Embed Size (px)
Citation preview

PRIMING IN THE BRAIN, AN IMMUNOLOGICALLY PRIVILEGED ORGAN,ELICITS ANTI-TUMOR IMMUNITYHassan M. FATHALLAH -SHAYKH *, Wei GAO, Michael CHO and Maria Alejandra HERRERA
Department of Neurology, University of Texas Southwestern Medical Center at Dallas, Dallas, TX, USA
A crucial question in the study of tumor neuro-immunologyconcerns the capacity of the central nervous system toinitiate and execute an immune response. In a 100% fatal ratmalignant glioma model, genetically modified tumors secret-ing INF-g intracerebrally generate an immune responseresulting in a substantial increase in survival time, tumorrejection and specific systemic immunity. Tumors modifiedto secrete IL-2 alone do not change the biologic behavior oftransfected gliomas. INF-g induces elevated expression ofmajor-histocompatibility-complex-class-I and -class-II mol-ecules in microglia throughout the brain and invokes en-hanced tumor infiltration by CD4, CD8 and NK cells. Thesefindings demonstrate successful immunization against a cen-tral-nervous-system tumor by direct priming in the brainwith a live growth-competent tumor vaccine. Int. J. Cancer75:266–276, 1998.r 1998 Wiley-Liss, Inc.
Malignant brain tumors progress rapidly and are almost invari-ably fatal. Over the past decade, despite aggressive combined-modality treatments, little improvement has been made in theprognosis and survival of patients suffering from malignant glialneoplasms. The location of tumors in the brain, an organ believedto be ‘‘immunologically privileged,’’ is thought to help them evadeimmune surveillance.
The classical idea that the mammalian brain is an immunologi-cally privileged site was initially based on observations that: (i)allografts of carcinomas are more successful in the brain than in thesub-cutaneous space (Barker and Billingham, 1971); (ii) a widevariety of tissues and organs from embryonic donors growsuccessfully in the brain (Barker and Billingham, 1971); (iii) thebrain lacks defined lymphatic drainage (Yoffey and Courtice,1970); (iv) the expression of major-histocompatibility-com-plex(MHC)-class I and -II molecules in the brain is low (Daaret al.,1984); and (v) only activated T lymphocytes cross the blood-brainbarrier (Hickeyet al., 1991). These facts are consistent with theconcept that the privileged status of the brain is related to aninterruption in the afferent pathway of the immune response(Barker and Billingham, 1971; Yoffey and Courtice, 1970).
In the study of systemic tumor immunology, a 3-stage process isrequired for the generation of effector T cells after peripheralvaccination: (i) tumor antigen uptake and processing at the site ofinjection by professional antigen-presenting cells (APC); (ii)migration of APC into regional draining lymph nodes, where T-cellpriming occurs; (iii) circulation of activated T cells that eitherperform or initiate effector mechanisms leading to tumor-celldestruction (Maasset al.,1995). Although, when transplanted to anaive brain, skin homografts survive, when transplanted to specifi-cally immunized animals they break down. This disparity ofgrowth suggests that the immune privileges of the brain are relatedto the absence of typical lymphatic drainage (Medawar, 1948).However, the privileges of the brain are far from complete, since,after irradiation of established experimental gliomas, treated ani-mals reject 37% of second intracerebral implants and 43% of s.c.implants (Scheinberget al.,1965).
Typical lymphatic vessels do not exist in the brain, and the majoroutflow pathway for extracellular fluid is passage through thearachnoid villi into blood of the dural sinus (Welleret al., 1992;Yamadaet al.,1991). Nevertheless, following injection of radiola-belled protein into the CNS, a significant fraction (14–47%) of thistracer leaving the CNS appears in the cervical lymph nodes (Weller
et al.,1992; Yamadaet al.,1991). Moreover, the injection of sheeperythrocytes into the lateral ventricles of rabbits (Jankovicet al.,1961) and the micro-infusion of human BSA into rat brains lead tothe production of specific antibodies in serum. The latter responsewas reduced about 10-fold by cervical lymph obstruction (Harling-Berget al.,1989).
Considerable gaps in our understanding of tumor neuro-immunology relate to the identity of the brain-resident cell able topresent antigens to T cells, and the site of antigen presentation.Recently, microglia have emerged as potential candidates for therole of APC of the CNS (Cash and Rot, 1994). When restingmicroglia are activated by treatment with interferon gamma(INF-g), they become fully competent to present antigens tolymphocytes in an appropriately-restricted manner and acquire thecapacity to activate an immune responsein vitro (Hickey andKimura, 1988). However, the site where microglia could presentantigens to T cellsin vivo has not been identified. It is not knownwhether antigen presentation takes place at a peripheral lymphaticorgan, in an intravascular compartment or within the CNS (Hartand Fabry, 1995).
In the study of peripheral immunology, both interleukin 2 (IL-2)and INF-g favor the development of Th1 cells. However, theintracerebral co-transplantation into syngeneic animals of fibro-blasts or of endothelial cells transduced with viral vectors to secreteIL-2 and/or INF-g with murine glioma cells does not elicit tumorrejection (Lichtoret al.,1995). Whether ‘‘high’’ amounts of INF-gand IL-2, secreted in the brain by the tumor itself, could inducetumor rejection and protective immunity remains unknown. Wehypothesized that, given appropriate stimulus, the brain is capableof mounting an anti-tumor immune response.
In this study, experiments were designed to investigate thebiologic effects of priming directly into the brain by implantinggenetically modified malignant glial cells secreting ‘‘high’’ quanti-ties of INF-g and IL-2. Responses were measured by: (i) survival,(ii) rejection and (iii) protective immunity against challenge by theparental line. This investigation led to successful immunizationagainst a CNS tumor by direct priming in the brain with a livegrowth-competent tumor vaccine.
MATERIALS AND METHODS
Plasmid construction
Human IL-2 cDNA, a generous gift from Dr. B. Cullen, (DukeUniversity, Durham, NC), was removed from the plasmid BC12/CMV/IL-2 (Cullen et al., 1988). Murine INF-g cDNA wasobtained from the plasmid pms 10 (ATCC, Rockville, MD).Sub-cloning was carried out by standard recombinant DNA tech-niques. The backbone of pGINF/IL2 was derived from pREP9(InVitrogen, San Diego, CA) after digestion with SalI. The
Contract grant sponsor: American Cancer Society; Contract grantnumber: 1RG-142L.
*Correspondence to: Department of Neurology, University of TexasSouthwestern Medical Center, Dallas, TX 75235-9036, USA.e-mail: [email protected]
Received 9 July 1997; Revised 9 September 1997
Int. J. Cancer:75,266–276 (1998)
r 1998 Wiley-Liss, Inc.
Publication of the International Union Against CancerPublication de l’Union Internationale Contre le Cancer

expression cassette was derived from the plasmid pCMVb (Clon-tech, Palo Alto, CA), where it extends from a 58 EcoRI site to a 38SalI site. pCLON was constructed by inserting a unique SpeI siteinto the EcoRI site of pCMVb. INF-g cDNA and IL-2 cDNA were
cloned separately into the NotI site of pCLON (pCLON-INF-g andpCLON-IL-2 respectively); the orientation was confirmed by DNAsequencing using the dideoxy termination method. pGINF/IL2 wasconstructed by sub-cloning both the SpeI-SalI fragments of PCLON-INF-g and pCLON-IL-2 in series into pREP9 digested with SalI.pGBGAL was constructed by sub-cloning the SpeI-SalI fragmentof pCLON into pREP9 digested with SalI. The plasmids pGIL2 andpGINF were constructed by cloning the SpeI-SalI fragments ofeither pCLON-IL-2 or pCLON-INF-g into pREP9 digested withSalI.
TransfectionRT2, an avian-sarcoma-virus-induced malignant glioma in an
F344 rat, was a generous gift from Dr. Y. Gillepsie (University ofAlabama, Birmingham, AL). Cells were grown in a 50:50 mixtureof Dulbecco’s and Ham’s F-10 media (GIBCO, Grand Island, NY)supplemented with 10% FCS at 37°C in a humidified incubator in5% CO2. RT2 cells were transfected with plasmids complexed tolipofectamine (GIBCO) following the manufacturer’s specifica-tions; then, 72 hr after transfection, cultures were grown in
FIGURE 1 – Plasmid maps of pGINF/IL2(a), pGIL2 (b), pGBGAL (c), pGINF (d). Abbreviations: IL2, human IL-2 cDNA; INF-g, murineINF-g cDNA; sigp, signal peptide; TK, thymidine kinase; Poly A, polyadenylation signal; SV40, simian virus 40; CMV, cytomegalovirus;enh/prom, enhancer/promoter; Neomycin, neomycin resistance; Ampicillin/ori, ampicillin resistance/ pUC origin of replication; EBNA-1,Epstein-Barr nuclear antigen 1; OriP, Epstein-Barr virus origin of replication.
TABLE I – AMOUNTS OF CYTOKINES SECRETED BY TRANSFECTED CLONALCULTURESIN VITRO
Culture Plasmid IL-2pg/M/D1
INF-gpg/M/D
RT2-pGBGAL pGBGAL 0 0RT2-hi-IL2 1 INF PGINF/IL2 192 171
2688 733382 437
RT2-lo-IL2 1 INF pGINF/IL2 62 014 2426 17
RT2-pGIL2 pGIL2 2065 0RT2-pGINF pGINF 0 473
1pg/M/D, pg per million cells per day.
267BIOLOGIC EFFECTS OF GLIOMA-SECRETED INF-g AND IL-2
selection media supplemented with Geneticin (GIBCO) at 4.0 g/l.Clonal transfected cultures were selected by cloning rings, sub-cloned at limiting dilution (0.5 cells/well, 96-well plates) andpropagated in selection media containing Geneticin.
ELISA measurement of secreted cytokinesSupernatants of transfected cultured cells were assayed for
human IL-2 (R&D, Minneapolis, MN) and murine INF-g (Gen-zyme, Cambridge, MA) by enzyme-linked immunosorbent assay(ELISA) following the manufacturer’s specifications.
Animal injectionsTransfected cells were implanted into the brain at the same
passage number as when their supernatants were assayed byELISA. Female Fisher 344 rats (Simonson, Gilrow, CA) weighing180 to 200 g were anesthetized i.p. by xylazine (10 mg/kg) andketamine (90 mg/kg), then placed in a stereotactic frame. Thebregma was identified, a burr hole was drilled and the dura
exposed. The dura was punctured by a Hamilton syringe and aneedle was inserted 3.5 mm (1 mm anterior and 2 mm to the right ofthe bregma) into the brain over 3 min. After 3 min, 2000 or 500tumor cells, suspended in 5 µl PBS, were injected over 3 min. Afteranother 3 min the needle was withdrawn slowly over 3 min.Animals challenged by the parental cells were reimplanted with2000 cells of wild-type(wt) RT2 in the contralateral hemisphere atleast 60 days after the first injection. All re-injected animals hadnegative magnetic resonance imaging of the brain.
Magnetic resonance imagingMagnetic resonance imaging (MRI) was performed at the
Rogers MRI Center using a General Electric Omega 4.7 T systemwith a 40-cm bore magnet. Eight images of the brain were obtained(TR 500 ms, TE 12 ms, with 2 excitations/phase coding step,3-mm-thick slices, and a gap of 1 mm between slices). The coil wasa 3-cm-diameter, home-built solenoid. The field of view was 80mm, and the acquisition matrix was 256X256. Imaging wasperformed within 10 min of contrast injection (Magnevist, 0.6ml/kg i.v.). This assay detected lesions as small as 1.1 mm indiameter.
Statistical analysisStatistical calculations were performed using JMP software
(SAS Institute, Cary, NC).
ImmunohistochemistryMonoclonal mouse anti-rat antibodies (OX-10, OX-6, OX34,
W3/24, OX-18, Mr. S. Lightfoot, University of Texas Southwest-ern, Dallas, TX; OX-42, Biosource International, Camarillo, CA)were reacted with biotin following the manufacturer’s specifica-tions (Vector, Burlingame, CA). Cryosections 10 µm thick wereplaced on microscope slides, air-dried and fixed in acetone. Thesections were then incubated in 0.3% H2O2 in methanol for 10 minat room temperature. After 3 washes, the sections were reacted withthe biotinylated antibodies (10–20 µg/ml) overnight at 4°C in ahumidified chamber. The sections were again washed 3 times,developed, then counterstained in 1% methyl green.
Non-specific esteraseThe cytochemical reaction for non-specific esterase activity was
performed as described (Yamet al., 1971). Briefly, cryostatsections of tissue were fixed for 30 sec in phosphate-bufferedacetone-formaldehyde. Sections were then incubated at roomtemperature in the following medium: 44.5 ml of 0.15 M PBS, 50
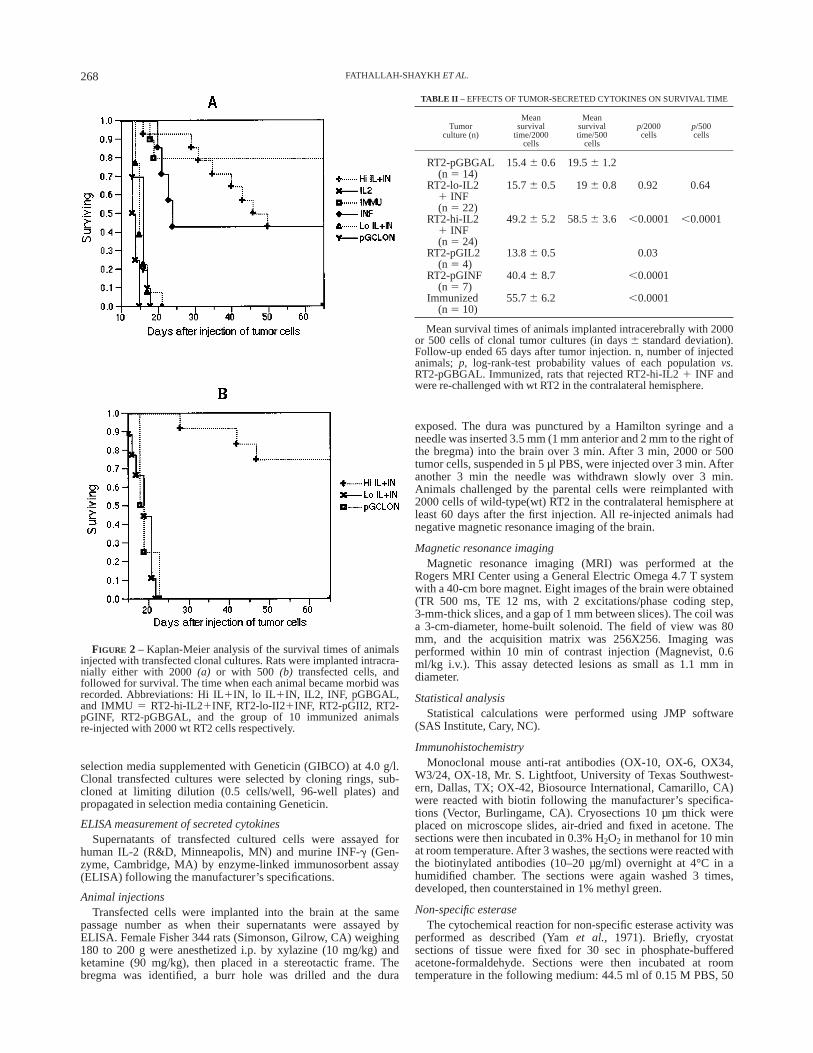
FIGURE 2 – Kaplan-Meier analysis of the survival times of animalsinjected with transfected clonal cultures. Rats were implanted intracra-nially either with 2000(a) or with 500 (b) transfected cells, andfollowed for survival. The time when each animal became morbid wasrecorded. Abbreviations: Hi IL1IN, lo IL 1IN, IL2, INF, pGBGAL,and IMMU 5 RT2-hi-IL21INF, RT2-lo-II21INF, RT2-pGII2, RT2-pGINF, RT2-pGBGAL, and the group of 10 immunized animalsre-injected with 2000 wt RT2 cells respectively.
TABLE II – EFFECTS OF TUMOR-SECRETED CYTOKINES ON SURVIVAL TIME
Tumorculture (n)
Meansurvival
time/2000cells
Meansurvivaltime/500
cells
p/2000cells
p/500cells
RT2-pGBGAL(n 5 14)
15.46 0.6 19.56 1.2
RT2-lo-IL21 INF(n 5 22)
15.76 0.5 196 0.8 0.92 0.64
RT2-hi-IL21 INF(n 5 24)
49.26 5.2 58.56 3.6 ,0.0001 ,0.0001
RT2-pGIL2(n 5 4)
13.86 0.5 0.03
RT2-pGINF(n 5 7)
40.46 8.7 ,0.0001
Immunized(n 5 10)
55.76 6.2 ,0.0001
Mean survival times of animals implanted intracerebrally with 2000or 500 cells of clonal tumor cultures (in days6 standard deviation).Follow-up ended 65 days after tumor injection. n, number of injectedanimals; p, log-rank-test probability values of each populationvs.RT2-pGBGAL. Immunized, rats that rejected RT2-hi-IL21 INF andwere re-challenged with wt RT2 in the contralateral hemisphere.
268 FATHALLAH-SHAYKH ET AL.
mg a-naphthyl acetate dissolved in 2.5 ml ethylene glycol mono-methyl ether and 3 ml hexazotized pararosaniline (Sigma, St.Louis, MO). After washing, the sections were counterstained with1% methyl green. Enzymic activity in spleen sections, used aspositive control, was seen as red granules in the cytoplasm ofmonocytic cells.
RESULTS
Generation of cytokine-secreting tumor cells
To achieve high levels of secreted cytokines, each expressioncassette was designed to carry the cytokine cDNA 38 to a CMV
promoter and SV40 splicer/donor site (Fig. 1). The plasmidpGINF/IL2 was constructed to drive the expression of human IL-2and murine INF-g in a rat brain-tumor cell line (RT2). Similarly,pGIL2, pGBGAL and pGINF drive the expression of IL-2,b-galactosidase and INF-g respectively. In mammalian cells,pGINF/IL2, pGBGAL, pGIL2 and pGINF induce expression of theneomycin resistance gene for selectionin vitro.
Six clonal RT2 cultures transfected with pGINF/IL2 wereisolated. Of these 6, 3 produced at least 192 pg/106 cells/day(pg/M/d) of IL-2 and 171 pg/M/d of INF-g (RT2-hi-IL2 1 INF);the other 3 secreted a maximum of 62 pg/M/d of IL-2 and 17pg/M/d of INF-g (RT2-Io-IL2 1 INF). To examine for synergy, 2
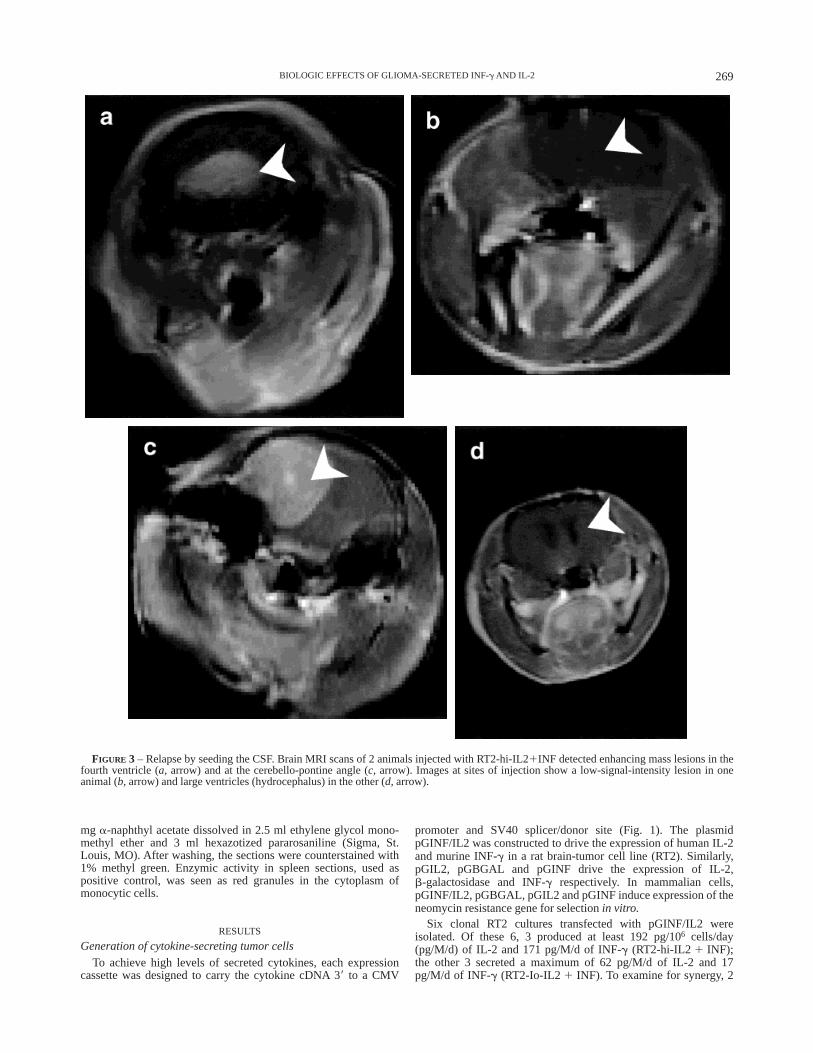
FIGURE 3 – Relapse by seeding the CSF. Brain MRI scans of 2 animals injected with RT2-hi-IL21INF detected enhancing mass lesions in thefourth ventricle (a, arrow) and at the cerebello-pontine angle (c, arrow). Images at sites of injection show a low-signal-intensity lesion in oneanimal (b, arrow) and large ventricles (hydrocephalus) in the other (d, arrow).
269BIOLOGIC EFFECTS OF GLIOMA-SECRETED INF-g AND IL-2

clonal cultures, secreting either 2065 pg/M/d of IL-2 (RT2-pGIL2)or 473 pg/M/d of INF-g (RT2-pGINF), were generated by transfec-tion with pGIL2 and pGINF respectively (Table I). As negativecontrol, a clonal culture transfected with pGBGAL was produced(RT2-pGBGAL).
Effect of tumor-secreted cytokines on survival timeTo study the biological effects of transfected cultures in immune-
competent syngeneic rats, animals were injected intracerebrallywith RT2-pGBGAL (n5 14) or with RT2-hi-IL21 INF (n 5 24).The maximum survival time of animals injected with RT2-pGBGAL was 21 days. RT2-hi-IL21 INF produced a substantialincrease in survival lengths; combining the 2 groups of ratsimplanted with 500 and 2000 cells, 23/24 rats survived longer than21 days and 15/24 (62%) survived longer than 65 days (p , 0.0001;Fig. 2; Table II). There was no difference in survival time amongthe 3 RT2-hi-IL21INF cultures. To investigate whether the bio-logic response was linked to the quantity of secreted cytokines, 22rats were injected with RT2-Io-IL21 INF. The survival times ofanimals implanted with RT2-Io-IL21 INF were comparable to therats injected with RT2-pGBGAL (Fig. 2; Table II); thus, culturessecreting ‘‘low’’ quantities of IL-2 and INF-g may serve as internalnegative controls.
To examine whether IL-2 and INF-g affected tumor growthinvivo, approximately 33 106 cells of RT2 (n5 4), RT2-pGBGAL(n 5 3) or RT2-hi-IL21 INF (n 5 3) were injected s.c. into nudemice. Three weeks later, the mean largest diameters of the tumorswere 1.75, 1.4 and 1.5 cm respectively. Therefore the secretedcytokines exert no direct effects on tumor growth in nude animalsin vivo.
Tumor rejection and relapse by seeding the cerebrospinal fluidParenchymal rejection was defined by the absence of a tumor
mass in the brain parenchyma at the site of implantation. Intracra-nial rejection was defined by the absence of enhancing mass lesionsin all slices of a brain examined by MRI at least 60 days afterimplantation.
Whereas 13/24 animals (54%) implanted with RT2-hi-IL21INF demonstrated intracranial rejection (p , 0.0001, Table III),negative control cultures such as RT2-pGBGAL and RT2-Io-IL21INF were 100% fatal (36/36, Fig. 2a,b). Of the 11 remaining rats, 9developed malignant gliomas at the injection site, while 2 rejectedthe parenchymal brain tumor, but relapsed by developing tumordeposits in the cerebrospinal fluid (CSF; Fig. 3). MRI of theirbrains showed contrast-enhancing lesions in the posterior fossa(Fig. 3a,c) without evidence of mass lesions at the site ofimplantation (Fig. 3b,d; Table III, parenchymal rejection). Patho-logical examination revealed malignant glial tumors in the poste-rior fossa, associated with necrosis of the primary tumor bed in thebrain parenchyma (data not shown).
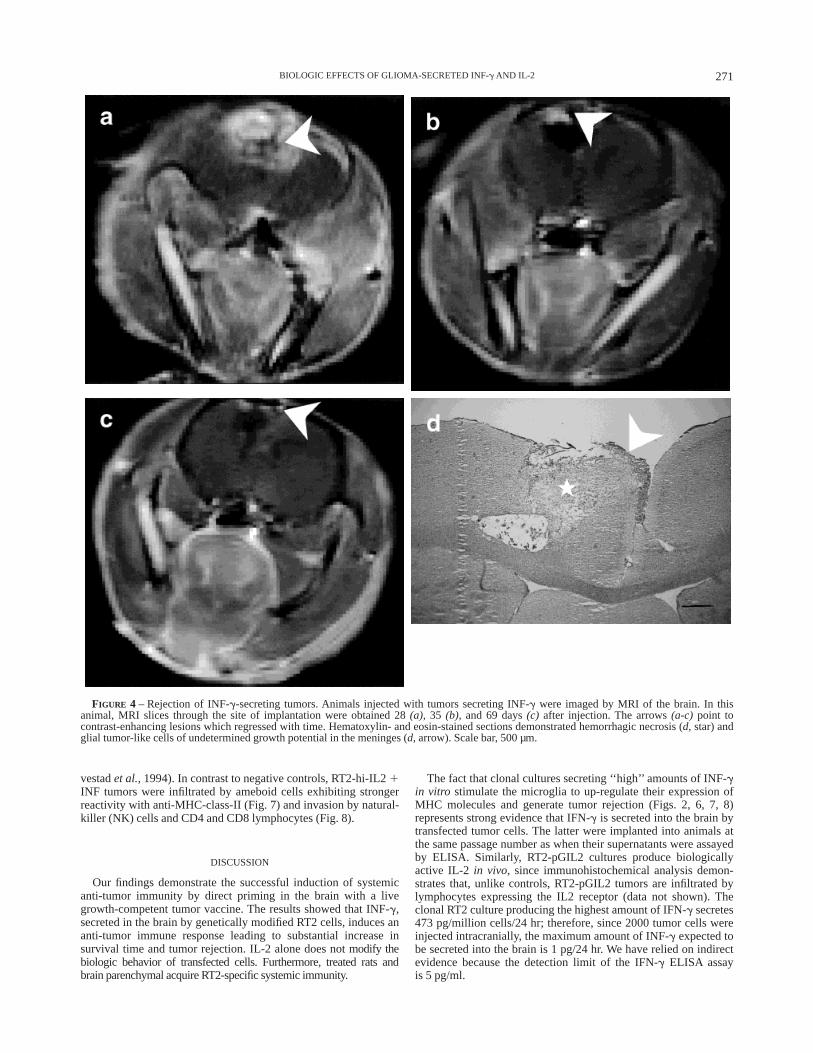
To determine whether the cytokines elicit a synergistic effect,animals were implanted with 2000 cells secreting either IL-2 orINF-g (RT2-pGIL2 or RT2-pGINF). The biologic behavior ofIL-2-secreting tumors was similar to pGBGAL (Fig. 2a); theseanimals showed a large parenchymal tumor mass at autopsy.However, 3/7 rats (43%) injected with INF-g-secreting cellssurvived longer than 65 days (p , 0.0001, Fig. 2). One of these 3animals had a negative MRI of the brain obtained 69 days afterimplantation, while the other 2 developed asymptomatic contrast-enhancing lesions at the injection site (Fig. 4a) which regressedover 41 days (Fig. 4b,c) into parenchymal necrosis (Fig. 4d, 69days after injection). Thus, a total of 3 animals showed intracranialrejection (p 5 0.0028, Table III). The differences in the biologiceffects of INF-g and IL2 1 INF-g on survival time (p 5 0.337,log-rank test) and tumor rejection (p 5 0.57, Fisher’s exact test)were not statistically significant. Moreover, the percentage survivalafter 65 days of the animals injected with 2000 cells of RT2-hi-IL2 1 INF (6/24, 50%) was similar to that of animals injected withRT2-pGINF (43%). Therefore, the biologic behavior of RT2-hi-IL2 1 INF was comparable with that of RT2-pGINF.
Tumor-specific immunityTo study the role of tumor-secreted cytokines in eliciting
protective immunity, 10 rats showing intracranial rejection ofRT2-hi-IL2 1 INF were re-challenged by wild-type(wt) RT2 cellsin the contralateral hemisphere (Fig. 2a). Out of 10 rats, 8 (80%)survived longer than 65 days (p , 0.0001; Fig. 2) and rejected theintracranial wt tumor. In 7 of these animals, the MRI scans werenegative, while one rat showed regression of a contrast-enhancinglesion into necrosis at the injection site (p , 0.0001; Table III).The scans of the latter were very similar to the data showingrejection of the INF-g-secreting tumors (Fig. 4). Two of the tendeveloped meningeal relapse; however, no tumor cells werepresent at the injection sites. Thus, 8/10 animals demonstratedintracranial rejection and 10/10 parenchymal rejection (Table III).
To examine whether priming in the brain elicited systemicimmunity, the animals that rejected intracerebrally implantedRT2-hi-IL2 1 INF were re-injected s.c. with approximately 1031010 wt RT2 cells. Unlike naive rats, the immunized animalsrejected the s.c. tumors (Fig. 5). This protective immunity wasspecific for RT2, since the growth rates of a syngeneic mammary-carcinoma line (MAT B III, ATCC) did not differ between naiveand immunized rats (Fig. 5).
Stimulation of microgliaTo characterize the immune cells associated with this anti-tumor
response, animals injected intracerebrally with RT2-pGBGAL(n 5 3) or RT2-hi-IL21 INF (n 5 5) were killed 2 to 3 weeks afterimplantation. Brains were sectioned and examined at multiple sitesby immunohistochemistry. Hematoxylin-and-eosin-stained sec-tions of the RT2-hi-IL21 INF brains revealed a unique inflamma-tory reaction in the brain parenchyma surrounding the tumor beds,consisting of lymphocytic cells cuffing blood vessels (data notshown).
Microglia are the only CNS-resident cells recognized by themonoclonal antibody OX-42, which also identifies monocytes,macrophages and dendritic cells. Compared with RT2-pGBGAL,parenchymal microglia throughout the brains implanted withRT2-hi-IL2 1 INF demonstrated up-regulation of MHC-class-Iand -class-II expression (Fig. 6). Moreover, brains implanted withRT2-pGIL2 showed anti-MHC-class-I and -class-II reactivity ofmicroglia similar to that shown by the brains injected withRT2-pGBGAL (data not shown).
Both RT2-pGBGAL and RT2-hi-IL21 INF tumors werediffusely infiltrated by round (ameboid) microglia (Fig. 7). TheseOX-42-reactive cells were differentiated from macrophages by theabsence of non-specific esterase activity (data not shown) (Ul-
TABLE III – GENERATION OF TUMOR REJECTION AND PROTECTIVEIMMUNITY
Tumor clones/animals
Intracranialrejection
Parenchymalrejection p
RT2-pGBGAL and RT2-lo-IL2 1 INF
0/36 0/36
RT2-hi-IL2 1 INF 13/24 15/24 ,0.0001RT2-pGINF 3/7 3/7 0.0028RT2-pGIL2 0/4 0/4Immunized 8/10 10/10 ,0.0001
Surviving animals were examined by MRI of the brain at least 60days after tumor implantation. Intracranial and parenchymal rejection,as defined in the text.p, Fisher’s exact probability values for the2-tailed test comparing the populations to RT2-pGBGAL and RT2-lo-IL2 1 INF. Tumor clones, intracerebrally-implanted transfected RT2cultures. Immunized, rats that rejected RT2-hi-IL21 INF and werere-challenged with wt RT2 in the contralateral hemisphere.
270 FATHALLAH-SHAYKH ET AL.
vestadet al.,1994). In contrast to negative controls, RT2-hi-IL21INF tumors were infiltrated by ameboid cells exhibiting strongerreactivity with anti-MHC-class-II (Fig. 7) and invasion by natural-killer (NK) cells and CD4 and CD8 lymphocytes (Fig. 8).
DISCUSSION
Our findings demonstrate the successful induction of systemicanti-tumor immunity by direct priming in the brain with a livegrowth-competent tumor vaccine. The results showed that INF-g,secreted in the brain by genetically modified RT2 cells, induces ananti-tumor immune response leading to substantial increase insurvival time and tumor rejection. IL-2 alone does notmodify thebiologic behavior of transfected cells. Furthermore, treated rats andbrain parenchymal acquire RT2-specific systemic immunity.
The fact that clonal cultures secreting ‘‘high’’ amounts of INF-gin vitro stimulate the microglia to up-regulate their expression ofMHC molecules and generate tumor rejection (Figs. 2, 6, 7, 8)represents strong evidence that IFN-g is secreted into the brain bytransfected tumor cells. The latter were implanted into animals atthe same passage number as when their supernatants were assayedby ELISA. Similarly, RT2-pGIL2 cultures produce biologicallyactive IL-2 in vivo, since immunohistochemical analysis demon-strates that, unlike controls, RT2-pGIL2 tumors are infiltrated bylymphocytes expressing the IL2 receptor (data not shown). Theclonal RT2 culture producing the highest amount of IFN-g secretes473 pg/million cells/24 hr; therefore, since 2000 tumor cells wereinjected intracranially, the maximum amount of INF-g expected tobe secreted into the brain is 1 pg/24 hr. We have relied on indirectevidence because the detection limit of the IFN-g ELISA assayis 5 pg/ml.
FIGURE 4 – Rejection of INF-g-secreting tumors. Animals injected with tumors secreting INF-g were imaged by MRI of the brain. In thisanimal, MRI slices through the site of implantation were obtained 28(a), 35 (b), and 69 days(c) after injection. The arrows(a-c) point tocontrast-enhancing lesions which regressed with time. Hematoxylin- and eosin-stained sections demonstrated hemorrhagic necrosis (d, star) andglial tumor-like cells of undetermined growth potential in the meninges (d, arrow). Scale bar, 500 µm.
271BIOLOGIC EFFECTS OF GLIOMA-SECRETED INF-g AND IL-2
Priming by s.c. immunization (Scheinberget al., 1965) orimplantation of genetically modified brain tumor cells is widelyreported to produce anti-glioma inflammatory effects (Trojanet al.,1992; Sampsonet al., 1996). When conditions for the peripheralpriming of tumor-specific responses are examined in chimericmice, tumor antigens are believed to be exclusively presented byhost bone-marrow-derived cells (Huanget al., 1994). In such amodel, priming of T cells is thought to occur in the lymph nodesdraining the vaccine site, after which activated T lymphocytesmigrate across the blood brain barrier to mediate effector immuno-logic functions within the CNS (Holladayet al.,1992).
The concept of utilizing genetically modified tumor cells tosecrete cytokines as peripheral tumor vaccines has shown effective-ness in producing tumor immunity in many animal models ofsystemic malignancy (Dranoffet al.,1993). Our results are novel,since they provide strong evidence of the existence of an induciblebrain-inherent capacity to initiate an immune response. Thesefindings further challenge the classical theory that the brain is‘‘immunologically privileged’’ and question the differences andsimilarities between tumor neuro-immunology and systemic tumorimmunology. The results raise the possibility that breach of theprivileged site is caused by breakdown of the blood brain barrier(BBB). The intracerebral injection into Fisher 344 rats of fibro-blasts transduced by retroviral vectors to secrete either 5 ng/M/d ofIL-2 or ,1 ng/M/d of INF-g produces brain edema and disruptionof the BBB (Tjuvajevet al.,1995). Furthermore, i.v. injection intoFisher 344 rats of 23 106 units/kg of IL-2 increases BBBdisruption in tumor-bearing brain, but does not affect the vascularpermeability of normal brain (Alexanderet al., 1989). In ourmodel, the potential cytokine-mediated breakdown of the BBB isprobably not the principal cause of destruction of the privilegedsite, because IL-2 disrupts the BBB and does not elicit tumorrejection (Tjuvajevet al.,1995; Alexanderet al.,1989).
Microglia comprise up to 13% of the cells in the human whitematter. Although the origin of microglia is unknown, the prevailingview is that they originate from bone-marrow-derived monocytesentering the CNS during fetal life (Hickey and Kimura, 1988).Microglia share common antigens with macrophages and dendriticcells, and recent evidence suggests that they are functionallyrelated to dendritic APC and to phagocytic macrophages (Ulvestadet al.,1994; Hickey and Kimura, 1988). Although resting microgliaappear to be quiescent, they are highly responsive to stimulation byINF-g (Cash and Rot, 1994; Ulvestadet al.,1994). Consistent withcurrent understanding of systemic immunology, INF-g-treatedmicroglia express the molecules necessary for antigen presentation;they up-regulate their expression of MHC-class-I and -class-IImolecules and several co-stimulatory molecules for T-cell activa-tion, including lymphocyte-function antigen-1 (LFA-1), LFA-3,
intercellular-adhesion molecule-1 (ICAM-1) and CD80/CD86 (Xuand Ling, 1994; Frohmanet al.,1989). The CD80/CD86 moleculeplays a functional role in the ability of microglia to serve as CNSantigen-presenting cells, and several studies have emphasized theimportance of CD80/CD86 and CD28 interactions in peripheralT-cell stimulation (Williamset al.,1994). The clustering of T cellsby dendritic cells appears to be mediated via bi-directional LFA-1/ICAM-1 interaction and unidirectional LFA-3/CD2 interaction. Further-more, INF-g-treated microglia function as competent APC, as evi-denced by their capacity to present recall antigen to T lymphocytesinvitro, antigen to unprimed CD41 T cells with resultant T cell activationand proliferation, and support the proliferation of previously sensitizedTcells (Williams et al., 1994). The notion that microglia performimmunologic effector functions is based on the fact that when stimulatedby INF-g, they mediate direct tumor-cell cytotoxicity and that they arephagocytic (Freiet al.,1987).
Our results are best explained by the hypothesis that INF-gmediates the transformation of microglia into an ‘‘immune-active’’state leading to T-cell priming and enhanced tumor-infiltration byeffector immune cells. In addition, unlike IL-2, INF-g mayinfluence the T-cell response by inhibiting the proliferation of Th2cells and favoring the development of Th1 cells; the latter isthought to be mediated by induction of expression of theb2sub-unit of the IL-12 receptor (Szaboet al.,1997). This study doesnot necessarily rule out the theory that tumor cells function as APC,nor the possibility that tumor antigens find their way to thelymphoid organs where they are processed by peripheral APC.
Our findings provide strong evidence that tumor-secreted INF-gelicits activation of microglia throughout the brain. Unlike thecontrol, tumors secreting INF-g 1 IL-2 stimulate the microglia toup-regulate their expression of MHC-class-I and class-II molecules(Fig. 6). Furthermore, the data support the notion that INF-g 1IL-2 elicit higher expression of MHC-class-II molecules in tumor-infiltrating microglia (Fig. 7). The effects of RT2-hi-IL21 INF aremost likely produced by INF-g, in view of the fact that tumorssecreting IL-2 only do not modify the expression of MHCmolecules by the microglia. The animal model presented in thisexperimental paradigm may help to define the site where CNS-derived antigens are presented to T cells and to understand thedifferential roles of microglia and peripheral lymphocytes inimmune surveillance, tumor-antigen presentation and tumor-cellcytotoxicity in the nervous system.
It is remarkable that, although parenchymal tumor rejection isassociated with systemic immunity (Fig. 5), some animals relapseby developing meningeal and intraventricular tumors consistent withseeding of the CSF (Fig. 3). In these animals, the CSF may have beeninoculated either by the injecting needle as it crosses the leptomeningesor because of the proximity of the lateral ventricles to the injection site.Potential causes of this relapse include low concentrations ofin situINF-g due to dilution by the CSF, and poor penetration of microglia andeffector cells into the ventricular portion of the tumor.
The novel strategy of vaccination by direct priming into the brainoffers hope of translation into treatment modalities for patientssuffering from primary or metastatic brain tumors. It is noteworthythat the immunized animals do not exhibit signs of neurologictoxicity. Our results suggest that, given appropriate stimulus, themammalian brain is capable of evolving into an ‘‘immunologicallyactive’’ organ.
ACKNOWLEDGEMENTS
We thank Dr. J. Foreman, Dr. P. Lipsky and Dr. S.C. Schold forhelpful discussions, also Dr. R. Rosenberg, Dr. P. Nisen, Dr. G.Smith, Dr. M. Bennett, Dr. J. Minna and Dr. D. Thiele for reviewingthis manuscript, and Dr. E. Babcock for guidance in imaging the ratbrains.
FIGURE 5 – Induction of RT2-specific systemic immunity. Immu-nized rats that rejected intracerebral RT2-hi-IL21INF (IMM) andnaive animals (N) were implanted S.C. either with 10 million cells ofwt RT2 (-RT2; n5 4 in each group) or with 5 million cells of MAT BIII, a syngeneic mammary-carcinoma line (-MATB, n5 3 in eachgroup). The mean largest diameters (cm) were measured using calipers.
272 FATHALLAH-SHAYKH ET AL.
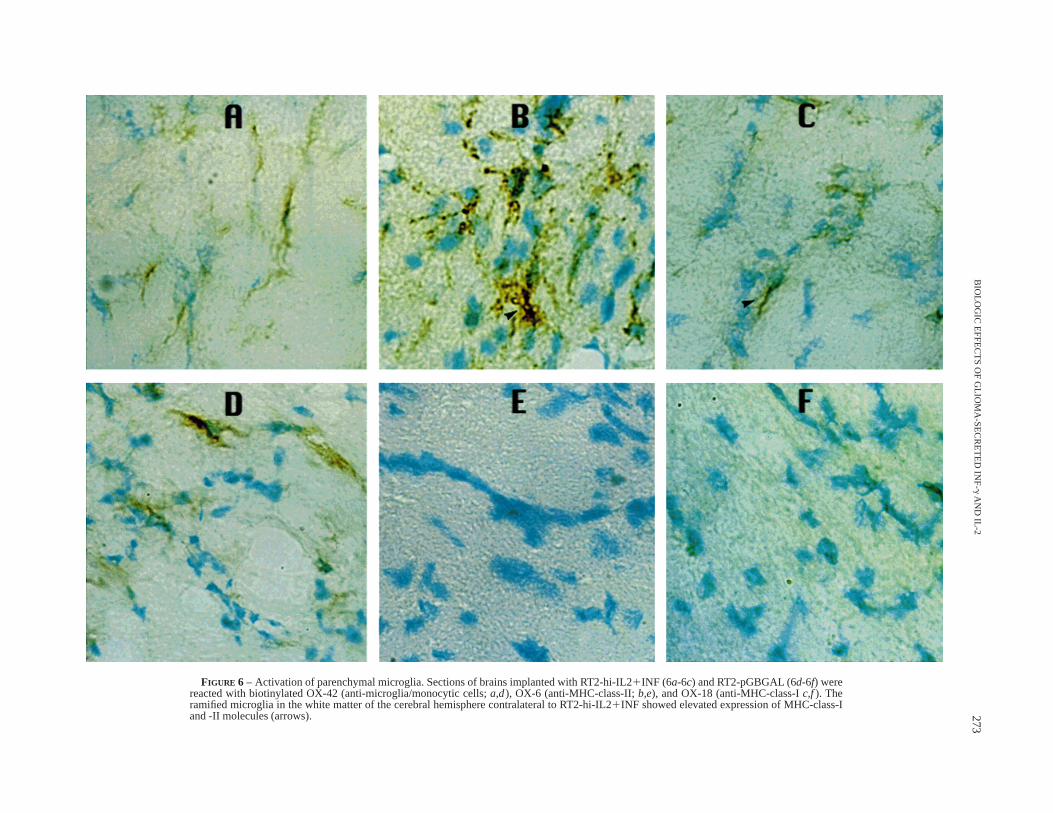
FIGURE 6 – Activation of parenchymal microglia. Sections of brains implanted with RT2-hi-IL21INF (6a-6c) and RT2-pGBGAL (6d-6f) werereacted with biotinylated OX-42 (anti-microglia/monocytic cells;a,d), OX-6 (anti-MHC-class-II;b,e), and OX-18 (anti-MHC-class-Ic,f). Theramified microglia in the white matter of the cerebral hemisphere contralateral to RT2-hi-IL21INF showed elevated expression of MHC-class-Iand -II molecules (arrows). 273
BIO
LOG
ICE
FF
EC
TS
OF
GLIO
MA
-SE
CR
ET
ED
INF
-gAN
DIL-2
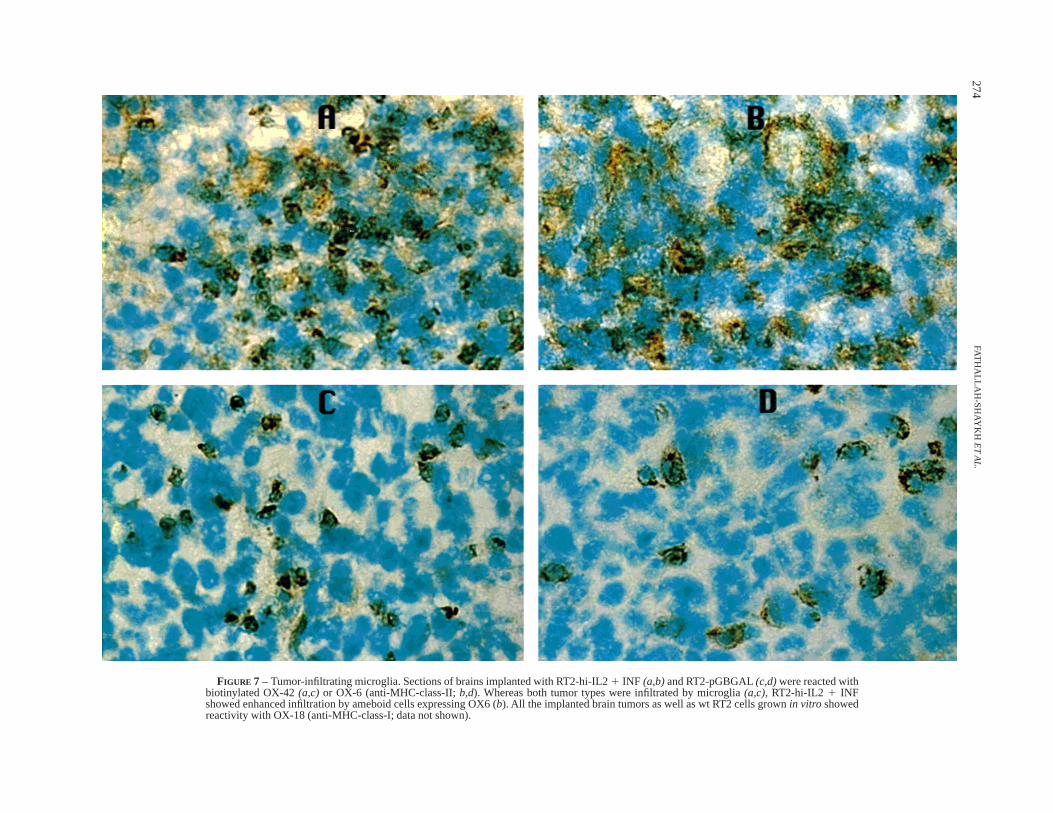
FIGURE 7 – Tumor-infiltrating microglia. Sections of brains implanted with RT2-hi-IL21 INF (a,b)and RT2-pGBGAL(c,d)were reacted withbiotinylated OX-42(a,c) or OX-6 (anti-MHC-class-II;b,d). Whereas both tumor types were infiltrated by microglia(a,c), RT2-hi-IL2 1 INFshowed enhanced infiltration by ameboid cells expressing OX6 (b). All the implanted brain tumors as well as wt RT2 cells grownin vitro showedreactivity with OX-18 (anti-MHC-class-I; data not shown).
274FAT
HA
LLAH
-SH
AY
KH
ET
AL
.
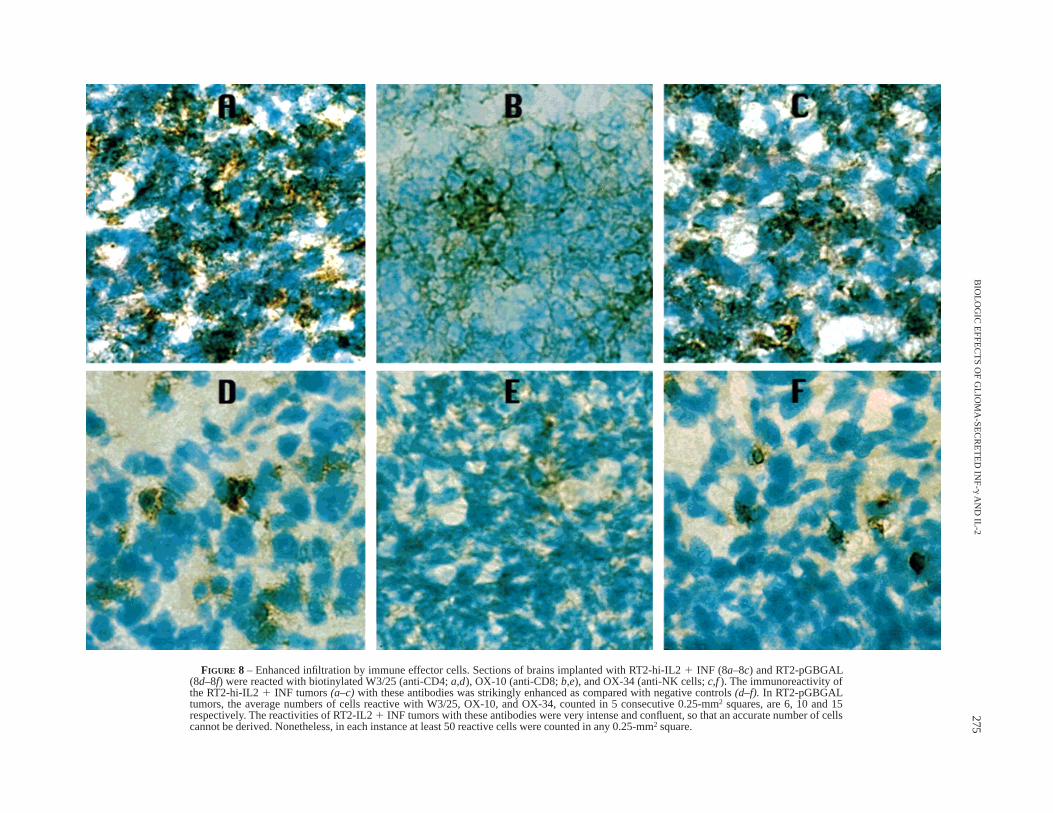
FIGURE 8 – Enhanced infiltration by immune effector cells. Sections of brains implanted with RT2-hi-IL21 INF (8a–8c) and RT2-pGBGAL(8d–8f) were reacted with biotinylated W3/25 (anti-CD4;a,d), OX-10 (anti-CD8;b,e), and OX-34 (anti-NK cells;c,f). The immunoreactivity ofthe RT2-hi-IL21 INF tumors(a–c)with these antibodies was strikingly enhanced as compared with negative controls(d–f). In RT2-pGBGALtumors, the average numbers of cells reactive with W3/25, OX-10, and OX-34, counted in 5 consecutive 0.25-mm2 squares, are 6, 10 and 15respectively. The reactivities of RT2-IL21 INF tumors with these antibodies were very intense and confluent, so that an accurate number of cellscannot be derived. Nonetheless, in each instance at least 50 reactive cells were counted in any 0.25-mm2 square.
275B
IOLO
GIC
EF
FE
CT
SO
FG
LIOM
A-S
EC
RE
TE
DIN
F-gA
ND
IL-2

REFERENCES
ALEXANDER, J.T., SARIS, S.C. and OLDFIELD, E.H., The effect of interleu-kin-2 on the blood-brain barrier in the 9L gliosarcoma rat model.J.Neurosurg.,70,92–96 (1989).BARKER, C.F. and BILLINGHAM , R.E., Immunologically privileged sites.Advanc. Immunol.,25,1–54 (1971).CASH, E. and ROT, O., Microglia cells qualify as the stimulators ofunprimed CD41 and CD81 T lymphocytes in the central nervous system.Clin. exp. Immunol.,98,313–318 (1994).CULLEN, B.R. Expression of a cloned human interleukin-2 cDNA isenhanced by the substitution of a heterologous mRNA leader region.DNA,7, 645–650 (1988).DAAR, A.S., FUGGLE, S.V., FABRE, J.W., TING, A. and MORRIS, P.J., Thedetailed distribution of MHC-class-II antigens in normal human organs.Transplantation,38,293–298 (1984).DRANOFF, G., JAFFEE, E., LAZENBY, A., GOLUMBECK, P., LEVITSKY, H.,BROSE, K., JACKSON, V., HIROFUMI, H., PARDOLL, D. and MULLIGAN , R.C.,Vaccination with irradiated tumor cells engineered to secrete murinegranulocyte-macrophage-colony-stimulating factor stimulates potent, spe-cific, and long-lasting anti-tumor immunity.Proc. nat. Acad. Sci. (Wash.),90,3539–3543 (1993).FREI, K., SIEPL, C., GROSCURTH, P., BODMER, S., SCHWERDEL, C. andFONTANA, A., Antigen presentation and tumor cytotoxicity by interferon-g-treated microglial cells.Europ. J. Immunol.,17,1271–1278 (1987).FROHMAN, E.M., FROHMAN, T.C., DUSTIN, M.L., VAYUVEGULA , B., CHOI, B.,GUPTA, A., VAN DER NOORT, S., GUPTA S. The induction of intercellularadhesion molecule 1 (ICAM-1) expression on human fetal astrocytes byinterferon-g, tumor necrosis factor-a, lymphotoxin, and interleukin-1:relevance to intracerebral antigen presentation.J. Neuroimmunol.,23,117–124 (1989).HARLING-BERG, C., KNOPF, P.M., MERRIAM, J. and CSERR, H.F., Role ofcervical lymph nodes in the systemic humoral immune response to humanserum albumin micro-infused into rat cerebrospinal fluid.J. Neuroimmu-nol.,25,185–193 (1989).HART, M.N. and FABRY, Z., CNS antigen presentation.Trends Neurosci.,18,475–481 (1995).HICKEY, W.F., HSU, B.L. and KIMURA, H., T-lymphocytes entry into thecentral nervous system.J. neurosci. Res.,28,254–260 (1991).HICKEY, W.F. and KIMURA, H., Perivascular microglia of the CNS arebone-marrow-derived and present antigenin vivo. Science,239, 290–292(1988).HOLLADAY, F.P., HEITZ, T., CHEN, Y.-L., CHIGA, M. and WOOD, G.W.,Successful treatment of malignant rat glioma with cytotoxic T lymphocytes.Neurosurgery,31,528–533 (1992).HUANG, A.Y.C., GOLUMBEK, P., AHMADZADEH, M., JAFFE, E., PARDOLL, D.and LEVITSKY, H., Role of bone-marrow-derived cells in presentingMHC-class-I-restricted tumor antigens.Science,264,961–965 (1994).JANKOVIC, B.D., DRASKOCI, M. and ISAKOVIC, K., Antibody response inrabbits following injection of sheep erythrocytes into the lateral ventriclesof brain.Nature (Lond.),191,288–289 (1961).LICHTOR, T., GLICK, R.P., KIM, T.S., HAND, R. and COHEN, E.P., Prolongedsurvival of mice with glioma injected intracerebrally with double cytokine-secreting cells.J. Neurosurg.,83,1038–1044 (1995).
MAASS, G., SCHMIDT, W., BERGER, M., SCHILCHER, F., KOSZIK, F., SCHNEE-BERGER, A., STINGL, G., BIRNSTIEL, M.L. and SCHWEIGHOFFER, T., Priming oftumor-specific T cells in the draining lymph nodes after immunization withinterleukin-2-secreting tumor cells: three consecutive stages may berequired for successful tumor vaccination.Proc. nat. Acad. Sci. (Wash.),92,5540–5544 (1995).
MEDAWAR, P.B., Immunity to homologous grafted skin. III. The fate of skinhomografts transplanted to the brain, to subcutaneous tissue, and to theanterior chamber of the eye.Brit. J. exp. Pathol.,29,58–69 (1948).
SAMPSON, J.H., ARCHER, G.E., ASHLEY, D.M., FUCHS, H.E., HALE, L.P.,DRANOFF, G. and BIGNER, D.D., Subcutaneous vaccination with irradiated,cytokine-producing tumor cells stimulates CD81-cell-mediated immunityagainst tumors located in the ‘‘immunologically privileged’’ central nervoussystem.Proc. nat. Acad. Sci. (Wash.),93,10399–10404 (1996).
SCHEINBERG, L.C., LEVY, A. and EDELMAN, F., Is the brain an ‘‘immunologi-cally privileged site’’? 2. Studies in induced host resistance to transplant-able mouse glioma following irradiation of prior implants.Arch. Neurol.,13,283–286 (1965).
SZABO, S.J., DIGHE, A.S., GUBLER, U. and MURPHY, K.M., Regulation of theinterleukin (IL)-12R b2 sub-unit expression in developing T-helper-1 (Th1)and Th2 cells.J. exp. Med.,185,817–824 (1997).
TJUVAJEV, J., GANSBACHER, B., DESAI, R., BEATTIE, B., KAPLITT, M., MATEI,C., KOUTCHER, J., GILBOA, E. and BLASBERG, R., RG-2-glioma growthattenuation and severe brain edema caused by local production of interleu-kin-2 and interferon-gamma.Cancer Res.,55,1902–1910 (1995).
TROJAN, J., BLOSSEY, B.K., JOHNSON, T.R., RUDIN, S.D., TYKOCINSKI, M. andILAN, J., Loss of tumorigenicity of rat glioblastoma directed by episome-based anti-sense cDNA transcription of insulin-like growth factor I.Proc.nat. Acad. Sci. (Wash.),89,4874–4878 (1992).
ULVESTAD, E., WILLIAMS , K., BJERKVIG, R., TIEKOTTER, K., ANTEL, J. andMATRE, R., Human microglia cells have phenotypic and functionalcharacteristics in common with both macrophages and dendritic antigen-presenting cells.J. Leukoc. Biol.,56,732–739 (1994).
WELLER, R.O., KIDA, S. and ZHANG, E.T., Pathways of fluid drainage fromthe brain: morphological aspects and immunological significance in rat andman.Brain Pathol.,2, 277–284 (1992).
WILLIAMS , K., ULVESTAD, E. and ANTEL, J.P., B7/BB-1 antigen expressionon adult human microglia studiedin vitro and in situ. Europ. J. Immunol.,24,3031–3037 (1994).
XU, J. and LING, E.A., Up-regulation and induction of major-histocompat-ibility-complex-class-I and -class-II antigens on microglial cells in earlypostnatal rat brain following intraperitoneal injections of recombinantinterferon-gamma.Neuroscience,60,959–967 (1994).
YAM, L.T., LI, C.J. and CROSBY, W.H., Cytochemical identification ofmonocytes and granulocytes.Amer. J. clin. Pathol.,55,283–290 (1971).
YAMADA , S., DEPASQUALE, M., PATLAK , C.S. and CSERR, H.F., Albuminoutflow into deep cervical lymph from different regions of rabbit brain.Amer. J. Physiol.,261,H1197–H11204 (1991).
YOFFEY, J.M. and COURTICE, F.C.,Lymphatics, lymph and the lymphomy-eloid complex,Academic Press, New York (1970).
276 FATHALLAH-SHAYKH ET AL.




























