
3)
–1.2
98
(Synth
etic
)/
0.7
15
(Bla
ck)
0.0
68
76
OL
AN
D.
pH
7.2
–7.7
,T
emp.
25.8
±0.4
�C,
DO
0.7
–1.1
g/m
3
Vla
emin
cket
al.
(2009)
FF
BR
8.1
910
-3
Synth
etic
?dig
este
r
effl
uen
t
Nit
rify
ing
bio
mas
s,
foll
ow
edby
anae
robic
gra
nula
rsl
udge
880/0
.15
–0.5
83
0.4
34
–T
emp.
22.4
±1.9
�CP
ynae
rtet
al.
(2004
)
SN
AP
5.4
39
10
-3
Synth
etic
Nit
rify
ing
bio
mas
s
10–100/0
0.4
8–
–60–80
Fil
led
wit
hhydro
phil
icnet
-
type
acry
lfi
ber
mat
eria
l.
pH
7.5
–7.7
,T
emp.
35�C
,
DO
2–3
g/m
3
Furu
kaw
aet
al.
(2006)
DE
MO
N�
500
Rej
ect
wat
erfr
om
dig
este
dsl
udge
––
0.6
–0.1
1–0.3
7
(CO
Dbas
e)
–S
BR
-type.
pH
contr
ol.
DO
0.3
g/m
3.
Str
asse
inA
ust
ria
Inner
ebner
etal
.(2
007
)
MA
BR
49
10
-3
Synth
etic
Anam
mox
bio
mas
s
200/0
0.8
70.7
7–
89
pH
7.9
,T
emp.
35�C
Gong
etal
.(2
007
)
MA
BR
2.4
910
-3
Synth
etic
Nit
rify
ing
bio
mas
s,
foll
ow
edby
anam
mox
bio
mas
s
530–780/0
1.1
30.7
8–
69
Seq
uen
tial
aera
tion,
Tem
p.
32�C
Pel
lice
r-N
acher
etal
.
(2010)
*A
bbre
via
tions
are
list
edbel
ow
:C
AN
ON
com
ple
teau
totr
ophic
nit
rogen
rem
oval
over
nit
rite
,D
EM
ON
�dea
mm
onifi
cati
on,
FB
Rfl
uid
ized
bed
reac
tor,
FF
BR
fixed
film
bio
reac
tor,
n.d
.not
det
erm
ined
,M
AB
Rm
embra
ne-
aera
ted
bio
film
reac
tor,
MB
Rm
embra
ne
bio
reac
tor,
MSB
Rm
embra
ne
sequen
cing
bat
chre
acto
r,R
BC
rota
ting
bio
logic
alco
nta
ctor,
SB
Rse
quen
cing
bat
chre
acto
r,SN
Asi
mult
aneo
us
nit
rita
tion
and
anam
mox,
SN
AP
single
-sta
ge
nit
rogen
rem
oval
usi
ng
anam
mox
and
par
tial
nit
rita
tion,
UA
SB
upfl
ow
anae
robic
sludge
bla
nket
,W
WT
Pw
aste
wat
ertr
eatm
ent
pla
nt,
–not
men
tioned
ina
man
usc
ript
A. Terada et al.
123

their detailed physiology would pave the way for new
paradigm of anammox research.
An extremely high nitrogen loading of more than 10 kg-
N/m3/day increases the likelihood of accumulating mono-
species anammox bacterium: e.g., a Ca. Brocadia-like
species with a 16S rRNA gene sequence similarity of
95.7% was enriched in an anammox cell aggregate known
as a granule with more than 80% dominance (Cho et al.
2010). These reports clearly indicate that the anammox
genera have unusual characteristics, not only utilizing both
ammonium and nitrite but also having a unique property
that is not outcompeted by the other anammox genera. This
type of enrichment strategy would ultimately be used for
genome analysis without isolating each bacterium.
The competition between Ca. Brocadia and Ca. Ku-
enenia, both of which are commonly found in engineered
systems for wastewater treatment, has been argued. The
affinity constants for nitrite are reported to be 2.8–42 mg-
N/L for Ca. Kuenenia (van der Star et al. 2008) and
\70 mg-N/L for Ca. Brocadia (Strous et al. 1999b). It has
been reported that Ca. Brocadia are less likely to grow at
high nitrite concentrations because of toxicity (Gaul et al.
2005), but recent publications disagree with that observa-
tion, with Ca. Brocadia detected even at high nitrite con-
centrations (Kindaichi et al. 2007; van der Star et al. 2007;
Cho et al. 2010). Given the population shift from a Ca.
Brocadia-dominant inoculum composition to Ca. Kuenenia
dominance in a membrane bioreactor (MBR) with a 16 day
SRT, van der Star et al. (2008) hypothesized that Ca.
Kuenenia is an affinity (K) strategist while Ca. Brocadia is
a growth rate (r) strategist, based on their observation of
MBR performance and FISH results (van der Star et al.
2008). The physiological properties of these two genera are
still being discussed, but the elucidation of such properties
may allow for control of the anammox community and
improvements in anammox process performance and
determine biokinetic parameters for the implementation of
a more accurate mathematical model for the prediction of
process performance.
In series partial nitritation and anammox processes
A prerequisite of the anammox process for partial nitrita-
tion and anammox (otherwise known as complete auto-
trophic nitrogen removal) is to create an environment
where the anammox bioreactor receives wastewater with
ammonium and nitrite concentrations appropriate to the
stoichiometry for the anammox reaction as shown in Eq. 1.
Thus, the ammonium has to be partly oxidized to nitrite
with an ammonium:nitrite ratio of 1:1.32 for the sub-
sequent anammox reaction (Strous et al. 1998). Hence, the
preliminary step prior to the anammox reaction, nitritation,
needs to occur in a stable process.
The single reactor system for high ammonium removal
over nitrite (SHARON) process is one possible process for
achieving the nitritation (Hellinga et al. 1998). This process
was originally developed for conventional nitrification and
heterotrophic denitrification via nitrite but has been applied
for complete autotrophic nitrogen removal. The SHARON
process takes advantage of the high-temperature of the
sludge digester after anaerobic digestion, allowing a higher
specific growth rate for AOB than NOB (Mulder et al.
2001). Because the growth rate of anammox bacteria
reaches at maximum at temperatures around 30–40�C
(Strous et al. 1999b; Egli et al. 2001; Dosta et al. 2008;
Isaka et al. 2008), the high-temperature would be particu-
larly beneficial for the anammox reaction. Generally,
investigations into the anammox reaction found in engi-
neered systems for wastewater treatment have been per-
formed at moderate temperatures of around 30–43�C as
shown in Table 2. Anammox bacteria in nature are typi-
cally detected when the temperature is well below 30�C
(Dalsgaard and Thamdrup 2002; Rysgaard et al. 2004), so
there is no apparent limit to the application of anammox at
approximately 20�C (Isaka et al. 2007b, 2008; Pathak et al.
2007). A minimum temperature of 18�C is reportedly a
threshold for the success or failure of a stable anammox
reaction (Dosta et al. 2008). Temperatures lower than 18�C
have been tested over short periods: for example, anammox
activity was reported at 11�C (Egli et al. 2001) and even at
6�C with anammox bacteria entrapped in a gel matrix
(Isaka et al. 2008). However, the nitrogen conversion rate
dramatically decreased with a decrease in temperature:
from 6.0 kg-N/m3/day at 32�C to 0.45 kg-N/m3/day at
6.3�C (Isaka et al. 2008); from 0.6 kg-N/m3/day at 30�C to
0.3 kg-N/m3/day at 15�C (Dosta et al. 2008). The anam-
mox activity loss caused by temperature decrease is
potentially mitigated by control of influent nitrite concen-
tration (Isaka et al. 2007b) or acclimatization of anammox
bacteria to low-temperature conditions (Dosta et al. 2008).
Strategy of the adaptation to and the long-term stability at
lower temperatures warrants future study.
Various types of reactors have been developed along
with the studies of anammox physiology and phylogeny
as shown in Table 2. These include fluidized bed reactors
(FBRs) (Mulder et al. 1995; Strous et al. 1997), gas-lift
reactors (Sliekers et al. 2003), upflow anaerobic sludge
blanket (UASB) reactors (Imajo et al. 2004; Tang et al.
2010), upflow column bed reactors (Fujii et al. 2002;
Kindaichi et al. 2007; Tsushima et al. 2007b), SBRs
(Strous et al. 1998; Dapena-Mora et al. 2004b, c),
membrane sequencing batch reactors (MSBRs) (Trigo
et al. 2006), and RBCs (Koch et al. 2000; Egli et al.
2001).
Given the extremely slow growth rate of anammox
bacteria, ranging from 2 to 17 days (normalized for the
Review of anammox detection, presence, and application
123

different methods used to estimate the rate) (Isaka et al.
2006; Tsushima et al. 2007a; van der Star et al. 2007,
2008), the immobilization of anammox bacteria is impor-
tant for engineering purposes to avoid washout. There are
many ways to immobilize anammox bacteria including in
an entrapped gel (Isaka et al. 2007a), a polyester non-
woven sheet as a fixed bed (Fujii et al. 2002), and sand
particles as a fixed bed (Strous et al. 1997). The immobi-
lization of anammox bacteria ensures an extremely long
SRT, and this is a good strategy to retain anammox bacteria
in a bioreactor, where the anammox bacteria are reportedly
grown in an aggregate form. However, the periodic with-
drawal of anammox biomass enhances the disaggregation
of the biomass while wastewater is permeated through a
membrane. This disaggregation ultimately eliminates the
diffusion limitation of ammonium and nitrite, ensuring
high enrichment of the anammox bacteria (van der Star
et al. 2008).
The inoculum source and composition differ according
to research (Table 2). From Table 2, the inoculum com-
position does not seem to be a determining factor for
triggering the success or failure of the anammox reaction,
but is a factor for determining the startup time. For
instance, Tsushima et al. (2007b) quantified the anammox
16S rRNA gene copy number from 11 inocula by qPCR
and have successfully reduced the anammox emergence
time to 37 days by initiating an inoculum with the highest
anammox density. This indicates the significance of inoc-
ulum selection in terms of startup time reduction. Once a
full-scale reactor has been operated for some time, the
dissemination of the anammox system should be acceler-
ated, as the inoculum from the full-scale plant can be used
as the inoculum for another plant. In addition to the engi-
neering perspective, it is also of ecological interest to
observe the effect of inoculum composition on the resultant
anammox community, stoichiometry, and performance.
Theoretically, an infinite SRT is reportedly a driver for
determining microbial populations in a bioreactor, and this
is an inherent property of a biofilm system (Terada et al.
2010). It has been recognized that, due to the non-linear
dynamics of complicated microbial communities, the
functional redundancy of bioreactors positively correlates
with the SRT (Curtis et al. 2003). It has also been reported
that different inocula lead to distinctive anammox com-
munities after long-term anammox reactor operation (Date
et al. 2009). Future studies should address the relationship
between the resultant microbial community and anammox
performance, determining whether a diversified anammox
community is useful or not, e.g., when there are fluctua-
tions in the nitrogen loadings/concentrations or changes in
environmental factors.
Since anammox reactors have been shown to be suc-
cessful at the laboratory scale, the operation of a full-size
plant with a volume of 70 m3 has commenced in Rotter-
dam, The Netherlands. The reactor aims to treat highly
concentrated ammonium effluent from the sludge digester
at the Dokhaven–Sluisjesdijk WWTP (Population Equiva-
lent 470,000, nitrogen loading 830 kg/day) (van der Star
et al. 2007). The reactor has been in full operation since
2006, and the volumetric nitrogen removal rate has reached
8–10 kg-N/m3/day, twice as high as its design capacity
(van der Star et al. 2007; Kuenen 2008). As mentioned
above, part of the anammox biomass from a reactor can be
transplanted for the startup of another full-scale WWTP.
The rapid dissemination of anammox biomass would be
expected in the near future.
Simultaneous nitritation–anammox processes
in biofilms and granules
Nitritation and anammox processes can be coupled in a
single reactor. In this situation, biofilms or aggregates
provide a redox-stratification where oxygen is either pres-
ent or absent, ensuring appropriate environments for both
nitritation and anammox (Terada et al. 2007). Thus, the
concept of a one-step autotrophic nitrogen removal process
from NH4? has been realized in processes known as
complete autotrophic nitrogen removal over nitrite
(CANON) (Sliekers et al. 2003, 2004; Third et al. 2005),
oxygen-limited autotrophic nitrification–denitrification
(OLAND) (Kuai and Verstraete 1998; Pynaert et al. 2003,
2004; Vlaeminck et al. 2009), single-stage nitrogen
removal using anammox and partial nitritation (Furukawa
et al. 2006) and aerobic deammonification (Hippen et al.
1997; Innerebner et al. 2007; Wett 2007; Wett et al. 2010).
The original definitions of OLAND and aerobic deammo-
nification assumed that AOB, especially Nitrosomonas
species, were able to denitrify, but these definitions were
corrected after the discovery of anammox bacteria (Helmer
et al. 2001; Pynaert et al. 2003). Thus, each process is
basically the same and is referred to hereafter as simulta-
neous nitritation and anammox (SNA). The spatiotemporal
control of a biofilm, i.e., control of the oxygen concentra-
tion, can be achieved by growing SNA granules in an SBR
system (Sliekers et al. 2002; Vlaeminck et al. 2008).
The application of SNA for wastewater treatment is
potentially beneficial as the SNA process has a small foot-
print and avoids the accumulation of nitrite in biofilms or
granules, which can inhibit the growth of anammox bac-
teria (van de Graaf et al. 1995; Kimura et al. 2010). Full-
scale SBR-type SNA systems with maximum volumes of
500 and 1400 m3 at nitrogen loading rates of 340 kg-N/day
and 700 kg-N/day have been operated in Strasse, Austria
(Hippen et al. 1997; Innerebner et al. 2007; Wett 2007;
Wett et al. 2010) and in Zurich, Switzerland (Joss et al.
2009), respectively, treating reject water from digested
A. Terada et al.
123

sludge at high nitrogen removal efficiency of above 85%. A
laboratory-scale RBC for SNA has demonstrated high
nitrogen removal performance for the treatment of black
water with a volumetric nitrogen removal rate of 0.715 kg-
N/m3/day (Vlaeminck et al. 2009). As can be seen in
Table 2, the SNA process appears to be suitable for treat-
ment of inorganic or low organic carbon/nitrogen ratio
wastewater.
The engineering challenge in a single-stage SNA pro-
cess is to control the redox-stratification in the biofilm or
granule (Terada et al. 2007; Vazquez-Padin et al. 2010).
Sufficient nitrogen removal in a single-stage SNA requires
fastidious micro-environmental control, as oxygen over-
load and subsequent nitrite accumulation are harmful to
anammox activity (van de Graaf et al. 1996; Hao et al.
2002; Dapena-Mora et al. 2007). Thus, complicated mon-
itoring is required for a single-stage SND, including control
of dissolved oxygen in the bulk liquid, nitrite, and biofilm
thickness (Nielsen et al. 2005; Vazquez-Padin et al. 2010).
Changes in the air flow rate, using a controller for dissolved
oxygen concentration in the bulk liquid, potentially leads to
significant shear of the bacterial aggregate, compromising
biofilm rigidity (Tsuneda et al. 2004). Therefore, careful
control of the air flow rate and choice of an appropriate
aerator are necessary when scaling up a reactor.
The use of a gas-permeable membrane may be a
potential solution for oxygen control. The membrane is
basically used as a carrier for the biofilm formation and as
an oxygen-supplying material (Syron and Casey 2008).
The oxygen supply by a gas-permeable membrane provides
bubbleless aeration, allowing independent control of both
ammonium and oxygen loadings. This biofilm has a
counter-diffusion geometry where oxygen is supplied from
the bottom of the biofilm while ammonium is supplied
from the top of the biofilm. Preliminary modeling studies
have indicated the controllability of redox-stratification in a
biofilm on a gas-permeable membrane. Thus, a membrane-
aerated biofilm (MAB) can theoretically operate under a
wider range of ammonium and oxygen loadings to achieve
sufficient nitrogen removal efficiencies than a conventional
biofilm where oxygen and ammonium are supplied from
the same direction (Terada et al. 2007; Lackner et al.
2008). The performance of MAB reactors (MABRs) have
been investigated, showing the feasibility of the MABRs
for SNA and the comparable SNA performance with con-
ventional biofilm or granule systems (Gong et al. 2007;
Pellicer-Nacher et al. 2010). The performance of MABRs
is governed by the membrane surface area (Brindle et al.
1998), so the optimization of the membrane surface area
and the fabrication of a membrane module configuration
are challenges for obtaining the highest removal perfor-
mance of SNA and long-term reactor stability (Syron and
Casey 2008).
Outlook for anammox research and concluding
remarks
Anammox research has been one of hottest topics in the
field of microbial ecology, microbiology and biotechnol-
ogy. Since the discovery of the anammox reaction and
anammox bacteria, the research has dramatically acceler-
ated. Much of this is due to the value of multidisciplinary
collaborations. Intensive microbiological research has
identified the genomic blueprint and biosynthesis of
anammox bacteria, which has provided clues to elucidate
the behavior of anammox bacteria in natural environments
and to harness anammox bacteria in engineered systems.
Conversely, the techniques for enriching specialized
anammox microorganisms have opened the door to studies
on environmental genomes.
The use of the state-of-the-art molecular techniques has
allowed investigations into zones where anammox reac-
tions takes place and into anammox bacterial density/
diversity. Anammox reactions have been detected all over
the world. However, there are some areas where anammox
reaction is not necessarily predominant in marine envi-
ronment (Ward et al. 2009) and soils (Humbert et al. 2010),
which illustrates that not all natural environments neces-
sarily harbor a niche where anammox reactions are pre-
valent over denitrification. Ward et al. (2009) suggested
that a critical factor is a difference in the magnitude and
timing of the supply of organic matter to an OMZ, which
has been discussed elsewhere (Voss and Montoya 2009).
Future studies should aim to elucidate a factor that deter-
mines the predominance of the denitrification or anammox
reactions, allowing for a more complete understanding of
the nitrogen cycle in oceans and other natural environ-
ments. In addition, recent molecular evidence of anammox
bacteria under thermophilic and hyperthermophilic condi-
tions (Byrne et al. 2009; Jaeschke et al. 2009a; Li et al.
2010a) potentially allows better comprehension of the
nitrogen cycle in natural environments and application of
these bacteria to engineered systems.
An anammox-based process for treating high-strength
nitrogenous wastewater has already been matured,
achieving a volumetric nitrogen removal rate of more than
25 kg-N/m3/day in an upflow column bed reactor (Tsu-
shima et al. 2007b) and 45 kg-N/m3/day in a UASB (Tang
et al. 2010) when suitable conditions are met, e.g., at
temperature of 35�C. As described in Chapter on Biore-
actor systems involving enrichment of anammox bacteria,
anammox activities drastically decrease at lower tempera-
ture (Strous et al. 1999b; Egli et al. 2001; Isaka et al. 2008;
Dosta et al. 2008). Hence, a startup strategy should be
developed in order to reduce time for anammox emergence
and to stabilize anammox performance at temperature
below 25�C. As proposed by Dosta et al. (2008), the
Review of anammox detection, presence, and application
123

adaptation of anammox bacteria enriched under mesophilic
conditions may be a promising strategy. The use of
anammox bacteria present in psychrophilic conditions as
inoculum could be an alternative counterpart. These
investigations should be a future challenge to accelerate
dissemination of the anammox process.
The anammox reaction is not mediated through nitrous
oxide (N2O), but nitrite accumulation through nitrification
potentially emits large amounts of N2O, known to be a
highly potent greenhouse gas (Kampschreur et al. 2009).
The amount of N2O emissions from the partial nitritation–
anammox pathway is slightly higher than that through the
conventional nitrification–denitrification pathway, which
offsets the reduced emissions of carbon dioxide (Joss et al.
2009). As high nitrite concentrations from limited oxy-
genation is a major cause of N2O emissions through the
nitrification process (Kampschreur et al. 2009), a single-
stage SNA process, e.g., CANON, OLAND, SNAP,
MABR, and deammonification, could be promising for
applications in wastewater treatment considering the
greenhouse gas emissions. Reportedly, an MABR system
successfully reduces nitrous oxide emission 100-fold lower
than two-stage nitritation and anammox reactors (Pellicer-
Nacher et al. 2010), supporting the promise of the SNA
process. However, recent reports indicate that the SNA
process encounters difficulty to maintain stable partial
nitrification (Terada et al. 2010; Cho et al. 2011). There-
fore, the stabilization of redox-stratification in a biofilm or
granule is contingent upon the replacement of two-stage
nitritation and anammox with a single-stage SNA. In
summary, an overall estimation of the energy consumption
and impact on global warming may be required when
designing the most appropriate nitrogen removal process in
the near future.
Acknowledgments This work was supported by the Environment
Research and Technology Development Fund (RF-1002) of the
Ministry of the Environment, Japan and in part by Grants-in-Aid for
Scientific Research (C) (22510005) and Grant-in-Aid for Young
Scientists (B) (22710072) from the Ministry of Education, Culture,
Sports, Science, and Technology of Japan.
References
Ahn YH (2006) Sustainable nitrogen elimination biotechnologies: a
review. Process Biochem 41(8):1709–1721
Ahn YH, Choi HC (2006) Autotrophic nitrogen removal from sludge
digester liquids in upflow sludge bed reactor with external
aeration. Process Biochem 41(9):1945–1950
Ahn JH, Yu R, Chandran K (2008) Distinctive microbial ecology and
biokinetics of autotrophic ammonia and nitrite oxidation in a
partial nitrification bioreactor. Biotechnol Bioeng 100(6):1078–
1087
Amann R (1995) In situ identification of microorganisms by whole
cell hybridization with rRNA-targeted nucleic acid probes. In:
Akkermans ADC, van Elsas JD, de Brujin FJ (eds) Molecular
microbial ecology manual. Kluwer Academic Publishers, Dordr-
echt, pp 1–15
Amann R, Fuchs BM (2008) Single-cell identification in microbial
communities by improved fluorescence in situ hybridization
techniques. Nat Rev Microbiol 6(5):339–348
Amano T, Yoshinaga I, Okada K, Yamagishi T, Ueda S, Obuchi A,
Sako Y, Suwa Y (2007) Detection of anammox activity and
diversity of anammox bacteria-related 16S rRNA genes in
coastal marine sediment in Japan. Microbes Environ 22(3):232–
242
Balmelle B, Nguyen KM, Capdeville B, Cornier JC, Deguin A (1992)
Study of factors controlling nitrite buildup in biological
processes for water nitrification. Water Sci Technol 26(5–6):
1017–1025
Bernet N, Sanchez O, Cesbron D, Steyer JP, Delgenes JP (2005)
Modeling and control of nitrite accumulation in a nitrifying
biofilm reactor. Biochem Eng J 24(2):173–183
Brindle K, Stephenson T, Semmens MJ (1998) Nitrification and
oxygen utilisation in a membrane aeration bioreactor. J Memb
Sci 144(1–2):197–209
Brochier C, Philippe H (2002) Phylogeny—a non-hyperthermophilic
ancestor for bacteria. Nature 417(6886):244
Broda E (1977) 2 kinds of lithotrophs missing in nature. Z Allg
Mikrobiol 17(6):491–493
Byrne N, Strous M, Crepeau V, Kartal B, Birrien JL, Schmid M,
Lesongeur F, Schouten S, Jaeschke A, Jetten M, Prieur D,
Godfroy A (2009) Presence and activity of anaerobic ammo-
nium-oxidizing bacteria at deep-sea hydrothermal vents. ISME J
3(1):117–123
Chavan PV, Dennett KE, Marchand EA (2008) Behavior of pilot-
scale constructed wetlands in removing nutrients and sediments
under varying environmental conditions. Water Air Soil Pollut
192(1–4):239–250
Cho S, Takahashi Y, Fujii N, Yamada Y, Satoh H, Okabe S (2010)
Nitrogen removal performance and microbial community anal-
ysis of an anaerobic up-flow granular bed anammox reactor.
Chemosphere 78(9):1129–1135
Cho S, Fujii N, Lee T, Okabe S (2011) Development of a
simultaneous partial nitrification and anaerobic ammonia oxida-
tion process in a single reactor. Bioresour Technol 102(2):
652–659
Cirpus IEY, de Been M, Op den Camp HJM, Strous M, Le Paslier D,
Kuenen GJ, Jetten MSM (2005) A new soluble 10 kDa
monoheme cytochrome c-552 from the anammox bacterium
Candidatus ‘‘Kuenenia stuttgartiensis’’. FEMS Microbiol Lett
252(2):273–278
Curtis TP, Head IM, Graham DW (2003) Theoretical ecology for
engineering biology. Environ Sci Technol 37(3):64A–70A
Daims H, Maixner F, Schmid MC (2009) The nitrifying microbes:
ammonia oxidizers, nitrite oxidizers, and anaerobic ammonium
oxidizers. In: Nielsen PH, Daims H, Lemmer H (eds) FISH
Handbook for Biological Wastewater Treatment: Identification
and quantification of microorganisms in activated sludge and
biofilms. FISH, IWA Publishing, London, pp 9–17
Dale OR, Tobias CR, Song BK (2009) Biogeographical distribution
of diverse anaerobic ammonium oxidizing (anammox) bacteria
in Cape Fear River Estuary. Environ Microbiol 11(5):1194–1207
Dalsgaard T, Thamdrup B (2002) Factors controlling anaerobic
ammonium oxidation with nitrite in marine sediments. Appl
Environ Microbiol 68(8):3802–3808
Dalsgaard T, Canfield DE, Petersen J, Thamdrup B, Acuna-Gonzalez
J (2003) N-2 production by the anammox reaction in the anoxic
water column of Golfo Dulce, Costa Rica. Nature 422(6932):
606–608
Damste JSS, Strous M, Rijpstra WIC, Hopmans EC, Geenevasen JAJ,
van Duin ACT, van Niftrik LA, Jetten MSM (2002) Linearly
A. Terada et al.
123

concatenated cyclobutane lipids form a dense bacterial mem-
brane. Nature 419(6908):708–712
Damste JSS, Rijpstra WIC, Schouten S, Fuerst JA, Jetten MSM,
Strous M (2004) The occurrence of hopanoids in planctomyce-
tes: implications for the sedimentary biomarker record. Org
Geochem 35(5):561–566
Dapena-Mora A, Arrojo B, Campos JL, Mosquera-Corral A, Mendez R
(2004a) Improvement of the settling properties of anammox
sludge in an SBR. J Chem Technol Biotechnol 79(12):1417–1420
Dapena-Mora A, Campos JL, Mosquera-Corral A, Jetten MSM,
Mendez R (2004b) Stability of the ANAMMOX process in a
gas-lift reactor and a SBR. J Biotechnol 110(2):159–170
Dapena-Mora A, Van Hulle SWH, Campos JL, Mendez R, Vanrol-
leghem PA, Jetten M (2004c) Enrichment of anammox biomass
from municipal activated sludge: experimental and modelling
results. J Chem Technol Biotechnol 79(12):1421–1428
Dapena-Mora A, Fernandez I, Campos JL, Mosquera-Corral A,
Mendez R, Jetten MSM (2007) Evaluation of activity and
inhibition effects on anammox process by batch tests based on
the nitrogen gas production. Enzyme Microb Technol 40(4):
859–865
Date Y, Isaka K, Ikuta H, Sumino T, Kaneko N, Yoshie S, Tsuneda S,
Inamori Y (2009) Microbial diversity of anammox bacteria
enriched from different types of seed sludge in an anaerobic
continuous-feeding cultivation reactor. J Biosci Bioeng 107(3):
281–286
Dong XL, Reddy GB (2010) Soil bacterial communities in con-
structed wetlands treated with swine wastewater using PCR-
DGGE technique. Bioresour Technol 101(4):1175–1182
Dosta J, Fernandez I, Vazquez-Padin JR, Mosquera-Corral A,
Campos JL, Mata-Alvarez J, Mendez R (2008) Short- and
long-term effects of temperature on the anammox process.
J Hazard Mater 154(1–3):688–693
Egli K, Fanger U, Alvarez PJJ, Siegrist H, van der Meer JR, Zehnder
AJB (2001) Enrichment and characterization of an anammox
bacterium from a rotating biological contactor treating ammo-
nium-rich leachate. Arch Microbiol 175(3):198–207
Erler DV, Eyre BD, Davison L (2008) The contribution of anammox
and denitrification to sedirnent N-2 production in a surface flow
constructed wetland. Environ Sci Technol 42(24):9144–9150
Francis CA, Beman JM, Kuypers MMM (2007) New processes and
players in the nitrogen cycle: the microbial ecology of anaerobic
and archaeal ammonia oxidation. ISME J 1(1):19–27
Fromin N, Hamelin J, Tarnawski S, Roesti D, Jourdain-Miserez K,
Forestier N, Teyssier-Cuvelle S, Gillet F, Aragno M, Rossi P
(2002) Statistical analysis of denaturing gel electrophoresis
(DGE) fingerprinting patterns. Environ Microbiol 4(11):634–643
Fujii T, Sugino H, Rouse JD, Furukawa K (2002) Characterization of
the microbial community in an anaerobic ammonium-oxidizing
biofilm cultured on a nonwoven biomass carrier. J Biosci Bioeng
94(5):412–418
Furukawa K, Rouse JD, Yoshida N, Hatanaka H (2003) Mass
cultivation of anaerobic ammonium-oxidizing sludge using a
novel nonwoven biomass carrier. J Chem Eng Jpn 36(10):
1163–1169
Furukawa K, Lieu PK, Tokitoh H, Fujii T (2006) Development of
single-stage nitrogen removal using anammox and partial
nitritation (SNAP) and its treatment performances. Water Sci
Technol 53(6):83–90
Fux C, Boehler M, Huber P, Brunner I, Siegrist H (2002) Biological
treatment of ammonium-rich wastewater by partial nitritation
and subsequent anaerobic ammonium oxidation (anammox) in a
pilot plant. J Biotechnol 99(3):295–306
Fux C, Huang D, Monti A, Siegrist H (2004) Difficulties in
maintaining long-term partial nitritation of ammonium-rich
sludge digester liquids in a moving-bed biofilm reactor (MBBR).
Water Sci Technol 49(11–12):53–60
Galan A, Molina V, Thamdrup B, Woebken D, Lavik G, Kuypers
MMM, Ulloa O (2009) Anammox bacteria and the anaerobic
oxidation of ammonium in the oxygen minimum zone off
northern Chile. Deep Sea Res Part II Top Stud Oceanogr 56(16):
1125–1135
Ganigue R, Lopez H, Balaguer MD, Colprim J (2007) Partial
ammonium oxidation to nitrite of high ammonium content urban
land fill leachates. Water Res 41(15):3317–3326
Gaul T, Marker S, Kunst S (2005) Start-up of moving bed biofilm
reactors for deammonification: the role of hydraulic retention
time, alkalinity and oxygen supply. Water Sci Technol 52(7):
127–133
Gong Z, Yang FL, Liu ST, Bao H, Hu SW, Furukawa KJ (2007)
Feasibility of a membrane-aerated biofilm reactor to achieve
single-stage autotrophic nitrogen removal based on anammox.
Chemosphere 69(5):776–784
Hamersley MR, Lavik G, Woebken D, Rattray JE, Lam P,
Hopmans EC, Damste JSS, Kruger S, Graco M, Gutierrez D,
Kuypers MMM (2007) Anaerobic ammonium oxidation in the
Peruvian oxygen minimum zone. Limnol Oceanogr 52(3):
923–933
Hamersley MR, Woebken D, Boehrer B, Schultze M, Lavik G,
Kuypers MMM (2009) Water column anammox and denitrifi-
cation in a temperate permanently stratified lake (Lake Rassn-
itzer, Germany). Syst Appl Microbiol 32(8):571–582
Hao XD, Heijnen JJ, van Loosdrecht MCM (2002) Sensitivity
analysis of a biofilm model describing a one-stage completely
autotrophic nitrogen removal (CANON) process. Biotechnol
Bioeng 77(3):266–277
Hellinga C, Schellen A, Mulder JW, van Loosdrecht MCM, Heijnen
JJ (1998) The SHARON process: an innovative method for
nitrogen removal from ammonium-rich waste water. Water Sci
Technol 37(9):135–142
Helmer C, Tromm C, Hippen A, Rosenwinkel KH, Seyfried CF,
Kunst S (2001) Single stage biological nitrogen removal by
nitritation and anaerobic ammonium oxidation in biofilm
systems. Water Sci Technol 43(1):311–320
Hippen A, Rosenwinkel KH, Baumgarten G, Seyfried CF (1997)
Aerobic deammonification: a new experience in the treatment of
wastewaters. Water Sci Technol 35(10):111–120
Hoshino T, Yilmaz LS, Noguera DR, Daims H, Wagner M (2008)
Quantification of target molecules needed to by fluorescence in
situ hybridization (FISH) and catalyzed reporter deposition-
FISH. Appl Environ Microbiol 74(16):5068–5077
Humbert S, Tarnawski S, Fromin N, Mallet MP, Aragno M, Zopfi J
(2010) Molecular detection of anammox bacteria in terrestrial
ecosystems: distribution and diversity. ISME J 4(3):450–454
Imajo U, Tokutomi T, Furukawa K (2004) Granulation of anammox
microorganisms in up-flow reactors. Water Sci Technol
49(5–6):155–163
Innerebner G, Insam H, Franke-Whittle IH, Wett B (2007) Identifi-
cation of anammox bacteria in a full-scale deammonification
plant making use of anaerobic ammonia oxidation. Syst Appl
Microbiol 30(5):408–412
Isaka K, Date Y, Sumino T, Yoshie S, Tsuneda S (2006) Growth
characteristic of anaerobic ammonium-oxidizing bacteria in an
anaerobic biological filtrated reactor. Appl Microbiol Biotechnol
70(1):47–52
Isaka K, Date Y, Sumino T, Tsuneda S (2007a) Ammonium removal
performance of anaerobic ammonium-oxidizing bacteria immo-
bilized in polyethylene glycol gel carrier—anammox bacteria
immobilized in gel carrier. Appl Microbiol Biotechnol 76(6):
1457–1465
Review of anammox detection, presence, and application
123

Isaka K, Sumino T, Tsuneda S (2007b) High nitrogen removal
performance at moderately low temperature utilizing anaerobic
ammonium oxidation reactions. J Biosci Bioeng 103(5):486–490
Isaka K, Date Y, Kimura Y, Sumino T, Tsuneda S (2008) Nitrogen
removal performance using anaerobic ammonium oxidation at
low temperatures. FEMS Microbiol Lett 282(1):32–38
Jaeschke A, den Camp H, Harhangi H, Klimiuk A, Hopmans EC,
Jetten MSM, Schouten S, Damste JSS (2009a) 16S rRNA gene
and lipid biomarker evidence for anaerobic ammonium-oxidiz-
ing bacteria (anammox) in California and Nevada hot springs.
FEMS Microbiol Ecol 67(3):343–350
Jaeschke A, Rooks C, Trimmer M, Nicholls JC, Hopmans EC,
Schouten S, Damste JSS (2009b) Comparison of ladderane
phospholipid and core lipids as indicators for anaerobic ammo-
nium oxidation (anammox) in marine sediments. Geochim
Cosmochim Acta 73(7):2077–2088
Jaeschke A, Abbas B, Zabel M, Hopmans EC, Schouten S, Damste
JSS (2010) Molecular evidence for anaerobic ammonium-
oxidizing (anammox) bacteria in continental shelf and slope
sediments off northwest Africa. Limnol Oceanogr 55(1):365–
376
Jensen MM, Thamdrup B, Dalsgaard T (2007) Effects of specific
inhibitors on anarnmox and denitrification in marine sedirnents.
Appl Environ Microbiol 73(10):3151–3158
Jetten MSM (2008) The microbial nitrogen cycle. Environ Microbiol
10(11):2903–2909
Jetten MSM, Horn SJ, Van Loosdrecht MCM (1997) Towards a more
sustainable municipal wastewater treatment system. Water Sci
Technol 35(9):171–180
Jetten MSM, Sliekers O, Kuypers M, Dalsgaard T, van Niftrik L,
Cirpus I, van de Pas-Schoonen K, Lavik G, Thamdrup B, Le
Paslier D, Op den Camp HJM, Hulth S, Nielsen LP, Abma W,
Third K, Engstrom P, Kuenen JG, Jorgensen BB, Canfield DE,
Damste JSS, Revsbech NP, Fuerst J, Weissenbach J, Wagner M,
Schmidt I, Schmid M, Strous M (2003) Anaerobic ammonium
oxidation by marine and freshwater planctomycete-like bacteria.
Appl Microbiol Biotechnol 63(2):107–114
Jetten M, Schmid M, van de Pas-Schoonen K, Damste JSS, Strous M
(2005) Anammox organisms: enrichment, cultivation, and envi-
ronmental analysis. In: Leadbetter JR (ed) Environmental
microbiology, vol 397. Methods in enzymology, Elsevier
Academic Press, California, pp 34–57
Jetten MSM, van Niftrik L, Strous M, Kartal B, Keltjens JT, Op den
Camp HJM (2009) Biochemistry and molecular biology of
anammox bacteria. Crit Rev Biochem Mol Biol 44(2–3):65–84
Joss A, Salzgeber D, Eugster J, Konig R, Rottermann K, Burger S,
Fabijan P, Leumann S, Mohn J, Siegrist H (2009) Full-scale
nitrogen removal from digester liquid with partial nitritation and
anammox in one SBR. Environ Sci Technol 43(14):5301–5306
Junier P, Molina V, Dorador C, Hadas O, Kim OS, Junier T, Witzel
KP, Imhoff JF (2010) Phylogenetic and functional marker genes
to study ammonia-oxidizing microorganisms (AOM) in the
environment. Appl Microbiol Biotechnol 85(3):425–440
Kampschreur MJ, Temmink H, Kleerebezem R, Jetten MSM, van
Loosdrecht MCM (2009) Nitrous oxide emission during waste-
water treatment. Water Res 43(17):4093–4103
Kartal B, Koleva M, Arsov R, van der Star W, Jetten MSM, Strous M
(2006) Adaptation of a freshwater anammox population to high
salinity wastewater. J Biotechnol 126(4):546–553
Kartal B, Kuypers MMM, Lavik G, Schalk J, den Camp H, Jetten
MSM, Strous M (2007a) Anammox bacteria disguised as
denitrifiers: nitrate reduction to dinitrogen gas via nitrite and
ammonium. Environ Microbiol 9(3):635–642
Kartal B, Rattray J, van Niftrik LA, van de Vossenberg J, Schmid
MC, Webb RI, Schouten S, Fuerst JA, Damste JSS, Jetten MSM,
Strous M (2007b) Candidatus ‘‘Anammoxoglobus propionicus’’
a new propionate oxidizing species of anaerobic ammonium
oxidizing bacteria. Syst Appl Microbiol 30(1):39–49
Kartal B, van Niftrik L, Rattray J, de Vossenberg J, Schmid MC,
Damste JSS, Jetten MSM, Strous M (2008) Candidatus ‘Broca-
dia fulgida’: an autofluorescent anaerobic ammonium oxidizing
bacterium. FEMS Microbiol Ecol 63(1):46–55
Kartal B, Kuenen JG, van Loosdrecht MCM (2010) Sewage treatment
with anammox. Science 328(5979):702–703
Kawagoshi Y, Nakamura Y, Kawashima H, Fujisaki K, Fujimoto A,
Furukawa K (2009) Enrichment culture of marine anaerobic
ammonium oxidation (anammox) bacteria from sediment of sea-
based waste disposal site. J Biosci Bioeng 107(1):61–63
Khin T, Annachhatre AP (2004) Novel microbial nitrogen removal
processes. Biotechnol Adv 22(7):519–532
Kim DJ, Lee DI, Keller J (2006) Effect of temperature and free
ammonia on nitrification and nitrite accumulation in landfill
leachate and analysis of its nitrifying bacterial community by
FISH. Bioresour Technol 97(3):459–468
Kimura Y, Isaka K, Kazama F, Sumino T (2010) Effects of nitrite
inhibition on anaerobic ammonium oxidation. Appl Microbiol
Biotechnol 86(1):359–365
Kindaichi T, Tsushima I, Ogasawara Y, Shimokawa M, Ozaki N,
Satoh H, Okabe S (2007) In situ activity and spatial organization
of anaerobic ammonium-oxidizing (anammox) bacteria in bio-
films. Appl Environ Microbiol 73(15):4931–4939
Kirschner C, Maquelin K, Pina P, Thi NAN, Choo-Smith LP,
Sockalingum GD, Sandt C, Ami D, Orsini F, Doglia SM,
Allouch P, Mainfait M, Puppels GJ, Naumann D (2001)
Classification and identification of enterococci: a comparative
phenotypic, genotypic, and vibrational spectroscopic study.
J Clin Microbiol 39(5):1763–1770
Klappenbach JA, Saxman PR, Cole JR, Schmidt TM (2001) rrndb: the
ribosomal RNA operon copy number database. Nucleic Acids
Res 29(1):181–184
Klotz MG, Stein LY (2008) Nitrifier genomics and evolution of the
nitrogen cycle. FEMS Microbiol Lett 278(2):146–156. doi:
10.1111/j.1574-6968.2007.00970.x
Klotz MG, Schmid MC, Strous M, den Camp H, Jetten MSM, Hooper
AB (2008) Evolution of an octahaem cytochrome c protein
family that is key to aerobic and anaerobic ammonia oxidation
by bacteria. Environ Microbiol 10(11):3150–3163
Knight RL, Kadlec RH, Ohlendorf HM (1999) The use of treatment
wetlands for petroleum industry effluents. Environ Sci Technol
33(7):973–980
Koch G, Egli K, Van der Meer JR, Siegrist H (2000) Mathematical
modeling of autotrophic denitrification in a nitrifying biofilm of
a rotating biological contactor. Water Sci Technol 41(4–5):
191–198
Konneke M, Bernhard AE, de la Torre JR, Walker CB, Waterbury JB,
Stahl DA (2005) Isolation of an autotrophic ammonia-oxidizing
marine archaeon. Nature 437(7058):543–546. doi:10.1038/
nature03911
Kuai LP, Verstraete W (1998) Ammonium removal by the oxygen-
limited autotrophic nitrification–denitrification system. Appl
Environ Microbiol 64(11):4500–4506
Kubota K, Ohashi A, Imachi H, Harada H (2006) Improved in situ
hybridization efficiency with locked-nucleic-acid-incorporated
DNA probes. Appl Environ Microbiol 72(8):5311–5317
Kuenen JG (2008) Anammox bacteria: from discovery to application.
Nat Rev Microbiol 6(4):320–326
Kuenen JG, Jetten MSM (2001) Extraordinary anaerobic ammonium-
oxidizing bacteria. ASM News 67(9):456
Kuypers MMM, Sliekers AO, Lavik G, Schmid M, Jorgensen BB,
Kuenen JG, Damste JSS, Strous M, Jetten MSM (2003)
Anaerobic ammonium oxidation by anammox bacteria in the
Black Sea. Nature 422(6932):608–611
A. Terada et al.
123

Kuypers MMM, Lavik G, Woebken D, Schmid M, Fuchs BM, Amann
R, Jorgensen BB, Jetten MSM (2005) Massive nitrogen loss
from the Benguela upwelling system through anaerobic ammo-
nium oxidation. Proc Natl Acad Sci USA 102(18):6478–6483
Lackner S, Terada A, Smets BF (2008) Heterotrophic activity
compromises autotrophic nitrogen removal in membrane-aerated
biofilms: results of a modeling study. Water Res 42(4–5):
1102–1112
Lam P, Lavik G, Jensen MM, van de Vossenberg J, Schmid M,
Woebken D, Dimitri G, Amann R, Jetten MSM, Kuypers MMM
(2009) Revising the nitrogen cycle in the Peruvian oxygen
minimum zone. Proc Natl Acad Sci USA 106(12):4752–4757
Lee CG, Fletcher TD, Sun GZ (2009a) Nitrogen removal in
constructed wetland systems. Eng Life Sci 9(1):11–22
Lee ZMP, Bussema C, Schmidt TM (2009b) rrnDB: documenting the
number of rRNA and tRNA genes in bacteria and archaea.
Nucleic Acids Res 37:D489–D493
Li XR, Du B, Fu HX, Wang RF, Shi JH, Wang Y, Jetten MSM, Quan
ZX (2009) The bacterial diversity in an anaerobic ammonium-
oxidizing (anammox) reactor community. Syst Appl Microbiol
32(4):278–289
Li H, Chen S, Mu BZ, Gu JD (2010a) Molecular detection of
anaerobic ammonium-oxidizing (anammox) bacteria in high-
temperature petroleum reservoirs. Microb Ecol 60(4):771–783
Li M, Hong YG, Klotz MG, Gu JD (2010b) A comparison of primer
sets for detecting 16S rRNA and hydrazine oxidoreductase genes
of anaerobic ammonium-oxidizing bacteria in marine sediments.
Appl Microbiol Biotechnol 86(2):781–790
Lopez H, Puig S, Ganigue R, Ruscalleda M, Balaguer MD, Colprim J
(2008) Start-up and enrichment of a granular anammox SBR to
treat high nitrogen load wastewaters. J Chem Technol Biotech-
nol 83(3):233–241
Loy A, Horn M, Wagner M (2003) probeBase: an online resource for
rRNA-targeted oligonucleotide probes. Nucleic Acids Res
31(1):514–516
Ludwig W, Strunk O, Westram R, Richter L, Meier H, Yadhukumar,
Buchner A, Lai T, Steppi S, Jobb G, Forster W, Brettske I,
Gerber S, Ginhart AW, Gross O, Grumann S, Hermann S, Jost R,
Konig A, Liss T, Lussmann R, May M, Nonhoff B, Reichel B,
Strehlow R, Stamatakis A, Stuckmann N, Vilbig A, Lenke M,
Ludwig T, Bode A, Schleifer KH (2004) ARB: a software
environment for sequence data. Nucleic Acids Res 32(4):
1363–1371
Meyer RL, Risgaard-Petersen N, Allen DE (2005) Correlation
between anammox activity and microscale distribution of nitrite
in a subtropical mangrove sediment. Appl Environ Microbiol
71(10):6142–6149
Mohamed NM, Saito K, Tal Y, Hill RT (2010) Diversity of aerobic
and anaerobic ammonia-oxidizing bacteria in marine sponges.
ISME J 4(1):38–48
Mulder A, Vandegraaf AA, Robertson LA, Kuenen JG (1995)
Anaerboc ammonium oxidation discovered in a denitrifying
fluidized-bed reactor. FEMS Microbiol Ecol 16(3):177–183
Mulder JW, van Loosdrecht MCM, Hellinga C, van Kempen R (2001)
Full-scale application of the SHARON process for treatment of
rejection water of digested sludge dewatering. Water Sci
Technol 43(11):127–134
Muyzer G (1999) DGGE/TGGE a method for identifying genes from
natural ecosystems. Curr Opin Microbiol 2(3):317–322
Neef A, Amann R, Schlesner H, Schleifer KH (1998) Monitoring
a widespread bacterial group: in situ detection of planctomyce-
tes with 16S rRNA-targeted probes. Microbiology 144:3257–
3266
Nielsen M, Bollmann A, Sliekers O, Jetten M, Schmid M, Strous M,
Schmidt I, Larsen LH, Nielsen LP, Revsbech NP (2005)
Kinetics, diffusional limitation and microscale distribution of
chemistry and organisms in a CANON reactor. FEMS Microbiol
Ecol 51(2):247–256
Okabe S, Satoh H, Watanabe Y (1999) In situ analysis of nitrifying
biofilms as determined by in situ hybridization and the use of
microelectrodes. Appl Environ Microbiol 65(7):3182–3191
Park H, Rosenthal A, Ramalingam K, Fillos J, Chandran K (2010)
Linking community profiles, gene expression and N-removal in
anammox bioreactors treating municipal anaerobic digestion
reject water. Environ Sci Technol 44(16):6110–6116
Pathak BK, Kazama F, Tanaka Y, Mori K, Sumino T (2007)
Quantification of anammox populations enriched in an immobi-
lized microbial consortium with low levels of ammonium
nitrogen and at low temperature. Appl Microbiol Biotechnol
76(5):1173–1179
Patzold R, Keuntje M, Anders-von Ahlften A (2006) A new approach
to non-destructive analysis of biofilms by confocal Raman
microscopy. Anal Bioanal Chem 386(2):286–292
Patzold R, Keuntje M, Theophile K, Muller J, Mielcarek E,
Ngezahayo A, Ahlften AAV (2008) In situ mapping of nitrifiers
and anammox bacteria in microbial aggregates by means of
confocal resonance Raman microscopy. J Microbiol Methods
72(3):241–248
Pavlekovic M, Schmid MC, Schmider-Poignee N, Spring S, Pilhofer
M, Gaul T, Fiandaca M, Loffler FE, Jetten M, Schleifer KH, Lee
NM (2009) Optimization of three FISH procedures for in situ
detection of anaerobic ammonium oxidizing bacteria in biolog-
ical wastewater treatment. J Microbiol Methods 78(2):119–126
Pellicer-Nacher C, Sun SP, Lackner S, Terada A, Schreiber F, Zhou
Q, Smets BF (2010) Sequential aeration of membrane-aerated
biofilm reactors for high-rate autotrophic nitrogen removal:
experimental demonstration. Environ Sci Technol 44(19):
7628–7634
Penton CR, Devol AH, Tiedje JM (2006) Molecular evidence for the
broad distribution of anaerobic ammonium-oxidizing bacteria in
freshwater and marine sediments. Appl Environ Microbiol
72(10):6829–6832
Pernthaler A, Pernthaler J, Amann R (2002) Fluorescence in situ
hybridization and catalyzed reporter deposition for the identifi-
cation of marine bacteria. Appl Environ Microbiol 68(6):
3094–3101
Perry-O’Keefe H, Rigby S, Oliveira K, Sorensen D, Slender H, Coull
J, Hyldig-Nielsen JJ (2001) Identification of indicator microor-
ganisms using a standardized PNA FISH method. J Microbiol
Methods 47(3):281–292
Pynaert K, Smets BF, Wyffels S, Beheydt D, Siciliano SD, Verstraete
W (2003) Characterization of an autotrophic nitrogen-removing
biofilm from a highly loaded lab-scale rotating biological
contactor. Appl Environ Microbiol 69(6):3626–3635
Pynaert K, Smets BF, Beheydt D, Verstraete W (2004) Start-up of
autotrophic nitrogen removal reactors via sequential biocatalyst
addition. Environ Sci Technol 38(4):1228–1235
Quan ZX, Rhee SK, Zuo JE, Yang Y, Bae JW, Park JR, Lee ST, Park
YH (2008) Diversity of ammonium-oxidizing bacteria in a
granular sludge anaerobic ammonium-oxidizing (anammox)
reactor. Environ Microbiol 10(11):3130–3139
Rattray JE, van de Vossenberg J, Jaeschke A, Hopmans EC,
Wakeham SG, Lavik G, Kuypers MMM, Strous M, Jetten
MSM, Schouten S, Damste JSS (2010) Impact of temperature on
ladderane lipid distribution in anammox bacteria. Appl Environ
Microbiol 76(5):1596–1603
Risgaard-Petersen N, Meyer RL, Schmid M, Jetten MSM, Enrich-
Prast A, Rysgaard S, Revsbech NP (2004) Anaerobic ammonium
oxidation in an estuarine sediment. Aquat Microb Ecol
36(3):293–304
Rittmann BE, McCarty PL (2001) Environmental biotechnology:
principles and application. McGraw-Hill Book Co, New York
Review of anammox detection, presence, and application
123

Rysgaard S, Glud RN, Risgaard-Petersen N, Dalsgaard T (2004)
Denitrification and anammox activity in arctic marine sediments.
Limnol Oceanogr 49(5):1493–1502
Sangolkar LN, Maske SS, Chakrabarti T (2006) Methods for
determining microcystins (peptide hepatotoxins) and microcy-
stin-producing cyanobacteria. Water Res 40:3485–3496
Schmid M, Twachtmann U, Klein M, Strous M, Juretschko S, Jetten
M, Metzger JW, Schleifer KH, Wagner M (2000) Molecular
evidence for genus level diversity of bacteria capable of
catalyzing anaerobic ammonium oxidation. Syst Appl Microbiol
23(1):93–106
Schmid M, Schmitz-Esser S, Jetten M, Wagner M (2001) 16S–23S
rDNA intergenic spacer and 23S rDNA of anaerobic ammonium-
oxidizing bacteria: implications for phylogeny and in situ
detection. Environ Microbiol 3(7):450–459
Schmid M, Walsh K, Webb R, Rijpstra WIC, van de Pas-Schoonen K,
Verbruggen MJ, Hill T, Moffett B, Fuerst J, Schouten S, Damste
JSS, Harris J, Shaw P, Jetten M, Strous M (2003) Candidatus
‘‘Scalindua brodae’’, sp nov., Candidatus ‘‘Scalindua wagneri’’,
sp nov., two new species of anaerobic ammonium oxidizing
bacteria. Syst Appl Microbiol 26(4):529–538
Schmid MC, Maas B, Dapena A, de Pas-Schoonen KV, de
Vossenberg JV, Kartal B, van Niftrik L, Schmidt I, Cirpus I,
Kuenen JG, Wagner M, Damste JSS, Kuypers M, Revsbech NP,
Mendez R, Jetten MSM, Strous M (2005) Biomarkers for in situ
detection of anaerobic ammonium-oxidizing (anammox) bacte-
ria. Appl Environ Microbiol 71(4):1677–1684
Schmid MC, Risgaard-Petersen N, van de Vossenberg J, Kuypers
MMM, Lavik G, Petersen J, Hulth S, Thamdrup B, Canfield D,
Dalsgaard T, Rysgaard S, Sejr MK, Strous M, den Camp H,
Jetten MSM (2007) Anaerobic ammonium-oxidizing bacteria in
marine environments: widespread occurrence but low diversity.
Environ Microbiol 9(6):1476–1484
Schmid MC, Hooper AB, Klotz MG, Woebken D, Lam P, Kuypers
MMM, Pommerening-Roeser A, op den Camp HJM, Jetten
MSM (2008) Environmental detection of octahaem cytochrome
c hydroxylamine/hydrazine oxidoreductase genes of aerobic and
anaerobic ammonium-oxidizing bacteria. Environ Microbiol
10(11):3140–3149
Schmidt I, Zart D, Bock E (2001) Gaseous NO2 as a regulator for
ammonia oxidation of Nitrosomonas eutropha. Anton Leeuw Int
J G 79(3–4):311–318
Schmidt I, Sliekers O, Schmid M, Bock E, Fuerst J, Kuenen JG, Jetten
MSM, Strous M (2003) New concepts of microbial treatment
processes for the nitrogen removal in wastewater. FEMS
Microbiol Rev 27(4):481–492
Schramm A, Larsen LH, Revsbech NP, Ramsing NB, Amann R,
Schleifer KH (1996) Structure and function of a nitrifying
biofilm as determined by in situ hybridization and the use of
microelectrodes. Appl Environ Microbiol 62(12):4641–4647
Schubert CJ, Durisch-Kaiser E, Wehrli B, Thamdrup B, Lam P,
Kuypers MMM (2006) Anaerobic ammonium oxidation in a
tropical freshwater system (Lake Tanganyika). Environ Micro-
biol 8(10):1857–1863
Schutte UME, Abdo Z, Bent SJ, Shyu C, Williams CJ, Pierson JD,
Forney LJ (2008) Advances in the use of terminal restriction
fragment length polymorphism (T-RFLP) analysis of 16S rRNA
genes to characterize microbial communities. Appl Microbiol
Biotechnol 80(3):365–380
Sharkey FH, Banat IM, Marchant R (2004) Detection and quantifi-
cation of gene expression in environmental bacteriology. Appl
Environ Microbiol 70(7):3795–3806
Shimamura M, Nishiyama T, Shigetomo H, Toyomoto T, Kawahara
Y, Furukawa K, Fujii T (2007) Isolation of a multiheme protein
with features of a hydrazine-oxidizing enzyme from an
anaerobic ammonium-oxidizing enrichment culture. Appl Envi-
ron Microbiol 73(4):1065–1072
Siegrist H, Reithaar S, Koch G, Lais P (1998) Nitrogen loss in a
nitrifying rotating contactor treating ammonium-rich wastewater
without organic carbon. Water Sci Technol 38(8–9):241–248
Sliekers AO, Derwort N, Gomez JLC, Strous M, Kuenen JG, Jetten
MSM (2002) Completely autotrophic nitrogen removal over
nitrite in one single reactor. Water Res 36(10):2475–2482
Sliekers AO, Third KA, Abma W, Kuenen JG, Jetten MSM (2003)
CANON and anammox in a gas-lift reactor. FEMS Microbiol
Lett 218(2):339–344
Sliekers AO, Haaijer S, Schmid M, Harhangi H, Verwegen K, Kuenen
JG, Jetten MSM (2004) Nitrification and anammox with urea as
the energy source. Syst Appl Microbiol 27(3):271–278
Smith CJ, Osborn AM (2009) Advantages and limitations of
quantitative PCR (Q-PCR)-based approaches in microbial ecol-
ogy. FEMS Microbiol Ecol 67(1):6–20
Strous M, VanGerven E, Zheng P, Kuenen JG, Jetten MSM (1997)
Ammonium removal from concentrated waste streams with the
anaerobic ammonium oxidation (anammox) process in different
reactor configurations. Water Res 31(8):1955–1962
Strous M, Heijnen JJ, Kuenen JG, Jetten MSM (1998) The sequencing
batch reactor as a powerful tool for the study of slowly growing
anaerobic ammonium-oxidizing microorganisms. Appl Micro-
biol Biotechnol 50(5):589–596
Strous M, Fuerst JA, Kramer EHM, Logemann S, Muyzer G, van de
Pas-Schoonen KT, Webb R, Kuenen JG, Jetten MSM (1999a)
Missing lithotroph identified as new planctomycete. Nature
400(6743):446–449
Strous M, Kuenen JG, Jetten MSM (1999b) Key physiology of
anaerobic ammonium oxidation. Appl Environ Microbiol 65(7):
3248–3250
Strous M, Pelletier E, Mangenot S, Rattei T, Lehner A, Taylor MW,
Horn M, Daims H, Bartol-Mavel D, Wincker P, Barbe V,
Fonknechten N, Vallenet D, Segurens B, Schenowitz-Truong C,
Medigue C, Collingro A, Snel B, Dutilh BE, Op den Camp HJM,
van der Drift C, Cirpus I, van de Pas-Schoonen KT, Harhangi HR,
van Niftrik L, Schmid M, Keltjens J, van de Vossenberg J, Kartal
B, Meier H, Frishman D, Huynen MA, Mewes HW, Weissenbach
J, Jetten MSM, Wagner M, Le Paslier D (2006) Deciphering the
evolution and metabolism of an anammox bacterium from a
community genome. Nature 440(7085):790–794
Sun SP, Nacher CPI, Merkey B, Zhou Q, Xia SQ, Yang DH, Sun JH,
Smets BF (2010) Effective biological nitrogen removal treat-
ment processes for domestic wastewaters with low C/N ratios: a
review. Environ Eng Sci 27(2):111–126
Syron E, Casey E (2008) Membrane-aerated biofilms for high rate
biotreatment: performance appraisal, engineering principles,
scale-up, and development requirements. Environ Sci Technol
42(6):1833–1844
Tang CJ, Zheng P, Hu BL, Chen JW, Wang CH (2010) Influence of
substrates on nitrogen removal performance and microbiology of
anaerobic ammonium oxidation by operating two UASB reactors
fed with different substrate levels. J Hazard Mater 181(1–3):19–26
Terada A, Lackner S, Tsuneda S, Smets BF (2007) Redox-stratifi-
cation controlled biofilm (ReSCoBi) for completely autotrophic
nitrogen removal: the effect of co- versus counter-diffusion on
reactor performance. Biotechnol Bioeng 97(1):40–51
Terada A, Lackner S, Kristensen K, Smets BF (2010) Inoculum
effects on community composition and nitritation performance
of autotrophic nitrifying biofilm reactors with counter-diffusion
geometry. Environ Microbiol 12(10):2858–2872
Thamdrup B, Dalsgaard T (2002) Production of N-2 through
anaerobic ammonium oxidation coupled to nitrate reduction in
marine sediments. Appl Environ Microbiol 68(3):1312–1318
A. Terada et al.
123

Third KA, Paxman J, Schmid M, Strous M, Jetten MSM, Cord-
Ruwisch R (2005) Enrichment of anammox from activated
sludge and its application in the CANON process. Microb Ecol
49(2):236–244
Trigo C, Campos JL, Garrido JM, Mendez R (2006) Start-up of the
anammox process in a membrane bioreactor. J Biotechnol
126(4):475–487
Trimmer M, Nicholls JC, Deflandre B (2003) Anaerobic ammonium
oxidation measured in sediments along the Thames estuary,
United Kingdom. Appl Environ Microbiol 69(11):6447–6454
Tsuneda S, Ejiri Y, Nagano T, Hirata A (2004) Formation mechanism
of nitrifying granules observed in an aerobic upflow fluidized
bed (AUFB) reactor. Water Sci Technol 49(11–12):27–34
Tsushima I, Kindaichi T, Okabe S (2007a) Quantification of
anaerobic ammonium-oxidizing bacteria in enrichment cultures
by real-time PCR. Water Res 41(4):785–794
Tsushima I, Ogasawara Y, Kindaichi T, Satoh H, Okabe S (2007b)
Development of high-rate anaerobic ammonium-oxidizing
(anammox) biofilm reactors. Water Res 41(8):1623–1634
Urakawa H, Martens-Habbena W, Stahl DA (2010) High abundance
of ammonia-oxidizing archaea in coastal waters, determined
using a modified DNA extraction method. Appl Environ
Microbiol 76(7):2129–2135
van de Graaf AA, Mulder A, Debruijn P, Jetten MSM, Robertson LA,
Kuenen JG (1995) Anaerobic oxidation of ammonium is a
biologically mediated process. Appl Environ Microbiol 61(4):
1246–1251
van de Graaf AA, deBruijn P, Robertson LA, Jetten MSM, Kuenen JG
(1996) Autotrophic growth of anaerobic ammonium-oxidizing
micro-organisms in a fluidized bed reactor. Microbiology
142:2187–2196
van de Vossenberg J, Rattray JE, Geerts W, Kartal B, van Niftrik L,
van Donselaar EG, Damste JSS, Strous M, Jetten MSM (2008)
Enrichment and characterization of marine anammox bacteria
associated with global nitrogen gas production. Environ Micro-
biol 10(11):3120–3129
van der Star WRL, Abma WR, Blommers D, Mulder JW, Tokutomi
T, Strous M, Picioreanu C, Van Loosdrecht MCM (2007) Startup
of reactors for anoxic ammonium oxidation: experiences from
the first full-scale anammox reactor in Rotterdam. Water Res
41(18):4149–4163
van der Star WRL, Miclea AI, van Dongen U, Muyzer G, Picioreanu
C, van Loosdrecht MCM (2008) The membrane bioreactor: a
novel tool to grow anammox bacteria as free cells. Biotechnol
Bioeng 101(2):286–294
Van Hulle SWH, Vandeweyer HJP, Meesschaert BD, Vanrolleghem
PA, Dejans P, Dumoulin A (2010) Engineering aspects and
practical application of autotrophic nitrogen removal from
nitrogen rich streams. Chem Eng J 162(1):1–20
van Loosdrecht MCM (2008) Innovative nitrogen removal. In: Henze
M, van Loosdrecht MCM, Ekama GA, Brdjanovic D (eds)
Biological wastewater treatment-principles, modelling and
design. IWA Publishing, London, pp 139–154
Vazquez-Padin J, Mosquera-Corral A, Campos JL, Mendez R,
Revsbech NP (2010) Microbial community distribution and
activity dynamics of granular biomass in a CANON reactor.
Water Res 44(15):4359–4370
Vlaeminck SE, Cloetens LFF, Carballa M, Boon N, Verstraete W
(2008) Granular biomass capable of partial nitritation and
anammox. Water Sci Technol 58(5):1113–1120
Vlaeminck SE, Terada A, Smets BF, Van der Linden D, Boon N,
Verstraete W, Carballa M (2009) Nitrogen removal from
digested black water by one-stage partial nitritation and anam-
mox. Environ Sci Technol 43(13):5035–5041
Voss M, Montoya JP (2009) Nitrogen cycle oceans apart. Nature
461(7260):49–50. doi:10.1038/461049a
Wagner M, Nielsen PH, Loy A, Nielsen JL, Daims H (2006) Linking
microbial community structure with function: fluorescence in
situ hybridization-microautoradiography and isotope arrays. Curr
Opin Biotechnol 17(1):83–91
Ward BB, Devol AH, Rich JJ, Chang BX, Bulow SE, Naik H, Pratihary
A, Jayakumar A (2009) Denitrification as the dominant nitrogen
loss process in the Arabian Sea. Nature 461(7260):78–81
Wett B (2007) Development and implementation of a robust
deammonification process. Water Sci Technol 56(7):81–88
Wett B, Hell M, Nyhuis G, Puempel T, Takacs I, Murthy S (2010)
Syntrophy of aerobic and anaerobic ammonia oxidisers. Water
Sci Technol 61(8):1915–1922
Wiesmann U (1994) Biological nitrogen removal from wastewater.
Berlin: Springer-Verlag 51 (In Advances in biochemical
engineering/biotechnology) pp 113–154
Woebken D, Fuchs BA, Kuypers MAA, Amann R (2007) Potential
interactions of particle-associated anammox bacteria with bac-
terial and archaeal partners in the Namibian upwelling system.
Appl Environ Microbiol 73(14):4648–4657
Woebken D, Lam P, Kuypers MMM, Naqvi SWA, Kartal B, Strous
M, Jetten MSM, Fuchs BM, Amann R (2008) A microdiversity
study of anammox bacteria reveals a novel Candidatus Scalindua
phylotype in marine oxygen minimum zones. Environ Microbiol
10(11):3106–3119
Wyffels S, Boeckx P, Pynaert K, Zhang D, Van Cleemput O, Chen G,
Verstraete W (2004) Nitrogen removal from sludge reject water
by a two-stage oxygen-limited autotrophic nitrification denitri-
fication process. Water Sci Technol 49(5–6):57–64
Zhang Y, Ruan XH, den Camp H, Smits TJM, Jetten MSM, Schmid
MC (2007) Diversity and abundance of aerobic and anaerobic
ammonium-oxidizing bacteria in freshwater sediments of the
Xinyi River (China). Environ Microbiol 9(9):2375–2382
Zhou S, Nakashimada Y, Hosomi M (2009) Nitrogen transformations
in vertical flow systems with and without rice (Oryza sativa)
studied with a high-resolution soil–water profiler. Ecol Eng
35(2):213–220
Zhu GB, Jetten MSM, Kuschk P, Ettwig KF, Yin CQ (2010) Potential
roles of anaerobic ammonium and methane oxidation in the
nitrogen cycle of wetland ecosystems. Appl Microbiol Biotech-
nol 86(4):1043–1055
Review of anammox detection, presence, and application
123





























