Embed Size (px)
Citation preview

Genes, Brain and Behavior (2013) doi: 10.1111/gbb.12112
Propensity for social interaction predictsnicotine-reinforced behaviors in outbred rats
T. Wang†, W. Han†, B. Wang‡, Q. Jiang†,L. C. Solberg-Woods§, A. A. Palmer¶,∗∗and H. Chen†,∗
†Department of Pharmacology, University of Tennessee HealthScience Center, Memphis, TN, USA, ‡College of Pharmacy,Shaanxi University of Chinese Medicine, Xi’an, PR China,§Human and Molecular Genetics Center, Medical College ofWisconsin, Milwaukee, WI, ¶Department of Human Genetics,and ∗∗Department of Psychiatry and Behavioral Neuroscience,The University of Chicago, Chicago, IL, USA††H.C., A.P. and L.C.S.-W. were responsible for the studyconcept and design. T.W., W.H., B.W. and Q.J. contributed tothe acquisition of animal data. H.C., T.W. and W.H. analyzed thedata and interpreted the findings. T.W. and W.H. drafted themanuscript. H.C., A.P. and L.C.S.-W. provided critical revision ofthe manuscript for important intellectual content. All authorscritically reviewed content and approved final version forpublication.
*Correspondence author: H. Chen, Department of Pharmacol-ogy, University of Tennessee Health Science Center, Suite 115,Crowe Building, 874 Union Ave., Memphis, TN 38163, USA.E-mail: [email protected]
Social and genetic factors can influence smoking
behavior. Using olfactogustatory stimuli as the sensory
cue for intravenous nicotine self-administration (SA),
we previously showed that social learning of nicotine
contingent odor cue prevented rats from developing
conditioned taste aversion and allowed them to instead
establish stable nicotine SA. We hypothesized that
genetic factors influenced socially acquired nicotine SA.
A heterogeneous stock (HS; N/NIH) of outbred rats was
trained to self-administer nicotine using the social learn-
ing protocol. Both male and female HS rats acquired
nicotine SA, but females self-administered more nico-
tine than males. After extinction, the context previously
paired with nicotine SA, in conjunction with socially
transmitted drug cues, was sufficient to cause reinstate-
ment of drug-seeking behavior. Wide variation in both
nicotine intake and reinstatement was observed. Using
multiple regression analysis, we found that measures of
social interaction were significant predictors of nicotine
intake and reinstatement of drug seeking in both males
and females. Furthermore, measures of depression were
predictors of nicotine intake in both males and females,
anxiety was a predictor only in males and response
to novelty was a predictor only in females. In males,
measures of both depression and anxiety predicted
nicotine reinstatement. Together, these data supported
the ideas that genetically determined propensities
for emotional and social phenotypes are significant
determinants for nicotine-reinforced behavior, and that
the HS rat is a suitable tool for dissecting genetic
mechanisms that may underlie the interaction between
social behavior, anxiety, depression and smoking.
Keywords: Nicotine, outbred, rats, self-administration, sociallearning
Received 25 August 2013, revised 14 October 2013 and 19November 2013, accepted for publication 26 November 2013
Cigarette smoking is one of the leading preventable causesof death worldwide; smoking results in approximately 5million deaths per year (Mathers & Loncar 2006). Manyaspects of smoking behavior are influenced by geneticfactors (Carmelli et al. 1992; Pergadia et al. 2006). Overall,the heritability of smoking is estimated to be 0.4–0.6 (Hallet al. 2002; Li et al. 2003). Genome-wide studies havefound that variants of many genes, such as the CHRNA5-CHRNA3-CHRNB4 cluster (Chen et al. 2012b; Saccone et al.2009; Thorgeirsson et al. 2008, 2010), CYP2A6 (Siedlinskiet al. 2011; Thorgeirsson et al. 2010) and brain-derivedneurotrophic factor (Tobacco and Genetics Consortium2010), contribute to smoking behavior (e.g. cigarettes perday, nicotine dependence and cessation). However, thesepolymorphisms account for a small amount of the heritablevariance in smoking behaviors (<1%), suggesting that mostof the genetic factors determining smoking behavior are stillunknown (Thorgeirsson et al. 2008, 2010).
Compared to human studies, rodent models can bet-ter control both environmental and genetic factors. Manyaspects of cigarette smoking have been modeled usingnicotine self-administration (SA) in rats, such as blood nico-tine levels, dose responses, drug dependence, withdrawal,relapse and response to pharmacological intervention (LeFoll & Goldberg 2006; Matta et al. 2007; O’Dell & Khroyan2009). The effect of genetic factors on nicotine SA has alsobeen shown. For example, large variation in nicotine intakewas found across 12 isogenic strains of rats, with heritabilityestimated to be 0.64 (Chen et al. 2012a), supporting theidea that rats are suitable tools for studying the genetics ofsmoking-related behaviors.
Social environment is also critical for smoking. In fact,peer smoking is one of the strongest predictors of smokinginitiation (Greenlund et al. 1997; White et al. 2008). Usingolfactogustatory (OG) stimuli as the contingent sensory cue
© 2013 John Wiley & Sons Ltd and International Behavioural and Neural Genetics Society 1

Wang et al.
for intravenous nicotine delivery, we demonstrated the roleof social learning in nicotine SA (Chen et al. 2011). In thismodel, a rat self-administered nicotine when accompaniedby a demonstrator rat. These two rats were separated by adivider that allowed orofacial interaction. Licking was usedas the operant behavior to contingently deliver OG cueand nicotine. Stable nicotine SA was established when thedemonstrator rat had access to the same OG cue as the SArat. In contrast, conditioned taste aversion was developedwhen the OG cue was withheld from the demonstrator rat.
Our long-term goal is to study the hypothesis thatgenetically determined propensity for social interactionpredicts nicotine-reinforced behaviors. In an effort to laythe groundwork for such studies, we examined the nicotineSA of a population of heterogeneous stock (HS) rats (officialdesignation: N/NIH) using the social learning protocol. TheHS rat was derived from eight inbred founder strains (Hansen& Spuhler 1984). After more than 60 generations of rotationaloutbreeding, each HS rat represents a unique random mosaicof the founders (Mott et al. 2000); these rats representthe most highly recombinant rat intercross available. Wealso assessed novelty exploration, anxiety, depression andsocial interaction phenotypes in each rat and used multipleregression to identify phenotypes that predicted nicotineintake and reinstatement of drug-seeking behavior.
Materials and methods
AnimalsHeterogeneous stock rats (52 males and 48 females) weretransferred from the Medical College of Wisconsin (NMcwi:HS) toUniversity of Tennessee Health Science Center. After quarantine,these rats were given at least 7 days of acclimation to a reversed12:12 h light–dark cycle (lights off at 0830 h). Eight adolescentSprague–Dawley rats (four for each sex) (Harlan Laboratories,Madison, WI, USA) were used as the stimulus rats in thesocial interaction test. An additional 30 female and 30 maleSprague–Dawley rats were used as demonstrators in the nicotineSA experiment. Standard rat chow and water were provided adlibitum. All rats were group housed with two to four same-sex peersthroughout the experiments to avoid social isolation. All procedureswere conducted in accordance with the NIH Guidelines concerningthe Care and Use of Laboratory Animals, as approved by theInstitutional Animal Care and Use Committee of the University ofTennessee Health Science Center.
Study designAll HS rats were young adults (55–65 days old) when tests began.Each HS rat was first tested in a series of behavioral tests, one testper day, conducted in the dark phase of the light cycle (0900–1600h). These tests were conducted in the following sequence: openfield test, novel object interaction test, social interaction test, tailsuspension test 1 and tail suspension test 2. Average body weightsat the beginning of the experiment were 201.4 ± 3.4 g for femalesand 312.4 ± 4.7 g for males. The first three tests were conductedin the same open field and recorded using the same video capturesystem. The sequence and procedures of these tests were adaptedfrom two protocols (Green et al. 2013; Vetter-O’Hagen & Spear 2012).
Open field testTwo open field test chambers constructed using black acrylic plastic,measuring 100 cm (l) × 100 cm (w) × 50 cm (h), were placed side by
side. The floors were painted with black or white acrylic paint (ART-Alternatives, ASTM D-4236, Emeryville, CA, USA) to contrast thecoat of the animals. The test chambers were illuminated by a long-range, 850-nm infrared illuminator (LIR850-70, LDP LLC, Carlstadt,NJ, USA) located 160 cm above the center of the two test chambers.No source of visible light was present during behavioral testingwith the exception of a flat panel monitor (Dell 1908FP). A digitalcamera (Panasonic WV-BP334) fitted with an 830-nm infrared filter(X-Nite830-M37, LTP LLC, Carlstadt, NJ, USA) located next to theinfrared light source was used to record the behavior of the rats.All rats were released at the same corner of the test chamber anddata were collected for 1 h on day 1 and 20 min on day 2 (novelobject) and day 3 (social interaction). Ethovision® XT video trackingsystem (Version 4.0; Noldus Information Technology, Wageningen,The Netherlands) was used to obtain the total distance traveled andthe duration the animal was present in the center of the test field(a circular region with a diameter of 20 cm). Distance traveled in thecenter zone was determined by analyzing exported data using the Rstatistical language (Version 3.0.2).
Novel object testThe novel object was a cylindrical cage constructed using 24 alu-minum rods (30 cm in length) that were spaced 1.7 cm apart. The bot-tom and top of the cage (15 cm in diameter) were manufactured usingMakerbot Replicator 2 (Makerbot Industries, Brooklyn, NY, USA).The design can be downloaded from http://www.thingiverse.com/haochen/designs. One novel object was placed into the center ofeach open field before testing. The duration and distance traveled bythe HS rat in the center zone, where the novel object was located,were used as an index of the novelty-seeking phenotype. Otheraspects of the test were the same as open field test with theexception that behavior was recorded only for 20 min.
Social interaction testA randomly selected stimulus Sprague–Dawley rat of the same sexas the HS test rat was placed into the cylindrical cage (describedabove) before the test rat was placed into the open field. The stimulusand test rats were never housed together and thus were unfamiliarto each other. No social isolation was conducted on either rat. Eachstimulus rat was used once per day. The test duration was 20 min.The duration and distance traveled in the center zone by the HS rats,where the stimulus rat was located, were used as an index of socialinteraction. Other aspects of the test were the same as the openfield test.
Elevated plus maze testThe elevated plus maze was constructed using black acrylic glass.The platform was 60 cm above the floor, with all four arms measuring12 cm (W ) × 50 cm (L). The two opposing closed arms had wallsmeasuring 30 cm (H). Rats were placed into the center of the mazefacing the closed arm. The behavior of the rat was recorded for6 min using the digital video system described above. Ethovisionwas used to extract the time the rats spent in each arm. Thepercent of time spent on the two open arms was then obtained foreach rat (Carobrez & Bertoglio 2005) and were used as a measureof anxiety.
Tail suspension testEach rat was hung upside-down by an adhesive tape, approximately2 cm from the tip of the tail, to a vertical surface. The positionwas adjusted so that the rat’s forepaws could lightly touch a sturdyplatform and thus minimize the weight sustained by its tail (Hinojosaet al. 2006). The rat was illuminated by backscatter infrared light. Thebehavior of the rat was recorded for 6 min. Ethovision was used toobtain the time the rat was immobile by setting the threshold at 5%pixel change. Data obtained using this threshold were in agreementwith those provided by a trained technician. Immobile time wasused as an index of depression. Rats were tested twice on twoconsecutive days.
2 Genes, Brain and Behavior (2013)

Propensity for social interaction predicts nicotine-reinforced behaviors
Figure 1: Illustration of the operant chamber. Each operantchamber contained a perforated divider that separated theself-administration and demonstrator rats to each side. Socialinteraction by orofacial contact through the divider wasmonitored by embedded sets of infrared sensors. Two drinkingspouts on the self-administration side were connected tolickometer controllers that recorded the licks. Licks on theactive spout meeting fixed-ratio 10 schedule triggered concurrentdelivery of an olfactogustatory cue and a intravenous nicotineinfusion. The demonstrator had unrestricted access to the sameolfactogustatory cue but did not receive nicotine.
Nicotine SA in social contextThe nicotine SA procedure in social context was conducted accordingto our published protocol (Chen et al. 2011). An illustrationof the operant chamber setup was provided in Fig. 1. Briefly,jugular catheters (Micro-Renathane® tubing; Braintree Scientific Inc.,Braintree, MA, USA) were implanted under isoflurane anesthesia.Ketoprofen (2 mg/kg, s.c.) was given for postoperative analgesia.After 3 days of recovery, rats were given access to nicotine SA3 h/day for 12 days in the dark phase of the light cycle. Eachoperant chamber (Med Associates, St. Albans, VT, USA) contained aperforated divider located in the middle that separated the SA rat anda randomly selected demonstrator to each side. The demonstratorand the SA rat were always the same sex. Social interaction byorofacial contact through six holes in the divider was monitoredby two sets of infrared sensors embedded in the divider, one foreach rat. Two drinking spouts were fitted on the same wall of theoperant chamber for the SA rat. Each spout connected to a contactlickometer controller allowing the number and timing of licks to berecorded. A solution of saccharin (0.4%) and unsweetened grapeKool-Aid (0.1%) was used as OG cue. The SA was conducted usinga fixed-ratio 10 (FR 10) schedule with 20-second timeout. Thus, 10licks on the active spouts elicited a computer-driven injection of60 μl OG cue to the active spout and nicotine (30 μg/kg, free base)via the jugular vein. Using a syringe pump to deliver the OG cueensured the contingency of the OG cue with nicotine. The inactivespout did not contain any liquid and licks on the inactive spoutproduced no programmed consequence. No prior food or waterdeprivation, operant training or priming nicotine injections, whichcould potentially affect the motivation for operant behaviors, wasconducted. The patency of the jugular catheters was tested using afast-acting anesthetic Methohexital (0.2 ml, 10 mg/ml) at the end of12 SA sessions for each rat. Three female rats had blocked jugularcatheter. Data from these rats were excluded from the analysis. Ourprevious data showed that in the absence of the demonstrators,rats developed conditioned aversion to the appetitive OG cue, butSA was established with the presence of the demonstrators whoconsumed the same OG cue (Chen et al. 2011). Therefore, OG cue
was also provided to the demonstrator rats used in this experiment.Demonstrators were not subjected to nicotine infusion.
Context-induced reinstatement of drug seekingContext extinction and reinstatement procedures were performedafter the SA sessions. Context extinction chambers were differentfrom SA operant chamber in many aspects, including the absenceof the divider, flat floor, distinct audiovisual cues and novel odor(apple cinnamon) (Xie et al. 2012). There were two clean dry spoutsin the chamber and the number of licks was recorded but hadno programmed consequence. The self-administering rats wereplaced in the extinction environment for 1 h/day, until the lick onthe ‘active’ spout was reduced to less than 50 for two consecutivesessions.
Reinstatement of drug-seeking behavior was tested the day afterthe last context extinction session. During the 1-h reinstatementtests, rats were placed in the original nicotine SA chamber.The demonstrator rats were present and were provided withthe OG cue because social environment is one of the strongestcomponents of the complex contextual cues that induces cravingfor cigarettes (Garcia-Rodriguez et al. 2011). Licking on neither spouthad programmed consequence.
Lick microstructure analysisThe microstructure of the licks on the active spout was analyzed toexamine potential sex difference in the appetitiveness of the OG cue.We focused on the size of lick clusters because previous studiesshowed that it corresponded to the appetitiveness of the solutionbeing consumed (Boughter et al. 2007; Dwyer 2008) and was notaffected by satiety (De Araujo et al. 2006). We analyzed the data onlyfrom the first day of SA because the licks were less likely affected bynicotine. Licks with interlick intervals smaller than 0.5 second weretreated as one cluster. The size of the cluster was defined as thenumber of licks it contained. Clusters with less than two licks wereexcluded from the analysis.
Statistical analysisData were presented as mean ± SD. Number of licks, infusions andnose pokes were analyzed by repeated-measures analysis of variance(ANOVA) with spout and day treated as within-subject variables. Pairedt-tests were used to compare the number of licks on the two spoutsduring reinstatement. The t-tests were used to compare the sexdifferences on the means shown in Table S1, Supporting Information.Statistical significance was assigned when P < 0.05. All statisticalanalyses were conducted using R statistical package. Because ofthe large number of behavioral phenotypes we measured and thecorrelation between some of these phenotypes, we used principalcomponent analysis (PCA) to obtain orthogonal components, whichwere then tested as predictors for nicotine intake in a multipleregression analysis. The ‘principal’ function available from the psychpackage was used for the PCA. The components were rotatedusing the varimax function. The ‘lm’ and ‘step’ functions wereused for multiple regression. Procedures used are described in theResults section.
Results
Socially acquired nicotine SA
HS rats were subjected to daily 3-h nicotine SA sessions for12 days. Licking on the active spout triggered the concurrentdelivery of i.v. nicotine (30 μg/kg/infusion, free base) and60 μl OG cue controlled by a FR10 schedule. Each SA ratwas accompanied by a demonstrator rat randomly selectedbefore each session. The demonstrator had unrestrictedaccess to the OG cue but did not receive nicotine. Femaledemonstrators consumed significantly more OG cue than
Genes, Brain and Behavior (2013) 3
Wang et al.
10 20 50 100 500 2000 5000
10
20
50
200
500
2000
Nose pokes by dyads
Nicotine self−administrators
Dem
on
stra
tors
FemalesMales
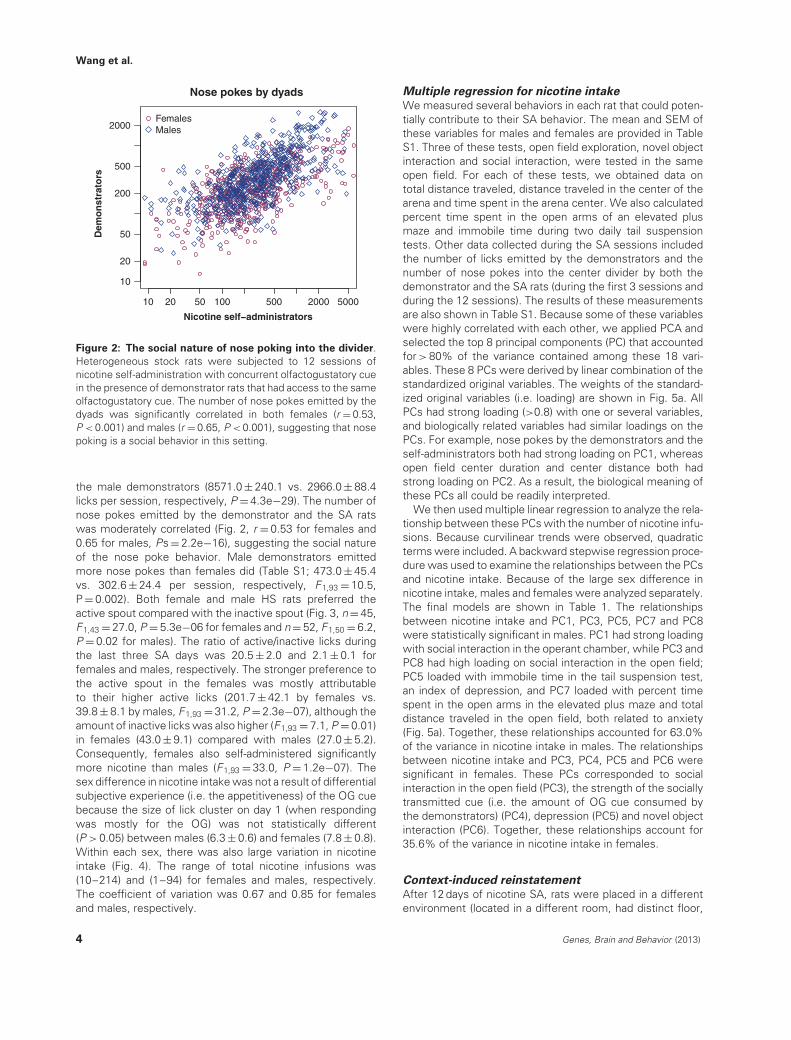
Figure 2: The social nature of nose poking into the divider.Heterogeneous stock rats were subjected to 12 sessions ofnicotine self-administration with concurrent olfactogustatory cuein the presence of demonstrator rats that had access to the sameolfactogustatory cue. The number of nose pokes emitted by thedyads was significantly correlated in both females (r = 0.53,P < 0.001) and males (r = 0.65, P < 0.001), suggesting that nosepoking is a social behavior in this setting.
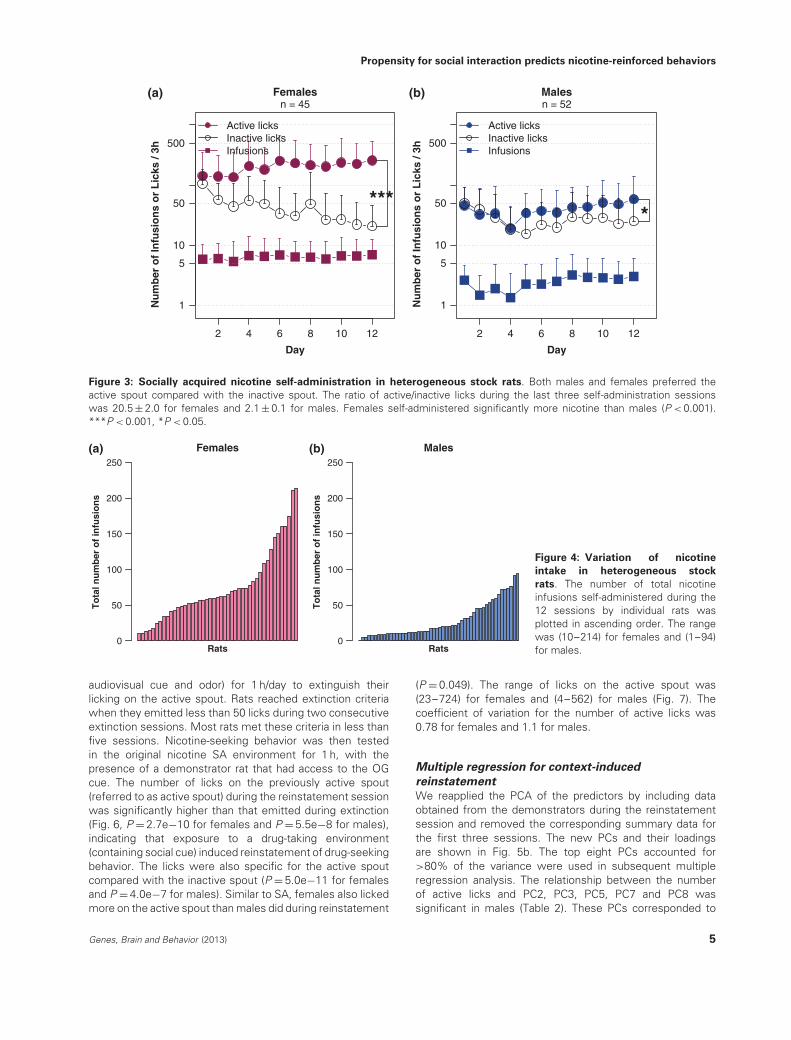
the male demonstrators (8571.0 ± 240.1 vs. 2966.0 ± 88.4licks per session, respectively, P = 4.3e−29). The number ofnose pokes emitted by the demonstrator and the SA ratswas moderately correlated (Fig. 2, r = 0.53 for females and0.65 for males, Ps = 2.2e−16), suggesting the social natureof the nose poke behavior. Male demonstrators emittedmore nose pokes than females did (Table S1; 473.0 ± 45.4vs. 302.6 ± 24.4 per session, respectively, F1,93 = 10.5,P = 0.002). Both female and male HS rats preferred theactive spout compared with the inactive spout (Fig. 3, n = 45,F1,43 = 27.0, P = 5.3e−06 for females and n = 52, F1,50 = 6.2,P = 0.02 for males). The ratio of active/inactive licks duringthe last three SA days was 20.5 ± 2.0 and 2.1 ± 0.1 forfemales and males, respectively. The stronger preference tothe active spout in the females was mostly attributableto their higher active licks (201.7 ± 42.1 by females vs.39.8 ± 8.1 by males, F1,93 = 31.2, P = 2.3e−07), although theamount of inactive licks was also higher (F1,93 = 7.1, P = 0.01)in females (43.0 ± 9.1) compared with males (27.0 ± 5.2).Consequently, females also self-administered significantlymore nicotine than males (F1,93 = 33.0, P = 1.2e−07). Thesex difference in nicotine intake was not a result of differentialsubjective experience (i.e. the appetitiveness) of the OG cuebecause the size of lick cluster on day 1 (when respondingwas mostly for the OG) was not statistically different(P > 0.05) between males (6.3 ± 0.6) and females (7.8 ± 0.8).Within each sex, there was also large variation in nicotineintake (Fig. 4). The range of total nicotine infusions was(10–214) and (1–94) for females and males, respectively.The coefficient of variation was 0.67 and 0.85 for femalesand males, respectively.
Multiple regression for nicotine intake
We measured several behaviors in each rat that could poten-tially contribute to their SA behavior. The mean and SEM ofthese variables for males and females are provided in TableS1. Three of these tests, open field exploration, novel objectinteraction and social interaction, were tested in the sameopen field. For each of these tests, we obtained data ontotal distance traveled, distance traveled in the center of thearena and time spent in the arena center. We also calculatedpercent time spent in the open arms of an elevated plusmaze and immobile time during two daily tail suspensiontests. Other data collected during the SA sessions includedthe number of licks emitted by the demonstrators and thenumber of nose pokes into the center divider by both thedemonstrator and the SA rats (during the first 3 sessions andduring the 12 sessions). The results of these measurementsare also shown in Table S1. Because some of these variableswere highly correlated with each other, we applied PCA andselected the top 8 principal components (PC) that accountedfor > 80% of the variance contained among these 18 vari-ables. These 8 PCs were derived by linear combination of thestandardized original variables. The weights of the standard-ized original variables (i.e. loading) are shown in Fig. 5a. AllPCs had strong loading (>0.8) with one or several variables,and biologically related variables had similar loadings on thePCs. For example, nose pokes by the demonstrators and theself-administrators both had strong loading on PC1, whereasopen field center duration and center distance both hadstrong loading on PC2. As a result, the biological meaning ofthese PCs all could be readily interpreted.
We then used multiple linear regression to analyze the rela-tionship between these PCs with the number of nicotine infu-sions. Because curvilinear trends were observed, quadraticterms were included. A backward stepwise regression proce-dure was used to examine the relationships between the PCsand nicotine intake. Because of the large sex difference innicotine intake, males and females were analyzed separately.The final models are shown in Table 1. The relationshipsbetween nicotine intake and PC1, PC3, PC5, PC7 and PC8were statistically significant in males. PC1 had strong loadingwith social interaction in the operant chamber, while PC3 andPC8 had high loading on social interaction in the open field;PC5 loaded with immobile time in the tail suspension test,an index of depression, and PC7 loaded with percent timespent in the open arms in the elevated plus maze and totaldistance traveled in the open field, both related to anxiety(Fig. 5a). Together, these relationships accounted for 63.0%of the variance in nicotine intake in males. The relationshipsbetween nicotine intake and PC3, PC4, PC5 and PC6 weresignificant in females. These PCs corresponded to socialinteraction in the open field (PC3), the strength of the sociallytransmitted cue (i.e. the amount of OG cue consumed bythe demonstrators) (PC4), depression (PC5) and novel objectinteraction (PC6). Together, these relationships account for35.6% of the variance in nicotine intake in females.
Context-induced reinstatement
After 12 days of nicotine SA, rats were placed in a differentenvironment (located in a different room, had distinct floor,
4 Genes, Brain and Behavior (2013)
Propensity for social interaction predicts nicotine-reinforced behaviors
2 4 6 8 10 12 2 4 6 8 10 12
1
5
10
50
500
1
5
10
50
500
Females
Day
Nu
mb
er o
f In
fusi
on
s o
r L
icks
/ 3h
n = 45
Active licksInactive licksInfusions
***
Males
Day
Nu
mb
er o
f In
fusi
on
s o
r L
icks
/ 3h
n = 52
Active licksInactive licksInfusions
*
(a) (b)
Figure 3: Socially acquired nicotine self-administration in heterogeneous stock rats. Both males and females preferred theactive spout compared with the inactive spout. The ratio of active/inactive licks during the last three self-administration sessionswas 20.5 ± 2.0 for females and 2.1 ± 0.1 for males. Females self-administered significantly more nicotine than males (P < 0.001).***P < 0.001, *P < 0.05.
Females
Rats
To
tal n
um
ber
of
infu
sio
ns
0
50
100
150
200
250
0
50
100
150
200
250
Males
Rats
To
tal n
um
ber
of
infu
sio
ns
(a) (b)
Figure 4: Variation of nicotine
intake in heterogeneous stock
rats. The number of total nicotineinfusions self-administered during the12 sessions by individual rats wasplotted in ascending order. The rangewas (10–214) for females and (1–94)for males.
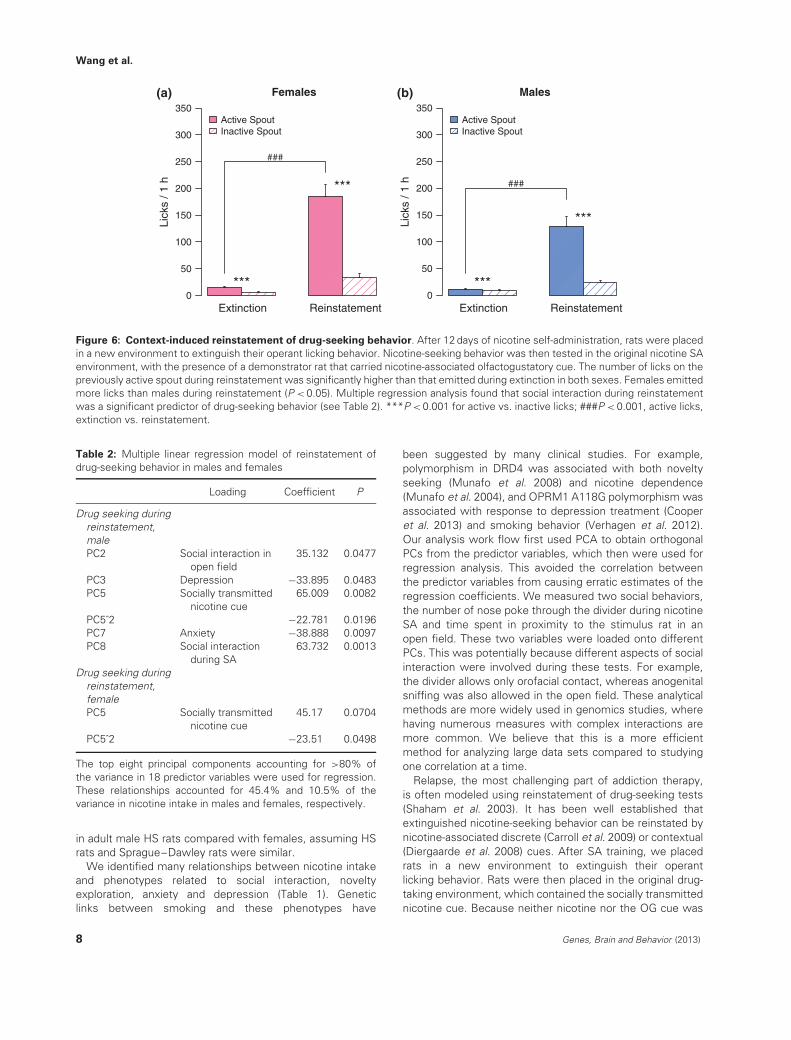
audiovisual cue and odor) for 1 h/day to extinguish theirlicking on the active spout. Rats reached extinction criteriawhen they emitted less than 50 licks during two consecutiveextinction sessions. Most rats met these criteria in less thanfive sessions. Nicotine-seeking behavior was then testedin the original nicotine SA environment for 1 h, with thepresence of a demonstrator rat that had access to the OGcue. The number of licks on the previously active spout(referred to as active spout) during the reinstatement sessionwas significantly higher than that emitted during extinction(Fig. 6, P = 2.7e−10 for females and P = 5.5e−8 for males),indicating that exposure to a drug-taking environment(containing social cue) induced reinstatement of drug-seekingbehavior. The licks were also specific for the active spoutcompared with the inactive spout (P = 5.0e−11 for femalesand P = 4.0e−7 for males). Similar to SA, females also lickedmore on the active spout than males did during reinstatement
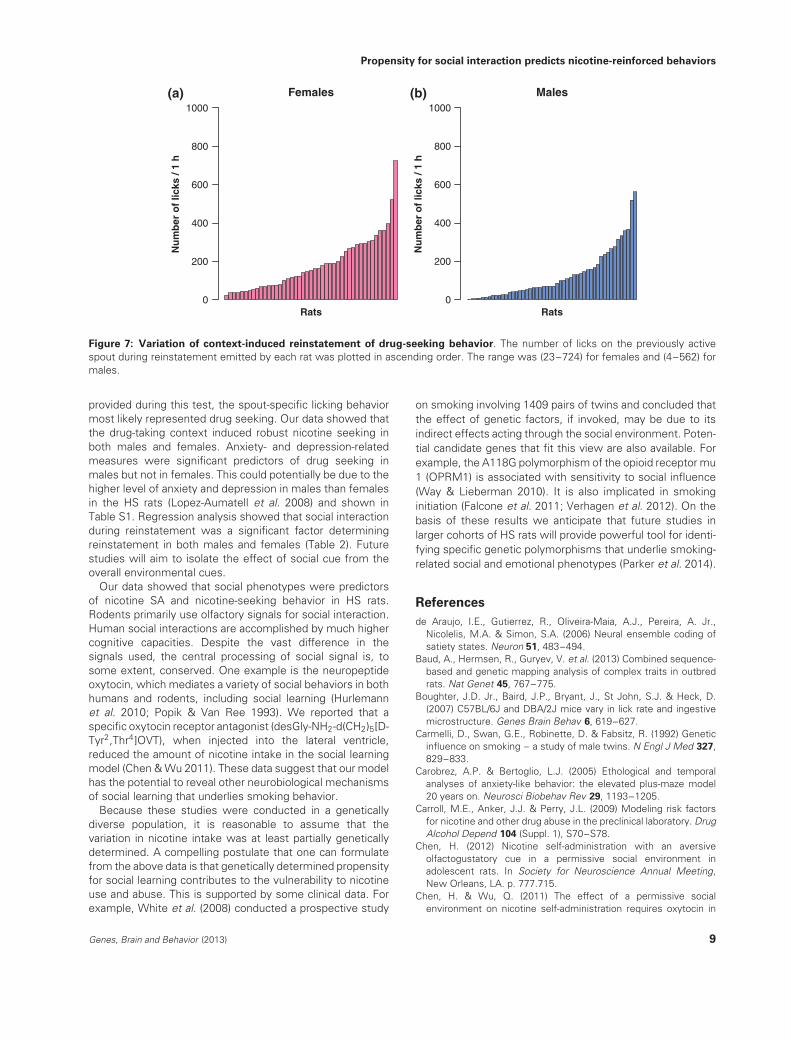
(P = 0.049). The range of licks on the active spout was(23–724) for females and (4–562) for males (Fig. 7). Thecoefficient of variation for the number of active licks was0.78 for females and 1.1 for males.
Multiple regression for context-induced
reinstatement
We reapplied the PCA of the predictors by including dataobtained from the demonstrators during the reinstatementsession and removed the corresponding summary data forthe first three sessions. The new PCs and their loadingsare shown in Fig. 5b. The top eight PCs accounted for>80% of the variance were used in subsequent multipleregression analysis. The relationship between the numberof active licks and PC2, PC3, PC5, PC7 and PC8 wassignificant in males (Table 2). These PCs corresponded to
Genes, Brain and Behavior (2013) 5

Wang et al.
0.84
0.81
−0.04
0.81
0.81
0.01
−0.24
−0.20
−0.12
−0.01
−0.01
0.08
0.00
0.16
0.12
−0.10
−0.19
0.02
0.23
−0.12
0.92
−0.01
−0.23
0.93
0.39
0.06
0.08
0.12
0.20
0.06
0.32
−0.02
−0.08
−0.12
0.03
−0.23
0.05
−0.06
0.17
0.10
−0.01
0.10
−0.01
−0.03
0.00
0.11
0.93
0.94
0.29
0.18
−0.09
−0.35
−0.13
0.08
0.09
0.14
−0.03
0.07
0.10
−0.10
0.24
−0.71
−0.04
−0.10
0.03
0.12
0.01
0.85
0.20
0.51
0.10
0.00
−0.15
0.08
0.08
−0.10
0.14
0.08
0.21
0.10
0.66
0.93
0.13
−0.01
0.27
−0.02
−0.04
−0.09
−0.08
0.04
0.09
−0.16
−0.12
−0.01
−0.16
−0.08
0.05
−0.09
0.00
−0.03
−0.03
−0.01
0.01
0.03
−0.10
−0.05
0.79
0.89
−0.03
0.32
−0.12
−0.12
0.24
−0.04
0.21
−0.29
−0.67
−0.05
−0.02
−0.06
0.08
0.00
0.82
0.08
−0.15
0.03
0.13
0.00
−0.03
−0.02
−0.02
0.08
−0.54
−0.01
−0.02
0.06
0.15
−0.03
0.73
−0.06
0.00
0.54
−0.22
0.16
EPM, open arm%
Lick, Demo, day 1−12
Lick, Demo, day 1−3
Nose poke, Demo, day 1−12
Nose Poke, Demo, day 1−3
Nose poke, SA rat, day 1−12
Nose Poke, SA rat, day 1−3
Novel Object, Center Distance
Novel Object, Center Duration
Novel Object, Total Distance
Open Field, Center Distance
Open Field, Center Duration
Open Field, Total Distance
Social Inter., Center Distance
Social Inter., Center Duration
Social Inter., Total Distance
Tail Suspension, day 1
Tail Suspension, day 2
PC1 PC2 PC3 PC4 PC5 PC6 PC7 PC8
−1.0
−0.5
0.0
0.5
1.0value
Variable loading for self−administration
0.08
−0.05
0.23
0.44
−0.21
0.03
−0.03
0.00
0.00
0.13
0.93
0.93
0.34
0.17
−0.06
−0.31
−0.12
0.08
0.12
0.29
−0.02
0.02
−0.08
−0.13
0.42
−0.73
−0.24
0.01
0.03
0.10
−0.03
0.76
0.54
0.37
0.11
0.05
0.12
−0.23
0.80
0.05
−0.14
0.85
0.45
0.08
0.16
−0.01
0.16
0.04
0.24
0.01
−0.29
−0.07
0.05
−0.23
−0.08
−0.04
0.18
−0.04
0.10
−0.02
0.12
0.16
0.83
0.90
0.13
0.01
0.24
0.01
−0.29
−0.13
−0.03
−0.01
0.02
−0.26
−0.16
−0.14
0.13
−0.01
−0.07
−0.11
0.15
−0.14
−0.03
0.04
−0.05
0.12
−0.40
−0.02
0.79
0.82
0.88
0.75
0.02
0.01
−0.05
−0.02
−0.33
−0.27
−0.09
−0.05
−0.01
0.05
0.10
0.14
−0.03
−0.13
−0.23
0.06
0.03
−0.09
−0.21
0.70
0.80
0.03
0.14
0.03
0.10
−0.02
−0.04
0.05
−0.11
−0.04
−0.05
0.15
0.01
0.01
0.03
−0.04
−0.05
0.14
−0.10
0.14
−0.41
−0.08
0.00
0.05
0.11
−0.08
0.69
0.06
0.05
0.69
−0.18
0.14
EPM, open arm%
Lick, Demo, day 1−12
Lick, Demo, reinstatement
Nose poke, Demo, day 1−12
Nose Poke, Demo, reinstatement
Nose poke, SA rat, day 1−12
Nose Poke, SA rat, reinstatement
Novel Object, Center Distance
Novel Object, Center Duration
Novel Object, Total Distance
Open Field, Center Distance
Open Field, Center Duration
Open Field, Total Distance
Social Inter., Center Distance
Social Inter., Center Duration
Social Inter., Total Distance
Tail Suspension, day 1
Tail Suspension, day 2
PC1 PC2 PC3 PC4 PC5 PC6 PC7 PC8
−1.0
−0.5
0.0
0.5
1.0value
Variable loading for reinstatement(a) (b)
Figure 5: Principle component analysis of the predictor variables. The top eight principal components accounting for approximately80% of the variance in the dataset were selected as predictors in subsequent multiple linear regression. The loading of the PCs withthe original variables were shown for (a) self-administration and (b) reinstatement. Most of the variables used for reinstatement werethe same as self-administration, with the exception that data for the first 3 days, critical for self-administration, were replaced by dataobtained during reinstatement. The size of the circles corresponded to the loading.
social interaction in open field (PC2), during SA (PC8) andreinstatement (PC5) as well as anxiety (PC7) and depression(PC3). These relationships account for 45.4% of the variancein the reinstatement data in males. In females, the number oflicks on active spout during reinstatement was only related toPC5, which had strong loading with social interaction duringreinstatement. This relationship accounted for 10.5% of thevariance in the reinstatement data.
Discussion
We studied socially acquired nicotine SA in outbred HS rats.Both males and females acquired nicotine SA; females self-administered more nicotine than males. After drug-seekingbehavior was extinguished in a new environment, the originaldrug-taking environment, which contained drug-related socialcues, triggered reinstatement of drug-seeking behavior.Large variation in both nicotine intake and reinstatementwas shown; these differences may be due to the geneticheterogeneity of the rats. Multiple regression analysis foundthat measures corresponding to social interaction were
significant predictors of nicotine intake and reinstatementof drug seeking; measures of depression (i.e. immobiletime) were predictors of nicotine intake in both males andfemales, anxiety (i.e. time spent in the open arms and totaldistance traveled in the open field) was a predictor onlyin males and response to novelty was a predictor only infemales. In males, both depression- and anxiety-relatedmeasures also predicted nicotine reinstatement. Largeportions (10.5–63.0%) of the variations in nicotine intakeand drug seeking were explained by these phenotypes.
The progression from initial experimentation withcigarettes to regular smoking is perplexing because mostfirst-time smokers experience little positive experience.Instead, negative effects such as nausea, dizziness, cough-ing and headache are much more common (Eissenberg &Balster 2000; Hahn et al. 1990). In spite of these negativeeffects, approximately 40% of teenagers who experimentwith cigarettes become regular smokers (Hofstetter et al.2007). One critical factor that has large influence on smokinginitiation is social environment (Hu et al. 2006; Schepis & Rao2005; White et al. 2008). In the socially acquired nicotine SAmodel, social transmission of the nicotine-associated odor
6 Genes, Brain and Behavior (2013)

Propensity for social interaction predicts nicotine-reinforced behaviors
Table 1: Multiple linear regression model of nicotine intake inmales and females
Loading Coefficient P
Total infusion, malePC1 Social interaction
during SA13.124 0.0003
PC1ˆ2 −4.126 0.0260PC3 Social interaction in
open field10.702 0.0001
PC3ˆ2 7.445 0.0012PC5 Depression −8.296 0.0016PC7 Anxiety −5.472 0.0154PC8 10.194 0.0007
Total infusion, femalePC3ˆ2 Social interaction in
open field−9.111 0.0397
PC4 Socially transmittednicotine cue
−100.307 0.0095
PC4ˆ2 47.238 0.0029PC5ˆ2 Depression −9.733 0.0200PC6ˆ2 Novelty −10.289 0.0352
The top eight principal components accounting for >80% ofthe variance in 18 predictor variables were used for regression.These relationships accounted for 63.0% and 35.6% of thevariance in nicotine intake in males and females, respectively.
cue enabled the transition from nicotine-conditioned tasteaversion to stable SA (Chen et al. 2011). This model thuscaptured two critical aspects of human smoking behavior:the initial aversive response to nicotine and the critical roleof the social environment and thus offered a unique oppor-tunity to identity the complex interaction between aversionand reward (both nicotine and social) in smoking behavior.
Genetic factors also strongly influence smoking behavior(Hall et al. 2002). This study used HS rats to investigatethe potential effect of genetic factors on socially acquirednicotine SA. The HS rats are the result of >60 generations ofintercrossing of eight inbred rat strains. The large number ofaccumulated recombinations has enabled many successfulgenetic studies (Baud et al. 2013; Solberg Woods et al.2010, 2012). The HS rats acquired stable nicotine SA in thepresence of a demonstrator rat carrying nicotine-associatedodor cue. This was shown by significant preference tothe active spout over the inactive spout (Fig. 3). We havepreviously shown that rats developed conditioned aversionto the OG cue in the absence of socially transmitted nicotine-associated odor cue. One indication of this aversion was thatSA rats avoided the active spout and thus more licks wereemitted on the inactive spout (Chen et al. 2011). Thus,significantly higher number of licks on the active spoutsuggested the reversal of conditioned nicotine aversion aftersocial learning and the establishment of nicotine SA.
Our previous data provided several lines of evidence thatthe active licks were specific for nicotine, rather than for theOG cue. For example, we reported an inverted U-shapednicotine dose-response curve in Sprague–Dawley rats (Chenet al. 2011). We also reported that the amount of nicotineintake was not affected by removing the OG cue in rats
that had already acquired nicotine SA, but mecamylamine,a nicotinic receptor antagonist, reduced nicotine SA (Chenet al. 2011). Further, replacing saccharin (sweet taste) withquinine (bitter) did not affect the amount of nicotine intake(Chen 2012). Last, we found that the number of activelicks increased by a factor of 2 when nicotine dose wasdecreased by a factor of 2 (Chen, unpublished observations);this observation was made using some of the same inbredstrains that are the founders for the HS. Taken together,these data strongly suggest that the active licks that weobserved were specific for nicotine.
Social interaction is also a reward, which can interact withdrug reward (Yates et al. 2013). Therefore, social interactionper se might support nicotine SA. For example, low dose ofnicotine interacted with social reward to enhance conditionedplace preference (Thiel et al. 2009). However, in our model,interaction with a demonstrator rat consuming the tastesolution without the odor cue was not sufficient to supportnicotine SA (Chen et al. 2011). This potentially could bedue to less reward that is provided by social interactionwhen a divider is in place than when the social interactionis not restricted by a divider. However, our previous datashowed that providing the demonstrator rats with nicotine-associated OG cue supported stable nicotine intake using thesame setup (Chen et al. 2011), indicating that social learning,rather than social reward, is critical for inducing nicotine SAin our model.
We found that female HS rats self-administered morenicotine than males. One potential source of sex differencein nicotine intake was the appetitiveness of the nicotine-associated sensory cue. Rats emit licks in clusters (i.e. boutsof licks with less than 0.5 second between licks). The numberof licks contained in these clusters reflects the subjectivevalue of the taste (Boughter et al. 2007; Dwyer 2008), withappetitive tastes associated with larger lick clusters. Weanalyzed the licks emitted by the HS rats on their first dayof SA and found that the size of the clusters produced bymales and females was not statistically different. Thus, theaffective value of the sensory cue was unlikely a sourceof differential nicotine intake. Another potential source ofsex difference was differential social learning. Similar tostudies reported by others (Valenstein et al. 1967), femaledemonstrators consumed more saccharine than males did.As a result, stronger social cues could have been presentedto the female SA rats. On the other hand, male demonstratorsinitiated more social interaction by nose poking into thedivider and spent more time in the center zone of the openfield, where the stimulus rat was placed (Table S1). The socialnature of these nose pokes was shown by the correlationbetween the demonstrators and the SA rats (Fig. 2). Thiscould potentially enhance social learning in males. Therefore,it was possible that different aspects of the behavior of thedemonstrator rats (i.e. consuming the OG cue vs. initiatingsocial contact) could contribute to the sex difference. Lastly,the interaction between age and sex could also contributeto the sex difference in nicotine intake. We have previouslyshown in Sprague–Dawley adolescent rats that males andfemales self-administered same amount of nicotine (Chenet al. 2011). It is thus possible that the sex difference wefound in HS rats was due to reduced social learning capability
Genes, Brain and Behavior (2013) 7
Wang et al.
Extinction Reinstatement
Females
Lick
s / 1
h
0
50
100
150
200
250
300
350
0
50
100
150
200
250
300
350Active SpoutInactive Spout
***
***
###
Extinction Reinstatement
Males
Lick
s / 1
h
Active SpoutInactive Spout
***
***
###
(a) (b)
Figure 6: Context-induced reinstatement of drug-seeking behavior. After 12 days of nicotine self-administration, rats were placedin a new environment to extinguish their operant licking behavior. Nicotine-seeking behavior was then tested in the original nicotine SAenvironment, with the presence of a demonstrator rat that carried nicotine-associated olfactogustatory cue. The number of licks on thepreviously active spout during reinstatement was significantly higher than that emitted during extinction in both sexes. Females emittedmore licks than males during reinstatement (P < 0.05). Multiple regression analysis found that social interaction during reinstatementwas a significant predictor of drug-seeking behavior (see Table 2). ***P < 0.001 for active vs. inactive licks; ###P < 0.001, active licks,extinction vs. reinstatement.
Table 2: Multiple linear regression model of reinstatement ofdrug-seeking behavior in males and females
Loading Coefficient P
Drug seeking duringreinstatement,malePC2 Social interaction in
open field35.132 0.0477
PC3 Depression −33.895 0.0483PC5 Socially transmitted
nicotine cue65.009 0.0082
PC5ˆ2 −22.781 0.0196PC7 Anxiety −38.888 0.0097PC8 Social interaction
during SA63.732 0.0013
Drug seeking duringreinstatement,femalePC5 Socially transmitted
nicotine cue45.17 0.0704
PC5ˆ2 −23.51 0.0498
The top eight principal components accounting for >80% ofthe variance in 18 predictor variables were used for regression.These relationships accounted for 45.4% and 10.5% of thevariance in nicotine intake in males and females, respectively.
in adult male HS rats compared with females, assuming HSrats and Sprague–Dawley rats were similar.
We identified many relationships between nicotine intakeand phenotypes related to social interaction, noveltyexploration, anxiety and depression (Table 1). Geneticlinks between smoking and these phenotypes have
been suggested by many clinical studies. For example,polymorphism in DRD4 was associated with both noveltyseeking (Munafo et al. 2008) and nicotine dependence(Munafo et al. 2004), and OPRM1 A118G polymorphism wasassociated with response to depression treatment (Cooperet al. 2013) and smoking behavior (Verhagen et al. 2012).Our analysis work flow first used PCA to obtain orthogonalPCs from the predictor variables, which then were used forregression analysis. This avoided the correlation betweenthe predictor variables from causing erratic estimates of theregression coefficients. We measured two social behaviors,the number of nose poke through the divider during nicotineSA and time spent in proximity to the stimulus rat in anopen field. These two variables were loaded onto differentPCs. This was potentially because different aspects of socialinteraction were involved during these tests. For example,the divider allows only orofacial contact, whereas anogenitalsniffing was also allowed in the open field. These analyticalmethods are more widely used in genomics studies, wherehaving numerous measures with complex interactions aremore common. We believe that this is a more efficientmethod for analyzing large data sets compared to studyingone correlation at a time.
Relapse, the most challenging part of addiction therapy,is often modeled using reinstatement of drug-seeking tests(Shaham et al. 2003). It has been well established thatextinguished nicotine-seeking behavior can be reinstated bynicotine-associated discrete (Carroll et al. 2009) or contextual(Diergaarde et al. 2008) cues. After SA training, we placedrats in a new environment to extinguish their operantlicking behavior. Rats were then placed in the original drug-taking environment, which contained the socially transmittednicotine cue. Because neither nicotine nor the OG cue was
8 Genes, Brain and Behavior (2013)
Propensity for social interaction predicts nicotine-reinforced behaviors
Females(a)
Rats
Nu
mb
er o
f lic
ks /
1 h
0
200
400
600
800
1000
0
200
400
600
800
1000
Males(b)
Rats
Nu
mb
er o
f lic
ks /
1 h
Figure 7: Variation of context-induced reinstatement of drug-seeking behavior. The number of licks on the previously activespout during reinstatement emitted by each rat was plotted in ascending order. The range was (23–724) for females and (4–562) formales.
provided during this test, the spout-specific licking behaviormost likely represented drug seeking. Our data showed thatthe drug-taking context induced robust nicotine seeking inboth males and females. Anxiety- and depression-relatedmeasures were significant predictors of drug seeking inmales but not in females. This could potentially be due to thehigher level of anxiety and depression in males than femalesin the HS rats (Lopez-Aumatell et al. 2008) and shown inTable S1. Regression analysis showed that social interactionduring reinstatement was a significant factor determiningreinstatement in both males and females (Table 2). Futurestudies will aim to isolate the effect of social cue from theoverall environmental cues.
Our data showed that social phenotypes were predictorsof nicotine SA and nicotine-seeking behavior in HS rats.Rodents primarily use olfactory signals for social interaction.Human social interactions are accomplished by much highercognitive capacities. Despite the vast difference in thesignals used, the central processing of social signal is, tosome extent, conserved. One example is the neuropeptideoxytocin, which mediates a variety of social behaviors in bothhumans and rodents, including social learning (Hurlemannet al. 2010; Popik & Van Ree 1993). We reported that aspecific oxytocin receptor antagonist (desGly-NH2-d(CH2)5[D-Tyr2,Thr4]OVT), when injected into the lateral ventricle,reduced the amount of nicotine intake in the social learningmodel (Chen & Wu 2011). These data suggest that our modelhas the potential to reveal other neurobiological mechanismsof social learning that underlies smoking behavior.
Because these studies were conducted in a geneticallydiverse population, it is reasonable to assume that thevariation in nicotine intake was at least partially geneticallydetermined. A compelling postulate that one can formulatefrom the above data is that genetically determined propensityfor social learning contributes to the vulnerability to nicotineuse and abuse. This is supported by some clinical data. Forexample, White et al. (2008) conducted a prospective study
on smoking involving 1409 pairs of twins and concluded thatthe effect of genetic factors, if invoked, may be due to itsindirect effects acting through the social environment. Poten-tial candidate genes that fit this view are also available. Forexample, the A118G polymorphism of the opioid receptor mu1 (OPRM1) is associated with sensitivity to social influence(Way & Lieberman 2010). It is also implicated in smokinginitiation (Falcone et al. 2011; Verhagen et al. 2012). On thebasis of these results we anticipate that future studies inlarger cohorts of HS rats will provide powerful tool for identi-fying specific genetic polymorphisms that underlie smoking-related social and emotional phenotypes (Parker et al. 2014).
References
de Araujo, I.E., Gutierrez, R., Oliveira-Maia, A.J., Pereira, A. Jr.,Nicolelis, M.A. & Simon, S.A. (2006) Neural ensemble coding ofsatiety states. Neuron 51, 483–494.
Baud, A., Hermsen, R., Guryev, V. et al. (2013) Combined sequence-based and genetic mapping analysis of complex traits in outbredrats. Nat Genet 45, 767–775.
Boughter, J.D. Jr., Baird, J.P., Bryant, J., St John, S.J. & Heck, D.(2007) C57BL/6J and DBA/2J mice vary in lick rate and ingestivemicrostructure. Genes Brain Behav 6, 619–627.
Carmelli, D., Swan, G.E., Robinette, D. & Fabsitz, R. (1992) Geneticinfluence on smoking – a study of male twins. N Engl J Med 327,829–833.
Carobrez, A.P. & Bertoglio, L.J. (2005) Ethological and temporalanalyses of anxiety-like behavior: the elevated plus-maze model20 years on. Neurosci Biobehav Rev 29, 1193–1205.
Carroll, M.E., Anker, J.J. & Perry, J.L. (2009) Modeling risk factorsfor nicotine and other drug abuse in the preclinical laboratory. DrugAlcohol Depend 104 (Suppl. 1), S70–S78.
Chen, H. (2012) Nicotine self-administration with an aversiveolfactogustatory cue in a permissive social environment inadolescent rats. In Society for Neuroscience Annual Meeting,New Orleans, LA. p. 777.715.
Chen, H. & Wu, Q. (2011) The effect of a permissive socialenvironment on nicotine self-administration requires oxytocin in
Genes, Brain and Behavior (2013) 9

Wang et al.
adolescent rats. In Society for Neuroscience Annual Meeting,Washington, DC. p. 686.619.
Chen, H., Sharp, B.M., Matta, S.G. & Wu, Q. (2011) Social interactionpromotes nicotine self-administration with olfactogustatory cuesin adolescent rats. Neuropsychopharmacology 36, 2629–2638.
Chen, H., Hiler, K.A., Tolley, E.A., Matta, S.G. & Sharp, B.M. (2012a)Genetic factors control nicotine self-administration in isogenicadolescent rat strains. PLoS One 7, e44234.
Chen, L.S., Baker, T.B., Piper, M.E., Breslau, N., Cannon, D.S.,Doheny, K.F., Gogarten, S.M., Johnson, E.O., Saccone, N.L.,Wang, J.C., Weiss, R.B., Goate, A.M. & Bierut, L.J. (2012b)Interplay of genetic risk factors (CHRNA5-CHRNA3-CHRNB4)and cessation treatments in smoking cessation success. Am JPsychiatry 169, 735–742.
Cooper, A.J., Rickels, K. & Lohoff, F.W. (2013) Association analysisbetween the A118G polymorphism in the OPRM1 gene andtreatment response to venlafaxine XR in generalized anxietydisorder. Hum Psychopharmacol 28, 258–262.
Diergaarde, L., de Vries, W., Raaso, H., Schoffelmeer, A.N. & DeVries, T.J. (2008) Contextual renewal of nicotine seeking in ratsand its suppression by the cannabinoid-1 receptor antagonistRimonabant (SR141716A). Neuropharmacology 55, 712–716.
Dwyer, D.M. (2008) Microstructural analysis of conditioned andunconditioned responses to maltodextrin. Learn Behav 36,149–158.
Eissenberg, T. & Balster, R.L. (2000) Initial tobacco use episodesin children and adolescents: current knowledge, future directions.Drug Alcohol Depend 59 (Suppl. 1), S41–S60.
Falcone, M., Jepson, C., Sanborn, P., Cappella, J.N., Lerman, C. &Strasser, A.A. (2011) Association of BDNF and COMT genotypeswith cognitive processing of anti-smoking PSAs. Genes BrainBehav 10, 862–867.
Garcia-Rodriguez, O., Ferrer-Garcia, M., Pericot-Valverde, I.,Gutierrez-Maldonado, J., Secades-Villa, R. & Carballo, J.L. (2011)Identifying specific cues and contexts related to smoking cravingfor the development of effective virtual environments. Cyberpsy-chol Behav Soc Netw 14, 91–97.
Green, M.R., Barnes, B. & McCormick, C.M. (2013) Social instabilitystress in adolescence increases anxiety and reduces socialinteractions in adulthood in male Long-Evans rats. Dev Psychobiol55, 849–859.
Greenlund, K.J., Johnson, C.C., Webber, L.S. & Berenson, G.S.(1997) Cigarette smoking attitudes and first use among third-through sixth-grade students: the Bogalusa Heart Study. Am JPublic Health 87, 1345–1348.
Hahn, G., Charlin, V.L., Sussman, S., Dent, C.W., Manzi, J., Stacy,A.W., Flay, B., Hansen, W.B. & Burton, D. (1990) Adolescents’first and most recent use situations of smokeless tobacco andcigarettes: similarities and differences. Addict Behav 15, 439–448.
Hall, W., Madden, P. & Lynskey, M. (2002) The genetics of tobaccouse: methods, findings and policy implications. Tob Control 11,119–124.
Hansen, C. & Spuhler, K. (1984) Development of the NationalInstitutes of Health genetically heterogeneous rat stock. AlcoholClin Exp Res 8, 477–479.
Hinojosa, F.R., Spricigo, L. Jr., Izidio, G.S., Bruske, G.R., Lopes, D.M.& Ramos, A. (2006) Evaluation of two genetic animal models inbehavioral tests of anxiety and depression. Behav Brain Res 168,127–136.
Hofstetter, C.R., Hovell, M.F., Jung, K.R., Raman, R., Irvin, V. &Ni, R. (2007) The first puff: forces in smoking initiation amongCalifornians of Korean descent. Nicotine Tob Res 9, 1277–1286.
Hu, M.C., Davies, M. & Kandel, D.B. (2006) Epidemiology andcorrelates of daily smoking and nicotine dependence among youngadults in the United States. Am J Public Health 96, 299–308.
Hurlemann, R., Patin, A., Onur, O.A., Cohen, M.X., Baumgartner,T., Metzler, S., Dziobek, I., Gallinat, J., Wagner, M., Maier, W.& Kendrick, K.M. (2010) Oxytocin enhances amygdala-dependent,socially reinforced learning and emotional empathy in humans. JNeurosci 30, 4999–5007.
Le Foll, B. & Goldberg, S.R. (2006) Nicotine as a typical drug of abusein experimental animals and humans. Psychopharmacology (Berl)184, 367–381.
Li, M.D., Cheng, R., Ma, J.Z. & Swan, G.E. (2003) A meta-analysis ofestimated genetic and environmental effects on smoking behaviorin male and female adult twins. Addiction 98, 23–31.
Lopez-Aumatell, R., Guitart-Masip, M., Vicens-Costa, E., Gimenez-Llort, L., Valdar, W., Johannesson, M., Flint, J., Tobena, A. &Fernandez-Teruel, A. (2008) Fearfulness in a large N/Nih geneticallyheterogeneous rat stock: differential profiles of timidity anddefensive flight in males and females. Behav Brain Res 188,41–55.
Mathers, C.D. & Loncar, D. (2006) Projections of global mortality andburden of disease from 2002 to 2030. PLoS Med 3, e442.
Matta, S.G., Balfour, D.J., Benowitz, N.L. et al. (2007) Guidelines onnicotine dose selection for in vivo research. Psychopharmacology(Berl) 190, 269–319.
Mott, R., Talbot, C.J., Turri, M.G., Collins, A.C. & Flint, J. (2000) Amethod for fine mapping quantitative trait loci in outbred animalstocks. Proc Natl Acad Sci U S A 97, 12649–12654.
Munafo, M., Clark, T., Johnstone, E., Murphy, M. & Walton, R. (2004)The genetic basis for smoking behavior: a systematic review andmeta-analysis. Nicotine Tob Res 6, 583–597.
Munafo, M.R., Yalcin, B., Willis-Owen, S.A. & Flint, J. (2008)Association of the dopamine D4 receptor (DRD4) gene andapproach-related personality traits: meta-analysis and new data.Biol Psychiatry 63, 197–206.
O’Dell, L.E. & Khroyan, T.V. (2009) Rodent models of nicotine reward:what do they tell us about tobacco abuse in humans? PharmacolBiochem Behav 91, 481–488.
Parker, C.C., Chen, H., Flagel, S.B., Geurts, A.M., Richards, J.B.,Robinson, T.E., Solberg Woods, L.C. & Palmer, A.A. (2014) Ratsare the smart choice: rationale for a renewed focus on rats inbehavioral genetics. Neuropharmacology 76 Pt B, 250–258.
Pergadia, M.L., Heath, A.C., Martin, N.G. & Madden, P.A. (2006)Genetic analyses of DSM-IV nicotine withdrawal in adult twins.Psychol Med 36, 963–972.
Popik, P. & Van Ree, J.M. (1993) Social transmission of flavoredtea preferences: facilitation by a vasopressin analog and oxytocin.Behav Neural Biol 59, 63–68.
Saccone, N.L., Wang, J.C., Breslau, N., Johnson, E.O., Hatsukami,D., Saccone, S.F., Grucza, R.A., Sun, L., Duan, W., Budde, J.,Culverhouse, R.C., Fox, L., Hinrichs, A.L., Steinbach, J.H., Wu,M., Rice, J.P., Goate, A.M. & Bierut, L.J. (2009) The CHRNA5-CHRNA3-CHRNB4 nicotinic receptor subunit gene cluster affectsrisk for nicotine dependence in African-Americans and in European-Americans. Cancer Res 69, 6848–6856.
Schepis, T.S. & Rao, U. (2005) Epidemiology and etiology ofadolescent smoking. Curr Opin Pediatr 17, 607–612.
Shaham, Y., Shalev, U., Lu, L., De Wit, H. & Stewart, J. (2003) Thereinstatement model of drug relapse: history, methodology andmajor findings. Psychopharmacology (Berl) 168, 3–20.
Siedlinski, M., Cho, M.H., Bakke, P., Gulsvik, A., Lomas, D.A.,Anderson, W., Kong, X., Rennard, S.I., Beaty, T.H., Hokanson, J.E.,Crapo, J.D. & Silverman, E.K. (2011) Genome-wide associationstudy of smoking behaviours in patients with COPD. Thorax 66,894–902.
Solberg Woods, L.C., Holl, K., Tschannen, M. & Valdar, W. (2010)Fine-mapping a locus for glucose tolerance using heterogeneousstock rats. Physiol Genomics 41, 102–108.
Solberg Woods, L.C., Holl, K.L., Oreper, D., Xie, Y., Tsaih, S.W. &Valdar, W. (2012) Fine-mapping diabetes-related traits, includinginsulin resistance, in heterogeneous stock rats. Physiol Genomics44, 1013–1026.
Thiel, K.J., Sanabria, F. & Neisewander, J.L. (2009) Synergisticinteraction between nicotine and social rewards in adolescentmale rats. Psychopharmacology (Berl) 204, 391–402.
Thorgeirsson, T.E., Geller, F., Sulem, P. et al. (2008) A variantassociated with nicotine dependence, lung cancer and peripheralarterial disease. Nature 452, 638–642.
10 Genes, Brain and Behavior (2013)

Propensity for social interaction predicts nicotine-reinforced behaviors
Thorgeirsson, T.E., Gudbjartsson, D.F., Surakka, I. et al. (2010)Sequence variants at CHRNB3-CHRNA6 and CYP2A6 affectsmoking behavior. Nat Genet 42, 448–453.
Tobacco and Genetics Consortium (2010) Genome-wide meta-analyses identify multiple loci associated with smoking behavior.Nat Genet 42, 441–447.
Valenstein, E.S., Cox, V.C. & Kakolewski, J.W. (1967) Polydipsiaelicited by the synergistic action of a saccharin and glucosesolution. Science 157, 552–554.
Verhagen, M., Kleinjan, M. & Engels, R.C. (2012) A systematicreview of the A118G (Asn40Asp) variant of OPRM1 in relation tosmoking initiation, nicotine dependence and smoking cessation.Pharmacogenomics 13, 917–933.
Vetter-O’Hagen, C.S. & Spear, L.P. (2012) The effects of gonadec-tomy on sex- and age-typical responses to novelty and ethanol-induced social inhibition in adult male and female Sprague-Dawleyrats. Behav Brain Res 227, 224–232.
Way, B.M. & Lieberman, M.D. (2010) Is there a genetic contributionto cultural differences? Collectivism, individualism and geneticmarkers of social sensitivity. Soc Cogn Affect Neurosci 5,203–211.
White, V.M., Byrnes, G.B., Webster, B. & Hopper, J.L. (2008) Doessmoking among friends explain apparent genetic effects on currentsmoking in adolescence and young adulthood? Br J Cancer 98,1475–1481.
Xie, X., Arguello, A.A., Reittinger, A.M., Wells, A.M. & Fuchs, R.A.(2012) Role of nicotinic acetylcholine receptors in the effects ofcocaine-paired contextual stimuli on impulsive decision making inrats. Psychopharmacology (Berl) 223, 271–279.
Yates, J.R., Beckmann, J.S., Meyer, A.C. & Bardo, M.T. (2013)Concurrent choice for social interaction and amphetamine usingconditioned place preference in rats: effects of age and housingcondition. Drug Alcohol Depend 129, 240–246.
Acknowledgments
We thank Ms. Kaitlyn Hill for analyzing the lick clusterdata. Funding was provided by NIDA grant DA-026894 (H.C.),DA021336 (A.A.P.), and the Chicago Biomedical Consortiumwith support from the Searle Funds at the Chicago CommunityTrust and the Conte Center for Computational NeuropsychiatricGenomics (NIH P50MH94267). The maintenance of the HS ratcolony is supported on grant DK-088975 (L.C.S.-W.).
Supporting Information
Additional supporting information may be found in the onlineversion of this article at the publisher’s web-site:
Behavioral measures potentially contributing to nicotineintake and drug-seeking behavior. Open field exploration,novel object interaction, social interaction, elevated plusmaze and tail suspension tests were conducted on eachrat before self-administration (SA). Data collected during theSA sessions included the number of licks emitted by thedemonstrators and the number of nose pokes into the centerdivider by both the demonstrator and SA rats (for the first 3sessions, for the 12 sessions and during reinstatement). Alldata were expressed as mean ± SEM. Statistical significanceof the sex difference was calculated using independentt-tests or repeated measures ANOVA (indicated by #). Pvalues smaller than 0.05 were highlighted in bold anditalicized font.
Genes, Brain and Behavior (2013) 11














![(S)-(-)-[11C]Nicotine Binding Assessed by PET](https://img.pdfslide.net/doc/110x75/635feb4eac0cd8fcb10e64d4/s-11cnicotine-binding-assessed-by-pet.jpg)














