Embed Size (px)
Citation preview

Brain Research, 485 (1989) 225-235 225 Elsevier
BRE 14413
Protein associated with the sensory cell layer of the rainbow trout saccular macula
Dennis G. Drescher 1'2, Khalid M. Khan 1, Richard L. Arden 1, Marian J. Drescher 1, Thomas P. Kerr 1"3 and James S. Hatfield 4
Laboratory of Bio-otology, Departments of lOtolaryngology, 2Biochemistry and 3Anatomy and Cell Biology, Wayne State University School of Medicine, Detroit, MI 48201 (U.S.A.) and 4Veterans Administration Medical Center,
Allen Park, MI 48101 (U.S.A.)
(Accepted 20 September 1988)
Key words: Saccular macula; Hair cell; Electrophoresis; Marker protein
A protein has been detected that is associated with the saccular hair cell layer of the rainbow trout, Salmo gairdnerii R. By one- and two-dimensional SDS polyacrylamide gel electrophoresis, the molecular weight and isoelectric point of this protein are estimated to be 13.6 and 8.8 kDa, respectively. The 13.6 kDa protein cannot be detected electrophoretically in brain, gill, liver, and fractions containing the basal lamina, non-sensory epithelium, and saccular nerve. This protein does not bind antibodies to bovine myelin basic protein, while trout myelin basic proteins in the same molecular weight range do. In addition, the protein does not bind concanavalin A or react with the periodic acid-Schiff reagent. The 13.6 kDa band represents about 1% of the total protein in saccular sensory epithelium, and may be a marker protein for the hair cell layer.
INTRODUCTION
The morphological and functional diversity of
living cells is manifested in their characteristic
complement of gene-directed proteins. For acousti- colateralis organs, several proteins have been de-
scribed that may be specific to mechanoreceptor systems. Thalmann et al. 24 reported the presence of
two proteins in guinea pig organ of Corti that were
absent from vestibular saccular macula and other inner ear tissues. Oberholtzer et al. 16 described a
protein specific to the basilar papilla in chick
cochlea, correlated with onset of sensory function. A neuromast associated protein has been detected in the lateral line of Xenopus laevis by Drescher 5. Wenthold and McGarvey 26 reported the presence of
a glycoprotein apparently specific to the guinea pig auditory nerve. The functions of the above proteins
are presently unknown, and none have yet been
isolated in pure form for analysis.
One of the challenges of acousticolateralis bio-
chemistry is the procurement of sufficient tissue for
biochemical studies. Fortunately, teleost fish provide
a source of a large number of hair cells of a single morphological type 1s-2°. In the present work, the
sensory epithelial sheet of the trout saccular macula,
containing 30,000-40,000 hair cells 7, was separated
from the basal lamina and other underlying struc-
tures with concomitant disruption of supporting
cells. This hair cell preparation was found to be suitable for analysis of proteins by one- and two- dimensional polyacrylamide gel electrophoresis 12'17
and other biochemical investigations on the mecha-
nosensory organs. We present here results for the identification of a protein associated with the puri- fied hair cell fraction. A preliminary report has been presented 6.
Correspondence: D.G. Drescher, Laboratory of Bio-otology, Wayne State University School of Medicine, 540 East Canfield Avenue, Detroit, MI 48201, U.S.A.
0006-8993/89/$03.50 t~) 1989 Elsevier Science Publishers B.V. (Biomedical Division)

226
MATERIALS AND METHODS
Materials Reagents for electrophoresis were of analytical
reagent grade or better. Protein standards (low molecular weight calibration kit) were obtained from Pharmacia, Piscataway, NJ. Ampholytes were from LKB, Bromma, Sweden. Phenylmethanesulfonyl fluoride (PMSF) was from Sigma, St. Louis, MO, and leupeptin and pepstatin were from Boehringer Mannheim, Indianapolis, IN. Nitrocellulose paper (0.45/~m pore size) was from Schleicher and Schuell, Keene, NH. Goat anti-rabbit IgG and rabbit per- oxisase-antiperoxidase complex were from Pel Freeze Immunochemicals, Rogers, AR. Biotinylated concanavalin A and avidin-biotin-peroxidase com- plex were from Vector, Burlingame, CA. 'Dissecting saline' was formulated as described previously 7. Rat myelin and antibodies to bovine myelin basic pro- teins (MBPs) 3, raised in rabbits, were generously provided by Dr. Joyce Benjamins.
Preparation of samples for electrophoresis Rainbow trout (Salmo gairdnerii R.), obtained
from Imlay City Fish Farms, Imlay City, MI, were maintained as previously described 7. All procedures were performed at 0-2 °C. The cranial cavity was opened dorsally and the membranous endolymphatic sacs were removed. Endolymphatic sacs, containing the saccular and lagenar maculae and their innerva- tion, are found in cartilagenous depressions in the floor of the cranium of the teleosts 7'18. Each sac measures approximately 3 x 5 mm, and the saccular epithelium measures about 1 x 4 mm. Saccular and lagenar nerves on the medial perilymphatic surface were gently teased away, and the sacs opened to remove the saccular and lagenar otoliths. With the sensory surface of the saccular macula facing up- wards, the hair cell layer (HC) of the macula was removed above the basement membrane, using a fine insect pin. The overlying otolithic membrane was peeled off. The basal lamina and underlying connective tissue, with some fragments of supporting cells and nerve fibers ('basal lamina fraction', BL), was excised about its perimeter. A non-sensory saccular wall fraction (NS) and a saccular nerve fraction (N), the latter including efferent and affer- ent fibers and associated Schwann cells, were also
dissected. Ten tissue samples for each of the frac- tions were pooled and homogenized in 100 ¢tl SDS sample buffer (62.5 mM Tris chloride, 2.3% SDS, 12.5% glycerol (wt/v), and 5% 2-mercaptoethanol (v/v), pH 7.0) for 5 min on ice. Sections of trout brain, gill, and liver (1 mm 3) were also homogenized in the SDS sample buffer. Each sample was heated at 100 °C for 3 min and allowed to return to room temperature. Bromphenol blue and sucrose were added to yield a final concentration of 0.074% and 1.85%, respectively, and the sample was centrifuged at 1450 g for 20 min. Supernatants were stored at -20 °C until analysis.
For some experiments, hair cell layers from ten maculae were pooled in 25 B1 of dissecting saline, an equal volume of sample buffer without SDS was added, and the tissues were homogenized. The homogenate was centrifuged at 10,000 g for 1 h, the supernatant was carefully withdrawn, and an equal volume of 4.6% SDS solution was added to the supernatant to provide a concentration of buffer components the same as that of the SDS sample buffer. The pellet (about 1 ktl in volume) was resuspended in 100 ¢tl of SDS sample buffer, and the supernatant and pellet samples were heated and prepared for electrophoresis.
For other experiments, the saccular tissue frac- tions (HC, BL, NS, and N) were homogenized in SDS sample buffer containing HEPES (in place of Tris), as well as the protease inhibitors leupeptin (0.5 /~g/ml), pepstatin (0.7 ~g/ml), and PMSF (200 mM).
Electrophoresis and electroblotting Polyacrylamide slabs (80 x 65 x 1.5 mm) were
prepared for one-dimensional electrophoresis utiliz- ing a miniature vertical slab gel unit (SE 250, Hoefer, San Francisco, CA). The separating gel was poured as a 10-20% exponential gradient (top to bottom), and a 4% acrylamide stacking gel was added. We chose the exponential gradient to obtain relatively distinct bands in the lower molecular weight region and to allow a comparison of molec- ular weights in this region. Electrophoresis was performed at 18 mA/gel, and proteins were visual- ized with a Kodavue electrophoresis visualization kit (Eastman Kodak Company, Rochester, NY) 11.
Two-dimensional electrophoresis was carried out according to the method of O'Farrel117, with minor

modifications. Isoelectric focusing was performed in cylindrical tubes (1.5 mm inner diameter) in the presence of 9 M urea, with a pH gradient of 3.5-10. We ran pI standard cylindrical gels for each experi- ment to establish pI axes for subsequent two- dimensional gels. The cylindrical gels were focused at 400 V for 16 h, followed by 800 V for 1 h. The gels were then equilibrated for 1 h in the SDS sample buffer in which the 2-mercaptoethanol concentration was reduced to 0.5%. Electrophoresis in the second dimension was carried out as described above.
Densitometric measurement of protein standard bands (phosphorylase B, 94 kDa, 64 ng; ovalbumin, 43 kDa, 83 ng; carbonic anhydrase, 30 kDa, 83 ng; a-lactalbumin, 14.4 kDa, 121 ng) was used to provide an estimate of the amount of protein in unknown bands. A Shimadzu CS-930 densitometer (Shimadzu, Tokyo, Japan) was employed to scan the gel lanes. A plot of densitometric area vs ng standard protein yielded a straight line with a correlation coefficient of 0.96.
Electroblotting of proteins from one-dimensional gel slabs to nitrocellulose sheets was accomplished in a Biorad electroblot cell according to the method of Towbin et al. 25, with minor modifications. Transfer occurred within 4 h at 10 V and 25 °C.
Detection of myelin basic proteins Nitrocellulose sheets were incubated overnight at
25 °C with anti-MBP diluted 1:200 in 0.5% gelatin in Tris buffered saline (TBS: 10 mM Tris-chloride, 150 mM NaCI, pH 7.4) according to the method of Sternberger et al. 23. The freshly prepared reaction mixture contained 0.5 mg/ml 4-chloro-l-naphthol, 20% methanol, 0.03% H202, in TBS. The reaction was terminated by washing the nitrocellulose sheets in deionized water. In control experiments, anti- MBP was replaced with normal rabbit serum.
Detection of glycoproteins One-dimensional slab gels were stained with
periodic acid-Schiff (PAS) reagent according to the method of Zacharius et al. aS. Nitrocellulose sheets were reacted with biotinylated concanavalin A as described by Heslip et al.8; in control experiments, either a-methyl-D-mannoside (200 mM) was added to the concanavalin A solution to block the binding of the lectin, or the nitrocellulose sheets were
227
incubated in TBS alone.
RESULTS
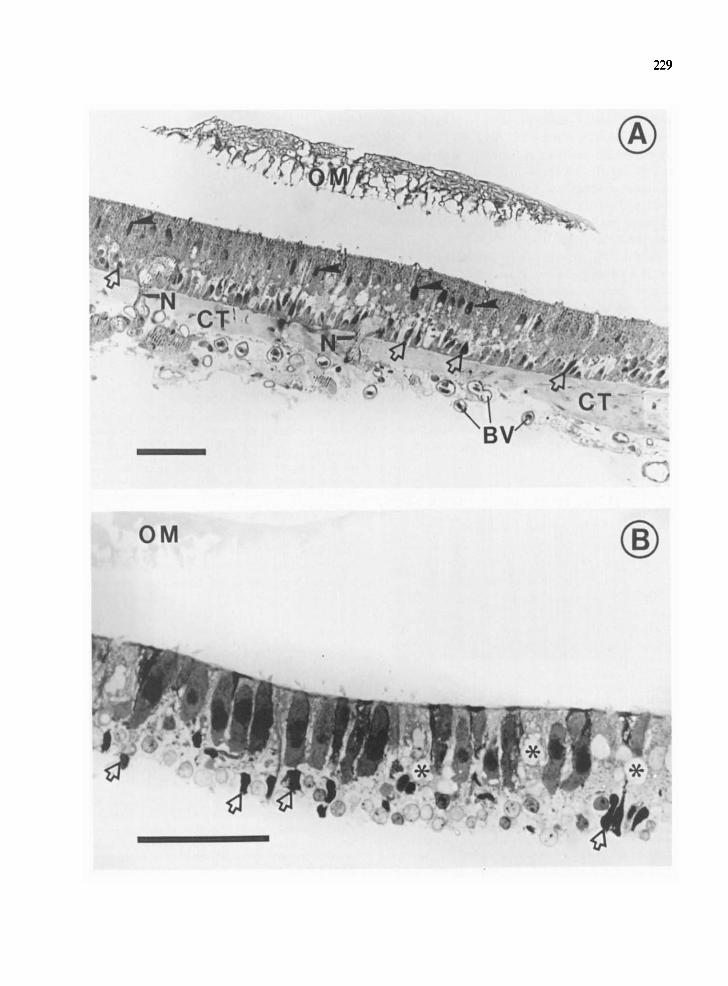
The morphology of samples used in the present study is illustrated in Fig. 1. Fig. 1A shows a section through the saccular sensory epithelium from which external saccular nerve fibers have been removed (cf. Materials and Methods). At the top of the field is the otolithic membrane (OM). The hair cell layer of the saccular epithelium is identified by fine stereociliary arrays on the free surface, and by the upper row of darkly stained nuclei. Fig. 1B illus- trates the isolated hair cell layer from which proteins were extracted. The sensory cell layer has been physically separated from the basal lamina, with hair cells remaining intact. The supporting cells have been disrupted at the level of their nuclei, and vacuolization of the supporting cell cytoplasm (as- terisks) is apparent. Proteins extracted from this tissue, therefore, originated primarily from hair cells, with a small contribution from supporting cell fragments. Total protein for the hair cell layer in Panel B averaged 10.8 + 1.1 pg protein/macula (mean + S.E.M., 7 determinations, 4 experiments, by the method of Lowry et al.14). This value may be compared to 38.6 pg dry weight (or approximately 19.3 pg protein) per macula determined previously for the tissue sample of Fig. 1A, representing a two-fold increase in purification with respect to hair cells.
Fig. 2 shows one-dimensional polyacrylamide gel electrophoretic patterns of the tissue fractions pre- pared in these studies. A protein band with an estimated molecular weight of 13.6 kDa was found to be characteristic of the HC fraction (lower arrow). There was no band at the same position, i.e., just below the 14.4 kDa standard, in any of the lanes representing the other fractions in Fig. 2 (except for N, which was diffusely and darkly stained in this region; discussed below). Another protein band in the high-molecular weight region of the gel, esti- mated at >100 kDa (upper arrow), was present in the other saccular fractions (BL, N, NS), but was virtually absent in the HC fraction.
The densitometric area of the HC band on the gel (Fig. 2, lower arrow) corresponded to approximately 60 ng HC protein in the aliquot applied to the gel

228
lane (5 ¢tl), or 1.2/~g of this protein in the 100/~1 of sample homogenate containing 10 HC layers. Since the average single HC layer (Fig. 1B) contained 10.6 ktg of total protein by Lowry measurements, the unknown HC protein was estimated to account for 0.12/xg/10.6/~g = 1.1% of the total protein in the sensory cell fraction.
Fig. 3 shows one-dimensional electrophoretic pat- terns of the HC fraction, initially homogenized and centrifuged at 10,000 g for 1 h in the absence of SDS to yield supernatant and pellet samples, which were applied to SDS gels. The band for the 13.6 kDa protein is prominent in the supernatant sample (arrow). The migration position of this band was identical to that of the 13.6 kDa band observed previously (Fig. 2) for the entire HC fraction applied to the SDS gel. The 13.6 kDa band was also observed for the pellet fraction.
Fig. 4 indicates that homogenization of the hair cell layer in a mixture of protease inhibitors does not alter the electrophoretic banding pattern of the proteins in the HC fraction (HC lanes 2 and 3). Although the banding patterns of the other saccular fractions (lanes 4-9) were not markedly changed by the presence of protease inhibitors, slight reduction of some lower molecular weight bands occurred.
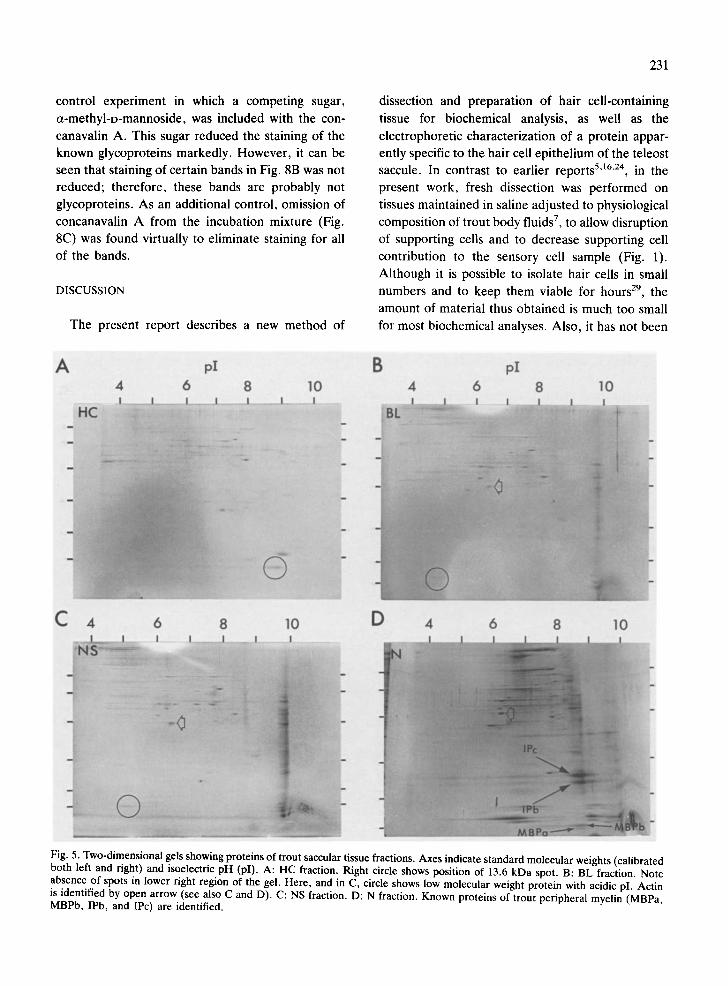
Fig. 5 shows two-dimensional electrophoretic sep- arations of proteins present in the 4 trout saccular tissues. In Fig. 5A (HC fraction), a single spot (lower right circle) with an estimated isoelectric pH of 8.8 was found at the level of the 13.6 kDa band, just below the 14.4 kDa standard, previously re- solved by one-dimensional electrophoresis (Fig. 2). This HC spot was absent in the BL and NS gels (Fig. 5), although an acidic protein of slightly higher molecular weight was detected in these fractions (lower left circles, B and C). For the N fraction, the previously dark, diffuse region below 14.4 kDa (Figs. 2 and 4) was resolved on the 2-dimensional gel
into MBPa, MBPb (11.5 and 13 kDa, res- pectively10), and a lower-molecular weight spot (Fig. 5D). A spot corresponding to the molecular weight of actin was seen in the BL, NS, and N fractions (open arrows), but was not prominent in the HC fraction. In all 4 fractions, the majority of proteins were clustered in the mid-to-high molecular weight range, in the neutral pl region. In particular, 12-15 proteins in this range formed a similar pattern for the BL and NS fractions. The N fraction exhibited a number of spots in the lower molecular weight region of the gel, representing basic proteins known to be present in trout nerve (see below).
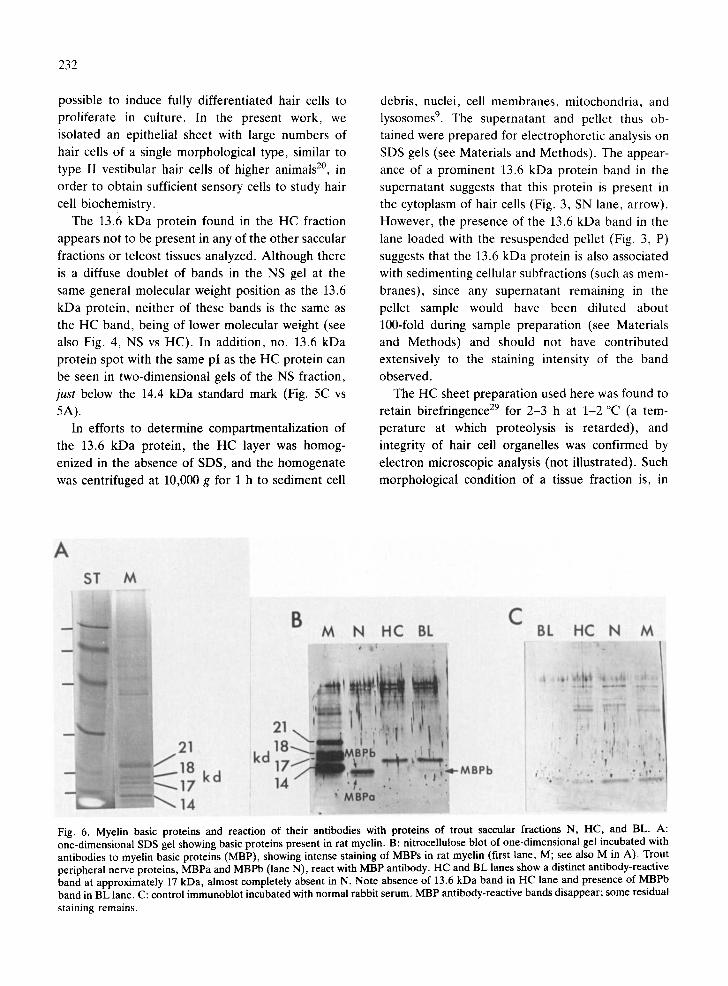
Fig. 6 shows the reaction of antibodies to MBPs with proteins of the 3 trout saccular fractions that are likely to contain MBPs. Fig. 6A illustrates a con- ventionally stained SDS polyacrylamide gel, with protein bands at 14, 17, 18, and 21 kDa, the known myelin basic proteins of the rat 3. Fig. 6B is an immunoblot of a one-dimensional gel of rat myelin and the trout fractions N, HC, and BL. The 4 rat MBP bands in the first lane (M) strongly reacted with MBP antibodies. In the N lane, two b a n d s - MBPa and MBPb of trout peripheral myelin 1° - - reacted strongly with antibody. A band at approxi- mately 17 kDa, present in the HC and BL fractions, demonstrated a pronounced antibody reaction, and in addition, the BL lane showed staining of a band at the position of MBPb. There was no reaction product at the position of the 13.6 kDa protein in the HC lane. When primary antibody was replaced with normal rabbit serum as a control (Fig. 6C), electro- blots showed a complete disappearance of bands representing the 4 rat MBPs (last lane, M), as well as MBPa (N), MBPb (BL and N), and the 17 kDa protein (HC and BL); however, some bands showed non-specific staining.
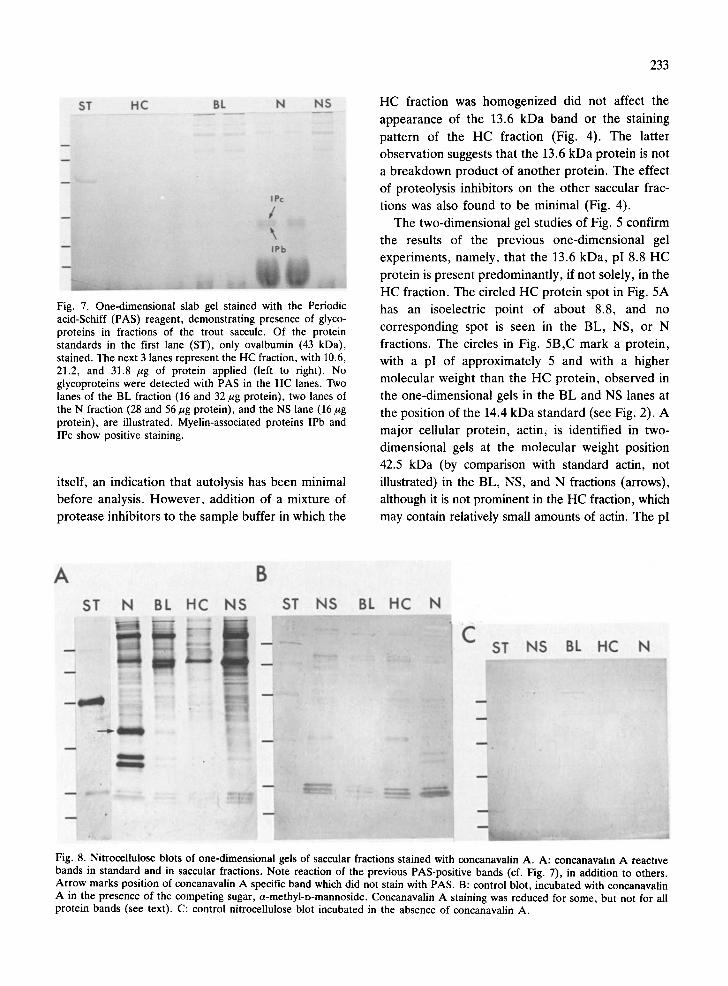
When one-dimensional gels were stained with the PAS reagent to detect glycoproteins, the 13.6 kDa
Fig. 1. A: light micrograph of a transverse section through the saccular macula of the rainbow trout. The displaced otolithic membrane (OM) appears at the top of the field. A row of hair cell nuclei lies beneath the free surface in the upper portion of the sensory epithelium (closed arrowheads) and a row of supporting cell nuclei lies just above the basal lamina in the lower portion of the epithelium. In the underlying connective tissue (CT), nerves (N) pass to and from the sensory epithelium. Blood vessels (BV) contain nucleated erythrocytes. (Cryofixation in liquid propane at liquid nitrogen temperature, Toluidine blue staining; scale = 50 /~m.) B: the saccular hair cell layer used for the extraction of proteins is shown, prepared as described in Materials and Methods. Elongated and darkly stained hair cells are seen distinctly, with arrays of stereocilia at their free surface. The hair cell layer has been detached from underlying structures at the level of the supporting cell nuclei (open arrows); disruption of supporting cell membranes results in vacuolization (asterisks) of supporting cell cytoplasm. (Trump's fixative with Toluidine blue staining; scale = 50 ~m.)
229
®

230
band in the HC fraction did not stain (Fig. 7). Of the
molecular weight s tandards, the glycoprotein, oval-
bumin, s tained positively (ST lane, 43 kDa). Two
glycoproteins, descr ibed for trout per ipheral nerve
as IPb and IPc "J (24 and 26 kDa, respectively, N
lanes), also were identif ied with PAS. In addit ion,
three to 5 protein bands at and above the 94 kDa
region of the BL, N, and NS lanes reacted weakly
with the PAS reagent (Fig. 7).
In o rder to further examine glycoproteins in the
saccular fractions, a more sensitive carbohydrate
reagent , concanavalin A, was employed. Fig. 8A
shows concanaval in A staining of a nitrocellulose
blot of a one-dimensional gel of the trout saccular
fractions. As observed for PAS staining, ovalbumin
(ST lane), IPb and IPc (intense double t in N lane),
and several high molecular weight proteins in the N,
BL, and NS lanes exhibi ted pronounced staining.
The HC fraction also exhibited concanavalin A-
reactive bands in the high molecular weight region.
In addit ion, a prote in band in the N lane which had
ST HC BL N NS B G L ST
kd
HC ST SN P
Fig. 3. One-dimensional SDS slab gel, showing electrophoretic banding patterns of supernatant (SN) and pellet (P) fractions of the HC layer, homogenized and centrifuged at 10,000 g for 1 h in the absence of SDS. Left tick marks indicate positions of standards (ST) in this and all subsequent illustrations, and correspond to those labeled in Fig. 2. Arrow shows position of 13.6 kDa protein band. Quantity of sample applied to SN and P lanes was 5.3 jtg protein in 5 ,ul.
not s tained previously with PAS (Fig. 8A, arrow),
having an es t imated molecular weight of 33 kDa,
reacted strongly with the lectin. Fig. 8B shows a
Fig. 2. One-dimensional slab gel showing protein band associated with hair cell layer (HC) of the trout saccular macula (lower left arrow), compared to proteins of other trout fractions. Note that HC protein lies just below the 14.4 kDa standard. Upper right arrow shows a band characteristic of the non-hair cell saccular fractions. The outermost lanes (ST) show molecular weight standard bands (phosphorylase B, 94 kDa, 321 ng; bovine serum albumin, 67 kDa, 416 ng; ovalbumin, 43 kDa, 738 ng, carbonic anhydrase, 30 kDa, 416 ng; trypsin inhibitor, 20 kDa, 401 ng; a-lactalbumin, 14.4 kDa, 607 ng); 5/~1 of standard protein mixture was applied to the ST lanes for this and all subsequent illustrations. Samples of supernatants from SDS solubilized tissue fractions were applied in the following amounts: hair cell (HC) fraction (5.3 /ag protein in 5/~1); basal lamina (BL) fraction (8/~g in 5/A); saccular nerve (N) fraction (14/~g in 5/A); non-sensory wall (NS) fraction (8/~g in 5 ktl); brain (B) fraction (2.7/~g in 3/~1); gill (G) fraction (2.6 k~g in 3/~1); liver (L) fraction (2.2/~g in 3/~l).
ST HC BL N N$
Fig. 4. One-dimensional slab gel showing electrophoretic banding patterns of proteins in SDS-solubilized saccular fractions prepared in the absence (-) or presence (+) of protease inhibitors (PMSF, leupeptin, and pepstatin). The 13.6 kDa protein (arrow) is present in both HC samples with and without the inhibitors. Sample amounts and volumes applied were the same as for Fig. 2.
control experiment in which a competing sugar,
a-methyl-D-mannoside, was included with the con-
canavalin A. This sugar reduced the staining of the known glycoproteins markedly. However, it can be seen that staining of certain bands in Fig. 8B was not
reduced; therefore, these bands are probably not
glycoproteins. As an additional control, omission of
concanavalin A from the incubation mixture (Fig. 8C) was found virtually to eliminate staining for all
of the bands.
DISCUSSION
The present report describes a new method of
231
dissection and preparation of hair cell-containing
tissue for biochemical analysis, as well as the
electrophoretic characterization of a protein appar-
ently specific to the hair cell epithelium of the teleost saccule. In contrast to earlier reports 5'16"24, in the
present work, fresh dissection was performed on
tissues maintained in saline adjusted to physiological
composition of trout body fluids 7, to allow disruption
of supporting cells and to decrease supporting cell
contribution to the sensory cell sample (Fig. 1).
Although it is possible to isolate hair cells in small
numbers and to keep them viable for hours 29, the
amount of material thus obtained is much too small
for most biochemical analyses. Also, it has not been
A pI B px 4 6 8 10 4 6 8 I0
C 4 6 8 10 D 4 6 8 10
Fig. 5. Two-dimensional gels showing proteins of trout saccular tissue fractions. Axes indicate standard molecular weights (calibrated both left and right) and isoelectric pH (pI). A: HC fraction. Right circle shows position of 13.6 kDa spot. B: BL fraction. Note absence of spots in lower right region of the gel. Here, and in C, circle shows low molecular weight protein with acidic pI. Actin is identified by open arrow (see also C and D). C: NS fraction. D: N fraction. Known proteins of trout peripheral myelin (MBPa, MBPb, IPb, and IPc) are identified.
232
possible to induce fully differentiated hair cells to
proliferate in culture. In the present work, we
isolated an epithelial sheet with large numbers of
hair cells of a single morphological type, similar to
type II vestibular hair cells of higher animals 2°, in
order to obtain sufficient sensory cells to study hair
cell biochemistry.
The 13.6 kDa protein found in the HC fraction
appears not to be present in any of the other saccular
fractions or teleost tissues analyzed. Although there
is a diffuse doublet of bands in the NS gel at the
same general molecular weight position as the 13.6
kDa protein, neither of these bands is the same as
the HC band, being of lower molecular weight (see
also Fig. 4, NS vs HC). In addition, no. 13.6 kDa protein spot with the same pI as the HC protein can
be seen in two-dimensional gels of the NS fraction,
just below the 14.4 kDa standard mark (Fig. 5C vs
5A). In efforts to determine compartmentalization of
the 13.6 kDa protein, the HC layer was homog-
enized in the absence of SDS, and the homogenate
was centrifuged at 10,000 g for 1 h to sediment cell
debris, nuclei, cell membranes, mitochondria, and lysosomes 9. The supernatant and pellet thus ob-
tained were prepared for etectrophoretic analysis on
SDS gels (see Materials and Methods). The appear-
ance of a prominent 13.6 kDa protein band in the
supernatant suggests that this protein is present in the cytoplasm of hair cells (Fig. 3, SN lane, arrow).
However, the presence of the 13.6 kDa band in the
lane loaded with the resuspended pellet (Fig. 3, P)
suggests that the 13.6 kDa protein is also associated
with sedimenting cellular subfractions (such as mem-
branes), since any supernatant remaining in the
pellet sample would have been diluted about
100-fold during sample preparation (see Materials and Methods) and should not have contributed
extensively to the staining intensity of the band
observed. The HC sheet preparation used here was found to
retain birefringence 29 for 2-3 h at 1-2 °C (a tem-
perature at which proteolysis is retarded), and
integrity of hair cell organelles was confirmed by
electron microscopic analysis (not illustrated). Such
morphological condition of a tissue fraction is, in
A ST M
B M N HC BL
C BL HC N M
kd
Fig. 6. Myelin basic proteins and reaction of their antibodies with proteins of trout saccular fractions N, HC, and BL. A: one-dimensional SDS gel showing basic proteins present in rat myelin. B: nitrocellulose blot of one-dimensional gel incubated with antibodies to myelin basic proteins (MBP), showing intense staining of MBPs in rat myelin (first lane, M; see also M in A). Trout peripheral nerve proteins, MBPa and MBPb (lane N), react with MBP antibody. HC and BL lanes show a distinct antibody-reactive band at approximately 17 kDa, almost completely absent in N. Note absence of 13.6 kDa band in HC lane and presence of MBPb band in BL lane. C: control immunoblot incubated with normal rabbit serum. MBP antibody-reactive bands disappear; some residual staining remains.
233
ST HC BL N NS
Fig. 7. One-dimensional slab gel stained with the Periodic acid-Schiff (PAS) reagent, demonstrating presence of glyco- proteins in fractions of the trout saccule. Of the protein standards in the first lane (ST), only ovalbumin (43 kDa), stained. The next 3 lanes represent the HC fraction, with 10.6, 21.2, and 31.8 /~g of protein applied (left to right). No glycoproteins were detected with PAS in the HC lanes. Two lanes of the BL fraction (16 and 32 ~g protein), two lanes of the N fraction (28 and 56/~g protein), and the NS lane (16/~g protein), are illustrated. Myelin-associated proteins IPb and IPc show positive staining.
itself, an indication that autolysis has been minimal
before analysis. However, addition of a mixture of
protease inhibitors to the sample buffer in which the
HC fraction was homogenized did not affect the
appearance of the 13.6 kDa band or the staining
pattern of the HC fraction (Fig. 4). The latter observation suggests that the 13.6 kDa protein is not
a breakdown product of another protein. The effect
of proteolysis inhibitors on the other saccular frac-
tions was also found to be minimal (Fig. 4).
The two-dimensional gel studies of Fig. 5 confirm
the results of the previous one-dimensional gel
experiments, namely, that the 13.6 kDa, pI 8.8 HC
protein is present predominantly, if not solely, in the
HC fraction. The circled HC protein spot in Fig. 5A
has an isoelectric point of about 8.8, and no
corresponding spot is seen in the BL, NS, or N
fractions. The circles in Fig. 5B,C mark a protein,
with a pI of approximately 5 and with a higher
molecular weight than the HC protein, observed in the one-dimensional gels in the BL and NS lanes at
the position of the 14.4 kDa standard (see Fig. 2). A
major cellular protein, actin, is identified in two-
dimensional gels at the molecular weight position
42.5 kDa (by comparison with standard actin, not
illustrated) in the BL, NS, and N fractions (arrows),
although it is not prominent in the HC fraction, which
may contain relatively small amounts of actin. The pI
A B ST N BL HC NS ST N S BL HC N
Fig. 8. Nitrocellulose blots of one-dimensional gels of saccular fractions stained with concanavalin A. A: concanavalin A reactive bands in standard and in saccular fractions. Note reaction of the previous PAS-positive bands (cf. Fig. 7), in addition to others. Arrow marks position of concanavalin A specific band which did not stain with PAS. B: control blot, incubated with concanavalin A in the presence of the competing sugar, a-methyl-D-mannoside. Concanavalin A staining was reduced for some, but not for all protein bands (see text). C: control nitrocellulose blot incubated in the absence of concanavalin A.

234
of the actin spot, about 6.3, does not correspond to that of muscle actin (pI = 5.4); however, its pI value is within the range of that of other actin species, such as the nonmuscle actins (pl values 6 and above27).
Two myelin basic proteins, MBPa and MBPb, with molecular weights of 11.5 and 13 kDa, respec- tively, have been reported for the peripheral nerve
of the rainbow trout m. These myelin proteins have isoelectric points in the range of 9-101. MBPa occurs
as a satellite band below MBPb on SDS gels of isolated peripheral nerve myelin 1°. It was of interest to determine whether the HC protein was a myelin basic protein. Although our trout HC protein is in the molecular weight range of MBPb, it does not appear to be a myelin basic protein, having a slightly
higher molecular weight, 13.6 kDa, and a lower pI, 8.8. However, the strongest evidence for the HC protein not being an MBP comes from its inability to react with antibody to MBPs (Fig. 6).
Jeserich and Waehneldt 1° have shown biochemical
similarities and antibody cross-reactivity among my- elin basic proteins across mammalian and fish spe- cies. Because of such known cross-reactivity, we incubated our one-dimensional gel electroblots with
antibodies against bovine myelin basic proteins. As expected, for the trout nerve fraction, a 13 kDa band, MBPb 1°, and a satellite band, MBPa (N lane, Fig. 6B), reacted with the antiserum. The basal lamina fraction, which lies below the hair cell layer and contains many nerve fibers (Fig. 1A), also
showed the presence of MBPb (BL, Fig. 6B) and trace amounts of MBPa. However, the 13.6 kDa HC
protein, a prominent band by conventional staining (Figs. 2-4), did not show immunoreactivity, suggest- ing that the HC protein is different from either MBPb or MBPa. (The lowest, nonspecifically stain- ing band in Fig. 6C lies well below the molecular weight range of MBPa, MBPb, and the HC protein.) Interestingly, a 17 kDa MBP-reactive band, identi- fied as a minor component in peripheral trout myelin by Jeserich and Waehneldt 1°, appeared as a major component in the trout HC and BL fractions (Fig. 6B). It is tempting to speculate that the distribution of the various MBP species in saccular nerve may change after the nerve enters the saccule, from a distribution containing a majority of MBPa and MBPb to one containing mainly 17 kDa MBP.
The present results clearly demonstrated that the HC protein is not a glycoprotein. The 13.6 kDa band did not stain with PAS reagent 28 (Fig. 7), and failed to bind concanavalin A s (Fig. 8).
We have also noted a protein band in the higher molecular weight region of our gels (Fig. 2, upper
arrow), which was found in the basal lamina, non-sensory epithelium, and saccular nerve, but not in the HC fraction. Although the exact molecular weight of this protein could not be determined because it migrated outside the linear resolving range of the gel (10-70 kDa) 2e, its molecular weight
was estimated to be >100 kDa. The presence of the >100 kDa protein in BL, NS and N, but not HC fractions, suggests that the HC layer is a separate population of cells and is relatively free of the other tissue fractions.
'Tissue specific' or 'marker ' proteins have been reported for various tissues of the central nervous system 4, visual system 13, and olfactory system 21. For
example, a 19 kDa olfactory marker protein, asso- ciated with the primary olfactory neurons as well as with certain brain neurons 2, has been isolated~5; this protein, of unknown function, has now been exten- sively characterized at the molecular level 21. When compared to tissue specific proteins previously re- ported for acousticolateralis organs (e.g., OCP I: 37 kDa24; OCP II: 22.5 kDa24; Xenopus neuromast
protein: 44 kDaS; chick basilar papilla protein: 23 kDa16; auditory nerve proteins: >25 kDa26), the HC
protein described here appears not to correspond, having a different molecular weight and isoelectric point. The HC protein could prove to be a 'marker protein' for the teleost saccular hair cell fraction. Further studies - - e.g., generation of specific anti- bodies for tissue localization, and micro amino acid analysis - - may help to suggest a role for the saccular HC protein described in the present report.
ACKNOWLEDGEMENTS
The authors thank Dr. J. Benjamins and Ms. D. Studinzski for assistance in staining electroblots of myelin basic proteins, and Dr. H. Maisel for advice regarding detection of glycoproteins. This investiga- tion was supported by NIH Grant NS 16166 to
D.G.D.

REFERENCES
1 Althaus, H.-H., Kl6ppner, S., Poehling, H.-M. and Neu- hoff, V., Two-dimensional gel electrophoresis of myelin and oligodendroglial proteins solubilized by a mixture of tetramethylurea and dimethylethyleneurea, Electropho- resis, 4 (1983) 347-353.
2 Baker, H. and Margolis, EL., Olfactory marker protein (OMP)-Iike immunoreactivity in discrete nuclei of rodent brain, Soc. Neurosci. Abstr., 13 (1987) 363.
3 Benjamins, J.A., Morell, P., Hartman, B.K. and Agrawal, H.C., Central nervous system myelin. In A. Lajtha (Ed.), Handbook of Neurochemistry, Vol. 7, 2nd edn., Plenum, New York, 1984, pp. 361-415.
4 Bock, E., Nervous system specific proteins, J. Neuro- chem., 30 (1978) 7-14.
5 Drescher, D.G., Comparison by electrophoresis of pro- teins characteristic of the lateral line and skin of Xenopus laevis, Comp. Biochem. Physiol., 77B (1984) 295-302.
6 Drescher, D.G., Arden, R.L., Drescher, M.J., Kerr, T.P., Hatfield, J.S. and Khan, K.M., Electrophoretic analysis of proteins characteristic of the saccular macula of the rainbow trout, Anat. Rec., 220 (1988) 33A.
7 Drescher, M.J., Drescher, D.G. and Hatfield, J.S., Po- tassium-evoked release of endogenous primary amine- containing compounds from the trout saccular macula and saccular nerve in vitro, Brain Research, 417 (1987) 39-50.
8 Heslip, J., Bagchi, M., Zhang, S., Alousi, S. and Maisel, H., An intrinsic membrane glycoprotein of the lens, Current Eye Res., 5 (1986) 949-958.
9 Hogeboom, G.H., Fractionation of cell components of animal tissues. In S.P. Colowick and N.O. Kaplan (Eds.), Methods in Enzymology, Vol. 1, Academic Press, New York, 1955, pp. 16-19.
10 Jeserich, G. and Waehneldt, T.V., Bony fish myelin: evidence for common major structural glycoproteins in central and peripheral myelin of trout, J. Neurochem., 46 (1986) 525-533.
11 Kodavue Electrophoresis Visualization Kit, Instructions 87-1, Eastman Kodak Company, Rochester, NY, 1987, pp. 1-4.
12 Laemmli, U.K., Cleavage of structural proteins during the assembly of the head of bacteriophage T4, Nature (Lond.), 227 (1970) 680-685.
13 Lai, Y.L., Wiggert, B., Lui, Y.P. and Chader, G.J., Interphotoreceptor retinol-binding proteins: possible trans- port vehicles between compartments of the retina, Nature (Lond.), 298 (1982) 848-849.
14 Lowry, O.H., Rosebrough, N.J., Farr, A.L. and Randall, R.J., Protein measurement with the Folin phenol reagent, J. Biol. Chem., 193 (1951)265-275.
15 Margolis, EL., A brain protein unique to the olfactory
235
bulb, Proc. Natl. Acad. Sci. U.S.A., 69 (1972) 1221-1224. 16 Oberholtzer, J.C., Schneider, M.E., Summers, M.C.,
Saunders, J.C. and Matschinsky, EM., The developmental appearance of a major basilar papilla-specific protein in the chick, Hearing Res., 23 (1986) 161-168.
17 O'Farrell, P.H., High resolution two-dimensional electro- phoresis of proteins, J. Biol. Chem., 250 (1975) 4007-4021.
18 Popper, A.N., Ultrastructure of the auditory regions in the inner ear of the lake whitefish, Science, 192 (1976) 1020-1023.
19 Popper, A.N., A scanning electron microscopic study of the sacculus and lagena of the ears of fifteen species of teleost fishes, J. Morphol., 153 (1977) 397-418.
20 Popper, A.N., Organization of the inner ear and auditory processing. In R.G. Northcutt and R.E. Davis (Eds.), Fish Neurobiology, Vol. 1: Brain Stem and Sense Organs, University of Michigan Press, Ann Arbor, 1983, pp. 125-178.
21 Rogers, K.E., Dasgupta, P., Gubler, U., Grillo, M., Khew-Goodall, Y.S. and Margolis, EL., Molecular cloning and sequencing of a cDNA for olfactory marker protein, Proc. Natl. Acad. Sci. U.S.A., 84 (1987) 1704-1708.
22 Shapiro, A.L., Vifiuela, E. and Maizel, Jr., J.V., Molec- ular weight estimation of polypeptide chains by electro- phoresis in SDS-polyacrylamide gels, Biochem. Biophys. Res. Commun., 28 (1967) 815-820.
23 Sternberger, L.S., Hardy, J.R., P.J., Cuculis, J.J. and Meyer, H.G., The unlabeled antibody enzyme method of immunohistochemistry. Preparation and properties of sol- uble antigen-antibody complex (horseradish peroxidase- antiperoxidase) and its use in identification of spirochetes, J. Histochem. Cytochem., 118 (1970) 315-333.
24 Thalmann, I., Rosenthal, H.L., Moore, B.W. and Thai- mann, R., Organ of Corti-specific polypeptides: OCP-I and OCP-II, Arch. Otorhinolaryngol., 226 (1980) 123-128.
25 Towbin, H., Staehelin, T. and Gordon, J., Electrophoretic transfer of proteins from polyacrylamide gels to nitrocel- lulose sheets: procedure and some applications, Proc. Natl. Acad. Sci. U.S.A., 76 (1979) 4350-4354.
26 Wenthold, R.J. and McGarvey, M.L., Different polypep- tides are rapidly transported in auditory and optic neurons, J. Neurochem., 39 (1982) 27-35.
27 Whalen, R.G., Butler-Browne, G.S. and Gros, F., Protein synthesis and actin heterogeneity in calf muscle cells in culture, Proc. Nat. Acad. Sci. U.S.A., 73 (1976) 2018- 2022.
28 Zacharius, R.M., Zell, T.E., Morrison, J.H. and Wood- lock, J.J., Glycoprotein staining following electrophoresis on acrylamide gels, Anal. Biochem., 30 (1969) 149-152.
29 Zajic, G. and Schacht, J., Comparison of isolated outer hair cells from five mammalian species, Hearing Res., 26 (1987) 249-256.





























